Sem Biodiversitas Universitas A ISBN : 978-602-14989-0-3 Prosiding minar Nasional dan Ekologi Tropika In ( BioETI ) Andalas, Padang, 27 September 20 i ndonesia 014

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

i
ProsidingSeminar Nasional
Biodiversitas dan Ekologi Tropika Indonesia
( BioETI )Universitas Andalas, Padang, 27 September 2014
ISBN : 978-602-14989-0-3
i
ProsidingSeminar Nasional
Biodiversitas dan Ekologi Tropika Indonesia
( BioETI )Universitas Andalas, Padang, 27 September 2014
ISBN : 978-602-14989-0-3
i
ProsidingSeminar Nasional
Biodiversitas dan Ekologi Tropika Indonesia
( BioETI )Universitas Andalas, Padang, 27 September 2014
ISBN : 978-602-14989-0-3

ii
Prosiding Seminar NasionalDalam rangka Ulang Tahun ke-52 Jurusan BiologiFakultas Matematika dan Ilmu Pengetahuan AlamUniversitas Andalas
Hak Cipta Dilindungi Undang-undangCopyright@2014ISBN : 978-602-14989-0-3
Editor :
Diterbitkan oleh :
Jurusan Biologi, FMIPA,Universitas Andalas
Dr. Erizal Mukhtar Universitas AndalasProf. Dr. Syamsuardi Universitas AndalasProf. Dr. Syafruddin Ilyas Universitas Sumatera UtaraDr. Revis Asra Universitas Jambi

iii
Kata Pengantar
Prosiding ini merupakan kumpulan makalah-makalah yang telah dipresentasikan di dalam
Seminar Nasional Biodiversitas dan Ekologi Tropika Indonesia di kampus Universitas
Andalas pada tanggal 27 September 2014 dengan tema “Pemanfaatan keanekaragaman
hayati tropika dalam menghadapi tantangan pasar bebas asean ”. Ada tiga topik utama
yang dibahas dalam seminar tersebut, yaitu Bioproses, Ekologi dan Biodiversitas.
Akhirnya kami berharap agar publikasi ini dapat dimanfaatkan bagi segala pihak demi
kemajuan bangsa.
Padang, Nopember 2014
Editors

iv
KATA SAMBUTAN KETUA PANITIA
Keanekaragaman hayati (biodiversitas) merupakan sumberdaya penting yang memberikan
manfaat baik langsung maupun tidak langsung bagi manusia dan lingkungan. Fakta bahwa
telah terjadi laju penurunan keanekaragaman hayati baik yang disebabkan oleh kehilangan
habitat, kebakaran hutan, eksplorasi yang berlebihan, introduksi jenis invasif baik sengaja
ataupun tidak sengaja, polusi dan perubahan iklim sangat mengkawatirkan kita semua.
Penelitian dibidang biologi seyogyanya mampu memberikan kontribusi untuk meng-
antisipasi dan/atau meminimalisasi keadaan tersebut.
Sejalan dengan visi dan misi utama Jurusan Biologi Universitas Andalas yakni pengkajian
dan penyelamatan sumber daya alam tropika dan sebagai institusi pengemban tridarma
perguruan tinggi maka Jurusan Biologi FMIPA UNAND dalam rangka Ulang Tahunnya
yang ke-51 mengadakan Seminar Nasional Biodiversitas dan Ekologi Tropika Indonesia.
Seminar Nasional Biodiversitas dan Ekologi Tropika Indonesia, dengan singkatan “Semnas
BioETI” dirancang sebagai forum untuk berbagi ilmu, menginformasikan dan
mendiskusikan hasil-hasil temuan ilmiah dalam bidang biodiversitas dan ekologi tropis
Indonesia. Seminar ini diharapkan menjadi forum untuk menginformasikan dan
mendiskusikan hasil-hasil temuan terbaru sehingga dapat diaplikasikan dalam kehidupan
nyata dan dapat menunjang kejayaan bangsa. Pada seminar ini dihadirkan tiga orang
pemakalah utama yang merupakan pakar-pakar dibidangnya masing-masing. Sesi paralel,
54 makalah dari berbagai bidang ilmu biologi yang akan dipresentasikan oleh peneliti-
peneliti dari 21 institusi di seluruh Indonesia.
Terakhir, Panitia mengucapkan terimakasih kepada semua pihak yang telah banyak
membantu sampai terlaksananya acara ini. Selanjutnya Panitia juga mengucapkan terima
kasih kepada Dekan Fakultas MIPA dn Rektor Universitas Andalas serta pihak sponsor
seperti Bank Nagari, PT Kencana Sawit Indonesia dan Wilmar International Plantation.
.
Padang, 27 September 2014Ketua Panitia
Dr. Syaifullah

v
KATA SAMBUTAN KETUA JURUSAN BIOLOGIFMIPA UNIVERSITAS ANDALAS
Assalamu’alaikum Wr.Wb.
Pertama-tama marilah kita bersama-sama mengucapkan puji dan syukur ke hadirat Allah
SWT yang selalu melimpahkan rahmat dan hidayhnya-Nya kepada kita semua, sehingga
kita berkumpul di ruangan ini untuk mengikuti acara seminar BioETI. Selawat beriring
salam kita kirimkan untuk junjungan kita Nabi Besar Muhammad SAW, pemimpin umat
sepanjang zaman.
Selanjutnya, perlu kami sampaikan bahwa Seminar Nasional Biologi sering kita lakukan,
baik yang dilakukan oleh Mahasiswa Biologi dengan Himabionya, Dosen bersama
Himabio. Namun seminar ini kami dikemas dalam satu tema yang direncanakan sebagai
seminar rutin setiap tahun (Seminar Tahunan Jurusan Biologi). Ucapan terima kasih kami
ucapkan kepada Ketua Jurusan Biologi sebelumnya (Dr. Anthoni Agustin, MS) yang telah
merancang kegiatan ini.
Seminar Kedua tahun ini yang diketuai oleh Dr. Syaifullah yang dirancang sesuai dengan
Visi Misi Jurusan Biologi FMIPA Universitas Andalas.Terimakasih kepada semua anggota
Panitia yang telah menyusun tema besar yaitu BioETI. Seminar BioETI diharapkan akan
menjadi agenda tahunan Jurusan Biologi FMIPA Unand dan kedepannya akan di
laksanakan setiap tahun dengan tema yang berbeda.
Seminar ini dapat terlaksana dengan baik tentulah berkat bantuan dan kerjasama semua
pihak. Oleh sebab itu,melalui forum ini saya mengucapkan terima kasih dan memberikan
penghargaan kepada semua pihak yang telah terlibat mulai dari persiapan sampai
pelaksanaan hari ini. Semoga Allah SWT memberi pahala atas jerih payahnnya.
Terimakasih.
Padang, 27 September 2014Ketua Jurusan BiologiFMIPA Univ. Andalas
Dr. Jabang Nurdin, M.Si

vi
DAFTAR ISI
Kata Pengantar ………………………………..……………………………………………………. iiiSambutan Ketua Panitia Seminar …………………………………………………………..…….… ivSambutan Ketua Jurusan Biologi …………….…………………………………………………..…. v
ALANIYAH SYAFAREN, RIDWAN SANTOSO, EGI YUDHA WINATA DANROFIZA YOLANDAKeanekaragaman jenis tumbuhan paku epifit di perkebunan kelapa sawit disekitar kampus Universitas Pasir Pengaraian …………………………………………………..….. 1
ANANDA, HERBERT SIPAHUTAR DAN MEIDA NUGRAHALIADaya fertilitas Mencit (Mus musculus) betina pasca pemberian air seduhan kopi peroral …….….. 6
ARYUDA YOZA SELFA, NASRIL NASIR DAN FUJI ASTUTI FEBRIAUji daya hambat formulasi minyak Piper aduncum sebagai pestisida nabati pengendalijamur Fusarium pada batang Hylocereus polyrhizus secara Invitro ……………………………….. 10
DEWI MURNI DAN YUHELSA PUTRAOptimalisasi produksi biogas Eceng Gondok dengan Hydrothermal pretreatment(production optimalisation of water hyacinth biogas with hydrothermal pretreatment) …..……….. 15
DITA OSRIANTI, NASRIL NASIR DAN FUJI ASTUTI FEBRIAUji daya hambat biopestisida formulasi minyak daun cengkeh dengan penambahanminyak Kayu Manis sebagai pengendali Colletotrichum pada Buah Naga secara Invitro ……….. 24
DIYONA PUTRI, HENNY HERWINA, RIJAL SATRIA DAN ALAN HANDRUJenis Semut (Hymenoptera: Formicidae) pada Macaranga spp. (Euphorbiaceae) diCagar Alam Bukit Barisan, Rimbo Panti dan Pangean, Sumatera Barat ........................................... 28
DWI ANINDITYA, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIAPemberian Fungi Mikoriza Arbuskula (FMA) pada tanaman Calopogonium muconoides(Desv.) dan Centrosema pubescens (Benth.) untuk bioremediasi lahan tercemar Merkuri …...…... 36
ENGGAR UTARIKearifan lokal masyarakat adat Baduy dalam pemanfaatan sumber daya hayati ……………..….. 42
FAUZIAH, RIZALDI DAN WILSON NOVARINOInteraksi interspesies tiga jenis Kuntul (Ardeidae) di Cagar Alam Baringin Sati,Sumatera Barat …………………………………………………………………………………….. 52
FAUZUR RAHMI, EFRIZAL DAN RESTI RAHAYUEfek ekstrak etanol rimpang Temu Mangga (Curcuma mangga Val.) terhadap kadar guladarah dan kolesterol Mencit Putih (Mus musculus) jantan yang diinduksi Aloksan ……………….58
FEBRI SEMBIRING DAN HERBERT SIPAHUTARAnalisis kualitas spermatozoa Mencit (Mus musculus) pasca pemberian air seduhan kopi ………..63
FITRA SUZANTI, RETNO WIDHYASTUTI, SUCI RAHAYU DAN AGUS SUSANTOIndeks keanekaragaman jenis serangga pada beberapa kelompok umur Kelapa Sawitdi kebun Aek Pancur (PPKS), Tanjung Morawa, Sumatera Utara …………………………………69
FITRI ROZA WIRANATA, MAIRAWITA DAN DAHELMIJenis-Jenis dan prevalensi soil transmitted helminth pada anak-anak di Olo BangauBatang Anai Kabupaten Padang Pariaman …………………………………………………...……75
FITRI WAHYUNI, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIAPotensi beberapa tanaman dalam mengakumulasi Merkuri pada tanah bekasTambang Emas ……………………………………………………………………………….…… 83

vii
FUJI ASTUTI FEBRIA, ANTHONI AGUSTIEN DAN S.P. RAHAYUIsolasi dan uji resistensi merkuri bakteri endogen tanah bekas tambang emasKabupaten Sijunjung ……………………………………………………………………………… 91
HAFIZATUR RAHMA, NURMIATI* DAN ANTHONI AGUSTIENKandungan Polifenol dan Aktivitas Antioksidan Jamur Tiram(Pleourotus spp.) Beraneka Warna ………………………………………………………………. . 96
HALIATUR RAHMA, MARTINIUS, RATNA WULANDARI DAN TRIMARYONODeteksi patogen terbawa benih pada tanaman Jagung ……………………………………………104
HARSUNA YUMNA, NURMIATI DAN PERIADNADIStudi komparatif Sagu (Metroxylon Rottb) sebagai media bibit produksi terhadap pertumbuhanmiselium dan aktifitas amilase dan selulase Jamur Merang (Volvariella volvacea (Bull.)Sing.) ...109
HASNI RUSLAN, PRIMA LADY DAN HILDA SILFIAKeanekaragaman serangga pada dua habitat berbeda di kawasan Cilintang,Taman Nasional Ujung Kulon, Banten …………………………………………………….……. 116
HAVIZA ANUGRA, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIAPotensi Monochoria vaginalis dalam mengakumulasi diperairan tercemar Merkuri (Hg) ….…. 122
INDAH FAJARWATI, EFRIZAL DAN RESTI RAHAYUPengaruh Gambir (Uncaria gambir Roxb) terhadap kadar gula darah padaMencit Putih Jantan (Mus musculus) yang diinduksi Aloksan ……………………………….…. 127
INDRA ANGGRIAWAN, PERIADNADI DAN NURMIATIInventarisasi Basidiomycetes di Gunung Singgalang Sumatera Barat ……………………….….134
IZMIARTI , JABANG NURDIN, MISREN AHYUNI DAN DEA RAHAYU SILVIANIKeanekaragaman dan penyebaran Kerang (Pelecypoda) di perairan Tanjung MutiaraDanau Singkarak Sumatera Barat ..................................................................................................140
JABANG NURDIN DAN IZMIARTIPerbandingan kepadatan populasi dan sebaran ukuran cangkang kerang Donax fabaGmelin, 1792 (Lamellibranchiata : Donacidae) berdasarkan kedalaman substrat diperairan pantai Bungus Teluk Kabung, Kota Padang …………………………………………. 145
MAIRAWITA, RESTI RAHAYU, DAHELMI DAN ROBBY JANNATANInventarisasi Kecoak di Pasar Tradisional dan Rumah Sakit di Kota Padang, Sumatera Barat…..149
MARDHA TILLAH, WILSON NOVARINO DAN RIZALDIStudi morfologi feses mamalia ……………………………………………………………………154
MARDHIYETTI, ZULFADLI SYARIF DAN NOVIRMAN JAMARUNInduksi kalus pada hipokotil tanaman Turi (Sesbania grandiflora) dengan menggunakanBAP yang dikombinasikan dengan beberapa konsentrasi auksin secara In-Vitro ……………..… 161
MELIYA WATI DAN ELZA SAFITRIKeanekaragaman makanan dan ukuran lambung Rana cancrivora Gravenhorst(Anura : Ranidae) pada dataran tinggi dan dataran rendah Sumatera Barat ……………………. 165
MILDAWATI, ARDINIS ARBAIN, MAHFUD HUDA DAN HERMANSAHMakromorfologi organ vegetatif dan mikromorfologi spora Asplenium tenerum G. Forstdari Gunung Marapi di Sumatera Barat …………………………………………………………. 171
NETTI ARYANI, EFAWANI DAN NUR ASIAHPengkayaan vitamin E pada pakan untuk pematangan gonad ikan mali(Labiobarbusfestivus , Heckel) ………………………………………………………………. 177

viii
NIKEN AYU PAMUKAS DAN MULYADIPenerapan sistem resirkulasi pada proses domestikasi dan pembesaran Ikan Juaro(Pangasius polyuranodon) ……………………………………………………………………… 183
NURUL ALIFAH, ZOZY ANELOI NOLI DAN SUWIRMENRespon tanaman Bayur (Pterospermum javanicum Jungh.) terhadap inokulan FungiMikoriza Arbuskula (FMA) pada lahan bekas Tambang Semen Padang ………………….…….193
PUTRI KUMALASARI, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIAPotensi tanaman Digitaria ciliaris (Retz.) Koeler dalam meremediasi tanah tercemarMerkuri (Hg) pada lahan bekas Tambang Emas di Sijunjung, Sumatera Barat …………………197
RAHMADHANI FITRIBeberapa jenis mikroorganisme probiotik dan manfaatnya dalam kehidupan …………………. 203
RELSAS YOGICAPotensi sektor pertanian dan perkebunan Kabupaten Pasaman Barat untukmenghadapi pasar bebas ASEAN ……………………………………………………………….212
REVIS ASRA, SYAMSUARDI DAN MANSYURDINKarakteristik morfologi polen Daemonorops draco (Willd.) Blume ……………………………218
RINI OKTAVIA, DAHELMI DAN HENNY HERWINAKupu-kupu pemakan buah di kawasan Taman Nasional Kerinci Seblat (TNKS)Wilayah IV Kabupaten Solok Selatan, Sumatera Barat ………………………………………… 223
ROFIZA YOLANDAJenis-jenis Gastropoda (Moluska) pada ekosistem Lamun di Pantai NirwanaPadang, Sumatera Barat ................................................................................................................ 230
SERLIAFRI SUSANTI, HENNY HERWINA DAN DAHELMIJenis Semut (Hymenoptera: Formicidae) di perkebunan Pisang Air Dingin,Lubuk Minturun, Sumatera Barat ……………………………………………………………….233
SHYNTIA HARSARI, NASRIL NASIR, FUJI ASTUTI FEBRIADaya hambat formulasi minyak daun kayu manis dengan penambahan minyakserai wangi sebagai pestisida nabati dalam menghambat Fusarium buah naga secara Invitro …240
SOLFIYENI, SYAMSUARDI, CHAIRUL, WELLA YURANTI DAN AFRIDA YULIAKeanekaragaman tumbuhan asing Invasif pada vegetasi semak belukar HutanPendidikan dan Penelitian Biologi Universitas Andalas ………………………………………. 245
SYAIFULLAH, ANAS SALSABILA DAN DENNY PUTRIThe diversity of Snakehead Fishes (Channa spp.) of West Sumatra andits morphological variation …………………………………………………………..………… 250
UCOP HAROENEkstraksi, identifikasi dan purifikasi limbah Jus Jeruk sebagai Feed Additive Alami ………….257
VIVI FITRIANI DAN ARMEIN LUSI ZESWITAAnalisis mikroba pada Kerang air tawar (Contradens contradens) di Danau SingkarakKabupaten Solok Sumatra Barat …………………………………………………………….…. 262
WARNETY MUNIR, INDRA JUNAIDI ZAKARIA DAN NELMIAnalisis tingkat kematangan gonad ikan mungkuih Sicyopterus macrostetholepis(Bleeker)hidup di Sungai Batang Kuranji Kota Padang berdasarkan umur, panjang dan berat tubuh …… 265

ix
WELLA YURANTI, SYAMSUARDI DAN SOLFIYENIJenis-jenis tumbuhan invasif di Hutan Pendidikan Dan Penelitian Biologi (HPPB) ………….. 274
WINCE HENDR DAN NAWIR MUHARInventarisasi jenis Kodok (Ranidae) sebagai komoditi ekspor di Sumatera Barat ……………..278
WITA PUSPITA SARI, HENNY HERWINA, DAHELMI DAN ERNIWATIJenis-jenis Hymenoptera sebagai Serangga Pengunjung pada Tanaman Mentimun(Cucumis sativus L. Cucurbitaceae) di Lubuk Minturun, Kota Padang danSungai Pua, Kabupaten Agam ………………………………………………………………….282
YEMPITA EFENDI DAN YUSRAStudi eksplorasi bakteri dari saluran pencernaan Ikan Nila (Oreochromis niloticus) yangdibudidayakan di Karamba Jaring Apung Danau Maninjau, Sumatera Barat ………………...289
YOSI RAHMAN RESTI RAHAYU DAN DAHELMIEfektivitas beberapa insektisida aerosol dengan metode glass jar dan semprot terhadapKecoak Jerman (Blattella germanica L.) Strain Plz-Smrd …………………………………….296
YUSRA DAN YEMPITA EFENDIKarakterisasi bakteri terseleksi Bacillus sp. 28 dari Budu, sebagai kandidat Biopreservatif ….302
ZA’AZIZA RIDHA JULIA, NURMIATI DAN PERIADNADIPenggunaan Air Kelapa, Air Cucian Beras dan Air Rendaman Jagung terhadappertumbuhan Miselium Jamur Kuping Hitam (Auricularia polytricha (mont.) Sacc)dalam media pembibitan dan poduksi …………………………………………………………310
ZUHRI SYAM , CHAIRUL DAN INDAH PRAFITRI YUSAKeanekaragaman Gulma pada kebun Kopi (coffea arabica l.) di Nagari Balingka,Kecamatan Ampek Koto, Kabupaten Agam …………………………………………………..315

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman jenis tumbuhan paku epifit di perkebunankelapa sawit di sekitar kampus Universitas Pasir Pengaraian
ALANIYAH SYAFAREN, RIDWAN SANTOSO, EGI YUDHA WINATA DAN ROFIZAYOLANDA
Program Studi Pendidikan Biologi, Fakultas Keguruan dan Ilmu Pendidikan, Universitas Pasir PengaraianE-mail: [email protected]
ABSTRAKTelah dilakukan penelitian mengenai keanekaragaman tumbuhan paku epifit pada bulan Februari hingga Maret 2014 diperkebunan kelapa sawit di sekitar kampus Universitas Pasir Pengaraian. Penelitian ini bertujuan untuk mengetahuikeanekaragaman tumbuhan paku epifit pada kelapa sawit dari 3 lokasi yaitu pada bagian gerbang masuk kampus, bagiantengah kampus dan di belakang kampus. Sampel dikoleksi secara purposive sampling dengan menggunakan plotberukuran 10 x 30 m. Didapatkan sebanyak 15spesies tumbuhan paku pada penelitian ini yaitu Antropyum callifolium, A.lineatum, A. nidus, A. platyneuron, Davalia denticulata, D. majuscula, D. trichomanoides, Drymo-glosum piloselloides,Goniophlebium persicifolium, Nephrolepis bisserata, N. falcata, Oleandra pistillaris, Polypodium verrucosum,Sellaginella willdenowii dan Thelypteris sp. Nilai indeks diversitas (H’) berkisar antara 1,25-1,74, nilai keseragaman (E)berkisar antara 0,67-0,84 dan nilai dominansi (C) berkisar antara 0,2-0,7. Dan dapat disimpulkan bahwa keanekaragamantumbuhan paku epifit pada batang kelapa sawit di sekitar kampus Universitas Pasir Pengaraian berada pada kategorirendah.
Key words: keanekaragaman, tumbuhan paku, perkebunan kelapa sawit
Pendahuluan
Indonesia merupakan salah satu negara yangterkenal akan negara hutan hujan tropis danmemiliki tingkat keanekaragaman jenistumbuhan yang terbanyak kedua setelah Brazil(Efendi, Haspari dan Nuraini, 2013). Salahjenis tumbuhan yang sering ditemukan adalahtumbuhan paku (Arini dan Kinho, 2012). DiIndonesia terdapat kurang lebih sebanyak 1.300jenis tumbuhan paku yang hidup pada berbagaihabitat (Sunarmi dan Sarwono, 2004). Salahsatu tempat hidup tumbuhan paku seringdijumpai adalah pada perkebunan kelapa sawit.Hal ini disebabkan pada perkebunan tersebutmerupakan ekosistem yang kompleks yangdapat ditumbuhi oleh berbagai jenis tumbuhan,yang bisa menyokong kehidupan daritumbuhan paku (Marisa, Erinda, Handayanidan Untari, 2013).
Pengkajian tumbuhan paku di Indonesiabelum begitu mendapat perhatian, padahaltumbuhan paku memiliki banyak manfaat.
Diantaranya tumbuhan paku dapatdimanfaatkan sebagai sayur-sayuran, obattradisional, serta tanaman hias yang bernilaiekonomis (Sastrapradja, Afriastini, Darnaedidan Witdjaja, 1979). Salah satu hasil penelitianmelaporkan bahwa tumbuhan paku khususnyaCyathea sp., mempunyai manfaat besar bagikeseimbangan ekosistem hutan antara lainsebagai pencegah erosi dan pengatur tata gunaair (Widhiastuti, Aththorick, dan Sari, 2006).Namun ada juga spesies yang bersifat sebagaigulma yaitu Nephrolepis sp. (Pribadi danAnggraini, 2011).
Beberapa penelitian mengenai tumbuhanpaku di Sumatera telah dilaporkan diantaranya:Arbain dan Chairul (1990) melaporkansebanyak 15 jenis dari 6 famili di beberapajalan utama Kotamadya Padang; Rahmat(1993) mengenai jenis-jenis paku epifit yangterdapat di HPPB;Hernawati (1995) 61 jenisdari 5 famili di Gunung Kerinci, TamanNasional Kerinci Seblat (TNKS);Mahfuz(1995) melaporkan 120 jenis dari 6 famili di

Alaniyah Syafaren, Ridwan Santoso, Egi Yudha Winata dan Rofiza Yolanda 2
Hutan Gunung Tujuh Kawasan Taman KerinciSeblat; Widhiastuti, Aththorick, dan Sari(2006) terdapat 44 spesies tumbuhan paku;Hartini (2006) meneliti tentang tumbuhan pakudi Cagar Alam Sago Malintang, Sumatera Baratdan Aklimatisasinya di Kebun Raya Bogormenemukan 17 spesies tumbuhan paku; Lamid(2007) 25 jenis dari 7 famili di KawasanWisata Perkampungan Minangkabau PadangPanjang; dan Mildawati dan Arbain (2011)melaporkan sebanyak 14 spesies tumbuhanpaku di taman Hutan Raya Bung Hatta KotaPadang. Sedangkan informasi mengenaitumbuhan paku di Kabupaten Rokan Hulubelum pernah dilakukan.
Di Rokan Hulu, Riau ditemukan banyakperkebunan kelapa sawit. Salah satunya adalahperkebunan kelapa sawit di sekitar kampusUniversitas Pasir Pengaraian, Rokan Hulu,Riau. Di perkebunan ini banyak sekali dijumpaitumbuhan paku yang hidup pada batang pohonkelapa sawit. Akan tetapi sampai saat ini belumada informasi mengenai keberadaan tumbuhanpaku di perkebunan kelapa sawit tersebut.Berdasarkan latar belakang diatas, makadilakukanlah penelitian mengenaikeanekaragaman jenis tumbuhan paku epifit diperkebunan kelapa sawit warga di sekitarkampus Universitas Pasir Pengaraian, RokanHulu, Riau.
BAHAN DAN METODE
Penelitian ini telah dilaksanakan pada bulanFebruari hingga Maret 2014 di perkebunankelapa sawit warga di sekitar kampusUniversitas Pasir Pengaraian.Alat dan bahan yang digunakan dalampenelitian ini adalah meteran, pancang, talirafia, kamera digital, pisau, dan alat-alat tulis.
Metode yang digunakan dalam penelitian iniadalah metode purposive sampling denganteknik pengambilan sampel menggunakan plotberukuran 10 x 30 m2 dengan pengulangansebanyak 3 kali pengulangan pada tiga stasiunyaitu pada gerbang kampus, pertengahankampus dan di belakang kampus. Semua jenis
tumbuhan paku yang berada pada batangkelapa sawit di dalam plot dihitung dan difotoserta diidentifikasi dengan menggunakan bukuacuan Sastrapradja, Afriastini, Darnaedi, danWitdjaja (1979) dan Olsen (2007).Data yang didapatkan kemudian dianalisadengan cara sebagai berikut :1. Indeks Deversitas (Krebs,1989)
H' = - ( ∑ pi ln pi )Keterangan :H' = Indeks keanekaragaman jenisPi = Probabilitas penting untuk tiap species = ni/Nni = Jumlah individu dari masing-masing speciesN = Jumlah seluruh individuKriteria hasil nilai indeks keanekaragamanadalah :H’ ≤ 3,32 : keanekaragaman rendah3,32 H’ < 9,97 : keanekaragaman sedangH’≥ 9,97 : keanekaragaman tnggi2. Indeks Keseragaman (Krebs, 1989)
E = H’/ H maks (ln S)Dimana :E : nilai indeks keseragamanH’ : nilai indeks keanekaragamanH maks : nilai ln dari jumlah spesies (S)Kriteria hasil nilai indeks keseragaman adalah :E < 0,4 : keseragaman rendah0,4 < E < 0,6 : keseragaman sedangE > 0,6 : keseragaman tinggi3. Indeks dominansi (Marrugaran, 1987)
C = ∑ (ni/N)2
Dimana :C = indeks dominansini = nilai individu tiap jenisN = jumlah individu seluruh jenisKriteria hasil nilai indeks dominansi:0< C < 0,5 : tidak ada jenis yang mendominansi0,5 < C < 1 : terdapat jenis yang mendominansi
HASIL DAN PEMBAHASAN
Dari penelitian yang telah dilakukan padalokasi penelitian didapatkan tumbuhan pakusebanyak 15 spesies, yaitu : Antropyumcallifolium, A. lineatum, A. nidus, A.platyneuron, Davalia denticulata, D. majuscula,D. trichomanoides, Drymoglosum pilo-
Alaniyah Syafaren, Ridwan Santoso, Egi Yudha Winata dan Rofiza Yolanda 3
1.251.6 1.74
0.7 0.670.84
0.320.7
0.2
0
0.5
1
1.5
2
Stasiun 1 Stasiun 2 Stasiun 3
H E C
selloides, Goniophlebium persicifolium,Nephrolepis bisserata, N. falcata, Oleandrapistillaris, Polypodium verrucosum,Sellaginella willdenowiidan Thelypterissp.,dengan jumlah sebanyak 3168 individu dandapat dilihat pada Tabel 1.
Tabel 1. Spesies dan jumlah tumbuhan pakuepifit yang didapatkan pada lokasipenelitian.
No SpesiesSta.1
Sta.2Sta.3
Jml(Ind)
1 Antropyum callifolium - 2 - 22 Asplenium lineatum - 1 - 13 Asplenium nidus 2 9 17 284 Asplenium platyneuron - 72 165 2375 Davalia denticulata 760 - - 7606 Davalia majuscula 1 139 - 1407 Davalia trichomanoides - 154 78 2328 Drymoglossum piloselloides - - 2 29 Goniophlebium persicifolium 141 263 - 404
10 Nephrolepis bisserata 398 1 144 54311 Nephrolepis falcata 352 255 137 74412 Oleandra pistillaris - 1 - 113 Polypodium verrucosum - - 46 4614 Sellaginella willdenowii - 1 - 115 Thelypteris sp. - - 27 27
Total 1654 898 616 3168
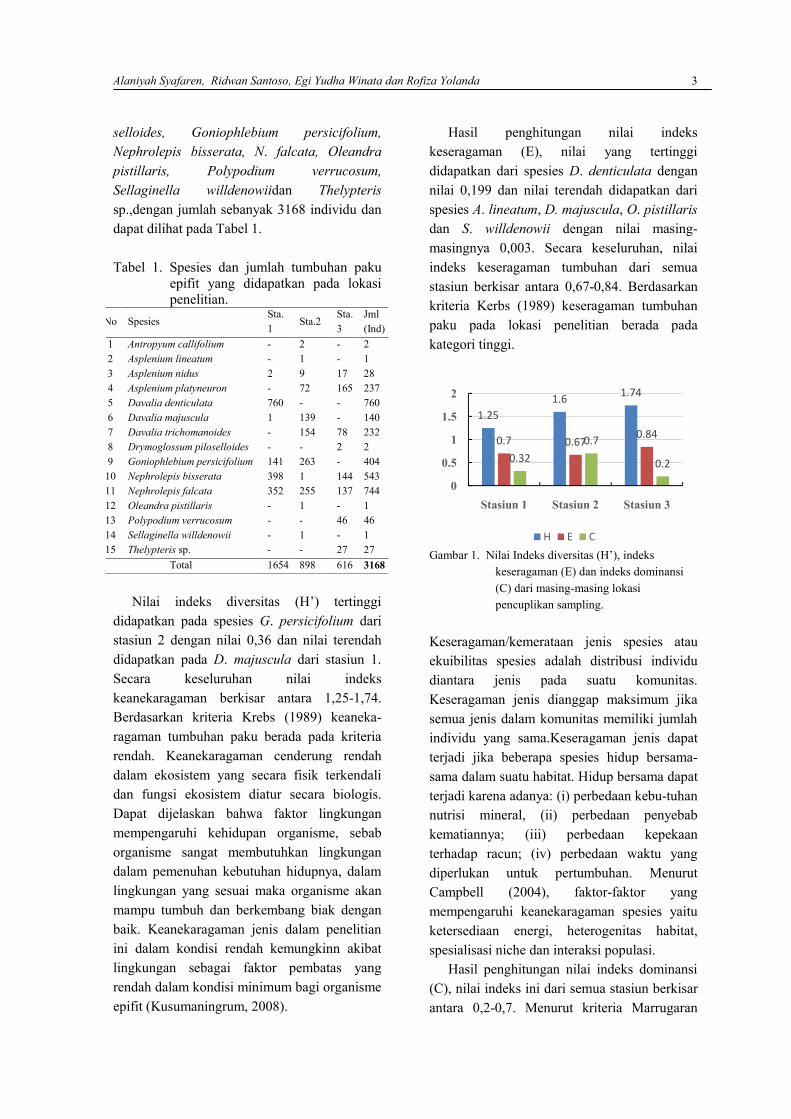
Nilai indeks diversitas (H’) tertinggididapatkan pada spesies G. persicifolium daristasiun 2 dengan nilai 0,36 dan nilai terendahdidapatkan pada D. majuscula dari stasiun 1.Secara keseluruhan nilai indekskeanekaragaman berkisar antara 1,25-1,74.Berdasarkan kriteria Krebs (1989) keaneka-ragaman tumbuhan paku berada pada kriteriarendah. Keanekaragaman cenderung rendahdalam ekosistem yang secara fisik terkendalidan fungsi ekosistem diatur secara biologis.Dapat dijelaskan bahwa faktor lingkunganmempengaruhi kehidupan organisme, sebaborganisme sangat membutuhkan lingkungandalam pemenuhan kebutuhan hidupnya, dalamlingkungan yang sesuai maka organisme akanmampu tumbuh dan berkembang biak denganbaik. Keanekaragaman jenis dalam penelitianini dalam kondisi rendah kemungkinn akibatlingkungan sebagai faktor pembatas yangrendah dalam kondisi minimum bagi organismeepifit (Kusumaningrum, 2008).
Hasil penghitungan nilai indekskeseragaman (E), nilai yang tertinggididapatkan dari spesies D. denticulata dengannilai 0,199 dan nilai terendah didapatkan darispesies A. lineatum, D. majuscula, O. pistillarisdan S. willdenowii dengan nilai masing-masingnya 0,003. Secara keseluruhan, nilaiindeks keseragaman tumbuhan dari semuastasiun berkisar antara 0,67-0,84. Berdasarkankriteria Kerbs (1989) keseragaman tumbuhanpaku pada lokasi penelitian berada padakategori tinggi.
Gambar 1. Nilai Indeks diversitas (H’), indekskeseragaman (E) dan indeks dominansi(C) dari masing-masing lokasipencuplikan sampling.
Keseragaman/kemerataan jenis spesies atauekuibilitas spesies adalah distribusi individudiantara jenis pada suatu komunitas.Keseragaman jenis dianggap maksimum jikasemua jenis dalam komunitas memiliki jumlahindividu yang sama.Keseragaman jenis dapatterjadi jika beberapa spesies hidup bersama-sama dalam suatu habitat. Hidup bersama dapatterjadi karena adanya: (i) perbedaan kebu-tuhannutrisi mineral, (ii) perbedaan penyebabkematiannya; (iii) perbedaan kepekaanterhadap racun; (iv) perbedaan waktu yangdiperlukan untuk pertumbuhan. MenurutCampbell (2004), faktor-faktor yangmempengaruhi keanekaragaman spesies yaituketersediaan energi, heterogenitas habitat,spesialisasi niche dan interaksi populasi.
Hasil penghitungan nilai indeks dominansi(C), nilai indeks ini dari semua stasiun berkisarantara 0,2-0,7. Menurut kriteria Marrugaran

Alaniyah Syafaren, Ridwan Santoso, Egi Yudha Winata dan Rofiza Yolanda 4
(1987) terdapat adanya spesies yangmendominansi, hingga menandakan strukturkomunitas tidak stabil dan terjadi tekananekologis. Hal ini disebabkan karena besarnyakeberadaan jenis paku tersebut, dan jugadikarenakan rendahnya keberadaan tumbuhanpaku yang lainnya. Dua hal tersebutdikarenakan kemampuan tumbuh denganpengaruh faktor abiotik yang baik. MenurutPramono (1992) pertumbuhan selaindipengaruhi oleh faktor genetic, juga sangatdipengaruhi oleh interaksinya denganlingkungan, seperti kondisi tanah, iklim,mikroorganisme, dan juga kompetisi denganorganisme lain.
Umumnya semakin ekstrim kondisilingkungan, baik karena iklim, tanah atauketinggian tempat yang bertambah, makan akansemakin berkurang keragaman komposisi jenisvegetasi dan satu atau dua jenis akan semakindominan. Tumbuh-tumbuan yang mempunyaiadaptasi tinggilah yang bisa hidup bahkanmendominasi di suatu daerah. Selain itudipengaruhi pula oleh pertumbuhan dari bibitatau kecambah dari suatu jenis (Efendi, Hansari,Nuraini, 2013).
KESIMPULAN
Dari penelitian yang telah dilaksanakan, makadapat diambil beberapa kesimpulan yaitusebanyak 15 spesies tumbuhan paku didapatkanpada penelitian ini, yaitu Antropyum callifolium,A. lineatum, A. nidus, A. platyneuron, Davaliadenticulata, D. majuscula, D. trichomanoides,Drymoglosum piloselloides, Goniophlebiumpersici-folium, Nephrolepis bisserata, N.falcata, Oleandra pistillaris, Polypodiumverrucosum, Sellaginella willdenowiidanThelypteris sp., dengan jumlah sebanyak 3168individu. Nilai indeks keanekaragamantertinggi terdapat pada stasiun 3 dengan nilai1,74 dan yang terendah pada stasiun 1 dengannilai 1,25. Dan keanekaragaman jenistumbuhan paku pada penelitian ini berada padakategori rendah.
UCAPAN TERIMA KASIHPenulis mengucapkan terima kasih kepadapihak Panitia Seminar Nasional Biodiversitasdan Ekologi Tropika Indonesia 2014 JurusanBiologi Universitas Andalas yang telahmemberikan kesempatan kepada penulis untukmempresentasikan hasil penelitian ini.Kemudian penulis juga mengucapkan terimakasih kepada Rektor Universitas PasirPengaraian Prof. Dr. H. Feliatra, DEA yangtelah mensupport untuk mengikuti SeminarNasional ini. Kepada Kaprodi PendidikanBiologi FKIP Universitas Pasir Pengaraianyang telah memberikan arahan dan motivasiuntuk mengikuti Seminar Nasional ini. DosenPendamping yang telah memberikan masukankepada penulis sehingga selesainya penelitianini, kepada Tim Peneliti yang telah ikut bekerjasama untuk menyelesaikan Laporan Penelitiandalam mengikuti Seminar Nasional ini sertateman-teman yang tidak dapat disebutkan satuper satu, ikut serta dalam kesempurnaanpenelitian ini.
DAFTAR PUSTAKA
Arbain A, Chairul. 1990. Paku-pakuan Epifitpada Pohon Pelindung di Beberapa JalanUtama Kotamadya Padang. LembagaPenelitian dan Pengabdian MasyarakatUniversitas Andalas. Padang.
Arini DID, Kinho J. 2012. KeanekaragamanJenis Tumbuhan Paku (Pteridophyta) diCagar Alam Gunung Ambang SulawesiUtara. Info BPK Manado 2(1): 17-40.
Campbell, N.A. 2004. Biologi. Jilid 3. Jakarta:Erlangga.
Efendi WW, Haspari FNP, Nuaini Z. 2013.Studi Inventarisasi KeanekaragamanTumbuhan Paku di Kawasan WisataCoban Rondo Kabupaten Malang. CogitoErgo Sum 2(3): 173-188.
Hartini S. 2006. Tumbuhan Paku di CagarAlam Sago Malintang Sumatera Barat danAklimatisasinya di Kebun Raya Bogor.Jurnal Biodiversitas 7(3): 230-236.
Hernawati. 1995. Studi Morfologi SporongiumPaku Epifit di Gunung Kerinci, TamanNasional Kerinci Seblat (TNKS). Skripsi.

Alaniyah Syafaren, Ridwan Santoso, Egi Yudha Winata dan Rofiza Yolanda 5
Jurusan Biologi FMIPA UniversitasAndalas. Padang.
Krebs CJ. 1989. Ecological Methodology.London: Harper and Row Publisher.
Kusumaningrum BD. 2008. Analisis VegetasiEpifit di Area Wana Wisata GonoharjoKaupaten Kendal Propinsi Jawa Tengah.Skripsi. Program Studi Pendidikan Biologi.Fakultas Pendidikan MIPA IKIP PGRISemarang. Semarang.
Lamit DM. 2007. Jenis-jenis Paku Epifit yangTerdapat di Kawasan WisataPerkampungan Minangkabau PadangPanjang. Skripsi. Jurusan Biologi FMIPAUniversitas Andalas. Padang.
Mahfuz M. 1995. Jenis-jenis Paku Epifit yangTerdapat di Hutan Gunung TujuhKawasan Taman Nasional Kerinci Seblat.Tesis. Program Studi Biologi ProgramPascasarjana. Universitas Andalas. Padang.
Marisa H, Erinda A, Handayani DP, Untari DP.2013. Inisiasi Pemanfaatan BawangMerah (Allium cepa) dan Bayam(Amaranthus hybridus) sebagaiPengendalian Epifit Secara Kultur padaPelepah Kelapa Sawit. www.academia.edu/3853285/JURNAL_Pengendalian_Epifit_Secara_Kultur_pada_Pelepah_Sawit. Diakses 25 Februari 2014.
Marrugaran AE. 1987. Ecologycal Diversityand Its Measurement. New Jersey:Princeton University Press.
Mildawati, Arbain A, Gusrianto. 2011.Tumbuhan Paku Epifit di Taman HutanRaya Bung Hatta Kota Padang.Abstrak.http://pustaka.pandani.web.id/2013/10/tumbuhan-paku-epifit-di-taman-hutan.html.Diakses 12 Februari 2014.
Olsen S. 2007. Encyclopedia of Garden Ferns.China: Timber Press, Inc.
Pramono HA. 1992. Tataguna Lahan danDeforestasi di Indonesia. Jakarta: YayasanObor Indonesia.
Rahmat R. 1993. Jenis-jenis Paku Epifit yangTerdapat di HPPB Universitas Andalaspadang. Skripsi. Jurusan Biologi FMIPAUniversitas Andalas. Padang.
SastrapradjaS, Afriastini JJ, DarnaediD,Witdjaja EA. 1979. Jenis Paku Indonesia.Bogor: Lembaga Biologi Nasional – LIPI.
Sunarmi, Sarwono. 2004. InventarisasiTumbuhan Paku di Daerah Malang.Berkala Pendidikan Hayati 10: 71-74.
Widhiastuti R, Aththorick TA, Sari WDP. 2006.Struktur dan Komposisi Tumbuhan Paku-pakuan di Kawasan Hutan GunungSinabung Kab. Karo. Jurnal BiologiSumatera 8(2): 38-41.

ISSN 978-602-14989-0-3 BioETI
Daya fertilitas Mencit (Mus musculus) betina pascapemberian air seduhan kopi peroralANANDA, HERBERT SIPAHUTAR DAN MEIDA NUGRAHALIA
Jurusan Biologi, Fakultas MIPA, Universitas Negeri Medan,Jln. Willem Iskandar Psr. V Medan Estate, Medan, 20221, Sumatera Utara, Indonesia.E-mail: [email protected]
ABSTRAKPenelitian ini dilakukan untuk mengetahui daya fertilitas mencit (Mus musculus) pasca pemberian air seduhan kopi secaraoral. Delapan ekor mencit betina berumur tiga bulan dikelompokkan berdasarkan Rancangan Acak lengkap dengan n=4,perlakuan terdiri dari kontrol (0ml/hari) dan perlakuan air seduhan kopi (0,5ml/hari) dengan konsentrasi setara dengantiga cangkir kopi pada manusia. Masing-masing kelompok diperlakukan selama 21 hari kemudian mencit dikawinkan danperlakuan diteruskan hingga usia kehamilan mencpai hari ke-19. Kemudian mencit dibedah untuk diambil ovariumnyaguna menghitung jumlah korpus luteum dan menghitung jumlah implantasi pada uterus sebagai parameter daya fertilisasi.Data dianalisis dengan menggunakan uji t. Hasil penelitian menunjukkan bahwa air seduahan kopi tidak menunjukkanperubahan yang signifikan terhadap daya fertilitas mencit (Mus musculus) betina.
Key words: daya fertilitas dan air seduhan kopi.
Pendahuluan
Kopi sering kali dipandang sebagai minumanyang sangat identik dengan laki-laki, tetapi bagiwanita yang bekerja dan memiliki jam kerjamelebihi delapan jam, peran kopi sangatdibutuhkan untuk menghilangkan kantuk.Namun kopi diduga memiliki efek tersendiriterhadap masalah kesehatan khususnya masalahkesehatan reproduksi wanita. Kandungankafein yang terdapat dalam kopi diduga dapatmempengaruhi kerja hormone estradiol yangberfungsi mengatur pengeluaran estrogen danprogesterone yang keberlanjutannya dapatmenunda terjadinya konsepsi, komplikasiendometris, serta bertambahnya kerentanan seltelur (Alderete et al., 1995).
Keberhasilan fungsi reproduksi mem-butuhkan serangkaian reaksi fisiologi kompleksyang saling bergantung baik seluler maupunmolekuler. Terdapat banyak peristiwakerentanan akibat gangguan senyawaxenobiotik yang mengarah kepada kegagalanovarium (Mattison et al., 1983). MenurutNawrot et al., (2003) kafein yang terkandungdalam kopi terbawa ke dalam tubuh melaluialiran darah dan menyebar ke seluruh jaringan
dengan cepat termasuk ke dalam jaringanreproduksi. Pengkonsumsian kafein merupakansalah satu dari banyak faktor penyebabkurangnya kesuburan manusia. Penelitian yangdilakukan Bech et al., (2007), mengatakankonsumsi kafein yang tinggi menyebabkanterjadinya peningkatnya cycle adenosinemonophospate (cAMP) yang akan menggangguperkembangan sel.
Penelitian lain dengan hewan eksperimentalmenunjukkan bahwa pemberian kafein secaraoral terhadap marmot menunjukkan bahwakafein merangsang produksi dan peningkatanprostaglandin uterus yang kemungkinanbertanggung jawab atau memberikan kontribusiterhadap gangguan menstruasi pada beberapawanita (Naderali dan Poyser, 1994).
Walaupun kafein dalam kopi telah diketahuimemiliki potensi untuk menginduksi disrupsiatau disfungsi sistem reproduksi betina tetapidampak aktual dari kopi itu sendiri terhadapperkembangan reproduksi akibat asupan oralsecara kronis belum begitu dipahami denganbaik. Penelitian ini dilakukan untuk memahamilebih menyeluruh pengaruh kopi terhadap dayafertilisasi pada hewan uji mencit (Musmusculus) betina.
Ananda, Herbert Sipahutar dan Meida 7
BAHAN DAN METODE
Mencit Betina galur DD Webster berumur 2-3bulan, sekam padi, pellet 202C, bubuk kopirobusta tanpa campuran, alcohol absolute,larutan bouin, NaCl 0,9%, spiritus, dan air.
Setiap 3,057 gr bubuk kopi dilarutkandalam 28 ml air mendidih dan dilanjutkandengan mengaduk larutan kopi selama 15 menit.Kemudian larutan kopi disaring denganmenggunakan saringan teh, sehingga didapatair seduhan kopi yang siap digunakan.Pemberian air seduhan kopi dilakukansebanyak 0,5 ml untuk setiap ekor mencitkelompok perlakuan yang diberikan selamasatu kali dalam satu hari, sisa kopi disimpandilemari pendingin dan jika ingin digunakankembali diaduk dan suhunya disesuaikandengan suhu kamar sebelum dilakukanpemberian.
Selama pemberian larutan kopi, mencitditimbang setiap hari. Penimbangan beratbadan juga sebelumnya dilakukan setiap haripada pagi hari (sebelum pemberian dosis danpembersihan kandang) dengan menggunakantimbangan elektronik (Portable ElectronikScale, Ohaus) dengan ketelitian 0,1 g.
Untuk pengamatan daya fertilisasi setelahperlakuan selama 21 hari mencit dikawinkandan ditentukan hari ke-0 kehamilan melaluisumbat vagina dan pemberian kopi diteruskanhingga kehamilan mencapai hari ke-19.Kemudian mencit dibedah untuk diambilovariumnya dan direndam didalam larutanbouin. Sementara uterus dibuka untukmengetahui jumlah janin yang implantasi.Pengamatan ovarium dilakukan setelahperendaman dengan bouin telah mencapaiwaktu 24 jam dengan menghitung jumlahbenjolan pada permukaan ovarium (corpusluteum) dengan menggunakan mikroskop. Datayang didapat dari hasil perbandingan jumlahcorpus luteum dan jumlah implantasi yangdinyatakan dalam persen (%).
HASIL DAN PEMBAHASAN
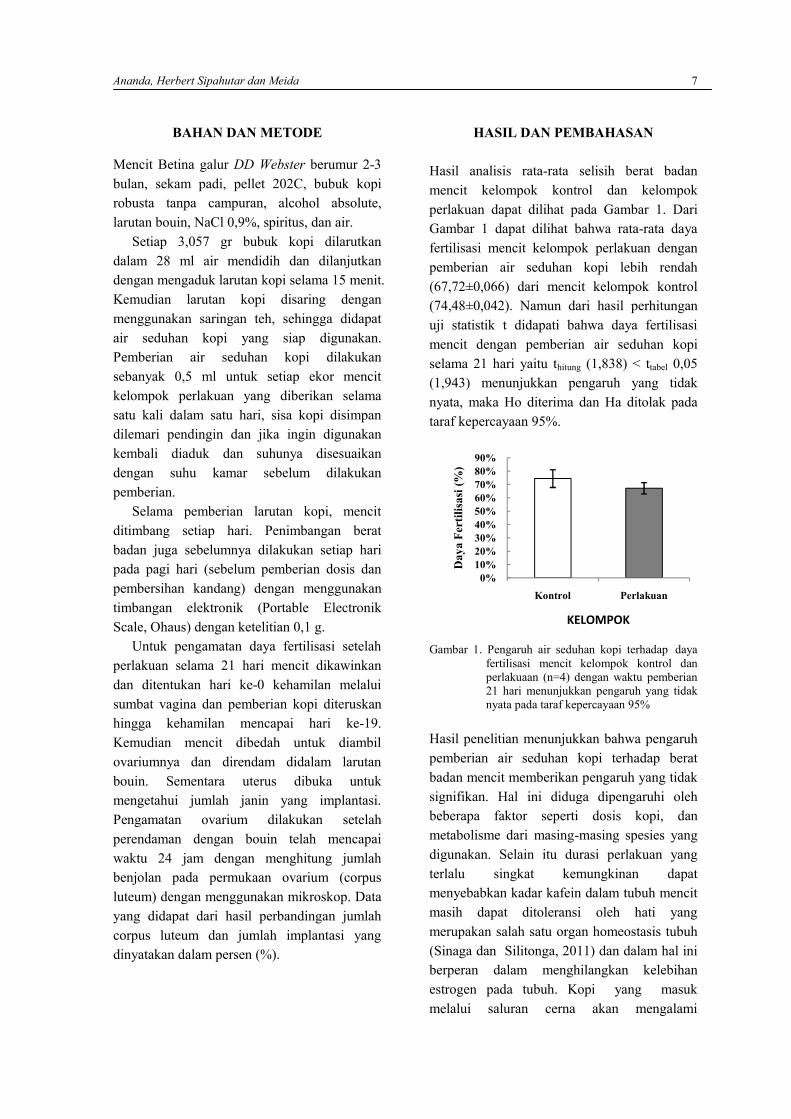
Hasil analisis rata-rata selisih berat badanmencit kelompok kontrol dan kelompokperlakuan dapat dilihat pada Gambar 1. DariGambar 1 dapat dilihat bahwa rata-rata dayafertilisasi mencit kelompok perlakuan denganpemberian air seduhan kopi lebih rendah(67,72±0,066) dari mencit kelompok kontrol(74,48±0,042). Namun dari hasil perhitunganuji statistik t didapati bahwa daya fertilisasimencit dengan pemberian air seduhan kopiselama 21 hari yaitu thitung (1,838) < ttabel 0,05(1,943) menunjukkan pengaruh yang tidaknyata, maka Ho diterima dan Ha ditolak padataraf kepercayaan 95%.
Gambar 1. Pengaruh air seduhan kopi terhadap dayafertilisasi mencit kelompok kontrol danperlakuaan (n=4) dengan waktu pemberian21 hari menunjukkan pengaruh yang tidaknyata pada taraf kepercayaan 95%
Hasil penelitian menunjukkan bahwa pengaruhpemberian air seduhan kopi terhadap beratbadan mencit memberikan pengaruh yang tidaksignifikan. Hal ini diduga dipengaruhi olehbeberapa faktor seperti dosis kopi, danmetabolisme dari masing-masing spesies yangdigunakan. Selain itu durasi perlakuan yangterlalu singkat kemungkinan dapatmenyebabkan kadar kafein dalam tubuh mencitmasih dapat ditoleransi oleh hati yangmerupakan salah satu organ homeostasis tubuh(Sinaga dan Silitonga, 2011) dan dalam hal iniberperan dalam menghilangkan kelebihanestrogen pada tubuh. Kopi yang masukmelalui saluran cerna akan mengalami
0%10%20%30%40%50%60%70%80%90%
Kontrol Perlakuan
Day
a Fe
rtili
sasi
(%)
KELOMPOK

Ananda, Herbert Sipahutar dan Meida 8
metabolisme awal di hati dikarenakan hatiadalah tempat metabolisme utama yang akanmendetoksifikasi dan mengeliminasi semuatoksin baik endogen ataupun eksogen (Bhara,2009).
Namun dilihat dari rata-rata daya fertilisasimencit betina yang disajikan pada Gambar 1.dapat dilihat bahwa rata-rata selisih berat badanmencit betina kelompok kontrol lebih tinggidibandingkan dengan mencit betina kelompokperlakuan. Perbedaan rata-rata yang terlihatdiduga dapat diakibatkan oleh beberapa hal,misalnya pencekokan dengan menggunakansonde untuk memasukkan air seduhan kopi kesaluran pencernaan dan menimbulkan strespada mencit yang diberi perlakuan, meskipuntingkat stres tidak diketahui. Efek lanjutan daristres ini kemungkinan juga diperantarai olehradikal bebas yang menurut Gunawan et al(2007) dapat mempengaruhi kondisipatofisiologis seperti kerusakan ataupunkelainan biokimia dan menimbulkanpenyimpangan metabolisme. Gangguanmetabolisme ini akan menurunkan sintesisbahan-bahan yang diperlukan untuk proliferasidan pematangan sel-sel tubuh (Gunawan et al,2007), sehingga secara tidak langsunggangguan metabolisme ini juga dapatmempengaruhi daya fertilisasi mencit.
Kemudian penurunan daya fertilisasi mencitjuga diduga disebabkan oleh pemberian airseduhan kopi. Kopi secara keseluruhan zatyang dikandungnya merupakan xenobiotikyang dapat menyebabkan kerusakan sel secaralangsung dengan mengganggu permeabilitasselaput, homeostasis osmosa, keutuhan enzimdan kofaktor yang selanjutnya membebani seltersebut, kemudian menyebabkan jejas danmengakibatkan perubahan morfologi sel(Robins, 1995). Kuper et al., (2000) jugamenyatakan bahwa kafein dapat mempengaruhistruktur DNA dan fungsinya denganmenghambat reaksi poli(ADP-rybosyl)ation,yang berperan penting dalam perbaikankerusakan DNA postreplication, hal itu akanmeningkatkan potensi terjadinya kerusakan
DNA melalui penghambatan DNA perbaikanselama fase S dan fase G2 pada siklus sel.Dengan demikian, kafein yang terkandungdalam air seduhan kopi tersebut diduga telahmengganggu pembelahan mitosis pada ovumyang baru saja dibuahi sperma (zygot).Sehingga pada akhirnya kondisi inimenyebabkan zigot gagal untuk berkembangmenjadi embrio (Supriati et al., 2010).
Selain itu, pada penelitian yang dilakukanoleh Dixon et al (2011), ditemukan bahwakafein menghambat kontraksi otot tuba fallopiyang merupakan efek peningkatan kadar cAMPsitosol ketika kafein menghambat pada PDEs.Saat hiperpolarisasi yang disebabkan olehpembukaan kanal Katp, membran potensialmengalami pergeseran dan juga membukakanal Ca2+ sehingga terjadi relaksasi.Hiperpolarisasi yang menyebabkan aktifasikanal Katp tersebut menyebabkanpenghambatan kontraksi otot tuba fallopi secaraspontan yang merupakan mekanisme pentinguntuk menghantar telur sepanjang tuba fallopi,sehingga dengan kata lain sel telur gagalmencapai uterus untuk proses implantasi.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Air seduhan kopi tidak memiliki pengaruh yangsignifikan terhadap daya fertilitas mencit (Musmusculus) betina.
DAFTAR PUSTAKA
Alderete, E., Eskenazi, B., and Sholtz,R. (1995).Effect of cigarette smoking and coffeedrinking on time to conception.Epidemiology, 6, 403–408.
Bech, H.B., Carsten O., Henriksen, B.T, Olsen,J, (2007), Efect of reducing caffeineintake on birth weihgt and lenght ofgestation: randomised controled trial,Departenment of Epidemiologi UniversityAarhus, Denmark.
Bhara, M. (2009). Pengaruh pemberian kopidosis bertingkat per oral 30 hari terhadap

Ananda, Herbert Sipahutar dan Meida 9
gambaran histologi hepar tikus wistar.Skripsi, Fakultas Kedokteran UniversitasDiponegoro, Semarang.
Dixon, R.E., Hwang, S.J., Britton, F.C.,Sanders, K.M., dan Ward, S.M. (2011).Inhibitory effect of caffeine on pacemakeractivity in the oviduct is mediated bycAMP-regulated conductances. BritishJournal of Pharmacology, 763, 745-754.
Gunawan, Setiatin E.T., Rosadi, B., Hine T.M.,dan Parakkasi, A. (2007). PerformansiReproduksi Tikus Betina denganPemberian Lendir Lidah Buaya. JurnalKedokteran Hewan, 1(1), 1-6.
Kuper, H., Titus-Ernstoff, L., Harlow B.L., danCramer, D.W. (2000). Population basedstudy of coffee, alcohol and tobacco useand risk of ovarian cancer. InternationalJournal of Cancer. 88, 313–318.
Mattison, D.R., Nightingale, M.S., andShiromizi, K. (1983). Effects of toxic
substances on female reproduction.Environmental Health Perspectives, 48,43-52.
Naderali, E.K. dan Poyser, N.L. (1994). TheEffect of caffeine on prostaglandin putputfrom the guinea-pig uterus. Journal ofPharmacology, 113, 103-110.
Nawrot, P., Jordan, S., Eastwood, J., Rotstein,J., Hugenholtz, A., dan Feeley, M. (2003).Effects of caffeine on human health. FoodAdditives and Contaminants, 20(1), 1–30.
Sinaga, E. dan Silitonga, M. (2011). AnatomiFisiologi Tubuh Manusia. FMIPAUNIMED.
Supriati, R., Karyadi B., dan Maherawati.Pengaruh pemberian getah buah pepaya(carica papaya l.) Terhadap daya fertilitasmencit (Mus musculus) balb/c betina.Konservasi Hayati: 6(2): 1-8.

ISSN 978-602-14989-0-3 BioETI
Uji daya hambat formulasi minyak Piper aduncum sebagaipestisida nabati pengendali Jamur Fusarium pada batangHylocereus polyrhizus secara InvitroARYUDA YOZA SELFA1 NASRIL NASIR1, FUJI ASTUTI FEBRIA1, JUMJUNIDANG2 DANNURMANSYAH3
1Laboratorium Mikrobiologi, Jurusan Biologi, FMIPA, Universitas Andalas, Padang, Sumatera Barat 251632Balai Penelitian Tanaman Buah Tropika, Jl. Raya Solok-Aripan Km. 7 Solok3 Balai Penelitian Tanaman Rempah dan Obat. KP. Laing. Solok. Sumatera Barat.E-mail: [email protected]
ABSTRACTPenelitian tentang uji daya hambat formulasi minyak Piper aduncum sebagai pestisida nabati pengendali jamur Fusariumsp. pada batang Hylocereus polyrhizus secara invitro telah dilakukan di Laboratorium Proteksi, Balai Penelitian TanamanBuah (Balitbu) Tropika, di Aripan, Solok dari bulan Juni sampai Agustus 2014. Penelitian dilakukan dengan metodeeksperimen menggunakan Rancangan Acak Lengkap (RAL) 4 perlakuan dan 6 ulangan untuk masing-masing perlakuan.Perlakuan terdiri dari kontrol (tanpa pemberian formulasi minyak Piper aduncum), formulasi minyak Piper aduncumdengan konsentrasi 500 ppm, 1000 ppm dan 2000 ppm. Hasil konsentrasi formulasi minyak yang efektif pada penelitianini dalam menghambat pertumbuhan koloni jamur Fusarium sp. adalah menggunakan formulasi minyak Piper aduncum2000 ppm dengan daya hambat > 50 %.
Key words: Fusarium sp., Hylocereus polyrhizus, pestisida nabati, Piper aduncum
Pendahuluan
Penggunaan pestisida kimia dalam mengendali-kan penyakit tanaman masih marak digunakandi Indonesia. Setiap tahunnya penggunaanpestisida kimia terus meningkat hingga meng-habiskan uang ± 6 triliun per tahun (Beritasatu,2012). Menurut Djunaedy (2009), rata-ratapeningkatan total konsumsi pestisida pertahun6,33% dan jika penggunaan pestisida kimiaterus meningkat dari tahun ke tahun, makabiaya produksi juga akan meningkat. Menurutperhitungan petani, biaya komponen pestisidamencapai 25-40% dari total biaya produksipertanian. Penggunaan pestisida kimia secaraterus menerus dapat menimbulkan dampaknegatif terhadap lingkungan dan makhluk hiduplainnya (Nurmansyah, 2010).
Salah satu alternatif yang cukup potensialdigunakan untuk menghindari dampak negatifpenggunaan pestisida kimia adalah denganpenggunaan pestisida nabati. Pestisida nabatimerupakan pestisida yang bahan aktifnyaberasal dari tumbuhan, ramah lingkungankarena tidak mencemari lingkungan, targetnya
lebih spesifik dan residu mudah terurai(Soehardjan, 1994). Salah satu pestisida nabatiyang mampu dan telah terbukti dapatmengendalikan penyakit tanaman akibatserangan jamur patogen adalah Piper aduncum(Nurmansyah, 1997a; Nurmansyah, 1997b).Formulasi minyak Piper aduncum memilikidaya fungisidal yang sangat baik dibandingkandengan sirih liar lain, terutama terhadap jamurSclleroticum rolfsii dan Fusarium oxysporumyang menyerang tanaman kacang tanah dancabai (Nurmansyah, 2012).
Formulasi minyak Piper aduncum terdiridari komponen utama yaitu phenylpropanoiddilapiole (32,9-61,8%), monoterpenoidspiperitone (2,2-13,5%), 1,8-cineole (0,1-8,6%),4-terpineol (1,6-5,4%), sesquiterpene dan b-caryophyllene (5,0-5,3%) (Clcclo dan Balletero,1997). Formulasi minyak P. aduncum padakonsentrasi 2000 ppm mampu menekanpertumbuhan jamur patogen Phytophthoracapsisi, Fusarium oxysporum dan Sclleroticumrolfsii yang di isolasi dari tanaman panili, tomatdan cabai. Kemampuan formulasi minyak

Aryuda Yoza Selfa, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 11
Piper aduncum dalam mengendalikanpertumbuhan jamur patogen pada tanamanyaitu: Phytophthora capsisi 85,16%, Fusariumoxysporum 72,39% dan Sclleroticum rolfsii80,75%. (Nurmansyah, 2004). Berdasarkanpotensi yang dimiliki tanaman Piper aduncum,formulasi minyak tersebut juga mampumenekan pertumbuhan jamur patogenFusarium sp. pada tanaman buah naga.
Tanaman buah naga saat ini menjadiprimadona bagi para petani untukdibudidayakan secara besar-besaran diIndonesia (Kristanto, 2009). Meskipunmemerlukan dana yang cukup besar untukmembudidayakan tanaman buah naga ini, tetapiminat untuk membuka lahan baru cukup besar.Hal ini dikarenakan nilai jual buah naga diIndonesia cukup tinggi dan potensi agribisnis-nya bagi para petani sangat menjanjikan. Saatini harga per kg buah naga ini adalah Rp.40.000 dan total produksi per hektar (ha) ± 11,2ton untuk tahun pertama produksi (Nasir, 2013).
Sejak lima sampai enam tahun terakhir inibanyak keluhan turunnya produksi buah nagaakibat serangan hama dan penyakit pada batangbuah naga merah. Lebih dari 50% perkebunanbuah naga di Riau dan Sumbar dilaporkandiserang gejala penyakit busuk batang, busukkuning dan busuk hitam (Nasir, 2013). MenurutBarthana (2013), gejala penyakit busuk kuningpada buah naga disebabkan salah satunya olehjamur Fusarium sp. Hingga saat ini belum adaditemukan pestisida nabati yang efektifmengendalikan penyakit pada datang buah nagayang disebabkan oleh patogen jamur Fusariumsp. Padahal buah naga merah saat ini mulaibanyak dibudidayakan di Indonesia setelahdiintroduksi pertama kali awal tahun 2000-an.
BAHAN DAN METODE
Penelitian ini dilaksanakan dari bulan Junisampai Agustus 2014 di Laboratorium Proteksi,Balai Penelitian Tanaman Buah (Balitbu)Tropika, di Aripan, Solok.
Penelitian ini dilakukan dengan metodeeksperimen menggunakan Rancangan Acak
Lengkap (RAL) 4 perlakuan dan 6 ulanganuntuk masing-masing perlakuan. Perlakuanterdiri dari kontrol (tanpa permberian formulasiminyak Piper aduncum), formulasi minyakPiper aduncum dengan konsentrasi 500 ppm,1000 ppm dan 2000 ppm. Koleksi isolat jamurFusarium sp. diperoleh dari Balai PenelitianTanaman Buah (Balitbu) Tropika, di Aripan,Solok dan formulasi minyak Piper aduncumdiperoleh dari Balai Tanaman Rempah danObat (Balitro) di Laing, Solok.
Kerja uji daya hambat formulasi minyakPiper aduncum terhadap pertumbuhan jamurFusarium sp yaitu medium PDA pada suhu ±50oC ditambahkan dengan masing-masingformulasi perlakuan dan dihomogenkan.Selanjutnya dituang ke cawan petri, tungguhingga dingin dan mengeras. Kemudian jamurFusarium sp diinokulasikan dengan carameletakkan fungal mat (yang telah dipotongdengan corkborer steril dengan diameter 5 mm)di tengah medium. Kemudian diinkubasidengan suhu ruang.
Parameter yang diamati berupapertumbuhan koloni jamur pada media PDApada masing-masing perlakuan yangdibandingkan dengan perlakuan kontrol.Pengamatan dilakukan sampai diameter jamurFusarium sp pada perlakuan kontrol memenuhicawan petri. Daya hambat pengaruh pemberianformulasi minyak Piper aduncum terhadappertumbuhan jamur Fusarium sp dihitungdengan rumus sebagai berikut:
HASIL DAN PEMBAHASAN
Berdasarkan hasil pengamatan yang telahdilakukan untuk melihat kemampuan dayahambat formulasi minyak Piper aduncumterhadap pertumbuhan koloni jamur Fusariumsp. yang diisolasi dari batang Hylocereuspolyrhizus, dapat dilihat pada Tabel 1 berikut.

Aryuda Yoza Selfa, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 12
Tabel 1. Daya hambat formulasi minyak Piperaduncum terhadap pertumbuhan JamurFusarium sp. pada hari ke-8 setelahinokulasi.
No Perlakuan DiameterKoloni (cm)
Persentase dayahambat (%)
1 Kontrol 9a 02 Piper aduncum
500 ppm 7,31b 18,80
3 Piper aduncum1000 ppm 5,46c 36,26
4 Piper aduncum2000 ppm 3,37d 62,59
Keterangan : Angka-angka yang dikuti oleh huruf kecil yangtidak sama menunjukkan perbedaan yang nyatamenurut hasil uji DNMRT 5 %
Dari Tabel 1 dapat dilihat bahwa pemberianformulasi minyak Piper aduncum denganbeberapa konsentrasi memberikan pengaruhterhadap pertumbuhan diameter koloni jamurFusarium sp. yang telah diisolasi dari batangHylocereus polyrhizus. Sedangkan denganperlakuan kontrol (tanapa pemberian formulasiminyak Piper aduncum) pertumbuhan jamurFusarium sp. sudah mencapai diameter 9 cmpada cawan petri pada hari ke-8 setalahinokulasi dilakukan. Pada pemberian formulasiminyak Piper aduncum 500 ppm menunjukkanpertumbuhan diameter koloni jamur 7,31 cmdan pemberian formulasi minyak Piperaduncum 1000 ppm menunjukkan pertumbuhandiameter koloni jamur 5,46 cm. Darikeseluruhan perlakuan yang diberikan terlihatabahwa konsentrasi yang efektif menekanpertumbuhan diameter koloni jamur Fusariumsp. pada batang Hylocereus polyrhizus secarainvitro yaitu pada konsentrasi 2000 ppmdengan diameter pertumbuhan koloni jamurterkcil 3,37 cm.
Persentase daya hambat terhadap formulasiminyak Piper aduncum terhadap pertumbuhankoloni jamur Fusarium sp. Pada tabel 1 terlihatbahwa pada perlakuan pemberian formulasiminyak Piper aduncum mampu menghambatpertumbuhan jamur Fusarium sp. dari 18,80-62,59% pada konsentrasi 500 sampai 2000 ppm.Hal ini sangan berbeda nyata dengan perlakuankontrol (tanpa pemberian formulasi minyakPiper aduncum) yaang memiliki persentasedaya hambat 0%. Pada Tabel 1 dapat dilihat
bahwa pemberian formulasi minyak Piperaduncum konsentrasi 500 ppm masih memilikipersentase daya hambat yang kecil dalammenekan pertumbuhan jamur Fusarium sp.yaitu 18,80%, sehingga konsentrasi 500 ppmbelum efektif digunakan sebagai pestisidanabati. Dari ketiga konsentrasi formulasiminyak Piper aduncum yang digunakan,persentase daya hambat tertinggi dalammenekan pertumbuhan jamur Fusarium sp.yaitu pada konsentrasi 2000 ppm dengan62,59%.
Gambar 1. Pertumbuhan diameter koloni jamurFusarium sp. dengan perlakuan (a)kontrol, (b) formulasi minyak P.aduncum 500 ppm, (c) formulasi minyakP. aduncum 1000 ppm, (d) formulasiminyak P. aduncum 2000 ppm
Komponen utama P. aduncum merupakansenyawa aktif dilapiol (golongan lignan).Senyawa dilapiol dapat menghambat prosesoksidasi yang terjadi didalam sel. Prosesoksidasi umumnya terjadi pada senyawa yangbersifat racun di dalam sel, sehingga dapatmengakibatkan terjadinya penurunan senyawaracun tersebut. Terhambatnya aktivitas enzimdapat mengakibatkan terjadinya penumpukansenyawa beracun di dalam sel yang selanjutnyadapat mengakibatkan kematian sel (Bernard etal., 1995).
Komponen kimia dari minyak atsiri yangbersifat antifungal dan anti bakteri memilikiperanan sebagai pestisida nabati yang dapatmenembus dinding sel jamur dan bakteri,sehingga mengganggu proses metabolisme
c
ba d
d

Aryuda Yoza Selfa, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 13
didalam sel. Mekanisme kerja dari golonganterpenoid yaitu dapat mereduksi pertumbuhanmiselium dan berakibat terjadinya pemendekanpada ujung hifa dan menghambat prosesmetabolisme dengan mengakumulasikanglobula lemak didalam sitoplasma sel,mengurangi jumlah mitokondria dan merusakmembran nukleus (Knoblock et al., 1989).
Dialam jamur Fusarium sp dapat mem-bentuk konidium dengan konidisfor bercabang-cabang dan makrokonidium berbentuk sabit,bertangkai kecil dan berpasangan. Miseliumnyaterdapat didalam sel khususnya didalampembuluh kayu (Ellis, 2007). Genus Fusariummerupakan patogenik menyebabkan penyakitlayu pada tanaman. Fusarium sp. yang terdapatdidalam tanah dapat bertahan dengan mem-bentuk klamidospora atau hifa pada sisatanaman dan bahan organik lainnya (Saragihdan Silalhi, 2006). Jamur Fusarium sp. untukmenginfeksi dan hidup didalam jaringanmampu berkembang pada suhu 28-300C(Windels, 1993).
KESIMPULAN
Berdasarkan hasil penelitian yang telahdilakukan, diperoleh kesimpulan sebagaiberikut: pemberian formulasi minyak Piperaduncum mampu menekan pertumbuhan jamurFusarium sp. pada batang Hylocereuspolyrhizus secara in vitro dengan persentasedaya hambat > 50% pada konsentrasi 2000 ppmdengan diameter pertumbuhan koloni jamur3,37 cm.
DAFTAR PUSTAKA
Barthana, D. 2013. Deskripsi Gejala danIdentifikasi Penyebab Penyakit BusukKuning Batang Tanaman Buah NagaMerah di Kecamata Batang Anai, PadangPariaman, Sumatera Barat. Skripsi SarjanaBiologi FMIPA Universitas Andalas.Padang.
Beritasatu. 2012. Petani KhawatirkanTingginya Penggunaan Pestisida.
http://www.beritasatu.com/bisnis/43463.19 April 2013 (14:30).
Bernard CB, Krishnamurty HG, Chauret D,Durst T, Philogene BJR, Vindas PS,Hasbun C, Poveda L, Roman LS, ArnasonJT. 1995. Insecticidal defenses ofPiperaceae from the Neotropics. Journal ofChemical Ecology 21 : 801-814.
Clcclo J.F. and C.M Ballestero. 1997.Constltuyentes Volatiles de Las Hojas yEspkjas de Piper adncum (Piperaceae) deCosta Rica. Rev. Biol. Trop. 45: 783-790.
Djunaedy, A. 2009. Biopestisida sebagaiPengendali Organisme PenggangguTanaman (OPT) yang Ramah Lingkungan.Embryo. vol. 6 No. 1.
Ellis, D. 2007. Fusarium. The University ofAdelaide.http://www.mycology.adelaide.edu.au/Fungal_Descriptions/Hyphomycetes_(hyaline)/Fusarium/. 24 Februari 2014.
Kristanto. 2009. Buah Naga: Pembudidayaan diPot dan di Kebun. Penebar Swadaya.Jakarta.
Knoblock, K., A, Pauli., B, Iberl, H, Weigandand N, Weis. 1989. Antibacterial andantifungal properties of essential oilcomponds. J. Ess, oil. Res 1p;119-128.
Nasir. N. 2013. Serangan Penayakit Pada BuahNaga Hylocereus polyrhizus DiKepulauan Riau. Laporan Penelitian.Jurusan Biologi FMIPA UniversitasAndalas. 5 hal.
Nurmansyah. 1997a. Kajian awal potensigulma sirih-sirih (Piper aduncum L)sebagai fungisida nabati. Journal biologika.Perhimpunan Biologi KomisariatSumatera Barat. 1 (2):48-56
Nurmansyah. 1997b. Pengaruh Tepung danMinyak Daun Gulma sirih-sirih (Piperaduncum L.) Terhadap PatogenSclerotium rolfsii dan Fusarium spp.Prosiding Kongres XIV dan SeminarNasional Fitopatologi Indonesia. 27-29Oktober 1997. Palembang. pp. 254-257.
Nurmansyah. 2004. Pengaruh PenambahanMinyak Serai Wangi dan LimbahKayumanis terhadap Daya AntifungalPestisida Nabati Sirih-Sirih. ProsidingSeminar Ekspose Teknologi Gambir,Kayumanis dan Atsiri. Pusat Penelitiandan Pengembangan. Bogor.
Nurmansyah. 2010. Efektivitas Minyak SeraiWangi dan Fraksi Sitronella TerhadapPertumbuhan Jamur Phytophthora

Aryuda Yoza Selfa, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 14
palmivora Penyebab Penyakit Busuk BuahKakao. Bul. Littro. 21 (1): 43-52.
Nurmansyah. 2012. Minyak Atsiri Piperaduncum Sebagai Bahan Baku PestisidaNabati Untuk Pengendalian JamurPenyakit Tanaman. Bunga Rampai InovasiTanaman Atsiri Indonesia.
Saragih, Y. S. dan F. H. Silalahi. 2006. Isolasidan Identifikasi Spesies FusariumPenyebab Penyakit Layu Pada TanamanMarkisa Asam. J. Hort. 16(4): 336-334.
Soehardjan, M. 1994. Konsepsi dan StrategiPenelitian dan Pengembangan PestisidaNabati. Prosiding Seminar HasilPenelitian dalam Rangka PemanfaatanPestisida Nabati. 1-2 Desember 1993.Balitro. Bogor. pp. 11-18.
Windels, C., E., 1993. Fusarium. In Singeletton,L. L., Mihail, J. D., and Rush, J. D., (Ed),Methods for Research on SoilbornePhytopathogenic Fungi. APS Press. TheAmerican Phytopathological Society, St.Paul Minnessota 115-126.

ISSN 978-602-14989-0-3 BioETI
Optimalisasi produksi biogas Eceng Gondok denganHydrothermal pretreatment (production optimalisation of waterhyacinth biogas with hydrothermal pretreatment)Dewi Murni dan Yuhelsa Putra
Universitas Sultan Ageng Tirtayasa, Serang, Taman Pesona, Blok E2 No.8, Serang, BantenE-mail: [email protected]/ [email protected].
ABSTRAKEceng gondok merupakan salah satu jenis keanekaragaman hayati yang selama ini dianggap merugikan karenadapat mencemari lingkungan. Namun, seiring dengan berkembangnya teknologi, ternyata eceng gondok bisadimanfaatkan sebagai bahan baku pembuatan biogas yang merupakan sumber energi alternatif berkelanjutan dan ramahlingkungan. Kandungan lignoselulosa yang tinggi menyebabkan produksi biogas kurang optimal. Untuk mengatasinyaperlu diberikan praperlakuan hidrotermal. Tujuan penelitian ini adalah untuk mengetahui lama (waktu) praperlakuanhidrotermal (hydrothermal pretreatment) yang tepat untuk memperoleh produksi biogas dan persentase metana yangpaling optimal. Penelitian ini dilakukan pada skala laboratorium dengan metode eksperimen di Laboratorium FakultasPertanian, Universitas Sultan Ageng Tirtayasa. Eceng gondok dicacah dengan alat pencacah (slicer) sampai ukuransekitar 6 mm, kemudian direbus pada suhu 1700C dengan variasi waktu 30, 60 dan 90 menit (hydrothermal pretreatment).Setelah diberi praperlakuan hidrotermal, eceng gondok dicampur dengan EM4. Campuran ini selanjutnya diinkubasipada biodigester anaerobic batch untuk difermentasi menjadi biogas. Hasil analisis data menunjukkan bahwa produksibiogas paling optimal dicapai pada praperlakuan hidrotermal 60 menit, yaitu 4,709 mL/g dengan laju produksi 13,47mL/jam dan mencapai titik tertinggi pada hari ke-39. Persentase metana tertinggi juga ditemukan pada praperlakuanhidrotermal 60 menit, yaitu 43,38%.
Key words: biogas, eceng gondok, praperlakuan hidrotermal, produksi, metana
Pendahuluan
Eceng gondok (Eichornia crassipes)merupakan spesies tropikal yang tergolong kedalam famili Ponterderiacea. Tanaman iniberasal dari Sungai Amazon yang terdapat diAmerika Selatan. Spesies ini selanjutnyatersebar ke lebih 50 negara dalam waktu limadasawarsa. Tanaman eceng gondok memilikitoleransi yang ekstrim terhadap variasi musim,ketersediaan unsur hara, pH, suhu dan substansiracun. Toleransi yang ekstrim ini menyebabkaneceng gondok menjadi salah satu polutanbiologis yang dapat mencemari ekosistemperairan (Gunnarsson and Petersen, 2007).
Menurut Villamagna and Murphy (2010),eceng gondok dapat tersebar luas dan mampumendominasi badan perairan dalam waktusingkat karena memiliki laju pertumbuhan yangsangat tinggi. Hal ini didukung olehkemampuan serapan hara dan produktivitasbiomassanya yang tinggi. Hasil penelitian
Gunnarsson & Petersen, (2007) menunjukkanbahwa rata-rata pertumbuhan eceng gondokmencapai 100-140 ton material kering perhektar per tahun.
Dominansi eceng gondok di suatu badanperairan bisa menyebabkan masalahlingkungan, seperti penurunan debit air sebagaisumber daya irigasi, memancing dan navigasi(Villamagna and Murphy, 2010). Ecenggondok juga dapat mengurangi aktivitasfotosintesis tumbuhan yang hidup di badanperairan karena menghalangi penetrasi cahayamatahari. Akibatnya terjadi penurunankonsentrasi oksigen yang merupakan variabelkualitas air paling penting (Perna and Burrows,2005).
Salah satu ekosistem perairan yang dicemarioleh eceng gondok adalah Sungai Ciujung yangmelintasi empat kecamatan di KabupatenSerang, Banten, yaitu Kecamatan Pontang,Carenang, Tirtayasa dan Tanara. Air SungaiCiujung dahulu menjadi sumber kehidupan

Dewi Murni dan Yuhelsa Putra 16
masyarakat. Sungai ini berperan sebagaisumber air irigasi persawahan. Selain itu, jugadimanfaatkan oleh warga yang tinggal disekitar sungai untuk kebutuhan sehari-hariseperti mencuci dan mandi. Saat ini, air SungaiCiujung sudah tidak dapat digunakan lagikarena telah tercemar oleh limbah pabrik.
Kandungan organik yang tinggi darilimbah pabrik menyebabkan eceng gondoktumbuh dengan sangat cepat dan menutupisebagian besar permukaan Sungai Ciujung.Keberadaan eceng gondok di sungai inimenyebabkan pendangkalan dan berkurangnyakecepatan arus sungai. Akibatnya, suplai airuntuk irigasi persawahan jadi berkurang.Permasalahan pencemaran ini harus segeradiatasi agar tidak berdampak buruk bagiekosistem perairan dan masyarakat sekitarnya.
Berbagai upaya bisa dilakukan untukmengatasi pencemaran badan perairan oleheceng gondok. Diantaranya melaluipemanfaatan eceng gondok sebagai bahanbaku pembuatan biogas. Biogas yangdihasilkan dari fermentasi eceng gondoksekaligus bisa menjadi solusi alternatif untukpermasalahan keterbatasan energi dari fosilyang keberadaannya semakin berkurang dantidak dapat diperbaharui. Biogas merupakansumber energi alternatif yang ramahlingkungan, terbarukan dan dapat dibakarseperti gas elpiji (LPG). Biogas juga dapatdigunakan sebagai sumber energi penggerakgenerator listrik. Gas ini dapat diperolehdengan biaya murah, karena diolah dari bahanyang selama ini dibuang atau dikategorikansebagai limbah.
Hasil penelitian Singhal and Rai (2003)menunjukkan bahwa rata-rata produksi biogasdari eceng gondok adalah 15,4-23,65 L/ kgberat kering eceng gondok yang diperolehsetelah 21 hari fermentasi. Patil et al., (2011)menyatakan bahwa penggunaan eceng gondoksebagai substrat pembuatan biogas merupakanstrategi pengendalian yang sangat menguntungkan karena bersifat produktif. Eceng gondok
selalu tersedia dan terbarukan sehingga bisadipanen setiap saat sesuai masa produksinya.
Permasalahan yang muncul dalampembuatan biogas berbahan baku eceng gondokadalah tingginya kandungan lignoselulosa yangmenyebabkan proses produksi biogas jaditerhambat. Taherzadeh dan Karimi (2008)menyatakan bahwa senyawa lignoselulosamerupakan senyawa penyusun sebagian besarbiomassa organik penghasil energi, sepertilimbah perkotaan, kotoran ternak, dan tanamanair. Hasil penelitian Sornvoraweat &Kongkiattikajorn (2010) menunjukkan bahwakandungan rata-rata pada daun eceng gondokadalah hemiselulosa (32,69 %), lignin (4,37%),selulosa (19,02%), protein (10,20%) danamilum (4,16%). Patil et al., (2011)menyatakan bahwa selulosa berikatan kuatdengan lignin. Menurut Mosier et al., (2005),selulosa merupakan bahan yang akan diubahmenjadi gas metana (CH4) melalui prosesmetanogenesis, namun keberadaan ligninmenghambat kerja enzim dalam menghidrolisisselulosa sehingga laju produktivitas biogasyang dihasilkan dari eceng gondok menjadirendah. Upaya untuk mengatasi permasalahantingginya kandungan lignoselulosa ecenggondok dapat dilakukan dengan menggunakanmetode praperlakuan (pretreatment) yang dapatmeningkatkan degradasi lignoselulosa secaraenzimatik. Taherzadeh & Karimi (2008)menyatakan bahwa hidrolisis enzimatiksenyawa lignoselulosa tanpa praperlakuanbiasanya tidak efektif karena senyawa tersebutmemiliki stabilitas yang tinggi terhadap kerjaenzim yang dihasilkan oleh bakteri. Mosier etal., (2005) menyatakan bahwa melaluipraperlakuan, biomassa selulosa akan lebihmudah diakses oleh enzim yang akanmengubah polimer karbohidrat menjadi gulasebagai bahan baku fermentasi.
Praperlakuan yang dapat meningkatkanefektifitas dan produksi biogas dari materiallignoselulosa diantaranya adalah hidrotermal.Zeng et al., (2007) menyatakan bahwaperebusan material lignoselulosa dalam air

Dewi Murni dan Yuhelsa Putra 17
panas merupakan salah satu metodehidrotermal yang telah digunakan beberapadekade, contohnya pada industri kertas. Airdengan tekanan yang tinggi dapat melakukanpenetrasi ke dalam biomassa, menghidrasiselulosa serta menghilangkan hemiselulosa danlignin. Keuntungan utama metode ini adalahtidak perlu penambahan senyawa kimia dantidak membutuhkan material tahan korosi untukreaktor hidrolisis pada proses ini. Menurut Qiao(2011), setelah praperlakuan 1700 C/jam,produksi biogas dari biomassa lignoselulosameningkat dari 210 mL/g menjadi 238 mL/gbahan baku.
Selain kandungan lignoselulosa yang tinggiShankar et al., (2013) menyatakan bahwaeceng gondok tidak memiliki mikroba esensialuntuk mengawali proses produksi biogas. Olehkarena itu perlu penambahan EM4 sebagaipenyedia mikroba untuk mengawali prosesfermentasi eceng gondok menjadi biogas.Menurut Herawati dan Wibawa (2010), hasilterbaik pada penelitian pembuatan biogasialah dengan penambahan EM4, dimana yieldbiogas rata-rata dihasilkan 0,030 L/g VS. EM4mengandung bakteri fermentasi dari genusLactobaccilus, jamur fermentasi, Actinomycetes,bakteri pelarut fosfat dan ragi (Herawati andWibawa, 2010).
Untuk memperoleh biogas dengan produksidan persentase metana yang tinggi, perludiketahui lama (waktu) pemberianpraperlakuan hidrotermal terhadap ecenggondok yang tepat. Berdasarkan permasalahantersebut, perlu dilakukan penelitian untukmengetahui lama (waktu) perlakuanhidrotermal yang tepat untuk memperolehproduksi biogas dan persentase metana yangpaling optimal.
BAHAN DAN METODE
Penelitian ini dilakukan di LaboratoriumFakultas Pertanian, Universitas Sultan AgengTirtayasa. Eceng gondok diambil dari SungaiCiujung, Kabupaten Serang. Penelitiandilakukan dalam skala laboratorium dengan
metode eksperimen. Sebagai kontrol(pembanding), dievaluasi juga produksi biogasdari eceng gondok tanpa praperlakuanhidrotermal (hydrothermal pretreatment).
Eceng gondok dicacah dengan alat pencacah(slicer) dengan ukuran sekitar 6 mm. Ecenggondok hasil pencacahan dicampur dengan air(1:1). Campuran ini kemudian dimasukkan kedalam autoklaf dan direbus (hydrothermalpretreatment) pada suhu 1700C dengan variasiwaktu 30 menit, 60 menit dan 90 menit.Selanjutnya, eceng gondok dikeluarkan dariautoklaf dan dibiarkan terbuka sampaimencapai suhu ruangan. Setelah itu,ditambahkan EM4 sebagai starter untukmenginisiasi proses fermentasi. Campuran inikemudian dimasukkan ke pembangkit/ reaktorbiodigester anaerobic batch. Selanjutnyadiinkubasi untuk memungkinkan terjadinyaproses fermentasi. Sebelum dan sesudah prosesfermentasi dilakukan pengamatan dan analisisparameter biokimia terkait dengan total solid(TS), kandungan oksigen (COD, BOD), pH,volatile fatty acid (VFA) dan volatile solid(VS).
Keberhasilan proses biokonversi ecenggondok menjadi biogas yang terjadi di dalambiogas digester diketahui dengan melakukananalisis produksi biogas, laju produksi biogasdan persentase gas metana (CH4). Produksibiogas diketahui melalui perbandingan volumebiogas yang dihasilkan dengan massa ecenggondok yang digunakan. Kadar metana diukurmelalui uji gas kromatografi (GC). Lajuproduksi biogas dihitung dari rata-rata volumebiogas yang dihasilkan per satuan waktu (jam).
Laju produksi =Rancangan percobaan yang dilakukantermasuk rancangan acak lengkap (RAL)factorial tunggal. Percobaan dilakukan dengan3 ulangan sehingga terdapat 12 unit percobaan.Data yang diperoleh dianalisis menggunakananalisis varians (ANOVA) yang akan dilakukanuji lanjut dengan uji DMRT (Duncan MultipleRange Test) pada taraf 1% dan 5%.
Dewi Murni dan Yuhelsa Putra 17
panas merupakan salah satu metodehidrotermal yang telah digunakan beberapadekade, contohnya pada industri kertas. Airdengan tekanan yang tinggi dapat melakukanpenetrasi ke dalam biomassa, menghidrasiselulosa serta menghilangkan hemiselulosa danlignin. Keuntungan utama metode ini adalahtidak perlu penambahan senyawa kimia dantidak membutuhkan material tahan korosi untukreaktor hidrolisis pada proses ini. Menurut Qiao(2011), setelah praperlakuan 1700 C/jam,produksi biogas dari biomassa lignoselulosameningkat dari 210 mL/g menjadi 238 mL/gbahan baku.
Selain kandungan lignoselulosa yang tinggiShankar et al., (2013) menyatakan bahwaeceng gondok tidak memiliki mikroba esensialuntuk mengawali proses produksi biogas. Olehkarena itu perlu penambahan EM4 sebagaipenyedia mikroba untuk mengawali prosesfermentasi eceng gondok menjadi biogas.Menurut Herawati dan Wibawa (2010), hasilterbaik pada penelitian pembuatan biogasialah dengan penambahan EM4, dimana yieldbiogas rata-rata dihasilkan 0,030 L/g VS. EM4mengandung bakteri fermentasi dari genusLactobaccilus, jamur fermentasi, Actinomycetes,bakteri pelarut fosfat dan ragi (Herawati andWibawa, 2010).
Untuk memperoleh biogas dengan produksidan persentase metana yang tinggi, perludiketahui lama (waktu) pemberianpraperlakuan hidrotermal terhadap ecenggondok yang tepat. Berdasarkan permasalahantersebut, perlu dilakukan penelitian untukmengetahui lama (waktu) perlakuanhidrotermal yang tepat untuk memperolehproduksi biogas dan persentase metana yangpaling optimal.
BAHAN DAN METODE
Penelitian ini dilakukan di LaboratoriumFakultas Pertanian, Universitas Sultan AgengTirtayasa. Eceng gondok diambil dari SungaiCiujung, Kabupaten Serang. Penelitiandilakukan dalam skala laboratorium dengan
metode eksperimen. Sebagai kontrol(pembanding), dievaluasi juga produksi biogasdari eceng gondok tanpa praperlakuanhidrotermal (hydrothermal pretreatment).
Eceng gondok dicacah dengan alat pencacah(slicer) dengan ukuran sekitar 6 mm. Ecenggondok hasil pencacahan dicampur dengan air(1:1). Campuran ini kemudian dimasukkan kedalam autoklaf dan direbus (hydrothermalpretreatment) pada suhu 1700C dengan variasiwaktu 30 menit, 60 menit dan 90 menit.Selanjutnya, eceng gondok dikeluarkan dariautoklaf dan dibiarkan terbuka sampaimencapai suhu ruangan. Setelah itu,ditambahkan EM4 sebagai starter untukmenginisiasi proses fermentasi. Campuran inikemudian dimasukkan ke pembangkit/ reaktorbiodigester anaerobic batch. Selanjutnyadiinkubasi untuk memungkinkan terjadinyaproses fermentasi. Sebelum dan sesudah prosesfermentasi dilakukan pengamatan dan analisisparameter biokimia terkait dengan total solid(TS), kandungan oksigen (COD, BOD), pH,volatile fatty acid (VFA) dan volatile solid(VS).
Keberhasilan proses biokonversi ecenggondok menjadi biogas yang terjadi di dalambiogas digester diketahui dengan melakukananalisis produksi biogas, laju produksi biogasdan persentase gas metana (CH4). Produksibiogas diketahui melalui perbandingan volumebiogas yang dihasilkan dengan massa ecenggondok yang digunakan. Kadar metana diukurmelalui uji gas kromatografi (GC). Lajuproduksi biogas dihitung dari rata-rata volumebiogas yang dihasilkan per satuan waktu (jam).
Laju produksi =Rancangan percobaan yang dilakukantermasuk rancangan acak lengkap (RAL)factorial tunggal. Percobaan dilakukan dengan3 ulangan sehingga terdapat 12 unit percobaan.Data yang diperoleh dianalisis menggunakananalisis varians (ANOVA) yang akan dilakukanuji lanjut dengan uji DMRT (Duncan MultipleRange Test) pada taraf 1% dan 5%.
Dewi Murni dan Yuhelsa Putra 17
panas merupakan salah satu metodehidrotermal yang telah digunakan beberapadekade, contohnya pada industri kertas. Airdengan tekanan yang tinggi dapat melakukanpenetrasi ke dalam biomassa, menghidrasiselulosa serta menghilangkan hemiselulosa danlignin. Keuntungan utama metode ini adalahtidak perlu penambahan senyawa kimia dantidak membutuhkan material tahan korosi untukreaktor hidrolisis pada proses ini. Menurut Qiao(2011), setelah praperlakuan 1700 C/jam,produksi biogas dari biomassa lignoselulosameningkat dari 210 mL/g menjadi 238 mL/gbahan baku.
Selain kandungan lignoselulosa yang tinggiShankar et al., (2013) menyatakan bahwaeceng gondok tidak memiliki mikroba esensialuntuk mengawali proses produksi biogas. Olehkarena itu perlu penambahan EM4 sebagaipenyedia mikroba untuk mengawali prosesfermentasi eceng gondok menjadi biogas.Menurut Herawati dan Wibawa (2010), hasilterbaik pada penelitian pembuatan biogasialah dengan penambahan EM4, dimana yieldbiogas rata-rata dihasilkan 0,030 L/g VS. EM4mengandung bakteri fermentasi dari genusLactobaccilus, jamur fermentasi, Actinomycetes,bakteri pelarut fosfat dan ragi (Herawati andWibawa, 2010).
Untuk memperoleh biogas dengan produksidan persentase metana yang tinggi, perludiketahui lama (waktu) pemberianpraperlakuan hidrotermal terhadap ecenggondok yang tepat. Berdasarkan permasalahantersebut, perlu dilakukan penelitian untukmengetahui lama (waktu) perlakuanhidrotermal yang tepat untuk memperolehproduksi biogas dan persentase metana yangpaling optimal.
BAHAN DAN METODE
Penelitian ini dilakukan di LaboratoriumFakultas Pertanian, Universitas Sultan AgengTirtayasa. Eceng gondok diambil dari SungaiCiujung, Kabupaten Serang. Penelitiandilakukan dalam skala laboratorium dengan
metode eksperimen. Sebagai kontrol(pembanding), dievaluasi juga produksi biogasdari eceng gondok tanpa praperlakuanhidrotermal (hydrothermal pretreatment).
Eceng gondok dicacah dengan alat pencacah(slicer) dengan ukuran sekitar 6 mm. Ecenggondok hasil pencacahan dicampur dengan air(1:1). Campuran ini kemudian dimasukkan kedalam autoklaf dan direbus (hydrothermalpretreatment) pada suhu 1700C dengan variasiwaktu 30 menit, 60 menit dan 90 menit.Selanjutnya, eceng gondok dikeluarkan dariautoklaf dan dibiarkan terbuka sampaimencapai suhu ruangan. Setelah itu,ditambahkan EM4 sebagai starter untukmenginisiasi proses fermentasi. Campuran inikemudian dimasukkan ke pembangkit/ reaktorbiodigester anaerobic batch. Selanjutnyadiinkubasi untuk memungkinkan terjadinyaproses fermentasi. Sebelum dan sesudah prosesfermentasi dilakukan pengamatan dan analisisparameter biokimia terkait dengan total solid(TS), kandungan oksigen (COD, BOD), pH,volatile fatty acid (VFA) dan volatile solid(VS).
Keberhasilan proses biokonversi ecenggondok menjadi biogas yang terjadi di dalambiogas digester diketahui dengan melakukananalisis produksi biogas, laju produksi biogasdan persentase gas metana (CH4). Produksibiogas diketahui melalui perbandingan volumebiogas yang dihasilkan dengan massa ecenggondok yang digunakan. Kadar metana diukurmelalui uji gas kromatografi (GC). Lajuproduksi biogas dihitung dari rata-rata volumebiogas yang dihasilkan per satuan waktu (jam).
Laju produksi =Rancangan percobaan yang dilakukantermasuk rancangan acak lengkap (RAL)factorial tunggal. Percobaan dilakukan dengan3 ulangan sehingga terdapat 12 unit percobaan.Data yang diperoleh dianalisis menggunakananalisis varians (ANOVA) yang akan dilakukanuji lanjut dengan uji DMRT (Duncan MultipleRange Test) pada taraf 1% dan 5%.

Dewi Murni dan Yuhelsa Putra 18
HASIL DAN PEMBAHASAN
Produksi BiogasProduksi biogas diukur setiap tiga hari sekali.Hasil pengukuran biogas menunjukkan bahwaproduksi biogas berbeda-beda pada masing-masing praperlakuan hidrotermal. Perbedaantersebut diantaranya dalam hal awal produksibiogas dan volume yang dihasilkan setiap 3hari (Gambar 1).
Gambar 1. Produksi biogas dengan variasi lamawaktu hidrotermal
Pada kontrol percobaan (tanpa praperlakuanhidrotermal), terbentuknya biogas dapat terukursetelah hari ke-9 yaitu sebesar 0,064 mL/g.Produksi biogas mulai meningkat di hari ke-18yaitu sebesar 0,530 mL/g. Produksi biogasselanjutnya menurun hingga hari ke-30 danmeningkat kembali serta mencapai puncakproduksi pada hari ke-39 sekitar 1,439 mL/g.Selanjutnya produksi turun drastis hingga akhirfermentasi. Penurunan tersebut disebabkan olehterganggunya aktivitas mikroorganisme akibatturunnya pH dan berkurangnya bahan organikyang dibutuhkan untuk pertumbuhan mikroorganisme.
Produksi biogas pada praperlakuanhidrotermal 30 menit dimulai pada hari ke-3.Hal ini mengindikasikan bahwa praperlakuanhidrotermal mampu meningkatkan aksesmikroorganisme terhadap selulosa sehinggamempercepat awal terbentuknya biogas. HasilpenelitianWinarni et al., (2011) menunjukkanbahwa, secara umum, awal terbentuknya biogas
eceng gondok dengan sistem batch adalahsetelah hari ke-5 fermentasi. Menurut Zeng etal., (2007), praperlakuan hidrotermalmemudahkan bakteri dalam menguraikanselulosa sehingga biogas bisa dihasilkan lebihcepat.
Produksi biogas pada praperlakuanhidrotermal 30 menit ini cenderung rendahhingga hari ke-15 yaitu berada dikisaran 0,31mL/g. Produksi yang kecil pada awalfermentasi dapat disebabkan oleh belumoptimalnya proses degradasi selulosa sehinggabahan organik yang dapat dikonversi menjadibiogas menjadi sedikit. Peningkatan produksiterjadi setelah hari ke-18 hingga mencapaipuncaknya pada hari ke-39 yakni 2,572 mL/g.Selanjutnya produksi biogas mengalami sedikitpenurunan hingga proses fermentasi selesai.Hasil penelitian Ferrer et al., (2010),menunjukkan bahwa praperlakuan hidrotermalselama 30 menit mampu meningkatkanhidrolisis eceng gondok dengan cara memicusolubiasi (pelarutan) eceng gondok sampaidengan 12%. Selulosa akan diuraikan menjadiglukosa yang merupakan bahan utamapembentukan asam asetat oleh bakteriasidogenik. Asam asetat inilah yang akandijadikan biogas oleh bakteri metanogenik.Pada akhirnya, pengaksesan selulosa olehenzim hidrolitik yang dipermudah akanmenghasilkan biogas yang lebih tinggidibanding tanpa praperlakuan.
Pada praperlakuan hidrotermal 60 menit,produksi biogas juga dimulai pada hari ke-3.Produksi biogas sampai hari 18 cenderungrendah dan berkisar sekitar 0,7 mL/g.Kemudian, mulai hari ke-21 terjadipeningkatan dan mencapai titik tertinggi padahari ke-39, yakni sekitar 4,709 mL/g. Puncakproduksi stabil hingga hari ke-48, kemudianterjadi penurunan hingga fermentasi berakhir.Produksi biogas pada praperlakuan ini lebihtinggi dibandingkan dengan praperlakuanlainnya. Hal tersebut menunjukkan bahwapraperlakuan hidrotermal 60 menit adalah lamawaktu yang paling tepat untuk produksi biogas

Dewi Murni dan Yuhelsa Putra 19
yang optimal. Praperlakuan ini mampumendegradasi unsur lignin yang terdapat padalignoselulosa eceng gondok dan memudahkanmikroba menghidrolisis selulosa sehinggamenghasilkan biogas lebih banyak. Pernyataantersebut sesuai dengan pendapat Zeng et al.(2007), yang mengatakan bahwa perebusanmaterial lignoselulosa dalam air panas(hidrotermal) merupakan salah satu metodehidrotermal yang telah digunakan beberapadecade. Penggunaan praperlakuan hidrotermalpada material lignoselulosa memudahkanbakteri dalam menguraikan selulosa. Airdengan tekanan yang tinggi dapat melakukanpenetrasi ke dalam biomassa, menghidrasiselulosa serta menghilangkan hemiselulosa danlignin. Mosier et al., (2005) juga menyatakanbahwa praperlakuan hidrotermal merupakansalah satu metode yang berpotensial tinggidalam meningkatkan kerja enzim hidrolisisbiomassa organik. Praperlakuan hidrotermalmampu merubah struktur biomassa lingoselulosa agar lebih mudah diakses oleh enzimyang akan mengubah polimer karbohidratmenjadi gula yang mudah untuk difermentasi.Caranya adalah memecah ikatan antara lignindan selulosa.
Produksi biogas pada praperlakuanhidrotermal pada 90 menit juga diawali padahari ke-3. Setelah terbentuknya biogas hinggahari ke-18, produksinya relatif rendah dankonstan, yakni berada pada kisaran 0,7 mL/g.Selanjutnya mulai hari ke-21, terjadipeningkatan secara signifikan hingga mencapaipuncak produksi pada hari ke-45, sebesar 3,332mL/g. Mulai hari ke-48 terjadi penurunansecara drastis hingga selesai proses fermentasi.
Penurunan produksi di akhir fermentasipada seluruh praperlakuan hidrotermaldisebabkan oleh terganggunya aktivitas bakteriakibat turunnya pH. Menurut Mara & Alit(2011), turunnya kadar pH menyebabkanmenurunnya produktivitas biogas. Hasilpenelitian Singhal & Rai (2003)3 menunjukkanbahwa rata-rata produksi biogas dari ecenggondok adalah 15,4-23,65 L/kg berat kering
eceng gondok yang diperoleh setelah 21 harifermentasi. Rata-rata produksi biogas tertinggidicapai pada hari ke 9-12 dari periodefermentasi, selanjutnya produksinya turunsecara bertahap.
Hasil analisis varians (ANOVA)menunjukkan bahwa F hitung (32,61) lebihbesar dari F tabel (4,07), artinya terdapatperbedaan yang nyata diantara kontrolpercobaan dengan kelompok praperlakuanhidrotermal. Hasil uji DMRT menunjukkanbahwa praperlakuan hidrotermal 60 menitmenghasilkan produksi biogas paling optimaldibandingkan dua praperlakuan lainnya.
Hasil analisis laju produksi biogasmenunjukkan bahwa praperlakuan hidrotermal60 menit menghasilkan biogas dengan lajuproduksi tertinggi yakni sekitar 13,467mL/jam. Hal tersebut semakin menguatkanbahwa pretreatment hidrotermal selama 60menit dengan suhu 170oC merupakanpraperlakuan yang paling optimal dalammeningkatkan laju produksi biogas. Lajuproduksi biogas pada masing-masingpraperlakuan hidrotermal dapat dilihat padaTabel 1.
No. Lama waktu PraperlakuanHidrotermal
Laju ProduksiBiogas
1. 0 menit 2,6659 mL/jam
2. 30 menit 8,1319 mL/jam
3. 60 menit 13,467 mL/jam
4. 90 menit 10,123 mL/jam
Laju produksi biogas rata-rata pada kontrolpercobaan (0 menit) lebih rendah dibandingkandengan laju produksi biogas denganpraperlakuan hidrotermal, yaitu hanya sebesar2,666 mL/jam. Hal tersebut disebabkan olehbelum terdegradasinya senyawa lignin,sehingga enzim yang dihasilkan olehmikroorganisme metanogenik tidak bisamengakses senyawa selulosa yang ada padaeceng gondok.
Pada perlakuan hidrotermal selama 90 menit,laju produksi biogasnya lebih kecil

Dewi Murni dan Yuhelsa Putra 20
dibandingkan 60 menit meskipun prosespendegradasian ligninnya lebih baik. Akantetapi, praperlakuan hidrotermal 90 menitmasih lebih baik dibandingkan denganpraperlakuan hidrotermal 30 menit. Lebihrendahnya laju produksi biogas padapraperlakuan hidrotermal 90 menitmenunjukkan bahwa dengan lama waktutersebut, tidak hanya senyawa lignin yangterdegradasi, namun juga senyawa lain yangmenghasilkan senyawa inhibitor. Selain itu,pemanasan yang terlalu lama jugamenyebabkan naiknya derajat disosiasi air danturunnya pH larutan.
Orchidea et al. (2010) menyatakan bahwabanyaknya produk monosakarida yangdiperoleh setelah perlakuan hidrotermalmemungkinkan terjadinya degradasimonosakarida menjadi senyawa turunannyayang bersifat inhibitor seperti furfural, HMFdan fenol. Menurut Orchidea et al., (2010),penggunaan suhu tinggi pada hidrotermaldengan waktu yang lama dapat mendegradasilignin namun hal itu dapat mempengaruhiderajat keasaman karena tingginya nilaikonstanta disosiasi air (Kw). Semakin lamaproses hidrotermal akan meningkatkan tekanan(kompresi) air sehingga pH larutan semakinrendah. Menurut Yonathan et al., (2013),produksi biogas dapat terbentuk secaramaksimal pada kondisi pH netral.
3.2 Persentase MetanaBiogas yang dihasilkan terdiri dari beberapakomponen yang dapat mempengaruhipersentase metana (CH4). Gas metanadiketahui terbentuk sebagai hasil dari aktivitasbakteri metanogenik dalam mengonversi asamasetat dan mereduksi CO2. Pada prosesmetanogenesis, selain CH4, terdapat hasilsampingan yang berupa CO2. Menurut Mara &Alit (2011), CH4 merupakan unsur pentingpada biogas yang menentukan terbakarnyabiogas dan berkaitan dengan energi yangdihasilkan biogas.
Untuk mengetahui kualitas biogas yangdihasilkan, dilakukan analisis GasChromatography (GC) yang menunjukkanpersentase gas metana (CH4) dibanding gaslain dalam campuran biogas. Hasil analisismenunjukkan bahwa biogas yang dihasilkanterdiri atas CH4, CO2, N2, dan O2 yangpersentasenya dapat dilihat pada Tabel 2.
Tabel 2. Komposisi biogas pada masing-masing praperlakuan hidrotermalPersentase (%)
Perlakuan CH4 CO2 N2 O20 menit 6,71 5,76 87,53 -30 menit 42,30 23,41 25, 88 8,4160 menit 43,38 24,50 32,12 -90 menit 30,57 15,12 54,31 -
Secara umum, persentase metana lebih besardibandingkan senyawa lainnya. Akan tetapi,persentase N2 pada kontrol percobaan (0menit) dan perlakuan 90 menit lebih besardibandingkan metana dan secara keseluruhanpersentase N2 cukup besar disbanding gassmapingan lainnya. Tingginya nilai N2mengakibatkan persentase metana belummencapai nilai standar umum biogas. Haltersebut kemungkinan terjadi akibat kebocoranpenampung gas ketika dalam perjalanan uji gaskromatografi. Faktor lainnya yangmenyebabkan rendahnya persentase metanapada kelompok kontrol adalah rendahnya nilaipH.
Nilai pH pada kontrol percobaan sesudahfermentasi (6,16) lebih rendah dibandingkannilai pH pada praperlakuan hidrotermal 60menit (6,49) dan 90 menit (6,27). Hal inimenandakan kurang optimalnya kerja bakterimetanogenik dalam mengubah asam organikmenjadi metana. Akibatnya, persentase metanayang didapat pada kontrol percobaan lebihkecil dibandingkan dengan praperlakuanhidrotermal. Yenni et al., (2012) menyatakanbahwa, perubahan nilai pH dapatmempengaruhi persentase metana dalam biogas.Kondisi pH yang tidak optimal akan
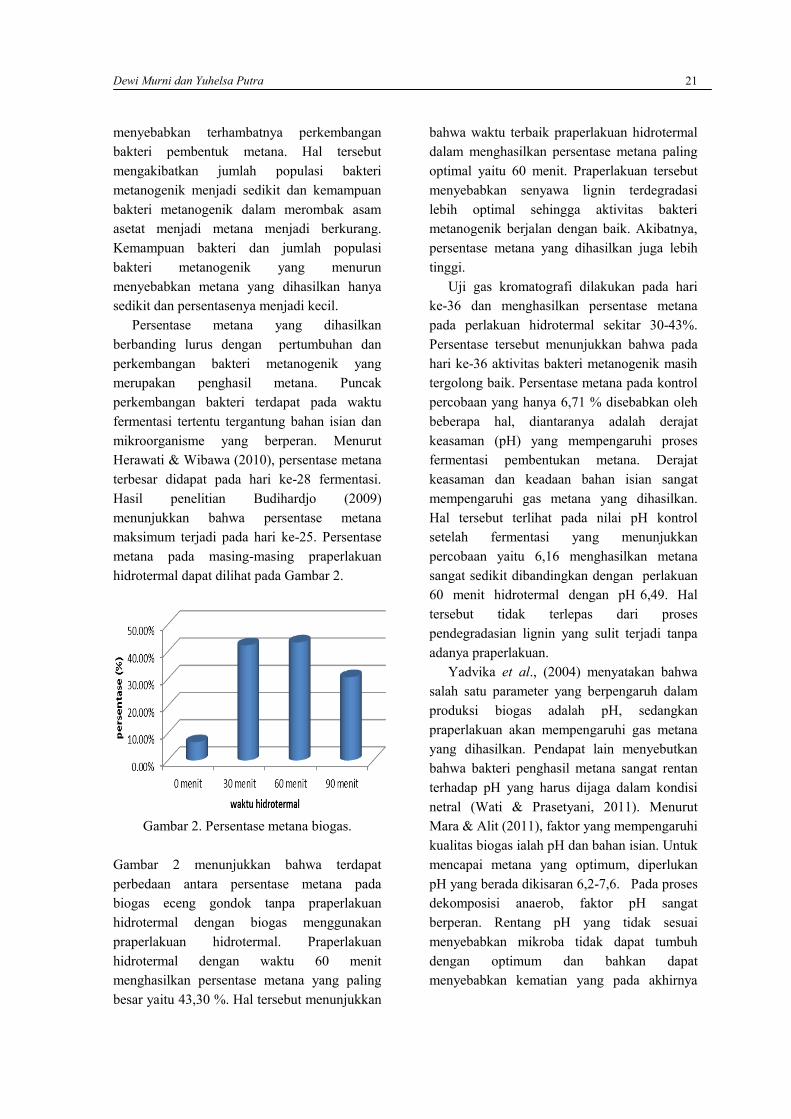
Dewi Murni dan Yuhelsa Putra 21
menyebabkan terhambatnya perkembanganbakteri pembentuk metana. Hal tersebutmengakibatkan jumlah populasi bakterimetanogenik menjadi sedikit dan kemampuanbakteri metanogenik dalam merombak asamasetat menjadi metana menjadi berkurang.Kemampuan bakteri dan jumlah populasibakteri metanogenik yang menurunmenyebabkan metana yang dihasilkan hanyasedikit dan persentasenya menjadi kecil.
Persentase metana yang dihasilkanberbanding lurus dengan pertumbuhan danperkembangan bakteri metanogenik yangmerupakan penghasil metana. Puncakperkembangan bakteri terdapat pada waktufermentasi tertentu tergantung bahan isian danmikroorganisme yang berperan. MenurutHerawati & Wibawa (2010), persentase metanaterbesar didapat pada hari ke-28 fermentasi.Hasil penelitian Budihardjo (2009)menunjukkan bahwa persentase metanamaksimum terjadi pada hari ke-25. Persentasemetana pada masing-masing praperlakuanhidrotermal dapat dilihat pada Gambar 2.
Gambar 2. Persentase metana biogas.
Gambar 2 menunjukkan bahwa terdapatperbedaan antara persentase metana padabiogas eceng gondok tanpa praperlakuanhidrotermal dengan biogas menggunakanpraperlakuan hidrotermal. Praperlakuanhidrotermal dengan waktu 60 menitmenghasilkan persentase metana yang palingbesar yaitu 43,30 %. Hal tersebut menunjukkan
bahwa waktu terbaik praperlakuan hidrotermaldalam menghasilkan persentase metana palingoptimal yaitu 60 menit. Praperlakuan tersebutmenyebabkan senyawa lignin terdegradasilebih optimal sehingga aktivitas bakterimetanogenik berjalan dengan baik. Akibatnya,persentase metana yang dihasilkan juga lebihtinggi.
Uji gas kromatografi dilakukan pada harike-36 dan menghasilkan persentase metanapada perlakuan hidrotermal sekitar 30-43%.Persentase tersebut menunjukkan bahwa padahari ke-36 aktivitas bakteri metanogenik masihtergolong baik. Persentase metana pada kontrolpercobaan yang hanya 6,71 % disebabkan olehbeberapa hal, diantaranya adalah derajatkeasaman (pH) yang mempengaruhi prosesfermentasi pembentukan metana. Derajatkeasaman dan keadaan bahan isian sangatmempengaruhi gas metana yang dihasilkan.Hal tersebut terlihat pada nilai pH kontrolsetelah fermentasi yang menunjukkanpercobaan yaitu 6,16 menghasilkan metanasangat sedikit dibandingkan dengan perlakuan60 menit hidrotermal dengan pH 6,49. Haltersebut tidak terlepas dari prosespendegradasian lignin yang sulit terjadi tanpaadanya praperlakuan.
Yadvika et al., (2004) menyatakan bahwasalah satu parameter yang berpengaruh dalamproduksi biogas adalah pH, sedangkanpraperlakuan akan mempengaruhi gas metanayang dihasilkan. Pendapat lain menyebutkanbahwa bakteri penghasil metana sangat rentanterhadap pH yang harus dijaga dalam kondisinetral (Wati & Prasetyani, 2011). MenurutMara & Alit (2011), faktor yang mempengaruhikualitas biogas ialah pH dan bahan isian. Untukmencapai metana yang optimum, diperlukanpH yang berada dikisaran 6,2-7,6. Pada prosesdekomposisi anaerob, faktor pH sangatberperan. Rentang pH yang tidak sesuaimenyebabkan mikroba tidak dapat tumbuhdengan optimum dan bahkan dapatmenyebabkan kematian yang pada akhirnya

Dewi Murni dan Yuhelsa Putra 22
dapat menghambat proses fermentasi sehinggaperolehan gas metana menjadi rendah.
Nilai pH pada praperlakuan 30 menit yanglebih kecil dari kontrol percobaan ternyatamenghasilkan metana yang lebih besar.Praperlakuan hidrotermal menyebabkanberkurangnya kadar lignin yang terkandungdalam eceng gondok sehingga pada perlakuan30 menit hidrotermal menghasilkan glukosalebih banyak dan menjadikan persentasemetana lebih besar. Hasil penelitian Orchidea etal. (2010) menunjukkan bahwa praperlakuanhidrotermal mampu mengurangi kadar ligninmencapai 22-23%. Pengurangan kadar ligninakan mengakibatkan laju hidrolisa meningkatdan juga akan berdampak terhadap daya serapdari enzim.
Persentase metana pada 90 menithidrotermal lebih kecil dari 60 dan 30 menithidrotermal (Gambar 2). Hal tersebut dapatdisebabkan praperlakuan hidrotermal selama 90menit menghasilkan glukosa yang tinggisehingga memungkinkan terbentuknya senyawaturunan glukosa yang bersifat inhibitor.Senyawa inhibitor itulah yang menyebabkanproses metanogenesis terhambat sehinggametana yang dihasilkan pretreatmenthidrotermal selama 90 menit lebih sedikit.Menurut Orchidea et al. (2010), proseshidrotermal dapat menghasilkan glukosa akibatpengaksesan terhadap selulosa yang lebihmudah. Semakin lama waktu hidrotermal makaproses degradasi lignin semakin besar. Selainitu, semakin lama waktu hidrotermal makakontak antara eceng gondok dan air semakinlama sehingga konversi selulosa menjadiglukosa semakin besar. Kandungan glukosadiharapkan hanya sedikit dihasilkan padaproses hidrotermal, karena proses maksimalkonversi selulosa menjadi glukosa diinginkanterjadi secara enzimatik Semakin kecil produkmonosakarida yang diperoleh setelah perlakuanhidrotermal maka semakin kecil kemungkinanterjadinya degradasi monosakarida menjadisenyawa turunannya yang bersifat inhibitorseperti furfural, HMF dan fenol. Pada
praperlakuan hidrotermal selama 60 menit,proses degradasi lignin berlangsung lebih baikdari praperlakuan hidrotermal selama 30 menit.Proses degradasi lignin tersebut akanmempermudah akses enzimatik terhadapselulosa pada proses fermentasi sehinggapersentase metana yang dihasilkan lebih baik.Pernyataan tersebut sesuai dengan hasilpenelitian Orchidea et al., (2010) bahwakerusakan struktur lignoselulosa yang palingoptimal didapatkan pada praperlakuanhidrotermal selama 60 menit. dan praperlakuanhidrotermal selama 60 menit tidakmenghasilkan senyawa inhibitor.
Taherzadeh & Karimi (2008) menyatakanbahwa praperlakuan hidrotermalmenyebabkan komponen lignoselulosabiomassa eceng gondok menjadi lebih mudahdiakses oleh bakteri melalui proses hidrolisisenzimatis. Pada akhir proses hidrolisis akandihasilkan glukosa. Senyawa ini selanjutnyaakan digunakan oleh bakteri asidogenik danmerubahnya menjadi senyawa asam asetat danalkohol. Senyawa asam asetat inilah yang akandijadikan sebagai bahan dasar pembentukanbiogas oleh bakteri metanogenik. Dengandemikian, peningkatan proses hidrolisis akanmeningkatkan aktivitas bakteri metanogenikdan pada akhirnya akan meningkatkan produksibiogas dan kandungan metana. Hadi dan El-Azeem (2008) menyatakan bahwa korelasisignifikan terlihat antara biogas dan metanayang dihasilkan dengan temperatur dan pH baikpada digester vertikal maupun horizontal.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Praperlakuan hidrotermal selama 60 menit
merupakan lama waktu paling tepat dalammenghasilkan produksi dan laju produksibiogas paling optimal. Produksinya adalah4,709 mL/g eceng gondok dengan lajuproduksi 13,467 mL/ jam dan mencapai titiktertinggi pada hari ke-39.

Dewi Murni dan Yuhelsa Putra 23
2. Praperlakuan hidrotermal selama 60 menitmenghasilkan biogas dengan persentasemetana tertinggi, yaitu 43, 38%.
DAFTAR PUSTAKA
Budihardjo, M.A. 2009. Pengaruh operasionalkombinasi feeding biostarter. JurnalPresipitasi, 6(2): 27-34.
Ferrer I., J. Palatsi, E. Campos, and X. Flotats.2010. Mesophilic and thermophilicanaerobic biodegradability of waterhyacinth pre-treated at 80oC. Wastemanagement, 30:1763-1767.
Gunnarsson, C. C. and C. M. Petersen. 2007.Water hyacinths as a resource inagriculture and energy production. WasteManagement, 27:117-129.
Hadi, A. M. A. and A. El-Azeem, S. A. M.(2008). Effect Of Heating, Mixing AndDigesterType On Biogas Production FromBuffalo Dung. Misr J. Ag. Eng.,25(4):1454-1477.
Herawati, D. A. and A. A. Wibawa. 2010.Pengaruh Pretreatment Jerami Padi padaProduksi Biogas dari Jerami Padi danSampah Sayur Sawi Hijau Secara Batch.Jurnal Rekayasa Proses, 4(1):25-29.
Mara, I.M. and I.B. Alit. 2011. Analisa kualitasdan kuantitas biogas dari kotoran ternak.
http://ejournal.ftunram.ac.id/FullPaper/Mara-Analisa-Kualitas dan Kuantitas-Biogas-dari Kotoran -Ternak.pdf. 8 hlm.16 agustus 2014 pk. 08.45.
Mosier N., C. Wyman, B. Dale, and R.Elander,Y.Y. Lee, M. Holtzapple, M.Ladisch. 2005. Features of promisingtechnologies for pretreatmentoflignocellulosic biomass. BioresourceTechnology, 96:673-686.
Orchidea R., A. Krishnanta, D. Ricardo, L.Febriyanti, K. Lazuardi, R. Pahlevi, andC.D. Mendila. 2010. Pengaruh metodepretreatment pada bahan lignosellulosaterhadap kualitas hidrolisat yangdihasilkan. Makalah seminar nasionalTeknik Kimia Soebardjo Brotohardjono:1-12.
Patil, J. H., A. MalourduAnd C. C. Gavimath.2011. Impact Of Dilution OnBiomethanation Of FreshWater Hyacinth.International Journal Of ChemicalSciences And Applications, 2:86-90.
Perna, C. and D. Burrows. 2005. Improveddissolved oxygen status following removalof exoticweed mats in important fishhabitat lagoons of the tropical BurdekinRiver floodplain, Australia, MarinePollution Bulletin. 51:138-148

ISSN 978-602-14989-0-3 BioETI
Uji daya hambat biopestisida formulasi minyak daun Cengkehdengan penambahan minyak Kayu Manis sebagai pengendaliColletotrichum pada buah Naga secara InvitroDITA OSRIANTI1, NASRILNASIR1*, FUJI ASTUTI FEBRIA1, JUMJUNIDANG2DANNURMANSYAH3
1Laboratorium Mikrobiologi, Jurusan Biologi, FMIPA, Universitas Andalas, Padang, Sumatera Barat 251632Balai Penelitian Tanaman Buah Tropika, Jl. Raya Solok-Aripan Km. 7 Solok3 Balai Penelitian Tanaman Rempah dan Obat. KP. Laing. Solok. Sumatera Barat.* koresponden : [email protected]
ABSTRACTPenelitian uji daya hambat formulasi minyak daun cengkeh dengan penambahan formulasi minyak kayu manis sebagaibiopestisida dalam pengendali jamur Colletotrichum sp. Secara invitro telah dilakukan di Balai Penelitian Tanaman Buahtropika (Balitbu) di Aripan Solok dilakukan dari bulan Mei sampai Juli 2014. Penelitian dilakukan secara eksperimendengan metode Rancangan Acak Lengkap (RAL) 5 perlakuan dan 5 ulangan untuk masing-masing perlakuan. Perlakuandengan kontrol (tanpa pemberian formulasi biopestisida), formulasi minyak cengkeh dengan penambahan formulasiminyak kayu manis dengan konsentrasi 250ppm, 500ppm, 1000ppm dan 1500ppm. Hasil efektif didapat pada perlakuanformulasi minyak cengkeh dengan penambahan formulasi minyak kayu manis dengan konsentrasi 1500ppm dapatmengendalikan jamur Colletorichum sp dengan daya hambat > 50%.
Kata Kunci : buah naga, Colletotrichum sp., biopestisida, cengkeh, kayu manis
Pendahuluan
Kebutuhan pasar yang menghendaki buah yangbebas pestisida kimia menjadi patokan yangpenting bagi petani Indonesia. Padahalserangan organisme penganggu tanaman (OPT)dapat mencapai diatas 20 persen (Djunaedy,2009), bahkan dalam beberapa kasus dapatlebih dari 50 % seperti pada buah naga (Nasir,2013). Kecenderungan pengendalian OPT saatini adalah menggunakan bahan kimia untukmengendalikan hama dan penyakit yangmenyerang tanaman mereka, karena cepat danefektif (Syahuti, 2012). Padahal harga pestisidakimia sangat tinggi mencapai 25-40 % daritotal biaya produksi pertanian. Dengandemikian harus ada upaya penguranganpenggunaan pestisida kimia dan beralih kepadajenis pestisida alami yang aman bagilingkungan (Djunaedy, 2009).
Di Indonesia, sangat banyak jenis tumbuhanpenghasil pestisida nabati, dan diperkirakan adasekitar 2400 jenis tanaman yang termasuk kedalam 235 famili (Kardinan, 1999). MenurutMorallo-Rijesus (1986) dalam Sastrosiswojo
(2002), jenis tanaman dari famili Asteraceae,Fabaceae dan Euphorbiaceae, dilaporkan palingbanyak mengandung bahan insektisida nabati.Salah satunya adalah daun cengkeh (Syzygiumaromaticum) yang dapat dijadikan alternatifyang bisa digunakan sebagai bahan bakupestisida alami (Mahardianto, 2007).
Salah satunya adalah daun cengkeh(Syzygium aromaticum) yang dapat dijadikanalternatif yang bisa digunakan sebagai bahanbaku pestisida alami (Mahardianto, 2007).Menurut Benes (1963), minyak cengkeh dapatmenghambat pertumbuhan jamur Fusicladiumeffusum penyebab penyakit kudis pada tanamankemiri. Ekstrak cengkeh atau biasa disebutminyak cengkeh mengandung bahan aktifterutama eugenol, eugenol asetat dan β-karyofilen. Eugenol adalah senyawa yangmenyebabkan aroma khas, sedangkan β-karyofilen adalah sesquiterpen yang mem-berikan rasa pahit dan merupakan bahan bakusintesis senyawa-senyawa lain. Besarnya kadarbahan-bahan aktif tersebut bervariasitregantung sumber ekstrak (bunga atau daun)

Dita Osrianti, Nasrilnasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 25
dan unsur tanaman (Furia dan Bellanca,1975;Suherdi dan Risfaheri,1992)
Minyak limbah kayu manis (Cinnamomumburmanii) daun, ranting dan kulit bersifatfungisida, minyak tanamana ini mengandungsenyawa sinemaldehid (Rusli dkk., 1985)minyak limbah kayu manis telah dijadikanformulasi pestisida nabati dan cukup efektifuntuk mengendalikan patogen Fusariumoxyforum, Phytopthora capsici, Sclerotiumrofsii dan Phytopthora cinnamomi(Nurmansyah, 2001 dan Nurmansyah dkk.,2003). Kandungan sinemaldehid yang terdapatpada kayu manis mencapai 49,60-75,95%(Rusli dan Hamid, 1990) juga mampumengendalikan jamur patogen tanaman, sepertiSclerotium rofsii, Fusarium sp. isolate cabai,kacang tanah dan tomat (Nurmansyah,1997).
Akhir-akhir ini buah naga banyak dibudi-dayakan oleh para petani termasuk di Indonesia,karena tanaman ini selain memiliki nilai jualyang tinggi juga mengandung gizi yang tinggi(Mahardianto, 2007). Menurut Mahsyait et al.,(2009) dan Nasir (2013) Colletotrichumgloeosporioides adalah salah satu penyebabpenyakit pada buah naga yang dilaporkanmenyerang perkebunan buah naga di Indonesiadan Malaysia. Di Malaysia serangan C.gloeosprioides mampu menekan produksidiatas 50 persen (masyahit, 2009).
Jamur Colletotrichum gloeosproides adalahjamur patogen yang menyerang banyaktanaman, gajalanya ditandai dengan bercakbulat yang berwarna coklat tua sampai hitamdan dibatasi halo berwarna kuning. Sejauh inibelum ada laporan keberhasilan pengendalianColletotrichum gloeosporioides pada buah naga,baik di Indonesia maupun di Malaysia.Sementara itu serangannya semakin meluas.Penelitian ini akan menggunkan kemampuanbiopestisida minyak daun Syzygiumaromaticum dengan penambahan Cinnamomumburmanii dalam menekan seranganColletotrichum sp. pada tanaman buah nagamerah.
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan Mei sampaiJuli 2014 di Labor Proteksi Balai PenelitianTanaman Buah (Balitbu) Tropika di Solok.
Penelitian ini menggunakan metodaeksperimen dengan tanpa pemberianbiopestisida sebagai control, formulasi minyakdaun cengkeh dengan penambahan formulasiminyak daun kayu manis pada konsentrasi250ppm, 500ppm, 1000ppm dan 1500ppmdengan 5 kali ulangan. Analisis data disajikansecara deskriptif. Koleksi isolat jamurColletotrichum sp. diperoleh dari BalaiPenelitian Tanaman Buah (Balitbu) Tropika, diAripan, Solok dan formulasi minyak Piperaduncum diperoleh dari Balai TanamanRempah dan Obat (Balitro) di Laing, Solok.
Parameter yang diamati berupa per-tumbuhan koloni jamur pada media PDA padamasing-masing perlakuan yang dibandingkandengan perlakuan kontrol. Pengamatandilakukan sampai diameter jamurColletotrichum sp. pada perlakuan kontrolmemenuhi cawan petri. Daya hambat pengaruhpemberian formulasi minyak cengkeh dengantambahan formulasi minyak kayu manisterhadap pertumbuhan jamur Colletotrichum sp.dihitung dengan rumus sebagai berikut:
Dari hasil uji daya hambat formulasi minyakcengkeh dengan tambahan minyak kayu manisterhadap pertumbuhan jamur Colletotrichumsp., dapat dilakukan uji resistensi patogen padaperlakuan yang memiliki daya hambat 100%(tidak tumbuh). Uji resistensi dilakukan dengancara memindahkan kembali inokulasi jamurpatogen yang pertumbuhannya terhambatkarena pengaruh pemberian formulasi minyakcengkeh dengan tambahan formulasi minyakkayu manis ke medium PDA tanpapenambahan formulasi minyak. Selanjutnyadiinkubasi pada suhu ruangan. Pertumbuhankoloni diketahui dengan mengukur diameterkoloni jamur yang tumbuh pada setiap
Dita Osrianti, Nasrilnasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 25
dan unsur tanaman (Furia dan Bellanca,1975;Suherdi dan Risfaheri,1992)
Minyak limbah kayu manis (Cinnamomumburmanii) daun, ranting dan kulit bersifatfungisida, minyak tanamana ini mengandungsenyawa sinemaldehid (Rusli dkk., 1985)minyak limbah kayu manis telah dijadikanformulasi pestisida nabati dan cukup efektifuntuk mengendalikan patogen Fusariumoxyforum, Phytopthora capsici, Sclerotiumrofsii dan Phytopthora cinnamomi(Nurmansyah, 2001 dan Nurmansyah dkk.,2003). Kandungan sinemaldehid yang terdapatpada kayu manis mencapai 49,60-75,95%(Rusli dan Hamid, 1990) juga mampumengendalikan jamur patogen tanaman, sepertiSclerotium rofsii, Fusarium sp. isolate cabai,kacang tanah dan tomat (Nurmansyah,1997).
Akhir-akhir ini buah naga banyak dibudi-dayakan oleh para petani termasuk di Indonesia,karena tanaman ini selain memiliki nilai jualyang tinggi juga mengandung gizi yang tinggi(Mahardianto, 2007). Menurut Mahsyait et al.,(2009) dan Nasir (2013) Colletotrichumgloeosporioides adalah salah satu penyebabpenyakit pada buah naga yang dilaporkanmenyerang perkebunan buah naga di Indonesiadan Malaysia. Di Malaysia serangan C.gloeosprioides mampu menekan produksidiatas 50 persen (masyahit, 2009).
Jamur Colletotrichum gloeosproides adalahjamur patogen yang menyerang banyaktanaman, gajalanya ditandai dengan bercakbulat yang berwarna coklat tua sampai hitamdan dibatasi halo berwarna kuning. Sejauh inibelum ada laporan keberhasilan pengendalianColletotrichum gloeosporioides pada buah naga,baik di Indonesia maupun di Malaysia.Sementara itu serangannya semakin meluas.Penelitian ini akan menggunkan kemampuanbiopestisida minyak daun Syzygiumaromaticum dengan penambahan Cinnamomumburmanii dalam menekan seranganColletotrichum sp. pada tanaman buah nagamerah.
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan Mei sampaiJuli 2014 di Labor Proteksi Balai PenelitianTanaman Buah (Balitbu) Tropika di Solok.
Penelitian ini menggunakan metodaeksperimen dengan tanpa pemberianbiopestisida sebagai control, formulasi minyakdaun cengkeh dengan penambahan formulasiminyak daun kayu manis pada konsentrasi250ppm, 500ppm, 1000ppm dan 1500ppmdengan 5 kali ulangan. Analisis data disajikansecara deskriptif. Koleksi isolat jamurColletotrichum sp. diperoleh dari BalaiPenelitian Tanaman Buah (Balitbu) Tropika, diAripan, Solok dan formulasi minyak Piperaduncum diperoleh dari Balai TanamanRempah dan Obat (Balitro) di Laing, Solok.
Parameter yang diamati berupa per-tumbuhan koloni jamur pada media PDA padamasing-masing perlakuan yang dibandingkandengan perlakuan kontrol. Pengamatandilakukan sampai diameter jamurColletotrichum sp. pada perlakuan kontrolmemenuhi cawan petri. Daya hambat pengaruhpemberian formulasi minyak cengkeh dengantambahan formulasi minyak kayu manisterhadap pertumbuhan jamur Colletotrichum sp.dihitung dengan rumus sebagai berikut:
Dari hasil uji daya hambat formulasi minyakcengkeh dengan tambahan minyak kayu manisterhadap pertumbuhan jamur Colletotrichumsp., dapat dilakukan uji resistensi patogen padaperlakuan yang memiliki daya hambat 100%(tidak tumbuh). Uji resistensi dilakukan dengancara memindahkan kembali inokulasi jamurpatogen yang pertumbuhannya terhambatkarena pengaruh pemberian formulasi minyakcengkeh dengan tambahan formulasi minyakkayu manis ke medium PDA tanpapenambahan formulasi minyak. Selanjutnyadiinkubasi pada suhu ruangan. Pertumbuhankoloni diketahui dengan mengukur diameterkoloni jamur yang tumbuh pada setiap
Dita Osrianti, Nasrilnasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 25
dan unsur tanaman (Furia dan Bellanca,1975;Suherdi dan Risfaheri,1992)
Minyak limbah kayu manis (Cinnamomumburmanii) daun, ranting dan kulit bersifatfungisida, minyak tanamana ini mengandungsenyawa sinemaldehid (Rusli dkk., 1985)minyak limbah kayu manis telah dijadikanformulasi pestisida nabati dan cukup efektifuntuk mengendalikan patogen Fusariumoxyforum, Phytopthora capsici, Sclerotiumrofsii dan Phytopthora cinnamomi(Nurmansyah, 2001 dan Nurmansyah dkk.,2003). Kandungan sinemaldehid yang terdapatpada kayu manis mencapai 49,60-75,95%(Rusli dan Hamid, 1990) juga mampumengendalikan jamur patogen tanaman, sepertiSclerotium rofsii, Fusarium sp. isolate cabai,kacang tanah dan tomat (Nurmansyah,1997).
Akhir-akhir ini buah naga banyak dibudi-dayakan oleh para petani termasuk di Indonesia,karena tanaman ini selain memiliki nilai jualyang tinggi juga mengandung gizi yang tinggi(Mahardianto, 2007). Menurut Mahsyait et al.,(2009) dan Nasir (2013) Colletotrichumgloeosporioides adalah salah satu penyebabpenyakit pada buah naga yang dilaporkanmenyerang perkebunan buah naga di Indonesiadan Malaysia. Di Malaysia serangan C.gloeosprioides mampu menekan produksidiatas 50 persen (masyahit, 2009).
Jamur Colletotrichum gloeosproides adalahjamur patogen yang menyerang banyaktanaman, gajalanya ditandai dengan bercakbulat yang berwarna coklat tua sampai hitamdan dibatasi halo berwarna kuning. Sejauh inibelum ada laporan keberhasilan pengendalianColletotrichum gloeosporioides pada buah naga,baik di Indonesia maupun di Malaysia.Sementara itu serangannya semakin meluas.Penelitian ini akan menggunkan kemampuanbiopestisida minyak daun Syzygiumaromaticum dengan penambahan Cinnamomumburmanii dalam menekan seranganColletotrichum sp. pada tanaman buah nagamerah.
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan Mei sampaiJuli 2014 di Labor Proteksi Balai PenelitianTanaman Buah (Balitbu) Tropika di Solok.
Penelitian ini menggunakan metodaeksperimen dengan tanpa pemberianbiopestisida sebagai control, formulasi minyakdaun cengkeh dengan penambahan formulasiminyak daun kayu manis pada konsentrasi250ppm, 500ppm, 1000ppm dan 1500ppmdengan 5 kali ulangan. Analisis data disajikansecara deskriptif. Koleksi isolat jamurColletotrichum sp. diperoleh dari BalaiPenelitian Tanaman Buah (Balitbu) Tropika, diAripan, Solok dan formulasi minyak Piperaduncum diperoleh dari Balai TanamanRempah dan Obat (Balitro) di Laing, Solok.
Parameter yang diamati berupa per-tumbuhan koloni jamur pada media PDA padamasing-masing perlakuan yang dibandingkandengan perlakuan kontrol. Pengamatandilakukan sampai diameter jamurColletotrichum sp. pada perlakuan kontrolmemenuhi cawan petri. Daya hambat pengaruhpemberian formulasi minyak cengkeh dengantambahan formulasi minyak kayu manisterhadap pertumbuhan jamur Colletotrichum sp.dihitung dengan rumus sebagai berikut:
Dari hasil uji daya hambat formulasi minyakcengkeh dengan tambahan minyak kayu manisterhadap pertumbuhan jamur Colletotrichumsp., dapat dilakukan uji resistensi patogen padaperlakuan yang memiliki daya hambat 100%(tidak tumbuh). Uji resistensi dilakukan dengancara memindahkan kembali inokulasi jamurpatogen yang pertumbuhannya terhambatkarena pengaruh pemberian formulasi minyakcengkeh dengan tambahan formulasi minyakkayu manis ke medium PDA tanpapenambahan formulasi minyak. Selanjutnyadiinkubasi pada suhu ruangan. Pertumbuhankoloni diketahui dengan mengukur diameterkoloni jamur yang tumbuh pada setiap

Dita Osrianti, Nasrilnasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 26
perlakuan. Jika koloni jamur tersebut tidaktumbuh, berarti formulasi minyak yangdiberikan dapat membunuh koloni jamur,namun jika koloni jamur tersebut tumbuhberarti formulasi minyak yang diberikan hanyadapat menghambat pertumbuhan koloni jamur.
HASIL DAN PEMBAHASAN
Dari pengamatan yang dilakukan didapatkanpersentase daya hambat biopestisida formulasiminyak daun cengkeh dengan penambahanformulasi minyak daun kayu manis beberapakonsentrasi dalam menghambat pertumbuhanjamur Colletotrichum sp. sebagai berikut:
Tabel 1. Persentase daya hambat biopestisida dalammenghambat pertumbuhan jamurColletotrichum sp pada hari ke-9 setelahinokulasi
Konsentrasi(ppm)
Diameterkoloni (cm)
Persentase Dayahambat (%)
0 9a 0250 7b 22,2500 2,79c 70,61000 2,16d 76,01500 1,96e 78,2
Keterangan: Angka-angka yang diikuti oleh notasi huruf yangsama, menunjukkan nilai yang berbeda tidaknyata dalam uji Duncan’s 5%.
Hasil analisis statistik menunjukkan bahwapemberian konsentrasi yang berbedaberpengaruh nyata terhadap pertumbuhanjamur Colletotrichum sp. Analisis Duncanmenunjukkan bahwa perlakuan 0ppm tidakberbeda nyata dengan perlakuan 250ppm,berbeda dengan perlakuan 500ppm, 1000ppmdan 1500ppm. Namun pada konsentrasi500ppm, 1000ppm, dan 1500ppm tidak berbedanyata dengan pertumbuhan jamurColletotrichum sp.
Pada konsentrasi minyak daun cengkehdengan tambahan minyak daun kayu manussebesar 500ppm, 1000ppm dan 1500ppmadalah konsentrasi efektif yang mampumenekan pertumbuhan jamur Colletotrichumsp., dengan demikian biopestisida formulasiminyak daun cengkeh dengan tambahanminyak daun kayu manis dapat digunakansebagai bahan anti jamur khususnya jamurColletotrichum sp.. Kandungan eugenol yangtinggi dalam cengkeh ditambahkan dengankayu manis yang mengandung sinemaldehidyang tinggi menunjukkan kesinergisan
sehingga dapat menghambat pertumbuhanjamur Colletotrichum sp. sebesar 78,22%.
Kandungan sinemaldehid pada kayu manismeningkatkan kemampuan eugenol padaminyak daun cengkeh, kandungan minyak padadaun kayu manis adalah 15,46% (Nurmansyahdkk, 2002). Menurut Suprianto (2008), senyawaantijamur dapat menghambat pertumbuhanmikroba melalui inaktivasi atau mengganggusatu atau lebih target subseluler seperti merusakdinding sel, mengganggu permeabilitasmembran, menghambat enzim-enzim metabolik,menghambat sintesis protein dan sintesis asamnukleat.
a b c d eGambar 1. Pertumbuhan diameter koloni jamur
Colletotrichum sp. dengan perlakuan (a)control, (b) Formulasi minyak cengkeh dankayu manis 250ppm, (c) Formulasi minyakcengkeh dan kayu manis 500ppm, (d)Formulasi minyak cengkeh dan kayu manis1000ppm, (e) Formulasi minyak cengkehdan kayu manis 1500ppm
Dari Gambar 1 dapat dilihat bahwa pengaruhaditif minyak daun kayu manis dapatmeningkatkan daya antifungal formulasibiopestisida minyak daun cengkeh.Meningkatkan daya antifungal ini didugakarena disebabkan karena kandungan utamasenyawa kimia minyak daun cengkeh adalaheugenol bersinergis dengan kandungan minyakkayu manis yang memiliki kandungan utamasinemaldehid. Selanjutnya minyak dauncengkeh juga efektif untuk mengendalikanjamur Aspergilus nidulans, Penicilliumfrequentans, P. cylopium dan Fusariumgraminaerum (Kurita dan Koike, 1982).
KESIMPULAN
Berdasarkan penelitian yang telah dilakukantentang uji daya hambat biopestisida formulasiminyak cengkeh dengan penambahan minyakdaun kayu manis dapat bersinergis sehinggadapat menekan pertumbuhan jamurColletotrichum sp dengan daya hambat sebesar78,22% pada konsentrasi 1500ppm

Dita Osrianti, Nasrilnasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 27
DAFTAR PUSTAKA
Djunaedy, A. 2009. Biopestisida sebagaiPengendalian Organisme PengangguTanaman (OPT) Yang Ramah Lingkungan.Embryo vol. 6 No. 1
Furia, T. E. and N. Bellanca. 1975. Fenaroli’sHandbook of Falvor Ingredients. Volume2. Cleveland: CRC Press
Kardinan, A., M. Iskandar dan E.A. Wikardi.1998. Pengaruh cara aplikasi minyaksuling Melaleuca bracteata dan metileugenol terhadap daya pikat lalat buahBactrocera dorsalis. Jurnal PerlindunganTanaman Indonesia 4(1) : 38-45.
Kurita, N., S. Koike. 1982. Synergisticantimicrobial effect of sodium chlorideand essential oil components. Agric. Biol.Chem., 46: 159-165.
Mahardianto, Nur. 2007. Budidaya Buah Naga(Dragon Fruit). http://agribisnis.deptan.go.id [12 November 2013].
Masyahit, M., Sijam, K,. Y. Awang andM.G.M Satar. 2009. The First Report ofthe Occurance of Anthracnose Diseases
Caused by Colletotrichum gloeosporoideson Dragon Fruit (Hylocereus spp.) inPeninsular Malaysia. Am. J. Applied Sci.6, 902-912.
Nasir, N. 2013. Serangan Penyakit PentingPada Buah Naga Hylocereus polyrhizus diKepulauan Riau. Laporan Penelitian.Jurusan Biologi FMIPA UniversitasAndalas. 5 hal.
Nurmansyah, Idris. H., Jamalius dan A. Asman.2003. Formulasi fungisida nabati minyaklimbah kayumanis dan pengaruhnyaterhadap pathogen. P. capsici, P.oxysporum dan S. rofsii. Kumpulan HasilPerkebunan. Kebun Percobaan LaingSolok. p. 1-5.
Rusli, S dan Hamid. 1990.. Kayumanis(Cinammomum burmanii) Perkembanganpenelitian tanaman penghasil minyak atsiri.Edisi khusus Penelitian Tanaman Rempahdan Obat vol VI No. 1 Balittro. Bogor.
Syahyuti. 2012. Sejarah Hama dan PestisidaIndonesia. http://syahyuti.blogspot.com/Sejarah Hama dan Pestisida Indonesia.

