Subscribe to PCMR and stay up-to-date with the only journal committed to publishing basic research in melanoma and pigment cell biology As a member of the IFPCS or the SMR you automatically get online access to PCMR. Sign up as a member today at www.ifpcs.org or at www.societymelanomaresarch.org If you wish to order reprints of this article, please see the guidelines here EMAIL ALERTS Receive free email alerts and stay up-to-date on what is published in Pigment Cell & Melanoma Research – click here The official journal of INTERNATIONAL FEDERATION OF PIGMENT CELL SOCIETIES · SOCIETY FOR MELANOMA RESEARCH PIGMENT CELL & MELANOMA Research To take out a personal subscription, please click here More information about Pigment Cell & Melanoma Research at www.pigment.org Adam10 haploinsufficiency causes freckle-like macules in Hairless mice Grace Tharmarajah, Laura Faas, Karina Reiss, Paul Saftig, Antony Young and Catherine D. Van Raamsdonk Submit your next paper to PCMR online at http://mc.manuscriptcentral.com/pcmr DOI: 10.1111/j.1755-148X.2011.01032.x Volume 25, Issue 5, Pages 555-565

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Subscribe to PCMR and stay up-to-date with the only journal committed to publishing basic research in melanoma and pigment cell biology
As a member of the IFPCS or the SMR you automatically get online access to PCMR. Sign up as a member today at www.ifpcs.org or at www.societymelanomaresarch.org
If you wish to order reprints of this article, please see the guidelines here
Email alErtsReceive free email alerts and stay up-to-date on what is published in Pigment Cell & Melanoma Research – click here
the official journal of
intErnational FEdEration oF PigmEnt CEll soCiEtiEs · soCiEty For mElanoma rEsEarCh
PIgMent Cell & MelAnoMAResearch
to take out a personal subscription, please click hereMore information about Pigment Cell & Melanoma Research at www.pigment.org
Adam10 haploinsufficiency causes freckle-like macules in Hairless mice
grace tharmarajah, laura Faas, Karina reiss, Paul saftig,
antony young and Catherine d. Van raamsdonk
submit your next paper to PCmr online at http://mc.manuscriptcentral.com/pcmr
doi: 10.1111/j.1755-148X.2011.01032.xVolume 25, issue 5, Pages 555-565

Adam10 haploinsufficiency causes freckle-like maculesin Hairless miceGrace Tharmarajah1, Laura Faas2, Karina Reiss3, Paul Saftig4, Antony Young5 andCatherine D. Van Raamsdonk1
1 Department of Medical Genetics, University of British Columbia, Vancouver, BC, Canada 2 Department ofBiology, University of York, Heslington, York, UK 3 Department of Dermatology, Christian-Albrecht University, Kiel,Germany 4 Biochemical Institute, Christian-Albrecht University, Kiel, Germany 5 St John's Institute ofDermatology, Division of Genetics and Molecular Medicine, King's College School of Medicine, King's CollegeLondon, Guy's Hospital, London, UK
CORRESPONDENCE Catherine D. Van Raamsdonk, e-mail: [email protected]
KEYWORDS Adam10 ⁄ Pied ⁄ Hairless ⁄ melanocytes ⁄pigmentation ⁄ freckles ⁄ Cd44
PUBLICATION DATA Received 10 August 2011,revised and accepted for publication 26 June 2012published online 26 July 2012
doi: 10.1111/j.1755-148X.2012.01032.x
Summary
The Hairless nuclear receptor co-repressor is required for hair follicle regeneration during the hair cycle.
The classical HairlessHr ⁄ HairlessHr mouse mutant loses all hair between 2 and 3 weeks of age. As the mice
age, their trunk skin develops epidermal pigmentation, a feature of human skin which is not found in
normal haired mice. In this report, we present a new, dominant mouse mutation, Pied, which arose within
a colony of HairlessHr ⁄ HairlessHr mice and causes freckle-like macules on the skin. The Pied macules
require HairlessHr homozygosity to form and are composed of localized clusters of epidermal melanocytes.
Through linkage analysis, we find that the Pied mutation is a 1914 base pair loss-of-function deletion in
the Adam10 zinc metalloprotease gene. The pathways that specifically maintain long-term pigmentation
patterns in adults are not well understood. We have identified Adam10 as an inhibitor of melanocyte
expansion in adult skin.
Introduction
Mammalian skin is pigmented by melanocytes that pro-
duce melanins. Melanocytes that are located in hair folli-
cles determine hair color, while those outside of hair
follicles in the epidermis and dermis determine skin
color. Melanocytes in the epidermis transfer pigment
containing organelles, called melanosomes, to surround-
ing keratinocytes. Abnormalities in melanocytes cause
pigmentary defects, such as albinism, vitiligo, and skin
hyperpigmentation.
The mouse trunk is densely populated with hair folli-
cles and lacks visible skin pigmentation. Melanocytes
are only occasionally observed in the interfollicular epi-
dermis and dermis (Hirobe, 1984). During development,
melanoblasts, immature melanocytes, reach individual
hair follicles by migrating first through the dermis and
then through the epidermis, collecting around and enter-
ing newly developing hair follicles beginning at E16.5
(Hirobe, 1984; Mayer, 1973; Yoshida et al., 1996). The
mechanism that restricts melanocyte growth in the
adult interfollicular trunk skin is unknown. However,
Significance
We have studied a new mouse mutant with age-related freckling, called Pied, and found that it is caused
by a deletion in the metalloprotease gene, Adam10. Our work has identified Adam10 as a regulator of
melanocyte progenitor cells in adult skin. As a membrane-linked metalloprotease, Adam10 likely sheds or
cleaves a cell surface molecule that regulates the attachment and ⁄ or proliferation of melanocytes in the
epidermis. It is possible that inhibition of Adam10 and Hairless might enhance repigmentation of human
skin in cases where it has been lost in adulthood.
ª 2012 John Wiley & Sons A/S 555
Pigment Cell Melanoma Res. 25; 555–565 ORIGINAL ARTICLE

pigmented trunk epidermis can be artificially produced
by expressing KitL, the ligand for the tyrosine kinase
receptor, C-kit, in the epidermis by way of the Cytokera-
tin 14 promoter (Kunisada et al., 1998). Melanocytes
normally persist in mouse tail, ear, and foot skin, possi-
bly because of the reduced density of hair follicles.
To remain in optimum condition, hair is continuously
grown and shed (Stenn et al., 1998). The hair cycle con-
sists of three phases: catagen, telogen, and anagen
(Paus and Cotsarelis, 1999). In catagen, the lower por-
tion of the hair follicle is deliberately destroyed by apop-
tosis (Botchkareva et al., 2006; Plikus and Chuong,
2008). During telogen, mesenchymal dermal papilla cells
at the base of the hair follicle signal to cells in the bulge
region of the permanent portion of the hair follicle,
where skin stem cells are located (Cotsarelis et al.,
1990). This triggers anagen, the regeneration of the
lower portion of the hair follicle, and the formation of a
new hair. In mice, there is a coordinate regulation of the
timing of the cycle among hair follicles, such that cycles
begin as waves moving across the body (Plikus and
Chuong, 2008). Melanocyte stem cells also reside in the
hair follicle bulge region (Nishimura et al., 2002). At the
start of each new cycle, melanocyte stem cells prolifer-
ate and populate the base of the hair follicle to contrib-
ute pigment to the keratinocytes of the newly growing
hair.
Hairless (Hr) is a retroviral insertion mutation in the
Hairless gene, which encodes a nuclear receptor co-
repressor (Potter et al., 2001; Stoye et al., 1988). Hair-
lessHr ⁄ HairlessHr mice undergo apparently normal hair
follicle development and grow a coat in their first
2 weeks (Sumner, 1924). Between 2 and 3 weeks of
age, beginning at the head and progressing toward the
tail, HairlessHr ⁄ HairlessHr mice become bald, possibly
through excessive apoptosis during catagen. Abnormal
remnants of the upper portion of the hair follicles, called
utricles, remain in the epidermis in HairlessHr ⁄ HairlessHr
mice (Panteleyev et al., 1999). HairlessHr ⁄ HairlessHr
mice also develop dermal cysts, which share some fea-
tures of sebaceous glands (Bernerd et al., 1996; Mann,
1971; Zarach et al., 2004).
