Journal of Fish Diseases 1995, 18, 105-115 Protozoan parasites of gilthead sea bream, Sparus aurata L,, from different culture systems in Spain p. ALVAREZ-PELLITERO, A. SITJA-BOBADILLA, A. FRANCO-SIERRA & O. PALENZUELA Instituto de Aadcultura Torre de la Sal (CSIC). Ribera de Cabanes, Castellon, Spain Abstract. In a parasitic survey of gilthead sea bream, Sparus aiircKa L., cultured in different systems in Spain, eight protozoan species were found. Ectoparasites included two ciliates {Trichodina sp. and a Cryptocaryon-Wkc species), one dinoflagellate {Amyloodinium sp.) and the zooflagcHatc Ichthyobodo sp. Endoparasites were represented by members of the Apicomplexa {Eimeria sp.) and Myxosporea {Polysporoplasma sparis. Ceratomyxa sp. and Leptotheca sp.). Ichthyobodo sp., Eimeria sp., Ceratomyxa sp. and Leptotheca sp. arc re- ported for the first time in 5. aurata. Amyloodinium sp. and Ichthyobodo sp. were associated with outbreaks of disease in larval and juvenile fish. Data on prevalence and intensity of in- feetion and the location in the host are provided. The potential importance of these protozoan parasites for gilthead sea bream culture is discussed. Introduction Gilthead sea bream. Spams aurata L., culture has significantly increased in recent years in Spain, reaching a production of 1600 tonnes in 1992 compared with only 127 tonnes in 1985 (Larrazabal 1992). In spite of the subsequent increase in pathological problems, references to parasitic protozoans of gilthead sea bream in culture conditions are scarce. Apart from our own data on a new myxosporean (Sitja-Bobadilla, Franco-Sierra & Alvarez-Pellitero 1992; Sitja-Bobadilla & Alvarez-Pellitero 1994), most of the available information deals only with occasional findings of protozoans, sometimes associated with outbreaks of disease (Blanc, Marques, Bouix, BrugeroUe & Breuil 1989; Paperna 1980, 1982, 1984, 1991; Diamant, Issar, Colorni & Paperna 1991; Diamant 1992; Mathieu-Daude, Faye, Coste, Monier, Marques & Bouix 1992). The present study analyses the protozoan fauna found in gilthead sea bream in a pathological survey carried out in differetit mariculture systems of Spain, The potential importance of these parasites for sea bream culture is also discussed. Materials and methods Fish From January 1990 to August 1993, different groups of gilthead sea bream from several culture systems were examined. Details of the fish and the culture systems are given in Table 1. In addition, some fish were examined during outbreaks of disease in groups G-1, G-2 and G-4, and in an intensive system on the Cantabric Sea. Correspondence: Dr P. Alvarez-Pellitero, Instituto de Aeuieultura Torre de la Sal (CSIC), Ribera de Cabanes, 12595 Castellon, Spain. 105

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Fish Diseases 1995, 18, 105-115
Protozoan parasites of gilthead sea bream, Sparusaurata L,, from different culture systems in Spain
p. ALVAREZ-PELLITERO, A. SITJA-BOBADILLA, A. FRANCO-SIERRA& O. P A L E N Z U E L A Instituto de Aadcultura Torre de la Sal (CSIC). Ribera de Cabanes, Castellon,
Spain
Abstract. In a parasitic survey of gilthead sea bream, Sparus aiircKa L., cultured in differentsystems in Spain, eight protozoan species were found. Ectoparasites included two ciliates{Trichodina sp. and a Cryptocaryon-Wkc species), one dinoflagellate {Amyloodinium sp.)and the zooflagcHatc Ichthyobodo sp. Endoparasites were represented by members of theApicomplexa {Eimeria sp.) and Myxosporea {Polysporoplasma sparis. Ceratomyxa sp. andLeptotheca sp.). Ichthyobodo sp., Eimeria sp., Ceratomyxa sp. and Leptotheca sp. arc re-ported for the first time in 5. aurata. Amyloodinium sp. and Ichthyobodo sp. were associatedwith outbreaks of disease in larval and juvenile fish. Data on prevalence and intensity of in-feetion and the location in the host are provided. The potential importance of these protozoanparasites for gilthead sea bream culture is discussed.
Introduction
Gilthead sea bream. Spams aurata L., culture has significantly increased in recent years inSpain, reaching a production of 1600 tonnes in 1992 compared with only 127 tonnes in 1985(Larrazabal 1992). In spite of the subsequent increase in pathological problems, references toparasitic protozoans of gilthead sea bream in culture conditions are scarce. Apart from ourown data on a new myxosporean (Sitja-Bobadilla, Franco-Sierra & Alvarez-Pellitero 1992;Sitja-Bobadilla & Alvarez-Pellitero 1994), most of the available information deals only withoccasional findings of protozoans, sometimes associated with outbreaks of disease (Blanc,Marques, Bouix, BrugeroUe & Breuil 1989; Paperna 1980, 1982, 1984, 1991; Diamant, Issar,Colorni & Paperna 1991; Diamant 1992; Mathieu-Daude, Faye, Coste, Monier, Marques& Bouix 1992).
The present study analyses the protozoan fauna found in gilthead sea bream in a pathologicalsurvey carried out in differetit mariculture systems of Spain, The potential importance of theseparasites for sea bream culture is also discussed.