ISSN 978-602-14989-0-3 BioETI 2014
Jenis Semut (Hymenoptera: Formicidae) padaMacaranga spp.(Euphorbiaceae) di Cagar Alam Bukit Barisan, Rimbo Panti,dan Pangean, Sumatera BaratDIYONA PUTRI1, HENNY HERWINA1, RIJAL SATRIA2 DAN ALAN HANDRU1
1Laboratorium Taksonomi Hewan, Jurusan Biologi, FMIPA, Universitas Andalas, Kampus Limau Manis Padang 251632Taxonomy Laboratory, Graduate School of Science and Engineering Tokyo Metropolitan Universityemail: [email protected]
ABSTRACTPenelitian ini bertujuan untuk mengetahui jenis-jenis semut (Hymenoptera: Formicidae) pada Macaranga spp.(Euphorbiaceae) di Cagar Alam Bukit Barisan, Rimbo Panti, dan Pangean Sumatera Barat. Pengambilan sampeldilakukan pada bulan Maret, April dan Juni 2014 dengan menggunakan metode Free Collection dan Colony Collection.Pada penelitian ini ditemukan sebanyak enam jenis semut yang tergolong ke dalam lima genera, empat tribe, tigasubfamili. Subfamili Formicinae memiliki jenis yang paling banyak (tiga jenis) diikuti Subfamili Myrmicinae (dua jenis)dan Subfamili yang paling sedikit adalah Ponerinae yang hanya terdiri dari satu jenis. Jenis semut yang paling banyakdidapatkan jumlah individunya adalah Crematogaster borneensis (Andre, 1896) (Myrmicinae) (414 individu). Jenis iniditemukan pada tiga jenis Macaranga (Macaranga bancana (Miq.) Mull.Arg.), Macaranga depressa (Mull. Arg), danMacaranga gigantea (Reichb.F. & Zoll.).
Key words: Semut, Macaranga, Cagar Alam, free collection, colony collection
Pendahuluan
Semut adalah salah satu kelompok seranggayang keberadaan dan habitatnya sangat umumdan tersebar luas. Semut juga merupakan salahsatu contoh serangga yang paling sukses darikelompok serangga lainnya (Borror et al.,1992). Semut dapat membuat sarang di sekitartempat tinggal kita misalnya di atas gundukantanah, sampah, pot bunga, pohon, sudut rumahdan lain-lain (Borror, 2005). Semut ada yangbersarang di dalam tanah, seperti pada genusOdontoponera, Solenopsis dan Pheidole. Adapula semut yang bersarang pada dedaunan(Oecophylla smaragdina). Ada yang bersarangpada kayu yang telah lapuk, seperti;Camponatus dan Dolichoderus. Sebagiansemut bersarang pada tumpukan serasah,seperti yellow crazy ants (Anoplolepisgracilipes) dan crazy ant (Paratrechinalongicornis) (Holldobler dan Wilson, 1990;Bolton, 1994; Borror et al., 1992; Agosti et al.,2000).
Beberapa jenis semut berasosiasi ataubersimbiosis dengan tumbuhan, seperti genus;
Crematogaster dan Iridomyrmex bersimbiosisdengan tumbuhan sarang semut (Myrmecodia),Crematogaster dengan Macaranga tribola dandengan tumbuh-tumbuhan yang dibudidayakanseperti; Theobroma cacao, kulit manis danlainnya (Holldobler dan Wilson, 1990; Bolton,1994; Borror et al., 1992; Agosti et al., 2000).Banyak semut yang mendiami akar tumbuhanepifit (misalnya: Anochetus dan Strumigenyssp.) (Agosti et al., 2000).
Interaksi semut dengan tumbuhan dapatdilihat pada tumbuhan Macaranga. Ada tigatipe utama dari asosiasi; non-myrmecophyticjenis, myrmecophytes dan myrmecophytesobligat. Macaranga (Euphorbiaceae) adalahtanaman yang hanya terdapat di Asia tropisdengan substansi dari myrmecophytes.Myrmecophytes berhubungan dengan semutspesifik yang hidup dalam batang ruas tanaman(Ong, 1978; Fiala and Maschwitz, 1990).
Di Malay Archipelago telah ditemukan 23jenis semut yang berasosiasi dengan tumbuhanMacaranga. Macaranga terdiri dari berbagaijenis, dari yang tidak dihuni oleh semut sampaiyang telah berasosiasi dengan semut

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 29
(myemecophytic) (Fiala and Maschwitz, 1991,1992 a, b; Fiala 1992). Macaranga merupakancontoh genus dari famili Euphorbiaceae yang didistribusikan dari Afrika ke Polinesia dansangat banyak terdapat di daerah Malesia. Jenisini banyak mendiami daerah bukaan dan tepihutan. Populasi umumnya ditemukan di semak-semak, tanah terlantar, di tepi hutan atau dihutan rawa (Corner, 1988).
Cagar Alam merupakan salah satu kawasanhutan konservasi di Sumatera Barat sepertiCagar Alam Bukit Barisan, Cagar Alam RimboPanti, dan Cagar Alam Pangean. Adanyadaerah bukaan lahan hutan di beberapa cagaralam ini sebagai akibat ulah manusia yangdijadikan areal perladangan menimbulkankerusakan di beberapa tempat. Kerusakan hutantersebut akan mengganggu kehidupan berbagaijenis satwa yang ada di Cagar Alam. Dengandemikian penelitian mengenai interaksi semutdengan tumbuhan perlu dilakukan, dimanasebagai studi penelitian pendahuluan terlebihdahulu kita harus mengetahui jenis-jenis semutyang mendiami tumbuhan yang dijadikansebagai tempat bersarang bagi semut. Adapunjenis tumbuhan Macaranga adalah sebagaisalah satu tumbuhan pionir yang bersimbiosisdengan semut.
Informasi tentang semut pada Macaranga diIndonesia belum banyak tersedia, termasuk diSumatera Barat, sehingga penelitian mengenaijenis-jenis semut yang terdapat pada tumbuhanMacaranga di beberapa Cagar Alam SumateraBarat perlu dilakukan.
BAHAN DAN METODE
Lokasi PenelitianPenelitian dilakukan pada tiga cagar alam diSumatera Barat, yaitu Cagar Alam BukitBarisan (Padang Pariaman, Tanah Datar, Solok,Kota Padang, Sumatera Barat), Cagar AlamRimbo Panti (Pasaman, Sumatera Barat) danCagar Alam Pangean (Sawah Lunto, Sijunjung,Solok, Sumatera Barat) (Gambar 1).
Pengoleksian SampelPenelitian dilakukan pada bulan Maret, April,dan Juni 2014 dengan menggunakan metodeFree Collection dan Colony Collection. MetodeFree Collection merupakan metodepengoleksian semut secara langsung dengantransek sepanjang 180 meter pada masing-masing cagar alam dengan menggunakan pinsetatau aspirator kemudian dimasukkan ke dalamvial yang telah berisi alkohol 96%. MetodeColony Collection merupakan pengambilansampel dengan melakukan pencarian kolonisemut yang berada pada tumbuhan Macarangaspp. (Euphorbiaceae) kemudian semutdimasukkan ke dalam plastik koleksi untukselanjutnya dibawa ke laboratorium. Semuasampel yang didapatkan diolah di LaboratoriumRiset Taksonomi Hewan, Jurusan Biologi,FMIPA, Universitas Andalas.
Pengolahan SampelPengolahan sampel dilakukan dengan beberapatahap, yaitu; penyortiran, identifikasimorfospesies, mounting sampel, pelabelan danpenyimpanan. Proses pengidentifikasiandiupayakan sampai pada tingkat spesies, namunsampel yang hanya teridentifikasi sampaitingkat genus diberi kode di belakang genussesuai kolektor {SKY: Seiki Yamane,Kagoshima University; HH: Henny Herwina,Universitas Andalas} (Ito et al., 2001). Semutyang didapatkan dikelompokkan berdasarkansubfamili, genus, jenis dan dihitung jumlahindividunya, lalu dibuat tabel daftar jenis danindividu per subfamili.
HASIL DAN PEMBAHASAN
Cendawan Secara keseluruhan didapatkanenam jenis semut yang tergolong ke dalam limagenera, empat tribe, tiga subfamili semut pada16 sampel pohon Macaranga spp. di beberapaCagar Alam Sumatera Barat. Semut tersebutditemukan pada tiga jenis tumbuhan Macaranga(Macaranga bancana (Miq.) Mull.Arg.),Macaranga depressa (Mull. Arg) dan

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 30
Tabel 1. Jumlah individu semut pada tiap jenis tumbuhan Macaranga yang dikoleksi dengan metode FreeCollection di beberapa Cagar Alam Sumatera Barat (BB: Cagar Alam Bukit Barisan, RP: CagarAlam Rimbo Panti, P: Cagar Alam Pangean; 1: Macaranga bancana, 2: Macaranga depressa, 3:Macaranga gigantea).
SubFamili Lokasi TotalTribe
JenisFormicinae
CamponotiniPolyrhachis (Myrma) proxima
LasiiniNylanderia sp. 1 of HHParaparatrechina sp. 2 of HH
MyrmicinaeCrematogastrini
Crematogaster borneensisCrematogaster modiglani
PonerinaePonerini
Pachycondyla (Brachyponera) sp. 28 of SKYTotal IndividuTotal JenisTotal GenusTotal TribeTotal Subfamili
BB1--1--1--376----3783332
-3----28---11--1314333
RP1--2-----5----72222
P1--------1130---412111
-2--------2116---372111
-3---------12---121111
---------------5066543
Gambar 1: Lokasi Pengambilan Sampel (A: Cagar Alam Bukit Barisan, B: Cagar Alam Rimbo Panti dan C:Cagar Alam Pangean).
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 30
Tabel 1. Jumlah individu semut pada tiap jenis tumbuhan Macaranga yang dikoleksi dengan metode FreeCollection di beberapa Cagar Alam Sumatera Barat (BB: Cagar Alam Bukit Barisan, RP: CagarAlam Rimbo Panti, P: Cagar Alam Pangean; 1: Macaranga bancana, 2: Macaranga depressa, 3:Macaranga gigantea).
SubFamili Lokasi TotalTribe
JenisFormicinae
CamponotiniPolyrhachis (Myrma) proxima
LasiiniNylanderia sp. 1 of HHParaparatrechina sp. 2 of HH
MyrmicinaeCrematogastrini
Crematogaster borneensisCrematogaster modiglani
PonerinaePonerini
Pachycondyla (Brachyponera) sp. 28 of SKYTotal IndividuTotal JenisTotal GenusTotal TribeTotal Subfamili
BB1--1--1--376----3783332
-3----28---11--1314333
RP1--2-----5----72222
P1--------1130---412111
-2--------2116---372111
-3---------12---121111
---------------5066543
Gambar 1: Lokasi Pengambilan Sampel (A: Cagar Alam Bukit Barisan, B: Cagar Alam Rimbo Panti dan C:Cagar Alam Pangean).
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 30
Tabel 1. Jumlah individu semut pada tiap jenis tumbuhan Macaranga yang dikoleksi dengan metode FreeCollection di beberapa Cagar Alam Sumatera Barat (BB: Cagar Alam Bukit Barisan, RP: CagarAlam Rimbo Panti, P: Cagar Alam Pangean; 1: Macaranga bancana, 2: Macaranga depressa, 3:Macaranga gigantea).
SubFamili Lokasi TotalTribe
JenisFormicinae
CamponotiniPolyrhachis (Myrma) proxima
LasiiniNylanderia sp. 1 of HHParaparatrechina sp. 2 of HH
MyrmicinaeCrematogastrini
Crematogaster borneensisCrematogaster modiglani
PonerinaePonerini
Pachycondyla (Brachyponera) sp. 28 of SKYTotal IndividuTotal JenisTotal GenusTotal TribeTotal Subfamili
BB1--1--1--376----3783332
-3----28---11--1314333
RP1--2-----5----72222
P1--------1130---412111
-2--------2116---372111
-3---------12---121111
---------------5066543
Gambar 1: Lokasi Pengambilan Sampel (A: Cagar Alam Bukit Barisan, B: Cagar Alam Rimbo Panti dan C:Cagar Alam Pangean).

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 31
Macaranga gigantea (Reichb dan Zoll).Subfamili Formicinae memiliki jenis yangpaling banyak yaitu terdiri dari tiga genera dantiga jenis dan subfamili Myrmicinae memilikijenis yang paling banyak jumlah individunyayaitu terdiri dari dua genera dan dua jenis.Subfamili yang paling sedikit adalah Ponerinaeyang hanya terdiri dari satu jenis (Tabel 1 danGambar 2). Pada penelitian ini SubfamiliFormicinae memiliki jumlah jenis yang palingbanyak. Subfamili Formicinae membuat sarangpada Macaranga dengan cara merekatkan daun.Hasil ini sesuai dengan yang dilaporkan olehJohnson et al., (2003) yang menyatakan bahwaSubfamili Formicinae adalah subfamili yangmembuat sarang dengan merekatkan dedauananpada pohon. Kebanyakan dari subfamili inibersifat arboreal.Pada penelitian ini ditemukan empat tribe.Tribe Crematogastrini dan Lasiini adalah tribeyang paling banyak ditemukan jenisnya,sedangkan untuk jumlah individu tribeCrematogastrini memiliki jumlah individusemut yang lebih banyak. Tribe yang palingsedikit ditemukan jenisnya adalah tribePonerini. Genus yang paling banyak didapatkanpada penelitian ini adlaah genus Crematogasteryang berjumlah dua jenis. Sedangkan genusyang paling sedikit ditemukan adalahPolyrhachis, Nylanderia, Paraparatrechina, danPachycondyla yang masing-masingnya hanyaditemukan satu jenis (Tabel 1 dan Gambar 2).
Penelitian ini menggunakan dua metodeyaitu metode Free Collection dan ColonyCollection. Jenis semut Polyrhachis (Myrma)proxima (Gambar 3.A), Nylanderia sp. 1 of HH(Gambar 3.B), Paraparatrechina sp. 2 of HH(Gambar 3.C) dan Pachycondyla(Brachyponera) sp. 28 of SKY (Gambar 3.F)adalah jenis semut yang ditemukan denganmenggunakan metode Free Collection.Sedangkan jenis semut Crematogasterborneensis (Gambar 3.D) dan Crematogastermodiglani (Gambar 3.E) dikoleksi denganmenggunakan metode Colony Collection. Hasilyang didapatkan telah dapat menjelaskan
bahwa ada interaksi antara semut denganMacaranga. Hasil penelitian ini didukung olehPutri, dkk (2013) yang menyatakan bahwametode direct colection/free collectionmerupakan metode yang lebih efektif dalammengoleksi koloni semut pada tumbuhanMacaranga.Kehadiran koloni semut dari genusCrematogaster (Gambar 4.D) juga didapatkanpada penelitian ini, dimana semutCrematogaster ditemukan sebanyak 473individu semut. Seperti yang juga dilaporkanoleh Feldhaar, Fiala, Hashimand danMaschwitz (2002) dan Feldhaar (2002) diPeninsular Malaysia serta Putri, dkk (2013) diHPPB Sumatera Barat, dimana merekamelaporkan bahwa genus Crematogastermerupakan semut yang berasosiasi secaralangsung dengan Macaranga. Beberapapenelitian di Peninsular Malaysia melaporkanbahwa genus Crematogaster merupakan semutyang berasosiasi dengan tumbuhanmyrmecophytic dan hidup berkoloni sertabersarang pada tumbuhan Macaranga. Semuajenis semut Crematogaster yang didapatkanpada penelitian ini terlihat bersarang padatumbuhan Macaranga spp.Crematogaster merupakan genus yang palingbanyak ditemukan pada ruas tumbuhanMacaranga. Seperti yang dilaporkan oleh Fialadan Maschwitz (1990) bahwa semut ini beradadi dalam ruas tumbuhan yaitu terletak di bagianpembuluh floem, dimana semut akanmemperoleh air dan garam mineral yangdihasilkan oleh tumbuhan ketika cairan tersebutmelewati kambium pembuluh. Pembelahan selkambium vaskular ke arah dalam membentukkulit kayu. Aktivitas tersebut mengakibatkanbagian kayu lebih besar daripada kulit kayu.Hal inilah yang menyebabkan pada ruas batangterdapat ruang kosong yang dapat dijadikansarang oleh semut.Jenis semut yang paling banyak ditemukanpada tumbuhan Macaranga yaitu padaMacaranga gigantea dan Macaranga bancanayaitu sebanyak empat jenis (Nylanderia sp. 1 of

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 32
HH, Crematogaster borneensis, Crematogastermodiglani dan Pachycondyla (Brachyponera)sp. 28 of SKY) yang ditemukan padaMacaranga gigantea sedangkan padaMacaranga bancana ditemukan empat jenisyaitu Polyrhachis (Myrma) proxima,Paraparatrechina sp. 2 of HH, Crematogasterborneensis dan Crematogaster modiglani. Padajenis Macaranga depressa hanya ditemukandua jenis yaitu Crematogaster borneensis danCrematogaster modiglani (Tabel 1 dan Gambar2). Perbedaan jenis semut yang didapatkantidak dipengaruhi oleh jumlah sampel pohonpada tiap jenis Macaranga dan diperkirakandipengaruhi oleh perbedaan morfologi dananatomi masing-masing Macaranga (tinggipohon dan diameter batang).
Jumlah individu semut yang paling banyakdidapatkan adalah pada jenis Macarangabancana dibandingkan dengan jenisMacaranga yang lainnya, yaitu 426 individu(Crematogaster borneensis 392 individu danCrematogaster modiglani 30 individu),Polyrhachis (Myrma) proxima ditemukan tigaindividu dan Paraparatrechina sp. 2 of HHhanya ditemukan satu individu (Tabel 1 danGambar 2).
Berdasarkan hasil yang didapatkandiperkirakan bahwa jenis Macaranga bancanaadalah tumbuhan yang berasosiasi dengansemut. Hal ini terlihat dengan banyaknyajumlah jenis dan individu semut yangditemukan pada tumbuhan ini. Penelitian lainyang dilakukan oleh Maschwitz et al., (2004) diMalaysia dan Thailand melaporkan bahwatumbuhan Macaranga bancana adalah jenistumbuhan yang dikunjungi semut, dimana jenisCrematogaster (Decacrema) bersarang didalam ruas batang. Macaranga bancana(Gambar 4.A) memiliki ciri-ciri yaitu pohondengan tinggi mencapai 16 m dan 17 cm dbh,ranting berongga, stipula merah dan membulat.Daun dengan tiga lobus dan bagian bawah daunberwarna hijau muda. Warna kulit batangnyapucat-coklat sampai keputihan, berbintik-bintik.(Whitmore, 1972). Dengan ciri-ciri yang
dimiliki Macaranga bancana tersebut makadiperkirakan semut dapat hidup berkoloni danmenjadikan batang dari tumbuhan M. bancanasebagai tempat bersarangnya. Hal ini terlihatdengan didapatkannya jumlah individu semutsebanyak 426 individu.
Pada penelitian ini ditemukan jenisMacaranga depressa yang berasosiasi dengansemut (myrmecophytes). Hasil ini terlihatdengan ditemukannya koloni dari jenisCrematogaster borneensis dan Crematogastermodiglani. Federle et al., (1997) melaporkanbahwa genus Crematogaster merupakan jenissemut yang bersarang di dalam ruas batang dariMacaranga depressa. M. depressa (Gambar4.B) ini memiliki ciri-ciri yaitu pohon dengantinggi 9 m dan 11 cm dbh, daun dengan tipepeltatus dan memiliki tiga lobus. JenisMacaranga ini dapat ditemukan pada daerahsemak, hutan sekunder dan daerah pinggi-pinggir hutan.
Macaranga gigantea adalah jenis tumbuhanyang tidak bersimbiosis dengan semut (non-myrmecophytes) karena batangnya tidakberongga sehingga semut tidak dapatmembangun sarang di dalam batang atauranting pohon. Berdasarkan penelitianKartasujana dan Martawijaya (1983) jenisMacaranga gigantea (Gambar 4.C) merupakantumbuhan yang tingginya dapat mencapai 25 mdan diameter 55 cm. Batang lurus, bulat, tidakberongga, berkulit halus dengan warna coklatmuda abu-abu. Tajuk agak melebar dan tidakseberapa lebat. Daun tunggal berbentuk bulattelur yang melebar dan bercagap dalam tiga.Permukaan bawah daun putih, berbuku halusdengan urat daun menjari. Dengan ciri-ciriyang dimiliki Macaranga gigantea tersebutmaka diperkirakan semut hanya dapat mencarimakanan pada pohon tersebut.
Beberapa genera yang mengindikasikansuatu kawasan telah terganggu oleh aktifitasmanusia ditemukan pada penelitian inidiantaranya adalah Paraparatrechina danNylanderia. Genera ini ditemukan pada lokasi

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 33
Gambar 2. Jumlah tribe, genera dan jenis yang ditemukan pada tiap subfamili semut padaMacaranga spp.
Gambar 3. Gambar sisi lateral semut pada Macaranga spp. di Cagar Alam Bukit Barisan, RimboPanti dan Pangean, Sumatera Barat. Jenis semut A: Polyrhachis (Myrma) proxima, B:Nylanderia sp. 1 of HH, C: Paraparatrechina sp. 2 of HH, D: Crematogasterborneensis, E: Crematogaster modiglani, dan F: Pachycondyla (Brachyponera) sp. 28of SKY.
00.511.522.533.5
Formicinae
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 33
Gambar 2. Jumlah tribe, genera dan jenis yang ditemukan pada tiap subfamili semut padaMacaranga spp.
Gambar 3. Gambar sisi lateral semut pada Macaranga spp. di Cagar Alam Bukit Barisan, RimboPanti dan Pangean, Sumatera Barat. Jenis semut A: Polyrhachis (Myrma) proxima, B:Nylanderia sp. 1 of HH, C: Paraparatrechina sp. 2 of HH, D: Crematogasterborneensis, E: Crematogaster modiglani, dan F: Pachycondyla (Brachyponera) sp. 28of SKY.
Formicinae Myrmicinae Ponerinae
TribeGeneraJenis
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 33
Gambar 2. Jumlah tribe, genera dan jenis yang ditemukan pada tiap subfamili semut padaMacaranga spp.
Gambar 3. Gambar sisi lateral semut pada Macaranga spp. di Cagar Alam Bukit Barisan, RimboPanti dan Pangean, Sumatera Barat. Jenis semut A: Polyrhachis (Myrma) proxima, B:Nylanderia sp. 1 of HH, C: Paraparatrechina sp. 2 of HH, D: Crematogasterborneensis, E: Crematogaster modiglani, dan F: Pachycondyla (Brachyponera) sp. 28of SKY.

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 34
Gambar 4. Gambar Jenis-Jenis Macaranga yang dikunjungi semut di Cagar Alam Bukit Barisan,Rimbo Panti dan Pangean, Sumatera Barat (A: Macaranga bancana, B: Macarangadepressa, C: Macaranga gigantea, D: Koloni semut pada ruas batang Macaranga)
Cagar Alam Bukit Barisan, dimana terlihatadanya kerusakan hutan yang ditandai denganbanyaknya penebangan hutan secara liar dilokasi ini. Semut biasanya dapat dijadikansebagai spesies indikator. Keberadaan jenisvegetasi lain yang berada di sekitar tumbuhanMacaranga diperkirakan juga dapat mem-pengaruhi jenis-jenis semut yang didapatkanpada saat pengambilan sampel misalnyaPolyrhachis (Myrma) proxima, dimana jenissemut ini tidak ditemukan bersarang padatumbuhan Macaranga. Kehadiran jenis semuttersebut diperkirakan untuk proses mencarimakanan (foraging). Pada penelitian ini jugaditemukan salah satu jenis semut yang bersifatscavenger yaitu Pachycondyla (Brachyponera)sp. 28 of SKY. Dejean (1991) melaporkanbahwa genus Pachycondyla merupakan semutyang umumnya bersifat predator dan scavenger.
KESIMPULANDari penelitian mengenai jenis semut(Hymenoptera: Formicidae) pada Macarangaspp. di Cagar Alam Bukit Barisan, Cagar AlamRimbo Panti dan Cagar Alam Pangean denganmenggunakan metode Free Collection dan
Colony Collection didapatkan enam jenis semutyang tergolong ke dalam lima genera, empatribe, tiga subfamili semut pada tiga jenisMacaranga spp. Jenis semut yang palingbanyak ditemukan pada Macaranga bancanadan Macaranga gigantea (masing-masingnyaempat jenis).
UCAPAN TERIMA KASIHUcapan terima kasih kepada Munetoshi Maruyama danKomatsu Takashi (Kyusu University, Japan) atasbantuan dalam pengidentifikasian sampel Macaranga.Terima kasih pada Dirjen Pendidikan Tinggi ataspendanaan bagi penelitian ini melalui HibahKerjasama Luar Negeri dan Publikasi Internasionaldengan ketua Dr. Henny Herwina.
DAFTAR PUSTAKAAgosti, D., Majer, J. D., Alonso, L. E. dan
Schultz, T.R. 2000. Ants StandardMethods For Measuring and MonitoringBiodiversity. Smithonian Institutio Press.Washington, U. S. A.
Bolton, B. 1994. Identification Guide to theAnt Genera of the World. HardvardUniversity Press. London.
Borror, J.D., Triplehorn, A.C. dan Johnson, F.N.1992. Pengenalan Pengajaran Serangga.Diterjemahkan oleh Partosoedjono, S dan
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 34
Gambar 4. Gambar Jenis-Jenis Macaranga yang dikunjungi semut di Cagar Alam Bukit Barisan,Rimbo Panti dan Pangean, Sumatera Barat (A: Macaranga bancana, B: Macarangadepressa, C: Macaranga gigantea, D: Koloni semut pada ruas batang Macaranga)
Cagar Alam Bukit Barisan, dimana terlihatadanya kerusakan hutan yang ditandai denganbanyaknya penebangan hutan secara liar dilokasi ini. Semut biasanya dapat dijadikansebagai spesies indikator. Keberadaan jenisvegetasi lain yang berada di sekitar tumbuhanMacaranga diperkirakan juga dapat mem-pengaruhi jenis-jenis semut yang didapatkanpada saat pengambilan sampel misalnyaPolyrhachis (Myrma) proxima, dimana jenissemut ini tidak ditemukan bersarang padatumbuhan Macaranga. Kehadiran jenis semuttersebut diperkirakan untuk proses mencarimakanan (foraging). Pada penelitian ini jugaditemukan salah satu jenis semut yang bersifatscavenger yaitu Pachycondyla (Brachyponera)sp. 28 of SKY. Dejean (1991) melaporkanbahwa genus Pachycondyla merupakan semutyang umumnya bersifat predator dan scavenger.
KESIMPULANDari penelitian mengenai jenis semut(Hymenoptera: Formicidae) pada Macarangaspp. di Cagar Alam Bukit Barisan, Cagar AlamRimbo Panti dan Cagar Alam Pangean denganmenggunakan metode Free Collection dan
Colony Collection didapatkan enam jenis semutyang tergolong ke dalam lima genera, empatribe, tiga subfamili semut pada tiga jenisMacaranga spp. Jenis semut yang palingbanyak ditemukan pada Macaranga bancanadan Macaranga gigantea (masing-masingnyaempat jenis).
UCAPAN TERIMA KASIHUcapan terima kasih kepada Munetoshi Maruyama danKomatsu Takashi (Kyusu University, Japan) atasbantuan dalam pengidentifikasian sampel Macaranga.Terima kasih pada Dirjen Pendidikan Tinggi ataspendanaan bagi penelitian ini melalui HibahKerjasama Luar Negeri dan Publikasi Internasionaldengan ketua Dr. Henny Herwina.
DAFTAR PUSTAKAAgosti, D., Majer, J. D., Alonso, L. E. dan
Schultz, T.R. 2000. Ants StandardMethods For Measuring and MonitoringBiodiversity. Smithonian Institutio Press.Washington, U. S. A.
Bolton, B. 1994. Identification Guide to theAnt Genera of the World. HardvardUniversity Press. London.
Borror, J.D., Triplehorn, A.C. dan Johnson, F.N.1992. Pengenalan Pengajaran Serangga.Diterjemahkan oleh Partosoedjono, S dan
Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 34
Gambar 4. Gambar Jenis-Jenis Macaranga yang dikunjungi semut di Cagar Alam Bukit Barisan,Rimbo Panti dan Pangean, Sumatera Barat (A: Macaranga bancana, B: Macarangadepressa, C: Macaranga gigantea, D: Koloni semut pada ruas batang Macaranga)
Cagar Alam Bukit Barisan, dimana terlihatadanya kerusakan hutan yang ditandai denganbanyaknya penebangan hutan secara liar dilokasi ini. Semut biasanya dapat dijadikansebagai spesies indikator. Keberadaan jenisvegetasi lain yang berada di sekitar tumbuhanMacaranga diperkirakan juga dapat mem-pengaruhi jenis-jenis semut yang didapatkanpada saat pengambilan sampel misalnyaPolyrhachis (Myrma) proxima, dimana jenissemut ini tidak ditemukan bersarang padatumbuhan Macaranga. Kehadiran jenis semuttersebut diperkirakan untuk proses mencarimakanan (foraging). Pada penelitian ini jugaditemukan salah satu jenis semut yang bersifatscavenger yaitu Pachycondyla (Brachyponera)sp. 28 of SKY. Dejean (1991) melaporkanbahwa genus Pachycondyla merupakan semutyang umumnya bersifat predator dan scavenger.
KESIMPULANDari penelitian mengenai jenis semut(Hymenoptera: Formicidae) pada Macarangaspp. di Cagar Alam Bukit Barisan, Cagar AlamRimbo Panti dan Cagar Alam Pangean denganmenggunakan metode Free Collection dan
Colony Collection didapatkan enam jenis semutyang tergolong ke dalam lima genera, empatribe, tiga subfamili semut pada tiga jenisMacaranga spp. Jenis semut yang palingbanyak ditemukan pada Macaranga bancanadan Macaranga gigantea (masing-masingnyaempat jenis).
UCAPAN TERIMA KASIHUcapan terima kasih kepada Munetoshi Maruyama danKomatsu Takashi (Kyusu University, Japan) atasbantuan dalam pengidentifikasian sampel Macaranga.Terima kasih pada Dirjen Pendidikan Tinggi ataspendanaan bagi penelitian ini melalui HibahKerjasama Luar Negeri dan Publikasi Internasionaldengan ketua Dr. Henny Herwina.
DAFTAR PUSTAKAAgosti, D., Majer, J. D., Alonso, L. E. dan
Schultz, T.R. 2000. Ants StandardMethods For Measuring and MonitoringBiodiversity. Smithonian Institutio Press.Washington, U. S. A.
Bolton, B. 1994. Identification Guide to theAnt Genera of the World. HardvardUniversity Press. London.
Borror, J.D., Triplehorn, A.C. dan Johnson, F.N.1992. Pengenalan Pengajaran Serangga.Diterjemahkan oleh Partosoedjono, S dan

Diyona Putri1, Henny Herwina, Rijal Satria dan Alan Handru 35
Djarubito, M.B. Edisi Keenam. GadjahMada University Press. Yogyakarta.
Borror, J.D., Triplehorn, A.C. and Johnson, F.N.2005. Borror and DeLong's Introduction tothe Study of Insects. 7th Edition.Thompson Brooks/Cole. Belmont,California. 864 pp.
Corner, E.J.H. 1988. Wayside trees of Malaya1. Kuala Lumpur. Malayan Nature Society,296–304.
Dejean, A. 1991. Le compertement predateurde Pachycondyla soror. Entomol, 58: 123-135.
Federle. W., Maschwitz, U., Fiala, B., Riederer,M. and Hölldöbler, B. 1997. Slippery ant-plants and skilful climbers: selection andprotection of specific ant partners byepicuticular wax blooms in Macaranga(Euphrbiaceae). Oecologia, 112: 217-224.
Federle, H., Fiala, B., Hashim Rosli B. andMaschwitz, U. 2002. Patterns of theCrematogaster-Macaranga association:The ant partner makes the difference.Insectes Soc, 50: 9–19.
Feldhaar, H. 2002. Structuring mechanisms ofthe Crematogaster-Macaranga ant-plantassociation: A combined ecological andphylogenetic approach. Dissertation at theFaculty of Biology Johann WolfgangGoethe-University Frankfurt. Germany.
Fiala, B. 1992. Domatia as most importantadaptations in the evolution ofmyrmecophytes in the paleotropical treegenus Macaranga (Euphorbiaceae). PlantSystematics and Evolution, 180: 53-64.
Fiala, B. and Maschwitz, U. 1990. Studies onthe South East Asian ant-plant associationCrematogaster borneensis/Macaranga:adaptations of the ant partner. InsectesSociaux, 37: 212-231.
Fiala, B. and Maschwitz, U. 1991. Food bodiesand tbeir significance for obligate ant-association in the tree genus Macaranga(Euphorbiaceae). Botanical Journal of theUnnean Soriel. 110: 61 – 75.
Fiala, B. and Maschwitz, U. 1992a. Domatia asmost important preadaptions in theevolution of myrmecophytes in apaleotropical tree genus. Plant Syst. Evol,180: 53-64.
Fiala, B. and U. Maschwitz. 1992b. Foodbodies in the genus Macaranga and theirsignificance for the evolution ofmyrmecophytism. Bot. J. Linn. Soc, 110:61-75.
Holldobler, B. and Wilson, O.E. 1990. TheAnts. Harvard University Press.Cambridge. U. S. A.
Ito, F., Yamane, S., Eguchi, K., Noerdjito,W.A., Kahono, S., Tsuji, K., Ohkawa, K.,Yamauchi, K. and Nishida, T. 2001. AntsSpecies Diversity in the Bogor BotanicalGarden, West Java, Indonesia, withDescription of Two New Species of theGenus Leptanilla (Hymenoptera:Formicidae). Tropics, 10 (3): 379-404.
Johnson, R.N., Agapow, P.M. and Crozier, R.H.2003. A tree island approach to inferringphylogeny in the ant subfamilyFormicinae, with especial reference to theevolution of weaving. Mol. Phylog, 29:317-330.
Maschwitz, U., Fiala, B., Dumpert, K. 2004.An Unusal Myrmecophytic MacarangaAssociation, Occuring in a Disjunct Areain The Mosoon Zone of South-East Asia:Phenology and The Description of A NewAnt Species. Ecotropica, 10: 33-49.
Ong, S.L. 1978. The ant-association in M.triloba. Ph. D. Thesis. Kuala Lumpur.
Putri, D., Herwina, H., Salmah, S. 2013. Jenis-Jenis Semut (Hymenoptera: Formicidae)pada Tumbuhan Macaranga spp. di HutanPendidikan dan Penelitian (HPPB)Universitas Andalas. pp. 217-222.Prosseding Semirata FMIPA UniversitasLampung.
Whitmore, T.C. 1972. Euphorbiaceae. In:Whitmore, T.C.(ed.). Tree Flora ofMalaya 2. Longman, Kuala Lumpur &London.

ISSN 978-602-14989-0-3 BioETI
Pengaruh Fungi Mikoriza Arbuskula (FMA) pada tanamanCentrosema pubescens (Benth.) untuk bioremediasi lahantercemar MerkuriDWI ANINDITYA SIREGAR , ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIA
Program Studi, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRACTThe research study the effect of Arbuscular Mycorrhiza Fungi (AMF) on the growth of plants Centrosema pubescens(Benth.) as phtyoremediation agent of mine contaminated by mercury (Hg). The research has been done from Decemberto February 2014 at the net house and the Laboratory of Plant Physiology and tissue culture, Department of Biology,Faculty of Mathematics and Natural Sciences Andalas University. As the treatment this research used C. pubescens(Benth.) inoculated with mycorrhiza and without mycorrhiza in soil contaminated of mercury. The results showed that C.pubescens was a potential plant as phtyoremediation agent (Hg) indicated by decreasing of mercury (Hg). C. pubescensinoculated with mycorrhiza have accumulated 0,05 ppm of mercury in shoot and 0,17 ppm in root, while withoutmycorrhiza accumulated 0.04 ppm of mercury in shoot and 0,10 ppm in root. Arbuscular Mycorrhiza Fungi (AMF)influenced on plants height, wet and dry weight of leaf, wet and dry weight of root during 8 weeks of observation.
Key words: Arbuscular Mycorrhiza Fungi (AMF), phtyoremediation, mercury, Centrosema pubescens
Pendahuluan
Kegiatan penambangan emas terutama yangdilakukan oleh pertambangan emas tanpa izin(PETI) umumnya menimbulkan kerusakanlingkungan. Salah satu bentuk kerusakan yangditimbulkan adalah pencemaran merkuri dariproses pengolahan emas secara amalgamasi.Pembuangan limbah berupa lumpurmengandung merkuri (Hg) dan berbagai logamberat lainnya biasanya dilakukan di lahansekitar lokasi penambangan sehingga men-cemari lahan pertanian yang berada di sekitarlokasi tersebut (Handayanto, 2012). Bentukkerusakan lainnya dari PETI adalah konta-minasi lahan, pemadatan tanah, berkurangnyaunsur hara, pH rendah, serta pencemaranlogam-logam berat pada lahan bekas tambang(Setyaningsih, 2007).
Salah satu upaya yang dapat dilakukanuntuk meremediasi lahan pertambangan emasyaitu dengan melakukan fitoremediasi untukmemperbaiki kerusakan. Fitoremediasi (phyto-remediation) merupakan suatu sistem di manatanaman tertentu, secara sendiri atau bekerjasama dengan mikroorganisme dalam media
tanam, dapat mengubah zat kontaminanmenjadi kurang atau tidak berbahaya (Benison,2004).
Dari penelitian diketahui beberapa tanamanberpotensi sebagai tanaman fitoremediasi,diantaranya adalah Centrosema pubescensmampu tumbuh dengan baik di media tailingtambang emas (Syarif, 2009). dan mampumemproduksi biomassa tinggi pada limbahyang terkontaminasi logam berat (Hidayati,Syarif, dan Juhaeti, 2005).
Untuk menunjang keberhasilan, fito-remediasi juga membutuhkan partum-buhantanaman yang cepat yang diikuti oleh perbaikansifat kimia tanah yang dapat menghasilkanproduktivitas tanaman yang baik. Oleh karenaitu dalam penelitian ini digunakan pupuk hayatiyaitu fungi mikoriza arbuskula (FMA). FMAmemiliki peranan yang sangat penting untukmelindungi tanaman dari serangan patogen, dankondisi tanah dan lingkungan yang kurangkondusif seperti : pH rendah, stress air,temperatur ekstrim, salinitas yang tinggi, dantercemar logam berat (Turjaman, 2005).
Hasil penelitian memperlihatkan bahwatanaman sengon yang diinokulasi mikoriza

Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 37
mampu menurunkan kandungan Pb, Zn dan Cudalam medium limbah lumpur minyak hasilekstraksi (Rossiana, 2003). Pemberian FMAmampu meningkatkan bobot kering Puerariajavanica dibandingkan tanpa pemberian FMApada lahan bekas tambang timah (Maulidesta,2005). Sedangkan Pemberian MA+BPP+Rhizobium memberikan hasil pertumbuhanyang baik bagi tanaman C. pubescens yangditanam pada media tailing (Safitri, 2008).
Penelitian ini bertujuan untuk mengetahuipengaruh pemberian Fungi Mikoriza Arbuskula(FMA) terhadap pertumbuhan tanaman C.pubescens sebagai agen fitoremediasi padalahan tercemar merkuri (Hg).
BAHAN DAN METODE
Penelitian ini dilaksanakan dari bulanDesember sampai Februari 2014 di RumahKawat dan dilanjutkan di LaboratoriumFisiologi Tumbuhan Jurusan Biologi, FMIPA,Universitas Andalas Padang, Sumatera Barat.
Penelitian ini menggunakan metodaeksperimen, sebagai perlakuan adalah inokulasimikoriza dan tanpa inokulasi mikoriza padamedia tanah tercemar, dengan 4 kali ulangan.Analisis data disajikan secara deskriptif.
Parameter yang diamati yaitu Tinggitanaman, Berat basah dan berat kering daun,Berat basah dan berat kering akar, sertaKandungan logam berat pada tanaman.
HASIL DAN PEMBAHASAN
Berdasarkan penelitian yang telah dilakukantentang pemberian Fungi Mikoriza Arbuskula(FMA) pada tanaman C. pubescens untukfitoremediasi lahan tercemar merkuri,didapatkan hasil sebagai berikut:Kolonisasi akar tanaman Centrosemapubescens (Benth.) oleh Fungi MikorizaArbuskula (FMA).Dari pengamatan terhadap kolonisasi FMAyang diinokulasikan pada akar tanaman C.pubescens yang ditanam pada media tercemar
merkuri dapat dilihat pada Gambar 1.Terjadinya infeksi FMA pada akar tanamanbiasanya ditandai dengan terbentuknya hifaeksternal, arbuskula dan vesikula (Santosa danAnas, 1992). Vesikula merupakan suatustruktur lonjong atau bulat yang mengandungcairan lemak dan berfungsi sebagai organpenyimpan makanan, sedangkan hifa eksternalmerupakan struktur dari FMA yangberkembang diluar akar yang berfungsi sebagaipenyerap hara dan air dalam tanah (Dewi,2007).Pertumbuhan tanaman.Hasil pengamatan terhadap parameterpertumbuhan tanaman Centrosema pubescens(Benth.) selama delapan minggu pengamatandidapatkan hasil seperti pada Tabel 1. Tabel 1menunjukkan bahwa pemberian mikorizamampu meningkatkan pertumbuhan tanaman,seperti tinggi tanaman, berat basah dan keringtanaman, berat basah dan berat kering akardibandingkan dengan tanaman tanpa inokulasimikoriza. Inokulasi FMA dapat meningkatkantinggi tanaman di bandingkan denganperlakuan tanpa inokulasi mikoriza.
Hal ini dikarenakan mikoriza yangdiinokulasi pada tanaman mampu membantupenyerapan unsur hara dan air sehingga dapatmeningkatkan pertumbuhan vegetatif tanaman.Hal ini sama dengan yang didapatkan olehJannah (2011) bahwa, inokulasi FMA dapatmeningkatkan tinggi tanaman kedelaibandingkan dengan yang tanpa inokulasi sertaIrianto (2009) Pemberian inokulan FMA padabibit jarak pagar dapat meningkatkanpertumbuhan tinggi tanaman.
Untuk melihat perbandingan pertambahantinggi tanaman yang diberi inokulasi mikorizadengan yang tidak diberi inokulasi mikoriza,dapat dilihat pada Gambar 2. Peningkatan rata-rata tinggi tanaman ini berhubungan dengantingginya persentase derajat infeksi mikorizapada akar tanaman (80%). Hifa eksternal padaakar tanaman yang terinfeksi mikoriza dapat
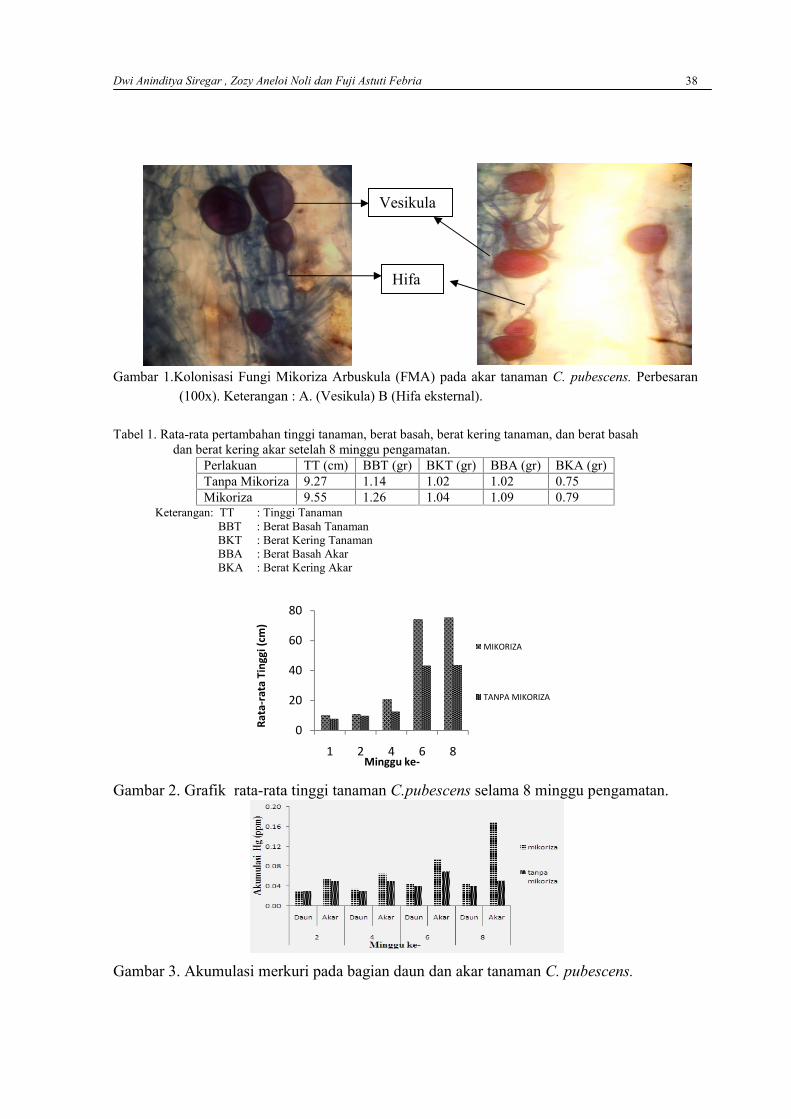
Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 38
Gambar 1.Kolonisasi Fungi Mikoriza Arbuskula (FMA) pada akar tanaman C. pubescens. Perbesaran(100x). Keterangan : A. (Vesikula) B (Hifa eksternal).
Tabel 1. Rata-rata pertambahan tinggi tanaman, berat basah, berat kering tanaman, dan berat basahdan berat kering akar setelah 8 minggu pengamatan.
Perlakuan TT (cm) BBT (gr) BKT (gr) BBA (gr) BKA (gr)Tanpa Mikoriza 9.27 1.14 1.02 1.02 0.75Mikoriza 9.55 1.26 1.04 1.09 0.79
Keterangan: TT : Tinggi TanamanBBT : Berat Basah TanamanBKT : Berat Kering TanamanBBA : Berat Basah AkarBKA : Berat Kering Akar
Gambar 2. Grafik rata-rata tinggi tanaman C.pubescens selama 8 minggu pengamatan.
Gambar 3. Akumulasi merkuri pada bagian daun dan akar tanaman C. pubescens.
20
40
60
80
Rata
-rat
aTi
nggi
(cm
)
Vesikula
Hifa
Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 38
Gambar 1.Kolonisasi Fungi Mikoriza Arbuskula (FMA) pada akar tanaman C. pubescens. Perbesaran(100x). Keterangan : A. (Vesikula) B (Hifa eksternal).
Tabel 1. Rata-rata pertambahan tinggi tanaman, berat basah, berat kering tanaman, dan berat basahdan berat kering akar setelah 8 minggu pengamatan.
Perlakuan TT (cm) BBT (gr) BKT (gr) BBA (gr) BKA (gr)Tanpa Mikoriza 9.27 1.14 1.02 1.02 0.75Mikoriza 9.55 1.26 1.04 1.09 0.79
Keterangan: TT : Tinggi TanamanBBT : Berat Basah TanamanBKT : Berat Kering TanamanBBA : Berat Basah AkarBKA : Berat Kering Akar
Gambar 2. Grafik rata-rata tinggi tanaman C.pubescens selama 8 minggu pengamatan.
Gambar 3. Akumulasi merkuri pada bagian daun dan akar tanaman C. pubescens.
0
20
40
60
80
1 2 4 6 8
MIKORIZA
TANPA MIKORIZA
Minggu ke-
Vesikula
Hifa
Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 38
Gambar 1.Kolonisasi Fungi Mikoriza Arbuskula (FMA) pada akar tanaman C. pubescens. Perbesaran(100x). Keterangan : A. (Vesikula) B (Hifa eksternal).
Tabel 1. Rata-rata pertambahan tinggi tanaman, berat basah, berat kering tanaman, dan berat basahdan berat kering akar setelah 8 minggu pengamatan.
Perlakuan TT (cm) BBT (gr) BKT (gr) BBA (gr) BKA (gr)Tanpa Mikoriza 9.27 1.14 1.02 1.02 0.75Mikoriza 9.55 1.26 1.04 1.09 0.79
Keterangan: TT : Tinggi TanamanBBT : Berat Basah TanamanBKT : Berat Kering TanamanBBA : Berat Basah AkarBKA : Berat Kering Akar
Gambar 2. Grafik rata-rata tinggi tanaman C.pubescens selama 8 minggu pengamatan.
Gambar 3. Akumulasi merkuri pada bagian daun dan akar tanaman C. pubescens.
Vesikula
Hifa

Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 39
mempengaruhi struktur tanah, seperti agregattanah dan ruang pori tanah. Melaluipembentukan agregat tanah akanmemungkinkan terjadinya aerasi yang lebih baikpada kondisi kadar air tanah yang rendah.Dengan semakin besarnya volume tanah yangdijadikan sebagai daerah serapan air tersebut,maka peluang tanamn untuk mengabsorbsiunsur-unsur hara dan air semakin besardibandingkan dengan bibit tanpa mikoriza(Irianto (2009).
Pemberian mikoriza dapat padameningkatkan berat basah tanaman, disebabkankarena ketersediaan air mempengaruhi prosespembentukan organ tanaman seperti daun,batang dan akar. Besarnya serapan air dalamjaringan tanaman akan berpengaruh terhadapberat basah tanaman. Muin (2002) menyatakanbahwa simbiosis tanaman dengan mikorizamampu meningkatkan penyerapan unsur haradan air sehingga meningkatkan laju prosesfotosintesis. Meningkatnya laju fotosintesismengakibatkan pertumbuhan produksi daritanaman tanaman meningkat (Dwidjoseputro,1994).
Pemberian inokulasi mikoriza selainmeningkatkan berat basah atas juga mampumeningkatkan berat kering tanaman, hal inikarena adanya inokulasi mikoriza yangmembantu dalam penyerapan unsur hara dan airakan berdampak pada berat kering tanaman.Ketersediaan air mempengaruhi prosespembentukan organ tanaman seperti daun,batang dan akar. Hasil ini sesuai dengan yangdikemukakan oleh Brundret (1966) bahwapengaruh mikoriza yang paling utama adalahdapat meningkatkan pengambilan unsur fosfatdari tanah dan meningkatkan berat kering.Lukiwati (1996) telah melakukan percobaanpada tanaman Centrosema pubescens danPueraria phaseoloides bermikoriza menunjuk-kan produksi bahan kering yang lebih tinggidiabandingkan tanaman yang tidak bermikoriza.
Dari Tabel 1 juga dapat dilihat bahwaperlakuan inokulasi mikoriza memperlihatkanpengaruh yang nyata terhadap berat basah akar
tanaman. Hal ini dikarenakan Fungi MikorizaArbuskula (FMA) yang bersimbiosis denganakar tanaman inang akan membentuk mikoriza.Mikoriza mampu meningkatkan penyerapanfosfat, air, dan zat hara lainnya danmeningkatkan penyerapan ion yang biasanyaberdifusi menuju akar. Hal ini dimungkinkankarena mikoriza memiliki jaringan hifa eksternalyang luas dan diameter yang lebih kecil daribulu-bulu akar, sehingga mampu memperluasbidang penyerapan air dan garam-garam mineraldi dalam tanah (Salisbury dan Ross, 1995). Halini didukung oleh Abdullah (2005), bahwapemanfaatan mikoriza pada tebu lahan keringmemberi dampak positif terhadap pertumbuhandan produksi tebu, dimana dengan penggunaanmikoriza sistem perakaran tebu akan lebih baik,dibandingkan dengan tebu yang tidakmenggunakan mikoriza.
Akumulasi Logam Berat Merkuri PadaTanamanHasil dari analisis kandungan merkuri padabagian akar dan daun tanaman C pubescensyang diberi inokulasi mikoriza dan yang tidakdiberi inokulasi mikoriza selama 8 minggupengamatan dapat dilihat pada Gambar 3.Gambar 3 menunjukkan bahwa logam berat Hglebih banyak diakumulasi pada akar tanaman.Tingginya kandungan logam berat Hg pada akardikarenakan akar merupakan organ yanglangsung menyerap logam berat dari tanah(Sagita, 2002). Logam berat Hg lebih banyakdiakumulasi oleh akar tanaman yang di-inokulasi mikoriza dibandingkan dengantanaman tanpa inokulasi mikoriza. Hal inidikarenakan akar yang terdapat hifa mikorizaberperan dalam mengakumulasi logam berat.
Pada akar tanaman yang diinokulasi olehmikoriza akan terdapat hifa yang berperandalam mengakumulasi logam berat. Subiska(2002) mengemukakan bahwa mekanismeperlindungan tanaman terhadap logam berat danunsur toksik dapat menggunakan mikoriza. Halini dikarenakan mikoriza memegang peranan

Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 40
penting dalam melindungi akar tanaman dariunsur toksik, diantaranya yaitu logam berat.
Mekanisme perlindungan terhadap logamberat dan unsur toksik oleh mikoriza dapatmelalui efek filtrasi, menonaktifkan secarakimiawi, atau akumulasi unsur tersebut dalamhifa. Tanaman yang diinokulasi mikorizamemiliki kemampuan menekan serapan Hg,karena mikoriza diketahui dapat mengikatlogam tersebut pada gugus karboksil dansenyawa pektak (hemiselulosa) pada matriksantar permukaan kontak mikoriza dan tanamaninang, pada selubung polisakarida dan dindingsel hifa. (Rossiana, N. 2003).
Mukarlina dkk., (2013) melaporkan logamberat Hg lebih banyak diakumulasi pada bagianakar tanaman jagung dibandingkan denganbagian daun dan batang. Hasil yang sama jugadidapatkan oleh Purwani dan Aprilia (2013)penambahan mikoriza pada tanama Euporbiamilii dan tanaman Dahlia pinnata mampumeningkatkan serapan logam berat Pb sertameningkatkan akumulasi logam berat Pb padaakar tanaman dan dibandingkan denganakumulasi pada batang dan daun.
Tanaman C. pubescens mampu menyerapmerkuri karena tanaman ini merupakan tanamanyang mampu beradaptasi dengan baik padamedia tanah tercemar. Hidayati dkk., (2006)mengatakan tanaman ini termasuk salah satujenis yang tumbuh dominan di lingkunganpembuangan limbah pengolahan emas yangtercemar merkuri dan dapat digunakan sebagaitanaman akumulator logam berat khususnyamerkuri. Dari penelitian Juhaeti dkk., (2006)mengatakan bahwa tanaman C. pubescensmemiliki kemampuan menyerap logam berat Hgsebanyak 0.480 ppm pada bagian akarnyasedangkan tanaman C. muconoides hanyamemiliki kemampuan menyerap logam Hgsebanyak 0.293 ppm pada bagian akarnya.Hasil yang sama juga didaptakan oleh Suherlina(2010) yang menyatakaan C. pubescens dapatmenyerap merkuri sebanyak 2.7 ppm padabagian akar,
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Tanaman C. pubescens diinokulasi mikoriza
berpotensi sebagai agen fitoremediasi padalahan tercemar merkuri dan mampumengakumulasi merkuri sebesar 0,05 ppmpada daun dan 0,17 ppm pada akar.
2. Pemberian Fungi Mikoriza Arbuskula(FMA) pada tanaman berpengaruh padapertambahan tinggi tanaman, berat basah dankering daun serta berat basah dan kering akar.
DAFTAR PUSTAKA
Abdullah, S,Y. 2005. Perbanyakan CendawanMikoriza Arbuskula (CMA) pada berbagaivarietas jagung (Zea mays L.) danPemanfaatannya pada dua varietas tebu(Saccharum officinarum L.). Jurnal Sains& Teknologi. 5(1): 12-20.
Benison,CG, Lello DP, Shokes EJ, Cosper JNdan Omichinski Gj. 2004. A stablemercuty-containing complex of theOrganon-Mercuryyase MerB: Catalysis,product release and direct tranfer to MerA.Biochemistry. Vol.43 : 8333-8345.
Brundrett, M. C., L. Melvillle and L. Peterson.1994. Practical Methods in MycorrhizaResearch. Mycology Publications. OntarioCanada. 161 pp.
Dewi, R.I.2007. Peran, Prospek dan KendalaDalam Pemanfaatan Endomikoriza.Makalah. Fakultas Pertanian. UniversitasPadjadjaran.
Dwidjoseputro,D. 1994. Pengantar FisiologiTumbuhan. PT. Gramedia Pustaka Utama.Jakarta.
Handayanto, 2012). Fitoremediasi TanahTercemar Merkuri Limbah Tambang EmasRakyat untuk Perbaikan Produksi Jagung.LPPM Universitas Brawijaya.
Hidayati N, Syarief F, Juhaeti T. 2005. PotensiCentrocema pubescence, Calopogoniummucunoides, dan Micania cordata dalamMembersihkan Logam Kontaminan padaLimbah Penambangan Emas. JurnalBiodiversitas. Nomor 1 Januari 2006Halaman: 4-6.
Hidayati N, Syarief F, Juhaeti T. 2006. PotensiTumbuhan Liar dari Lokasi Penampungan

Dwi Aninditya Siregar , Zozy Aneloi Noli dan Fuji Astuti Febria 41
Limbah Tailing PT ANTAM CikotokUntuk Fitoremediasi Lahan TerdegradasiSianida. Jurnal Teknologi Lingkungan.Pusat Teknologi Lingkungan. DeputiBidang Teknologi PengembanganSumberdaya Alam BPPT.
Irianto, S, B. 2005. Pengaruh Inokulasi FungiMikoriza Arbuskula TerhadapPertumbuhan Bibit Jarak Pagar DiPesemaian. Jurnal penelitian dankonservasi alam. vol VI.No 2 :195-201.
Jannah, H. 2011. Respon Tanaman KedelaiTerhadap Asosiasi Fungi MikorizaArbuskular Di lahan Kering. Jurnal GaneSwara Vol. 5 No.2 September 2011
Juhaeti T, Syarif F, Hidayati N. 2006.Inventarisasi tumbuhan potensial untukfitoremediasi lahan dan air terdegradasipenambangan emas. Jurnal Biodiversitas.
Lukiwati, D,R, 1996. Pemingkatan Produksi danNilai Nutrisi Legume Pakan denganpemupukan batuan fosfat dan inokulasimikoriza vesicular-arbuskular [Disertasi].Bogor. Program Pascasarjana. InstitutPertanian Bogor.
Maulidesta, N. 2005. Efek pemberian mikorizadan pembenah tanah terhadap produksileguminosa pada media tailing liat daripasca penambangan timah. [Skripsi].Fakultas Peternakan. IPB. Bogor.
Muin, A, 2002, Uji Efektivitas CendawanMikoriza Arbuskula lokal Hasil Isolasi dariLokasi Hutan Tanaman Industri PTInhutani 111 di Sanggau Kalimantan Barat.Kalimantan Barat
Mukarlina, Linda, R, Leskona, D. 2013.Pertumbuhan Jagung (Zea mays L).Dengan Pemberian Glomus aggregatumdan Biofertilizer pada Tanah BekasPertambahan Emas. Jurnal Protobiont. vol2 (3): 176-180.
Purwani, I, K dan Aprilia, D. 2013. PengaruhPemberian Mikoriza Glomus fasciculatumTerhadap Akumulasi Logam Timbal (Pb)Pada Tanaman Euphorbia milii. JurnalSains Dan Seni Pomits Vol.
Rossiana, N. 2003. Penurunan KandunganLogam Berat Dan Pertumbuhan TanamanSengon (Paraserianthes falcataria L(Nielsen) Bermikoriza Dalam MediumLimbah Lumpur Minyak Hasil Ekstraksi.Laboratorium Mikrobiologi dan BiologiLingkungan Jurusan Biologi FakultasMatematika dan Ilmu Pengetahuan AlamUniversitas Padjadjaran.
Safitri, R. 2008). Pemberian mikroorganismedan asam humik pada tanah latosol dantailing untuk memperbaiki pertumbuhandan produksi Centrosema pubescens Benth.[skripsi] Program Studi Ilmu Nutrisi DanMakanan Ternak.Fakultas Peternakan.Institut Pertanian Bogor.
Sagita, W.A. 2002. Uji Kemampuan AkumulasiLogam Kadmium dar Media oleh RumputGagajahan (Panicum maximum Jacq)[Skripsi ] S1 Biologi ITB.
Santosa, D. A. dan I, Anas.1992. Mikorizavesikular asbuskular dalam S. Harran danN. Ansori. Bioteknologi Pertanian 2. PusatAntar Universitas Pangan dan Gizi-IPB.Bogor. Hal: 285-327.
Salisbury, F. B. dan Ross, C. W. 1995. FisiologiTumbuhan Jilid 3. ITB. Bandung.
Suherlina, T. 2010. Potensi AmmoniumTiosulfat Dalam Meningkatkan SerapanMerkuri Pada Tanaman Sentro(Centrosema pubescens Benth.) SebagaiAgen Fitoremediasi. [Skripsi] JurusanBiologi. Intitut Teknologi Sepuluh Maret
Suharno dan Sancayaningsih, R.2013. PotensiTeknologi Mokorizoremediasi LogamBerat Dalam Rehabilitasi Lahan Tambang.Bioteknologi 10 (1): 31-42. ISSN: 0216-6887.
Setyaningsih, L. 2007. Pemanfaatan cendawanmioriza arbuskula dan kompos aktif untukmeningkatkan pertumbuhan semai mindi(Melia azedarach Linn) pada media tailingtambang emas Pongkor. [Tesis] SekolahPascasarjana IPB. Bogor.
Syarif, F. 2009. Serapan Sianida (Cn) PadaMikania cordata (Burm.f) B.L. Robinson,Centrosema pubescens Bth Dan Leersiahexandra Swartz Yang Ditanam PadaMedia Limbah Tailing Terkontaminasi Cn.Peneliti di Bidang Botani, Pusat PenelitianBiologi Lembaga Ilmu PengetahuanIndonesia. Jurnal Teknik Lingkungan No. 1Hal. 69 – 76.
Subiksa, IGM, 2002, Pemanfaatan Mikorizauntuk Penanggulangan Lahan Kritis,Makalah Falsafah Sains, IPB, Bogor.
Turjaman, Maman., Yana Sumarba Winarto.Erdy Santoso. 2005. Prospek aplikasiteknologi Cendawan Ektomikoriza (ECM)untuk mempercepat rehabilitasi hutan danlahan tergredasi. Seminar Nasional danWorkshop Cendawan Mikoriza.Universitas Jambi. Jambi.

ISSN 978-602-14989-0-3 BioETI
Kearifan lokal masyarakat adat Baduy dalam pemanfaatansumber daya hayatiENGGAR UTARI
Universitas Sultan Ageng Tirtayasa, SerangJl. Raya Jakarta Km 4 Pakupatan SerangE-mail: enggar.utari@yahoo. co.id
ABSTRACTMasyarakat Adat Baduy telah sejak lama dalam konservasi lingkungan yang dilaksanakan sebagai kearifan lokal merekadalam menjaga dan melindungi kelestarian serta keseimbangan alam. Konservasi dalam pengertian sekarang, seringditerjemahkan sebagai the wise use of nature resource yaitu pemanfaatan sumber daya alam secara bijaksana. Tujuanpenelitian ini untuk mengetahui bagaimana masyarakat adat Baduy dalam memanfaatkan sumber daya hayati. Populasidalam penelitian ini adalah seluruh masyarakat Adat Baduy yang ada di Desa Kanekes, kecamatan Leuwidamar,Kabupaten Lebak, Provinsi Banten. Penelitian ini dilaksanakan pada bulan Februari sampai Mei 2014 dengan metodekualitatif deskriptif. Pada tahap ini dilakukan langkah-langkah yang secara khusus menggali informasi mengenaipemanfaatan sumber daya hayati yang telah, sedang dan akan dilakukan. Informasi diperoleh dari data primer dan datasekunder. Data primer meliputi observasi, wawancara mendalam (Indepth Interview) kepada beberapa key persons sertadokumentasi mengenai kondisi alam dan beberapa jenis organisme yang dimanfaatkan. Data Sekunder diperoleh melaluistudi literatur. Hasil Penelitian menunjukkan bahwa dalam pemanfaatannya masyarakat tetap memperhatikan aturan adatuntuk tidak mengambil secara berlebihan. Masyarakat adat baduy mengambil seperlunya saja keperluan bahan tambahanpangan pokok, bahan obat-obat tradisional, bahan upacara adat, bahan bangunan, kayu bakar dan kerajinan rumah tangga.Dengan demikian disimpulkan bahwa dalam memanfaatkan sumber daya hayati, masyarakat melakukan norma-norma,nilai-nilai yang telah berlaku turun temurun yang merupakan kearifan lokal setempat.
Key words: Kearifan Lokal, Masyarakat adat Baduy, Sumber daya hayati
Pendahuluan
Masyarakat suku Baduy sangat menghormatikawasan hutan mereka, sehingga merekamenganggap hutan memiliki peran danfungsinya serta memiliki tingkat kesakralannya.Menurut Iskandar (2009) membagi kawasanBaduy kedalam beberapa zonasi, Kawasantersebut dibedakan menjadi 3 zonasi yanganalogi dengan zona inti/area inti, zonapenyagga, dan zona transisi/area transisi padasistem cagar biosfer. Zona inti/area inti, adalahkawasan yang dianggap paling sakral, disebutpula Daerah Kabuyutan, yaitu daerah SasakaPusaka Buana dan Sasaka Domas. Sementara,zona penyagga, merupakan daerah kurangsakral dibandingkan dengan zona inti, berada diluar zona inti, yaitu Baduy Dalam (tanahlarangan). Kemudian zona transisi berada diluar zona penyangga yang merupakan daerahBaduy Luar/Panamping dan daerah Dangk.
Berdasarkan pembagian zonasi dari utara keselatan, zona paling terluar adalah DaerahBaduy Luar/Panamping dan Derah Dangkayang merupakan bagian zona transisi, karenamerupakan daerah terluar yang fleksibel danberdampingan dengan zona penyangga.Sementara pada daerah penyangga merupakandaerah bagian Baduy Dalam/Tangtu karenadihuni oleh masyarakat Baduy Dalam yangmemiliki adat masih kuat dan tempat bermukimPuun. Seorang Puun dapat melindungikeberadaan zona inti (Daerah Kabuyutan) olehadanya daerah penyangga disekitarnya.Sedangkan zona yang paling sakral terletakpada zona inti, karena pada daerah tersebutmerupakan kawasan konservasi, yang tidakboleh dikunjungi bebas oleh setiap orang, tetapihanya digunakan untuk ziarah (Puun) pimpinanadat masyarakat Baduy pada waktu yangkhusus.

Enggar Utari 43
Dalam konteks pemanfaatan sumber daya yangada lingkungan masyarakat Adat Baduy,terdapat beberapa aturan yang wajib dijalankanoleh seluruh masyarakat adat Baduy. Aturantersebut merupakan sebuah kearifan lokalmasyarakat adat Baduy dalam memanfaatkansumber daya alam. Oleh karena itu, dalampenelitian ini telah dilakukan analisis mengenaipemanfaatan sumber daya hayati yang terikatdalam sebuah kearifan lokal masyarakat AdatBaduy.
BAHAN DAN METODE
Penelitian ini menggunakan metode penelitiandeskriptif kualitatif yang berguna melihatsecara detil dan mendalam bagaimanaMasyarakat Adat Baduy dalam memanfaatkansumber daya hayati.
Data diperoleh berdasar 2 sumber yaitu dataprimer yang meliputi pengamatan (observasi),wawancara mendalam (in depth interview) dandata sekunder yaitu studi literatur. Pengamatandilakukan secara cermat untuk mengamatibagaimana masyarakat sebagai sebuah sistemyang memiliki aspek-aspek tertentu, hubungan-hubungan yang khas dalam memanfaatkansumberdaya dayati. Wawancara mendalamdilakukan pada informan-informan yangberasal dari masyarakat Baduy Dalam maupunBaduyLuar baik anggota masyarakat maupun
pemimpin masyarakat. Studi literaturedilakukan pada sumber-sumber teori dan kasus-kasus penelitian lain yang terkait denganpenelitian ini.
Data kualitatif dianalisis melalui tiga jalan,yaitu reduksi data, penyajian data, danpenarikan kesimpulan. Reduksi data berupaproses pemilihan, pemilahan, pemusatanperhatian pada penyederhanaan, pengabstrakandan transformasi data kasar yang muncul secaratertulis di lapangan. Penyajian dimaksudkanpada sekumpulan informasi tersusun yangmember kemungkinan adanya penarikankesimpulan dan pengambilan tindakan.Sedangkan penarikan kesimpulan dalam hal inimencakup juga verifikasi atas kesimpulan itu.
HASIL DAN PEMBAHASAN
Deskripsi Masyarakat Adat baduyMasyarakat suku Baduy tinggal secaramengelompok pada suatu kampung danmenyebar di wilayah Kanekes. Sebagaiantisipasi terhadap masa depan kesukuannya,dibentuklah dua komunitas generasi peneruskesukuan mereka yaitu lahirlah kelompokpewaris yang disebut Baduy Dalam danpewaris yang disebut Baduy Luar. Keduapewaris ini memiliki aturan atau ciri-ciritertentu yang spesifik dalam melaksanakan
Gambar 1. Zonasi Praktik Konservasi Masyarakat Suku Baduy[Sumber: Iskandar, 2009: 96, Suparmini et.al: 2013: 21]
U
Daerah Muslim
Semakin keSelatan semakin
Sakral
Daerah Muslim (Daerah tidakSakral)
Daerah Baduy Luar / Panamping
Daerah Dangka
BaduyDalam/Tangtu
SasakaDomas
Zona IntiZona PenyanggaZona Transisi

Enggar Utari 44
amanat leluhurnya, dengan aturan hukumadatnya masing-masing yang sarat dengan cirikhas dan perbedaan yang jelas, namun mampumengikat menjadi satu kesatuan Baduy yangutuh. Inilah yang kemudian menjadikan merekasebagai satu kesukuan yang unik.
Komunitas yang menamakan suku BaduyDalam (Tangtu) atau disebut Baduy asli dimanapola kehidupan sehari-harinya benar-benarsangat kuat memegang hukum adat serta kukuhpengkuh dalam melaksanakan amanat leluhur.Baduy Dalam lebih menunjukkan pada replikaBaduy masa lalu. Ada tiga kampung yangmereka tinggali, yaitu Cikeusik, Cikartawana,dan Cibeo. Kelompok Baduy Dalam tidakpernah menambah jumlah kampung yang ada,wilayahnya hanya ada di tiga kampung tersebut.Komunitas yang menamakan dirinya SukuBaduy Luar (panamping) yang artinya adalahpendamping, karena mereka bermukim dibagian luar wilayah Baduy dan mendampingimasyarakat Baduy Dalam. Pada kegiatankehidupan sehari-harinya mereka diberikansuatu kebijakan atau kelonggaran dalammelaksanakan ketentuan-ketentuan hukum adat,tetapi ada batas-batas tertentu yang tetapmengikat mereka sebagai suatu komunitas adatkhas Suku Baduy (Kurnia & Sihabudin, 2010:9). Baduy Luar dari tahun ketahun jumlahkampungnya bertambah seiring denganpertambahan populasi disana. Berikut disajikanmatrik perbedaan masyarakat adat Baduy luardan dalam.
Berdasarkan data yang ada di lembagapamarentahan, laporan mengeni jumlahpenduduk masyarakat suku Baduy terhitungsampai tanggal 2 Mei 2014 Baduy adalah11.280 orang, yaitu 5.633 laki-laki dan 5.647perempuan, dengan total 2.886 KK. Jumlahtersebut tersebar di 62 kampung, denganrincian 3 kampung berada di Baduy Dalam dan59 kampung lainnya berada di wilayah BaduyLuar. Ada satu perkampungan khusus SukuBaduy yang berada di luar tanah ulayat Baduyyang diakui sebagai bagaian dari kesukuanmereka yaitu di daerah Kompol Desa
Sangkanwangi Kecamatan Leuwidamar yangsering disebut Baduy Kompol (Gambar 1).
Penduduk yang berada di Baduy Dalamberjumlah 1.150 orang (240 kepala keluarga)dan penduduk yang berada di Baduy Luarberjumlah 10.130 orang (2.646 kepalakeluarga). Jika dilihat perbedaan jeniskelaminnya, maka laki-laki yang berada diBaduy Dalam dan Baduy Luar masing-masingberjumlah 543 orang dan 5.190 orang.Sedangkan perempuan yang berada di BaduyDalam dan Baduy Luar masing-masingberjumlah 592 orang dan 5.055 orang.Perbandingan masyarakat adat baduy dalamdan baduy luar dapat dilihat pada gambarberikut.Pemanfaatan sumber daya hayatiSeluruh masyarakat suku Baduy tinggal dilembah, lereng bukit dan pegunungan yangdiapit oleh Hutan Larangan dan Hutan Titipan.Hutan Larangan adalah hutan yang sangat luas,ditumbuhi oleh berbagai tumbuhan tanpacampur tangan manusia, namun berkembangsecara alami. Di dalam hutan hidup secaraalami berbagai jenis hewan. Manusia tidakdiperkenankan untuk mengambil sebagian atauseluruh hasil dari hutan kecuali mengambilmadu, buah dan tanaman obat secukupnya. Ciridari hutan larangan yang paling mudah dikenaladalah tidak adanya jalan untuk dilalui,dikarenakan memang dilarang untuk dimasukimanusia kecuali orang Baduy Dalam itu sendiridengan tujuan mengontrol dan mengawasi sertamemanfaatkan beberapa bagian tanamanseperlunya saja.Hutan Titipan adalah hutan diluar hutan larangan yang berdasarkankeyakinan masyarakat suku Baduy hutantersebut telah dititipkan oleh leluhur/nenekmoyang mereka secara turun temurun untuktetap dijaga keutuhan dan kelestariannya,karena sangat menentukan bagi stabilitasekosistemnya. Hutan titipan disebut juga hutanlindung, dimana hutan yang keberadaannyatidak boleh dirubah dan dilarang untuk dikelolaserta dilindungi secara ketat oleh masyarakatmaupun pemerintah.
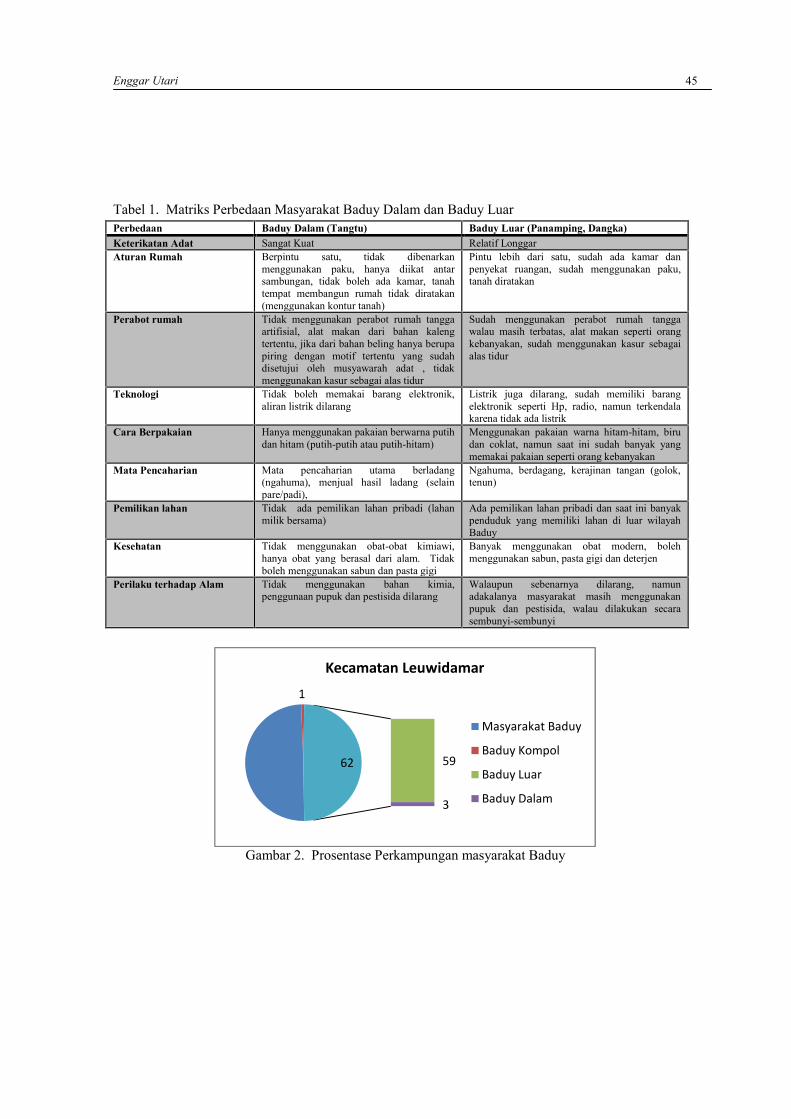
Enggar Utari 45
Tabel 1. Matriks Perbedaan Masyarakat Baduy Dalam dan Baduy LuarPerbedaan Baduy Dalam (Tangtu) Baduy Luar (Panamping, Dangka)Keterikatan Adat Sangat Kuat Relatif LonggarAturan Rumah Berpintu satu, tidak dibenarkan
menggunakan paku, hanya diikat antarsambungan, tidak boleh ada kamar, tanahtempat membangun rumah tidak diratakan(menggunakan kontur tanah)
Pintu lebih dari satu, sudah ada kamar danpenyekat ruangan, sudah menggunakan paku,tanah diratakan
Perabot rumah Tidak menggunakan perabot rumah tanggaartifisial, alat makan dari bahan kalengtertentu, jika dari bahan beling hanya berupapiring dengan motif tertentu yang sudahdisetujui oleh musyawarah adat , tidakmenggunakan kasur sebagai alas tidur
Sudah menggunakan perabot rumah tanggawalau masih terbatas, alat makan seperti orangkebanyakan, sudah menggunakan kasur sebagaialas tidur
Teknologi Tidak boleh memakai barang elektronik,aliran listrik dilarang
Listrik juga dilarang, sudah memiliki barangelektronik seperti Hp, radio, namun terkendalakarena tidak ada listrik
Cara Berpakaian Hanya menggunakan pakaian berwarna putihdan hitam (putih-putih atau putih-hitam)
Menggunakan pakaian warna hitam-hitam, birudan coklat, namun saat ini sudah banyak yangmemakai pakaian seperti orang kebanyakan
Mata Pencaharian Mata pencaharian utama berladang(ngahuma), menjual hasil ladang (selainpare/padi),
Ngahuma, berdagang, kerajinan tangan (golok,tenun)
Pemilikan lahan Tidak ada pemilikan lahan pribadi (lahanmilik bersama)
Ada pemilikan lahan pribadi dan saat ini banyakpenduduk yang memiliki lahan di luar wilayahBaduy
Kesehatan Tidak menggunakan obat-obat kimiawi,hanya obat yang berasal dari alam. Tidakboleh menggunakan sabun dan pasta gigi
Banyak menggunakan obat modern, bolehmenggunakan sabun, pasta gigi dan deterjen
Perilaku terhadap Alam Tidak menggunakan bahan kimia,penggunaan pupuk dan pestisida dilarang
Walaupun sebenarnya dilarang, namunadakalanya masyarakat masih menggunakanpupuk dan pestisida, walau dilakukan secarasembunyi-sembunyi
Gambar 2. Prosentase Perkampungan masyarakat Baduy
1
59
3
62
Kecamatan Leuwidamar
Masyarakat Baduy
Baduy Kompol
Baduy Luar
Baduy Dalam
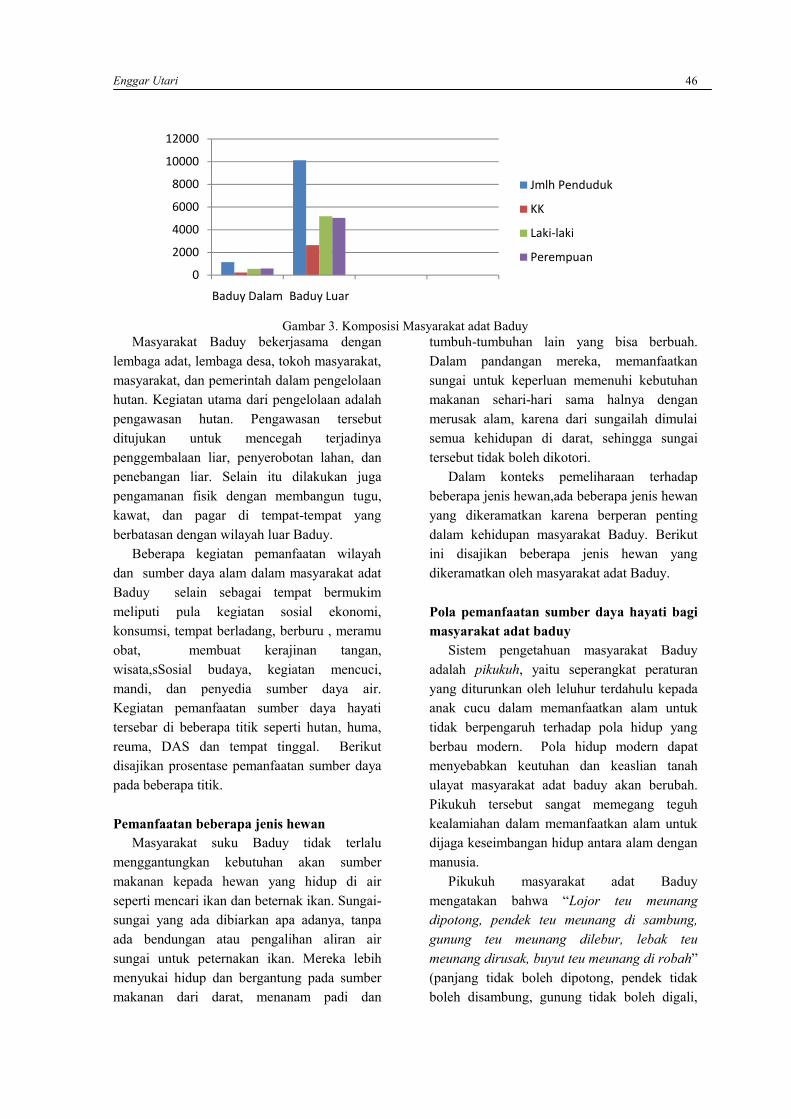
Enggar Utari 46
Gambar 3. Komposisi Masyarakat adat BaduyMasyarakat Baduy bekerjasama dengan
lembaga adat, lembaga desa, tokoh masyarakat,masyarakat, dan pemerintah dalam pengelolaanhutan. Kegiatan utama dari pengelolaan adalahpengawasan hutan. Pengawasan tersebutditujukan untuk mencegah terjadinyapenggembalaan liar, penyerobotan lahan, danpenebangan liar. Selain itu dilakukan jugapengamanan fisik dengan membangun tugu,kawat, dan pagar di tempat-tempat yangberbatasan dengan wilayah luar Baduy.
Beberapa kegiatan pemanfaatan wilayahdan sumber daya alam dalam masyarakat adatBaduy selain sebagai tempat bermukimmeliputi pula kegiatan sosial ekonomi,konsumsi, tempat berladang, berburu , meramuobat, membuat kerajinan tangan,wisata,sSosial budaya, kegiatan mencuci,mandi, dan penyedia sumber daya air.Kegiatan pemanfaatan sumber daya hayatitersebar di beberapa titik seperti hutan, huma,reuma, DAS dan tempat tinggal. Berikutdisajikan prosentase pemanfaatan sumber dayapada beberapa titik.
Pemanfaatan beberapa jenis hewanMasyarakat suku Baduy tidak terlalu
menggantungkan kebutuhan akan sumbermakanan kepada hewan yang hidup di airseperti mencari ikan dan beternak ikan. Sungai-sungai yang ada dibiarkan apa adanya, tanpaada bendungan atau pengalihan aliran airsungai untuk peternakan ikan. Mereka lebihmenyukai hidup dan bergantung pada sumbermakanan dari darat, menanam padi dan
tumbuh-tumbuhan lain yang bisa berbuah.Dalam pandangan mereka, memanfaatkansungai untuk keperluan memenuhi kebutuhanmakanan sehari-hari sama halnya denganmerusak alam, karena dari sungailah dimulaisemua kehidupan di darat, sehingga sungaitersebut tidak boleh dikotori.
Dalam konteks pemeliharaan terhadapbeberapa jenis hewan,ada beberapa jenis hewanyang dikeramatkan karena berperan pentingdalam kehidupan masyarakat Baduy. Berikutini disajikan beberapa jenis hewan yangdikeramatkan oleh masyarakat adat Baduy.
Pola pemanfaatan sumber daya hayati bagimasyarakat adat baduy
Sistem pengetahuan masyarakat Baduyadalah pikukuh, yaitu seperangkat peraturanyang diturunkan oleh leluhur terdahulu kepadaanak cucu dalam memanfaatkan alam untuktidak berpengaruh terhadap pola hidup yangberbau modern. Pola hidup modern dapatmenyebabkan keutuhan dan keaslian tanahulayat masyarakat adat baduy akan berubah.Pikukuh tersebut sangat memegang teguhkealamiahan dalam memanfaatkan alam untukdijaga keseimbangan hidup antara alam denganmanusia.
Pikukuh masyarakat adat Baduymengatakan bahwa “Lojor teu meunangdipotong, pendek teu meunang di sambung,gunung teu meunang dilebur, lebak teumeunang dirusak, buyut teu meunang di robah”(panjang tidak boleh dipotong, pendek tidakboleh disambung, gunung tidak boleh digali,
0
2000
4000
6000
8000
10000
12000
Baduy Dalam Baduy Luar
Jmlh Penduduk
KK
Laki-laki
Perempuan

Enggar Utari 47
lebak tidak boleh dirusak), artinya bahwasegala sesuatu yang diciptakan oleh Sang MahaPencipta di alam tidak boleh diubah dalamkeadaan apapun baik ditambahkan maupundikurangi, karena hal tersebut dapatmenimbulkan kerusakan yang akanmenyebabkan kerugian yang akan dirasakanoleh masyarakat luas. Akibat kerusakan dapatmenimbulkan kehancuran dari sebuahkhususnya di Provinsi Banten
Kesederhanaan hidup tersebut menjadikanmereka untuk selalu bersatu dengan alamdengan menjaga stabilitas alam agar tetapseimbang. Menurut masyarakat adat baduy,alam dapat memberikan respon positif biladijaga dengan baik sedangkan jikadiperlakukan dengan tidak sesuai maka akanmenyebabkan dampak negatif yang akandirasakan oleh semua masyarakat yangselanjutnya akan menimbulkan bencana alam.
Pemanfaatan hutan tua sebagian besardilakukan oleh masyarakat Baduy dalam karena
Gambar 4. Prosentase wilayah pemanfaatan SDH olehMasyarakat Baduy
Tabel 2.Jenis-jenis hewan yang dikeramatkanJenis hewan Tujuan PemeliharaanAyam Diperlakukan dengan sebaik-baiknya dan
tidak boleh disiksa atau ditelantarkan bilatidak ingin mendapat hukuman dankutukan Tuhan.
Kucing Rumah Menjaga padi yang ada di lumbung dariserangan hama tikus
Anjing Menjaga rumah penduduk, kebun,ladang/humaserta ayam yang dipelihara dikebun dari serangan hama seperti musang,meongsisi, lasun, babi hutan
Burung burung yang hidup liar di sekitar hutanproduksi/hutan garapan sebagai pembasmihama berupa ulat-ulat yang menggerogotihasil bumi seperti padi, sayur-sayuran,maupun kayu produksinya.
aksesibilitasnya lebih dekat menuju hutan tua.Dalam pemanfaatannya masyarakat tetapmemperhatikan aturan adat yang tentunya harusseizin Puun, kemudian tidak mengambil secaraberlebihan, mereka mengambil hanyaseperlunya saja ketika menemukan hasil hutan.Selain itu hutan sangat dikeramatkan, sehinggatidak dimanfaatkan hasil hutannya, namunmemiliki fungsi sosial budaya untukkepentingan kepercayaan masyarakat Baduy,sebagai tempat muja
Berdasarkan hasil pencatatan yangdilakukan oleh Iskandar (2009: 102), flora yangterdapat di dukuh lembur Baduy Luar terdapat79 jenis tumbuhan. Berbagai jenis tumbuhandominan (khas) di dukuh lembur diantaranyaadalah: Aren/Kawung (Arenga pinnata),Kelapa (Cocos nucifera), Durian/Kadu (Duriozibethinus), Andul (Elaeocarpus obtusus), AwiGombong (Gigantochola verticilata), Cariang(Homolaena odorata), Pisitan (Lansiumdomesticum), Kiray (Metroxylon sagu),Rambutan (Nephelium lappaceum), danKecibeling (Sericocalyx crispus).
Secara umum adanya aneka ragam jenistumbuhan, buah-buahan dan kayu-kayuan, didaerah kampung dan dukuh lembur, bantaransungai yang sangat rimbun, serta memilikikualitas air yang baik bagi habitat berbagaijenis ikan dapat memiliki fungsi konservasiyang sangat penting diantaranya: (1) konservasijenis/varietas tumbuhan, (2) sebagai habitathewan, seperti jenis-jenis serangga, burung danmamalia kecil, (3) sebagai habitat bagi jenis-jenis ikan yang hidup di sungai, (4) konservasitanah dan mengatur sistem hidrologi air, (5)iklim mikro, memberi keteduhan dan kesejukanlingkungan pemukiman, dan (6) fungsi sosialekonomi, dengan menghasilkan buah-buahandan hasil industri rumah tangga, seperti gulaaren dan hasil sadapan karangan bunga pohonaren.
Zona Tengah/ PemanfaatanZona tengah berada diatas lembah-lembah
bukit (diatas kawasan pemukiman) atau bagian
57
7.27.2
7.2
21.4
Prosentase Wilayah Pemanfaatan
Hutan
Huma
Reuma
DAS
Pemukiman

Enggar Utari 48
luar pemukiman/kampung dan dukuh lembur,merupakan daerah hutan garapan. Hutangarapan adalah wilayah hutan yang difungsikansebagai kebun, tempat ngahuma (berladang).Lahan ini dapat disebut juga sebagai lahanabadi untuk tanaman tumpang sari terutamauntuk tanaman pangan, yaitu padi dankomoditas kebun. Oleh karena itu, padawilayah ini lahan huma (ladang) dan reuma(hutan sekunder) bekas ladang yang sedangdiistirahatkan (diberakan) dengan berbagaiumur. Hutan sekunder yang diberakan 2-3tahun dinamakan reuma ngora (hutan sekundermuda), lahan ini tidak akan sempat untukmenjadi hutan, melainkan hanya menjadisemak belukar. Jika lahan tersebut telahdiberakan lebih dari 3 tahun dinamakan reumakolot (hutan sekunder tua).
Pada Reuma sedikit sekali didapatkantumbuhan bermanfaat yang ditanam pada saatberhuma. Kebanyakan tumbuhan yang ada diReuma yaitu belukar yang dimanfaatkan untukkayu bakar. Namun, ketika daerahreumadiberakan, banyak ditemukan jenis-jenistumbuhan. Diantaranya adalah tumbuhancampuran buah-buahan dan kayu-kayuan, sisa-sisa perladangan dan jenis-jenis tumbuhanhutan sekunder yang sedang mengalami suksesialami. Sedangkan pada lahan huma biasanyamasyarakat menanam padi, selain tanaman padibiasanya ditanami pula aneka ragam tanamanlain.
Hutan garapan terbagi-bagi menjadi milikperkeluarga dengan batas yang jelas sertadikelola oleh masing-masing keluarga. Namun,penggarapannya tetap harus disiplin menurutcara dan aturan-aturan adat Baduy yang berlaku.Tujuannya untuk mencegah kerusakan,memelihara dan melestarikan potensinya untukmenghasilkan panen yang memuaskan.Erwinantu (2012: 84) menjelaskan bahwa haltersebut telah tersirat makna untuk memeliharakeharmonisan hubungan manusia dengan alamsebagai sahabat dan pasangannya, yangmemeluk, memangku, mengayomi danmemberikan semua daya potensinya. Tanpa
pamrih kecuali meminta untuk dihormatidengan cara memelihara dan janganmerusaknya, serta jangan meminta darinyamelebihi kekuatan dan kemampuannya.
Pencatatan mengenai keanekaragamanhayati (beberapa jenis tumbuhan dan burung) didaerah hutan garapan telah dilakukan olehIskandar melalui penelitinnya pada tahun 2009,hasil pencatatan tersebut dapat dilihat padaTabel 3.
Secara umum huma memiliki fungsi pentingbagi pelestarian alam dan sosial ekonomipenduduk Baduy, diantaranya (1) bahantambahan pangan pokok, (2) bahan lalab/sayur,(3) bahan bumbu masak, (4) bahan obat-obattradisional, (5) bahan upacara adat, (6) bahanbangunan dan kayu bakar, dan (7) bahanindustri dan kerajinan rumah tangga.Sedangkan fungsi budaya, berladang dianggapsuatu kewajiban dalam agama mereka yaituSlam Sunda Wiwitan.
Sementara itu, reuma secara umummempunyai fungsi, (1) fungsi konservasikeanekaan jenis/varietas tumbuhan, (2) sebagaihabitat fauna, (3) fungsi perlindungan tanahdan hidrologi, (4) sebagai fungsi sosialekonomi, dapat dikonsumsi aneka ragam hasilbuah-buahan (durian, pisitan, rambutan) danlalap/sayur (seperti petai, jengkol).
Hutan Lindung adalah hutan yang beradadiluar hutan larangan/hutan sakral SasakaDomas. Daerah ini berada di puncak-puncakbukit, dan lembah tempat keluarnya mata air,kemudian diperuntukkan bagi konservasi hutan.Masyarakat Baduy menyebut kawasan inisebagai “leuweung kolot (hutan tua), leuweungtitipan (hutan titipan) atau leuweung gede(hutan besar/luas)” artinya hutan tua atau hutantitipan yang berdasarkan keyakinan masyarakatsuku Baduy, hutan tersebut telah dititipkan olehleluhur/nenek moyang mereka secara turuntemurun untuk tetap dijaga keutuhan dankelestariannya, karena sangat menentukan bagistabilitas ekosistemnya yang harus dijagakelestariannya. Mereka sangat patuh terhadaplarangan untuk tidak masuk ke wilayah hutan

Enggar Utari 49
tua tanpa seizin Puun. Beberapa hutan tua yangmasih cukup luas ditemukan di daerah BaduyDalam, khusunya di Cibeo dan Cikeusik.
Hutan lindung tidak boleh dibuka dandijadikan ladang. Sehingga, lahan hutan dipuncak-puncak bukit tersebut cukup rimbun.Hutan Titipan tumbuh dan berkembang secaraalamiah, larangan-larangan masyarakat padaHutan Titipan diantaranya adalah tidakdiperbolehkan kepada siapapun yang masuk ke
dalam hutan untuk mengambil atau menebangpohon dan tumbuhan lain maupun menangkaphewan, atau potensi sumber daya alam yangada di dalamnya, lebih-lebih dalam halmengeksploitasi untuk alasan apapun. Olehkarena itu, wilayah ini merupakan daerahkonservasi yang tidak boleh dibuat untukladang, dimana keberadaannya tidak bolehdiubah dan dilarang untuk dikelola serta
Tabel 3. Hasil pencatatan jenis keanekaragaman hayatidi Hutan GarapanHuma ReumaTercatat : 79 jenis tumbuhan Tercatat : 93 jenis tumbuhanTumbuhan khas (dominan):Padi (Oryza sativa) dengan memiliki 89 varietaslokal, Kacang Hiris (Cajanus cajan), Pisang/Cau(Musa paradisiaca), Ubi Jalar (Ipomoea batatas),Jagung (Zea mays), Ubi Manis (Dioscorea alata),Talas (Colocasia esculenta), Lengkuas/Laja(Languas galaga), Turubuk/Tiwu Endog (Saccharumedule), Mentimun/Bonteng (Cucumis sativus),Kunyit (Panicum viride), Cabe Rawit (Capsicumfrutescens), Hanjeli (Coix lacrima jobi).
Tumbuhan khas (dominan):Jeungjing (Albizia chinensis), Aren/Kawung (Arengapinnata), Sempur (Dillenea aurea), Beunying (Ficusbrevicuspis), Seuhang (Ficus grassularoides),Hamerang (Ficus fulva), Babakoan/Ki Tambaga(Flemingia lineata), Mara (Macaranga tanarius),Harendong (Melastoma malabatricum), Ki Seureuh(Piper aduncum), Glagah/Kaso (Saccharumspontaneum), dan Jengkol (Pithecelobium jeringa).
Huma ReumaTercatat : 48 jenis burung Tercatat : 74 jenis burungBurung khas (dominan):Tikukur (Streptopelia chinensis), Burung layang-layang/Striti (Collocalia linchi), Kuricang(Pycnonotus atriceps), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris).
Burung khas (dominan):Kuricang (Pycnonotus atriceps), Kutilang(Pycnonotus aurigaster), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris), Burung Cabe-cabe Gunung (Dicaeum trigonstigma), Striti(Collocalia linchi).
Gambar 5 . Daerah hutan garapan (a) reuma (semak belukar), (b) lahan huma, (c) Pemandangan di hutangarapan [Sumber: Dokumentasi Pribadi]
(b)(a)
(c)(c)
Enggar Utari 49
tua tanpa seizin Puun. Beberapa hutan tua yangmasih cukup luas ditemukan di daerah BaduyDalam, khusunya di Cibeo dan Cikeusik.
Hutan lindung tidak boleh dibuka dandijadikan ladang. Sehingga, lahan hutan dipuncak-puncak bukit tersebut cukup rimbun.Hutan Titipan tumbuh dan berkembang secaraalamiah, larangan-larangan masyarakat padaHutan Titipan diantaranya adalah tidakdiperbolehkan kepada siapapun yang masuk ke
dalam hutan untuk mengambil atau menebangpohon dan tumbuhan lain maupun menangkaphewan, atau potensi sumber daya alam yangada di dalamnya, lebih-lebih dalam halmengeksploitasi untuk alasan apapun. Olehkarena itu, wilayah ini merupakan daerahkonservasi yang tidak boleh dibuat untukladang, dimana keberadaannya tidak bolehdiubah dan dilarang untuk dikelola serta
Tabel 3. Hasil pencatatan jenis keanekaragaman hayatidi Hutan GarapanHuma ReumaTercatat : 79 jenis tumbuhan Tercatat : 93 jenis tumbuhanTumbuhan khas (dominan):Padi (Oryza sativa) dengan memiliki 89 varietaslokal, Kacang Hiris (Cajanus cajan), Pisang/Cau(Musa paradisiaca), Ubi Jalar (Ipomoea batatas),Jagung (Zea mays), Ubi Manis (Dioscorea alata),Talas (Colocasia esculenta), Lengkuas/Laja(Languas galaga), Turubuk/Tiwu Endog (Saccharumedule), Mentimun/Bonteng (Cucumis sativus),Kunyit (Panicum viride), Cabe Rawit (Capsicumfrutescens), Hanjeli (Coix lacrima jobi).
Tumbuhan khas (dominan):Jeungjing (Albizia chinensis), Aren/Kawung (Arengapinnata), Sempur (Dillenea aurea), Beunying (Ficusbrevicuspis), Seuhang (Ficus grassularoides),Hamerang (Ficus fulva), Babakoan/Ki Tambaga(Flemingia lineata), Mara (Macaranga tanarius),Harendong (Melastoma malabatricum), Ki Seureuh(Piper aduncum), Glagah/Kaso (Saccharumspontaneum), dan Jengkol (Pithecelobium jeringa).
Huma ReumaTercatat : 48 jenis burung Tercatat : 74 jenis burungBurung khas (dominan):Tikukur (Streptopelia chinensis), Burung layang-layang/Striti (Collocalia linchi), Kuricang(Pycnonotus atriceps), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris).
Burung khas (dominan):Kuricang (Pycnonotus atriceps), Kutilang(Pycnonotus aurigaster), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris), Burung Cabe-cabe Gunung (Dicaeum trigonstigma), Striti(Collocalia linchi).
Gambar 5 . Daerah hutan garapan (a) reuma (semak belukar), (b) lahan huma, (c) Pemandangan di hutangarapan [Sumber: Dokumentasi Pribadi]
(b)(a)
(c)(c)
Enggar Utari 49
tua tanpa seizin Puun. Beberapa hutan tua yangmasih cukup luas ditemukan di daerah BaduyDalam, khusunya di Cibeo dan Cikeusik.
Hutan lindung tidak boleh dibuka dandijadikan ladang. Sehingga, lahan hutan dipuncak-puncak bukit tersebut cukup rimbun.Hutan Titipan tumbuh dan berkembang secaraalamiah, larangan-larangan masyarakat padaHutan Titipan diantaranya adalah tidakdiperbolehkan kepada siapapun yang masuk ke
dalam hutan untuk mengambil atau menebangpohon dan tumbuhan lain maupun menangkaphewan, atau potensi sumber daya alam yangada di dalamnya, lebih-lebih dalam halmengeksploitasi untuk alasan apapun. Olehkarena itu, wilayah ini merupakan daerahkonservasi yang tidak boleh dibuat untukladang, dimana keberadaannya tidak bolehdiubah dan dilarang untuk dikelola serta
Tabel 3. Hasil pencatatan jenis keanekaragaman hayatidi Hutan GarapanHuma ReumaTercatat : 79 jenis tumbuhan Tercatat : 93 jenis tumbuhanTumbuhan khas (dominan):Padi (Oryza sativa) dengan memiliki 89 varietaslokal, Kacang Hiris (Cajanus cajan), Pisang/Cau(Musa paradisiaca), Ubi Jalar (Ipomoea batatas),Jagung (Zea mays), Ubi Manis (Dioscorea alata),Talas (Colocasia esculenta), Lengkuas/Laja(Languas galaga), Turubuk/Tiwu Endog (Saccharumedule), Mentimun/Bonteng (Cucumis sativus),Kunyit (Panicum viride), Cabe Rawit (Capsicumfrutescens), Hanjeli (Coix lacrima jobi).
Tumbuhan khas (dominan):Jeungjing (Albizia chinensis), Aren/Kawung (Arengapinnata), Sempur (Dillenea aurea), Beunying (Ficusbrevicuspis), Seuhang (Ficus grassularoides),Hamerang (Ficus fulva), Babakoan/Ki Tambaga(Flemingia lineata), Mara (Macaranga tanarius),Harendong (Melastoma malabatricum), Ki Seureuh(Piper aduncum), Glagah/Kaso (Saccharumspontaneum), dan Jengkol (Pithecelobium jeringa).
Huma ReumaTercatat : 48 jenis burung Tercatat : 74 jenis burungBurung khas (dominan):Tikukur (Streptopelia chinensis), Burung layang-layang/Striti (Collocalia linchi), Kuricang(Pycnonotus atriceps), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris).
Burung khas (dominan):Kuricang (Pycnonotus atriceps), Kutilang(Pycnonotus aurigaster), Jog-jog (Pycnonotusgoiavier), Pacikrak (Prinia familiaris), Burung Cabe-cabe Gunung (Dicaeum trigonstigma), Striti(Collocalia linchi).
Gambar 5 . Daerah hutan garapan (a) reuma (semak belukar), (b) lahan huma, (c) Pemandangan di hutangarapan [Sumber: Dokumentasi Pribadi]
(b)(a)
(c)(c)

Enggar Utari 50
dilindungi secara ketat oleh masyarakatmaupun pemerintah, karena selain untukmengukur iklim mikro juga berfungsi untukmengatur tata air di Desa Kanekes.
Menurut informasi, masyarakat disekitarnya (Baduy Dalam) boleh memanfaatkandan mengambil hasil-hasil hutan lindungseperti diambil kayunya dan buah-buahansecara terbatas, namun harus dengan seizin adat(Puun), dimana mereka boleh menanamtanaman jika diketahui masih ada lahan yangkosong atau ada pohon yang roboh dengansendirinya, kemudian diganti dengan tanamanpohon baru seperti pohon Durian. Setelah itu,hasilnya menjadi hak yang menanam danmerawatnya, dalam artian boleh memanfaatkanpotensinya tanpa mengganggu kondisi aslinya.Oleh karena itu barang siapa yang merusak ataumemanfaatkan wilayah ini secara komersilakan mendapatkan musibah, sehingga daerahini dikenal oleh masyarakat sebagai PunggungBumi (Sanghyang Pundak).
Di daerah-daerah hutan tersebut tumbuhaneka ragam tumbuhan khas hutan, beberapajenis buah-buahan yang tumbuh secara liar atausetengah liar. Pohon-pohon tersebut dibiarkantumbuh bebas, berkembang secara alami sejakpuluhan hingga ratusan bahkan ribuan tahunsilam. Seperti adanya pohon durian yangbatangnya berdiameter lebih dari 2-3 meterdengan tinggi batang menjulang ke langit lebihdari 50-100 meter dan masih menghasilkanbuah durian berkualitas. Kemudian masihterdapat tumbuhan jenis kayu langka yangumumnya memiliki ukuran yang sangat besardan tinggi, dikarenakan tumbuhan ini sangatdilindungi keberadaannya dari sejak dulu(nenek moyang) hingga sekarang baik secaraadat maupun dari pemerintah.
Pencatatan mengenai keanekaragamanhayati di daerah hutan garapan telah dilakukanoleh Iskandar melalui publikasi penelitinnyapada tahun 2009, hasil pencatatan tersebutdapat dilihat pada Tabel 4.
Masyarakat suku Baduy memahamibagaimana menjaga tatanan kehidupanmakhluk hidup yang berada di hutan lindung,berbagai macam hewan maupun tumbuhanharus diamankan dan dijaga kelestariannya agarkeseimbangan alam dan ekosistemnya tetapterjaga dengan utuh. Menurut penuturanmereka bahwa terdapat satwa harimau diwilayah hutannya namun satwa tersebutdikatakan sebagai hewan ghaib yang dapatterlihat maupun tidak, sebagai tata tertibnyahewan ini tidak boleh disombongkan karenapada dasarnya mereka adalah makhluk ghaibyang menghuni dan menjaga hutan lindung dariancaman luar. Bahkan jika masyarakat adayang melihat, hal tersebut dapat berakibatbahwa mereka telah lalai terhadap aturan dansebagai teguran terhadap pelanggaran.
Masyarakat suku Baduy biasanya me-manfaatkan hutan tua untuk mencari bahan-bahan kerajinan tangan, buah-buahan, mencarimadu, jamur, mengambil kayu bakar daripohon yang sudah roboh, dan lain-lain.Peamanfaatan hutan tua ini terutama bagimasyarakat Baduy Dalam yang aksesibilitasnyalebih dekat menuju hutan tua. Dalampemanfaatannya masyarakat tetap memper-hatikan aturan adat yang tentunya harus seizinPuun, kemudian tidak mengambil secaraberlebihan, mereka mengambil hanyaseperlunya saja ketika menemukan hasil hutan.Oleh karena itu, secara umum fungsi hutan tuasebagai konservasi alam adalah (1) konservasikeanekaragaman jenis/varietas tumbuhan, (2)
Gambar 6. Daerah hutan lindung di puncak bukit [Sumber: Dokumentasi Pribadi]
Enggar Utari 50
dilindungi secara ketat oleh masyarakatmaupun pemerintah, karena selain untukmengukur iklim mikro juga berfungsi untukmengatur tata air di Desa Kanekes.
Menurut informasi, masyarakat disekitarnya (Baduy Dalam) boleh memanfaatkandan mengambil hasil-hasil hutan lindungseperti diambil kayunya dan buah-buahansecara terbatas, namun harus dengan seizin adat(Puun), dimana mereka boleh menanamtanaman jika diketahui masih ada lahan yangkosong atau ada pohon yang roboh dengansendirinya, kemudian diganti dengan tanamanpohon baru seperti pohon Durian. Setelah itu,hasilnya menjadi hak yang menanam danmerawatnya, dalam artian boleh memanfaatkanpotensinya tanpa mengganggu kondisi aslinya.Oleh karena itu barang siapa yang merusak ataumemanfaatkan wilayah ini secara komersilakan mendapatkan musibah, sehingga daerahini dikenal oleh masyarakat sebagai PunggungBumi (Sanghyang Pundak).
Di daerah-daerah hutan tersebut tumbuhaneka ragam tumbuhan khas hutan, beberapajenis buah-buahan yang tumbuh secara liar atausetengah liar. Pohon-pohon tersebut dibiarkantumbuh bebas, berkembang secara alami sejakpuluhan hingga ratusan bahkan ribuan tahunsilam. Seperti adanya pohon durian yangbatangnya berdiameter lebih dari 2-3 meterdengan tinggi batang menjulang ke langit lebihdari 50-100 meter dan masih menghasilkanbuah durian berkualitas. Kemudian masihterdapat tumbuhan jenis kayu langka yangumumnya memiliki ukuran yang sangat besardan tinggi, dikarenakan tumbuhan ini sangatdilindungi keberadaannya dari sejak dulu(nenek moyang) hingga sekarang baik secaraadat maupun dari pemerintah.
Pencatatan mengenai keanekaragamanhayati di daerah hutan garapan telah dilakukanoleh Iskandar melalui publikasi penelitinnyapada tahun 2009, hasil pencatatan tersebutdapat dilihat pada Tabel 4.
Masyarakat suku Baduy memahamibagaimana menjaga tatanan kehidupanmakhluk hidup yang berada di hutan lindung,berbagai macam hewan maupun tumbuhanharus diamankan dan dijaga kelestariannya agarkeseimbangan alam dan ekosistemnya tetapterjaga dengan utuh. Menurut penuturanmereka bahwa terdapat satwa harimau diwilayah hutannya namun satwa tersebutdikatakan sebagai hewan ghaib yang dapatterlihat maupun tidak, sebagai tata tertibnyahewan ini tidak boleh disombongkan karenapada dasarnya mereka adalah makhluk ghaibyang menghuni dan menjaga hutan lindung dariancaman luar. Bahkan jika masyarakat adayang melihat, hal tersebut dapat berakibatbahwa mereka telah lalai terhadap aturan dansebagai teguran terhadap pelanggaran.
Masyarakat suku Baduy biasanya me-manfaatkan hutan tua untuk mencari bahan-bahan kerajinan tangan, buah-buahan, mencarimadu, jamur, mengambil kayu bakar daripohon yang sudah roboh, dan lain-lain.Peamanfaatan hutan tua ini terutama bagimasyarakat Baduy Dalam yang aksesibilitasnyalebih dekat menuju hutan tua. Dalampemanfaatannya masyarakat tetap memper-hatikan aturan adat yang tentunya harus seizinPuun, kemudian tidak mengambil secaraberlebihan, mereka mengambil hanyaseperlunya saja ketika menemukan hasil hutan.Oleh karena itu, secara umum fungsi hutan tuasebagai konservasi alam adalah (1) konservasikeanekaragaman jenis/varietas tumbuhan, (2)
Gambar 6. Daerah hutan lindung di puncak bukit [Sumber: Dokumentasi Pribadi]
Enggar Utari 50
dilindungi secara ketat oleh masyarakatmaupun pemerintah, karena selain untukmengukur iklim mikro juga berfungsi untukmengatur tata air di Desa Kanekes.
Menurut informasi, masyarakat disekitarnya (Baduy Dalam) boleh memanfaatkandan mengambil hasil-hasil hutan lindungseperti diambil kayunya dan buah-buahansecara terbatas, namun harus dengan seizin adat(Puun), dimana mereka boleh menanamtanaman jika diketahui masih ada lahan yangkosong atau ada pohon yang roboh dengansendirinya, kemudian diganti dengan tanamanpohon baru seperti pohon Durian. Setelah itu,hasilnya menjadi hak yang menanam danmerawatnya, dalam artian boleh memanfaatkanpotensinya tanpa mengganggu kondisi aslinya.Oleh karena itu barang siapa yang merusak ataumemanfaatkan wilayah ini secara komersilakan mendapatkan musibah, sehingga daerahini dikenal oleh masyarakat sebagai PunggungBumi (Sanghyang Pundak).
Di daerah-daerah hutan tersebut tumbuhaneka ragam tumbuhan khas hutan, beberapajenis buah-buahan yang tumbuh secara liar atausetengah liar. Pohon-pohon tersebut dibiarkantumbuh bebas, berkembang secara alami sejakpuluhan hingga ratusan bahkan ribuan tahunsilam. Seperti adanya pohon durian yangbatangnya berdiameter lebih dari 2-3 meterdengan tinggi batang menjulang ke langit lebihdari 50-100 meter dan masih menghasilkanbuah durian berkualitas. Kemudian masihterdapat tumbuhan jenis kayu langka yangumumnya memiliki ukuran yang sangat besardan tinggi, dikarenakan tumbuhan ini sangatdilindungi keberadaannya dari sejak dulu(nenek moyang) hingga sekarang baik secaraadat maupun dari pemerintah.
Pencatatan mengenai keanekaragamanhayati di daerah hutan garapan telah dilakukanoleh Iskandar melalui publikasi penelitinnyapada tahun 2009, hasil pencatatan tersebutdapat dilihat pada Tabel 4.
Masyarakat suku Baduy memahamibagaimana menjaga tatanan kehidupanmakhluk hidup yang berada di hutan lindung,berbagai macam hewan maupun tumbuhanharus diamankan dan dijaga kelestariannya agarkeseimbangan alam dan ekosistemnya tetapterjaga dengan utuh. Menurut penuturanmereka bahwa terdapat satwa harimau diwilayah hutannya namun satwa tersebutdikatakan sebagai hewan ghaib yang dapatterlihat maupun tidak, sebagai tata tertibnyahewan ini tidak boleh disombongkan karenapada dasarnya mereka adalah makhluk ghaibyang menghuni dan menjaga hutan lindung dariancaman luar. Bahkan jika masyarakat adayang melihat, hal tersebut dapat berakibatbahwa mereka telah lalai terhadap aturan dansebagai teguran terhadap pelanggaran.
Masyarakat suku Baduy biasanya me-manfaatkan hutan tua untuk mencari bahan-bahan kerajinan tangan, buah-buahan, mencarimadu, jamur, mengambil kayu bakar daripohon yang sudah roboh, dan lain-lain.Peamanfaatan hutan tua ini terutama bagimasyarakat Baduy Dalam yang aksesibilitasnyalebih dekat menuju hutan tua. Dalampemanfaatannya masyarakat tetap memper-hatikan aturan adat yang tentunya harus seizinPuun, kemudian tidak mengambil secaraberlebihan, mereka mengambil hanyaseperlunya saja ketika menemukan hasil hutan.Oleh karena itu, secara umum fungsi hutan tuasebagai konservasi alam adalah (1) konservasikeanekaragaman jenis/varietas tumbuhan, (2)
Gambar 6. Daerah hutan lindung di puncak bukit [Sumber: Dokumentasi Pribadi]

Enggar Utari 51
habitat satwa liar, (3) fungsi konservasi tanahdan hidrologi sungai, (4) fungsi iklim mikro,(5) fungsi sosial ekonomi, untuk pengambilananeka ragam hasil hutan.
Tabel 4. Hasil pencatatan jenis keanekaragaman hayatidi Hutan Lindung
Tercatat : 165 jenis tumbuhanTumbuhan khas (dominan):Keranji, Pisitan, Durian, Picung (Tumbuhan liar yangmenghasilkan buah untuk dimanfaatkan), Kayu Garu, Kiara,Huru, Leles, Rasamala(kayu langka), Leungsir, Kalimborot,Tapos, Lolot, Kaneungay, Kibuluh, Haraghag, Jirak, Hantap,Puspa, Kileho, Bubuay, Putat, Trembeusi, Paku Kapal,Bayur/Cayur (Pterosperum javanicum), Burahol atau Turalak(Stechocarpus buraholhook), Caringin/Beringin (Ficusbenyamin lnn), Kiara (Ficus Indica), Kianggir (Otoporaspectabilis), Kokosan atau Pisitan (Lansium domesticum),Jeunjing (Albizzia fakata backer), Rotan (Korthalisia laciosa),Tubaleleur atau Areuy Kawao (Milletia servicea), Kedoya(Dysoxylum caulostachym miq), Bambu Apus (Gigantocholaapus kurz), Bambu Betung (Dendrocalamus asper), BambuHitam.Tercatat : 90 jenis burung (sebagian dilindungi Undang-undang di Indonesia)Burung khas (dominan):Elang Hitam (Ictinaetus malayensis), Elang Ruyuk (Spilornischeela), Manuk Paok (Pitta guajana), Manuk Sapu (Riphidurajavanica), Manuk Madu (Arachnothera braziliensis), ManukMadu Beureum (Aetopyga mystacalis), dan Klaces(Arachnothera longirostra), Gagak, Jalak, Kacer,Cangkurileung, Bincarung.Tercatat : 14 jenis mammalia (dilindungi Undang-undang diIndonesia)Mammalia khas (dominan):Surili (Presbytis aygularis), Owa/Kueung (Hylobates moloch),Landak (Hystrix javanica), Sigung (Mydaus javanensis),Peusing (Manis javanica), Peucang (Tragulus javanicus),Monyet Ekor Panjang, Kukang (Nycticebus coucang) danMencek (Mutiacus muntjak), kucing hutan, macan tutul,harimau (dikatakan sebagai hewan ghaib yang dapat terlihatmaupun tidak)10 jenis hewan melataUlar Kobra, Ular Tanah, Ular Pohon, Biawak
KESIMPULAN
1. Masyarakat adat memiliki kearifan lokaldalam memanfaatkan dan memeliharalingkungan hidupnya sehingga tetap mampubertahan hingga saat ini.
2. Masyarakat adat baduy terbagi menjadi duayaitu Baduy dalam dan luar dengan berbagaikarakteristik yang menyertainya.
3. Kearifan Lokal tertuang dalam beberapaaturan adat dan pikukuhkaruhun sebagaipedoman hidupnya dengan segala sanksi adatyang diberlakukan terhadap seluruhmasyarakatadat baduy. Kearifan lokal inidipegang teguh oleh masyarakat adat BaduyDalam.
4. Masyarakat adat Baduy membagi dalambeberapa zonasi dalam pemanfaatan sumberdaya hayati. Zona inti adalah hutan sakralyang dikeramatkan sehingga tidak boleh samasekali memanfaatkan SDA nya. Zonapenyangga merupakan wilayah hutan yangboleh dimanfaatkan Sumber daya alamnyadengan tetap memegang teguh pada pikukuhkaruhun
Daftar Pustaka
Daryanto dan A. Suprihatin. 2013. PengantarPendidikan Lingkungan Hidup. Gava media.Yogyakarta.
Erwinantu. 2012. Saba baduy. Sebuah PerjalananWisata Budaya Inspiratif. Gramedia.
Soedjito, H. Y. Purwanto,. E. Sukara (eds). Situskeramat alami: Peran Budaya DalamKonservasi Keanekaragaman hayati.Yayasan obor Indonesia. Jakarta.
Koentjaraningrat. 2009. Pengantar IlmuAntopologi. Rineka Cipta. Jakarta.
Kurnia, A. dan A. Sihabudin. 2010. Saatnyabaduy Bicara. Bumi Aksara bekerjasamadengan Untirta. Jakarta.
Moleong. L.J. 2004. Metodologi PenelitianKualitatif. Edisi Revisi. PT. RemajaRosdakarya. Bandung.
Rachman, M. 2012. Konservasi Nilai dan warisanBudaya. Indonesian Journal of Conservation.
Senoaji, G. 2011. Perilaku Masyarakat Baduydalam Mengelola hutan, lahan danlingkungan di Banten Selatan. JurnalHumaniora 23 (1):14-25
Senoaji, G. 2011. Pengelolaan Hutan dan SistemAgroforestyoleh masyarakat Baduy diBanten Selatan. Jurnal Bumi lestari 12(2):283—293
Sutendy, U. 2010. Kearifan Hidup Orang baduy.Damai dengan Alam. Media Komunika.Tangerang.
Gambar 7. Jenis tegakkan pohon khas Huta Tua (HutanLindung) [Sumber: Dokumentasi pribadi diambil dariperbatasan zona tengah]
Enggar Utari 51
habitat satwa liar, (3) fungsi konservasi tanahdan hidrologi sungai, (4) fungsi iklim mikro,(5) fungsi sosial ekonomi, untuk pengambilananeka ragam hasil hutan.
Tabel 4. Hasil pencatatan jenis keanekaragaman hayatidi Hutan Lindung
Tercatat : 165 jenis tumbuhanTumbuhan khas (dominan):Keranji, Pisitan, Durian, Picung (Tumbuhan liar yangmenghasilkan buah untuk dimanfaatkan), Kayu Garu, Kiara,Huru, Leles, Rasamala(kayu langka), Leungsir, Kalimborot,Tapos, Lolot, Kaneungay, Kibuluh, Haraghag, Jirak, Hantap,Puspa, Kileho, Bubuay, Putat, Trembeusi, Paku Kapal,Bayur/Cayur (Pterosperum javanicum), Burahol atau Turalak(Stechocarpus buraholhook), Caringin/Beringin (Ficusbenyamin lnn), Kiara (Ficus Indica), Kianggir (Otoporaspectabilis), Kokosan atau Pisitan (Lansium domesticum),Jeunjing (Albizzia fakata backer), Rotan (Korthalisia laciosa),Tubaleleur atau Areuy Kawao (Milletia servicea), Kedoya(Dysoxylum caulostachym miq), Bambu Apus (Gigantocholaapus kurz), Bambu Betung (Dendrocalamus asper), BambuHitam.Tercatat : 90 jenis burung (sebagian dilindungi Undang-undang di Indonesia)Burung khas (dominan):Elang Hitam (Ictinaetus malayensis), Elang Ruyuk (Spilornischeela), Manuk Paok (Pitta guajana), Manuk Sapu (Riphidurajavanica), Manuk Madu (Arachnothera braziliensis), ManukMadu Beureum (Aetopyga mystacalis), dan Klaces(Arachnothera longirostra), Gagak, Jalak, Kacer,Cangkurileung, Bincarung.Tercatat : 14 jenis mammalia (dilindungi Undang-undang diIndonesia)Mammalia khas (dominan):Surili (Presbytis aygularis), Owa/Kueung (Hylobates moloch),Landak (Hystrix javanica), Sigung (Mydaus javanensis),Peusing (Manis javanica), Peucang (Tragulus javanicus),Monyet Ekor Panjang, Kukang (Nycticebus coucang) danMencek (Mutiacus muntjak), kucing hutan, macan tutul,harimau (dikatakan sebagai hewan ghaib yang dapat terlihatmaupun tidak)10 jenis hewan melataUlar Kobra, Ular Tanah, Ular Pohon, Biawak
KESIMPULAN
1. Masyarakat adat memiliki kearifan lokaldalam memanfaatkan dan memeliharalingkungan hidupnya sehingga tetap mampubertahan hingga saat ini.
2. Masyarakat adat baduy terbagi menjadi duayaitu Baduy dalam dan luar dengan berbagaikarakteristik yang menyertainya.
3. Kearifan Lokal tertuang dalam beberapaaturan adat dan pikukuhkaruhun sebagaipedoman hidupnya dengan segala sanksi adatyang diberlakukan terhadap seluruhmasyarakatadat baduy. Kearifan lokal inidipegang teguh oleh masyarakat adat BaduyDalam.
4. Masyarakat adat Baduy membagi dalambeberapa zonasi dalam pemanfaatan sumberdaya hayati. Zona inti adalah hutan sakralyang dikeramatkan sehingga tidak boleh samasekali memanfaatkan SDA nya. Zonapenyangga merupakan wilayah hutan yangboleh dimanfaatkan Sumber daya alamnyadengan tetap memegang teguh pada pikukuhkaruhun
Daftar Pustaka
Daryanto dan A. Suprihatin. 2013. PengantarPendidikan Lingkungan Hidup. Gava media.Yogyakarta.
Erwinantu. 2012. Saba baduy. Sebuah PerjalananWisata Budaya Inspiratif. Gramedia.
Soedjito, H. Y. Purwanto,. E. Sukara (eds). Situskeramat alami: Peran Budaya DalamKonservasi Keanekaragaman hayati.Yayasan obor Indonesia. Jakarta.
Koentjaraningrat. 2009. Pengantar IlmuAntopologi. Rineka Cipta. Jakarta.
Kurnia, A. dan A. Sihabudin. 2010. Saatnyabaduy Bicara. Bumi Aksara bekerjasamadengan Untirta. Jakarta.
Moleong. L.J. 2004. Metodologi PenelitianKualitatif. Edisi Revisi. PT. RemajaRosdakarya. Bandung.
Rachman, M. 2012. Konservasi Nilai dan warisanBudaya. Indonesian Journal of Conservation.
Senoaji, G. 2011. Perilaku Masyarakat Baduydalam Mengelola hutan, lahan danlingkungan di Banten Selatan. JurnalHumaniora 23 (1):14-25
Senoaji, G. 2011. Pengelolaan Hutan dan SistemAgroforestyoleh masyarakat Baduy diBanten Selatan. Jurnal Bumi lestari 12(2):283—293
Sutendy, U. 2010. Kearifan Hidup Orang baduy.Damai dengan Alam. Media Komunika.Tangerang.
Gambar 7. Jenis tegakkan pohon khas Huta Tua (HutanLindung) [Sumber: Dokumentasi pribadi diambil dariperbatasan zona tengah]
Enggar Utari 51
habitat satwa liar, (3) fungsi konservasi tanahdan hidrologi sungai, (4) fungsi iklim mikro,(5) fungsi sosial ekonomi, untuk pengambilananeka ragam hasil hutan.
Tabel 4. Hasil pencatatan jenis keanekaragaman hayatidi Hutan Lindung
Tercatat : 165 jenis tumbuhanTumbuhan khas (dominan):Keranji, Pisitan, Durian, Picung (Tumbuhan liar yangmenghasilkan buah untuk dimanfaatkan), Kayu Garu, Kiara,Huru, Leles, Rasamala(kayu langka), Leungsir, Kalimborot,Tapos, Lolot, Kaneungay, Kibuluh, Haraghag, Jirak, Hantap,Puspa, Kileho, Bubuay, Putat, Trembeusi, Paku Kapal,Bayur/Cayur (Pterosperum javanicum), Burahol atau Turalak(Stechocarpus buraholhook), Caringin/Beringin (Ficusbenyamin lnn), Kiara (Ficus Indica), Kianggir (Otoporaspectabilis), Kokosan atau Pisitan (Lansium domesticum),Jeunjing (Albizzia fakata backer), Rotan (Korthalisia laciosa),Tubaleleur atau Areuy Kawao (Milletia servicea), Kedoya(Dysoxylum caulostachym miq), Bambu Apus (Gigantocholaapus kurz), Bambu Betung (Dendrocalamus asper), BambuHitam.Tercatat : 90 jenis burung (sebagian dilindungi Undang-undang di Indonesia)Burung khas (dominan):Elang Hitam (Ictinaetus malayensis), Elang Ruyuk (Spilornischeela), Manuk Paok (Pitta guajana), Manuk Sapu (Riphidurajavanica), Manuk Madu (Arachnothera braziliensis), ManukMadu Beureum (Aetopyga mystacalis), dan Klaces(Arachnothera longirostra), Gagak, Jalak, Kacer,Cangkurileung, Bincarung.Tercatat : 14 jenis mammalia (dilindungi Undang-undang diIndonesia)Mammalia khas (dominan):Surili (Presbytis aygularis), Owa/Kueung (Hylobates moloch),Landak (Hystrix javanica), Sigung (Mydaus javanensis),Peusing (Manis javanica), Peucang (Tragulus javanicus),Monyet Ekor Panjang, Kukang (Nycticebus coucang) danMencek (Mutiacus muntjak), kucing hutan, macan tutul,harimau (dikatakan sebagai hewan ghaib yang dapat terlihatmaupun tidak)10 jenis hewan melataUlar Kobra, Ular Tanah, Ular Pohon, Biawak
KESIMPULAN
1. Masyarakat adat memiliki kearifan lokaldalam memanfaatkan dan memeliharalingkungan hidupnya sehingga tetap mampubertahan hingga saat ini.
2. Masyarakat adat baduy terbagi menjadi duayaitu Baduy dalam dan luar dengan berbagaikarakteristik yang menyertainya.
3. Kearifan Lokal tertuang dalam beberapaaturan adat dan pikukuhkaruhun sebagaipedoman hidupnya dengan segala sanksi adatyang diberlakukan terhadap seluruhmasyarakatadat baduy. Kearifan lokal inidipegang teguh oleh masyarakat adat BaduyDalam.
4. Masyarakat adat Baduy membagi dalambeberapa zonasi dalam pemanfaatan sumberdaya hayati. Zona inti adalah hutan sakralyang dikeramatkan sehingga tidak boleh samasekali memanfaatkan SDA nya. Zonapenyangga merupakan wilayah hutan yangboleh dimanfaatkan Sumber daya alamnyadengan tetap memegang teguh pada pikukuhkaruhun
Daftar Pustaka
Daryanto dan A. Suprihatin. 2013. PengantarPendidikan Lingkungan Hidup. Gava media.Yogyakarta.
Erwinantu. 2012. Saba baduy. Sebuah PerjalananWisata Budaya Inspiratif. Gramedia.
Soedjito, H. Y. Purwanto,. E. Sukara (eds). Situskeramat alami: Peran Budaya DalamKonservasi Keanekaragaman hayati.Yayasan obor Indonesia. Jakarta.
Koentjaraningrat. 2009. Pengantar IlmuAntopologi. Rineka Cipta. Jakarta.
Kurnia, A. dan A. Sihabudin. 2010. Saatnyabaduy Bicara. Bumi Aksara bekerjasamadengan Untirta. Jakarta.
Moleong. L.J. 2004. Metodologi PenelitianKualitatif. Edisi Revisi. PT. RemajaRosdakarya. Bandung.
Rachman, M. 2012. Konservasi Nilai dan warisanBudaya. Indonesian Journal of Conservation.
Senoaji, G. 2011. Perilaku Masyarakat Baduydalam Mengelola hutan, lahan danlingkungan di Banten Selatan. JurnalHumaniora 23 (1):14-25
Senoaji, G. 2011. Pengelolaan Hutan dan SistemAgroforestyoleh masyarakat Baduy diBanten Selatan. Jurnal Bumi lestari 12(2):283—293
Sutendy, U. 2010. Kearifan Hidup Orang baduy.Damai dengan Alam. Media Komunika.Tangerang.
Gambar 7. Jenis tegakkan pohon khas Huta Tua (HutanLindung) [Sumber: Dokumentasi pribadi diambil dariperbatasan zona tengah]

ISSN 978-602-14989-0-3 BioETI
Interaksi interspesies tiga jenis Kuntul (Ardeidae) di Cagar AlamBaringin Sati, Sumatera BaratFAUZIAH1), RIZALDI2) DAN WILSON NOVARINO3)
1)Jurusan Biologi, FMIPA, Universitas Andalas, Kampus UNAND Limau Manis, Padang, 251632)Labor Riset Ekologi, Jurusan Biologi, FMIPA, Universitas Andalas, Kampus UNAND Limau Manis, Padang, 251633)Museum Zoologi, Jurusan Biologi, FMIPA, Universitas Andalas, Kampus UNAND Limau Manis, Padang, 25163E-mail: [email protected]
ABSTRAKPenggunaan sumber daya yang sama diduga memicu terjadinya interaksi interspesies yang cukup tinggi. Salah satupenggunaan sumber daya yang sama oleh jenis yang berbeda terjadi di Cagar Alam (CA) Baringin Sati. Penelitian inibertujuan untuk mengetahui intensitas interaksi antara tiga jenis burung Kuntul (Ardea purpurea, Bubulcus ibis, danEgretta garzetta), yang memanfaatkan pohon Beringin di CA Baringin Sati secara bersama. Penelitian dilakukan padaJuni hingga Agustus 2014 di CA Baringin Sati dengan total waktu pengamatan 150 jam. Pengamatan dilakukan denganmenggunakan metoda scan sampling terhadap seluruh koloni dan pencatatan secara instantaneous recording denganpengambilan poin pengamatan setiap lima menit, selama enam jam setiap harinya yang dibedakan atas tiga sesi; pagi,siang, dan sore hari. Pengamatan dilakukan satu arah dan mengasumsikan tidak terjadinya pengamatan berulang terhadapindividu yang sama. Interaksi interspesies yang teramati adalah Displacement dan Agonistik. Interaksi lebih sering terjadipada sore hari. Jenis yang paling sering melakukan interaksi interspesies adalah Ardea purpurea dengan Egretta garzetta.
Key words: Interaksi, interspesies, Ardeidae, scan sampling, instantaneous recording
Pendahuluan
Pulau Sumatera, sebagai salah satu darikepulauan Sunda Besar, memiliki banyak CagarAlam untuk mengamati keindahan burung.Salah satunya adalah Cagar Alam Baringin Sati.Cagar Alam ini merupakan cagar alam terkecildi dunia dengan luas hanya 300 meter persegi(BKSDA, 2012). Cagar Alam ini hanya berupapohon Ficus benjamina berumur ratusan tahunyang berdaun rimbun dan dilindungi olehpemerintah setempat. Ficus benjamina ataudikenal sebagai pohon beringin termasuk kedalam famili Moraceae dan sering dimanfaatkanoleh beberapa jenis burung hantu, kutilang,rangkong, dan ayam-ayaman untuk bersarang dihutan (Aniger dan Hasyim, 1985).
Keunikan Cagar Alam Baringin Sati adalahhadirnya koloni ratusan ekor burung kuntul yangmenjadikan pohon beringin sebagai satu-satunyatempat bertengger dan bersarang. Berdasarkanhasil survei pendahuluan, diketahui bahwakoloni kuntul yang menghuni Cagar AlamBaringin Sati terdiri dari tiga spesies kuntul,yaitu Cangak Merah (Ardea purpurea), Kuntul
Kerbau (Bubulcus ibis), dan Kuntul Kecil(Egretta garzetta). Menurut Gause (1934),kebiasaan hidup bersama dalam koloni yangcukup besar ini memberikan dampak yangsangat berpengaruh terhadap perilaku sehari-hariburung. Ketika individu dari spesies yang samamaupun berbeda memanfaatkan sumber dayayang sama, maka akan timbul interaksi antarindividu dalam koloni. Setiap interaksi yangterjadi baik intraspesies maupun interspesiesakan memberikan dampak positif dan negatifterhadap masing-masing individu. Interaksiyang terjadi dapat dilihat dari wilayah yangdijadikan teritori oleh masing-masing spesies.
Kemungkinan adanya aktivitas yangmenimbulkan interaksi interspesies kuntul yanghidup di Cagar Alam Baringin Sati besertaadanya dominansi jenis tertentu dari ketiga jeniskuntul yang hanya menempati satu pohonberingin secara bersama dalam jumlah yangbanyak merupakan hal yang menarik untukdipelajari.
Adapun tujuan dari penelitian ini adalahuntuk mengetahui jenis interaksi interspesiesyang terjadi antara ketiga jenis kuntul (Ardea

Fauziah, Rizaldi dan Wilson Novarino 53
purpurea, Bubulcus ibis, dan Egretta garzetta)yang menempati pohon Beringin di CA.Baringin Sati dan mengetahui dominansi spesies.
Burung dalam ordo Ciconiiformes terbagimemiliki ciri khas rentang sayap lebar, kakilurus panjang, serta paruh yang menunjukkankelompok ini sebagai pemakan hewan kecil diperairan. Salah satu famili dalam ordo ini, yaitufamili Ardeidae memiliki leher yang bisamelengkung atau dilipat membentuk simpul “S”ketika terbang. Cangak Merah (Ardea purpurea),Kuntul Kerbau (Bubulcus ibis), dan KuntulKecil (Egretta garzetta) termasuk ke dalamfamili Ardeidae ditandai oleh kaki dan leheryang panjang, serta paruh yang panjang danlurus (MacKinnon, et al., 2010).
Ardea purpurea, lebih dikenal sebagaiCangak Merah, berukuran besar dengan tinggirata-rata 80 cm, berwarna abu-abu, coklatberangan, dan hitam. Pada bagian atas kepalaterdapat topi hitam dengan jambul menjuntaidengan setrip hitam menurun sepanjang leher.Ketika terbang posisi leher ditekuk. Punggungdan penutup sayap berwana abu-abu, sementarabulu terbang hitam dan bulu lainnya coklatkemerahan. Bagian iris kuning berwarna, paruhcoklat, dan kaki coklat kemerahan. Cangak inibiasa mengeluarkan bunyi “Uak” yang keras(Ayat, 2011).
Berbeda dari Ardea purpurea yang warnabulunya didominasi oleh abu-abu, Egrettagarzetta dan Bubulcus ibis justru memiliki buluputih bersih di seluruh tubuhnya. Bubulcus ibismerupakan spesies kuntul yang sering kitajumpai di areal persawahan dan berkeliarandekat kerbau, sehingga dikenal juga dengansebutan Kuntul Kerbau. Perbedaannya terletakpada warna paruh kuning pada Bubulcus ibisdan hitam pada Egretta garzetta (Ayat, 2011;MacKinnon, et al., 2010).
Burung beraktivitas dalam kesehariannyadalam dua fungsi yaitu tingkah laku perawatandiri (self-maintanance behavior) seperti makan,menelisik, dan mandi yang bertujuan untukmerawat kondisi fisik individu, dan tingkah lakusosial (social behavior) yang bertujuan
menyampaikan informasi kepada individu lainseperti bersuara dan kawin. Terkadang, batasantara dua kategori tingkah laku ini menjadikabur. Sebagai contohnya, makan yang awalnyatermasuk ke dalam kategori tingkah lakuperawatan diri, yang kemudian terpengaruh olehinteraksi sosial ketika spesies tersebut makansecara bersama dalam sebuah koloni (Sibley,2001).
Semua tingkah laku bersifat adaptif. Tingkahlaku pengabaian-penyerangan (aggressiveneglect) merupakan kecendrungan satu spesiesuntuk mengabaikan sarang atau anak agartampak menghasilkan sikap menyerang yangberlebihan terhadap spesies lain yang hadir.Tingkah laku agonistik (agonistic behavior)didefenisikan sebagai semua bentuk tingkahlaku yang muncul ketika terjadi konflik antarhewan, termasuk perkelahian dan kabur (vanTyne dan Berger, 1975).
Pemanfaatan sumber makanan maupuntempat bertengger yang sama pastimenimbulkan interaksi. Gause (1934)menyatakan bahwa kompetisi banyak terjadipada hewan yang bersarang pada satu tempatyang sama. Kompetisi ini dapat terjadi dalamspesies yang sama, disebut intraspesies maupunterjadi antara spesies yang berbeda, disebutinterspesies. Dalam kondisi dimana burungbertengger bersama dalam satu pohon (mixed-flock) kompetisi interspesies biasanya akan lebihbesar dibandingkan dengan intraspesies.
Menurut Wiens (1989), penjelasan yangseringkali diberikan tentang pola komunitasadalah bahwa mereka merupakan hasil darikompetisi interspesies. Sebuah tantangan yangmengundang kontroversi dalam ekologikomunitas terpusat pada ada atau tidaknyakompetisi menghasilkan pola yang kita lihat danseberapa aplikatif sebuah teori tentangkompetisi secara alami. Perbedaan antaramunculnya kompetisi antar individu danefeknya terhadap individu, populasi, ataukomunitas tergambarkan dalam perbedaanantara intensitas kompetisi dan pentingnyakompetisi. Kompetisi bisa saja terjadi secara

Fauziah, Rizaldi dan Wilson Novarino 54
intensif tetapi tidak relatif penting apabilakesuksesan bertahannya individu atau komunitastergantung kepada faktor lain yang lebih besar.Kebalikannya, kompetisi bisa jadi tidak terlalusering muncul, tetapi hasilnya memberikandampak yang sangat besar terhadap ketahananindividu dalam populasi atau ruang relung yangditempati oleh suatu spesies.
BAHAN DAN METODE
Alat-alat yang digunakan dalam penelitian iniadalah binokuler Nikon, kamera Nikon D-40,stopwatch, dan alat tulis. Metode yangdigunakan dalam penelitian ini adalah scansampling terhadap populasi Ardea purpurea,Egretta garzetta, dan Bubulcus ibis kemudiandilakukan pencatatan aktivitas tingkah lakudengan metode pencatatan instantaneousrecording (Martin dan Bateson, 1993).Pengamatan burung kuntul dilakukan denganmenggunakan teropong. Waktu pengamatandibagi 3 sesi pengamatan, yaitu pagi (06.30-08.30), siang (11.00-13.00), dan sore (16.30-18.30). Setiap sesi pengamatan dibagi ke dalamtitik pengamatan dalam 1 x 5 menitinstantaneous recording dengan lamapengamatan 60 detik. Scan sampling dilakukandengan mengamati seluruh jenis, denganmenghitung jumlah individu, mengamatitingkah laku dan interaksi yang terjadi, danpreferensi tempat berlangsungnya aktivitastersebut. Preferensi tempat diamati denganmembagi pohon menjadi kanopi luar dan dalam.Kanopi luar adalah apabila aktivitas kuntulterlihat jelas dari arah luar kanopi tanpahambatan oleh dedaunan atau cabang pohon dibagian luar pohon. Sedangkan kanopi dalammerupakan kondisi aktivitas kuntul terlihat daricelah-celah daun atau cabang pohon. Kategoritingkah laku yang diamati dari hewan tersebutadalah (dimodifikasi dari Khuslan, 2011):1. Aktivitas menelisik (Grooming) (G)
Aktivitas menelisik bulu dengan paruh ataudengan kaki, dan menggesek-gesekkan paruh.
2. Aktivitas pergantian tempat (Displacement)(D)
Peristiwa berpindah tempat satu individu keposisi lain karena kedatangan individu lainyang mengambil alih posisi semula.
3. Aktivitas agonistik (A)Semua kejadian yang termasuk kategorimengancam atau menyerang sepertimembuka paruh, saling mengadu paruhsambil terkadang mengepakkan sayapkemudian naik ke punggung individu lawan,dan memburu.
4. Aktivias seksual (S)Aktivitas kopulasi atau kawin antar duaindivdu berbeda jenis.
5. Aktivitas bersarang (N)Aktivitas di sarang seperti membawa rantingdari luar ke sarang, merapikan sarang, ataumemberikan ranting kepada pasangan yangmenunggu di sarang.
6. Aktivitas istirahat (R)Posisi tegak atau menekuk leher.
7. Aktivitas berpindah tempat (M)Berpindah dari suatu tempat ke tempat lain
dalam pohon yang sama, tanpa stimulasiakibat kedatangan individu lain.
8. Aktivitas makan (F)Aktivitas mengambil makanan dengan paruhkemudian menelannya.
9. Courtship display (C)Aktivitas atraksi seperti memamerkan buludi hadapan individu lawan jenis.Dalam pencatatan tingkah laku dibedakanketika interaksi yang terjadi berupaintraspesifik atau interspesifik.Data yang didapatkan selama pengamatanditerangkan secara deskriptif dan dianalisadengan cara menghitung indeks aktivitasmasing-masing jenis dengan kategori tingkahlaku dan jenis interaksinya, dengan rumussebagai berikut:
Indeks aktivitas suatu sesi =∑Keterangan :n : Hari pengamatan (1-25)m : Poin pengamatan (1-24)Indeks aktivitas dibandingkan antar tiga sesi pengamatansehingga didapatkan pada sesi kapan dan aktivitas apayang sangat intens dilakukan dan oleh jenis apa.