The Hairless protein is transiently expressed in
keratinocytes of the outer root sheath, including the
bulge region, during telogen (Beaudoin et al., 2005;
Panteleyev et al., 2000). It has been postulated that
Hairless downregulates the expression of Soggy and
Wise, two negative regulators of Wnt signaling
(Beaudoin et al., 2005). Because Wnt signaling can ini-
tiate hair regrowth in mouse models (Lo Celso et al.,
2004; Van Mater et al., 2003), it is possible that Hair-
less plays a role in triggering anagen via a transient
de-repression of Wnt.
A number of groups have used HairlessHr ⁄ HairlessHr
mice to study the effects of UV radiation and various
carcinogens on trunk skin pigmentation (Epstein and
Epstein, 1963; Faas et al., 2008; Naganumaa et al.,
2001; Sato and Kawada, 1972). These studies have sug-
gested that the loss of the Hairless gene somehow per-
mits melanocyte survival and ⁄ or proliferation in the
trunk skin. In this article, we describe a new, spontane-
ous mouse mutation, Pied, that produces freckle-like
macules in the skin of HairlessHr ⁄ HairlessHr mice.
Results
Pied mutant mice have freckle-like macules
In a colony of inbred HRA ⁄ Skh HairlessHr ⁄ HairlessHr
mice maintained at Kings College London, we discov-
ered an animal with light brown, freckle-like spots, or
macules, on the trunk (Figure 1A). We bred this mouse
to HRA ⁄ Skh HairlessHr ⁄ HairlessHr animals without ma-
cules and found that the macules are heritable and that
they appear on the skin beginning at approximately
20 weeks of age. This mutant line of mice was named
Pied. At 5 months of age, the macules appear small and
discrete and randomly scattered (Figure 1D). Upon clo-
ser inspection, the macules have a striated appearance
(Figure 1B). The striation is the key feature that distin-
guishes Pied macules from other trunk pigmentation in
HairlessHr ⁄ HairlessHr mice.
To map the location of the Pied mutation, we crossed
5-month-old HairlessHr ⁄ HairlessHr mice with macules to
Castaneous mates. We then backcrossed each of the
F1 offspring to a HairlessHr ⁄ + mouse, produced by
crossing HRA ⁄ Skh HairlessHr ⁄ HairlessHr mice to + ⁄ +(C3HeB ⁄ FeJ) inbred mice. In 53% of the backcross
cages that were set up, HairlessHr ⁄ HairlessHr mapping
progeny with macules were produced. We selected 10
of the HairlessHr ⁄ HairlessHr mapping progeny with ma-
cules for a genome scan to look for SSLP markers with
a decreased frequency of the Castaneous allele. A defi-
ciency was found at markers on chromosome 14,
where the Hairless gene is located, as well as at mark-
ers on chromosome 9. By analyzing more densely
spaced SSLP markers on chromosome 9 in 128 more
HairlessHr ⁄ HairlessHr mapping progeny with macules,
we narrowed the Pied physical interval to 6.6 megabas-
es, close to D9Mit259 (Figure 1E). None of the 138
mapping progeny with macules inherited the castaneous
allele at both CVR4 and D9Mit123, indicating that there
is no phenocopy of the macule phenotype in wild-type
animals.
We next intercrossed HairlessHr ⁄ +; Pied ⁄ + F1 animals
that gave rise to progeny with macules in the mapping
cross. Of 48 progeny, we recovered no animals that
were solely HRA ⁄ Skh throughout the interval that flanks
Pied (CVR4-D9Mit123) (expected number = 12,
P < 0.0001, chi-square test). This shows that homozy-
gosity of Pied is lethal prior to weaning and confirms
that the macule phenotype of Pied is dominant.
We then used the Pied flanking markers to genotype
HairlessHr ⁄ HairlessHr progeny from the mapping cross
that did not develop macules. We determined that the
Tharmarajah et al.
556 ª 2012 John Wiley & Sons A/S

penetrance of the macule phenotype in HairlessHr ⁄ Hair-
lessHr; Pied ⁄ + animals is �80% at 5 months of age.
A deletion within the metalloprotease, Adam10,
causes Pied
According to the Ensembl mouse genome database,
there are 47 genes in the interval flanked by the mark-
ers CVR4 and D9Mit123 (Figure 1E). Three of the genes
in the interval are known to be embryonic lethal:
Adam10 (Hartmann et al., 2002), Tcf12 (Zhuang et al.,
1996), and Nedd4 (Fouladkou et al., 2008). We decided
to characterize the Pied homozygous phenotype to nar-
row down potential candidate genes. We intercrossed
HairlessHr ⁄ +; Pied ⁄ + F1 animals and dissected pregnant
females at E9.5. Embryonic membranes were used to
genotype the embryos using the flanking markers,
CVR4 and D9Mit123. At E9.5, we recovered several
Pied ⁄ Pied homozygotes (Figure 1F). We noted that the
Pied ⁄ Pied embryos have the following features: they
are small, their caudal regions are truncated, their som-
ites are not clearly delineated, their neural tubes are
irregularly shaped, their branchial arches are small, their
hearts are still linear, their pericardial sacs are enlarged,
and their telencephalons are small. In short, their phe-
notype closely matches the previously reported pheno-
type of E9.5 Adam10 knockout mice (Hartmann et al.,
2002).
We attempted to PCR amplify each of the 16 exons
of Adam10 in Pied ⁄ Pied DNA from E9.5-dissected
embryos. We successfully amplified all exons except
exon 11. Suspecting a deletion, we analyzed the region
around exon 11 with additional primer pairs to narrow
the non-amplifiable region and then designed pairs to
span the hypothesized deletion. One set produced a
Pied-specific product. Upon sequencing this PCR prod-
uct, we determined that there is a deletion of 1914
bases in Pied DNA, which removes all of exon 11 of
Adam10 (Figure 2A). Unexpectedly, we found a 50 base
pair sequence inserted in between the two Pied dele-
tion break points, as compared to HRA ⁄ Skh DNA (Fig-
ure 2B). The inserted sequence shows homology to
mouse L1 Line elements. Interestingly, the Pied proxi-
A B
C D
E F
Figure 1. Pied causes skin
hyperpigmentation and maps to
chromosome 9. (A) Appearance of
HairlessHr ⁄ HairlessHr; Pied ⁄ + mouse at
6 months of age. (B) Enlargement
showing striations within the macules of a
HairlessHr ⁄ HairlessHr; Pied ⁄ + mouse at
6 months of age. (C, D) Comparison of 6-
month-old HairlessHr ⁄ HairlessHr; + ⁄ + (C)
and HairlessHr ⁄ HairlessHr; Pied ⁄ + (D) trunk
skin. An example of a macule is indicated
with an arrow in D. (E) A map of the
6.6 megabase physical interval of Pied on
chromosome 9, flanked by the markers,
CVR4 and D9Mit123. For each marker, the
recombination fraction is shown above the
line and the position in megabases
according to Ensembl assembly NCBI
m37 is shown below. (F) Appearance of
Pied ⁄ Pied homozygous embryos at day
9.5. Pied ⁄ Pied embryos show abnormal
features also found in Adam10
homozygous knockout embryos: (1) small
size, (2) truncated caudal region, (3)
somites not clearly delineated, (4) irregular
neural tube, (5) small branchial arch, (6)
linear heart, (7) enlarged pericardial sac,
(8) small telencephalon.
Adam10 is mutated in Pied mice
ª 2012 John Wiley & Sons A/S 557

mal break point occurs within a 173 base pair Line-type
repetitive element. The 50 base pair insertion suggests
that either a retroviral insertion ⁄ removal event caused
the Pied deletion (Mine et al., 2007) or that an L1 ele-
ment was used to repair a double-strand break at the
Adam10 locus (Mager et al., 1985).
