Rapid Communication Protective Effect of Sanguinarine on Ultraviolet B-mediated Damages in SKH-1 Hairless Mouse Skin: Implications for Prevention of Skin Cancer Haseeb Ahsan, Shannon Reagan-Shaw, David M. Eggert, Thomas C. Tan, Farrukh Afaq, Hasan Mukhtar and Nihal Ahmad* Department of Dermatology, University of Wisconsin, School of Medicine and Public Health, Madison, WI Received 30 March 2007; accepted 27 April 2007; DOI: 10.1111 ⁄ j.1751-1097.2007.00156.x ABSTRACT Excessive exposure of solar ultraviolet (UV) radiation, partic- ularly its UVB component (280–320 nm), to human skin is the major cause of skin cancers. UV exposure also leads to the development of precancerous conditions such as actinic kera- tosis and elicits a variety of other adverse effects such as sunburn, inflammation, hyperplasia, immunosuppression and skin aging. Therefore, there is a need to intensify our efforts towards the development of novel mechanism-based approa- ches ⁄ agents for the protection of UVB-mediated damages. Chemoprevention is being investigated as a potential approach for the management of UV damages including skin cancer. We have earlier shown that sanguinarine, a benzophenanthridine alkaloid, inhibits UVB exposure-mediated damages in HaCaT keratinocytes. In this study, to determine the relevance of our in vitro findings to in vivo situations, we assessed the effects of sanguinarine on UVB-mediated damages in SKH-1 hairless mice. Our data demonstrated that a topical application of sanguinarine (5 lmol 0.3 mL )1 ethanol per mouse), either as a pretreatment (30 min prior to UVB) or posttreatment (5 min after UVB), resulted in a significant decrease in UVB-mediated increases in skin edema, skin hyperplasia and infiltration of leukocytes. Further, sanguinarine treatments (pre and post) also resulted in a significant decrease in UVB mediated (1) generation of H 2 O 2 and (2) increases in the protein levels of markers of tumor promotion ⁄ proliferation viz. ornithine decarboxylase (ODC), proliferating cell nuclear antigen (PCNA) and Kiel antigen-67. Based on this data, we suggest that sanguinarine could be developed as an agent for the management of conditions elicited by UV exposure including skin cancer. However, further detailed studies are needed to support this suggestion. INTRODUCTION Ultraviolet (UV) radiation from sun, particularly its UVB component (290–320 nm), is the major cause of skin cancer that accounts for more than a million new cases each year in the USA alone (1). Further, exposure to UVB may lead to precancerous lesions such as actinic keratoses and may also elicit a variety of other adverse effects including erythema, sunburn, inflammation, hyperplasia, skin aging, hyperpigmen- tation and immunosuppression (2–5). UVB radiation causes tumor initiation through DNA damage and its promoting activity includes transcriptional modulation of genes involved in tumor promotion as well as activation of several signal transduction pathways (2–5). Whereas low doses of UVB cause DNA mutation leading to tumor initiation, high doses result in irreparable DNA damage causing apoptosis (sun- burn) and eventually cell death (2–5). UVB also indirectly damages DNA by increasing levels of reactive oxygen species (ROS) which facilitate DNA oxidation (6). Cells in the epidermis with common UV mutations represent ‘‘initiated cells’’ which can then undergo clonal expansion to become tumors (7). The available prevention strategies have not been able to effectively manage the increasing incidence of skin cancers and other UV-related conditions. This warrants immediate attention towards developing novel mechanism-based approaches for the prevention as well as treatment of skin cancer. Chemoprevention by naturally occurring plant-based agents is being investigated as a potential approach for prevention as well as treatment of skin cancer as well as other UV exposure related conditions. Sanguinarine, derived from the root of Sanguinaria canadensis and other poppy- fumaria species, is a benzophenanthridine alkaloid and a structural homolog of chelerythrine and has been shown to possess antimicrobial, antioxidant and anti-inflammatory properties (8,9). It is often used in toothpastes and mouth- washes for its anti-inflammatory effects (9–11). We have shown previously that sanguinarine induces apoptosis in human squamous carcinoma A431 cells but not in normal human epidermal keratinocytes at micromolar doses (12). In addition, sanguinarine was shown to induce apoptosis via activation of Bcl-2 family proteins and the mitochondrial pathway in immortalized human HaCaT keratinocytes (13). In a recent study, we have shown that the plant alkaloid sanguinarine protects HaCaT skin keratinocytes from UVB- mediated damages via enhancing the apoptotic elimination of UVB-damaged cells (14). This study was designed to investigate the relevance of these findings to in vivo situations in SKH-1 hairless mouse skin that possesses *Corresponding author email: [email protected] (Nihal Ahmad) Ó 2007 The Authors. Journal Compilation. The American Society of Photobiology 0031-8655/07 Photochemistry and Photobiology, 2007, 83: 986–993 986

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Rapid Communication
Protective Effect of Sanguinarine on Ultraviolet B-mediated Damages inSKH-1 Hairless Mouse Skin: Implications for Prevention of Skin Cancer
Haseeb Ahsan, Shannon Reagan-Shaw, David M. Eggert, Thomas C. Tan, Farrukh Afaq, Hasan Mukhtarand Nihal Ahmad*
Department of Dermatology, University of Wisconsin, School of Medicine and Public Health, Madison, WI
Received 30 March 2007; accepted 27 April 2007; DOI: 10.1111 ⁄ j.1751-1097.2007.00156.x
ABSTRACT
Excessive exposure of solar ultraviolet (UV) radiation, partic-
ularly its UVB component (280–320 nm), to human skin is the
major cause of skin cancers. UV exposure also leads to the
development of precancerous conditions such as actinic kera-
tosis and elicits a variety of other adverse effects such as
sunburn, inflammation, hyperplasia, immunosuppression and
skin aging. Therefore, there is a need to intensify our efforts
towards the development of novel mechanism-based approa-
ches ⁄ agents for the protection of UVB-mediated damages.
Chemoprevention is being investigated as a potential approach
for the management of UV damages including skin cancer. We
have earlier shown that sanguinarine, a benzophenanthridine
alkaloid, inhibits UVB exposure-mediated damages in HaCaT
keratinocytes. In this study, to determine the relevance of our
in vitro findings to in vivo situations, we assessed the effects of
sanguinarine on UVB-mediated damages in SKH-1 hairless
mice. Our data demonstrated that a topical application of
sanguinarine (5 lmol 0.3 mL)1 ethanol per mouse), either as a
pretreatment (30 min prior to UVB) or posttreatment (5 min
after UVB), resulted in a significant decrease in UVB-mediated
increases in skin edema, skin hyperplasia and infiltration of
leukocytes. Further, sanguinarine treatments (pre and post)
also resulted in a significant decrease in UVB mediated (1)
generation of H2O2 and (2) increases in the protein levels of
markers of tumor promotion ⁄ proliferation viz. ornithine
decarboxylase (ODC), proliferating cell nuclear antigen
(PCNA) and Kiel antigen-67. Based on this data, we suggest
that sanguinarine could be developed as an agent for the
management of conditions elicited by UV exposure including
skin cancer. However, further detailed studies are needed to
support this suggestion.
