Accepted by P. Velazco: 14 Oct. 2013; published: 1 Nov. 2013 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2013 Magnolia Press Zootaxa 3731 (4): 589–598 www.mapress.com/zootaxa/ Article 589 http://dx.doi.org/10.11646/zootaxa.3731.4.10 http://zoobank.org/urn:lsid:zoobank.org:pub:117F6046-A2A9-4743-8B87-DE18DDA1C404 Is Leopoldamys neilli (Rodentia, Muridae) a synonym of Leopoldamys herberti? A reply to Balakirev et al. (2013) ALICE LATINNE 1 , YANNICK CHAVAL 2 , SURACHIT WAENGSOTHORN 3 , PRATEEP ROJANADILOK 4 , KRAIRAT EIAMAMPAI 5 , KRIANGSAK SRIBUAROD 6 , VINCENT HERBRETEAU 7 , SERGE MORAND 8,9 & JOHAN R. MICHAUX 1, 2 1 Conservation Genetics Unit, University of Liege, Liège, Belgium. E-mail: [email protected] 2 INRA, UMR1062 CBGP, Campus international de Baillarguet, Montferrier-sur-Lez, France 3 Environment and Resources Technology Department, Thailand Institute of Scientific and Technological Research, Pathum Thani, Thailand 4 Doi Chiangdao Wildlife Research Station, Chiang Mai, Thailand 5 Bung Boraphet Wildlife Research Station, Nakhon Sawan, Thailand 6 Khlong Saeng Wildlife Research Station, Surat Thani, Thailand 7 IRD, UMR ESPACE-DEV (IRD, UM2, UAG, UR), Station SEAS-OI, F-97410 Saint-Pierre, France 8 Institut des Sciences de l’Evolution, CNRS-IRD-UM2, Université de Montpellier 2, F-34093, Montpellier, France 9 Walai Rukhavej Botanical Research Institute, Mahasarakham University, Maha Sarakham 44150 Thailand Abstract Recently, Balakirev et al. (2013) presented a taxonomic revision of the genus Leopoldamys based on phylogenetic analy- ses. They identified five main Leopoldamys genetic lineages and suggested to rename several of them. According to these authors, the genetic lineage previously thought to belong to L. edwardsi (lineage L1) should be assigned to L. revertens while L. neilli (lineage L2) should be considered as a junior synonym of L. herberti. Using molecular and morphological data from a large sampling of Leopoldamys specimens, the aim of the present study was to investigate the taxonomic status of L. herberti and L. neilli. This study reveals that, contrary to Balakirev et al.’s statement, both genetic lineages L1 and L2 occur in Nakhon Ratchasima Province, close to the type locality of L. herberti. We also show that the external mea- surements and color pattern of L. herberti are highly similar to those of L1 specimens but are not consistent with the mor- phology of L2 specimens. Therefore these results strongly suggest that L. herberti should be assigned to the genetic lineage L1. Consequently L. neilli should not be considered as a junior synonym of L. herberti and this study confirms that the appropriate name of the genetic lineage L2 is L. neilli. Moreover, as our results show that L. herberti should be assigned to the lineage L1, this name has nomenclatural priority over L. revertens, the species name suggested by Bala- kirev et al. (2013) for this lineage. Key words: Leopoldamys, long-tailed giant rats, Thailand, Indochina, Murinae rodents Introduction The genus Leopoldamys Ellerman, 1941 comprises several species of long-tailed giant rats living in forests of Southeast Asia, and extending into northeast India and southern China. Recent studies of the taxonomy of Murinae rodents in Thailand based on molecular data recovered three main Leopoldamys phylogenetic lineages that were assigned to the three Leopoldamys species traditionally recognized in Thailand (Latinne et al. 2012, 2013; Pages et al. 2010). One of these lineages is distributed in southern Thailand and has been identified as L. sabanus (Thomas, 1887), another one is present in northern Thailand and has been assigned to L. edwardsi (Thomas, 1882) whereas the last one has been found exclusively in limestone karsts of continental Thailand and was identified as L. neilli (Marshall, 1977). The last-mentioned identification was supported by Pages et al. (2010) using sequences obtained from the holotype specimen of this species. Recently, Balakirev et al. (2013) presented a quite different taxonomic arrangement of the genus. They identified five main Leopoldamys genetic lineages and suggested to rename several TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2013 Magnolia Press
Zootaxa 3731 (4): 589–598
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3731.4.10
http://zoobank.org/urn:lsid:zoobank.org:pub:117F6046-A2A9-4743-8B87-DE18DDA1C404
Is Leopoldamys neilli (Rodentia, Muridae) a synonym of Leopoldamys herberti?