Fauziah, Rizaldi dan Wilson Novarino 55
HASIL DAN PEMBAHASAN
Interaksi interspesies yang terjadi antara ketigajenis kuntul di pohon Beringin, CA. BaringinSati adalah agonistik dan displacement.Agonistik lebih banyak dilakukan pada sore harioleh Egretta garzetta dengan rata-rata 9% daritotal individunya yang beraktivitas agonistik,menyusul Ardea purpurea (5%) dan Bubulcusibis (1%) (Gambar 1). Pola serupa jugaditemukan pada pagi hari dan siang hari. Padasiang hari, jumlah rata-rata individu Ardeapurpurea dan Bubulcus ibis sama.
0.0000
0.0100
0.0200
0.0300
0.0400
0.0500
0.0600
0.0700
0.0800
0.0900
0.1000
sesi 1 sesi 2 sesi 3
Inde
ks R
ata-
rata
Jum
lah
Indi
vidu
Sesi pengamatan
Ardea purpurea
Bubulcus ibis
Egretta garzetta
Gambar 1. Indeks Rata-rata Individu Tiga JenisKuntul dalam Aktivitas Agonistik
Tingginya persentase individu Egretta garzettayang melakukan aktivitas agonistik pada setiapsesi menunjukkan bahwa Egretta garzettamerupakan jenis yang paling agresif di antaraketiga jenis kuntul. Pada sore hari menunjukkanbahwa antara individu-individu Egretta garzettadan Ardea purpurea mengalami interaksi yangcukup besar dalam upaya mendapatkan tempatuntuk bertengger setelah kembali ke sarang.
Agonistik merupakan aktivitas dimanaindividu menunjukkan kewaspadaan, menyerangatau bertahan dari serangan individu lainnya.Dalam hal ini, kuntul biasanya melakukannyadengan cara menghadapkan bagian paruh luruhke depan lawan menunjukkan sikap pertahananakan daerah teritori (Kushlan, 2013).
Tingginya presentase ini mengikutibanyaknya jumlah individu, sehingga semakinbanyak individu maka semakin tinggipersentasenya. Jumlah individu Egretta garzettalebih banyak dibandingkan individu lainnyameskipun pada siang hari relatif sedikit karenabanyak individu yang meninggalkan sarang.Berbeda halnya dengan Egretta garzetta,individu Ardea purpurea terlihat jarangmeninggalkan sarang sehingga sangat jelasmemiliki titik-titik tertentu yang dikuasai padapohon. Ardea purpurea mengawasi titik-titikwilayahnya dengan ketat. Ketika ada individulain yang mecoba masuk dan mengambil alihtempatnya, Ardea purpurea akan menunjukkansikap menyerang balik dengan mencondongkanparuhnya ke arah lawan. Peristiwa ini sangatjelas terlihat ketika rombongan burung yangpulang ke sarang datang dan mengitari pohonuntuk mencari sarang.
Ardea purpurea memiliki ukuran tubuh yangbesar untuk mengancam Egretta garzettamaupun Bubulcus ibis yang memiliki ukurantubuh yang lebih kecil. Egretta garzettamemiliki jumlah yang banyak untuk melakukanagresi dan membutuhkan lebih banyak ruang,sementara Bubulcus ibis meskipun memilikiukuran tubuh yang relatif sama dengan Egrettagarzetta hanya berjumlah sedikit.
Beberapa dari aktivitas agonistik inikemudian berlanjut menjadi aktivitasdisplacement atau tidak. Aktivitas displacementdalam hal ini diartikan sebagai pindahnyaindividu karena kedatangan individu lain.Interaksi displacement tidak terlalu seringdilakukan terlihat dari persentase tertinggi padasore hari hanya 0,86% dari individu Egrettagarzetta (Gambar 2), kemudian menyusul Ardeapurpurea (0,44%) dan Bubulcus ibis (0,38%).Pola serupa juga terjadi pada sesi pagi.Perbedaannyaa dengan sesi siang hari adalahEgretta garzetta tetap lebih tinggi persentasenyakemudian diikuti oleh Bubulcus ibis.

Fauziah, Rizaldi dan Wilson Novarino 56
0.0000
0.0010
0.0020
0.0030
0.0040
0.0050
0.0060
0.0070
0.0080
0.0090
0.0100
sesi 1 sesi 2 sesi 3
Inde
ks Ra
ta-ra
ta Ju
mlah
Indiv
idu
Sesi pengamatan
Ardea purpureaBubulcus ibisEgretta garzetta
Gambar 2. Indeks rata-rata individu tiga jenisKuntul dalam aktivitas displacement
Individu yang datang bisa dikategorikansebagai individu yang lebih memiliki dominansidi kawasan tersebut. Namun, apabila individupertama melakukan perlawanan (agonistik) dankemudian dapat mempertahankan teritorinyamaka aktivitas displacement tidak terjadi yangmenunjukkan individu pertama memilikidominansi yang lebih tinggi.
Ardea purpurea memiliki ukuran tubuh yanglebih besar dibandingkan dengan Egrettagarzetta dan Bubulcus (MacKinnon, et.al, 2010).Ardea purpurea di CA Baringin Sati jarangmeninggalkan pohon dan kebiasaan inimemudahkan mereka untuk menguasai titik-titiktertentu pada pohon. Ukurannya yang besarkemudian memberikan dukungan untukmelakukan ancaman dan pengambil alihanposisi bertengger dari individu jenis lain.Sehingga, meskipun jumlahnya lebih sedikitdibandingkan Egretta garzetta, individu Ardeapurpurea tetap bisa mempertahankan daerahnya.
Bubulcus ibis memiliki tubuh yang hampirsama ukurannya dengan Egretta garzetta akantetapi dengan jumlah individu yang lebih sedikitdibandingkan dengan Egrettta garzetta danArdea purpurea. Oleh karena itu, individuBubulcus ibis tidak terlalu intens terlibat dalamkedua kategori inteaksi interspesies ini padapagi hari dimana masih banyak individu Egretta
garzetta yang belum meninggalkan sarang, dansore hari dimana burung mulai pulang kembalike sarang. Berbeda halnya dengan siang hari.Pada siang hari, individu Egretta garzetta hanyatertinggal sebagian kecil di pohon sementaraindividu Ardea purpurea tetap banyak. Olehkarena itu, interaksi antara Egretta garzetta danBubulcus ibis tinggi pada siang hari ketika duajenis ini terlihat seimbang baik dari segi ukuranmaupun jumlah.
Gill (2007) menyatakan bahwa peristiwatersebut termasuk ke dalam kompetisi dalampersaingan yang merupakan gabungan yangkompleks dari agresi dan menghindar, dandisebut dengan agonistik. Ketika burungmempertahankan sesuatu seperti pasangan,makanan, ataupun teritori, mereka cenderungmenghindari kontak langsung dan resiko terlukadengan menggunakan ancaman danpengendalian ketenangan. Telah disampaikansebelumnya bahwa pemanfaatan pohon secarabersama oleh koloni burung ini tentunya akanmenimbulkan adanya pengaturan akan daerahuntuk bertengger dan bersarang.
Kompetisi mungkin terjadi dengan intensitasyang cukup tinggi tetapi tidak relatif pentingjika fitness dari individu atau komunitasditentukan oleh faktor lain secara luas.Kebalikannya, kompetisi mungkin munculdalam intensitas yang rendah, akan tetapimenjadi sangat penting jika ini merupakan carautama yang mempengaruhi fitness individudalam populasi atau relung yang ditempati olehsatu spesies (Wiens, 1989). Kuntul yang hidupdi CA Baringin Sati memanfaatkan relung yangsama. Aktivitas yang dilakukan pada relungyang sama ini sangat berpengaruh terhadapketahanan hidup masing-masing individumaupun spesies. Kompetisi interspesiesmempengaruhi ukuran populasi saatmemanfaatkan sumber daya yang sama (Sibley,2001).
Interaksi agonistik memiliki persentase yangcukup signifikan dari persentase displacement.Ini menunjukkan bahwa meskipun saling serang,atau saling mengancam antar spesies sering

Fauziah, Rizaldi dan Wilson Novarino 57
terjadi, hanya sedikit yang berrlanjut menjadipengambil alihan tempat. Dengan demikian,dapat dikatakan bahwa masing-masing spesiestelah memiliki pengaturan tertentu pada pohonsehingga meminimialisir adanya kompetisibesar-besaran yang berdampak terhadapkeberadaan spesies tersebut di pohon.Pengaturan ini tampak melalui dominansi olehmasing-masing spesies. Ardea purpureamerupakan jenis yang paling dominan di CA.Baringin Sati. Dominansi ini sangat terlihat padasore hari ketika banyak individu Egrettagarzetta yang pulang ke sarang dan mencobauntuk mengambil alih tempat bertengger.Sementara antara Egretta garzetta dan Bubulcusibis tidak terlalu berpengaruh karena rendahnyainteraksi Bubulcus ibis dengan individu lainyang juga dipengaruhi oleh ukuran danjumlahnya lebih sedikit.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Interaksi interspesies yang teramati adalahDisplacement dan Agonistik. Interaksi lebihsering terjadi pada sore hari. Jenis yang palingsering melakukan interaksi interspesies adalahArdea purpurea dengan Egretta garzetta.
DAFTAR PUSTAKA
Aniger, A. dan Hasyim, Syed Z. 1985.Lingkungan Hidup Pohon Beringin SeriPedoman Pengamatan di Lapangan No. 3.PT. Aksara Indira Harapan.
Ayat A. 2011. Burung-burung Agroforest diSumatera. World Agroforestry Centre -ICRAF, SEA Regional Office. Bogor,Indonesia.
BKSDA. 2012. Buku Informasi KawasanKonservasi Balai KSDA Sumatera Barat.BKSDA Sumatera Barat. Padang.
Gause, G.F. 1934. The Struggle for Existence.The Williams & Wilkins Company.Baltimore.
Gill, Frank B. 2007. Ornithology 3rd ed. W.HFreeman and Company. New York.
Kushlan, J.A 2011. The Terminology ofCourtship, Nesting, Feeding, andMaintenance in Herons. [online]www.HeronConservation.org
MacKinnon, John, Philips, Karen dan van Balen,Bas. 2010. Burung-burung di Sumatera,Jawa, Bali, dan Kalimantan. LIPI. Bogor.
Martin, Paul dan Bateson, Patrick. 1993.Measuring Behaviour: An IntrocductoryGuide. Cambridge University Press.Cambridge.
Sibley, D.A. 2001. The Sibley Guide to Bird Lifeand Behavior. Chanticler Press, Inc. NewYork.
Van Tyne, Josselyn dan Berger, Andrew John.1975. Fundamentals of Ornithology. JohnWiley & Sons, Inc. USA.
Wiens, J.A. 1989. The Ecology of BirdCommunity Vol. 2 Processes and Variation.Cambridge University Press. Cambridge.

ISSN 978-602-14989-0-3 BioETI
Efek ekstrak etanol rimpang Temu Mangga (Curcuma manggaVal.) terhadap kadar gula darah dan kolesterol Mencit Putih(Mus musculus) jantan yang diinduksi Aloksan
FAUZUR RAHMI, EFRIZAL DAN RESTI RAHAYU
Laboratorium Riset Fisiologi Hewan, Jurusan Biologi FMIPA Universitas Andalas, Kampus UNAND Limau ManisPadang – 25163, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian ini dilakukan untuk mengetahui efek ekstrak etanol rimpang temu mangga terhadap kadar gula darah dankolesterol mencit putih jantan yang diinduksi aloksan. Rancangan yang digunakan pada penelitian ini adalah RancanganAcak Lengkap (RAL) yang terdiri dari lima perlakuan yaitu kontrol (P0 dan P1) dan perlakuan dengan pemberianbeberapa dosis ekstrak temu mangga 200 (P2); 400 (P3); dan 800 mg/kg bb (P4). Masing-masing perlakuan diuji sebanyakdua sampai empat kali ulangan dengan lama pemeliharaan selama 28 hari. Hasil penelitian ini menunjukkan bahwa dosisekstrak etanol rimpang temu mangga yang berbeda memberikan pengaruh yang nyata (p < 0,05) terhadap kadar guladarah dan kolesterol mencit putih jantan yang diinduksi aloksan. Nilai persentase penurunan kadar gula darah tertinggidiperoleh pada perlakuan P4 yaitu -69,71% pada hari ke 28. Sedangkan kadar rata-rata kolesterol yang terendah diperolehpada perlakuan P3 (98,00 mg/dL), kemudian diikuti dengan perlakuan P2 (106,50 mg/dL), P4 (109,33 mg/dL), P1 (135,00mg/dL), P0 (150,00 mg/dL).
Key words: temu mangga (Curcuma mangga Val.), kadar gula darah, kolesterol
Pendahuluan
Obat-obatan tradisional telah dipakai olehmasyarakat sejak zaman dahulu hinggasekarang. Dunia farmakologi modern saat inijuga semakin mengarahkan penelitiannyakepada penggunaan bahan-bahan alamiterutama yang berasal dari tumbuh-tumbuhan(Rukmana, 2004). Obat-obat alami memilikibeberapa kelebihan seperti tingkat bahaya yanglebih rendah dari pada obat-obatan kimia,penerimaan tubuh terhadap obat yang berasaldari tumbuh-tumbuhan lebih mudah (Muhlisah,1999) dan efek sampingnya dapat ditekanseminimal mungkin (Agromedia, 2008).
Menurut Winarti dan Nurdjannah (2005),berbagai tanaman rempah termasuk familiZingiberaceae (jahe-jahean) sudah lama dikenalmengandung komponen fitokimia yangberperan penting untuk pencegahan danpengobatan berbagai penyakit. Salah satutanaman tersebut adalah temu mangga(Curcuma mangga Val.). Temu manggamengandung senyawa antioksidan seperti
kurkuminoid, flavonoid dan polifenol (Sudewo,2004). Berdasarkan penelitian yang dilakukanoleh Susmiati et al., (2010), melalui pengujiansecara in vitro senyawa dalam temu manggadapat berperan sebagai penurun kadarkolesterol yang berlebih di dalam darah.
Saat ini, penyakit diabetes mellitus dankolesterol merupakan penyakit yang banyakdiderita oleh masyarakat diberbagai belahandunia. Berdasarkah laporan WHO (2008),terhitung di Indonesia terdapat 700 jiwa per100.000 penduduk meninggal dunia akibatpenyakit diabetes mellitus dan kardiovaskularyang pada umumnya diakibatkan olehmeningkatnya kadar gula darah dan kolesteroldalam darah.
Diabetes mellitus dicirikan oleh tingginyakadar gula darah (hiperglikemia) (Winarsi,2010). Diabetes yang tidak terkontrol dengankadar gula darah yang tinggi cenderungmeningkatkan kadar kolesterol di dalam tubuh(Pusat Jantung Nasional, 2011).

Fauzur Rahmi, Efrizal dan Resti Rahayu 59
Salah satu penyebab naiknya kadar guladarah adalah karena terjadinya kerusakan sel βpankreas yang berfungsi sebagai penghasilhormon insulin. Menurut Suarsana (2010),kerusakan sel β pankreas dapat disebabkan olehbeberapa faktor yaitu faktor genetik, infeksioleh kuman, faktor nutrisi, radikal bebas (stresoksidatif) dan zat diabetogenik (penyebabdiabetes). Salah satu zat diabetogenik yangdapat merusak pankreas adalah aloksan.Menurut Szkudelski (2001), aloksan di dalamtubuh mengalami metabolisme oksidasi reduksimenghasilkan radikal bebas yang dapatmerusak sel β pankreas. Efek diabetogenikaloksan dapat dicegah oleh senyawa penangkapradikal hidroksil (Studiawan dan Santosa,2005). Berdasarkan pemaparan tesebut,penelitian ini dilakukan untuk mengetahuikemampuan ekstrak etanol rimpang temumangga untuk menurunkan kadar gula darahdan kolesterol mencit yang diinduksi aloksan.
BAHAN DAN METODE
Penelitian ini menggunakan metodeeksperimen Rancangan Acak Lengkap berbedaulangan dengan perlakuan yaitu mencit normaldan mencit diinduksi aloksan sebagai kontrol(P0 dan P1), serta mencit diinduksi aloksanyang diberikan beberapa dosis ekstrak temumangga 200 (P2); 400 (P3); dan 800 mg/kg bb(P4) (Yuandani et al., 2011).
Mencit yang digunakan berumur 2,5-3bulan dengan berat badan berkisar 25-35 gram.Sebelum penginduksian aloksan, mencitdipuasakan terlebih dahulu selama 16 jam.Aloksan diinduksikan dengan dosis 150 mg/kgbb secara intraperitoneal kemudian mencitdiberi makan pellet dan minum yangmengandung gula 10% selama 2 hari setelahpenginduksian aloksan. Pada hari ke-2 danseterusnya, air gula 10% diganti dengan airbiasa, setelah 7 hari kadar gula darah mencitdiukur (Chairunnisa, 2011). Mencit dengankadar gula darah lebih dari 150 mg/dldinyatakan diabetes dan siap untuk digunakansebagai hewan percobaan.
Parameter yang diukur adalah kadar gula darahdan kadar kolesterol mencit. Kadar gula darahdiukur setelah 7, 14 dan 21 hari pemberianekstrak temu mangga menggunakanglucosemeter (Nesco multicheck) dan kadarkolesterol total diukur setelah pemberianekstrak selama 21 hari menggunakan metodeCHOD-PAP. Persentase perubahan kadar guladarah mencit (PGn-0) dihitung denganmenggunakan rumus berikut:Gn − G0G0 100%Keterangan:Gn : Kadar gula darah hari ke-n (mg/dL)G0 : Kadar gula darah hari ke-0 (setelah
penginduksian aloksan) (mg/dL)(Dimodifikasi dari Pujilestari dan Pratiwi,2009)Data persentase perubahan kadar gula darahdan kadar kolesterol total mencit dianalisamenggunakan uji ANOVA dan diuji lanjutdengan uji Duncan.
HASIL DAN PEMBAHASAN
Kadar gula darah setelah diinduksi aloksanHasil analisis sidik ragam terhadap perubahankadar gula darah mencit menunjukkan terdapatperbedaan kadar gula darah antara kelompokmencit yang tidak diinduksi aloksan dengankelompok mencit yang diinduksi aloksan(Tabel 1).
Berdasarkan Tabel 1, dapat dilihat bahwapada pengamatan hari ke-0 yaitu sebelumpenginduksian aloksan (Gn) rata-rata kadar guladarah mencit berada dalam rentang kadar guladarah normal. Menurut Malole dan Pramono(1989 cit Utami et al., 2009) rentang kadar guladarah mencit normal adalah 62 – 175 mg/dL.Setelah penginduksian aloksan (G0), rata-ratakadar gula darah mencit mengalamipeningkatan mencapai lebih dari 300 mg/dL.Menurut Szkudelski (2001), aloksan dapatmenaikkan kadar gula darah ketika zat tersebutdiinduksikan secara intravena, intraperitoneal,atau subkutan. Jika penginduksian aloksandilakukan secara intraperitoneal atau subkutan,

Fauzur Rahmi, Efrizal dan Resti Rahayu 60
Tabel 1. Perubahan kadar gula darah mencit setelah diinduksi aloksan
Perlakuan nRata-rata kadar gula darah(mg/dL) PG0 (%)Gn G0
P0 4 103,50 103,50 -0,96P1 2 120,50 486,00 304,60P2 2 111,50 339,50 199,88P3 3 94,67 377,00 346,76P4 3 112,00 404,00 252,99
Keterangan: Gn : rata-rata kadar gula darah mencit normal; G0 : rata-rata kadar gula darah mencitsetelah diinduksi aloksan; PG0: persentase perubahan kadar gula darah setelah diinduksi aloksan
Tabel 2. Perubahan kadar gula darah mencit setelah pemberian ekstrak temu mangga
Perlakuan n PG1 (%) PG2 (%) PG3 (%)
P0 4 -8,14a 6,54a -9,25a
P1 2 0,76a 1,39a 16,77a
P2 2 -19,93a -5,13a -4,75a
P3 3 -14,84a 7,58a -9,73a
P4 3 -51,00a -46,79a -69,71b
Keterangan: PG1: perubahan kadar gula darah setelah 7 hari pemberian ekstrak temu mangga; PG2 :perubahan kadar gula darah setelah 14 hari pemberian ekstrak temu mangga; PG2 : perubahan kadar guladarah setelah 21 hari pemberian ekstrak temu mangga
Tabel 3. Rata-rata kadar kolesterol total mencit setelah pemberian ekstrak temu mangga selama 21 hari
Perlakuan N Rata-rata kadar kolesterol (mg/dL)
P0 4 150,00a
P1 2 135,00ab
P2 2 106,50b
P3 3 98,00b
P4 3 109,33b
maka dosis penginduksian dijadikan 2 -3 kalilipat dari dosis yang diinduksikan secaraintravena. Szkudelski (1998) berhasilmeningkatkan kadar gula darah pada tikusWistar jantan dengan menginduksikan aloksandengan dosis 75 mg/kg bb secara intravena.
Kadar gula darah setelah pemberian ekstraketanol rimpang temu manggaData rata-rata kadar gula darah mencit setelahpemberian ekstrak etanol rimpang temu manggaselama 7, 14 dan 21 hari diperlihatkan padaGambar 1. Berdasarkan grafik yang ditunjukkanpada Gambar 1, terlihat bahwa kelompok mencit
perlakuan yang hanya diinduksi aloksan 150mg/kg bb (P1) tidak menunjukkan penurunankadar gula darah selam tiga minggu perlakuan.Sedangkan untuk kelompok perlakuan mencitdiabetes yang diberi ekstrak etanol rimpangtemu mangga dosis 200, 400 dan 800 mg/kg bbmemperlihatkan kadar gulad arah yangfluktuatif (naik turun) setelah pemberian ekstrakselama tujuh, 14 dan 21 hari, hal ini juga terjadipada kelompok mencit tanpa perlakuan aloksandan temu mangga. Turun naiknya kadar guladarah ini terjadi karena pola makan dan aktivitasfisik yang berbeda-beda pada masing-masingmencit. Menurut Tandra (2007), kadar gula
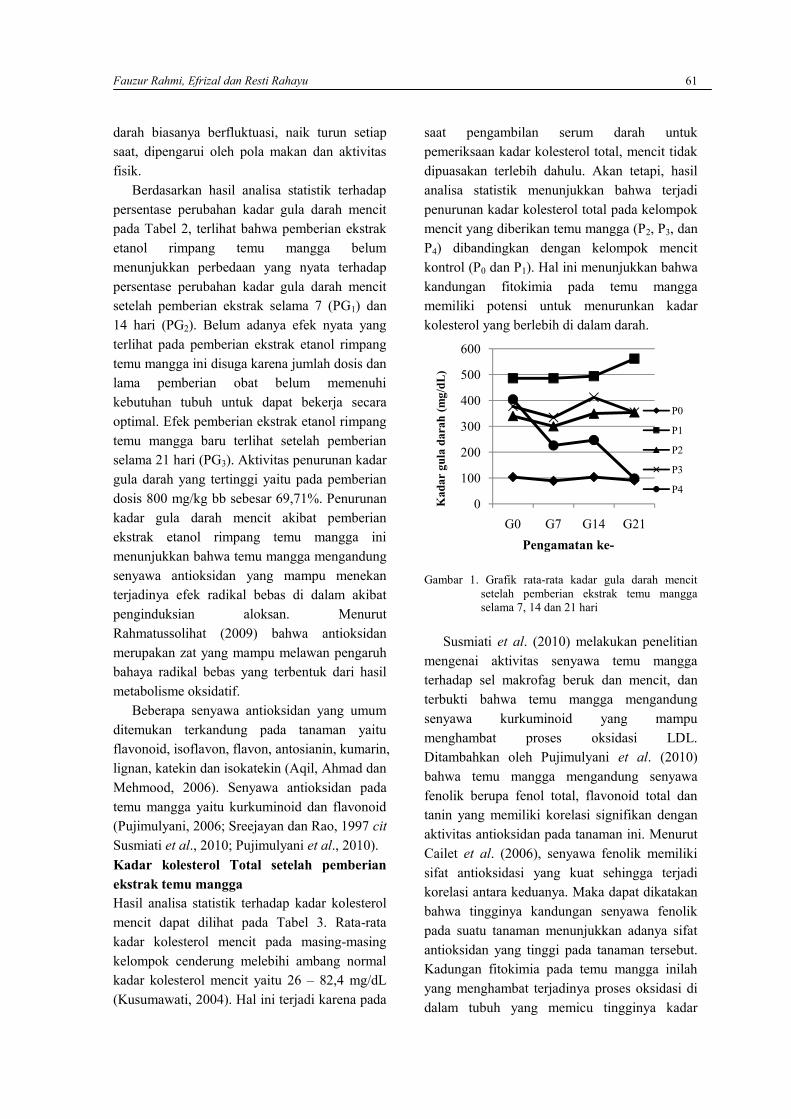
Fauzur Rahmi, Efrizal dan Resti Rahayu 61
darah biasanya berfluktuasi, naik turun setiapsaat, dipengarui oleh pola makan dan aktivitasfisik.
Berdasarkan hasil analisa statistik terhadappersentase perubahan kadar gula darah mencitpada Tabel 2, terlihat bahwa pemberian ekstraketanol rimpang temu mangga belummenunjukkan perbedaan yang nyata terhadappersentase perubahan kadar gula darah mencitsetelah pemberian ekstrak selama 7 (PG1) dan14 hari (PG2). Belum adanya efek nyata yangterlihat pada pemberian ekstrak etanol rimpangtemu mangga ini disuga karena jumlah dosis danlama pemberian obat belum memenuhikebutuhan tubuh untuk dapat bekerja secaraoptimal. Efek pemberian ekstrak etanol rimpangtemu mangga baru terlihat setelah pemberianselama 21 hari (PG3). Aktivitas penurunan kadargula darah yang tertinggi yaitu pada pemberiandosis 800 mg/kg bb sebesar 69,71%. Penurunankadar gula darah mencit akibat pemberianekstrak etanol rimpang temu mangga inimenunjukkan bahwa temu mangga mengandungsenyawa antioksidan yang mampu menekanterjadinya efek radikal bebas di dalam akibatpenginduksian aloksan. MenurutRahmatussolihat (2009) bahwa antioksidanmerupakan zat yang mampu melawan pengaruhbahaya radikal bebas yang terbentuk dari hasilmetabolisme oksidatif.
Beberapa senyawa antioksidan yang umumditemukan terkandung pada tanaman yaituflavonoid, isoflavon, flavon, antosianin, kumarin,lignan, katekin dan isokatekin (Aqil, Ahmad danMehmood, 2006). Senyawa antioksidan padatemu mangga yaitu kurkuminoid dan flavonoid(Pujimulyani, 2006; Sreejayan dan Rao, 1997 citSusmiati et al., 2010; Pujimulyani et al., 2010).Kadar kolesterol Total setelah pemberianekstrak temu manggaHasil analisa statistik terhadap kadar kolesterolmencit dapat dilihat pada Tabel 3. Rata-ratakadar kolesterol mencit pada masing-masingkelompok cenderung melebihi ambang normalkadar kolesterol mencit yaitu 26 – 82,4 mg/dL(Kusumawati, 2004). Hal ini terjadi karena pada
saat pengambilan serum darah untukpemeriksaan kadar kolesterol total, mencit tidakdipuasakan terlebih dahulu. Akan tetapi, hasilanalisa statistik menunjukkan bahwa terjadipenurunan kadar kolesterol total pada kelompokmencit yang diberikan temu mangga (P2, P3, danP4) dibandingkan dengan kelompok mencitkontrol (P0 dan P1). Hal ini menunjukkan bahwakandungan fitokimia pada temu manggamemiliki potensi untuk menurunkan kadarkolesterol yang berlebih di dalam darah.
Gambar 1. Grafik rata-rata kadar gula darah mencitsetelah pemberian ekstrak temu manggaselama 7, 14 dan 21 hari
Susmiati et al. (2010) melakukan penelitianmengenai aktivitas senyawa temu manggaterhadap sel makrofag beruk dan mencit, danterbukti bahwa temu mangga mengandungsenyawa kurkuminoid yang mampumenghambat proses oksidasi LDL.Ditambahkan oleh Pujimulyani et al. (2010)bahwa temu mangga mengandung senyawafenolik berupa fenol total, flavonoid total dantanin yang memiliki korelasi signifikan denganaktivitas antioksidan pada tanaman ini. MenurutCailet et al. (2006), senyawa fenolik memilikisifat antioksidasi yang kuat sehingga terjadikorelasi antara keduanya. Maka dapat dikatakanbahwa tingginya kandungan senyawa fenolikpada suatu tanaman menunjukkan adanya sifatantioksidan yang tinggi pada tanaman tersebut.Kadungan fitokimia pada temu mangga inilahyang menghambat terjadinya proses oksidasi didalam tubuh yang memicu tingginya kadar
0
100
200
300
400
500
600
G0 G7 G14 G21
Kad
ar g
ula
dara
h (m
g/dL
)
Pengamatan ke-
P0
P1
P2
P3
P4

Fauzur Rahmi, Efrizal dan Resti Rahayu 62
kolesterol akibat adanya radikal bebas daripenginduksian aloksan.
KESIMPULAN
Berdasarkan hasil penelitian yang dilakukanmengenai efek ekstrak etanol rimpang temumangga (C. mangga Val.) terhadap kadar guladarah dan kolesterol mencit putih yang diinduksialoksan, maka diperoleh kesimpulan sebagaiberikut : ekstrak etanol rimpang temu manggamemberikan pengaruh terhadap kadar guladarah dan kolesterol mencit yang diinduksialoksan. Pemberian ekstrak etanol rimpang temumangga dosis 800 mg/kg bb merupakakn dosisefektif untuk menurunkan kadar gula darahmencit yang diinduksi aloksan
DAFTAR PUSTAKA
AgroMedia. 2008. Buku Pintar Tanaman Obat.Agromedia Pustaka. Jakarta.
Caillet, S., S. Salmieri dan M. Lacroix. 2006.Valuation of Free Radical-ScavengingProperties Of Comercial Grape PhenolExtract by a Fast Colorimetric Method.Journal of Food Chemistry 95 (1) : 1-8.
Chairunnisa, R. 2011. Pengaruh Jumlah PastaTomat Terhadap Penurunan Kadar GulaDarah pada Mencit Diabetes. JurnalTeknologi Industri Pertanian.
Kusumawati, D. 2004. Bersahabat denganHewan Coba. Gajah Mada Press.Yogyakarta.Muhlisah, F. 1999. Temu-temuan dan Empon-empon Budi Daya danManfaatnya. Kanisius. Yogyakarta.
Pujimulyani, D. 2006. Sifat Oksidatif EkstrakKunir Putih (Curcuma mangga Val.)dengan Pelarut Aseton, Etanol atauMetanol. BIOTA 11 (1) : 14-19.
Pujimulyani, D., S. Raharjo, Y. Marsono dan U.Santoso. 2010. Aktivitas Antioksidan danKadar Senyawa Fenolik Kunir Putih(Curcuma mangga Val.) Segar dan SetelahBlanching. AGRITECH 30 (2) : 68-74.
Pusat Jantung Nasional. 2011. Kolesterol danDiabetes (Kencing Manis).http://www.pjnhk.go.id/content/view/3549/31/ Diakses Rabu, 25 September 2013.
Rukmana, R. 2004. Temu-temuan Apotik hidupdi Pekarangan. Yogyakarta. Kanisius.
Studiawan, H. dan M. H. Santosa. 2005. UjiAktivitas Kadar Glukosa Darah EkstrakDaun Eugenia polynatha pada Mencit yangDiinduksi Aloksan. Majalah KedokteranHewan. (21) : 62-65.
Suarsana, I. N., B. P. Priosoeryanto, M. Bintang,dan T. Wresdiyati. Profil Glukosa Darahdan Ultrastruktur Sel Beta Pankreas Tikusyang Diinduksi Senyawa Aloksan. JITV 15(2) : 118-123.
Sudewo, B. 2004. Tanaman Obat PopulerPenggempur Aneka Penyakit. AgromediaPustaka. Jakarta.
Susmiati, T., Sulistiyani, D. Sajuthi, dan L. K.Darusman. 2010. Kemampuan EkstrakTemu Mangga (Curcuma mangga) dalamMenghambat Proses Oksidasi Low DensityLipoprotein. Forum Pascasarjana 33 (1) :25-34.
Szkudelski, T. 2001. The Mechanism of Alloxanand Streptozotocin Action in B Cells ofThe Rat Pancreas. Physiological Research50 : 536-546.
Tandra, H. 2007. Segala sesuatu yang HarusAnda Ketahui Tentang Diabetes. GramediPustaka Utama. Jakarta.
Utami, E. T., R. Fitrianti, Mahriani, dan S.Fajariyah. 2009. Efek KondisiHiperglikemik terhadap Struktur Ovariumdan Siklus Estrus Mencit (Mus musculusL.). Jurnal Ilmu Dasar 10 (2) : 219-224.
WHO. 2008. Mortality: Cardiovascular Diseaseand Diabetes, Deaths per 100.000.apps.who.int/gho/data/?vid=2469#.Diakses Rabu, 30 Januari 2013.
Winarsi, H. 2010. Protein Kedelai & KecambahManfaatnya bagi Kesehatan. Kanisius.Yogyakarta.
Winarti, C., dan N. Nurdjanah. 2005. PeluangTanaman Rempah dan Obat SebagaiSumber Pangan Fungsional. Jurnal LitbangPertanian 24 (4) : 47-55.
Yuandani, A. Dalimunthe, P. A. Z. Hasibuan,dan A. W. Septama. 2011. Uji AktivitasAntikanker (Preventif dan Kuratif) EkstrakEtanol Temu Mangga (Curcuma manggaVal.) pada Mencit yang DiinduksiSiklofosfamid. Majalah KesehatanPharmaMedika 3 (2) : 255-259.

ISSN 978-602-14989-0-3 BioETI
Analisis kualitas spermatozoa Mencit (Mus musculus)pasca pemberian air seduhan KopiFEBRI SEMBIRING DAN HERBERT SIPAHUTAR
Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Negeri Medan,Jln. Willem Iskandar Psr. V Medan Estate, Medan, 20221, Sumatera UtaraE-mail: [email protected]
ABSTRACTThe Coffee is one of most favorite drinks in the world. But the effect of coffee on reproduction quality are still debated.The aims of this study was to describe the effects of coffee concentrate giving on sperm concentration of epididymis, andspem qualities, consisted of motility, viability, and normal morphology. Fourteen male mice which aged three monthswere grouped based on completely randomized design (n=7), consisted of control (0ml/day) and the coffee concentratetreatment (0,5ml/day) with a concentration equivalent for three cups of coffee in humans which converted to a mice. Eachgroup was treated for 36 days. Based on the data analysis using t-test (α= 0.05) showed that the coffee concentrate givinghas significantly increased the sperm qualities of epididymis, are motility, viability, and normal morphology, and numberof sperm concentration of epididymis (p<0.05).
Key words: coffee concentrate giving, sperm qualities, male mice
Pendahuluan
Kopi merupakan salah satu alternatif minumanpilihan yang sangat digemari masyarakat duniaselain teh. Terbukti dengan rata-rata konsumsikopi dunia per tahun periode 2000-2003mencapai 4,9 juta ton (Gardjito et al., 2011).Kemudian tercatat 136,5 juta kantung kopidikonsumsi pada tahun 2011 menurutOrganisasi Kopi Internasional. Angka itu naikdua persen dari total hitungan 2010.Kopikemudian terus berkembang hingga saat inimenjadi salah satu minuman paling populer didunia yang dikonsumsi oleh berbagai kalanganmasyarakat (US National Coffee Association,1999).
Penelitian yang dilakukan oleh Khazaei(2012) terhadap mencit yang diberikan kafein1% meningkatkan motilitas sperma. Hal yangsama dikemukakan oleh Špaleková et al. (2011),Dlugosz and Bracken (1992), dan Nawrot et al.(2002) dengan melihat peningkatan motilitasdan kualitas sperma terhadap kafein. Efekmerugikan lainnya terhadap reproduksi adalahmenurunnya produksi sperma dengan carakafein yang berlebihan akan meningkatkankadar hormon testosteron dari jumlah normal
kemudian menghambat spermatogenesis(Flexstaf, 2012; Handlesman 2000).
BAHAN DAN METODE
Tanah Hewan coba dalam penelitian ini adalahMencit (Mus musculus) jantan Strain DDWebster sebanyak 14 ekor. Bahan yangdigunakan adalah biji kopi robusta dari daerahSidikalang Sumatera Utara yang digiling duakali kemudian diseduh dengan air mendidih.
Penentuan dosis kopi dalam penelitianmenggunakan tabel perbandingan luaspermukaan tubuh hewan coba dimana dosismencit dengan berat badan 20 gram adalah0,0026 kali dosis manusia menurut caraLaurance dan Bacharach (1964). MenurutNational Coffee Association’s Survey (2003),masyarakat Amerika Serikat (US) rata-ratamengkonsumsi kopi sebanyak 3 cangkir per hari.Dalam 1 cangkir kopi terdapat 7 gram bubukkopi. Apabila 3 cangkir kopi yang dikonsumsimaka terdapat 21 gram kopi yang terlarut untukmanusia. Kemudian di-konversikan 21 gramkopi tersebut dengan hewan coba mencit,dimana 21 gram kopi tersebut dikalikan dengan0,0026 sehingga menjadi 0,0546 gram kopi.0,0546 gram kopi ini dilarutkan dalam air

Febri Sembiring dan Herbert Sipahutar 64
mendidih sebanyak 0,5 ml. 0,5 ml ini dipilihkarena daya konsumsi mencit terhadap air. Stokdibuat untuk 280 ekor mencit dan 1 ekornyadiberikan 0,0546 gram kopi. Sehingga total stokkopi bubuk yang dilarutkan adalah 15,28 gram.Begitu pula dengan pelarutnya sebesar 140 ml.Sehingga 15,28 gram kopi dilarutkan dalam 140ml air mendidih. Air seduhan kopi dipisahkandari ampasnya.Pemberian air seduhan kopidilakukan setelah suhu optimum. Stok diguna-kan untuk 4 hari dan disimpan dalampendingin.Setiap empat hari air seduhan kopiharus diganti.
Jenis penelitian yang digunakan adalaheksperimental menggunakan Rancangan AcakLengkap.Empat belas ekor mencit jantan dibagimenjadi dua kelompok, yaitu kelompok P0 danP1.Kelompok P0 sebagai kontrol dan kelompokP1 sebagai perlakuan.Pada kelompok perlakuan,hari pertama sampai hari ke 36 diberi airseduhan kopi secara oral.Pemberian kopi padasetiap mencit dilakukan secara bertahap.
Parameter pengamatan dalam penelitian iniadalah konsentrasi sperma epididimis, motilitas,viabilitas, dan morfologi spermatozoaepididimis.
Pada hari ke 37, mencit dibedah dan diambilepididimis.Pembuatan suspensi spermamenggunakan epididimis yang dimasukkan kewell plate berisi 250 µl NaCl 0,9 % bersuhu 37-40o C.. Kemudian epididimis dipotong dengangunting hingga halus dan diaduk dengan jarumpengaduk sehingga diperoleh suspensispermatozoa yang homogen (Ilyas, 1997;Nugraheni et al., 2003).
Suspensi spermatozoa dari epididimisditetesi pada kamar hitung haemocytometerNeubeur 2 slide kaca sebanyak 20 µl. Padahaematocytometer Neubeur diamatispermatozoa bergerak dan tidak bergerak(motilitas) (Nugraheni et al., 2003). Suspensispermatozoa pada slide kaca kemudianditeteskan Eosin Y 0.5% sebanyak 20 µl (1 : 1)dan ditutup cover glass. Diamati spermatozoahidup dan mati (viabilitas). Suspensi sperma-tozoa mati akan terwarnai merah bagian
kepalanya dan hidup yang tidak terwarnai /bening (Ilyas, 1997; Cao et al., 2011). Suspensispermatozoa pada slide kaca lainnya dibuatapusan spermatozoa dan dikeringkan.Selanjutnya ditambahkan alkolohol absolutkeatas apusan spermatozoa kering.Dibiarkan 10menit, kemudian dibuang alkohol absolt yangtersisa diatas apusan. Dikeringkan dan kemudianapusan spermatozoa ditetesi Giemsa hinggapermukaan slide kaca tertutup. Dibiarkan hingga60 menit.Setelah itu, dibilas dengan airmengalir.Selanjutnya diamati morfologi normaldan abnormal spermatozoa.Spermatozoaabnormal mempunyai bentuk kepala tidakberaturan, terlalu membengkok, dan lipatan-lipatan ekor yang abnormal (Washington, et al.,1983; Zohreet al., 2010).Untuk parameterkualitas sperma dihitung 100 spermatozoaberdasarkan karakteristik masing-masingdibawah mikroskop cahaya dengan perbesaran400 kali dan 3 kali pengulangan.
Perhitungan konsentrasi spermatozoaepididimis dilakukan dengan mencampurkansuspensi tersebut dengan larutan George ditabung reaksi dengan perbandingan 1:1 yaitu 50µl suspensi spermatozoa : 50 µl larutan George.Kemudian diaduk dengan vortex selama 0,5menit. Selanjutnya dengan mikro pipet, teteskan10 µl pada kamar hitung Neubeur lalu dihitungdibawah mikroskop cahaya jumlah spermatozoa(Polakoski et al., 1977). Perhitungan dilakukandengan cara menentukan bagian kotak besar ditengah kamar hitung Neubeur denganmenggunakan mikroskop cahaya perbesaran 10kali. Selanjutnya, dihitung 10 kotak kecil secarazig zag yang terletak dalam kotak besar tersebut,masing-masing 5 kotak kecil pada sisi kiri dan 5kotak kecil pada sisi kanan. Hasil perhitunganspermatozoa yang didapat dalam 10 kotak keciltersebut dijumlahkan (=Y) dan dimasukkandalam rumus berikut:× 2,5 × 20 × 10000 × 4Dimana 2,5 adalah 25 kotak kecil dalam 1 kotakbesar pada kamar hitung Neubeur dan yang
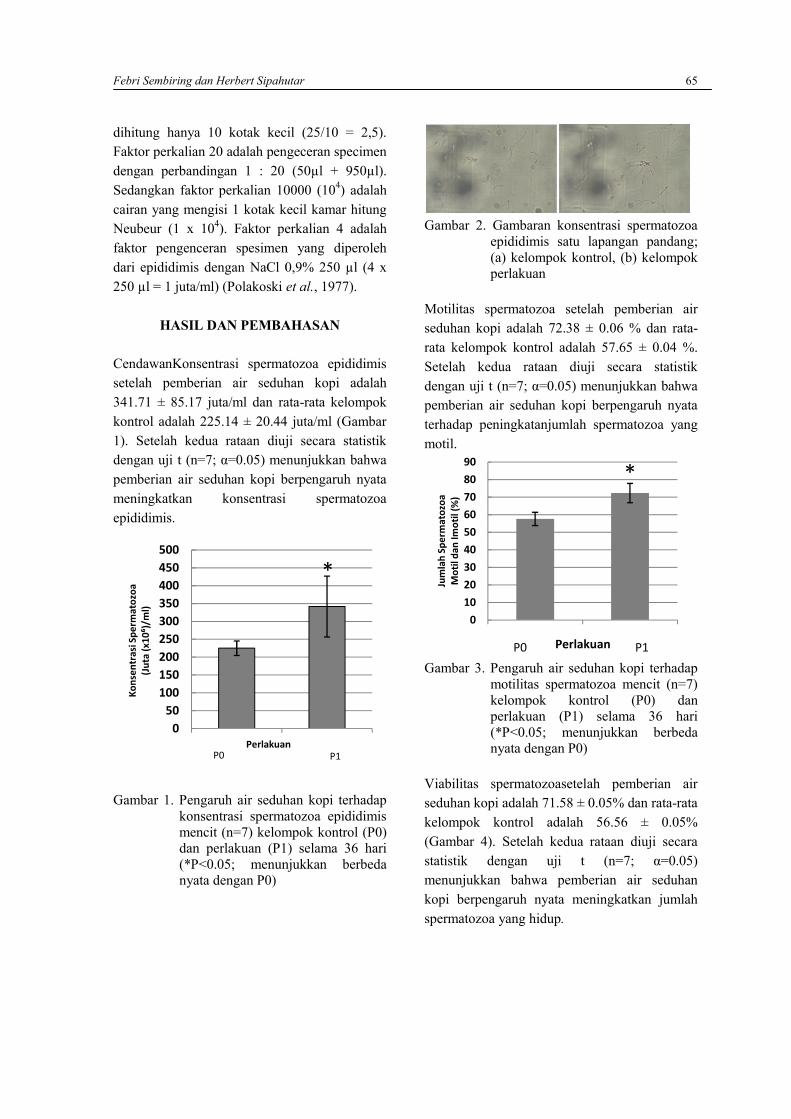
Febri Sembiring dan Herbert Sipahutar 65
dihitung hanya 10 kotak kecil (25/10 = 2,5).Faktor perkalian 20 adalah pengeceran specimendengan perbandingan 1 : 20 (50µl + 950µl).Sedangkan faktor perkalian 10000 (104) adalahcairan yang mengisi 1 kotak kecil kamar hitungNeubeur (1 x 104). Faktor perkalian 4 adalahfaktor pengenceran spesimen yang diperolehdari epididimis dengan NaCl 0,9% 250 µl (4 x250 µl = 1 juta/ml) (Polakoski et al., 1977).
HASIL DAN PEMBAHASAN
CendawanKonsentrasi spermatozoa epididimissetelah pemberian air seduhan kopi adalah341.71 ± 85.17 juta/ml dan rata-rata kelompokkontrol adalah 225.14 ± 20.44 juta/ml (Gambar1). Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyatameningkatkan konsentrasi spermatozoaepididimis.
Gambar 1. Pengaruh air seduhan kopi terhadapkonsentrasi spermatozoa epididimismencit (n=7) kelompok kontrol (P0)dan perlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Gambar 2. Gambaran konsentrasi spermatozoaepididimis satu lapangan pandang;(a) kelompok kontrol, (b) kelompokperlakuan
Motilitas spermatozoa setelah pemberian airseduhan kopi adalah 72.38 ± 0.06 % dan rata-rata kelompok kontrol adalah 57.65 ± 0.04 %.Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyataterhadap peningkatanjumlah spermatozoa yangmotil.
Gambar 3. Pengaruh air seduhan kopi terhadapmotilitas spermatozoa mencit (n=7)kelompok kontrol (P0) danperlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Viabilitas spermatozoasetelah pemberian airseduhan kopi adalah 71.58 ± 0.05% dan rata-ratakelompok kontrol adalah 56.56 ± 0.05%(Gambar 4). Setelah kedua rataan diuji secarastatistik dengan uji t (n=7; α=0.05)menunjukkan bahwa pemberian air seduhankopi berpengaruh nyata meningkatkan jumlahspermatozoa yang hidup.
050
100150200250300350400450500
1
Kons
entr
asi S
perm
atoz
oa(J
uta
(x10
⁶)/m
l)
P0Perlakuan
b
Febri Sembiring dan Herbert Sipahutar 65
dihitung hanya 10 kotak kecil (25/10 = 2,5).Faktor perkalian 20 adalah pengeceran specimendengan perbandingan 1 : 20 (50µl + 950µl).Sedangkan faktor perkalian 10000 (104) adalahcairan yang mengisi 1 kotak kecil kamar hitungNeubeur (1 x 104). Faktor perkalian 4 adalahfaktor pengenceran spesimen yang diperolehdari epididimis dengan NaCl 0,9% 250 µl (4 x250 µl = 1 juta/ml) (Polakoski et al., 1977).
HASIL DAN PEMBAHASAN
CendawanKonsentrasi spermatozoa epididimissetelah pemberian air seduhan kopi adalah341.71 ± 85.17 juta/ml dan rata-rata kelompokkontrol adalah 225.14 ± 20.44 juta/ml (Gambar1). Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyatameningkatkan konsentrasi spermatozoaepididimis.
Gambar 1. Pengaruh air seduhan kopi terhadapkonsentrasi spermatozoa epididimismencit (n=7) kelompok kontrol (P0)dan perlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Gambar 2. Gambaran konsentrasi spermatozoaepididimis satu lapangan pandang;(a) kelompok kontrol, (b) kelompokperlakuan
Motilitas spermatozoa setelah pemberian airseduhan kopi adalah 72.38 ± 0.06 % dan rata-rata kelompok kontrol adalah 57.65 ± 0.04 %.Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyataterhadap peningkatanjumlah spermatozoa yangmotil.
Gambar 3. Pengaruh air seduhan kopi terhadapmotilitas spermatozoa mencit (n=7)kelompok kontrol (P0) danperlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Viabilitas spermatozoasetelah pemberian airseduhan kopi adalah 71.58 ± 0.05% dan rata-ratakelompok kontrol adalah 56.56 ± 0.05%(Gambar 4). Setelah kedua rataan diuji secarastatistik dengan uji t (n=7; α=0.05)menunjukkan bahwa pemberian air seduhankopi berpengaruh nyata meningkatkan jumlahspermatozoa yang hidup.
0102030405060708090
1
Jum
lah
Sper
mat
ozoa
Mot
il da
n Im
otil
(%)
P0 Perlakuan
*
2P1Perlakuan
b
Febri Sembiring dan Herbert Sipahutar 65
dihitung hanya 10 kotak kecil (25/10 = 2,5).Faktor perkalian 20 adalah pengeceran specimendengan perbandingan 1 : 20 (50µl + 950µl).Sedangkan faktor perkalian 10000 (104) adalahcairan yang mengisi 1 kotak kecil kamar hitungNeubeur (1 x 104). Faktor perkalian 4 adalahfaktor pengenceran spesimen yang diperolehdari epididimis dengan NaCl 0,9% 250 µl (4 x250 µl = 1 juta/ml) (Polakoski et al., 1977).
HASIL DAN PEMBAHASAN
CendawanKonsentrasi spermatozoa epididimissetelah pemberian air seduhan kopi adalah341.71 ± 85.17 juta/ml dan rata-rata kelompokkontrol adalah 225.14 ± 20.44 juta/ml (Gambar1). Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyatameningkatkan konsentrasi spermatozoaepididimis.
Gambar 1. Pengaruh air seduhan kopi terhadapkonsentrasi spermatozoa epididimismencit (n=7) kelompok kontrol (P0)dan perlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Gambar 2. Gambaran konsentrasi spermatozoaepididimis satu lapangan pandang;(a) kelompok kontrol, (b) kelompokperlakuan
Motilitas spermatozoa setelah pemberian airseduhan kopi adalah 72.38 ± 0.06 % dan rata-rata kelompok kontrol adalah 57.65 ± 0.04 %.Setelah kedua rataan diuji secara statistikdengan uji t (n=7; α=0.05) menunjukkan bahwapemberian air seduhan kopi berpengaruh nyataterhadap peningkatanjumlah spermatozoa yangmotil.
Gambar 3. Pengaruh air seduhan kopi terhadapmotilitas spermatozoa mencit (n=7)kelompok kontrol (P0) danperlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Viabilitas spermatozoasetelah pemberian airseduhan kopi adalah 71.58 ± 0.05% dan rata-ratakelompok kontrol adalah 56.56 ± 0.05%(Gambar 4). Setelah kedua rataan diuji secarastatistik dengan uji t (n=7; α=0.05)menunjukkan bahwa pemberian air seduhankopi berpengaruh nyata meningkatkan jumlahspermatozoa yang hidup.
*
2P1Perlakuan
b
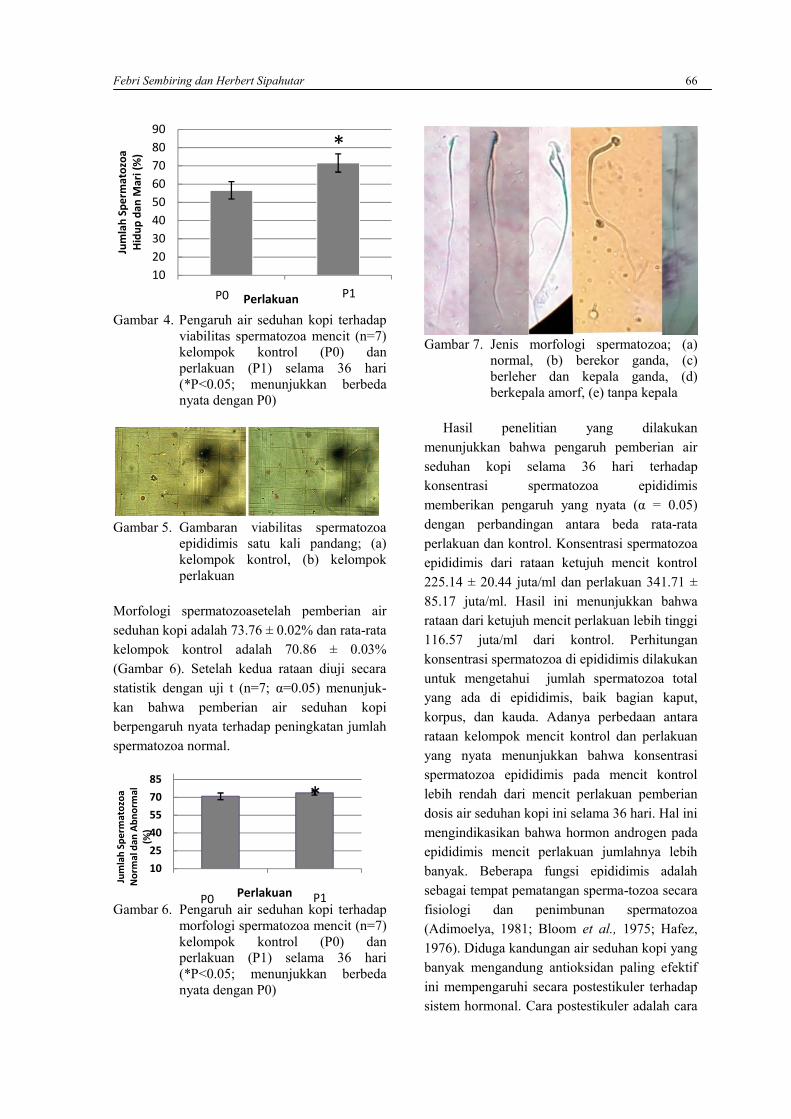
Febri Sembiring dan Herbert Sipahutar 66
*
102540557085
1 2
Jum
lah
Sper
mat
ozoa
Nor
mal
dan
Abn
orm
al(%
)
P0 P1Perlakuan
Gambar 4. Pengaruh air seduhan kopi terhadapviabilitas spermatozoa mencit (n=7)kelompok kontrol (P0) danperlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Gambar 5. Gambaran viabilitas spermatozoaepididimis satu kali pandang; (a)kelompok kontrol, (b) kelompokperlakuan
Morfologi spermatozoasetelah pemberian airseduhan kopi adalah 73.76 ± 0.02% dan rata-ratakelompok kontrol adalah 70.86 ± 0.03%(Gambar 6). Setelah kedua rataan diuji secarastatistik dengan uji t (n=7; α=0.05) menunjuk-kan bahwa pemberian air seduhan kopiberpengaruh nyata terhadap peningkatan jumlahspermatozoa normal.
Gambar 6. Pengaruh air seduhan kopi terhadapmorfologi spermatozoa mencit (n=7)kelompok kontrol (P0) danperlakuan (P1) selama 36 hari(*P<0.05; menunjukkan berbedanyata dengan P0)
Gambar 7. Jenis morfologi spermatozoa; (a)normal, (b) berekor ganda, (c)berleher dan kepala ganda, (d)berkepala amorf, (e) tanpa kepala
Hasil penelitian yang dilakukanmenunjukkan bahwa pengaruh pemberian airseduhan kopi selama 36 hari terhadapkonsentrasi spermatozoa epididimismemberikan pengaruh yang nyata (α = 0.05)dengan perbandingan antara beda rata-rataperlakuan dan kontrol. Konsentrasi spermatozoaepididimis dari rataan ketujuh mencit kontrol225.14 ± 20.44 juta/ml dan perlakuan 341.71 ±85.17 juta/ml. Hasil ini menunjukkan bahwarataan dari ketujuh mencit perlakuan lebih tinggi116.57 juta/ml dari kontrol. Perhitungankonsentrasi spermatozoa di epididimis dilakukanuntuk mengetahui jumlah spermatozoa totalyang ada di epididimis, baik bagian kaput,korpus, dan kauda. Adanya perbedaan antararataan kelompok mencit kontrol dan perlakuanyang nyata menunjukkan bahwa konsentrasispermatozoa epididimis pada mencit kontrollebih rendah dari mencit perlakuan pemberiandosis air seduhan kopi ini selama 36 hari. Hal inimengindikasikan bahwa hormon androgen padaepididimis mencit perlakuan jumlahnya lebihbanyak. Beberapa fungsi epididimis adalahsebagai tempat pematangan sperma-tozoa secarafisiologi dan penimbunan spermatozoa(Adimoelya, 1981; Bloom et al., 1975; Hafez,1976). Diduga kandungan air seduhan kopi yangbanyak mengandung antioksidan paling efektifini mempengaruhi secara postestikuler terhadapsistem hormonal. Cara postestikuler adalah cara
*
102030405060708090
1 2
Jum
lah
Sper
mat
ozoa
Hidu
p da
n M
ari (
%)
P0 P1Perlakuan

Febri Sembiring dan Herbert Sipahutar 67
yang berpengaruh pada spermatozoa setelahberada pada epididimis. Sebab menurut Ilyas(1997), kebutuhan epididimis akan androgenuntuk menjalankan fungsinya lebih tinggidaripada testis, hingga penurunan kadarandrogen sedikit saja dapat mengganggu fungsiepididimis, tetapi tidak menggangguspermatogenesis. Dimana kandungan kimiatersebut memperngaruhi kerja hormon androgensehingga meningkatkan fungsi kerja epididimisseperti pematangan. Proses tersebut sangatbergantung pada hormon androgen (Tadjudin,1988). Sehingga total konsentrasi di epididimisperlakuan lebih tinggi dari kontrol.
Selanjutnya, hasil penelitian yang dilakukanmenunjukkan bahwa pengaruh pemberian airseduhan kopi selama 36 hari terhadap motilitasspermatozoa memberikan pengaruh yang nyata(α = 0.05) dengan perbandingan antara bedarata-rata perlakuan dan kontrol. Motilitasspermatozoa kontrol dari rataan tujuh mencit57.65 ± 0.04 % dan kelompok perlakuan 72.38 ±0.06 %.Hasil ini menunjukkan bahwa kelompokperlakuan 14.73 % lebih tinggi dari kelompokkontrol. Penelitian mengenai viabilitasspermatozoa mencit terhadap pemberian airseduhan kopi selama 36 hari juga memberikanpengaruh yang nyata (α = 0.05) denganperbandingan antara beda rata-rata perlakuandan kontrol. Dengan viabilitas kelompok kontroldari tujuh mencit 56.56 ± 0.05 % dan perlakuan71.58 ± 0.05 %.Ini menunjukkan bahwakelompok perlakuan juga lebih tinggi 15.02 %dari kontrol. Kedua hasil penelitian ini baikmotilitas, viabilitas menunjukkan bahwakomposisi senyawa kimia dari air seduhan kopiyang diberikan bekerja secara postestikuler.Artinya mempengaruhi spermatozoa yangberada di epididimis. Motilitas dan viabilitasditentukan oleh komposisi kimia danperbandingan ion-ion anorganik dalam plasmasemen misalnya: Na+, Ca2+ , Mg2+, dan Zn 2+,serta keseimbangan ATP (Hafez, 1976;Adimoelya, 1981; Mitchell et al., 1976). Dansenyawa kimia kopi salah satunya kafein dapatmenjadi agen stimulasi motilitas sperma dengan
fungsi sebagai penghambat fosfodiesterase yangmenghasilkan akumulasi cAMP yang akhirnyamengaktifkan respirasi dan motilitas sperma.Kafein dapat menstimulasi Ca2+ padaspermatozoa.Ca2+ berperan dalam reaksiakrosom spermatozoa mamalia, sehingga kafeindapat menstimulasi Ca2+ pada spermatozoamenyebabkan reaksi akrosom (Rahayu,2001).Lebih jauh, kafein menstimulasi Ca2+
pada spermatozoa (Rahayu, 2001).Peningkatanmotilitas terjadi karena kafein yangmeningkatkan aktifitas enzim ATP-ase padamembrane sel spermatozoa (Kong et al.,1985).Enzim ATP-ase tersebut berfungsimempertahankan homeostasis internal untuk ionnatrium dan kalium. Jika aktivitas enzim ATP-ase meningkat, maka homeostasis ion natriumdan kalium akan meningkat sehinggakonsentrasi Na+ intrasel menurun, gradien Na+melintasi membran sel akan meneningkatsehingga pengeluaran Ca2+ juga akanmengalami peningkatan (Ganong, 2001).Apabila ion Ca2+ meningkat maka membranakan meningkatkan kemampuannya untukmengangkut bahan-bahan terlarut ke dalamsitoplasma (Salisbury dan Ross, 1995). Denganmeningkatnya permeabilitas membran spermaakan menyebabkan meningkatnya transpornutrien yang diperlukan oleh spermatozoa untukpergerakannya dan daya hidupnya (viabilitas).Seperti yang telah dijelaskan sebelumnya,bahwa kebutuhan epididimis akan hormonandrogen yaitu testosterone lebih besar daritestis, dimana testosteron juga berperan dalampematangan spermatozoa. Selain itu polipenolyang terkandung dalam kopi memiliki fungsisebagai antioksidan yang mampu meningkatkankualitas spermatozoa.
Hasil penelitian mengenai morfologi normalspermatozoa mencit terhadap pemberian airseduhan kopi selama 36 hari juga memberikanpengaruh yang nyata (α = 0.05) denganperbandingan antara beda rata-rata perlakuandan kontrol. Dengan morfologi rataan kelompokkontrol dari tujuh mencit 70.86 ± 0.03 danperlakuan 73.76 ± 0.02%.Ini menunjukkan

Febri Sembiring dan Herbert Sipahutar 68
bahwa kelompok perlakuan lebih tinggi 2.9 %dari kontrol.Hal ini dapat terjadi karena airseduhan kopi banyak mengandungantioksidan.Penelitian Mangoli et al., (2012)mengemukakan bahwa pemberian antioksidanseperti vitamin C dan E dapat meningkatkanjumlah morfologi normal daripada kelompokyang tidak diberikan.Antioksidan dapatmemelihara integritas DNA dari selspermatozoa dan meminimalisir resiko darimutasi di sel-sel germinal.Sehingga didugadapat memaksimalkan jumlah morfologi normaldari spermatozoa.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Air seduhan kopi dengan dosis yang diberikanselama 36 berpengaruh nyata meningkatkankonsentrasi dan kualitas spermatozoa epididimisyang meliputi motilitas, viabilitas, sertamorfologi.
DAFTAR PUSTAKA
Adimoelya A. 1981. Epididimis dan kelenjaraksesoria genitalia pria. PentalokaAndrologi Fakultas Kedokteran UniveritasNegeri Se Indonesia
Bloom W, Fawcett D. 1975. Textbook ofhistology. Philadelphia: Saunders
Cao XW, Lin K, Li CY, Yuan CW. 2011.Areview of WHO laboratory manual for theexamination andprocessing of humansemen (5th edition).Zhonghua Nan Ke Xue17: 1059-63.
Dlugosz L, Bracken MB. 1992. Reproductiveeffects of caffeine: a review and theoreticalanalysis. EpidemiologicReviews 14, 83–100
Ganong WF. 2001. Buku ajar FisiologiKedokteran. Jakarta: Penerbit BukuKedokteran EGC
Hafez ESE. 1976. Transport and survival ofspermatozoa in the female reproductivetract, dalam: human semen anf fertilityregulation in men. Hafez ESE. St Louis.Mosby
Handelsman DJ. 2000. A hormonal malecontraceptive: from wish to reality.Medical Journal of Australia 176: 204-205
Ilyas S. 1997. Pengaruh ekstrak air teh hijau(Camellia sinensis L.) terhadap fertilitasmencit jantan (Mus musculus Strain CBR)dan anak hasil perkawinannya. UniversitasSumatera Utara
Khazaei M. 2012. Protective effects ofsubchronic caffeine administration oncisplatin induced urogenital toxity in malemice. Indian Journal of ExperimantalBiology 50: 638-644
Mangoli E, Pourentezari M, Anvari M, TalebiAR, Nahangi H. 2012. The improvement ofsperm parameters and chomatin quality byvitamin C. Researcher 4: 43-49
Nawrot P, Jordan J, Eastwood, Rotstein A. 2003.Effects of caffeine on human health.FoodAdditives and Contaminants 20: 1–30
Nugraheni T. 2003. Pengaruh vitamin Cterhadap perbaikan spermatogenesis dankualitas spermatozoa mencit (Musmusculus) setelah pemberian ekstraktembakau (Nicotiana tabacum). Biofarmasi1: 13-19
Polakoski KL, Syner FN, Zanveld LJD. 1976.Biochemistry of human seminal plasma,dalam: human semen and fertilityregulation in man. Hafez ESE (ed.). CVMosby Company. Saint Louis
Špaleková E, Makarevich AV, Pivko J.2011.Effect of cafeineon parameters of ramsperm motility.Slovakis Journal of AnimalScience 44:78-83
Tadjudin MK. 1978. Arah Penelitian BiologiSpermatozoa. Surabaya: SimponsiumSpermatologi
Washington WT, Murthy RC, Doye A., EugeneK, Brown D, Bradley I. 1983. Introductionmorphologically abnormal sperm in ratsexposed to oxylene. Arch Androl 11: 233-237
ZohreZare, Hossein E, Moslem M, Mahmood M,Hossein D. 2010. The effect of orallyadministered l-carnitine on testis tissue,sperm parameters and dailyspermproduction in adult mice.Yakhtehmedical journal 11: 382-389.

ISSN 978-602-14989-0-3 BioETI
Indeks keanekaragaman jenis Serangga pada beberapakelompok umur Kelapa Sawit di Kebun Aek Pancur (PPKS),Tanjung Morawa, Sumatera UtaraFITRA SUZANTI1, RETNO WIDHYASTUTI2, SUCI RAHAYU2 DAN AGUS SUSANTO3
1 Mahasiswa S3 FMIPA Pascasarjana Biologi USU2Dosen Pascasarjana FMIPA Biologi USU3Ketua Kelompok Peneliti Proteksi Tanaman PPKS Marihat SumutJl. Bioteknologi no. 1 kampus USU Medan.E-mail: [email protected]
ABSTRAKTPerkebunan kelapa sawit merupakan salah satu ekosistem binaan manusia yang rentan terhadap serangan hama danpenyakit. Keanekaragaman serangga yang ada di ekosistem ini sangat penting diketahui untuk mencegah serangan hamaataupun untuk menciptakan kondisi yang seimbang sehingga kehadiran serangga-serangga tersebut tidak merugikan.Penelitian dilaksanakan dari bulan Februari sampai Juni 2013. Pengambilan sampel serangga dilakukan di kebun kelapasawit milik Pusat Penelitian Kelapa Sawit (PPKS) di Aek Pancur, Tanjung Morawa, Medan, Sumatera Utara. Identifikasiserangga dilakukan di Lembaga Ilmu Pengetahuan Indonesia (LIPI). Penelitian ini dilakukan dengan mengelompokankelapa sawit dalam tiga kelompok umur yaitu umur 1-5 thn, 6-15 thn dan > 15 thn. Pada masing-masing kelompok umurdipilih 10 pohon untuk diambil serangganya dengan cara menyemprotkan insektisida Deltametrin 25 g/l dan menampungserangga yang jatuh. Penyemprotan dilakukan pada siang dan malam hari. Hasil penelitian menunjukan bahwa seranggayang tertangkap seluruhnya adalah 4115 ekor terdiri dari 11 ordo dan 78 famili. Serangga paling banyak ditemukan padakelompok umur 6-15 thn yaitu 2569 ekor, diikuti umur >15 thn sebanyak 881 ekor dan umur 1-5 thn 665 ekor. Indekskeanekaragaman jenis tertinggi adalah pada kelapa sawit berumur >15 thn, diikuti umur 1-5 thn dan 6-15 thn dengan nilaiberturut-turut 1,918; 1,389 dan 0,42.
Kata kunci: keanekaragaman, serangga, kelapa sawit.
Pendahuluan
Kelapa sawit (Elaeis guineensis Jacq.)(Palmaceae), berasal dari (Guinea) Afrika Barat.Tanaman ini memiliki rentang hidup dari 25sampai 30 tahun dan tumbuh sampai dengan 12sampai 15 meter (Owusu-Appiah 2007).Kelapa sawit memberikan kontribusi terhadappembangunan ekonomi dengan meningkatkandevisa, menyediakan lapangan kerja terutamauntuk masyarakat pedesaan dapatmeningkatkan kondisi hidup dan dengandemikian mengurangi kemiskinan dan migrasidesa-kota. Di Negara-negara tropis dan Negara-negara yang tingkat kesejahteraannya rendahperkebunan kelapa sawit merupakan pendorongpembangunan ekonomi yang kuat (Casson,2000; McCarthy dan Zen, 2010; Sheil et al.,2009.; World Growth, 2011), sehingga sering
disebut sebagai '' emas hijau '' (Friends of theEarth, 2008).
Di sisi lain, Kelapa Sawit seringkalidikaitkan dengan deforestasi, degradasi gambut,hilangnya keanekaragaman hayati, kebakaranhutan, dan berbagai masalah sosial (Danielsenet al., 2009.; Koh dan Wilcove, 2008, 2009;Sheil et al., 2009.; Sodhi et al., 2010). Banyakperdebatan tentang dampak kelapa sawitterhadap keanekaragaman hayati, meskipunhanya sebagian kecil dari penelitian yangdipublikasikan berkaitan dengan kelapa sawittelah difokuskan pada dampak tanaman initerhadap keanekaragaman hayati danlingkungan yang lebih luas (Turner et al., 2008).
Serangga merupakan bagian dari ke-anekaragaman hayati yang harus dijagakelestariannya dari kepunahan maupunpenurunan keanekaragaman jenisnya. Seranggamemiliki nilai penting antara lain nilai ekologi,

Fitra Suzant, Retno Widhyastuti, Suci Rahayu dan Agus Susanto 70
endemisme, konservasi, pendidikan, budaya,estetika dan ekonomi (Little, 1957).Penyebaran serangga dibatasi oleh faktor-faktorgeologi dan ekologi yang cocok, sehinggaterjadi perbedaan keragaman jenis serangga.Perbedaan ini disebabkan adanya perbedaaniklim, musim, ketinggian tempat, serta jenismakanannya (Borror & Long, 1998).
Konversi hutan menjadi perkebunan kelapasawit mengakibatkan terjadinya perubahansecara drastis terhadap ekosistem tersebut.Sangatlah penting bagi kita untuk mendapatkanpemahaman yang lebih mendalam tentang efekdari perluasan perkebunan kelapa sawit padaserangga karena sebagian serangga dapatmenjadi hama penting di perkebunan kelapasawit (Mariau et al., 1991). Akibat yang di-timbulkan hama ini sangat besar, sepertipenurunan produksi bahkan kematian tanaman.
Keanekaragaman serangga diyakini dapatdigunakan sebagai salah satu bioindikatorkondisi suatu ekosistem. Data keanekaragamanserangga ini merupakan salah satu langkahawal untuk menciptakan manajemen per-kebunan kelapa sawit yang lebih baik.Pengelolaan perkebunan yang lebih baikdengan mengembangkan performa ekonomitanpa mengabaikan tanggung jawab sosial danlingkungan merupakan model yang ingindicapai oleh perkebunan yang berkelanjutan.
BAHAN DAN METODE
Penelitian ini dilaksanakan pada bulan meisampai Desember 2013. Pengambilan datalapangan dilakukan di kebun milik PusatPenelitian Kelapa Sawit (PPKS) di Desa AekPancur Kab. Deli Serdang, Sumatera Utara.Identifikasi serangga dilakukan di Labor PusatPenelitian Kelapa Sawit (PPKS) Marihatbekerjasama dengan LIPI Cibinong.
Kebun Aek Pancur, secara administratifterletak di Desa Sei Pancur, Tanjung Morawa,Deli Serdang, Sumatera Utara. Jarak Medan –Aek Pancur ± 30 km. Secara geografis terletakN= 3027’0”- 3029’0” dan E = 98046’30” -98048’0”, Ketinggian: 40– 70.m dpl. Curah
hujan rata-rata 2,302 mm dan jumlah hari hujan121 hari.
Untuk keseragaman data dan memudahkanpengolahan data, kelapa sawit dikelompokanmenjadi tiga kelompok umur yang berbedayaitu umur 1-5 tahun, 6-15 tahun dan >15 tahun.Pada masing-masing kelompok umur dipilih 10pohon secara acak untuk diambil sampelserangganya. Masing-masing pohon dipasangkain penampung serangga berukuran 1x1 msebanyak dua buah setiap pohon. Kemudiandilakukan penyemprotan insektisida berbahanaktif Deltamethrin 25 g/ l (senyawa pyretroidsintetik) dengan dosis sesuai anjuran. Alat yangdigunakan adalah Mistblower jenis MaruyamaMD 1800X. Setelah 30 menit kain tampungandiambil dan serangga yang jatuh dimasukan kedalam botol berisi alcohol 70 % untukdiidentifikasi.
Penyemprotan serangga dilakukan padapagi hari sekitar jam 09.00 dan malam hari jam19.00. Bersamaan dengan pengambilan sampelserangga dilakukan pencatatan suhu dankelembaban udara.
Struktur komunitas serangga dianalisisdengan menghitung Indeks keanekaragaman,dan indeks kemerataan. Keanekaragamandihitung dengan indeks Shannon ( H’ = -∑ Piln Pi) dan indeks kemerataan E= H‘/ln(S).Besaran H’<1.5 menunjukkan keanekaragamanjenis tergolong rendah, H’=1.5–3.5 menunjuk-kan keanekaragaman jenis tergolong sedangdan H’>3.5 menunjukkan keanekaragamantergolong tinggi. Besaran E’< 0.3 menunjukkankemerataan jenis tergolong rendah, E’= 0.3 –0.6 kemerataan jenis tergolong sedang danE’>0.6 maka kemerataaan jenis tergolongtinggi (Magurran, 1988).
HASIL DAN PEMBAHASAN
Serangga yang ditemukan di Kebun AekPancur keseluruhannya berjumlah 4115individu yang termasuk dalam 11 ordo dan 78famili. Famili yang terbanyak adalah dari ordoDiptera sebanyak 26 famili, diikuti oleh ordo
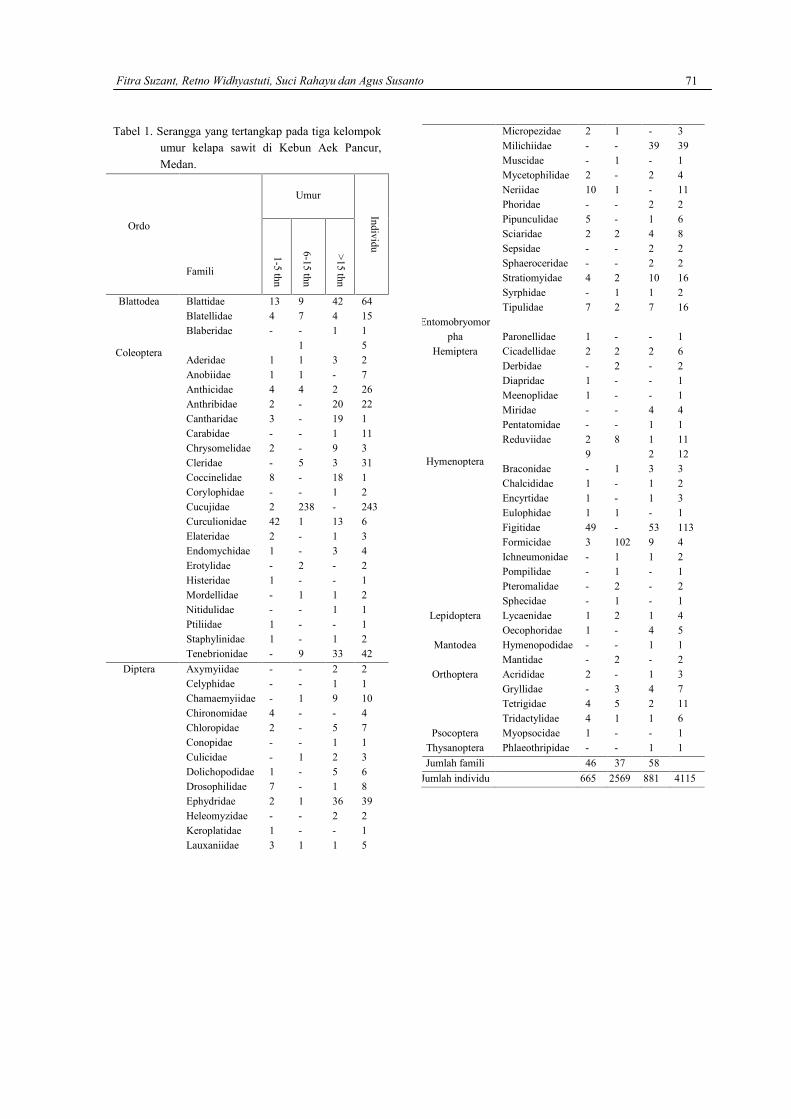
Fitra Suzant, Retno Widhyastuti, Suci Rahayu dan Agus Susanto 71
Tabel 1. Serangga yang tertangkap pada tiga kelompokumur kelapa sawit di Kebun Aek Pancur,Medan.
Ordo
Famili
Umur
Individu
1-5 thn
6-15 thn
>15 thn
Blattodea BlattidaeBlatellidaeBlaberidae
134-
97-
4241
64151
ColeopteraAderidaeAnobiidaeAnthicidaeAnthribidaeCantharidaeCarabidaeChrysomelidaeCleridaeCoccinelidaeCorylophidaeCucujidaeCurculionidaeElateridaeEndomychidaeErotylidaeHisteridaeMordellidaeNitidulidaePtiliidaeStaphylinidaeTenebrionidae
11423-2-8-24221-1--11-
1114----5--2381--2-1---9
3-22019193181-1313--11-133
52726221113311224363421211242
Diptera AxymyiidaeCelyphidaeChamaemyiidaeChironomidaeChloropidaeConopidaeCulicidaeDolichopodidaeDrosophilidaeEphydridaeHeleomyzidaeKeroplatidaeLauxaniidae
---42--172-13
--1---1--1--1
219-51251362-1
211047136839215
MicropezidaeMilichiidaeMuscidaeMycetophilidaeNeriidaePhoridaePipunculidaeSciaridaeSepsidaeSphaeroceridaeStratiomyidaeSyrphidaeTipulidae
2--210-52--4-7
1-1-1--2--212
-39-2-214221017
33914112682216216
Entomobryomorpha Paronellidae 1 - - 1
Hemiptera CicadellidaeDerbidaeDiapridaeMeenoplidaeMiridaePentatomidaeReduviidae
2-11--2
22----8
2---411
62114111
HymenopteraBraconidaeChalcididaeEncyrtidaeEulophidaeFigitidaeFormicidaeIchneumonidaePompilidaePteromalidaeSphecidae
9-111493----
1--1-1021121
2311-5391---
12323111342121
Lepidoptera LycaenidaeOecophoridae
11
2-
14
45
Mantodea HymenopodidaeMantidae
--
-2
1-
12
Orthoptera AcrididaeGryllidaeTetrigidaeTridactylidae
2-44
-351
1421
37116
PsocopteraThysanoptera
MyopsocidaePhlaeothripidae
1-
--
-1
11
Jumlah famili 46 37 58Jumlah individu 665 2569 881 4115

Fitra Suzant, Retno Widhyastuti, Suci Rahayu dan Agus Susanto 72
Coleoptera 21 famili dan ordo Hymenoptera 10famili. Individu terbanyak ditemukan darifamili Curculionidae sebanyak 2436 individu,diikuti oleh family Formicidae sebanyak 1134individu dan family Blattidae 64 individu(Tabel 1.)
Perbedaan umur kelapa sawit mengakibat-kan terjadinya perubahan komposisi seranggadimana jumlah family terbanyak yaitu padakelompok umur >15 tahun, diikuti oleh umur 1-5 tahun dan 6-15 tahun yaitu 58, 46 dan 37famili. Sementara itu dari jumlah individu yangterbanyak adalah pada kelompok umur 6-15tahun, diikuti oleh umur >15 tahun dan 1-5tahun yaitu sebanyak 2569, 881 dan 665individu.
Tabel 2. Indeks Keanekaragaman dan IndeksKemerataan
Umur 1-5 tahun 6-15tahun
>15tahun
H’ 1,39 (R) 0,42 (R) 1,918 (S)E 0.36 (S) 0,12 (R) 0,47 (S)
Indeks keanekaragaman serangga di kebunAek Pancur berkisar antara 0,42 sampai 1,918.Indeks keanekaragaman tertinggi yaitu padakelompok umur >15 tahun termasuk kriteriasedang, pada kelompok umur 1-5 tahun dan 6-15 tahun termasuk kriteria rendah. Indekskemerataan serangga di perkebunan kelapasawit Aek Pancur berkisar antara 0,12 sampai0,47. Indeks kemerataan tertinggi ditemukanpada kelompok umur >15 tahun diikuti olehkelompok umur 1-5 tahun termasuk kriteriasedang, kelompok umur 6-15 tahun indekskemerataannya termasuk kriteria rendah.
Kelapa sawit memiliki rentang hidup dari25 sampai 30 tahun dengan tinggi batangmencapai 12 sampai 15 meter. Semakin tuaumur kelapa sawit maka semakin berfariasimikro habitat yang disediakan untuk serangga.Berdasarkan hasil penelitian didapatkan bahwajumlah famili terbanyak ditemukan padakelompok umur > 15 tahun. Sedangkan jumlahindividu terbanyak pada kelompok umur 6-15tahun. Hal ini karena pada sampel serangga
kelompok umur 6-15 tahun ditemukan banyaksekali jenis serangga penyerbuk Elaeidobiuskamerunicus yang termasuk famili curcu-lionidae.
Indeks keanekaragaman serangga di kebunAek Pancur termasuk kriteria sedang sampairendah. Sementara di hutan alami indekskeanekaragaman serangga biasanya tinggi( Pelawi, 2009). Hal ini berhubungan denganrendahnya keanekaragaman tumbuhan diperkebunan kelapa sawit. Gillison dan Liswanti,(1999) menyatakan bahwa keanekaragamantumbuhan 75% lebih sedikit di perkebunankelapa sawit dibandingkan hutan alam. Hal inimengakibatkan kekayaan spesies di lokasihutan primer adalah rata-rata lima kali lebihtinggi dibandingkan dengan lokasi perkebunan(Bruhl dan Eltz, 2010).
Angka tertinggi untuk indekskeanekaragaman yaitu pada kelapa sawit umur>15 tahun. Hal ini disebabkan pada umur inilingkungan perkebunan menyediakan habitatyang lebih beragam dengan sumber daya yangbanyak untuk mendukung kehidupan serangga-serangga tersebut. Misalnya dengan semakinberagamnya jenis gulma terestrial maupunepifit yang tumbuh, juga adanya sisa-sisapelepah sawit yang mempunyai mikro habitatberagam sehingga memungkinkan untukditempati oleh serangga yang beragam pula.Indeks keanekaragaman tertinggi kedua adalahpada kelompok umur 1-5 tahun. Padakelompok umur ini biasanya tanaman penutuptanah (cover crop) masih tumbuh dengan subur,kemungkinan kondisi ini disukai oleh serangga,sedangkan pada umur 6-15 tahun indekskeanekaragamannya paling rendah. Kondisi inibarangkali disebabkan lahan perkebunankebanyakan dibersihkan secara berkala, baikmenggunakan zat kimia ataupun secara manual,sehingga mikro habitat yang bisa dimanfaatkanoleh seranggapun berkurang.
Indeks keanekaragaman dapat dipergunakansebagai salah satu petunjuk ataupun indikatorkestabilan suatu ekosistem. Dalam hal inisemakin tinggi indeks keanekaragaman

Fitra Suzant, Retno Widhyastuti, Suci Rahayu dan Agus Susanto 73
menunjukan semakin rumit atau kompleknyajaring-jaring makanan, yang berarti jugasemakin stabil atau semakin baik suatulingkungan.
Secara umum, nilai keanekaragaman hayatiperkebunan kelapa sawit sangatsedikit dibandingkan dengan hutan alam danjuga sering lebih rendah dari hutan terganggudan tanaman perkebunan lainnya ( Fitzherbertet al. 2008; Danielsen et al., 2009). Alasanutama untuk nilai keanekaragaman hayati yanglebih rendah pada monokultur kelapa sawitadalah tidak adanya komponen utama darivegetasi hutan, termasuk pohon hutan, lianadan anggrek epifit (Danielsen dkk.,. 2009) .
Adanya perluasan monokultur tanamanyang mengorbankan vegetasi alami sehinggamengurangi keragaman habitat lokal, akhirnyamenimbulkan ketidakstabilan agroekosistemdan meningkatnya serangan hama. Komodititanaman yang dimodifikasikan untukmemenuhi kebutuhan manusia rusak karenatingginya serangan hama. Umumnya semakinintensif tanaman tersebut dimodifikasi makaakan semakin intensif pula hama yangmenyerangnya karakteristik sifat-sifatpengaturan sendiri komoditi alami akan hilangbila manusia memodifikasi komoditi tersebutdengan memecah interaksi kehidupan tanamandan akhirnya menjadi rapuh. Pemecahan inidapat diperbaiki dengan pemulihan komponenkomoditi melalui penambahan atau pening-katan keanekaragaman hayati.
Khusus untuk kelapa sawit, jika manajemenperkebunan dapat disesuaikan sehinggaekosistem ini mendukung sebagian besarspesies hutan sambil mempertahankan hasilyang tinggi, upaya konservasi harus fokus padacara-cara untuk meningkatkan keanekaragamanhayati di perkebunan. Misalnya denganmembiarkan epifit yang tumbuh di pohonkelapa sawit, sehingga keanekaragaman hayatidi perkebunan semakin meningkat, diharapkankeseimbangan ekosistempun semakin baik.Namun demikian hal ini masih memerlukanpenelitian lebih lanjut.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Serangga yang berhasil ditangkap di kebun AekPancur pada kelompok umur 1-5 tahun, 6-15tahun dan >15 tahun berjumlah 4115 individuterdiri 11 ordo dan 78 famili. Serangga palingbanyak ditemukan pada kelompok umur 6-15thn yaitu 2569 ekor, diikuti umur >15 thnsebanyak 881 ekor dan umur 1-5 thn 665 ekor.Indeks keanekaragaman jenis tertinggi adalahpada kelapa sawit berumur >15 thn, diikutiumur 1-5 thn dan 6-15 thn dengan nilaiberturut-turut 1,918 (Sedang); 1,389 dan 0,42(Rendah). Indeks kemerataan serangga diperkebunan kelapa sawit Aek Pancur berkisarantara 0,12 sampai 0,47.
DAFTAR PUSTAKA
Brühl, C. A. and Eltz, T. (2009). "Fuelling thecrisis: Species loss of ground-dwellingforest ants in oil palm plantations in Sabah,Malaysia (Borneo)." Biodiversity&Conservation. .
Casson A (2000) The Hesitant Boom:Indonesia’s Oil Palm Sub-Sector in an Eraof Economic
Crisis and Political Change, CIFOR OccasionalPaper No. 29. Bogor, Indonesia: Centerfor International Forestry Research.
Fitzherbert, E. B., M. J. Struebig, A. Morel, F.Danielsen, C.A. Bruhl, P.F. Donald dan B.Phalan. 2008. "How will oil palmexpansion affect biodiversity? ." Trends inEcology and Evolution 23: 538-45.
Friends of the Earth (2008) Malaysian Palm oil– Green Gold or Green Wash? ACommentary on the Sustainability Claimsof Malaysia’s Palm Oil Lobby, WithaSpecial Focus on the State of Sarawak.Amsterdam, The Netherlands: Friends ofthe Earth.www.foeeurope.org/publications/2008/malaysian-palm-oil-report.pdf
Gillison A and Liswanti N (1999) Impact of oilpalm plantations on biodiversity in Jambi,
Central Sumatra, Indonesia. Bogor, Indonesia:Center for International Forestry Research.

Fitra Suzant, Retno Widhyastuti, Suci Rahayu dan Agus Susanto 74
Magurran, A. E. 1988. Ecological Diversityand Its Measurement. New Jersey:Princeton University Press.
McCarthy J and Zen Z (2010) Regulating theoil palm boom: Assessing theeffectiveness of environmentalgovernance approaches to agro-industrialpollution in Indonesia. Law & Policy 32:153–179.
Meijaard, E., & International, N. C. 2013. Oil-Palm Plantations in the Context ofBiodiversity Conservation.5: 600–612.
Owusu-Appiah, S (2007).Pests of oil palmElaeis guineensis. In Major pest of foodand selected fruit and industrial crops inWest Africa, D. Obeng Ofori, (ed.),CitiPrinters Ltd. 159pp.
Persey S. dan Anhar S. 2010. BiodiversityInformation for Oil Palm. InternationalConference on Oil Palm and Environment,Bali, INDONESIA
Sheil D, Casson A, Meijaard E, et al. (2009)The impacts and opportunities of oil palmin Southeast Asia. What do we know andwhat do we need to know? CIFOROccasional Paper No. 51. Bogor,Indonesia: Center for InternationalForestry Research.
Sodhi, N. S., Koh, L. P., Clements, R., Wanger,T. C., Hill, J. K., Hamer, K. C., … Lee, T.M. 2010. Conserving Southeast Asianforest biodiversity in human-modifiedlandscapes. Biological Conservation.143(10):2375–2384.doi:10.1016/j.biocon.2009.12.029
World Growth (2011) World Bank’s New AntiPoor Palm Oil Policy. Green Paper IssueVIII. World Growth.

ISSN 978-602-14989-0-3 BioETI
Jenis-jenis dan prevalensi Soil Transmitted Helminth pada anak-anak di Olo Bangau Batang Anai Kabupaten Padang Pariaman
FITRI ROZA WIRANATA, MAIRAWITA DAN DAHELMI
Labor Riset Taksonomi Hewan, Jurusan Biologi, FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian tentang Jenis-Jenis dan Prevalensi Soil Transmitted Helminths pada Anak-Anak di Olo Bangau Batang AnaiKabupaten Padang Pariaman telah dilakukan dari bulan Maret sampai April 2014. Penelitian ini dilakukan denganmetode survei. Penentuan lokasi dilakukan dengan cara purposive sampling. Sampel tinja diperiksa menggunakan metodepengapungan. Pada penelitian ini didapatkan 3 jenis parasit dari kelas nematoda, ordo Ascaridorida famili Ascarididaedan ordo Strongylidea famili Strongyloidea yaitu : Ascaris lumbricoides, Necator americanus dan Strongyloidesstercoralis. Prevalensi paling tinggi adalah pada cacing Ascaris lumbricoides yaitu sebesar 70%, kemudian Necatoramericanus sebesar 23%, Strongyloides stercoralis sebesar 6% dan prevalensi gabungan cacing A. lumbricoides dan N.americanus sebesar 16,6%. Anak-anak yang kebiasaannya sehari-hari kurang baik terinfeksi oleh cacing nematoda.
Key words: Anak-anak, infeksi, nematoda, prevalensi
Pendahuluan
Nematoda merupakan anggota dari filumNemathelminthes. Nematoda ada yang hidupbebas di air, di tanah dan ada juga yang bersifatparasit pada hewan maupun tumbuhan.Beratus-ratus ekor cacing dan beberapa spesiessering menghuni satu tubuh inang. Seranggadan avertebrata lain dapat mengandung cacingyang memang sebagai parasit mereka sendirimaupun hanya bertindak sebagai inangperantara. Nematoda sangat baikperkembangannya pada daerah yang beriklimtropis (Noble and Noble, 1989).
Indonesia mempunyai iklim tropis dengankelembaban tinggi dan sangat baik untukberkembangnya penyakit parasit terutamapenyakit cacing. Penyakit cacing parasit iniditularkan melalui tanah yang tercemar telurcacing, lingkungan tempat tinggal yang kumuhdan kurangnya kesadaran akan menjagakebersihan diri. Penyakit cacing parasit yangpaling banyak menyerang anak balita dan usiasekolah dasar (Irawati, 1994).
Kabupaten Padang Pariaman pada daerahpinggir pantai tepatnya di Olo Bangau
mempunyai sanitasi yang masih kurang baik.Kondisi geografis daerah dan lingkungansebagian kecil masyarakat mengakibatkanmereka masih memanfaatkan sungai, kolam,tanah dan pinggiran pantai sebagai saranaMCK. Kondisi sanitasi yang kurang baikmenyebabkan Padang Pariaman masukkedalam 5 besar kota/kab yang mengalamipenyakit karena lingkungan yang kurang baikuntuk wilayah Sumatera Barat (Profil SanitasiKota, 2012). Sanitasi lingkungan yang kurangbaik serta kebiasaan masyarakat yang kurangmenjaga kebersihan diri pribadi memungkinkanuntuk terserang penyakit seperti penyakitparasit yang disebabkan oleh cacing nematoda.Berdasarkan uraian di atas perlu dilakukanpenelitian tentang jenis-jenis dan prevalensinematoda usus pada anak-anak di Olo BangauBatang Anai Kabupaten Padang Pariaman.
Manusia merupakan inang defenitifbeberapa nematoda usus (cacing perut), yangdapat mengakibatkan masalah bagi kesehatanmanusia itu sendiri. Contoh nematoda ususyang menginfeksi manusia antara lain : cacinggelang (Ascaris lumbricoides), cacing tambang(Ancylostoma duodenale dan Necator

Fitri Roza Wiranata, Mairawita dan Dahelmi 76
americanus) dan cacing cambuk (Trichiuristrichiura). Jenis-jenis cacing tersebut banyakditemukan di daerah tropis seperti Indonesia.Pada umumnya telur cacing bertahanpada tanahyang lembab, tumbuh menjadi telur yanginfektif dan siap untuk masuk ke tubuh manusiayang merupakan hospes defenitifnya (Sitorus,2008).
Prevalensi infeksi cacing yang ditularkanmelalui tanah (soil transmitted helminth) masihcukup tinggi dan infeksi cacing ini dapatmenyebabkan masalah kesehatan masyarakat,khususnya pada anak yang masih dalam usiasekolah dasar. Golongan anak sekolah dasarmerupakan kelompok usia yang rentan terhadapinfeksi cacing. Hal ini disebabkan olehkebiasaan bermain anak yang tidakmemperhatikan kebersihan diri danlingkungannya. Demikian pula dengan meng-konsumsi makanan yang dijual di sekolah,tanpa memperhatikan hygiene serta sanitasimakanan dan lingkungan (Silitongga, 2008).Penelitian ini bertujuan untuk mengetahuijenis-jenis dan prevalensi nematoda usus padaanak-anak di Olo Bangau Batang AnaiKabupaten Padang Pariaman.
BAHAN DAN METODE
MetodePenelitian ini dilaksanakan pada bulan Maretsampai April 2014 di Olo Bangau Batang AnaiKabupaten Padang Pariaman denganmenggunakan metode survei. Pemeriksaan tinjadan tanah menggunakan metode pengapungan(Soedarto, 2008). Penentuan lokasi dilakukandengan cara Purposive Sampling. Lokasi yangdipilih merupakan pemukiman pendudukpinggir pantai. Sebanyak 30 orang anakdiberikan satu lembar kuisioner kemudiandiambil sampel tinja dan juga sampel tanahdisekitar rumahnya. Sampel diperiksa denganmetode pengapungan menggunakan sentrifus(Soedarto, 2008).Alat dan bahanAlat yang digunakan dalam penelitian iniadalah daftar kuisioner, mikroskop, kamera
digital, mikrometer, botol film, sendok, kertaslabel, tabung sentrifus, cawan petri, batangpengaduk, objek glass, cover glass, timbangan,rak tabung reaksi, sarung tangan, masker,termos es dan alat-alat tulis. Bahan yangdigunakan dalam penelitian ini adalah tinja darianak-anak dan tanah yang didugaterkontaminasi tinja, NaCl jenuh dan aquades.Cara Kerja
a. Koleksi sampel di lapanganSampel tinja diambil kira-kira setengah sendokteh atau seujung jari tangan. Masing-masinganak diberi satu botol sampel yang telah diberilabel. Sampel tinja yang telah dikoleksidisimpan dalam termos es yang telah diisi batues, untuk mencegah tinja agar tidak menjadikeras dan berbau disimpan di dalam kulkasselanjutnya sampel tanah diambil disekitarpekarangan rumah anak yang diambil tinjanyadan dibawa ke Laboratorium TaksonomiHewan Universitas Andalas Padang.
b. Identifikasi sampel di laboratoriumPemeriksaan sampel tinja dilakukan denganmenggunakan sentrifus dengan metodepengapungan. Tinja diambil sebanyak 5 gram,diencerkan dengan air sebanyak 100 ml dandiaduk hingga homogen, kemudian diambilsebanyak 10 ml dan dimasukkan ke dalamtabung sentrifus dan disentrifus dengankecepatan 2500 rpm selama 5 menit sampaifiltratnya bening, selanjutnya filtrat dalamtabung dibuang dan ditambahkan larutan NaCljenuh sebanyak 10 ml kemudian di sentrifusdengan kecepatan 2500 rpm selama 5 menit.Semakin tinggi kecepatan dan semakin lamawaktu yang diberikan maka akan semakin cepatfiltratnya menjadi bening. Hasil sentrifusditambahkan larutan jenuh NaCl hinggapermukaan larutan mendekati mulut tabung laluditutup dengan cover glass pada mulut tabungdan ditunggu selama 5 menit. Cover glassdiambil dan diletakkan diatas object glasskemudian diperiksa dengan menggunakanmikroskop dengan perbesaran 10 sampai 400kali. Pemeriksaan sampel tanah sama denganpemeriksaan sampel tinja.

Fitri Roza Wiranata, Mairawita dan Dahelmi 77
Pemeriksaan dilakukan sampai tidakditemukan lagi telur cacing pada masing-masing tinja anak yang diperiksa. Jika tidakditemukan lagi telur atau larva cacing padasatu anak, maka pemeriksaan dilanjutkan lagike tinja anak selanjutnya dan begitu jugaseterusnya. Larva atau telur cacing parasit yangditemukan difoto menggunakan kamera digitalkemudian diidentifikasi dengan menggunakanbeberapa literatur seperti Brown (1983),Purnomo, Magdalena, Ayda dan Harijani(1987), Soedarto (2008). Telur yang ditemukandihitung jumlahnya dan diperhatikan ciri-ciriserta warnanya.Analisis DataData hasil identifikasi nematoda usus yangdianalisis secara deskriptif dan disajikan dalambentuk gambar dan tabel. Nilai prevalensidihitung untuk setiap spesies parasit. Nilaiprevalensinya dihitung merujuk kepada Bush etal, 1997.Prevalensi = n x 100%
NKeterangan :n = jumlah inang yang terinfeksi parasit (orang).N = jumlah inang yang diamati (orang).
HASIL DAN PEMBAHASAN
Cendawan Dari pemeriksaan sampel tinja yangtelah dilakukan didapatkan jenis cacing yangmenginfeksi anak-anak terdiri dari kelasnematoda, dua ordo (Ascaridorida danStrongylidae), tiga famili (Ascarididae,Necatoritidae dan Strongyloidea), tiga jenisyaitu : Ascaris lumbricoides, Necatoramericanus dan Strongyloides stercoralis. Daripemeriksaan sampel tanah yang telah dilakukandidapatkan larva rhabditiform dari cacing N.americanus dan S. stercoralis. Masing-masingdeskripsi dari jenis nematoda usus yangditemukan adalah sebagai berikut :
Ascaris lumbricoides (Linnaeus, 1758)Telur cacing A. lumbricoides yang didapatkanpada tinja berbentuk oval dengan warna
kecoklatan. Bagian pinggir telur terdapatbenjol-benjol atau bergerigi. Ukuran telur yangdidapatkan bervariasi yaitu : panjang 45 - 62,5µ dan diameter 30 - 47,5 µ (Gambar 1).
Berdasarkan dari bentuk, ukuran dan warnadari telur cacing A. lumbricoides yangdidapatkan, sesuai dengan ciri-ciri yangdikemukakan oleh Purnomo et al (1987), Nobleand Noble (1989) dan Brown (1983) bahwatelur A. lumbricoides berbentuk oval, berwarnakecoklatan dan mempunyai dinding yangbergerigi, panjang telur A. lumbricoides 45-70µ dan diameter 35-50 µ. Mohr (1957), telurcacing A. lumbricoides yang dibuahi berbentukoval, berkulit dua lapis, panjang telur berkisar40-70 µ dan diameter telur berkisar 35-50 µ.Cacing ini terutama menyerang anak-anak usia5-9 tahun, sedangkan menurut jenis kelamintidak menunjukkan perbedaan yang nyata,artinya laki-laki dan perempuan memilikikemungkinan terinfeksi yang sama. Cacing A.lumbricoides ini tersebar kosmopolit terutamadi daerah tropis dengan udara yang lembabserta sangat erat hubungannya dengan keadaanhigien dan sanitasi yang buruk. Dengankelembapan tinggi dan suhu berkisar antara250C-300C sangat baik untuk perkembangantelur A. lumbricoides menjadi bentuk infektif(Brown, 1983).
Necator americanus (Dubini, 1983).Telur cacing N. americanus yang didapatkanpada tinja berbentuk bulat lonjong dengandinding yang tipis dan terdapat 2-8 sel didalamtelur, ukuran panjang telur cacing tambangbervariasi berkisar antara 65-69 µ dan diameternya 36-40 µ (Gambar 2a). Berdasarkan bentukdan ukuran telur cacing N. americanus yangdidapatkan sesuai dengan ciri-ciri yangdikemukakan oleh Purnomo et al (1987), Nobleand Noble (1989) dan Brown (1983), bahwatelur cacing tambang panjangnya 56-76 µ dandiameter 30-40 µ, berbentuk bulat lonjongdengan dinding yang tipis dan di dalam telurterdapat 2-8 sel.

Fitri Roza Wiranata, Mairawita dan Dahelmi 78
Larva rhabditiform cacing N. americanusyang ditemukan pada tanah mempunyaipanjang 305µ dan diameter 13µ (Gambar 2b).Ekor dari larva tersebut meruncing. MenurutLevine (1990), Larva filariform panjangesophagusnya setengah dari panjang badan,dengan tubuh agak sedikit langsing, ukuranpanjang badannya kira-kira 450-700µ dandiameternya 10-15µ. Larva rhabditiformukuran panjangnya adalah 300-400µ dandiameternya 15-20µ, dan esophagusnya pendek.Cacing tambang yang hidup pada manusiaselain N. americanus, juga terdapat A.duodenale. Telur A. duodenale hampir samadengan telur N. americanus, hanya ukurannyayang berbeda, dimana A. duodenale ukuranpanjangnya berkisar 56-60µ dan diameter 36-40µ. Larvanya juga berbeda, larva filariformdari A. duodenale tombak esophagus tidakmenonjol, tertutup pada ujung anterior danmemiliki sarung yang halus, sedangkan N.americanus memiliki tombak esophagus yangmenonjol, terbuka pada ujung anterior danmemiliki sarung bergaris nyata pada ujungposterior (Zaman, 1997). Pada mulut larvafilariform N. americanus terdapat otot mulutyang berbentuk lanset berwarna putihmengkilat, sedangkan pada mulut larvafilariform A. duodenale otot mulutnya tidakberbentuk lanset (Noble dan Noble, 1989).
Strongyloides stercoralis (Linneus, 1758)Telur cacing S. stercoralis yang didapatkanpada tinja berbentuk bulat lonjong dan ujungnya lebih runcing dari pada cacing tambang,berdinding tipis dan juga terdapat beberapa seldi dalamnya. Ukuran telur cacing S. stercoralisini panjang nya 50-67µ dan diameter 32,5-35µ(Gambar 3a). Berdasarkan dari bentuk danukuran telur S. stercoralis yang telahdidapatkan, hal ini sesuai dengan ciri-ciri yangdikemukakan oleh Purnomo et al (1987),Brown (1983), dan Noble and Noble (1989),bahwa telur S. stercoralis panjangnya 50-70µdan diameter 30-40µ.
Larva rhabditiform cacing S. stercoralisyang ditemukan pada tanah mempunyaipanjang 377µ dan diameter 17µ (Gambar 3b).Ekor dari larva tersebut agak membulat.Menurut Levine (1990), Larva filariform danlarva rabdhitiform agak sedikit susah dibedakan,karena dibedakan berdasarkan panjangesophagusnya. Larva filariform panjangesophagusnya setengah dari panjang badan danmulutnya terbuka, ukuran panjang badannyakira-kira 450-630µ dan diameternya 13-16µ.Larva rhabditiform ukuran panjangnya adalah350-380µ dan diameternya 15-20µ, danesophagusnya pendek.
PREVALENSIPada Tabel 1 dapat dilihat bahwa prevalensicacing Ascaris lumbricoides merupakanprevalensi paling tinggi yaitu sebesar 70%,kemudian prevalesi cacing tambang sebesar23% dan yang paling rendah adalah prevalensicacing Strongyloides stercoralis yaitu sebesar6% dan prevalensi gabungan antara cacing A.lumbricoides dengan N. americanus sebesar16,6%. Tingginya prevalensi telur cacing A.lumbricoides disebabkan karena cacing betinadewasa A. lumbricoides dapat menghasilkantelur sebanyak 200.000 butir per harinya.Prevalensi tertinggi kedua yaitu cacing N.americanus. Cacing betina dewasa N.americanus dapat menghasilkan telur 9000butir per harinya. Prevalensi yang palingrendah adalah cacing S. stercoralis, cacingbetina dewasanya dapat menghasilkan telur6000 butir per hari (Brown, 1983).
Penyebab tingginya prevalensi gabungandari cacing A. lumbricoides dengan N.americanus adalah kebersihan diri danlingkungan yang kurang terjaga dengan baik,bermain tidak memakai alas kaki, membuangtinja di sembarangan tempat seperti dipekarangan rumah, dan makan tidak mencucitangan sampai bersih sehingga memungkinkantelur cacing yang menempel pada tangan akanterbawa bersama makanan yang dimakan.Menurut Rahayau (2006), telur cacing yang

Fitri Roza Wiranata, Mairawita dan Dahelmi 79
Gambar 1. Telur cacing A. lumbricoides ditemukan pada tinja manusia.
(a) (b)
Gambar 2. a. Telur cacing N. americanus ditemukan pada tinja manusia. Gambar. b. Larvafilariform N. americanus ditemukan pada tinja.
(a) (b)
Gambar 3. a. Telur cacing S. stercoralis ditemukan pada tinja manusia. Gambar. b. Larvafilariform cacing S. stercoralis ditemukan pada tanah.

Fitri Roza Wiranata, Mairawita dan Dahelmi 80
Tabel 1. Prevalensi Soil Transmitted Helminthpada Anak-anak di Olo Bangau BatangAnai Kab. Padang Pariaman.
No
Jenis cacing parasit Prevalensi(%)
1 Ascaris lumbricoides 70%2 Necator. Americanus 23%3 Strongyloides stercoralis 6%4 Gabungan (A. lumbricoides +
N. americanus)16,6%
mencemari seseorang akan dapat tertelan jikaorang tersebut memegang makanan dan tidakmencuci tangan terlebih dahulu sebelum makan.Tingginya prevalensi Soil TransmittedHelminth ini mungkin disebabkan karenasanitasi lingkungan yang kurang baik,kebiasaan berdefekasi disembarang tempat dankebersihan diri. Telur cacing ini juga bisamenginfeksi anak melalui oral yaitu telurinfektif dari cacing yang telah mengontaminasitanah dan terbawa oleh kaki lalat sehinggamakanan yang dihinggapi lalat makanantermakan dan larva dari cacing itu juga bisamenebus kulit seperti telapak kaki karena saatbermain anak-anak tidak memakai alas kaki(Rahayu, 2006).
Seperti yang dikemukakan oleh Brown(1983), bahwa orang-orang yang terinfeksiparasit yang berdefekasi di tanah ataudisembarang tempat menyebabkan terinfeksioleh parasit. Ukuran telur cacing yang sangatkecil dan tidak terlihat dan mudah berpindahakan sangat mudah untuk terinfeksi parasit.Menurut Irianti (1999), kebiasaan orang-orangmembuang kotoran dan sampah sertaperumahan yang kurang bersih sehingga dapatmenimbulkan penyakit.
Menurut Margono (2004), kebiasaanpenduduk dengan BAB di tanah dan pemakaiantinja sebagai pupuk kebun (dibeberapa daerahtertentu) juga lebih memudahkan dalampenyebaran serangan cacingan, karenakebiasaan seperti berdefekasi disekitar rumah,makan tanpa cuci tangan, bermain-main ditanah sekitar rumah tidak menggunakan alaskaki, maka khususnya anak balita terusmenerus mendapatkan serangan.
Anak kecil yang terinfeksi parasitmerupakan sumber terpenting untukterkontaminasi oleh tanah karena merekaberdefekasi dimana-mana di halaman rumahdan juga sering bermain tanah di sekitarpekarangan rumah, tempat telur-telur yangresisten dapat hidup dalam waktu yang lama(Brown, 1983). Penyakit cacingan dapatditularkan melalui tangan dan kebanyakan telurcacing parasit bertebaran dipermukaan tanah,debu dan menempel di karpet perumahan. Telurcacing yang mencemari tangan seseorang dapattertelan jika orang tersebut memegang makanandan tidak mencuci tangan sebelum makan(Rahayu, 2006).
Menurut Faust dan Russel (1964), bahwatelur cacing A. lumbricoides tahan terhadaplingkungan yang buruk dan memiliki telur yangbanyak dimana seekor A. lumbricodes betinamampu menghasilkan telur sebanyak 200.000butir telur per harinya. Telur cacing A.lumbricoides juga tahan terhadap desinfektankimiawi dan terhadap ancaman rendamansementara didalam berbagai bahan kimia yangkeras (Brown, 1983), dibandingkan dengantelur cacing lainnya, sehingga telur A.lumbricoides dapat hidup berbulan-bulan dalamair, tanah atau tinja.
Prevalensi anak-anak yang terinfeksi cacingN. americanus adalah sebanyak 23%, lebihsedikit jika dibandingkan dengan yangterinfeksi A. lumbricoides, maka dapatdikatakan bahwa lokasi penelitian di daerahOlo Bangau ini bukan lingkungan yang sesuaiuntuk pertumbuhan telur cacing N. americanus.Infeksi cacing N. americanus merupakan salahsatu penyakit yang sering diderita masyarakatyang tidak memperhatikan kebersihan diri dankebersihan lingkungan. Jika di dalam tinjaditemukan telur cacing N. americanusmeskipun hanya satu, keadaan seperti itudikatakan bahwa anak-anak sudah terinfeksicacing N. americanus.
Brown (1983), menyatakan bahwapenyebaran cacing tambang banyak ditemukanpada lingkungan yang paling cocok dengan

Fitri Roza Wiranata, Mairawita dan Dahelmi 81
suhu dan kelembapan yang tinggi terutama didaerah pertanian dan pertambangan dimanacacing tambang dapat menghasilkan telursebanyak 9000 butir perharinya. Tanah berpasirdan kelembaban yang tinggi juga merupakanbeberapa faktor yang mendukungberkembangnya telur cacing tambang menjadibentuk larva infektif (filariform) dan juga tanahyang baik untuk pertumbuhan larva ialah tanahgembur (berpasir, humus).
Tangan yang bersentuhan langsung dengankotoran manusia dan binatang ataupun cairantubuh lain seperti ingus, makanan atauminuman yang terkontaminasi saat tidak dicucidengan sabun dapat memindahkan bakteri,virus dan parasit pada orang lain yang tidaksadar bahwa dirinya sedang tertular. Haltersebut yang memungkinkan terjadinyapenularan cacing terutaman nematoda ususyang penularannya dengan tanah (Tjitra, 1991).Anak-anak paling sering terserang penyakitcacingan karena biasanya jari-jari tanganmereka dimasukkan ke dalam mulut, ataumakan tanpa cuci tangan, namun demikiansesekali orang dewasa juga bisa terinfeksicacing. Cacing yang paing sering ditemui ialahcacing A. lumbricoides, cacing tambang, T.trichiura (Ariyasta, 2010). Perilaku individualsangat mempengaruhi infeksi cacingan,intensitas prevalensi yang tinggi pada anak-anak juga disebabkan oleh kebiasaanmemasukan jari-jari tangan yang kotor kedalam mulut atau makan tanpa cuci tangan.Menurut Yulianto (2007), menyatakan bahwausaha pencegahan penyakit cacingan ataralain : menjaga kebersihan badan, kebersihanlingkungan dengan baik, makanan danminuman yang baik dan bersih, memakai alaskaki, membuang air besar di jamban (kakus),memelihara kebersihan diri dengan baik sepertimemotong kuku dan mencuci tangan sebelummakan.
Kebiasaan makan sehari-hari juga dapatmenyebabkan terjadinya penularan penyakitcacing tertentu, misalnya, kebiasaan makanmakanan yang mentah, ikan, kerang, daging
dan sayuran. Jika dalam makanan tersebutterdapat kista atau larva cacing, maka siklushidup cacingnya menjadi lengkap sehinggaterjadi infeksi pada manusia (Ariyasta, 2010).
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Ditemukan tiga jenis cacing nematoda ususyaitu Ascaris lumbricoides, Necatoramericanus dan Strongyloides stercoralis.Prevalensi yang paling tinggi didapatkan daricacing Ascaris lumbricoides yaitu 63,3%,cacing Necator americanus yaitu 23% sertacacing Strongyloides stercoralis sebesar 6%.Prevalensi gabungan antara cacing A.lumbricoides dan N. americanus adalah 16,6%.
DAFTAR PUSTAKA
Noble, E. R. and G. A. Noble. 1989. BiologiParasit Hewan. Terjemahan Wardianto.Gajah Mada University Press: Yogyakarta.
Irawati, N.1994. Nematoda Usus pada AnakUsia Sekolah Di Sekitar Lokasi TempatPembuangan Sampah Akhir LubukMinturun. Lembaga Penelitian UniversitasAndalas. Padang.
Profil Sanitasi Kota. 2012. Profil Sanitasi Kota.http//.Ppsp.nawasis.info/sanitasi/profil/2012. Diakses pada 22Maret 2014.
Soedarto. 2008. Buku Ajar ParasitologiKedokteran. Sagung Seto. Jakarta.
Brown, H. W. 1983. Dasar Parasitologi Klinis.Terjemahan B. Rukmono. Gramedia.Jakarta.
Purnomo, W., Gunawan., L. Magdalena., R.Ayda dan Harijani. 1987. AtlasHelminthologi Kedokteran. Gramedia.Jakarta.
Bush, A. O., K. D. Lafferty., J. M. Lotz, and A.W. Shostak. 1997. Parasitology MeetsEcology on Its Own Terms: Margolis EtAl. Revisited. Journal of Parasitology 83:575-583.
Levine, N. D. 1990. Parasitologi Veteriner.Terjemahan Gatut Ashadi. UGM.Yogyakarta.

Fitri Roza Wiranata, Mairawita dan Dahelmi 82
Zaman, V. 1997. Atlas ParasitologiKedokteran Edisi II. Hipokrates. Jakarta.
Rahayu, S. E. 2006. Keberadaan Telur CacingParasit pada Siswa SD di Sekitar InstalasiPengolahan Air Limbah (IPAL) TerpaduKota Malang dan Hubungannya denganKepadatan Telur Cacing pada Air LimbahPerumahan di IPAL Terpadu. PenelitianHayati 11:105-112.
Irianti, I. S. Zalbawi, Supraptini. 1999.Penelitian dalam Rangka PenerapanSistem Pembuangan Tinja dan SampahTepat Guna Desa Pantai di KabupatenRembang dan Kabupaten Lamongan.Buletin Penelitian Kesehatan 27 (34) :346-361.
Margono. 2004. Parasitologi Kedokteran.Fakultas Kedokteran UniversitasIndonesia. Jakarta.
Faust, E.C. and P. F. Russell. 1964. Craig &Fausts Clinical Parasitology. 7th ed. Lea7 Febiger Philadelphia. USA. 341-429.
Tjitra, E. 1991. Penelitian-penelitian SoilTransmitted Helminth di Indonesia.Cermin Dunia Kedokteran 72 : 12-16.
Yulianto, Evi, 2007. Hubungan HygieneSanitasi dengan Kejadian PenyakitCacingan pada Siswa Sekolah DasarNegeri Rowosari 01 KecamatanTembalang Kota Semarang. Skripsi.Jurusan Ilmu Kesehatan Masyarakat),Unversitas Negeri Semarang.
Ariyasta, Lupiana, 2010. Faktor-faktor yangBerhubungan dengan Kejadian PenyakitKecacingan di SD 02 Desa TerentangKecamatan Banyuasin III KabupatenBanyuasin. Skripsi. Program Studi IlmuKesehatan Masyarakat STIK Bina HusadaPalembang.