INTRODUCTION
Ultraviolet (UV) radiation from sun, particularly its UVB
component (290–320 nm), is the major cause of skin cancerthat accounts for more than a million new cases each year inthe USA alone (1). Further, exposure to UVB may lead to
precancerous lesions such as actinic keratoses and may also
elicit a variety of other adverse effects including erythema,sunburn, inflammation, hyperplasia, skin aging, hyperpigmen-tation and immunosuppression (2–5). UVB radiation causestumor initiation through DNA damage and its promoting
activity includes transcriptional modulation of genes involvedin tumor promotion as well as activation of several signaltransduction pathways (2–5). Whereas low doses of UVB
cause DNA mutation leading to tumor initiation, high dosesresult in irreparable DNA damage causing apoptosis (sun-burn) and eventually cell death (2–5). UVB also indirectly
damages DNA by increasing levels of reactive oxygen species(ROS) which facilitate DNA oxidation (6). Cells in theepidermis with common UV mutations represent ‘‘initiatedcells’’ which can then undergo clonal expansion to become
tumors (7).The available prevention strategies have not been able to
effectively manage the increasing incidence of skin cancers
and other UV-related conditions. This warrants immediateattention towards developing novel mechanism-basedapproaches for the prevention as well as treatment of skin
cancer. Chemoprevention by naturally occurring plant-basedagents is being investigated as a potential approach forprevention as well as treatment of skin cancer as well as
other UV exposure related conditions. Sanguinarine, derivedfrom the root of Sanguinaria canadensis and other poppy-fumaria species, is a benzophenanthridine alkaloid and astructural homolog of chelerythrine and has been shown to
possess antimicrobial, antioxidant and anti-inflammatoryproperties (8,9). It is often used in toothpastes and mouth-washes for its anti-inflammatory effects (9–11). We have
shown previously that sanguinarine induces apoptosis inhuman squamous carcinoma A431 cells but not in normalhuman epidermal keratinocytes at micromolar doses (12). In
addition, sanguinarine was shown to induce apoptosis viaactivation of Bcl-2 family proteins and the mitochondrialpathway in immortalized human HaCaT keratinocytes (13).
In a recent study, we have shown that the plant alkaloidsanguinarine protects HaCaT skin keratinocytes from UVB-mediated damages via enhancing the apoptotic eliminationof UVB-damaged cells (14). This study was designed
to investigate the relevance of these findings to in vivosituations in SKH-1 hairless mouse skin that possesses
*Corresponding author email: [email protected] (Nihal Ahmad)� 2007TheAuthors. JournalCompilation.TheAmericanSociety ofPhotobiology 0031-8655/07
Photochemistry and Photobiology, 2007, 83: 986–993
986
relevance to human situations. Our data demonstrated that atopical application of sanguinarine resulted in a significantdecrease in UVB-mediated (1) increases in skin edema, skinhyperplasia and infiltration of leukocytes, (2) generation of
H2O2 and (3) increases in the protein levels of markers oftumor promotion ⁄proliferation viz. ornithine decarboxylase(ODC), proliferating cell nuclear antigen (PCNA) and Kiel
antigen-67 (Ki-67). To our knowledge, this is the first studyshowing prevention of UVB-mediated damages in skin bysanguinarine.
MATERIALS AND METHODS
Materials. Sanguinarine chloride (>99.0% pure) was purchased fromSigma Chemical Co. (St. Louis, MO). The following antibodies wereused: PCNA (Biosource International, Camarillo, CA), ODC (Neo-markers ⁄Lab Vision, Fremont, CA), Ki-67 (Santa Cruz Biotechno-logy, Santa Cruz, CA) and b-actin (Sigma-Aldrich, St. Louis, MO). Allother chemicals were of analytical grade.
Animals and UVB light source. Female SKH-1 hairless mice (6-weeks old) obtained from Charles River Laboratories (Wilmington,MA) were used in this study. The mice were allowed to acclimatizefor 2 weeks prior to the start of the experiments. The animals had anad libitum access to Purina Chow diet and tap water. Throughout theexperimental protocols, the mice were maintained at standardconditions (temperature 24 ± 2�C; relative humidity 50 ± 10%;12 ⁄ 12 h light ⁄dark cycle). A ‘‘Daavlin Research Irradiator’’ obtainedfrom Daavlin Company (Bryan, OH) was used for UVB exposure.This equipment consists of a fixture mounted on fixed legs andcontains four UVA and four UVB lamps. The exposure system iscontrolled using two Daavlin Flex Control Integrating Dosimeters.The dose units, in this equipment, could be entered as mJ cm)2 (forUVB) or Joules (for UVA). For accuracy, the machine is periodicallycalibrated using International Light IL 1400, digital light meter(Daavlin Company).