Materials and methods
Fish
From January 1990 to August 1993, different groups of gilthead sea bream from several culturesystems were examined. Details of the fish and the culture systems are given in Table 1. Inaddition, some fish were examined during outbreaks of disease in groups G-1, G-2 and G-4,and in an intensive system on the Cantabric Sea.
Correspondence: Dr P. Alvarez-Pellitero, Instituto de Aeuieultura Torre de la Sal (CSIC), Ribera de Cabanes,12595 Castellon, Spain.
105

1*»<̂ /*• Alvarez-Pelliicro ct al.
' I. I'JctMils of ihe j;ri)ups of fish ;inil the culUirc systems surveyed
Toial Culture Temperature Salinity SamplingGroup Location no. lish conditions Age/size (cm) (°C)' (%o) frequency
Ci-1
G-2
G-3
G 4
SouthernAtlantic C'oaslof Spain
208 Intensive open 8—60 dayshatchery
18 21
Nursery 45-100 days 20 20
2-3 monthsuntil 9 monthsold
6-12 months
WesternMediterraneanCoast(Valencia)
WesternMediterraneanCoast (RiverEbro delta)
WesternMediterraneanCoast (lATS)"^
200
48
127
Semi-intensiveand extensivegrowing stocks
Intensiveelosed system
Semi-intensivegrowing system
Intensive opensystem
46-296
3-5-31
6-3-31
10-2-33-0
10-28-5 25-40
16-27 16 2-3 months
8-1-26-7 28-36
9-27 37-8
4 occasionalsamplings
10 occasionalsamplings
' Range of monthly averages, with the minimum values in the period from December to February, and themaximum values in the period from July to September. For the nursery and hatehery systems, well water tem-perature fluctuated very slightly.' [ATS = fnstituto de Acuieultura de Torre de la Sal facilities (Castellon).
Sampling procedure
Fish were killed by overexposure to the anaesthetic MS-222 (Sigma, St Louis, MO, USA). Freshsmears of the different organs were examined for protozoan parasites and the presence of anystage of a protozoan was semiquantitatively evaluated following a scale from + to + + + + + + .This intensity scale was based on the number of parasitic stages per microscope field at 125xfor Trichodina sp. and Amyloodinium sp., and at 300x for the remaining protozoans, withthe range: ( + ) 1-5; ( + + ) 6-10; (+ + + ) 11-25; ( + + + +) 26-50; (+ + + + +) 51-100;( + + + + + + ) > 100.
Histological processing
Thin sections {1-3/im) were obtained from material fixed in 10% buffered formalin andembedded in Historesin (Leica, Spain). They were stained with toluidine blue (TB).
Results
Ectoparasites
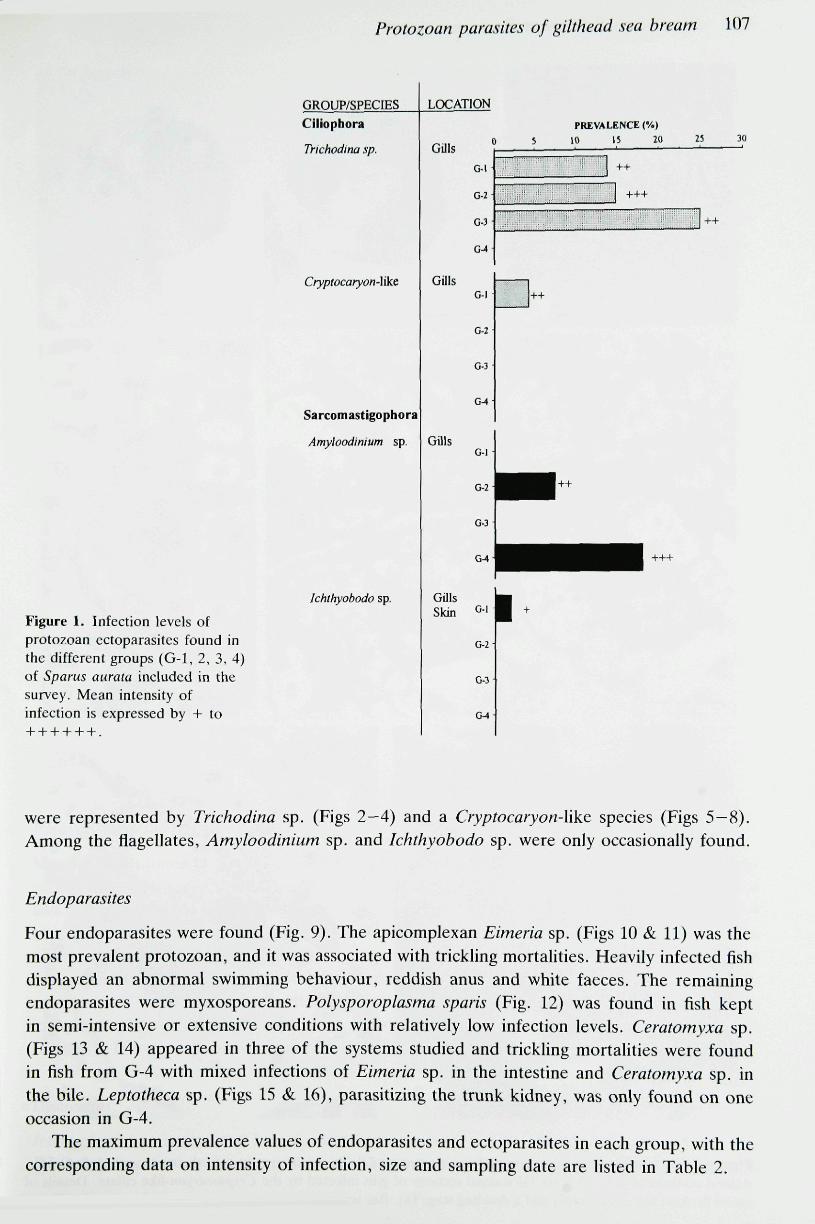
Four species of ectoparasites were found on the fish examined in the survey (Fig. 1). Cifiophora
Protozoan parasites of gilthead sea bream 107
GROUP/SPECIES
Ciliophora
Trichodina sp.