ISSN 978-602-14989-0-3 BioETI
Potensi beberapa tanaman dalam mengakumulasi Merkuri padatanah bekas Tambang EmasFITRI WAHYUNI, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIA
Labor Riset Fisiologi Tumbuhan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRACTThis research studied about potention of some plants for mercury accumulation on gold mine soil has been done fromNovember 2013 until Maret 2014 in Screen House and Laboratory of Plant Phsiology and tissue culture, BiologyDepartment, Faculty of Mathematics and Natural Science, Andalas University Padang. This research used experimentalmethod with five replications. The research used Ageratum conyzoides, Cyperus kyllingia dan Digitaria ciliaris asmercury plants accumulator. The result of this research showed that A.conyzoides, C.kyllingia and D.ciliaris havepotential for phytoremediation in mercury contaminated soil. Accumulated mercury was found in some parts of plantespecially in shoot and root. A.conyzoides was found 0.01 ppm and 0.015 ppm. C.kyllingia 0.0135 ppm and 0.015 ppmand D.ciliaris 0.016 ppm and 0.022 ppm. Digitaria ciliaris can be considered as phytoremediation agent for mercurycontaminated soil.
Key words: Phytoremediation, mercury, plants accumulator
Pendahuluan
Provinsi Sumatera Barat mengandung potensisumber daya mineral seperti emas dan mangan.Menurut laporan Dinas Pertambangan danEnergi Provinsi Sumbar (2004), lokasipenambangan emas terdapat pada wilayahdaerah Kabupaten Sijunjung, 50 Kota, Pasamandan Pesisir Selatan. Pada wilayah KabupatenSijunjung, deposit emas diperkirakan terdapatdi sejumlah lokasi dan salah satu lokasi tersebutadalah Tanjung Ampalu. Dengan adanyakegiatan pertambangan memberikan dampakpositif bagi masyarakat karena dapatmeningkatkan pendapatan bagi masyarakat. Dilain pihak juga memberikan dampak negatifbagi kesehatan masyarakat dan lingkungan.Kegiatan pertambangan ini berpotensimencemari lokasi dan lingkungan karenaadanya penggunaan merkuri dalam prosesamalgamasi sebagai bahan untuk mengikat danmemisahkan biji emas (Setiabudi, 2005).
Merkuri termasuk dalam kategori sangatberacun untuk makhluk hidup maupunlingkungan. Merkuri merupakan logam beratyang berbahaya karena memiliki rapat massatinggi dan dalam konsentrasi kecil dapat
bersifat racun dan berbahaya (Aminudin, 2006).Adapun dampak lingkungan akibat kegiatanpertambangan emas antara lain berupapenurunan produktivitas tanah, pemadatantanah, terjadinya erosi dan sedimentasi,terjadinya gerakan tanah dan longsoran, pHtanah yang rendah, terganggunya flora danfauna, terganggunya keamanan dan kesehatanmasyarakat.
Hasil penelitian pendahuluan pada lokasipertambangan emas Tanjung Ampalu diSijunjung didapatkan kandungan merkuri(Hg) sebesar 1,100 ppm (Wahyuni, Noli danFebria, 2013, Unpublished).
Salah satu metode aplikatif diharapkanmampu menangani masalah pencemaran logamberat merkuri (Hg) pada tanah dengan carafitoremediasi. Fitoremediasi adalah peng-gunaan tumbuhan untuk menghilangkanpolutan dari tanah atau perairan yangterkontaminasi. Salah satu karakter tumbuhanyang potensial sebagai fitoremediator adalahmempunyai sifat hipertoleran yang tinggiterhadap lingkungan yang tercemar dankemampuannya dalam menyerap logam berat.Fitoremediator tersebut dapat berupa herba,

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 84
semak, perdu atau jenis rerumputan bahkanpohon (Juhaeti, Syarif dan Hidayati, 2003).
Beberapa penelitian yang dilakukan di lahanbekas penambangan emas ditemukan tanamanDigitaria radicosa (Presl) Miq. yangmengandung Hg 50,93 mg/kg (Juhaeti, Syarifdan Hidayati, 2005). Hasil penelitian lainbeberapa tanaman mampu mampumengakumulasi Hg Lindernia crustacean (2,96mg/kg), Digitaria radicosa (1,65 mg/kg),Zingiber purpurium (0.85 mg/kg), Paspalumconjugatum (8,82 mg/kg), Cyperus kylingia(3,97 mg/kg) dan Caladium bicolor (0,14mg/kg) selama pertumbuhan 9 minggu(Handayanto, 2012).
Mekanisme biologis dari hiperakumulasiunsur logam pada dasarnya meliputi proses-proses: (1) Interaksi rizosferik, yaitu prosesinteraksi akar tanaman dengan media tumbuh(tanah dan air). Dalam hal ini tumbuhanhiperakumulator memiliki kemampuan untukmelarutkan unsur logam pada rizosfer danmenyerap logam sehingga menjadikanpenyerapan logam oleh tumbuhan hiper-akumulator melebihi tumbuhan normal; (2)Proses penyerapan logam oleh akar padatumbuhan hiperakumulator lebih cepatdibandingkan tumbuhan normal, terbuktidengan adanya konsentrasi logam yang tinggipada akar (Lasat 1996). Akar tumbuhanhiperakumulator memiliki daya selektifitasyang tinggi terhadap unsur logam tertentu; (3)Sistem translokasi unsur dari akar ke tajuk padatumbuhan hiperakumulator lebih efisien di-bandingkan tanaman normal (Hidayati, 2005).Tanaman Ageratum conyzoides, Digitariaciliaris dan Cyperus kyllingia didugaberpotensi sebagai agen fitoremediasi. Namunsejauh ini belum banyak data mengenai potensitanaman A. conyzoides, D. ciliaris dan C.kyllingia dalam meremediasi lahan tambangemas yang tercemar merkuri (Hg). Penelitianini bertujuan untuk mengetahui potensitanaman A. conyzoides, D. ciliaris dan C.kyllingia dalam meremediasi lahan bekastambang emas yang tercemar merkuri (Hg)
sehingga dapat berpotensi sebagai fitoremediasidan untuk mengetahui tanaman terbaik dalammeremediasi lahan bekas tambang emas yangtercemar merkuri (Hg).
BAHAN DAN METODE
Penelitian ini dilaksanakan pada bulanNovember 2013 sampai Maret 2014 di RumahKaca dan Labor Fisiologi Tumbuhan JurusanBiologi, FMIPA, Universitas Andalas, Padang.
Penelitian ini menggunakan metodeeksperimen dan sebagai perlakuan adalahbeberapa jenis tanaman yang berpotensisebagai tanaman akumulator Hg yaituA.conyzoides, D.ciliaris dan C.kyllingia,masing-masing diulang 5 kali.
Alat yang digunakan dalam penelitian iniadalah cangkul, polybag, timbangan, mistar,ICPE9000 Shimadzu, oven, baki, karung,kantong plastik, camera digital, ember plastikdan alat tulis.
Bahan yang digunakan adalah bibitA.conyzoides, D.ciliaris, C.kyllingia dan tanahtambang emas yang diambil di lokasipenambangan emas di daerah Sijunjung yaituTanjung Ampalu.Pengambilan sampel tanah :Pengambilan tanah bekas tambang emasdilakukan di lokasi daerah Sijunjung yaituTanjung Ampalu. Tanah dibersihkan dariserasah dan dikering anginkan (Raharjo,Mustamir dan Suryadi, 2012). Hasil penelitianpendahuluan pada lokasi pertambangan emasTanjung Ampalu di Sijunjung didapatkankandungan merkuri (Hg) sebesar 1,100 ppm(Wahyuni, Noli dan Febria, 2013,Unpublished).Persiapan media tanam :Tanah tambang emas diambil di lokasi daerahSijunjung yaitu Tanjung Ampalu. Selanjutnyatanah dibersihkan dari serasah dan dikeringanginkan. Lalu, tanah ditimbang sebanyak 3 kgdan dimasukkan kedalam polybag (Syarif et al,2005).

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 85
Penanaman :Anakan tanaman A.conyzoides, D.ciliaris danC.kyllingia ditanam kedalam polibag yangtelah berisi tanah. Satu polibag berisi satuanakan/polibag. Selanjutnya lubang tanam ditutup tanah (Ekawati, Badruzsaufari dan Saidy,2009).
Pemeliharaan meliputi penyiraman danpengendalian gulma. Penyiraman dilakukansetiap pagi atau sore hari. Penyiramandilakukan 1-2 kali sehari. Pengendalian gulmadilakukan dengan cara membersihkan ataumencabut rumput/gulma yang tumbuh disekitartanaman (Ekawati, Badruzsaufari dan Saidy,2009).
Pengamatan meliputi pertambahan tinggitanaman, berat basah tanaman, berat keringtanaman.
Analisis kandungan merkuri (Hg) padatanah dan tanaman : setelah dilakukanpenelitian, perlakuan diuji kandungan merkuripada tanah dan tanaman dengan menggunakanICPE9000 Shimadzu.Analisis data :Data pertambahan tinggi tanaman disajikandalam bentuk grafik. Analisis data tinggitanaman, berat basah dan berat kering tanamandisajikan secara deskriptif.
HASIL DAN PEMBAHASAN
Dari penelitian yang telah dilakukan tentangpotensi beberapa tanaman dalammengakumulasi merkuri pada tanah bekastambang emas didapatkan hasil sebagaiberikut :
Pertambahan tinggi tanaman, berat basahtanaman dan berat kering tanamanDari penelitian yang telah dilakukan tentangpotensi beberapa tanaman dalammengakumulasi merkuri pada tanah bekastambang emas didapatkan hasil sebagaiberikut :
Tabel 1 menunjukkan bahwa tanamanAgeratum conyzoides, Digitaria ciliaris dan
Cyperus kyllingia mampu beradaptasi danmampu tumbuh baik meskipun tumbuh padamedia tercemar merkuri sehinggamemperlihatkan pertumbuhan yang baik yaituterhadap pertambahan tinggi tanaman.Pertambahan tinggi tanaman tertinggi yaituD.cliaris dibandingkan C. kyllingia danA.conyzoides. Hal ini disebabkan ketigatanaman mempunyai kemampuan untukberadaptasi pada lahan yang tercemar dan tiaptanaman memiliki respon yang berbeda dalammeningkatkan tinggi tanaman. Masing-masingtanaman tersebut memiliki daya toleran yangtinggi sehingga mampu tumbuh dan memilikikemampuan adaptasi terhadap tanah yangtercemar oleh kandungan merkuri. Hal inisesuai dengan pendapat Sambas (2002) yangmenyatakan bahwa populasi tumbuhan darifamili Poacea, Cyperacea dan Asteraceamerupakan jenis populasi tanaman yangmampu hidup pada lahan marginal.
Salah satu spesies dari famili Cyperaceaeadalah C. kyllingia. Tumbuhan ini memilikidaya adaptasi yang tinggi, distribusi luas, danmampu tumbuh pada lahan kering maupuntergenang (Rukmana & Saputra, 1999).Menurut Hidayati et al., (2009) menunjukkanbahwa C. kyllingia merupakan spesies tanamanyang mampu mengakumulasi sampai dengan20 ppm Hg.
Tabel 1. Rata-rata pertambahan tinggi tanaman,berat basah dan berat kering tanamanpada tanah tercemar merkuri
PerlakuanParameter
T.T(cm)
BBA(g)
BBB(g)
BKA(g)
BKB(g)
A. conyzoides 12.78 0.23 0.20 0.09 0.05
C. kyllingia 14.26 0.28 0.25 0.10 0.14
D. ciliaris 14.80 0.68 0.26 0.17 0.15
Ket : T.T : Tinggi TanamanBBA : Berat Basah Atas (Tajuk)BBB : Berat Basah Bawah (Akar)BKA : Berat Kering Atas (Tajuk)BKB : Berat Kering Bawah (Akar)

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 86
Famili Poaceae jenis rumput yang mamputumbuh dengan baik di tempat yang miskinhara bahkan di tempat yang banyakmengandung merkuri. Hal ini dikarenakanspesies tersebut memiliki kemampuanmengakumulasi logam. Menurut Hidayati(2005) bahwa merkuri yang dapat diserap olehD. radicosa adalah 50,93 mg/kg bobotkeringnya.
Salah satu spesies dari anggota familiAsteraceae adalah A. conyzoides. Spesies iniditemukan tumbuh subur di kawasanpembuangan limbah atau tailing. Hal inidisebabkan spesies tersebut memiliki sifathipertoleran terhadap lingkungan yangtercemar. Pengamatan pertambahan rata-ratatinggi tanaman sampai akhir pengamatan (8minggu setelah penanaman), dapat dilihatseperti Gambar 1.
Gambar 1. Histogram pertambahan rata-ratatinggi tanaman Ageratumconyzoides, Cyperus kyllingia danDigitaria ciliaris yang ditanampada tanah yang tercemar merkuriselama 8 minggu penanaman
Gambar 1, ketiga tanaman memilikipertambahan tinggi yang berbeda-beda danmempunyai kemampuan yang berbeda untukdapat beradaptasi terhadap tanah tercemarmerkuri dan mampu tumbuh baik. Pertambahantinggi tanaman berjalan cukup lambat padaawal penanaman. Hal ini disebabkan tanamanmasih menyesuaikan diri dengan lingkunganbaru (media tanam) Pada tanaman D. ciliaris,C.kyllingia dan A.conyzoides mengalamipeningkatan pertambahan tinggi setiap
minggunya. Peningkatan pertumbuhan tanamanakan meningkatkan pertumbuhan akar danbatang sehingga kemampuan beradaptasi yangberbeda-beda akan mempengaruhi berat basahtanaman. Pada berat basah atas tanaman, D.cliaris memiliki nilai berat basah atas tertinggi,lalu di ikuti C. kyllingia dan A.conyzoides . Jadi,D. ciliaris mempunyai kemampuan beradaptasiyang lebih baik dibanding jenis tanaman yanglain meskipun tumbuh pada tanah tercemarmerkuri.
Tabel 1 menunjukkan bahwa tanaman A.conyzoides, C. kyllingia dan D. ciliaris yangditanam pada tanah tercemar merkurimempengaruhi berat basah atas dan bawahtanaman. Hal ini diduga tanaman dapatmenyerap air dan hara sesuai dengankebutuhannya sehingga dapat mengakibatkanproses fotosintesis dapat berjalan dengan lancar.Selain itu, tanaman tersebut diduga mampumenyerap dan mengakumulasi logam beratmerkuri pada tanah. Produksi biomas tinggimerupakan salah satu karakteristik yangdiharapkan untuk tanaman hiperakumulator.Diharapkan dengan produksi biomasa yangtinggi tanaman dapat mengakumulasi polutandalam jumlah lebih besar sehingga dapat lebihefektif untuk membersihkan polutan dari dalamtanah.
Tabel 1 juga dijelaskan pada parameterberat basah bawah dapat dilihat bahwaperlakuan A. conyzoides, C. kyllingia dan D.ciliaris berpengaruh terhadap berat basahbawah tanaman. D. ciliaris memiliki beraatbasah bawah tertinggi dibandingkan A.conyzoides, C. kyllingia. Hal ini diduga karenapertumbuhan tanaman dipengaruhi faktorlingkungan tempat tumbuh dan mediatumbuhnya. D. ciliaris mampu menyerap unsurhara beserta logam dalam tanah melalui akar.Akar berfungsi sebagai organ penyerap unsur-unsur hara dan logam berat danmengalirkannya ke bagian batang dan daun.Tanaman yang mampu tumbuh pada tanahdengan kandungan logam berat yang tinggiharus mempunyai daya tolerasi yang tinggi dan
0
1
2
3
4
5
1 2 4 6 8
Ting
gi ta
nam
an (c
m)
minggu ke-
A. conyzoides
C. kyllingia
D. ciliaris

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 87
memiliki struktur perakaran yang panjang dandalam (Hidayati, 2005).
Tabel 1 juga dijelaskan bahwa A.conyzoides,C. kyllingia dan D. ciliaris memilikikemampuan beradaptasi yang tinggi meskipunditanam pada tanah tercemar merkurimenunjukkan pertumbuhan yang baik sehinggadapat mendukung fungsi fisiologis denganbaik dan dapat meningkatkan berat keringtanaman. Hal ini diduga karena kemampuansetiap tanaman berbeda-beda dapat menyeraphara mineral dan logam berat yang terkandungdalam tanah sehingga menghasilkan produkfotosintesis yang berbeda pula. Selain itu, akartanaman merupakan organ tanaman yangberfungsi sebagai penyerap unsur hara dansekaligus organ yang kontak langsung denganmedia tanam yang tercemar merkuri. Hal inisesuai dengan pernyataan Dwidjoseputro(1994), yang menyatakan bahwa pertumbuhanorgan-organ tanaman seperti akar, batang, dandaun akan menentukan bobot kering tanaman.Tabel 1 juga dapat dijelaskan bahwa D.ciliarismemiliki berat kering bawah tanaman tertinggi,lalu diikuti C.kyllingia dan A.conyzoides.Masing-masing tanaman mampu menyeraphara dalam media tumbuhnya sehingga dapatmeningkatkan metabolisme tanaman danmenunjukkan adanya akumulasi senyawaorganik yang berhasil disintesis tanaman danmengakibatkan terjadinya penumpukan dalamtanaman sehingga berpengaruh terhadap beratkering tanaman. Goldsworthy dan Fisher(1992) menyatakan berat kering tanamanmerupakan gambaran tumbuhan dalammemanfaatkan nutrisi, air dan cahaya. Beratkering tanaman erat kaitannya dengankemampuan akar dalam menyerap air danunsur yang terdapat dalam media tumbuhnya.Pengangkutan hasil fotosintesis ke akarmenentukan kemampuan akar untuk menyerapdan memperoleh hara (Fitter & Hay, 1991).
Menurut Salisbury dan Ross (1995),besarnya berat kering menggambarkan statusnutrisi yang diserap, dimana unsur tersebutakan dimanfaatkan dalam proses metabolisme
atau akumulasi senyawa organik yang berhasildisintesis. Sarief (1986) menyatakan bahwatanaman yang mengalami pertumbuhan yangbaik dapat mengaktifkan proses fotosintesissehingga perpanjangan, pembelahan dandiferensiasi sel akan lebih baik dan akanmempengaruhi pertumbuhan seperti pening-katan berat kering tanaman.
Kandungan Merkuri Pada Tanah DanTanaman
Setelah dilakukan penelitian maka dapatdiketahui kandungan merkuri pada tanah dantanaman. Kandungan merkuri yang terdapatdalam tanah semakin menurun. Hal ini didugatanaman mampu menyerap kandungan Hg lebihbanyak sehingga terjadi penurunan kandunganHg dalam tanah dan tanaman A.conyzoides,C.kyllingia dan D.ciliaris mampumengakumulasi merkuri pada bagian tanamanyaitu tajuk dan akar. Menurut Chaney (1995)bahwa semua tumbuhan memiliki kemampuanmenyerap logam dalam jumlah yang bervariasi.Hal ini dapat dilihat pada gambar 2, 3 dan 4.Tanaman A. conyzoides, C. kyllingia dan D.ciliaris tumbuh diketahui tidak hanya menyeraphara dan air didalam tanah, akan tetapi tanamantersebut ternyata juga mampu menyerap Hgpada tanah tercemar merkuri, sehingga terjadipenurunan kandungan Hg didalam tanah. Halini berarti bahwa tanaman tersebut memilikipotensi sebagai tanaman hiperakumulator.Dari Gambar 2, 3 dan 4 dapat dilihat bahwanilai serapan Hg pada tanaman A.conyzoides,C.kyllingia dan D.ciliaris memberikan nilaiyang bervariasi pada bagian tajuk dan akar.Adapun rata-rata kadar Hg pada akar di akhirpengamatan (minggu ke- 8) pada A.conyzoides0.012 ppm , C.kyllingia 0.015 ppm danD.ciliaris 0.022 ppm. Sedangkan rata-ratakadar Hg pada tajuk di akhir pengamatan(minggu ke- 8) pada A.conyzoides 0.01 ppm , C.kyllingia 0.0135 ppm dan D. ciliaris 0.016 ppm.Hal ini berarti tanaman mampu menyerap danmengakumulasi logam berat merkuri dan masih
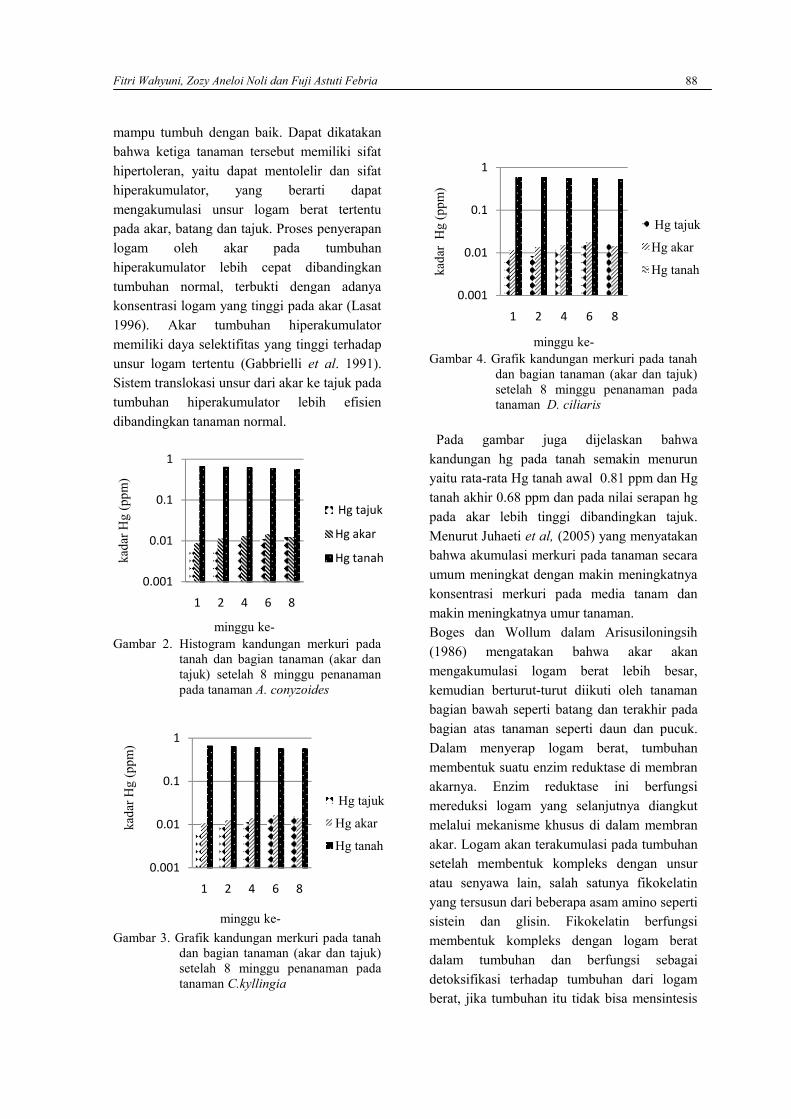
Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 88
mampu tumbuh dengan baik. Dapat dikatakanbahwa ketiga tanaman tersebut memiliki sifathipertoleran, yaitu dapat mentolelir dan sifathiperakumulator, yang berarti dapatmengakumulasi unsur logam berat tertentupada akar, batang dan tajuk. Proses penyerapanlogam oleh akar pada tumbuhanhiperakumulator lebih cepat dibandingkantumbuhan normal, terbukti dengan adanyakonsentrasi logam yang tinggi pada akar (Lasat1996). Akar tumbuhan hiperakumulatormemiliki daya selektifitas yang tinggi terhadapunsur logam tertentu (Gabbrielli et al. 1991).Sistem translokasi unsur dari akar ke tajuk padatumbuhan hiperakumulator lebih efisiendibandingkan tanaman normal.
Gambar 2. Histogram kandungan merkuri padatanah dan bagian tanaman (akar dantajuk) setelah 8 minggu penanamanpada tanaman A. conyzoides
Gambar 3. Grafik kandungan merkuri pada tanahdan bagian tanaman (akar dan tajuk)setelah 8 minggu penanaman padatanaman C.kyllingia
Gambar 4. Grafik kandungan merkuri pada tanahdan bagian tanaman (akar dan tajuk)setelah 8 minggu penanaman padatanaman D. ciliaris
Pada gambar juga dijelaskan bahwakandungan hg pada tanah semakin menurunyaitu rata-rata Hg tanah awal 0.81 ppm dan Hgtanah akhir 0.68 ppm dan pada nilai serapan hgpada akar lebih tinggi dibandingkan tajuk.Menurut Juhaeti et al, (2005) yang menyatakanbahwa akumulasi merkuri pada tanaman secaraumum meningkat dengan makin meningkatnyakonsentrasi merkuri pada media tanam danmakin meningkatnya umur tanaman.Boges dan Wollum dalam Arisusiloningsih(1986) mengatakan bahwa akar akanmengakumulasi logam berat lebih besar,kemudian berturut-turut diikuti oleh tanamanbagian bawah seperti batang dan terakhir padabagian atas tanaman seperti daun dan pucuk.Dalam menyerap logam berat, tumbuhanmembentuk suatu enzim reduktase di membranakarnya. Enzim reduktase ini berfungsimereduksi logam yang selanjutnya diangkutmelalui mekanisme khusus di dalam membranakar. Logam akan terakumulasi pada tumbuhansetelah membentuk kompleks dengan unsuratau senyawa lain, salah satunya fikokelatinyang tersusun dari beberapa asam amino sepertisistein dan glisin. Fikokelatin berfungsimembentuk kompleks dengan logam beratdalam tumbuhan dan berfungsi sebagaidetoksifikasi terhadap tumbuhan dari logamberat, jika tumbuhan itu tidak bisa mensintesis
0.001
0.01
0.1
1
1 2 4 6 8
kada
r Hg
(ppm
)
minggu ke-
Hg tajuk
Hg akar
Hg tanah
0.001
0.01
0.1
1
1 2 4 6 8
kada
r Hg
(ppm
)
minggu ke-
Hg tajuk
Hg akar
Hg tanah
0.001
0.01
0.1
1
1 2 4 6 8
kada
r H
g (p
pm)
minggu ke-
Hg tajuk
Hg akar
Hg tanah

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 89
fikokelatin menyebabkan terhambatnyapertumbuhan dan berakhir pada kematian,kadar tinggi fikokelatin ditemukan padatumbuhan yang toleran terhadap logam berat.Mekanisme penyerapan dan akumulasi logamberat oleh tumbuhan dibagi menjadi tiga prosesyang berkesinambungan yaitu (1) penyerapanoleh akar lewat pembentukan suatu zat khelatyang akan mengikat logam dan membawanyake dalam sel akar; (2) translokasi logam dariakar ke bagian lain tumbuhan melalui jaringanpengangkut yaitu xylem dan floem dan (3)lokalisasi logam pada bagian sel tertentu untukmenjaga agar tidak menghambat metabolismtumbuhan tersebut (Syahputra, 2005).
KESIMPULAN
Berdasarkan penelitian yang telah dilakukantentang potensi beberapa tanaman dalammengakumulasi merkuri pada tanah bekastambang emas didaerah Tanjung Ampalu dapatdisimpulkan sebagai berikut :1. Tanaman Ageratum conyzoides, Digitaria
ciliaris dan Cyperus kyllingia berpotensidalam meremediasi lahan tambang emasyang tercemar merkuri (Hg). Adapun kadarHg pada akar di akhir pengamatan pada A.conyzoides 0.012 ppm , C. kyllingia 0.015ppm dan D. ciliaris 0.022 ppm. Kadar Hgpada tajuk di akhir pengamatan pada A.conyzoides 0.01 ppm , C. kyllingia 0.0135ppm dan D. ciliaris 0.016 ppm.
2. Digitaria ciliaris merupakan tanamanterbaik dalam meremediasi lahan tambangemas yang tercemar merkuri (Hg). D.ciliarismempunyai kemampuan mengakumulasimerkuri yang lebih baik dibanding jenistanaman yang lain.
DAFTAR PUSTAKA
Aminudin S. 2006. Polutan Logam Berat.Tesis:Institut Pertanian Bogor. Bogor.
Arisusiloningsih, E. 1986. Pengaruh Timbal(Pb) dan Cadmium (Cd) TerhadapPertumbuhan Tanaman Kedelai (Gleicyne
max (L.). (Merr), Tesis S2, jurusanBiologi. ITB, Bandung.
Chaney RL et al. 1995. Potential use of metalhyperaccumulators. Mining EnvironManag (3):9-11.
Dinas Pertambangan dan Energi ProvinsiSumatera Barat, 2004. Potensi BahanGalian Sumatera Barat, Padang.
Dwidjoseputro, D. 1994. Pengantar FisiologiTumbuhan. PT Gramedia Pustaka Utama.Jakarta.
Ekawati, Y, Badruzsaufari dan Saidy. R. A.2009. Pengaruh Gypsum dan Pupuk FosfatTerhadap Penyerapam Logam Cromiumdan Nikel Serta Pertumbuhan TanamanJarak (Jatropha curcas L.) Di TanahSerpentin. Bioscientiae. 6 (1): 16-25.
Fitter, A.H & Hay, R.K.M. 1991. FisiologiLingkungan Tanaman. Terjemahan olehSri Andani dan E.D. Purbayanti.Universitas Gadjah Mada Press.Yogyakarta.
Gabbrielli R, Mattioni C, Vergnano O. 1991.Accumulation mechanisms and heavymetal tolerance of a nickelhyperaccumulator. J Plant Nutr.(14):1067-1080.
Goldsworthy, P. R. dan N. M. Fisher. 1992.Fisiologi Tanaman Budidaya Tropik.Gajah Mada University Press :Yogyakarta.
Handayanto, E. 2012. Fitoremediasi TanahTercemar Merkuri Limbah TambangEmas Rakyat untuk Perbaikan ProduksiJagung. Lembaga Penelitian DanPengabdian Kepada Masyarakat.Universitas Brawijaya.
Hidayati N. 2005. Fitoremediasi dan PotensiTumbuhan Hiperakumulator. Hayati. 12(1):35-40.
Hidayati, N., Juhaeti, T. and Syarif, F. 2009.Mercury and Cyanide Contaminations inGold Mine Environment and PossibleSolution of Cleaning Up by UsingPhytoextraction. Hayati Journal ofBiosciences 16: 88-94.
Juhaeti, T. dan F. Syarif. 2003. Studi PotensiBeberapa Jenis Tumbuhan Air UntukFitoremediasi. Pusat Penelitian BiologiLipi. Bogor.
Juhaeti, T., F. Syarif dan N. Hidayati. 2005.Inventarisasi Tumbuhan Potensial UntukFitoremediasi Lahan Dan AirTerdegradasi Penambangan Emas.Biodiversitas. 6 (1): 31-33.

Fitri Wahyuni, Zozy Aneloi Noli dan Fuji Astuti Febria 90
Lasat MM, Baker AJM, dan Kochian LV.1996. Physiological characterization ofroot Zn2+ absorption and translocation toshoot in Zn hyperaccumulator andnonaccumulator species of Thlaspi. PlantPhysiol. (112) :1715-1722.
Raharjo, D. Mustamar, E dan Suryadi, U, D.2012. Uji fektivitas Beberapa Jenis ArangAktif dan Tanaman Akumulator LogamPada Lahan Bekas Penambangan Emas.Jurnal Perkebunan dan Lahan Tropika.Vol 2 (2) : 15 – 22.
Rukmana, H.R. dan U.S. Saputra. 1999. Gulmadan Tehnik Pengendalian. Kanisius.Jakarta
Salisbury, F. B. dan C. W. Ross. 1995. PlantFisiology. Wadswosth PublishingCompany, Inc. California.
Sambas, E. N. 2002. Analisis Vegetasi Tumbuhan Bawah Pada Areal Tailing Dam PT.Aneka Tambang (Antam) Pongkor.[Laporan Teknik]. Bogor: Pusat PenelitianBiologi, LIPI. Bogor.
Sarief. E.S. 1986. Kesuburan dan PemupukanTanah Pertanian. Pustaka Buana.Bandung.
Setiabudi, B. T. 2005. Penyebaran MerkuriAkibat Usaha Pertambangan Emas DiDaerah Sangon, Kabupaten Kulon Progo,D.I. Yogyakarta. Subdit Konservasi.Kolokium Hasil Lapangan.
Syahputra R, 2005. Fitoremediasi Logam Cudan Zn dengan Tanaman Enceng Gondok.Jurnal Logika. Vol 2 (2) : 57– 67.
Syarif., T. Juhaeti, N, Hidayati, E, Komarudindan Suwanto. 2005. Screening BeberapaJenis Tumbuhan yang ditanam PadaMedia Limbah Penambangan EmasCikotok Untuk Fitoremediasi. LaporanTeknik. Bidang Botani. Pusat PenelitianBiologi-LIPI.
Wahyuni, F., Z. A. Noli., F. A. Febria. 2013.Survey Pendahuluan Kandungan LogamBerat Merkuri (Hg) Pada Tanah BekasTambang Emas di Sijunjung SumateraBarat. Unpublished.

ISSN 978-602-14989-0-3 BioETI
Isolasi dan uji resistensi merkuri bakteri endogen tanah bekastambang emas Kabupaten Sijunjung
FUJI ASTUTI FEBRIA, ANTHONI AGUSTIEN DAN S.P. RAHAYU
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKKegiatan penambangan emas menyisakan merkuri di lingkungan sekitarnya. Akumulasi merkuri melampaui ambangbatas mengancam kesehatan dan lingkungan. Diperlukan upaya remediasi lingkungan, salah satunya menggunakanbakteri. Pada tanah bekas tambang emas secara alamiah terdapat sekelompok bakteri endogen toleran terhadap merkuri.Penelitian ini bertujuan untuk menemukan isolat bakteri resisten merkuri. Penelitian menggunakan metoda eksperimen.Sampel tanah diambil langsung pada salah satu lokasi bekas penambangan emas di Kabupaten Sijunjung yang kandunganmerkuri tanah telah dianalisa sebelumnya. Isolasi menggunakan teknik dillution series, penanaman isolat secara pourplate. Uji resitensi isolat dilakukan pada medium yang ditambahkan merkuri pada konsentrasi bertingkat, mulai 50, 100,150 dan 200 ppm. Hasil penelitian ditemukan 7 isolat bakteri endogen tanah bekas tambang emas. Uji resistensi isolatterhadap merkuri menunjukkan bahwa 5 dari 7 isolat yang ditemukan resisten pada konsentrasi merkuri tertinggi padaperlakuan 200 ppm.
Key words: isolasi, bakteri, resisten, merkuri, tambang emas
Pendahuluan
Cabai Propinsi Sumatera Barat memilikipotensi sumber daya mineral seperti emas. DataDinas Pertambangan dan Energi PropinsiSumbar (2004) deposit emas tersebar padaKabupaten Lima Puluh Kota, Pasaman, PesisirSelatan, dan Sijunjung (Refles, 2012).Perkembangan penambangan emas diKabupaten Sijunjung sepuluh tahun terakhirsangat pesat dan mendapat perhatian banyakkalangan, karena umumnya kegiatan tersebuttanpa dilengkapi ijin operasional (illegalminning). Kekhawatiran illegal minning yangpaling mendasar adalah tidak diketahui secarapasti jumlah merkuri yang digunakan dalamproses pemisahan biji emas dengan teknikamalgamasi. Akibatnya merkuri terpapar dilingkungan tanah. Merkuri di tanah bisaberubah menjadi merkuri anorganik danmerkuri organik oleh aktifitas mikrobia. Logammerkuri mudah menguap, dan bisa kembali kebumi melalui hujan asam. Merkuri organik dananorganik dapat masuk ke lingkungan perairandan terakumulasi dalam tubuh biota air melaluirantai makanan (Barkay, 2001). Menurut Roger
(1984) merkuri yang masuk ke dalam tubuhterus menerus akan menyebabkan kerusakanpermanen pada otak, hati dan ginjal.
Hasil analisis kandungan merkuri padatanah bekas tambang emas di salah satu lokasipenambangan emas di Kabupaten Sijunjungadalah 1,11 ppm (Siregar dkk., 2013),melampaui nilai baku mutu kandungan merkuriditanah menurut WHO 0,001 ppm (Utomo etal., 2014), sehingga dibutuhkan suatu cara yangtepat dan ramah lingkungan dalam upayapemulihan lahan tercemar merkuri. Salah satuupaya pemulihan lingkungan dilakukan denganmenggunakan bakteri resisten merkuri. Padakawasan tercemar merkuri secara alamiahterdapat bakteri endogen yang toleran terhadapmerkuri. Bakteri ini memiliki kemampuandalam mendetoksifikasi merkuri menjadi tidakberbahaya. Mekanisme resistensi mikrobadapat dilakukan melalui biotransformasi(melalui oksidasi – reduksi), biopresipitasi (ionlogam dipresipitasi pada permukaan sel melaluimekanisme mikrobial seperti efflux kation ataumengubah pH) dan biosorpsi (menggunakanbiomass mikroba alami atau rekombinan untukadsorpsi ion metal) (Hughes & Poole, 1989).

Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 92
Beberapa penelitian seperti Immamudin (2010)bahwa bakteri Ochrobactrum sp. s79 resistenmerkuri konsentrasi 90 ppm. Manampiring(2011) mendapatkan satu isolat bakteri resistenmerkuri pada konsentrasi 100 ppm.
Sejauh ini belum ada informasi isolatbakteri resisten merkuri pada tanah di kawasanbekas tambang emas Kabupaten Sijunjung.Untuk itu dilakukan penelitian dengan tujuanuntuk menemukan isolat bakteri resistenmerkuri endogen pada tanah bekas tambangemas dan mengetahui kemampuannya tumbuhpada konsentrasi merkuri secara bertingkat.Penelitian ini bermanfaat sebagai basis kajianbioremediasi kawasan tercemar merkuriterutama pada lahan bekas tambang emas.
BAHAN DAN METODE
Bahan yang digunakan dalam penelitian iniantara lain : Medium Nutrient Agar (NA),aquadest steril, Merkuri (HgCl2), alkohol, danspritus. Alat-alat yang digunakan untukmelakukan penelitian ini antara lain : palstiksampel 2 x 2 kg, petri dish, tabung reaksi,mikropipete, alumunium foil, plastik wrap,erlenmeyer, jarum ose, bunsen, kapas, kainkasa, box alat, wadah plastik, tissue, GPS, pipettetes, gelas piala, autoklaf, vortex.
Penelitian menggunakan metodaeksperimen. Tahapan penelitian meliputi:pengambilan sampel tanah pada salah satulokasi bekas tambang emas di KabupatenSijunjung yang sebelumnya telah dilakukananalisis kandungan merkurinya.
Selanjutnya, isolasi bakteri menggunakanteknik dilution series. Inokulasi dilakukansecara pour plate pada medium NA modifikasimerkuri 10 ppm. Setelah 24 jam inkubasi,koloni yang tumbuh diamati dan dilanjutkanpemurnian isolat bakteri dengan teknik streakplate secara kuadran sampai diperoleh kulturmurni (Cappuchino&Sherman, 2005; Zulaika,2011).
Tahap terakhir dari penelitian adalah ujiresistensi isolat bakteri endogen tanah bekastambang emas terhadap kandungan merkuri
secara bertingkat. Konsentrasi yang digunakanadalah konsentrasi 50, 100, 150, dan 200 ppm.Pengujian dilakukan dengan metoda pour plate.Pengatan bakteri resisten merkuri dilakukansetelah inkubasi pada suhu ruang selama 24jam (Zulaika, 2011). Data yang diperoleh daripenelitian disajikan dalam bentuk tabel dangambar dan disajikan secara deskriptif.
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa padatahap isolasi bakteri endogen tanh bekastambang emas ditemukan 7 isolat bakteri. Salahsatu foto proses pengisolasian bakteriditunjukkan pada Gambar 1.Ditemukannya koloni bakteri yang tumbuhpada tahap isolasi menujukkan bahwa kolonibakteri dan seluruh aktifitas metabolismenyateradaptasi terhadap cekaman merkuri. MenurutGadd (2000) bahwa koloni bakteri yangditemukan pada tahap isolasi merupakan kolonibakteri yang memiliki kemampuan adaptasisecara fisiologis dan genetis terhadap merkuripada lingkungan alamiahnya (Gadd, 2000).Menurut Canstein et al (2002) bakteri yangdapat tumbuh pada medium sintetis denganpenambahan HgCl2 dengan konsentrasiminimal 5 ppm merupakan bakteri resistenmerkuri. Dalam hal ini bakteri yang didapatkandari pengisolasian dalam penelitian inimerupakan bakteri resisten merkuri dengankode isolat BRM 1, BRM 2, BRM 3, BRM 4,BRM 5, BRM 6, BRM 7, karena isolat bakteritersebut diisolasi dengan menggunakanmedium NA modifikasi merkuri 10 ppm. Setiapkoloni bakteri yang didapatkan memilikikarakter morfologi yang berbeda (Tabel 1).
Uji resistensi isolat bakteri endogen padakonsentrasi merkuri secara bertingkat diketahuibahwa kemampuan masing-masing isolat tidaksama dalam menghadapi cekaman merkuri.Dari 7 isolat yang diujikan 5 diantaranyamampu tumbuh hingga konsentrasi merkuripada konsentrasi 200 ppm, yaitu ; Isolat BRM 1,BRM 2, BRM 3, BRM 5 dan BRM 6 (Tabel 2).

Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 93
Gambar 1. Salah satu koloni bakteri hasil isolasi menggunakan NA modifikasi merkuri
Tabel 1. Karakter makroskopis isolat bakteri resisten merkuri endogen pada tanah bekas tambang emas
Kode isolat Pengamatan Secara MakroskopisBentuk Pinggiran Elevasi Warna koloni
BRM 1 Bulat Rata Timbul orangeBRM 2 Tidak beraturan Bergelombang dalam Timbul orangeBRM 3 Bulat Rata Timbul merah bataBRM 4 Bulat Bergelombang Datar kremBRM 5 Tidak beraturan Rata Datar putih kekuninganBRM 6 Bulat Bergelombang Timbul Putih susuBRM 7 Bulat Bergelombang Timbul kuning
Gambar 2. Tujuh isolat bakteri hasil isolasi menggunakan media NA modifikasi penambahan merkuri yangmemiliki karakter morfologi berbeda
Tabel 2. Uji resistensi isolat bakteri indigen pada tanah bekas tambang emas pada konsentrasi merkuri secarabertingkat
Kode Isolat Bakteri Konsentrasi Merkuri50 ppm 100 ppm 150 ppm 200 ppm
BRM 1 +++ +++ +++ +++BRM 2 +++ +++ +++ ++BRM 3 +++ +++ ++ +BRM 4 +++ ++ - -BRM 5 +++ +++ ++ +BRM 6 +++ +++ ++ +BRM 7 +++ ++ + -
Ket : +++(tunbuh banyak) ++ (tumbuh sedang), + (tumbuh) dan - (tidak tumbuh)
BRM 1 BRM 2 BRM 3
BRM 6BRM 5
BRM 4
BRM 7
Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 93
Gambar 1. Salah satu koloni bakteri hasil isolasi menggunakan NA modifikasi merkuri
Tabel 1. Karakter makroskopis isolat bakteri resisten merkuri endogen pada tanah bekas tambang emas
Kode isolat Pengamatan Secara MakroskopisBentuk Pinggiran Elevasi Warna koloni
BRM 1 Bulat Rata Timbul orangeBRM 2 Tidak beraturan Bergelombang dalam Timbul orangeBRM 3 Bulat Rata Timbul merah bataBRM 4 Bulat Bergelombang Datar kremBRM 5 Tidak beraturan Rata Datar putih kekuninganBRM 6 Bulat Bergelombang Timbul Putih susuBRM 7 Bulat Bergelombang Timbul kuning
Gambar 2. Tujuh isolat bakteri hasil isolasi menggunakan media NA modifikasi penambahan merkuri yangmemiliki karakter morfologi berbeda
Tabel 2. Uji resistensi isolat bakteri indigen pada tanah bekas tambang emas pada konsentrasi merkuri secarabertingkat
Kode Isolat Bakteri Konsentrasi Merkuri50 ppm 100 ppm 150 ppm 200 ppm
BRM 1 +++ +++ +++ +++BRM 2 +++ +++ +++ ++BRM 3 +++ +++ ++ +BRM 4 +++ ++ - -BRM 5 +++ +++ ++ +BRM 6 +++ +++ ++ +BRM 7 +++ ++ + -
Ket : +++(tunbuh banyak) ++ (tumbuh sedang), + (tumbuh) dan - (tidak tumbuh)
BRM 1 BRM 2 BRM 3
BRM 6BRM 5
BRM 4
BRM 7
Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 93
Gambar 1. Salah satu koloni bakteri hasil isolasi menggunakan NA modifikasi merkuri
Tabel 1. Karakter makroskopis isolat bakteri resisten merkuri endogen pada tanah bekas tambang emas
Kode isolat Pengamatan Secara MakroskopisBentuk Pinggiran Elevasi Warna koloni
BRM 1 Bulat Rata Timbul orangeBRM 2 Tidak beraturan Bergelombang dalam Timbul orangeBRM 3 Bulat Rata Timbul merah bataBRM 4 Bulat Bergelombang Datar kremBRM 5 Tidak beraturan Rata Datar putih kekuninganBRM 6 Bulat Bergelombang Timbul Putih susuBRM 7 Bulat Bergelombang Timbul kuning
Gambar 2. Tujuh isolat bakteri hasil isolasi menggunakan media NA modifikasi penambahan merkuri yangmemiliki karakter morfologi berbeda
Tabel 2. Uji resistensi isolat bakteri indigen pada tanah bekas tambang emas pada konsentrasi merkuri secarabertingkat
Kode Isolat Bakteri Konsentrasi Merkuri50 ppm 100 ppm 150 ppm 200 ppm
BRM 1 +++ +++ +++ +++BRM 2 +++ +++ +++ ++BRM 3 +++ +++ ++ +BRM 4 +++ ++ - -BRM 5 +++ +++ ++ +BRM 6 +++ +++ ++ +BRM 7 +++ ++ + -
Ket : +++(tunbuh banyak) ++ (tumbuh sedang), + (tumbuh) dan - (tidak tumbuh)
BRM 1 BRM 2 BRM 3
BRM 6BRM 5
BRM 4
BRM 7

Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 94
Kemampuan tumbuh yang dimiliki oleh limaisolat bakteri dalam menghadapi cekamanmerkuri diduga karena memiliki kemampuanbaik dalam transformasi, imobilisasi, metilasibahkan mereduksi merkuri secara enzimatisyang bisa merubah merkuri toksik (Hg2+ )menjadi Hg0 yang tidak toksik sehingga bakteritetap bertahan hidup hingga konsentrasi 200ppm. Pada isolat bakteri kode BRM 4 danBRM 7 hanya mampu hidup sampaikonsentrasi 100 ppm dan 150 ppm. Didugakemampuan yang dimiliki kedua isolat tersebutsama dengan lima isolat bakteri yang hiduppada konsentrasi 200 ppm, namun kemampuanyang dimiliki oleh bakteri tersebut, tidak bisamerubah seluruh Hg2+ menjadi Hg0 yang tidaktoksik.
Jika dilihat dari segi penghasilan kadarprotein enzim, kelima isolat yang mamputumbuh hingga konsentrasi tertinggimenghasilkan enzim yang lebih banyakdibandingkan dengan dua isolat yang hanyamampu hidup hingga konsentrasi 100 dan 150ppm. Sehingga masih ada kemungkinan sisa-sisa dari Hg2+ yang tidak diubah sempurna danmenyebabkan bakteri tidak mampu bertahanhidup dalam cekaman merkuri yangkonsentrasinya besar dari 100 ppm dan 150ppm. Madigan (2006), menyatakan bahwabakteri resisten merkuri memiliki mekanismeenzimatis yang dapat mengubah senyawamerkuri menjadi merkuri tidak toksik (Hg0),agar senyawa merkuri tersebut tidak meracunitubuh bakteri. Mekanisme enzimatismerupakan salah satu bagian dari siklus dasarreaksi merkuri yang dilakukan oleh bakteri.Dimana reaksi antara merkuri dengan bakterimeliputi transformasi aerobik dan anaerobikyaitu dari Hg2+ ke monometil Hg danselanjutnya menjadi metana dan Hg0. Prosesmetilisasi ini merupakan awal prosesdetoksifikasi bagi bakteri bersangkutan yangkemudian dilanjutkan dengan proses reduksiHg2+ menjadi Hg0 yang melibatkan duamacam enzim yaitu merkuri reduktase danorganomerkuri liase (Barkay, 2001).
KESIMPULAN
Berdasarkan hasil penelitian dapat disimpulkanbahwa :1. Ditemukan isolat dan kemampuannya
tumbuh pada konsentrasi merkuri secarabertingkat.
2. Ditemukan 7 isolat bakteri resisten merkuriendogen pada tanah bekas tambang emasmenggunakan medium NA modifikasimerkuri
3. Didapatkan 5 dari 7 isolat bakteri resistenmerkuri endogen pada tanah bekas tambangemas yang mampu bertahan hinggakonsentrasi merkuri tertinggi yaitu 200 ppmpada uji resistensi merkuri.
DAFTAR PUSTAKA
Barkay, T. and Miller S.M. 2003. Bacterialmercury resistance from atoms toecosystems. FEMS Microbiology Reviews27:355-384.
Canstein, H.V., Li, Y., Leonhauser, J., Haase,E., Felske, A., Deckwer, W.D. and Dobler,I.W,. 2002. Spatially Oscillating Activityand Microbial Succession of Mercury-Reducing Biofilms in a Technical-ScaleBioremediation System. Appl. Environ.Microbiol. 68: 1938- 1946 .
Cappuccino, J.G. and Sherman. 2005.Microbiolgy a Laboratory Manual. 7th Ed.Pearson Education, Inc., Pubhlishing asBenjamin Cummings. San Fransisco.
Gadd, G.M. 2000. Heavy Metal Pollutants;Environmental and BiotechnologicalEspect. Encyclopedia of Microbiology 2nd.Ed 2 : 607 –617.
Hughes, M.N. and Poole. 1989. Metals andmicroorganism. Chapman and Hall.London.
Immamudin, H. 2010. Pola Pertumbuhan danToksisitas Bakteri Resisten HgCl2Ochrobactrum sp. s79 dari Cikotok,Banten. Mikrobiologi. LIPI. Vol. II.
Madigan, M.T. and Martinko, J.M. 2006. BrockBiology of Microorganism 11th. PearsonEducation Inc. New Jersey.
Manampiring, A.E. dan Keppel, B.J. 2011.Studi Populasi Bakteri Resisten MerkuriDi Daerah Aliran Sungai Tondano,

Fuji Astuti Febria, Anthoni Agustien dan S.P. Rahayu 95
Kelurahan Ketang Baru, Manado. JurnalIlmiah Sains. Vol.11.(1).
Nascimento, A.M.A. and Chartone Souza E.2003. Operon mer: Bacterial resistance tomercury and potential for bioremediationof contaminated environments. Genet MolRes 2:92-101.
Roger A.R, and Lawrence H.K. 1984. WaterAnalysis: Inorganic Spesies 2nd Ed.:Academic Press. Orlando Florida.
Siregar, D.A. 2013. Pengaruh Pemberian FMA(Mikoriza Arbuskula) Pada TanamanCalopogonium muconoides danCentrosema pubescens UntukBioremediasi Lahan Tercemar Merkuri.Jurusan Biologi. FMIPA UNAND. Padang(Unpublish).
Utomo, W.H., Retno, S., Novi, A., Suhartini.dan Handayanto. 2014. Rehabilitation ofArtisanal Small-Scale Gold Mining Land
in West Lombok, Indonesia, Explorationof Endogenous Plant Species and TheAssociated Mycorrhiza ForPhytomycoremediation of MercuryContaminated Soils. American-EurasianJournal of Sustainable Agriculture. ISSN:1995-0748.
Vouck. 1986. General Chemistry of Metal.Handbook on the Toxicology of Metal.Elsivier. New York.
Widiyanti, A., Maya, S. dan Nengah, D. 2011.Isolasi Dan Karakterisasi Bakteri ResistenMerkuri Di Hilir Kalimas Surabaya.Institute Teknologi Surabaya Press.Surabaya.
Zulaika, E., Atik, W. dan Maya, S. 2011.Bakteri Resisten Merkuri Endogenik HilirKalimas Surabaya. Institute TeknologiSurabaya Press. Surabaya.

ISSN 978-602-14989-0-3 BioETI
Kandungan Polifenol dan Aktivitas Antioksidan Jamur Tiram(Pleourotus spp.) Beraneka Warna
HAFIZATUR RAHMA, NURMIATI* DAN ANTHONI AGUSTIEN
Laborat Riset Mikrobiologi, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: *[email protected]
ABSTRAK
Penelitian ini dilakukan untuk menentukan pengaruh pencucian media tanam terhadap keberadaan polifenol dan aktifitasantioksidan tubuh buah masing-masing jamur tiram (Pleurotus ostreatus, P.sajor-caju, P. flabellatus, P.cystidiosus) danmengetahui jenis jamur tiram yang mempunyai kandungan polifenol dan aktifitas antioksidan tertinggi.Penelitiandilakukan dalam Rancangan Acak Lengkap (RAL) pola Nested dengan 2 faktor perlakuan dan 4 ulangan. Faktor Aadalah perlakuan media tanam yaitu dengan pencucian dan tanpa pencucian media. Sedangkan faktor B adalah beberapajenis jamur tiram yaitu tiram putih (P. ostreatus), tiram pink (P. flabellatus), tiram kelabu (P.sajor-caju)dan tiram cokelat(P. cystidiosus).Hasil penelitian menunjukkan bahwa perlakuan tanpa pencucian media serbuk gergaji memberikan rata-rata total polifenol tertinggi dengan jenis Jamur tiram kelabu (1901,25 mg GAC/g). Sedangkan dalam aktivitasantioksidan, perlakuan dengan pencucian justru memberikan nilai terbaik pada jenis Jamur tiram pink 11.607,64 (mg/mL).
Key words: Pencucian, Polifenol, Aktivitas Antioksidan dan Jamur Tiram
Pendahuluan
Dewasa ini, dunia kesehatan dan kedokterantelah banyak membahas tentang keberadaanradikal bebas (free radical) dan antioksidan.Hal ini dikarenakan sebagian besar penyakityang diderita diawali oleh adanya reaksioksidasi yang berlebihan pada dalam tubuh.Reaksi oksidasi ini akan menyebabkanterbentuknya radikal bebas yang sangat aktifyang dapat merusak fungsi dan stuktur sel.Radikal bebas merupakan salah satu bentuksenyawa oksigen reaktif yang secara umumdiketahui sebagai senyawa yang memilikielektron yang tidak berpasangan (Halliwell etal., 1992). Selain itu radikal bebas juga bisaterbentuk dari senyawa lain yang sebenarnyabukan radikal bebas tetapi mudah berubahmenjadi radikal bebas. Dinegara tropis danberkembang seperti Indonesia, terjadinyapenyakit infeksi oleh virus, bakteri dan parasitdisebabkan karena oksigen atau terganggunyarespirasi selama aktifitas sistem pertahananseluler dalam tubuh. Oleh sebab itu,
meningkatnya kejadian penyakit kardiovaskuler,aterosklerosis, diabetes melitus dan kankerdiduga berkaitan erat dengan radikal bebas dansenyawa oksigen reaktif (Winarsi, 2007).
Meningkatnya animo masyarakat untukmengkonsumsi makanan sehat secara tidaklangsung menyebabkan semakin bervariasinyamakanan yang diditawarkan dengan praktis dancepat saji. Tak jarang, makanan yangditawarkan dapat memicu keracunan dankerusakan pada organ tubuh dan berakhir padakematian. Semakin maju dan modernya zamanmenuntut konsumen untuk lebih bijak danpintar dalam memilih makanan yangdibutuhkan tubuh sekaligus menyehatkan tanpaproses pengolahan yang panjang sehinggadapat mengurangi nilai gizi makanan tersebut.Disisi lain, terjadinya booming produkmakanan dan minuman yang berlabelantioksidan dan dikatakan dapat melawan kerjaradikal bebas juga telah banyak dipasaran.Produk-produk yang tersedia dijual denganharga yang cukup mahalsehingga tidak semuakalangan mampu mengkonsumsinya. Padahal

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 97
komponen antioksidan yang terdapat dialammelimpah dan sangatlah banyak, misal dalamsayur-sayuran, buah-buahan bahkan jamursekalipun. Banyak orang yang belummenyadari hal ini karena belum paham apayang dimaksud dengan antioksidan, jenis,kegunaan, dan bahan apa saja yangmenggandung radikal bebas.
Pada saat sekarang ini, jamur diakui sebagaibahan makanan yang mempunyai nutrisi yangbaik serta sangat penting sebagi sumberalternatif pengobatan dan kesehatan masyarakat.Bahkan bagi masyarakat Jepang jamurdigunakan sebagai pelengkap menu harian.Tidak hanya sedap, jamur juga memilikikualitas gizi yang tinggi (Anonymous,2009).Jamur dapat dimanfaatkan sebagai bahanpangan (edible mushroom) karena memilikirasa yang enak juga memiliki nilai gizi yangtinggi dengan kandungan lemak yang rendahsehingga baik untuk dikonsumsi, apalagi bagiorang- orang yang sedang melakukan diet.Kandungan proteinnya dilaporkan mencapai10-30% (Anonymous, 2009). Selain itu jamurjuga berpotensi untuk pengobatan berbagaipenyakit karena mengandung senyawa yangbersifat antibakteri, antikanker, antitumor (Josedan Janardhanan, 2000), sumber senyawabioaktif (Lindequiest, et al., 2005) dan sumberantioksidan, mengurangi kolesterol danstimulant imun tubuh (Barros, et al., 2007).
Jamur tiram (Pleourotus) merupakan jamurdari kelas Basidiomycetes yang dikenal jugadengan sebutan oyster mushroom atau dikenaljuga dengan jamur kayu. Muchroji (2001),menambahkan Jamur tiram memiliki berbagainama, di Jepang jamur tiram dikenal dengannama Shimeji, sedangkan di Eropa danAmerika jamur tiram lebih dikenal dengannama Abalone mushroom atau Aystermushroom, dan di Indonesia populer dengannama jamur tiram dikarenakan tudungnya yangmenyerupai cangkang tiram. Diantara semuajenis jamur tiram, jamur tiram putih (P.ostreatus) merupakan jenis yang paling banyakdibudidayakan di Indonesia, tetapi akhir-akhir
ini beberapa jenis jamur tiram lainnya jugasudah mulai dibudidayakan dan bahkan sudahmemasuki pasaran
Sejauh ini, belum ditemukan adanya penelitiIndonesia yang meneliti tentang adanya totalpolifenol dan aktifitas antioksidan yangterdapat pada jamur tiram. Penelitian akan totalpolifenol dan antioksidan pada umumnyadilakukan oleh peneliti asing yang dimulaitahun 2001 sampai sekarang. Diantaranya Mauet al., 2001; Hsu et al., 2002; Mau et al., 2002;Yang et al.,2002; Lakshmi et al., 2004; Lo andCheung, 2005; Choi et al., 2006; Ferreira et al.,2007; Yoon et al., 2011, Saha et al., 2012 danGan et al., 2013. Oleh karena itu, penelitianakan total polifenol dan antioksidan perludilakukan agar masyarakat Indonesiamengetahui bahwa pada jamur tiram jugaterkandung antioksidan dan dapat digunakansekaligus sebagai pangan fungsional yangmenyehatkan.
BAHAN DAN METODE
Metode penelitian adalah metoda eksperimendalam Rancangan Acak Lengkap (RAL) Nesteddengan 2 faktor, faktor pertama: tanpapencucian serbuk gergaji dan dengan pencucianserbuk gergaji. Faktor kedua: Jenis jamurtiram;Tiram putih (Pleurotus ostreatus), tirampink (P. flabellatus), tiram kekabu (P. sajor-caju), dan tiram cokelat (P.cystidiosus) dengan4 kali ulangan. Hasil yang diperoleh akandianalisa secara statistik dengan menggunakanSPSS 15.0.Prosedur KerjaPersiapan bibit jamur. Bibit jamur tiram yangdigunakan merupakan bibit F1tiram putih,tiram pink, tiram kelabu dan tiram cokelat yangdiperoleh dari Agro Jamur PabuwaranPurwokerto, Banyumas Jawa Tengah.Persiapan media serbuk gergaji danpencampuran media dengan perbandingan15%, 1:1 dolomit dan kapur pertanian.Pelapukan (pengomposan) media ditutupdengan terpal kemudian dilapukan selama 3hari untuk menguraikan senyawa senyawa yang

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 98
terdapat pada media tanam agar diserap olehjamur. Pembuatan Baglog dan Sterilisai,media tanam dimasukkan kedalam kantongplastik polipropilen yang telah di lipat padabagian ujungnya masing- masing sebanyak 700g/baglog. Bentuk log menyerupai tabung dandipasangkan ring, ditutup dengan kertas koran,diikat dengan karet kemudian disterilisasidalam drum yang dimodifikasi pada suhu 121o
C selama 6 jam. Penanaman (Inokulasi) danInkubasi dibuka tutup baglog yang telahdisteril dari sumbatan kertas, kemudian diambildan ditaburkan bibit sebanyak 6 biji pada media.Ditutup baglog dengan sumbat dan diikatdengan karet gelang, kemudian di inkubasipada suhu 22-28oC hingga miselium tumbuhmerata pada baglog.Persiapan Sampel dan EkstraksiJamur segar yang telah dipanen dihancurkandengan lumpang dan diperas sehinggadidapatkan ekstrak murni dari jamur.Penentuan Kadar PolifenolSenyawa fenolik dalam ekstrak sampelmenggunakan Folin-Ciocalteu assay denganbeberapa modifikasi berdasarkan prosedur yangdijelaskan oleh Parrilla, et al.,(2007), Barros, etal., (2008), Gan,et al., (2013). Diambi 1 mlsampel dicampur dengan 1 ml Folin-Ciocalteu(Fenol reagen dibuat dengan perbandinganFolin-Ciocalteu reagen: air suling ; 1:9).Setelah 5 menit, ditambahkan 1 ml larutanNatrium karbonat 13 % kedalam campuran dandicukupkan dengan aquadest hinggavolumenya mencapai 10 ml. Reaksi disimpangelap selama 90 menit dan diukur nilaiabsorban pada 725 nm. Sebagai standardigunakan larutan asam galat.Penentuan Aktivitas Antioksidan denganMetode DPPHMetode DPPH yang digunakan mengacukepada Molyneux (2004).Dilarutkan 4 ml larutan DPPH 0,05 mMdengan 1 ml larutan uji. Campuran didiamkanselama waktu operating time yang telahdiperoleh. Diukur serapan larutan denganspektofotometer pada panjang gelombang 517
nm. Sebagai pembanding digunakan AsamAskorbat pada konsentrasi 10, 20, 30, 40 dan50 ppm dengan perlakuan yang sama denganlarutan uji. Sebagai blangko digunakan etanol.Penentuan Persentase PerendamanPersen Perendaman = A1-A2 x 100%
A1A1= Absorbansi kontrolA2= Absorbansi sampel
HASIL DAN PEMBAHASAN
1. Total PolifenolTotal polifenol didapatkan dari hasil analisaekstrak tubuh buah yang dilakukan denganmetoda Follin-ciocalteu. Adapun hasil yangdiperoleh dapat dilihat pada Tabel1.
Tabel 1. Rata-rata total polifenol tubuh buahberdasarkan perlakuan pencucian dantanpa pencucian media tanam padamasing-masing jamur tiram
Perlakuan Media Tanam Kode Rata-rataTotal
Polifenol(mg GAC/g)
Notasi
Tanpa Pencucian, Tiram KelabuPencucian, Tiram KelabuTanpa Pencucian, Tiram PutihTanpa Pencucian, Tiram CokelatTanpa Pencucian, Tiram PinkPencucian, Tiram PinkPencucian, Tiram CokelatPencucian, Tiram Putih
A1B3A2B3A1B1A1B4A1B2A2B2A2B4A2B1
1901,251700,001205,461200,00914,08826,25803,75443,75
aabbbcbcbcc
Keterangan : angka-angka pada tabel yang diikuti hurufkecil yang sama berbeda tidak nyata pada α= 0,05
Berdasarkan hasil uji statistik Tabel 1,terlihat bahwa perlakuan yang mempunyai totalpolifenol tertinggi yaitu perlakuan media tanpapencucian Jamur tiramkelabu (1901,25 mgGAC/g), dan perlakuan yang mempunyai totalpolifenol terendah yaitu perlakuan denganpencucian media Jamur tiram putih (443,75 mgGAC/g). Sehingga dapat diketahui bahwarentang total polifenol pada perlakuan mediatanam berkisar antara 443,75 mg GAC/g-1901,25 mg GAC/g.
Terjadinya perbedaan kadar polifenol antarperlakuan pencucian dan tanpa pencucianmedia tanam diduga erat hubungannya dengan

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 99
berkurangnya kadar tanin seiiring dilakukannyapencucian media tanam. Serbuk gergaji denganperlakuan pencucian akan kehilangan tanindalam jumlah yang banyak. Tanin yang tersisapada media tanam dalam jumlah yang sedikitakan terserap sedikit, tapi sebaliknya tanin yangtersedia dalam jumlah banyak juga akanterserap dalam jumlah yang lebih. Keadaan iniberhubungna erat dengan cara hidup jamuryang saprofit yang menyerap semua komponendalam pemenuhan nutrisi. Hal ini sesuai denganDjarijah (2001), yang menyatakan kehidupanjamur mengambil makanan atau nutrisi yangtelah dibuat oleh organisme lain yang telahmati (saprofit), karena jamur tidak memilikiklorofil yang mampu berfotosintesis untukmenghasilkan nutrisi.
Keberadaan tanin pada media tanammempengaruhi keadaan tubuh buah yangdibentuk dikarenakan tanin merupakan bagiandari polifenol yang ikut terserap pada saatpemenuhan nutrisi tumbuh. Hal ini dikuatkanoleh Parilla (2007), yang menyatakan jikakadar tanin semakin tinggi pada substrat, makatotal fenolik yang terdapat pada tubuh buahakan tinggi dikarenakan kadar air yang rendahdan dehidrasi. Keadaan ini berhubungna eratdengan cara hidup jamur yang saprofit yangmenyerap semua komponen dalam pemenuhannutrisi. Hal ini sesuai dengan Djarijah (2001),yang menyatakan kehidupan jamur mengambilmakanan atau nutrisi yang telah dibuat olehorganisme lain yang telah mati (saprofit),karena jamur tidak memiliki klorofil yangmampu berfotosintesis untuk menghasilkannutrisi.
Disamping pengamatan total polifenolberdasarkan perlakuan media tanam pada tubuhjamur, juga diamati perbedaan total polifenolpada masing-masing jenis jamur tiram. DariGambar 1 diatas, terlihat bahwa pada masing-masing jamur tiram mempunyai total polifenolyang berbeda. Jamur tiram kelabu merupakanjenis yang mempunyai total polifenol tertinggi(1800,63 mg GAC/g ) dan berbeda nyatadengan jenis lainnya kemudian diikuti oleh
Jamur tiram cokelat (1001,88mg GAC/g),Jamur tiram pink (870,17 mg GAC/g).Sedangkan Jamur tiram putih yang seringdibudidayakan mempunyai total polifenol yangterendah (824,62 mg GAC/g) diantara jenislainnya. Rentang total polifenol pada beberapajenis jamur tiram adalah 824,62 mg GAC/g-1800,63 mg GAC/g.
Gambar 1. Total polifenol pada beberapa jenis jamurtiram ( mg GAC/g)
Tingginya kandungan polifenol pada jenistiram kelabu menunjukkan bahwa jamur inimempunyai toleransi yang tinggi dalammenyerap nutrisisekalipun berada dalam kadartanin yang tinggi. Tingginya perbedaankandungan polifenol pada jenis jamur tiramlainnya diduga karena kandungan polifenolpada masing-masing jamur tiram sebagianbesar teroksidasi sehingga dapat mengurangkankadar polifenol didalam tubuh jamur. Sesuaidengan Shahidi dan Naczk (1995), yangmenyatakan bahwa kandungan utama polifenoladalah flavanol (katekin, galokatekin,epikatekin, epikatekin galat, epigalokatekin,dan epigalokatekin galat), flavonol (quercetin,kaemferol, dan glikosidanya), flavone (vixetindan iso vixetin), asam fenolik (asam galat danasam klorogenat) yang merupakan senyawayang mudah teroksidasi.
Total polifenol yang terserap pada masing-masing jamur tiram diduga erat kaitannyadengan waktu yang dibutuhkan selamaproduksi. Jamur tiram kelabu mempunyaiwaktu terlama untuk produksi dibandingkanjenis lainnya. Selain itu secara morfologikeadaan jamur tiram kelabu mempunyai
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 99
berkurangnya kadar tanin seiiring dilakukannyapencucian media tanam. Serbuk gergaji denganperlakuan pencucian akan kehilangan tanindalam jumlah yang banyak. Tanin yang tersisapada media tanam dalam jumlah yang sedikitakan terserap sedikit, tapi sebaliknya tanin yangtersedia dalam jumlah banyak juga akanterserap dalam jumlah yang lebih. Keadaan iniberhubungna erat dengan cara hidup jamuryang saprofit yang menyerap semua komponendalam pemenuhan nutrisi. Hal ini sesuai denganDjarijah (2001), yang menyatakan kehidupanjamur mengambil makanan atau nutrisi yangtelah dibuat oleh organisme lain yang telahmati (saprofit), karena jamur tidak memilikiklorofil yang mampu berfotosintesis untukmenghasilkan nutrisi.
Keberadaan tanin pada media tanammempengaruhi keadaan tubuh buah yangdibentuk dikarenakan tanin merupakan bagiandari polifenol yang ikut terserap pada saatpemenuhan nutrisi tumbuh. Hal ini dikuatkanoleh Parilla (2007), yang menyatakan jikakadar tanin semakin tinggi pada substrat, makatotal fenolik yang terdapat pada tubuh buahakan tinggi dikarenakan kadar air yang rendahdan dehidrasi. Keadaan ini berhubungna eratdengan cara hidup jamur yang saprofit yangmenyerap semua komponen dalam pemenuhannutrisi. Hal ini sesuai dengan Djarijah (2001),yang menyatakan kehidupan jamur mengambilmakanan atau nutrisi yang telah dibuat olehorganisme lain yang telah mati (saprofit),karena jamur tidak memiliki klorofil yangmampu berfotosintesis untuk menghasilkannutrisi.
Disamping pengamatan total polifenolberdasarkan perlakuan media tanam pada tubuhjamur, juga diamati perbedaan total polifenolpada masing-masing jenis jamur tiram. DariGambar 1 diatas, terlihat bahwa pada masing-masing jamur tiram mempunyai total polifenolyang berbeda. Jamur tiram kelabu merupakanjenis yang mempunyai total polifenol tertinggi(1800,63 mg GAC/g ) dan berbeda nyatadengan jenis lainnya kemudian diikuti oleh
Jamur tiram cokelat (1001,88mg GAC/g),Jamur tiram pink (870,17 mg GAC/g).Sedangkan Jamur tiram putih yang seringdibudidayakan mempunyai total polifenol yangterendah (824,62 mg GAC/g) diantara jenislainnya. Rentang total polifenol pada beberapajenis jamur tiram adalah 824,62 mg GAC/g-1800,63 mg GAC/g.
Gambar 1. Total polifenol pada beberapa jenis jamurtiram ( mg GAC/g)
Tingginya kandungan polifenol pada jenistiram kelabu menunjukkan bahwa jamur inimempunyai toleransi yang tinggi dalammenyerap nutrisisekalipun berada dalam kadartanin yang tinggi. Tingginya perbedaankandungan polifenol pada jenis jamur tiramlainnya diduga karena kandungan polifenolpada masing-masing jamur tiram sebagianbesar teroksidasi sehingga dapat mengurangkankadar polifenol didalam tubuh jamur. Sesuaidengan Shahidi dan Naczk (1995), yangmenyatakan bahwa kandungan utama polifenoladalah flavanol (katekin, galokatekin,epikatekin, epikatekin galat, epigalokatekin,dan epigalokatekin galat), flavonol (quercetin,kaemferol, dan glikosidanya), flavone (vixetindan iso vixetin), asam fenolik (asam galat danasam klorogenat) yang merupakan senyawayang mudah teroksidasi.
Total polifenol yang terserap pada masing-masing jamur tiram diduga erat kaitannyadengan waktu yang dibutuhkan selamaproduksi. Jamur tiram kelabu mempunyaiwaktu terlama untuk produksi dibandingkanjenis lainnya. Selain itu secara morfologikeadaan jamur tiram kelabu mempunyai
jenis jamur tiram
824.62b
870.17b
1800.63a
Total Polifenol pada beberapa jenisjamur tiram (mg GAC/g)tiram putihtiram kelabu
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 99
berkurangnya kadar tanin seiiring dilakukannyapencucian media tanam. Serbuk gergaji denganperlakuan pencucian akan kehilangan tanindalam jumlah yang banyak. Tanin yang tersisapada media tanam dalam jumlah yang sedikitakan terserap sedikit, tapi sebaliknya tanin yangtersedia dalam jumlah banyak juga akanterserap dalam jumlah yang lebih. Keadaan iniberhubungna erat dengan cara hidup jamuryang saprofit yang menyerap semua komponendalam pemenuhan nutrisi. Hal ini sesuai denganDjarijah (2001), yang menyatakan kehidupanjamur mengambil makanan atau nutrisi yangtelah dibuat oleh organisme lain yang telahmati (saprofit), karena jamur tidak memilikiklorofil yang mampu berfotosintesis untukmenghasilkan nutrisi.
Keberadaan tanin pada media tanammempengaruhi keadaan tubuh buah yangdibentuk dikarenakan tanin merupakan bagiandari polifenol yang ikut terserap pada saatpemenuhan nutrisi tumbuh. Hal ini dikuatkanoleh Parilla (2007), yang menyatakan jikakadar tanin semakin tinggi pada substrat, makatotal fenolik yang terdapat pada tubuh buahakan tinggi dikarenakan kadar air yang rendahdan dehidrasi. Keadaan ini berhubungna eratdengan cara hidup jamur yang saprofit yangmenyerap semua komponen dalam pemenuhannutrisi. Hal ini sesuai dengan Djarijah (2001),yang menyatakan kehidupan jamur mengambilmakanan atau nutrisi yang telah dibuat olehorganisme lain yang telah mati (saprofit),karena jamur tidak memiliki klorofil yangmampu berfotosintesis untuk menghasilkannutrisi.
Disamping pengamatan total polifenolberdasarkan perlakuan media tanam pada tubuhjamur, juga diamati perbedaan total polifenolpada masing-masing jenis jamur tiram. DariGambar 1 diatas, terlihat bahwa pada masing-masing jamur tiram mempunyai total polifenolyang berbeda. Jamur tiram kelabu merupakanjenis yang mempunyai total polifenol tertinggi(1800,63 mg GAC/g ) dan berbeda nyatadengan jenis lainnya kemudian diikuti oleh
Jamur tiram cokelat (1001,88mg GAC/g),Jamur tiram pink (870,17 mg GAC/g).Sedangkan Jamur tiram putih yang seringdibudidayakan mempunyai total polifenol yangterendah (824,62 mg GAC/g) diantara jenislainnya. Rentang total polifenol pada beberapajenis jamur tiram adalah 824,62 mg GAC/g-1800,63 mg GAC/g.
Gambar 1. Total polifenol pada beberapa jenis jamurtiram ( mg GAC/g)
Tingginya kandungan polifenol pada jenistiram kelabu menunjukkan bahwa jamur inimempunyai toleransi yang tinggi dalammenyerap nutrisisekalipun berada dalam kadartanin yang tinggi. Tingginya perbedaankandungan polifenol pada jenis jamur tiramlainnya diduga karena kandungan polifenolpada masing-masing jamur tiram sebagianbesar teroksidasi sehingga dapat mengurangkankadar polifenol didalam tubuh jamur. Sesuaidengan Shahidi dan Naczk (1995), yangmenyatakan bahwa kandungan utama polifenoladalah flavanol (katekin, galokatekin,epikatekin, epikatekin galat, epigalokatekin,dan epigalokatekin galat), flavonol (quercetin,kaemferol, dan glikosidanya), flavone (vixetindan iso vixetin), asam fenolik (asam galat danasam klorogenat) yang merupakan senyawayang mudah teroksidasi.
Total polifenol yang terserap pada masing-masing jamur tiram diduga erat kaitannyadengan waktu yang dibutuhkan selamaproduksi. Jamur tiram kelabu mempunyaiwaktu terlama untuk produksi dibandingkanjenis lainnya. Selain itu secara morfologikeadaan jamur tiram kelabu mempunyai
jenis jamur tiram
1800.63a
1001.88b
Total Polifenol pada beberapa jenisjamur tiram (mg GAC/g)
tiram pinktiram cokelat

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 100
tangkai yang lebih panjang diantara jenislainnya. Keadaan seperti ini menyebabkantransportasi air dan nutrisi berlangsung dengancepat dalam pemenuhan nutrisi.
Adapun jenis jamur tiram yang digunakandalam pengukuran total polifenol dan aktifitasantioksidan pada tubuh buah seperti terlihatpada Gambar 2.
a b c dGambar 2. Jenis jamur tiram yang diukur total
polifenol dan aktifitas antioksidantubuh buah a).Tiram putih ( b).tirampink (c). Tiram kelabu (d). Tiramcokelat
2. Aktifitas Antioksidan Tubuh BuahAntioksidan merupakan senyawa yang dapatmenunda atau mencegah terjadinya reaksioksidasi radikal bebas dalam oksidasi lipid.Aktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanambeberapa jenis jamur tiram dilakukan denganmetode DPPH. Hasil analisis yang diperolehdapat dilihat pada Tabel 2.
Tabel 2. Rata-rata aktivitas antioksidan tubuh buahberdasarkan perlakuan pencucian dan tanpapencucian media tanam pada masing-masingjamur tiram
Perlakuan Media TanamKode
Rata-rataAktifitas
AntioksidanTubuhBuah
(mg/ml)
Notasi
Pencucian, Tiram PinkTanpa Pencucian, Tiram PinkPencucian, Tiram PutihPencucian, Tiram CokelatPencucian, Tiram KelabuTanpa Pencucian, Tiram KelabuTanpa Pencucian, Tiram PutihTanpa Pencucian, Tiram Cokelat
A2B2A1B2A2B1A2B4A2B3A1B3A1B1A1B4
11.607,6411.459,4311.418,7211.358,0011.220.8611.058,0010.690,8610.585,14
abcdefgh
Ket.: angka-angka pada tabel yang diikuti huruf kecilyang sama berbeda tidak nyata pada α= 0,05
Dari Tabel 2 diatas, terlihat bahwa pencuciandan tanpa pencucian media tanam serbuk gergajipada beberapa jenis jamur tiram menunjukkan
terjadi perbedaan nyata setiap perlakuan.Perlakuan yang mempunyai aktifitasantioksidan tertinggi yaitu dengan pencucianmedia tanam Jamur tiram pink (11.607,64mg/ml) sedangkan perlakuan yang mempunyaiaktifitas antioksidan terendah yaitu perlakuantanpa pencucian media Jamur tiram cokelat(10.585,14 mg/ml). Dapat diketahui bahwaaktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanamberkisar antara 10.585,14 mg/ml - 11.607,64mg/ml. Hal ini menunjukkan bahwa jenispencucian media tanam memberikan pengaruhyang signifikan pada aktifitas antioksidan tubuhbuah jamur tiram.
Aktivitas antioksidan pada jamur yangsejenis menunjukkan perbedaan yang nyata.Perlakuan tanpa pencucian media jenis tiramputih mempunyai aktivitas antioksidan10.690,86 mg/ml yang lebih rendah jikadibandingkan dengan perlakuan denganpencucian media (11.418,72 mg/ml). Begitupunhalnya dengan perlakuan tanpa pencucian mediajenis tiram pink yang juga mempunyai aktifitasantioksidan (11.459,43 mg/ml) yang lebihrendah jika dibandingkan dengan pencucianmedia (11.607,64 mg/ml). Hal itu juga terjadipada perlakuan tanpa pencucian media jenistiram kelabu yang mempunyai aktifitasantioksidan (11.058,00 mg/ml) juga lebihrendah jika dibandingkan dengan pencucianmedia (11.220.86 mg/ml). Pada perlakuan tanpapencucian media Jamur tiram cokelat (10.585,14mg/ml), aktifitas antioksidan juga lebih rendahdari pada dengan pencucian media (11.358,00mg/ml). Dari hal ini maka dapat diketahuibahwa aktifitas antioksidan tubuh buah padabeberapa jenis jamur tiram mempunyai aktifitasyang tinggi pada perlakuan dengan pencucianmedia tanam.
Tingginya aktifitas antioksidan tubuh buahpada media tanam yang dilakukan pencuciandiduga karena pada media tanam tidak terdapatbanyak tanin yang dapat menghambat prosespembentukan tubuh buah. Dari hasil yangdiperoleh menunjukkan bahwa aktifitas
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 100
tangkai yang lebih panjang diantara jenislainnya. Keadaan seperti ini menyebabkantransportasi air dan nutrisi berlangsung dengancepat dalam pemenuhan nutrisi.
Adapun jenis jamur tiram yang digunakandalam pengukuran total polifenol dan aktifitasantioksidan pada tubuh buah seperti terlihatpada Gambar 2.
a b c dGambar 2. Jenis jamur tiram yang diukur total
polifenol dan aktifitas antioksidantubuh buah a).Tiram putih ( b).tirampink (c). Tiram kelabu (d). Tiramcokelat
2. Aktifitas Antioksidan Tubuh BuahAntioksidan merupakan senyawa yang dapatmenunda atau mencegah terjadinya reaksioksidasi radikal bebas dalam oksidasi lipid.Aktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanambeberapa jenis jamur tiram dilakukan denganmetode DPPH. Hasil analisis yang diperolehdapat dilihat pada Tabel 2.
Tabel 2. Rata-rata aktivitas antioksidan tubuh buahberdasarkan perlakuan pencucian dan tanpapencucian media tanam pada masing-masingjamur tiram
Perlakuan Media TanamKode
Rata-rataAktifitas
AntioksidanTubuhBuah
(mg/ml)
Notasi
Pencucian, Tiram PinkTanpa Pencucian, Tiram PinkPencucian, Tiram PutihPencucian, Tiram CokelatPencucian, Tiram KelabuTanpa Pencucian, Tiram KelabuTanpa Pencucian, Tiram PutihTanpa Pencucian, Tiram Cokelat
A2B2A1B2A2B1A2B4A2B3A1B3A1B1A1B4
11.607,6411.459,4311.418,7211.358,0011.220.8611.058,0010.690,8610.585,14
abcdefgh
Ket.: angka-angka pada tabel yang diikuti huruf kecilyang sama berbeda tidak nyata pada α= 0,05
Dari Tabel 2 diatas, terlihat bahwa pencuciandan tanpa pencucian media tanam serbuk gergajipada beberapa jenis jamur tiram menunjukkan
terjadi perbedaan nyata setiap perlakuan.Perlakuan yang mempunyai aktifitasantioksidan tertinggi yaitu dengan pencucianmedia tanam Jamur tiram pink (11.607,64mg/ml) sedangkan perlakuan yang mempunyaiaktifitas antioksidan terendah yaitu perlakuantanpa pencucian media Jamur tiram cokelat(10.585,14 mg/ml). Dapat diketahui bahwaaktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanamberkisar antara 10.585,14 mg/ml - 11.607,64mg/ml. Hal ini menunjukkan bahwa jenispencucian media tanam memberikan pengaruhyang signifikan pada aktifitas antioksidan tubuhbuah jamur tiram.
Aktivitas antioksidan pada jamur yangsejenis menunjukkan perbedaan yang nyata.Perlakuan tanpa pencucian media jenis tiramputih mempunyai aktivitas antioksidan10.690,86 mg/ml yang lebih rendah jikadibandingkan dengan perlakuan denganpencucian media (11.418,72 mg/ml). Begitupunhalnya dengan perlakuan tanpa pencucian mediajenis tiram pink yang juga mempunyai aktifitasantioksidan (11.459,43 mg/ml) yang lebihrendah jika dibandingkan dengan pencucianmedia (11.607,64 mg/ml). Hal itu juga terjadipada perlakuan tanpa pencucian media jenistiram kelabu yang mempunyai aktifitasantioksidan (11.058,00 mg/ml) juga lebihrendah jika dibandingkan dengan pencucianmedia (11.220.86 mg/ml). Pada perlakuan tanpapencucian media Jamur tiram cokelat (10.585,14mg/ml), aktifitas antioksidan juga lebih rendahdari pada dengan pencucian media (11.358,00mg/ml). Dari hal ini maka dapat diketahuibahwa aktifitas antioksidan tubuh buah padabeberapa jenis jamur tiram mempunyai aktifitasyang tinggi pada perlakuan dengan pencucianmedia tanam.
Tingginya aktifitas antioksidan tubuh buahpada media tanam yang dilakukan pencuciandiduga karena pada media tanam tidak terdapatbanyak tanin yang dapat menghambat prosespembentukan tubuh buah. Dari hasil yangdiperoleh menunjukkan bahwa aktifitas
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 100
tangkai yang lebih panjang diantara jenislainnya. Keadaan seperti ini menyebabkantransportasi air dan nutrisi berlangsung dengancepat dalam pemenuhan nutrisi.
Adapun jenis jamur tiram yang digunakandalam pengukuran total polifenol dan aktifitasantioksidan pada tubuh buah seperti terlihatpada Gambar 2.
a b c dGambar 2. Jenis jamur tiram yang diukur total
polifenol dan aktifitas antioksidantubuh buah a).Tiram putih ( b).tirampink (c). Tiram kelabu (d). Tiramcokelat
2. Aktifitas Antioksidan Tubuh BuahAntioksidan merupakan senyawa yang dapatmenunda atau mencegah terjadinya reaksioksidasi radikal bebas dalam oksidasi lipid.Aktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanambeberapa jenis jamur tiram dilakukan denganmetode DPPH. Hasil analisis yang diperolehdapat dilihat pada Tabel 2.
Tabel 2. Rata-rata aktivitas antioksidan tubuh buahberdasarkan perlakuan pencucian dan tanpapencucian media tanam pada masing-masingjamur tiram
Perlakuan Media TanamKode
Rata-rataAktifitas
AntioksidanTubuhBuah
(mg/ml)
Notasi
Pencucian, Tiram PinkTanpa Pencucian, Tiram PinkPencucian, Tiram PutihPencucian, Tiram CokelatPencucian, Tiram KelabuTanpa Pencucian, Tiram KelabuTanpa Pencucian, Tiram PutihTanpa Pencucian, Tiram Cokelat
A2B2A1B2A2B1A2B4A2B3A1B3A1B1A1B4
11.607,6411.459,4311.418,7211.358,0011.220.8611.058,0010.690,8610.585,14
abcdefgh
Ket.: angka-angka pada tabel yang diikuti huruf kecilyang sama berbeda tidak nyata pada α= 0,05
Dari Tabel 2 diatas, terlihat bahwa pencuciandan tanpa pencucian media tanam serbuk gergajipada beberapa jenis jamur tiram menunjukkan
terjadi perbedaan nyata setiap perlakuan.Perlakuan yang mempunyai aktifitasantioksidan tertinggi yaitu dengan pencucianmedia tanam Jamur tiram pink (11.607,64mg/ml) sedangkan perlakuan yang mempunyaiaktifitas antioksidan terendah yaitu perlakuantanpa pencucian media Jamur tiram cokelat(10.585,14 mg/ml). Dapat diketahui bahwaaktifitas antioksidan tubuh buah pada perlakuanpencucian dan tanpa pencucian media tanamberkisar antara 10.585,14 mg/ml - 11.607,64mg/ml. Hal ini menunjukkan bahwa jenispencucian media tanam memberikan pengaruhyang signifikan pada aktifitas antioksidan tubuhbuah jamur tiram.
Aktivitas antioksidan pada jamur yangsejenis menunjukkan perbedaan yang nyata.Perlakuan tanpa pencucian media jenis tiramputih mempunyai aktivitas antioksidan10.690,86 mg/ml yang lebih rendah jikadibandingkan dengan perlakuan denganpencucian media (11.418,72 mg/ml). Begitupunhalnya dengan perlakuan tanpa pencucian mediajenis tiram pink yang juga mempunyai aktifitasantioksidan (11.459,43 mg/ml) yang lebihrendah jika dibandingkan dengan pencucianmedia (11.607,64 mg/ml). Hal itu juga terjadipada perlakuan tanpa pencucian media jenistiram kelabu yang mempunyai aktifitasantioksidan (11.058,00 mg/ml) juga lebihrendah jika dibandingkan dengan pencucianmedia (11.220.86 mg/ml). Pada perlakuan tanpapencucian media Jamur tiram cokelat (10.585,14mg/ml), aktifitas antioksidan juga lebih rendahdari pada dengan pencucian media (11.358,00mg/ml). Dari hal ini maka dapat diketahuibahwa aktifitas antioksidan tubuh buah padabeberapa jenis jamur tiram mempunyai aktifitasyang tinggi pada perlakuan dengan pencucianmedia tanam.
Tingginya aktifitas antioksidan tubuh buahpada media tanam yang dilakukan pencuciandiduga karena pada media tanam tidak terdapatbanyak tanin yang dapat menghambat prosespembentukan tubuh buah. Dari hasil yangdiperoleh menunjukkan bahwa aktifitas

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 101
antioksidan pada perlakuan pencucian mediatanam dengan tanpa pencucian media tanamberbanding terbalik dengan kadar polifenol padatubuh buah. Jika dalam substrat media tanammempunyai banyak kadar tanin, maka akanmeningkatkan kadar polifenol dan menurunkanaktifitas antioksidan. Tapi sebaliknya jika kadartanin berkurang, maka kadar polifenol akanrendah sementara aktifitas antioksidan akantinggi. Hal ini sesuai dengan Furham danAviram (2002), di bawah kondisi tertentu,seperti konsentrasi antioksidan fenolik yangtinggi, pH yang tinggi, atau keberadaan ion besi,antioksidan fenolik dapat menginisiasi prosesauto-oksidasi dan lebih bersifat seperti pro-oksidan dibandingkan antioksidan.
Berbanding terbaliknya total polifenol danaktifitas antioksidan tubuh buah beberapa jenisjamur tiram pada perlakuan pencucian dan tanpapencucian media tanam diduga erat kaitannyadengan kemampuan terlarutnya masing-masingkomponen selama ekstraksi sampel. Dalam halantioksidan terdapat dua komponen yangmenentukan nilai total antioksidan secarakeseluruhan yaitu antioksidan yang dapat larutair dan larut lemak. Dalam hal ini aktifitasantioksidan yang diukur hanyalah larut dalamair saja. Antioksidan yang larut dalam airhanyalah sebagian kecil dari total antioksidanyang terdapat dalam tubuh jamur. Berbedahalnya dengan total polifenol yang memanglarut dalam air dimana nilai total akan diperolehsecara sempurna.
Selain aktifitas antioksidan tubuh buahdengan perlakuan pencucian media tanam, jugadilakukan pengamatan aktifitas antioksidan padamasing-masing jamur tiram. Hasil analisisaktifitas antioksidan pada beberapa jenis jamurtiram dapat dilihat pada Gambar 3.
Berdasarkan Gambar 3 diatas, maka dapatdiketahui bahwa pada jamur tiram mempunyaiaktifitas antioksidan yang berbeda-beda. Tirampink merupakan jenis jamur yang mempunyaiaktifitas antioksidan tertinggi dengan nilai11.533,54 mg/ml dan berbeda nyata denganjenis lainnya, diikuti oleh Jamur tiram kelabu
(11.139,43 mg/ml), dan tiram putih (11.054,79mg/ml). Sedangkan jenis jamur yangmempunyai aktifitas antioksidan terendahadalah jenis tiram cokelat dengan nilai10,971,57 mg/ml. Dapat diketahui rentangaktifitas antioksidan pada beberapa jenis jamurtiram berkisar antara 10,971,57 mg/ml -11.533,54 mg/ml.
Gambar 3. Aktivitas antioksidan tubuhbuah masing-masing jamurtiram (mg/ml)
Dari nilai aktifitas antioksidan yangdiperoleh, maka dapat diketahui bahwa padatubuh buah jamur tiram yang tidak mempunyaiklorofil mempunyai aktifitas antioksidan yangtinggi. Hal ini sesuai dengan Gregori, Mirjanand Pohleven (2007), yang menyatakan jamurmengandung antioksidan atau dapatmeningkatkan aktivitas enzim antioksidansehingga digunakan untuk mengurangikerusakan oksidatif pada manusia. Saha,Sandeep and Aparajita (2012), menambahkansecara umum jamur yang dikonsumsi di negara-negara Asia ditemukan memiliki aktivitasantioksidan dan korelasi dengan total senyawafenolik.Pertimbangan efek dari kandungan fenolik totalterhadap aktivitas antioksidan dan folifenol yangakan diuji dilakukan pada ekstrak jamur yangdiambil pada bagian tudung buah. Adapun jenisekstrak tubuh buah jamur tiram yang dihasilkandapat diperhatikan pada Gambar 4.
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 101
antioksidan pada perlakuan pencucian mediatanam dengan tanpa pencucian media tanamberbanding terbalik dengan kadar polifenol padatubuh buah. Jika dalam substrat media tanammempunyai banyak kadar tanin, maka akanmeningkatkan kadar polifenol dan menurunkanaktifitas antioksidan. Tapi sebaliknya jika kadartanin berkurang, maka kadar polifenol akanrendah sementara aktifitas antioksidan akantinggi. Hal ini sesuai dengan Furham danAviram (2002), di bawah kondisi tertentu,seperti konsentrasi antioksidan fenolik yangtinggi, pH yang tinggi, atau keberadaan ion besi,antioksidan fenolik dapat menginisiasi prosesauto-oksidasi dan lebih bersifat seperti pro-oksidan dibandingkan antioksidan.
Berbanding terbaliknya total polifenol danaktifitas antioksidan tubuh buah beberapa jenisjamur tiram pada perlakuan pencucian dan tanpapencucian media tanam diduga erat kaitannyadengan kemampuan terlarutnya masing-masingkomponen selama ekstraksi sampel. Dalam halantioksidan terdapat dua komponen yangmenentukan nilai total antioksidan secarakeseluruhan yaitu antioksidan yang dapat larutair dan larut lemak. Dalam hal ini aktifitasantioksidan yang diukur hanyalah larut dalamair saja. Antioksidan yang larut dalam airhanyalah sebagian kecil dari total antioksidanyang terdapat dalam tubuh jamur. Berbedahalnya dengan total polifenol yang memanglarut dalam air dimana nilai total akan diperolehsecara sempurna.
Selain aktifitas antioksidan tubuh buahdengan perlakuan pencucian media tanam, jugadilakukan pengamatan aktifitas antioksidan padamasing-masing jamur tiram. Hasil analisisaktifitas antioksidan pada beberapa jenis jamurtiram dapat dilihat pada Gambar 3.
Berdasarkan Gambar 3 diatas, maka dapatdiketahui bahwa pada jamur tiram mempunyaiaktifitas antioksidan yang berbeda-beda. Tirampink merupakan jenis jamur yang mempunyaiaktifitas antioksidan tertinggi dengan nilai11.533,54 mg/ml dan berbeda nyata denganjenis lainnya, diikuti oleh Jamur tiram kelabu
(11.139,43 mg/ml), dan tiram putih (11.054,79mg/ml). Sedangkan jenis jamur yangmempunyai aktifitas antioksidan terendahadalah jenis tiram cokelat dengan nilai10,971,57 mg/ml. Dapat diketahui rentangaktifitas antioksidan pada beberapa jenis jamurtiram berkisar antara 10,971,57 mg/ml -11.533,54 mg/ml.
Gambar 3. Aktivitas antioksidan tubuhbuah masing-masing jamurtiram (mg/ml)
Dari nilai aktifitas antioksidan yangdiperoleh, maka dapat diketahui bahwa padatubuh buah jamur tiram yang tidak mempunyaiklorofil mempunyai aktifitas antioksidan yangtinggi. Hal ini sesuai dengan Gregori, Mirjanand Pohleven (2007), yang menyatakan jamurmengandung antioksidan atau dapatmeningkatkan aktivitas enzim antioksidansehingga digunakan untuk mengurangikerusakan oksidatif pada manusia. Saha,Sandeep and Aparajita (2012), menambahkansecara umum jamur yang dikonsumsi di negara-negara Asia ditemukan memiliki aktivitasantioksidan dan korelasi dengan total senyawafenolik.Pertimbangan efek dari kandungan fenolik totalterhadap aktivitas antioksidan dan folifenol yangakan diuji dilakukan pada ekstrak jamur yangdiambil pada bagian tudung buah. Adapun jenisekstrak tubuh buah jamur tiram yang dihasilkandapat diperhatikan pada Gambar 4.
jenis jamur tiram
11054.79b
11533.54a 11139.43
b
Aktifitas Antioksidan Tubuh BuahBeberapa Jenis jamur tiram (mg/ml)tiram putihtiram kelabu
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 101
antioksidan pada perlakuan pencucian mediatanam dengan tanpa pencucian media tanamberbanding terbalik dengan kadar polifenol padatubuh buah. Jika dalam substrat media tanammempunyai banyak kadar tanin, maka akanmeningkatkan kadar polifenol dan menurunkanaktifitas antioksidan. Tapi sebaliknya jika kadartanin berkurang, maka kadar polifenol akanrendah sementara aktifitas antioksidan akantinggi. Hal ini sesuai dengan Furham danAviram (2002), di bawah kondisi tertentu,seperti konsentrasi antioksidan fenolik yangtinggi, pH yang tinggi, atau keberadaan ion besi,antioksidan fenolik dapat menginisiasi prosesauto-oksidasi dan lebih bersifat seperti pro-oksidan dibandingkan antioksidan.
Berbanding terbaliknya total polifenol danaktifitas antioksidan tubuh buah beberapa jenisjamur tiram pada perlakuan pencucian dan tanpapencucian media tanam diduga erat kaitannyadengan kemampuan terlarutnya masing-masingkomponen selama ekstraksi sampel. Dalam halantioksidan terdapat dua komponen yangmenentukan nilai total antioksidan secarakeseluruhan yaitu antioksidan yang dapat larutair dan larut lemak. Dalam hal ini aktifitasantioksidan yang diukur hanyalah larut dalamair saja. Antioksidan yang larut dalam airhanyalah sebagian kecil dari total antioksidanyang terdapat dalam tubuh jamur. Berbedahalnya dengan total polifenol yang memanglarut dalam air dimana nilai total akan diperolehsecara sempurna.
Selain aktifitas antioksidan tubuh buahdengan perlakuan pencucian media tanam, jugadilakukan pengamatan aktifitas antioksidan padamasing-masing jamur tiram. Hasil analisisaktifitas antioksidan pada beberapa jenis jamurtiram dapat dilihat pada Gambar 3.
Berdasarkan Gambar 3 diatas, maka dapatdiketahui bahwa pada jamur tiram mempunyaiaktifitas antioksidan yang berbeda-beda. Tirampink merupakan jenis jamur yang mempunyaiaktifitas antioksidan tertinggi dengan nilai11.533,54 mg/ml dan berbeda nyata denganjenis lainnya, diikuti oleh Jamur tiram kelabu
(11.139,43 mg/ml), dan tiram putih (11.054,79mg/ml). Sedangkan jenis jamur yangmempunyai aktifitas antioksidan terendahadalah jenis tiram cokelat dengan nilai10,971,57 mg/ml. Dapat diketahui rentangaktifitas antioksidan pada beberapa jenis jamurtiram berkisar antara 10,971,57 mg/ml -11.533,54 mg/ml.
Gambar 3. Aktivitas antioksidan tubuhbuah masing-masing jamurtiram (mg/ml)
Dari nilai aktifitas antioksidan yangdiperoleh, maka dapat diketahui bahwa padatubuh buah jamur tiram yang tidak mempunyaiklorofil mempunyai aktifitas antioksidan yangtinggi. Hal ini sesuai dengan Gregori, Mirjanand Pohleven (2007), yang menyatakan jamurmengandung antioksidan atau dapatmeningkatkan aktivitas enzim antioksidansehingga digunakan untuk mengurangikerusakan oksidatif pada manusia. Saha,Sandeep and Aparajita (2012), menambahkansecara umum jamur yang dikonsumsi di negara-negara Asia ditemukan memiliki aktivitasantioksidan dan korelasi dengan total senyawafenolik.Pertimbangan efek dari kandungan fenolik totalterhadap aktivitas antioksidan dan folifenol yangakan diuji dilakukan pada ekstrak jamur yangdiambil pada bagian tudung buah. Adapun jenisekstrak tubuh buah jamur tiram yang dihasilkandapat diperhatikan pada Gambar 4.
jenis jamur tiram
11139.43b 10971.57
b
Aktifitas Antioksidan Tubuh BuahBeberapa Jenis jamur tiram (mg/ml)
tiram pinktiram cokelat

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 102
Gambar 4. Perbandingan tingkat kekeruhanekstrak yang dihasilkan dariperlakuan pencucian media padabeberapa jenis jamur tiram
Dari Gambar 4 diatas, terlihat bahwa perbedaanyang signifikan pada masing-masing ekstrakyang dihasilkan. Kepekatan ekstrak padamasing-masing jamur pada perlakuan tanpapencucian lebih tinggi jika dibandingkan denganekstrak dengan pencucian. Terjadinya perbedaantingkat kekeruhan diduga berkaitan dengankadar tanin yang diserap oleh jamur sehinggaturut mempengaruhi aktifitas antioksidan dantotal polifenol yang terdapat dalam tubuh buah.Vamanu (2012), menjelaskan bahwa dalamekstrak jamur kapasitas antioksidan ditentukanoleh jumlah senyawa fenolik yangdikandungnya. Kuantitas ekstrak juga mem-pengaruhi kapasitas penghambatan radikal bebas.Yoon et al., (2011), menambahkan ekstrakjamur memiliki tingkat senyawa fenolik yangterdiri dari satu atau lebih cincin aromatik dansatu atau lebih gugus hidroksil, sehinggamenunjukkan pengurangan radikal sebagaidonor hidrogen atau agen menyumbangkanelektron, serta sifat ion logam.
KESIMPULAN
Dari penelitian yang telah dilakukan, makadapat disimpulkan perlakuan tanpa pencucianmedia serbuk gergaji memberikan rata-rata totalpolifenol tertinggi dengan jenis Jamur tiramkelabu Sedangkan dalam aktifitas antioksidan,perlakuan dengan pencucian justru memberikannilai terbaik pada jenis Jamur tiram pink.
DAFTAR PUSTAKA
Anonymous. 2009. Bertanam Jamur Konsumsi(Tiram, Kuping, Shiitake, Merang danChampignon). PT. Agromedia Pustaka.Jakarta.
Barros, L., S.Falcao., P.Baptista., C.Freire,.M.Vilas-Boas and I.C.F.R. Ferreira. 2008.Antioxidant activity of Agaricus sp.mushrooms by chemical, biochemical andelectrochemical assays. Journal FoodChemistry .111 : 61-66.
Choi, Y., S.M Lee., J.Chun., H.B.Lee and J.Lee.2006. Influence of heat treatment on theantioxidant activities and polyphenoliccompounds of Shiitake (Lentinus edodes)mushroom. Journal Food Chemistry. 99 :381-387.
Djarijah, N.M . 2001. Budidaya Jamur Tiram.Penerbit Kanisius. Yogyakarta.
Ferreira, I. C. F. R., P. Baptista., M.Vilas-Boasand L.Barros. 2007. Free-radicalscavenging capacity and reducing power ofwild edible mushrooms from northeastPortugal: Individual cap and stipe activity.Journal Food Chemistry. 100: 1511-1516.
Furham, B. dan M. Aviram. 2002. Polyphenolsand flavonoids protect LDLagainstatherogenic modification. Di dalam:Cadenas, E. dan L. Packer(Eds.) Handbookof Antioxidant 2nd Edition Revised andexpanded. MarcelDekker, inc. New York.
Gan,C.H., N.B. Amira and R.Asmah. 2013.Antioxidant analysis of different types ofedible mushrooms (Agaricus bisporous andAgaricus brasiliensis). International FoodResearch Journal 20(3): 1095-1102.
Gregori, A., S. Mirjan and J. Pohleven. 2007.Cultivation Techniques and MedicinalProperti es of Pleourotus spp. FoodTechnology and Biotechnology 45 (3): 238-249.
Halliwell, B., J.M.C.Gutteridge, dan C.E. Cros.1992. Free Radicals, Antioxidants andHuman Disease . Journal of LaboratoryClinical Medicine 119(6) : 598-620.
Hsu, T.H.,L.H.Shiao., C.Hsieh., and D.M.Chang.2002. A comparison of the chemicalcomposition and bioactive ingredients ofthe Chinese medicinal mushroom DongChong Xia Cao, its counterfeit and mimic,and fermented mycelium of Cordycepssinensis. Journal Food Chemistry 78: 463-469.
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 102
Gambar 4. Perbandingan tingkat kekeruhanekstrak yang dihasilkan dariperlakuan pencucian media padabeberapa jenis jamur tiram
Dari Gambar 4 diatas, terlihat bahwa perbedaanyang signifikan pada masing-masing ekstrakyang dihasilkan. Kepekatan ekstrak padamasing-masing jamur pada perlakuan tanpapencucian lebih tinggi jika dibandingkan denganekstrak dengan pencucian. Terjadinya perbedaantingkat kekeruhan diduga berkaitan dengankadar tanin yang diserap oleh jamur sehinggaturut mempengaruhi aktifitas antioksidan dantotal polifenol yang terdapat dalam tubuh buah.Vamanu (2012), menjelaskan bahwa dalamekstrak jamur kapasitas antioksidan ditentukanoleh jumlah senyawa fenolik yangdikandungnya. Kuantitas ekstrak juga mem-pengaruhi kapasitas penghambatan radikal bebas.Yoon et al., (2011), menambahkan ekstrakjamur memiliki tingkat senyawa fenolik yangterdiri dari satu atau lebih cincin aromatik dansatu atau lebih gugus hidroksil, sehinggamenunjukkan pengurangan radikal sebagaidonor hidrogen atau agen menyumbangkanelektron, serta sifat ion logam.
KESIMPULAN
Dari penelitian yang telah dilakukan, makadapat disimpulkan perlakuan tanpa pencucianmedia serbuk gergaji memberikan rata-rata totalpolifenol tertinggi dengan jenis Jamur tiramkelabu Sedangkan dalam aktifitas antioksidan,perlakuan dengan pencucian justru memberikannilai terbaik pada jenis Jamur tiram pink.
DAFTAR PUSTAKA
Anonymous. 2009. Bertanam Jamur Konsumsi(Tiram, Kuping, Shiitake, Merang danChampignon). PT. Agromedia Pustaka.Jakarta.
Barros, L., S.Falcao., P.Baptista., C.Freire,.M.Vilas-Boas and I.C.F.R. Ferreira. 2008.Antioxidant activity of Agaricus sp.mushrooms by chemical, biochemical andelectrochemical assays. Journal FoodChemistry .111 : 61-66.
Choi, Y., S.M Lee., J.Chun., H.B.Lee and J.Lee.2006. Influence of heat treatment on theantioxidant activities and polyphenoliccompounds of Shiitake (Lentinus edodes)mushroom. Journal Food Chemistry. 99 :381-387.
Djarijah, N.M . 2001. Budidaya Jamur Tiram.Penerbit Kanisius. Yogyakarta.
Ferreira, I. C. F. R., P. Baptista., M.Vilas-Boasand L.Barros. 2007. Free-radicalscavenging capacity and reducing power ofwild edible mushrooms from northeastPortugal: Individual cap and stipe activity.Journal Food Chemistry. 100: 1511-1516.
Furham, B. dan M. Aviram. 2002. Polyphenolsand flavonoids protect LDLagainstatherogenic modification. Di dalam:Cadenas, E. dan L. Packer(Eds.) Handbookof Antioxidant 2nd Edition Revised andexpanded. MarcelDekker, inc. New York.
Gan,C.H., N.B. Amira and R.Asmah. 2013.Antioxidant analysis of different types ofedible mushrooms (Agaricus bisporous andAgaricus brasiliensis). International FoodResearch Journal 20(3): 1095-1102.
Gregori, A., S. Mirjan and J. Pohleven. 2007.Cultivation Techniques and MedicinalProperti es of Pleourotus spp. FoodTechnology and Biotechnology 45 (3): 238-249.
Halliwell, B., J.M.C.Gutteridge, dan C.E. Cros.1992. Free Radicals, Antioxidants andHuman Disease . Journal of LaboratoryClinical Medicine 119(6) : 598-620.
Hsu, T.H.,L.H.Shiao., C.Hsieh., and D.M.Chang.2002. A comparison of the chemicalcomposition and bioactive ingredients ofthe Chinese medicinal mushroom DongChong Xia Cao, its counterfeit and mimic,and fermented mycelium of Cordycepssinensis. Journal Food Chemistry 78: 463-469.
Hafizatur Rahma, Nurmiati dan Anthoni Agustien 102
Gambar 4. Perbandingan tingkat kekeruhanekstrak yang dihasilkan dariperlakuan pencucian media padabeberapa jenis jamur tiram
Dari Gambar 4 diatas, terlihat bahwa perbedaanyang signifikan pada masing-masing ekstrakyang dihasilkan. Kepekatan ekstrak padamasing-masing jamur pada perlakuan tanpapencucian lebih tinggi jika dibandingkan denganekstrak dengan pencucian. Terjadinya perbedaantingkat kekeruhan diduga berkaitan dengankadar tanin yang diserap oleh jamur sehinggaturut mempengaruhi aktifitas antioksidan dantotal polifenol yang terdapat dalam tubuh buah.Vamanu (2012), menjelaskan bahwa dalamekstrak jamur kapasitas antioksidan ditentukanoleh jumlah senyawa fenolik yangdikandungnya. Kuantitas ekstrak juga mem-pengaruhi kapasitas penghambatan radikal bebas.Yoon et al., (2011), menambahkan ekstrakjamur memiliki tingkat senyawa fenolik yangterdiri dari satu atau lebih cincin aromatik dansatu atau lebih gugus hidroksil, sehinggamenunjukkan pengurangan radikal sebagaidonor hidrogen atau agen menyumbangkanelektron, serta sifat ion logam.
KESIMPULAN
Dari penelitian yang telah dilakukan, makadapat disimpulkan perlakuan tanpa pencucianmedia serbuk gergaji memberikan rata-rata totalpolifenol tertinggi dengan jenis Jamur tiramkelabu Sedangkan dalam aktifitas antioksidan,perlakuan dengan pencucian justru memberikannilai terbaik pada jenis Jamur tiram pink.
DAFTAR PUSTAKA
Anonymous. 2009. Bertanam Jamur Konsumsi(Tiram, Kuping, Shiitake, Merang danChampignon). PT. Agromedia Pustaka.Jakarta.
Barros, L., S.Falcao., P.Baptista., C.Freire,.M.Vilas-Boas and I.C.F.R. Ferreira. 2008.Antioxidant activity of Agaricus sp.mushrooms by chemical, biochemical andelectrochemical assays. Journal FoodChemistry .111 : 61-66.
Choi, Y., S.M Lee., J.Chun., H.B.Lee and J.Lee.2006. Influence of heat treatment on theantioxidant activities and polyphenoliccompounds of Shiitake (Lentinus edodes)mushroom. Journal Food Chemistry. 99 :381-387.
Djarijah, N.M . 2001. Budidaya Jamur Tiram.Penerbit Kanisius. Yogyakarta.
Ferreira, I. C. F. R., P. Baptista., M.Vilas-Boasand L.Barros. 2007. Free-radicalscavenging capacity and reducing power ofwild edible mushrooms from northeastPortugal: Individual cap and stipe activity.Journal Food Chemistry. 100: 1511-1516.
Furham, B. dan M. Aviram. 2002. Polyphenolsand flavonoids protect LDLagainstatherogenic modification. Di dalam:Cadenas, E. dan L. Packer(Eds.) Handbookof Antioxidant 2nd Edition Revised andexpanded. MarcelDekker, inc. New York.
Gan,C.H., N.B. Amira and R.Asmah. 2013.Antioxidant analysis of different types ofedible mushrooms (Agaricus bisporous andAgaricus brasiliensis). International FoodResearch Journal 20(3): 1095-1102.
Gregori, A., S. Mirjan and J. Pohleven. 2007.Cultivation Techniques and MedicinalProperti es of Pleourotus spp. FoodTechnology and Biotechnology 45 (3): 238-249.
Halliwell, B., J.M.C.Gutteridge, dan C.E. Cros.1992. Free Radicals, Antioxidants andHuman Disease . Journal of LaboratoryClinical Medicine 119(6) : 598-620.
Hsu, T.H.,L.H.Shiao., C.Hsieh., and D.M.Chang.2002. A comparison of the chemicalcomposition and bioactive ingredients ofthe Chinese medicinal mushroom DongChong Xia Cao, its counterfeit and mimic,and fermented mycelium of Cordycepssinensis. Journal Food Chemistry 78: 463-469.

Hafizatur Rahma, Nurmiati dan Anthoni Agustien 103
Jose, N and K.K. Janardhanan. 2000.Antioxidant and Antitumour Activity ofPleurotus florida. Journal Current Science.(79): 941-943.
Lakshmi, B., J.C. Tilak., S. Adhikari., T.P.Decasagayam and K.K. Janardhanan. 2004.Avaluation of antioxidant activity ofselected indian mushrooms.Pharmaceutical Biology Journal 42: 179-185.
Lindequiest, U., T.H.J Viedermeyer and W.D.Julich. 2005. The Pharmacologicalpotential of mushroom Ediv. BasedComplement Alternat Med 2(3): 285-299.
Lo, K. M and P.C.K.Cheung. 2005. Antioxidantactivity of extracts from the fruiting bodiesof Agrocybe aegerita var. Alba. JournalFood Chemistry 89(4): 533-539.
Mau, J. L., G.R.Chao and K.T.Wu. 2001.Antioxidant properties of methanolicextracts from several ear mushrooms.Journal of Agricultural and FoodChemistry 49: 5461-5467.
Mau, J.L., H.C. Lin and S.F.Song. 2002.Antioxidant properties of several specialtymushrooms. Food Research International.35: 519-526.
Molyneux, P. 2004. The use of the stable freeradical diphenylpicrylhydrazyl (DPPH) forestimating antioxidant activity.Songklanakarin Journal Scince andTechnology. 26(2) :211- 219.
Muchrodi. 2001. Jamur Tiram Putih. PenebarSwadaya. Jakarta
Parrilla,A E., L.A. de-la Rosa., N.R. Martinezand G.A.G.Aguilar. 2007. Total PhenolsAnd Antioxidant Activity Of CommercialAnd Wild Mushrooms From Chihuahua,Mexico. Cienc. Tecnol. Aliment. 5 (5) :329-334.
Saha, A.K., A.Sandeep and R. Aparajita. 2012.Antioxidant level ofwild edible mushroom:Pleurotus djamor (Fr.) Boedijn.Journal ofAgricultural Technology. Vol. 8(4): 1343-1351.
Vamanu, E. 2012. In Vitro Antimicrobial andAntioxidant Activities of EthanolicExtractof Lyophilized Mycelium of Pleurotusostreatus. Molecules Journal 17: 3653-3671.
Winarsy,H. 2007. Antioksidan Alami danRadikal Bebas. Kanasius. Yogyakarta.
Yang, J.H., H.C.Lin and J.L.Mau. 2002.Antioxidant properties of severalcommercial mushrooms. Journal FoodChemistry 77(2): 229-235.
Yoon, K.N., A.Nuhu., R.L.Kyung., G.S.Pyung.,C.C. Jong Chun., B.Y.Young And S.Ltae,2011. Antioxidant and AntityrosinaseActivities of Various Extractsfrom theFruiting Bodies of Lentinus lepideus.Molecules Journal 16 :2334-2347.

ISSN 978-602-14989-0-3 BioETI
Studi deteksi patogen terbawa benih pada tanaman Jagung
HALIATUR RAHMA1*, MARTINIUS1, RATNA WULANDARI2 DAN TRIMARYONO3
1Program Studi Agroekoteknologi Faperta Universitas Andalas, Kampus Unand Limau Manis Padang 251632Balai Pengkajian Teknologi Pertanian (BPTP) Sumatera Barat. Jl. Raya Padang-Solok Km. 40,Solok 25001 - Sumbar3Program Studi Agroekoteknologi Fakultas Pertanian Universitas Lampung. Bandar Lampung 35145
E-mail: *[email protected]
ABSTRAKPenelitian ini bertujuan untuk mendeteksi patogen tular benih pada tanaman jagung. Penelitian dilakukan dengan tekniksurvei pada tiga propinsi di Sumatera, yaitu Sumatera Barat, Sumatera Utara dan Lampung yang berlangsung dari bulanMaret-September 2014. Patogen terbawa benih yang ditemukan di pada lokasi survei adalah penyakit layu stewart danbusuk tongkol dengan kejadian penyakit 14-45% dan 10-50%. Dari tanaman yang bergejala layu stewart berhasildiisolasi 34 isolat murni bakteri yang berwarna krem kekuningan, koloni bulat, elevasi datar hingga cembung dan gramnegatif dengan rincian (9 dari Sumatera Barat, 10 dari Sumatera Utara dan 15 isolat dari Lampung) yang mencirikansebagai bakteri Pnss. Sementara hasil isolasi dari gejala busuk tongkol diantaranya adalah cendawan Fusarium sp,Aspergillus sp, Penicillium sp, dan Trichoderma sp.
Key words: busuk tongkol, jagung, patogen tular benih, penyakit layu stewart
Pendahuluan
Impor benih jagung selain mampu mencukupikebutuhan dalam negeri, juga akanmeningkatkan masuknya patogen penyebabpenyakit yang belum terdapat di NegaraKesatuan Republik Indonesia, hal ini disebabkanbenih yang diimpor terdapat kemungkinanterinfenksi oleh patogen tular benih. Hal inidisebabkan karena benih merupakan salah satubahan perbanyakan tanaman yang merupakantempat yang baik bagi patogen untuk bertahandan sangat efektif sebagai wahana penyebaranpatogen tanaman dari satu tempat ke tempat lain.Menurut Shurtleff 1980), berbagai kelompokpatogen yang menyerang tanaman jagungdiantaranya adalah cendawan, bakteri, virus,serta nematoda, diketahui beberapa diantaranyabersifat sebagai patogen yang terbawa benihyang menjadi sumber inokulum pada tanamanjagung di lapangan .
Beberapa penyakit penting yang meng-infeksi tanaman jagung diantaranya penyakitlayu stewart (Pantoea stewartii subsp. stewartii,karat daun yang disebabkan oleh Pucciniasorghi dan P. Polysora, penyakit bulai(Peronoscerospora maydis), seedling blight
yang disebabkan oleh Aspergillus spp danPenicillium sp, penyakit busuk batang jagungdisebabkan oleh delapan spesies cendawan yaituColletotrichum graminearum, Diplodia maydis,Gibberella zeae, Fusarium moniliforme,Macrophomina phaseolina, Pythiumapanidermatum, Cephalosporium maydis, danCephalosporium acremonium (Shurtleff 1980),Maize Dwarf Mosaic Virus (White 1999).Sebagian besar patogen yang menginfeksijagung dapat ditularkan melalui benih,diantaranya Pantoea stewartii subsp. stewartii,Peronosclerospora maydis, Aspergillus flavus,Fusarium spp, dan Penicillium sp (Mardinus2003).
Pengujian kesehatan benih untuk mendeteksikeberadaan patogen pada benih mempunyai artipenting dalam menjamin pendistribusian benihyang menjadi alat transportasi bagi patogen.Disamping itu uji kesehatan benih berfungsisebagai sarana pengendalian mutu yangmenjamin kualitas benih dalam pertukaran benihbaik untuk tujuan penelitian maupunperdagangan. Hanya saja, sampai saat inipengujian kesehatan benih di Indonesia belumbersifat wajib, hanya berdasarkan permintaandari konsumen (Ilyas 2008). Untuk memenuhi

Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 105
persyaratan benih bermutu, maka diperlukanketersediaan metode standar untuk mendeteksiorganisme pengganggu tanaman (OPT)khususnya patogen terbawa benih, yangdibutuhkan adalah sistem deteksi dini yangdapat diterima secara luas. Tujuan penelitian iniadalah untuk deteksi dan identifikasi patogenterbawa benih dari sentra pertanaman jagung diSumbar, Sumut dan Lampung.
BAHAN DAN METODE
Penelitian dilakukan dari bulan Maret-September 2014, penelitian dilakukan diLaboratorium Mikrobiologi Program StudiAgroekoteknologi Faperta Unand danLaboratorium BPTPH Sumatera Barat.
Survei dilakukan di beberapa sentra produksijagung di Sumatera yaitu Propinsi SumateraBarat (Pasaman Barat), Sumatera Utara(Sidikalang-Dairi) dan Propinsi Lampung(Sukadana-Lampung Timur), penentuan daerahsentra berdasarkan data BPS (2013). Padamasing-masing daerah sentra dipilih 3 desa dandari masing-masing desa diambil 2 contohkebun jagung yang akan dijadikan petak contohpengambilan sampel. Sampel yang diambilberupa benih, daun, batang (5-10 helai), tongkolserta malai, serta sampel tanah yang ditujukanuntuk mendeteksi patogen tular tanah yang jugabersifat terbawa benih.Isolasi bakteri Pantoea stewartii subsp.stewartii pada tanaman jagung bergejalapenyakit layu stewartIsolasi bakteri dilakukan berdasarkan protokolThai Agricultural Standard (2008). EkstraksiPnss dari tanaman jagung bergejala secaralangsung dilakukan menggunakan media TrypticSoy Agar (TSA). Sampel tanaman sakitdisterilisasi permukaan dengan air leding sampaibersih. Sampel daun dipotong 1 x 1 cmkemudian dimasukkan ke dalam erlenmeyer 250ml yang berisi 100 ml media TSA. Sampeldiinkubasi pada inkubator bergoyang 110 rpmsemalam. Suspensi bakteri hasil ekstraksidipindahkan ke tabung eppendorf steril,diencerkan 10 atau 100 kali menggunakan
akuades steril. Kemudian disebar pada mediaTSA diratakan dengan glass beads steril.Inkubasi dilakukan pada suhu 25-27 oC selama2-3 hari. Koloni Pnss pada media TSA agarakan berwarna kuning cerah, berkilat namuntidak berlendir. Koloni yang memiliki ciri-cirisebagai Pnss dimurnikan pada media TSA.Deteksi cendawan terbawa benih bergejalabusuk tongkolUntuk merangsang pertumbuhan cendawan yangterbawa pada benih jagung, dilakukan platingbenih pada cawan petri yang telah dilapisi tigalembar kertas saring steril basah. Setiap petriditanam 10 biji. Kemudian benih diinkubasiselama 6, 12, 18 dan 24 jam pada suhu ruang.Setelah masa inkubasi masing-masing benihtersebut diekstraksi, yaitu benih dimasukkankedalam 250 ml air steril, kemudian dikocokselama 12 jam. Cendawan yang tumbuhdiisolasi dan dimurnikan pada media PDA.
HASIL DAN PEMBAHASAN
Penyakit Layu StewartGejala penyakit layu stewart ditemukan berupatanaman kerdil (pada saat fase vegetative),bercak kuning disepanjang pertulangan daun.Kejadian penyakit yang ditemukan berkisar 15 –45%. Gejala penyakit layu stewart hamper miripdengan penyakit bulai yang disebabkan olehcendawan Peronosclerospora maydis, yangmembedakannya adalah bercak me-manjangpada penyakit layu stewart berwana kuningkehijauan, sedangkan pada bulai berwarna putih.Pada penyakit layu stewart tidak ada batasanyang jelas antara bercak kuning dengan warnahijau pada daun, sementara pada penyakit bulaibatasan antara garis putih dan hijau terlihatdengan jelas.Penyakit Busuk TongkolGejala busuk tongkol ditandai dengan matinyasebagian tanaman, kelobot tongkol terlihatmenguning seperti kelobot yang sudah matang,diantara biji ditemukan adanya massa berwarnaputih-pink, ada juga yang massa berwarna hijauyang diduga sebagai miselia cendawan. Penyakit

Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 106
busuk tongkol ditemukan pada hampir semualahan tapi dengan intensitas 10-50%, namun diPasaman Barat gejala busuk tongkol ditemukandengan kejadian penyakit yang sangat tinggi(90%) pada jagung Pasific 105, sehingga jagungini tidak bisa dipanen. Di Lampung kejadianpenyakit tongkol yang paling tinggi ditemukanpada jagung Pacific 105 dengan intensitas 30%,sementara di Sumatera Utara kejadian penyakittertinggi 25%. Dari hasil survei, penyakit busuktongkol ditemukan pada semua varietas dengankejadian penyakit yang paling tinggi (100%)ditemukan pada varietas Pacific 105. Berdasar-kan gejala yang ditemukan di lapangan makatanaman sakit serta sampel tongkol yangbergejala diambil untuk analisis di laboratorium.Sampel benih yang ditanam petani juga diambiluntuk melihat keberadaan pathogen pada benih.Isolasi, Identifikasi dan Koleksi PatogenPantoea stewartii subsp. stewartii
Pada media TSA koloni bakteri Pnssberwarna kuning, krem kekuningan, elevasicembung, dan tidak berlendir (Gb.1). Dari hasilisolasi diperoleh 34 isolat murni (9 dariSumatera Barat, 10 dari Sumatera Utara dan 15isolat dari Lampung) yang mencirikan sebagaibakteri Pnss.
Berdasarkan informasi dari petani diPasaman Barat, gejala layu stewart dikenalsebagai penyakit “ular” karena tanamanbergejala biasanya daun agak melintir.Sementara petani di Karo (Sumatera Utara)menyebut gejala ini dengan penyakit “musing”(berputar). Berdasarkan survei di lapangangejala penyakit ini dapat ditemukan pada lahanyang menunjukkan pertumbuhan tanaman tidakseragam, hal ini disebabkan pathogen penyebabpenyakit ini merupakan pathogen tular benih.Kejadian penyakit berkisar 15 - 40% danditemukan pada semua jenis jagung yangditanaman. Intensitas tertinggi ditemukan padavarietas Pioneer 23 dan Pioneer 27.Isolasi Cendawan dari Gejala Busuk TongkolGejala busuk tongkol pada tanaman jagungditandai dengan matinya sebagian tanaman,diantara biji ditemukan adanya massa berwarna
putih-pink yang diduga sebagai miseliacendawan. Pada pengamatan di lapangan petanidi Pasaman Barat cenderung menggunakanvarietas jagung yang sama, NK 99, NK 22,Pioneer 23 dan Pacific 105. Dari pengamatanpenyakit busuk tongkol ditemukan pada semuavarietas, kejadian penyakit yang paling tinggi(100%) ditemukan pada varietas Pacific 105.
Cendawan yang berhasil diisolasi dari gejalabusuk tongkol dan benih jagung diperoleh 5jenis cendawan. Isolat tersebut adalah Fusariumsp, Aspergillus flavus, A.niger, Trichoderma spdan Penicllium sp.
Gejala penyakit layu stewart yang ditemukanpada lokasi survei, baik di sumatera Barat,Sumatera Utara maupun Lampung, miripdengan gejala penyakit layu stewart yangditemukan di Sumatera Barat (Rahma danArmansyah 2008). Kejadian penyakit layustewart pada ketiga lokasi survei tergolong lebihrendah bila dibandingkan dengan kejadianpenyakit yang ditemukan di Jawa Barat yaitu23,67% - 52,41%, hal ini disebabkan perbedaanjenis jagung yang digunakan. Pada umumnyapetani jagung di ketiga lokasi survei di Sumateramenanam jagung hibrida, sementara di JawaBarat lebih banyak menanam jagung manis danJawa Barat (Rahma 2013). Menurut (Freeman &Pataky 2001), jagung manis sangat rentanterhadap Pantoea stewartii subsp. stewartiidengan tingkat keparahan penyakit bisamencapai hingga 100%, sementara jagunghibrida dan bersari bebas lebih tahan terhadapPnss.
Penyakit busuk tongkol ditemukan padasemua jenis jagung. Dari sampel benih yangdiambil dari lapangan ternyata ditemukan lebihdari 1 jenis cendawan. Data menunjukkanbahwa Fusarium sp (2 jenis), Aspergillus sp (2jenis) merupakan cendawan yang selalu adapada setiap sampel benih yang diuji danditemukan pada setiap lokasi survei, sementaraPenicillium sp dan Trichoderma sp ditemukanpada beberapa sampel benih dan ada pada setiaplokasi survei.
c

Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 107
Gambar 1. Isolat murni Pnss pada media TSA.(a) Isolat dari Laumeciho, Karo(Sumatera Utara), (b) Isolat dariPerbesi Karo (Sumatera Utara), (c)Isolat Ketapang (Lampung Selatan),Isolat Pasaman Barat (SumateraBarat)
Berdasarkan ciri-ciri morfologisnya cendawanFusarium sp menunjukkan miselium yangberwarna putih, semakin tua warna merah mudaagak ungu ditemukan 2 jenis Fusarium spberdasarkan karakteristik mikroskopis yangpertama mikrokonidia insitu yang berbentukseperti rantai tanpa disertai denganmakrokonidia, isolat Fusarium yang keduakarakter mikroskopisnya hanya adamakrokonidia yang memiliki sekat (Gb. 2A).Aspergillus flavus koloni berwarna hijaukekuningan, tekstur agak kasar, konidiaberwarna hijau, fialid memenuhi seluruhpermukaan vesikel, dan vesikel yang berbentukbulat. Koloni cendawan Aspergillus nigerberwarna hitam, konidia berwarna hitam, fialidmemenuhi permukaan vesikel, miselia berwarnacoklat kehitaman (Gb. 2B).
Penicillium sp, hampir mirip denganAspergillus sp, namun memiliki konidiofor yangbercabang (Gb. 3A). Sementara Trichoderma sp,memiliki ciri-ciri warna koloni hijau, teksturseperti bulu, konidiofor memiliki banyak cabangdan hialin (Gb. 3B)
Menurut Niaz dan Dawar (2009), cendawanFusarium sp dan Aspergillus sp merupakanpatogen utama pada benih jagung, persentaseinfeksinya lebih tinggi bila dibandingkandengan inang lainnya seperti kacang tanah dansorgum. Akibat dari keberadaan patogen ini
pada benih jagung adalah penurunan baik darisegi kuantitatif maupun kualitatif, mem-pengaruhi kualitas benih jagung melaluipeningkatan asam lemak, penurunanperkecambahan, kelapukan dan pembusukanpada benih, mikotoksin sebagai hasil metabolitsekunder sangat berbahaya bagi kesehatanmanusia maupun hewan ternak.
Gambar 2. Fusarium sp. A. Fusarium 1 denganmikrokonidia insitu (a) danmikrokonidia (b). B. Fusarium 2 denganmakrokonidia insitu (a), makrokonidiayang bersekat (b). C-D Aspergillus sp.fialid (a), vesikel (b), konidiofor (c).
Gambar 3. A. Cendawan Penicillium sp, dengankonidia (a), fialid (b), konidiofor(c), dan cabang konidiofor (d). B.Trichoderma sp, dengan konidia(a), fialid (b) dan konidiofor (c).
Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 107
Gambar 1. Isolat murni Pnss pada media TSA.(a) Isolat dari Laumeciho, Karo(Sumatera Utara), (b) Isolat dariPerbesi Karo (Sumatera Utara), (c)Isolat Ketapang (Lampung Selatan),Isolat Pasaman Barat (SumateraBarat)
Berdasarkan ciri-ciri morfologisnya cendawanFusarium sp menunjukkan miselium yangberwarna putih, semakin tua warna merah mudaagak ungu ditemukan 2 jenis Fusarium spberdasarkan karakteristik mikroskopis yangpertama mikrokonidia insitu yang berbentukseperti rantai tanpa disertai denganmakrokonidia, isolat Fusarium yang keduakarakter mikroskopisnya hanya adamakrokonidia yang memiliki sekat (Gb. 2A).Aspergillus flavus koloni berwarna hijaukekuningan, tekstur agak kasar, konidiaberwarna hijau, fialid memenuhi seluruhpermukaan vesikel, dan vesikel yang berbentukbulat. Koloni cendawan Aspergillus nigerberwarna hitam, konidia berwarna hitam, fialidmemenuhi permukaan vesikel, miselia berwarnacoklat kehitaman (Gb. 2B).
Penicillium sp, hampir mirip denganAspergillus sp, namun memiliki konidiofor yangbercabang (Gb. 3A). Sementara Trichoderma sp,memiliki ciri-ciri warna koloni hijau, teksturseperti bulu, konidiofor memiliki banyak cabangdan hialin (Gb. 3B)
Menurut Niaz dan Dawar (2009), cendawanFusarium sp dan Aspergillus sp merupakanpatogen utama pada benih jagung, persentaseinfeksinya lebih tinggi bila dibandingkandengan inang lainnya seperti kacang tanah dansorgum. Akibat dari keberadaan patogen ini
pada benih jagung adalah penurunan baik darisegi kuantitatif maupun kualitatif, mem-pengaruhi kualitas benih jagung melaluipeningkatan asam lemak, penurunanperkecambahan, kelapukan dan pembusukanpada benih, mikotoksin sebagai hasil metabolitsekunder sangat berbahaya bagi kesehatanmanusia maupun hewan ternak.
Gambar 2. Fusarium sp. A. Fusarium 1 denganmikrokonidia insitu (a) danmikrokonidia (b). B. Fusarium 2 denganmakrokonidia insitu (a), makrokonidiayang bersekat (b). C-D Aspergillus sp.fialid (a), vesikel (b), konidiofor (c).
Gambar 3. A. Cendawan Penicillium sp, dengankonidia (a), fialid (b), konidiofor(c), dan cabang konidiofor (d). B.Trichoderma sp, dengan konidia(a), fialid (b) dan konidiofor (c).
Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 107
Gambar 1. Isolat murni Pnss pada media TSA.(a) Isolat dari Laumeciho, Karo(Sumatera Utara), (b) Isolat dariPerbesi Karo (Sumatera Utara), (c)Isolat Ketapang (Lampung Selatan),Isolat Pasaman Barat (SumateraBarat)
Berdasarkan ciri-ciri morfologisnya cendawanFusarium sp menunjukkan miselium yangberwarna putih, semakin tua warna merah mudaagak ungu ditemukan 2 jenis Fusarium spberdasarkan karakteristik mikroskopis yangpertama mikrokonidia insitu yang berbentukseperti rantai tanpa disertai denganmakrokonidia, isolat Fusarium yang keduakarakter mikroskopisnya hanya adamakrokonidia yang memiliki sekat (Gb. 2A).Aspergillus flavus koloni berwarna hijaukekuningan, tekstur agak kasar, konidiaberwarna hijau, fialid memenuhi seluruhpermukaan vesikel, dan vesikel yang berbentukbulat. Koloni cendawan Aspergillus nigerberwarna hitam, konidia berwarna hitam, fialidmemenuhi permukaan vesikel, miselia berwarnacoklat kehitaman (Gb. 2B).
Penicillium sp, hampir mirip denganAspergillus sp, namun memiliki konidiofor yangbercabang (Gb. 3A). Sementara Trichoderma sp,memiliki ciri-ciri warna koloni hijau, teksturseperti bulu, konidiofor memiliki banyak cabangdan hialin (Gb. 3B)
Menurut Niaz dan Dawar (2009), cendawanFusarium sp dan Aspergillus sp merupakanpatogen utama pada benih jagung, persentaseinfeksinya lebih tinggi bila dibandingkandengan inang lainnya seperti kacang tanah dansorgum. Akibat dari keberadaan patogen ini
pada benih jagung adalah penurunan baik darisegi kuantitatif maupun kualitatif, mem-pengaruhi kualitas benih jagung melaluipeningkatan asam lemak, penurunanperkecambahan, kelapukan dan pembusukanpada benih, mikotoksin sebagai hasil metabolitsekunder sangat berbahaya bagi kesehatanmanusia maupun hewan ternak.
Gambar 2. Fusarium sp. A. Fusarium 1 denganmikrokonidia insitu (a) danmikrokonidia (b). B. Fusarium 2 denganmakrokonidia insitu (a), makrokonidiayang bersekat (b). C-D Aspergillus sp.fialid (a), vesikel (b), konidiofor (c).
Gambar 3. A. Cendawan Penicillium sp, dengankonidia (a), fialid (b), konidiofor(c), dan cabang konidiofor (d). B.Trichoderma sp, dengan konidia(a), fialid (b) dan konidiofor (c).