The spanning PCR assay was subsequently used to
genotype for Pied in all animals of the colony. To
determine whether the mutation in Adam10 is a null,
we performed a test cross between Adam10Pied ⁄ + mice
and heterozygous Adam10 knockout mice,
Adam10tm1Psa ⁄ + (Hartmann et al., 2002) (Figure 2C).
We dissected the resulting embryos at E9.5. All of the
Adam10Pied ⁄ Adam10tm1Psa embryos exhibited small
size, caudal truncation, reduced somite development,
irregular neural tubes, small branchial arches, linear
hearts, enlarged pericardial sacs, and small telencepha-
lons, phenotypes identical to those observed previously
in Adam10Pied ⁄ Adam10Pied embryos (Figure 1F) and
Adam10tm1Psa ⁄ Adam10tm1Psa embryos (Hartmann et al.,
2002).
In addition, we crossed Adam10tm1Psa ⁄ + animals to
HRA ⁄ Skh HairlessHr ⁄ HairlessHr mice and then
backcrossed the resulting HairlessHr ⁄ +; Adam10tm1Psa ⁄ +
A
B
C
E F
G
D
Figure 2. Haploinsufficiency of Adam10 causes Pied (A) Vertical lines indicate the break points that remove 1914 base pairs of the Adam10
locus in the Pied allele. (B) Sequences flanking the Pied deletion are black and underlined, and the 50 base pair novel sequence inserted
between the break points is red. (C) The outcome of a test cross between Adam10tm1Psa ⁄ + and Adam10Pied ⁄ + mice. Obs., Observed, Exp.,
Expected, N, Number. (D) Examples of E9.5 embryos from the test cross in C. All 9 of the Adam10tm1Psa ⁄ Adam10Pied E9.5 embryos exhibited
the following features: (1) small size, (2) truncated caudal region, (3) somites not clearly delineated, (4) irregular neural tube, (5) small branchial
arch, (6) linear heart, (7) enlarged pericardial sac, (8) small telencephalon. (E) Comparison of HairlessHr ⁄ HairlessHr; + ⁄ + (left) and
HairlessHr ⁄ HairlessHr; Adam10tm1Psa ⁄ + (right) animals at 5 months of age. The HairlessHr ⁄ HairlessHr; Adam10tm1Psa ⁄ + mouse exhibits striated
macules, one of which is indicated with a black arrow. (F) By RT-PCR, Adam10 is expressed in neonatal mouse trunk skin and in cultured
mouse melanocytes (melan-a cells). Controls include the ubiquitously expressed glyceraldehydes-3 phosphate dehydrogenase (Gapdh), the
melanocyte specific, dopachrome tautomerase (Dct), and the keratinocyte specific, cytokeratin 5 (Keratin 5). (G) The Pied mutation is
predicted to delete amino acids 455–505 of the Adam10 protein, comprising half of the disintegrin domain. s, signal sequence; tm,
transmembrane domain.
Tharmarajah et al.
558 ª 2012 John Wiley & Sons A/S

animals to HairlessHr ⁄ HairlessHr; + ⁄ +. We aged the
progeny for 6 months and observed that the
HairlessHr ⁄ HairlessHr; Adam10tm1Psa ⁄ + mice developed
macules (Figure 2E). These macules exhibited the same
unique striations as those on HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + mice. We quantitated the density of the
macules in HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + and
HairlessHr ⁄ HairlessHr; Adam10tm1Psa ⁄ + animals at
5 months of age and found no significant difference
between the two groups (8.5 ± 3 versus 8.0 ± 5 ma-
cules ⁄ cm2, respectively). From this data, we conclude
that haploinsufficiency of Adam10 causes macules in
HairlessHr ⁄ HairlessHr mice.
Adam10 encodes a disintegrin and metalloprotease
(ADAM) family member (Hartmann et al., 2002). Adams
are zinc-binding metalloproteases that are membrane
bound and have a large number of potential cleavage
targets (Klein and Bischoff, 2010). Using RT-PCR, we
found that Adam10 is expressed in whole neonatal
mouse skin, as well as in an immortalized mouse
melanocyte cell line, melan-a (Bennett et al., 1987)
(Figure 2F).
The deletion of exon 11 in the Adam10Pied allele is
predicted to eliminate amino acids 455 to 505, which
comprise half of the cysteine-rich disintegrin domain
(from 458 to 552) (Figure 2G). The disintegrin domain,
present in all Adam family members, is named after
its ability to bind to integrin receptors; however, the
disintegrin domain of Adam10 lacks aspartic acid resi-
dues and a Rx6DEVF sequence that are thought to
mediate interactions (Seals and Courtneidge, 2003).
Previously it was shown that deletion of the disintegrin
domain does not abrogate alpha-secretase activity of
Adam10 in vitro (Fahrenholz et al., 2000). Splicing from
exon 10 to exon 12, detected by RT-PCR (data not
shown), would cause a frameshift in the mRNA, result-
ing in the replacement of the remaining disintegrin
domain, the transmembrane domain, and the cytoplas-
mic tail with 95 aberrant amino acids, before a stop
codon is encountered. However, non-sense-mediated
mRNA decay probably would prevent the production of
such a protein.
Macules are produced by localized clusters
of melanocytes
To characterize the macule phenotype, we first per-
formed histological analysis of skin of HairlessHr ⁄ Hair-
lessHr; + ⁄ + and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +littermates at 6 months of age. We sectioned trunk
skin, noting the location of macules, and counterstained
the sections with hematoxylin and eosin. In both the
HairlessHr ⁄ HairlessHr; + ⁄ + and HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + mice, most of the epidermis was unpig-
mented (Figure 3A,B). Melanin in the epidermis was
observed in some sections of HairlessHr ⁄ HairlessHr; + ⁄ +mouse skin, as expected (Naganumaa et al., 2001). In
the macules of HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + ani-
mals, heavier concentrations of melanin were observed
mainly at the junction between the dermis and the epi-
dermis, where melanocytes typically reside (Figure 3C).
Thus, microscopically, the macules are associated with
areas of increased melanin deposition.
We next addressed whether the hyperpigmentation
of HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + skin is the result
of an increased number of melanocytes. We crossed
HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + mice to a melano-
cyte reporter line, Dct-LacZ, which expresses the mar-
ker, beta-galactosidase, in melanocytes beginning at
E10 of development (Mackenzie et al., 1997). We sacri-
ficed animals at 16 days, 3 months, and 5 months of
age, split the dermis from the epidermis using sodium
bromide and incubated the epidermal sheets in X-gal
overnight to stain melanocytes blue. At P16, there
was no difference between HairlessHr ⁄ HairlessHr; + ⁄ +and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + animals (Fig-
ure 3D, E). At 3 months of age, we observed small
groups of LacZ-positive cells in HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + skin, but not in HairlessHr ⁄ HairlessHr; + ⁄ +skin (Figure 3F,G). At 5 months of age, these groups of
LacZ-positive cells were larger and were accompanied
by melanin deposition in the surrounding epidermis
(Figure 3I,N).
We noticed that utricles in HairlessHr ⁄ HairlessHr skin
are arranged in parallel rows and that the melanocytes
of the macules are restricted to the intervening aisles
between the rows (Figure 3I,K). This might explain why
the macules appear striated by gross examination
(Figure 1B). We also found that the utricles of
HairlessHr ⁄ HairlessHr; + ⁄ + and HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + animals exhibit LacZ staining (Fig-
ure 3H,I). In control HairlessHr ⁄ HairlessHr; + ⁄ + and Hair-
lessHr ⁄ HairlessHr; Adam10Pied ⁄ + mice that do not carry
Dct-LacZ, we also observed blue staining in the utricles,
indicating that it is non-specific (Figure 3L,M).