Cryplocaryon-] ike
Sarcomastigophora
Amyloodinium sp.
Ichthyobodo sp.
Figure 1. Infection levels ofprotozoan ectoparasites found inthe different groups (G-l, 2, 3, 4)of Sparus aurata included in thesurvey. Mean intensity ofinfection is expressed by + to
LOCATION
Gills
Gills
GUIs
GillsSkin
0
G-l
G-2
G-3i
G4
G-l
G-2
G-3
G4
G-J
G-2
G-3
G-3
G-A
PREVALENCE (%)
10 IS 20 25 30
L
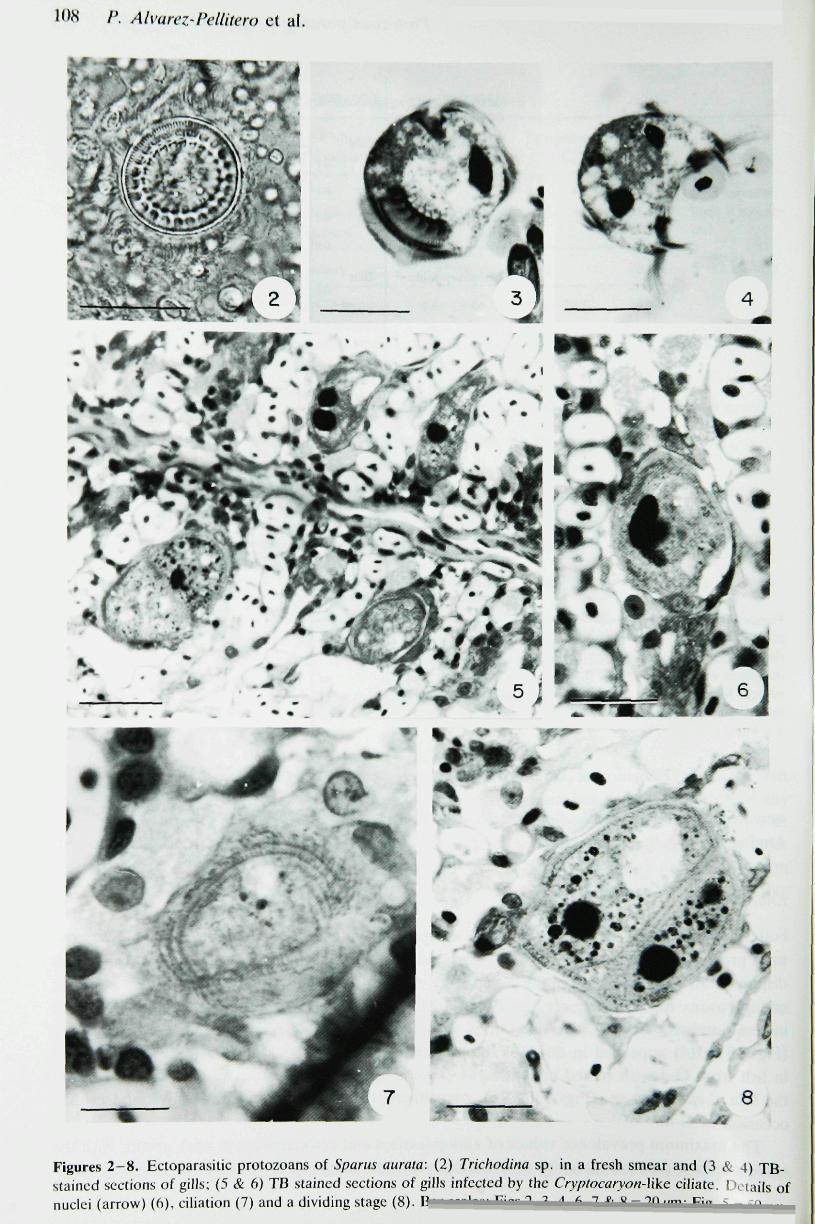
were represented by Trichodina sp. (Figs 2—4) and a Cryptocaryon-Ukc species (Figs 5—8).Among the flagellates, Amyloodinium sp. and Ichthyobodo sp. were only occasionally found.