Haliatur Rahma, Martinius, Ratna Wulandari dan Trimaryono 108
Berdasarkan data diatas dapat dilihat bahwabaik penyakit layu stewart maupun busuktongkol ditemukan pada setiap lokasi surveipada beberapa jenis jagung yang ditanam olehpetani. Kejadian penyakit dari masing-masingpenyakit terlihat cukup tinggi sehingga perludiantisipasi supaya penyakit ini tidak menjadipenyakit endemic didaerah survei.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :
Dari tanaman yang bergejala layu stewartberhasil diisolasi 34 isolat murni bakteri yangberwarna krem kekuningan, koloni bulat, elevasidatar hingga cembung dan gram negatif denganrincian (9 dari Sumbar, 10 dari Sumut dan 15isolat dari Lampung). Sementara hasil isolasidari gejala busuk tongkol diantaranya adalahcendawan Fusarium sp, Aspergillus sp,Penicillium sp, dan Trichoderma sp.
UCAPAN TERIMA KASIHKami mengucapkan terima kasih kepada BadanPenelitian dan Pengembangan Pertanian(Balitbangtan), atas Hibah Penelitian KerjaSama Kemitraan Penelitian dan PengembanganPertanian Nasional (KKP3N) TA 2014 denganNomor Kontrak 101/PL.220/I.1/3/2014.K.Tanggal 10 Maret 2014.
DAFTAR PUSTAKA
BPS. 2013. Produktivitas dan produksi tanamanpangan. http://www.bps.go.id/tnmn_pgn.php?kat=3 [8 Januari 2014].
Freeman ND and Pataky JK. 2001. Levels ofstewart’s wilt resistance necessary topreven reductions in yield of sweet cornhybrids. Plant Dis. 85:1278-1284.
Ilyas, S. 2010. Ilmu dan Teknologi Benih: Teoridan Hasil-hasil Penelitian. InstitutPertanian Bogor
Mardinus. 2003. Patologi dan Benih JamurGudang. Andalas University Press. 342 hal.
Niaz, I and S. Dawar. 2009. Detection Of SeedBorne Mycoflora In Maize (Zea Mays L.).Pak. J. Bot., 41(1): p. 443-451.
Rahma, H dan Armansyah. 2008. DeteksiPenyakit Layu Stewart oleh BakteriPantoea stewartii sebagai Penyakit Barupada Tanaman Jagung (Zea mays): StudiKasus di Pasaman Barat. Vol. 9. No. 2 p. 1-5. November 2008. Jurnal Manggaro ISSN1410-9719.
Rahma, H. 2013. Penyakit Layu Stewart(Pantoea stewartii subsp. stewartii) padaJagung dan Upaya Pengendaliannya.Disertasi. Institut Pertanian Bogor.
Shurtleff, M.C. 1980. Compendium of CornDiseases. Second Edition. APS Press. TheAmerican Phytopathological Sociaty.
Thai Agricultural Standard. 2008. Diagnosticprotocols for Pantoea stewartii subsp.stewartii bacterial wilt of maize. NationalBureau of Agricultural Commodity AndFood Standards Ministry of Agricultureand Cooperatives.
White, D.G. 1999. Compendium of CornDiseases, Third Edition. The AmericanPhytopathological Society, USA. 128 p

ISSN 978-602-14989-0-3 BioETI
Studi komparatif Sagu (Metroxylon Rottb) sebagai media bibitproduksi terhadappertumbuhan Miselium dan aktifitas Amilasedan SelulaseJamur Merang (Volvariella volvacea(Bull.)Sing.)HARSUNA YUMNA, NURMIATI* DAN PERIADNADI
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: nurmiati @ fmipa.unand.ac.id
ABSTRAKJamur merang (Volvariella volvacea. (Bull). Sing.) memiliki potensi yang tinggi baik bagi kesehatan maupun secaraekonomis. Jamur ini sangat bermanfaat untuk kesehatan karena kualitas gizinya yang komplit. Bibit merupakan salah satusarana yang sangat penting bagi keberhasilan budidaya. Bibit harus berasal dari biakan murni, bebas dari kontaminasi danmemiliki sifat-sifat genetik unggul sehingga mampu memberikan hasil yang optimal. Penelitian ini bertujuan untukmengetahui perbandingan beberapa Media Bibit Produksi (terdiri dari kombinasi sagu-jerami:(0:4),(1:3),(2:2),(3:1),(4:0))terhadap kecepatan pertumbuhan miselium jamur merang. Hasil penelitian menunjukankecepatan pertumbuhan miselium jamur merang pada Media Bibit Produksi yang terbaik didapat pada dosis sagu:jerami(3:1) (direndam) dengan kecepatan pertumbuhan miselium yaitu 11.66 hari setelah inokulasi serta Aktifitas Amilasetertinggi (0.1088 unit/g) dan Aktifitas Selulase tertinggi (0.0657 unit/g)
Key words: Media Bibit Produksi, Sagu-Jerami, Jamur merang, Miselium
Pendahuluan
Jamur merang memiliki potensi yang tinggisebagai jamur kosumsi maupun karenamemiliki daging buah yang lezatdengannilaigizi yang tinggi, terutama kaya akanprotein, fosfor, kalsium dan mengandungberbagai zat esensial yang berguna bagimetabolism tubuh dan pertukaran sel (Trimurti,2009).
Berdasarkan kandungannya jamur inimengandung banyak zat yang bermanfaat.Menurut Suriawiria, (2007), zat yangterkandung V. volvacea berkhasiat obat sepertiVolvatoksin dan flummuttoksin (cardiac tonic)yang dapat memacu kerja jantung sehinggabermanfaat bagi penderita gangguan fungsijantung. Selain itu menurut Parjimo danAndoko (2007), Jamur merang mengandungsenyawa eritadenin yang berkhasiat sebagaianti racun dan mengandung sejenis antibiotikyang dapat mencegah anemia serta menurunkantekanan darah tinggi. Sedangkan menurut Chenet al., 2010, ekstrak jamur merang dapatmenurunkan viabilitas sel kanker. Beberapafaktor diatas menyebakan jamur merang
sebagai komoditi holtikultura yang bernilaiekonomis tinggi.
Kualitas bibit merupakan salah satu saranayang sangat penting bagi keberhasilan budidayajamur merang. Bibit yang berkualitas terlihatpada pertumbuhan miselium yang padat danmerata. Bibit harus berasal dari biakan murni,bebas dari kontaminasi dan memiliki sifat-sifatgenetik unggul sehingga mampu memberikanhasil yang optimal (Oei dalam Permatasari,2007). Media Bibit Produksi jamur merangselama ini disediakan dalam botol denganviabilitas hanya sampai 2-3 minggu setelahdiinokulasi. Pertumbuhan miselium MediaBibit Produksi yang tidak merata menyebabkanproduksi jamur merang tiap bedengan mediajerami tidak sama dimana setiap 1 M2 jeramimembutuhkan bibit yang relatif banyak yaitudua botol bibit atau lebih sehingga secaraekonomi tidak efisien (Sinaga, 2008)
Sagu merupakan tanaman tropik yang kayaakan kandungan pati dan merupakan sumberpangan alternatif setelah beras. Secara kimiawipati sagu memiliki kandungan karbohitrat lebihtinggi dari pada jagung dan beras tetapikandungan lemak dan proteinnya rendah. Pati

Harsuna Yumna, Nurmiati dan Periadnadi 110
sagu mengandung 28 % amilosa dan 72 %amilopeptin (Harsanto, 1989 dalam Ade, 2008).Karena kandungan pati sagu yang sangat tinggibila ditambahkan kedalam Media BibitProduksi maka diharapkan dapat menambahkecepatan pertumbuhan miselium danproduktifitas jamur merang dan secaraekonomis dapat menghemat jumlah bibitsehingga lebih menguntungkan.
Tujuan penelitian ini adalah untukmengetahui pertumbuhan miselium jamurmerang pada beberapa Media Bibit Produksidan untuk mengetahui Aktifitas Amilase danSelulase jamur merang (V. volvacea) padabeberapa Media Bibit Produksi
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan Juni sampaiDesember 2013 di Labor Mikrobiologi JurusanBiologi FMIPA, Universitas Andalas Padangdan di kebun budidaya jamur tiram” NUBEJA“ di Komp Pemda blok A no 24 Padang SaraiLubuk Buaya Padang.Bahan-bahan yang digunakan dalam penelitianini adalah : limbah sagu, Potongan jerami,bekatul, dolomit, Na2CO3anhidrat, KnaTartarat,NaHCO3, Na2SO4anhidrat, CuSO4.5H2O,H2SO4, Glukosa, natriumasetat, asamasetat,CMC, pati, Aquades steril, Alkohol.
Biakan murni jamur merang (Volvariellavolvaceae (Bull). Sing.) berasal dari hasilisolasi tubuh buah jamur yang dibiakkan padaagar miring PDA (Potatoes Dectrose Agar).Dari biakan murni tersebut miselium bibitjamur merang dipindahkan ke media biji-bijian.Penelitian inimenggunakan Rancangan AcakLengkap (RAL) Faktorialdengan dua faktor,faktor pertama terdiri dari duaperlakuan,tanpaperendaman jerami dan dengan perendamanjerami. Faktor kedua terdiri dari lima dosissagu:jerami: (0:4), (1:3), (2:2), (3:1) dan (4:0).Masing-masing perlakuan diulang sebanyak 3kali sehingga diperoleh 30 unit percobaan.Parameter yang diamati adalah kecepatanpertumbuhan miselium dan aktivitas Amilasedan Selulase.
Penyediaan Biakan MurniBiakan murni merupakan miselium jamur yangtumbuh pada agar miring. Untuk mendapatkanbiakan murni, hifa yang diisolasi dari tubuhbuah jamur merang dibiakkan pada mediumdasar yaitu Potato Dekstrosa Agar (PDA) sterildalam petridish.
Pembuatan Media Bibit IndukBahan baku Media Bibit Induk berupa biji-bijian (jagung) direndam selama 12 jam,kemudian biji–bijian dilakukan perebusanselama 30 menit, Selanjutnya biji-bijiandimasukan kedalam botol saus tomat volume400 g dan disterilkan dalam Autoclave suhu121o C tekanan 1 ATM selama 15 menit.Kemudian Miselium biakan murni jamurmerang pada testube diinokulasikan pada botolsaus tomat yang berisi Media Bibit Induk(terdiri dari biji-bijian). Botol ditutup kapas dandiinkubasi pada Incubator temperatur 30o Csampai miselium bibit jamur merang tumbuhsecara maksimum.
Pembuatan Media Bibit ProduksiBahan-bahan yang dijadikan Media BibitProduksi adalah jerami yang diperoleh daritempat pemanenan di kenagarian Anak Air KecKoto Tangah Padang. Sedangkan bekatuldiperoleh dari penggilingan padi di kenagarianTabek Kec Pariangan, Tanah Datar dan kapuryang didapatkan dari pabrik pengolahan kapurPadang Panjang. Setelah media jeramidiperoleh, sebagian direndam dengan airselama 1 hari, lalu ditiriskan dan sebagian lagitidak direndam tapi hanya dialiri air saja. Padakedua media ditambahkan kapur 1% danbekatul 15%. Diaduk sampai rata laluditambahkan limbah sagu sesuai dosis : (0:4),(1:3), 2:2), (3:1), (4:0). Semua bahan dicampur,kemudian media dikompos selama 10 hari.Media yang telah dikomposkan kemudiandimasukan kedalam plastik PP ukuran 17 x 10lalu disterilkan dengan menggunakan drumpada suhu 1000C selama 7 jam.

Harsuna Yumna, Nurmiati dan Periadnadi 111
Inokulasi Media Bibit Induk ke Media BibitProduksiMiselium dari Media Bibit Induk jamur merangyang tumbuh pada media biji-bijian, laludipindahkan ke botol yang berisi Media BibitProduksi. Botol ditutup dengan kapas daningkubasi pada Inkubator temperatur 32 oCsampai miselium bibit jamur merang tumbuhsecara maksimum.Uji Aktifitas Amilase dan Selulase
Media Bibit Produksi yang telah dipenuhimiselium jamur merang, diuji AktifitasAmilase dan Selulase dengan cara mediamasing-masing perlakuan ditimbang sebanyaklima gram lalu ditambahkan aquades sampai 50ml diaduk sampai homogen. Diambil cairanlalu disentrifuge selama 15 menit sehinggadidapat filrat enzim.
Larutan Pati 1% untuk amylase danCMC1% untukselulasesebanyak 1 ml dimasukkan kedalam tabung reaksi dan dipreinkubasikan padasuhu 40oC selama 5 menit kemudiandimasukkan larutan ekstrak filtrat enzimsebanyak 1 ml dan diinkubasikan pada suhu40oC selama 30 menit. Campuran tersebutkemudian dimasukkan ke dalam air mendidihselama 20 menit, lalu tambahkan denganSomogy-Nelson sebanyak1 ml, selanjutnyadivortex dan dipanaskan lagi pada air mendidihselama 20 menit. Larutan didinginkan segeradalam air es hingga suhunya mencapai 26oClalu tambahkan ragen arsenomolibdat sebanyak1 ml dan dikocok sampai tidak terlihat adanyagas keluar. Selanjutnya, dicukupkan volumelarutan menjadi 10 ml dengan aquades.Campuran larutan tersebut dikocok kembalisehingga tidak ada gelembung udara lagi danselanjutnya dilakukan pengukuran absorbandengan spektrofotometer pada panjanggelombang 540 nm. Pengukuran kontrol samadengan perlakuan sampel, hanya pada kontrolenzim, 1 ml filtrat enzim yang digunakanterlebih dahulu dinonaktifkan dengan caramemanaskan air mendidih selama 20 menittanpa penambahan substrat Pati 1% atauCMC1% sedangkan untuk kontrol substrat, 1 larutan
Pati atau CMC yang telah dipreinkubasian padasuhu 40oC selama 5 menit, kemudiandiinkubasi kembali pada suhu ruangan selama30 menit, kemudian perlakuan dilanjutkantetapi tanpa penambahan filtrate enzim.Aktivitas selulase (unit/ml) dapat dihitungDimana =
AE = aktivitas enzim (Unit/ml)MG = Berat glukosaBM = berat molekul glukosa (180)t = lama inkubasi
satu unit dari aktivitas enzim dinyatakansebagai jumlah yang dibutuhkan untukmenghasilkan glukosa sebanyak 1 µgmol/mlsubstrat PatiatauCMC per menit denganperlakuan inkubasi 30 menit selama 40oC.Pembuatan Kurva Standar GlukosaKurva standar glukosa dibuat denganmemvariasikan konsentrasi glukosa menjadi 0,20, 40, 60, 80, dan 100 µg/ml. Masing-masingperlakuan di atas diambil 1 ml danditambahkan 1 ml reagen Somogy-Nelson danreagen Arsenomolibdat, kemudian dicukupkanvolumenya menjadi 10 ml dengan penambahanair suling. Lalu diukur absorbannya padapanjang gelombang 540 nm. Kemudian dibuatkurva standarnyaParameter yang diukur meliputi :1. Lama pertumbuhan miselium jamur merang
pada Media Bibit Produksi. Yang diamatiadalah lama (hari) yang dibutuhkan olehmiselium jamur merang untuk mencapai dasarbotol.
2. Aktifitas Amilase dan Selulose pada MediaBibit Produksi dan hubungan AktifitasAmilase dan Selulase dengan kecepatanpertumbuhan miselium jamur merang.
HASIL DAN PEMBAHASAN
Pengamatan kecepatan pertumbuhan miseliumjamur merang pada media bibit produksisetelah dianalisis dg statistik dapat dilihat padaTabel 1 berikut.
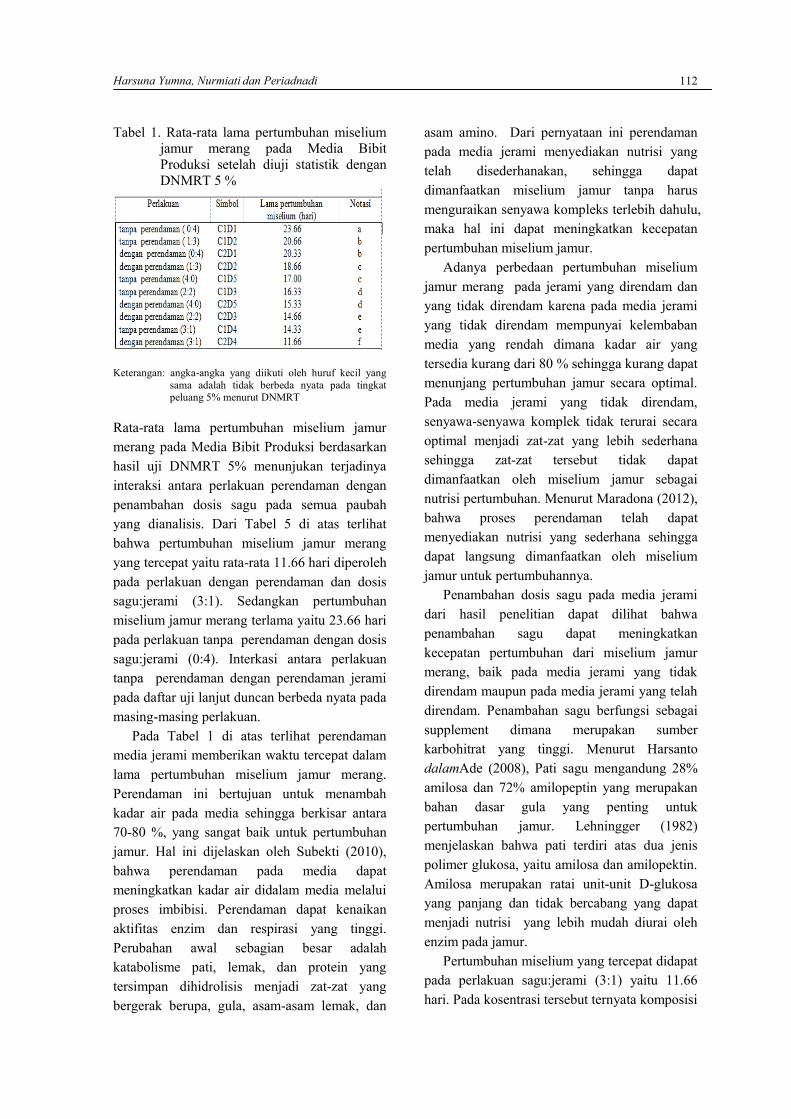
Harsuna Yumna, Nurmiati dan Periadnadi 112
Tabel 1. Rata-rata lama pertumbuhan miseliumjamur merang pada Media BibitProduksi setelah diuji statistik denganDNMRT 5 %
Keterangan: angka-angka yang diikuti oleh huruf kecil yangsama adalah tidak berbeda nyata pada tingkatpeluang 5% menurut DNMRT
Rata-rata lama pertumbuhan miselium jamurmerang pada Media Bibit Produksi berdasarkanhasil uji DNMRT 5% menunjukan terjadinyainteraksi antara perlakuan perendaman denganpenambahan dosis sagu pada semua paubahyang dianalisis. Dari Tabel 5 di atas terlihatbahwa pertumbuhan miselium jamur merangyang tercepat yaitu rata-rata 11.66 hari diperolehpada perlakuan dengan perendaman dan dosissagu:jerami (3:1). Sedangkan pertumbuhanmiselium jamur merang terlama yaitu 23.66 haripada perlakuan tanpa perendaman dengan dosissagu:jerami (0:4). Interkasi antara perlakuantanpa perendaman dengan perendaman jeramipada daftar uji lanjut duncan berbeda nyata padamasing-masing perlakuan.
Pada Tabel 1 di atas terlihat perendamanmedia jerami memberikan waktu tercepat dalamlama pertumbuhan miselium jamur merang.Perendaman ini bertujuan untuk menambahkadar air pada media sehingga berkisar antara70-80 %, yang sangat baik untuk pertumbuhanjamur. Hal ini dijelaskan oleh Subekti (2010),bahwa perendaman pada media dapatmeningkatkan kadar air didalam media melaluiproses imbibisi. Perendaman dapat kenaikanaktifitas enzim dan respirasi yang tinggi.Perubahan awal sebagian besar adalahkatabolisme pati, lemak, dan protein yangtersimpan dihidrolisis menjadi zat-zat yangbergerak berupa, gula, asam-asam lemak, dan
asam amino. Dari pernyataan ini perendamanpada media jerami menyediakan nutrisi yangtelah disederhanakan, sehingga dapatdimanfaatkan miselium jamur tanpa harusmenguraikan senyawa kompleks terlebih dahulu,maka hal ini dapat meningkatkan kecepatanpertumbuhan miselium jamur.
Adanya perbedaan pertumbuhan miseliumjamur merang pada jerami yang direndam danyang tidak direndam karena pada media jeramiyang tidak direndam mempunyai kelembabanmedia yang rendah dimana kadar air yangtersedia kurang dari 80 % sehingga kurang dapatmenunjang pertumbuhan jamur secara optimal.Pada media jerami yang tidak direndam,senyawa-senyawa komplek tidak terurai secaraoptimal menjadi zat-zat yang lebih sederhanasehingga zat-zat tersebut tidak dapatdimanfaatkan oleh miselium jamur sebagainutrisi pertumbuhan. Menurut Maradona (2012),bahwa proses perendaman telah dapatmenyediakan nutrisi yang sederhana sehinggadapat langsung dimanfaatkan oleh miseliumjamur untuk pertumbuhannya.
Penambahan dosis sagu pada media jeramidari hasil penelitian dapat dilihat bahwapenambahan sagu dapat meningkatkankecepatan pertumbuhan dari miselium jamurmerang, baik pada media jerami yang tidakdirendam maupun pada media jerami yang telahdirendam. Penambahan sagu berfungsi sebagaisupplement dimana merupakan sumberkarbohitrat yang tinggi. Menurut HarsantodalamAde (2008), Pati sagu mengandung 28%amilosa dan 72% amilopeptin yang merupakanbahan dasar gula yang penting untukpertumbuhan jamur. Lehningger (1982)menjelaskan bahwa pati terdiri atas dua jenispolimer glukosa, yaitu amilosa dan amilopektin.Amilosa merupakan ratai unit-unit D-glukosayang panjang dan tidak bercabang yang dapatmenjadi nutrisi yang lebih mudah diurai olehenzim pada jamur.
Pertumbuhan miselium yang tercepat didapatpada perlakuan sagu:jerami (3:1) yaitu 11.66hari. Pada kosentrasi tersebut ternyata komposisi

Harsuna Yumna, Nurmiati dan Periadnadi 113
media bibit paling optimal untuk pertumbuhanjamur. Dimana kombinasi antara sagu yangmengandung kadar amilosa dan amilopeptintinggi dan jerami yang mengandung sellulosadan hemiselulosa telah dapat menunjangpertumbuhan jamur. Penambahan nutrisi dan zatmakanan penting seperti pati dan protein sertakondisi yang baik untuk pertumbuhan awalmiselium jamur merang. Pendapat ini didukungoleh pendapat Sinaga (2005) bahwa jamurmerang merupakan organisme heterotropikdimana untuk keperluan hidupnya tergantungsumber nutrisi. Sumber nutrisi penting untukpertumbuhan jamur khususnya perkembanganmiselium karena nutrisi diperoleh dari media.Menurut Gunawan (2010) bahwa senyawa ataunutrisi tersebut berupa selullosa, glukosa, lignin,protein dan senyawa pati.
Penambahan sumber energi dan sumber Ndari media tumbuh dan sagu dapat mempercepatpertumbuhan jamur yang merata dan kompakkarena penggunaan zat-zat makanan dapatmaksimal. Hal ini sesuai pendapat Chang andHayes dalam Trimurti (2009) bahwa sumber Cdan N berfungsi mensintesis sel-sel tumbuhjamur sehingga miselium akan cepat tumbuhtanpa pelepasan N. Nilai C dan N menurunkarena sejumlah amoniak dan senyawa Nlainnya mudah menguap dan hilang. Chang andMiles (2004) menyatakan senyawa komplekyang terdapat dalam media akan diuraikanmenjadi senyawa yang lebih sederhana selamapengomposan. Selulosa dan hemiselulosamerupakan sumber carbon utama yang dapatdigunakan untuk pertumbuhan miselium jamurmerang.
Pengunaan media sagu secara keseluruhansebagai media bibit ternyata menurunkanpembentukan miselium jamur merang. Karenadidalam media sagu hanya terdapat kadaramilosa dan amilopeptin tinggi tampa dibarengidenganadanya selullosa dan hemisellulosa darijerami sehingga kurang baik untuk pertumbuhanjamur. Dari hasil penelitian kombinasi dosissagu:jerami dari kosentrasi (0:4), (1:3),(2:2)
ternyata juga menurunkan pembentukanmiselium jamur merang.Nilai Aktifitas Amilase pada Beberapa MediaBibit Produksi jamur merangDari Gambar 1 terlihat bahwa aktifitas Amilasejamur merang yang tertinggi yaitu 0.1088 unit/gdiperoleh pada perlakuan dengan perendamandan dosis sagu:jerami (3:1). Sedangkan aktifitasAmilase jamur merang terendah yaitu 0.0874unit/g pada perlakuan tanpa perendamandengan dosis sagu:jerami (0:4). Interkasi antaraperlakuan tanpa perendaman denganperendaman jerami ternyata masing-masingperlakuan memperlihatkan perbedaan.
Aktifitas Amilase tertinggi pada dosissagu:jerami (1:3), hal ini disebabkan olehsejumlah pati sagu yang terdapat dalamcampuran media. Sagu merupakan golongan patiyang mengandung karbohitrat yang terdapatpada bagian batang sebagai cadangan makanan.Pati sagu merupakan butiran atau granul yangbewarna putih mengkilat yang tidak berbau danberasa serta mengandung kadar amilosa danamilopektin yang tinggi yang merupakan faktorpenting untuk pertumbuhan jamur. Menurut Ade(2009), media empelur sagu terdiri dari pati danserat tentu dimana enzim yang berperan dalamperombakan substrat tersebut adalah enzim-enzim dari amilase, disamping enzim-enzimlainnya. Leatham (1989), menyatakan Enzimamilase berperan dalam perombakan pati yangterkandung dalam media. Dan selamapertumbuhan miselium jamur merang meng-gunakan sumber dari carbon seperti gula bebas,pati dan hemiselulosa.
Gambar 1. Aktifitas Amilase (u/g) jamurmerang

Harsuna Yumna, Nurmiati dan Periadnadi 114
Hubungan Aktifitas Amilase denganKecepatan Pertumbuhan Miselium.Pada Gambar 2 dapat dilihat bahwa setiapperlakuan baik tanpa perendaman ataupundengan perendaman jerami, Aktifitas Amilasesangat berpengaruh terhadap pertumbuhanmiselium jamur merang. Berdasarkanpersamaan garis y =-1436,x_106,1 (tanpaperendaman) dan y=-1101,x +85,08 (denganperendaman) dapat dilihat bahwa kecepatanpertumbuhan miselium jamur merangberkorelasi negatif dengan aktifitas Amilase.Semakin tinggi aktifitas enzim maka semakincepat juga pertumbuhan miselium jamur merang.Namun disini terlihat perlakuan denganperendaman memberikan nilai aktifitas enzimtertinggi. Peristiwa perendaman telah mem-berikan kadar air yang cukup pada mediasehingga dapat menyediakan nutrisi dan sumberenergi yang telah disederhanakan. Sumberenergi ini berpengaruh pada pertumbuhanmiselium jamur merang dan aktifitas enzimamilase. Menurut Ade (2009), banyaknyasumber energi yang dapat diserap oleh sel darisubstrat menyebabkan pertumbuhan jamur terusmeningkat sehingga memungkinkan hifa jamurlebih banyak mengeluarkan enzim-enzimekstraseluler seperti amilase dalam meng-hasilkan gula.
Sagu yang terdapat dalam media bibitproduksi jamur merang mengandung pati danselulosa berupa serat kasar. Jamur merang dapatmengkonversi empelur sagu terlebih dahuludengan merombak amilosa sehingga didapatgula sebelum merombak selulase. Adanya gulaterbentuk dapat digunakan oleh miselium jamurmerang dalam pertumbuhannya. Menurut Ade(2009), yang berperan dalam perombakan patisagu adalah enzim amilase disamping enzim-enzim lainnya. Sedangkan menurut Winarno(1983), terjadinya proses perombakan ataureaksi biokimia dalam media dikarenakanadanya enzim yang terkandung dalam mediaseperti amilase.
Gambar 2. Hubungan perbandingan AktifitasAmilase dengan kecepatanpertumbuhan miselium. Kiri:Tanpa perendaman Kanan: Denganperendaman
KESIMPULAN
Berdasarkan hasil penelitian yang telahdilakukan tentang Studi Komparatif Sagu(Metroxylon Rottb) sebagai Media BibitProduksi Terhadap Lama PertumbuhanMiselium dan Aktifitas Amilase dan SelulaseJamur Merang (Volvariella volvacea(Bull).Sing.) dapat disimpulkan bahwa:1. Pertumbuhan miselium jamur merang pada
Media BibitProduksi yang terbaik didapatpada dosis sagu:jerami (3:1) (direndam)dengan lama pertumbuhan miselium yaitu11.66 hari setelah inokulasi.
2. Nilai Aktifitas Amilase tertinggi (0.1088unit/g) dan nilai Aktifitas Selulase tertinggi(0.0657 unit/g) didapat pada media jeramiyang direndam dengan dosis sagu:jerami(3:1)
DAFTAR PUSTAKA
Ade, F.Y. 2008. Potensi Jamur-JamurPendegradasi Sagu (Metroxylon saguRottb). Skripsi Jurusan Biologi. FMIPA.Universitas Andalas. Padang.
Ambriyanto, K. S., 2010, Isolasi danKarakterisasi Bakteri Aerob PendegradasiSelulosa dari Serasah Daun Rumput Gajah(Pennisetum purpureum schaum), InstitutTeknologi Sepuluh Nopember (Skripsi).
Astuti, Y., D.N.A. Makalew. 2000. EfektifitasPengomposan Limbah Pertanian Sebagai

Harsuna Yumna, Nurmiati dan Periadnadi 115
Media Tanam Jamur merang (Volvariellavolvacea). Jakarta. Universitas Mercubuana.Tajuk Edisi Istimewa. Tahun VI.
Chang, S and P.G Miles, 2004. Mushrooms:Cultivation, Nutritional Value, MedicinalEffect and Environmental. New York:CRC Press.
Chen, C.H., J.Y. Wu, Chen C.H, W.H. Chang,K.T. Chung, Y.W. Liu, F.J. Lu and C.H.Chen. 2010. Anti-Cancer Effects of ProteinExtracts from Calvatiali lacina, Pleurotusostreatus and Volvariell avolvacea.Evidence-Based Complementary andAlternative Medizine. Vo. 2011, Artikel ID982368, 10 pp.
Dewi. 2002. Hidrolisis Limbah Hasil PertanianSecara Enzimatik, AktaAgrosia, Vol. 5, No.2,hal. 67 – 71.
Gunawan,A.W. 2010. Usaha Pembibitan Jamur.Penebar Swadaya: Jakarta.
Lechninger, A.L. 1993. Dasar-dasar Biokimia.GadjahMada University. Yogyakarta
Leatham, G.F., T.J. Leonard, 1989. Biologi andphysiology of Shitake MushroomCultivation.In: Shitake Mushroom. TheProceedings of National Symposium andtrade show.
Parjimo dan Andoko,A. 2008. Budidaya Jamur.PT. Agromedia Pustaka: Jakarta
Patmasari, U., Suharni, T.T., Permana, D.J,2007. Pengaruh Penambahan Zeolitterhadap Viabilitas Bibit Jamur Merang.Lembaga Ilmu Pengetahuan Indonesia(LIPI), Cibinong-Bogor 16911.
Sinaga, M. 2008. Jamur Merang danBudidayanya. Penebar Swadaya. Jakarta.
Suriawiria, H, 2007. Bioteknologi Penjamuran.Bandung. Angkasa.

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman serangga pada dua habitat berbeda di kawasanCilintang, Taman Nasional Ujung Kulon, BantenHASNI RUSLAN, PRIMA LADY DAN HILDA SILFIA
Fakultas Biologi Universitas Nasional, JakartaE-mail: [email protected]
ABSTRAKTaman Nasional Ujung Kulon memiliki kekayaan alamhayati yang tinggi, ditinjau dari keberadaan flora dan fauna yangterdapat di taman nasional, salahsatu fauna yang adalah serangga. Serangga merupakan fauna yang jumlahnya banyak dimuka bumi ini, mempunyai perananyang sangat penting di suatu ekosistem baik secara langsung maupun tidak langsung.Peranan serangga di dalam ekosistem diantaranya adalah sebagai pollinator, dekomposer, pemangsa, parasitoid danbioindikator bagi suatu ekosistem.Tujuan daripada penelitian ini untuk mengetahui keanekaragaman berbagai jenisserangga yang terdapat di Taman Nasional Ujung Kulon. Penelitian dilakukan pada tanggal 25 April – 1 Mei 2014, dikawasan Cilintang, Desa Taman Jaya, Kabupaten Pandeglang, Ujung Kulon. Penelitian serangga dilakukan denganmetode purposive sampling yang berlangsung selama 4 hari untuk 2 habitat. Pengamatandilakukan di dua habitat yangberbeda yaitu habitat hutandan habitat padangrumput. Di Taman Nasional Ujung Kulon ditemukan 10 ordo, 34 famili,127 jenis dengan total individu 725. Indeks keanekaragaman serangga di Taman Nasional Ujung Kulon tergolong tinggi(H’= 4,45). Berdasarkan uji hutchinson terhadap perbanding indeks keanekaragaman di habitat padang rumput dan hutan,tidak terdapat perbedaan yang bermakna. Kelimpahan relatif dan frekuensi relatif serangga yang tertinggi didapatkanpada Junonia atlites, Papilio peranthus dan Semuthitam (Formicidae). Faktor lingkungan, suhu dan kelembaban yangdidapat sesuai dengan kehidupan serangga.
Key words: Keanekaragaman, Serangga , Taman Nasional Ujung Kulon
Pendahuluan
Taman Nasional Ujung Kulon memilikikekayaan alam hayati yang tinggi, ditinjau darikeberadaan flora dan fauna yang terdapat ditaman nasional, salah satu fauna yang adaadalah serangga. Serangga memiliki perananyang sangat penting dalam ekosistem.Tanpaserangga, kehidupan ekosistem terganggu dantidak mencapai keseimbangan. Peranan dariserangga diantaranya: sebagai penyerbuktanaman, pengurai bahan organik, sebagai salahsatu rantai makanan dalam ekosistem(pemangsa), sebagai pakan dan ada juga perandari serangga ini yang bersifat merugikan :sebagai hama pada tanaman, sebagai perusakbahan bangunan, sebagai vektor penyakit, dansebagai penganggu (nyamuk).
Serangga merupakan fauna yang jumlahnyamelimpah atau dominan di muka bumi ini(Borroret al, 1992). Dominansi dari serangga inidikarenakan ukurannya yang kecilsehinggadapat menempati suatu habitat dengan jumlah
yang banyak, kemampuan reproduksi yang cepat,kemampuan adaptasi tinggi yaitu dengan adanyakutikula berlapis lilin sebagai pelindung danpakan melimpah.
Serangga termasuk dalam FilumArthropoda,yang bertungkai enam (tiga pasang) dan dikenaldengan heksapoda. Tubuh serangga terdiri atastiga bagian : kepala, toraks dan abdomen. Padakepala terdapat mata majemuk dan oseli,sepasang antena, satu pasang mandibula, dansatu pasang maksila beserta modifikasi-modifikasi, sehingga terdapat ber-macam-macam tipe mulut serangga : tipe mulutpengunyah, pengisap – penjilat, pengisap,pengisap - penusuk. Toraks serangga terdiri daritiga segmen, masing-masing terdapat sepasangkaki. Berbeda dengan hewan Arthropodalainnya, pada toraks serangga terdapat duapasang sayap, masing-masing di segmen torakskedua dan ketiga. Adanya sayap memungkinkankelompok serangga dapat terbang dan berpindahke tempat yang jauh. Abdomen pada dasarnyaterdiri dari 12 ruas, tetapi pada beberapa

Hasni Ruslan, Prima Lady dan Hilda Silfia 117
serangga hanya mempunyai 6-8 ruas karena adaruas-ruas abdomen yang mereduksi (Triplehornand Johnson 2005).
Kawasan Taman nasional Ujung Kulonsecara administratif terletak di KecamatanSumur dan Cimanggu, Kabupaten Pandeglang,Propinsi Banten. Secara geografis TamanNasional Ujung Kulon terletak antara 102º02’32”- 105º37’37” BT dan 06º30’43” - 06º52’17” LS.Berdasarkan SK Menteri Kehutanan No.284/Kpts-II/1992 tentang Perubahan FungsiCagar Alam Gunung Honje, Cagar Alam PulauPanaitan, Cagar Alam Pulau Peucang, dan Cagaralam Ujung Kulon seluas 78.619 Ha danPenunjukan perairan laut di sekitarnya seluas44.337 Ha yang terletak di KabupatenPandeglang, Jawa Barat menjadi TamanNasional dengan nama TN Ujung Kulon makaluas kawasan TN Ujung Kulon adalah 122.956Ha (www.ujungkulon.org).
Dokumentasi daftar spesies kupu-kupu di TNUjung Kulon telah dilakukan oleh Peggie, 2012,dan Keanekaragaman kupu-kupu SuperfamiliPapilionoidea di kampung Paniis, Desa TamanJaya, Sekitar TN Ujung Kulon telah dilakukanoleh Ruslan dkk (2012). Tujuan penelitian iniadalah untuk memperkaya dan mengetahuikeanekaragaman berbagai jenis serangga yangterdapat di Taman Nasional Ujung Kulon.Hasilpenelitian ini diharapkan bermanfaat sebagaisalah satu tambahan informasi bagi duniapendidikan, berguna bagi pengelolaan TamanNasional dan konservasi keanekaragaman hayati.
BAHAN DAN METODE
Penelitian dilakukan pada tanggal 25 April – 1Mei 2014, di kawasan Cilintang, Desa TamanJaya, Kabupaten Pandeglang, Ujung Kulon.Pengamatan dilakukan di dua habitat yangberbeda yaitu habitat hutan dan habitat padangrumput. Masing-masing habitat terdiri dari duaplot dan memiliki empat titik GPS sehinggadapat membentuk suatu plot (Gambar 1 danGambar 2).
Gambar 1.Peta lokasi penelitian
Gambar 2. Lokasi penelitian, (a) lokasi padangrumput (plot 1), (b) lokasi padangrumput (plot 2), (c) lokasi hutan (plot 3)dan (d) lokasi hutan (plot 4)(Dokumentasi pribadi, 2014).

Hasni Ruslan, Prima Lady dan Hilda Silfia 118
Pengamatan Keanekaragaman SeranggaPengamatan serangga dilakukan dengan metodepurposive sampling, 4 plot dibuat masing-masing dengan ukuran 200 m x 200 m, 2 plotterdapat di habitat padang rumput dan 2 plot dihabitat hutan. Pengamatan diawali denganmengukur titik koordinat dengan menggunakanGPS pada setiap plot. Pengamatan dilakukanpada pagi hari (08.00-12.00 WIB). Pada saatpengamatan, dilakukan juga pengukuranparameter lingkungan, yaitu suhu dankelembaban di setiap tipe habitat yang diamati.Serangga yang didapat di simpan dalam kertaspapilot dan plastik untuk keperluan identifikasidi laboratorium.Identifikasi SeranggaSpesimen yang telah diambil dari lapangandiidentifikasi menggunakan buku identifikasiBorror et al 1992, Triplehorn 2005, Peggie &Amir 2006, D’abrera, 2005 dan Neo, 2001.Setelah diidentifikasi, serangga di awetkan dandisimpan dalam kotak serangga.Analisis DataIndeks Keanekaragaman jenis seranggaIndeks keanekaragaman jenis serangga dihitungdengan menggunakan indeks keanekaragamanShannon-Wiener (H’) dengan rumus berikut :
Keterangan:H’ = Indeks Keanekaragaman Shannon-WienerPi = Proporsi kelimpahan jenisni = Jumlah individu ke-iN = Jumlah total individuKriteria nilai indeks keanekaragaman jenis berdasarkanShannon-Wiener adalah sebagai berikut :Nilai H ≤ 1,5 : Keanekaragaman rendahNilai H >1,5 – 3,5 : Keanekaragaman sedangNilai H > 3,5 : Keanekaragaman tinggi
Untuk membedakan nilai indekskeanekaragaman pada kedua hutan digunakanuji Hutchinson yang dilengkapi dengan uji t :
Var H’ =
Keterangan :Var = Varians yaitu perbedaan keanekaragaman jenis
antar hutanS = Jumlah spesies satu hutan
Uji ini menggunakan uji “t” dengan peluang95% (=0.05). Rumus-rumus yang digunakanberdasarkan Magurran (1988) adalah :
t =
df =
Hipotesis :t hit < t tabel, tolak Ho (terdapat perbedaan yangbermakna)t hit > t tabel, terima Ho (tidak terdapat perbedaanbermakna)Indeks Kemerataan Spesies
Keterangan :H’ = Indeks Keanekaragaman Shannon-WienerS = Jumlah spesies yang ditemukan (kekayaan jenis)
Indeks kesamaan jenis antar habitat (IndeksSorensen)Indeks kesamaan jenis antar habitat dihitunguntuk mengetahui kesamaan komunitas padadua tipe habitat yang dihitung berdasarkan jenisyang ditemukan.Indeks yang digunakan adalahIndeks Sorensen (IS). Adapun rumus IndeksSorensen (IS) adalah sebagai berikut :(Magguran 1988).
Keterangan :a = Jumlah jenis pada tipe habitat Ab = Jumlah jenis pada tipe habitat Bj = Jumlah jenis yang ditemukan pada kedua tipehabitat tersebut
4. Kelimpahan, Frekuensi dan Indeks NilaiPenting(INP)( Fachrul, 2012):Nilai kelimpahan relatif (KR) ditetapkanmenggunakan rumus,
100%Nilai frekuensi Relatif (FR) ditetapkanmenggunakan rumus,Frekuensi individu suatu jenisJumlah frekuensi seluruh jenis 100%
H’ = -∑ pi ln pi dengan pi =
E =
IS = x 100%
Hasni Ruslan, Prima Lady dan Hilda Silfia 118
Pengamatan Keanekaragaman SeranggaPengamatan serangga dilakukan dengan metodepurposive sampling, 4 plot dibuat masing-masing dengan ukuran 200 m x 200 m, 2 plotterdapat di habitat padang rumput dan 2 plot dihabitat hutan. Pengamatan diawali denganmengukur titik koordinat dengan menggunakanGPS pada setiap plot. Pengamatan dilakukanpada pagi hari (08.00-12.00 WIB). Pada saatpengamatan, dilakukan juga pengukuranparameter lingkungan, yaitu suhu dankelembaban di setiap tipe habitat yang diamati.Serangga yang didapat di simpan dalam kertaspapilot dan plastik untuk keperluan identifikasidi laboratorium.Identifikasi SeranggaSpesimen yang telah diambil dari lapangandiidentifikasi menggunakan buku identifikasiBorror et al 1992, Triplehorn 2005, Peggie &Amir 2006, D’abrera, 2005 dan Neo, 2001.Setelah diidentifikasi, serangga di awetkan dandisimpan dalam kotak serangga.Analisis DataIndeks Keanekaragaman jenis seranggaIndeks keanekaragaman jenis serangga dihitungdengan menggunakan indeks keanekaragamanShannon-Wiener (H’) dengan rumus berikut :
Keterangan:H’ = Indeks Keanekaragaman Shannon-WienerPi = Proporsi kelimpahan jenisni = Jumlah individu ke-iN = Jumlah total individuKriteria nilai indeks keanekaragaman jenis berdasarkanShannon-Wiener adalah sebagai berikut :Nilai H ≤ 1,5 : Keanekaragaman rendahNilai H >1,5 – 3,5 : Keanekaragaman sedangNilai H > 3,5 : Keanekaragaman tinggi
Untuk membedakan nilai indekskeanekaragaman pada kedua hutan digunakanuji Hutchinson yang dilengkapi dengan uji t :
Var H’ =
Keterangan :Var = Varians yaitu perbedaan keanekaragaman jenis
antar hutanS = Jumlah spesies satu hutan
Uji ini menggunakan uji “t” dengan peluang95% (=0.05). Rumus-rumus yang digunakanberdasarkan Magurran (1988) adalah :
t =
df =
Hipotesis :t hit < t tabel, tolak Ho (terdapat perbedaan yangbermakna)t hit > t tabel, terima Ho (tidak terdapat perbedaanbermakna)Indeks Kemerataan Spesies
Keterangan :H’ = Indeks Keanekaragaman Shannon-WienerS = Jumlah spesies yang ditemukan (kekayaan jenis)
Indeks kesamaan jenis antar habitat (IndeksSorensen)Indeks kesamaan jenis antar habitat dihitunguntuk mengetahui kesamaan komunitas padadua tipe habitat yang dihitung berdasarkan jenisyang ditemukan.Indeks yang digunakan adalahIndeks Sorensen (IS). Adapun rumus IndeksSorensen (IS) adalah sebagai berikut :(Magguran 1988).
Keterangan :a = Jumlah jenis pada tipe habitat Ab = Jumlah jenis pada tipe habitat Bj = Jumlah jenis yang ditemukan pada kedua tipehabitat tersebut
4. Kelimpahan, Frekuensi dan Indeks NilaiPenting(INP)( Fachrul, 2012):Nilai kelimpahan relatif (KR) ditetapkanmenggunakan rumus,
100%Nilai frekuensi Relatif (FR) ditetapkanmenggunakan rumus,Frekuensi individu suatu jenisJumlah frekuensi seluruh jenis 100%
H’ = -∑ pi ln pi dengan pi =
E =
IS = x 100%
Hasni Ruslan, Prima Lady dan Hilda Silfia 118
Pengamatan Keanekaragaman SeranggaPengamatan serangga dilakukan dengan metodepurposive sampling, 4 plot dibuat masing-masing dengan ukuran 200 m x 200 m, 2 plotterdapat di habitat padang rumput dan 2 plot dihabitat hutan. Pengamatan diawali denganmengukur titik koordinat dengan menggunakanGPS pada setiap plot. Pengamatan dilakukanpada pagi hari (08.00-12.00 WIB). Pada saatpengamatan, dilakukan juga pengukuranparameter lingkungan, yaitu suhu dankelembaban di setiap tipe habitat yang diamati.Serangga yang didapat di simpan dalam kertaspapilot dan plastik untuk keperluan identifikasidi laboratorium.Identifikasi SeranggaSpesimen yang telah diambil dari lapangandiidentifikasi menggunakan buku identifikasiBorror et al 1992, Triplehorn 2005, Peggie &Amir 2006, D’abrera, 2005 dan Neo, 2001.Setelah diidentifikasi, serangga di awetkan dandisimpan dalam kotak serangga.Analisis DataIndeks Keanekaragaman jenis seranggaIndeks keanekaragaman jenis serangga dihitungdengan menggunakan indeks keanekaragamanShannon-Wiener (H’) dengan rumus berikut :
Keterangan:H’ = Indeks Keanekaragaman Shannon-WienerPi = Proporsi kelimpahan jenisni = Jumlah individu ke-iN = Jumlah total individuKriteria nilai indeks keanekaragaman jenis berdasarkanShannon-Wiener adalah sebagai berikut :Nilai H ≤ 1,5 : Keanekaragaman rendahNilai H >1,5 – 3,5 : Keanekaragaman sedangNilai H > 3,5 : Keanekaragaman tinggi
Untuk membedakan nilai indekskeanekaragaman pada kedua hutan digunakanuji Hutchinson yang dilengkapi dengan uji t :
Var H’ =
Keterangan :Var = Varians yaitu perbedaan keanekaragaman jenis
antar hutanS = Jumlah spesies satu hutan
Uji ini menggunakan uji “t” dengan peluang95% (=0.05). Rumus-rumus yang digunakanberdasarkan Magurran (1988) adalah :
t =
df =
Hipotesis :t hit < t tabel, tolak Ho (terdapat perbedaan yangbermakna)t hit > t tabel, terima Ho (tidak terdapat perbedaanbermakna)Indeks Kemerataan Spesies
Keterangan :H’ = Indeks Keanekaragaman Shannon-WienerS = Jumlah spesies yang ditemukan (kekayaan jenis)
Indeks kesamaan jenis antar habitat (IndeksSorensen)Indeks kesamaan jenis antar habitat dihitunguntuk mengetahui kesamaan komunitas padadua tipe habitat yang dihitung berdasarkan jenisyang ditemukan.Indeks yang digunakan adalahIndeks Sorensen (IS). Adapun rumus IndeksSorensen (IS) adalah sebagai berikut :(Magguran 1988).
Keterangan :a = Jumlah jenis pada tipe habitat Ab = Jumlah jenis pada tipe habitat Bj = Jumlah jenis yang ditemukan pada kedua tipehabitat tersebut
4. Kelimpahan, Frekuensi dan Indeks NilaiPenting(INP)( Fachrul, 2012):Nilai kelimpahan relatif (KR) ditetapkanmenggunakan rumus,
100%Nilai frekuensi Relatif (FR) ditetapkanmenggunakan rumus,Frekuensi individu suatu jenisJumlah frekuensi seluruh jenis 100%
H’ = -∑ pi ln pi dengan pi =
E =
IS = x 100%
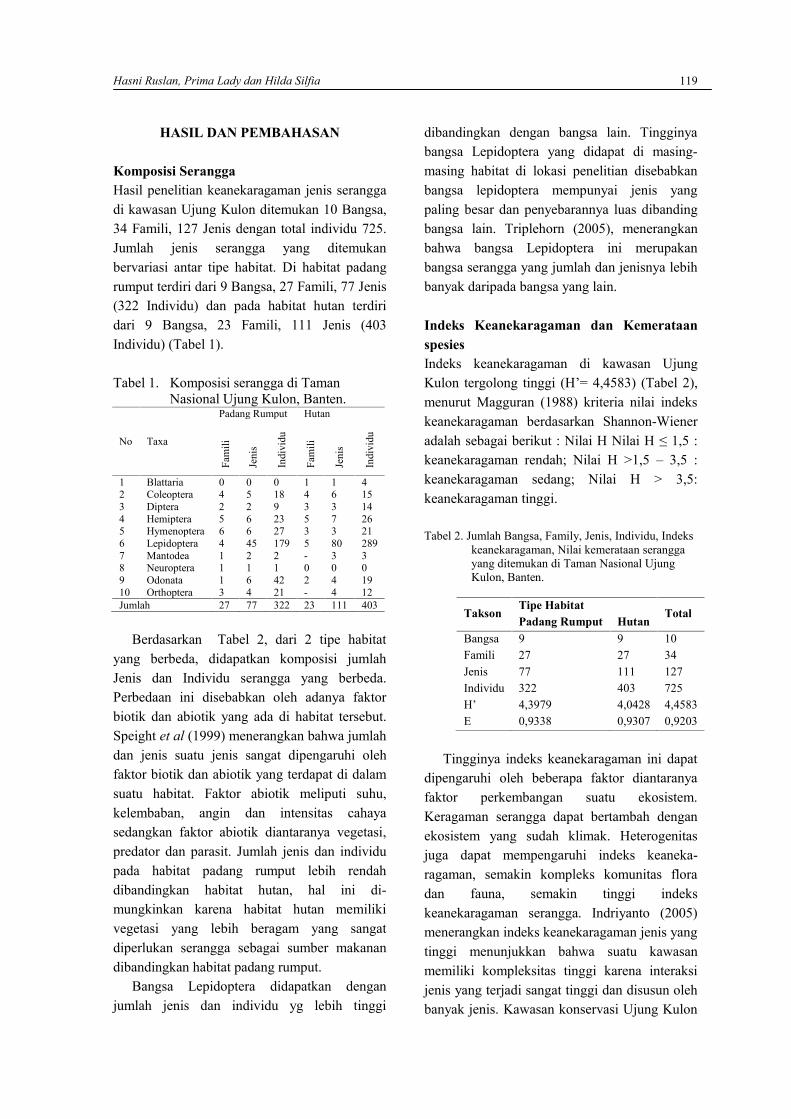
Hasni Ruslan, Prima Lady dan Hilda Silfia 119
HASIL DAN PEMBAHASAN
Komposisi SeranggaHasil penelitian keanekaragaman jenis seranggadi kawasan Ujung Kulon ditemukan 10 Bangsa,34 Famili, 127 Jenis dengan total individu 725.Jumlah jenis serangga yang ditemukanbervariasi antar tipe habitat. Di habitat padangrumput terdiri dari 9 Bangsa, 27 Famili, 77 Jenis(322 Individu) dan pada habitat hutan terdiridari 9 Bangsa, 23 Famili, 111 Jenis (403Individu) (Tabel 1).
Tabel 1. Komposisi serangga di TamanNasional Ujung Kulon, Banten.
No Taxa
Padang Rumput Hutan
Fam
ili
Jeni
s
Indi
vidu
Fam
ili
Jeni
s
Indi
vidu
12345678910
BlattariaColeopteraDipteraHemipteraHymenopteraLepidopteraMantodeaNeuropteraOdonataOrthoptera
0425641113
05266452164
01892327179214221
143535-02-
16373803044
415142621289301912
Jumlah 27 77 322 23 111 403
Berdasarkan Tabel 2, dari 2 tipe habitatyang berbeda, didapatkan komposisi jumlahJenis dan Individu serangga yang berbeda.Perbedaan ini disebabkan oleh adanya faktorbiotik dan abiotik yang ada di habitat tersebut.Speight et al (1999) menerangkan bahwa jumlahdan jenis suatu jenis sangat dipengaruhi olehfaktor biotik dan abiotik yang terdapat di dalamsuatu habitat. Faktor abiotik meliputi suhu,kelembaban, angin dan intensitas cahayasedangkan faktor abiotik diantaranya vegetasi,predator dan parasit. Jumlah jenis dan individupada habitat padang rumput lebih rendahdibandingkan habitat hutan, hal ini di-mungkinkan karena habitat hutan memilikivegetasi yang lebih beragam yang sangatdiperlukan serangga sebagai sumber makanandibandingkan habitat padang rumput.
Bangsa Lepidoptera didapatkan denganjumlah jenis dan individu yg lebih tinggi
dibandingkan dengan bangsa lain. Tingginyabangsa Lepidoptera yang didapat di masing-masing habitat di lokasi penelitian disebabkanbangsa lepidoptera mempunyai jenis yangpaling besar dan penyebarannya luas dibandingbangsa lain. Triplehorn (2005), menerangkanbahwa bangsa Lepidoptera ini merupakanbangsa serangga yang jumlah dan jenisnya lebihbanyak daripada bangsa yang lain.
Indeks Keanekaragaman dan KemerataanspesiesIndeks keanekaragaman di kawasan UjungKulon tergolong tinggi (H’= 4,4583) (Tabel 2),menurut Magguran (1988) kriteria nilai indekskeanekaragaman berdasarkan Shannon-Wieneradalah sebagai berikut : Nilai H Nilai H ≤ 1,5 :keanekaragaman rendah; Nilai H >1,5 – 3,5 :keanekaragaman sedang; Nilai H > 3,5:keanekaragaman tinggi.
Tabel 2. Jumlah Bangsa, Family, Jenis, Individu, Indekskeanekaragaman, Nilai kemerataan seranggayang ditemukan di Taman Nasional UjungKulon, Banten.
Tingginya indeks keanekaragaman ini dapatdipengaruhi oleh beberapa faktor diantaranyafaktor perkembangan suatu ekosistem.Keragaman serangga dapat bertambah denganekosistem yang sudah klimak. Heterogenitasjuga dapat mempengaruhi indeks keaneka-ragaman, semakin kompleks komunitas floradan fauna, semakin tinggi indekskeanekaragaman serangga. Indriyanto (2005)menerangkan indeks keanekaragaman jenis yangtinggi menunjukkan bahwa suatu kawasanmemiliki kompleksitas tinggi karena interaksijenis yang terjadi sangat tinggi dan disusun olehbanyak jenis. Kawasan konservasi Ujung Kulon
TaksonTipe Habitat
TotalPadang Rumput Hutan
BangsaFamiliJenisIndividuH’E
927773224,39790,9338
9271114034,04280,9307
10341277254,45830,9203
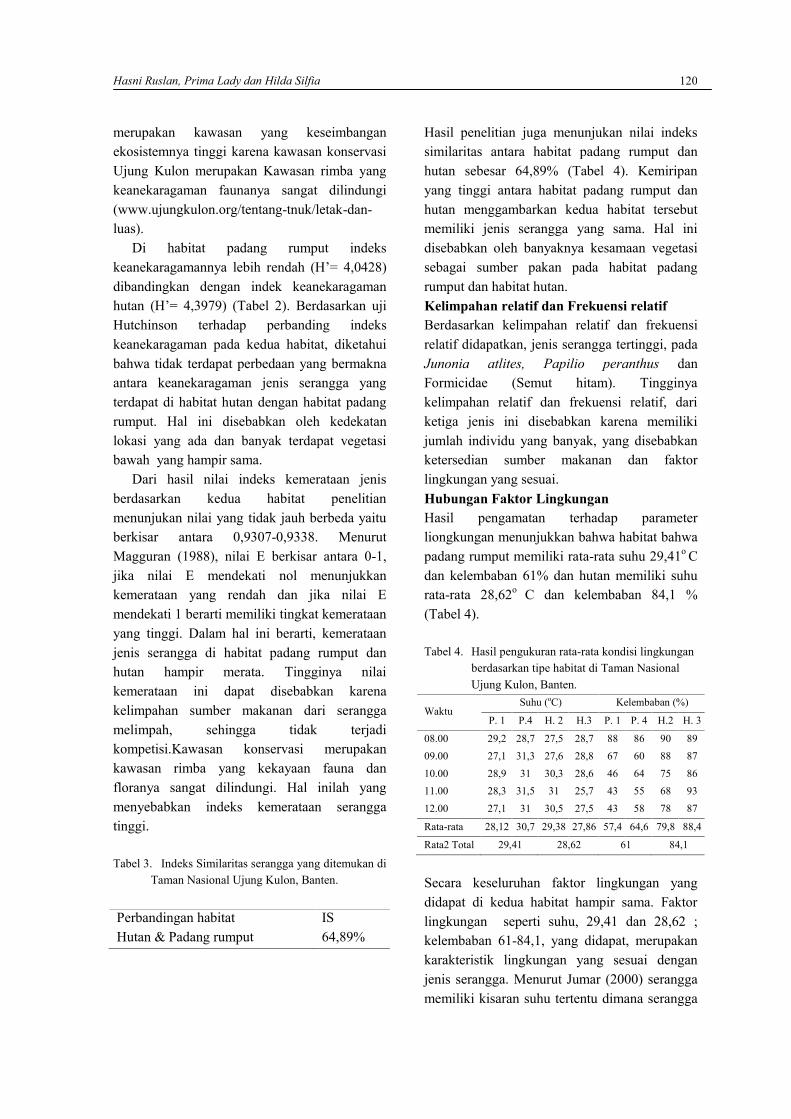
Hasni Ruslan, Prima Lady dan Hilda Silfia 120
merupakan kawasan yang keseimbanganekosistemnya tinggi karena kawasan konservasiUjung Kulon merupakan Kawasan rimba yangkeanekaragaman faunanya sangat dilindungi(www.ujungkulon.org/tentang-tnuk/letak-dan-luas).
Di habitat padang rumput indekskeanekaragamannya lebih rendah (H’= 4,0428)dibandingkan dengan indek keanekaragamanhutan (H’= 4,3979) (Tabel 2). Berdasarkan ujiHutchinson terhadap perbanding indekskeanekaragaman pada kedua habitat, diketahuibahwa tidak terdapat perbedaan yang bermaknaantara keanekaragaman jenis serangga yangterdapat di habitat hutan dengan habitat padangrumput. Hal ini disebabkan oleh kedekatanlokasi yang ada dan banyak terdapat vegetasibawah yang hampir sama.
Dari hasil nilai indeks kemerataan jenisberdasarkan kedua habitat penelitianmenunjukan nilai yang tidak jauh berbeda yaituberkisar antara 0,9307-0,9338. MenurutMagguran (1988), nilai E berkisar antara 0-1,jika nilai E mendekati nol menunjukkankemerataan yang rendah dan jika nilai Emendekati 1 berarti memiliki tingkat kemerataanyang tinggi. Dalam hal ini berarti, kemerataanjenis serangga di habitat padang rumput danhutan hampir merata. Tingginya nilaikemerataan ini dapat disebabkan karenakelimpahan sumber makanan dari seranggamelimpah, sehingga tidak terjadikompetisi.Kawasan konservasi merupakankawasan rimba yang kekayaan fauna danfloranya sangat dilindungi. Hal inilah yangmenyebabkan indeks kemerataan seranggatinggi.
Tabel 3. Indeks Similaritas serangga yang ditemukan diTaman Nasional Ujung Kulon, Banten.
Hasil penelitian juga menunjukan nilai indekssimilaritas antara habitat padang rumput danhutan sebesar 64,89% (Tabel 4). Kemiripanyang tinggi antara habitat padang rumput danhutan menggambarkan kedua habitat tersebutmemiliki jenis serangga yang sama. Hal inidisebabkan oleh banyaknya kesamaan vegetasisebagai sumber pakan pada habitat padangrumput dan habitat hutan.Kelimpahan relatif dan Frekuensi relatifBerdasarkan kelimpahan relatif dan frekuensirelatif didapatkan, jenis serangga tertinggi, padaJunonia atlites, Papilio peranthus danFormicidae (Semut hitam). Tingginyakelimpahan relatif dan frekuensi relatif, dariketiga jenis ini disebabkan karena memilikijumlah individu yang banyak, yang disebabkanketersedian sumber makanan dan faktorlingkungan yang sesuai.Hubungan Faktor LingkunganHasil pengamatan terhadap parameterliongkungan menunjukkan bahwa habitat bahwapadang rumput memiliki rata-rata suhu 29,41o Cdan kelembaban 61% dan hutan memiliki suhurata-rata 28,62o C dan kelembaban 84,1 %(Tabel 4).
Tabel 4. Hasil pengukuran rata-rata kondisi lingkunganberdasarkan tipe habitat di Taman NasionalUjung Kulon, Banten.
WaktuSuhu (oC) Kelembaban (%)
P. 1 P.4 H. 2 H.3 P. 1 P. 4 H.2 H. 3
08.00 29,2 28,7 27,5 28,7 88 86 90 8909.00 27,1 31,3 27,6 28,8 67 60 88 87
10.00 28,9 31 30,3 28,6 46 64 75 8611.00 28,3 31,5 31 25,7 43 55 68 93
12.00 27,1 31 30,5 27,5 43 58 78 87
Rata-rata 28,12 30,7 29,38 27,86 57,4 64,6 79,8 88,4
Rata2 Total 29,41 28,62 61 84,1
Secara keseluruhan faktor lingkungan yangdidapat di kedua habitat hampir sama. Faktorlingkungan seperti suhu, 29,41 dan 28,62 ;kelembaban 61-84,1, yang didapat, merupakankarakteristik lingkungan yang sesuai denganjenis serangga. Menurut Jumar (2000) seranggamemiliki kisaran suhu tertentu dimana serangga
Perbandingan habitat ISHutan & Padang rumput 64,89%

Hasni Ruslan, Prima Lady dan Hilda Silfia 121
dapat hidup. Pada umumnya kisaran suhuserangga 15-45 °C, kelembaban 50-90%. Faktorlingkungan seperti suhu dan kelembaban yangdidapat sesuai dengan kehidupan serangga.
KESIMPULANBerdasarkan penelitian yang telah dilakukandapat disimpulkan beberapa hal antara lainsebagai berikut :Di Taman Nasional Ujung Kulom ditemukan 10ordo, 34 famili, 127 jenis dengan total individu725. Indeks keanekaragaman serangga di TamanNasional Ujung Kulon tergolong tinggi(H=4.45). Berdasarakn uji hutchinson terhadapperbandingan indeks keanekaragaman di habitatpadang rumput dan hutan, tidak terdapatperbedaan yang bermakna. Bangsa Lepidopteramerupakan bangsa yang memiliki jumlah jenisdan individu yang tinggi dari bangsa seranggalain. Kelimpahan dan frekunsi relatif yang tinggididapatkan pada jenis Junonia atlites, Papilioperanthus dan Semut hitam (Formicidae).Faktor lingkungan, suhu dan kelembaban yangdidapat sesuai dengan kehidupan serangga.
DAFTAR PUSTAKA
Borror, D.J., C.A. Triplehorn dan N. F. Johnson.Pengenalan Pelajaran Serangga. Edisikeenam. Soetiono Porto Soejono.Gajahmada university Press.Yogyakarta. 1992.
d’ Abrera, B. World Butterflies. Hill HousePublisher. Australia. 2005.http://www.ujungkulon.org/tentang-tnuk/letak-dan-luas
Fachrul, M.F. Metode Sampling Bioekologi. PTBumi Aksara. Jakarta. 2012
Indriyanto.2005. Ekologi Hutan.PT BumiAksara. Jakarta.p 145-146.
Neo, Steven SH. 2001. A Guide To CommonButterflies Of Singapore. Singapore ScieneCentre. Singapore.
MagurranAE. 1988.Ecological Diversity and ItsMeasurement. Princeton University Press.New Jersey
Peggie D, Amir M. 2006. Practical Guide to theButterflies of Bogor Botanical Garden –Panduan Praktis Kupu-kupu di Lahanpertanian Raya Bogor. Bidang Zoologi,Pusat Penelitian Biologi, LIPI Cibinong
dan Nagao Natural EnvironmentFoundation.
Peggie, D. 2012. A list of the butterfly of UjungKulon National Park, Java, Indonesia.Research Center for Biology IndonesianInstitute of Sciences Bogor Indonesia
Ruslan, H. Febrian E, Anggoro, E.S. danPranoto, C.H. 2012. Keanekaragamankupu-kupu Superfamili Papilionoidea dikampung Paniis, Desa Taman jaya, sekitarTaman Nasional Ujung Kulon.ProcecdingSeminar Nasional BiodiversitasIV.Departemen Biologi Fakultas Sains danTeknologi Universitas Airlangga. Surabaya.Hal 289.
Speight M.R, Hunter M.D dan Watt A.D. 1999.Ecology of Insects Consepts andApplication Blackwell Science, Ltd
Triplehorn CA, Johnson NF.2005. Borror andDelong’s Introduction to the Study ofInsects.Ed ke-7. Belmont: ThomsonBrooks/Cole..

ISSN 978-602-14989-0-3 BioETI
Potensi Monochoria vaginalis dalam mengakumulasi diperairantercemar Merkuri (Hg)HAVIZA ANUGRA, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIA
Labor Riset Fisiologi Tumbuhan dan Kultur Jaringan, Jurusan Biologi FMIPA, Universitas Andalas,Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAK
Penelitian tentang daya akumulasi tanaman eceng gondok (Eichornia crassipes), eceng padi (Monochoria vaginalis) dangenjer (Lymnocharis flava) dalam meremediasi perairan tercemar merkuri (Hg), dilakukan pada bulan Desember 2013sampai bulan Januari 2014 di rumah kaca dan Laboratorium Fisiologi Tumbuhan dan Kultur Jaringan Jurusan Biologi,Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Andalas Padang. Penelitian ini menggunakan metodeeksperimen RAL Faktorial, dengan perlakuan jenis tanaman : eceng gondok (Eichornia crassipes), eceng padi(Monochoria vaginalis) dan genjer (Lymnocharis flava) dan konsentrasi merkuri (Hg) yaitu : kontrol, 0,005 ppm, 0,1 ppm,0,2 ppm. Dari penelitian ini memperlihatkan Tanaman eceng gondok (Eichornia crassipes) lebih efektif sebagai agenfitoremediasi dibandingkan tanaman eceng padi (Monochoria vaginalis) dan genjer (Lymnocharis flava) dalammeremediasi cemaran merkuri (Hg). Daya akumulasi tanaman eceng gondok (Eichornia crassipes) lebih besar daritanaman eceng padi (Monochoria vaginalis) dan genjer (Lymnocharis flava)
Key words: Merkuri (Hg), Fitoremediasi, Tanaman Akumulator.
Pendahuluan
Saat ini fenomena kerusakan lingkungan terjadidi seluruh sektor, salah satunya adalah sektorpertambangan. Pertambangan emas tanpa ijin(PETI) menjadi sumber pencemaran, karenamerkuri digunakan penambang emas untukpengolahan bijih melalui proses amalgamasisebagai media pengikat emas (Setiabudi, 2005).
Kegiatan pertambangan emas tanpa ijin(PETI) menyebabkan hilangnya keanek-aragaman hayati, terjadinya degradasi padadaerah aliran sungai, perubahan bentuk lahandan terlepasnya logam-logam berat yang dapatmasuk ke lingkungan perairan (Rahmawaty,2002) Jika Hg masuk kedalam perairan makayang lebih membahayakan adalah setelah Hgtersebut berada di perairan, karena Hg tersebutakan diubah oleh mikroorganisme menjadi metilmerkuri (Tulalessy, 2005). Senyawametilmerkuri ini dapat menyerang syarafmanusia melalui peredaran darah karenagastrointestine manusia mampu menyerapsekitar 95% (Rugh, 2000 dalam: Nofiani danGuzrisal, 2004).
Salah satu upaya untuk memulihkan lingkunganyang tercemar merkuri adalah melalui upayaremediasi, dalam hal ini menggunakan tanamanyang mengakumulasi merkuri dari lapangan(Phytoremediasi). Mekanisme tanamanmengakumulasi merkuri adalah tanaman padasaat menyerap logam berat, akan membentuksuatu enzim reduktase di membran akarnya.Reduktase ini berfungsi mereduksi logam yangselanjutnya diangkut melalui mekanisme khususdi dalam membran akar. Pada saat terjaditranslokasi di dalam tubuh tanaman, logam yangmasuk ke dalam sel akar, selanjutnya diangkutke bagian tumbuhan yang lain melalui jaringanpengangkut yaitu xylem dan floem. Prosestanaman mengakumulasi merkuri yaitu :Pertama, penyerapan oleh akar, senyawa-senyawa yang larut dalam air biasanya diambiloleh akar bersama air, sedangkan senyawa-senyawa hidrofobik diserap oleh permukaanakar. Kedua, translokasi logam dari akar kebagian tanaman lain. Setelah logam menembusendodermis akar, logam atau senyawa asing lainmengikuti aliran transpirasi ke bagian atastanaman melalui jaringan pengangkut (xylem

Haviza Anugra, Zozy Aneloi Noli dan Fuji Astuti Febria 123
dan floem) ke bagian tanaman lainnya. Ketiga,lokalisasi logam pada sel dan jaringan, hal inibertujuan untuk menjaga agar logam tidakmenghambat metabolisme tanaman. Sebagaiupaya untuk mencegah peracunan logamterhadap sel, tanaman mempunyai mekanismedetoksifikasi, misalnya dengan menimbunlogam di dalam organ tertentu seperti akar(Priyanto dan Prayitno 2004).
Dari hasil penelitian pendahuluanpenambangan emas tanpa izin (PETI), diketahuikandungan merkuri (Hg) pada perairanSijunjung dilokasi Jorong Silokek cukup tinggiyaitu 0,074 ppm (Anugra, Noli dan Febria,2013). Berdasarkan Peraturan PemerintahRepublik Indonesia No. 82 Tahun 2001, tentangkadar maksimum merkuri (Hg) dalam air sungaiadalah 0,005 ppm, sehingga perlu dilakukanupaya remediasi.
Menurut Priyanto dan Suryati (2003)beberapa tanaman diketahui mampumengakumulasi merkuri (Hg) diantaranyakelas liliopsida seperti eceng gondok dan genjer,hasil penelitian Hudori, Hakim dan Purbata(2009) diketahui bahwa, tanaman eceng gondokdapat menurunkan limbah yang mengandung Hgbuatan dengan efisiensi penurunan optimal padakonsentrasi 0,5 ppm sebesar 99,72% . penelitianJuhaeti, Syarif , Hidayati, dan Hidayat (2009)tanaman Lymnocharis flava pada panen ketigaselama 1 tahun persentase penyerapan merkuri(Hg) sekitar 25 % yang diakumulasi di akar dantajuk tanaman.
Pada penelitian ini, digunakan tanaman yangtergolong dalam kelas yang sama dengan E.crassipes yaitu : M. vaginalis (kelas liliopsida) ,dengan asumsi tanaman ini mampumengakumulasi merkuri (Hg) di perairan.Penelitian ini bertujuan mengetahui potensitanaman air eceng padi (M. vaginalis)berpotensi sebagai agen fitoremediasi terhadapkonsentrasi merkuri (Hg).
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan Desember2013 sampai bulan januari 2014 dirumah kaca
Laboratorium Fisiologi Tumbuhan JurusanBiologi FMIPA Universitas Andalas Padang.Penelitian ini dilakukan dengan metodeeksperimen yang menguji kemampuan ecengpadi (M. vaginalis) dalam mengakumulasimerkuri (Hg). Sebagai perlakuan konsentrasiMerkuri yaitu : 0 ppm (kontrol), 0,05 ppm, 0,1ppm, 0,2 ppm. Pada percobaan ini pengamatandilakukan setiap 2 minggu selama 2 bulan.
Alat yang digunakan dalam penelitian iniadalah ICP (Inductively Coupled Plasma),wadah, alat tulis, plastik, sedangkan bahan yangdigunakan adalah air, tanaman dan eceng padi(M. vaginalis), merkuri (Hg) sintetis.Data yang diperoleh disajikan secara deskripsi,dengan parameter pengamatan : tinggi tanaman,berat basah akar, berat basah tajuk, berat keringakar, berat kering tajuk, kadar merkuri akar, dankadar merkuri tajuk
HASIL DAN PEMBAHASAN
Tabel 1 menunjukkan pengaruh perlakuanmerkuri (Hg) terhadap pertambahan tinggitanaman, berat basah akar, berat basah tajuk,berat kering akar, dan berat kering tajuk dapatdilihat bahwa, pada pertambahan tinggi tanamanyang paling tinggi terdapat pada perlakuankonsentrasi merkuri (Hg) 0,2 ppm, pertambahanberat basah akar yang paling tinggi terdapatpada perlakuan konsentrasi merkuri (Hg) 0,005ppm, pertambahan berat basah tajuk yang palingtinggi terdapat pada perlakuan konsentrasimerkuri (Hg) 0,1 ppm, pertambahan berat keringakar yang paling tinggi terdapat pada perlakuankonsentrasi merkuri (Hg) 0,2 ppm, pertambahanberat kering tajuk yang paling tinggi terdapatpada perlakuan kontrol.
Dari hasil parameter menunjukkan bahwatanaman M. vaginalis dapat tumbuh dengan baikdengan konsentrasi merkuri (Hg) yang terendahsampai dengan konsentrasi merkuri (Hg) yangtertinggi, dan tanaman M. vaginalis mampuberadaptasi pada lingkungan yang tercemarmerkuri (Hg). Tanaman M. vaginalis berasaldari kelas Liliopsida dapat mencapai tinggi
Haviza Anugra, Zozy Aneloi Noli dan Fuji Astuti Febria 124
sekitar 5 - 50 cm (Juhaeti, Syarif , Hidayati, danHidayat, 2009).
Tabel 1. Rata-rata pertambahan tinggi tanaman,berat basah akar, berat basah tajuk, berat keringakar dan berat kering tajuk M. vaginalis
konsentrasimerkuri(Hg)
Tinggitanaman(cm)
Beratbasahakar(gr)
Beratbasahtajuk(gr)
Beratkeringakar(gr)
Beratkeringtajuk(gr)
Kontrol 8,40 5,49 8,84 5,02 7,330,005 ppm 8,40 6,17 8,41 5,25 3,800,1 ppm 7,26 4,77 9,59 5,65 4,050,2 ppm 9,50 4,44 7,30 5,92 4,42
Dari hasil berat basah akar dan tajuktanaman M. vaginalis diperkirakan bahwakonsentrasi dari merkuri (Hg) tidakmenghambat aktivitas metabolik tanaman. Beratbasah merupakan total berat tanaman yangmenunjukkan hasil metabolik tanaman(Salisbury dan Ross, 1995). Hal ini jugamenujukkan bahwa M. vaginalis toleranterhadap merkuri (Hg), dimana tumbuhanhiperakumulator mempunyai kemampuan dalammengakumulasi dan mentranslokasi dalam halini logam berat merkuri. Merkuri dilokalisasipada bagian jaringan tertentu untuk menjagaagar tidak menghambat metabolisme tumbuhantersebut. Pada masing-masing organ, polutanyang diserap segera diuraikan melalui prosesmetabolisme tumbuhan secara enzimatik. Prosesini disebut fitodegradasi dimana kontaminanorganik diserap kedalam tanaman dalam prosesmetabolisme tanaman dan dapat merombakkontaminan didalam jaringan tanaman menjadimolekul yang tidak bersifat toksis. Enzim yangberperan pada proses ini biasanya adalahdehaloganases, oxygenases, dan reductases(Wang et al.,2004).
Dari hasil berat kering akar dan tajuktanaman M. vaginalis menunjukkan bahwakonsentrasi merkuri tidak mengganggu danmenghambat dalam proses respirasi danfotosintesis pada tanaman sehingga tanaman airini memiliki pertumbuhan yang baik pada mediayang tercemar merkuri (Hg). Hal ini sesuaidengan pernyataan Gardner et al, (1991)
menyatakan bahwa hasil berat kering tanamanmerupakan keseimbangan pengambilan C02
melalui respirasi. Hal ini juga diperkirakanbahwa bobot kering tanaman erat sekalikaitannya dengan proses fotosintesis sertapenyimpanan fotosintat. Sebagian dari hasilfotosintesis digunakan untuk respirasi danasimilasi, kemudian kelebihannya disimpanpada bagian-bagian tertentu dari tanamanterutama batang dan akar.
Bobot kering biasanya dijadikan indikatorbahwa semakin baik pertumbuhan tanamanmakin baik pula terhadap bobot kering tanaman(Sitompul dan Guritno, 1995). Hal inimembuktikan bahwa tanaman air yaitu M.vaginalis adalah tanaman hiperakumulator yangdapat tumbuh baik dengan konsentrasi merkurisebesar 0,005 ppm, 0,1 ppm dan 0,2 ppm.
Gambar 1. Pertambahan Rata-rata Tinggi TanamanM. vaginalis selama 2 bulanpengamatan
Gambar 1 menunjukkan pertambahan rata-rata tinggi tanaman dari minggu pertama sampaidengan minggu ke delapan, dapat dilihat bahwatanaman M. vaginalis pada minggu pertamalebih tinggi dari minggu kedua, keempat,keenam dan kedelapan. Hal ini menunjukkanbahwa pada minggu pertama tanaman inimampu beradaptasi dengan baik dengan air yangtercemar merkuri (Hg), sedangkan pada minggukedua, keempat, keenam dan kedelapanpertambahan rata-rata tanaman hanyamengalami sedikit pertambahan, hal ini diduga
02468101214161820
1 2 4 6 8
pengamatan ke
Ting
gi T
anam
an (c
m)
kontrol
0,005 ppm
0,1 ppm
0,2 ppm

Haviza Anugra, Zozy Aneloi Noli dan Fuji Astuti Febria 125
bahwa merkuri mempengaruhi pertumbuhantanaman dimana pada minggu kedua, keempat,keenam dan kedelapan interaksi tanaman danmerkuri dalam proses meremedisi danmengakumulasi merkuri (Hg) lebih banyak, Halini dapat terjadi karena respons pertumbuhanyang diberikan tanaman M. vaginalis terhadapkadar merkuri pada media tanam, tetapipertumbuhan tanaman sampai minggukedelapan tetap mampu tumbuh dengan baik,Menurut Supriyanto (1999), perkembanganyang seimbang antara bagian pucuk tanamandan akar menunjukkan pertumbuhan yang baik.
Gambar 2 : Akumulasi Merkuri Pada BagianAkar dan Tajuk Tanaman
ket : a = kontrolb = 0,005 ppmc = 0,1 ppmd = 0,2 ppm
Gambar 2 menunjukkan akumulasi merkuripada bagian tanaman M. vaginalis akar dantajuk, tanaman M. vaginalis berpotensi sebagaibioremediator ditunjukkan oleh konsentrasimerkuri akar dan tajuk pada minggu terakhirpengamatan, akumulasi pada akar padakonsentrasi 0,005 ppm akumulasi merkurisebesar 0,003 ppm, pada konsentrasi 0,1 ppmakumulasi merkuri sebesar 0,025 dan padakonsentrasi 0,2 ppm akumulasi merkuri sebesar0,027. Sedangkan akumulasi pada tajuk padakonsentrasi 0,005 ppm akumulasi merkurisebesar 0,0013 ppm, pada konsentrasi 0,1 ppmakumulasi merkuri sebesar 0,016 dan padakonsentrasi 0,2 ppm akumulasi merkuri sebesar
0,016. Bayu et al (2010) yang menyatakanbahwa semakin besar konsentrasi Hg dalammedia, maka tanaman akan semakin banyakmenyerap Hg.
Adanya kecenderungan akumulasi merkuri diakar yang lebih tinggi dari pada di tajukdisebabkan karena akar merupakan organtanaman yang berfungsi menyerap unsur haradari media tanam dan sekaligus organ yangkontak langsung dengan media tanam yaitu airyang terkontaminasi logam merkuri (Hg).Hal ini menunjukkan bahwa tanaman M.vaginalis memiliki kemampuan sebagai tanamanhiperakumulator, Rugh (2000) menyatakanbahwa merkuri dapat diserap oleh tumbuhan dankemudian menguap melalui daun dalam bentukHg0. Sebagian tumbuhan mengakumulasimerkuri lebih banyak terdapat di bagian akardari pada di bagian atas tumbuhan, artinya adakemungkinan merkuri terserap dari tanah atauair melalui akar atau xylem kemudianmengendap di dalam akar tumbuhan. Logamberat termasuk merkuri yang ada dalam tanahatau air tidak baik untuk pertumbuhan tanaman.Pada kondisi tercemar, secara umum logamberat yang diangkut terbatas hanya sampai padaakar tanaman. Kemungkinan hal ini terjadikarena beberapa mekanisme yang mencegahpemuatan logam ke dalam xylem secara berlebih,yaitu mekanisme dari pengikat logam spesifik didalam akar untuk melindungi tanaman darikonsentrasi logam yang tinggi. (Liao et al.,2000dalam Reichman, 2002).
Dari Gambar 2 juga membuktikan bahwatanaman M. vaginalis dapat digunakan sebagaitanaman fitoremediasi pada proses rizofiltrasidan fitoekstraksi. Rizofiltrasi yaitu prosespengendapan zat kontaminan oleh akar untukmenempel di akar (Khan AG,2005). Sedangkandalam proses fitoekstraksi yaitu mekanismeyang terjadi ketika akar tumbuhan mengabsorpsilarutan polutan sekitar akar ke dalam akar, yangselanjutnya ditranslokasi ke dalam organtumbuhan melalui pembuluh xylem(Erakhrumen & Agbontalor, 2007).
0
0.005
0.01
0.015
0.02
0.025
0.03
a b c dKad
ar M
erku
ri Ta
nam
an (p
pm)
Konsentrasi Merkuri (ppm)
akar
tajuk

Haviza Anugra, Zozy Aneloi Noli dan Fuji Astuti Febria 126
KESIMPULAN
Berdasarkan penelitian yang telah dilakukantentang potensi monochoria vaginalis dalammengakumulasi diperairan tercemar merkuri(Hg) , dapat disimpulkan bahwa :Tanaman eceng padi (M. vaginalis) adalahtanaman yang yang berpotensi sebagai agenfitoremediasi terhadap konsentrasi merkuri (Hg).Akumulasi merkuri lebih banyak di bagian akardibandingkan pada bagian tajuk.
DAFTAR PUSTAKA
Bloom Anugra. H. Z. A.Noli. F.A. Febria. NonPublished. Biologi FMIPA, Univ. Andalas.Padang
Bambang Tjahjono Setiabudi., 2005,Pendataan Penyebaran Merkurikecamatan Kokap. Badan Konservasi.Yogyakarta.
Bayu MI, Roosmini D, Tjahaja P I. 2010.Akumulasi Logam Kobalt dari TanahAndosol Menggunakan Tanaman SawiIndia (Brassica juncea).Program StudiTeknik Lingkungan FTSL ITB. Bandung.
Erakhrumen dan Agbontalor, A. 2007.Phytoremediation: An EnvironmentallySound Technology for Pollution Prevention,Control and Remediation in DevelopingCountries, Educational Research andReview Vol. 2 (7)
Gardner, F.P., Pearce, P. R. B., Mitchell, R. L.1991. Fisiologi Tanaman Budidaya. UIPress: Jakarta.
Hudori. Hakim L dan Purbata F. A. 2009.Pengolahan Logam Berat Merkuri DariEffluent Karbon Aktif MenggunakanTanaman Eceng Gondok (Eichhorniacrassipes). Fakultas Teknik Sipil danPerencanaa. Universitas Islam Indonesia :Yogyakarta.
Khan AG. 2005 . Rule of Soil Microbes in theRhizospheres of Plants Growing on TraceMetal Contaminated Soils inPhytoremediation. Journal of TraceElements in Medicine and Biology.
Nofiani dan Gusrizal. 2004. Bakteri ResistenMerkuri Spektrum Sempit dari DaerahBekas Penambangan Emas Tanpa Izin(PETI) Mandor, Kalimantan Barat. JurnalNatur Indonesia. 6(2): 67-74
Priyanto. B dan Suryati . T. 2003.
Eliminasi Logam Berat Dalam AirLimbah Menggunakan Tanaman Air.Balai Teknologi Lingkungan BadanPengkajian dan Penerapan Teknologi.Tek. Ling, P3TL BPPT.4(3): 143-147
Priyanto B., Prayitno J. 2004. Fitoremediasisebagai Sebuah Teknologi PemulihanPencemaran Khusus Logam Berat.http : //ltl.bppt.tripod. com/sublab/1flora1.htm.
Rahmawati. 2002. Restorasi Lahan Pascatambang Berdasarkan Kaidah Ekologi.Fakultas Pertanian. Universitas SumateraUtara
Rugh CL, Bizily SP, Meagher. 2000.Phytoreduction of Enviromental MerkuriPollution, (di dalam) Raski, I., dan Ensley,B. D (penyunting), Phytoreduction ofToxicMetal Using Plants to Clean Up TheEnviroment.New York: Wiley IntersciencePublication, Jhon Wiley and Sons.Inc
Reichman SM. 2002 . The Responses of Plantsto Metal Toxicity: A Review Focusing onCopper, Manganese and Zinc. TheAustralian Minerals Energy EnvironmentFoundation Published as Orcasional PaperNo.14
Salisbury, B.F. dan C.C.W. Ross. 1995.Fisiologi Tumbuhan. Jilid 3. ITB:Bandung.
Sitompul, S. M. dan Guritno, B. 1995.Analisis Pertumbuhan Tanaman. GadjahMada University Press, Yogyakarta.
Supriyanto. 1999. The effectiveness of someectomycorrhizal fungi in alginate beads inpromoting the growth of severalDipterocarp seedlings. Biotropika 12: 59-77.
Juhaeti,T. N. Hidayati, Syarif dan S. Hidayat.2009. Uji Potensi TumbuhanAkumulatorMerkuri untuk Fitoremediasi LingkunganTercemar Akibat Kegiatan PenambanganEmas Tanpa Izin (PETI) di KampungLeuwi Bolang, DesaBantar Karet,Kecamatan Nanggung, Bogor : PusatKonservasi Tumbuhan Kebun Raya Bogor.
Tulalessy A. H, 2005. Studi PencemaranMerkuri Pada Kawasan PenambanganEmas Rakyat Tatelu Sulawesi Utara. IPB :Bogor.
Wang, L.W, J. Y. Wu, Z. Z. Xia, K Wang,2004. Anew Type Adsorber ForAdsorption Ice Maker On Fishing Boats,School Of Mechanical Engineering,Institute Of Refrrigeration and Cryogenics,Shanghai Jiao Tong University, Shanghai.

ISSN 978-602-14989-0-3 BioETI
Pengaruh Gambir (Uncaria gambir Roxb) terhadap kadar guladarah pada Mencit Putih Jantan (Mus musculus) yang diinduksiAloksanINDAH FAJARWATI, EFRIZAL DAN RESTI RAHAYU
Laboratorium Riset Fisiologi Hewan, Jurusan Biologi FMIPA, Universitas Andalas, Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian mengenai pengaruh gambir (Uncaria gambir Roxb) terhadap kadar gula darah pada mencit putih jantan (Musmusculus) yang diinduksi Aloksan telah dilakukan pada bulan November 2013 hingga April 2014 di Laboratorium RisetFisiologi Hewan, Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Andalas, Padang.Penelitian ini bertujuan untuk mengetahui kemampuan serta dosis efektif dari gambir dalam menurunkan kadar guladarah. Sampel penelitian dikelompokkan menjadi 6 perlakuan yang terdiri dari mencit normal yang diberi aquadessebagai kontrol negatif (P1), mencit yang diinduksi aloksan sebagai kontrol positif (P2), aloksan 200 mg/kg BB danUncaria gambir 200 mg/kg BB (P3), aloksan 200 mg/kg BB dan Uncaria gambir 300 mg/kg BB (P4), aloksan 200 mg/kgBB dan Uncaria gambir 400 mg/kg BB (P5), aloksan 200 mg/kg BB dan Uncaria gambir 500 mg/kg BB (P6). Hasilpenelitian menunjukan bahwa gambir dapat menurunkan kadar gula darah. Dosis terbaik dari gambir dalam menurunkankadar gula darah adalah 200 mg/kg BB setelah pemberian selama 21 hari berturut-turut, dosis ini mampu mengembalikankadar gula darah ke kisaran normal yaitu 98,3 mg/dl dengan penurunan sebesar 64,2% dari kadar gula darah setelahdiinduksi aloksan.
Key words: Aloksan, gambir (Uncaria gambir Roxb), kadar gula darah
Pendahuluan
Gambir (Uncaria gambir Roxb) merupakansalah satu komoditas ekspor utama Indonesiadari sektor perkebunan. Sumatera Barat sebagaisentra produksi gambir, mampu memasok 80persen kebutuhan pasar dunia dengan negaratujuan utama adalah India, Bangladesh, Taiwan,Jepang, dan Perancis (Dhalimi, 2006). Luaskebun gambir di Sumatera Barat pada tahun2008 adalah 19.575 ha dengan total produksi13.956 ton (Idrus, 2012).
Secara tradisional gambir memiliki banyakmanfaat. Menurut Laus (2004) gambirdigunakan untuk mengatasi diare, sariawan,sakit kepala, demam, luka bakar, dan berbagaigangguan gastrointestinal. Baharuddin danTaskirawati (2009) menyatakan bahwa gambirbermanfaat sebagai penyamak kulit, bahancampuran pelengkap makanan serta bahan bakudalam berbagai industri, seperti industri farmasi,kosmetik, dan tekstil.
Gambir diduga bermanfaat untuk menjagastabilitas gula darah. Kemampuan gambirdalam menjaga stabilitas gula darah berasaldari senyawa aktif yang terkandung dalamgambir. Penelitian Anggraini et al., (2011)menunjukkan bahwa gambir yang diekstraksisecara tradisional memiliki aktivitasantioksidan yang tinggi. Adanya aktivitasantioksidan yang tinggi pada gambirdiharapkan mampu membantu mengatasi stresoksidatif yang mendasari patogenesis diabetesmellitus (Astiyandani dkk., 2010).
Sejumlah penelitian yang telah dilakukanbelakangan ini bergerak dalam evaluasipengaruh aktivitas antioksidan terhadappengontrolan kadar glukosa darah (Nishikawaand Araki, 2013). Penelitian Domingues et al.,(2011) juga membuktikan bahwa tumbuhangambir amazon (Uncaria tomentosa) dapatmengatasi gangguan gula darah melaluimekanisme perbaikan kerusakan sistematikpada pankreas.

Indah Fajarwati, Efrizal dan Resti Rahayu 128
Diabetes mellitus merupakan suatu penyakitmetabolik yang ditandai dengan tingginyakadar gula darah baik karena tubuh tidakmemproduksi insulin yang cukup atauberkurangnya efektivitas biologis dari insulin(Guyton and Hall, 2007). Penyakit ini telahdikategorikan sebagai penyakit global olehWorld Health Organization (WHO) denganjumlah penderita di dunia mencapai 199 jutajiwa pada tahun 2009. Menurut data statistikdari studi Global Burden of Disease WHOtahun 2004, Indonesia menempati peringkatpertama di Asia Tenggara, dengan prevalensipenderita sebanyak 8.426.000 jiwa di tahun2000 dan diproyeksi meningkat 2,5 kali lipatsebanyak 21.257.000 penderita pada tahun2030 (WHO, 2009).
Berbagai upaya dilakukan untuk menekanjumlah penderita diabetes mellitus, salahsatunya melalui terapi herbal. Terapi ini lebihdiarahkan untuk menjaga tingkat gula darahsedekat mungkin dengan batas normal. Validasiilmiah membuktikan beberapa spesies tanamanmemiliki khasiat dalam mengurangi kadar guladarah, sehingga minat dalam penelitian obatherbal terus berlanjut. Harapan kedepan kitamampu membawa senyawa yang lebih amandan lebih efektif pengganti obat-obatan sintetik(Jerald, Joshi, and Jain, 2008).
Penelitian terhadap penyakit diabetesmellitus biasanya dilakukan denganmenginduksi agar hewan uji menderita diabetes.Penginduksian ini dapat dilakukan denganbeberapa cara salah satunya denganmenggunakan senyawa kimia Aloksan (King,2012). Aloksan bekerja spesifik pada pankreasdan dengan cepat diserap oleh sel-sel βpankreas membentuk senyawa radikal bebasdan menghasilkan sel-sel yang nekrosis. Selainitu, aloksan juga terserap di hati akan tetapijaringan pada hati lebih tahan (resisten)terhadap radikal bebas dibandingkan dengan selβ pankreas (Szkudelski, 2001).
Berdasarkan uraian yang telah dipaparkanmaka perlu dilakukan penelitian mengenaipengaruh gambir dalam menurunkan kadar gula
darah mencit putih jantan (Mus musculus) yangdiinduksi aloksan. Dengan adanya penelitianini, diharapkan dapat menjadi bagian pentingdari upaya pemanfaatan tumbuhan lokalsebagai produk obat andalan untuk mengatasipenyakit diabetes mellitus serta mendukungpenggalan kalimat “back to herbal” yang sudahmenjadi suatu trend kesehatan dunia saat ini.
BAHAN DAN METODE
Penelitian ini menggunakan metodaeksperimental. Sampel penelitian dikelompokkan menjadi 6 perlakuan yang terdiri darimencit normal yang diberi aquades sebagaikontrol negatif (P1), mencit yang diinduksialoksan sebagai kontrol positif (P2), aloksan200 mg/kg BB dan Uncaria gambir 200 mg/kgBB (P3), aloksan 200 mg/kg BB dan Uncariagambir 300mg/kg BB (P4), aloksan 200 mg/kgBB dan Uncaria gambir 400 mg/kg BB (P5),aloksan 200 mg/kg BB dan Uncaria gambir500 mg/kg BB (P6).
Mencit dikondisikan untuk menderitadiabetes dengan menginduksikan aloksanintraperitonial dosis tunggal 200 mg/kg BB(King, 2012; Jing, 2009). Setelah dilakukanpenginduksi aloksan, pada hari ke-7 dilakukanpengukuran kadar gula darah. Kriteria mencitpositif diabetes mellitus apabila kadar guladarah puasa ≥150 mg/dl. Mencit yang sudahmemenuhi kriteria dimasukkan ke dalamkelompok sampel.
Gambir diberikan pada tingkatan dosis yangberbeda-beda sesuai level yang telah ditentukansetiap hari selama 21 hari berturut-turut.Gambir pertama kali diberikan adalah pada harike-7 setelah penginduksian aloksan. Gambiryang diberikan merupakan gambir yang telahdisuspensikan dalam aquades sehinggadihasilkan konsentrasi larutan yang sesuaidengan tingkatan dosis per masing-masingkelompok perlakuan. Pemberian suspensikepada hewan uji dilakukan secara oral denganmenggunakan jarum sonde.
Kadar gula darah diukur sebanyak 5 kali.Sebelum pengambilan darah, semua hewan uji

Indah Fajarwati, Efrizal dan Resti Rahayu 129
dipuasakan selama ±16 jam (Ayala et al., 2010).Hal ini bertujuan untuk menghilangkan faktor-faktor lain yang mempengaruhi perhitunganpada pengukuran kadar gula darah (Padilah,2009). Pengukuran pertama adalah sebelumhewan uji diberi perlakuan, hal ini bertujuanuntuk memastikan kadar gula darah beradadalam rentang kadar gula darah normal.Pengukuran kadar gula darah selanjutnyaadalah setelah hewan uji diinduksi aloksannamun belum diberikan gambir (awal),pengukuran ini untuk memastikan keberhasilaninduksi aloksan. Pengukuran kadar gula darahpada hari ke 7, 14, dan 21 adalah untuk melihatpengaruh dari gambir. Semua sampel darahdiambil dari ekor mencit yang telah dibersihkandengan alkohol 70%. Selanjutnya ekor mencitdipotong maksimal 0,2 cm dari ujung ekor,dilakukan pemijatan perlahan terhadap ekoragar darah keluar. Kemudian diukur kadar guladarah hingga diperoleh kadar gula mencitdalam satuan mg/dl. Adapun parameter yangdiukur pada penelitian ini kadar gula darahhewan uji. Kadar gula darah hewan uji setelahpemberian gambir dibandingkan dengan kadargula darah hiperglikemia awal, dihitungsebagai PGl:
PGl =– 100%
(Dimodifikasi dari Pujilestari dan Pratiwi, 2009).
Keterangan:PGl = Persentase perubahan kadar gula darahminggu ke-n terhadap kadar gula darah awalsetelah diinduksi aloksan.
HASIL DAN PEMBAHASAN
Penginduksian AloksanTelah dilakukan penelitian tentang pengaruhgambir terhadap kadar gula darah pada mencitputih jantan yang diinduksi aloksan. Jumlahmencit yang memenuhi kriteria inklusi padaawal penelitian yaitu sebanyak 24 individuyang terbagi ke dalam kelompok kontrol (P1,
P2) dan kelompok perlakuan (P3, P4, P5, P6).Namun, dalam perjalanan penelitian didapatkanmencit yang mati sehingga terjadi penguranganhewan uji pada masing-masing kelompok.
Mencit yang mati diduga disebabkan olehkadar gula darah yang terlalu tinggi setelahdilakukan penginduksian aloksan sebagai zatdiabetogenik. Hal ini mengakibatkan terjadinyagangguan homeostasis yang besar sehinggaberdampak pada kerja fisiologi tubuh. Kadargula darah darah mencit yang berhasil diukursebelum kematian menunjukkan simbol HIpada alat ukur yang berarti high glucose, HI inididapatkan jika kadar gula darah hewan uji>500 mg/dl sehingga kadar gula darah ini tidakmampu dibaca oleh alat ukur yang digunakan.Perbedaan sensitivitas beberapa ekor hewan ujipada penelitian ini terkait dengan responfisiologi tubuh dan ketahanan yang belumdapat dikontrol dan dimodifikasi untuk dapatpersis sama antara satu dan yang lainnya. Halini didukung oleh pernyataan Goodman &Gilman (2012) bahwa beberapa individumemiliki perbedaan respon terhadap suatu zat.Penelitian pada kembar identik dan nonnonidentik menunjukkan bahwa genotipmerupakan penentu yang sangat penting dalamperbedaan laju metabolisme suatu zat danbiotransformasinya memiliki keragaman yangbesar diantara individu.
Aloksan 200 mg/kg mampu menimbulkankondisi diabetes mellitus dengan kadar guladarah mencit pada penelitian ini berkisar di atas200 mg/dl. Kadar gula darah mencit sesudahdan sebelum diinduksi aloksan sebagaimanadisajikan pada Gambar 1.Setelah kondisi diabetes mellitus didapatkan,selanjutnya hewan uji diperlakukanberdasarkan kelompok perlakuan. Gambirdiberikan selama 21 hari dengan pengecekangula darah sebanyak 3 kali yang dilakukan padamasing-masing minggu (Hari ke-7, 14, 21).Mencit diabetes yang digunakan dalampenelitian ini memiliki rentang kadar guladarah 203 mg/dl – 478 mg/dl.
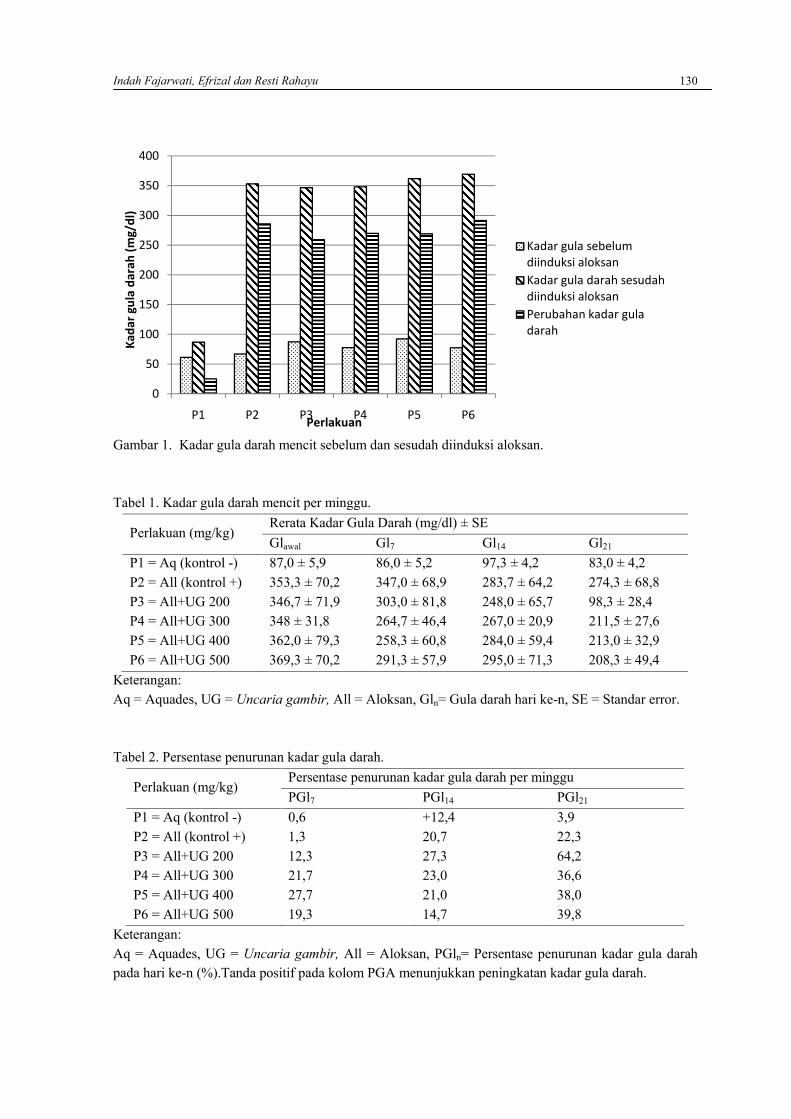
Indah Fajarwati, Efrizal dan Resti Rahayu 130
Gambar 1. Kadar gula darah mencit sebelum dan sesudah diinduksi aloksan.
Tabel 1. Kadar gula darah mencit per minggu.
Perlakuan (mg/kg)Rerata Kadar Gula Darah (mg/dl) ± SEGlawal Gl7 Gl14 Gl21
P1 = Aq (kontrol -) 87,0 ± 5,9 86,0 ± 5,2 97,3 ± 4,2 83,0 ± 4,2P2 = All (kontrol +) 353,3 ± 70,2 347,0 ± 68,9 283,7 ± 64,2 274,3 ± 68,8P3 = All+UG 200 346,7 ± 71,9 303,0 ± 81,8 248,0 ± 65,7 98,3 ± 28,4P4 = All+UG 300 348 ± 31,8 264,7 ± 46,4 267,0 ± 20,9 211,5 ± 27,6P5 = All+UG 400 362,0 ± 79,3 258,3 ± 60,8 284,0 ± 59,4 213,0 ± 32,9P6 = All+UG 500 369,3 ± 70,2 291,3 ± 57,9 295,0 ± 71,3 208,3 ± 49,4
Keterangan:Aq = Aquades, UG = Uncaria gambir, All = Aloksan, Gln= Gula darah hari ke-n, SE = Standar error.
Tabel 2. Persentase penurunan kadar gula darah.
Perlakuan (mg/kg)Persentase penurunan kadar gula darah per mingguPGl7 PGl14 PGl21
P1 = Aq (kontrol -) 0,6 +12,4 3,9P2 = All (kontrol +) 1,3 20,7 22,3P3 = All+UG 200 12,3 27,3 64,2P4 = All+UG 300 21,7 23,0 36,6P5 = All+UG 400 27,7 21,0 38,0P6 = All+UG 500 19,3 14,7 39,8
Keterangan:Aq = Aquades, UG = Uncaria gambir, All = Aloksan, PGln= Persentase penurunan kadar gula darahpada hari ke-n (%).Tanda positif pada kolom PGA menunjukkan peningkatan kadar gula darah.
0
50
100
150
200
250
300
350
400
P1 P2 P3 P4 P5 P6
Kada
r gul
a da
rah
(mg/
dl)
Perlakuan
Kadar gula sebelumdiinduksi aloksanKadar gula darah sesudahdiinduksi aloksanPerubahan kadar guladarah

Indah Fajarwati, Efrizal dan Resti Rahayu 131
Pengaruh Gambir terhadap Kadar Gula DarahMencitDari Tabel 1 dan Tabel 2 dapat diketahuibahwa gambir dosis 400 mg/kg BBmemberikan persentase penurunan paling besarterhadap kadar gula darah mencit yangdiinduksi aloksan setelah pemberian selama 7hari. Ketika pemberian gambir dilanjutkanmenjadi 14 hari, gambir dosis 400 mg/kg BByang pada mulanya memberikan persentasepenurunan paling besar, mengalami penurunankinerja. Dapat disimpulkan bahwa peningkatandosis gambir tidak memberikan pengaruh yangsebanding terhadap penurunan kadar gula darahmencit yang diinduksi aloksan. Pemberiangambir dosis tinggi dalam waktu yang lamadiduga dapat menyebabkan akumulasi gambiryang tinggi di dalam tubuh melebihi jumlahyang diharapkan sehingga menghambat kinerjagambir dalam menurunkan kadar gula darahmencit yang diinduksi aloksan. MenurutMutschler (1991), beberapa jenis antidiabetestidak akan memperlihatkan pengaruh yanglebih besar ketika dosis dinaikkan di atas dosismaksimum karena dapat menghambatmetabolisme insulin dan menurunkan ikataninsulin pada protein plasma.
Gambir dosis 200 mg/kg BB merupakandosis terbaik dalam menurunkan kadar guladarah dibandingkan dengan perlakuan lainnya.Dosis ini mampu mempertahankan danmeningkatkan kinerjanya dengan pemberiandalam waktu yang lebih panjang yaitu selama14 dan 21 hari. Pada pemberian selama 14 hari,gambir dosis 200 mg/kg BB memberikanpersentase penurunan kadar gula darah palingbesar diantara dosis lainnya. Dosis 200 mg/kgBB mampu menurunkan kadar gula darahsebesar 27,3% dari kadar gula darah setelahdiinduksi aloksan. Ketika gambir dosis 200mg/kg BB diberikan selama 21 hari, terjadipeningkatan kemampuan gambir dalammenurunkan kadar gula darah yaitu denganmengembalikan kadar gula darah menujurentang kadar gula darah normal yaitu 98,3
mg/dl dengan penurunan sebesar 64,2% darikadar gula darah setelah diinduksi aloksan.Gambir dosis 200 mg/kg BB memberikanpersentase penurunan gula darah paling besardiduga karena waktu serta jumlah gambir yangdibutuhkan untuk dapat bekerja maksimalsudah tercapai dengan pemberian dosis iniselama 21 hari ini. Lamanya pemberian sertadosis yang digunakan menjadi salah satu faktorpenentu karena hal tersebut berkaitan eratdengan jumlah gambir yang tersedia di dalamtubuh. Hal ini didukung oleh pernyataanMutschler (1991) bahwa untuk menghasilkanpengaruh yang spesifik, suatu obat harustersedia dalam konsentrasi yang tepat di tempatkerjanya.
Kemampuan gambir dalam menurunkankadar gula darah berasal dari zat aktif yangterkadung dalam gambir. Kandungan terbesardari gambir adalah katekin. Katekin merupakansenyawa polifenol yang memiliki aktivitasantioksidan yang tinggi (Anggraini, 2011).Selain dari katekin, aktivitas antioksidan padagambir juga dihasilkan oleh quersetin (Coskun,2004). Menurut Amos (2010), kandunganquersetin yang terdapat pada gambir beradadalam jumlah yang lebih sedikit. Selainmemiliki aktivitas antioksidan, gambir jugamengandung senyawa tannin yang bersifatsebagai astringen (Anggraeni, 2006).
Kemampuan antioksidan yang tinggi darigambir dapat menghambat stres oksidatif yangterjadi pada penderita diabetes mellitus.Penghambatan stres oksidatif dapat mencegahterjadinya penurunan insuline-sensitive glucosetransporter (GLU-4) dan mencegah resistensiinsulin yang terjadi pada penderita sehinggakadar gula darah dapat kembali ke ambangnormal (Widowati, 2008).
Selain melalui peran antioksidan,kemampuan gambir dalam menurunkan kadargula darah diduga berasal aktifitas astringendari gambir. Astringen merupakan senyawayang dapat mengendapkan protein selaputlendir di permukaan usus halus dan membentuk

Indah Fajarwati, Efrizal dan Resti Rahayu 132
suatu lapisan yang melindungi usus, sehinggamenghambat penyerapan gula darah di usus danlaju peningkatan gula darah tidak terlalu tinggi(Anggraeni, 2006). Menurut Carmona (1996)tannin dapat menghambat aktivitas sukrase.Sukrase merupakan enzim yang berperanmengkonversi sukrosa menjadi glukosa danfruktosa sebelum kedua monosakarida tersebutdiserap usus kecil. Dengan demikian,keberadaan tannin berpotensi menghambatpenyerapan glukosa dan fruktosa sertamenurunkan respon glikemik, khususnya padamakanan yang banyak mengandung sukrosa(Hoerudin, 2012).
Menurut Karasov et al., (1992) tannin jugadapat secara langsung menghambat sistempenyerapan glukosa yang tergantungketersediaan natrium (sodium-dependentglucose uptakesystem). Welsch et al., (1989)menemukan bahwa pada konsentrasi 0,5 mg/mLtannic acid dapat menghambat 50% penyerapanglukosa pada potongan usus kecil tikus.
KESIMPULAN
Berdasarkan penelitian yang telah dilakukandiperoleh kesimpulan sebagai berikut:1. Gambir menurunkan kadar gula darah pada
mencit putih jantan yang diinduksi aloksan.2. Dosis terbaik dari gambir yang dapat
menurunkan kadar gula darah pada mencitputih jantan yang diinduksi aloksan adalah200 mg/kg BB setelah pemberian selama 21hari berturut-turut.
Ucapan TerimakasihPenulis mengucapkan terimakasih kepadaKemendikbud, Dikti yang telahmembiayai penelitian ini melalui danahibah Program Kreatifitas MahasiswaPenelitian (PKM-P) 2014.
DAFTAR PUSTAKA
Amos. 2010. Kandungan Katekin Gambir SentraProduksi di Indonesia. Jurnal standardisasiISJD.
Anggraeni, A.D. 2006. Pemberian Infusa BijiAlpukat (Persea Americana Mill.) terhadapKadar Glukosa Darah Tikus Wistar yangDiberi Beban Glukosa. Artikel Karya TulisIlmiah Program Pendidikan SarjanaFakultas Kedokteran UniversitasDiponegoro Semarang.
Anggraini, T., T. Akihiro, Y. Tomoyuki, I.Tomio. 2011. Antioxidative Activity andCatechin Content of Four Kinds of Uncariagambir Extracts From West Sumatra,Indonesia. Afr. J. Biochem Res.
Astiyandani, P.G., G.A, Permana., A.W.P.D.Vedayanti., I.D. Larayanthi., P. Windasari.,I.A.Wahyuniari. 2010. Uji Klinis In VivoKonsumsi Daluman (Cycllea Barbata)Terhadap Penurunan Kadar Gula DarahPada Tikus Wistar Jantan Dengan DiabetesMellitus Tipe 2. IPTEKMA
Ayala,J.E.,V.T. Samuel., G.J. Morton., S. Obici.,C.M. Croniger., G.I. Shulman., D.H.Wasserman., O.P. McGuinnes. 2010.Standard operating procedures fordescribing andperforming metabolic testsof glucose homeostasisin mice. Specialarticle Disease Models & Mechanisms 3
Baharuddin dan I. Taskirawati. 2009. Buku AjarHasil Hutan Bukan Kayu. FakultasKehutanan Universitas Hasanudin.Makasar.
Carmona, A., L. Borgudd, G. Borges, and A.Levy-Benshimol. 1996. Effect of blackbean tannins onin vitro carbohydratedigestion and absorption. The Journal ofNutritional Biochemistry.
Coskun, O., M. Kanter., F. Armutcu., K. Cetin.,B. Kaybolmaz., O. Yazgan. 2004.Protective Effects of Quercetin, AFlavonoid Antioxidant, in AbsoluteEthanol-Induced Acut Gastric Ulcer. Eur JGen Me.
Dhalimi, A. 2006. Permasalahan Gambir(Uncaria gambir.) di Sumatera Barat danAlternatif Pemecahannya. Perspektif. .
Domingues, A.,A. Sartori., M.A. Golim., L.M.Valente., L.C. da Rosa., L.LW. Ishikawa.,A.C. Siani and R.M. Viero. 2011.Prevention of experimental diabetes byUncaria tomentosa extract: Th2polarization, regulatory T cell preservationor both? Journal of Ethnopharmacology.
Goodman and Gilman. 2012. DasarFarmakologi Terapi Vol 1 Ed 10. EGC.Jakarta
Guyton, A. C., and J. E. Hall. 2007. Buku AjarFisiologi Kedokteran Ed 11. EGC. Jakarta.

Indah Fajarwati, Efrizal dan Resti Rahayu 133
Hoerudin. 2012. Indeks Glikemik Buah danImplikasinya dalam Pengendalian KadarGlukosa Darah. Buletin TeknologiPascapanen Pertanian
Idrus, R. K. 2012. Tren PerkembanganKomoditi Ungulan Perkebunan Rakyat DiSumatera Barat. Jurnal Ekonomi STIE HajiAgus Salim Bukittinggi
Jerald, E.,S.B. Joshi., D.C. Jain. 2008. Diabetesand Herbal Medicines. Iranian Journal OfPharmacology & Therapeutics.
Jing, L., G. Cui., Q. Feng., F.Xiao. 2009.Evaluation of Hypoglycemic Activity ofthe Polysaccharides Extracted fromLycium Barbarum. Afr.J.Trad.
Karasov, W.H., M.W. Meyer, and B.W. Darken.1992. Tannic acid inhibition of amino acidandsugar absorption by mouse and voleintestine: Tests following acute andsubchronic exposure. Journal of ChemicalEcology
King, A. 2012. The use of animal modelsindiabetes research. British Journal ofPharmacology.
Laus, G. 2004. Advances in Chemistry andBioactivity of the Genus Uncaria. Phytoher.
Mutschler, E. 1991. Dinamika Obat. PenerbitITB. Bandung.
Nishikawa, T and E. Araki. 2013. Mechanism-based antioxidant therapies promise to
prevent diabetic complications?Journal ofDiabetes Investigation.
Padilah, I. 2009. Uji Pengaruh HipoglikemiaFraksi Etil Asetat Biji Jinten Hitam(Nigella sativa Linn) Pada Tikus PutihJantan Dengan Metode Induksi AloksanDan Toleransi Glukosa. Skripsi UniversitasIslam Negeri (Uin) Syarif Hidayatullah.Jakarta.
Pujilestari, B.I. dan R. Pratiwi. 2009.Pemanfaatan Tanaman Brotowali(Tinospora Crispa L.)Sebagai Antidiabetik.Prosiding Biteknologi Fakultas BiologiUniversitas Gadjah Mada.
Szkudelski. 2001. The Mechanism of Aloksanand Streptozotocin Action in B Cells of theRat Pancreas. MinireviewDepartment ofAnimal Physiology and Biochemistry,University of Agriculture, Poznan, Polan.
Welsch, C.A., P.A. Lachance, and B.P.Wasserman. 1989. Dietary phenoliccompounds: inhibition of Na+-dependentD-glucose uptake in rat intestinal brushborder membrane vesicles. Journal ofNutrition
WHO Country and Regional Data: World. 2009.http://www.who.int/diabetes/facts/world_figures/en/index.html. Diakses 2 Februari2013
Widowati, W. 2008. Potensi Antioksidansebagai antidiabetes. JKM.

ISSN 978-602-14989-0-3 BioETI
Inventarisasi jamur tingkat tinggi (Basidiomycetes) di GunungSinggalang Sumatera BaratINDRA ANGGRIAWAN, PERIADNADI* DAN NURMIATI
Labor Riset Mikrobiologi Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRACTThe macrofungi of Basidiomycetes of Singgalang Moutain was collected and identificated of Microbiology Laboratory,Department of Biology, Faculty of Sciences, Andalas University. The aim of this study was to identify the macrofungifrom Basidiomycetes. This study used direct survey to collect macrofungi from the field. This study identified 22 generawhich belong to 10 family and 6 orders. Most of the fungi were belong to subclass of Hymenomycetidae.
Key words: basidiomycetes, inventarisation, macrofungi.
Pendahuluan
Indonesia dengan 129 gunung merupakannegara terkaya akan keberadaan gunung(Sudradjat, 2011), salah satunya GunungSinggalang. Gunung Singgaalang termasuk kedalam Cagar Alam Singgalang Tandikat,terletak pada tiga kabupaten yaitu Tanah Datar,Agam dan Padang Pariaman. GunungSinggalang yang memiliki ketinggian 2.877mdpl merupakan gunung api yang sudah tidakaktif lagi (Anonimous, 2002). GunungSinggalang memiliki kelembaban relativetinggi, dan tersedianya nutrisi dari serasah dankayu mati sehingga hal ini menjadi suatuindikasi baik untuk pertumbuhan jamur tingkattinggi (Suriawiria, 1986). Jamur merupakanorganisme yang memegang peranan pentingdalam penguraian unsur-unsur alam (Armawi,2009). Jamur Basidiomycetes merupakanpengurai utama dari serasah daun dan kayu-kayu mati di hutan (Musyafa, 2005).
Jamur memiliki peran penting dalam siklusbiogeokimia tanah, siklus hara, dekomposer,fungi simbion pada tanaman yang bersifatsaling menguntungkan atau bersifat merugikansebagai parasit tumbuhan. Dalam ekosistemhutan siklus hara akan terhambat jika serasahtidak terurai dengan baik. Proses penguraiandilakukan oleh enzim yang terdapat pada
miselium jamur (Musyafa, 2005; Hesti, 2010;Okabe dan Thompson, 2010).
Kelas Basidiomycetes sering disebut jamurtingkat tinggi karena jamur ini lebih maju darikelas lainnya karena dilihat dari strukturnyayang sudah lengkap dan jelas terlihat bagian-bagiannya seperti caps, hymenium, stipe, ringdan volva (Suriawiria, 1986; Alexopoulos,1962). Selain itu suatu jamur dikelompokkanke dalam jamur tingkat tinggi karena tidak adafase motil, sedangkan jamur tingkat rendahmemiliki fase motil (Alexopoulos dan Mims,1979). Penelitian ini bertujuan meng-inventarisasi jamur Basidiomycetes yangterdapat di Gunung Singgalang.
BAHAN DAN METODE
Pengambilan sampel di Gunung Singgalangdengan menggunakan metode survey denganteknik kolesi langsung di lapangan. Sampelyang ditemukan difoto bagian-bagian yangmenjadi ciri pembeda dari jamur seperti : caps,hymenium, stipe, tempat tumbuh dan dicatatkarakter seperti : tipe caps, warna caps, warnahymenium, bentuk dan warna stipe, sertatempat tumbuh. Kemudian dilakukanidentifikasi di Labor Riset MikrobiologiJurusan Biologi, FMIPA, Universitas Andalas.dengan menggunakaan buku identifikasi jamur.
Analisis Data

Anggriawan, Periadnadi dan Nurmiati 135
Sampel yang didapatkan akan ditampilkandalam bentuk tabel karakter morfologipembeda masing-masing genus (Kibby, 1979;Polese,2000; Del Conte and Læssøe, 2008).
HASIL DAN PEMBAHASAN
Hasil dari penelitian yang telah dilakukan,didapatkan jamur Basidiomycetes sebanyak 22genus jamur yang terdiri dari 6 ordo dan 10famili (Tabel 1). Pada waktu pengambilansampel suhu berkisar dari 8o C – 27o C dengankelembaban 90-100 %. Menurut Suriawiria,(1986); Swapna et al., (2008), suhu dankelembaban menjadi faktor dalam keberadaandan pertumbuhan jamur tingkat tinggi.
Roth et al., (1990), membagiBasidiomycetes menjadi 3 sub-kelas yaitu
Phragmobasidiomycetidae, Hymenomycetidaedan Gasteromycetidae. Pembagian iniberdasarkan sporanya, spora pada Phragmo-basidiomycetidae bersegmen. Kibby (1979),menambahkan bahwa Phragmobasidio-mycetidae memiliki caps bergelatin, lunak danseperti karet. Hymenomycetidae danGasteromycetidae dibedakan berdasarkanposisi sporanya, pada Hymenomycetidaesporanya terdapat diluar yaitu padahymeniumnya (gills, pori dan seperti duri),sedangkan pada Gasteromycetidae sporanyaterdapat di dalam caps yang memiliki rongga.Gasteromycetidae kebanyakan jamur yangbulat atau seperti bola, biasa dikenal jamurpuffball (Roth, Frank dan Kormann, 1990).
Tabel 1. Klasifikasi Jamur Basidiomycetes di Gunung Singgalang Sumatera BaratNo. GENUS FAMILI ORDO SUB-CLASS1 Lycoperdon Lycoperdaceae Lycoperdales Gasteromycetidae2 Ganoderma Ganodermataceae Aphyllophorales Hymenomycetidae3 Fomes Polyporaceae Aphyllophorales Hymenomycetidae4 Meripilus Polyporaceae Aphyllophorales Hymenomycetidae5 Microporus Polyporaceae Aphyllophorales Hymenomycetidae6 Phaeolus Polyporaceae Aphyllophorales Hymenomycetidae7 Polyporus Polyporaceae Aphyllophorales Hymenomycetidae8 Trametes Polyporaceae Aphyllophorales Hymenomycetidae9 Stereum Stereaceae Aphyllophorales Hymenomycetidae
10 Boletus Boletaaceae Boletales Hymenomycetidae11 Cortinarius Cortinariaceae Cortinariales Hymenomycetidae12 Galerina Cortinariaceae Cortinariales Hymenomycetidae13 Inocybe Cortinariaceae Cortinariales Hymenomycetidae14 Hygrophorus Hygroporaceae Tricholomatales Hymenomycetidae15 Strobilurus Strobilomycetaceae Tricholomatales Hymenomycetidae16 Collybia Tricholomataceae Tricholomatales Hymenomycetidae17 Laccaria Tricholomataceae Tricholomatales Hymenomycetidae18 Lichenomphalia Tricholomataceae Tricholomatales Hymenomycetidae19 Marasmius Tricholomataceae Tricholomatales Hymenomycetidae20 Panellus Tricholomataceae Tricholomatales Hymenomycetidae21 Pleurotus Tricholomataceae Tricholomatales Hymenomycetidae22 Auricularia Auriculariaceae Auriculariales Phragmobasidiomycetidae
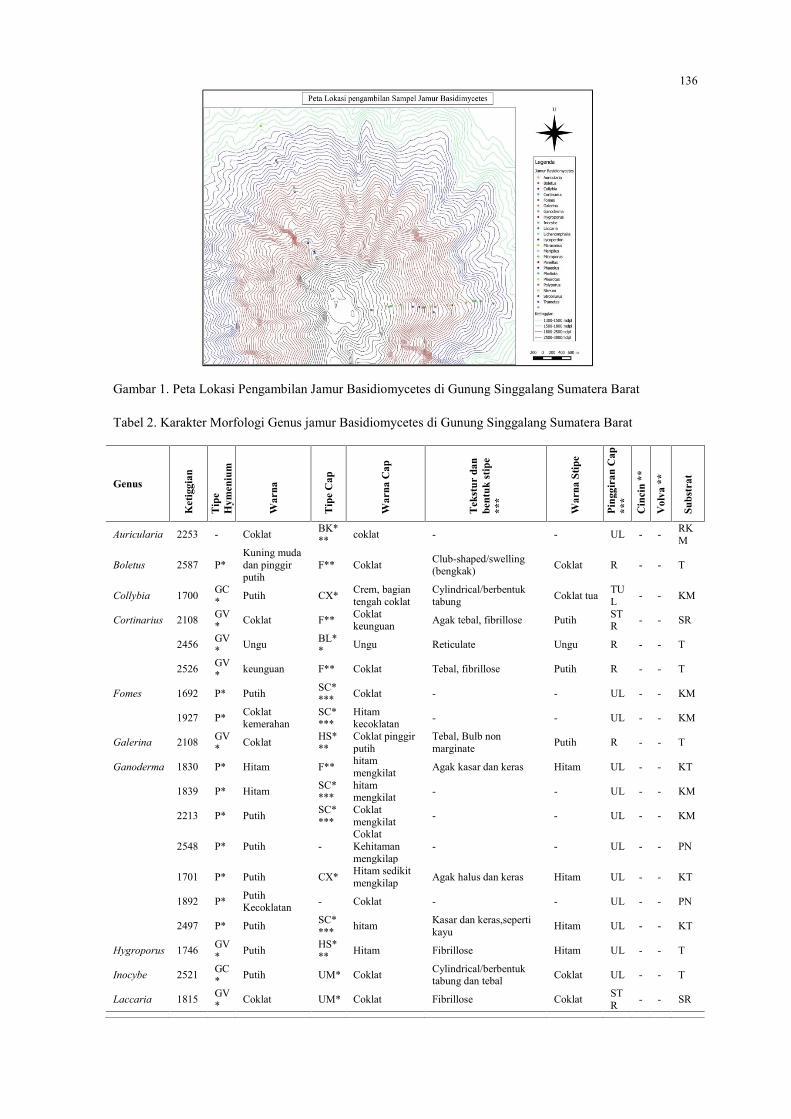
136
Gambar 1. Peta Lokasi Pengambilan Jamur Basidiomycetes di Gunung Singgalang Sumatera Barat
Tabel 2. Karakter Morfologi Genus jamur Basidiomycetes di Gunung Singgalang Sumatera Barat
Genus
Ket
iggi
an
Tip
eH
ymen
ium
War
na
Tip
e C
ap
War
na C
ap
Tek
stur
dan
bent
uk st
ipe
***
War
na S
tipe
Ping
gira
n C
ap**
*C
inci
n**
Vol
va**
Subs
trat
Auricularia 2253 - Coklat BK*** coklat - - UL - - RK
M
Boletus 2587 P*Kuning mudadan pinggirputih
F** Coklat Club-shaped/swelling(bengkak) Coklat R - - T
Collybia 1700 GC* Putih CX* Crem, bagian
tengah coklatCylindrical/berbentuktabung Coklat tua TU
L - - KM
Cortinarius 2108 GV* Coklat F** Coklat
keunguan Agak tebal, fibrillose Putih STR - - SR
2456 GV* Ungu BL*
* Ungu Reticulate Ungu R - - T
2526 GV* keunguan F** Coklat Tebal, fibrillose Putih R - - T
Fomes 1692 P* Putih SC**** Coklat - - UL - - KM
1927 P* Coklatkemerahan
SC****
Hitamkecoklatan - - UL - - KM
Galerina 2108 GV* Coklat HS*
**Coklat pinggirputih
Tebal, Bulb nonmarginate Putih R - - T
Ganoderma 1830 P* Hitam F** hitammengkilat Agak kasar dan keras Hitam UL - - KT
1839 P* Hitam SC****
hitammengkilat - - UL - - KM
2213 P* Putih SC****
Coklatmengkilat - - UL - - KM
2548 P* Putih -CoklatKehitamanmengkilap
- - UL - - PN
1701 P* Putih CX* Hitam sedikitmengkilap Agak halus dan keras Hitam UL - - KT
1892 P* PutihKecoklatan - Coklat - - UL - - PN
2497 P* Putih SC**** hitam Kasar dan keras,seperti
kayu Hitam UL - - KT
Hygroporus 1746 GV* Putih HS*
** Hitam Fibrillose Hitam UL - - T
Inocybe 2521 GC* Putih UM* Coklat Cylindrical/berbentuk
tabung dan tebal Coklat UL - - T
Laccaria 1815 GV* Coklat UM* Coklat Fibrillose Coklat ST
R - - SR
7
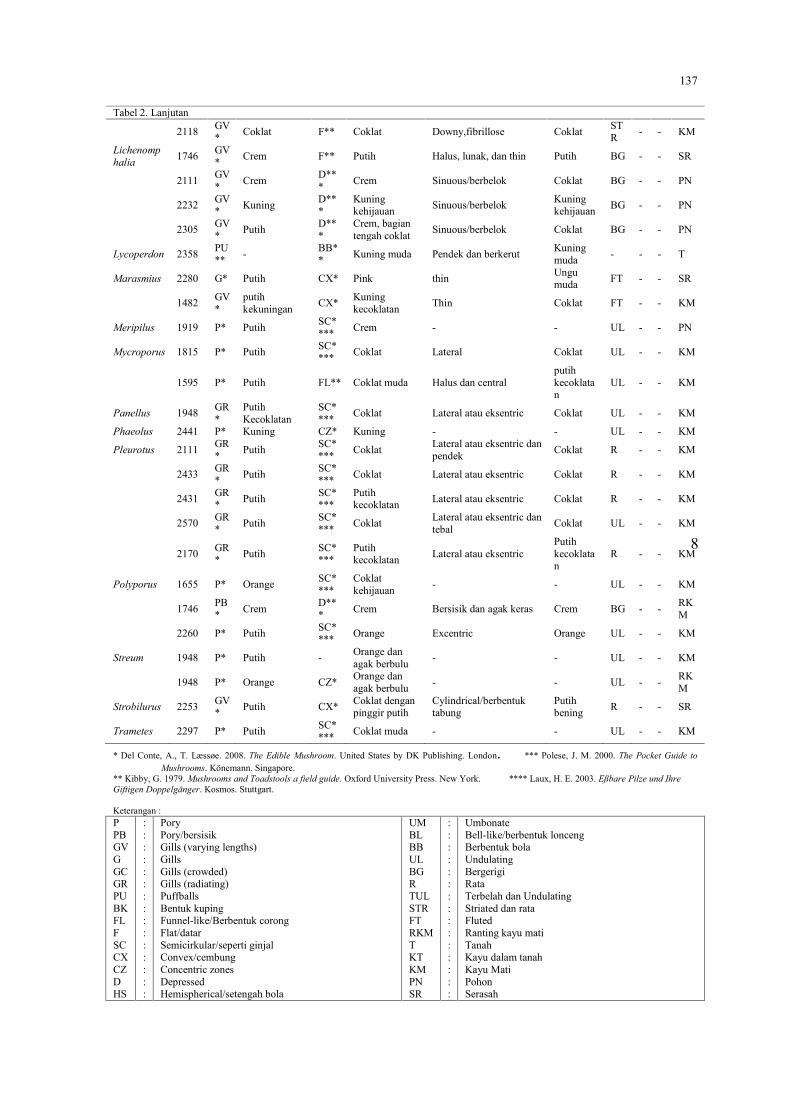
137
Tabel 2. Lanjutan
2118 GV* Coklat F** Coklat Downy,fibrillose Coklat ST
R - - KM
Lichenomphalia 1746 GV
* Crem F** Putih Halus, lunak, dan thin Putih BG - - SR
2111 GV* Crem D**
* Crem Sinuous/berbelok Coklat BG - - PN
2232 GV* Kuning D**
*Kuningkehijauan Sinuous/berbelok Kuning
kehijauan BG - - PN
2305 GV* Putih D**
*Crem, bagiantengah coklat Sinuous/berbelok Coklat BG - - PN
Lycoperdon 2358 PU** - BB*
* Kuning muda Pendek dan berkerut Kuningmuda - - - T
Marasmius 2280 G* Putih CX* Pink thin Ungumuda FT - - SR
1482 GV*
putihkekuningan CX* Kuning
kecoklatan Thin Coklat FT - - KM
Meripilus 1919 P* Putih SC**** Crem - - UL - - PN
Mycroporus 1815 P* Putih SC**** Coklat Lateral Coklat UL - - KM
1595 P* Putih FL** Coklat muda Halus dan centralputihkecoklatan
UL - - KM
Panellus 1948 GR*
PutihKecoklatan
SC**** Coklat Lateral atau eksentric Coklat UL - - KM
Phaeolus 2441 P* Kuning CZ* Kuning - - UL - - KM
Pleurotus 2111 GR* Putih SC*
*** Coklat Lateral atau eksentric danpendek Coklat R - - KM
2433 GR* Putih SC*
*** Coklat Lateral atau eksentric Coklat R - - KM
2431 GR* Putih SC*
***Putihkecoklatan Lateral atau eksentric Coklat R - - KM
2570 GR* Putih SC*
*** Coklat Lateral atau eksentric dantebal Coklat UL - - KM
2170 GR* Putih SC*
***Putihkecoklatan Lateral atau eksentric
Putihkecoklatan
R - - KM
Polyporus 1655 P* Orange SC****
Coklatkehijauan - - UL - - KM
1746 PB* Crem D**
* Crem Bersisik dan agak keras Crem BG - - RKM
2260 P* Putih SC**** Orange Excentric Orange UL - - KM
Streum 1948 P* Putih - Orange danagak berbulu - - UL - - KM
1948 P* Orange CZ* Orange danagak berbulu - - UL - - RK
M
Strobilurus 2253 GV* Putih CX* Coklat dengan
pinggir putihCylindrical/berbentuktabung
Putihbening R - - SR
Trametes 2297 P* Putih SC**** Coklat muda - - UL - - KM
* Del Conte, A., T. Læssøe. 2008. The Edible Mushroom. United States by DK Publishing. London. *** Polese, J. M. 2000. The Pocket Guide toMushrooms. Kӧnemann. Singapore.
** Kibby, G. 1979. Mushrooms and Toadstools a field guide. Oxford University Press. New York. **** Laux, H. E. 2003. Eßbare Pilze und IhreGiftigen Doppelgänger. Kosmos. Stuttgart.
Keterangan :PPBGVGGCGRPUBKFLFSCCXCZDHS
:::::::::::::::
PoryPory/bersisikGills (varying lengths)GillsGills (crowded)Gills (radiating)PuffballsBentuk kupingFunnel-like/Berbentuk corongFlat/datarSemicirkular/seperti ginjalConvex/cembungConcentric zonesDepressedHemispherical/setengah bola
UMBLBBULBGRTULSTRFTRKMTKTKMPNSR
:::::::::::::::
UmbonateBell-like/berbentuk loncengBerbentuk bolaUndulatingBergerigiRataTerbelah dan UndulatingStriated dan rataFlutedRanting kayu matiTanahKayu dalam tanahKayu MatiPohonSerasah
8

138
Auricularia Boletus Collybia Cortinarius Fomes Galerina
Ganoderma Hygroporus Inocybe Laccaria Lichenomphalia Lycoperdon
Marasmius Meripilus Microporus Panellus Phaeolus
Pleurotus Polyporus Stereum Strobilurus Trametes

139
Jamur yang sering ditemukan di GunungSinggalang yaitu dari sub-kelasHymenomycetidae, sedangkan dari sub-kelasPhragmobasidiomycetidae dan Gastero-mycetidae ditemukan 1 genus.Hymenomycetidae banyak ditemukan karenaterdiri dari genus yang lebih banyak sertakondisi lingkungan yang cocok untukpertumbuhan jamur dari sub-kelas ini. JamurBasidiomycetes di Gunung Singgalang iniditemukan pada ketinggian 1482 mdpl – 2570mdpl. Kondisi lingkungan dengan terdapatnyabanyak pohon-pohon besar serta serasah dankayu atau ranting kayu yang mati menjadisumber nutrisi bagi pertumbuhan jamurBasidiomycetes (Syriawiria, 1986).
KESIMPULAN
Berdasarkan hasil penelitan yang telahdilakukan di Gunung Singgalang SumateraBarat didapatkan jamur Basidiomycetessebanyak 22 genus yang merupakan bagian dari10 famili dan 6 ordo. Jamur yang seringditemukan yaitu dari sub-kelas Hymeno-mycetidae sebanyak 20 genus.
DAFTAR PUSTAKA
Bloom Anonimous. 2002. Rencana PengelolaanCagar Alam Singgalang Tandikat PropinsiSumatera Barat. Balai KSDA SumateraBarat. Padang.
Alexopoulos, C. J. 1962. Introductory Mycologysecond edition. John Wiley & Sons, Inc.New York.
Alexopoulos, C. J., and C. W. Mims. 1979.Introductory Mycology third edition. JohnWiley and Sons, Inc. New York.
Armawi. 2009. Pengaruh Tingkat KemasakanBuah Kelapa dan Konsentrasi Air Kelapapada Media Tanam TerhadapPertumbuhan Jamur Tiram Putih(Pleurotus ostreatus). Universitas IslamNegeri. Malang.
Del Conte, A., and T. Læssøe. 2008. The EdibleMushroom. United States by DKPublishing. London.
Fergus, C. L. 1960. Illustrated Genera of WoodDecay Fungi. Burgess PublishingCompany. America.
Gem, C. 1999. Mushrooms and Toadstools.Harper Collins Publisher. Itali.
Gerhardt, E. 2000. Pilze, mit Schnellbestimm-System. BLV Verlagsgessellschaft mbH.München Wien Zürich
Kibby, G. 1979. Mushrooms and Toadstools afield guide. Oxford University Press. NewYork.
Laux, H. E. 2003. Eßbare Pilze und IhreGiftigen Doppelgänger. Kosmos. Stuttgart.
Musyafa. 2005. Peranan Makrofauna Tanahdalam Proses Dekomposisi Serasah Acaciamangium willd. Biodiversitas. 6(1) : 63-65
Polese, J. M. 2000. The Pocket Guide toMushrooms. Kӧnemann. Singapore.
Roth, L., H. Frank and K. Kormann. 1990.Giftpilze-pilsgifte. Nicol. Hamburg.
Sudradjat, A. 2011. Gunung Api Di TatarSunda : Antisipasi Hidup di DaerahBencana. Konferensi Internasional BudayaSunda II. Jakarta.
Suriawiria, U. 1986. Pengantar Untuk Mengenaldan Menanam Jamur. Angkasa. Bandung.
Swapna, S., S. Abrar and M. Krishnappa. 2008.Diversity of Macrofungi in Semi-Evergreen and Moist Deciduous Forest ofShimoga Distric-Karnataka India. JournalMycology Plant Pathology. 38(1) : 21-26
Zoberi, M. H. 1972. Tropical Macrofungi.Macmillan. London.