We quantitated melanocyte density in 5-month-old
HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +; Dct-LacZ ⁄ + and
HairlessHr ⁄ HairlessHr; + ⁄ +; Dct-LacZ ⁄ + trunk epidermis
(Figure 3O). Even in the areas of the epidermis with no
obvious macule, there was a 9-fold increase in melano-
cyte density in HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +mu-
tants as compared to HairlessHr ⁄ HairlessHr; + ⁄ +(P < 0.013, student’s t-test). We conclude that Adam10
haploinsufficiency increases melanocyte numbers in
HairlessHr ⁄ HairlessHr skin.
Adam10 haploinsufficiency affects select
melanocytes in the skin
We next addressed whether the basic skin color of the
tail is increased by Adam10 haploinsufficency. We dis-
sected and split the skin from the tails of HairlessHr ⁄HairlessHr; + ⁄ + and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +3-week-old animals to examine pigmentation of the
epidermis. There was no significant difference in the
average pixel intensity of group photographed HairlessHr ⁄
Adam10 is mutated in Pied mice
ª 2012 John Wiley & Sons A/S 559

HairlessHr; + ⁄ + epidermis as compared to HairlessHr ⁄HairlessHr; Adam10Pied ⁄ + epidermis (116 ± 2.9 versus
118 ± 2.9, respectively) (Figure 4A).
In an older cohort of HairlessHr ⁄ HairlessHr; + ⁄ + and
HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + animals, we discov-
ered that macules also form on the less hairy glabrous
skin (tail, ears, and feet) of HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + animals, beginning at around 10 months
of age (Figure 4B–D). The basic color of the skin around
the macules appears unchanged in the HairlessHr ⁄HairlessHr; Adam10Pied ⁄ + animals. The macules on the
HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + glabrous skin do
not have the striated appearance of the trunk macules,
possibly because the density of utricles is less in the
glabrous skin. Altogether, these observations suggest
that Adam10 haploinsufficiency causes localized expan-
sions of select melanocytes in the skin of Hair-
lessHr ⁄ HairlessHr mice.
AD E
F G
H I
J K
L M
N
B
C
O
Figure 3. Adam10 haploinsufficiency causes localized clusters of melanocytes. (A–C) Hematoxylin and eosin stained trunk sections from
HairlessHr ⁄ HairlessHr; + ⁄ + and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + non-macule skin (A and B) and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +macule skin (C) at 6 months of age. The dermal–epidermal boundary is indicated by a black dotted line. Melanin deposition at the dermal–
epidermal junction in a macule is indicated with a white dotted circle. ep, epidermis; der, dermis. (D–M) X-gal staining (D–I, L, M) or unstained
control (J–K) epidermal sheets from HairlessHr ⁄ HairlessHr; + ⁄ + (left column) and HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + (right column) trunk
epidermis at the indicated ages. Examples of interfollicular LacZ-positive melanocytes (blue) are indicated with black arrows. Asterisks label
example utricles. In G and I, localized clusters of melanocytes are indicated with a dotted white circle. In K, the size and shape of melanin
deposition within the macule is shown. In L and M, mice that lack the Dct-LacZ transgene show that X-gal positive staining of utricles is non-
specific. (N) Enlargement of LacZ-positive cells within a macule in HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + epidermis, showing dark X-gal positive
staining in the melanocytes and melanin deposition in surrounding cells. (O) Quantitation of the number of interfollicular LacZ-positive cells in
epidermal sheets from Dct-LacZ ⁄ +; HairlessHr ⁄ HairlessHr; + ⁄ + and Dct-LacZ ⁄ +; HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + mice at 5 months of age.
Scale bars: (A–C) 25 lM, (D–M) 200 lM, (N) 25 lM.
Tharmarajah et al.
560 ª 2012 John Wiley & Sons A/S
Requirement for Hairless and CD44 in the macule
phenotype
To determine whether the HairlessHr mutation is
required for the expression of the Adam10Pied pheno-
type, we plucked the hair from the trunks of 6-month-
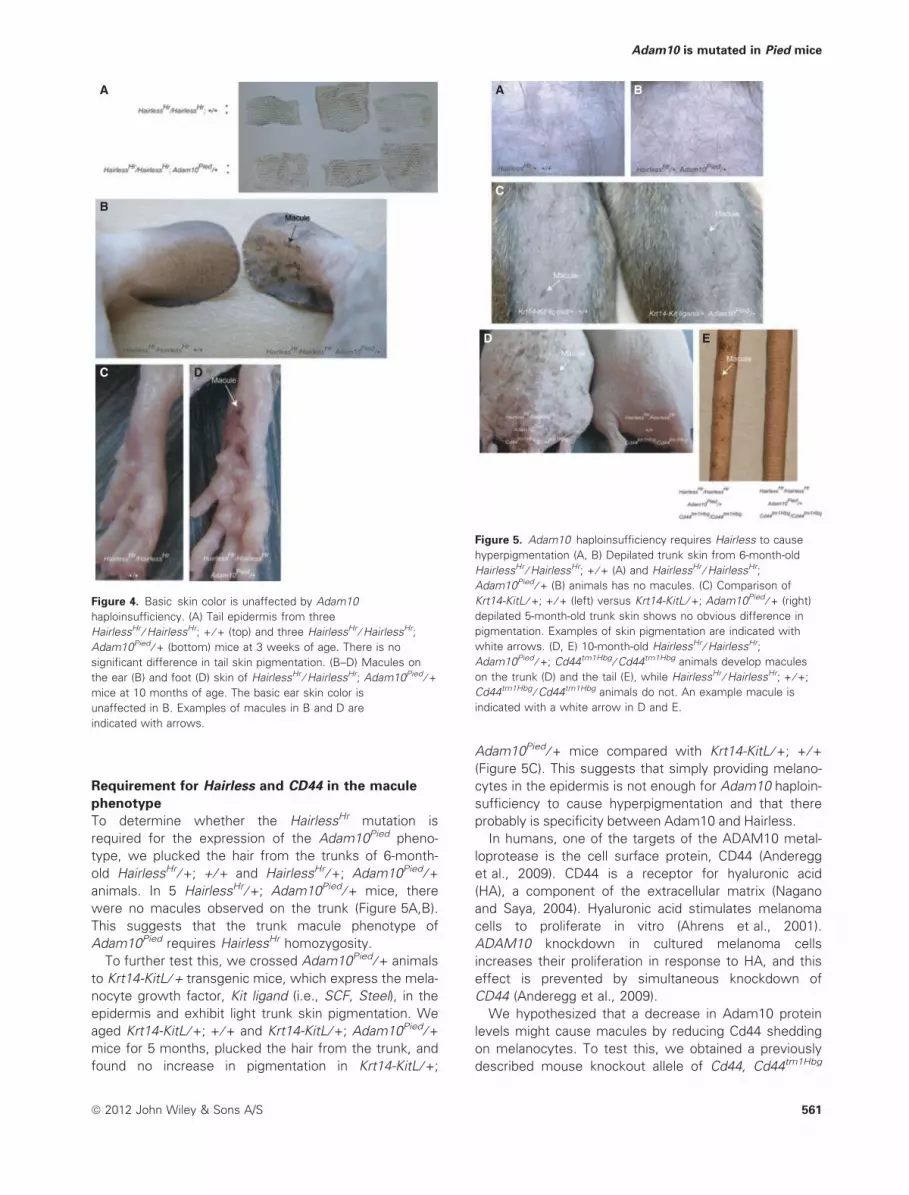
old HairlessHr ⁄ +; + ⁄ + and HairlessHr ⁄ +; Adam10Pied ⁄ +animals. In 5 HairlessHr ⁄ +; Adam10Pied ⁄ + mice, there
were no macules observed on the trunk (Figure 5A,B).
This suggests that the trunk macule phenotype of
Adam10Pied requires HairlessHr homozygosity.