Endoparasites
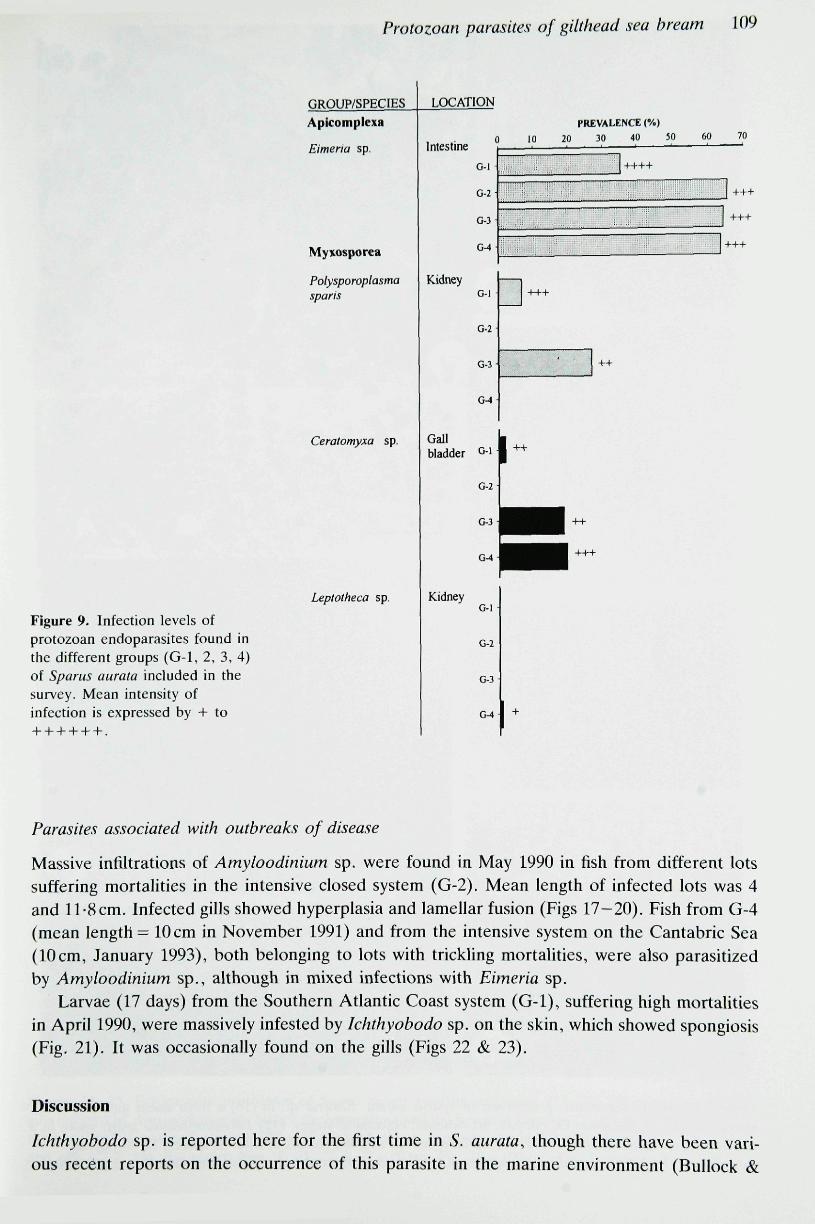
Four endoparasites were found (Fig. 9). The apicomplexan Eimeria sp. (Figs 10 & 11) was themost prevalent protozoan, and it was associated with trickling mortahties. Heavily infected fishdisplayed an abnormal swimming behaviour, reddish anus and white faeces. The remainingendoparasites were myxosporeans. Polysporoplasma sparis (Fig. 12) was found in fish keptin semi-intensive or extensive conditions with relatively low infection levels. Ceratomvxa sp.(Figs 13 & 14) appeared in three of the systems studied and trickling mortalities were foundin fish from G-4 with mixed infections of Eimeria sp. in the intestine and Ceratomvxa sp. inthe bile. Leptotheca sp. (Figs 15 & 16), parasitizitig the trunk kidney, was only found on oneoccasion in G-4.
The maximum prevalence values of endoparasites and ectoparasites in each group, with thecorresponding data on intensity of infection, size and sampling date are listed in Table 2.
108 l\ Alvarez-Pellitero c( al.
8
Figures 2-8. Eetoparasitic protozoans of Sparus aurata: (2) Trichodina sp. in a fresh smear and (3 t̂stained sections of gills; (5 & 6) TB stained sections of gills infected by the Cryptocaryon-Wkc ciliate. Inuclei (arrow) (6), eiliation (7) and a dividing stage (8). Uimnlnm Fiiir 1 7 1 ft 7 fr V-70 irnr Fin -
4)'.-laiis of
Protozoan parasites of gilthead sea bream 109
GROUP/SPECIES
Apicomplcxa
Eimeria sp.
Myxosporea
Poiysporoplasmasparis
Ceratomyxa Sp.
Leptotheca sp.