A reply to Balakirev et al. (2013)
ALICE LATINNE1, YANNICK CHAVAL2, SURACHIT WAENGSOTHORN3, PRATEEP ROJANADILOK4,
KRAIRAT EIAMAMPAI5, KRIANGSAK SRIBUAROD6, VINCENT HERBRETEAU7, SERGE MORAND8,9
& JOHAN R. MICHAUX1, 2
1Conservation Genetics Unit, University of Liege, Liège, Belgium. E-mail: [email protected], UMR1062 CBGP, Campus international de Baillarguet, Montferrier-sur-Lez, France3Environment and Resources Technology Department, Thailand Institute of Scientific and Technological Research, Pathum Thani,
Thailand4Doi Chiangdao Wildlife Research Station, Chiang Mai, Thailand5Bung Boraphet Wildlife Research Station, Nakhon Sawan, Thailand6Khlong Saeng Wildlife Research Station, Surat Thani, Thailand7IRD, UMR ESPACE-DEV (IRD, UM2, UAG, UR), Station SEAS-OI, F-97410 Saint-Pierre, France8Institut des Sciences de l’Evolution, CNRS-IRD-UM2, Université de Montpellier 2, F-34093, Montpellier, France9Walai Rukhavej Botanical Research Institute, Mahasarakham University, Maha Sarakham 44150 Thailand
Abstract
Recently, Balakirev et al. (2013) presented a taxonomic revision of the genus Leopoldamys based on phylogenetic analy-
ses. They identified five main Leopoldamys genetic lineages and suggested to rename several of them. According to these
authors, the genetic lineage previously thought to belong to L. edwardsi (lineage L1) should be assigned to L. revertens
while L. neilli (lineage L2) should be considered as a junior synonym of L. herberti. Using molecular and morphological
data from a large sampling of Leopoldamys specimens, the aim of the present study was to investigate the taxonomic status
of L. herberti and L. neilli. This study reveals that, contrary to Balakirev et al.’s statement, both genetic lineages L1 and
L2 occur in Nakhon Ratchasima Province, close to the type locality of L. herberti. We also show that the external mea-
surements and color pattern of L. herberti are highly similar to those of L1 specimens but are not consistent with the mor-
phology of L2 specimens. Therefore these results strongly suggest that L. herberti should be assigned to the genetic
lineage L1. Consequently L. neilli should not be considered as a junior synonym of L. herberti and this study confirms
that the appropriate name of the genetic lineage L2 is L. neilli. Moreover, as our results show that L. herberti should be
assigned to the lineage L1, this name has nomenclatural priority over L. revertens, the species name suggested by Bala-
kirev et al. (2013) for this lineage.
Key words: Leopoldamys, long-tailed giant rats, Thailand, Indochina, Murinae rodents
Introduction
The genus Leopoldamys Ellerman, 1941 comprises several species of long-tailed giant rats living in forests of
Southeast Asia, and extending into northeast India and southern China. Recent studies of the taxonomy of Murinae
rodents in Thailand based on molecular data recovered three main Leopoldamys phylogenetic lineages that were
assigned to the three Leopoldamys species traditionally recognized in Thailand (Latinne et al. 2012, 2013; Pages et
al. 2010). One of these lineages is distributed in southern Thailand and has been identified as L. sabanus (Thomas,
1887), another one is present in northern Thailand and has been assigned to L. edwardsi (Thomas, 1882) whereas
the last one has been found exclusively in limestone karsts of continental Thailand and was identified as L. neilli
(Marshall, 1977). The last-mentioned identification was supported by Pages et al. (2010) using sequences obtained
from the holotype specimen of this species. Recently, Balakirev et al. (2013) presented a quite different taxonomic
arrangement of the genus. They identified five main Leopoldamys genetic lineages and suggested to rename several
Accepted by P. Velazco: 14 Oct. 2013; published: 1 Nov. 2013 589

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
of them. According to these authors, the Indochinese genetic lineage previously thought to belong to L. edwardsi
(lineage L1) should be assigned to L. revertens (Robinson & Kloss, 1922) while L. neilli (lineage L2) should be
considered as a junior synonym of L. herberti (Kloss, 1916).
Leopoldamys herberti, described from a single specimen from Nakhon Ratchasima Province (central
Thailand), was considered by Musser & Carleton (2005) as a synonym of L. sabanus. However, both Balakirev et
al. (2013) and Latinne et al. (2013) have showed that L. sabanus is a Sundaic species that is restricted in Indochina
to southern Thailand (with a northern range limit in Kanchanaburi Province). According to these results, L. herberti
should therefore not be considered as a synonym of L. sabanus. Furthermore, Balakirev et al. (2013) suggested that
the external characteristics of L. herberti are strongly consistent with the morphological description of L. neilli.
They also claimed that the genetic lineage assigned to L. neilli (lineage L2) was the sole Leopoldamys lineage
occurring in Nakhon Ratchasima Province where the holotype of L. herberti was collected. Accordingly, these
authors speculated that L. neilli was a junior synonym of herberti. However, Balakirev et al. (2013) did not
compare the external morphology of the Leopoldamys lineages occurring in Thailand with the holotype of L.
herberti neither did they check whether other genetic Leopoldamys genetic lineages might exist in Nakhon
Ratchasima Province. For these reasons, the decision of Balakirev et al. (2013) to consider L. neilli as a junior
synonym of L. herberti appears premature.
Using molecular and morphological data from a large sampling of Leopoldamys specimens from Thailand and
Indochina, the aim of the present study was to investigate the taxonomic status of L. herberti and L. neilli by
answering the following questions: Is the lineage L2 the sole Leopoldamys lineage occurring in Nakhon
Ratchasima Province? Is the external morphology of the type specimen of L. herberti consistent with the
morphology of L2 specimens and the holotype of L. neilli specimens?