To further test this, we crossed Adam10Pied ⁄ + animals
to Krt14-KitL ⁄ + transgenic mice, which express the mela-
nocyte growth factor, Kit ligand (i.e., SCF, Steel), in the
epidermis and exhibit light trunk skin pigmentation. We
aged Krt14-KitL ⁄ +; + ⁄ + and Krt14-KitL ⁄ +; Adam10Pied ⁄ +mice for 5 months, plucked the hair from the trunk, and
found no increase in pigmentation in Krt14-KitL ⁄ +;
Adam10Pied ⁄ + mice compared with Krt14-KitL ⁄ +; + ⁄ +(Figure 5C). This suggests that simply providing melano-
cytes in the epidermis is not enough for Adam10 haploin-
sufficiency to cause hyperpigmentation and that there
probably is specificity between Adam10 and Hairless.
In humans, one of the targets of the ADAM10 metal-
loprotease is the cell surface protein, CD44 (Anderegg
et al., 2009). CD44 is a receptor for hyaluronic acid
(HA), a component of the extracellular matrix (Nagano
and Saya, 2004). Hyaluronic acid stimulates melanoma
cells to proliferate in vitro (Ahrens et al., 2001).
ADAM10 knockdown in cultured melanoma cells
increases their proliferation in response to HA, and this
effect is prevented by simultaneous knockdown of
CD44 (Anderegg et al., 2009).
We hypothesized that a decrease in Adam10 protein
levels might cause macules by reducing Cd44 shedding
on melanocytes. To test this, we obtained a previously
described mouse knockout allele of Cd44, Cd44tm1Hbg
A
B
C D
Figure 4. Basic skin color is unaffected by Adam10
haploinsufficiency. (A) Tail epidermis from three
HairlessHr ⁄ HairlessHr; + ⁄ + (top) and three HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + (bottom) mice at 3 weeks of age. There is no
significant difference in tail skin pigmentation. (B–D) Macules on
the ear (B) and foot (D) skin of HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +mice at 10 months of age. The basic ear skin color is
unaffected in B. Examples of macules in B and D are
indicated with arrows.
A
C
D E
B
Figure 5. Adam10 haploinsufficiency requires Hairless to cause
hyperpigmentation (A, B) Depilated trunk skin from 6-month-old
HairlessHr ⁄ HairlessHr; + ⁄ + (A) and HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ + (B) animals has no macules. (C) Comparison of
Krt14-KitL ⁄ +; + ⁄ + (left) versus Krt14-KitL ⁄ +; Adam10Pied ⁄ + (right)
depilated 5-month-old trunk skin shows no obvious difference in
pigmentation. Examples of skin pigmentation are indicated with
white arrows. (D, E) 10-month-old HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ +; Cd44tm1Hbg ⁄ Cd44tm1Hbg animals develop macules
on the trunk (D) and the tail (E), while HairlessHr ⁄ HairlessHr; + ⁄ +;
Cd44tm1Hbg ⁄ Cd44tm1Hbg animals do not. An example macule is
indicated with a white arrow in D and E.
Adam10 is mutated in Pied mice
ª 2012 John Wiley & Sons A/S 561

(Protin et al., 1999) and crossed it to Adam10Pied ani-
mals. HairlessHr ⁄ HairlessHr; Adam10Pied ⁄ +; Cd44tm1Hbg ⁄Cd44tm1Hbg and HairlessHr ⁄ HairlessHr; + ⁄ +; Cd44tm1Hbg ⁄Cd44tm1Hbg mice were aged for 10 months and
examined. Macules were evident on the trunk (Fig-
ure 5D) and tails (Figure 5E) of HairlessHr ⁄ HairlessHr;
Adam10Pied ⁄ +; Cd44tm1Hbg ⁄ Cd44tm1Hbg mice, but not of
the HairlessHr ⁄ HairlessHr; + ⁄ +; Cd44tm1Hbg ⁄ Cd44tm1Hbg
animals. Thus, Adam10 haploinsufficiency increases
melanocyte numbers through a Cd44-independent
mechanism.
Discussion
Adam10 is a ubiquitously expressed gene with many
potential cleavage targets and has roles in cellular prolif-
eration, adhesion, migration, and cell signaling (Chantry
and Glynn, 1990; Mochizuki and Okada, 2007; Seals and
Courtneidge, 2003). Adam10 is membrane bound and
can release cleavage products both intracellularly and to
the extracellular space. For example, Adam10 cleaves
ligand-activated Notch receptors, causing the Notch
intracellular domain (NICD) to translocate to the nucleus
and regulate gene expression (Hartmann et al., 2002;
Lieber et al., 2002). Adam10 is also an alpha-secretase
for the amyloid precursor protein (APP), which is impli-
cated in the formation of plaques in Alzheimer’s disease
(Postina et al., 2004).
We have found that haploinsufficiency of Adam10
causes flat, freckle-like macules on HairlessHr ⁄ Hair-
lessHr mice, beginning on the trunk at 5 months of age
and on the glabrous skin at 10 months of age. The ma-
cules are comprised of localized clusters of melano-
cytes that are able to transfer pigment to surrounding
keratinocytes. It appears that the macules grow slowly,
which could explain why there is a latency for the
visible phenotype. The basic skin color of the glabrous
skin around the macules is unaffected. This sug-
gests that Adam10 haploinsufficiency only acts on
certain melanocytes, either through a stochastic
process or through localized differences in the skin
microenvironment.
The macules require loss of the Hairless receptor co-
repressor to form. Supplying interfollicular melanocytes
by way of the Krt14-KitL transgene is not sufficient
to induce Adam10-mediated skin hyperpigmentation in
the presence of the Hairless protein. This suggests
that Adam10 and Hairless might act within a shared
pathway.
Role of Hairless and Adam10 in pigmentation
Mouse trunk skin is normally unpigmented. Melano-
cytes migrate through the epidermis to reach hair folli-
cles, but do not persist in the interfollicular epidermis in
adults (Hirobe, 1984; Mayer, 1973; Yoshida et al., 1996).
Expression of Kit ligand in the epidermis by way of the
Keratin 14 promoter causes trunk skin hyperpigmenta-
tion (Kunisada et al., 1998). Postnatal treatment with an
antibody against the C-kit receptor in these mice
eliminates all melanocytes except the C-kit-independent
melanocyte stem cells in the hair follicle bulge (Nishim-
ura et al., 2002). Once antibody treatment is ceased,
melanocytes can be observed leaving hair follicles and
repigmenting the skin, beginning with small concentric
circles, which then expand (Nishimura et al., 2002).
Thus, transgenic Kit ligand may provide a path on
which melanocytes can ectopically migrate out of
hair follicles and also supports their growth in the
interfollicular skin.
In this study, we report that the trunk pigmentation in
HairlessHr ⁄ HairlessHr animals is associated with Dct-
LacZ-positive cells in the skin. Therefore, one role of the
Hairless protein is to restrict the presence of melano-
cytes in the trunk skin to hair follicles. Dct-LacZ-positive
cells are present at P16 and could either be lingering
migratory melanoblasts (Hirobe, 1984) or melanocytes
previously localized to hair follicles. A conditional Hair-
less allele could address this.
The expression of the Hairless protein in hair follicles
fluctuates with the hair cycle (Beaudoin et al., 2005).
Anagen hair follicles do not express Hairless. When hair
follicles enter catagen, Hairless can be detected in the
outer root sheath, including the bulge region, and in Ker-
atin-14-negative bulb cells, but not in the dermal papilla.
Hairless protein is maintained in this pattern until the
next anagen. Hairless RNA was detected in the supra-
basal cell layers of the epidermis, the hair follicle infun-
dibulum, the hair follicle matrix, and the inner root
sheath (Panteleyev et al., 2000). Furthermore, Hairless
RNA expression was detected in a microarray analysis
of primary human melanocytes and melanoma cell lines
(Ryu et al., 2007).