Figure 9. Infection levels ofprotozoan endoparasites found inthe different groups (G-l, 2, 3, 4)of Sparus aurata included in thesurvey. Mean intensity ofinfection is expressed by + to
LOCATION
Intestine
Kidney
Gallbladder
Kidney
PREVALENCE(%)
0 10 20 30 40 50 60 70
G-l
G-2
G-3
G-t
G-l
G-2
G-3
G-4
G-t
G-l
G-2
G-3
G-4
Parasites associated with outbreaks of disease
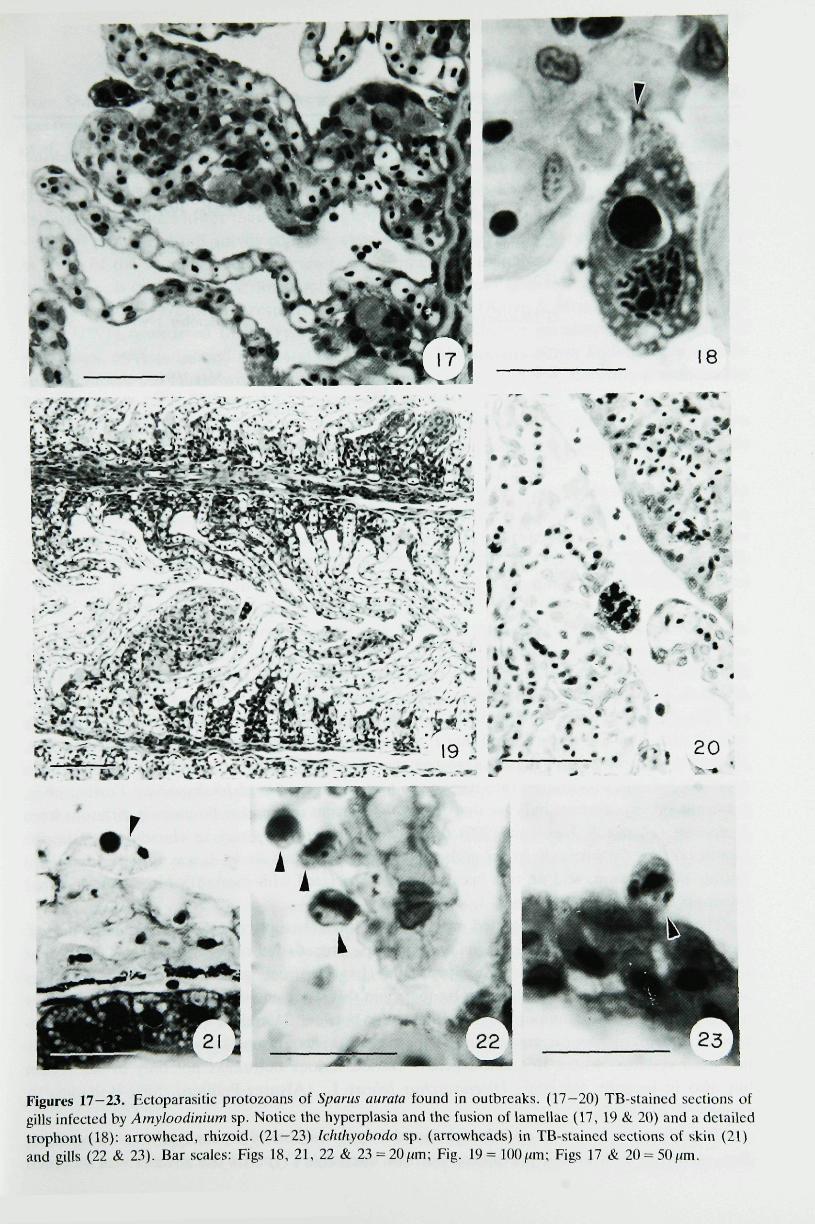
Massive infiltrations of Amyloodinium sp. were found in May 1990 in fish from different lotssuffering mortalities in the intensive closed system (G-2). Mean length of infeeted lots was 4and ll-8cm. Infeeted gills showed hyperplasia and lamellar fusion (Figs 17-20). Fish from G-4(mean length = 10cm in November 1991) and from the intensive system on the Cantabric Sea(lDcm, January 1993), both belonging to lots with trickling mortalities, were also parasitizedby Amyloodinium sp., although in mixed infections with Eimeria sp.
Larvae (17 days) from the Southern Atlantic Coast system (G-l), suffering high mortalitiesin April 1990, were massively infested by Jchthyobodo sp. on the skin, whieh showed spongiosis(Fig. 21). It was occasionally found on the gills (Figs 22 & 23).
Discussion
Ichthyobodo sp. is reported here for the first time in S. aurata, though there have been vari-ous recent reports on the occurrence of this parasite in the marine environment (Bulloek &
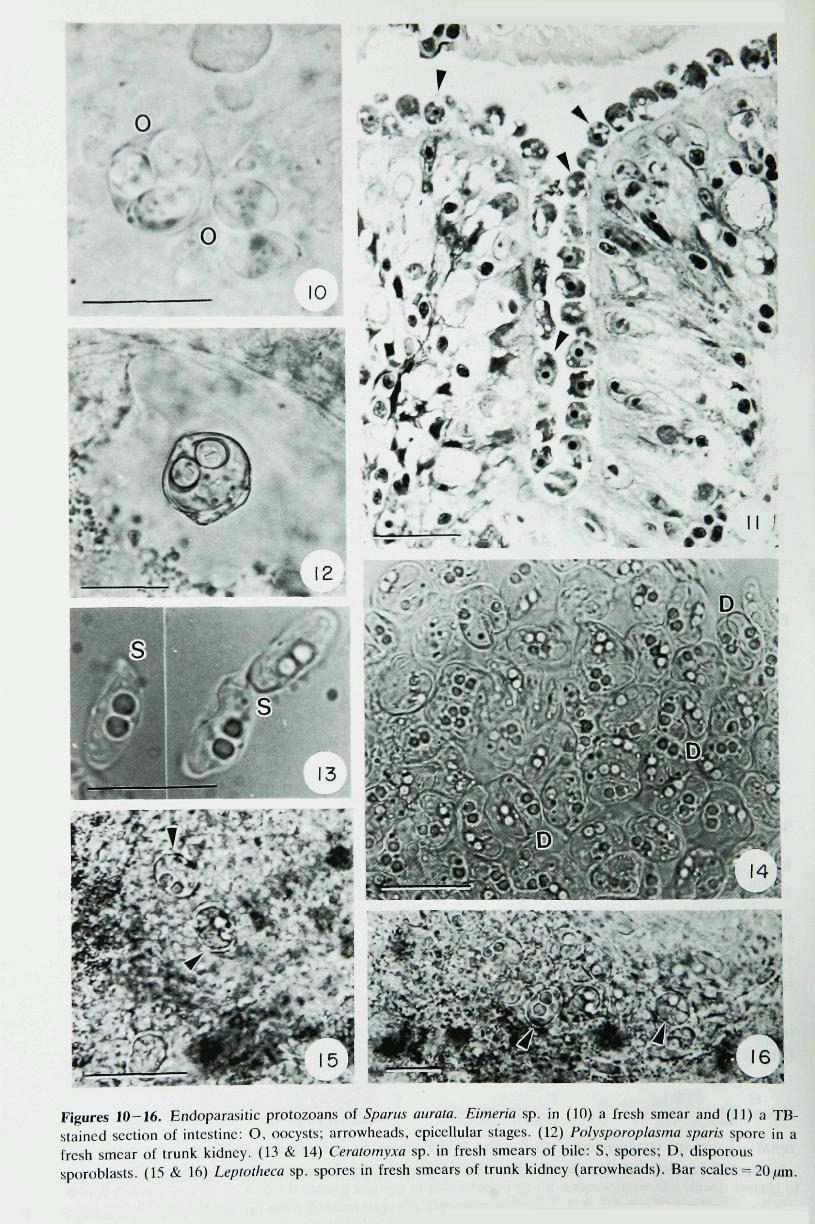
Figures 10-16. Endoparasitic protozoans of Spams aurata. Eimeria sp. in (10) a fresh smear and (11) a TB-staincd section of intestine: O, ooeysts; arrowlieads, epicclUilar stages. (12) Polysporoplasma sparis spore in afresh smear of trunk kidney. (13 & 14) Ceratomyxa sp. in fresh smears of bile: S, spores; D, disporoussporoblasts. (15 & 16) Leptotheca sp. spores in fresh smears of trunk kidney (arrowheads). Bar scales = 20
4 f̂ • .•