Treatments of animals. Female SKH-1 hairless mice were dividedinto six groups of eight animals each. The mice in the first group didnot receive any treatment and served as a control. The second groupreceived a topical application of 0.3 mL ethanol (vehicle control).The mice in the third group received a topical treatment ofsanguinarine (5 lmol in 0.3 mL ethanol per mouse) on their dorsalskin. The mice in the fourth group received UVB (180 mJ cm)2)treatment. The mice in the fifth group received a topical application
5
10
15
Unt Veh UVB UVB +Sang
UVB Sang
Sang +Ski
n p
un
ch w
eig
ht
(mg
mm
–1 d
iam
eter
)
*
##
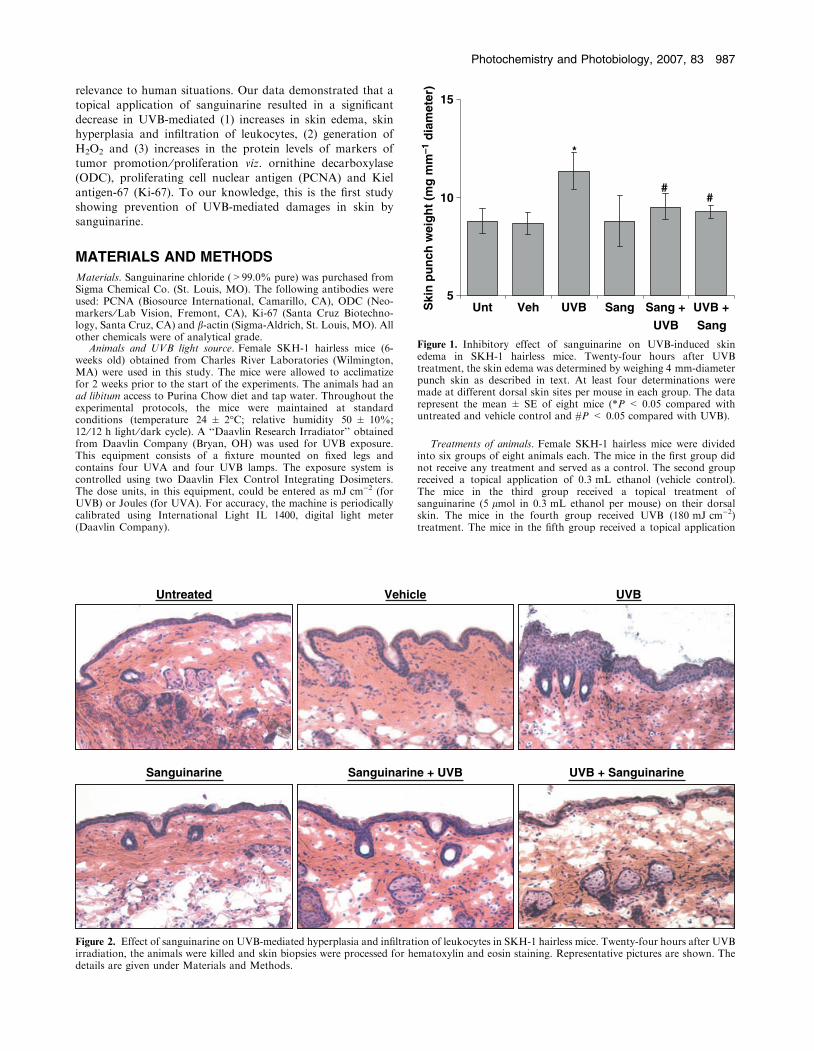
Figure 1. Inhibitory effect of sanguinarine on UVB-induced skinedema in SKH-1 hairless mice. Twenty-four hours after UVBtreatment, the skin edema was determined by weighing 4 mm-diameterpunch skin as described in text. At least four determinations weremade at different dorsal skin sites per mouse in each group. The datarepresent the mean ± SE of eight mice (*P < 0.05 compared withuntreated and vehicle control and #P < 0.05 compared with UVB).
Untreated Vehicle UVB
Sanguinarine Sanguinarine + UVB UVB + Sanguinarine
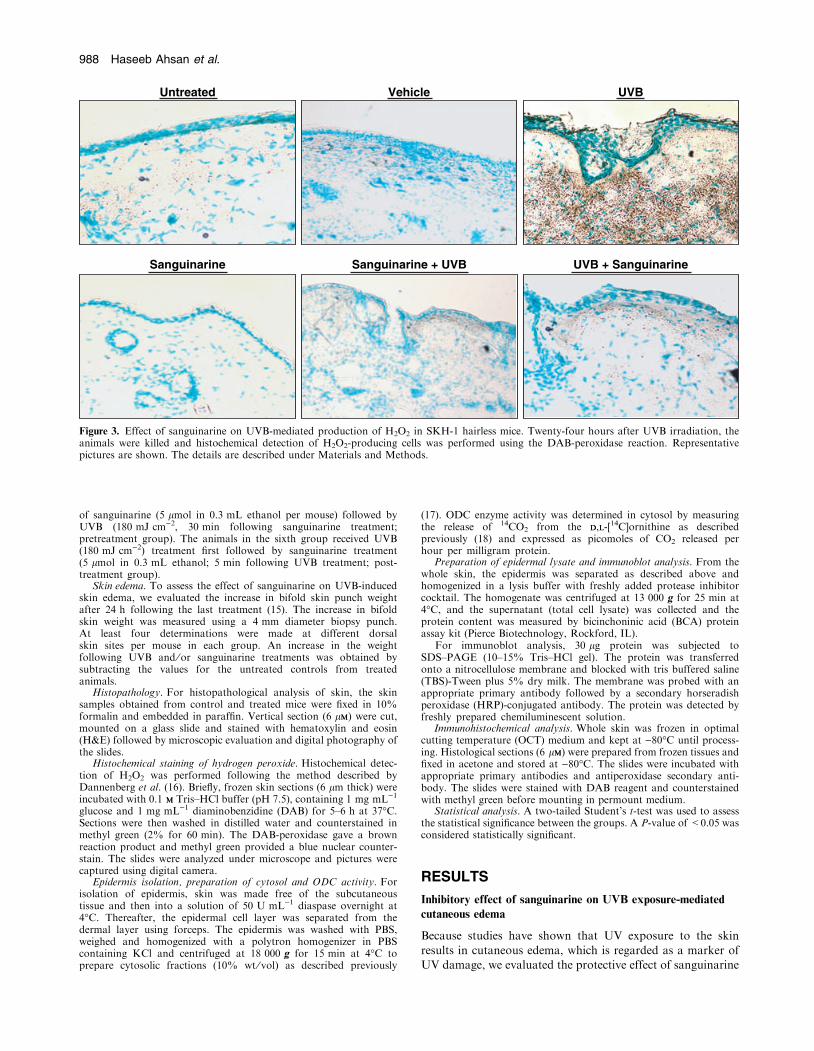
Figure 2. Effect of sanguinarine on UVB-mediated hyperplasia and infiltration of leukocytes in SKH-1 hairless mice. Twenty-four hours after UVBirradiation, the animals were killed and skin biopsies were processed for hematoxylin and eosin staining. Representative pictures are shown. Thedetails are given under Materials and Methods.
Photochemistry and Photobiology, 2007, 83 987
of sanguinarine (5 lmol in 0.3 mL ethanol per mouse) followed byUVB (180 mJ cm)2, 30 min following sanguinarine treatment;pretreatment group). The animals in the sixth group received UVB(180 mJ cm)2) treatment first followed by sanguinarine treatment(5 lmol in 0.3 mL ethanol; 5 min following UVB treatment; post-treatment group).
Skin edema. To assess the effect of sanguinarine on UVB-inducedskin edema, we evaluated the increase in bifold skin punch weightafter 24 h following the last treatment (15). The increase in bifoldskin weight was measured using a 4 mm diameter biopsy punch.At least four determinations were made at different dorsalskin sites per mouse in each group. An increase in the weightfollowing UVB and ⁄ or sanguinarine treatments was obtained bysubtracting the values for the untreated controls from treatedanimals.
Histopathology. For histopathological analysis of skin, the skinsamples obtained from control and treated mice were fixed in 10%formalin and embedded in paraffin. Vertical section (6 lMM) were cut,mounted on a glass slide and stained with hematoxylin and eosin(H&E) followed by microscopic evaluation and digital photography ofthe slides.