Material and methods
Sampling. One hundred twenty-one Leopoldamys samples collected by our team in continental Thailand,
Cambodia and Lao PDR were used in this study. Several of them were previously included in the studies of Pages
et al. (2010), Latinne et al. (2012, 2013), and Balakirev et al. (2013). Two museum specimens from Nakhon
Ratchasima Province (Sakaerat, Pak Thong Chai district) belonging to the rodent collection of the Centre for Thai
National Reference Collections (CTNRC) (reference numbers: CTNRC 54-1549 and CTNRC 54-1550) were also
added to our dataset. To allow an accurate morphological comparison with the L. herberti holotype specimen (an
adult male), all samples included in our morphological study were adult males that had reached sexual maturity.
Molecular identification of Leopoldamys specimens. To avoid species misidentifications, all our samples
were unambiguously identified using DNA sequencing and phylogenetic analysis.
Fresh samples (tissues in ethanol): The cytochrome b gene (Cytb) and/or the cytochrome c oxidase subunit I
gene (COI) were amplified for all the fresh samples as described in Pages et al. (2010) and Latinne et al. (2013).
Haplotypes were identified using ARLEQUIN 3.11 (Excoffier et al. 2005). Phylogenetic reconstructions were
performed on the combined dataset using the Maximum Likelihood (ML) and Bayesian inference (BI) approaches.
Maxomys and Niviventer sequences were used as outgroups in our phylogenetic trees. The most suitable model of
DNA substitution for each locus and dataset was determined using MODELTEST 3.0 (Posada & Crandall 1998)
according to the Akaike Information Criterion (AIC). ML analyses were performed using PhyML 3.0 (Guindon et
al. 2010). Robustness of the tree was assessed by 1000 bootstrap replicates. MRBAYES 3.1.1. (Ronquist &
Huelsenbeck 2003) was used to perform Bayesian analyses. Metropolis-coupled Markov chain Monte Carlo
(MCMC) sampling was performed with 5 chains run for 3 million generations with one tree sampled every 1000
generations, using default parameters as starting values. A 50% Majority-rule consensus tree was then generated in
PAUP 4.0b10 with burn-in values of 100,000 generations (Swofford 1998).
Museum samples (dried skins): As museum samples contained only small quantity of degraded DNA, we
amplified a 85 base pairs (bp) fragment of the Cytb gene that proved to be suitable for amplification of ancient
DNA and to allow to discriminate among most vertebrate species (Teletchea et al. 2008) and more specifically
Rattini species (Galan et al. 2012; Latinne et al. 2013; Pages et al. 2010). DNA extraction was performed using the
QIAamp DNA Micro Kit (Qiagen) following the protocol for isolation of genomic DNA from tissues in a
physically remote area in ancient DNA laboratory where Leopoldamys samples were never processed before this
LATINNE ET AL.590 · Zootaxa 3731 (4) © 2013 Magnolia Press
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
study. The precautions and criteria pertinent for ancient DNA studies were observed during all laboratory
procedures (Gilbert et al. 2005). Amplifications were carried out using the primers LLedw (5’-
GATAAAATTCCATTCCACCC-3’) and HLedw (5’-TAATTGTCTGGGTCTCC-3’) following the protocol
described in Latinne et al. (2013). The 85 bp Cytb fragment obtained from our museum samples was then
compared with other Leopoldamys sequences to identify diagnostically informative attributes allowing
discrimination between Leopoldamys lineages.
Morphological analysis. In order to investigate the morphological differences between the Leopoldamys
lineages L1 and L2, size differences among them were assessed using analyses of variance (one-way ANOVA) on
three linear measurements (head+body length, tail length, hind foot length) of adult males. Our sampling for this
morphological analysis comprises 33 L1 specimens (including the two museum samples from Nakhon Ratchasima)
and 92 L2 specimens (including two paratypes of L. neilli from the United States National Museum of Natural
History [NMNH], catalogue numbers: NMNH 533481, NMNH 533482). The measurements of the L. herberti
holotype specimen (available in Kloss [1916]) and those of the L. neilli holotype specimen (deposited in the
collection of the CTNRC [Waengsothorn et al. 2009]) were then compared with those of L1 and L2 specimens
using box-and-whisker plots.