Figures 17-23. Ectoparasitic protozoans of Spartis aurata found in outbreaks. (17-20) TB-staincd sections ofgills infccled by Amyioodinium sp. Notice the hyperplasia and the fusion of lamellae (17, 19 & 20) and a detailedtrophonl (18): arrowhead, rhizoid. (21-23) Ichthyobodo sp. (arrowheads) in TB-staincd seetions of skin (21)and gills (22 & 23), Bar scales: Figs 18, 21, 22 & 23 = 20/im; Fig. 19= 100/an; Figs 17 & 20-50/mi.

tl2 r, Alvarez-reUitcro ct al.
' 2 !»)• s;imp!os of brc;im wilh m;ixiimiin prcv.'ilcfu'c values for each prolozoan parasite
r . i i . is iu-
Trirhndind sp.
Cr\pl<H'(tryi>n sp.
AniyUHhluiiiini sp.
/(liihvnhoilo sp.
l.uucriu sp.
Polvsporoplusnui sparis
Ceratomyxa sp.
Group/syslcm
G-l/scmi intensiveG-3
G-1/semi-inlcnsivc
G-2
G-1/hateliery
G-l/scmi-iniensiveG-2G 3G-4
G-1/semi-intensiveG-3
G-1/semi-intensiveG-3G-4
Prevalence(%)
80100
80
75
30
10085
10080
10050
2310063-6
Meanintensity
- t - - l - - l -
-I- + +
+ +
+ +
+
+ -\- + ++ + + -h-I--I- + -I-
+ +
-I- + -I--I--I- + -I--I--I- +
Sampling date
Oetober 1990May 1992
November 1991
May 1992
Oetober 1990
November 1992May 1991April 1993March 1993
Aprii 1990September 1993
May 1993April 1993June 1993
Age/size*
10-87-8
23-4
23-7
30 days
25-723-71519-8
2317-6
15-615-321-4
* Mean Icncth of the lot (em).
Robertson 1982; Cone & Wiles 1984; Morrison & Cone 1986; Diamant 1987; Urawa, Ueki,Nakai & Yamasaki 1991; Kusakari & Urawa 1990; Bruno 1992).
Ellis Sc Wootten (1978) noted the survival of /. necator on the gills of farmed Atlanticsalmon post-smolts and considered that /. necator could be transferred from fresh to sea water.However, several authors have suggested that the Ichthyobodo sp. found in marine hostsprobably belongs to a marine race or even to a separate species of Ichthyobodo. Furthermore,transmission experiments indicate that Ichthyobodo from the marine flounder is different from/. necator (Urawa & Kusakari 1990). Further studies are necessary to elucidate the specificidentity of the Ichthyobodo sp. of gilthead sea bream. In our study, it was found in larvae inSpring and Autumn, and on one occasion, was associated with mortalities in which no otherinfection could be demonstrated.