Histochemical staining of hydrogen peroxide. Histochemical detec-tion of H2O2 was performed following the method described byDannenberg et al. (16). Briefly, frozen skin sections (6 lm thick) wereincubated with 0.1 MM Tris–HCl buffer (pH 7.5), containing 1 mg mL)1
glucose and 1 mg mL)1 diaminobenzidine (DAB) for 5–6 h at 37�C.Sections were then washed in distilled water and counterstained inmethyl green (2% for 60 min). The DAB-peroxidase gave a brownreaction product and methyl green provided a blue nuclear counter-stain. The slides were analyzed under microscope and pictures werecaptured using digital camera.
Epidermis isolation, preparation of cytosol and ODC activity. Forisolation of epidermis, skin was made free of the subcutaneoustissue and then into a solution of 50 U mL)1 diaspase overnight at4�C. Thereafter, the epidermal cell layer was separated from thedermal layer using forceps. The epidermis was washed with PBS,weighed and homogenized with a polytron homogenizer in PBScontaining KCl and centrifuged at 18 000 g for 15 min at 4�C toprepare cytosolic fractions (10% wt ⁄ vol) as described previously
(17). ODC enzyme activity was determined in cytosol by measuringthe release of 14CO2 from the DD,LL-[14C]ornithine as describedpreviously (18) and expressed as picomoles of CO2 released perhour per milligram protein.
Preparation of epidermal lysate and immunoblot analysis. From thewhole skin, the epidermis was separated as described above andhomogenized in a lysis buffer with freshly added protease inhibitorcocktail. The homogenate was centrifuged at 13 000 g for 25 min at4�C, and the supernatant (total cell lysate) was collected and theprotein content was measured by bicinchoninic acid (BCA) proteinassay kit (Pierce Biotechnology, Rockford, IL).
For immunoblot analysis, 30 lg protein was subjected toSDS–PAGE (10–15% Tris–HCl gel). The protein was transferredonto a nitrocellulose membrane and blocked with tris buffered saline(TBS)-Tween plus 5% dry milk. The membrane was probed with anappropriate primary antibody followed by a secondary horseradishperoxidase (HRP)-conjugated antibody. The protein was detected byfreshly prepared chemiluminescent solution.
Immunohistochemical analysis. Whole skin was frozen in optimalcutting temperature (OCT) medium and kept at )80�C until process-ing. Histological sections (6 lMM) were prepared from frozen tissues andfixed in acetone and stored at )80�C. The slides were incubated withappropriate primary antibodies and antiperoxidase secondary anti-body. The slides were stained with DAB reagent and counterstainedwith methyl green before mounting in permount medium.
Statistical analysis. A two-tailed Student’s t-test was used to assessthe statistical significance between the groups. A P-value of <0.05 wasconsidered statistically significant.
RESULTS
Inhibitory effect of sanguinarine on UVB exposure-mediated
cutaneous edema
Because studies have shown that UV exposure to the skin
results in cutaneous edema, which is regarded as a marker ofUV damage, we evaluated the protective effect of sanguinarine
Untreated Vehicle UVB
Sanguinarine Sanguinarine + UVB UVB + Sanguinarine
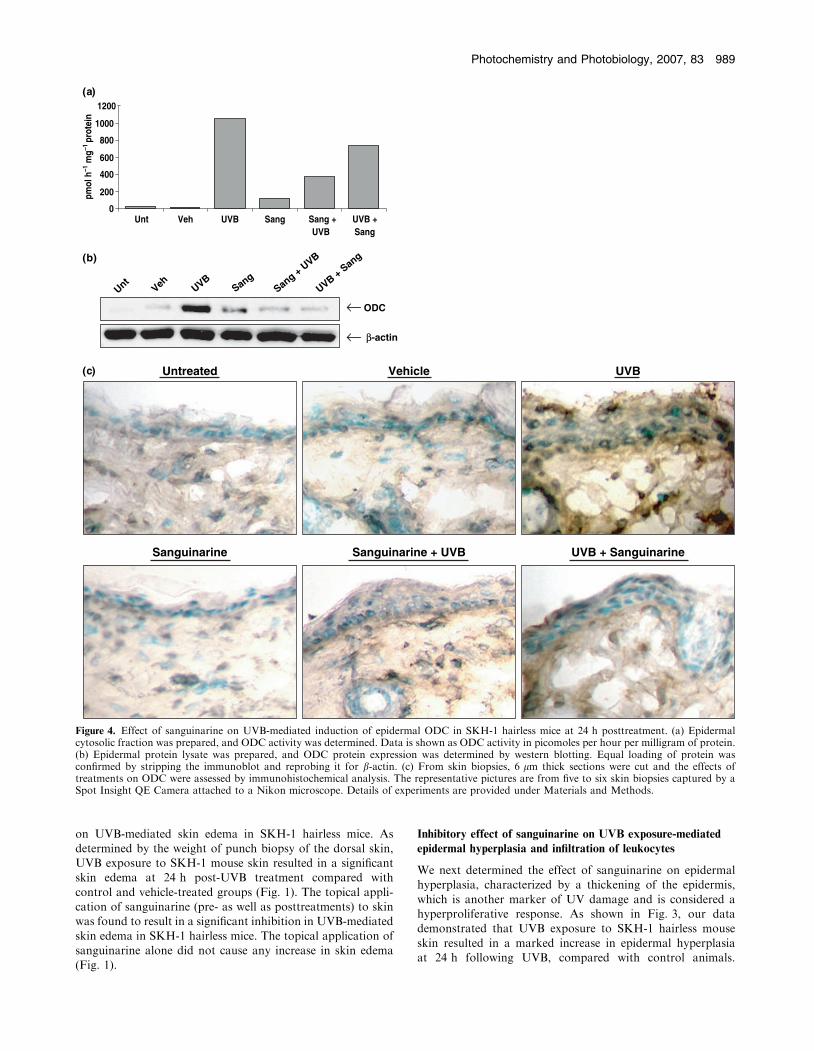
Figure 3. Effect of sanguinarine on UVB-mediated production of H2O2 in SKH-1 hairless mice. Twenty-four hours after UVB irradiation, theanimals were killed and histochemical detection of H2O2-producing cells was performed using the DAB-peroxidase reaction. Representativepictures are shown. The details are described under Materials and Methods.
988 Haseeb Ahsan et al.
on UVB-mediated skin edema in SKH-1 hairless mice. Asdetermined by the weight of punch biopsy of the dorsal skin,UVB exposure to SKH-1 mouse skin resulted in a significantskin edema at 24 h post-UVB treatment compared with
control and vehicle-treated groups (Fig. 1). The topical appli-cation of sanguinarine (pre- as well as posttreatments) to skinwas found to result in a significant inhibition in UVB-mediated
skin edema in SKH-1 hairless mice. The topical application ofsanguinarine alone did not cause any increase in skin edema(Fig. 1).
Inhibitory effect of sanguinarine on UVB exposure-mediated
epidermal hyperplasia and infiltration of leukocytes
We next determined the effect of sanguinarine on epidermalhyperplasia, characterized by a thickening of the epidermis,
which is another marker of UV damage and is considered ahyperproliferative response. As shown in Fig. 3, our datademonstrated that UVB exposure to SKH-1 hairless mouse
skin resulted in a marked increase in epidermal hyperplasiaat 24 h following UVB, compared with control animals.