Results
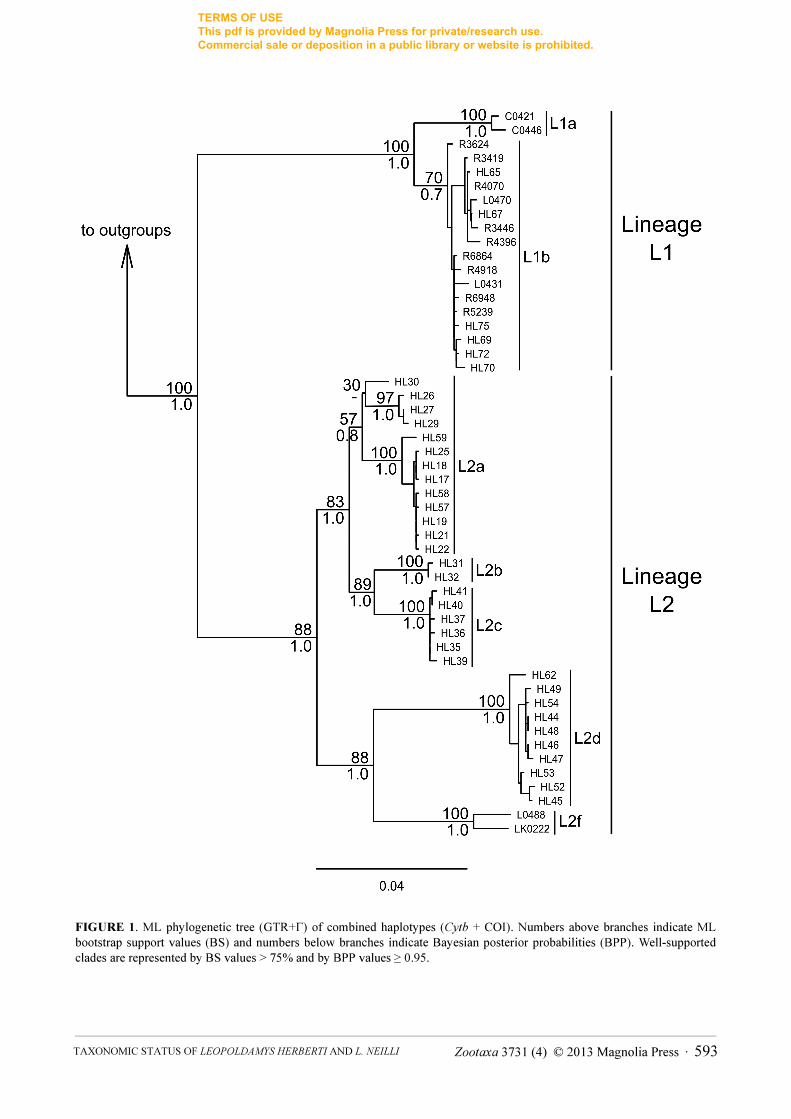
Phylogenetic analysis. Cytochrome b and COI sequences were obtained for 103 and 119 Leopoldamys samples,
respectively. The final alignment includes 52 combined haplotypes (Cytb + COI) (see Table 1 for GenBank
accession numbers of haplotypes). The ML and BI trees of the combined dataset give congruent results and retrieve
two main lineages, the lineages L1 and L2 already evidenced by Balakirev et al. (2013) (Fig. 1). These well-
supported lineages are subdivided into the sublineages L1a (southern Indochina), L1b (northern Indochina), L2a
(northern and northeastern Thailand), L2b (central Thailand), L2c (central Thailand), L2d (western Thailand) and
L2f (Lao PDR) following the nomenclature of Balakirev et al. (2013).
TABLE 1. Lineage, haplotype ID numbers, and GenBank accession numbers for the Leopoldamys samples used in this
study.
Lineage Haplotype Country Cytb COI Source
L1a C0421 Cambodia / KF577939 This study
C0446 Cambodia / KF577940 This study
L1b R3419 Thailand HM217414 HM217543 Pages et al. 2010
HL65 Thailand KC010034 KC010175 Latinne et al. 2013
HL67 Thailand KC010036 KC010175 Latinne et al. 2013
HL69 Thailand KC010038 KC010177 Latinne et al. 2013
HL70 Thailand KC010039 KC010178 Latinne et al. 2013
HL72 Thailand KC010041 KC010177 Latinne et al. 2013
HL75 Thailand KC010043 KC010177 Latinne et al. 2013
R3446 Thailand / KF577944 This study
R3624 Thailand KF577948 KF577934 This study
R4070 Thailand KF577949 KF577935 This study
R4396 Thailand / KF577945 This study
R4918 Thailand / KF577946 This study
R5239 Thailand KF577950 KF577936 This study
R6864 Thailand KF577951 KF577937 This study
R6948 Thailand KF577952 KF577938 This study
L0431 Lao PDR / KF577941 This study
......continued on the next page
Zootaxa 3731 (4) © 2013 Magnolia Press · 591TAXONOMIC STATUS OF LEOPOLDAMYS HERBERTI AND L. NEILLI

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Museum samples identification. The sequences of the 85 bp fragment amplified for the two museum samples
from Nakhon Ratchasima Province (CTNRC 54-1549, 54-1550) are similar and identical to one of the four L1
variants of this fragment (L1b-1) (Fig. 2). This variant is distributed in northern and northeastern Thailand. The
other three L1 variants (L1b-2, L1b-3, L1b-4) differ from the museum sequences by only one base pair. The L2
variants are much more divergent and differ from the museum sequences by at least 10 base pairs (Fig. 2). The
sequence from the L. neilli holotype is identical to one of the L2 variants (L2b) distributed in central Thailand.
Three pure characteristic attributes (attributes that exist across all variants of a single clade and never in any other
clade according to the definition of Sarkar et al. [2008]) have been identified between the lineages L1 and L2 and
allow unambiguous discrimination of these lineages (Fig. 2).