It has been postulated that Hairless downregulates
the expression of Soggy and Wise, two negative regu-
lators of Wnt signaling in hair follicles (Beaudoin et al.,
2005). Similarly, a Dickkopfs-related Wnt inhibitor was
identified in a screen for genes upregulated in Hair-
lessHr ⁄ HairlessHr skin (Yamaguchi et al., 2008; Zarach
et al., 2004). Because Wnt signaling can initiate hair
regrowth in mouse models (Lo Celso et al., 2004; Van
Mater et al., 2003), it is possible that Hairless plays a
role in triggering anagen via a transient de-repression
of Wnt. Wnt signaling also coordinates the activation
of epithelial and melanocyte stem cells during hair
regeneration (Rabbani et al., 2011). However, as a
direct effect, inhibition of Wnt signaling would be
expected to decrease melanocyte proliferation ⁄ growth,
not increase it. Interestingly, inhibition of Wnt signaling
was shown to downregulate Adam10 expression in
primary cortical neurons (Wan et al., 2012). Inhibited
Wnt signaling in HairlessHr ⁄ HairlessHr mice might
therefore result in decreased Adam10 expression,
which might exaggerate the effects of Adam10 hap-
loinsufficiency.
Tharmarajah et al.
562 ª 2012 John Wiley & Sons A/S

In Hairless mutant mice, the utricle appears to be
composed of an enlarged and widened infundibulum,
the uppermost portion of the hair follicle that is con-
nected to the epidermis (Zarach et al., 2004). The utri-
cles also express Keratin 10, which is normally only
expressed in the interfollicular epidermis (Zarach et al.,
2004). Thus, the utricle might ‘bridge’ the hair follicle
with the interfollicular epidermis.
Like Hairless, Adam10 also plays a critical role in
keratinocytes. Using Keratin 14-cre, the effects of
Adam10 loss after E9.5 were studied using a conditional
Adam10 knockout allele (Weber et al., 2011). Keratin
14-cre ⁄ +; Adam10tm2Psa ⁄ Adam10tm2Psa homozygous
knockout mice die soon after birth because of a skin
barrier defect in the spinous layer. Sebaceous glands
are not visible in the skin. Knockout beginning at P21
causes hair follicles to be replaced with cysts, through
disturbed differentiation. There were no obvious
changes in skin pigmentation reported following
Adam10 knockout in keratinocytes.
Lethality at E9.5 is likely due to a failure of notch sig-
naling in Adam10 homozygous mutant embryos (Hart-
mann et al., 2002). The Notch signaling pathway is also
required for maintaining melanocyte stem cells in the
hair follicle bulge region (Aubin-Houzelstein et al., 2008).
Mutations in Notch receptors or ligands cause a loss of
melanocytes (premature hair graying), the opposite
effect of the Pied mutation, which is to increase mela-
nocyte numbers in the skin (Moriyama et al., 2006;
Schouwey et al., 2007). Therefore, it does not seem
likely that Adam10Pied causes macules through
decreased production of NICD.
Interestingly, Adam10 also act as a sheddase for
Delta proteins, which serve as Notch ligands (Muraguchi
et al., 2007). Shedding of ligand from the cell membrane
reduces Notch receptor activation, and we hypothesized
that decreased Adam10 activity might actually increase
Notch signaling. However, expressing the NICD in mela-
nocytes on a HairlessHr ⁄ HairlessHr background did not
cause macules to form (C. Van Raamsdonk, personal
observation).
In the future, tissue-specific Adam10 haploinsufficien-
cy could be made on a HairlessHr ⁄ HairlessHr background
to determine whether Adam10 acts cell autonomously
in melanocytes or keratinocytes to cause macules. Simi-
larly, tissue-specific knockouts of the Hairless gene
could address whether Hairless is required in the outer
root sheath, the suprabasal layer, or melanocytes to per-
mit hyperpigmentation. These findings could help nar-
row down the search for the target of Adam10 cleavage
that mediates macule formation and clarify the role of
Hairless in regulating melanocytes.
The hair follicle serves as a reservoir for melanocytes
in the skin because it supplies a niche for melanocyte
stem cells in the outer root sheath bulge (Nishimura
et al., 2002). Repigmentation of human skin from hair
follicle melanocytes has been described in a few cases
of vitiligo, a complex disease in which epidermal mela-
nocytes are destroyed (Cui et al., 1991). Our results
indicate that Hairless and Adam10 might be useful tar-
gets with which to stimulate the expansion of melano-
cytes in the adult epidermis.
Methods
Mouse strains and genotypingPied arose spontaneously on the HRA ⁄ Skh genetic background,
which is homozygous for HairlessHr (Stoye et al., 1988; Sumner,
1924). Krt14-KitL transgenic mice (Tg(Krt14-KitL)4XTG2Bjl ⁄ J),
Adam10Tm1Psa (Hartmann et al., 2002) and Cd44tm1Hbg (Protin et al.,
1999) knockout mice were maintained on a C57Bl ⁄ 6J background,
and Dct-LacZ transgenic mice (Tg(Dct-LacZ)A12Jkn), a gift of Ian
Jackson (Mackenzie et al., 1997; Takemoto et al., 2006), were
maintained on a C3HeB ⁄ FeJ background. Ear notch sample DNA
was purified using columns (Qiagen DNeasy) and amplified
using PCR (Qiagen, Hilden, Germany). The reaction conditions
were 0.25 mM each dNTPs, 1 U Hotstar Taq (Qiagen), 1· Hotstar
Taq buffer (Qiagen), and 0.5 lM each primer. PCR consisted
of 35 cycles of 95� (30 s), 58� (1 min), and 72� (1 min). The
following primers were used for genotyping: Adam10Pied2L, 5¢-GGGAACTTTCGGGATAGCAT, Adam10Pied2R, 5¢-TTAGTGTTTTCC
CCCACAGC, Hairless mutant L, 5¢-GGGTCTCCTCAGATTGATTGAC,
Hairless mutant R, 5¢-CCATGGCCAGATTTAACACA, Hairless wild
type L, 5¢-CAGGGAGAAAACAGGTGAGG, Hairless wild type R, 5¢-ACTTGCTGTTGAGGGTCCAG, Cd44 wild type L, 5¢-TTCAACCCA-
GAGGCATCC, Cd44 wild type R, 5¢-GGCGACTAGATCCCTCCGTT,
Cd44 mutant L, 5¢-CTTGGGTGGAGAGGCTATTC, Cd44 mutant R,
5¢-AGGTGAGATGACAGGAGATC, Dct-LacZ DRp (Takemoto et al.,
2006), 5¢-CATTCATCGTCTCTCAGGAAT, Dct-LacZ LFp, 5¢-CAG-
GACACGGCTTGTCATCAT, Dct-LacZ LRp, 5¢-CCACACAGACACC-
TACCACATGCGT, Adam10 knockout neo kiel 3, 5¢-GTTG
TCACTGAAGCGGGAAGGGACTGGCTG, Adam10 knockout neo kiel
4, 5¢-GCGAACAGTTCGGCTGGCGCGAGCCCCTGA.
Positional cloning of PiedHairlessHr ⁄ HairlessHr; Adam10Pied ⁄ + animals were crossed to Cas-
t ⁄ EiJ (Jackson Laboratories) and backcrossed to HairlessHr ⁄ + ani-
mals, which were produced by crossing HRA ⁄ Skh mice to
C3HeB ⁄ FeJ (Jackson Laboratories, Bar Harbor, ME, USA). Ten Hair-
lessHr ⁄ HairlessHr; Adam10Pied ⁄ + progeny with macules were
selected for a genome scan using SSLP makers spaced every 30 cM,
and subsequently, 138 total mapping progeny were typed with 18
additional SSLP markers in the region showing linkage on chromo-
some 9. Marker sequences are available upon request. For sequenc-
ing, PCRs were purified using columns and then used as templates
for sequencing reactions using Big Dye (ABI), which were performed
in both directions.