Untreated Vehicle UVB
Sanguinarine Sanguinarine + UVB UVB + Sanguinarine
ODC
β-actin
Unt
Veh UVBSang
Sang + UVB
UVB + Sang
0
200
400
600
800
1000
1200(a)
(b)
(c)
Unt Veh UVB Sang Sang +UVB
UVB +Sang
pmol
h–1
mg–1
prot
ein
←
←
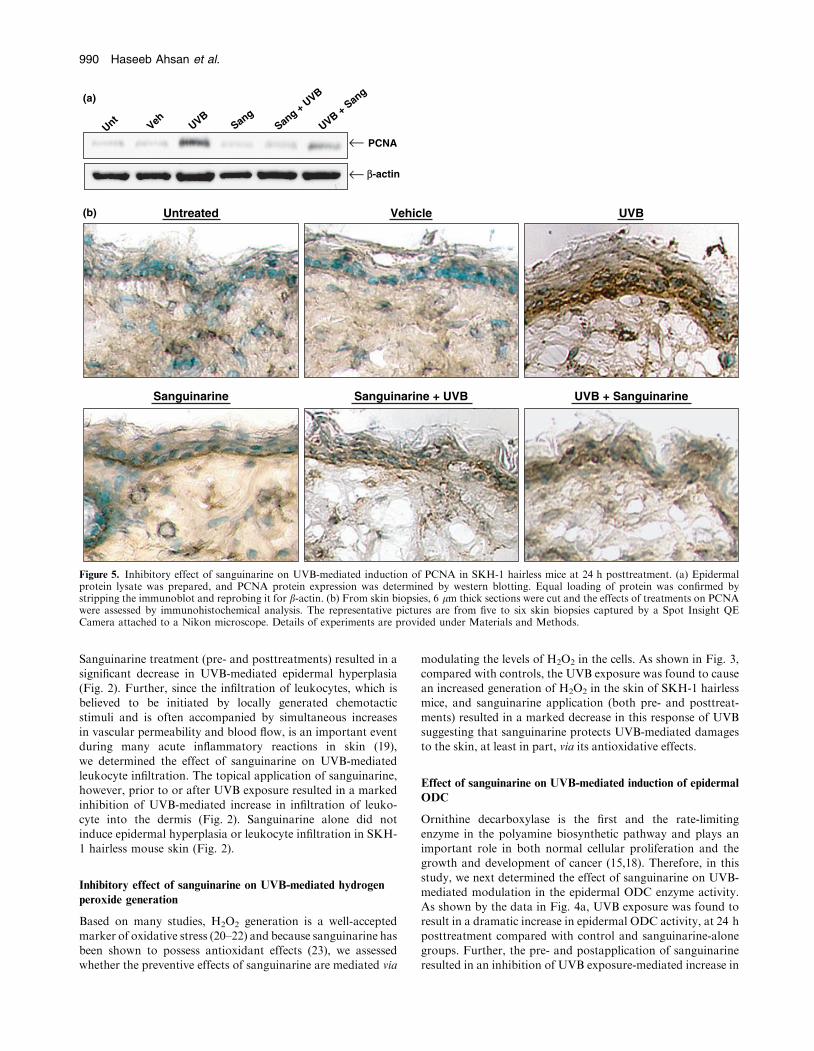
Figure 4. Effect of sanguinarine on UVB-mediated induction of epidermal ODC in SKH-1 hairless mice at 24 h posttreatment. (a) Epidermalcytosolic fraction was prepared, and ODC activity was determined. Data is shown as ODC activity in picomoles per hour per milligram of protein.(b) Epidermal protein lysate was prepared, and ODC protein expression was determined by western blotting. Equal loading of protein wasconfirmed by stripping the immunoblot and reprobing it for b-actin. (c) From skin biopsies, 6 lm thick sections were cut and the effects oftreatments on ODC were assessed by immunohistochemical analysis. The representative pictures are from five to six skin biopsies captured by aSpot Insight QE Camera attached to a Nikon microscope. Details of experiments are provided under Materials and Methods.
Photochemistry and Photobiology, 2007, 83 989
Sanguinarine treatment (pre- and posttreatments) resulted in asignificant decrease in UVB-mediated epidermal hyperplasia(Fig. 2). Further, since the infiltration of leukocytes, which isbelieved to be initiated by locally generated chemotactic
stimuli and is often accompanied by simultaneous increasesin vascular permeability and blood flow, is an important eventduring many acute inflammatory reactions in skin (19),
we determined the effect of sanguinarine on UVB-mediatedleukocyte infiltration. The topical application of sanguinarine,however, prior to or after UVB exposure resulted in a marked
inhibition of UVB-mediated increase in infiltration of leuko-cyte into the dermis (Fig. 2). Sanguinarine alone did notinduce epidermal hyperplasia or leukocyte infiltration in SKH-
1 hairless mouse skin (Fig. 2).
Inhibitory effect of sanguinarine on UVB-mediated hydrogen
peroxide generation
Based on many studies, H2O2 generation is a well-accepted
marker of oxidative stress (20–22) and because sanguinarine hasbeen shown to possess antioxidant effects (23), we assessedwhether the preventive effects of sanguinarine are mediated via
modulating the levels of H2O2 in the cells. As shown in Fig. 3,compared with controls, the UVB exposure was found to causean increased generation of H2O2 in the skin of SKH-1 hairlessmice, and sanguinarine application (both pre- and posttreat-
ments) resulted in a marked decrease in this response of UVBsuggesting that sanguinarine protects UVB-mediated damagesto the skin, at least in part, via its antioxidative effects.
Effect of sanguinarine on UVB-mediated induction of epidermal
ODC
Ornithine decarboxylase is the first and the rate-limiting
enzyme in the polyamine biosynthetic pathway and plays animportant role in both normal cellular proliferation and thegrowth and development of cancer (15,18). Therefore, in this
study, we next determined the effect of sanguinarine on UVB-mediated modulation in the epidermal ODC enzyme activity.As shown by the data in Fig. 4a, UVB exposure was found toresult in a dramatic increase in epidermal ODC activity, at 24 h
posttreatment compared with control and sanguinarine-alonegroups. Further, the pre- and postapplication of sanguinarineresulted in an inhibition of UVB exposure-mediated increase in
Untreated Vehicle UVB
Sanguinarine Sanguinarine + UVB UVB + Sanguinarine
Unt
Veh UVBSang
Sang + UVB
UVB + Sang
PCNA
(a)
(b)
β-actin
←
←
Figure 5. Inhibitory effect of sanguinarine on UVB-mediated induction of PCNA in SKH-1 hairless mice at 24 h posttreatment. (a) Epidermalprotein lysate was prepared, and PCNA protein expression was determined by western blotting. Equal loading of protein was confirmed bystripping the immunoblot and reprobing it for b-actin. (b) From skin biopsies, 6 lm thick sections were cut and the effects of treatments on PCNAwere assessed by immunohistochemical analysis. The representative pictures are from five to six skin biopsies captured by a Spot Insight QECamera attached to a Nikon microscope. Details of experiments are provided under Materials and Methods.