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman dan penyebaran kerang (pelecypoda)di perairan Tanjung Mutiara Danau Singkarak Sumatera BaratIZMIARTI, JABANG NURDIN, MISREN AHYUNI DAN DEA RAHAYU SILVIANI
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian tentang keanekaragaman dan penyebaran kerang (Pelecypoda) di perairan Tanjung Mutiara Danau SingkarakSumatera Barat telah dilakukan bulan Juni 2013. Penelitian dilakukan dengan metode survey dan teknik pengambilansampel purposive stratified sampling. Sampel diambil pada tiga stasiun, masing-masing stasiun dibagi atas tiga stratakedalaman <5m, 5-10m, >10-15m, masing-masing strata dikoleksi 3 sampel dengan menggunakan alat penangkap kerangyang digunakan penduduk (dauah). Kerang air tawar yang ditemukan di Tanjung Mutiara Danau Singkarak sebanyak 3jenis yaitu Rectidens sumatraensis, Contradens ascia dan Corbicula sumatrana dengan indeks keanekaragaman yangrendah (0,25). Kepadatan total masing-masing jenis diketiga stasiun berkisar dari 4.75 - 452, 33 ind/m2 yangdidominasi oleh Corbicula sumatarana sebesar 98,18%. Ketiga jenis kerang ditemukan pada ketiga stasiun tetapipenyebarannya tidak diseluruh kedalaman. R. sumatraensis dan C. ascia terkonsentrasi pada kedalaman 5-15 ind/m2
sedangkan C. sumatrana cenderung tersebar diseluruh kedalaman dengan kepadatan yang lebih tinggi dari dua jenislainnya.
Key words: keanekaragaman, penyebaran, kerang, Danau Singkarak
Pendahuluan
Sumatera Barat mempunyai beberapa buahdanau yang cukup luas dan dalam, salah satunyaadalah Danau Singkarak dengan luas permukaan13.011 ha dan kedalaman maksimum 268 m(PSLH, 1984). Danau ini berpotensi untukmenghasilkan ikan, sumber air minum, irigasi,tempat rekreasi dan untuk pembangkit tenagalistrik air. Dari segi ekositem Danau Singkarakmerupakan habitat bagi berbagai organisme,seperti plankton, bentos, ikan, dan kerang-kerangan. Kerang-kerangan (Kelas Pelecypoda)berperan sebagai kunsumen yangmengkonsumsi organisme kecil lainnya dengancara menyaring (filter feeder).
Keberadaan kerang dapat mengambarkankondisi lingkungan sekitarnya karena itu dandapat dijadikan sebagai indikator (Abaychi andMustafa, 1988). Kerang banyak ditangkap dandimakan oleh masyarakat untuk memenuhikebutuhan akan protein hewani karenamengandung protein yang tinggi dan rasadagingnya enak.
Kerang air tawar menyukai air tenang danmengalir lamban, membenamkan diri dalamsubstrat berpasir dan berlumpur. Saat ini 37jenis kerang terancam punah karena kerusakanhabitat, penurunan kualitas air, pencemaranlimbah domestik dan pupuk dan eutrofikasi(Master et al., 2000 cit. Grabarkiewicz andWayne,2008). Penelitian tentang keberadaankerang air tawar di Sungai di Indoneia telahbanyak dilakukan seperti Junaidi, Sagala danSukotjo (2010) dan Ramadani (2011). Hasilpenelitian menunjukan bahwa pada perairanyang berbeda ditemukan jenis- jenis kerangyang berbeda dengan keanekaragaman yangrendah. Penelitian krang air tawar diperairantergenang masih sedikit informasinya.
Tanjung Mutiara terletak disebelah UtaraDanau Singkarak pada kordinat 0037’12”LS dan 100032’24” BT. Tanjung Mutiaramerupakan salah satu wilayah pariwisatasekaligus tempat pemukiman penduduk, namunpada lokasi tertentu ada yang tidak tersentuholeh aktifitas manusia termasuk penangkapankerang karena dasar perairan lebih curam daripada daerah lainnya. Pada umumnya masyarakat

Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 141
yang tinggal didaerah T. Mutiara ini menangkapkerang sebagai mata pencaharian dan untukdikonsumsi sendri. Penangkapan kerang yangtidak terkendali dan kerusakan habitat atauperubahan kualitas air tempat hidupnya akibatberbagai aktivitas manusia disekitar perairanakan menjadi ancaman terhadapkeanekaragaman jenis dan populasi kerang dialam. Sehubungan dengan itu dilakukanpenelitian dengan tujuan untuk mengetahuikeanekargaman dan penyebaran kerangPelecypoda di perairan T. Mutiara DanauSingkarak. Kajian ini penting sebagai langkahawal untuk usaha konservasi kerang di DanauSingkarak.
BAHAN DAN METODE
Penelitian ini dilaksanakan pada bulan Juni 2013.Lokasi penelitian adalah perairan danau didaerah Tanjung Mutiara, Kecamatan Batipuh,Kabupaten Tanah Datar, Provinsi SumateraBarat. Pengerjaan sampel lebih lanjut dilakukandi Laboratorium Riset Ekologi, Jurusan Biologi,FMIPA, Universitas Andalas, Padang.
Penelitian ini dilakukan dengan metodesurvey dengan teknik pengambilan sampelpurposive stratified sampling. Sampel diambilpada tiga stasiun, dan setiap stasiun dibagi atastiga strata kedalaman <5m, 5-10m, >10-15m.Pada setiap strata dikoleksi tiga sampel denganmenggunakan alat penangkap kerang yangdigunakan penduduk (dauah). Stasiun I terletakpada perairan dekat pemukiman dan banyakaktifitas penduduk, Stasiun II lokasi wisata danStasiun III tidak ada aktivitas penduduk. PadaStasiun I dan II penduduk melakukanpenangkapan kerang, tidak demikian halnyapada stasiun III. Pada setiap stasiun dankedalaman dilakukan pengukuran faktor fisikakimia air meliputi: suhu air, kecerahan air, pH,O2 terlarut, CO2 bebas, BOD, TSS, KO substratdan komposisi partikel substrat. Dilaboratoriumdilakukan identifikasi jenis kerang danpenghitungan jumlah individu masing-masingjenis. Untuk identifikasi jenis kerang digunakan
buku acuan yang terkait Analisis data dilakukanterhadap kepadatan populasi yang dinyatakandengan jumlah ind/m2. Indeks keanekaragamanjenis dihitung dengan menggunakan Indekskeanekaragaman Shannon- wiener.
HASIL DAN PEMBAHASAN
Berdasarkan penelitian yang telah dilakukandidapatkan hasil sebagai berikut:1. Keanekaragaman jenis kerangDari hasil penelitian yang telah dilakukan diTanjung Mutiara Danau Singkarak didapatkan 3jenis kerang yaitu Rectidens sumatraensis,Contradens ascia. Keduanya tergolog familiUnionidae dan Corbicula sumatrana dari familiCorbiculidae. Ketiga jenis tersebut dapat dilihatpada Gambar 1. Indeks keanekaragaman jeniskerang tergolong rendah (H’ = 0,25). MenurutEstradivani dkk., (2007) bahwa indekskeanekaragaman jenis dapat dikelompokan atas3 yaitu kenekaragam tergolong rendah apabila H’< 2, sedang 2<H’<3 dan tinggi bila H”>3.Kerang Rectidens sumatraensis dan Contradensascia termasuk kerang air tawar yang terancampunah karena pelumpuran akibat penggundulanhutan (Ubaidillah dkk., 2013)
Secara keseluruhan komunitas kerang diTanjung Mutiara Danau Singkarak didominasioleh Corbicula sumatrana dengan persentase98,18 %, yang paling sedikit ditemukan adalahRectidens sumatraensis hanya sebesar 0,67 %.Salah satu faktor yang mempengaruhi distribusidan kelimpahan Corbicula adalah tipe substratSubstrat yang cocok untuk kerang ini adalahpasir bercampur lumpur dan lempung (Karatajevet al., 2003). Faktor lain yang menyebabkantingginya populasi Corbicula sumatranadibandingkan dua jenis lainnya mungkinberkaitan dengan karakter biologinya. MenurutHubenov et al., (2013) beberapa karakter biologiyang menentukan kelimpahan Corbiculafluminea adalah pertumbuhan yang cepat,matang seksual lebih awal, fekunditasnya tinggi,strategi reproduksinya yang beragam.

Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 142
Rectidens sumatraensis
Contradens ascia
Corbicula sumatrana
Gambar 1. Foto jenis kerang (Pelecypoda) diTanjung Mutiara Danau Singkarak
Sebaran kerang Pelecypoda di Tanjung MutiaraDanau singkarakDari Tabel 2 diatas dapat dilihat bahwaKepadatan kerang rata-rata 51,72 ind/m2,masing-masing jenis berkisar dari 0,53 – 50,26ind/m2. Pada Stasiun III diperoleh kepadatanpopulasi yang paling tinggi diantara ketigastasiun. Hal ini disebabkan karena pada stasiunini tidak ada aktivitas penangkapan kerang danaktivitas lainnya. Tidak adanya aktifitaspenangkapan kerang di Stasiun III karena dasar
perairan yang relatif curam, dengan demikianmemberikan kesempatan pada kerangberkembang lebih baik pada stasiun tersebut.Pada Stasiun II dan III terjadi aktivitaspenangkapan kerang yang terus menerus,akibatnya dapat menurunkan populasi kerang.
Jenis R. sumatraensis dan C. asciaditemukan pada ketiga stasiun cendrung tersebarpada kedalaman 5-15 m, sedangkan C.sumatrana tersebar hampir diseluruhkedalaman dengan kepadatan yang jauh lebihtinggi dibandingkan dengan 2 jenis lainnya.Pada kedalaman <5 m pada ketiga stasiun tidakditemukan R. sumtraensis dan C. ascia. Tidakditemukannya kedua jenis kerang tersebut dikedalaman <5m berkaitan dengan substrat yangtidak sesuai untuk kehidupan kerang. Substratdasar pada kedalaman <5 m sebagian besarterdiri dari batu dan kerikil, kurang cocok untukkehidupan kerang ini. Selain itu strata II teletakpada bagian pinggir danau relatif dangkalsehingga sering dilakukan penangkapan kerangoleh penduduk dan terganggu oleh aktivitaslainnya.
C. sumatrana tersebar disetiap kedalamanyang diamati bahkan dengan kepadatan yangjauh lebih tinggi dari kedua jenis yang lainnya.Spesies yang berbeda membutuhkan persyaratansubstrat yang berbeda pula. Junaidi (2010)menemukan sejenis Corbicula yang hidup diSungai Borang Banyuasin lebih banyak padastasiun yang mempunyai substrat kasar sepertipasir dan kerikil atau campuran pasir denganmaterial lain. Kerang Corbiculidae umumnyamenyukai substrat berlumpur dengan sedikitpasir (Pennak, 1978), Substrat yang terdiri darilumpur lempung dapat menyumbat insangkerang menghambat pernafasan, pencernaan danreproduksi (Aldridge, 1987). Menurut Hubenovet al. (2013) C. fluminea yang hidup di sungaidapat ditemukan pada substrat yang bervariasiyaitu pasir, pasir kasar, kerikil dan batu. Selainitu dapat pula ditemukan pada substrat yangterdiri dari pasir, kerikil, lempung dan lumpur,namun kelimpahan yang paling tinggiditemukan pada substrat yang didominasi oleh
Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 142
Rectidens sumatraensis
Contradens ascia
Corbicula sumatrana
Gambar 1. Foto jenis kerang (Pelecypoda) diTanjung Mutiara Danau Singkarak
Sebaran kerang Pelecypoda di Tanjung MutiaraDanau singkarakDari Tabel 2 diatas dapat dilihat bahwaKepadatan kerang rata-rata 51,72 ind/m2,masing-masing jenis berkisar dari 0,53 – 50,26ind/m2. Pada Stasiun III diperoleh kepadatanpopulasi yang paling tinggi diantara ketigastasiun. Hal ini disebabkan karena pada stasiunini tidak ada aktivitas penangkapan kerang danaktivitas lainnya. Tidak adanya aktifitaspenangkapan kerang di Stasiun III karena dasar
perairan yang relatif curam, dengan demikianmemberikan kesempatan pada kerangberkembang lebih baik pada stasiun tersebut.Pada Stasiun II dan III terjadi aktivitaspenangkapan kerang yang terus menerus,akibatnya dapat menurunkan populasi kerang.
Jenis R. sumatraensis dan C. asciaditemukan pada ketiga stasiun cendrung tersebarpada kedalaman 5-15 m, sedangkan C.sumatrana tersebar hampir diseluruhkedalaman dengan kepadatan yang jauh lebihtinggi dibandingkan dengan 2 jenis lainnya.Pada kedalaman <5 m pada ketiga stasiun tidakditemukan R. sumtraensis dan C. ascia. Tidakditemukannya kedua jenis kerang tersebut dikedalaman <5m berkaitan dengan substrat yangtidak sesuai untuk kehidupan kerang. Substratdasar pada kedalaman <5 m sebagian besarterdiri dari batu dan kerikil, kurang cocok untukkehidupan kerang ini. Selain itu strata II teletakpada bagian pinggir danau relatif dangkalsehingga sering dilakukan penangkapan kerangoleh penduduk dan terganggu oleh aktivitaslainnya.
C. sumatrana tersebar disetiap kedalamanyang diamati bahkan dengan kepadatan yangjauh lebih tinggi dari kedua jenis yang lainnya.Spesies yang berbeda membutuhkan persyaratansubstrat yang berbeda pula. Junaidi (2010)menemukan sejenis Corbicula yang hidup diSungai Borang Banyuasin lebih banyak padastasiun yang mempunyai substrat kasar sepertipasir dan kerikil atau campuran pasir denganmaterial lain. Kerang Corbiculidae umumnyamenyukai substrat berlumpur dengan sedikitpasir (Pennak, 1978), Substrat yang terdiri darilumpur lempung dapat menyumbat insangkerang menghambat pernafasan, pencernaan danreproduksi (Aldridge, 1987). Menurut Hubenovet al. (2013) C. fluminea yang hidup di sungaidapat ditemukan pada substrat yang bervariasiyaitu pasir, pasir kasar, kerikil dan batu. Selainitu dapat pula ditemukan pada substrat yangterdiri dari pasir, kerikil, lempung dan lumpur,namun kelimpahan yang paling tinggiditemukan pada substrat yang didominasi oleh
Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 142
Rectidens sumatraensis
Contradens ascia
Corbicula sumatrana
Gambar 1. Foto jenis kerang (Pelecypoda) diTanjung Mutiara Danau Singkarak
Sebaran kerang Pelecypoda di Tanjung MutiaraDanau singkarakDari Tabel 2 diatas dapat dilihat bahwaKepadatan kerang rata-rata 51,72 ind/m2,masing-masing jenis berkisar dari 0,53 – 50,26ind/m2. Pada Stasiun III diperoleh kepadatanpopulasi yang paling tinggi diantara ketigastasiun. Hal ini disebabkan karena pada stasiunini tidak ada aktivitas penangkapan kerang danaktivitas lainnya. Tidak adanya aktifitaspenangkapan kerang di Stasiun III karena dasar
perairan yang relatif curam, dengan demikianmemberikan kesempatan pada kerangberkembang lebih baik pada stasiun tersebut.Pada Stasiun II dan III terjadi aktivitaspenangkapan kerang yang terus menerus,akibatnya dapat menurunkan populasi kerang.
Jenis R. sumatraensis dan C. asciaditemukan pada ketiga stasiun cendrung tersebarpada kedalaman 5-15 m, sedangkan C.sumatrana tersebar hampir diseluruhkedalaman dengan kepadatan yang jauh lebihtinggi dibandingkan dengan 2 jenis lainnya.Pada kedalaman <5 m pada ketiga stasiun tidakditemukan R. sumtraensis dan C. ascia. Tidakditemukannya kedua jenis kerang tersebut dikedalaman <5m berkaitan dengan substrat yangtidak sesuai untuk kehidupan kerang. Substratdasar pada kedalaman <5 m sebagian besarterdiri dari batu dan kerikil, kurang cocok untukkehidupan kerang ini. Selain itu strata II teletakpada bagian pinggir danau relatif dangkalsehingga sering dilakukan penangkapan kerangoleh penduduk dan terganggu oleh aktivitaslainnya.
C. sumatrana tersebar disetiap kedalamanyang diamati bahkan dengan kepadatan yangjauh lebih tinggi dari kedua jenis yang lainnya.Spesies yang berbeda membutuhkan persyaratansubstrat yang berbeda pula. Junaidi (2010)menemukan sejenis Corbicula yang hidup diSungai Borang Banyuasin lebih banyak padastasiun yang mempunyai substrat kasar sepertipasir dan kerikil atau campuran pasir denganmaterial lain. Kerang Corbiculidae umumnyamenyukai substrat berlumpur dengan sedikitpasir (Pennak, 1978), Substrat yang terdiri darilumpur lempung dapat menyumbat insangkerang menghambat pernafasan, pencernaan danreproduksi (Aldridge, 1987). Menurut Hubenovet al. (2013) C. fluminea yang hidup di sungaidapat ditemukan pada substrat yang bervariasiyaitu pasir, pasir kasar, kerikil dan batu. Selainitu dapat pula ditemukan pada substrat yangterdiri dari pasir, kerikil, lempung dan lumpur,namun kelimpahan yang paling tinggiditemukan pada substrat yang didominasi oleh

Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 143
pasir kasar dan pasir dan kerikil. Corbiculasumatrana di Tanjung Mutiara ditemukan padasubstrat kerikil, pasir dan lumpur, namun lebihbanyak ditemukan pada substrat lumpur danpasir. Kemampuan dari C. sumatrana mendiamihabitat dengan berbagai ukuran partikel substratmenyebabkan jenis kerang ini lebih dominandari 2 jenis kerang yang lainnya. Hasilpengukuran faktor fisika kimia air seperti suhu,oksigen terlarut, CO2, BOD, pH, TSS tidak jauhberbeda pada masing-masing stasiun dan masihberada dalam kisaran toleransi kerang untukhidup. Perbedaan hanya terlihat pada stasiun III,bahwa substrat dasar pada kedalaman 5-10 mdan >10-15 m komposisinya sebagian besarterdiri dari lumpur 71,08 -77,85 % dengankandungan organik substrat 2,90 – 6,01 %(Tabel 3). Pada stasiun ini ditemukankepadatan populasi C. sumatrana yang lebihtinggi dari pada dua jenis yang lainnya. Bahanorganik didalam substrat merupakan salah satusumber bahan makanan bagi kerang (Pennak,1978).
Perkembangbiakan Corbicula yang cepatmenyebabkan populasi kerang ini seringditemukan dalam jumlah besar, Hasil penelitianmenunjukan bahwa Corbicula memiliki 2periode spawning setiap tahun, dimanawaktunya berbeda dengan spesies yang berbeda.Corbicula malinensis Philippi periode spawningyang pertama April sampai Juli dan periodekedua Agustus sampai desember (Aldridge andMc Mahon, 1978). Corbicula flumineahermaprodit membebaskan larvaperdiveligernya bulan May-September denganpuncak spawningnya Mai/Juni dan September,sementara C. fluminalis dioceus (3%hermaprodit) membebaskan gametnya bulanOktober sampai Desember dan bulan Maretsampai April memperlihatkan puncak spawningOktober/Nopember dan Maret (Rajagopal et al.,2000). Hasil penelitian Mouthon andParghentanian (2004) menyatakan bahwa C.fluminea periode reproduksinya dimulai bulanMaret dan berakhir pada bulan September
Tabel 1. Indeks keanekaraman dan dominansijenis kerang Pelecypoda di TanjungMutiara di Danau Singkarak
Jenis Dominansi(%)
Indekskeanekaragaman
Rectidens sumatraensis 0.670.25Contradens ascia 1.15
Corbicula sumatrana 98.18
Tabel 2. Kepadatan populasi (ind/m2) kerangPelecypoda di Tanjung MutiaraDanau Singkarak
Jenis Stasiun I Stasiun II Stasiun III
Rat
a-ra
ta
<5 m
5-10
m
>10-
15m <5
m
5-10
m
>10-
15m <5
m
5-10
m
>10-
15m
Ract
iden
ssp
. 0 0,16 0,11 0,03 0,06 0,05 0 1,46 2,88 0,53
Con
trde
nsas
cia
0 0,32 0,14 0 0,09 0,07 0 2,18 5,61 0,93
Cor
bicu
lasu
mat
rana
5,02 55,19 28,09 0 10,73 12,34 79,50 88,73 172,73 50,26
Tota
lkep
adat
an(in
d/m
2 )
5,02 55,67 28,34 0 10,88 12,46 79,50 92,37 181,22 51,72
Tabel 3. Kondisi Fisika Kimia Air Perairan TanjungMutiara, Danau Singkarak
ParameterStasiun I Stasiun II Stasiun III
<5m 5-10m
>10-15m <5m 5-
10m>10-15m <5m 5-
10m>10-15m
Temperatur (oC)Kecerahan (m)
pHDO (ppm)
BOD5 (ppm)CO2
292,57
6,450,470,88
2838
5,940,531,65
253,27
5,701,730,88
292,77
7,980,830,88
2638
7,171,000,88
2447
6,971,101,32
271,88
7,980,430,88
252,58
7,370,570,88
232,77
6,910,770,88
Kadar Organik(%)
0 0 0 0 0 0 0 2,90 6,01
KomposisiSubstrat (%)Batu / Kerikil
PasirLumpur
77,922,10,0
78,421,60,0
71,6028,40
0,0
77,522,50,0
86,313,70,0
65,1234,880,0
40,259,80,0
0,0028,971,1
0,0022,1577,85
Keterangan :Stasiun I = perairan dekat pemukiman penduduk dan banyakaktivitas pendudukStasiun II = perairan yang di jadikan sebagai objek wisataStasiun III = perairan yang tidak ada aktivitas penduduk.
sampai Oktober dengan puncaknya pada bulanJuni dan Agustus. Corbicula fluminalis perodereproduksinya terjadi selama musim dingin danbulan Mai sampai Oktober. Puncak

Izmiarti, Jabang Nurdin, Misren Ahyuni dan Dea Rahayu Silviani 144
maksimumnya adalah bulan juni dan Juli.Namun periode reproduksi dari Corbiculasumatrana belum diketahui.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Kerang (Pelecypoda) yang ditemukan
sebanyak 3 jenis yaitu Rectidenssumatraensis, Contradens ascia (Unionidae)dan Corbicula sumatrana (Corbiculidae).Indeks keanekaragaman jenis kerang initergolong rendah ( H’ = 0,25). Komunitasdidominasi oleh Corbicula sumatranadengan persentase 98,18 %.
2. Kepadatan kerang rata-rata 51,72 ind/m2,masing-masing jenis berkisar dari 0,53 –50,26 ind/m2.
3. Ketiga jenis kerang ditemukan pada ketigastasiun tetapi penyebarannya tidak diseluruhkedalaman. Rectidens. sumatraensis danContradens. ascia tersebar pada kedalaman5-15 m sedangkan Corbicula. sumatranatersebar hampir diseluruh kedalaman dengankepadatan yang lebih tinggi dari dua jenislainnya.
3. Fisika kimia air disetiap stasiun dankedalaman tidak jauh berbeda danmendukung untuk kehidupan kerang
DAFTAR PUSTAKA
Abaychi, J. K. And Y.Z. Mustafa. 1988. TheAsiatic clam, Corbicula fluminea: AnIntroduction of metal pollution in the Shattal-Arab River, Iraq. Enviromental Pollution54(1):109-122
Aldridge, D.W. and R.F. McMahon .1978.Growth, fecundity and bioenergetics in anatural population of the aciatic freshwaterclam, Corbicula manilensis Philippi fromnorth Texas. Journal of Molluscan Studies.44 (1): 49-70
Djajasasmita, M. 1999. Keong dan kerangsawah. Puslitbang Biologi-LIPI. Bogor
Estradivani., Syahrir, M., Susilo, N.., Yusri, Sdan Timotius, S. 2007. Terumbu KarangJakarta. Laporan Pengamatan JangkaPanjang Terumbu Karang KepulauanSeribu 2004 – 2005. Yayasan TerumbuKarang Indonesia (Terangi). Jakarta
Grabarkiewicz, J.D dan S. D. Wayne. 2008. Anintroduction to freshwater Mussel asBiological Indicator: Including Account ofInterior Basin, Cumberlandian and AtlanticSlope spesies, US Enviromental ProtectionAgancy, Washington D.C.
Hubenov, Z.,. Trichkova, T.,. Kenderov, L andD. Kozuharov. 2013. Distribution ofCorbicula fluminea (Mollusca:Corbiculidae) over an Elaven-year Periodof its invation in Bulgaria. Acta zool. bulg.65 (3) : 315 - 326
Junaidi, E., E.P. Sagala dan Joko. 2010.Kelimpahan Populasi dan Pola DistribusiRemis (Corbicula sp.) di Sungai BorangKabupaten Banyuasin. Jurnal PenelitianSains. 13 (3D): 13310-50 – 13310-54
Karatajev, A.Y., Burlakova, L.E., Kesterson, Tand D.K. Padilla. 2003. Dominanceof theAsiatic clam, Corbicula fluminea (Muller)in the benthic community of a reservoir.Journal of Shellfish Research 22 (2). 487-493
Mouthon, J. and T. Parghentanian. 2004.Comparison of the life cycle andpopulation dinamic of two Corbiculaspecies, C. Fluminea and C. Fluminalis(Bivalva: Corbiculidae in two Frenchcanals. Archiv fur Hydrobiologie. 161 (2):267 – 287
Pennak, R. W. 1978. Freshwater Invertebratesof the United States. Second ed. JohnWiley and Sons. New York. Chichester.Brisbane. Toronto
Rajagopal, S., Van Der Velde, G.and A. Bij DeVaate, 2000. Reproductive biology of theAsiatic Clams Corbicula fluminalis danCorbicula fluminea in the River.Archiv furHydrobiologie 149: 403-420 .
Ramadani, A,H. 2011. Keanekaragaman danPola Distribusi Longitudinal Kerang AirTawar di Perairan Sungai Berantas. Skripsi.FSAINTEK. UNAIR. Surabaya
Ubaidillah, R., Marwoto R.M.., Hadiaty, R.K.,Fahmi.,Wowor D.,Mumpuni., Pratiwi, R.,Tjakrawidjaja, A. H., Mudjiono., HartatiS.T. Heryanto.,R R.M.iyanto, A., Mujiono,N. 2013. Biota Perairan Terancam Punah diIndonesia. Direktorat Konservasi Kawasandan Jenis Ikan, Ditjen Kelautan, Pesisir danPulau-Pulau Kecil Kemetrian Kelautan danPerikanan – LIPI
Pusat Studi Lingkungan Hidup. 1984. Penelitianair dan Biota Akuatik Danau-Danau diSumatera Barat. Laporan Penelitian PSLHUniversitas Andalas.

ISSN 978-602-14989-0-3 BioETI
Perbandingan kepadatan populasi dan sebaran ukurancangkang kerang Donax faba Gmelin, 1792 (Lamellibranchiata :Donacidae) berdasarkan kedalaman substrat di perairan pantaiBungus Teluk Kabung, Kota PadangJABANG NURDIN DAN IZMIARTI
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKTelah dilakukan penelitian tentang perbandingan kepadatan populasi dan sebaran ukuran cangkang kerang Donax fabaGmelin, 1792 (Lamellibranchiata : Donacidae) berdasarkan kedalaman substrat di perairan pantai Bungus Teluk Kabung,Kota Padang dari September hingga Desember 2010. Kerang dikoleksi di daerah intertidal dengan metode survei danteknik pengambilan sampel menggunakan stratified sistimatik sampling. Masing-masing strata (Upper, middle dan lower)diambil dua ulangan menggunakan petak kuadrat ukuran 1x1 m2 dan digali substratnya berdasarkan kedalaman(permukaan, 0-2 cm, >2-4 cm, >4-6 cm, >6-8 cm dan > 8-10 cm). Hasil penelitian didapatkan 972 individu kerangdengan ukuran panjang cangkang berkisar antara 1,9 – 31 mm dengan distribusi dari permukaan substrat sampaikedalaman substrat > 6-8 cm. Kepadatan populasi kerang D. faba tertinggi pada strata II yaitu 4,06 ind./m2 dan terendahstrata I yaitu 1,2 ind./m2. Berdasarkan kedalaman substrat bahwa kepadatan tertinggi ditemukan pada kedalaman >2-4cm dan kepadatan terendah pada permukaan substrat dan diikuti kedalaman substrat >6-8 cm. Sebaran ukuran panjangcangkang ditemukan tiga fase yaitu Juvenil, panjang cangkang (1,9-11mm), muda (14-21 mm) dan dewasa (25-31 mm).Kerang juvenile lebih banyak ditemukan pada kedalaman > 2-4 mm dan kerang muda dan dewasa terdistribusi berimbangdari kedalaman 0-2 mm sampai >8-10 mm. Kedalaman substrat dan ketidakstabilan daerah intertidal sangatmempengaruhi kepadatan dan sebaran ukuran panjang cangkang D. faba.
Key words: Donax faba, Donacidae, Teluk Kabung, kepadatan
Pendahuluan
Kerang remih (Donax faba) merupakan hewanyang menyerap makanan secara filter feeder.Sehubungan hal tersebut,, kerang ini jugadimanfaatkan sebagai pembersih lingkunganperairan yang tercemar oleh logam berat.Mekanisme sifat penyerapan logan pencemaroleh kerang diunjang oleh sifat kantung mantelyang dimiliki termasuk kerang Donax faba..
Manfaat lain dari kerang ini adalah sebagaisumber protein bagi penduduk karena memilikiprotein yang sangat tinggi. Biasanya, pendudukmengkonsumsi kerang Donax faba sebagaisayuran oleh penduduk pantai. Sedangkancangkangnya dapat dimanfaatkan sebagai hiasan(Nontji, 1987 dan Nybakken, 1988).
Distribusi kerang Donax faba pada daerahintertidal pantai dipengaruhi oleh ukuranpartikel substrat, kemiringan pantai, arus
gelombang dan kandungan organik substrat(Moosa, Kastoro dan Romimohtarto. 1980).Kerang Donax faba hidup membenamkan diridalam substrat. Gerakan membenamkan diridilakukan oleh gerakan otot kaki dandipengaruhi oleh faktor eksternal seperti arus,ombak, dan aktivitas pasang surut (Jabang, 1995dan Kastoro, 2001). Kerang ini dapat menggalisubstrat hingga kedalaman 10-15 cm (Nontji,1993). Penyebaran kerang Donax faba padasubstrat ada dua tipe yaitu secara vertical danhorizontal. Penyebaran secara verticalberhubungan dengan kedalaman kerang inimenggali substrat, sedangkan secara horizontalmerupakan gerakan berpindah pada permukaansubstrat pada rentang daerah intertidal yangmempengaruhi kelimpahan kerang ini.Kelimpahan ini terkait dengan jumlah individuyang tersebar di daerah tersebut baik juvenile,muda dan dewasa.

Jabang Nurdin dan Izmiarti 146
Tingkatan umur kerang berkaitan dengansebaran ukuran panjang cangkang tersebut(Nurdin, dan Izmiarti, 2013, Kerang Donax
faba memiliki ukuran juvenile dengan ukuranpanjang cangkang 1,9-11 mm, muda (14-21mm) dan dewasa (25-31 mm). Sebaran umurkerang Donax faba pada setiap kedalamanmaupun pada permukaan substrat di daerahintertidal akan menunjukkan kesukaannya padasubstrat. Tetapi ukuran berapa yang dominanpada setiap kedalaman substrat belum adapenelitian pada daerah Bungus Teluk Kabung,Kota Padang. Bungus Teluk Kabung merupakandaerah Teluk yang memiliki daerah intertidalyang luas dengan panjang daerah garis pantarkearah plat karang lebih kuran 30-40 meter,kemudian daerah plat karang yang sudahtertutupi lumpur hingga kearah tubir. Arusgelombang agak kurang karena terhalang pulanSetan yang tidak terlalu jauh dari daerah tubir.Dengan latar belakang tersebut, telah dilakukanpenelitian tentang perbandingan kepadatanpopulasi dan sebaran ukuran cangkang kerangDonax faba Gmelin, 1792 (Lamellibranchiata :Donacidae) berdasarkan kedalaman substrat diperairan pantai Bungus Teluk Kabung. Tujuanpenelitian ini untuk mengetahui perbandingankepadatan populasi dan sebaran ukurancangkang kerang Donax faba.
BAHAN DAN METODE
Penelitian ini dilakukan pada perairan pantaiBungus Teluk Kabung, Kota Padang dariSeptember hingga Desember 2010. Kerangdikoleksi di daerah intertidal dengan metodesurvei dan teknik pengambilan sampelmenggunakan stratified sampling dandiidentifikasi (Marshal, dan Williams. 1972;Jutting, 1953). Masing-masing strata dibagi atastiga (Upper, middle dan lower) dan diambil duaulangan menggunakan petak kuadrat ukuran 1x1m2 dan digali substratnya berdasarkankedalaman (permukaan, 0-2 cm, >2-4 cm, >4-6cm, >6-8 cm dan > 8-10 cm) (Gambar 1 dan 2).
Sungai Pisang Teluk Kabung
Pengambilan SampelGambar 1. Lokasi dan sketsa pengambilan
sampel kerang Donax faba dilapangan
Strataatas tengah tepi Daerah flat karang
Gambar 2. Strata pengambilan sampel kerangD.faba (atas, tengah dan tepi) padadaerah intertidal
Di lapangan, sampel kerang diambil dalam plot1x1 m2 pada masing-masing strata yang sudahditentukan. Pada masing-masing plot, sampelkerang D. faba diambil pada daerah permukaankemudian dimasukan dalam katung plastik dandikasih alkohol 70% kemudian diikat sertadikasih label. Selanjutnya diambil padakedalaman >2-4 cm, >4-6 cm, >6-8 cm dan > 8-10 cm) dari permukaan substrat. Semua sampelkerang D. faba yang didapatkan dimasukandalam kantung plastik dari setiap kedalamansubstrat. Kemudian dibawa ke laboratorium.
Di laboratoriun, sampel dari lapangandikeluarkan dari kantung plastik untuk dihitungjumlah individu yang didapatkan dan diukurpanjang dengan menggukan kaliver vernier (0.1mm). Data yang dari hasil penghitungan

Jabang Nurdin dan Izmiarti 147
individu dan panjang cangkang kerang D. fabadimasukan dalam tabel data. Kemudian datadiolah berdasarkan kepadatan (ind./m2)(Michael, 1986) dan sebaran ukuran panjangcangkang setiap kedalaman substrat (mm)berdasarkan kelompok umur (junvenil, mudadan dewasa).
HASIL DAN PEMBAHASAN
Cendawan Hasil penelitian didapatkan 972individu kerang Donax faba dengan ukuranpanjang cangkang berkisar antara 1,9 – 31 mm.Kerang yang didapatkan memberikan variasiwarna cangkang pada setiap kelompok ukuran,mulai putih transparan pada umumnya padakelompok yang lebih kecil (juvenil), putihkeunguan pada kelompok muda dan dewasa danputih orange pada umumnya ditemukan padayang dewasa (Gambar 3).
Gambar 3. Jenis kerang D. faba yang ditemukan
Distribusi kerang D. faba ditemukan daripermukaan substrat sampai kedalaman substrat> 6-8 cm. Pada kedalaman >6-8 cm jumlahindividu yang ditemukan sangat sedikit danumumnya dari kelompok dewasa. Pada sampleyang didapat bahwa distribusi vertical tertinggipada kedalaman 8 cm dalam substrat dan sudahsangat jarang, Nontji (1987) bahwa kerangDonax faba mampu hidup pada kedalaman 10-15 cm. Secara umum, bahwa kepadatan kerangD. faba tertinggi pada kedalaman >0-2 cmdengan kelompok umur juvenile dan muda yanglebih dominan, sedangkan pada permukaan darikelompok muda dan dewasa, pada kedalaman>2-4 cm umumnya kelompok dewasa. Hal ini
dapat dilihat pada kerang yang didapatkan padadaerah middle plot pertama (Gambar 4).
Prosentase kepadatan (%) kerang D. faba
Pemukaan
>0-2 cm
> 2-4 cm
Gambar 4. Prosentase kepadatan (%) kerang D.faba pada daerah middle plotpertama
Kepadatan populasi kerang D. faba tertinggipada strata II yaitu 4,06 ind./m2 dan terendahstrata I yaitu 1,2 ind./m2. Berdasarkankedalaman substrat bahwa kepadatan tertinggiditemukan pada kedalaman >2-4 cm dankepadatan terendah pada permukaan substratdan diikuti kedalaman substrat >6-8 cm. Hal inijuga dipengaruhi oleh Posisi kerang D. fabapada kedalaman substrat serta model siphonyang dimiliki oleh kerang D. faba (Gambar 5).
3 cm
3.4 cm
7 cm
Gambar 5. Posisi cangkang kerang D. fabadalam substrat
Hasil pengamatan bahwa siphon kerang D. faba
dijulurkan kepemukaan substrat ada yang sejajardengan permukaan substrat dan ada juga yangkeluar dari permukaan substrat. Variasi panjangsiphon sangat mempengaruhi distribusi kerangini di dalam substrat.
Sebaran ukuran panjang cangkang kerang D.faba ditemukan tiga fase yaitu Juvenil, panjang
Jabang Nurdin dan Izmiarti 147
individu dan panjang cangkang kerang D. fabadimasukan dalam tabel data. Kemudian datadiolah berdasarkan kepadatan (ind./m2)(Michael, 1986) dan sebaran ukuran panjangcangkang setiap kedalaman substrat (mm)berdasarkan kelompok umur (junvenil, mudadan dewasa).
HASIL DAN PEMBAHASAN
Cendawan Hasil penelitian didapatkan 972individu kerang Donax faba dengan ukuranpanjang cangkang berkisar antara 1,9 – 31 mm.Kerang yang didapatkan memberikan variasiwarna cangkang pada setiap kelompok ukuran,mulai putih transparan pada umumnya padakelompok yang lebih kecil (juvenil), putihkeunguan pada kelompok muda dan dewasa danputih orange pada umumnya ditemukan padayang dewasa (Gambar 3).
Gambar 3. Jenis kerang D. faba yang ditemukan
Distribusi kerang D. faba ditemukan daripermukaan substrat sampai kedalaman substrat> 6-8 cm. Pada kedalaman >6-8 cm jumlahindividu yang ditemukan sangat sedikit danumumnya dari kelompok dewasa. Pada sampleyang didapat bahwa distribusi vertical tertinggipada kedalaman 8 cm dalam substrat dan sudahsangat jarang, Nontji (1987) bahwa kerangDonax faba mampu hidup pada kedalaman 10-15 cm. Secara umum, bahwa kepadatan kerangD. faba tertinggi pada kedalaman >0-2 cmdengan kelompok umur juvenile dan muda yanglebih dominan, sedangkan pada permukaan darikelompok muda dan dewasa, pada kedalaman>2-4 cm umumnya kelompok dewasa. Hal ini
dapat dilihat pada kerang yang didapatkan padadaerah middle plot pertama (Gambar 4).
Prosentase kepadatan (%) kerang D. faba
Pemukaan
>0-2 cm
> 2-4 cm
Gambar 4. Prosentase kepadatan (%) kerang D.faba pada daerah middle plotpertama
Kepadatan populasi kerang D. faba tertinggipada strata II yaitu 4,06 ind./m2 dan terendahstrata I yaitu 1,2 ind./m2. Berdasarkankedalaman substrat bahwa kepadatan tertinggiditemukan pada kedalaman >2-4 cm dankepadatan terendah pada permukaan substratdan diikuti kedalaman substrat >6-8 cm. Hal inijuga dipengaruhi oleh Posisi kerang D. fabapada kedalaman substrat serta model siphonyang dimiliki oleh kerang D. faba (Gambar 5).
3 cm
3.4 cm
7 cm
Gambar 5. Posisi cangkang kerang D. fabadalam substrat
Hasil pengamatan bahwa siphon kerang D. faba
dijulurkan kepemukaan substrat ada yang sejajardengan permukaan substrat dan ada juga yangkeluar dari permukaan substrat. Variasi panjangsiphon sangat mempengaruhi distribusi kerangini di dalam substrat.
Sebaran ukuran panjang cangkang kerang D.faba ditemukan tiga fase yaitu Juvenil, panjang
Jabang Nurdin dan Izmiarti 147
individu dan panjang cangkang kerang D. fabadimasukan dalam tabel data. Kemudian datadiolah berdasarkan kepadatan (ind./m2)(Michael, 1986) dan sebaran ukuran panjangcangkang setiap kedalaman substrat (mm)berdasarkan kelompok umur (junvenil, mudadan dewasa).
HASIL DAN PEMBAHASAN
Cendawan Hasil penelitian didapatkan 972individu kerang Donax faba dengan ukuranpanjang cangkang berkisar antara 1,9 – 31 mm.Kerang yang didapatkan memberikan variasiwarna cangkang pada setiap kelompok ukuran,mulai putih transparan pada umumnya padakelompok yang lebih kecil (juvenil), putihkeunguan pada kelompok muda dan dewasa danputih orange pada umumnya ditemukan padayang dewasa (Gambar 3).
Gambar 3. Jenis kerang D. faba yang ditemukan
Distribusi kerang D. faba ditemukan daripermukaan substrat sampai kedalaman substrat> 6-8 cm. Pada kedalaman >6-8 cm jumlahindividu yang ditemukan sangat sedikit danumumnya dari kelompok dewasa. Pada sampleyang didapat bahwa distribusi vertical tertinggipada kedalaman 8 cm dalam substrat dan sudahsangat jarang, Nontji (1987) bahwa kerangDonax faba mampu hidup pada kedalaman 10-15 cm. Secara umum, bahwa kepadatan kerangD. faba tertinggi pada kedalaman >0-2 cmdengan kelompok umur juvenile dan muda yanglebih dominan, sedangkan pada permukaan darikelompok muda dan dewasa, pada kedalaman>2-4 cm umumnya kelompok dewasa. Hal ini
dapat dilihat pada kerang yang didapatkan padadaerah middle plot pertama (Gambar 4).
Prosentase kepadatan (%) kerang D. faba
Pemukaan
>0-2 cm
> 2-4 cm
Gambar 4. Prosentase kepadatan (%) kerang D.faba pada daerah middle plotpertama
Kepadatan populasi kerang D. faba tertinggipada strata II yaitu 4,06 ind./m2 dan terendahstrata I yaitu 1,2 ind./m2. Berdasarkankedalaman substrat bahwa kepadatan tertinggiditemukan pada kedalaman >2-4 cm dankepadatan terendah pada permukaan substratdan diikuti kedalaman substrat >6-8 cm. Hal inijuga dipengaruhi oleh Posisi kerang D. fabapada kedalaman substrat serta model siphonyang dimiliki oleh kerang D. faba (Gambar 5).
3 cm
3.4 cm
7 cm
Gambar 5. Posisi cangkang kerang D. fabadalam substrat
Hasil pengamatan bahwa siphon kerang D. faba
dijulurkan kepemukaan substrat ada yang sejajardengan permukaan substrat dan ada juga yangkeluar dari permukaan substrat. Variasi panjangsiphon sangat mempengaruhi distribusi kerangini di dalam substrat.
Sebaran ukuran panjang cangkang kerang D.faba ditemukan tiga fase yaitu Juvenil, panjang

Jabang Nurdin dan Izmiarti 148
cangkang (1,9-11mm), muda (14-21 mm) dandewasa (25-31 mm). Kerang juvenile lebihbanyak ditemukan pada kedalaman > 2-4 mmdan kerang muda dan dewasa terdistribusiberimbang dari kedalaman 0-2 mm sampai >8-10 mm. Kedalaman substrat dan ketidakstabilandaerah intertidal sangat mempengaruhikepadatan dan sebaran ukuran panjangcangkang D. faba.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Kepadatan populasi dan sebaran ukurancangkang kerang Donax faba Gmelin, 1792(Lamellibranchiata : Donacidae) berdasarkankedalaman substrat di perairan pantai BungusTeluk Kabung, cukup bervariasi pada setiapkedalaman. Variasi tersebut didukung oleh tipehidup didalam substrat dan fase umur sertapengaruh ombak pada daerah intertidal.
DAFTAR PUSTAKA
Jabang, 1995. Kepadatan populasi dan poladistribusi kerang (Pelecypoda) di estuaryBatang Masang Tiku Kabupaten Agam.Skripsi Biologi FMIPA UniversitasAndalas. (Tidak dipublikasikan).
Jutting Van Benthem, W.S.S. 1953. Systematicstudies on the marine molusca of Indo-Australia Archipelago. Treubia. 22 ( 1 ):47-65.
Kastoro. W.W. 2001. Dukungan IPTEK untukusaha budidaya jenis-jenis kerang laut diIndonesia. Oseana LIPI. Jakarta.
Marshal, A.J. dan W.D. Williams. 1972. Texbook of zoology invertebrate Englishlanguage book society and Mc. Millan.London.
Michael, P. 1986. Ecological methods for fieldand laboratory investigation. Tata Mc.Graw-hill publishing ltd. New Delhi.
Moosa, M.K., W. Kastoro dan K. Romimohtarto.1980. Peta sebaran Geografi beberapa biotalaut di perairan Indonesia. LON-LIPI.Jakarta.
Nybakken, J.W. 1988. Biologi laut suatupendekatan ekologis. PT. Gramedia.Jakarta.\
Nurdin, J. dan Izmiarti. 2013. Struktur populasikerang Polymesoda bengalensis danBatissa violacea pada ukuran partikelsubstrat berbeda di perairan batang JawiJawi Bandara Internasional Minang KabauKabupaten Padang Pariaman. Maklahseminar semirata-Bogor. 2014.
Nontji. A. 1987. Laut Nusantara. PenerrbitJambatan. Jakarta.

ISSN 978-602-14989-0-3 BioETI
Inventarisasi Kecoak (Dictyoptera) di pasar tradisional danrumah sakit di kota Padang Sumatera BaratMAIRAWITA, RESTI RAHAYU, DAHELMI DAN ROBBY JANNATAN
Labor Taksonomi Hewan Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian tentang inventarisasi kecoak di pasar tradisional dan rumah sakit di Kota Padang Sumatera Barat telahdilakukan pada bulan Agustus sampai September 2014.Metoda yang digunakan adalah metoda umpan dan tangkaplangsung. Kecoak dikoleksi di dua pasar tradisional dan dua rumah sakit swasta di kota padang, selanjutnya sampeldiidentifikasi di Laboratorium Taksonomi Hewan Jurusan Biologi Universitas Andalas. Kecoak yang dikoleksi terdiridari tiga spesies yaitu Periplaneta americana (Linn.), Blattella gemanica (Linn.)dan Nauphoeta cinerea(Olivier), kecoaktersebut termasuk ke dalam tiga subfamili dan tiga famili. Jumlah spesies terbanyak didapatkan pada famili Blattellidae.Ketiga jenis kecoak tersebut merupakan hama, dan dua jenis diantaranya telah terdistribusi secara global, yaitu P.americana dan B. germanica.
Key words: kecoak, rumah sakit, pasar tradisional, Padang.
Pendahuluan
Kecoak umum ditemukan di perumahan,perkantoran, rumah makan, restoran, swalayandan rumah sakit. Kebiasaannya suka mencarimakanan di dapur, tempat penyimpananmakanan bahkan di sampah-sampah, sehinggahal ini meresahkan masyarakat. Serangga iniseringkali masuk hunian manusia secara tidaksengaja melalui kardus berisi bahan makanan,sayuran, minuman, atau masuk bersama bahan-bahan furnitur atau alat-alat lainnya. Kecoakjuga bisa bermigrasi dari satu tempat ke tempatterdekat lainnya. Kecoak umumnya mendiamidapur dan kamar mandi. Pada siang hari,kecoak dewasa dan nimfa bisa ditemukan ditempat-tempat tersembunyi secara ber-kelompok di sekitar atau bahkan di dalamdinding alat-alat seperti pemanas, kulkas,tempat cuci piring, sekitar tempat pembuangan,di bawah atau sekitar alat pemanas air, kloset,pantri dan di bagian belakang dinding papan(Hadi, 2006).
Dari 3500 spesies kecoak, yang palingbanyak ditemukan dan menjadi hama adalahSupella longipalpa (kecoak bergaris coklat),Periplaneta americana (kecoak amerika) dan
Blattella germanica (kecoak jerman)(Layton,1914; Gillot, 2005; Bell, Roth dan Nalepa,2007). Namun laporan ilmiah tentang jenis-jenis kecoak hama di Indonesia terutama diSumatera Barat sepengetahuan penulis belumpernah ada. Selama ini penelitian tentangkecoak lebih banyak kepada kasus-kasusresistensi dan upaya pengendaliannya (Rahayudkk., 2012; Ahmad dkk., 2009; Ahmad danSuliyat, 2011).
Sehubungan dengan uraian di atas, sebagailangkah awal sangat perlu dilakukaninventarisasi jenis-jenis kecoak (Dictyoptera) dibeberapa tempat umum di Sumatera Barat,seperti pasar dan rumah sakit serta rumahpenduduk.Inventarisasi keragaman spesiessangat diperlukan sebagai penambah khasanahilmu pengetahuan dan membantu dalammempermudah pengendalian populasinya jikajenis tersebut tergolong merugikan (hama).Penelitian ini bertujuan untuk mengetahui,membandingkan dan mengelompokkan jenis-jenis kecoak di pasar tradisional dan rumahsakit yang tergolong kepada hama di kotaPadang.

Mairawita, Resti Rahayu dan Robby Jannatan 150
BAHAN DAN METODE
Pengoleksian sampel kecoak dilakukan di duapasar tradisional dan dua rumah sakit di KotaPadang; Pasar I, Pasar II, Rumah Sakit I danRumah Sakit II. Selanjutnya, sampel diolah diLaboratorium Taksonomi Hewan, JurusanBiologi FMIPA Universitas Andalas, PadangSumatera Barat.Pengoleksian SampelPengoleksian sampel kecoak dilakukan padabulan Agustus sampai September 2014 denganmenggunakan metode Tangkap Langsung danUmpan.Metode Tangkap Langsung merupakanmetode pengoleksian kecoak secara langsungmenggunakan pinset atau tangan dan kemudiandimasukkan ke dalam botol koleksi dalamkeadaan hidup. Metode Umpan merupakanmetode pengoleksian kecoak menggunakanumpan, umpan yang digunakan adalahmakanan kucing (pedigree) yang diletakkan didalam sebuah wadah berdiameter +7 cm,kemudian umpan ditunggu selama 2-3 jam dankecoak yang masuk ke wadah dikoleksi dandisimpan di dalam botol koleksi dalam keadaanhidup.
Pengolahan SampelPengolahan sampel kecoak dilakukan diLaboratorium Taksonomi Hewan JurusanBiologi FMIPA UNAND Padang. Pengolahansampel terdiri dari penyortiran, pematiansampel dengan cara disimpan di dalam freezersampai sampel mati, pengukuran (jika jumlahindividu lebih dari 10 pengukuran dilakukansebanyak n= 10), identifikasi morfospesies,mounting sampel, pengovenan, pelabelan danpenyimpanan. Identifikasi kecoak merujukkepada Lee dan Ng (2009).Kecoak yangdidapatkan dikelompokkan berdasarkan famili,subfamili, genus, jenis dan dihitung jumlahindividunya serta dibuat tablel daftar jenis.
HASIL DAN PEMBAHASAN
Berdasarkan identifikasi sampel kecoak diLaboratoriumdidapatkan tiga jenis kecoak yangtermasuk ke dalam tiga genera, tiga subfamili
dan tiga famili. Famili yang didapatkan yaituBlattidae dengan jenis Periplaneta americana(Linn.), Blaberidae dengan jenis Nauphoeta
cinerea (Olivier) dan Blattellidae dengan jenisBlattella germanica (Linn.) (Tabel 1). Jenisyang terbanyak ditemukan pada pasartradisional yaitu dua jenis dan yang palingsedikit ditemukan pada rumah sakit yaitu satujenis.
P. americana atau kecoak Americamerupakan kecoak yang tergolong ke dalamsubfamili Blattinae dan famili Blattidae.Kecoak ini mempunyai panjang total +35,7 mm,berwarna coklat terang kemerahan (Gambar. 1A dan B). Menurut Lee dan Ng (2009), P.
americana berwarna merah terang sampaicoklat, cercinya panjang dan meruncing, padainstar pertama tidak ditemukan tanda putihpada antenna dan nimpanya berwarna coklat. P.americana merupakan kecoak hama terbesaryang paling banyak ditemukan di daerahterbuka (Layton, 1914; Gillot, 2005; Bell, Rothdan Nalepa, 2007), hidup di tempat sampah,saluran got, bangunan komersil seperti pasardan pemukiman penduduk (Lee dan Ng, 2009),sesuai dengan pendapat Lee dan Ng (2009)kemungkinan inilah penyebab P. americana
ditemukan di Pasar I dan II serta got rumahsakit II. Pasar tradisional tempat pengoleksiansampel kecoak merupakan pasar yang masihbelum mempunyai sanitasi yang baik. Kecoakini banyak ditemukan di tempat penjualanhewan potong, pendapat ini juga didukung olehWhitworth (2007) bahwa P. americana banyakditemukan di fasilitas peternakan hewanmaupun di kandang hewan.
B. germanicaatau kecoak jermanmerupakan kecoak yang tergolong ke dalamsubfamili Blattellinae dan famili Blattellidae.Kecoak ini mempunyai panjang total +11,4 mm,berwarna coklat terang kekuningan,mempunyai garis hitam memanjang dipronotum dan garis kuning diantara garis hitamtersebut. Cerci kecil dan memanjang, sayapmenutupi seluruh bagian abdomen (Gambar 1E dan F). Menurut Lee dan Ng (2009),

Mairawita, Resti Rahayu dan Robby Jannatan 151
Tabel 1. Jumlah individu Kecoak yang dikoleksi pada pasar tradisional dan rumah sakit di kota Padang.
No
Lokasi
TotalFamili Pasar I Pasar II Rumah Sakit I Rumah Sakit II
SubfamiliSpesies
Hs Bt Hs Bt Hs Bt Hs Bt
BlattidaeBlattinae
1 Periplaneta americana 16 4 3 23BlaberidaeOxyhaloinae
2 Nauphoeta cinerea 6 7 5 18BlattellidaeBlattellinae
3 Blattella germanica 31 31Total Individu 22 11 5 31 3 72
(%) 31.89 15.95 7.24 44.92 4,17 100Total jenis per lokasi 2 2 1 1
(%) 40 40 20 20Total genus per lokasi 2 2 1 1
(%) 40 40 20 20Keterangan: Hs; metoda koleksi langsung
Bt; metoda umpan
(A) (B)
(C) (D)
(E) (F)
Gambar 1. (A) P. americana dorsal, (B) P. americana ventral, (C) N. cinerea dorsal, (D) N. cinerea ventral,(E) B. germanica dorsal, (F) B. germanica ventral.
Mairawita, Resti Rahayu dan Robby Jannatan 151
Tabel 1. Jumlah individu Kecoak yang dikoleksi pada pasar tradisional dan rumah sakit di kota Padang.
No
Lokasi
TotalFamili Pasar I Pasar II Rumah Sakit I Rumah Sakit II
SubfamiliSpesies
Hs Bt Hs Bt Hs Bt Hs Bt
BlattidaeBlattinae
1 Periplaneta americana 16 4 3 23BlaberidaeOxyhaloinae
2 Nauphoeta cinerea 6 7 5 18BlattellidaeBlattellinae
3 Blattella germanica 31 31Total Individu 22 11 5 31 3 72
(%) 31.89 15.95 7.24 44.92 4,17 100Total jenis per lokasi 2 2 1 1
(%) 40 40 20 20Total genus per lokasi 2 2 1 1
(%) 40 40 20 20Keterangan: Hs; metoda koleksi langsung
Bt; metoda umpan
(A) (B)
(C) (D)
(E) (F)
Gambar 1. (A) P. americana dorsal, (B) P. americana ventral, (C) N. cinerea dorsal, (D) N. cinerea ventral,(E) B. germanica dorsal, (F) B. germanica ventral.
Mairawita, Resti Rahayu dan Robby Jannatan 151
Tabel 1. Jumlah individu Kecoak yang dikoleksi pada pasar tradisional dan rumah sakit di kota Padang.
No
Lokasi
TotalFamili Pasar I Pasar II Rumah Sakit I Rumah Sakit II
SubfamiliSpesies
Hs Bt Hs Bt Hs Bt Hs Bt
BlattidaeBlattinae
1 Periplaneta americana 16 4 3 23BlaberidaeOxyhaloinae
2 Nauphoeta cinerea 6 7 5 18BlattellidaeBlattellinae
3 Blattella germanica 31 31Total Individu 22 11 5 31 3 72
(%) 31.89 15.95 7.24 44.92 4,17 100Total jenis per lokasi 2 2 1 1
(%) 40 40 20 20Total genus per lokasi 2 2 1 1
(%) 40 40 20 20Keterangan: Hs; metoda koleksi langsung
Bt; metoda umpan
(A) (B)
(C) (D)
(E) (F)
Gambar 1. (A) P. americana dorsal, (B) P. americana ventral, (C) N. cinerea dorsal, (D) N. cinerea ventral,(E) B. germanica dorsal, (F) B. germanica ventral.

Mairawita, Resti Rahayu dan Robby Jannatan 152
B. germanica jantan berwarna kuningkecoklatan, sedangkan yang betina berwarnaagak gelap daripada yang jantan. Mempunyaidua pita hitam paralel longitudinal dipronotumnya dan dibatasi oleh garis terang.Nimpa berwarna hitam dengan strip terangpada setengah bagian pertama tubuhnya dibagian dorsal. B. germanica selamapengoleksian sampel hanya ditemukan diRumah Sakit I dan tidak ditemukan di pasartradisional. Kecoak ini hanya ditemukan ditempat-tempat modern. Lee dan Ng (2009)mengatakan bahwa B. germanica biasanyaditemukan di tempat penyimpanan makanan,dapur komersial, toko makanan dan restoran.Hal ini sesuai dengan yang ditemukan dilapangan bahwa B. germanica juga ditemukandi dapur, tempat penyimpanan makanan,restoran dan kafe yang ada di Rumah Sakit Itersebut (Gambar. 2 D).
N. cinerea atau kecoak lobster merupakankecoak yang tergolong ke dalam subfamiliOxyhaloinae famili Blaberidae. Kecoak inimempunyai panjang total +27,8 mm, berwarnacoklat muda terang dan mempunyai corak padapronotumnya serta sayapnya tidak menutupiseluruh bagian abdomen (Gambar. 1 C dan D).Menurut Lee dan Ng (2009), N. cinereaberwarna seperti abu dan mempunyai polaseperti lobster di bagian pronotumnya,sayapnya pendek dan tidak menutupi seluruhabdomen serta kecoak betina lebih besardaripada kecoak jantan.N. cinerea banyakditemukan di fasilitas penjualan hewan di pasartradisional, seperti ayam potong atau sapipotong. Selama pengoleksian sampel, hanya N.cinerea yang menyukai umpan pada perangkap,karena umpan yang diberikan adalah makanankucing yaitu pedigree (Gambar. 2 A). Lee danNg (2009) juga mengatakan bahwa N. cinereamemakan makanan yang dimakan oleh hewan,memakan tumbuhan dan makanan yangmengandung minyak ikan.
Alasan penting kecoak sebagi hama bahwakecoak ini telah menginvasi bangunan,mengkontaminasi makanan, terdistribusi secara
global, 25 sampai 30 spesies merupakan hamadan 4 sampai 5 spesies merupakan hama yangmendominasi secara global, termasuk P.americana dan B. germanica (Cochran, 2003).P. americana dan B. germanica merupakankecoak hama yang paling besar dan palingbanyak ditemukan di pemukiman pendudukserta kecoak ini juga telah resisten terhadapinsektisida (Layton, 1914; Gillot, 2005; Bell,Roth dan Nalepa, 2007).
KESIMPULAN
Berdasarkan hasil yang didapatkan maka dapatdisimpulkan bahwa kecoak yang ditemukan dipasar tradisional dan rumah sakit di kotaPadang tergolong ke dalam tiga famili, tigasubfamili dan tiga spesies, yaitu P. americana,N. cinerea dan B. germanica. Jenis yang palingbanyak ditemukan di pasar tradisional dan yangpaling sedikit pada rumah sakit di kota Padang.Semua kecoak yang ditemukan merupakanhama bagi manusia dan diantaranyamendominasi secara global yaitu P. americanadan B. germanica.
UCAPAN TERIMA KASIHPenulis mengucapkan terima kasih kepadaProgram DIPA Universitas Andalas 2014Nomor: 14/UN.16/PL/DM/i/2014 yang telahmembiayai penelitian ini serta Hirzan Riyandidan Annisa Izmi Aulia sebagai tim lapangandalam pengoleksian sampel kecoak.
DAFTAR PUSTAKA
Ahmad, I. dan Suliyat. 2011.Adaptasi Seranggadan Dampaknya bagi KehidupanManusia.Pidato Ilmiah Guru Besar ITB.ITB Bandung 21 Oktober 2011.
Ahmad, I., Sriwahjuningsih, Astari, S., Putra,R.E., dan A.D. Permana. 2009.Monitoring Pyrethroid Resistance in FieldCollected B. germanica (Dictyoptera:Blattellidae) in Indonesia, EntomologicalResearch.39: 114-118.
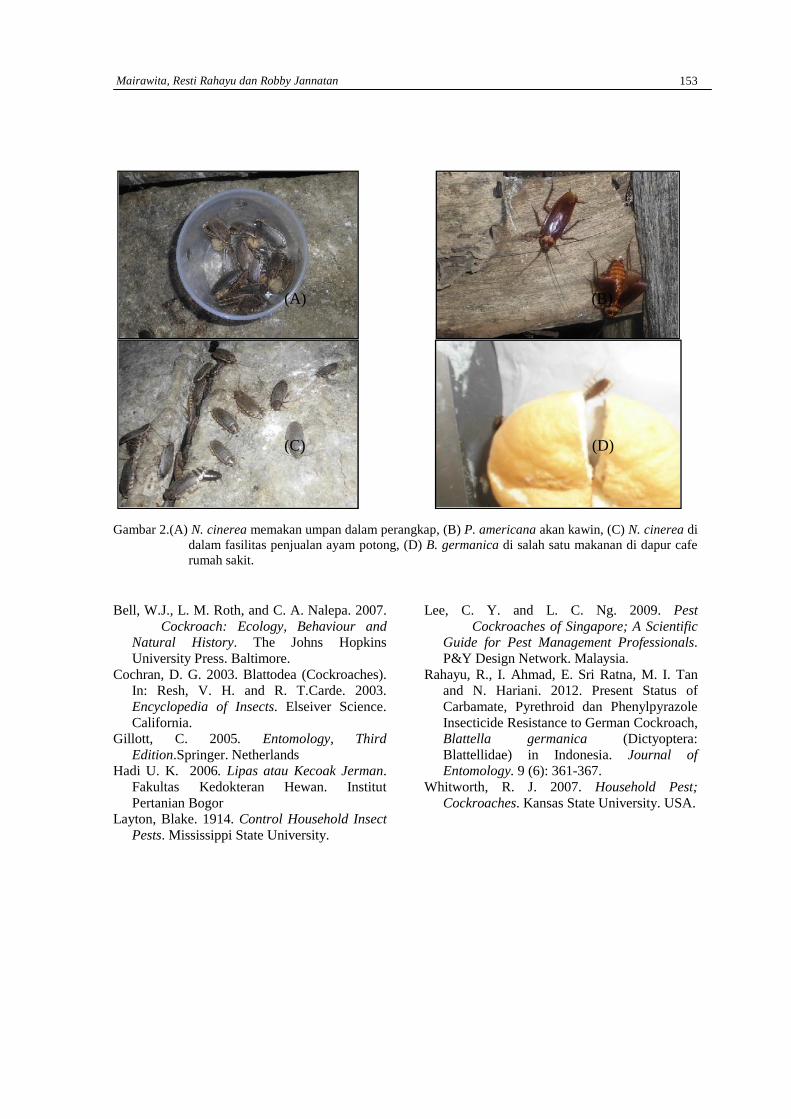
Mairawita, Resti Rahayu dan Robby Jannatan 153
(A) (B)
(C) (D)
Gambar 2.(A) N. cinerea memakan umpan dalam perangkap, (B) P. americana akan kawin, (C) N. cinerea didalam fasilitas penjualan ayam potong, (D) B. germanica di salah satu makanan di dapur caferumah sakit.
Bell, W.J., L. M. Roth, and C. A. Nalepa. 2007.Cockroach: Ecology, Behaviour and
Natural History. The Johns HopkinsUniversity Press. Baltimore.
Cochran, D. G. 2003. Blattodea (Cockroaches).In: Resh, V. H. and R. T.Carde. 2003.Encyclopedia of Insects. Elseiver Science.California.
Gillott, C. 2005. Entomology, ThirdEdition.Springer. Netherlands
Hadi U. K. 2006. Lipas atau Kecoak Jerman.Fakultas Kedokteran Hewan. InstitutPertanian Bogor
Layton, Blake. 1914. Control Household InsectPests. Mississippi State University.
Lee, C. Y. and L. C. Ng. 2009. PestCockroaches of Singapore; A Scientific
Guide for Pest Management Professionals.P&Y Design Network. Malaysia.
Rahayu, R., I. Ahmad, E. Sri Ratna, M. I. Tanand N. Hariani. 2012. Present Status ofCarbamate, Pyrethroid dan PhenylpyrazoleInsecticide Resistance to German Cockroach,Blattella germanica (Dictyoptera:Blattellidae) in Indonesia. Journal ofEntomology. 9 (6): 361-367.
Whitworth, R. J. 2007. Household Pest;Cockroaches. Kansas State University. USA.

ISSN 978-602-14989-0-3 BioETI
Studi Morfologi Feses MamaliaMARDHA TILLAH 1, WILSON NOVARINO 1 DAN RIZALDI 2
1)Laboratorium Riset Taksonomi Hewan,2)Laboratorium Riset Ekologi HewanJurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian tentang studi morfologi feses mamalia telah dilakukan pada bulan Maret sampai dengan Mei 2014, di TamanMargasatwa Budaya Kinantan Kota Bukittinggi dengan menggunakan metode observasi secara langsung di lapangan.Dari hasil pengamatan setiap tingkat taksa mamalia yang lebih dekat memiliki kemiripan feses yang lebih banyak.Komposisi feses mamalia tersusun dari serat kasar dan serat halus, dengan persentase yang lebih tinggi yaitu serat kasar.
Key words: morfologi, feses, mamalia, komposisi
Pendahuluan
Pada Feses merupakan salah satu tanda darihewan yang jelas dan paling mudah dikenali(Liebenberg, 2000). Cabang ilmu yangmempelajari tentang feses disebut Scatology(Seton, 1925). Pelacakan feses merupakansalah satu metode non invasif untukmempelajari spesies yang terancam, jenishewan yang sulit ditemukan dan jenis hewannokturnal (Chame, 2003). Menurut Gormandan Trownbridge (1989) pelacakan feses bisadigunakan untuk mengetahui daerah jelajahhewan dan daerah teritorinya. Penghitungsnjumlah feses bisa memberikan informasimengenai distribusi serta kelimpahan suatujenis hewan dalam suatu habitat.
Feses tidak hanya membantu dalampengamatan jenis dan komposisi makananhewan, namun juga dapat memberikaninformasi tetang kondisi kesehatan dan jenis-jenis endoparasit dari hewan tersebut. Melaluipengamatan feses dapat diketahui perilakumakan dan mengetahui jenis-jenis hewanmangsa hewan tersebut (Halfpenny, 2008).
Namun ada beberapa spesies yang sulitdiamati fesesnya, akibat beberapa faktor seperti,adanya feses yang dikubur atau perilakumembuang feses dalam air atau di cabang-cabang pohon. Untuk mengidentifikasi, bentukfeses asli harus dipertahankan. Beberapa faktor
dapat menimbulkan kerusakan pada bentukfeses yaitu akibat lingkungan yang panas dankekeringan, atau dekomposisi yang cepatuntuk daerah lembab dengan curah hujantinggi. Selain itu kesulitan dalam mengamatifeses yaitu adanya fragmentasi oleh hewan lainseperti kumbang tinja dan rayap yang seringmengkonsumsi feses herbivora (Stuart andStuart 1998).
Ukuran dan jumlah feses yang dihasilkanoleh setiap individu bervariasi tergantung usia,jenis makanan yang dimakan, dan kapasitaspenyerapannya (Bang and Dahlstrom 1975).Hewan herbivora berbeda bentuk sertakomponen penyusun fesesnya dengan hewankarnivora dan omnivora. Selain itu musim dankelimpahan sumber makanan jugamempengaruhi struktur dan komposisi darifeses mamalia. Fesessuatu spesies memiliki bauyang khas dan kompleks (Gorman andTrowbridge, 1989). Feses terdiri dari materialyang dicerna keseluruhan dicerna sebagian dantidak tercerna. Komponen dari fesesdiantaranya bulu, tulang, gigi, cakar, sisik, kitinarthropoda, biji dari tanaman, serbuk sari danmaterial lainya baik dari tumbuhan maupunhewan mangsa (Bang dan Dahlstrom, 1975;Bjune, 2000).
Sejak tahun 1970-an jumlah penelitianmengenai scatology mulai meningkat (Seton,1925). Khusus di Indonesia penelitian tentang

Mardha Tillah, Wilson Novarino dan Rizaldi 155
feses sudah dilakukan seperti analisis variasi D-Loop DNA mitokondria pada populasi gajahsumatera menggunakan sampel feses (Savira,2008). Penelitian lainnya adalah efektivitaspenggunaan metode penghitungan feses untukmenilai distribusi dan kelimpahan relatifmonyet dan musang di taman nasional komodo(Jessop et al., 2006). Hal ini mengarah kepadapemanfaatan feses sebagai salah satu metodedalam pelacakan hewan untuk melihatdistribusi dan menghitung kelimpahan hewantersebut. Terlebih untuk jenis mamalia yangdilindungi, susah diamati, populasi rendah danmendiami hutan luas. Karenanya dilakukanpenelitian ini dengan harapan dapat membantudalam identifikasi lapangan. Pengamatanmorfologi feses baru dilakukan di Afrika (Cillie,2003) dan Amerika Selatan (Chame, 2003)untuk Asia terutama Indonesia info tentangmorfologi feses belum banyak atau sangatjarang dilakukan. Padahal morfologi fesesberpeluang untuk pemantaun dan pelacakansatwa mamalia.
BAHAN DAN METODE
Penelitian ini menggunakan metode observasidan koleksi secara langsung di lapangan, fesesmamalia diamati secara langsung di TamanMargasatwa dan Budaya Kinantan Bukittinggi.
HASIL DAN PEMBAHASAN
Pada saat penelitian dilakukan, tercatatsebanyak 18 jenis mamalia di TamanMargasatwa dan Budaya Kinantan Bukittinggi.Jenis-jenis tersebut tergolong kedalam 7 ordodan 15 famili. Sebanyak 16 jenis merupakansatwa asli Indonesia dan 2 jenis didatangkandari luar Indonesia. Feses dari mamalia tersebutmempunyai bentuk yang bervariasi dari bulat,lonjong, tabung, dan lain-lain. Ukuran fesesantara 0,54 cm sampai dengan 13,96 cm (Tabell ).
Morfologi feses mamalia mempunyai polatertentu pada tingkat ordo. Ordo proboscideabentuk umum feses silinder ditemukan pada
Elephas maximus (Gajah Sumatera). Fesesgajah mempunyai ciri morfologi sebagaiberikut: warna hijau kecoklatan untuk yangsegar sampai kecoklatan untuk feses yang telahkering (Gambar 1). Menurut (Cillie, 2003) padagajah afrika feses segar berwarna hijau zaitundan warna akan semakin gelap setelah lebihdari 6 jam. Bentuk feses gajah yaitu silinder.Permukaan feses segar berlendir dan agak kesatkarena masih terdapat serat-serat tumbuhan.Berat feses basah yaitu mencapai 1164 gramdan feses kering 381 gram.
Gambar 1. Elephas maximus a. feses segarb. feses kering
Ordo primata bentuk umum feses bulatmemanjang tidak beraturan dan terdapat sekat-sekat pada bagian tengah. Sekat pada ordoprimata ada yang berupa garis dan ada yangsudah terdiferensiasi menjadi bulatan-bulatanyang menyatu.
Feses Pongo pygmaeus mempunyai cirimorfologi sebagai berikut: Feses yang segarberwarna coklat dan feses kering berwarnakehitaman. Bentuk feses segar bulatmemanjang tersususun dari kepingan-kepingan.Feses yang kering berbentuk bulat tabungseperti batang kayu yang telah kering (Gambar2a). Permukaan feses agak licin. Berat fesesbasah yaitu mencapai 59 gram dan feses kering34 gram.
Feses Nycticebus coucang mempunyai cirimorfologi sebagai berikut: Feses segarberwarna kuning kecoklatan, semi keringberwarna coklat keputihan dan feses keringberwarna keputihan. Feses tersusun daribulatan-bulatan yang memanjang dengan salahsatu ujung meruncing dan terdapat sekat-sekatyang jelas (Gambar 2b). Permukaan feses agaklicin. Berat feses basah yaitu mencapai 4 gramdan feses kering 1 gram.
a b

Mardha Tillah, Wilson Novarino dan Rizaldi 156
Gambar 2. a. Pongo pygmaeus b. Nycticebuscoucang c. Symphalangussyndactylus d. Hylobates agilis e.Macaca nemestrina f. Presbytismelalophos
Feses Symphalangus syndactylusmempunyai ciri morfologi sebagai berikut:Feses segar berwana hijau kecoklatan dan feseskering berwarna kehitaman. Bentuk feses segarseperti bulatan bulatan yang tersusunmemanjang seperti untian. Untuk feses keringbentuknya bulat pipih memanjang dan terdapatsekat-sekat (Gambar 2c). Panjang fesesberkisar antara 4,24 cm sampai dengan 16,97cm dengan panjamg rata-rata 9,57 cm.Diameter feses berkisar antara 1,43 cm sampaidengan 3,27 cm. Permukaan feses agak licinkarena masih terdapat serat dan sisa makananlainya yang belum tercerna. Berat feses basahyaitu mencapai 15 gram dan feses kering 8gram.
Feses Hylobates agilis mempunyai cirimorfologi sebagai berikut: warna feses segarcoklat sedangkan feses semi kering hitamkecoklatan. Bentuk feses bulat memanjang danterdapat sekat-sekat (Gambar 2d). Permukaanfeses agak licin karena masih terdapat sisamakanan yang belum tercerna. Panjang fesesberkisar antara 6,28 cm sampai dengan 7,18 cmdengan panjamg rata-rata 6,73 cm. Diameterfeses berkisar antara 1,32 cm sampai dengan2,24 cm dengan diameter rata-rata 1,78 cm.
Berat feses basah yaitu mencapai 10 gram danfeses kering 5 gram.
Feses Macaca nemestrina mempunyai cirimorfologi sebagai berikut: berwarna coklatuntuk yang masih segar dan hitam kecoklatanuntuk yang kering (Gambar 2e). Bentuk bulatmemanjang salah satu ujung runcing danterdapat sekat sekat. Permukaan feses kuranglicin karena masih ada bagian makanan yangtidak tercerna dengan baik. Panjang fesesMacaca nemestrina berkisar antara 3,05 cmsampai dengan 4,27 cm dengan panjang rata-rata 3,68 cm. Diameternya berkisar antara 1,94sampai dengan 2,25 dengan diameter rata-rata2,02 cm. Berat feses basah yaitu mencapai 4,15gram dan feses kering 3 gram.
Feses Presbytis melalophos mempunyai cirimorfologi sebagai berikut: Feses segarberwarna coklat kehijauan sedangkan feseskering berwarna coklat keputihan. Bentuk fesesbulat panjang bersekat sekat (Gambar 2f).Permukaan feses agak licin karena masih adamaterial dari feses yang belum tercernasempurna. Panjang feses berkisar antara 7,44cm sampai dengan 8,65 cm dengan panjamgrata-rata 8,18 cm. Diameter feses berkisarantara 1,43 cm sampai dengan 2,34 cm dengandiameter rata-rata 1,87 cm. Berat feses basahyaitu mencapai 46 gram dan feses kering 8gram.
Feses ordo cetartiodactyla bentuk umumfeses bulatan. Bulatannya ada yang menyatudan ada yang berupa butiran. Ukuran fesesbervariasi dari ukuran sedang sampai denganukuran kecil. Untuk rusa totol dan rusa sambarukuran dan bentuknya hampir sama denganbagian salah satu ujung meruncing.Perbedaanya rusa totol meruncingnya hanyapada bagian ujungnya saja namun rusa sambarseperti peluru. Permukaan feses rusa sambarlebih licin dibandingkan rusa totol.Feses Sus scrofa mempunyai ciri morfologisebagai berikut: Feses yang ditemukan diTaman Margasatwa dan Budaya Kinantanberwarna coklat sedangkan feses yangditemukan di kandang milik warga berwarna
ab
c d
e f

Mardha Tillah, Wilson Novarino dan Rizaldi 157
kehitaman. Bentuk feses bulat memanjang yangtersusun dari bulatan-bulatan kecil yangmenyatu dengan salah satu ujung agakmeruncing (Gambar 3a). Permukaan feses agakkasar karena masih terdapat serat – serat darisisa makanan. Panjang feses berkisar antara8,06 cm sampai dengan 12,65 cm denganpanjamg rata-rata 9,67 cm. Diameter fesesberkisar antara 3,24 cm sampai dengan 4,12 cmdengan diameter rata-rata 3,66 cm. Berat fesesbasah yaitu mencapai 75gram dan feses kering27 gram.
Gambar 3. a. Sus scrofa b. Camelusdromedarius c. Rusa unicolor d.Axis axis
Feses dari Camelus dromedariusmempunyai ciri morfologi sebagai berikut:warna coklat kehijauan. Bentuk feses membulat(Gambar 3b). Permukaan feses agak licinkarena masih terdapat sisa makanan yangbelum tercerna. Panjang feses Camelus
dromedarius berkisar dari 3,44 cm sampaidengan 6,97 cm dengan panjang rata-rata 5,20cm. Diameter feses berkisar dari 3,18 cmsampai dengan 3,82 cm dengan diameter rata-rata 3,5 cm. Berat feses basah yaitu mencapai52 gram dan feses kering 17 gram.
Feses Rusa sambar mempunyai cirimorfologi sebagai berikut: Feses berwarnacoklat. Feses berbentuk butiran silinder dengansalah satu ujung agak meruncing (sepertipeluru) (Gambar 3c). Permukaan feses licin.Panjang feses berkisar antara 1,54 cm sampai
dengan 2,22 cm dengan panjamg rata-rata 1,92cm. Diameter feses berkisar antara 0,92 cmsampai dengan 1,14 cm dengan diameter rata-rata 0,97 cm. Berat feses basah yaitu mencapai1,5 gram dan feses kering 0,3 gram.
Feses rusa totol mempunyai ciri morfologisebagai berikut: Feses berwarna kehitaman.Feses berbentuk butiran silinder dengan salahsatu ujung meruncing (Gambar 3d). Permukaanfeses kurang licin dibandingkan dengan rusasambar. Panjang feses berkisar antara 1,44 cmsampai dengan 2,07 cm dengan panjamg rata-rata 1,60 cm. Diameter feses berkisar antara0,67 cm sampai dengan 0,96 cm dengandiameter rata-rata 0,75 cm. Berat feses basahyaitu mencapai 1,9 gram dan feses kering 0,77gram.
Feses ordo rodentia bentuk umumnyabutiran silindris, satu-satu, kedua ujungmembulat atau salah satu ujung agakmeruncing. Ukuran bervariasi dari sangat kecil,seperti pada Muridae (tikus) and Sciuridae(bajing) sampai ukuran sedang seperti landak(Hystricidae) (Chame, 2003).
Gambar 4. a. Hystrix javanica b. Callosciurus
prevostii
Feses Hystrix javanica mempunyai cirimorfologi sebagai berikut: Feses segarberwarna coklat sedangkan feses semi keringberwarna hitam coklat keputihan. Bentuk fesesseperti butiran silindris, ujung membulat danyang satunya agak meruncing. Pada bagianujung yang membulat tersusun atas bulatan-bulatan kecil yang menyatu (Gambar 4a).Permukaan feses agak licin. Panjang fesesberkisar antara 7,25 cm sampai dengan 12,24cm dengan panjamg rata-rata 9,92 cm.Diameter feses berkisar antara 3,05 cm sampai
a b
c d
a
b
b

Mardha Tillah, Wilson Novarino dan Rizaldi 158
dengan 4,26 cm dengan diameter rata-rata 3,66cm. Berat feses basah yaitu mencapai 113gramdan feses kering 31 gram.
Feses Callosciurus prevostii mempunyaiciri morfologi sebagai berikut: feses berwarnakuning kemerahan. Bentuk feses bulatmemanjang (Gambar 4b). Permukaan fesesagak licin. Feses tersusun dari materil bubukdengan berat 23,8 % dari berat feses kering danserat makanan yang tidak tercerna beratnya76,2 % dari berat feses kering. Panjang fesesberkisar antara 0,98 cm sampai dengan 1,83 cmdengan panjamg rata-rata 1,37 cm. Diameterfeses berkisar antara 0,54 cm sampai dengan0,77 cm dengan diameter rata-rata 0,67 cm.Berat feses basah yaitu mencapai 1 gram danfeses kering 0,8 gram. Feses dari ordocarnivora bentuk umum feses juga hampir samayaitu bulat silindris dengan salah satu bagianujung meruncing.
Gambar 5. a. Helarctos malayanus b. Arctictisbinturong c. Panthera tigrissumatrae
Feses Helarctos malayanus (beruang)mempunyai ciri morfologi sebagai berikut:warna feses bervariasi dari warna hitamkecoklatan, kuning kehijauan, coklat keorenan.Bentuk feses silindris (bentuk sosis) dengansalah satu bagian ujung meruncing (Gambar5a). Panjang feses berkisar antara 4,76 cmsampai dengan 7,24 cm dengan panjamg rata-rata 6,23 cm. Diameter feses berkisar antara2,18 cm sampai dengan 3,21 cm dengandiameter rata-rata 2,55 cm. Berat feses basahyaitu mencapai 53 gram dan feses kering 22gram.
Feses Arctictis binturong (binturong)mempunyai ciri morfologi sebagai berikut:Feses segar berwarna hitam, sedangkan feseskering berwarna putih kecoklatan. Bentuk fesesbulat panjang dan salah satu ujung meruncing(Gambar 5b). Permukaan feses agak kasarkarena masih terdapat sisa makanan yangbelum tercerna sempurna. Panjang fesesberkisar antara 6,28 cm sampai dengan 12,66cm dengan panjamg rata-rata 9,47 cm.Diameter feses berkisar antara 1,46 cm sampaidengan 3,54 cm dengan diameter rata-rata 2,5cm. Berat feses basah yaitu mencapai 73 gramdan feses kering 9 gram.
Feses Panthera tigris sumatrae (harimausumatera) mempunyai ciri morfologi sebagaiberikut: Feses segar berwarna hitam dan feseskering berwarna coklat keputihan. Bentuk fesessilindris dengan salah satu bagian ujungmeruncing (Gambar 5c). Untuk feses harimauyang sebelumnya hidup liar seluruh permukaanfeses licin dan dilapisi oleh bulu padapermukaanya. Feses harimau sejak kecil dipenangkaran permukaan lebih kasar. Panjangfeses berkisar antara 6,69 cm sampai dengan12,51 cm dengan panjamg rata-rata 10,29 cm.Diameter feses berkisar antara 2,55 cm sampaidengan 5,41 cm dengan diameter rata-rata 3,94cm. Berat feses basah yaitu mencapai 97 gramdan feses kering 25 gram.
Feses ordo diprodontia bentuk umumnyabulat memanjang beraturan dan terdapat sekat-sekat pada bagian tengah ditemukan padaThylogale brunii (kangguru tanah). Feseskanguru tanah mempunyai ciri morfologisebagai berikut: Feses segar berwarna hijaucoklat kehitaman. Bentuk feses bulat panjangsatu ujung agak runcing dan yang satunyamembulat tumpul. Terdapat sekat - sekat padabagian tengahnya (Gambar 6). Panjang fesesberkisar antara 5,78 cm sampai dengan 6,52 cmdengan panjamg rata-rata 6,24 cm. Diameterfeses berkisar antara 1,88 cm sampai dengan 1cm dengan diameter rata-rata 2,31 cm. Beratfeses basah yaitu mencapai 16 gram dan feseskering 8 gram.
a b
c
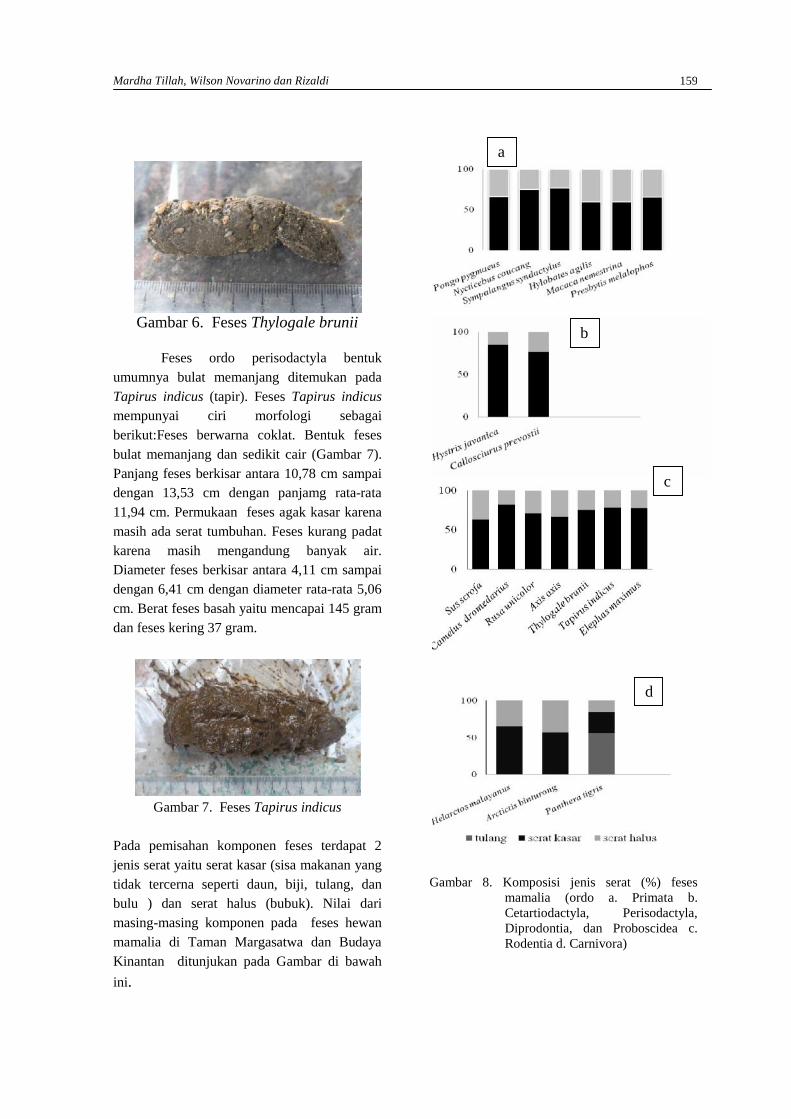
Mardha Tillah, Wilson Novarino dan Rizaldi 159
Gambar 6. Feses Thylogale brunii
Feses ordo perisodactyla bentukumumnya bulat memanjang ditemukan padaTapirus indicus (tapir). Feses Tapirus indicusmempunyai ciri morfologi sebagaiberikut:Feses berwarna coklat. Bentuk fesesbulat memanjang dan sedikit cair (Gambar 7).Panjang feses berkisar antara 10,78 cm sampaidengan 13,53 cm dengan panjamg rata-rata11,94 cm. Permukaan feses agak kasar karenamasih ada serat tumbuhan. Feses kurang padatkarena masih mengandung banyak air.Diameter feses berkisar antara 4,11 cm sampaidengan 6,41 cm dengan diameter rata-rata 5,06cm. Berat feses basah yaitu mencapai 145 gramdan feses kering 37 gram.
Gambar 7. Feses Tapirus indicus
Pada pemisahan komponen feses terdapat 2jenis serat yaitu serat kasar (sisa makanan yangtidak tercerna seperti daun, biji, tulang, danbulu ) dan serat halus (bubuk). Nilai darimasing-masing komponen pada feses hewanmamalia di Taman Margasatwa dan BudayaKinantan ditunjukan pada Gambar di bawah
ini.
Gambar 8. Komposisi jenis serat (%) fesesmamalia (ordo a. Primata b.Cetartiodactyla, Perisodactyla,Diprodontia, dan Proboscidea c.Rodentia d. Carnivora)
a
b
c
d
Mardha Tillah, Wilson Novarino dan Rizaldi 159
Gambar 6. Feses Thylogale brunii
Feses ordo perisodactyla bentukumumnya bulat memanjang ditemukan padaTapirus indicus (tapir). Feses Tapirus indicusmempunyai ciri morfologi sebagaiberikut:Feses berwarna coklat. Bentuk fesesbulat memanjang dan sedikit cair (Gambar 7).Panjang feses berkisar antara 10,78 cm sampaidengan 13,53 cm dengan panjamg rata-rata11,94 cm. Permukaan feses agak kasar karenamasih ada serat tumbuhan. Feses kurang padatkarena masih mengandung banyak air.Diameter feses berkisar antara 4,11 cm sampaidengan 6,41 cm dengan diameter rata-rata 5,06cm. Berat feses basah yaitu mencapai 145 gramdan feses kering 37 gram.
Gambar 7. Feses Tapirus indicus
Pada pemisahan komponen feses terdapat 2jenis serat yaitu serat kasar (sisa makanan yangtidak tercerna seperti daun, biji, tulang, danbulu ) dan serat halus (bubuk). Nilai darimasing-masing komponen pada feses hewanmamalia di Taman Margasatwa dan BudayaKinantan ditunjukan pada Gambar di bawah
ini.
Gambar 8. Komposisi jenis serat (%) fesesmamalia (ordo a. Primata b.Cetartiodactyla, Perisodactyla,Diprodontia, dan Proboscidea c.Rodentia d. Carnivora)
a
b
c
d
Mardha Tillah, Wilson Novarino dan Rizaldi 159
Gambar 6. Feses Thylogale brunii
Feses ordo perisodactyla bentukumumnya bulat memanjang ditemukan padaTapirus indicus (tapir). Feses Tapirus indicusmempunyai ciri morfologi sebagaiberikut:Feses berwarna coklat. Bentuk fesesbulat memanjang dan sedikit cair (Gambar 7).Panjang feses berkisar antara 10,78 cm sampaidengan 13,53 cm dengan panjamg rata-rata11,94 cm. Permukaan feses agak kasar karenamasih ada serat tumbuhan. Feses kurang padatkarena masih mengandung banyak air.Diameter feses berkisar antara 4,11 cm sampaidengan 6,41 cm dengan diameter rata-rata 5,06cm. Berat feses basah yaitu mencapai 145 gramdan feses kering 37 gram.
Gambar 7. Feses Tapirus indicus
Pada pemisahan komponen feses terdapat 2jenis serat yaitu serat kasar (sisa makanan yangtidak tercerna seperti daun, biji, tulang, danbulu ) dan serat halus (bubuk). Nilai darimasing-masing komponen pada feses hewanmamalia di Taman Margasatwa dan BudayaKinantan ditunjukan pada Gambar di bawah
ini.
Gambar 8. Komposisi jenis serat (%) fesesmamalia (ordo a. Primata b.Cetartiodactyla, Perisodactyla,Diprodontia, dan Proboscidea c.Rodentia d. Carnivora)
a
b
c
d

Mardha Tillah, Wilson Novarino dan Rizaldi 160
Persentase untuk serat kasar lebih dominandisebabkan karena sumber makan mamalia diTMSBK berasal dari tumbuh-tumbuhan yangmengandung selulosa. Selulosa tidak dapatdicerna oleh tubuh karena tidak ada enzimuntuk memecah selulosa. Meskipun tidak dapatdicerna, selulosa berfungsi sebagai sumberserat sehinnga dapat memperbesar volume darifeses (Hutagalung, 2004).
KESIMPULAN
Dari penelitian yang telah dilaksanakan dapatdisimpulkan setiap tingkat taksa mamalia yanglebih dekat memiliki kemiripan yang lebihbanyak. Komposisi feses mamalia terbagi atasserat kasar (sisa serat tumbuhan, biji, tulang)dan serat halus (bubuk) dengan persentasekomposisinya lebih dominan serat kasardibandingkan dengan serat halus (bubuk)
UCAPAN TERIMAKASIHTerimakasih kepada kepala Taman Margasatwadan Budaya Kinantan Bukittinggi dan stafkandang mamalia di Taman Margasatwa danBudaya Kinantan Bukittinggi atas bantuan dankerja samanya.
DAFTAR PUSTAKA
Bang P, Dahlström P 1975. Huellas y Señalesde los Animales de Europa, Omega,Barcelona, 239 pp.
Bujne A.E 2000. Pollen analysis of faeces as amethod of demonstrating seasonalvariations in the diet of Svalbard reindeer
(Rangifer tarandus platyrhynchus). PolarRes19: 183-192.
Chame, M. 2003. Terrestrial Mammal Feces: aMorphometric Summary and Description.Mem Inst Oswaldo Cruz, Rio de Janeiro,Vol. 98(Suppl. I): 71-94, 2003
Cillie, B. 2003. The Pocket Photoguide toMammals of Southern Africa. Sunbirdpublishing. South africa
Gorman ML, Trowbridge BJ 1989. The roleodor in the social lives carnivores. In JLGittleman, Carnivore Behaviour, Ecologyand Evolution. Chapman & Hall Ltd, NewYork, p. 57-88.
Halfpenny J, Biesiot 1986. A Field Guide toMammal Tracking in North America, 2nded., Johnson Publishing, New york, 161pp.
Hutagalung, H. 2004. Karbohidrat. UniversitasSumatera Utara. Medan
Jessop, T.S., Forsyht, D.M., Purwandana, D.,Seno, A. 2006. Efektivitas Penggunaan
Metode Penghitungan Kotoran Untuk MenilaiDistribusi dan Kelimpahan Relatif Monyetdan Musang di Taman Nasional Komodo,Indonesia. Terjemahan. Ariefiandy, A.,Purwandana, D. Imansyah, M.J. CRES-ZSSD/BTNK/TNC. Labuan Bajo, Flores,Indonesia.
Liebenberg L 2000. Tracks and Tracking inSouthern Africa, Struik Publishers, CapeTown, 144 pp.
Savira, M. 2012. Analisis variasi D-Loop DNAmitokondria pada populasi gajah Sumatera.Universitas Indonesia. Jakarta
Seton ET. 1925. On the study of scatology. JMamm 6: 47-49.
Stuart C, Stuart T 1998. A Field Guide to theTracks and Signs of Southern and EastAfrican Wildlife, Southern BooksPublishers, Cape Town, 310 pp.

ISSN 978-602-14989-0-3 BioETI
Induksi kalus pada hipokotil tanaman Turi (Sesbaniagrandiflora) dengan menggunakan bap yang dikombinasikandengan beberapa konsentrasi Auksin secara In VitroMARDHIYETTI1, ZULFADLY SYARIF2, NOVIRMAN JAMARUN1 DAN IRFANSULIANSYAH2
1Fakultas Peternakan, Universitas Andalas, Kampus Limau Manis Padang 251632Fakultas Pertanian, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKTuri merupakan leguminosa pohon, mempunyai banyak fungsi dan digunakan sebagai pakan ternak. Perbanyakan turisecara vegetatif sulit dilakukan, karena kemampuan turi untuk tumbuh kembali setelah dilakukan pemotongan sangatrendah. Penelitian dalam upaya perbanyakan turi melalui metode regenerasi tanaman turi secara in vitro telah dilakukandi Laboratorium Kultur Jaringan, Pertanian, Universitas Andalas. Eksplan yang digunakan adalah hipokotil steril tanamanturi. Media dasar yang digunakan adalah MS (Murashige skoog) .Penelitian ini bertujuan untuk menemukan konsentrasiBAP dan Auksin yang terbaik untuk induksi kalus. Auksin yang digunakan terdiri dari NAA, IAA dan IBA. Hasilpenelitian menunjukkan bahwa media induksi kalus terbaik pada hipokotil adalah kombinasi BAP 1ppm dan NAA 0.08ppm.
Key words: turi, callus, auksin, benzil adenin purin, in vitro
Pendahuluan
Turi sudah dikenal sebagai hijauan pakanternak, namun saat ini ketersediaan turi terbatasdan sulit dijumpai. Menurut (Heyne, 1987) turitidak ditemukan tumbuh liar di pulau Jawa.Tanaman ini sangat berpotensi sebagai pakandan tidak ada faktor pembatas bila diberikanke ternak (NAS,1979). Sutikno (2002)menyatakan turi merupakan leguminosa pohongolongan kacang-kacangan yang cukupberharga bila dikembangkan karena hampirsemua bahagian tanaman dapat dimanfaatkansebagai bahan pakan, pernyataan ini jugadidukung oleh Heyna (1987) . Disamping ituTuri juga memiliki kandungan protein kasaryang tinggi, Siregar (1992) menyatakan bahwabahwa hasil analisis kadar protein kasar padabeberapa bahan pakan ternak seperti kaliandra(Caliandra calothyrsus), lamtoro (leucaena
leucepala), gamal (Gliricidia maculate) danbeberapa jenis rumput menunjukkan bahwa turimemiliki kandungan protein kasar tertinggi.Permasalahan yang ditemukan dalam budidayaturi adalah sulitnya perbanyakan secara
konvensional yaitu menggunakan stek. Johndan Mannetje (1992) menyatakan bahwarespon tanaman turi untuk tumbuh kembalisetelah ternak digembalakan atau terhadappemupukan belum didapatkan informasinya.Sampai saat ini belum banyak yang terungkaptentang pemanfaatan turi dan kapan waktu yangtepat untuk pemberian turi ke ternak. Demikianjuga dengan umur pemotongan untuk dijadikansebagai bahan tanam, sehingga tidakmenganggu pertumbuhannya dan diharapkandapat tumbuh kembali dengan cepat.Pemangkasan turi secara rutin akanmenyebabkan tingkat kematian yang tinggi,selanjutnya pemotongan teratur (5 kali setahun)untuk membentuk pagar yang rendah (denganketinggian 1 meter) menyebabkan kematianhampir 100% dalam suatu percobaan di TimurLaut Thailand.Untuk mengatasi permasalahandi atas dapat dilakkukan dengan perbanyakansecara in vitro, penggunaan teknik in vitro
dengan metode kultur jaringan akan mengatasikendala-kendala yang umum dijumpai padateknik – teknik budidaya dengan pembiakan

Mardhiyetti, Zulfadly Syarif, Novirman Jamarun dan Irfan Suliansyah 162
generatif ataupun vegetatif yang dapatdikatakan sebagai teknik konvensional.
Penelitian ini merupakan lanjutan daripenelitian sebelumnya.Penelitian yang telahdilakukan adalah menginduksi tunas denganmenggunakan hipokotil, daun dan kotiledon.Hasil yang didapat rata-rata tunas yangterbentuk 1 shoodlet per eksplan. Selanjutnyapada penelitian ini menggunakan hipokotilsebagai eksplan dengan perlakuan beberapaauksin dengan komposisi media yang samadengan tujuan menghasilkan kalus, diharapkandari kalus yang diinduksi mampu menghasilkantunas dalan jumlah yang lebih banyak.
BAHAN DAN METODE
Tanah Penelitian ini dilaksanakan diLaboratorium Kultur Jaringan TumbuhanJurusan Budidaya Pertanian Fakultas PertanianUniversitas Andalas. Media induksi yangdigunakan adalah BAP dan dikombinasikandengan beberapa auksin yaitu NAA, IAA danIBA. Selanjutnya perlakuan dipindahkan kemedia regenerasi, Media yang digunakan dapatdilihat pada Tabel 1.
Tabel 1. Komposisi hormon tumbuh yangdigunakan pada media induksi kalus
Media Kode media Konsentrasi hormon tumbuhInduksi kalus IK-1 1 mg/l BAP+0,08 mg/l NAA
IK-2 1 mg/l BAP+ 0,08 mg/l IAAIK-3 1 mg/1 BAP+ 0,08 mg/l IBA
Regenerasi RG-1 1 mg/l BAP+0,08 mg/l NAARG-2 1 mg/l BAP + 0,08 mg/l IAARG-3 1 mg/l BAP + 0,08 mg/l IBARG-4 2 mg/l BAP+0,08 mg/l NAARG-5 2 mg/l BAP + 0,08 mg/l IAA
RG-6 2 mg/l BAP + 0,08 mg/l IBA
Bahan tanam yang digunakan adalah biji Turi(Sesbania grandiflora) yang didapatkan dariBalai Penelitian Peternakan Bogor (Balitnak,Bogor). Zat – zat pengatur diperoleh dariLaboratorium Kultur Jaringan Pertanian Unand.Eksplan yang digunakan adalah hipokotil Turi,nutrisi penyusun media basal yaitu MurashigeSkoog (MS), vitamin, agar – agar powder 8
gr/L, sukrosa 30 gr/L, innomyositol 0,1 gr/L,NaOH 0,1 N, Hcl 0,1 N, zat pengatur tumbuhNAA (Naftalen Acetat Acid), IAA (Indol acetat
acid), IBA (Indol Butiric Acid) dan BAP(Benzil Amino Purin), alkohol 70%, bayclin,deterjent, spritus dan aquades steril.
Alat – alat yang digunakan ; scapel, botolkultur dengan tinggi 9,3 cm dan diameternya6,5 cm, LAFC (Laminar Air Flow Cabinet),gelas beker, gelas ukur, labu semprot,autoclave, hot plate dengan magnetic stirer,pengaduk kaca, petridish, hansprayer,timbangan analitik, label, plastik kaca, lakbanbening, karet gelang, tissue, plastik wrap,gunting, pinset, bunsen, aluminium foil,kompor gas, pH meter, lemari es untukpenyimpanan media, oven untuk penyimpananalat, rak kultur yang dilengkapi dengan lampudan ruangan yang dilengkapi pengatur suhu,kamera digital.
Kegiatan awal pembuatan larutan stok, yaitularutan garam – garam an organik yang didalamnya terkandung hara makro, sebagaimedia dasar pembuatan media MS dan mediaWPM, larutan vitamin dan zat pengatur tumbuh.Larutan – larutan yang telah dibuat disipandalam refrigerator. Masing – masing larutanstok dipipet sesuai dengan media perlakuan,selanjutnya dicampur dengan sukrosa 30 g/l,myoinositol 0,1 g kedalam labu takar danditambahkan aquades hingga mencapai 1000ml (untuk pembuatan medium 1 liter).
Pada pembuatan media perlakuanditambahkan zat pengatur tumbuh (NAA, IBA,IAA dan BAP) sesuai dengan kombinasi dantaraf konsentrasi perlakuan. Pengukuran pHdilakukan sebelum ditambah agar –agar sebagaibahan pemadat sebanyak 8 gr/L, pH yangdiinginkan adalah sekitar 5,6 – 5,8. Apabila pH< 5,6 larutan ditambahkan NaOH 1N danapabila pH > 5,8 larutan ditambahkan larutanHCL 1N. Larutan media berisi agar – agartersebut kemudian dimasak sampai mendidih,kemudian dituangkan ke dalam botol kulturyang steril sebanyak 20 ml/liter. Botol – botolkultur tersebut ditutup rapat dengan aluminium
Mardhiyetti, Zulfadly Syarif, Novirman Jamarun dan Irfan Suliansyah 163
foil, selanjutnya diautoclaf pada tekanan 17,5psi dengan suhu 120ºC selama 15 menit. Mediadisimpan dalam ruang inkubator. Data diamatisecara deskriptif.
HASIL DAN PEMBAHASAN
Respon hipokotil terhadap komposisi mediabasal dengan menggunaan sitokinin yaitu BAPdan beberapa jenis auksin untuk membentukkalus dapat dilihat pada Tabel 1. Pembentukankalus diawali dengan terjadinya pembengkakanekplan, selanjutnya,kalus mulai terbentuk padabagian pelukaan. Kalus yang dihasilkanberwarna putih dengan tekstur kompak agakremah.
Tabel 1. Respon hipokotil pada media induksikalus
Media induksikalus
Jumlah eksplanyang ditanam
Jumlah kalusyang terbentuk
Tekstur kalus
IK BAP/NAAIk BAP/IAAIK BAP/IBA
100100100
100 %95 %93%
Kompak agak remahKompak agak remahKompak agak remah
Pada Tabel 1 dapat dilihat bahwa hampirsemua eksplan yang berasal dari hipokotilmembentuk kalus, terbentuknya kalus padamedia perlakuan karena hormon endogen daneksogen berada dalam komposisi yangseimbang dan mencukupi untuk menginduksikalus. Morfogenesis eksplan tergantung kepadakeseimbangan auksin dan sitokinin didalammedia dan interaksi antara zat pengatur tumbuhendogen di dalam tanaman dan zat pengaturtumbuh eksogen yang diserap dari mediatumbuh (Wattimena 1992). Pembentukan kalusdiawali pada bagian eksplan yang dilukai,kemudian berkembang sampai menutupipermukaan eksplan. Zat pengatur tumbuh yangdiberikan bila sesuai dengan kebutuhantanaman, maka proses pembentukan kalusdapat lebih cepat, apalagi dengan menggunakanjaringan meristem.
Dari penelitian sebelumnya juga telahdilakukan dengan menggunakan eksplan daundan kotiledon pada perlakuan yang sama,membentuk kalus hanya pada ujung eksplan
dan mampu membentuk tunas (Mardhiyetti etal., 2014). Hasil ini menunjukkan bahwarespon setiap eksplan membentuk kalusberbeda-beda pada media yang sama. Responeksplan pada media induksi kalus dapat dilihatpada Tabel 2.
Tabel 2. Perkembangan hipokotil pada mediainduksi kalus
Pengamatan IAA IBA NAA
Hari inisiasi kalus 2,24 2.44 3.94Warna kalus 4.51 4.68 4.46Hari Induksi tunas - - 4.92
Respon kalus pada media induksi terhadaphari inisiasi kalus menunjukkan bahwa padaNAA 3.94 lebih cepat pada IAA dan IBA, yaitu2.24 dan 2.44. Pengamatan terhadap warnakalus dilakukan secara scoring, warna kalusyang didapat berkisar 4.46-6.68, inimenunjukkan bahwa warna kalus didominasiputih/tak berwarna. Hari induksi terdapat padaNAA sedangkan pada IAA dan IBA belumterlihat karena eksplan mencoklat (Gambar 1).
Tabel 3. Respon kalus pada media RegenerasiMediaregenerasi
JumlahkalusAwal
Jumlahkalusdenganspothijau
Jumlahkalusberegenerasi
JumlahTunas
RG-1 75 40% 22 22RG-2 75 23% 11 11RG-3 75 12% 10 10RG-4 75 15% - -RG-5 75 3% - -RG-6 75 7% - -
Pada Tabel 3 dapat dilihat bahwa kalus padamedia regenerasi mampu menghasilkan kalus.Menurut Wattimena (1992), zat pengaturtumbuh dari golongan auksin berperan antaralain dalam pembentukan kalus, morfogenesisakar dan tunas serta embriogenesis. Pemilihankonsentrasi dan jenis auksin ditentukan antaralain oleh tipe pertumbuhan dan perkembangan.Kalus yang didapatkan spot hijau meskipundalam jumlah yang terbatas. Kalus dengan spot

Mardhiyetti, Zulfadly Syarif, Novirman Jamarun dan Irfan Suliansyah 164
hijau berpotensi membentuk tunas, tunas yangdihasilkan sebahagian besar satu tunas pereksplannya, dari hasil temuan ada beberapaeksplan yang menghasilkan tunas lebih darisatu tunas, namun terjdai vitrifikasi dan eksplanmencoklat (Gambar 1).
Gambar 1. Perkembangan kalus pada hipokotilturi
Setiap tanaman berbeda kebutuhannya terhadapzat pengatur tumbuh, kebutuhannya tergantungpada zat pengatur tumbuh yang sudah ada padatanaman tersebut. Pada tanaman melinjo(Rozen, 2010) menunjukkan bahwapembentukan kalus dengan menggunakan NAAyang dikombinasikan dengan arang aktif padakonsentrasi 1 ppm. Gautam et al (1993)melaporkan bahwa rasio auksin dan sitokiniyang tidak sama diperlukan bagi keberhasilaninduksi kalus azadirachta indica.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Melalui kalus diharapkan dapat menginduksitunas dalam jumlah yang banyak Hipokotilmampu membentuk kalus pada BAP yangdikombinasikan dengan IAA, NAA dan IBA.Warna kalus hipokotil adalah putih/takberwarna dengan tekstur kompak agak remah..Dari hasil penelitian ini tunas dapat terbentuk,namun eksplan cepat mencoklat, akibatnnyapertumbuhan tunas tidak maksimal.
DAFTAR PUSTAKA
AAK, 1983. Hijauan makanan ternak potong,kerja dan perah. YayasanKanisius.Yokyakarta.
Gautam VK, Nanda J, Gupta SC. 1993.Development of shoots and root in anther:derived callus of Azadirachta indica A.Juss.-a medical tree. Plant Cell, Tissue.And Organ Culture 34: 13-18.
Gunawan, L, W. 1988. Teknik Kultur JaringanTumbuhan. PAU Bioteknologi. IPB.Bogor. 252 hal.
Heyne. 1987. Tumbuhan berguna Indonesia II.Badan Peneliti dan PengembanganKehutanan Departemen Kehutanan.Jakarta. Cetakan ke-1. Hal 971.
Mardhiyetti, Zulfadly, S, Novirman, J, Irfan. S.2014. Response Same Explant of Turi(Sesbania grandiflora) Shoot InductionMedium. Journal international. SAFE.
Mukhri Z, Baihaki A, Soedigdo P. 1985. KulturJaringan temulawak (Curcumaxanthorrhiza Roxb., Zingiberaceae) danstudi awal kemungkinan penggunaanmutagen untuk meningkatkankadarkurkuminnya. Dalam ProsidingSimposium Nasional TemulawakBandung: Lembaga Penelitian Univ.Padjajaran.
NAS. 1979. Forages. National Academy ofSciences. Washington, DC.
Siregar, S. B. 1994. Ransum TernakRuminasia . Pusat Penelitian danPengembangan Peternakan Bogor.
Rozen. 2010. Inisiasi kalus eksplan daunmelinjo pada berbagai konsentrasi arangaktif dengan komposisi konsentrasi BAPdan NAA secara in vitro. FakultasPertanian. Unand.
Susetyo, S, 1980. Padang Pengembalaan.Departemen Ilmu makanan teknak.Fakultas Peternakan. IPB Bogor.
Sutikno, I. 2002. Pengolahan biji turi (sesbaniagrandiflora) untuk mengurangi senyawaanti nutrisi. Balai Penelitian Ternak.Bogor.

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman makanan dan ukuran lambung Ranacancrivora Gravenhorst (Anura : Ranidae) pada dataran tinggidan dataran rendah Sumatera BaratMELIYA WATI DAN ELZA SAFITRI
Program Studi Pendidikan BiologiSTKIP PGRI SumbarE-mail:
ABSTRAKAmfibia memiliki berbagai peranan penting bagi kehidupan manusia, terutama peranan ekologis. Amfibia juga berfungsisebagai bio-indikator bagi kondisi lingkungan karena memiliki respon terhadap perubahan lingkungan. Makanan utamaAmfibia adalah serangga sehingga dapat membantu keseimbangan ekosistem terutama dalam pengendalian populasiserangga. Rana cancrivora merupakan jenis katak sawah yang berperan penting untuk pengendalian hama. Penelitian inibertujuan untuk mengetahui keanekaragaman makanan dan ukuran lambung Rana cancrivora pada dataran tinggi danrendah di Sumatera Barat. Hasil penelitian menunjukkan bahwa katak Rana cancrivora di lokasi dataran rendah (Padang)lebih beragam yaitu 7 ordo dari kelas Insecta dan 1 dari Gastropoda serta ditemukan larva dan pupa serangga. Keragamanmakanan pada lokasi dataran tinggi (Batu Sangkar) lebih sedikit yaitu 5 ordo dari kelas Insecta dan ditemukan jenis dariGastropoda. Hasil pengukuran morfometrik menunjukkan perbedaan yang nyata pada parameter lebar lambung, panjangdan lebar moncong. Parameter panjang lambung dan berat menunjukkan perbedaan yang tidak nyata.
Key words: Keanekaragaman makanan, ukuran lambung, ketinggian, Rana cancrivora
Pendahuluan
Amfibia memiliki berbagai peranan penting bagikehidupan manusia, terutama peranan ekologis.Secara ekologis, Amfibia memiliki perananpenting dalam rantai makanan sebagaikonsumen sekunder. Amfibia memakanserangga sehingga dapat membantukeseimbangan ekosistem terutama dalampengendalian populasi serangga. Selain itu,Amfibia juga dapat berfungsi sebagai bio-
indikator bagi kondisi lingkungan karenamemiliki respon terhadap perubahan lingkungan.Peranan Amfibia dari segi ekonomis dapatditinjau dari pemanfaatan untuk kepentingankonsumsi. Beberapa jenis Amfibia dari OrdoAnura diketahui memiliki nilai ekonomis yangtinggi seperti Fejervarya cancrivora, Fejervaryalimnocharis, dan Limnonectes macrodon. Selainuntuk tujuan konsumsi, Amfibia memilikikegunaan yang lain yaitu sebagai binatangpeliharaan, binatang percobaan dan bahan obat-obatan (Stebbins and Cohen 1997).
Keberadaan katak di alam semakinmenunjukkan penurunan, karena penangkapanliar yang berlebihan dan beberapa faktor laindiperkirakan telah memperbesar penurunanpopulasi katak di alam adalah adanya kerusakanhabitat, intensifikasi pertanian, pembukaanlahan dan adanya industri beserta limbahnya.Pada masa yang akan datang, tekanan terhadappopulasi katak akan terus berlanjut dan bukantidak mungkin pada suatu saat spesies ini akanpunah (Nasaruddin, 2008).
Kerusakan habitat dan pemakaian insektisidauntuk pembasmi hama termasuk faktor terbesaryang dihadapi saat ini. Hal ini mempengaruhipelestarian katak dalam habitat, karenakeragaman serangga suatu tempatmempengaruhi makanan katak. Hal ini menjadibagi penulis untuk mengamati komposisimakanan. Suatu lokasi memiliki komposisisumber makanan yang menunjang kehidupankatak, tetapi katak memiliki kemampuan yangberbeda berdasarkan distribusinya di alam(Teynie et al., 2010).

Meliya Wati dan Elza Safitri 166
Beberapa hasil penelitian menunjukkanbahwa sebagian besar jenis makanan katakadalah spesies-spesies Arthropoda terutamainsekta. Berdasarkan penelitian Hirai and Matsui(2001), komposisi diet Rana (Fejervarya)limnocharis 94,8 % adalah Artrhopoda. Menurutpenelitian Qingqing et.al., (2004) indeksdiversitas Insekta yang ditemukan dalam katakR. limnocharis adalah 0,88 dan antara individujantan dan betina memiliki diversitas yangberbeda signifikan.
Faktor jenis-jenis makanan yang dikonsumsikatak dipengaruhi oleh faktor musim dan faktorketinggian dan faktor abiotik lainnya, sepertisuhu, temperatur dan kelembaban. Perbedaankomposisi makan yang memperlihatkanperbedaan pada iklim yang berbeda, berdasarkanpenelitian Maneyro dan Rosa (2004) dietmakanan katak Hyla pulchella berbeda padamusim panas dan dingin serta iklim mikro (suhu,kelembaban dan intensitas cahaya). MenurutNaya et al., (2009) bahwa perbedaan jenismakanan bisa terjadi pada spesies yang sama,akibat faktor abiotik yaitu ketinggian yangberasosiasi dengan temperatur. Berdasarkanukuran lambungnya, ditemukan adanyahubungan panjang lambung dengan kemampuanmakanan katak tersebut dan ketersediaanmakanan. Ketersediaan makanan dipengaruhioleh faktor abiotik seperti keadaan lingkungan,kondisi tanah, kepadatan populasi sumbermakanan serta perubahan lingkungan.
Sumatera Barat sebagai bagian dari pulauSumatera memiliki geografis yang bervariasi,mulai dari dataran rendah sampai dataran tinggi.Faktor geografis sangat memungkinkanterbentuk perbedaan faktor abiotik terutamatemperatur, kelembaban, dan intensitas cahaya.Faktor abiotik mempengaruhi komposisikomunitas spesies yang saling hidupberdampingan secara ekologis. Faktor abiotikmenjadi salah satu faktor perbedaan komposisimakanan termasuk pada katak.
BAHAN DAN METODE
Bahan yang digunakan adalah etanol 70%,aquadest, tissue gulung, formalin, sarung tangan.Alat-alat yang digunakan adalahheadlamp/senter, karung, kantung plastik, karetgelang, sarung tangan, kamera digital, jangkasorong digital, label gantung, alat bedah, bakbedah, jarum injeksi, botol specimen, masker,botol sampel, mikroskop binokuler dan alat tulis.
Metode Penelitian ukuran lambung dankomposisi makanan menggunakan metodeNaya , Veloso, Bozinovic (2009), Sole’ et. al.(2005) dan pengkoleksian sampel langsung dilapangan (Mistar dan Iskandar, 2003). Sampelditangkap di daerah Padang (ketinggian 0-200mdpl) dan daerah Kabupaten Tanah Datar(ketinggian 500-1000 mdpl). Selain pengamatanukuran lambung dan komposisi lambung,dilakukan pengamatan pengukuran faktorabiotik: suhu, kelembaban dan ketinggiantempat.
Pengkoleksian sampel dilakukan secaralangsung di sekitar pemukiman penduduk dansepanjang aliran sungai. Pengkoleksiandilakukan pada malam hari dimulai pukul 19.00– 22.00 WIB. Penangkapan katak dilakukan disekitar dan dalam sawah.
Spesimen kodok dibius denganmenggunakan alkohol 70% minimal 2 jamsetelah penangkapan. Selain itu , alkohol 70%diinjeksikan keorgan dalam atau dibawah kulitperut. Lalu diawetkan dalam botol specimendalam botol specimen dengan alkohol 70%.Pemeriksaan isi lambung dilanjutkan diLaboratorium.
Spesimen kodok dibedah pada bagianabdomen dan dipisahkan lambungnya,kemudian diletakkan kedalam petridish dandisortir dengan menggunakan mikoskopbinokuler.

Meliya Wati dan Elza Safitri 167
Isi lambung yang diidentifikasikan adalahhewan-hewan yang dimakan katak. Identifikasidilakukan pada tingkat ordo terutama serangga,dengan menngunakan buku identifikasi Borroret al., (1996).
Pengukuran morfometrik yang dilakukanterhadap spesimen yaitu panjang dan lebarlambung, panjang moncong sampai anus, lebarmulut, jenis kelamin, dan juga ditimbang beratbadan.
Perbedaan pengukuran morfometrik danberat antar lokasi dianalisis dengan uji t. Hewanyang ditemukan dalam lambung kodok dihitungperindividu dan dikelompokkan berdasarkanOrdo. Penghitungan jumlah individu mengikuticara Berry, 1965 dalam Kurniati, 1998; yaitukategori bentuk mangsa dalam lambung terbagidua:1. Mangsa berada dalam bentuk utuh atau
hampir utuh2. Potongan kaki, sclerit, sayap, kepala elytra,
ovipositor dan jumlah pasangan sayap
HASIL DAN PEMBAHASAN
Keanekaragaman Makanan Rana cancrivoraBerdasarkan penelitian yang telah dilaksanakan,kelas Insecta merupakan kelas terbanyak yangditemukan dalam lambung katak R. cancrivorayang dijelaskan pada Tabel 1. Sebagian besarkelompok serangga yang dimakan oleh seranggaadalah ordo Orthoptera yaitu kelompok belalang,baik ditemukan di dataran rendah maupundataran tinggi. Berdasarkan lokasinya penelitiankeanekaragaman serangga makanan terbesarterdapat pada lokasi dataran rendah dan datarantinggi. Hasil penelitian ini berbeda daripenelitian lain. Menurut Kurniati (1998) bahwaFormicidae merupakan makanan utama kataklebih dari 75 %. Makanan umum katak adalahsemua jenis serangga yang ada di dalam habitat.Komponen lain yang ditemukan mungkin masuktertelan bersamaan dengan mangsanya, misalnyadaun-daun kecil. Hasil penelitian Rogerio et al.,(2007) bahwa Serangga yang dominan dimakan
oleh katak S. argyreornatus adalah Isoptera,Formicidae dan Homoptera.
Tabel 1. Komposisi serangga yang ditemukan didalam lambung Katak Rana cancrivora
Kelas dan Ordo Dataran Rendah(Padang)
Dataran Tinggi(Batu sangkar)
Jumlah % Jumlah %
1. InsectaOrthoptera 11 36,66 3 25Hemiptera 6 20 2 16,67Arachnida 7 23,33 1 8,33Coleoptera 3 10 - -Dermoptera 1 3,33 - -Diptera 1 3,33 2 16,67Phthiraptera 1 3,33 - -Hymenoptera - - 2 16,67III. MoluscaGastropoda 1 3,33 2 16,67Total 30 100 12 100
Berdasarkan lokasi geografis terlihat kodok daridataran rendah memakan serangga yang lebihberagam dibandingkan dengan katak datarantinggi. Keanekaragaman serangga yang dimakandipengaruhi dengan komposisi serangga yangdimana habitat kodok.
Hasil pengukuran Morfometrik dan BeratBadanPengukuran morfometrik dan berat badan kataktelah dilakukan pada panjang lambung, lebarlambung, panjang moncong sampai anus danlebar mulut serta berat badan. Data hasilpengkuran dapat dilihat pada Tabel 2.Berdasarkan Tabel 2, Jumlah individu totalkatak adalah 14 ekor, yang mana dari Padangsebanyak 8 ekor, dan dari Batu sangkar 6 ekor.Hasil pengkuran panjang lambung antara lokasiketiga lokasi menunjukkan bahwa antara lokasiPadang dan Batu Sangkar memiliki perbedaan(Tabel 3.).
Perbedaan morfometrik didukung olehperbedaan faktor lingkungan dan kondisimakanan yang tersedia di suatu tempat. MenurutShaubeble (2004) bahwa ukuran tubuhmerupakan karakter morfometrik yangfundamental dalam hal yang berkaitan denganfisiologi, ekologi dan tingkahlaku spesies. Wienet al., (2009) menyatakan bahwa variasi dan
Meliya Wati dan Elza Safitri 168
diferensiasi yang ditunjang faktor genetik dalamwaktu yang lama akan menuju evolusi.
Tabel 2. Hasil pengukuran MorfometrikLambung dan Badan Katak Sawah Ranacancrivora dari Lokasi PenelitianPadang dan Batu Sangkar
Lok
asi
JenisKelamin dan
Jumlah(ekor)
PanjangLambung (mm)
LebarLambung (mm)
PanjangMoncong
-anus(mm)
LebarMulut(mm)
BeratBadan
(gr)
Pada
ng
Jant
an (
4)
17,65-21,70
6,00-10,00
45,60-64,10
13,55-
22,50
8,20-25,84
Bet
ina
(4)
13,50-25,55
8,40-14,50
38,60-67,20
11,35-
23,80
4,57-31,73
Bat
u Sa
ngka
r
Jant
an (
1)
18,90 7,60 33,30 11,70 2,53
Bet
ina
(5)
13,45-18,20
5,40-6,60
35,60-47,45
12,50-
14,60
3,40-9,46
Tabel 3. Uji t untuk melihat perbedaan rata-ratapengukuran morfometrik dan beratbadan antara lokasi Padang danBatusangkar
Parameter Padang(rata-rata)
BatuSangkar
(rata-rata)Db t hitung t tabel
PanjangLambung
19,78 16,82 12 1,43ns 2,179
LebarLambung
9,91 6,92 12 2,18* 2,179
PanjangMoncong
54,80 38,68 12 3,72* 2,179
LebarMoncong
17,89 12,43 12 2,70* 2,179
Berat 13,80 8,95 14 1,09ns 2,145
Keterangan : ns = nonsignificant * = significant
Perbedaan morfometrik didukung olehperbedaan faktor lingkungan dan kondisimakanan yang tersedia di suatu tempat. MenurutShaubeble (2004) bahwa ukuran tubuh
merupakan karakter morfometrik yangfundamental dalam hal yang berkaitan denganfisiologi, ekologi dan tingkahlaku spesies. Wienet al., (2009) menyatakan bahwa variasi dandiferensiasi yang ditunjang faktor genetik dalamwaktu yang lama akan menuju evolusi. Faktorlingkungan yang mendukung dalam habitatkatak R. cancrivora pada tiga lokasimenunjukkan perbedaan, dapat dilihat padaTabel 4.
Tabel 4. Perbedaan Habitat dan Faktor AbiotikKatak Sawah Rana cancrivora dariLokasi Padang dan Batu Sangkar
Parameter DataranRendah
(Padang)
Dataran Tinggi(Batu sangkar)
Suhu oC 28-29 26-28Kelembaban (%) 71-81 86-90Ketinggian (mdpl) 1-2 500-800Kondisi Habitat Habitat sawah
air tawar, airtergenang,kondisi sawahpascapanen
Habitat sawah airtawar, air tergenang,kondisi sawahpascatanam
Perbedaan habitat akan memperlihatkan jenismakanan yang dimakan oleh katak. Makanankatak terutama jenis Insecta atau serangga,seperti penelitian yang telah dilakukan olehWati dan Hidayat (2013), meneliti diet kodokBufo melanostictus dan diperoleh perbedaankomposisi makanan, antara dataran rendah dandataran tinggi yaitu di dataran rendah ditemukanordo Diptera, sedangkan di dataran rendah tidakditemukan.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Penelitian terhadap keanekaragaman makanandan ukuran lambung Rana cancrivora yangberasal dari dua lokasi, Padang dan Batusangkarterdiri dari dua kelas yaitu Insecta dan Molusca.Ordo terbanyak adalah Ordo Orthoptera, untukkedua lokasi. Hasil pengukuran lambung yangmenunjukan perbedaan yang nyata adalah lebarlambung, lebar lambung dan panjang lambung,sedangkan panjang lambung dan berat badanmenunjukan perbedaan yang tidak nyata.