The other ectoparasites detected in this survey have been previously reported in S, aurata.According to Paperna (1980), Amyloodinium sp. caused outbreaks of disease in differentperiods of the year in gilthead sea bream at Eilat (Israel), affecting larvae, fingerlings, juvenilesand even breeders. The low infection frequency in this study could be caused by a lower watertemperature, as winter temperatures in Eilat range between 21 and 26°C. The reported import-ance oi Amyloodinium for mariculture (Paperna 1980) is confirmed by our results, particularlyin closed systems, where this dinofiagellate caused serious outbreaks not only in sea breambut also in juvenile sea bass, Dicentrarchus labrax L. (Alvarez-Pellitero, Sitja-Bobadilla &
Franco-Sierra 1993).The ciliate Cryptocaryon sp., though found only in two lots of bream on the same sampling
date, occurred with a high prevalence but low intensity. Cryptocaryon irritans was recorded

Protozoan parasites of gilthead sea bream 113
from Sparus aurata from Israel (Colorni 1985), and more recently, a Cryptocaryon-Wkz ciliatewas reported to be the cause of severe epizootics in gilthead sea bream culture in Israel (Diamantet al. 1991); fingerlings were affected in September (water temperature = 27 °C) and breedersin November (22 °C). It is well known that the life-cycle of this ciliate is temperature dependent(Lom & Dykova 1992), and that it is sensitive to saline concentrations, as tomonts undergocytolysis in salinities lower than 20%o (Colorni 1985). The lack of disease outbreaks associatedwith this parasite in this survey could be caused by the lower water temperature and salinitiesin some of the systems studied. Cryptocaryon irritans. an ubiquitous eiliate with a very widehost range, is considered a serious problem in marine aquaria and mariculture (Paperna 1984;Lom & Dykova 1992), and is responsible for striking losses (Kaige & Miyazaki 1985). Diamantet al. (1991) considered their Cryptocaryon species a coldwater counterpart of the tropicalC. irritans, as they differed in several characteristics. The scarcity of our findings do not allowus to compare our ciliate with the other Cryptocaryon species, though it mostly resembles theMediterranean form described by Diamant et al. (1991).
The prevalence and intensity of infection by ectoparasites in the closed culture systemswere lower than expected. This might be caused by routine chemical disinfections carried outprior to the introduction of fish to the system.
Eimeria sp. is recorded for the first time in gilthead sea bream. The wide distribution in thedifferent culture systems studied and the associated mortalities suggest the potential importanceof this coccidiosis for sea bream culture. The data obtained indicate Spring and Autumn as themost favourable periods for infection. Although fish of different ages were affected by thiscoecidian, it was never found in fish smaller than 5 cm. Taxonomic and histopathologicalstudies on this parasite will be presented elsewhere.
The myxosporean previously reported (Sitja-Bobadilla et al. 1992) in the Southern AtlanticCoast system was also found in the Mediterranean (River Ebro delta) in semi-intensive cultureconditions. The parasite caused noticeable histopathological damage in the kidney, as well asskin ulcerations in some affected fish. The parasite has recently been described as a new genusand species, Poiysporoplasma sparis^ because of the presence of several sporoplasms in thespore (Sitja-Bobadilla & Alvarez-Pellitero 1994).
Two other myxosporeans have been found in gilthead sea bream in this survey. Ceratomvxasp. appeared frequently in mixed infections with Eimeria sp. in fish stocks with trickling mor-talities and abdominal swelling. Although total prevalences were not very high, all fish examinedin some periods were parasitized and at a high intensity. The genus Ceratomyxa is reported forthe first time in S. aurata and possibly represents a new species. Leptotheca sp. was only foundin one fish from G-4.
In conclusion, various protozoan species have been recorded in cultured gilthead sea bream.Spring and Autumn seem to be the main periods of infection for most of these parasites, whenwater temperature ranges were 13-26 and 24-13°C, respectively, which are close to theoptimum temperature for the growth of gilthead sea bream (23-25""C) (Larrazabal 1992). Therichest protozoan fauna was found in semi-intensive conditions, perhaps because of the poss-ible presence of putative intermediate hosts for the transmission of myxosporeans (Wolf &Markiw 1984) and Apicomplexa (Dykova & Lom 1981; Fournie & Overstreet 1983).
Acknowledgments
This work was supported by research grants MAR78970557 from the Spanish CICYT andFAR/AQ/M78 from the EC. O. Palenzuela was recipient of a grant from the Departamento

114 /'. Alvarez-Pellitcro ct al.
dc Hducacion, Lhiivcrsidatlcs e Investigacion del (iobierno Vasco. We are grateful to M.J.Sanclcto and J. Monfort for the histological processing.
References
Alvarcz-Pcllitcro P., Silja-Bobadilla A. & Franco-Sierra A. (1993) Protozoan parasites of wild and eultured seabass, Dicentrarchus lahrax (L.), from the Mediterranean area. Aquaculture and Fisheries Management 24,101-108.
Blane E., Marques A., Bouix G., Brugerolle G. & Breuil G. (1989) Cryptohia sp. from the gills of the giltheadsea hrcam Sparus aurata. Btilletin of ihe European Association of Fish Palhologisls 9, 81-82.
Bruno D. W. (1992) khihyobodo sp. on farmed Atlanlie salmon, Salmo salar L., reared in the marine environ-ment. Journal of Ftsh Diseases 15, 349-351.
Bulloek A. M. & Robertson D. A. (1982) A note on the oecurrenee of Ichthyobodo necator (Henneguy, 1883) ina wild population of juvenile plaiee, Pleuronectes platessa L. Journal of Fish Diseases 5, 531 — 533.