990 Haseeb Ahsan et al.
ODC enzyme activity. Similar effects of sanguinarine on ODC
protein expression (Fig. 4b) and immunohistochemical analy-sis (Fig. 4c) were also observed.
Inhibitory effect of sanguinarine on the protein levels of the
markers of tumor promotion viz. PCNA and Ki-67
In our next set of experiments, we determined the effect ofsanguinarine on UVB exposure-mediated modulations in theprotein levels of proliferation ⁄ tumor promotion markers
PCNA and Ki-67. PCNA is a widely accepted qualitative aswell as quantitative molecular marker of cellular proliferation.As expected, UVB exposure resulted in a marked increase in
the protein levels and immunohistochemical staining of PCNAin mouse skin epidermis (Fig. 5). Topical pre- and postappli-cation of sanguinarine were found to result in an appreciabledecrease in PCNA (Fig. 5). Immunohistochemical staining of
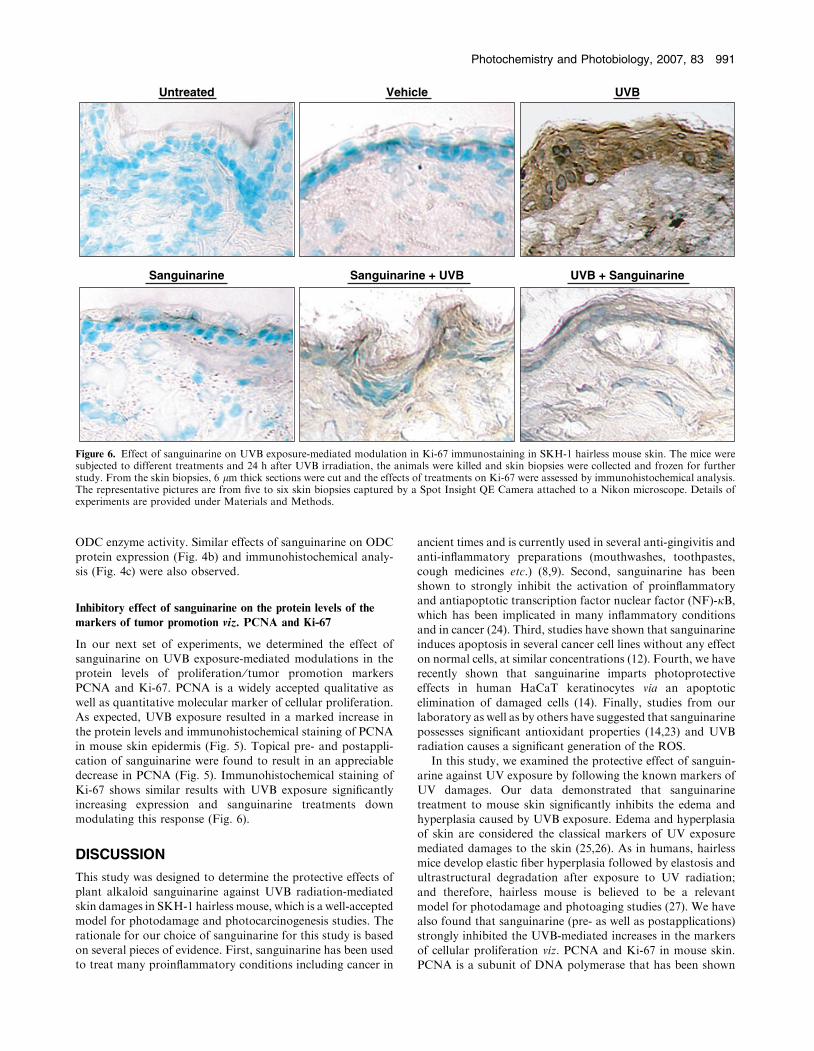
Ki-67 shows similar results with UVB exposure significantlyincreasing expression and sanguinarine treatments downmodulating this response (Fig. 6).
DISCUSSION
This study was designed to determine the protective effects ofplant alkaloid sanguinarine against UVB radiation-mediatedskin damages in SKH-1 hairless mouse, which is a well-acceptedmodel for photodamage and photocarcinogenesis studies. The
rationale for our choice of sanguinarine for this study is basedon several pieces of evidence. First, sanguinarine has been usedto treat many proinflammatory conditions including cancer in
ancient times and is currently used in several anti-gingivitis and
anti-inflammatory preparations (mouthwashes, toothpastes,cough medicines etc.) (8,9). Second, sanguinarine has beenshown to strongly inhibit the activation of proinflammatoryand antiapoptotic transcription factor nuclear factor (NF)-jB,which has been implicated in many inflammatory conditionsand in cancer (24). Third, studies have shown that sanguinarineinduces apoptosis in several cancer cell lines without any effect
on normal cells, at similar concentrations (12). Fourth, we haverecently shown that sanguinarine imparts photoprotectiveeffects in human HaCaT keratinocytes via an apoptotic
elimination of damaged cells (14). Finally, studies from ourlaboratory as well as by others have suggested that sanguinarinepossesses significant antioxidant properties (14,23) and UVBradiation causes a significant generation of the ROS.
In this study, we examined the protective effect of sanguin-arine against UV exposure by following the known markers ofUV damages. Our data demonstrated that sanguinarine
treatment to mouse skin significantly inhibits the edema andhyperplasia caused by UVB exposure. Edema and hyperplasiaof skin are considered the classical markers of UV exposure
mediated damages to the skin (25,26). As in humans, hairlessmice develop elastic fiber hyperplasia followed by elastosis andultrastructural degradation after exposure to UV radiation;
and therefore, hairless mouse is believed to be a relevantmodel for photodamage and photoaging studies (27). We havealso found that sanguinarine (pre- as well as postapplications)strongly inhibited the UVB-mediated increases in the markers
of cellular proliferation viz. PCNA and Ki-67 in mouse skin.PCNA is a subunit of DNA polymerase that has been shown
Untreated Vehicle UVB
Sanguinarine Sanguinarine + UVB UVB + Sanguinarine
Figure 6. Effect of sanguinarine on UVB exposure-mediated modulation in Ki-67 immunostaining in SKH-1 hairless mouse skin. The mice weresubjected to different treatments and 24 h after UVB irradiation, the animals were killed and skin biopsies were collected and frozen for furtherstudy. From the skin biopsies, 6 lm thick sections were cut and the effects of treatments on Ki-67 were assessed by immunohistochemical analysis.The representative pictures are from five to six skin biopsies captured by a Spot Insight QE Camera attached to a Nikon microscope. Details ofexperiments are provided under Materials and Methods.