Meliya Wati dan Elza Safitri 169
UCAPAN TERIMAKASIHTerimakasih penulis ucapkan kepada perguruantinggi STKIP PGRI Sumbar Padang dan DIPADP2M Kementrian Pendidikan dan KebudayaanTahun 2014 yang telah membantu danmendukung penelitian ini.
DAFTAR PUSTAKA
Almeria, L. M. dan O. M., Nuheza. 2013. Dietof Seven Anuran Species (Amphibia :Anura) in Agusan Marsh, Mindanao,Philippines. Animal : Biology and AnimalHusbandry International Journal of TheBioflux Society. ABAH Bioflux. Vol. 5issue.
Borror, D.J., S. A. Triplehorn, N. F. Jhonson.1996. Pengenalan Pelajaran Serangga.Edisi Bahasa Indonesia. Yogyakarta: GajahMada University press.
Dure’, M. I., A. I. Kehrand and E. F. Schaefer.2009. Niche Overlap and ResourcePartitioning among Five SympatricBufonids (Anura, Bufonidae) fromNortheaster Argentina. Phyllomedusa. Vol.VII (1):27-39.
Hartmann, P.A. dan Marques, O.A.V. 2005.Diet and Habitat use of two SympatricSpecies of Philodryas (Colubridae) inSouth Brazil. Amphibia Reptilia. XXVI:25-21
Hirai, T and M. Matsui. 2001. Diet Compositionof The Indian Rice Frog, Rana limnocaris,In Rice Field of Central Japan. CurentHerpetology. XX (2): 97-103
Iskandar, D. T. 2003. Amphibi Jawa dan Bali.Puslitbang Biologi LIPI. Jakarta.
Kurniati, H. 1998. Kebiasaan Makan EmpatJenis Katak Rana Asal Kelila, KabupatenJayawijaya, Irianjaya. Biota, Vol. III(2):58-62.
Kurniati. H. 2006. Jenis-jenis amfibi di TamanNasional Gunung Halimun, Jawa Barat.Zoo Indonesia. Vol. 15 (2): 107-120
Maneyro, R. and I. Rosa. 2004. Temporal andSpatial changes in The Diet of Hylapulchella (Anura, Hylidae) in SouthermUruguay. Phyllomedusa. Vol. III (2): 101-113.
Mistar dan D.T Iskandar. 2003. PanduanLapangan Amphibi Kawasan Ekosistem
Leuser. PILI-NGO Movement. Jakarta.
Marcelino, J. 2006. A Information on AmphibianBiology and Conservation.http;//amphibiaweb.org
McKay, J.L. (2006). A Field Guide to theAmphibians and Reptiles of Bali. KriegerPublishing Company, Malabar, Florida.
Nasaruddin. 2008. Karakteristik Habitat danBeberapa Aspek Biologi Kodok Raksasa(Limnonectes cf. grunniens). Vol.9 No.4 :182-187.
Naya, D. E., C. Veloso, F. Bozinovic. 2009. GutSize variation among Bufo spinulosuspopulations along a altitudinal (and dietary)Gradient. Ann. Zool. Fennici . 46:16-20.
Qing-qing X., Y. Dan, T. Chen, G. Bao-rong, Z.Qiu-jin, R. Xiao-zhen. 2004. Study on theDiversity of Foods of Rana limnocharis inFuzhou. Journal of Fujian NormalUniversity(Natural Science Edition).X(3):67-69,89.
Rahman, N. L, M. D. Kusrini and N. F. Haneda.2013. Food Preference of The Javan TreeFrog (Rhacophorus margaritifer) in MountGede Pangrango National. Park andCibodas Botanical Garden, West Jawa.Journal of Indonesian Natural History. Vol.1 No. 1.
Ren, Z., Zhu, B., Ma, E., Wen, J., Tu, T., Cao,Y., Hasegawa, M., and Zhong, Y. (2009).''Complete nucleotide sequence and genearrangement of the mitochondrial genomeof the crab-eating frog Fejervaryacancrivora and evolutionary implications .''Gene, 441, 148-155.
Schauble, C. 2004. Variation in The Body Sizeand Sexual Dimorphism AcrossGeographical and Environmental Space inthe Frogs Limnodynastes tasmaniensis andL. peronii. Biol J Linn Soc 82:39-54
Sole’, M; Beckmann, O; Pelz, B; Kwet, A;Engels, Wolf. 2005: Stomach-flushing fordiet analysis in anurans: an improvedprotocol evaluated in a case study inAraucaria forests, southern Brazil. Studieson Neotropical Fauna and Environment.40(1): 23-28.
Sugai, J. L. M. M., et all. 2012. Diet ofLeptodactylus fuscus (Amphibia : Anura :Lepidopter) in The Pantanal of MirandaRiver, Brazil. Biota Neotrop, 12 (1) : 99-104
Stebbins RC, Cohen NW. 1997. A NaturalHistory of Amphibians. New Jersey:Princeton Univ. Pr.
Tyynie, A., P. David, A. Ohler. 2010. Note an aCollection of Amphibians and Reptiles

Meliya Wati dan Elza Safitri 170
from Western Sumatra (Indonesia) with theDescription of a new Species of GenusBufo. Zootaxa: 1-43.
Wati, M. dan Hidayat, Y. 2013. Diet danMikrohabitat Dua Spesies Kodok B.melanostictus, Schneider (1799) dan B.asper, Gravenhorst (1829) Di dataranTinggi dan Dataran Rendah SumateraBarat. Hasil Penelitian Dosen Pemula.
Wien, J. J., J. Sukumaran, R. A. Pyron, dan R.M., Brown. 2009. Evolutionary andBiogeographic Origins of High TropicalDiversity in Old world Frogs (Ranidae).Original Article. The society for the studyof Evolution. 63-65:1217-1231.
Yu, T., dan Y. Guo. 2012. Trophic Ecology andMicrohabitat Utilization by The BufoGargarizans, Rana Guetheri and Ranalimnocharis in Southwestern China.Zoologia. XXIX (1):54-58.
Zhigang, Y., Zhao, E., Haitao, S., Diesmos, A.,Alcala, A., Brown, R., Afuang, L., Gee, G.,Sukumaran, J., Yaakob, N., Ming, L. T.,Chuaynkern, Y., Thirakhupt, K., Das, I.,Iskandar, D., Mumpuni, and Inger, R.(2009). Fejervarya cancrivora. In: IUCN2011. IUCN Red List of ThreatenedSpecies. www.iucnredlist.org. Downloadedon 11 November 2011.
Zug, G.R. 1993. Herpetology and IntroductoryBiology of Amphibians and Reptiles.Academic Press. Washington D

ISSN 978-602-14989-0-3 BioETI
Studi makromorfologi organ vegetatif dan mikromorfologi sporaAsplenium tenerum G. Forst dari Gunung Marapi di SumateraBaratMILDAWATI, ARDINIS ARBAIN, MAHFUD HUDA DAN HERMANSYAH
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKAsplenium tenerum G Forst merupakan salah satu spesies tumbuhan paku yang termasuk kedalam genus Asplenium.Pada studi ini akan dilakukan pengamatan karakteristik morfologi organ vegetatif serta karakteristik mikromorfologispora dari genus Asplenium tenerum G.Forst yang ditemukan pada Gunung Marapi di Sumatera Barat. Metode yangdigunakan pada penelitian ini yaitu metode koleksi langsung di lapangan dan selanjutnya dilakukan penelitian diLaboratorium Taksonomi Tumbuhan Jurusan Biologi Universitas Andalas dan Pengamatan SEM (Scanning ElectronMicroscope) di Laboratorium Teknik Mesin Universitas Andalas. Hasil yang didapatkan karakteristik organ vegetatifAsplenium tenerum G. Forst secara morfologi yaitu habitat epifit pada tumbuhan berbiji, bentuk rhizom bulat, stipesberbentuk pipih kadang-kadang bulat dan tidak berambut, frond majemuk, Pinna; daun tersusun pinnatus, pinnamenempel secara akroscopic pada basal lobus, Lamina ; macrophylloid, warna hijau muda sampai hijau tua, apektridentate, basis acuminatus, margin crenate, bentuk vena free, memiliki vena sejati dan tidak memiliki vena semu,dengan susunan lamina alternate. Sorus submarginal tersusun pada tulang daun dan memiliki indusia berbentuk elongatesedangkan berdasarkan data palinologi ditemukan tipe spora monolet, dengan diameter polar 35.51 µm – 44.62 µm dandiameter equatorialnya 26.12 µm – 28.21 µm dan rasio P/E 1.4 – 1.6 sehingga berdasarkan rasio tersebut maka bentukspora Asplenium tenerum G. Forst yang dapat dikemukakan pada penelitian ini adalah Prolate.
Key words: Asplenium, Morfologi, Spora, Monolet
Pendahuluan
Asplenium tenerum G. Forst termasuk kedalamordo Polypodiales, family Aspleniaceae,genus: Asplenium L (Holttum, 1967). A.tenerum G Forst merupakan tumbuhan yanghidup pada daerah-daerah hutan pegunungandengan ketinggian 400 m - 1500 m dpl (LIPI,1979, Lin, Y. X. & R. Viane, 2013). Tumbuhpaku ini epifit dengan perawakan yang kecildan berumpun. Rhizomnya lurus dan pendek,dengan beberapa rambut yang bercabang padabagian akarnya, membentuk spon yang mampumempertahankan kelembaban selama musimpanas, apek roset, pinnanya simple. Pinna 20 –30 pasang. Asplenium tenerum G. Forster,memiliki tinggi 30–65 cm, fronds caespitose;stipe hijau dengan panjang 12–30 cm, laminanarrowly triangular sampai linear, 20–38 cm,apek caudate, 1-pinnate, pinnae 15–25(–35)pasang, subopposite sampai alternate, tangkaipendek, basis asymmetrical, acroscopic.
Tumbuhan paku ini ditemukan di beberapadaerah seperti Hainan, Taiwan, India, Indonesia,Japan, Korea, Malaysia, Myanmar, Philippines,Sri Lanka, Vietnam; dan kepulauan Pasifik.Asplenium tenerum memiliki takson yangmasih beranekaragam. Berdasarkancircumscripsinya, spesies ini diyakini masihsangat diperlukan penelitian tentangmonographnya (Piggout, 1989 dan Holttum,1959 dan Lin, Y. X. & R. Viane, 2013).
Berdasarkan bentuk frondnya tumbuhan inimemiliki fronds yang pinnate-pinnatisect, miripdengan A. Sampsonii tetapi basal pinnanyamenyempit, disamping itu data tentang bentukmorfologi spora pada spesies ini belumditemukan laporannya. Tumbuhan pakuAsplenium tenerum G. Fosrt ditemukan padabeberapa kawasan di Sumatera Barat sepertiGunung Tandikek (Mildawati, Arbain danFitrah, 2013). Berdasarkan studi literatur danpengamatan Lapangan yang dilakukan padakawasan konservasi alam Gunung Marapi

Mildawati, Ardinis Arbain, Mahfud Huda dan Hermansyah 172
Sumatera Barat maka ditemukan spesies inipada beberapa tempat. Sebagai salah satu upayadalam mendokumentasikan flora dan faunayang ada di Sumatera Barat khususnya GunungMarapi serta mendukung upaya konservasijenis terutama spesies Asplenium tenerum G.Forst serta melengkapi data monograph baikdari aspek makromorfologi maupunmikromorfologinya maka dilakukanlahpenelitian tentang penggunaan karaktermorfologi organ vegetatif dan karaktermikromorfologi spora spesies Asplenium
tenerum dari Gunung Marapi di Sumatera Baratdengan tujuan 1. Mengetahui karaktermakromorfologi organ vegetatif padaAsplenium tenerum G. Forst 2. Mengetahuikarakter mikromorfologi spora genusAsplenium tenerum.
BAHAN DAN METODE
Pengambilan sampel Asplenium tenerum yangdigunakan pada penelitian ini dilakukan diGunung Marapi Sumatera Barat. GunungMarapi terdapat di Kabupaten Agam denganketinggian 2891 mdpl. Metode yang dipakaipada penelitian ini yaitu metode survey denganmelalukan survey pada Gunung Marapi.Penelitian survei botani umum dilakukan.Survei ini mencakup semua kelompok tanaman,dengan penekanan khusus pada Tumbuhanpaku genus Asplenium tenerum. Tanamandikumpulkan sepanjang perjalanan kemudian,diberi label, diukur (diameter dan tinggi),bentuk spora, karakteristik habitat, dll), difoto,dan posisinya ditentukan dengan GPS.Tanaman yang dikumpulkan disimpan padakantong plastik dengan alkohol 70% (metodeSchweinfurt). Selanjutnya dilakukanpemprosesan di Laboratorium Takonomitumbuhan dan Herbarium Universitas Andalas(ANDA) Jurusan Biologi Padang. untukpembuatan spesimen herba dan prosesidentifikasi. Identifikasi spesimen dilakukandengan menggunakan literature sesuai denganacuan sebagai berikut Copeland (1947),Holltum (1967), Piggott (1988), Harris dan
Harris (1994), Hickey dan King (2000).Selanjutnya dilakukan pengamatan spora dilaboratorium teknik mesin, Fakultas Teknik,Universitas Andalas mengacu kepada metodeyang digunakan oleh Herrero, A., Aedo,C.,Velayos, M and Viane, R.L.L (2001).
HASIL DAN PEMBAHASAN
Gunung Marapi merupakan sebuah kawasanSuaka Alam pegunungan yang terbentang padajajaran pegunungan yang ada di Sumatera Baratdengan ketinggian mencapai 2.891 meter diatas permukaan laut (m.dpl). Gunung initerletak di antara Kabupaten Tanahdatar danAgam, Sumatra Barat, dan merupakan gunungapi aktif yang hampir setiap tahunmengeluarkan letusannya. Abu vulkanik yangdikeluarkan oleh letusan Gunung Marapi iniakan menyuburkan berbagai jenis tumbuhanyang terdapat disekitarnya seperti halnyatumbuhan paku spesies A. tenerum. Keberadaanflora dari spesies A. tenerum didaerah tropisdiperkirakan sangat melimpah seperti halnyadi temukan di Philiphine, Thailand danIndonesia (Langenberger, G. 2000,Langenberger, et al. 2006, Boonkerd, et.al.2008, Mildawati et. al., 2013). Hal ini terbuktidari beberapa penelitian yang dilakukan diSumatera Barat spesies ini ditemukan padahabitat dan ketinggiaan yang berbedadiantaranya dari Gunung Tandikek dan GunungMarapi. Pada Gunung Marapi Asplenium
tenerum G. Forst ditemukan mulai dariketinggian 1.500 m.dpl – 2.109 m. dpl. Dataini menunjukkan bahwa distribusi spesies inicukup luas di Gunung Marapi.
Dalam rangka mendapatkan data yang lebihdetail tentang deskripsi jenis tumbuhan paku A.
tenerum maka sangat diperlukan data yanglebih detail tentang karakteristik yangditemukan pada spesies tersebut sehingga dapatmemudahkan pengenalan jenis baik dari segimakromorfologi organ vegetatif maupun darimikromorphologi sporanya.
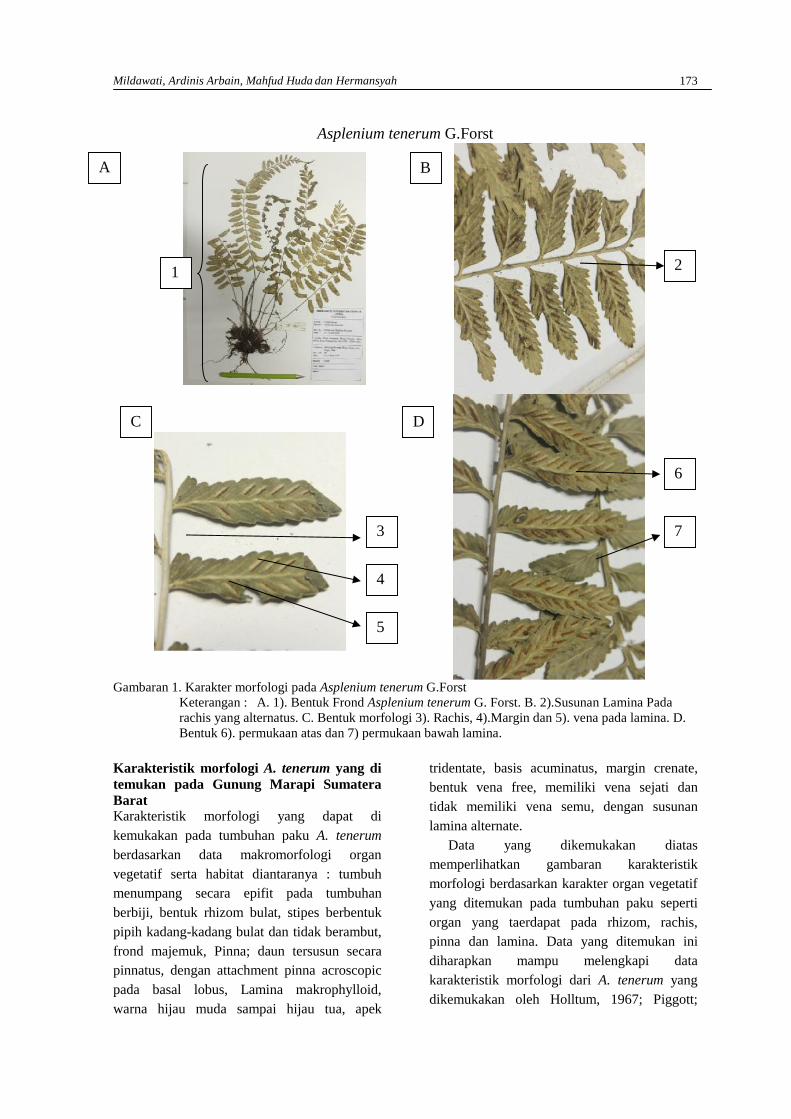
Mildawati, Ardinis Arbain, Mahfud Huda dan Hermansyah 173
Asplenium tenerum G.Forst
Gambaran 1. Karakter morfologi pada Asplenium tenerum G.ForstKeterangan : A. 1). Bentuk Frond Asplenium tenerum G. Forst. B. 2).Susunan Lamina Padarachis yang alternatus. C. Bentuk morfologi 3). Rachis, 4).Margin dan 5). vena pada lamina. D.Bentuk 6). permukaan atas dan 7) permukaan bawah lamina.
Karakteristik morfologi A. tenerum yang ditemukan pada Gunung Marapi SumateraBaratKarakteristik morfologi yang dapat dikemukakan pada tumbuhan paku A. tenerum
berdasarkan data makromorfologi organvegetatif serta habitat diantaranya : tumbuhmenumpang secara epifit pada tumbuhanberbiji, bentuk rhizom bulat, stipes berbentukpipih kadang-kadang bulat dan tidak berambut,frond majemuk, Pinna; daun tersusun secarapinnatus, dengan attachment pinna acroscopicpada basal lobus, Lamina makrophylloid,warna hijau muda sampai hijau tua, apek
tridentate, basis acuminatus, margin crenate,bentuk vena free, memiliki vena sejati dantidak memiliki vena semu, dengan susunanlamina alternate.
Data yang dikemukakan diatasmemperlihatkan gambaran karakteristikmorfologi berdasarkan karakter organ vegetatifyang ditemukan pada tumbuhan paku sepertiorgan yang taerdapat pada rhizom, rachis,pinna dan lamina. Data yang ditemukan inidiharapkan mampu melengkapi datakarakteristik morfologi dari A. tenerum yangdikemukakan oleh Holltum, 1967; Piggott;
A
DC
B
1 2
7
6
3
4
5
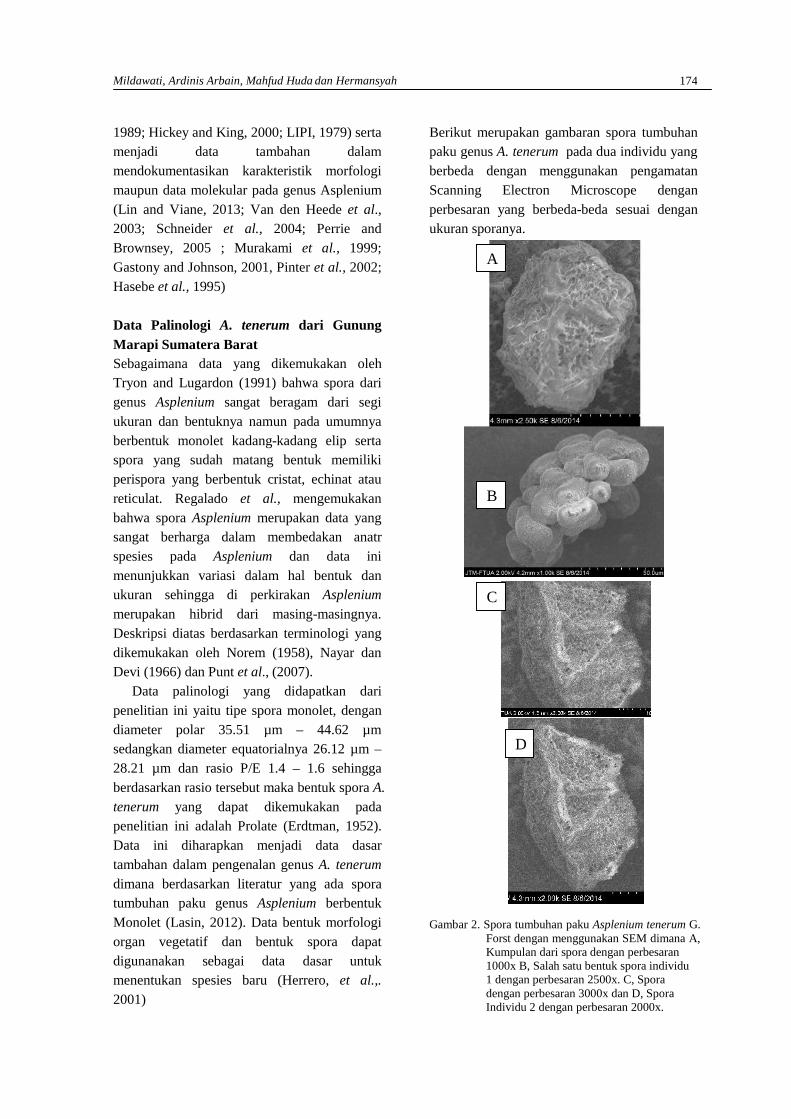
Mildawati, Ardinis Arbain, Mahfud Huda dan Hermansyah 174
1989; Hickey and King, 2000; LIPI, 1979) sertamenjadi data tambahan dalammendokumentasikan karakteristik morfologimaupun data molekular pada genus Asplenium(Lin and Viane, 2013; Van den Heede et al.,2003; Schneider et al., 2004; Perrie andBrownsey, 2005 ; Murakami et al., 1999;Gastony and Johnson, 2001, Pinter et al., 2002;Hasebe et al., 1995)
Data Palinologi A. tenerum dari GunungMarapi Sumatera BaratSebagaimana data yang dikemukakan olehTryon and Lugardon (1991) bahwa spora darigenus Asplenium sangat beragam dari segiukuran dan bentuknya namun pada umumnyaberbentuk monolet kadang-kadang elip sertaspora yang sudah matang bentuk memilikiperispora yang berbentuk cristat, echinat ataureticulat. Regalado et al., mengemukakanbahwa spora Asplenium merupakan data yangsangat berharga dalam membedakan anatrspesies pada Asplenium dan data inimenunjukkan variasi dalam hal bentuk danukuran sehingga di perkirakan Aspleniummerupakan hibrid dari masing-masingnya.Deskripsi diatas berdasarkan terminologi yangdikemukakan oleh Norem (1958), Nayar danDevi (1966) dan Punt et al., (2007).
Data palinologi yang didapatkan daripenelitian ini yaitu tipe spora monolet, dengandiameter polar 35.51 µm – 44.62 µmsedangkan diameter equatorialnya 26.12 µm –28.21 µm dan rasio P/E 1.4 – 1.6 sehinggaberdasarkan rasio tersebut maka bentuk spora A.
tenerum yang dapat dikemukakan padapenelitian ini adalah Prolate (Erdtman, 1952).Data ini diharapkan menjadi data dasartambahan dalam pengenalan genus A. tenerum
dimana berdasarkan literatur yang ada sporatumbuhan paku genus Asplenium berbentukMonolet (Lasin, 2012). Data bentuk morfologiorgan vegetatif dan bentuk spora dapatdigunanakan sebagai data dasar untukmenentukan spesies baru (Herrero, et al.,.
2001)
Berikut merupakan gambaran spora tumbuhanpaku genus A. tenerum pada dua individu yangberbeda dengan menggunakan pengamatanScanning Electron Microscope denganperbesaran yang berbeda-beda sesuai denganukuran sporanya.
Gambar 2. Spora tumbuhan paku Asplenium tenerum G.Forst dengan menggunakan SEM dimana A,Kumpulan dari spora dengan perbesaran1000x B, Salah satu bentuk spora individu1 dengan perbesaran 2500x. C, Sporadengan perbesaran 3000x dan D, SporaIndividu 2 dengan perbesaran 2000x.
A
D
C
B

Mildawati, Ardinis Arbain, Mahfud Huda dan Hermansyah 175
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Karakteristik khusus secara morfologi yang
membedakan Asplenium tenerum G. Forstdengan spesies lain dapat dilihat dari bentukrhizom yang bulat, stipes berbentuk pipihkadang-kadang bulat dan tidak berambut,frond majemuk, Pinna dengan attachmentacroscopic pada basal lobus, apek tridentate,basis acuminatus, margin crenate, bentukvena free, memiliki vena sejati dan tidakmemiliki vena semu, dengan susunanlamina alternate.
2. Karakteristik palinologi yang menjadi cirikhas Asplenium tenerum G. Forst adalahtipe spora monolet, diameter polar 35.51 µm– 44.62 µm dan diameter equatorial 26.12µm – 28.21 µm dan rasio P/E 1.4 – 1.6,berdasarkan rasio tersebut maka bentukspora Asplenium tenerum G. Forst padapenelitian ini adalah Prolate
DAFTAR PUSTAKA
Bloom Copeland, A. C. 1947. Genera Filicium,The Genera of Ferns. The CronicalBotanica Company. Waltham. USA.
Erdtman, G. 1952. Pollen Morphology danPlant Taxonomy Angiosperm. Almquistdan Wiksell Stockhom. Sweden.
Gastony, G. J. & Johnson, W. P. 2001.Phylogenetic placements of Loxoscaphethecifera (Aspleniaceae) andActiniopteris radiata (Pteridaceae) basedon analysis of rbcL nucleotide sequences.Amer. Fern J. 91: 197–213.
Harris, J.G and M.W. Harris. 1994. PlantIdentification Terminology. An IllustratedGlossary. Spring lake Publishing. UnitedStates of America.
Hasebe, M., Wolf, P. G., Pryer, K. M., Ueda,K., Ito, M., Sano, R., Gastony, G. J.,Yokoyama, J., Manhart, J. R.,Murakami, N., Crane, E. H., Haufler, C. H.& Hauk, W. D. 1995. Fern phylogenybased on rbcL nucleotide sequences. Amer.Fern J. 85: 134–181.
Herrero, A., Aedo,C.,Velayos, M & Viane,R.L.L.2001 ; A new species of Asplenium
(Aspleniaceae, Pteridophyta) fromEquatorial Guinea. Ann. Bot. Fennici 38 ;175 - 180
Hickey, M dan C. King. 2000. The CambridgeIllustrated Glossary of Botanical Term.CambridgeUniversity Press. UnitedKingdom.
Holttum, R. E. 1967. A Revised Flora ofMalaya Volume II. Ferns of Malaya.Government Printing Office. Singapore.
Lashin, G.M, 2012. Palynological Studies ofSome Species of Aspleniaceae-Pteridophyta. American Journal ofPlant Sciences, 2012, 3, 397-402
Lin, Y. X. & R. Viane. 2013. Aspleniaceae. Pp.267–316 in Z. Y. Wu, P. H. Raven & D. Y.
Hong, eds., Flora of China, Vol. 2–3(Pteridophytes). Beijing: Science Press; St.Louis: Missouri Botanical GardenPress
LIPI. 1980. Jenis Paku Indonesia. LembagaBiologi Nasional. LIPI. Bogor.
Mildawati, Ardinis Arbain, Hary Fitrah. 2013.Aspleniaceae ofTandikek Mountain,West
Sumatra. The Journal Of Tropical LifeScience. Vol. 3, 202 – 206
Murakami, N., Nogami, S., Watanabe, M. &Iwatsuki, K. 1999. Phylogeny ofAspleniaceae inferred from rbcLnucleotide sequences. Amer. Fern J. 89:232–243.
Nayar, B. K. & Devi, S. (1966). Sporemorphology of the Pteridoid Ferns. I. ThePteridoid ferns. Grana Palynol., 6,476–503.
Norem, W. L. 1958. Keys for the classificationof spores and pollen. J. Paleontol., 32,666-676.
Pinter, I., Bakker, F., Barrett, J., Cox, C., Gibby,M., Henderson, S., Morgan-Richards, M.,
Rumsey, F., Russell, S., Trewick, S.,Schneider, H. & Vogel, J. 2002.Phylogenetic and biosystematicrelationships in four highly disjunctpolyploid complexes in the subgeneraCeterach and Phyllitis in Asplenium(Aspleniaceae). Org. Diver. Evol. 2: 299
311.Punt, W., Hoen, P. P., Blackmore, S., Nilsson,
S. & Le Thomas, A. 2007. Glossary ofpollen and spore terminology. Rev.2nd ed. Rev. Paleobot. Palynol., 143
Schneider, H., Russell, S. J., Cox, C. J., Bakker,F., Henderson, S., Gibby, M. & Vogel, J.

Mildawati, Ardinis Arbain, Mahfud Huda dan Hermansyah 176
C. 2004. Chloroplast phylogeny ofasplenioid ferns based on rbcL and trnL-Fspacer sequences (Polypodiidae,Aspleniaceae) and its implications for thebiogeography. Syst. Bot. 29: 260–274.
Piggott, A.G. 1988. Ferns of Malaysia inColour. Tropical Press SDN.BHD.Malaysia
Tryon, R. M. & Lugardon, B. (1991). Spores ofthe Pteridophyta: Surface, wall structure
and diversity based on electronmicroscope studies. New York: Springer.
Van den Heede, C. J., Viane, R. L. L. & Chase,M. W. 2003. Phylogenetic analysis ofAsplenium subgenus Ceterach(Pteridophyta: Aspleniaceae) based onplastid and nuclear ribosomal ITSDNA sequences. Amer. J. Bot. 90: 481–493

ISSN 978-602-14989-0-3 BioETI
Pengkayaan vitamin E pada pakan untuk pematangan gonadIkan Mali (Labiobarbus festivus , Heckel)NETTI ARYANI, EFAWANI DAN NUR ASIAH
Departement of Aquaculture Faculty Fisheries and Marine Science, Riau University, Campus Binawidya Km 12,5Panam, Indonesia.E-mail: [email protected]
ABSTRACTThe aim the research is to increase the reproductive potential of L. festivus female that istime matured gonads, somaticovi index, fecundity, egg diameter, hatching rate and time to hatching in the control group (feed without enrichment ofvitamin E) and exposed to 150mg/kg, 300 mg/kg, 450mg/kg feed of vitamin E. Treatment of vitamin E levels weresignificant (p <0.05) different with respect to time ripe gonads, ovi somatic index, fecundity and egg diameter. Femalesexposed to 300 µg/kg feed of vitamin E can increased time matured gonads 67 days, somatic ovi index 14.95%, absolutefecundity 13547egss/spaw, relatif fecundity 90 eggs/ g gonadoweight and egg diameter 1.16 mm.
Key words: Labiobarbus festivus, vitamin, gonadal maturation, fecundity, eggs diameter
Pendahuluan
Sungai Kampar Kanan adalah salah satu sungaiyang terdapat di Provinsi Riau yang berperansangat besar untuk mendukung aktivitasmanusia yang tinggal di daerah sekitarnya.Aktivitas di sempadan sungai tersebut adalahperkebunan kelapa sawit dan industry,sedangkan aktifitas di badan air sungai adalahtransportasi, pertambangan pasir danpenangkapan ikan yang tidak selektif (Aryaniet al, 2013).
Ikan mali (L. festivus, CYPRINIDAE)merupakan salah satu spesies ikan asli yanghidup di Sungai Kampar (Fithra dan Siregar,2010), bernilai ekonomi tinggi dandiperdagangkan secara luas sebagai ikankonsumsi dengan nilai jual Rp. 30.000,-/kg(Warsaet al, 2009. Keistimewaan ikan malidisukai oleh konsumen karena memiliki rasayang gurih. Permasalahannya pada saat ini ikanmali sudah mulai berkurang di Sungai KamparKanan, karena penangkapan dilakukan secarakontinue dengan beragam alat tangkap yangtidak selektif. Oleh karena itu penting dimulaiupaya konservasi ikan mali secara insitu yaitudengan melakukan domestikasi dalam rangkamemproduksi benih secara massal.
Kontinuitas benih merupakan salah satufaktor pembatas dalam kegiatan budidaya.Menurut (Izquierdo et al, 2001; Aryani, 2001dan Aryani et al, 2010). Vitamin E dapatmempercepat waktu pencapaian matang gonaddan meningkatkan jumlah telur yangdiovulasikan (Aryani, 2001, Basri, 2002,Syandri et al, 2004). Fernandez et al., (1995)menyatakan ikan yang kekurangan vitamin E didalam pakannya akan berpengaruh terhadapperkembangan gonad, fekunditas dan daya tetastelur. Kebutuhan vitamin Edi dalam pakan ikanuntuk proses reproduksi bervariasi. Untuk ikanpatin (Pangasiussutchii) dibutuhkan 190 mg/kgpakan (Mokoginta et al, 2000), ikan baung(Mystusnemurus Bagridae)) adalah150 mg/kgpakan (Aryani, 2002), dan ikan garing (Tor
douronensis Cyprnidae) adalah 439,29 mg/kgpakan (Syandri et al, 2004). Pengayaan vitaminE ke dalam pakan dosis berbeda dalam rangkameningkatkan daya reproduksi ikan malipenting dilakukan agar larva dapat diproduksisecara massal.
BAHAN DAN METODE
Induk ikan mali L.festivus diperoleh darinelayan di Sungai Kampar Kanan, Desa PadangLuas, Kecamatan Kampar Provinsi Riau. Indukikan mali diperlihara selama tiga bulan dengan

Netti Aryani, Efawani dan Nur Asiah 178
berat rata-rata 132-140 g/ekor. Induk ikandiberi pakan dengan cara ditebar secara meratakedalam karamba dengan jumlah 12 buah yangberukuran 200×60×50 cm dengan padat tebar10 ekor/karamba. Karamba ditempatkan dipinggir sungai dengan kedalaman rata-rata duameter. Temperatur air sungai antara 26-28o C.Pakan diberikan dua kali sehari dengan jumlah5 % dari berat biomass. Komposisi Proximatepakan terdiri atas kadar air (% berat kering)12,0 %, protein kasar 38.0 %, lipid 2.0 %,karbohidrat 5.6 % dan kadar abu 13.0 %.
Kematangan induk ikan diperiksa dengancara mengambil oosit secara invivo menurutmetoda Syandri (1997). Oosit dimasukkan kedalam larutan Sera (6:3:1,70 % ethanol, 40 %formaldehyde dan 99.5 % acetic acid) yangakan digunakan untuk klasifikasi cytoplasma.Setelah lima menit, posisi dari nucleus diditermiansi 4 tahap yaitu :stage 1 germinal vesicle in berada di intinucleus, stage 2 awal migrasi dari germinalvesicle (less than half of radius), stage 3 akhirmigrasi dari germinal vesicle (more than half ofradius), stage 4 Inti nuclues berada dalamposisi germinal vesicle breakdown (GVBD)
Vitamin E yang digunakan untuk pengayaanpakan adalah dalam bentuk soft kapsulmengandung d-αtokoferol 100 IU vitamin E.Vitamin E terlebih dahulu dilarutkan dalamminyak jagung, kemudian dicampurkankedalam pakan secara merata dengan dosis 0,150, 300, 450 mg/kg pakan, selanjutnya pakantersebut dikeringkan selama 15 menit diruangan terbuka tanpa cahaya matahari.Penelitian terdiri dari tiga perlakuan dan tigakali ulangan. Perlakuan pakan kontrol tanpadiberi vitamin E. Perlakuan ke dua, ke tiga danke empat dosis vitamin E masing-masing 150,300, 450 mg/kg pakan. Pakan diberikan duakali sehari sebanyak 5 % dari berat biomass.Pemijahan L. festivus menggunakan GnRHayang mengandung dopamin antagonis dosis 0,5ml/kg berat induk. Sampel telur masing-masingperlakuan diberi ulangan. Sebelum dilakukanovulasi induk ditimbang dan ovulasi dilakukanpada pagi hari dengan cara stripping dan telurditampung dalam mangkuk plastik. LarutanGilson dengan komposisi ( 60% alkohol, 880ml air suling, 15 ml asam nitric, 18 ml asamacetic dan 20 gram mercury chlorida) dalam100 ml digunakan untuk mengawetkan sampeltelur. Diameter telur masing-masing perlakuansebanyak 30 butir diukur dengan menggunakanmikroskop Olympus Cx21. Fertilisasi telurdihitung setelah sepuluh jam dengan
menggunakan stereomikroskop. Telur ditebarsecara merata ke dalam 12 akurium ukuran40x20x20 cm dengan volume air 10 liter,masing–masing tiga ulangan untuk kontrol dantiga ulangan untuk perlakuan pakan yangdikayakan dengan vitamin E. Jumlah telur padakontrol dan masing-masing perlakuan sebanyak200 butir. Temperatur air, DO dan pH diukursecara berkala yaitu satu kali dalam tiga hari.Waktu pencapaian matang gonad dihitungmulai dari induk ikan diberi perlakuan pakanhingga mencapai matang gonad (dalam hari).Indek Ovi Somatik diukur dengan meng-gunakan formula IOS = BTO/BI x100%; BTO:berat telur yang dihasilkan pada saat ovulasi;BI= berat induk; Fekunditas Absolut(FA)=Jumlah telur per gram x berat gonad,Fekunditas relatif diukur dengan menggunakanformula FR=Jumlah telur per gram bobotgonad.
Data dianalisis dengan One Way Anova dandilanjutkan dengan Uji . Seluruh data dirancangdan dianalisis dengan program SPSS versi 13.1.
HASIL DAN PEMBAHASAN
Waktu pencapaian matang gonad danIndeks Ovi SomatikRata-rata waktu pencapaian matang gonad padakontrol adalah 93 hari, sedangkan padaperlakuan pengayaan pakan dengan vitamin Erata-rata berkisar antara 67-87 hari (Tabel 1).Terdapat perbedaan nyata (p< 0.05) antarperlakuan pakan yang dikayakan denganvitamin E dan lebih cepat matang gonad.. rata-rata jumlah telur ovulasi pada group kontroladalah5,60%, sedangkan pada group perlakuanvitamin E berkisar antara 7.12% to 14.95%.Terdapat perbedaan nyata (p <0.05) antarakontroldan group perlakuan vitamin E.
Table 1-Time matured the gonads and SomaticOviIndeksLevel vitamin E
(mg/kg feed)Time maturedthe gonadal
(days)
SomaticOviIndeks (%)a
Control150300450
93±6a
87±4b
67±4c
81±7b
5,60±0,89a
7,12±1,85b
14,95±1,91b
12,77±2,37b
a Weight of eggs ∙100%/female weight.abc Values with the different superscript in each columnare significantly different from each other (P < 0.05).

Netti Aryani, Efawani dan Nur Asiah 179
Fekunditas dan diameter telurRata-rata fekunditas absolu ikan L.festivuspadagroup kontrol adalah 8.497 telur/pemijahan,sedangkan pada group perlakuan berkisarantara 9.903-13.547 telur/pemijahan (Tabel 2).Fekunditas absolut memperlihatkanpeningkatan dengan meningkatnya dosisvitamin E, walaupun peningkatan terjadi padadosis 150 dan 300 mg / kg feed. Pada dosis450 mg/kg pakan tidak meningkatkanfekunditas jika dibandingkan dengan dosis 300mg/kg pakan, tetapi lebih tinggi daripada dosis150mg/kg pakan. Fekunditas absolut padakontrol dengan dosis 150 mg/kg pakan tidakberbeda nyata (p>0,05), tetapi berbeda nyata(p<0,05) pada dosis 300 dan 450 mg/kg feed.Rata-rata fekunditas relatif (jumlah telur/ gberat gonad) untuk empat group (Tabel 2).Fekunditas relatif pada group kontrol adalah 60eggs / g berat gonad .On the grouptwo,threeandfour, respectively 47, 90 and 60eggs/g gonadal weight.The relative fecunditywas significant (p<0,05) differencesbetweenthe control and exposure groups. Tidak terdapatperbedaan (P < 0.05) diameter telur untukseluruh level dosis vitamin E termasuk kontrol(Table 2),Diameter telur pada group kontroladalah 0.95 mm, sedangkan dengan perlakuanvitamin Eberkisar antara 1.06-1.16 mm.
Tabel 2. Fecundity and eggs diameter in thecontrol and treatment groupsexposure to various levels of vitaminE
DosevitaminE(mg/kgfeed)
Averagebodyweight(g)
Fecundity(number ofeggs perspawn )
Relatiffecundity(number of eggsper g gonadalweight )
Eggs diameter(mm)
Control 140±46 8497±1234a 61±2.0a 0.95±0.02a
(n=50)150 132±25 9903±668a 75±2.0b 1.06±0.09b(50)300 138±36 13547±2166ab98±5.0c 1.16±0.00b
(50)450 136±50 12462±1865ab91±6.0d 1.13±0.02b
(50)
Values with the different superscript in eachcolumn are significantly different from eachother (P < 0.05).
Pengkayaan vitamin E dalam pakan indukberpengaruh terhadap waktu pencapaianmatang gonad. Waktu matang gonad yangpaling cepat diperoleh pada perlakuan P3(dosisvitamin E 300 mg/kg pakan) berpengaruh nyataterhadap waktu pencapaian matang gonad (P>0,05) merupakan dosis yang terbaik biladibandingkan dengan perlakuan lainnya. Hasilperhitungan jumlah hari yang dibutuhkan untukmencapai matang gonad yang paling cepatterdapat pada perlakuan P3 (dosis vitamin E300 mg/kg berat badan) yaitu selama67,00±4,35 hari (Tabel 1). Lebih cepatnyawaktu pencapaian matang gonad padaperlakuan P3 diduga vitamin E dibutuhkanuntuk pematangan gonad ikan dan mempunyaiperanan sebagai antioksidan sehingga dapatmempertahankan asam lemak essensial didalam telur. Hal ini sesuai dengan hasilpenelitian Aryani (2001) pengkayaan vitamin E100 mg/kg pakan waktu pencapaian matanggonad yang diperoleh 73 hari, sedangkan darihasil penelitian Basri (2002) pengkayaanvitamin E sebesar 338,72 mg/kg pakan waktumatang gonad yang terbaik diperoleh selama 58hari. Hasil penelitian Syandri (2004)pengkayaan vitamin E dengan dosis 439,29mg/kg dapat meningkatkan potensi reproduksiikan Garing (Tor douronensis Blkr) denganwaktu pencapaian matang gonad diperoleh 116hari. Dari beberapa hasil penelitian tersebutperbedaan lama waktu pencapaian matanggonad yang diperoleh disebabkan perbedaanspecies ikan yang digunakan.
Indek Ovi SomatikPerhitungan nilai Indeks Ovi Somatik
dilakukan untuk mengetahui persentase bobotgonad berbanding bobot tubuh untuk setiapinduk ikan. Hasil yang diperoleh untuk IOS(Tabel 2) menunjukkan bahwa semuaperlakuan pakan yang diperkaya denganvitamin E memiliki nilai IOS yang lebih tinggidibandingkan dengan kontrol. Nilai IOS yangterbesar diperoleh pada perlakuan pengkayaanvitamin E sebesar 300 mg/kg pakan sebesar14,95±1,91 % (P> 0,05). Hal ini sesuai dengan

Netti Aryani, Efawani dan Nur Asiah 180
hasil peneltian Basri (2002) pengkayaanvitamin E sebesar 338,72 mg/kg pakan ikangurami (O. gouramy) menghasilkan IOS3,17 %, sedangkan perlakuan pakan tanpavitamin E nilai IOS sebesar 0,90%. Dari hasilpenelitian Eriza dan Syandri (2001) pemberianvitamin E pada pakan ikan jambal siam (P.
hypoptahlmus) sebesar 152,38 mg/kg pakanmenghasilkan nila IOS sebesar 8,98 % dantanpa pengkayaan vitamin E nilai IOS yangdiperoleh sebesar 6,14 %. Selanjutnya darihasil penelitian Aryani (2002) pengkayaanvitamin E pada pakan ikan Baung sebesar 100mg/kg pakan nilai IOS yang diperoleh sebesar9,16 %. Diduga besarnya nilai IOS yangdiperoleh akibat peranan vitamin E pada prosesperkembngan gonad yaitu vitamin Emempengaruhi biosintesis vitellogenin di hati.Oksidasi lemak yang terjadi pada vitellogenindicegah dengan vitamin E sebagai antioksidanterhadap lemak. Hal ini akan menyebabkanpertambahan jumlah vitellgenin pada oosit danmeningkatkan bobot gonad sehingga persentaseIOS menjadi besar (Arfah et al., 2013)FekunditasFekunditas adalah jumlah total telur yangdiproduksi pada setiap pemijahan atau jumlahtelur / berat tubuh (Fernandez-Palacios et al.,1995). Fekunditas yang dihasilkan dalam satusiklus reproduksi dapat dipengaruhi olehketersediaan pakan, vitamin E dan C di dalampakan (Izquierdo et al., 2001), ukuran tubuh(Syandri et al, 2013) dan lingkungan(Hardjamulia, 1987; Khal et al., 2008). Hasilpenelitian ini membuktikan bahwa perlakuanpakan dengan vitamin E berpengaruh nyata(p< 0,05) terhadap fekunditas ikan Mali (Tabel1). Fekunditas tertinggi terdapat pada pakanyang dikayakan dengan vitamin E sebesar 300mg/kg pakan dengan jumlah fekunditassebanyak 13.547±2166 butir dan fekunditasterkecil terdapat pada control yaitu sebanyak8497±1234 butir. Perbedaan fekunditas ikanmali (L. festivus) antar perlakuan terjadi akibatperbedaan dosis vitamin E di dalam pakan.Penurunan jumlah fekunditas, dilaporkan pada
beberapa spesies ikan laut, bisa disebabkanoleh pengaruh ketidakseimbangan nutrisi padasistem endokrin otak-hipofisis-gonad ataudengan pembatasan pada ketersediaankomponen biokimia untuk pembentukan telur(Izquierdo et al., 2001). Semakin banyakvitellogenin yang dibawa ke ovarium, makasemakin banyak butir-butir telur yang dibentukdalam ovarium. Fekunditas juga dapatdipengaruhi oleh nilai gonad somatic indek(GSI), Tang dan Affandi (2004) menyatakanbahwa semakin besar persentase GSI, makasemakin banyak telur telur yang dihasilkan olehinduk. Aryani et al., (2013) melaporkan bahwafekunditas ikan mali yang berasal dari perairanWaduk Koto Panjang adalah 6.902-18.756 butirdengan kisaran bobot tubuh51,9–122,33 g.Diameter telurDarihasil penelitian pengkayaan vitamin Eberpengaruh nyata terhadap diameter telur ((P<0,05). Diameter telur terbesar terdapat padaperlakuan pengkayaan pakan dengan vitamin Esebesar 300 mg/kg pakan dengan nilai1,16±0,00 mm. Hasil penelitian Aryani et al.,(2013) diameter telur ikan mali yang berasaldari perairanWaduk Koto Panjang berkisarantara 0,85–1,13 mm. Besarnya diameter telurakibat pengkayaan vitamin E ke dalam pakandiduga pada proses vitellogenesis menyebakanakumulasi kuning telur dan menyebabkankeberadaan asam lemak di dalam telur dapatdipertahankan. Aktivitas ini membuat jumlahdan ukuran granula kuning telur bertambahbesar sehingga volume dan diameter kuningtelur meningkat (Sumantri, 2006 dalam Arfahet al, 2013). Menurut Yulfiperius (2003)hubungan vitamin E dengan vitellogenin dalamperkembangan oosit ternyata melaluiprostaglandin. Dalam hal ini prostaglandindisintesis secara enzimatik dengan meng-gunakan asam lemak essensial. Vitamin Eberfungsi sebagai antioksidan sehingga asamlemak dapat dipertahankan. Oleh karena ituvitamin E memberikan pengaruh terhadapperkembangan oosit induk betina. MenurutArfah et al., (2013)perkembangan telur dan

Netti Aryani, Efawani dan Nur Asiah 181
penyerapan vitellogenin ini berhenti ketikaoosit mencapai ukuran maksimal, selanjutnyadiduga ketika penyerapan vitellogenin berhenti,maka aktivitas vitamin E yang membantudalam proses vitellogenesis juga berhenti.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Perlakukan pengkayaan vitamin E 300 mg/kgpakan merupakan dosis terbaik untukpematangan gonad ikan mali (L. festivus)dengan waktu pencapaian matang gonadselama 67,00±4,35 hari, Indeks Ovi Somatik14,95±1,91%, fekunditas 13.547±2166 butirdan diameter telur 1,16±0,00 mm.
UCAPAN TERIMAKASIHUcapan terimakasih disampaikan kepada KetuaLembaga Penelitian Universitas Riau yangtelah mendanai penelitian ini melalui danaHibah Bersaing tahun anggaran 2014. Ucapanini juga ditujukan kepada rekan dan mahasiswayang telah membantu pelaksanaan penelitian dilapangan dan dilaboratorium.
DAFTAR PUSTAKA
Arfah H, Melati dan Setiawati M. 2013.Suplementasi Vitamin E dengan dosisberbeda pada pakan terhadap kinerjareproduksi induk betina ikan komet(Carassius auratus auratus). JurnalAkuakultur Indonesia 12(1) : 14-18.
Aryani N., 2001. Penggunaan vitamin E padapakan untuk pematangan gonad ikanbaung (Mystusnumerus CV). JurnalPerikanan dan IlmuKelautan, 6 (1): 28-36.
Aryani N., Pamungkas dan Adelina. 2010.Kajian kadar nutrisi telur ikan baung(Mystus nemurus) sebagai dasar untukpengkayaan pakan buatan. ProsidingSeminar Nasional Fakultas PerikananIlmu kalautan Universitas RiauPekanbaru.
Aryani N., Efawani dan Nurasiah, 2013.Kebiasaan makan dan penampilanreproduksi ikan Mali (Labiobarbusfestivus , Heckel) di Waduk Koto Panjang.Prosiding Seminar 2nd National and
International Fakultas Perikanan dan IlmuKelautan Univesitas Riau.
Basri, Y. 2002. Penambahan vitamin E padapakan buatan induk dalam usahapeningkatan kecepatan kematangan gonad,fekunditas, kondisi telur, fertilitas dandaya tetas telur ikan gurami(Osphronemus gourami Lacepede).Fisheries Journal Garing, 1 (11) : 56-82.
Eriza Mdan H. Syandri. 2001. Penambahanvitamin E dalam pakan buatan untukmeningkatkan potensi reproduksi ikanjambal siam (Pangasius hypthalmus).Fiheries Journal Garing 2 (10): 57-73
Fernadez P, Izquirdo M, Robaina L, ValenciaA, Salhi M, Jose M, 1995. Effect of n-3HUFA level in broodstock diets on eggquality on gilthead sea bream,Sparatusauratus L. Aquaculture 132: 325-337.
Fithra, RY., dan Siregar, Y.I., 2010.Keanekaragaman ikan sungai Kampar.Inventarisasi dari sungai Kampar Kanan.Journal of Environmental Science 2 (4) :139-147.
Hardjamulia A, 1987. Beberpa Aspek pengaruhpenundaan dan frekwensi pemijahanterhadap potensi reproduksi ikan Mas(Cyprinus carpio L). Disertasi ProgramPascasarjana Institut Pertanian Bogor.
Izquierdo, M.S; Fernandezs-Palacios.H;Tacon.A.G.J. 2001. Effect of broodstocknutrition on repductive performance of.Fish. Aquacultur197 : 25-42.
Kahl. U., Stephan H,I,, Robert J,R., Jurgen. B.2008. The impact of water levelfluctuations on the year class strength ofRoach: Implications for fish stockmanagement . Limnologica 38 : 258–268
Mokoginta, Syahrizal M, Zairin MJR. 2002.Pengaruh kadar vitamin E(α-tokopherol)pakan terhadap kadar lemak, asam lemakessensial telur dan derajat tetas telur ikanlele Clariasbatrachus Linn. JurnalAkuakultur Indonesia 1 : 75-59.
Syandri, H .1997. The development of oocytesand testes of Bilih (Mystacoleucuspadangensis Bleeker) in Singkarak Lake.Journal ofFisheries Garing 2 (6):1-8.
Syandri, H. 2004. Penggunaan vitamin E untukpeningkatkan potensi reproduks iikangaring (Tor douronensis Blkr). JurnalDinamika Pertanian XIX (1) : 141 – 151.
Tang U dan Affandi, R. 2004. BiologiReproduksi Ikan Pekanbaru; Pusat

Netti Aryani, Efawani dan Nur Asiah 182
Penelitian Kawasan Pantai an PerairanUniversitas Riau.
Warsa, A. Nastiti.A.S, Krismono danNurfiarini.A. 2009. Sumberdaya perikanantangkap di waduk Koto Panjang . Bawal2 (3) :93-97.
Yulfiperius. 2003. Penambahan Vitamin Edalam formulasi pakan induk dapatmemperbaiki kualitas reproduksinya.Makalah Falsafah Sains. Bogor ; InstitutPertanian Bogor.

ISSN 978-602-14989-0-3 BioETI
Penerapan sistem resirkulasi pada proses domestikasi danpembesaran Ikan Juaro (Pangasius polyuranodonNIKEN AYU PAMUKAS 1) DAN MULYADI 2)
Fisheries and Marine Sciences Faculty, Riau UniversityE-mail: [email protected] 1); [email protected] 2)
ABSTRACTThe study on the implementation of resirculation system to domesticate and to grow out of juaro juveniles (Pangasius
polyuranodon) has been carried out. Several filter materials were used into four kinds of treatment, namely aquariumusing aerators (control), aquarium using spon, carcoal and palm fiber as well as zeolite. Results of the study showed thatdifferent filter materials were significantly affecting in the juaro juveniles culture media. As long adaptation period (oneweek), juaro juveniles (Pangasius polyuranodon) could wean to consume fish meal by involving other fish species in thesame culture media. It was invented that treatment of resirculation system combining with zeolite could increased waterquality parameters such as DO2 (3,9-4,56 mg/L), CO2 (8,6-9,15 mg/L), NH3 (0,02-0,10 mg/L), NO2 (0,01-0,08 mg/L),NO3 (0,01-0,09 mg/L) respectively, but the other water quality parameters (pH and temperature ) were not differsignificantly. The best results were achieved at the same treatment namely absolute growth rates (9,24 grams), absolutelength rate (5,14 cm), specific growth rates (1,76 %), biomass growth rate (62,23 gram), food efficiency (28,07%), FCR(3,45%) and survival rates (86,67 %) respectively.
Key words: catfish (Mystus nemurus C.V), filter materials, resirculation, aquaponic
Pendahuluan
Ikan Juaro (Pangasius polyuranodon) termasukke dalam keluarga Pangasidae (Saanin, 1984)memiliki ciri-ciri yaitu tidak memiliki sisik,sirip punggung berjari-jari keras dan tajam(Kottelat et al, 1993). Ikan juaro ini, memilikitekstur daging yang lembut, rasanya gurih danharganya cukup mahal, sehingga sangatprospektif untuk dikembangkan sebagai salahsatu jenis ikan budidaya masa depan. Saat iniuntuk memenuhi permintaan terhadap ikanjuaro (Pangasius polyuranodon) masihmengandalkan hasil tangkapan dari alam.Penangkapan yang tidak terkendali olehmanusia mengakibatkan terganggunya habitatikan-ikan di perairan, sehingga ikan-ikantersebut dikhawatirkan mengalami kepunahan.Untuk mencegah kepunahan ikan ini usahabudidaya merupakan suatu langkah strategisyang dapat dilakukan. Kegiatan budidaya dapatberlangsung dengan baik apabila kegiatandomestikasi dikuasai terlebih dahulu.Selanjutnya, proses domestikasi dapatdipercepat jika ikan yang akan didomestikasi
kan diberikan pakan yang sesuai untukmenopang kehidupannya.
Penelitian mengenai studi kebiasaanmakanan ikan Juaro (Pangasius polyuranodon)di Perairan Sungai Musi, Sumatera Selatantelah dilakukan oleh Ramadhan (2008) danBiologi Reproduksi Ikan Juaro (Pangasiuspolyuranodon) di Daerah Aliran Sungai Musi,Sumatera Selatan telah dilakukan olehErnawati dkk., (2009). Dari penelitian Ernawatidkk., (2009) dilaporkan bahwa, perlu dilakukanupaya pengelolaan ikan Juaro untuk meningkatkan populasi dan produksi ikan ini melaluikegiatan domestikasi dan pembudidayaan.
Untuk menjadikan ikan juaro (P.
polyuranodon) sebagai komoditas budidaya,langkah-langkah penjinakan (domestikasi)harus dituntaskan. Ikan juaro tersebut harusbisa hidup serta dapat tumbuh dan berkembangbiak pada kondisi terkontrol. Untuk itu,langkah-langkah domestikasi tersebut harusdimulai dengan melakukan kajian-kajianbiologi, ekologi dan penangkaran awal denganmelakukan uji coba untuk mendapatkan kondisilingkungan optimal yang dapat menopang

Niken Ayu Pamukas dan Mulyadi 184
kehidupan dan pertumbuhannya. Disamping itu,usaha domestikasi perlu mendapat prioritasguna menjaga kelestarian ikan ini. Melaluiusaha domestikasi, diharapkan ikan dapatberadaptasi pada lingkungan yang terkontroldan dapat menerima pakan buatan yangdiberikan serta bertahan hidup pada kepadatanyang tinggi dan tahan terhadap penanganan.Untuk itu, perlu ditemukan teknologi yangterbaik dengan mengoptimalkan pemanfaatansumberdaya pakan buatan disamping pakanalami untuk aktivitas domestikasi danpembesaran sehingga dihasilkan benih ikanjuaro yang berkualitas dan ketersediaannyamencukupi untuk dijadikan input dalam usahabudidaya dalam rangka menghasilkan ikankonsumsi.
Teknik budidaya secara intensif untukmemacu pertumbuhan ikan juaro juga ditujukanpada perbaikan manajemen kualitas air yangharus diterapkan mulai dari tahap domestikasi,pemijahan, pemeliharaan larva, pendederan danpembesaran. Untuk meningkatkan danmenjadikan kualitas air yang ideal sesuaidengan kondisi yang diinginkan oleh ikan danbiota akuatik lainnya, berbagai metode dapatdilakukan seperti teknik penyaringan,pengendapan dan penyerapan. Bahan yangdapat digunakan untuk meningkatkan kualitasair tersebut juga beranekaragam seperti pasir,kerikil, arang batok, ijuk, bubur kapur, tawas,batu, zeolit dan lain-lain (Mulyadi, Hasibuandan Romiantoyo, 2010). Selanjutnya beberapateknik yang juga dapat digunakan untukmemperbaiki kualitas air atau menghilangkanpengaruh buruk air kotor agar menjadi layakdan sehat untuk kehidupan ikan dalambudidaya yaitu aerasi, sirkulasi air,penggunaan pemanas air, pergantian air segardan filtrasi.
Inovasi teknologi sistem resirkulasi denganmenggunakan berbagai macam bahan filterdiharapkan mampu mengurangi limbah danmeningkatkan produktifitas. Teknologi ini padaprinsipnya disamping menghemat penggunaanlahan dan air juga meningkatkan efisiensi usaha
melalui pemanfaatan hara dari sisa pakan danmetabolisme ikan, serta merupakan salah satusistem budidaya ikan yang ramah lingkungan.Oleh karena itu penelitian ini dimaksudkanuntuk mengungkap jenis pakan yang efektifdiberikan pada ikan juaro pada tahapdomestikasi dan pembesaran denganmenerapkan sistem resirkulasi tertutup.
Penelitian ini bertujuan untuk menemukanjenis pakan yang sesuai pada tahap domestikasidan pembesaran serta sistem resirkulasidengan menggunakan berbagai jenis filterterhadap kualitas air serta menemukanteknologi resirkulasi terbaik dalam domestikasidan pembesaran ikan juaro. Adapun manfaatdari penelitian ini adalah memberikaninformasi kepada masyarakat tentang teknikdomestikasi dan pembesaran ikan juaro padalahan dan sumberdaya air terbatas dengansistem resirkulasi.
BAHAN DAN METODE
Penelitian ini dilaksanakan selama sembilanbulan dari bulan Maret sampai September 2014di Laboratorium UPT Kolam Pembenihan,Fakultas Perikanan dan Ilmu KelautanUniversitas Riau.
Penempatan wadah sistem resirkulasidilakukan di Laboratorium UPT KolamPembenihan, Fakultas Perikanan dan IlmuKelautan. Wadah pemeliharaan ikan digunakanadalah akuarium yang berukuran (60 x 40 x 40)cm3 dengan volume air yang diisi sebanyak 48liter dilengkapi pompa air dengan kekuatan 20watt untuk mengalirkan air ke bakpemeliharaan ikan. Bak filter yang digunakanterbuat dari talang air dengan volume 24 liter.Selanjutnya air dari bak filter akan mengalirkembali melalui pipa PVC dengan diameter 2,5cm ke bak pemeliharaan benih ikan juaro.Perbandingan antara wadah filter denganwadah pemeliharaan ikan adalah 1 : 2. Hewanuji yang digunakan untuk penelitian ini adalahbenih ikan juaro berukuran 3-5 cm yang berasaldari aliran Sungai Siak, Desa Tampan. Pakanyang diberikan pada pemeliharaan adalah pelet
2

Niken Ayu Pamukas dan Mulyadi 185
ikan terapung buatan pabrik FF-999 dan pakanalami (cacing sutera).
Penelitian ini dilakukan dengan metodeeksperimen menggunakan Rancangan AcakLengkap (RAL) dua faktor, 3 taraf perlakuandan 3 kali ulangan. Sebagai perlakuan padapenelitian ini : 5 ekor ikan juaro per akuariumtanpa substrat filter (P0), 5 ekor ikan juaro perakuarium menggunakan substrat filter spons(P1), 5 ekor ikan juaro per akuariummenggunakan substrat filter ijuk dan arangaktif (P2) dan 5 ekor ikan juaro per akuariummenggunakan substrat filter batu zeolit (P3).
Penelitian utama dilakukan setelah ikanJuaro berhasil beradaptasi dengan baik padawadah budidaya dan telah dapat menerimapellet 100%. Penelitian utama dilakukanselama 2 bulan dengan variabel kerja yangdiamati pada penelitian ini adalah parameterutama yaitu ; pertumbuhan ikan (pertumbuhanbobot mutlak, pertumbuhan panjang mutlak,laju pertumbuhan spesifik dan pertambahanbiomassa), Efisiensi Pakan, konversi pakan(FCR), kelangsungan hidup ikan (SR),ammonia (NH3), nitrit (NO2-) dan nitrat (NO3).Parameter penunjang yaitu kualitas air (suhu,pH, oksigen terlarut dan karbondioksida bebas).Data yang diperoleh berupa parameter utamadilakukan uji keragaman (ANOVA) apabilaterjadi perbedaan yang nyata antar perlakuandilakukan uji lanjut SNK dengan menggunakanprogram SPSS.
HASIL DAN PEMBAHASAN
Benih ikan Juaro sebagai hewan uji diperolehdari hasil tangkapan nelayan di Desa Tampanpada aliran sungai Siak, dimana habitat tempatikan ini hidup pada perairan yang mengalir,relatif asam dengan tingkat kesadahan (pH 6),suhu berkisar antara 27-30 0C dan warna airkecoklatan. Pada pinggiran sungai dijumpaiperumahan penduduk yang kegiatan MCKnyadilakukan di sungai tersebut. Pinggiran sungaisebagian ditutupi oleh tanaman eceng gondok(Gambar 1).
Gambar 1. Lokasi fishing ground ikan Juaro diSungai Siak
Benih ikan ditangkap dengan menggunakanseser, dimana hasil tangkapan kemudiandikumpulkan oleh nelayan pada keramba yangditambatkan di pinggiran sungai tanpa diberipakan. Menurut nelayan setempat Sungai Siakmerupakan fishing ground dari ikan juaro ini,namun sangat sulit untuk mempertahankan ikanjuaro tetap hidup pada wadah budidaya sepertibak terpal maupun akuarium karena ikan inisangat mudah stress.
Pengangkutan benih ikan dilakukan denganmenggunakan wadah terbuka berupa drumplastik berukuran diameter 75 cm dan tinggi125 cm yang dilubangi sampingnya. Drumdilengkapi dengan aerator batterai dan diberi esuntuk mempertahankan suhu air selamadiperjalanan tetap stabil.
Selama pengangkutan dari Sungai Siak keLaboratorium UPT Pembenihan FAPERIKAUNRI dibutuhkan waktu lebih kurang 1 jam,dan pada saat pengangkutan tersebut terjadikematian ikan sebanyak 5 ekor, hal inidisebabkan ikan mengalami luka akibatpenanganan pada saat pemindahan darikeramba ke perahu nelayan.
Adaptasi benih ikan juaro sebelum diberiperlakuan dilakukan selama 2 bulan. Adaptasidilakukan pada bak semen, akuarium dan bakfiber yang dilengkapi dengan sistem resirkulasi,daun ketapang dan tanaman eceng gondokuntuk meniru habitat alaminya. Pada bak semendan akuarium seluruh ikan mati karena
3

Niken Ayu Pamukas dan Mulyadi 186
mengalami stress, sedangkan pada bak fiberikan dapat bertahan tetap hidup. Hal inidisebabkan kondisi pada bak fiber yang palingmendekati habitat alaminya, dimana warna airkecoklatan karena adanya daun ketapang(Gambar 2). Menurut Akbar (2013) daunketapang mengandung asam humic dan tanninyang menyebabkan air berwarna agak gelapkecoklatan, disamping itu juga dapatmenghambat pertumbuhan berbagai jenisbakteri yang membahayakan kesehatan ikan.
Gambar 2. Adaptasi Ikan Juaro di Bak Fiber
Pada hari pertama adaptasi, ikan juaroterlihat bergerombol di bawah tanaman air didekat batu aerasi dan cenderung berdiam diri.Pada saat itu ikan juaro tidak diberi makandengan tujuan apabila ikan dalam keadaanlapar akan lebih mudah menerima pakan yangdiberikan. Pada hari kedua adaptasi terjadikematian ikan pada bak semen, akuarium danbeberapa ekor pada bak fiber. Hal ini karenapada tubuh ikan terdapat luka, sehinggamenimbulkan jamur pada permukaan kulitnyadan juga oleh kanibalisme dari ikan juarotersebut. Ikan yang masih hidup pada bak fiber
direndam dengan Kalium Permanganat(KMnO4) dengan dosis 0,2 g/L. Pada hariketiga adaptasi ikan Juaro diberi pakan Tubifex,ikan juaro terlihat belum mau merespon pakanyang diberikan. Pada hari selanjutnya prosespembelajaran ikan (Weaning) dilakukan denganmenggunakan ikan pendamping yaitu ikan nila.Ikan terlihat sudah mulai mau merespon pakanTubifex yang diberikan. Kemudian berangsur-angsur sedikit demi sedikit ikan diberi peletsampai benih ikan dapat menerima pellet.
Setelah ikan juaro mulai menerima pakanyang diberikan, ikan terlihat sehat dan mulaiberenang aktif. Selama adaptasi ikan diberipakan Tubifex dan pellet secara adlibitum,dengan frekuensi pemberian 3 kali sehari yaitupagi, siang dan sore hari. Setelah 1 minggumasa adaptasi, ikan sudah bisa menerima pellet100%. Pada bak fiber dilakukan penyiponansetiap pagi hari untuk menjaga kualitas airtetap baik. Selama adaptasi suhu air pada bakfiber berkisar antara 27-30 0C, pH 5,5 danOksigen terlarut 4 – 4,5 mg/L.
Hasil pengamatan terhadap suhu, pH,oksigen terlarut (DO), karbon dioksida (CO2),amoniak (NH3), nitrit (NO2
-) dan nitrat (NO3-)
selama penelitian dapat dilihat pada Tabel 1.Tabel 1 menunjukkan kisaran rata-rata suhupada semua perlakuan selama penelitian relatifhampir sama, yaitu berkisar antara 28,50-29,330C, suhu pada semua wadah penelitian masihdalam kisaran yang baik untuk mendukungpertumbuhan ikan Juaro berdasarkan nilaistandar bakumutu untuk kegiatan budidaya airtawar menurut PP No. 82 tahun 2001. MenurutBoyd (1982) perbedaan suhu tidak melebihi 100C masih tergolong baik dan kisaran suhu yangbaik untuk organisme di daerah tropis adalah25-320C. Surya Mina (2014) menyatakanbahwa suhu yang ideal bagi budidaya ikanadalah suhu yang stabil di kisaran 28-30 0Cserta tidak terjadi perbedaan suhu air yangmencolok antara siang dan malam tidak lebihdari 50C. Pada kondisi ini ikan akanmemberikan respon maksimal ketika diberi
4
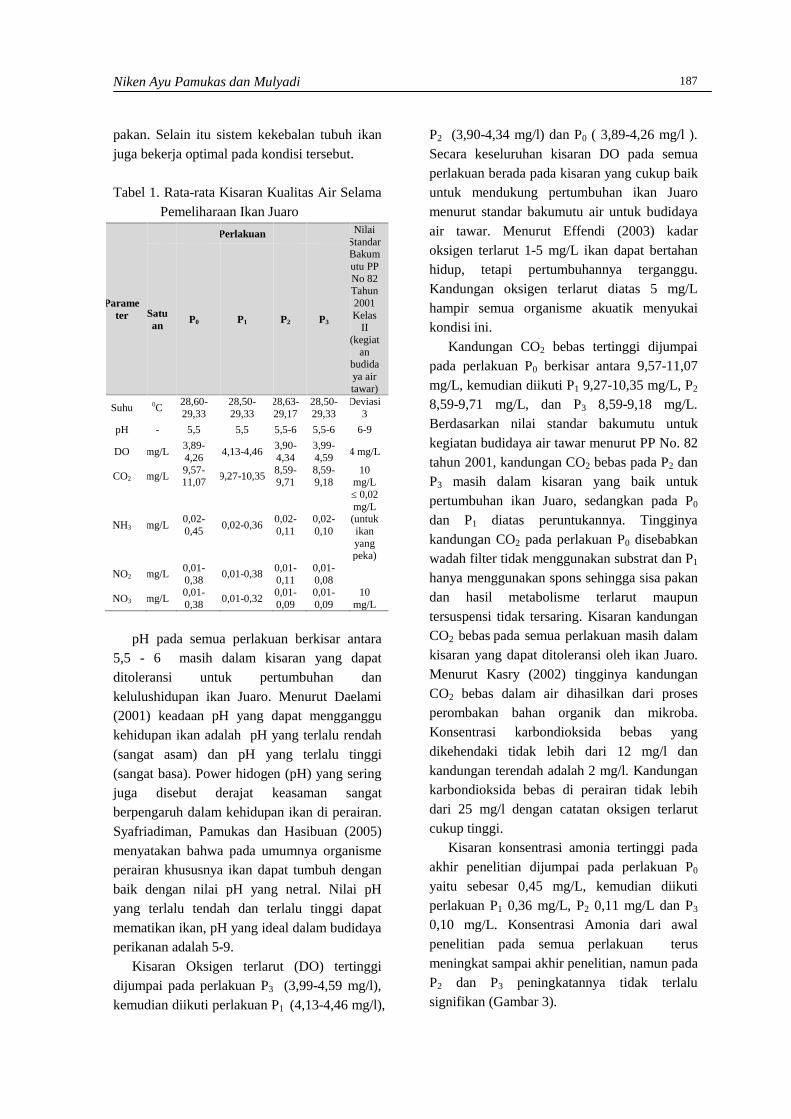
Niken Ayu Pamukas dan Mulyadi 187
pakan. Selain itu sistem kekebalan tubuh ikanjuga bekerja optimal pada kondisi tersebut.
Tabel 1. Rata-rata Kisaran Kualitas Air SelamaPemeliharaan Ikan Juaro
Parameter
Perlakuan NilaiStandarBakumutu PPNo 82Tahun2001Kelas
II(kegiat
anbudidaya airtawar)
Satuan
P0 P1 P2 P3
Suhu 0C28,60-29,33
28,50-29,33
28,63-29,17
28,50-29,33
Deviasi3
pH - 5,5 5,5 5,5-6 5,5-6 6-9
DO mg/L3,89-4,26
4,13-4,463,90-4,34
3,99-4,59
4 mg/L
CO2 mg/L9,57-11,07
9,27-10,358,59-9,71
8,59-9,18
10mg/L
NH3 mg/L0,02-0,45
0,02-0,360,02-0,11
0,02-0,10
≤ 0,02mg/L(untukikanyangpeka)
NO2 mg/L0,01-0,38
0,01-0,380,01-0,11
0,01-0,08
NO3 mg/L0,01-0,38
0,01-0,320,01-0,09
0,01-0,09
10mg/L
pH pada semua perlakuan berkisar antara5,5 - 6 masih dalam kisaran yang dapatditoleransi untuk pertumbuhan dankelulushidupan ikan Juaro. Menurut Daelami(2001) keadaan pH yang dapat mengganggukehidupan ikan adalah pH yang terlalu rendah(sangat asam) dan pH yang terlalu tinggi(sangat basa). Power hidogen (pH) yang seringjuga disebut derajat keasaman sangatberpengaruh dalam kehidupan ikan di perairan.Syafriadiman, Pamukas dan Hasibuan (2005)menyatakan bahwa pada umumnya organismeperairan khususnya ikan dapat tumbuh denganbaik dengan nilai pH yang netral. Nilai pHyang terlalu tendah dan terlalu tinggi dapatmematikan ikan, pH yang ideal dalam budidayaperikanan adalah 5-9.
Kisaran Oksigen terlarut (DO) tertinggidijumpai pada perlakuan P3 (3,99-4,59 mg/l),kemudian diikuti perlakuan P1 (4,13-4,46 mg/l),
P2 (3,90-4,34 mg/l) dan P0 ( 3,89-4,26 mg/l ).Secara keseluruhan kisaran DO pada semuaperlakuan berada pada kisaran yang cukup baikuntuk mendukung pertumbuhan ikan Juaromenurut standar bakumutu air untuk budidayaair tawar. Menurut Effendi (2003) kadaroksigen terlarut 1-5 mg/L ikan dapat bertahanhidup, tetapi pertumbuhannya terganggu.Kandungan oksigen terlarut diatas 5 mg/Lhampir semua organisme akuatik menyukaikondisi ini.
Kandungan CO2 bebas tertinggi dijumpaipada perlakuan P0 berkisar antara 9,57-11,07mg/L, kemudian diikuti P1 9,27-10,35 mg/L, P2
8,59-9,71 mg/L, dan P3 8,59-9,18 mg/L.Berdasarkan nilai standar bakumutu untukkegiatan budidaya air tawar menurut PP No. 82tahun 2001, kandungan CO2 bebas pada P2 danP3 masih dalam kisaran yang baik untukpertumbuhan ikan Juaro, sedangkan pada P0
dan P1 diatas peruntukannya. Tingginyakandungan CO2 pada perlakuan P0 disebabkanwadah filter tidak menggunakan substrat dan P1
hanya menggunakan spons sehingga sisa pakandan hasil metabolisme terlarut maupuntersuspensi tidak tersaring. Kisaran kandunganCO2 bebas pada semua perlakuan masih dalamkisaran yang dapat ditoleransi oleh ikan Juaro.Menurut Kasry (2002) tingginya kandunganCO2 bebas dalam air dihasilkan dari prosesperombakan bahan organik dan mikroba.Konsentrasi karbondioksida bebas yangdikehendaki tidak lebih dari 12 mg/l dankandungan terendah adalah 2 mg/l. Kandungankarbondioksida bebas di perairan tidak lebihdari 25 mg/l dengan catatan oksigen terlarutcukup tinggi.
Kisaran konsentrasi amonia tertinggi padaakhir penelitian dijumpai pada perlakuan P0
yaitu sebesar 0,45 mg/L, kemudian diikutiperlakuan P1 0,36 mg/L, P2 0,11 mg/L dan P3
0,10 mg/L. Konsentrasi Amonia dari awalpenelitian pada semua perlakuan terusmeningkat sampai akhir penelitian, namun padaP2 dan P3 peningkatannya tidak terlalusignifikan (Gambar 3).
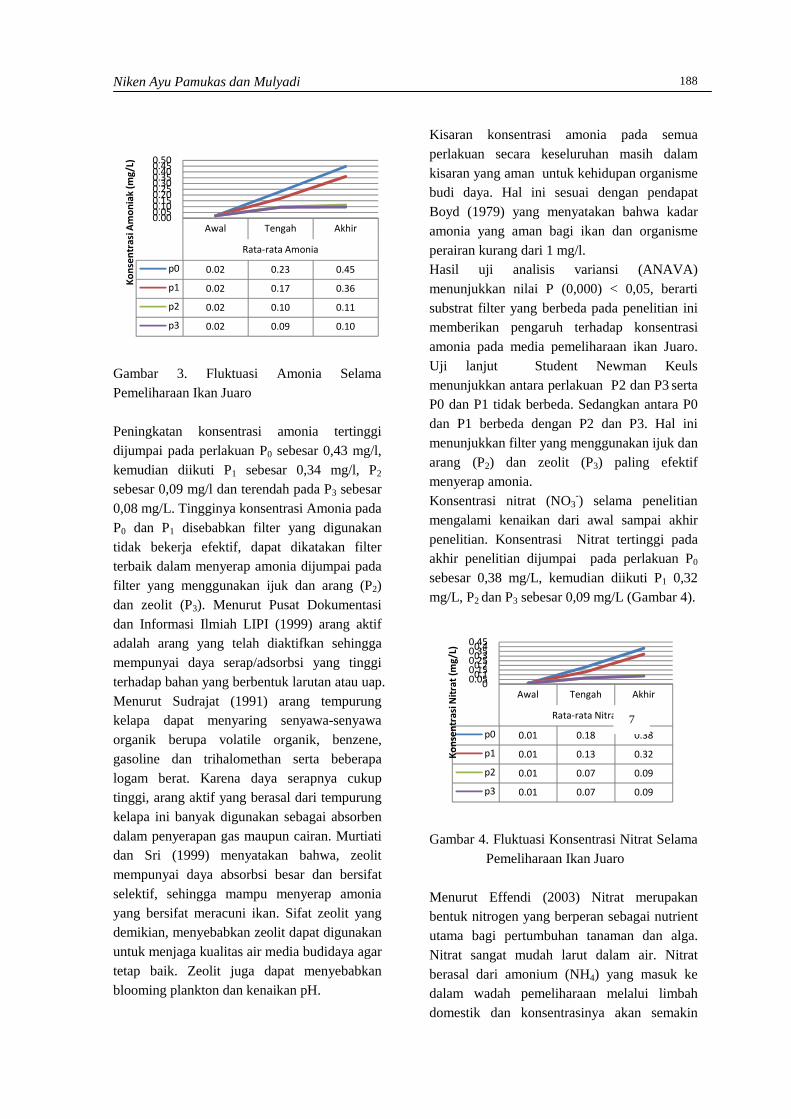
Niken Ayu Pamukas dan Mulyadi 188
Gambar 3. Fluktuasi Amonia SelamaPemeliharaan Ikan Juaro
Peningkatan konsentrasi amonia tertinggidijumpai pada perlakuan P0 sebesar 0,43 mg/l,kemudian diikuti P1 sebesar 0,34 mg/l, P2
sebesar 0,09 mg/l dan terendah pada P3 sebesar0,08 mg/L. Tingginya konsentrasi Amonia padaP0 dan P1 disebabkan filter yang digunakantidak bekerja efektif, dapat dikatakan filterterbaik dalam menyerap amonia dijumpai padafilter yang menggunakan ijuk dan arang (P2)dan zeolit (P3). Menurut Pusat Dokumentasidan Informasi Ilmiah LIPI (1999) arang aktifadalah arang yang telah diaktifkan sehinggamempunyai daya serap/adsorbsi yang tinggiterhadap bahan yang berbentuk larutan atau uap.Menurut Sudrajat (1991) arang tempurungkelapa dapat menyaring senyawa-senyawaorganik berupa volatile organik, benzene,gasoline dan trihalomethan serta beberapalogam berat. Karena daya serapnya cukuptinggi, arang aktif yang berasal dari tempurungkelapa ini banyak digunakan sebagai absorbendalam penyerapan gas maupun cairan. Murtiatidan Sri (1999) menyatakan bahwa, zeolitmempunyai daya absorbsi besar dan bersifatselektif, sehingga mampu menyerap amoniayang bersifat meracuni ikan. Sifat zeolit yangdemikian, menyebabkan zeolit dapat digunakanuntuk menjaga kualitas air media budidaya agartetap baik. Zeolit juga dapat menyebabkanblooming plankton dan kenaikan pH.
Kisaran konsentrasi amonia pada semuaperlakuan secara keseluruhan masih dalamkisaran yang aman untuk kehidupan organismebudi daya. Hal ini sesuai dengan pendapatBoyd (1979) yang menyatakan bahwa kadaramonia yang aman bagi ikan dan organismeperairan kurang dari 1 mg/l.Hasil uji analisis variansi (ANAVA)menunjukkan nilai P (0,000) < 0,05, berartisubstrat filter yang berbeda pada penelitian inimemberikan pengaruh terhadap konsentrasiamonia pada media pemeliharaan ikan Juaro.Uji lanjut Student Newman Keulsmenunjukkan antara perlakuan P2 dan P3 sertaP0 dan P1 tidak berbeda. Sedangkan antara P0dan P1 berbeda dengan P2 dan P3. Hal inimenunjukkan filter yang menggunakan ijuk danarang (P2) dan zeolit (P3) paling efektifmenyerap amonia.Konsentrasi nitrat (NO3
-) selama penelitianmengalami kenaikan dari awal sampai akhirpenelitian. Konsentrasi Nitrat tertinggi padaakhir penelitian dijumpai pada perlakuan P0
sebesar 0,38 mg/L, kemudian diikuti P1 0,32mg/L, P2 dan P3 sebesar 0,09 mg/L (Gambar 4).
Gambar 4. Fluktuasi Konsentrasi Nitrat SelamaPemeliharaan Ikan Juaro
Menurut Effendi (2003) Nitrat merupakanbentuk nitrogen yang berperan sebagai nutrientutama bagi pertumbuhan tanaman dan alga.Nitrat sangat mudah larut dalam air. Nitratberasal dari amonium (NH4) yang masuk kedalam wadah pemeliharaan melalui limbahdomestik dan konsentrasinya akan semakin
Awal Tengah Akhir
Rata-rata Amonia
p0 0.02 0.23 0.45
p1 0.02 0.17 0.36
p2 0.02 0.10 0.11
p3 0.02 0.09 0.10
0.000.050.100.150.200.250.300.350.400.450.50
Kons
entr
asi A
mon
iak
(mg/
L)
Awal Tengah Akhir
Rata-rata Nitrat
p0 0.01 0.18 0.38
p1 0.01 0.13 0.32
p2 0.01 0.07 0.09
p3 0.01 0.07 0.09
00.050.10.150.20.250.30.350.40.45
Kons
entr
asi N
itrat
(mg/
L)
8
7
Niken Ayu Pamukas dan Mulyadi 189
berkurang bila semakin jauh dari titikpembuangan yang disebabkan adanya aktifitasmikroorganisme di dalam air, contohnya ;bakteri nitrozomonas. Mikroba tersebut akanmengoksidasi ammonium menjadi nitrat.Kisaran konsentrasi nitrat tertinggi selamapenelitian dijumpai pada P0 yaitu sebesar 0,01-0,38 mg/L, kemudian diikuti P1 0,01-0,32 mg/Ldan P2 dan P3 yaitu sebesar 0,01-0,09 mg/L.
Hasil uji analisis variansi (ANOVA)menunjukkan nilai P (0,000) < 0,05, berartisubstrat filter yang berbeda memberikanpengaruh terhadap Konsentrasi Nitrat padamedia pemeliharaan ikan juaro. Uji lanjutStudent-Newman-Keuls menunjukkan antaraperlakuan P2 dan P3 serta P0 dan P1 tidakberbeda. Sedangkan antara P0 dan P1 berbedadengan P2 dan P3.
Konsentrasi nitrit (NO2-) pada semua
perlakuan terus meningkat dari awal sampaiakhir penelitian. Konsentarsi nitrit tertinggipada akhir penelitian dijumpai pada perlakuanP0 dan P1 sebesar 0,38 mg/L, kemudian diikutiperlakuan P2 0,11 mg/L dan P3 0,08 mg/L(Gambar 5).
Gambar 5. Fluktuasi Konsentrasi Nitrit SelamaPemeliharaan Juaro
Senyawa nitrit merupakan hasil reduksisenyawa nitrat juga oksidasi senyawa amoniakoleh mikroorganisme. Selain itu senyawa nitritjuga berasal dari ekskresi fitoplankton. Nitritmemuncak pada akhir penelitian disebabkanoleh oksidasi amoniak yang tidak lengkap atau
karena menurunnya nitrat (NO3-) menjadi nitrit
(NO2-). Menyebabkan terganggunya proses
metabolik dalam organisme, yang akhirnyadapat menyebabkan kematian pada ikan(Effendi, 2003).
Kisaran Nitrit tertinggi selama penelitiandijumpai pada perlakuan P0 dan P1 yaituberkisar antara 0,01-0,38 mg/L, kemudiandiikuti P2 0,01-0,11 mg/l dan P3 0,01– 0,08mg/L. Kisaran konsentrasi Nitrit pada semuaperlakuan masih dalam batas yang dapatditoleransi oleh ikan Juaro, hal ini sesuaidengan pendapat Siikavuopio dan Saether(2006) yang menyatakan bahwa konsentrasinitrit pada level 16 mg/L merupakankonsentrasi lethal dosis, 1-5 mg/L sudahmembahayakan bagi ikan dan batas amannyaadalah kecil dari 1 mg/L. Sedangkan menurutSyafriadiman, Pamukas dan Hasibuan (2005)konsentrasi nitrit di atas 2 mg/l untuk jangkawaktu yang lama bersifat mematikan bagi ikan.
Hasil uji analisis variansi (ANOVA)menunjukkan nilai P (0,001) < 0,05, berartisubstrat filter yang berbeda memberikanpengaruh terhadap Konsentrasi Nitrit padamedia pemeliharaan ikan Juaro. Uji lanjutStudent-Newman-Keuls menunjukkan antaraperlakuan P2 dan P3 serta P0 dan P1 tidakberbeda. Sedangkan antara P0 dan P1 berbedadengan P2 dan P3.Hasil pengamatan terhadap pertumbuhan bobotmutlak, pertumbuhan panjang mutlak, lajupertumbuhan spesifik, pertumbuhan bobotbiomassa, efisiensi pakan, konversi pakan dankelulushidupan ikan Juaro dapat dilihat padaTabel 2.
Tabel 2 menunjukkan Pertambahan bobotmutlak ikan Juaro tertinggi selama penelitiandijumpai pada perlakuan P3 yaitu sebesar 9,24gram, kemudian diikuti perlakuan P1 (3,76gram), P3 (3,60 gram) dan P0 (3,11 gram). Halini disebabkan karena pada perlakuan P3 ikandapat memanfaatkan pakan secara efektif untukpertumbuhan, disamping itu juga dipengaruhioleh kualitas airnya yang lebih bagusdibandingkan perlakuan lainnya, sehingga
Awal Tengah Akhir
Rata-rata Nitrit
p0 0.01 0.15 0.38
p1 0.01 0.14 0.38
p2 0.01 0.08 0.11
p3 0.01 0.07 0.08
00.050.10.150.20.250.30.350.40.45
Kons
entr
asi N
itrit
(mg/
L)
9
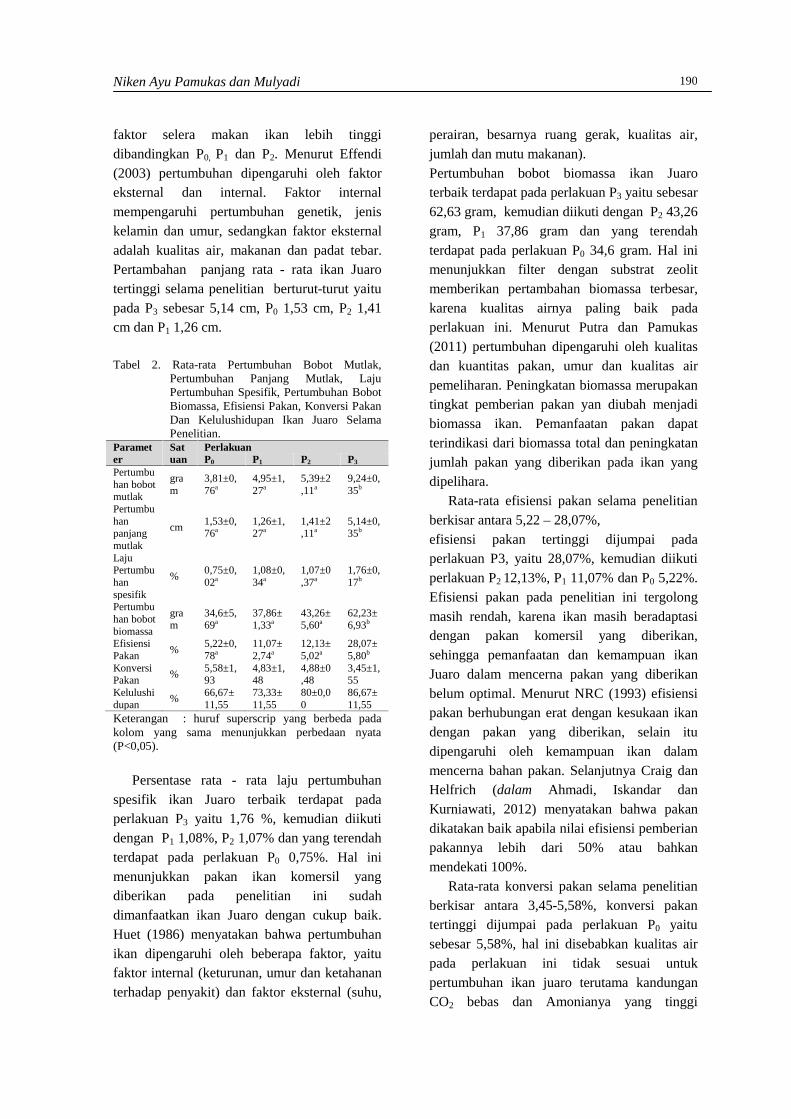
Niken Ayu Pamukas dan Mulyadi 190
faktor selera makan ikan lebih tinggidibandingkan P0, P1 dan P2. Menurut Effendi(2003) pertumbuhan dipengaruhi oleh faktoreksternal dan internal. Faktor internalmempengaruhi pertumbuhan genetik, jeniskelamin dan umur, sedangkan faktor eksternaladalah kualitas air, makanan dan padat tebar.Pertambahan panjang rata - rata ikan Juarotertinggi selama penelitian berturut-turut yaitupada P3 sebesar 5,14 cm, P0 1,53 cm, P2 1,41cm dan P1 1,26 cm.
Tabel 2. Rata-rata Pertumbuhan Bobot Mutlak,Pertumbuhan Panjang Mutlak, LajuPertumbuhan Spesifik, Pertumbuhan BobotBiomassa, Efisiensi Pakan, Konversi PakanDan Kelulushidupan Ikan Juaro SelamaPenelitian.
Parameter
Satuan
PerlakuanP0 P1 P2 P3
Pertumbuhan bobotmutlak
gram
3,81±0,76a
4,95±1,27a
5,39±2,11a
9,24±0,35b
Pertumbuhanpanjangmutlak
cm1,53±0,76a
1,26±1,27a
1,41±2,11a
5,14±0,35b
LajuPertumbuhanspesifik
%0,75±0,02a
1,08±0,34a
1,07±0,37a
1,76±0,17b
Pertumbuhan bobotbiomassa
gram
34,6±5,69a
37,86±1,33a
43,26±5,60a
62,23±6,93b
EfisiensiPakan
%5,22±0,78a
11,07±2,74a
12,13±5,02a
28,07±5,80b
KonversiPakan
%5,58±1,93
4,83±1,48
4,88±0,48
3,45±1,55
Kelulushidupan
%66,67±11,55
73,33±11,55
80±0,00
86,67±11,55
Keterangan : huruf superscrip yang berbeda padakolom yang sama menunjukkan perbedaan nyata(P<0,05).
Persentase rata - rata laju pertumbuhanspesifik ikan Juaro terbaik terdapat padaperlakuan P3 yaitu 1,76 %, kemudian diikutidengan P1 1,08%, P2 1,07% dan yang terendahterdapat pada perlakuan P0 0,75%. Hal inimenunjukkan pakan ikan komersil yangdiberikan pada penelitian ini sudahdimanfaatkan ikan Juaro dengan cukup baik.Huet (1986) menyatakan bahwa pertumbuhanikan dipengaruhi oleh beberapa faktor, yaitufaktor internal (keturunan, umur dan ketahananterhadap penyakit) dan faktor eksternal (suhu,
perairan, besarnya ruang gerak, kualitas air,jumlah dan mutu makanan).Pertumbuhan bobot biomassa ikan Juaroterbaik terdapat pada perlakuan P3 yaitu sebesar62,63 gram, kemudian diikuti dengan P2 43,26gram, P1 37,86 gram dan yang terendahterdapat pada perlakuan P0 34,6 gram. Hal inimenunjukkan filter dengan substrat zeolitmemberikan pertambahan biomassa terbesar,karena kualitas airnya paling baik padaperlakuan ini. Menurut Putra dan Pamukas(2011) pertumbuhan dipengaruhi oleh kualitasdan kuantitas pakan, umur dan kualitas airpemeliharan. Peningkatan biomassa merupakantingkat pemberian pakan yan diubah menjadibiomassa ikan. Pemanfaatan pakan dapatterindikasi dari biomassa total dan peningkatanjumlah pakan yang diberikan pada ikan yangdipelihara.
Rata-rata efisiensi pakan selama penelitianberkisar antara 5,22 – 28,07%,efisiensi pakan tertinggi dijumpai padaperlakuan P3, yaitu 28,07%, kemudian diikutiperlakuan P2 12,13%, P1 11,07% dan P0 5,22%.Efisiensi pakan pada penelitian ini tergolongmasih rendah, karena ikan masih beradaptasidengan pakan komersil yang diberikan,sehingga pemanfaatan dan kemampuan ikanJuaro dalam mencerna pakan yang diberikanbelum optimal. Menurut NRC (1993) efisiensipakan berhubungan erat dengan kesukaan ikandengan pakan yang diberikan, selain itudipengaruhi oleh kemampuan ikan dalammencerna bahan pakan. Selanjutnya Craig danHelfrich (dalam Ahmadi, Iskandar danKurniawati, 2012) menyatakan bahwa pakandikatakan baik apabila nilai efisiensi pemberianpakannya lebih dari 50% atau bahkanmendekati 100%.
Rata-rata konversi pakan selama penelitianberkisar antara 3,45-5,58%, konversi pakantertinggi dijumpai pada perlakuan P0 yaitusebesar 5,58%, hal ini disebabkan kualitas airpada perlakuan ini tidak sesuai untukpertumbuhan ikan juaro terutama kandunganCO2 bebas dan Amonianya yang tinggi
10
23
22
20
19
18
17
16
15
4
12

Niken Ayu Pamukas dan Mulyadi 191
sehingga menyebabkan ikan stress. Akhirnyapakan yang diberikan tidak bisa dimanfaatkanikan secara maksimal untuk meningkatkanpertumbuhannya. Sedangkan konversi pakanterendah dijumpai pada perlakuan P3 sebesar3,45%, hal ini berarti dibandingkan denganfilter lainnya, filter yang menggunakan zeolitmemberikan lingkungan yang paling kondusifuntuk pertumbuhan ikan Juaro.
Secara keseluruhan konversi pakan padapenelitian ini tergolong masih tinggi, hal inidisebabkan karena ikan masih beradaptasidengan pakan komersil yang diberikan,sehingga pemanfaatan dan kemampuan ikanJuaro dalam mencerna pakan yang diberikanbelum optimal. Konversi pakan yang baikapabila nilainya ≥ 2%, bahkan mendekati 1% .Kelulushidupan tertinggi ikan Juaro dijumpaipada perlakuan P3 yaitu sebesar 86,67%,sedangkan tingkat kelulushidupan terendahterjadi pada perlakuan P0 yaitu 66,67% (Tabel10). Kematian pada ikan Juaro yang terjadiselama penelitian secara keseluruhandiakibatkan ikan stress dan belum bisaberadaptasi pada wadah budidaya dengan baik.Pada perlakuan P0 dan P1 kematian ikan didugadisebabkan karena kualitas air terutamakonsentrasi amonia pada hari ke -60 yangcukup tinggi mencapai 0,45 mg/L dan 0,36mg/L. Menurut Lakshmana (2010) faktor yangmempengaruhi tinggi rendahnya kelangsunganhidup ikan adalah faktor biotik antara lainkompetitor, kepadatan, populasi, umur, dankemampuan organisme beradaptasi terhadaplingkungan.
Uji analisis variansi (ANAVA) P < 0,05menunjukkan filter yang berbeda memberikanpengaruh terhadap pertumbuhan bobot mutlak,pertumbuhan panjang mutlak, lajupertumbuhan spesifik, pertumbuhan bobotbiomassa, efisiensi pakan dan kelulushidupanikan Juaro. Uji lanjut Student-Newman Keulsmenunjukkan perlakuan terbaik dijumpai padaP3. Filter yang berbeda tidak memberikanpengaruh terhadap konversi pakan dankelulushidupan Ikan Juaro.
KESIMPULAN
Berdasarkan hasil penelitian yang telahdilakukan dapat disimpulan bahwa ikan Juaro(Pangasius polyuranodon) sudah dapatdipelihara dalam lingkungan terkontrol, yaitupada akuarium dengan sistem resirkulasi sertapemberian pakan buatan (pellet). Penggunaansistem resirkulasi dengan substrat filter yangberbeda dapat menjaga kualitas air terutamasuhu, pH, Oksigen terlarut, kandungan CO2
bebas, Amonnia (NH3), Nitrit (NO2) dan Nitrat(NO3) tetap sesuai untuk pertumbuhan ikanJuaro.
Perlakuan terbaik dijumpai pada P3
menggunakan sistem resirkulasi dengansubstrat filter batu zeolit, dimana suhu berkisar28,50-29,330C, pH 5,5-6, konsentrasi Oksigenterlarut 3,99-4,59 mg/L, konsentrasi CO2 bebas8,59-9,18 mg/L, konsentrasi NH3 berkisarantara 0,02 - 0,10 mg/L, NO2 0,01-0,08 mg/L,NO3 0,01-0,09 mg/L, rata-rata pertambahanbobot mutlak ikan Juaro 9,24 gram, rata-ratapertambahan panjang mutlak 5,14 cm, lajupertumbuhan spesifik 1,76%, pertumbuhanbobot biomassa 62,23 gram, efisiensi pakan28,07%.
Sistem resirkulasi menggunakan filter yangberbeda tidak memberikan pengaruh terhadapkonversi pakan dan tingkat kelulushidupan ikanJuaro. Namun secara deskriptif hasil terbaikpada penelitian ini yaitu pada perlakuan P3
dengan konversi pakan 3,45% dan rata-ratatingkat kelulushidupan 86,67%.
DAFTAR PUSTAKA
Ahmadi, H., Iskandar dan N. Kurniawati. 2012.Pemberian Probiotik dalam Pakanterhadap Pertumbuhan Lele Sangkuriang(Clarias gariepinus) Pada Pendederan II.Jurnal Perikanan dan Kelautan UNPAD.3,4 (2012) : 99-107
Akbar, H. 2013. Manfaat Daun Ketapang BagiIkan Cupang. http://Aquariumhias.blogspot.com. Diakses tanggal 28 Juni2014.
11

Niken Ayu Pamukas dan Mulyadi 192
Boyd, C.E. 1982. Water Quality in Ponds forAquaculture. Alabama AgriculturalExperiment Station. Auburn University,Alabama. 482 pp.
Daelami, D.A.S. 2001. Agar Ikan Sehat.Penebar Swadaya. Jakarta. 80 hal.
Effendi, H. 2003. Telaah Kualitas Air. Kanisius.Yogyakarta. 273 pp.
Ernawati, Y., E. Prianto dan A. Ma’suf. 2009Biologi Reproduksi Ikan Juaro (Pangasiuspolyuranodon) di Daerah Aliran SungaiMusi, Sumatera Selatan. BerkalaPenelitian Hayati. 15: 45 – 52.
Huet, M. 1986. Text Book of Fish Culture.Breeding and Cultivation of Fish 2nd Ed.Fishing News (books). Oxford. 438 p.
Kasry. A, Sedana, I. P, Feliatra, Syahrul,Nugroho. F. and Sofyan. I. 2002.Pengantar Perikanan dan Ilmu Kelautan.Universitas Riau. Faperika Press.136 pp.
Kottelat, M, A. J. Whitten, S. N. Kartika dan S.Wirjoatmodjo. 1993. Ikan-ikan Air TawarIndonesia Bagian Barat dan Sulawesi.Periplus Edition (HK), Ltd., kerjasamadengan proyek EMBI, Kantor MenteriNegara Kependudukan dan LingkunganHidup Republik Indonesia. Jakarta. 293hal.
Mulyadi, Hasibuan N dan Romiantoyo. 2010.Sistem Resirkulasi Dengan MenggunakanFilter Berbeda Dalam Media PemeliharaanBenih Ikan Mas (Cyprinus carpio L).Laporan Penelitian. 48 halaman.
Murtiati dan Sri, E. A. 1999. PengaruhBerbagai Kadar Zeolit dalam Filter SistemResirkulasi Terhadap Pertumbuhan danKelangsungan Hidup Ikan Lele Dumbo(Clarias gariepinus, Buechell).Institutional Repository. UniversitasDiponegoro.
NRC. 1993. Nutrition and Requirement ofWarmwater Fishes. National Academic ofScience. Washington, D. C. 248 p.
Peraturan Pemerintah Republik IndonesiaNomor 82. 2001. Tentang PengelolaanKualitas Air dan PengendalianPencemaran Air.
Putra dan N. A. Pamukas. 2011. Pemeliharaanikan selais (Ompok sp) dengan resirkulasi,sistem aquaponik. Jurnal Perikanan danKelautan 16,1 (2011) : 125-131
Pusat Dokumentasi dan Informasi Ilmiah LIPI.1999. Arang Aktif dari Tempurung Kelapa.Pusat Dokumentasi dan Informasi Ilmiah-Lembaga Ilmu Pengetahuan Indonesia(tidak diterbitkan).
Ramadhan, P. P. 2008. Studi KebiasaanMakanan Ikan Juaro (Pangasiuspolyuranodon) di Perairan Sungai Musi,Sumatera Selatan. Repository IPB.
Saanin, H. 1984. Taksonomi dan KunciIdentifikasi Ikan. Bina Cipta, Jakarta. 131hal.
Siikavuopio S.I and Saether BS. 2006. Effectsof chronic nitrite exposure on growth injuvenile Atlantic cod Gadus morhua.Aquaculture 255 : 351–356
Sudradjat, E. S. B. 1991. Aktivasi ArangTempurung Kelapa dengan MenggunakanSeng Klorida. Jurnal Teknologi Indonesia.14 (1) : 14-15.
Surya Mina. 2014. Air Terlalu Dingin, IkanJadi Malas Makan. www.bibitikan.net.Diakses tanggal 21 September 2014.
Syafriadiman. N. A. Pamukas.. S. Hasibuan..2005. Dasar-dasar Kualitas Air. MinaMandiri Press. Pekanbaru. 131 pp.
13

ISSN 978-602-14989-0-3 BioETI
Respon tanaman Bayur (Pterospermum javanicum Jungh.)terhadap inokulan Fungi Mikoriza Arbuskula (FMA) pada lahanbekas Tambang Semen PadangNURUL ALIFAH, ZOZY ANELOI NOLI DAN SUWIRMEN
Labor Riset Fisiologi Tumbuhan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRACTThe research about responses of Plants Pterospermum javanicum (Jungh.) inoculated with Arbuscular Mycorrhiza Fungi(AMF) on mined land soil Padang Cement has been done on August to November 2013 in the net house and Laboratoryof Plant Physiology, Departement of Biology, Faculty of Mathematics and Natural Science, Andalas University. As thetreatment this research used P. javanicum were planted on topsoil, land mines soil without inoculant of mycorrhiza andland mines with inoculant of mycorrhiza. The results showed indicated that giving of inoculant of mycorrhiza affected thepercentage degree of infection up to 76.66% with very high criteria and increased in leaf but not increased in plant height,the number of stem diameter and total dry weight during 12 weeks of observation.
Key words: Arbuscular Mycorhizza Fungi (AMF), cement mined, Pterospermum javanicum
Pendahuluan
Industri semen merupakan salah satu sektoryang berperan penting bagi perekonomianIndonesia (Asmarahman, 2008). Kegiatandalam proses pertambangan sangatmempengaruhi kondisi lahan dan menyebabkankerusakan yang signifikan terutama kerusakanpada lapisan tanah atas (top soil) yang memilikibanyak unsur hara (Margarettha, 2007).
Lahan yang mengalami degradasi karenaaktivitas pertambangan bahan baku semenmenimbulkan banyak kendala pada upayarevegetasi lahan karena tingkat kesuburan lahanyang rendah, lahan berupa hamparan tanahkapur (CaO), silica (SiO2), aluminium oksida(Al2O3), pasir besi (Fe2O3), gips dan tanah liatsehingga tumbuhan sulit untuk tumbuh di lahanbekas tambang semen tersebut dan hanya akanmenjadi lahan tidur yang tidak termanfaatkan(Asmarahman, 2008).
Untuk membantu pertumbuhan danmeningkatkan daya hidup tanaman pada lahanbekas tambang semen diperlukan tekniksilvikultur yang tepat, pemilihan jenis tanamanyang cocok, input energi yang tinggi sepertisaturasi fosfat, pemupukan yang lengkap danmanajemen bahan organik. Namun teknik-
teknik tersebut memerlukan biaya yang tinggi(Asmarahman, 2008). Alternatif yang dapatdilakukan yaitu berupa pendekatan secarabioteknologi dengan memanfaatkan potensimikroorganisme yang mampu memperbaikistruktur tanah dan berperan dalam siklus harasalah satunya yaitu Fungi Mikoriza Arbuskula(FMA). Fungi Mikoriza Arbuskula (FMA)merupakan srtruktur yang terbentuk karenaasosiasi berupa simbiosis mutualisme antarafungi tanah dengan akar tanaman. Dalamsimbiosis ini tanaman inang memperoleh haradengan bantuan fungi sedangkan fungimendapat hasil fotosintat dari inang (Brundrett,et al., 1996 ).
Penggunaan FMA menjadi strategi yangmenguntungkan karena penyebarannya lebihluas dan mempunyai kemampuan berasosiasiyang tinggi, yaitu mencapai 90% jenis tanamanbersasosiasi dengan mikoriza (Cruz et al.,2000). FMA mampu menarik perhatian parapeneliti lingkungan dan biologis karenacendawan ini dapat membantu tanaman dalammeningkatkan kualitas dan produktivitastanaman terutama bagi tanaman yang di tanampada lahan-lahan marginal yang kurang suburatau areal bekas tambang (Delvian, 2006).

Nurul Alifah, Zozy Aneloi Noli dan Suwirmen 194
Pada umumnya tanaman yang digunakan untukupaya revegetasi adalah tanaman jenis pohondan tanaman jenis fast growing spesies (FGS)(Ardanari, 2011). Dari beberapa penelitiandiketahui beberapa tanaman yang dapatdigunakan untuk upaya revegetasi diantaranyaadalah tanaman Pterospermum javanicum. P.
javanicum merupakan tanaman jenis pohonyang mampu berasosiasi dengan mikoriza danmampu beradaptasi pada lahan kritis (Contesa,2012; Herdina, 2013). Penelitian ini dilakukanuntuk mengetahui respon tanaman bayur(Pterospermum javanicum Jungh.) yangdiinokulasi Fungi Mikoriza Arbuskula (FMA)Pada Lahan Bekas Tambang Semen Padang.
BAHAN DAN METODE
Penelitian ini dilaksanakan dari bulan Agustussampai November 2013 di rumah kawat dandilanjutkan di Labor Fisiologi Tumbuhan,Jurusan Biologi, Fakultas Matematika dan IlmuPengetahuan Alam Universitas Andalas Padang,Sumatera Barat.
Penelitian ini menggunakan metodaeksperimen sebagai media tanam perlakuanyaitu topsoil (kontrol), tanah tambang tanpainokulasi mikoriza dan tanah tambang diberiinokulasi mikoriza dengan 3 kali ulangan.Analisa data disajikan secara deskriptif.
Parameter yang diamati yaitu Pertambahantinggi tanaman, Pertambahan jumlah daun,Pertambahan diameter Batang, Bobot keringtanaman, dan Derajat infeksi FMA.
HASIL DAN PEMBAHASAN
Derajat Infeksi Fungi Mikoriza Arbuskula(FMA)Dari pengamatan yang dilakukan didapatkanrata-rata derajat infeksi mikoriza pada akartanaman P. javanicum seperti pada Tabel 1.Berdasarkan Tabel 1, tanaman P. javanicummemiliki kriteria derajat infeksi yang sangattinggi pada media tambang + inokulasi, hal inimengindikasikan bahwa tanaman P. javanicum
memiliki kemampuan berasosiasi dengan FMA.
Tingginya derajat infeksi pada perakarantanaman menunjukkan adanya kecocokanantara FMA dengan tanaman inangnya.Kecocokan FMA dengan tanaman inangnyajuga dipengaruhi oleh jenis FMA yangdigunakan (Delvian, 2006).
Tabel 1. Persentase derajat infeksi FMA padaakar tanaman P. javanicum.
Perlakuan Persentase Derajat Infeksi(%)
Topsoil 53,33 (Tinggi)Tanpa Mikoriza 46,66 (Sedang)Mikoriza 76,66 (Sangat Tinggi)
Pada perlakuan tanpa inokulasi, infeksiyang terjadi pada akar di sebabkan oleh adanyaFMA indigenous dari media tanam karena padapenelitian ini media tanam yang di gunakantidak di sterilisasi terlebih dahulu.Kemungkinan lain yang terjadi adalah akartanaman telah terinfeksi FMA ketika bibitmasih dalam media pembibitan karenapemindahan tanaman ke media perlakuanmembawa sedikit tanah dari media pembibitanuntuk menghindarkan tanaman dari stres ataumati.
Infeksi FMA terjadi karena adanyasimbiosis dan interaksi antara FMA denganakar tanaman. Perakaran tanaman yangterinfeksi FMA dicirikan dengan adanyabeberapa organ khas dari FMA yang terdapatpada jaringan akar tanaman akan terlihat sepertipada Gambar 1.
Gambar 1. Kolonisasi Fungi Mikoriza Arbuskula(FMA) pada akar tanaman P. javanicum.Keterangan : A (Vesikula),
B (Hifa internal)
A
BB

Nurul Alifah, Zozy Aneloi Noli dan Suwirmen 195
Pada penelitian ini tidak ditemukan adanyaarbuskula. Hal ini di duga karena siklus hiduparbuskula yang sangat singkat yaitu antara 1-3minggu. Pada umumnya arbuskula terbentuksebelum vesikula, namun adapula vesikulayang dibentuk tanpa pembentukan arbuskulaterlebih dahulu (Pattimahu, 2004). Hal yangsama di peroleh pada hasil penelitian Contesa(2010), dimana pada akar bibit tanaman pisangFHIA-25 yang diinokulasi multispora (Glomussp. + Acaulospora sp.) tidak ditemukan adanyaarbuskula.Pertumbuhan TanamanDari Tabel 2 dapat dilihat bahwa padaparameter pertambahan tinggi tanaman (TT)kombinasi antara tanaman P. Javanicumdengan topsoil menunjukkan interaksi yangpaling baik jika di bandingkan dengan mediayang lain. Hal ini diduga karena tanaman P.Javanicum merupakan salah satu jenis tanamanyang mampu tumbuh dengan sangat baik padatanah yang memiliki kemampuan mengikat airyang tinggi. Seperti pada habitat aslinya,tanaman bayur biasanya ditemukan tumbuhsecara lokal pada hutan primer atau sekunderdengan tempat tumbuh selalu berdekatandengan sumber air seperti sungai. Yamada,Ngakan dan Suzuki (2007) mengemukakanbahwa tanaman P. javanicum dapat di jumpaipada tanah datar hingga lereng hutan yangberdekatan dengan sumber air dan sedikitternaung.
Pada pertambahan jumlah daun (JD),tanaman P. javanicum yang diinokulasi FMAmenunjukkan hasil tertinggi jika dibandingkandengan kedua media yang lain, hal inidikarenakan mikoriza yang diinokulasikanberpotensi mengoptimalkan suplai hara bagitanaman sehingga mampu meningkatkanpertumbuhan vegetatif tanaman. Mikorizadapat membantu tanaman menyerap unsur Ndan P yang sangat berperan dalampembentukan organ daun bagi tanaman(Tuheteru dan Husna, 2011).
Pada pertambahan diameter batang (DB)dan bobot kering tanaman (BK), tanaman P.javanicum menunjukkan respon tertbaik padamedia tanpa inokulan FMA, hal ini dikarenakanadanya kecocokan antara mikoriza indigenouspada media yang digunakan dengan tanaman.Delvian (2006) mengemukakan bahwamikoriza indigenous lebih berpotensi dalammembentuk asosiasi karena telah mengenalitanaman inangnya. Sedangkan bobot keringtanaman (BK) erat kaitannya dengan nutrisiyang diserap tanaman, laju fotosintesis danrespirasi serta akumulasi senyawa organikterutama air, dan CO2 (Lakitan, 1995). Didugakarena mikoriza indigenous memiliki potensilebih tinggi dalam membentuk asosiasisehingga suplai hara bagi tanaman pun menjadilebih optimal.
Tabel 2. Rata-rata pertambahan tinggi tanaman,Pertambahan jumlah daun,Pertambahan diameter Batang, Bobotkering tanaman setelah 12 minggupengamatan.
Perlakuan TT(cm)
JD(helai)
DB(mm)
BK(gr)
Top Soil 5,03 3,28 1,19 2.88TanpaMikoriza
4,61 4,11 1.39 3,08
+Mikoriza 4,22 4,36 1,29 2,84
Keterangan : TT : Tinggi TanamanJD : Jumlah DaunDB : Diameter BatangBK : Berat Kering
KESIMPULAN
Berdasarkan hasil penelitian yang dilakukantentang respon tanaman bayur (Pterospermum
javanicum Jungh.) yang diinokulasi FungiMikoriza Arbuskula (FMA) Pada Lahan BekasTambang Semen Padang, maka diperolehkesimpulan sebagai berikut : tanaman P.
javanicum memberikan respon yang baikterhadap inokulasi Fungi Mikoriza Arbuskula(FMA), ditunjukkan dengan persentase derajatinfeksi yang sangat tinggi namun hanyaberpengaruh pada pertambahan jumlah daun.

Nurul Alifah, Zozy Aneloi Noli dan Suwirmen 196
DAFTAR PUSTAKA
Ardanari, C. Y. 2011. Status Penggunaan FMAPada Tanaman Fast Growing SpeciesDalam Pembangunan Hutan TanamanIndustri dan Rehabilitasi Lahan Kritis.Skripsi Sarjana Kehutanan InstitutPertanian Bogor. Bogor.
Asmarahman, C. 2008. Pemanfaatan Mikorizadan Rhizobium Untuk MeningkatkanPertumbuhan Semai Kayu Energi PadaMedia Tanah Bekas TambangSemen.[Tesis]. Bogor. Institut PertanianBogor.
Brundrett, N., B. Bougher, T. Dell, Grove andN. Malajazuk. 1996. Working WithMycorrhizas In Forestry And Agriculture.Australian Centre for InternationalAgriculture Research (ACIAR). Canberra.Pp. 162-171.
Contesa, E. 2010. Pertumbuhan Bibit TanamanPisang (Musa paradisiaca L.) FHIA-25.yang Diinokulasi dengan beberapa DosisFMA Glomus sp. + Acaulospora sp.Skripsi Sarjana Biologi FMIPA.Universitas Andalas. Padang.
Contesa, E. 2012. Isolasi Dan Potensi FungiMikoriza Arbuskula (Fma) IndigenousDari Tanaman Pionir Di HutanPendidikan Dan Penelitian Biologi(Hppb). Tesis Pascasarjana UniversitasAndalas. Padang.
Cruz, A. F., T. Ishii and Kadoya. 2000. Effectof rbuskular mycorrhizal fungi on treegrowth, leaf water potensial, and leaf of 1-Aminochyclopropane-1-carboxylic acidand Ethylene in the roots of papaya under
water stress condition. Mycorrhiza. 20:121-205.
Delvian. 2006. Peranan ekologi dan agronomicendawan mikoriza arbuskula.Departemen kehutanan. Fakultaspertanian. Universitas Sumatera Utara.Medan.
Herdina, J. 2013. Pertumbuhan BeberapaTanaman Untuk Revegetasi yangdiinokulasi Ektomikoriza Pada LahanBekas Tambang Batu Bara Ombilin. TesisPascasarjana Universiatas Andalas.Padang.
Lakitan, B. 1995. Fisiologi Tumbuhan danPerkembangan Tanaman. RajagrafindoPersada. Jakarta.
Margarettha. 2007. Pemanfaatan Tanah BekasTambang Batubara Dengan Pupuk HayatiMikoriza Sebagai Media Tanam JagungManis. Jurnal Hidrolitan 1(3):1-10.
Pattimahu, D. V. 2004. Restorasi Lahan KritisPasca Tambang Sesuai Kaidah Ekologi.Makalah Mata Kuliah Falsafah Sains.Pasca sarjana IPB. Bogor.
Tuheteru, F.D dan Husna. 2011. Pertumbuhandan Biomassa Albizia saponaria yangDiinokulasi Fungi Arbuskula MikorizaLokal Sulawesi Tenggara. JurnalSilvikultur Tropika 2(03):143-148.
Yamada, T., O. P. Ngakan., E. Suzuki. 2007.Habitat Differences Between TwoCongeneric Canopy Trees, Pterospermumjavanicum and P. diversifolium(Sterculiaceae) In an IndonesianFloodplain Forest. Tropics 16 (2) 165-169.

ISSN 978-602-14989-0-3 BioETI
Potensi tanaman Digitaria ciliaris dalam meremediasi tanahtercemar Merkuri (Hg) pada lahan bekas Tambang Emasdi Sijunjung, Sumatera Barat
PUTRI KUMALASARI, ZOZY ANELOI NOLI DAN FUJI ASTUTI FEBRIA
Labor Riset Fisiologi Tumbuhan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian tentang potensi tanaman Digitaria ciliaris dalam meremediasi tanah tercemar merkuri (Hg) lahan bekastambang emas telah dilakukan dari bulan November 2013 sampai Februari 2014 di Rumah Kawat dan LaboratoriumFisiologi Tumbuhan, Jurusan Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Andalas Padang.Penelitian ini menggunakan metode eksperimen, sebagai perlakuan adalah media tanam meliputi A: tanah subur (kontrol),B: tanah tercemar 100 %, C: tanah tercemar merkuri 90 % + kompos 10 %. Hasil penelitian ini menunjukkan bahwaperlakuan media tanam berpengaruh nyata terhadap pertumbuhan tinggi tanaman, bobot basah dan bobot kering (tajukdan akar). Kombinasi tanah tercemar 90 % + kompos 10 % merupakan media terbaik dalam meningkatkan bobot basahakar 0,185 g, bobot kering tajuk 0,190 g dan bobot kering akar 0,121 g. Penyerapan merkuri (Hg) pada tanaman D.ciliaris paling banyak diakumulasi di bagian akar tanaman yaitu 0,132 ppm pada perlakuan media tanah tercemar +kompos 10 %.
Key words: fitoremediasi, Digitaria ciliaris, merkuri (Hg), kompos
Pendahuluan
Aktivitas pertambangan sering dianggapmemiliki dua sisi yang saling berlawanan,disatu sisi berpotensi meningkatkan pemasukandaerah namun di sisi lain memberikan dampaknegatif bagi lingkungan. Penggunaan Hg padaproses amalgamasi sebagai media untukmengikat emas pada lahan tambang emassecara terus menerus akan menyebabkankerusakan dan pencemaran terhadaplingkungan (Setiabudi, 2005). Hasil penelitianpendahuluan terhadap sampel tanah bekastambang emas di daerah Tanjung AmpaluSijunjung, Sumatera Barat didapatkankandungan logam berat merkuri (Hg) yangcukup tinggi yaitu sebesar 1,1 mg/l dan kadarhara N, P dan K masing- masingnya adalah0,22 %, 313,43 ppm dan 0,534 ml/100 g(Kumalasari, Noli, Febria, 2013, (unpublished).Sedangkan ambang batas konsentrasi merkuriyang masih bisa ditoleransi untuk media tanahyaitu < 1 mg/l (Departemen Pertanian, 2005).
Upaya pemulihan (remediasi) perludilakukan agar lahan bekas dapat digunakanuntuk berbagai kegiatan secara aman. Salahsatu metode untuk memulihkan lahan tersebutadalah metode fitoremediasi, yaitu penggunaantumbuhan untuk menghilangkan polutan daritanah atau perairan yang terkontaminasi(Priyanto dan Prayitno, 2005). Menurut Goshand Sing (2005) fitoremediasi adalah teknologipembersihan, penghilangan atau penguranganpolutan berbahaya, seperti logam berat,pestisida, dan senyawa organik beracun dalamtanah atau air dengan menggunakan bantuantanaman (hiperakumulator plant).
Mekanisme penyerapan dan akumulasilogam berat oleh tanaman dibagi menjadi tigaproses yang bersinambungan, ialah: 1.Penyerapan oleh akar (Interaksi rizosferik) 2.Translokasi logam dari akar ke bagian asinglain mengikuti aliran transpirasi ke bagian atastanaman melalui jaringan pengangkut (xylemdan floem). 3. Lokalisasi logam pada sel danjaringan. Sebagai upaya untuk mencegahperacunan logam terhadap sel, tanaman

Putri Kumalasari, Zozy Aneloi Noli dan Fuji Astuti Febria 198
mempunyai mekanisme detoksifikasi, denganmenimbun logam di dalam organ tertentuseperti akar, batang dan daun (Priyanto danPrayitno, 2005).
Menurut Hidayati (2004), beberapa jenistumbuhan terbukti mampu beradaptasi padalingkungan pembuangan limbah penambanganemas rakyat yang terkontaminasi merkuri (Hg)hingga 21,66 ppm, di antaranya Linderniacrustacea (family Poaceae) yang mampumenyerap Hg hingga 89,13 mg per kg beratkeringnya. Penelitian Syarif (2009), tanamanLeersia (family Poaceae) mampumengakumulasi merkuri di jaringan tanamanyang lebih tinggi dibandingkan ketiga tanamanlainnya (Mikania dan Centrocema).
Keberhasilan dalam meremediasi lahanbekas juga ditentukan oleh penambahan bahanorganik dalam tanah. Bahan organik seringkalidijadikan sebagai indikator umum kesuburantanah. Kombinasi penggunaan kompos dalamrevegetasi tailing sangat efektif dalammenentukan dan menjaga daya penutupanvegetasi, meningkatkan produksi biomasa sertamembantu pertumbuhan tanaman (Kelly et al.,2003). Menurut Triastuti (2012) diperolehbahwa komposisi media tanam 90 % tanahtercemar + 10 % kompos lebih efisien dalammembantu tanaman akar wangi (Vetiveriazizanioides) memulihkan tanah yang tercemarmerkuri yaitu sebesar 65,252%.
Penelitian ini dilakukan bertujuan untukmengetahui pengaruh tanaman D. ciliaris
(family Poaceae) sebagai akumulator merkuri(Hg) pada lahan bekas tambang emas di daerahSijunjung dalam meremediasi tanah tercemarmerkuri (Hg) dan untuk mengetahui pengaruhkompos terhadap pertumbuhan tanamanakumulator. Digunakannya D. ciliaris sebagaitanaman akumulator karena dari surveypendahuluan tanaman ini dominan ditemukandi lokasi pengambilan sampel.
BAHAN DAN METODE
Penelitian ini dilakukan pada bulan November2013 sampai Februari 2014 di Rumah Kawat
dan Laboratorium Fisiologi Tumbuhan, JurusanBiologi Fakultas Matematika dan IlmuPengetahuan Alam, Universitas AndalasPadang. Penelitian ini menggunakan metodeeksperimen, sebagai perlakuan adalah mediatanam meliputi A : Tanah subur (kontrol), B :Tanah tercemar merkuri 100 %, C : Tanahtercemar merkuri 90 % + kompos 10 %.
Bahan yang digunakan dalam penelitian iniialah bibit tanaman Digitaria ciliaris, tanahbekas tambang emas, kompos, tanah subur danaquadest. Alat yang digunakan ialah timbangananalitik, pH meter, GPS (Global PositioningSystem), ICPE (Inductively Coupled PlasmaEmission) Spectrometer 9000 Shimadzu, oven,polibag, plastik sampel, kertas label, koran,ember, parang, gunting, kamera dan alat tulis.
Cara kerja meliputi : 1) Pengambilansampel tanah bekas tambang emas, yangdiambil di sekitar lokasi bekas penambanganemas rakyat di nagari Tanjung Ampalu,Sijunjung Sumatera Barat pada koordinat S :00038’05.6 dan E : 100050’58.0 dengan elevasi181 mdpl. Tanah dibersihkan dari serasah, dikeringanginkan dan di ukur pH tanah. Tahapanpengukuran pH tanah menurut Van Reeuwijk(1993). 2) Tahap persiapan meliputi penyiapanbibit tanaman dan persiapan media tanam 3)Pemberian kompos, dilakukan sebelumpenanaman sesuai perlakuan yaitu tanpakompos (kontrol) dan pemberian kompossebesar 10 %. Kompos dicampur meratadengan tanah dan di keringanginkan. 4)Penanaman tanaman, bibit D. ciliaris yangsudah siap tanam dipindahkan ke dalampolybag yang telah berisi media tanam. Satupolybag masing-masing hanya diisi oleh satubibit D. ciliaris. 5) Pemeliharaan tanaman,meliputi penyiraman dan pengendalian gulma.6) Panen dilakukan dengan interval setiapumur 2 minggu (1, 2, 4, 6 dan 8 minggu setelahtanam).
Parameter yang diamati meliputi tinggitanaman, bobot basah tajuk, bobot basah akar,bobot kering tajuk, bobot kering akar dankandungan logam merkuri (Hg) pada tanah dan

Putri Kumalasari, Zozy Aneloi Noli dan Fuji Astuti Febria 199
jaringan tanaman (tajuk dan akar). KandunganHg pada tanah dan jaringan tanaman dianalisisdengan menggunakan metode ICPE 9000Shimadzu.
Data yang diperoleh dari hasil penelitiandisajikan secara deskriptif. Kandungan merkuripada tanah dan jaringan tanaman diperolehberdasarkan hasil analisis Laboratorium Tanah,Teknik Lingkungan dengan menggunakanICPE.
HASIL DAN PEMBAHASAN
Pertumbuhan tanaman D. ciliarisDari hasil pemberian perlakuan media tanamterhadap pertambahan tinggi, bobot basah danbobot kering (tajuk dan akar) pada tanaman D.ciliaris didapatkan hasil seperti pada Tabel 1.Tabel 1 menunjukkan bahwa rata - ratapertambahan tinggi tanaman (TT) dan bobotbasah tajuk (BBT) tanaman D. ciliaris padamedia tanah subur (A) sebagai kontrol lebihtinggi dibandingkan media tanah lainnya.Sedangkan untuk pertambahan BBA, BKT danBKA media tanam dengan kombinasi tanahtercemar + kompos 10 % memiliki nilai yanglebih tinggi dibandingkan perlakuan mediatanah subur dan tanah tercemar 100 %.
Tabel 1. Rata- rata pertambahan tinggi, Bobot basahdan Bobot kering (tajuk dan akar) tanamanD. ciliaris dengan perlakuan media tanam
Media tanamRata-rata di akhir pengamatan
TT(cm)
BBT (g) BBA(g)
BKT(g)
BKA(g)
T. subur (b1) 34,1 1,271 0,169 0,183 0,099
T. tercemar 100 %(b2)
21,3 0,983 0,122 0,155 0,042
T. tercemar 90 %+ kompos 10 %
(b3)
26,9 1,129 0,185 0,190 0,121
Keterangan : A: t. subur (kontrol), B: t. tercemar100 %, C: t. tercemar 90 % + kompos 10 %. TT:Tinggi Tanaman, BBT: Bobot Basah Tajuk, BBA:Bobot Basah Akar, BKT: Bobot Kering Tajuk, BBA:Bobot Kering Akar
Perlakuan media tanam dengan kombinasitanah tercemar + kompos 10 % (C)menunjukkan pertumbuhan yang lebih baikjika dibandingkan dengan perlakuan media
tanah tercemar (B) yang memiliki daya dukungtanah yang rendah dan miskin akan hara. Mediatanah tercemar (B) memiliki struktur tanahyang liat, sehingga tanaman mengalamikesulitan dalam penyerapan mineral dan harayang terdapat di dalam tanah yang dibutuhkanuntuk pertumbuhan dan perkembanganhidupnya. Hal ini ditunjukkan denganperlakuan B memiliki nilai rata-ratapertumbuhan terendah untuk semua parameterpertumbuhan.
Menurut Fauziah (2009) dari hasil analisakarakteristik hara tailing tambang emas tampakbahwa tekstur tanah bekas tambang emasdidominasi oleh fraksi liat berpasir, hal inidapat mengakibatkan tanaman sulit untukmenyerap (menahan) air dan unsur hara, tetapidengan pemberian pupuk kompos terlihatbahwa komposisi fraksi liat dapat meningkat,sehingga dapat meningkatkan kemampuantanah dalam menahan air. Peranan bahanorganik (kompos) dalam pertumbuhan tanaman,umumnya secara langsung atau sebagian besarmempengaruhi tanaman melalui perubahansifat dan ciri tanah.
Menurut Dharmawan (2003) komposmerupakan bahan organik yang telahmengalami dekomposisi oleh mikroorganismepengurai sehingga dapat dimanfaatkan untukmemperbaiki sifat- sifat tanah, disamping itu didalam kompos terkandung hara-hara mineralyang berfungsi untuk membantu pertumbuhantanaman. Menurut Hanafiah (2005) bahanorganik merupakan perekat butiran lepas ataubahan pemantap agregat, sebagai sumber haratanaman dan sumber energi dari sebagian besarorganisme tanah. Bahan organik jugamenjadikan fluktuasi suhu tanah lebih kecil dandapat membantu akar tanaman menembustanah lebih dalam dan luas sehingga tanamanlebih kokoh dan lebih mampu menyerap unsurhara dan air dalam jumlah banyak.
Penelitian Triastuti (2012) pemberiankompos yang dikombinasikan dengan tanahtailing dapat meningkatkan pertumbuhantanaman akar wangi (Vetiveria zizanioides)

Putri Kumalasari, Zozy Aneloi Noli dan Fuji Astuti Febria 200
sebesar 65,3 %. Kompos bersifat hidrofiliksehingga dapat meningkatkan kemampuantanah dalam memegang air dan mengandungunsur C menjadi sumber energi mikroba.Pengamatan pertambahan rata- rata tinggitanaman D. ciliaris selama 8 minggupengamatan, dapat dilihat pada Gambar 1.
Gambar 1. Grafik pertambahan rata-rata tinggi tanamanD. ciliaris dengan perlakuan media tanamselama 8 minggu pengamatan.Ket: A : t. subur (kontrol), B : t. tercemar100 %, C : t. tercemar + kompos 10 %
Pada Gambar 1, rata-rata pertambahan tinggitanaman D. ciliaris terus mengalamipeningkatan dari pengamatan minggu pertamahingga minggu ke- 8. Pertambahan tinggitanaman D. ciliaris terbaik terdapat pada mediatanah subur (A) yaitu 34,1 cm di akhirpengamatan. Sedangkan perlakuan kombinasitanah tercemar + kompos 10 % (C) yaitu 26,9cm dan tanah tercemar (B) yaitu 21,3 cm diakhir pengamatan. Struktur tanah yang baik danketersediaan hara di dalam tanah sangatmempengaruhi pertumbuhan tanaman, jikaterjadi defisiensi unsur hara maka dapatmenyebabkan pertumbuhan tanaman terganggu,seperti halnya pada tanah bekas tambang emas.Tanah bekas tambang memiliki kandungan harayang rendah, untuk itu diperlukan pemupukan.Ladranada (2009) menjelaskan untukmemperbaiki kesuburan tanah perlunyapemberian bahan organik hingga batas yangoptimal untuk tanaman tertentu, sehingga dapatmembantu dan merangsang prosespertumbuhan vegetatif tanaman. Yunus (2004)menambahkan, struktur tanah menentukan sifataerasi, permeabilitas, kapasitas menahan airdan kerapatan isi tanah. Struktur yang
beragregat baik dapat menciptakan ruang poritanah lebih besar, sehingga air dan minerallainnya dapat dengan mudah masuk ke dalamtanah.
Sebagai media tumbuh bahan tailingmempunyai banyak kendala fisik maupunkimia. Secara fisik bahan tailing relatifbertekstur kasar, berbutir tunggal tidakmembentuk agregat seperti tanah, akibatnyadaya menahan hara rendah. Secara kimia,bahan tailing tidak mengandung koloid samasekali, akibatnya kapasitas tukar kation (KTK)sangat rendah, kandungan unsur hara rendah,kemampuan menahan air juga rendah karenamerupakan bahan sisa tambang.
Kadar merkuri (Hg) pada tanah danjaringan tanamanHasil analisis terhadap kadar logam beratmerkuri (Hg) dalam tanah dan akumulasi padajaringan tanaman D. ciliaris (tajuk dan akar)selama 8 minggu pengamatan ditunjukkan padaGambar 2. Dari Gambar 2 menunjukkan kadarmerkuri (Hg2+) tanah mengalami (Hg2+)penurunan hingga minggu pengamatan ke- 8untuk masing-masing perlakuan pada perlakuanC (tanaman D. ciliaris pada media tanamtanah tercemar + 10 % kompos). Sedangkanperlakuan B yaitu pada media tanah tercemar100 % hanya mengalami sedikit penurunankonsentrasi merkuri (Hg2+) untuk setiapminggunya.
Gambar 2. Kadar merkuri (Hg) pada tanah dan jaringantanaman D. ciliaris (tajuk dan akar) selama8 minggu pengamatan. Ket: B : t. tercemar100 %, C : t. tercemar + kompos 10 %
0.0
10.0
20.0
30.0
40.0
1 2 4 6 8
00.10.20.30.40.5
tanah
tajuk
akar
tanah
tajuk
akar
tanah
tajuk
akar
tanah
tajuk
akar
tanah
tajuk
akar
1 2 4 6 8
Kad
ar H
g (p
pm)
Pengamatan ke
BC

Putri Kumalasari, Zozy Aneloi Noli dan Fuji Astuti Febria 201
Gambar 2 juga menunjukkan terjadinyapenurunan kadar merkuri tanah seiring denganpeningkatan serapan merkuri pada jaringantanaman, baik itu di bagian tajuk maupun akartanaman.
Kadar merkuri pada jaringan tanaman (tajukdan akar) mengalami peningkatan untuk setiapminggu selama 8 minggu pengamatan.Penyerapan kadar merkuri oleh jaringan hanyamengalami sedikit penurunan konsentrasimerkuri (Hg2+) untuk setiap tanaman tajuk danakar terbaik terdapat pada media tanahtercemar + kompos 10 % (C). Hal ini didugadisebabkan kombinasi tanah tercemar + tanahkompos 10 % memberikan pengaruh terhadapperbaikan struktur tanah yang baik sehinggatanaman dengan leluasa dapat menyerap harayang terkandung dalam tanah. Sedangkanuntuk akumulasi merkuri (Hg) pada D. ciliarislebih banyak pada bagian akar dibandingkanbagian tajuk. Tanaman D. ciliaris didugabersifat toleran terhadap media tanah tercemarmerkuri (Hg), sehingga tanaman ini tetapmampu tumbuh dan mampu meyerap merkuri,yang diperlihatkan dengan terjadinyapenurunan kadar merkuri tanah danpeningkatan penyerapan merkuri pada jaringantanaman (akar dan tajuk) untuk setiapminggunya walaupun pada tanah yang tercemarmerkuri.
Tanaman D. ciliaris merupakan salah satujenis tanaman yang relatif mudah tumbuh dantoleransinya tinggi terhadap berbagai jenistanah dan iklim. Menurut Sagita (2002)tanaman D. ciliaris yang berasal dari familyPoaceae merupakan spesies ruderal (spesiesyang mampu berkembang dalam lingkungantercemar serta mempunyai siklus hidup yangrelatif cepat), dapat mengakumulasi pencemardalam jumlah yang besar tanpa menampakkangejala kerusakan eksternal. Famili Poaceaememiliki kemampuan mengkhelat logam(logam diikat oleh molekul khelat) danmembawanya kedalam sel akar melalui prosestransport aktif. Menurut Priyanto dan Prayitno(2005) untuk mencegah peracunan logam
terhadap sel, tumbuhan mempunyai mekanismedetoksifikasi, dengan menimbun logam didalam organ tertentu seperti akar, batang dandaun.
Beberapa spesies rumput-rumputan(Poaceae) telah diuji pula kemampuannyadalam mengakumulasi logam. Sesuai denganpenelitian Syarif (2009), tanaman Leersia(Poaceae) memiliki akumulasi merkuri di akaryang lebih tinggi dibandingkan ketiga tanamanlainnya (Mikania dan Centrocema). Akumulasiterjadi lebih tinggi pada akar dibandingkantajuk sekitar (3.63 – 9.91 kali). Kandungan CNdi akar tertinggi pada Leersia hexandra 0.144mg pada konsentrasi 2.5 ppm CN.
Ketersediaan unsur logam dan penyerapanoleh tanaman ditentukan oleh konsentrasi totaldan bentuk logam di dalam tanah selain faktorgeokimia pada zona perakaran. Faktor genetikdan jenis tumbuhan menentukan penyerapanlogam pada zona perakaran dan akar/ tajukpada tingkat yang bervariasi. Penyerapan jugaditentukan oleh tipe jaringan tanaman danperlakuan yang diberikan pada tanah (Knox etal., 2000;Vangronsveld et al,. 2000).
KESIMPULAN
Berdasarkan penelitian yang telah dilakukantentang potensi tanaman Digitaria ciliarisdalam meremediasi tanah tercemar merkuri(Hg) pada lahan bekas tambang emas di daerahSijunjung, dapat disimpulkan sebagai berikut :1. Tanaman D. ciliaris memiliki kemampuan
sebagai fitoremediator tanah tercemarmerkuri (Hg) dengan akumulasi logammerkuri (Hg) paling tinggi di bagian akarpada perlakuan kombinasi media tanahtercemar 90 % + kompos 10 % yaitu 0,132ppm.
2. Pemberian kompos 10 % berpengaruh nyatadalam meningkatkan pertumbuhan tanamanD. ciliaris, meliputi tinggi tanaman 26,9 cm,bobot basah tajuk 1,129 g, bobot basah akar0,185 g, bobot kering tajuk 0,190 g danbobot kering akar 0,121 g.