TABLE 1. (Continued)
Lineage Haplotype Country Cytb COI Source
L0470 Lao PDR / KF577942 This study
L2a HL17 Thailand HM219591 HM219573 Latinne et al. 2013
HL18 Thailand HM219592 HM219573 Latinne et al. 2013
HL19 Thailand HM219593 HM219573 Latinne et al. 2013
HL21 Thailand HM219594 HM219573 Latinne et al. 2013
HL22 Thailand HM219595 HM219573 Latinne et al. 2013
HL25 Thailand HM219592 HM219573 Latinne et al. 2013
HL26 Thailand HM219597 HM219577 Latinne et al. 2013
HL27 Thailand HM219598 HM219578 Latinne et al. 2013
HL29 Thailand JQ081357 HM219578 Latinne et al. 2013
HL30 Thailand HM219599 HM219579 Latinne et al. 2013
HL57 Thailand HM219593 JQ081314 Latinne et al. 2013
HL58 Thailand HM219593 JQ081315 Latinne et al. 2013
HL59 Thailand JQ081365 JQ081316 Latinne et al. 2013
L2b HL31 Thailand HM219600 HM219580 Latinne et al. 2013
HL32 Thailand HM219601 HM219580 Latinne et al. 2013
L2c HL35 Thailand HM219603 HM219583 Latinne et al. 2013
HL36 Thailand HM219603 HM219581 Latinne et al. 2013
HL37 Thailand HM219604 HM219583 Latinne et al. 2013
HL39 Thailand HM219603 HM219582 Latinne et al. 2013
HL40 Thailand HM219605 HM219583 Latinne et al. 2013
HL41 Thailand HM219605 JQ081310 Latinne et al. 2013
L2d HL44 Thailand HM219606 HM219586 Latinne et al. 2013
HL45 Thailand HM219607 HM219587 Latinne et al. 2013
HL46 Thailand HM219610 HM219586 Latinne et al. 2013
HL47 Thailand JQ081362 HM219588 Latinne et al. 2013
HL48 Thailand HM219606 HM219584 Latinne et al. 2013
HL49 Thailand HM219608 HM219586 Latinne et al. 2013
HL52 Thailand HM219607 HM219585 Latinne et al. 2013
HL53 Thailand HM219611 HM219586 Latinne et al. 2013
HL54 Thailand HM219612 HM219586 Latinne et al. 2013
HL62 Thailand JQ081368 JQ081318 Latinne et al. 2013
L2f LK0222 Lao PDR KF577947 KF577933 This study
L0488 Lao PDR / KF577943 This study
LATINNE ET AL.592 · Zootaxa 3731 (4) © 2013 Magnolia Press
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 1. ML phylogenetic tree (GTR+Γ) of combined haplotypes (Cytb + COI). Numbers above branches indicate ML
bootstrap support values (BS) and numbers below branches indicate Bayesian posterior probabilities (BPP). Well-supported
clades are represented by BS values > 75% and by BPP values ≥ 0.95.
Zootaxa 3731 (4) © 2013 Magnolia Press · 593TAXONOMIC STATUS OF LEOPOLDAMYS HERBERTI AND L. NEILLI

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 2. Comparison of the 85 bp Cytb fragment obtained from our museum samples (CTNRC 54-1549, 54-1550) with
lineages L1 and L2 and the L. neilli holotype specimen (sequence from Pages et al. [2010], GenBank accession number:
HM235947). Identical positions as those of the sequences from museum samples are indicated by dots. Pure characteristic
attributes allowing discrimination between lineages L1 and L2 are outlined in black.
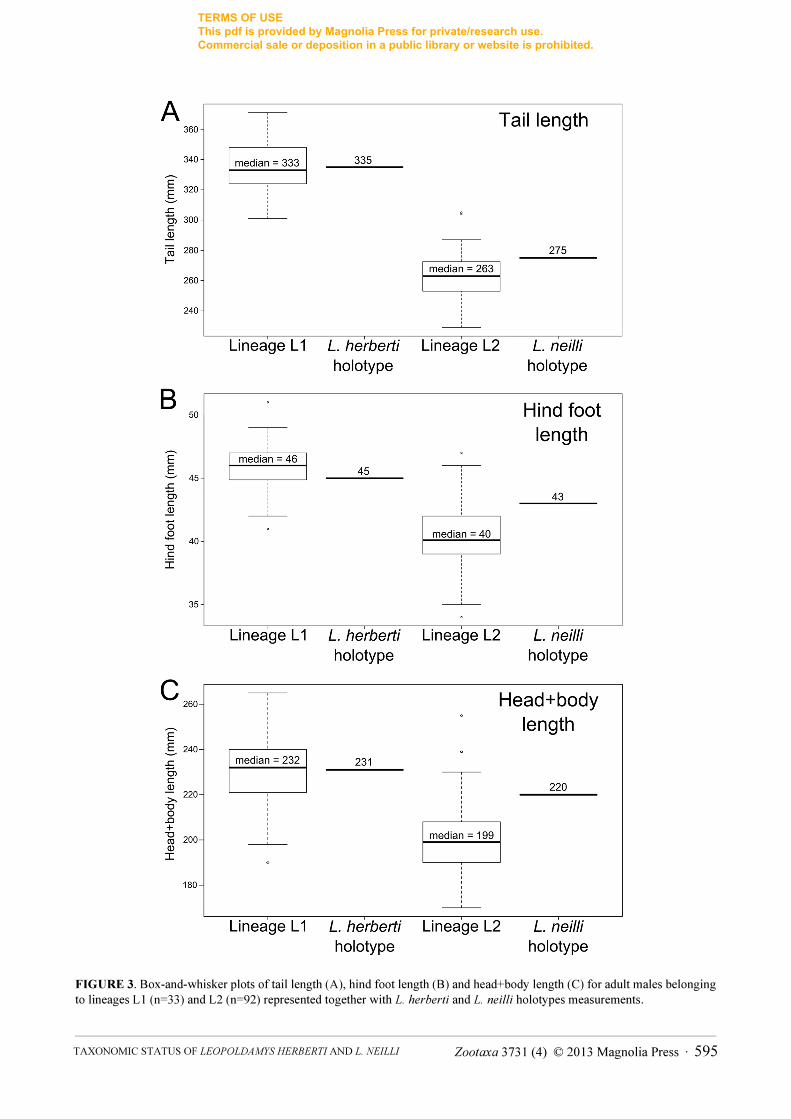
Morphological analysis. All external measurements of adult males show highly significant differences
between the lineages L1 and L2 according to the ANOVAs of tail length (F = 495.03 – p < 0.0001 – df = 1), hind
foot length (F = 146.43 – p < 0.0001 – df = 1) and head+body length (F = 83.54 – p < 0.0001 – df = 1). For each of
these three measurements, individuals of lineage L1 average significantly larger than those of lineage L2.