Photochemistry and Photobiology, 2007, 83 991

to play a role in DNA replication and repair and serves as abiomarker of proliferation (28). Studies have shown that UVradiation induces Ki-67 and epidermal hyperplasia (29) andaberrant accumulation of p53 has been associated with Ki-67
in benign skin lesions (30). Similarly, UVB radiation has alsobeen shown to induce PCNA and p53 in the skin (31).Further, PCNA promoter has been shown to be transcrip-
tionally activated via wild-type p53 as a mechanism of cellularresponse to DNA damage (32). Furthermore, PCNA plays acritical role in cell cycle progression where the cyclin kinase
inhibitor (cki) WAF1 ⁄ p21 can inhibit cell cycle progression byforming a quaternary complex with cyclin, cyclin-dependentkinase (cdk) and PCNA (33). Thus, the observed inhibition
of UVB-mediated increase in PCNA by sanguinarine isimportant and it also suggests the involvement of p53signaling and a cki-cyclin-cdk network in the protective effectsof sanguinarine.
It is now established that UV exposure causes an oxidativestress in the skin that may be involved in a wide range ofconditions including immunosuppression, inflammation, can-
cer and skin aging (34–36). Studies have suggested thatsanguinarine possesses strong antioxidant activity(14,23,37,38). In this study, we have found that sanguinarine
markedly inhibits UVB exposure-mediated generation of H2O2
suggesting that the photoprotective effects of sanguinarinemay be mediated via its antioxidant effects. Also, studies haveshown that UVB exposure may cause an additional outburst of
ROS in the skin cells via infiltration of leukocytes, therebyinflicting further cellular damages (39). In this study, we havefound that both pre- and post-UVB treatment of sanguinarine
causes a decrease in the infiltration of leukocytes that furthersubstantiate our suggestion that the observed effect of san-guinarine could be through its antioxidant activity that reduces
leukocyte-mediated oxidative stress. This data is consistentwith our previous in vitro findings that sanguinarine imparts aphotoprotective effect via modulation of oxidative stress
pathways in vitro (14). Our data have also demonstrated thatthe topical application of sanguinarine dramatically inhibitedUVB-mediated induction of ODC activity as well as proteinexpression. This is an interesting observation because ODC
has been shown to be associated with cellular transformationthat is regarded as a marker of tumor promotion (40–43). Thisdata suggests that sanguinarine may be able to inhibit tumor
promotion which makes it an interesting and potential agentfor the management of UVB-induced skin cancers.
Thus, taken together, our results suggest that sanguina-
rine may provide protection against the damages caused byUVB exposure and these protective effects may be mediatedvia its antioxidant properties. Interestingly, the posttreat-ment of sanguinarine was found to impart equal protection
compared with the pretreatment suggesting that sanguina-rine-mediated responses are not sunscreen effects. Based onour study, we suggest that sanguinarine could be developed
as an agent for the management of skin cancer and otherhyperproliferative skin conditions. However, detailed in vivostudies in relevant animal models are needed to support this
suggestion.
Acknowledgements—This work was supported by funding from the
National Cancer Institute (CA114060 and CA099076) and from the
Department of Defense (W81XWH-04-1-0220).
REFERENCES1. Jemal, A., R. Siegel, E. Ward, T. Murray, J. Xu and M. J. Thun
(2007) Cancer statistics, 2007. CA Cancer J. Clin. 57, 43–66.2. Melnikova, V. O. and H. N. Ananthaswamy (2005) Cellular and
molecular events leading to the development of skin cancer.Mutat. Res. 571, 91–106.
3. Ortonne, J. P. (2002) From actinic keratosis to squamous cellcarcinoma. Br. J. Dermatol. 146 (Suppl. 61), 20–23.
4. Mukhtar, H. and C. A. Elmets (1996) Photocarcinogenesis:Mechanisms, models and human health implications. Photochem.Photobiol. 63, 356–357.
5. Cleaver, J. E. and E. Crowley (2002) UV damage, DNA repair andskin carcinogenesis. Front Biosci. 7, d1024–d1043.
6. de Gruijl, F. R. (2002) Photocarcinogenesis: UVA vs UVB radi-ation. Skin Pharmacol. Appl. Skin Physiol. 15, 316–320.
7. Bowden, G. T. (2004) Prevention of non-melanoma skin cancer bytargeting ultraviolet-B-light signalling. Nat. Rev. Cancer. 4, 23–35.
8. Lenfeld, J., M. Kroutil, E. Marsalek, J. Slavik, V. Preiningerand V. Simanek (1981) Antiinflammatory activity of quaternarybenzophenanthridine alkaloids from Chelidonium majus. PlantaMed. 43, 161–165.
9. Mandel, I. D. (1988) Chemotherapeutic agents for controllingplaque and gingivitis. J. Clin. Periodontol. 15, 488–498.
10. Walterova, D., J. Ulrichova, I. Valka, J. Vicar, C. Vavreckova, E.Taborska, R. J. Harjrader, D. L. Meyer, H. Cerna and V. Simanek(1995) Benzo[c]phenanthridine alkaloids sanguinarine andchelerythrine: Biological activities and dental care applications.Acta Univ. Palacki. Olomuc. Fac. Med. 139, 7–16.
11. Mandel, I. D. (1994) Antimicrobial mouthrinses: Overview andupdate. J. Am. Dent. Assoc. 125 (Suppl. 2), 2S–10S.
12. Ahmad, N., S. Gupta, M. M. Husain, K. M. Heiskanen andH. Mukhtar (2000) Differential antiproliferative and apoptoticresponse of sanguinarine for cancer cells versus normal cells. Clin.Cancer Res. 6, 1524–1528.
13. Adhami, V. M., M. H. Aziz, H. Mukhtar and N. Ahmad (2003)Activation of prodeath Bcl-2 family proteins and mitochondrialapoptosis pathway by sanguinarine in immortalized humanHaCaT keratinocytes. Clin. Cancer Res. 9, 3176–3182.
14. Reagan-Shaw, S., J. Breur and N. Ahmad (2006) Enhancement ofUVB radiation-mediated apoptosis by sanguinarine in HaCaThuman immortalized keratinocytes. Mol. Cancer Ther. 5, 418–429.
15. Pegg, A. E. (2006) Regulation of ornithine decarboxylase. J. Biol.Chem. 281, 14529–14532.
16. Dannenberg, A. M., Jr, B. H. Schofield, J. B. Rao, T. T. Dinh,K. Lee, M. Boulay, Y. Abe, J. Tsuruta and M. J. Steinbeck (1994)Histochemical demonstration of hydrogen peroxide production byleukocytes in fixed-frozen tissue sections of inflammatory lesions.J. Leukoc. Biol. 56, 436–443.