Reverse transcription-PCRWe isolated total RNA from cultured melan-a mouse immortalized
melanocytes (Bennett et al., 1987) and neonatal trunk skin dis-
sected from C3HeB ⁄ FeJ mice using Rneasy (Qiagen). 350 ng of
RNA was reverse transcribed and amplified using One Step RT-
PCR (Life Technologies, Carlsbad, CA, USA). Primers spanning
introns were used as follows: Adam10L, 5¢-TGAAGGATTAT
CTTACAATGTGGA, Adam10R, 5¢-AAAGGGAAGTGTCCCTCTTCA,
Krt5L, 5¢-ACCTTCGAAACACCAAGCAC, Krt5R, 5¢-TCCTCCAG
CTCTGTCAGCTT, DCT-L, 5¢-AGCAGACGGAACACTGGACT, DCT-R,
5¢-GCATCTGTGGAAGGGTTGTT, Gapdh-L, 5¢-AACTTTGGCATTGTG-
GAAGG, Gapdh-R, 5¢-ATGTAGGCCATGAGGTCCAC.
Adam10 is mutated in Pied mice
ª 2012 John Wiley & Sons A/S 563

HistologyTrunk skin from 6-month-old animals was removed, fixed in 4%
paraformaldehyde, and embedded in paraffin. Eight-micron sec-
tions were cut, stained with hematoxylin and eosin, and exam-
ined by light microscopy. To assess the number of melanocytes
in the epidermis of Dct-LacZ transgenic mice, we dissected trunk
skin and adhered it epidermis-side down on a glass slide with
vacuum grease. The samples were incubated in 2 M sodium bro-
mide for 1 h at 37 Celsius. The dermis was peeled away, and the
epidermis was rinsed in PBS and stained for 16 h at 25�C in buf-
fer containing 100 mM sodium phosphate (pH 7.3), 2 mM MgCl2,
0.01% sodium deoxycholate, 0.02% NP-40, 5 mM potassium fer-
ricyanide, 5 mM potassium ferrocyanide, 1 mg ⁄ ml 5-bromo-4-
chloro-3-indolyl-b-D-galactopyranoside (X-gal), and 2.5% dimethyl-
formamide. Epidermal sheets were photographed using stereomi-
croscopy, and areas were calculated using IMAGE J (National
Institutes of Health, Bethesda, MD, USA). For tail scale
pigmentation, pieces of 3-week-old tail skin were removed,
adhered epidermis-side down to a glass slide with vacuum
grease, and incubated in 2 M sodium bromide for 2 h at 37�C.
The dermis was removed, the epidermis was photographed, and
the pixel intensity of each tail skin sample was determined using
Image J. For all histological examinations, at least three individu-
als of each genotype were examined.
Statistical analysisError bars represent the standard error of the mean. P values were
calculated using Student’s t-test (melanocyte density, tail scale
analysis) or chi-square analysis (embryonic lethality).
Acknowledgements
We thank Dr. Dorothy Bennet for melan-a cells (Bennett et al.,
1987). Funding for this research was provided by a Canadian
Cancer Society grant to Dr. Van Raamsdonk (019055). Paul Saftig
was supported through a grant from the Deutsche Forschungs-
gemeinschaft (DFG, SFB877-A3) and the Interuniversity Attraction
Poles Program IUAP P6 ⁄ 58 of the Belgian Federal Science Policy
Office.
References
Ahrens, T., Assmann, V., Fieber, C., Termeer, C., Herrlich, P., Hof-
mann, M., and Simon, J.C. (2001). CD44 is the principal mediator
of hyaluronic-acid-induced melanoma cell proliferation. J. Invest.
Dermatol. 116, 93–101.
Anderegg, U., Eichenberg, T., Parthaune, T. et al. (2009). ADAM10
is the constitutive functional sheddase of CD44 in human mela-
noma cells. J. Invest. Dermatol. 129, 1471–1482.
Aubin-Houzelstein, G., Djian-Zaouche, J., Bernex, F., Gadin, S., Del-
mas, V., Larue, L., and Panthier, J.J. (2008). Melanoblasts’ proper
location and timed differentiation depend on Notch ⁄ RBP-J signal-
ing in postnatal hair follicles. J. Invest. Dermatol. 128, 2686–2695.
Beaudoin 3rd, G.M., Sisk, J.M., Coulombe, P.A., and Thompson,
C.C. (2005). Hairless triggers reactivation of hair growth by pro-
moting Wnt signaling. Proc. Natl Acad. Sci. USA 102, 14653–
14658.
Bennett, D.C., Cooper, P.J., and Hart, I.R. (1987). A line of non-
tumorigenic mouse melanocytes, syngeneic with the B16 mela-
noma and requiring a tumour promoter for growth. Int. J. Cancer
39, 414–418.
Bernerd, F., Schweizer, J., and Demarchez, M. (1996). Dermal
cysts of the rhino mouse develop into unopened sebaceous
glands. Arch. Dermatol. Res. 288, 586–595.
Botchkareva, N.V., Ahluwalia, G., and Shander, D. (2006). Apopto-
sis in the hair follicle. J. Invest. Dermatol. 126, 258–264.
Chantry, A., and Glynn, P. (1990). A novel metalloproteinase origi-
nally isolated from brain myelin membranes is present in many
tissues. Biochem. J. 268, 245–248.
Cotsarelis, G., Sun, T.T., and Lavker, R.M. (1990). Label-retaining
cells reside in the bulge area of pilosebaceous unit: implications
for follicular stem cells, hair cycle, and skin carcinogenesis. Cell
61, 1329–1337.
Cui, J., Shen, L.Y., and Wang, G.C. (1991). Role of hair follicles
in the repigmentation of vitiligo. J. Invest. Dermatol. 97, 410–
416.
Epstein, J.H., and Epstein, W.L. (1963). A study of tumor types
produced by ultraviolet light in hairless and hairy mice. J. Invest.
Dermatol. 41, 463–473.
Faas, L., Venkatasamy, R., Hider, R.C., Young, A.R., and Sou-
myanath, A. (2008). In vivo evaluation of piperine and syn-
thetic analogues as potential treatments for vitiligo using a
sparsely pigmented mouse model. Br. J. Dermatol. 158, 941–
950.
Fahrenholz, F., Gilbert, S., Kojro, E., Lammich, S., and Postina, R.
(2000). Alpha-secretase activity of the disintegrin metalloprotease
ADAM 10. Influences of domain structure. Ann. N. Y. Acad. Sci.
920, 215–222.
Fouladkou, F., Landry, T., Kawabe, H., Neeb, A., Lu, C.,
Brose, N., Stambolic, V., and Rotin, D. (2008). The ubiquitin
ligase Nedd4-1 is dispensable for the regulation of PTEN
stability and localization. Proc. Natl Acad. Sci. USA 105,
8585–8590.
Hartmann, D., De Strooper, B., Serneels, L. et al. (2002). The disin-
tegrin ⁄ metalloprotease ADAM 10 is essential for Notch signalling
but not for alpha-secretase activity in fibroblasts. Hum. Mol.
Genet. 11, 2615–2624.
Hirobe, T. (1984). Histochemical survey of the distribution of the
epidermal melanoblasts and melanocytes in the mouse during
fetal and postnatal periods. Anat. Rec. 208, 589–594.
Klein, T., and Bischoff, R. (2010). Active metalloproteases of the a
disintegrin and metalloprotease (ADAM) family: biological func-
tion and structure. J. Proteome Res. 10, 17–33.
Kunisada, T., Yoshida, H., Yamazaki, H., Miyamoto, A., Hemmi, H.,
Nishimura, E., Shultz, L.D., Nishikawa, S., and Hayashi, S.
(1998). Transgene expression of steel factor in the basal layer of
epidermis promotes survival, proliferation, differentiation and
migration of melanocyte precursors. Development 125, 2915–
2923.
Lieber, T., Kidd, S., and Young, M.W. (2002). kuzbanian-mediated
cleavage of Drosophila Notch. Genes Dev. 16, 209–221.
Lo Celso, C., Prowse, D.M., and Watt, F.M. (2004). Transient acti-
vation of beta-catenin signalling in adult mouse epidermis is suffi-
cient to induce new hair follicles but continuous activation is
required to maintain hair follicle tumours. Development 131,
1787–1799.
Mackenzie, M.A., Jordan, S.A., Budd, P.S., and Jackson, I.J.