Colorni A. (1985) Aspects of the biology of Cryptocaryon irritans. and hyposalinity as a control measure incultured gilthead sea bream Sparus aurata. Diseases of Aquatic Organisms I, 19—22.
Cone D. K. & Wiles M. (1984) Ichthyobodo necator (Henneguy, 1883) from winter flounder, Pseudopleuronectesamericanus (Walbaum), in the north-west Atlantic Ocean. Journal of Fish Diseases 7. 87-89.
Diamant A. (1987) UUrastructure and pathogenesis of Ichthyobodo sp. from wild common dab, Limanda limandaL.. in the North Sea. Journal of Fish Diseases 10, 241-247.
Diamant A. (1992) A new pathogcnie histozoic Myxidium (Myxosporea) in eultured gilthead sea bream Spartisaurata. Bulletin of the European Association of Fish Pathologists 12, 64 — 66.
Diamant A.. Issar G., Colorni A. & Paperna I. (1991) A pathogenic Cryptocaryon-\\\iQ eiliate from the Mediter-ranean Sea. Bulletin of the European Association of Fish Pathologists 11, 122—124.
Dykova I. & Lom J. (1981) Fish coecidia: critical notes on life cycles, classification and pathogenicity. Journal ofFish Diseases 4. 487-505.
Ellis A. E. & Wootten R. (1978) Costiasis of Atlantie salmon Salmo salar L. smoits in sea water. Journal of FishDiseases 1. 389-393.
Fournie J. W. & Overstreet R. M. (1983) True intermediate hosts for Eimeria funduh (Apicomplexa) fromestuarinc fishes. Journal of Protozoology 30, 672—675.
Kaigc N. & Miyazaki T. (1985) A histopathological study of white spot disease in Japanese flounder. Fish Path-ology 20, 61-64.
Kusakari M. &. Urawa S. (1990) Histopathology of the skin oi yearling Japanese flounder, Paralichthys oUvaceus,\nfccic6 with the flagellate Ichthyobodo sp. Fish Pathology 25, 59-68.
Larrazabal G. (1992) Fish aquaeulture in Spain. World Aquaculture 23, 10-16.Lom J. & Dykova 1. (1992) Protozoan parasites of fishes. In: Developments in Aquaculture and Fisheries Science,
Vol. 26, p. 315. Elsevier, Amsterdam.Mathicu-Daude F.. Faye N., Coste F., Monier J. F., Marques A. & Bouix G. (1992) Occurrence of a micro-
sporidiosis in marine eultured gilthead sea bream from the Languedoc Coast: a problem of speeifieity in thegenus Glugea (Protozoa, Microspora). Bulletin of the European Association of Fish Pathologists 12, (sl-lO.
Morrison C. M. & Cone D. K. (1986) A possible marine form of Ichthyobodo sp. on haddock, Melanogrammusaeglefinus (L.), in the north-west Atlantic Ocean. Journal of Fish Diseases 9, 141-142.
Paperna I. (1980) Amyloodinium ocellatum (Brown, 1931) (Oinoflagellida) infestations in cultured marine fish atEilat, Red Sea: epizootioiogy and pathology. Journal of Fish Diseases 3, 363—372.
Paperna L (1982) Kudoa infections in the glomerulus, mesentery and peritoneum of eultured Sparus aurata L.Journal of Fish Diseases 5, 539-544.
Paperna L (1984) Review of diseases affecting cultured Sparus aurata and Dicentrarchus labrax. In: L'Aquaculluredu bar et des sparides (ed. by G. Barnabe & R. Billard), pp. 465-482. INRA Publications, Paris.
Paperna L (1991) Diseases caused by parasites in the aquaculture of warm water fish. Annual Review of FishDisea.ses I, 155-194.
Sitja-Bobadilla A., Franco-Sierra A. & Alvarcz-Pellitero P. (1992) Sphaerospora (Myxosporea: Bivalvulida)infeetion in cultured gilthead sea bream, Sparus aurata L.: a preliminary report. Journal of Fish Diseases15, 339-343.
Sitja-Bobadilla A. & Alvarez-Pellitcro P. (1994) Light and electron mieroseopie description of Poiysporoplasman. g. (Myxosporea: Bivalvulida), Poiysporoplasma sparis n. sp. from Sparus aurata (L.) and Poiysporoplasmamugilis n. sp. from Liza aurata L. European Journal of Protistology 30, in press.
Urawa S. & Kusakari M. (1990) The survivability of the ectoparasitie flagellate Ichthyobodo necator on chum

Protozoan parasites of gilthead sea bream 115
salmon fry {Oncorhynchus keta) in sea water and comparison to Ichthyobodo sp. on Japanese flounder{Paralichthvs olivaceus). Journal of Parasitology 76, 33-40.
Drawa S., Ueki N., Nakai T. & Yamasaki H. (1991) High mortality of cultured Japanese flounder, Paralichthysolivaceus (Temminek & Sehlegel), caused by the parasitic flagellate Ichthyobodo sp. Journal of Ftsh Diseases14. 489-494.
Wolf K. & Markiw M. E. (1984) Biology eontravenes taxonomy in the Myxozoa: new diseoveries show alter-nation of invertebrate and vertebrate hosts. Science 225, 1449-1452.

Related Documents