17. Wang, Z. Y., R. Agarwal, D. R. Bickers and H. Mukhtar (1991)Protection against ultraviolet B radiation-induced photocarcino-genesis in hairless mice by green tea polyphenols. Carcinogenesis12, 1527–1530.
18. Ahmad, N., A. C. Gilliam, S. K. Katiyar, T. G. O’Brien and H.Mukhtar (2001) A definitive role of ornithine decarboxylase inphotocarcinogenesis. Am. J. Pathol. 159, 885–892.
19. Gillitzer, R. and M. Goebeler (2001) Chemokines in cutaneouswound healing. J. Leukoc. Biol. 69, 513–521.
20. Katiyar, S. K., F. Afaq, A. Perez and H. Mukhtar (2001) Greentea polyphenol ())-epigallocatechin-3-gallate treatment of humanskin inhibits ultraviolet radiation-induced oxidative stress.Carcinogenesis 22, 287–294.
21. Charron, R. A., J. C. Fenwick, D. R. Lean and T. W. Moon(2000) Ultraviolet-B radiation effects on antioxidant status andsurvival in the zebrafish, Brachydanio rerio. Photochem. Photobiol.72, 327–333.
22. Muramatsu, S., Y. Suga, Y. Mizuno, T. Hasegawa, S. Matsuba,Y. Hashimoto, A. Volkl, J. Seitz and H. Ogawa (2005) Differen-tiation-specific localization of catalase and hydrogen peroxide,and their alterations in rat skin exposed to ultraviolet B rays.J. Dermatol. Sci. 37, 151–158.
23. Firatli, E., T. Unal, U. Onan and P. Sandalli (1994) Antioxidativeactivities of some chemotherapeutics. A possible mechanism inreducing gingival inflammation. J. Clin. Periodontol. 21, 680–683.
992 Haseeb Ahsan et al.

24. Chaturvedi, M. M., A. Kumar, B. G. Darnay, G. B. Chainy, S.Agarwal and B. B. Aggarwal (1997) Sanguinarine (pseudochel-erythrine) is a potent inhibitor of NF-kappaB activation, Ikap-paBalpha phosphorylation, and degradation. J. Biol. Chem. 272,30129–30134.
25. Soter, N. A. (1990) Acute effects of ultraviolet radiation on theskin. Semin. Dermatol. 9, 11–15.
26. Sterenborg, H. J., F. R. de Gruijl and J. C. van der Leun (1986)UV-induced epidermal hyperplasia in hairless mice. Photoderma-tology 3, 206–214.
27. Kligman, L. H. (1989) The ultraviolet-irradiated hairless mouse: Amodel for photoaging. J. Am. Acad. Dermatol. 21, 623–631.
28. Ouhtit, A., H. K. Muller, D. W. Davis, S. E. Ullrich,D. McConkey and H. N. Ananthaswamy (2000) Temporal eventsin skin injury and the early adaptive responses in ultraviolet-irradiated mouse skin. Am. J. Pathol. 156, 201–207.
29. El-Abaseri, T. B., S. Putta and L. A. Hansen (2006) Ultravioletirradiation induces keratinocyte proliferation and epidermalhyperplasia through the activation of the epidermal growth factorreceptor. Carcinogenesis 27, 225–231.
30. Soini, Y., D. Kamel, P. Paakko, V. P. Lehto, A. Oikarinen and K.V. Vahakangas (1994) Aberrant accumulation of p53 associateswith Ki67 and mitotic count in benign skin lesions. Br. J.Dermatol. 131, 514–520.
31. Hall, P. A., P. H. McKee, H. D. Menage, R. Dover and D. P.Lane (1993) High levels of p53 protein in UV-irradiated normalhuman skin. Oncogene 8, 203–207.
32. Morris, G. F., J. R. Bischoff and M. B. Mathews (1996) Tran-scriptional activation of the human proliferating-cell nuclearantigen promoter by p53. Proc. Natl Acad. Sci. USA 93, 895–899.
33. Liebermann, D. A., B. Hoffman and R. A. Steinman (1995)Molecular controls of growth arrest and apoptosis: p53-dependentand independent pathways. Oncogene 11, 199–210.
34. Halliday, G. M. (2005) Inflammation, gene mutation and photo-immunosuppression in response to UVR-induced oxidative dam-age contributes to photocarcinogenesis. Mutat. Res. 571, 107–120.
35. Wenk, J., P. Brenneisen, C. Meewes, M. Wlaschek, T. Peters, R.Blaudschun, W. Ma, L. Kuhr, L. Schneider and K. Scharffetter-Kochanek (2001) UV-induced oxidative stress and photoaging.Curr. Probl. Dermatol. 29, 83–94.
36. Sander, C. S., H. Chang, F. Hamm, P. Elsner and J. J. Thiele(2004) Role of oxidative stress and the antioxidant network incutaneous carcinogenesis. Int. J. Dermatol. 43, 326–335.
37. Vrba, J., J. Hrbac, J. Ulrichova and M. Modriansky (2004)Sanguinarine is a potent inhibitor of oxidative burst in DMSO-differentiated HL-60 cells by a non-redox mechanism. Chem. Biol.Interact. 147, 35–47.
38. Vavreckova, C., J. Ulrichova, M. Hajduch, F. Grambal, E. Weigland V. Simanek (1994) Effect of quaternary benzo[c]phenanthri-dine alkaloids sanguinarine, chelerythrine and fagaronine on somemammalian cells. Acta Univ. Palacki. Olomuc. Fac. Med. 138, 7–10.
39. Katiyar, S. K. and H. Mukhtar (2001) Green tea polyphenol())-epigallocatechin-3-gallate treatment to mouse skin preventsUVB-induced infiltration of leukocytes, depletion of antigen-presenting cells, and oxidative stress. J. Leukoc. Biol. 69, 719–726.
40. Kubota, S., H. Kiyosawa, Y. Nomura, T. Yamada and Y. Seyama(1997) Ornithine decarboxylase overexpression in mouse 10T1 ⁄ 2fibroblasts: Cellular transformation and invasion. J. Natl CancerInst. 89, 567–571.
41. Shantz, L. M., C. S. Coleman and A. E. Pegg (1996) Expression ofan ornithine decarboxylase dominant-negative mutant reverseseukaryotic initiation factor 4E-induced cell transformation.Cancer Res. 56, 5136–5140.
42. Shantz, L. M. and A. E. Pegg (1994) Overproduction ofornithine decarboxylase caused by relief of translational repres-sion is associated with neoplastic transformation. Cancer Res. 54,2313–2316.
43. Ahmad, N., A. C. Gilliam, S. K. Katiyar, T. G. O’Brien and H.Mukhtar (2001) A definitive role of ornithine decarboxylase inphotocarcinogenesis. Am. J. Pathol. 159, 885–892.
Photochemistry and Photobiology, 2007, 83 993
Related Documents