(1997). Activation of the receptor tyrosine kinase Kit is required
for the proliferation of melanoblasts in the mouse embryo. Dev.
Biol. 192, 99–107.
Mager, D.L., Henthorn, P.S., and Smithies, O. (1985). A Chinese G
gamma + (A gamma delta beta) zero thalassemia deletion: com-
parison to other deletions in the human beta-globin gene cluster
and sequence analysis of the breakpoints. Nucleic Acids Res. 13,
6559–6575.
Mann, S.J. (1971). Hair loss and cyst formation in hairless and rhino
mutant mice. Anat. Rec. 170, 485–499.
Mayer, T.C. (1973). The migratory pathway of neural crest cells into
the skin of mouse embryos. Dev. Biol. 34, 39–46.
Tharmarajah et al.
564 ª 2012 John Wiley & Sons A/S

Mine, M., Chen, J.M., Brivet, M. et al. (2007). A large genomic
deletion in the PDHX gene caused by the retrotranspositional
insertion of a full-length LINE-1 element. Hum. Mutat. 28, 137–
142.
Mochizuki, S., and Okada, Y. (2007). ADAMs in cancer cell prolifer-
ation and progression. Cancer Sci. 98, 621–628.
Moriyama, M., Osawa, M., Mak, S.S. et al. (2006). Notch sig-
naling via Hes1 transcription factor maintains survival of mel-
anoblasts and melanocyte stem cells. J. Cell Biol. 173, 333–
339.
Muraguchi, T., Takegami, Y., Ohtsuka, T. et al. (2007). RECK modu-
lates Notch signaling during cortical neurogenesis by regulating
ADAM10 activity. Nat. Neurosci. 10, 838–845.
Nagano, O., and Saya, H. (2004). Mechanism and biological signifi-
cance of CD44 cleavage. Cancer Sci. 95, 930–935.
Naganumaa, M., Yagi, E., and Fukuda, M. (2001). Delayed induction
of pigmented spots on UVB-irradiated hairless mice. J. Dermatol.
Sci. 25, 29–35.
Nishimura, E.K., Jordan, S.A., Oshima, H., Yoshida, H., Osawa,
M., Moriyama, M., Jackson, I.J., Barrandon, Y., Miyachi, Y.,
and Nishikawa, S. (2002). Dominant role of the niche in
melanocyte stem-cell fate determination. Nature 416, 854–
860.
Panteleyev, A.A., Botchkareva, N.V., Sundberg, J.P., Christiano,
A.M., and Paus, R. (1999). The role of the hairless (hr) gene in
the regulation of hair follicle catagen transformation. Am. J.
Pathol. 155, 159–171.
Panteleyev, A.A., Paus, R., and Christiano, A.M. (2000). Patterns of
hairless (hr) gene expression in mouse hair follicle morphogene-
sis and cycling. Am. J. Pathol. 157, 1071–1079.
Paus, R., and Cotsarelis, G. (1999). The biology of hair follicles. N.
Engl. J. Med. 341, 491–497.
Plikus, M.V., and Chuong, C.M. (2008). Complex hair cycle domain
patterns and regenerative hair waves in living rodents. J. Invest.
Dermatol. 128, 1071–1080.
Postina, R., Schroeder, A., Dewachter, I. et al. (2004). A disintegrin-
metalloproteinase prevents amyloid plaque formation and
hippocampal defects in an Alzheimer disease mouse model.
J. Clin. Invest. 113, 1456–1464.
Potter, G.B., Beaudoin 3rd, G.M., Derenzo, C.L., Zarach, J.M.,
Chen, S.H., and Thompson, C.C. (2001). The hairless gene
mutated in congenital hair loss disorders encodes a novel nuclear
receptor corepressor. Genes Dev. 15, 2687–2701.
Protin, U., Schweighoffer, T., Jochum, W., and Hilberg, F. (1999).
CD44-deficient mice develop normally with changes in subpopu-
lations and recirculation of lymphocyte subsets. J. Immunol. 163,
4917–4923.
Rabbani, P., Takeo, M., Chou, W., Myung, P., Bosenberg, M., Chin,
L., Taketo, M.M., and Ito, M. (2011). Coordinated activation of
Wnt in epithelial and melanocyte stem cells initiates pigmented
hair regeneration. Cell 145, 941–955.
Ryu, B., Kim, D.S., Deluca, A.M., and Alani, R.M. (2007). Com-
prehensive expression profiling of tumor cell lines identifies
molecular signatures of melanoma progression. PLoS ONE 2,
e594.
Sato, T., and Kawada, A. (1972). Mitotic activity of hairless
mouse epidermal melanocytes: its role in the increase of mela-
nocytes during ultraviolet radiation. J. Invest. Dermatol. 58,
392–395.
Schouwey, K., Delmas, V., Larue, L., Zimber-Strobl, U., Strobl, L.J.,
Radtke, F., and Beermann, F. (2007). Notch1 and Notch2
receptors influence progressive hair graying in a dose-dependent
manner. Dev. Dyn. 236, 282–289.
Seals, D.F., and Courtneidge, S.A. (2003). The ADAMs family of
metalloproteases: multidomain proteins with multiple functions.
Genes Dev. 17, 7–30.
Stenn, K., Parimoo, S., and Prouty, S. (1998). Growth of the
hair follicle: a cycling and regenerating biological system. In
Molecular Basis of Epithelial Appendage Morphogenesis, C.,
Chuong, ed. (Austin, TX: RG Landes Company), 111–130.
Stoye, J.P., Fenner, S., Greenoak, G.E., Moran, C., and Coffin, J.M.
(1988). Role of endogenous retroviruses as mutagens: the
hairless mutation of mice. Cell 54, 383–391.
Sumner, F. (1924). Hairless mice. J. Hered. 15, 475–481.
Takemoto, Y., Keighren, M., Jackson, I.J., and Yamamoto, H.
(2006). Genomic localization of a Dct-LacZ transgene locus: a
simple assay for transgene status. Pigment Cell Res. 19,
644–645.
Van Mater, D., Kolligs, F.T., Dlugosz, A.A., and Fearon, E.R. (2003).
Transient activation of beta -catenin signaling in cutaneous kerati-
nocytes is sufficient to trigger the active growth phase of the
hair cycle in mice. Genes Dev. 17, 1219–1224.
Wan, X.Z., Li, B., Li, Y.C., Yang, X.L., Zhang, W., Zhong, L., and
Tang, S.J. (2012). Activation of NMDA receptors upregulates a
disintegrin and metalloproteinase 10 via a Wnt ⁄ MAPK signaling
pathway. J. Neurosci. 32, 3910–3916.
Weber, S., Niessen, M.T., Prox, J. et al. (2011). The disinte-
grin ⁄ metalloproteinase Adam10 is essential for epidermal
integrity and Notch-mediated signaling. Development 138, 495–
505.
Yamaguchi, Y., Passeron, T., Hoashi, T. et al. (2008). Dickkopf 1
(DKK1) regulates skin pigmentation and thickness by affecting
Wnt ⁄ beta-catenin signaling in keratinocytes. FASEB J. 22, 1009–
1020.
Yoshida, H., Kunisada, T., Kusakabe, M., Nishikawa, S., and Nishik-
awa, S.I. (1996). Distinct stages of melanocyte differentiation
revealed by anlaysis of nonuniform pigmentation patterns. Devel-
opment 122, 1207–1214.
Zarach, J.M., Beaudoin 3rd, G.M., Coulombe, P.A., and Thomp-
son, C.C. (2004). The co-repressor hairless has a role in epi-
thelial cell differentiation in the skin. Development 131, 4189–
4200.
Zhuang, Y., Cheng, P., and Weintraub, H. (1996). B-lymphocyte
development is regulated by the combined dosage of three basic
helix-loop-helix genes, E2A, E2-2, and HEB. Mol. Cell. Biol. 16,
2898–2905.
Adam10 is mutated in Pied mice
ª 2012 John Wiley & Sons A/S 565
Related Documents