Differences in tail length, hind foot length and head+body length distinguish 96.8%, 91.2% and 87.2% of adult
males for the two genetic lineages, respectively. Therefore, these three measurements and especially tail length, can
be used to discriminate the lineages L1 and L2 and assign the holotype L. herberti to one of them.
According to these results, the external measurements of the L. herberti holotype specimen are undoubtedly
consistent with those of the lineage L1 rather than lineage L2 (Fig. 3). Indeed tail length of the L. herberti holotype
clearly falls within the range of L1 specimens and is much larger than observed for L2 specimens. Hind foot length
and head+body length of L. herberti are also highly similar to the median values of L1 specimens and larger than
the median values of L2 specimens. All measurements of the L. neilli holotype specimen are similar to those of L2
specimens.
Discussion
Is the lineage L2 the sole Leopoldamys lineage occurring in Nakhon Ratchasima province?
Balakirev et al. (2013) claimed that genetic lineage L2 (assigned to L. neilli) was the sole Leopoldamys lineage
occurring in Nakhon Ratchasima Province because no other genetic lineages have been recorded in this region
despite the considerable number of localities sampled in Latinne et al. (2011). However, the present study clearly
reveals the presence of L1 specimens in Nakhon Ratchasima Province, based on two museum samples identified
using a small Cytb fragment (Fig. 2). These specimens were captured in Sakaerat area, in the Pak Thong Chai
district of Nakhon Ratchasima Province, about sixty kilometers away from Pak Chong where the holotype of L.
herberti was collected (Kloss 1916). Therefore, our results demonstrate that both lineages L1 and L2 occur in the
vicinity of the L. herberti type locality.
Is the external morphology of Leopoldamys herberti consistent with the morphology of L. neilli specimens
(lineage L2)?
The external measurements of the holotype of Leopoldamys herberti and especially its tail length clearly
demonstrate its morphological similarity to specimens of lineage L1 and distinction from specimens of lineage L2
(Fig. 3). The external description of L. herberti also closely matches the color pattern of our L1 specimens.
According to Kloss (1916), the upperparts of L. herberti are “clay-colour streaked with black by the tips of the
hairs. This grizzling is most pronounced dorsally. The limbs are duller and browner. Top of muzzle and a ring
round the eye are pale clove-brown.” Its underparts are “white, this colour extending over the upper lip to include
the tip of the nose and part of the vibrissae roots, projecting upwards to reach the dark eye-ring.” Its feet are “white
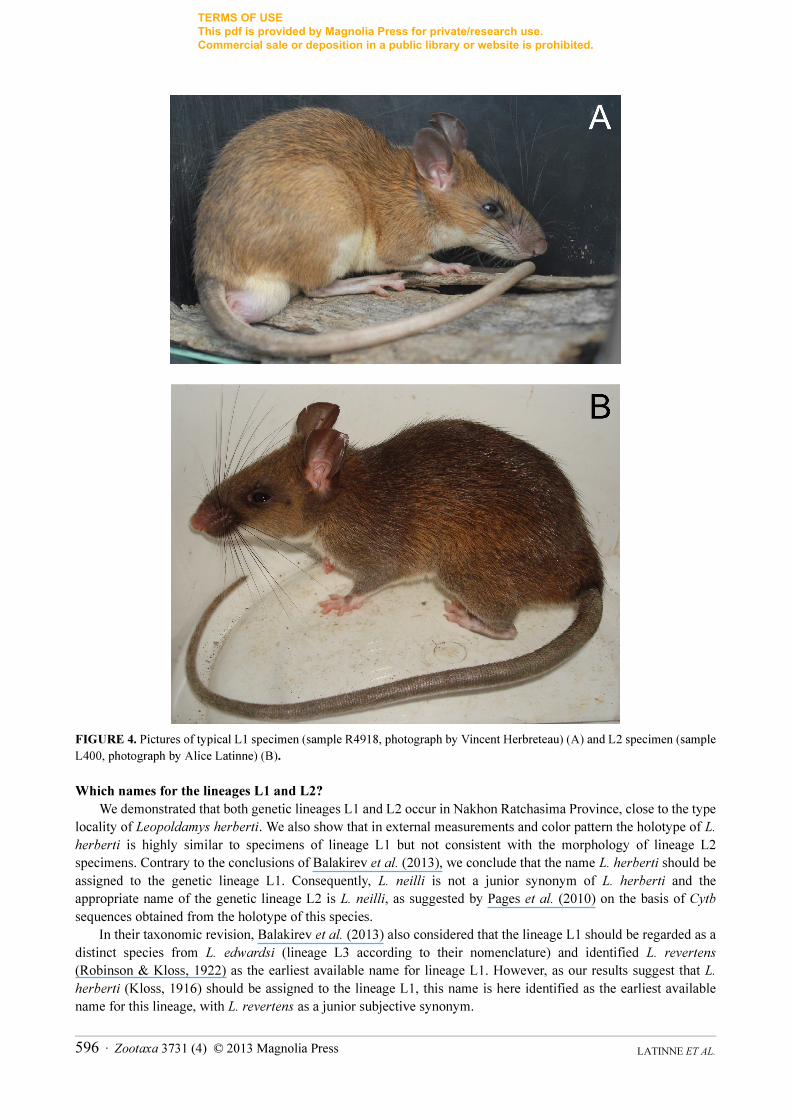
with brown centres” and its tail is bicolored (Kloss 1916). Exactly similar color pattern has been observed on our
L1 specimens while the typical L2 specimens are duller and darker brown and the white fur on their upper lip does
not reach their vibrissae or eye-ring (Fig. 4). Therefore, based on its external measurements and color pattern, it is
clear that the holotype of L. herberti should be associated with lineage L1 rather than with lineage L2.
LATINNE ET AL.594 · Zootaxa 3731 (4) © 2013 Magnolia Press
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 3. Box-and-whisker plots of tail length (A), hind foot length (B) and head+body length (C) for adult males belonging
to lineages L1 (n=33) and L2 (n=92) represented together with L. herberti and L. neilli holotypes measurements.
Zootaxa 3731 (4) © 2013 Magnolia Press · 595TAXONOMIC STATUS OF LEOPOLDAMYS HERBERTI AND L. NEILLI
TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
FIGURE 4. Pictures of typical L1 specimen (sample R4918, photograph by Vincent Herbreteau) (A) and L2 specimen (sample
L400, photograph by Alice Latinne) (B).
Which names for the lineages L1 and L2?
We demonstrated that both genetic lineages L1 and L2 occur in Nakhon Ratchasima Province, close to the type
locality of Leopoldamys herberti. We also show that in external measurements and color pattern the holotype of L.
herberti is highly similar to specimens of lineage L1 but not consistent with the morphology of lineage L2
specimens. Contrary to the conclusions of Balakirev et al. (2013), we conclude that the name L. herberti should be
assigned to the genetic lineage L1. Consequently, L. neilli is not a junior synonym of L. herberti and the
appropriate name of the genetic lineage L2 is L. neilli, as suggested by Pages et al. (2010) on the basis of Cytb
sequences obtained from the holotype of this species.
In their taxonomic revision, Balakirev et al. (2013) also considered that the lineage L1 should be regarded as a
distinct species from L. edwardsi (lineage L3 according to their nomenclature) and identified L. revertens
(Robinson & Kloss, 1922) as the earliest available name for lineage L1. However, as our results suggest that L.
herberti (Kloss, 1916) should be assigned to the lineage L1, this name is here identified as the earliest available
name for this lineage, with L. revertens as a junior subjective synonym.
LATINNE ET AL.596 · Zootaxa 3731 (4) © 2013 Magnolia Press

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Conclusion
Five main Leopoldamys genetic lineages have been delimitated and named by Balakirev et al. (2013) in their
taxonomic revision of the genus. Based on the results of the present study, we propose to revise their nomenclature
as follows: lineage L1 = L. herberti (instead of L. revertens), L2 = L. neilli (instead of L. herberti), L3 = L.
edwardsi, L4 = L. milleti, and L5 = L. sabanus. Leopoldamys ciliatus and L. siporanus, two additional
Leopoldamys species recognized by Musser & Carleton (2005), were not included in the study of Balakirev et al.
(2013) or in our study. Future taxonomic studies based on independent data (mitochondrial and nuclear markers,
morphology) and geographically broader large sampling will be required to confirm these propositions and further
improve the taxonomy of the Leopoldamys genus.
Acknowledgements
First of all, we warmly thank all our collaborators who made sample collection in the field possible and especially
Boonchai Tontan, Sathaporn Jittapalapong and Kittipong Chaisiri. We also thank Marie Pages for her advices about
extraction and amplification of DNA from museum samples. This work was supported by a Belgian FRS-FNRS
(Fonds de la Recherche Scientifique) fellowship to A. Latinne (“Aspirant”) and to J.R. Michaux (“Maître de
recherches”), and a financial grant from the Belgian FRS-FNRS (“crédits pour brefs séjours à l’étranger” to A.
Latinne and J.R. Michaux and credits from the “Fonds de la Recherche Fondamentale Collective (FRFC)” to J.R.
Michaux), from the University of Liège (Patrimoine) and from the Communauté française de Belgique. A. Latinne
also thanks VOCATIO (Belgian Foundation of Vocation) for its financial support. This study is part of the
“CERoPath project” (Community Ecology of Rodents and their Pathogens in South-East Asia: effects of
biodiversity changes and implications in health ecology, www.ceropath.org), ANR Biodiversity ANR 07 BDIV
012, and the “BiodivHealthSEA project”, ANR CP&ES 11 CPEL 002, both funded by the French National Agency
for Research.
References
Balakirev, A.E., Abramov, A.V. & Rozhnov, V.V. (2013) Revision of the genus Leopoldamys (Rodentia, Muridae) as inferred
from morphological and molecular data, with a special emphasis on the species composition in continental Indochina.
Zootaxa, 3640, 521–549.
http://dx.doi.org/10.11646/zootaxa.3640.4.2
Ellerman, J.R. (1941) The families and genera of living rodents. Vol. 2. Family Muridae. British Museum (Natural History),
London, 690 pp.
Excoffier, L., Laval, G. & Schneider, S. (2005) Arlequin (version 3.0): An integrated software package for population genetics
data analysis. Evolutionary Bioinformatics, 1, 47–50.
Galan, M., Pagès, M. & Cosson, J.-F. (2012) Next-Generation sequencing for rodent barcoding: species identification from
fresh, degraded and environmental samples. PLoS ONE, 7, e48374.
http://dx.doi.org/10.1371/journal.pone.0048374
Gilbert, M.T.P., Bandelt, H.J., Hofreiter, M. & Barnes, I. (2005) Assessing ancient DNA studies. Trends in Ecology &
Evolution, 20, 541–544.
http://dx.doi.org/10.1016/j.tree.2005.07.005
Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W. & Gascuel, O. (2010) New algorithms and methods to
estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Systematic Biology, 59, 307–321.
Kloss, C.B. (1916) On a collection of mammals from Siam. Journal of the Natural History Society of Siam, 2, 1–32.
Latinne, A., Waengsothorn, S., Herbreteau, V. & Michaux, J.R. (2011) Evidence of complex phylogeographic structure for the
threatened rodent Leopoldamys neilli, in Southeast Asia. Conservation Genetics, 12, 1495–1511.
http://dx.doi.org/10.1007/s10592-011-0248-3
Latinne, A., Waengsothorn, S., Rojanadilok, P., Eiamampai, K., Sribuarod, K. & Michaux, J.R. (2012) Combined
mitochondrial and nuclear markers revealed a deep vicariant history for Leopoldamys neilli, a cave-dwelling rodent of
Thailand. PLoS ONE, 7, e47670.
http://dx.doi.org/10.1371/journal.pone.0047670
Latinne, A., Waengsothorn, S., Rojanadilok, P., Eiamampai, K., Sribuarod, K. & Michaux, J.R. (2013) Diversity and endemism
of Murinae rodents in Thai limestone karsts. Systematics and Biodiversity, 11, 323–344.
http://dx.doi.org/10.1080/14772000.2013.818587
Zootaxa 3731 (4) © 2013 Magnolia Press · 597TAXONOMIC STATUS OF LEOPOLDAMYS HERBERTI AND L. NEILLI

TERMS OF USE This pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website is prohibited.
Marshall, J.D. (1977) Rats and mice of Thailand. In: Lekagul, B. & McNeely, J.A. (Eds.), Mammals of Thailand. Saha Karn
Bhaet Bangkok, Thailand, pp. 395–490.
Musser, G.G. & Carleton, M. (2005) Superfamily Muroidea. In: Wilson, D.E. & Reeder, D.M. (Eds.), Mammal species of the
World: a taxonomic and geographic reference. Johns Hopkins University Press, Baltimore, pp. 894–1531.
Pages, M., Chaval, Y., Herbreteau, V., Waengsothorn, S., Cosson, J.F., Hugot, J.P., Morand, S. & Michaux, J. (2010) Revisiting
the taxonomy of the Rattini tribe: a phylogeny-based delimitation of species boundaries. BMC Evolutionary Biology, 10,
184.
http://dx.doi.org/10.1186/1471-2148-10-184
Posada, D. & Crandall, K.A. (1998) MODELTEST: testing the model of DNA substitution. Bioinformatics, 14, 817–818.
http://dx.doi.org/10.1093/bioinformatics/14.9.817
Robinson, H.C. & Kloss, C.B. (1922) V.—New mammals from French Indo-China and Siam. Journal of Natural History Series
9, 9, 87–99.
http://dx.doi.org/10.1080/00222932208632642
Ronquist, F. & Huelsenbeck, J.P. (2003) MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics, 19,
1572–1574.
http://dx.doi.org/10.1093/bioinformatics/btg180
Sarkar, I.N., Planet, P.J. & Desalle, R. (2008) CAOS software for use in character-based DNA barcoding. Molecular Ecology
Resources, 8, 1256–1259.
http://dx.doi.org/10.1111/j.1755-0998.2008.02235.x
Swofford, D.L. (1998) PAUP*. Phylogenetic Analysis Using Parsimony, (*and other methods). Version 4. Sinauer Associates,
Sunderland, Massachusetts.
Teletchea, F., Bernillon, J., Duffraisse, M., Laudet, V. & Hanni, C. (2008) Molecular identification of vertebrate species by
oligonucleotide microarray in food and forensic samples. Journal of Applied Ecology, 45, 967–975.
http://dx.doi.org/10.1111/j.1365-2664.2007.01415.x
Thomas, O. (1882) Description of a new species of rat from China. Proceedings of Zoological Society of London, 50, 587–588.
Thomas, O. (1887) Description of a new rat from North Borneo. Annals and Magazine of Natural History, series 5, 20, 269–
270.
http://dx.doi.org/10.1080/00222938709460053
Waengsothorn, S., Kenthao, A., Latinne, A. & Hugot, J.P. (2009) Rodents within the Centre for Thai National Reference
Collections (CTNRC), past, present and future. Kasetsart Journal Natural Sciences, 43, 118–124.
LATINNE ET AL.598 · Zootaxa 3731 (4) © 2013 Magnolia Press
Related Documents