Putri Kumalasari, Zozy Aneloi Noli dan Fuji Astuti Febria 202
DAFTAR PUSTAKA
Bloom Dharmawan, I. W. 2003. Pemanfaatanendomikoriza dan pupuk organik dalammemperbaiki pertumbuhan Gmelinaarborea LINN pada tanah tailing [Tesis].Bogor: Program Pasca Sarjana. InstitutPertanian Bogor.
Departemen Pertanian. 2005. Petunjuk TeknisAnalisis Kimia Tanah, Tanaman, Air danPupuk. Balai Penelitian danPengembangan Pertanian.
Fauziah, A. B. 2009. Pengaruh asam humatdan kompos aktif untuk memperbaiki sifatdengan indikator pertumbuhan tinggisemai Enterolobium cyclocarpum Grisebdan Altingia excelsa Noronhae. [Skripsi].Departemen Silvikultur. Bogor: InstitutPertanian Bogor.
Ghosh, M and Singh, S. P. 2005. A Review onPhytoremediation of Heavy Metals andUtilization of Its by-Products. AppliedEcology and Environmental Research,3(1):1–18.
Hanafiah, K.A. 2005. Dasar-dasar Ilmu Tanah.Rajawali Press. Jakarta.
Hidayati, N. 2004. Environmental DegradationAnd Biological Reclamation Of MinedLand: Case Of Gold Mining In Jampang-West Jawa. Di dalam: ProsidingWorkshop Vegetation Recovery inDegraded land Areas. Kalgoorlie,Western Australia, 6 Juni 2013.
Kelly, D.B, J. Cornish, R. Gordon And I. Licis.2003. Revegetation Of Mining WasteUsing Organic Amendments AndEvaluating The Potential For CreatingAttractive Nuisance For Wildlife. NationalMeeting of the American Society ofMining and Reclamation and the 9thBillings and Reclamation Symposium,Billings MT, June 3 – 6, 2003. Publishedby ASMR, 3134 Montevesta Rd.,Lexington, KY 40502. http://www. Billingslandreclamation symposium. org/tailings_abstr acts.htm. (6 Juni 2013).
Knox, A.S., Seaman, J., Andriano D.C.,Pierzynski, G. 2000. Chemostabilizationof metals in contaminated soils. Di dalam:Wise DL, Trantolo DJ, Cichon EJ, InyangHI, Stottmeister U (ed). Bioremediation of
Cotaminated Soils. New York: MarcekDekker Inc. hlm 811-836.
Kumalasari P., Z. A. Noli, F. A. Febria. 2013.Kandungan Logam Berat Merkuri (Hg)Pada Tanah Bekas Tambang Emas diSijunjung Sumatera Barat. Unpublish
Ladranada, H. K. 2009. Pengolahan KesuburanTanah. PT. Bina Aksara. Jakarta.
Priyanto, B dan Prayitno, J. 2005.Fitoremediasi sebagai Sebuah TeknologiPemulihan Pencemaran, KhususnyaLogam Berat. Makalah. IPB. Bogor.
Sagita, W.A. 2002. Uji Kemampuan AkumulasiLogam Kadmium dar Media oleh RumputGagajahan (Panicum maximum Jacq).Skripsi. S1 Biologi. ITB
Setiabudi, B Tjahjono. 2005. PenyebaranMerkuri Akibat Usaha PertambanganEmas Di Daerah Sangon, KabupatenKulon Progo, D.I. Yogyakarta. JurnalBiodiversitas 2(1) : 34-39.
Syarif F, N. Hidayati dan T. Juhaeti. 2009.Toleransi dan Akumulasi Sianida padaCentrosema pubescens Benth yangTumbuh di Media Limbah Tailing denganPerlakuan Tingkat Konsentrasi Sianidadan pH. BIOTA Jurnal Ilmiah Ilmu-ilmuHayati, Fakultas Teknologi UniversitasAtmajaya. Yogyakarta (Reefree)
Triastuti, Y. 2012. Fitoremediasi TanahTercemar Merkuri (Hg2+) MenggunakanTanaman Akar Wangi (VetiverZizanioides) Pada Lahan Eks-Tpa Keputih,Surabaya. Teknik Lingkungan- FTSP-ITS.Surabaya
Vangronsveld J et al. 2000. In situ inactivationand phytoremediation of metal-andmetalloid-contaminated soils: fieldexperiments. Di dalam: Wise DL,Trantolo DJ, Cichon EJ, Inyang HI,Stottmeister U (ed). Bioremediation ofContaminated Soils. New York: MarcekDekker Inc. hlm 859-884.
Van Reeuwijk, L.P. 1993. Procedures for SoilAnalysis. 4th ed. Technical Paper,International Soil Reference andInformation Centre. Wageningen, TheNetherlands
Yunus, Y. 2004. Tanah dan Pengelolaan.Alfabeta. Bandung

ISSN 978-602-14989-0-3 BioETI
Beberapa jenis mikroorganisme probiotik dan manfaatnyadalam kehidupanRAHMADHANI FITRI
Jurusan Biologi FMIPA Universitas Negeri PadangJalan Prof. Dr. Hamka, Air Tawar, PadangE-mail: [email protected]
ABSTRAKPenggunaan probiotik untuk memperkuat daya tahan tubuh dan melawan penyakit semakin banyak menarik perhatian.Faktor pendorong lainnya adalah adanya ketertarikan manusia yang mulai ingin hidup sehat dengan cara alami.Perkembangan informasi tentang probiotik lebih lanjut didorong oleh informasi adanya efek samping dari penggunaanantibiotik untuk terapi maupun growth promoter. Metode penelitian yang digunakan adalah metode deskriptif. Data-datadikumpulkan melalui studi kepustakaan. Berdasarkan hasil studi kepustakaan yang dilakukan didapatkan bahwamikroorganisme yang dipilih untuk penggunaan probiotik memiliki ciri: dapat menempel pada mukosa usus inangnya,mudah dibiakkan, tidak menjadi racun dan patogen bagi inangnya, memberikan efek yang menguntungkan bagi inangnya,dapat mentolerir HCl yang dihasilkan lambung inang dan garam empedu di usus kecil. Contoh mikroorganisme probiotikyang bermanfaat diantaranya tergolong pada Bakteri Asam Laktat, beberapa jenis khamir dan jamur. Mikroorganismeprobiotik ini banyak dimanfaatkan bagi kesehatan manusia dan ternak serta sebagai agen bioremidiasi yang bergunauntuk memperbaiki kualitas lingkungan.
Key words: BAL, Manfaat Probiotik, Mikroorganisme Probiotik
Pendahuluan
Penggunaan probiotik untuk memperbaikiproduktivitas ternak semakin banyak menarikperhatian para peneliti. Probiotik didefinisikansebagai substrat mikroorganisme, yangdiberikan kepada manusia atau ternak lewatpakan dan memberikan efek positif dengan caramemperbaiki keseimbangan mikroorganismealami di dalam saluran pencernaan (Estrada,1997 dalam Pamungkas dan Anggraeny, 2006).Probiotik berasal dari bahasa Latin yang artinyauntuk hidup dan mengalami perkembanganpengertiannya selama beberapa tahun ini.Probiotik adalah mikroorganisme dan substansiikutannya yang berperanan dalamkeseimbangan mikroorganisme saluranpencernaan. Definisi probiotik menurutHavenaar dan Huist dalam Pamungkas danAnggraeny (2006) adalah kultur tunggal ataucampuran dari mikrorganisme yang diberikankepada ternak atau manusia danmenguntungkan induk semang (inang) dengancara memperbaiki sifat-sifat mikroorganismealami dalam saluran pencernaan. Havenaar danHuist dalam Pamungkas dan Anggraeny (2006)
memberikan batasan pada istilah probiotikyaitu: (a) produk yang mengandungmikroorganisme dalam bentuk freeze dried atauproduk fermentasinya, (b) dapat memperbaikikesehatan pada manusia dan ternak dan targetorgannya adalah mulut atau saluran pencernaan(pemberian dapat lewat campuran pakan ataukapsul), bagian atas saluran pernafasan(pemberian dalam bentuk aerosol) dan saluranurogenitalia.
Sejak 1974, istilah probiotik mulaidihubungkan dengan feed supplement, namunsejarah penggunaan feed supplement yangberupa mikroorganisme hidup telah ada sejakribuan tahun yang lalu. Elie Metchnikof yanghidup tahun 1845–1916 membuat hipotesisbahwa, pada sebagian mikroorganisme saluranpencernaan memproduksi racun yang dapatmenyebabkan auto intoxication. Racun tersebutdapat merusak jaringan, akibatnya dapatmempercepat proses penuaan. Proses penuaanakibat auto intoxication tersebut dapat dicegahdengan implantasi bakteri asam laktat (sejenisdengan genus bakteri asam laktat yang terdapatdalam yoghurt). Diduga bakteri dapatmenghasilkan zat tertentu yang dapat

Rahmadhani Fitri 204
menghilangkan reaksi patologi pada prosesauto intoxication sehingga dapatmemperlambat penuaan denganmemperpanjang umur sel pada sebuah jaringan(Pamungkas dan Anggraeny, 2006).
Menurut Firmansyah (2001) penggunaanantibiotik dapat merusak keseimbanganmikroflora usus sehingga dapat menimbulkanefek samping yang tidak diinginkan.Pamungkas dan Anggraeny (2006) jugamenyampaikan bahwa perkembangan informasitentang probiotik lebih lanjut didorong olehinformasi adanya efek samping daripenggunaan antibiotik untuk terapi maupunsebagai growth promoter. Tahun 1969 Swann
Committe membatasi penggunaan antibiotiksebagai growth promoter. Akhirnya, kelompokanti aditif membatasi penggunaan antibiotikpada skala pengobatan. Beberapa supermarketdi Eropa saat ini telah menjual produk dagingbebas antibiotik dan di negara Skandinavia,antibiotik tidak digunakan lagi sebagai growthpromoter.
Mikroorganisme yang sering dimanfaatkansebagai probiotik diantaranya adalah beberapajenis bakteri. Bakteri yang biasa digunakansebagai probiotik pada umumnya berasal darigenus Lactobasillus dan Bifidobacterium.Kedua bakteri ini digolongkan ke dalamBakteri Asam Laktat (BAL). Selain darigolongan bakteri, beberapa jenis jenis khamirjuga dimanfaatkan sebagai probiotik (Suparjo,2008; Soeharsono 2010).
Berdasarkan hal tersebut di atas, penulistertarik untuk lebih mengetahui jenismikroorganisme yang dimanfaatkan sebagaiprobiotik dan manfaat penggunaanya dalamkehidupan sehari-hari. Jadi, tujuan daripenelitian ini adalah untuk mengetahuibeberapa jenis mikroorganisme probiotik danmanfaatnya dalam kehidupan.
BAHAN DAN METODE
Metode penelitian yang digunakan oleh penulisadalah metode deskriptif analisis. Metodedeskriptif analisis merupakan metode penelitian
dengan cara mengumpulkan data-data sesuaidengan yang sebenarnya kemudian data-datatersebut disusun, diolah, dan dianalisis untukdapat memberikan gambaran mengenaimasalah yang ada (Sugiyono, 2008). Metodedeskriptif analisis, merupakan metode yangdipergunakan untuk meneliti gagasan atauproduk pemikiran manusia yang telah tertuangdalam bentuk media cetak, baik yang berbentuknaskah primer maupun naskah sekunder denganmelakukan studi kritis terhadapnya.
Karena penulis menggunakan metodepenelitian deskriptif, maka penulismengumpulkan objek yang akan diteliti sertadata-data yang diperlukan untuk mendukungpenelitian melalui studi kepustakaan. MenurutHasan (2002) studi kepustakaan adalah teknikpengumpulan data dengan mengadakan studipenelaahan terhadap buku-buku, literatur-literatur, catatan-catatan, dan laporan-laporanyang ada hubungannya dengan masalah yangdipecahkan. Sumber-sumber kepustakaan dapatdiperoleh dari: buku, jurnal, majalah, hasil-hasil penelitian (tesis dan disertasi), dansumber-sumber lainnya yang sesuai (internet,koran, dan sebagainya). Data-data tentangmikroorganisme probiotik dan manfaatnyadalam kehidupan diperoleh dari studi literatureberupa buku, jurnal, dan hasil-hasil penelitianyang telah ada.
HASIL DAN PEMBAHASAN
A. Probiotik dan Ciri-ciri MikroorganismeProbiotik
Menurut Willey et.al (2008) dan Tortora et.al(2010) probiotik merupakan mikroba yangdiinokulasi ke dalam tubuh inang untukmenempati tubuh inang dan mencegahpertumbuhan pathogen pada tubuh inang.Mikroorganisme probiotik yang hidup dalamtubuh inang ini dimanfaatkan untukmeningkatkan kesehatan dan pertumbuhan, danjuga memiliki potensi untuk membangunkembali keseimbangan dalam tubuh inang danmengembalikan kesehatan normal dan gizi bagi

Rahmadhani Fitri 205
inang. Menurut Tortora et.al (2010) probiotikberasal dari kata pro yang berarti untuk danbios yang berarti hidup. Probiotik dapatdiaplikasikan dengan prebiotik, yangmerupakan bahan kimia yang selektifmeningkatkan pertumbuhan bakterimenguntungkan.
Menurut Abdulrahim, Haddadin,Haslamound dan Robinson dalam Trisna(2012) probiotik merupakan bahan tambahanberupa mikroorganisme yang berpengaruhterhadap peningkatan keseimbanganmikroorganisme dalam usus apabiladikonsumsi dalam jumlah yang cukup,probiotik mempunyai kemampuan untukmenurunkan kadar kolesterol serum darah(Kusumawati, Bettysri, Siswa, Ratihdewantidan Hariadi, 2003 dalam Trisna, 2012) dansebelumnya dikatakan bahwa, kandungankolesterol telur.
Probiotik merupakan feed additive yangmengandung mikroorganisme hidup yangmenguntungkan induk semang (inang), denganmemperbaiki keseimbangan mikroorganisme didalam saluran pencernaan. Berdasarkan hasilperkembangan ilmu pengetahuan, "probiotik"berkembang dari keyakinan bahwa,mikroorganisme saluran pencernaan terlibatdalam perlindungan inang terhadap kolonisasibakteri patogen pada saluran pencernaan.Keyakinan tersebut berdasarkan bukti-buktibahwa:1. Germ free animals sangat rentan terhadap
infeksi saluran pencernaan dibandingkandengan ternak konvensional yangmempunyai komposisi mikroflora saluranpencernaan lebih lengkap.
2. Pengobatan menggunakan antibiotik padaternak menyebabkan ternak lebih rentanterhadap infeksi saluran pencernaan.
3. Pengaruh perlindungan olehmikroorganisme asli saluran pencernaanpada ternak yang telah diberi antibiotikdapat dikembalikan dengan preparat yangberasal dari feses ternak sehat (Num dan
Rantala dalam Pamungkas dan Anggraeny,2006).
Menurut Willey et.al (2008) mikroorganismeyang dipilih untuk penggunaan probiotikharus menunjukkan ciri-ciri sebagai berikut.
1. Dapat menempel pada mukosa ususinangnya.
2. Dapat diisolasi dari inang dan mudahdibiakkan.
3. Mengandung sejumlah besar sel hidup ataumampu membentuk koloni yang banyak.
4. Mampu bertahan dan melakukan kegiatanmetabolisme dalam usus.
5. Tidak menjadi racun dan patogen bagiinangnya.
6. Memberikan efek yang menguntungkanbagi inangnya.
7. Menghasilkan enzim yang berguna atauproduk akhir fisiologis yang dapatdigunakan oleh inangnnya.
8. Mampu hidup dan tinggal pada tubuh inangpada jangka waktu yang lama. Selain ituprobiotik juga dapat bertahan hidup, mampubersaing, tidak hanya sekedar tumbuh dalamsaluran pencernaan.
9. Dapat mentolerir atau tahan terhadap HClyang dihasilkan lambung inang dan garamempedu di usus kecil.
Lisal dalam Anonim a (Tanpa tahun)menyampaikan karakterisitik probiotik yangdiinginkan dari satu strain spesifik, misalnya:1. Mempunyai kapasitas untuk bertahan hidup
(survive), untuk melakukanm kolonisasi(colonize), serta melakukan metabolisme(metabolize) dalam saluran cerna.
2. Mampu mempertahankan suatukeseimbangan mikroflora usus yang sehatmelalui kompetisi dan inhibisi kuman-kuman patogen.
3. Dapat menstimulasi bangkitnya pertahananimunitas, bersifat non-patogenik, dan non-toksik.
4. Harus mempunyai karakteristik teknologikyang baik, yaitu mampu bertahan hidupsecara optimal dan stabil selamapenyimpanan (storage) dan penggunaan

Rahmadhani Fitri 206
(use) dalam bentuk preparat makanan yangdidinginkan dan dikeringkan, agar dapatdisediakan secara massal dalam industri.
B. Mikroorganisme Penyusun ProbiotikMenurut Chen et al. dalam Pamungkas danAnggraeny (2006) mikroorganisme penyusunprobiotik yang aktif di saluran pencernaanbagian belakang pada umumnya adalah bakteriyang berasal dari genus Streptococcus,
Leuconostoc, Pediococcus, Propionicbacterium,Bacillus dan Enlerococcu. Spesies dari genusStreptococcus yang digunakan sebagaiprobiotik adalah Streptococcus salivarius,Streptococcus lactis, Streptococcus cremoris,
Streptococcus diacetilactis dan Streptococcusinterinedius. Mikroorganisme lain yang bisadijadikan probiotik adalah khamir dan jamur.Menurut Theodorou et al. dalam Pamungkasdan Anggraeny (2006) spesies khamir yangdapat digunakan sebagai probiotik adalahSaccharomyces cereviseae dan Candidapentolopesii, sedangkan spesies jamur yangdigunakan sebagai probiotik adalah Aspergillusniger dan Aspergillus oryzae. Neocallimastix sp.adalah jamur rumen yang digunakan sebagaiprobiotik yang terbukti efekif dapatmeningkatkan konsumsi pakan danpertambahan bobot badan.
Alternatif untuk menanggulangi penyakityang disebabkan bakteri patogen dapat melaluipendekatan biokontrol, yaitu penggunaanbakteri probiotik yang terdapat pada ususudang vanname (Litopenaeus vannamei). Daripenelitian yang pernah dilakukan Suminto dkk(2007 dalam Suminto, 2008) terdapat sepuluhjenis bakteri yang berasal dari usus udangvanname yaitu Enterobacter, Acinetobacter,
Bacillus, Lactobasillus, Mycobacterium,
Flavobacterium, Clostradium, Desulvabrio,Alkaligenes, dan Vibrio. Dari bakteri tersebut,terdapat empat bakteri yang berpotensi sebagaibakteri probiotik yaitu Flavobacterium,
Bacillus, Lactobasillus, dan Alkaligenes telahdiujikan dan mempunyai kemampuan untukmendegradasi lemak, protein, dan karbohidrat.
Menurut Trisna (2012) Bakteri Asam Laktat(BAL) adalah mikroorganisme yang dapatmembunuh dan menghambat pertumbuhanbakteri patogen yang ada disekitarnya. Suparjomendefinisikan BAL sebagai bakteripembentuk asam laktat dalam metabolismekarbohidrat dan terdiri dari berbagai macamkelompok bakteri gram positif.
Menurut Axelsson dalam Anonim a (Tanpatahun) deskripsi secara umum dari BAL iniadalah termasuk dalam bakteri gram positif,tidak berspora, berbentuk bulat maupun batang,dan menghasilkan asam laktat sebagaimayoritas produk akhir selama memfermentasikarbohidrat. Fardiaz dalam Anonim a (Tanpatahun) juga menambahkan bahwa berdasarkantipe fermentasinya, bakteri asam laktat terbagimenjadi heterofermentatif dan homofermentatif.Kelompok homofermentatif menghasilkanasam laktat sebagai produk utama darifermentasi gula, sedangkan kelompokheterofermentatif menghasilkan asam laktatdan senyawa lain yaitu CO2, etanol,asetaldehida, diasetil, serta senyawa lainnya.
Gambar 1. Bifidobacterium dari kultur susu.
Willey (2008) juga menyampaikan bahwamikroorganisme yang digunakan sebagaiprobiotik seperti dari Lactobacillusacidophillus dan genus Bifidobacterium
(Gambar 1). Menurut Abdulrahim, Haddadin,Haslamound dan Robinson (1996 dalam Trisna,2012) tidak semua bakteri baik dapat dijadikansebagai probiotik, salah satu bakteri yangberperan sebagai probiotik adalah BakteriAsam Laktat (BAL). BAL terbagi menjadi

Rahmadhani Fitri 207
delapan genus antara lain Lactobacillus,Streptococcus, Lactococcus, Pediococcus,Enterococcus, Leuconostoc, Bifidobacterium
dan Corinobacterium. Beberapa contohnyadapat di lihat pada Gambar 2.
Seperti yang disampaikan oleh beberapaahli di atas, bakteri probiotik yang digunakansecara komersial dewasa ini berasal dari genusLactobasillus dan Bifidobacterium (Heller2001; Suskovic dkk. 2001; Touhy 2003 dalamSuparjo, 2008). Beberapa spesies bakteri yangbiasa digunakan sebagai probiotik ditampilkanpada Tabel 1.
Menurut Shin et al., 1989 dalam Asmarasaridan Zain (Tanpa tahun) penggunaan probiotiksaat ini telah menunjukkan perkembangan yangpesat pada hewan ternak, sebagai penggantiantibiotik. Probiotik yang umum dan amandigunakan untuk ternak, yaitu: Aspergillusniger, A. oryzae, Bacillus coagulans, B. lentus,B. pumilus, Bacteriodes amylophilus, B.ruminocola, Lactobacillus acidophilus, L.brevis, Leuconostoc mesenteroides,Pediococcus acidolacticii, Propionibacteriumshemanii, Sacharomyces cereviseae,Streptococcus cremoris, S. faecium, S. lactisdan S. thermophilus. Gilliland, 2004 dalamAsmarasari dan Zain (Tanpa tahun) jugamenyampaikan bahwa probiotik campuranyang mengandung Enterococcus faecium danLactobacillus acidophilus ditambah kulturkapang, digunakan untuk meningkatkanproduksi susu pada ternak sapi perah.S eperti yang telah disampaikan di atas, adabeberapa contoh mikroorganisme lain yangterdapat dari beberapa produk yangmengandung probiotik seperti yang terdapatdalam Majalah Supermarket Savvy (2007) yaituLactobasillus bulgaricus, Streptococcus
thermophilus, L. acidophilus, Bifidobacteriumbifidus, B. longum, L. casei, dan L. reuteri, L.
rhamnosus, L. lactis, L. plantarum,Leuconostoc cremoris, B. breve, S.
diacetylactis, dan S. florentinus.Berdasarkan hasil penelitian Santoso dkk.
(2013) BAL yang dapat dijadikan kandidat
probiotik pada ternak dapat diisolasi rumputraja (Pennisetum purpureophoides). Isolat BALyang teridentifikasi dari ekstrak Pennisetum
purpureophoides adalah strain Lactobacillus
plantarum. L. plantarum masih mampubertahan pada pH ekstrim 2,0 dan garamempedu dengan konsentrasi 0,3% sertamembutuhkan suhu 30°C untuk pertumbuhanyang optimum. L. plantarum juga mampumenghambat pertumbuhan bakteri patogenStaphylococcus aureus, Salmonella typhi danEscherecia coli. L. plantarum. mempunyaipotensi untuk digunakan sebagai probiotik padaternak.
Selain itu, berdasarkan hasil penelitianMutmainnah dkk. (Tanpa tahun) bakteriprobiotik juga dapat di isolasi dari saluranpencernaan ayam. Hasil isolasi bakteriprobiotik dari saluran pencernaan ayamkampung didapatkan 11 isolat. Berdasarkan ujifisiologi dan biokimia kesebelas isolatmenunjukkan karakteristik probiotik.Keseluruhan isolat tersebut tergolong bakterigram positif dimana isolat A, B, G, J, dan Kselnya berbentuk bulat (coccus) didugatergolong Lactococcus, Streptococcus,Leuconostoc, Enterococcus dan Pediococcus.
Isolat C, D, E, F, H, dan I berbentuk batang(basil) diduga Lactobacillus. Keseluruhanisolat mampu tumbuh pada pH rendah maupunpada konsentrasi ox bile 1% dan 5% serta padasuhu rendah dan suhu tinggi. Mampumenghambat pertumbuhan bakteri ujiEscherichia coli dan Staphylococcus aureus.
Goldin dan Gorbach dalam Hardiningsihdkk. (2006) mengatakan bahwa beberapasubstansi antimikroba yang dihasilkan bakteriprobiotik, misalnya L. acidophilus
menghasilkan acidotin, acidophilin, bacteriocin,lactocidin, L. bulgaricus (bulgarican), L.plantarum (lactolin), L. brevis (lactobullin,lactobrevin), dan L. reuteri (rauterin).
C. Manfaat dan Peranan ProbiotikProbiotik merupakan mikroorganisme yang
bila dikonsumsi akan memberikan dampakpositif bagi kesehatan dan merupakan galur

Rahmadhani Fitri 208
flora usu normal yang dapat diisolasi dari fesesyang sehat. Kaitan ilmiah antara probiotik danmanfaatnya bagi keshatan manusia pertama kalidiungkapkan oleh ahli mikrobiologi Rusiabernama Metchnikoff. Ia menyatakan bahwaasam laktat yang dihasilkan oleh Lactobasillusdalam yogurt dapat menghambat pertumbuhanbeberapa spesies bakteri pathogen. Probiotikjuga bermanfaat dalam pencegahan danpengobatan beberapa penyakit saluran cerna,termasuk diare infeksi, diare karena antibiotic,travellers diarrhea, dan intoleransi laktosa.Penggunaan probiotik sejauh ini aman tetapidianjurkan berhati-hati pada anakimunokompromais (Firmansyah, 2001).
Menurut Haryabriansyah (2004) probiotikselain sebagai growth promotor ternayata jugadipercaya memiliki efek imunomodulasi karenadapat mempengaruhi sistem kekebalan tubuhnon spesifik dan sistem kekebalan spesifik.Pada sistem kekebalan non spesifik, probiotikmampu menciptakan suasana asam yang akanmenghambat pertumbuhan orgnaisme patogen,melapisi sel epitel mukosa usus sehingga dapatmenghambat perlekatan mikroorganismepathogen. Selain itu probiotik mampumeningkatkan penyerapan nutrisi dari pakanoleh usus. Pada sistem kekebalan spesifik,probiotik dapat meningkatkan sirkulasi antibodidelapan kali dari kondisi normal dan jugamampu mengingkatkan aktivitas sertaproliferasi sel limfosit. Beberapa spesies dariLactobasillus menunjukkan kemampuan dalammenginduksi pengeluaran sitokin. Probiotikjuga dapat mendegradasi protein susu danmampu membebaskan molekul peptide yangdapat merangsang respon imun.
Selain bakteri, khamir dan jamur jugadigunakan sebagai probiotik. Khamir dan jamursering digunakan bersama sebagai probiotikpada ternak ruminansia dan sering disebutfungal feed additives atau jamur probiotik. Adabeberapa probiotik yang tidak begitu dikenaltetapi telah diteliti seperti Saccaromycesboulardii (Oezteurk et.al dalam Pamungkasdan Anggraeny, 2006) dan Issatcenkia
orientalis (Lee et.al dalam Pamungkas danAnggraeny, 2006). Penggunaan mikrobatersebut memberikan keuntungan padapeningkatan efisiensi fermentasi di dalamrumen, peningkatan kecernaan hijauan, danpeningkatan laju aliran protein mikroba darirumen (Wallace dan Newbold dalamPamungkas dan Anggraeny, 2006).
Pemberian probiotik ke ternak ruminansiadapat memperbaiki laju pertumbuhan,menstabilkan produksi pada ternak, efisiensikonversi ransum, meningkatkan penyerapannutrisi, kesehatan hewan, menambahkan nafsumakan sehingga mempercepat peningkatanberat badan dan memperbaiki kualitas feces(Zakariah, 2012).
Menurut Pamungkas dan Anggraeny (2006)pengaruh penggunaan probiotik ternakruminansia belum konsisten meskipunbeberapa penelitian menghasilkan pengaruhnyata baik pada produksi daging dan susu, sertaketahanan terhadap penyakit. MenurutAsmarasari dan Zain (Tanpa tahun)menyampaikan bahwa pemberian probiotikLactobasillus acidophilus pada sapi perahmeningkatkan produksi susu. MenurutGarriques dan Arevalo dalam Murtiati dkk.(2007) peranan bakteri probiotik sebagaicontrol biologis pada sistem budi daya sebagaiberikut.1. Menekan pertumbuhan bakteri patogen.2. Mempercepat degradasi bahan organik dan
limbah.3. Meningkatkan ketersediaan nutrisi esensial.4. Meningkatkan aktivitas mikroorganisme
indigenus yang menguntungkan padatanaman, missal Mikoriza, Rhizobium, danbakteri pelarut posfat.
5. Memfiksasi nitrogen.6. Mengurangi penggunaan pupuk dan
pestisida.Menurut Murtiati dkk. (2007) dengan
adanya probiotik, maka proses degradasi bahanorganik pada dasar tambak akan lancar,sehingga menghasilkan zat-zat yangbermanfaat bagi pertumbuhan plankton. Bahan

Rahmadhani Fitri 209
organik yang mengalami mineralisasi olehjasad pengurai (probiotik) akan diubah menjadibahan anorganik seperti nitrat dan posfat.Bahan anorganik ini dapat digunakan secaralangsung oleh fitoplankton dalam air untukkelangsungan hidupnya.
Dari hasil penelitian yang dilakukanSaputri (2012) didapatkan hasil bahwapemberian probiotik bakteri Pediococcuspentosaceus mampu menurunkan kadarTrigleserida pada daging itik pitalah denganpenurunan optimal yaitu pada pemberian 3 mldengan persentase 61,06%. Selain itu denganpemberian 3 ml probiotik Pediococcuspentosaceus meningkatkan jumlah koloniBakteri Asam Laktat (BAL) yaitu sebanyak 5,2x107 cfu/g, dan meningkatkan keseimbanganmikroflora usus dengan persentaseperbandingan BAL 95,9% dan patogen 4,1%.Hal yang sama juga dilakukan oleh Trisna(2012) yang juga melakukan penelitian denganhasil bahwa pemberian probiotik bakteriPediococcus pentosaceus 2 ml mampumenurunkan kadar kolesterol daging itik pitalahdari 39,50 menjadi 32,19.
Hal ini didukung oleh hasil penelitianNapitupulu et al., (2003 dalam Yulinery dkk.,2006) menunjukkan bahwa Lactobacillus sp.Mar 8 relatif lebih tepat guna sebagai probiotikpenurun kolesterol dilihat dari daya ikatkolesterol dan ketahanan terhadap garamempedu (Na-taurokolat). Penelitian secara invivo menunjukkan bahwa pemberian suspensiprobiotik Lactobacillus sp. Mar 8 dapatmenurunkan kolesterol pada hari ke-28(Kurniawati, 2003), terjadi penurunan LowDensity Lipoprotein (LDL) kolesterol dan tidakterjadi penurunan High Density Lipoprotein
(HDL) kolesterol (Yulinery et al., 2004 dalamYulineri dkk., 2006).
Menurut Avnimelech (1999 dalam Sartika,dkk., 2012) probiotik dapat digunakan sebagaiagen bioremediasi yang berguna untukmemperbaiki kualitas lingkungan budidaya.Penggunaan probiotik sangat bermanfaat dalammeningkatkan populasi bakteri agen
bioremediasi karena bakteri probiotik dapatmencegah bakteri patogen agar tidakmemperbanyak diri dalam media hidup hewanbudidaya dengan melawan permunculan kolonibakteri lain sehingga diharapkan bakteri yangtumbuh merupakan bakteri agen bioremediasi.
Menurut Trisna (2012) Bakteri Asam Laktat(BAL) adalah mikroorganisme yang dapatmembunuh dan menghambat pertumbuhanbakteri patogen yang ada disekitarnya. Selaindapat membunuh dan menghambatpertumbuhan bakteri patogen, juga dapatmenurunkan kadar kolesterol pada dagingternak apabila diberi BAL tersebut. BAL jugadapat menekan penyakit yang terkenal padaunggas yaitu penyakit AI, BAL berpengaruhbaik terhadap peningkatan keseimbanganmikroflora usus bila dikonsumsi dalam jumlahyang cukup. Hal ini didukung oleh Suparjo(2008) bahwa BAL mempunyai perananpenting dalam pengawetan bahan pangan danmelawan bakteri patogen melalui senyawapeptida antimikroba.
Menurut Abdulrahim dkk., (1996 dalamTrisna, 2012) probiotik dapat meningkatkankesehatan ternak, meningkatkan produksi telur,serta dapat menghilangkan sifat reservoar AIpada itik. Menurut Tortora et al., (2010) JikaBAL membentuk koloni di usus besar, asamlaktat dan bakteriosin yang dihasilkan dapatmenghambat pertumbuhan patogen tertentu.Para peneliti juga menguji penggunaan BALuntuk mencegah infeksi luka bedah yangdisebabkan oleh Stapylococcus aureus daninfeksi vagina yang disebabkan oleh E. coli.Dalam sebuah penelitian di University Stanford,infeksi HIV berkurang pada wanita yangdiobati dengan BAL yang secara genetikdimodifikasi untuk menghasilkan protein CD4yang mengikat HIV.
Penggunaan BAL atau mikroorganismeprobiotik lainnya juga merupakan suatu carapendekatan untuk mengurangi atau mencegahterjadinya kontaminasi penyakit terutamapenyakit thipus terhadap produk-produk unggasyaitu daging dan telur, sehingga daging dan

Rahmadhani Fitri 210
telur yang dihasilkan higienis dan aman untukdikonsumsi sesuai dengan standar kesehatan(Patterson dan Burkholder, 2003 dalam Trinsa,2012). Selain itu BAL sering digunakansebagai kultur probiotik dalam produk-produkfermentasi susu seperti dadih, buah-buahan,daging dan ikan.
Menurut Saputri (2012) BAL bisamenghasilkan enzim Bile Salt Hydrolase yangbisa menurunkan kadar kolesterol dan enzimlipase yang bisa menurunkan trigliserida karenakemampuannya dalam memutuskan asamlemak rantai panjang menjadi asam lemakrantai sedang dan rantai pendek sehinggamudah diserap dalam usus. BAL sepertiLactobacillus disamping menurunkan kadarkolesterol dan trigliserida juga berfungsi untukmemelihara kesehatan dan meningkatkan dayatahan tubuh ternak. Menurut Purwati danSyukur (2006 dalam Saputri, 2012) pemberianblondo jamur akan menciptakan keseimbanganmikroflora usus, karena adanya bakteri asamlaktat dalam usus yang dapat menciptakansuasana asam sehingga menekan pertumbuhanbakteri patogen dalam usus halus.
Dari hasil penelitian yang dilakukanWidanarni dkk. (2010) di dapatkan bahwaVibrio SKT-b merupakan salah satu jenisbakteri kandidat probiotik yang telah diujidapat menekan pertumbuhan bakteri patogenVibrio harveyi dan dapat meningkatkankelangsungan hidup larva udang windu.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Mikroorganisme yang dipilih untukpenggunaan probiotik memiliki ciri-ciri sebagaiberikut: dapat menempel pada mukosa ususinangnya, mudah dibiakkan, tidak menjadiracun dan patogen bagi inangnya, memberikanefek yang menguntungkan bagi inangnya,menghasilkan enzim yang berguna atau produkakhir fisiologis yang dapat digunakan olehinangnya, mampu hidup dan tinggal pada tubuh
inang pada jangka waktu yang lama, dapatmentolerir atau tahan terhadap HCl yangdihasilkan lambung inang dan garam empedudi usus kecil. Mikroorganisme probiotik inibanyak dimanfaatkan bagi kesehatan manusiadan ternak serta sebagai agen bioremidiasi yangberguna untuk memperbaiki kualitaslingkungan.
Mikroorganisme yang termasuk padaprobiotik yang bermanfaat diantaranya adalahbakteri asam laktat yang terdiri dari beberapagenus, khamir, dan juga beberapa jenis jamur.Contoh mikroorganisme probiotik itudiantaranya adalah adalah Lactobasillusbulgaricus, Streptococcus thermophilus, L.
acidophilus, Bifidobacterium bifidus, B.longum, L. casei, dan L. lactis, L. plantarum,Leuconostoc cremoris, B. breve, S.
diacetylactis, S. florentinus, Aspergillus niger,A. oryzae, dan Saccharomyces cereviseae.
DAFTAR PUSTAKA
Anonim. Tanpa tahun. Tinjauan Pustaka.Bogor: IPB.
Asmarasari, S. A., Zain, W. N. H. Tanpa tahun.“Respon Pemberian Probiotik dalamPakan terhadap Produksi Susu Sapi Perah”.Artikel disampaikan pada SemilokaNasional Prospek Industri Sapi PerahMenuju Perdagangan Bebas 2020,hlm192-195.
Firmansyah, A. 2001. “Terapi Probiotik danPrebiotik pada Penyakit Saluran CernaAnak”. Sari Pediatri, Vol. 2, No. 4: 210-214.
Hardiningsih, R., Napitupulu, R. N. F.,Yulinery, T. 2006. “Isolasi dan UjiResistensi Beberapa Isolat Lactobasilluspada pH Rendah. Biodiversitas, Volume 1,Nomor 7: 15-17.
Haryabriyansah, K. 2004. “Prospek ProbiotikGolongan Bakteri Asam Laktat sebagaiImunomudulator”. Skripsi. Bogor: FKHIPB.
Hasan, M. Iqbal. 2002. Pokok-pokok MateriMetodologi Penelitian dan Aplikasinya.Bogor: Ghalia Indonesia.
McDonald L. (Ed.). 2007. “Eat Your Bacteria:Understanding Probitics and Live ActiveCultures”. Supermarket Savvy, pages: 1-9.

Rahmadhani Fitri 211
Murtiani, Simbolon K., Wahyuni, T., Juyana.2007. “Penggunaan Biokatalisator padaBudidaya Udang Galah”. Jurnal BudidayaAir Tawar, Volume 4, Nomor 1: 19-26.
Mutmainnah, Heni, Risco B. Gobel, NatsirDjide, Zaraswati Dwyana. Tanpa tahun.“Isolasi dan Karakterisasi BakteriProbiotik dari Saluran Pencernaan AyamKampung Gallus Domesticus”. Artikel,Makasar: Universitas Hasanuddin
Pamungkas, D., Anggraeny, Y. N. 2006.“Probiotik dalam Pakan TernakRuminansia”. Wartazoa, Vol. 16, No. 2:82-91.
Santoso B., Maunatin A., Hariadi BT.,Abubakar H. 2013. Isolasi dan IdentifikasiBakteri Asam Laktat Asal Rumput Raja(Pennisetum purpureophoides) sebagaiKandidat Probiotik pada Ternak. JITV,Vol.18 No.2: 131-137.
Saputri F. 2012. “Pengaruh PemberianProbiotik Bakteri Asam Laktat (BAL)Pediococcus pentosaceus terhadapKeseimbangan Mikroflora Usus danTrigleserida Daging Itik Pitalah. ArtikelProgram Pascasarjana UniversitasAndalas: 1-7.
Sartika, D., Esti H., Rara D. 2012. “PemberianMolase pada Aplikasi Probiotik terhadapKualitas Air, Pertumbuhan, dan TingkatKelangsungan Hidup Benih Ikan Mas(Cyprinus carpio)”. e-Jurnal Rekayasadan Teknologi Budidaya Perairan, Vol. 1No.1: 57-64.
Soeharsono. 2010. Probiotik Basis Ilmiah.Bandung: Widya Padjajaran.
Sugiyono. 2008. Metode Penelitian Kuantitatif,Kualitatif, dan R & D. Bandung: Alfabeta.
Suminto. 2008. “Pertumbuhan BakteriProbiotik Alkaligenus sp. dan
Flavobacterium sp. yang Diisolasi dariUsus Udang pada Media Kultur Molasedan Kaolin”. Jurnal Saintek Perikanan,Vol. 4, No. 1: 21-27.
Suparjo. 2008. Bakteriosin dan Perannyadalam Ekologi Mikroba Rumen. Jambi:Laboratorium Makanan Ternak FakultasPeternakan Universitas Jambi.
Tortora, G. J., Funke, B. R., Case, C. L. 2010.Microbiology: An Introduction TenthEdition. New York: Benjamin Cummings.
Trisna, W. N. 2012. “Identifikasi Molekulerdan Pengaruh Pemberian Probiotik BakteriAsam Laktat (BAL) Asal Dadih dariKabupaten Sijunjung terhadap KadarKolesterol Daging pada Itik PitalahSumber Daya Genetik Sumatera Barat.Artikel Program Pascasarjana UniversitasAndalas: 1-31.
Wirdanarni, Lidaenni, M. A., Wahjuningrum,D. 2010. “Pengaruh Pemberian BakteriProbiotik Vibrio SKT-b dengan Dosisyang Berbeda terhadap KelangsunganHidup dan Pertumbuhan Larva UdangWindu (Penaeus monodon) Fab.”. JurnalAkuakultur Indonesia 9(1): 21-29.
Willey, J. M., Sherwood, L. M., Woolverton, C.J. 2008. Prescott, Harley, and Klein’sMicrobiology. New York: McGraw Hill.
Yulinery, T., Yulianto, E., Nurhidayat, N. 2006.“Uji Fisiologis Probiotik Lactobacillus sp.Mar 8 yang Telah Dienkapsulasi denganMenggunakan Spray Dryer untukMenurunkan Kolesterol”. Biodiversitas,Volume 7, Nomor 2: 118-122.
Zakariah, M. A. 2012. Penggunaan Probiotikuntuk Meningkatkan Produktivitas TernakRuminansia. Yogyakarta: FakultasPeternakan UGM.

ISSN 978-602-14989-0-3 BioETI
Potensi sektor pertanian dan perkebunan Kabupaten PasamanBarat untuk menghadapi pasar bebas aseanRELSAS YOGICA
Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Negeri PadangE-mail: [email protected]
ABSTRAKDalam rangka mewujudkan visi 2020 ASEAN akan diterapkan pasar tunggal untuk wilayah Asia Tenggara. Pasar tunggalini akan membentuk jalur bebas barang, jasa, tenaga kerja dan investasi. Sebagai anggota ASEAN, Indonesia tentu sajaakan ikut serta dalam proses ini. Dalam rangka memperkuat integritas Indonesia dibutuhkan dasar perekonomian yangkuat, tentu saja ini didukung penuh oleh potensi unggulan daerah.Kabupaten Pasaman Barat merupakan salah satuwilayah pertanian dan perkebunan di Sumatera Barat.Komoditas unggulan Pasaman Barat adalah kelapa sawit, jagung,padi, pisang dan ubi jalar.Makalah ini membahas tentang potensi Kabupaten Pasaman Barat dari sektor komoditasunggulan pertanian dan perkebunan. Diharapkan dengan pembahasan yang telah dibuat maka akan diketahui secara lebihjelas mengenai potensi pertanian dan perkebunan Pasaman Barat untuk dapat dikembangkan demi keikutsertaanmasyarakat daerah menghadapi pasar bebas ASEAN.
Key words: pasar bebas, perkebunan, pertanian, potensi
Pendahuluan
Pasar bebas di wilayah Asia Tenggaramerupakan salah satu visi ASEAN 2020 yangditetapkan pada KTT ASEAN ke-2 di Malaysia,dengan adanya pasar bebas ini maka akanterbentuk sebuah pasar tunggal di AsiaTenggara yang menyediakan jalur bebas untukarus barang, jasa, tenaga kerja dan investasi[1].Pasar bebas ASEAN akan terlaksana padatahun 2015, lebih cepat dari tahun yangdirencanakan awalnya yaitu 2020, tujuannyaadalah untuk memperkuat daya saing ASEANdalam menghadapi kompetisi global khususnyadengan China dan India.Keuntungan yangdiharapkan adalah keseimbangan sektorperekonomian negara-negara anggota.Dalampelaksanaan pasar bebas, terdapat 12 sektoryang menjadi prioritas liberalisasi. Sektor-sektor tersebut adalah produk pertanian,angkutan udara, otomotif, e-ASEAN,elektronik, perikanan, kesehatan, produk karet,tekstil, pariwisata, produk kayu dan jasalogistik[1].
Sebagai salah satu negara ASEAN,Indonesia tidak akan lepas dari proses ini.Indonesia akan menjadi salah satu negara
ASEAN yang menerapkan perdagangan bebas,oleh karena itu Indonesia dituntut untukmempersiapkan diri dalam beberapa sektorpotensial. Tahun 2007 telah dibuat kartu skoryang bertujuan untuk menghitung angkakesiapan negara ASEAN dalam menghadapipasar bebas. Pada periode 1 Januari 2008sampai 30 September 2009, sekretariat ASEANmelaporkan kesiapan Indonesia menghadapipasar bebas ASEAN berdasarkan kartu skortersebut adalah 80,37%[1]. Angka ini dibawahMyanmar (85,05%), Vietnam (88,13%) danSingapura (93,52%).
Sebagai negara agraris, Indonesiamempunyai sektor unggulan untuk menguasaipasar bebas ASEAN yaitu sektor pertanian,kayu dan kulit[3]. Bahkan pada bidang obat-obatan tradisional, Presiden DirekturSidomuncul menegaskan bahwa Indonesia jauhlebih siap dibandingkan Malaysia, Vietnam danThailand[4]. Kementerian Pertanianmengemukakan bahwa produksi komoditaspangan strategis dalam empat tahun terakhirumumnya menunjukkan kinerja yang baikseperti produksi padi mengalami peningkatanrata-rata 2,6 persen per tahun dari 64,4 juta tongabah kering gilingpada 2009 menjadi 71,29

Relsas Yogica 213
juta ton gabah kering giling pada 2013.Produksi jagung juga meningkat rata-rata 1,39persen per tahun dari 17,63 juta ton jagungpipilan kering tahun 2009 menjadi 18,51 jutaton tahun 2013.
Indonesia memiliki potensi yang bagusuntuk bidang pertanian dan perkebunan, hal initentu saja didukung oleh daerah-daerahsentra.Salah satu daerah tersebut adalahKabupaten Pasaman Barat, SumateraBarat.Pasaman Barat terletak 175 km ke arahutara pusat kota Provinsi, Padang[2]. PasamanBarat memiliki 11 kecamatan dengan luasdaratan total adalah 3.887,77 km2 atau sekitar9,29% total wilayah Provinsi Sumatera Barat,dan luas lautan total adalah 800,47 km2 denganpanjang garis pantai 152 km. Bentukanwilayahnya bergelombang dengan beberapabukit dan gunung, ketinggian wilayah jugabervariasi dari 0mdpl sampai 2.912 mdpl.Kondisi wilayah yang seperti ini mendukungbanyak sektor kehidupan masyarakat,termasuksektor pertanian dan perkebunan.Sektor ini diharapkan berperan dalammemperkuat integritas Indonesia dalam pasarbebas ASEAN 2015 dengan carameningkatkankualitas diri melalui penguasaan teknologi danmeningkatkan produksi barang atau jasadengan memperhatikan efektifitas danefisiensinya. Tujuannya adalah agar produkyang dihasilkan menjadi salah satu produkyang diminati di pasar ASEAN.
Berdasarkan penjelasan di atas, makapenulis merasa tertarik untuk menyajikanmakalah mengenai potensi sektor kehidupanmasyarakat Pasaman Barat untuk menghadapipasar bebas ASEAN.Dalam makalah inidibatasi pada sektor pertanian dan perkebunanunggulan, yaitu sawit, jagung, padi, pisang danubi jalar.Komoditas unggulan yang dimaksuddalam makalah ini adalah komoditas yangmemiliki nilai produksi tinggi dan ketersediaanlahan yang cukup. Sebagai bahan penunjangjuga akan dibahas mengenai geografis,kependudukan dan kesempatan investasi diPasaman Barat. Diharapkan dengan adanya
kajian ini maka juga akan memberikangambaran dan peluang investasi pada masayang akan datang.
BAHAN DAN METODE
Penulisan makalah ini menggunakanpendekatan penelitian deskriptif dengan sumberdata primer.Data primer merupakan data yangdiperoleh langsung dari pihak pertama, data inibersifat masa lampau dan tidak dilakukanperlakuan khusus terhadap variabel yangdiamati.Metode yang digunakan dalampengumpulan data adalah studiliteratur.Tahapan pengumpulan dan penyajiandata makalah adalah:1. menemukan informasi mengenai kondisi
wilayah Pasaman Barat,2. menemukan informasi mengenai kehidupan
sektor pertanian dan perkebunan masyarakatPasaman Barat melalui situs pemerintahkabupaten yaitu www.pasamanbarat.com,
3. mengoleksi informasi yang dibutuhkan,4. memilih 5 komoditas pertanian dan
perkebunan unggulan masyarakat,5. menyajikan dalam bentuk tabel hasil yang
telah diperoleh.Metode pemilihan sampel penulisan makalahini adalah dengan teknik purposive
sampling.Penulis memiliki tujuan untukmenambah nilai daerah Pasaman Barat darisektor pertanian dan perkebunan kepadamasyarakat Indonesia khususnya danmasyarakat dunia pada umumnya.
HASIL DAN PEMBAHASAN
A. GeografiKabupaten Pasaman Barat merupakan salahsatu kabupaten di Provinsi Sumatera Baratyang lahir pada 7 Januari 2004 berdasarkan UUNo.38 tahun 2003.Pasaman Barat memiliki luaswilayah yang cukup besar yakni lebih dari 9%luas wilayah provinsi, dengan bentukpermukaan tanah yang bergelombang. Jarakkabupaten dari Kota Padang adalah 180km.Pasaman Barat memiliki pantai sepanjang

Relsas Yogica 214
152 km dan deretan pegunungan bukit barisan,dua gunung terbesar adalah Gunung Talamaudan Gunung Pasaman.Pasaman Barat terletakpada 00°33” LU - 00°11 LS dan 99°10” BT -100°04 BT. Batas wilayah bagian utara adalahKabupaten Mandailing Natal, bagian selatanadalah Kabupaten Pasaman dan KabupatenAgam, bagian barat adalah Samudera Hindiadan bagian timur adalah KabupatenPasaman[2].Kabupaten Pasaman Barat memiliki11 kecamatan yang berperan sebagai pusatbeberapa komoditas.
Letak dan kondisi tanah akan memberikankeuntungan bagi masyarakat Pasaman Barat.Sektor perikanan dan pariwisata dapatdikembangkan dengan cepat dan mudah karenaberbatasan langsung dengan SamuderaHindia.Kondisi tanah yang berada disekitarpegunungan dapat digunakan secara baik untuksektor pertanian dan perkebunan, karenamenyediakan tekstur yang pas dan mineralyang cukup untuk berbagai komoditas panganmaupun komoditas ekspor.Ketersediaan lahancukup luas untuk dapat dikembangkan.B. KependudukanPenduduk di Pasaman Barat memilikikeanekaragaman suku dan pekerjaan yangberbeda.Kabupaten ini memiliki pendudukyang sebagian berasal dari Kabupaten Pasaman,mengingat bahwa Kabupaten Pasaman Barathasil pemekaran Kabupaten Pasaman tahun2004.Pertumbuhan penduduk berdasarkan datadari BPS 2010 mengalami peningkatan sebesar3%. Penduduk berasal dari suku berbedadengan pekerjaan yang yang beranekaragampula. Suku-suku yang ada di Pasaman Baratantara lain adalah minang, jawa, batak danmandahiling. Pekerjaan yang dikerjakan antaralain adalah bertani, berkebun, pegawai negerisipil dan pegawai pemerintahan dan masih adapekerjaan lainnya.
Lahan yang mendukung untukmengembangkan sektor pertanian danperkebunan membuat penduduk mengarahkanusaha ke arah pertanian dan perkebunan,termasuk pegawai negeri sipil dan pegawai
pemerintahan. Saat pagi mereka bekerja dikantor, kemudian sore harinya bekerja di lahan.Beberapa juga memberikan upah kepada oranglain untuk mengurus lahan yang mereka punya.Pada tahun ini, petaniPasaman Barat mewakiliProvinsi Sumatera Barat ke kompetisi tingkatnasional pada sektor tanaman jeruk,holtikultura, sayur dan tanaman hias[5]. Inimerupakan suatu kesempatan promosi yangluas bagi sektor pertanian dan perkebunan, danjuga membuktikan bahwa selain sektorunggulan yang dikelola masyarakat juga adatanaman pertanian dan perkebunan lain yangpatut untuk diperhitungkan secaraglobal.Contohnya jeruk tersebut, tahun 2011perkebunan jeruk mengalami kemundurandrastis karena kerusakan lahan oleh banjir danrusaknya batang karena virus. Namun padatahun 2014 ini pemerintah kembalimenggiatkan penanaman jeruk, usaha yangdilakukan pemerintah salah satunya adalahpemberian bibit jeruk unggul sebanyak 12.000batang kepada petani disertai dengan pelatihanpengelolaan perkebunan jeruk.C. Komoditas pertanian dan perkebunan
unggulanProduksi pertanian dan perkebunan masyarakatantara lain adalah sawit, jagung, padi, pisang,ubi jalar, jeruk, pepaya, kelapa, nilam danbeberapa jenis kayu. Komoditas unggulandaerah adalah sawit, jagung, padi, pisang danubi jalar dengan sebaran produksi ada pada 11kecamatan, yaitu Kecamatan Talamau,Pasaman, Luhak Nan Duo, Sasak Ranah Pasisie,Kinali, Gunung Tuleh, Sungai Aua, LembahMelintang, Koto Balingka, Sungai Beremas danRanah Batahan.1. SawitPerkebunan sawit di Pasaman Barat memilikiluas lahan 150.785 Ha pada tahun 2013. Lahanini menghasilkan 222.489 ton sawit pada tahunyang sama dengan angka produktivitasnyaadalah 1,4. Sekitar 80% masyarakatmenggantungkan hidupnya pada sektorperkebunan yang luasnya mencapai 39% luaskabupaten[2], namun sebagian besar masyarakat

Relsas Yogica 215
hanya berperan sebagai pekerja di perkebunandan bukan sebagai pemilik. Produksi sawit diPasaman Barat pada tahun 2013 jikadibandingkan dengan tahun sebelumnyamengalami peningkatan, pada tahun 2012 hasilproduksi sawit adalah 114.619 ton. Perkebunansawit di Pasaman Barat dikendalikan olehpihak swasta dalam kapasitas makro dan mikrodan juga BUMN (PTP).Di Kecamatan GunungTulehterdapat pabrik pengolahan sawit.
Perkebunan sawit di Pasaman Barat dapatdikelompokkan pada usia tua yang nyaris tidakproduktif lagi, sehingga membutuhkanperemajaan. Hal ini sudah dilakukan padabeberapa tahun terakhir di beberapa pusatperkebunan PTPN VI Ophir.Minyak sawitmerupakan barang ekspor utama Indonesia keASEAN dan dunia[1], ini dapat menjadipertimbangan menarik investor untukmenanamkan kepercayaan pada perkebunansawit di Indonesia umumnya dan PasamanBarat khususnya.2. JagungLuas pertanian jagung di Pasaman Barat padatahun 2013 adalah 42.386 Ha (10% luaskabupaten) dengan hasil produksi pada tahuntersebut adalah 290.344 ton. Hampir 70%jagung yang ada di Sumatera Barat berasal dariKabupaten Pasaman Barat.Lahan untukpertanian jagung bagi masyarakat PasamanBarat sebagian tidak disediakan secara khusus.Masyarakat menanam jagung dengan diselingitanaman lain, seperti sawit, karet, pepaya ataubeberapa jenis tanaman sayur. Lahan yangdikelilingi oleh perkebunan sawit memilikihama yang merusak pertanian jagung, yaitukera dan babi hutan. Hama ini menyerang padasaat usia jagung masih muda, bahkan padalahan yang cukup luas hama babi hutanmembuat sarang di tengan lahan. Berbagaiupaya dilakukan masyarakat untuk mengatasihama pertanian jagung mereka, antara lainadalah memasang perangkap disekeliling lahandan melakukan pengawasan rutin.Teknikpenanaman juga masih didominasi tekniktradisional.
Pemerintah kabupaten terus memperhatikanmengenai pertanian jagung ini.Berdasarkandata dari antarasumbar.news diperolehinformasi bahwa pemerintah melakukansekolah lapangan pertanian teknologi terpadu(SLPTT) kepada 40 unit kelompok tani yangmewakili setidaknya 1000 Ha lahanjagung.Tujuan kegiatan ini adalah untukmeningkatkan mutu petani jagung dalampengelolaan pertanian, mulai dari pemilihanbibit, pemupukan sampai kepadapemanenan.Hasil pertanian jagung juga masihdalam bentuk barang belum jadi, ini merupakankesempatan investasi untuk pengolahan jagungmenjadi barang siap jadi, misalnya pop corn
dan tepung.Dari segi luas lahan,pengembangannya mengarah kepada target55.724 Ha pada tahun 2015.3. PadiLuas pertanian padi pada tahun 2013 adalah27.856 Ha dengan hasil produksi pada lahantersebut adalah 120.704 Ton. Angkaproduktivitas padi adalah 4,3.Target produksipadi adalah 5 ton per hektar setiap tahun,namun angka ini belum terpenuhi. Sekarangrata-rata hasil pertanian padi per hektarnya tiaptahun adalah 4,8 ton. Pemerintah DinasPertanian Tanaman Pangan Holtikultura danPeternakan menyebutkan bahwa jika angka 5ton per hektar tiap tahun itu tercapai, makaakanterbentuk masyarakat yang tahan pangan(swadaya pangan).
Padi yang ditanam di Kabupaten PasamanBarat beraneka varietas.Salah satunya adalahpadi gogo sigudang, padi yang dapatdikembangkan pada lahan kering.BerdasarkanSK Menteri No.5001/KPTS/Sr.120/2013, padigogo sigudang yang berasal dari Pasaman Baratmendapatkan pengakuan dari menteripertanian.Pertanian padi dari varietas inimerupakan peluang investasi yang cukup besar,mengingat sekarang sedang dilakukanperemajaan tanaman sawit.Jadi lahan tinggalbekas perkebunan sawit dapat digunakan untukpengembangan jagung.
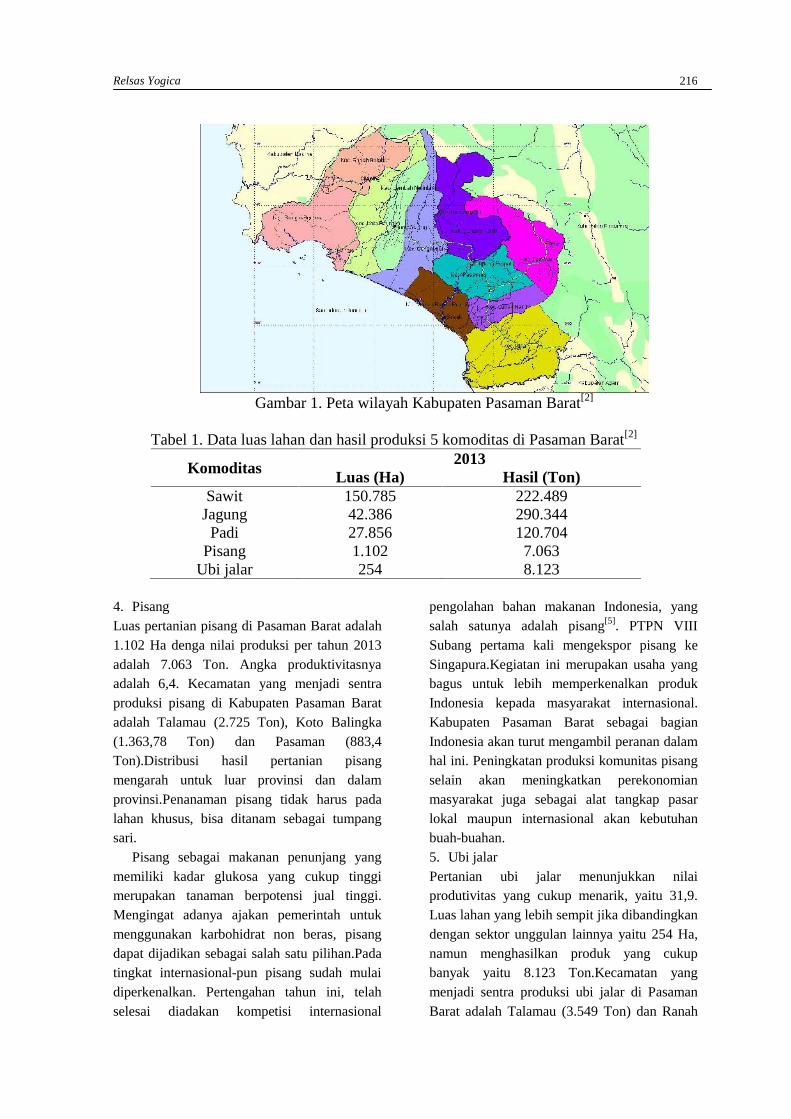
Relsas Yogica 216
Gambar 1. Peta wilayah Kabupaten Pasaman Barat[2]
Tabel 1. Data luas lahan dan hasil produksi 5 komoditas di Pasaman Barat[2]
Komoditas2013
Luas (Ha) Hasil (Ton)Sawit 150.785 222.489
Jagung 42.386 290.344Padi 27.856 120.704
Pisang 1.102 7.063Ubi jalar 254 8.123
4. PisangLuas pertanian pisang di Pasaman Barat adalah1.102 Ha denga nilai produksi per tahun 2013adalah 7.063 Ton. Angka produktivitasnyaadalah 6,4. Kecamatan yang menjadi sentraproduksi pisang di Kabupaten Pasaman Baratadalah Talamau (2.725 Ton), Koto Balingka(1.363,78 Ton) dan Pasaman (883,4Ton).Distribusi hasil pertanian pisangmengarah untuk luar provinsi dan dalamprovinsi.Penanaman pisang tidak harus padalahan khusus, bisa ditanam sebagai tumpangsari.
Pisang sebagai makanan penunjang yangmemiliki kadar glukosa yang cukup tinggimerupakan tanaman berpotensi jual tinggi.Mengingat adanya ajakan pemerintah untukmenggunakan karbohidrat non beras, pisangdapat dijadikan sebagai salah satu pilihan.Padatingkat internasional-pun pisang sudah mulaidiperkenalkan. Pertengahan tahun ini, telahselesai diadakan kompetisi internasional
pengolahan bahan makanan Indonesia, yangsalah satunya adalah pisang[5]. PTPN VIIISubang pertama kali mengekspor pisang keSingapura.Kegiatan ini merupakan usaha yangbagus untuk lebih memperkenalkan produkIndonesia kepada masyarakat internasional.Kabupaten Pasaman Barat sebagai bagianIndonesia akan turut mengambil peranan dalamhal ini. Peningkatan produksi komunitas pisangselain akan meningkatkan perekonomianmasyarakat juga sebagai alat tangkap pasarlokal maupun internasional akan kebutuhanbuah-buahan.5. Ubi jalarPertanian ubi jalar menunjukkan nilaiprodutivitas yang cukup menarik, yaitu 31,9.Luas lahan yang lebih sempit jika dibandingkandengan sektor unggulan lainnya yaitu 254 Ha,namun menghasilkan produk yang cukupbanyak yaitu 8.123 Ton.Kecamatan yangmenjadi sentra produksi ubi jalar di PasamanBarat adalah Talamau (3.549 Ton) dan Ranah

Relsas Yogica 217
Batahan (2.660 Ton).Data yang diperoleh untukluas tanaman ubi jalar ada perbedaan antaradata dari Pemerintah Kabupaten Pasaman Baratdengan Badan Pusat Statistik.BPSmenyebutkan bahwa pada tahun 2013 luastanaman ubi jalar adalah 3.304 Ha.Perbedaanini tidak harus dipermasalahkan karena nilaiproduksi masih cukup besar.
D. Peluang investasiBanyak hal yang harus dipertimbangkan
untuk melaukan investasi pada suatu daerah.Pertimbangan-pertimbangan ini lah yangsedang disiapkan oleh Kabupaten PasamanBarat. Secara tidak langsung dampak yang akanmuncul adalah semakin kuatnya integritasIndonesia dimata dunia. Penelitian danpengembangan berbagai sektor masyarakanakan terus menambah nilai positifperekonomian masyarakat tersebut.
Pengembangan sektor pertanian danperkebunan yang ada di Kabupaten PasamanBarat terus diusahakan, seperti pemberianpelatihan pengelolaan lahan, pemilihan bibittanam sampai kepada bagaimana caramendistribusikan hasil. Sekarang ini investasiyang sedang diusahakan adalah pada sektorpeternakan. Kabupaten Pasaman Baratmembuka peluang investasi kepada Australiauntuk melakukan pembibitan sapi denganjumlah mencapai 400.000 ekor dengan kisarandana mencapai 30 triliun rupiah.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Pasaman barat merupakan kabupaten diSumatera Barat yang terletak dibagian utaraprovinsi.Lahan yang dimiliki memiliki peluangbesar untuk membantu integritas Indonesiamenghadapi pasar bebas ASEAN.Potensi lahanyang ada menunjang kehidupan masyarakatpertanian dan perkebunan.Pengembangannyajuga membutuhkan perhatian khusus oleh
pemerintah, agar arah kemajuan dapat bernilaipositif. Produksi unggulan yang dihasilkan darisektor pertanian dan perkebunan adalah sawit,jagung, padi, pisang dan ubi jalar.
DAFTAR PUSTAKA
[1]Departemen Perdagangan RepublikIndonesia.2014. Menuju Asean EconomicCommunity 2015. Jakarta: DepartemenPerdagangan Republik Indonesia.
[2]Dinas Pemerintah Kabupaten Pasaman Barat.2014. Kabupaten Pasaman Barat(pasamanbarat.com). Diakses tanggal 13Agustus 2014.
[3]Yogatama, Benediktus Krisna. 2014. HadapiMEA 2015, ini sektor unggulan Indonesia(kontan.co.id).Diakses tanggal 12 Agustus2014.
[4]Ramdan, Dadan M. 2014. Industri JamuTradisional Ketar Ketir Hadapi MEA(kontan.co.id). Diakses tanggal 12Agustus 2014.
[5]Antara. 2014. Kabupaten Pasaman Barat(antarasumbar.news). Diakses tanggal 12Agustus 2014.

ISSN 978-602-14989-0-3 BioETI
Karakteristik morfologi polen Daemonorops draco (Willd.)BlumeREVIS ASRA1 SYAMSUARDI2 MANSYURDIN3 DAN JOKO RIDHO WITONO4
1Jurusan Biologi, Fakultas Sains dan Teknologi, Universitas Jambi, Jambi Indonesia Kampus Pinang Masak, JalanJambi-Muara Bulian KM. 15 Mendalo Darat, Jambi 36361email: 2Herbarium
2Universitas Andalas (ANDA) Jurusan Biologi FMIPA Universitas AndalasKampus Limau Manih, Padang Sumatra Barat 25163, Indonesia
3Laboratorium Genetika Jurusan Biologi FMIPA Universitas Andalas Kampus Limau Manih,Padang Sumatra Barat 25163, Indonesia
4Pusat Konservasi Tumbuhan-Kebun Raya Bogor, Lembaga Ilmu Pengetahuan Indonesia (LIPI)Jalan Ir. H. Juanda 13 Bogor 16003, Indonesia
E-mail: [email protected]
ABSTRAKKajian morfologi polen penting dalam taksonomi tumbuhan, karena ciri morfologi polen konsisten untuk menentukanstatus genera dalam suatu famili. Observasi polen secara ultrastruktur dengan menggunakan Scanning Electron
Microscopy (SEM) membantu mendefinisikan karakteristik polen seperti bentuk, ukuran, dan ada atau tidak adanya eksin.Pengambilan sampel perbungaan jantan D. draco untuk pengkoleksian polen dilakukan di Mandiangin,Jambi.Pengamatan polen dengan Scanning Electron Microscope (SEM) dilakukan di Laboratorium SEM, Bidang Zoologi,Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia. Berdasarkan ukuran dari diameter ekuatorial, makapolen D. draco termasuk ke dalam kelompok polen yang berukuran kecil (21 µm). Polen mempunyai apertura danmemiliki lesura, tipe ornamentasi eksin rata. Tipe aperture pada polen D. draco adalah monocolpate. Bentuk aperturemonocolpate dianggap sebagai starting point dari seluruh bentuk lainnya. Tipe aperture ini, dipertimbangkan menjadikarakter primitif. Ukuran polen Daemonorops draco yaitu Panjang Sumbu Polar (PSP) = 26.08 µm , Panjang SumbuEquatorial (PSE) = 21µm dan Polar / Equatorial (P/E) = 1.24µm . Berdasarkan ukuran polen, yang kecil danornamentasi eksin yang rata maka polinasi D. draco melalui angin/anemogami.
Key words: Daemonorops draco, polen, ukuran, ornamentasi , polinasi
Pendahuluan
Daemonorops adalah salah satu genus rotan
yang berasal dari bahasa Greek, yaitu “daemon”(setan) dan “rhops” (semak) ( Beccari, 1911dalam Rustiami dkk., 2004). Hal inidisebabkan oleh banyaknya duri yang terdapatpada tumbuhan ini. Walaupun dikenal dengannama semak setan, namun tumbuhan inimemiliki nilai ekonomi tinggi, terutama untukjenis-jenis Daemonorops yang menghasilkanresin merah, sehingga jenis ini dikenal denganpalem darah naga. Menurut Rustiami et al.(2004), spesies palem darah naga ini endemicuntuk wilayah Asia Tenggara bagian barat.Salah satu spesies darah naga yang ditemukandi Sumatera adalah Daemonorops draco(Willd.) Blume.
Daemonorops draco termasuk ke dalamkelompok tumbuhan berumah dua (dioecious),dimana bunga jantan dan bunga betina terpisahpada individu yang berbeda. Pada tumbuhandioecious sering gagal dalam melakukanperkawinan (polinasi), karena kematanganbunga jantan dan betina yang sering tidak sama.Hambatan ini akan berdampak terhadapkeberhasilan polinasi sehingga dapatmenurunkan produksi buah.
Upaya untuk dapat melakukan polinasibuatan, maka perlu diketahui karakteristikmorfologi polen Daemonorops draco, karenaberdasarkan karakterikstik polen dapatdiketahui bagaimana system polinasi tumbuhantersebut. Disamping itu karakteristik polen jugamerupakan pengetahuan mendasar yang akanmembantu kegiatan pemuliaan (breeding) daribeberapa jenis palem darah naga di masa

Revis Asra, Syamsuardi, Mansyurdin dan Joko Ridho Witono 219
mendatang. Karakteristik polen yang pentingdiketahui adalah bentuk (morfologi) polen,ukuran pollen, ada/tidak apertur, jumlah apertur,ada/tidak laesura, bentuk laesura, bentukornamentasi jumlah polen.
BAHAN DAN METODE
Koleksi SampelSampel bunga jantan D. draco diperoleh dariMandiangin, Kecamatan Pauh, KabupatenSarolangun, Jambi.
Prosedur PenelitianPengamatan polen dengan Scanning Electron
Microscope (SEM) dilakukan di LaboratoriumSEM, Bidang Zoologi, Pusat Penelitian Biologi,Lembaga Ilmu Pengetahuan Indonesia. Untukmengamati morfologi polen dilakukan denganmenggunakan Metode Scanning ElectronMicroscope (SEM), dengan cara kerja sebagaiberikut :
Polen yang telah direndam dalam larutanFAA dicuci dengan caccodylate buffer selama±2 jam pada suhu 4oC. kemudian dilakukanprefiksasi dengan cara memasukkan sampelpolen ke dalam larutan glutaraldehida 2,5%pada suhu 4oC selama ±2 jam, lalu difiksasidengan larutan asam tannic 2% selamasemalam juga pada suhu 4oC. Setelah direndamdengan larutan asam tannic 2% selamasemalam sampel dicuci dengan buffercaccodilat dingin 4 kali selama masing-masing5 menit. Setelah itu dilanjutkan dengandehidrasi dengan cara merendam polen dalamalkohol 50% sebanyak 4 kali selama masing-masing 5 menit, kemudian direndam lagidengan alkohol 70%, 85% dan 95% selamamasing-masing 20 menit dan kemudiandirendam lagi dengan alkohol absolut 2 kaliselama masing-masing 10 menit. Setelahseluruh tahapan dehidrasi selesai sampel polendirendam dalam tert butanol 2 kali selamamasing-masing 10 menit, kemudian dibekukandalam freezer refrigerator sampai beku selamasemalam. Setelah dibekukan semalamkemudian dilakukan pengeringan denganCritical Point Drying sampai kering pada suhu
-50 oC pada tekanan 25 Pa). setelah kering lalusampel dipasang pada holder denganmelekatkan pada selotip dua sisi rekat yangdiatur dengan menjepitkan holder pada penjepitsehingga bagian sampel menghadap ke dalam.Kemudian dilakukan pelapisan dengan emasmurni dengan alat Ion sputtering JEOL IB2selama 15 menit. Setelah selesai holder diambillalu dipasang pada Scanning Electron
Microscope (SEM) JEOL JSM 5310 LV dandiambil gambar polen yang paling baikkemudian dipotret untuk diidentifikasi (Wang,2003, cit Susanti, 2009).
Analisis DataAnalisis data secara deskriptif, meliputipengamatan terhadap bentuk pollen, ukuranpollen, ada/tidak apertur, jumlah apertur,ada/tidak laesura, bentuk laesura dan bentukornamentasi. Penentuan bentuk pollenberdasarkan indeks rasio panjang sumbu polar(PSP) dan panjang sumbu ekuatorial (PSE)Ukuran morfologi pollen diperoleh denganmelakukan pengukuran pada panjang sumbupolar (PSP), panjang sumbu ekuatorial (PSE).Penentuan karakter morfologi pollen inimengacu kepada Erdtman (1963).
HASIL DAN PEMBAHASAN
Hasil pengamatan morfologi polen denganmenggunakan Scanning Electron Microscope
(SEM), diperoleh beberapa karakter morfologipolen. Ukuran morfologi polen dihitungberdasarkan panjang sumbu polar dan sumbuekuatorial dalam skala mikrometer. Penentuanbentuk polen dapat diketahui denganmembandingkan antara panjang sumbu polardan sumbu ekuatorial, hal ini dapat dinyatakandalam bentuk polen menurut Halbritter et al.(2008), Kapp (1960) dan Erdtman (1952). Hasilpengukuran sumbu polar (P) dan sumbuekuatorial (E) serta bentuk polen disajikandalam Tabel 1.

Revis Asra, Syamsuardi, Mansyurdin dan Joko Ridho Witono 220
Tabel 1. Ukuran polen jernang (Daemonorops
draco)
BentukPolen
Rata-rataPSP(µm)
Rata-rataPSE(µm)
RasioP/E(µm)
Ornamentasi
Prolate1,Prolatesperoidal2,Prolatesperoidal3
26.08 21 1.24 Rata
Keterangan : 1Halbritter et al. (2008), 2Kapp (1969), 3Erdtman(1952);PSP: Panjang Sumbu PolarPSE: Panjang Sumbu Equatorial;P/E: Polar / Equatorial.
Polen mempunyai ukuran sumbu polar 20 -30 µm dengan rata-rata 26,08 µm dan sumbuekuatorial 20 - 25 µm dengan rata-rata 21 µm,indeks atau rasio P/E 1,24 µm, bentuk polenprolate (Halbritter et al. 2008), prolatesperoidal (Kapp 1969), prolate speroidal(Erdtman 1952). Berdasarkan ukuran daridiameter ekuatorial, maka polen D. draco (21µm), termasuk ke dalam kelompok polen yangberukuran kecil (10-25 µm). Kisaran ukuranpolen pada family Palmae beragam, dengandiameter ekuatorial berkisar dari 20 µm hingga75 µm (Sawunmi, 1968).
Polen mempunyai apertura dan memilikilesura, tipe ornamentasi eksin rata (Gambar 1)Tipe aperture pada polen D. draco adalahmonocolpate. Menurut Sowunmi (1968)aperture monocolpate merupakan tipe apertureyang paling dominan dalam family Palmae danjuga merupakan karakteristik terhadapkebanyakan monokotiledon. Bentuk aperturemonocolpate dianggap sebagai starting pointdari seluruh bentuk lainnya yang mencapaiperkembangan lanjutan melalui modifikasi daneliminasi (Gambar 2). Tipe aperture inidipertimbangkan menjadi karakter primitif,yang juga ditemukan pada jenis Elaeis
guineensis.Dari hasil pengamatan terlihat polen D.
draco berupa polen tunggal. Hal ini diperkuatoleh Knox (1985) dalam Aprianty danKriswiyanti (2008) yang menyatakan bahwa
sebagian besar polen Angiospermae merupakanpolen yang soliter dan bebas, masing-masingberkembang dari mikrospora tunggal. Padapolen dari Caulokaempferia coenobialis yangmerupakan salah satu jenis dari Zingiberaceaemempunyai struktur penghubung antar polenyang seperti benang (Wang, Zhang, Renner,and Chen, 2005).
Dari semua pengukuran morfologi polenyang telah dilakukan, ukuran polen yangterkecil didapatkan dengan ukuran rata-ratapolar dan equatorial 20 µm. Ukuran polen yangterbesar didapatkan dengan rata-rata ukuranpolar 30 dan rata-rata equatorial 25 µm. Rata-rata rasio P/E tertinggi pada D. draco yaitu1,375 µm. Menurut Erdtman (1952)menyebutkan bentuk, ukuran ataupun tipepolen bisa juga bervariasi menurut tahapkematangannya. Penelitian polen dari beberapaahli terhadap beberapa jenis tumbuhan di Eropamenurut Faegri dan Iversen (1989) dalamAprianty dan Kriswiyanti (2008) menunjukkanadanya variasi ukuran berdasarkan letakgeografisnya. Akan tetapi usaha untukmenghubungkan ukuran polen yang bervariasidalam menentukan adanya faktor lingkunganbelum memberi hasil yang memuaskan.
Ukuran polen, dan ornamentasi eksin eratkaitannya dengan sistem polinasi, dimanabiasanya ukuran polen yang kecil, danornamentasi eksinnya rata polinasinya melaluiangin/anemogami karena polen yang ringandan kecil. Hubungan antara polinator dan polenini memastikan bahwa polen tidak akanterbuang sia-sia apabila dipindahkan dari satubunga ke bunga yang lain. Senada denganGojmerac (1983) proses polinasi dipengaruhioleh temperatur, kelembaban, dan adanyapollinator yang dapat dilakukan oleh seranggaataupun angin. Penyerbukan yang dilakukanoleh angin biasanya memiliki serbuk sari yangsangat banyak, ringan dan kecil, sedangkanpenyerbukan yang dilakukan oleh seranggapada umumnya memiliki warna bunga yangmencolok, serbuk sari relatif berat, besar,lengket, dan kaya protein.
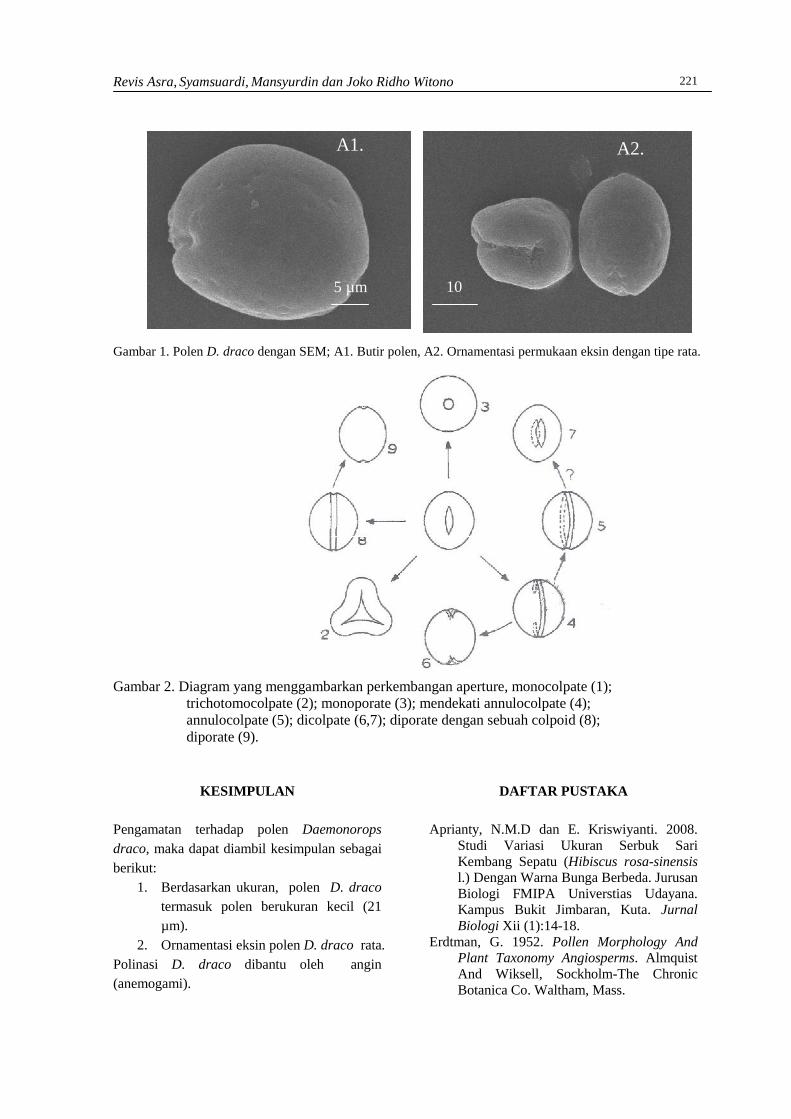
Revis Asra, Syamsuardi, Mansyurdin dan Joko Ridho Witono 221
Gambar 1. Polen D. draco dengan SEM; A1. Butir polen, A2. Ornamentasi permukaan eksin dengan tipe rata.
Gambar 2. Diagram yang menggambarkan perkembangan aperture, monocolpate (1);trichotomocolpate (2); monoporate (3); mendekati annulocolpate (4);annulocolpate (5); dicolpate (6,7); diporate dengan sebuah colpoid (8);diporate (9).
KESIMPULAN
Pengamatan terhadap polen Daemonorops
draco, maka dapat diambil kesimpulan sebagaiberikut:
1. Berdasarkan ukuran, polen D. dracotermasuk polen berukuran kecil (21µm).
2. Ornamentasi eksin polen D. draco rata.Polinasi D. draco dibantu oleh angin(anemogami).
DAFTAR PUSTAKA
Aprianty, N.M.D dan E. Kriswiyanti. 2008.Studi Variasi Ukuran Serbuk SariKembang Sepatu (Hibiscus rosa-sinensisl.) Dengan Warna Bunga Berbeda. JurusanBiologi FMIPA Universtias Udayana.Kampus Bukit Jimbaran, Kuta. JurnalBiologi Xii (1):14-18.
Erdtman, G. 1952. Pollen Morphology AndPlant Taxonomy Angiosperms. AlmquistAnd Wiksell, Sockholm-The ChronicBotanica Co. Waltham, Mass.
A2.A1.
10µm
5 µm

Revis Asra, Syamsuardi, Mansyurdin dan Joko Ridho Witono 222
Halbritter, H, M. Weber, R. Zetter, A. Frosch-Radivo, R. Buchner, M. Hesse. 2008.PalDat-Illustrated Handbook on PollenTerminology. University of Vienna.Austria.
Kapp, R.O. 1969. How To Know Pollen andSpores. WM. C, Brown CompanyPublisher. Dubuoue. Lowa.
Rustiami, H., Setyowatii, F.M. , Kartawinata K.,2004. Taxonomy and uses ofDaemonorops draco (Willd.) Blume.Journal of Tropical Ethnobiology Vol I(2): 65 – 75.
Sowunmi, M. A. 1968. Pollen morphology inthe palmae, with special reference to
trends in aperture development. Rev.Palaeobotan. Palynol. 45-53
Susanti, T. 2009. Klasifikasi Numerik GenusGlobba (Zingiberaceae) Sumatera Barat.Tesis Pascasarjana Biologi. ProgramPasca Sarjana Universitas Andalas Padang.
Wang, Y-Q., D-X Zhang, S. S. Renner, Z-Y.Chen. 2005. Self-Pollination by SlidingPollen in Caulokaempferia coenobialis(Zingiberaceae). Int. J. Plant Sci. 166(5):753-759. 2005.

ISSN 978-602-14989-0-3 BioETI
Kupu-kupu pemakan buah di kawasan Taman Nasional KerinciSeblat (TNKS) Wilayah IV Kabupaten Solok Selatan, SumateraBaratRINI OKTAVIA, DAHELMI DAN HENNY HERWINA
Laboratorium Taksonomi Hewan, Jurusan Biologi, FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163email: [email protected]
ABSTRACTPenelitian mengenai spesies kupu-kupu pemakan buah yang terdapat di Taman Nasional Kerinci Seblat (TNKS) wilayahIV Kabupaten Solok Selatan, Sumatera Barat telah dilakukan pada bulan Februari sampai Juni 2014. Kupu-kupudikoleksi dengan menggunakan perangkap Cylindrical Gauze dengan umpan buah Nenas. Sebanyak 13 spesies kupu-kupu telah didapatkan, yang terdiri dari 3 famili (Amathusiidae, Nymphalidae dan Satyridae), 12 genera dan 33 individu.Spesies yang paling banyak ditemukan adalah dari famili Nymphalidae (7 spesies), diikuti famili Satyridae (4 spesies)dan famili Amathusiidae (2 spesies). Jumlah spesies dan individu lebih banyak ditemukan pada perangkap yang dipasangpada understorey (11 spesies) dibandingkan dengan canopy (2 spesies).
Key words: kupu-kupu, buah, Taman Nasional, Cylindrical gauze, spesies
Pendahuluan
Indonesia memiliki jumlah spesies kupu-kupusekitar 4.000-5.000 spesies (Tsukada andNishiyama, 1982). Jenis kupu-kupu yangditemukan pada wilayah barat Indonesiapenyebarannya berasal dari benua Asia,sedangkan kupu-kupu yang terdapat bagiantimur Indonesia berasal dari benua Australia(Corbet and Pendlebury, 1956). Kupu-kuputermasuk kedalam ordo lepidoptera.Lepidoptera berasal dari bahasa Yunani, yaitulepis yang artinya sisik dan pteron yang artinyasayap (Kunte, 2006). Kupu-kupu mempunyaiciri-ciri memiliki kaki tiga pasang, bersayap,tubuh ditutupi sisik dan mempunyai warnayang menarik (Suharto dan Zulkarnain, 2005).Kupu-kupu tergolong holometabola yaitumemiliki metamorfosis sempurna dengansiklus: telur, larva, pupa dan dewasa (Peggiedan Amir, 2006).
Kupu-kupu merupakan komponen biotikyang mudah dikenal dan mempunyai perananekologi pada suatu ekosistem yaitu sebagaipolinator serta berfungsi sebagai bio-indikatorlingkungan (Tati-Subahar et al, 2007). Kupu-kupu juga bagus digunakan sebagai subjekpengamatan ilmu pengetahuan dan studi ilmiah
(Kunte, 2006). Selain itu kupu-kupu memilikinilai ekonomis karena bentuknya yang indahsehingga banyak diminati dan sering dijadikankoleksi (Suharto dan Zulkarnain, 2005).
Kupu-kupu hanya meletakkan telur ditumbuhan inang yang dimakan oleh larvanya,ini bisa menandai habitat suatu spesies kupu-kupu (Soekardi, 2007). Kebanyakan larvakupu-kupu hanya memakan satu jenistumbuhan saja tetapi ada beberapa spesieskupu-kupu yang larvanya bisa memakan lebihdari satu jenis tumbuhan. Spesies kupu-kupukadang hanya kita temukan pada suatu daerahtertentu saja yang dikenal spesies endemik.Kondisi ini terjadi karena adanya pembatasankarena kondisi lokasi geografis yang terisolasi(Peggie dan Amir, 2006).
Berdasarkan makanan (Feeding guild),kupu-kupu dikelompokkan kedalam tiga guildyaitu pertama kupu-kupu pemakan buah (fruitfeeder) biasanya buah yang disukai adalah buahyang mulai busuk dan sudah busuk. Keduakupu-kupu pemakan madu atau nektar (nectarfeeder) pada bunga yang sedang mekar danyang ketiga kupu-kupu pemakan berbagaimacam-macam (omni feeder) yaitu pemakanbuah, nektar dan lainnya seperti feses burungdan mamalia serta berbagai kotoran dan

Rini Oktavia, Dahelmi dan Henny Herwina 224
bangkai (Kunte, 2006). Kupu-kupu pemakanbuah dapat ditemukan di hutan hujan tropis danhutan tropis di Sabah, Borneo (Tangah et al.,
2004). Di hutan Sulawesi telah dilakukanbeberapa penelitian tentang stratifikasi vertikalkupu-kupu di hutan menggunakan CylindricalGauze dengan umpan buah yang sudahdibusukkan (Schulze, et al., 2001; Fermon etal., 2005).
Taman Nasional Kerinci Seblat (TNKS)merupakan Taman Nasional kedua terluas diIndonesia. Taman Nasional Kerinci Seblat(TNKS) memiliki luas 1.386.000 hamemanjang 350 km barat laut kearah tenggaradengan lebar rata-rata 50 km yang terletak diempat wilayah propinsi yaitu Sumatera Barat,Jambi, Bengkulu dan Sumatera Selatan.Sebagian besar kawasan Taman Nasional inimerupakan rangkaian pegunungan BukitBarisan Selatan di pulau Sumatera bagiantengah. TNKS wilayah IV terletak di PropinsiSumatera Barat. Pentingnya TNKS telah diakuidunia internasional dengan ditetapkannyaTNKS sebagai situs warisan ASEAN(Departemen Kehutanan, 2010). Kepala SeksiPengolahan Taman Nasional Wilayah IV SolokSelatan mengatakan jumlah Polisi Hutan belumideal dengan luas TNKS, sehingga kerusakanhutan bisa saja dilakukan oleh oknum yangtidak bertanggung jawab. Kegiatan ini bisamempegaruhi flora dan fauna yang ada dihutantersebut bahkan bisa punah. Salah satu faunatersebut adalah kupu-kupu.
Penelitian kupu-kupu yang telah dilakukandi TNKS diantaranya adalah Salmah dkk.,
(1999) melaporkan 230 spesies Papilionidae.Selanjutnya Dahelmi dkk., (2009) mendapatkan90 spesies Papilionidae di Muara Imat danGunung Tujuh. Andrianti (2010) di ResortGunung Tujuh mendapatkan 38 spesies dengan6 famili yaitu Acraeidae, Lycaenidae,Nymphalidae, Papilionidae, Pieridae danSatyridae dan Prayuda (2010) di TNKS ResortBukit Reges, Kabupaten Lebong, PropinsiBengkulu mendapatkan 47 spesies dengan 6famili yaitu Danaidae, Lycaenidae,
Nymphalidae, Papilionidae, Pieridae danSatyridae. Namun penelitian kupu-kupupemakan buah belum pernah dilakukan dikawasan TNKS Wilayah IV ini.
BAHAN DAN METODE
Lokasi PenelitianPengambilan sampel dilakukan di TNKSwilayah IV Kabupaten Solok Selatan, SumateraBarat pada tanggal 01-10 Februari 2014.Identifikasi dilakukan di Lab. TaksonomiHewan, Jurusan Biologi, FMIPA, UniversitasAndalas, Padang.
Pengoleksian SampelKupu-kupu dikoleksi dengan menggunakanperangkap Cylindrical Gauze pada dua lokasi(masing-masing 10 perangkap perlokasi).Perangkap dipasang secara vertikal di canopy(20 m dari tanah) dan di understorey (1 m daritanah). Pada tegakan canopy diletakkan 5 buahperangkap dan di tegakan understorey pohonsebanyak 5 buah perangkap. Perangkapdipasang didalam hutan, jarak masing-masingperangkap ±100 m. Pemeriksaan perangkapdilakukan setiap 1 x 24 jam. Sampel yangdikoleksi hanya satu jantan dan satu betina, jikasampel kupu-kupu tersebut belum diketahuijantan atau betinanya, sampel dikoleksi 1sampai 3 invidu setiap spesies, kemudian jikatertangkap spesies yang sama hanya dicatatjumlahnya kemudian kupu-kupu tersebutdilepas kembali.
Gambar 1. Perangkap Cylindrical Gauze yangdigunakan untuk menangkap kupu-kupupemakan buah di Kawasan TNKSWilayahIV Kabupaten Solok Selatan.

Rini Oktavia, Dahelmi dan Henny Herwina 225
IdentifikasiKupu-kupu diidentifikasi dengan menggunakanbuku panduan Corbet and Pendlebury (1956),Tsukada (1982, 1991), Tsukada and Nishiyama(1982; 1985). Jumlah spesies dan jumlahindividu dihitung lalu data ditampilkan dalambentuk tabel dan grafik.
HASIL DAN PEMBAHASAN
Dari penelitian kupu-kupu pemakan buah yangtelah dilakukan pada dua lokasi (Lokasi I danII) di Taman Nasional Kerinci Seblat (TNKS)Wilayah IV Kabupaten Solok Selatan,Sumatera Barat ditemukan kupu-kupusebanyak 13 spesies yang terdiri dari 3 famili(Amathusiidae, Nymphalidae dan Satyridae),12 genera dan 33 individu (Tabel 1.). Kupu-kupu lebih banyak tertangkap pada lokasi satu(9 spesies, 3 famili, 8 genera dan 17 individu)dibandingkan lokasi dua (5 spesies, 3 famili, 5genera dan 16 individu) (Tabel 1.). Perbedaanjumlah spesies, famili, genera dan jumlahindividu yang didapatkan pada dua lokasi inidikarenakan pada lokasi satu hutannya masihsedikit alami dan kerusakan hutan yang terjadibelum terlalu parah. Walaupun disekitar lokasisatu terdapat perladangan warga tapiperladangan dan hutan mempunyai sedikitbatas yang tidak boleh di garap olehmasyarakat sekitar sedangkan pada lokasi duayang hutannya sudah mulai dirusak(penebangan hutan secara liar di sekitar hutanbahkan sudah masuk ke pinggir hutan) danperladangan masyarakat sekitar sudahberbatasan langsung dengan hutan. MenurutDeVries and Walla (2001), kupu-kupu yangdikoleksi di hutan yang tidak dirusak dan hutanyang dirusak memiliki perbedaan padakeragaman spesies, jumlah individu dan warnaspesies.
Kupu-kupu di dalam hutan lebih banyakdijumpai terbang rendah dari pada terbangtinggi sehingga lebih banyak masuk perangkapyang di pasang di understorey dari pada dicanopy. Dari 10 perangkap Cylindrical Gauze
yang dipasang (5 di canopy dan 5 diunderstorey) kupu-kupu lebih banyaktertangkap di understorey dari pada di canopy.Pada undersotrey pohon tertangkap 11 spesiesyang terdiri dari 2 famili, 10 genera dan 31individu sedangkan pada canopy pohondidapatkan 2 spesies yang terdiri dari 1 famili,2 genera dan 2 individu. Menurut Prayuda(2010), kupu-kupu juga banyak tertangkap diunderstorey pohon dari pada di canopy pohondi Taman Nasional Kerinci Seblat Resort BukitReges, Kecamatan Tapus Kabupaten Lebong,Propinsi Bengkulu.
Amathuxidia amythaon dan Zeuxidiaaurelius (Gambar 2. A dan B) merupakan kupu-kupu dari famili Amathusiidae. Menurut Corbetand Pendlebury (1956) famili ini memiliki ciri-ciri sayap bergerigi dan bergelombang. Padabagian ventral sayap belakang terdapat bintikmata (eyespot). Famili Amathusiidae hampirsama bentuknya dengan famili Nymphalidae,perbedaannya terletak pada ukuran sayap,famili ini memiliki sayap lebih kecil dari padaNymphalidae.
Cynitia cocytina, Doleschallia bisaltide,Euthalia adonia, Junonia atlites, Kallimalimborgi, Rhinopalpa polynice dan Tanaeciaaruna (Gambar 2. C, D, E, F, G, H dan I)merupakan famili Nymphalidae. MenurutCorbet and Pendlebury (1956) famili Satyridaememiliki bentuk sayap depan seperti segitigadan sayap belakang memanjang ke depan danmembengkok. Pada umumnya sayap kupu-kupu dari famili Nymphalidae berwarna coklatatau coklat kekuningan. Prayuda (2010) jugamenemukan Doleschallia bisaltide di TamanNasional Kerinci Seblat Resort Bukit RegesKecamatan Tapus Kabupaten Lebong, PropinsiBengkulu.
Elymnias dara, Elymnias nesaea, Melanitisleda dan Mycalesis oroatis (Gambar 2. J, K, Ldan M) merupakan kupu-kupu dari familiSatyridae. Menurut Amir dan Kahono (2003)Ciri dari famili Satyridae yaitu ukuran tubuhkecil sampai sedang, sayap lebar dan agaksedikit membulat. Kupu-kupu ini memiliki
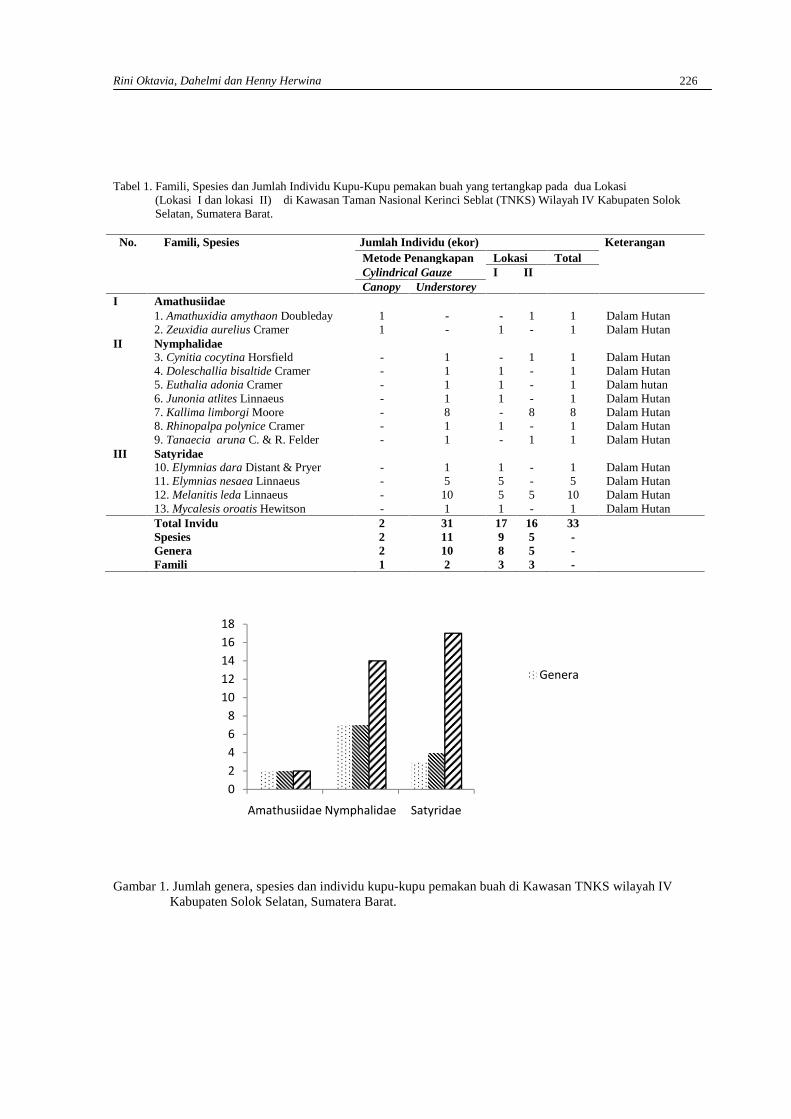
Rini Oktavia, Dahelmi dan Henny Herwina 226
Tabel 1. Famili, Spesies dan Jumlah Individu Kupu-Kupu pemakan buah yang tertangkap pada dua Lokasi(Lokasi I dan lokasi II) di Kawasan Taman Nasional Kerinci Seblat (TNKS) Wilayah IV Kabupaten SolokSelatan, Sumatera Barat.
No. Famili, Spesies Jumlah Individu (ekor) KeteranganMetode Penangkapan Lokasi TotalCylindrical Gauze I IICanopy Understorey
I Amathusiidae1. Amathuxidia amythaon Doubleday2. Zeuxidia aurelius Cramer
11
--
-1
1-
11
Dalam HutanDalam Hutan
II Nymphalidae3. Cynitia cocytina Horsfield4. Doleschallia bisaltide Cramer5. Euthalia adonia Cramer6. Junonia atlites Linnaeus7. Kallima limborgi Moore8. Rhinopalpa polynice Cramer9. Tanaecia aruna C. & R. Felder
-------
1111811
-111-1-
1---8-1
1111811
Dalam HutanDalam HutanDalam hutanDalam HutanDalam HutanDalam HutanDalam Hutan
III Satyridae10. Elymnias dara Distant & Pryer11. Elymnias nesaea Linnaeus12. Melanitis leda Linnaeus13. Mycalesis oroatis Hewitson
----
15
101
1551
--5-
15
101
Dalam HutanDalam HutanDalam HutanDalam Hutan
Total InviduSpesiesGeneraFamili
2221
3111102
17983
16553
33---
Gambar 1. Jumlah genera, spesies dan individu kupu-kupu pemakan buah di Kawasan TNKS wilayah IVKabupaten Solok Selatan, Sumatera Barat.
024681012141618
Amathusiidae Nymphalidae Satyridae
Genera

Rini Oktavia, Dahelmi dan Henny Herwina 227
A B C D
E F G H
I J K L
M
Gambar 2. Spesies Kupu-Kupu pemakan buah yang tertangkap di dua lokasi di Taman Nasional kerinciSeblat (TNKS) Wilayah IV Kabupaten Solok Selatan, Sumatera Barat. Spesies A: Amathuxidiaamythaon, B: Zeuxidia aurelius, C: Cynitia cocytina, D: Doleschallia bisaltide, E: Euthaliaadonia, F: Junonia atlites, G: Kallima limborgi, H: Rhinopalpa polynice, I: Tanaecia aruna,J: Elymnias dara, K: Elymnias nesaea, L: Melanitis leda, M: Mycalesis oroatis.
daya terbang yang lambat dan biasa kitatemukan didekat tanah dan juga di bawahnaungan. Sayap berwarna abu-abu, coklat danmemiliki eyespot pada bagian sub marginal.Memiliki antena yang berbulu dan membesarpada ujungnya (Tsukada, 1982). Melanitis ledajuga ditemukan oleh Prayuda (2010) di TNKSResort Bukit Reges Kecamatan TapusKabupaten Lebong, Propinsi Bengkulu.
Spesies yang paling banyak ditemukanindividunya adalah Melanitis leda (10 individu),diikuti oleh Kallima limborgi (8 individu),Elymnias nesaea (5 individu). Spesies yangpaling sedikit didapatkan jumlah individunyayaitu Amathuxidia amythaon, Zeuxidia aurelius,
Cynitia cocytina, Doleschallia bisaltide,Euthalia adonia, Junonia atlites, Rhinopalpa
polynic, Tanaecia aruna, Elymnias dara danMycalesis oroatis yang masing-masingdidapatkan satu individu. Menurut Shalihah et
al., (2014), tanaman inang larva Melanitis ledaadalah tumbuhan dari famili Arecaceae danPoaceae. Di Kawasan Taman Nasional KerinciSeblat (TNKS) Wilayah IV banyak ditemukantumbuhan dari famili Arecaceae sehinggajumlah individu dari Melanitis leda banyakditemukan.
Famili yang mempunyai spesies terbanyakyaitu famili Nymphalidae yaitu 13 spesies,diikuti oleh famili Satyridae (4 spesies) dan

Rini Oktavia, Dahelmi dan Henny Herwina 228
yang paling sedikit adalah famili Amathusiidae(hanya 2 spesies). Famili Nymphalidaemerupakan salah satu famili terbesar jumlahnyasesudah famili Papilionidae (Smart, 1975).Famili Nymphalidae umumnya mempunyaipenyebaran yang luas seperti hutan, ladangpenduduk. Famili ini menyukai kotoran hewandan buah busuk.
KESIMPULAN
Dari penelitian kupu-kupu pemakan buah yagtelah dilakukan pada dua lokasi di TamanNasional kerinci Seblat (TNKS) Wilayah IVKabupaten Solok Selatan, Sumatera Baratdapat disimpulkan bahwa kupu-kupu yangditemukan yaitu sebanyak 13 spesies yangtergolong kedalam 3 famili, 12 genera dan 33individu. Kupu-kupu tersebut tergolong padaFamili Amathusiidae (2 spesies dan 2 genera),Famili Nymphalidae (7 spesies dan 7 genera)dan Famili Satyridae (4 spesies dan 3 genera).Amathuxidia amythaon, Zeuxidia aurelius,Cynitia cocytina, Euthalia adonia, Junoniaatlites, Kallima limborgi, Rhinopalpa polynice,Tanaecia aruna, Elymnias dara, Elymniasnesaea dan Mycalesis oroatis merupakanspesies yang belum pernah ditemukan diTaman Nasional Kerinci Seblat (TNKS) padapenelitian sebelumnya.
UCAPAN TERIMA KASIHTerima kasih penulis ucapkan kepada KepalaTaman Nasional Kerinci Seblat (TNKS) yangtelah memberikan izin penulis untukmelakukan penelitian.
DAFTAR PUSTAKA
Andrianti, T. 2010. Kupu- Kupu (Butterflies) diKawasan Resort Gunung Tujuh TamanNasional Kerinci Seblat (TNKS)Kabupaten Kerinci Propinsi SumateraBarat. Skripsi Sarjana Biologi. FakultasMatematika dan Ilmu Pengetahuan Alam.Universitas Andalas.
Corbet, A.S. and H.M. Pendlebury. 1956. TheButteflies of Malaya Peninsula. OliverBoyd Edinburg. London.
Dahelmi., Salmah, S. dan H. Herwina. 2009.Diversitas Kupu-Kupu (Butterflies) PadaBeberapa Taman Nasional di Sumatera.Laporan Penelitian Hibah StrategisNasional. Universitas Andalas. Padang.
Departemen Kehutanan. 2010.http://kerinciseblat.dephut.go.id/. Di aksestanggal 25 September 2013.
DeVries, P.J. and T.R.. Walla. 2001. SpeciesDiversity and Community Structure inNeotropical Fruit-Feeding Butterflies.Biological Journal of the Linneas Society,74: 1-15.
Fermon, H., Waltert, M., Vane-Wright, R.I. andM. Muhlenberg. 2005. Forest Use andVertical Stratification in Fruit-FeedingButterflies of Sulawesi, Indonesia:Impacts for Conservation. Biodiversityand Conservation, 14: 333–350.
Fleming, W.A. 1991. Butterflies of WestMalaysia and Singapore. Secon Edition.Vinlin Press Sdn. Bhd. Sri Petaling. KualaLumpur.
Kunte, K. 2006. Butterflies of Peninsular India.Indian Academy of Sciences. UniversitiesPress. India.
Peggie, D. dan M. Amir. 2006. Practical Guideto the Butterflies of Bogor Botanic Garden(Panduan Praktis Kupu-kupu di KebunRaya Bogor). Pusat Penelitian Biologi,LIPI Bogor dan Nagao NaturalEnvironment Foundation Shitaya, Japan.
Prayuda, D. 2010. Jenis-Jenis Kupu-Kupu(Butterflies) Di Taman Nasional KerinciSeblat Resort Bukit Reges KecamatanTapus Kabupaten Lebong PropinsiBengkulu. Skripsi Sarjana Biologi.Fakultas Matematika dan IlmuPengetahuan Alam. Universitas Bengkulu.
Salmah, S., Abbas, I. dan Dahelmi. 1999.Keanekaragaman Kupu-kupu (Butterflies)dan Tanaman Pakan dari Beberapa JenisFamili Papilionidae di Taman NasionalKerinci Seblat. Laporan Penelitian BBI.Universitas Andalas.
Shalihah, A., Pamula, G., Cindy, R., Rizkawati,V. dan Z.I. Anwar. 2014. Kupu-Kupu diKampus Universitas Padjajaran Jatinangor.https://www.cbd.int/undb/countries/id/undb-id-butterflies.book.pdf. DepartemenKeilmuan Divisi Entomologi. HimpunanMahasiswa Biologi Universitas Padjajaran.

Rini Oktavia, Dahelmi dan Henny Herwina 229
Jatinangor. Di akses tanggal 23 September2014.
Schulze, C.H., Linsenmair, K.E. and K. Fiedler.2001. Understory versus canopy: Patternsof Vertical Stratification and Diversityamong Lepidoptera in a Bornean RainForest. Plant Ecology, 153: 133-152.
Smart, P. 1975. The Illustrated Encyclopedia ofthe Butterflies World. Salamander BooksLtd. London.
Soekardi, H. 2007. Kupu-Kupu di KampusUNILA. Press. Lampung. UniversitasLampung.
Suharto, W. dan R. Zulkarnain. 2005. A SurveyOf the Butterflies. (Rhopalocera:Lepidopetra) In Ireng-Ireng Forest OfBromo Tengger Semeru National Park.Jurnal Ilmu Dasar, 6: 62-65.
Tangah, J., Hill, J.K., Hamer, K.C. and M.M.Dawood. 2004. Vertikal Distributions ofFruit-Feeding Butterflies in Sabah, Borneo.Sepilok Bulletin, 1: 17-27.
Tati-Subahar, S.S., Amasya, A.F. dan D. N.Choesin. 2007. Butterfly (Lepidoptera:
Rhopalocera) Distribution Along AnAltitudinal Gradient On MountTangkuban Perahu, West Java, Indonesia.The Raffles Bulletin Of Zoology 2007, 55(1): 175-178.
Tsukada, E. 1982. Butterflies of the South EastAsian Vol. III. Satyrinae, Libythiidae.Plapac. Ltd. Tokyo. Japan.
Tsukada, E. 1991. Butterflies of the South EastAsian Vol. V. Nymphalidae II. Plapac. Ltd.Tokyo. Japan.
Tsukada, E. and Y. Nishiyama. 1982.Butterflies of the South East Asian IslandVol I. Papilionidae. Plapac. Ltd. Tokyo.Japan.
Tsukada, E. and Y. Nishiyama. 1985.Butterflies of the South East Asian IslandVol IV. Nymphalidae 1. Plapac. Ltd.Tokyo. Japan.

ISSN 978-602-14989-0-3 BioETI
Jenis-jenis Gastropoda (Moluska) pada ekosistem Lamun dipantai Nirwana Padang, Sumatera BaratROFIZA YOLANDA
Program Studi Pendidikan Biologi FKIP Universitas Pasir Pengaraian, RiauE-mail: [email protected]
ABSTRAKTelah dilakukan penelitian mengenai jenis-jenis gastropoda yang terdapat pada ekosistem lamun di pantai Nirwana,Padang, Sumatera Barat pada bulan Januari hingga April 2014. Penelitian ini dilaksanakan dengan metoda survei pada 5stasiun dengan menggunakan line transect yang ditarik dari bibir pantai mengarah laut sepanjang 100 m. Sampeldikoleksi dengan menggunakan kerangka kuadrat 1 x 1 meter. Didapatkan sebanyak 15 spesies gastropoda, yaituCerithium sp.,Cypraea annulus, C. arabica, C. interupta, C. lynx,C. tigris,Monodonta labio, Morula fusca, Morulasp.,Polinices tumidus, Tectus pyramis, Trochus maculatus, T. niloticus, Turbo argyrostomus,dan T. cinereus. Spesiesyang mendominasi adalah C. arabica. Sedikitnya jumlah spesies gastropoda yang didapatkan pada ekosistem inidisebabkan karena pemanfaatan organisme ini sebagai sumber makanan, akibat pencemaran dan rusaknya ekosistemlamun akibat kapal-pakal nelayan yang berlabuh pada ekosistem ini.
Key words: Gastropoda, Ekosistem Lamun, Pantai Nirwana
Pendahuluan
Sebagian besar (70%) wilayah duniamerupakan lautan. Namun, hanya sebagiankecil wilayah lautini yang produktif yaituwilayah laut dangkal, yang memilikikeanekaragaman hayati yang tinggisertabeberapa ekosistem yang sangat penting,sepertimangrove, estuaria, terumbu karang danpadang lamun (Hendriks, 2005).
Lamun (seagrass) merupakan salah satutumbuhan berbunga (Angiospermae) yanghidup di wilayah perairan laut dan perairanestuaria (Björk, Short, Mcleod, dan Beer,2006)dengan tutupan area yang cukup luassehingga sering juga disebut dengan padanglamun(Green dan Short, 2003) yang berjumlahsebanyak 60 spesies di seluruh dunia (Björk etal., 2006). Di Indonesia, ditemukan sebanyak13 spesies (Green dan Short, 2003; Kuo, 2007;Azkab, 2010) dengan luas tutupan kurang lebih30.000 km2dari Pulau Weh di Aceh hinggaMerauke di Papua.Walaupun jumlah spesieslamun sedikit, namun ekosistem ini memilikibentuk fisik yang lengkap dan produktifitasyang tinggi yang memungkinkan lamun dapatmenghasilkan biomassa yang tinggi dan
beranekararagam spesies lain yang berasosiasidengan lamun(Green dan Short, 2003).
Padang lamun sangat berperan dalammenstabilisasi sedimen, mencegah erosi danmenyaring nutrien-nutrien yang masuk kedalam air, sehingga menjaga perairan pantaitetap bersih (Björk et al., 2006). Tumbuhan inijuga berperan sebagai sumber makanan bagiduyung (Dugong dugon), penyu hijau(Chelonia mydas), ikan-ikan, burung laut daninvertebrata lainnya. (Menez, Phillips danCalumpong, 1983; Green dan Short, 2003;Björk et al., 2006).Moluska merupakan salahsatu hewan inver-tebrata yang paling banyakjumlahnya di lautan (Brusca dan Brusca, 2003)dan di ekosistem ini, hewan moluska yangpaling sering ditemukan adalah Gastropoda(Kusnadi, Triandiza dan Hernawan, 2008).
Gastropoda memanfaatkan lamun sebagaitempat perlindungan, tempat tinggal dan jugaberkembangbiak serta mendekomposisi serasahlamun yang jatuh ke lantai laut (Arbi, 2011).Dengan adanya ekosistem ini, tentunya akanmemberikan keanekaragaman pula padagastropoda yang terdapat pada ekosistem ini.

Rofiza Yolanda 230
Di pantai Nirwana Padang, Sumatera Baratmasih dijumpai ekosistem lamun yang terdiriatas 2 spesies, yaitu Thalassia hemprichii danEnhalus acoroides (Purnama, 2011). Hasilsurvei di lokasi ini masih ditemukan beberapajenis dari gastropoda yang terdapat padaekosistem ini. Akan tetapi, masih banyakditemukan cangkang-cangkang gastropoda yangterdapat di sekitar lokasi ini yang merupakansisa dari peman-faatannya sebagai sumbermakanan bagi masyarakat sekitar. Ditambah lagidengan berlabuhnya kapal-kapal nelayan di ataspadang lamun yang mengakibatkan rusaknyaekosistem ini. Hal ini tentunya mempengaruhikeanekaragaman dari gastropoda yang beradapada ekosistem lamun tersebut. Sampai saat inibelum ada data yang melaporkan mengenaigastropoda pada ekosistem lamun di pantaiNirwana ini, maka dilakukanlah penelitian iniyang bertujuan untuk mengetahui jenis-jenisgastropoda yang terdapat pada ekosistem lamundi pantai Nirwana Padang, Sumatera Barat.
BAHAN DAN METODE
Penelitian ini telah dilakukan pada bulan Januarihingga April 2014 pada ekosistem lamun dipantai Nirwana Padang, Sumatera Barat dandilanjutkan di laboratorium Pendidikan BiologiUniversitas Pasir Pengaraian. Beberapaperalatan yang digunakan selama penelitian iniadalah masker, snorkel, fins, coral boot, meteran,kerangka kuadrat 1 x 1 m, plastik, botol koleksi,label, botol spesimen, kamera digital, GPS danalat tulis. Sedangkan bahan yang akandigunakan adalah alkohol 70%.Penelitian ini dilaksanakan dengan metodasurvei dengan teknik pencuplikan sampelmenggunakan line transect yang ditarik daribibir pantai mengarah laut sepanjang 100 mpada 5 stasiun. Kemudian sampel dicuplikdengan cara melemparkan kerangka kuadratsepanjang transek secara acak. Gastropoda yangdicuplik adalah gastropoda yang hidup (bukancangkang kosong). Semua gastropoda yangberada di dalam kerangka kuadrat kemudian
dikoleksi dan dimasukkan ke dalam botolkoleksi yang berisi alkohol 70% dan dibawa kelaboratorium untuk diidentifikasi lebih lanjut.Selanjutnya sampel akan diidentifikasi denganmenggunakan acuan Carpenter dan Niem (1998),Abbott dan Dance (2000) serta Dharma,Schwabe dan Schrödl (2005) dan difotomenggunakan kamera digital dan disimpan didalam botol spesimen.
HASIL DAN PEMBAHASAN
Dari penelitian yang telah dilaksanakandidapatkan gastropoda dari 7 famili, 15 jenis danberjumlah sebanyak 177 individu. Spesies yangpaling banyak ditemukan adalah C. arabica (46individu), diikuti Cerithium sp. (24 individu), C.annulus(20 individu), C. interrupta(19 individu),Turbo cinereus(13 individu), Morula fusca(11individu), Monodonta labio(8 individu),Trochus niloticus(8 individu), Turboargyrostomus(8 individu), Morula sp. (6individu), Polinices tumidus(5 individu), C.lynx(2 individu), C. tigris(1 individu) dan Tectus
pyramis(1 individu) dan secara lengkap dapatdilihat pada Tabel 1.
Cypraeidae merupakan salah satu kelompokgastropoda yang memiliki penyebaran yangpaling banyak pada ekosistem lamun, karenaorganisme ini hidup melekat pada substrat,kebanyakan warna cangkang gelap, memilikiaperture yang kecil dan sempit dan aktif padamalam hari (Carpenter dan Niem, 1998; Arbi,2012).
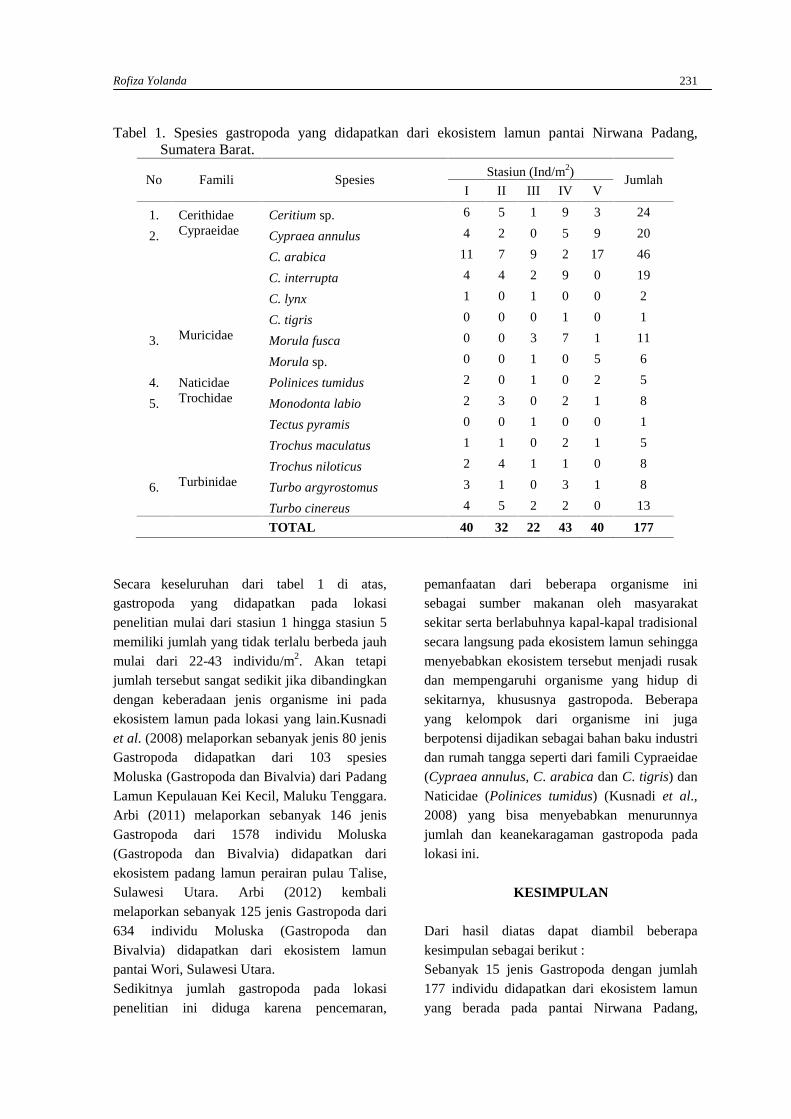
Rofiza Yolanda 231
Tabel 1. Spesies gastropoda yang didapatkan dari ekosistem lamun pantai Nirwana Padang,Sumatera Barat.
No Famili SpesiesStasiun (Ind/m2)
JumlahI II III IV V
1. Cerithidae Ceritium sp. 6 5 1 9 3 24
2. Cypraeidae Cypraea annulus 4 2 0 5 9 20
C. arabica 11 7 9 2 17 46
C. interrupta 4 4 2 9 0 19
C. lynx 1 0 1 0 0 2
C. tigris 0 0 0 1 0 1
3. Muricidae Morula fusca 0 0 3 7 1 11
Morula sp. 0 0 1 0 5 6
4. Naticidae Polinices tumidus 2 0 1 0 2 5
5. Trochidae Monodonta labio 2 3 0 2 1 8
Tectus pyramis 0 0 1 0 0 1
Trochus maculatus 1 1 0 2 1 5
Trochus niloticus 2 4 1 1 0 8
6. Turbinidae Turbo argyrostomus 3 1 0 3 1 8
Turbo cinereus 4 5 2 2 0 13
TOTAL 40 32 22 43 40 177
Secara keseluruhan dari tabel 1 di atas,gastropoda yang didapatkan pada lokasipenelitian mulai dari stasiun 1 hingga stasiun 5memiliki jumlah yang tidak terlalu berbeda jauhmulai dari 22-43 individu/m2. Akan tetapijumlah tersebut sangat sedikit jika dibandingkandengan keberadaan jenis organisme ini padaekosistem lamun pada lokasi yang lain.Kusnadiet al. (2008) melaporkan sebanyak jenis 80 jenisGastropoda didapatkan dari 103 spesiesMoluska (Gastropoda dan Bivalvia) dari PadangLamun Kepulauan Kei Kecil, Maluku Tenggara.Arbi (2011) melaporkan sebanyak 146 jenisGastropoda dari 1578 individu Moluska(Gastropoda dan Bivalvia) didapatkan dariekosistem padang lamun perairan pulau Talise,Sulawesi Utara. Arbi (2012) kembalimelaporkan sebanyak 125 jenis Gastropoda dari634 individu Moluska (Gastropoda danBivalvia) didapatkan dari ekosistem lamunpantai Wori, Sulawesi Utara.Sedikitnya jumlah gastropoda pada lokasipenelitian ini diduga karena pencemaran,
pemanfaatan dari beberapa organisme inisebagai sumber makanan oleh masyarakatsekitar serta berlabuhnya kapal-kapal tradisionalsecara langsung pada ekosistem lamun sehinggamenyebabkan ekosistem tersebut menjadi rusakdan mempengaruhi organisme yang hidup disekitarnya, khususnya gastropoda. Beberapayang kelompok dari organisme ini jugaberpotensi dijadikan sebagai bahan baku industridan rumah tangga seperti dari famili Cypraeidae(Cypraea annulus, C. arabica dan C. tigris) danNaticidae (Polinices tumidus) (Kusnadi et al.,2008) yang bisa menyebabkan menurunnyajumlah dan keanekaragaman gastropoda padalokasi ini.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Sebanyak 15 jenis Gastropoda dengan jumlah177 individu didapatkan dari ekosistem lamunyang berada pada pantai Nirwana Padang,

Rofiza Yolanda 232
Sumatera Barat. Secara umum Gastropoda yangberada didapatkan lokasi ini dibandingkandengan lokasi lain memiliki jumlah yang rendah.Hal ini diduga akibat pengaruh masyarakatsekitar yang memanfaatkan organisme ini danberlabuhnya kapal-kapal tradisional padaekosisitem lamun yang menyebabkan ekosistemtersebut menjadi rusak dan mempengaruhiorganisme yang hidup di sekitarnya, khususnyagastropoda.
Ucapan Terima KasihPenulis mengucapkan terima kasih kepada Dr.Jabang Nurdin, M.Si yang telah memberikanbantuan dan arahan kepada penulis. Kemudianpenulis juga mengucapkan terima kasih kepadaRektor Universitas Pasir Pengaraian.
DAFTAR PUSTAKA
Abbott, R.T dan S.P. Dance. 2000. Compendiumof Seashells. 8th Printing. OdysseyPublishing. China.
Arbi, U.Y. 2011. Struktur Komunitas MoluskaDi Padang Lamun Perairan Pulau Talise,Sulawesi Utara. Oseanologi dan LimnologiIndonesia 37(1): 71-89.
Arbi, U.Y. 2012. Komunitas Moluska DiPadang Lamun Pantai Wori, SulawesiUtara. Jurnal Bumi Lestari 12(1): 55-65.
Azkab, H. 2010. Panduan Penelitian untuk
Lamun. Pusat Penelitian Oseanografi.Lembaga Ilmu Pengetahuan Indonesia.Jakarta.
Björk, M., F. Short, E. Mcleod dan S.Beer .2006. Managing Seagrasses forResilience to Climate Change. IUCN,Gland, Switzerland. 56pp.
Brusca, R.C dan G.J. Brusca. 2003.Invertebrates. 2nd Edition. SinauerAssociates, Inc. Sunderland. USA.
Carpenter, K.Edan V.H. Niem. 1998. FAOSpecies Identification Guide for FisheryPurposes. The Living Marine Resources of
the Western Central Pacific Volume 1
Seaweeds, Corals, Bivalves andGastropods. Food and AgricultureOrganization of the United Nations. Roma.
Dharma, B., E. Schwabe dan M. Schrödl. 2005.Recent and Fossil Indonesian Shells.University of California. United State ofAmerica.
Green, E.P dan F. Short. 2003. World Atlas of
Seagrasses. Prepared by UNEP WorldConservation Monitoring Centre.University California Press, Berkeley. USA.
Hendriks, D. 2005. Jenis-jenis Moluska KelasGastropoda di Tanjung Bunaken Siau,Pulau Gunatin, Pulau-pulau Sangihe,Sulawesi Utara. Warta OseanografiXIX(4): 17-20.
Kuo, J. 2007. New Monoceious Seagrass ofHalophila sulawesii (Hydrocharitaceae)from Indonesia. Aquatic Botany 2: 171-175.
Kusnadi, A., T. Triandiza dan U.E. Hernawan.Inventariasi Jenis dan Potensi MoluskaPadang Lamun di Kepulauan Kei Kecil,Maluku Tenggara. Biodiversitas 9(1): 30-34.
Menez, E.G., R.C Phillips dan H.P Calumpong.1983. Seagrasses from Philippines.Smithsonian Institution Press. City ofWashington. USA.
Purnama, A.A. 2011. Pemetaan dan KajianBeberapa Aspek Ekologi KomunitasLamun di Perairan Pantai Karang TirtaPadang. Tesis. Program Studi BiologiProgram Pascasasrjana Universitas Andalas.Padang.

ISSN 978-602-14989-0-3 BioETI
Jenis Semut (Hymenoptera: Formicidae) di perkebunan PisangAir Dingin, Lubuk Minturun, Sumatera Barat
SERLI AFRI SUSANTI, HENNY HERWINA*DAN DAHELMI
Labor Riset Taksonomi Hewan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail *: [email protected]
ABSTRAKPenelitian tentang jenis semut (Hymenoptera: Formicidae) di Perkebunan Pisang Air Dingin, Lubuk Minturun, SumateraBarat dikoleksi dengan menggunakan metode “Quadra protocol" (free collection, honey bait, soil core sampling dan leaf
litter sampling) pada tanggal 15 Maret 2014 pada transek sepanjang 180 m yang dibagi menjadi 3 subtransek. Ditemukan20 jenis semut yang tergolong ke dalam empat subfamili, 12 tribe, 14 genera dan 901 individu. Subfamili Formicinaeditemukan dengan jumlah yang paling banyak (sembilan jenis dan lima genera) diikuti oleh Myrmicinae (6 jenis dan 4genera), Dolichoderinae (tiga jenis dan 3 genera) dan Ponerinae (dua jenis dan dua genera). Genus yang paling banyakditemukan yaitu Pheidole, Camponotus dan Polyrhachis (masing-masing tiga jenis). Jenis yang paling banyak ditemukanindividunya adalah Tapinoma melanocephalum (367 individu). Anoplolepis gracilipes dan Oecophylla smaragdina
adalah jenis semut yang paling sering ditemukan (masing-masing 12%) diikuti oleh Iridomyrmex anceps danOdontoponera denticulata (10%). Indeks Diversitas Shannon-Wiener semut di perkebunan pisang Air Dingin adalah 1,90.
Key words: Semut, jenis, pisang, perkebunan, Air Dingin
Pendahuluan
Indonesia merupakan salah satu negara yangmemiliki keanekaragaman hayati sangat tinggi.Hal ini disebabkan karena Indonesia terletak dikawasan tropik yang mempunyai iklim yangstabil dan secara geografi adalah negarakepulauan yang terletak diantara dua benuayaitu Asia dan Australia (Primack, Supriatna,Indrawan, dan Kramadibrata, 1988). Hal inibisa dibuktikan dengan banyaknya jenisorganisme yang ditemukan di Indonesia dengankarakter yang khas, salah satunya adalahserangga (Bestia, 2011). Serangga yang palingtinggi tingkat kepadatannya dibandingkan jenisserangga lain adalah semut. Semut merupakankelompok serangga pada daerah terestrial yangbersifat eusosial yang ditandai dengan adanyakerjasama diantara anggota koloni dalammemelihara serangga pradewasa, adanyasistem-sistem kasta dan generasi yang tumpangtindih (Wilson, 1979; Borror et al., 1992).
Menurut jenisnya semut dibagi menjadi ratusemut, semut pejantan, semut prajurit dansemut pekerja. Tubuh semut terbagi atas tiga
bagian yaitu kepala (head), mesosoma (thorak),dan metasoma (abdomen). Morfologi semutsangat jelas dibandingkan dengan serangga lainyang memiliki antena dengan tipe genikulat(berbentuk siku, dengan ruas pertama panjangdan ruas-ruas berikutnya kecil membengkokpada satu sudut dengan yang pertama). Segmenkedua dari abdomen semut mengalamipengecilan atau penggentingan yang berbentuksub-sylindricalform, nodiform, squamiformatau sessile, bagian ini disebut dengan petiole(Mohamed, 2003; Bolton, 1994). Pada sebagianjenis semut memiliki dua buah petiole. Segmenpertama tetap dinamakan petiole, sedangkansegmen yang kedua disebut dengan post petiole(Bolton, 1994; Borror et al., 1992).
Indonesia merupakan salah satu sentraprimer keragaman pisang, baik pisang segar,olahan dan pisang liar. Lebih dari 200 jenispisang terdapat di Indonesia. Luas panen danproduksi pisang selalu menempati posisipertama. Pada tahun 2002 produksinyamencapai 4.384.384 ton dengan nilai ekonomisebesar Rp 6,5 triliun. Produksi tersebutsebagian besar dipanen dari pertanaman kebun

Serli Afri Susanti, Henny Herwina*dan Dahelmi 234
rakyat seluas 269.000 ha. Pisang banyakmengandung vitamin dan mineral esensial yangsangat bermanfaat bagi kesehatan. Bahkan dibeberapa daerah di Papua pisang merupakansubsitusi makanan pokok, seperti di beberapanegara di Afrika (Depertemen Pertananian,2007). Namun sampai saat ini intensitasserangan penyakit seperti penyakit layu ataupenyakit darah dan Banana Bunchy Top-Virus(BBTV) mengancam dan belum ditemukanmetode yang efektif sehingga menurunkanproduksi pisang di Indonesia.
Penelitian tentang jenis-jenis semut dipisang masih terbatas sebelumnya Herwina,Nasir, Jumjunidang dan Yaherwandi (2013)dimana ditemukan 24 jenis semut padatanaman pisang dengan gejala BBTV danselanjutnya Herwina, Nasir, Mairawita danErniwati (2014) tentang jenis-jenis semutpermukaan pada dua perkebunan pisang diSumatera Barat (2014) ditemukan 33 jenis yangdiantaranya berperan sebagai predator, seedharvester dan generalized forager. Informasitentang semut pada perkebunan pisang di AirDingin, Lubuk Minturun, Sumatera Barat,belum tersedia. Penelitian ini bertujuan untukMengetahui Jenis-jenis Semut (Hymenoptera:Formicidae) yang terdapat di Perkebunanpisang Air Dingin, Lubuk Minturun, SumateraBarat.
BAHAN DAN METODE
Tanah Penelitian ini dilakukan pada bulanMaret sampai Juni 2014 di perkebunan pisangAir Dingin, Lubuk Minturun, Sumatera Barat.Pada lokasi dibuat satu transek sepanjang 180m yang dibagi menjadi tiga subtransek, masing-masing subtransek berjarak 60 m denganmenggunakan metode survei dan pengambilansampel dilakukan dengan teknik“quadraprotocol” (Hashimoto, Yamane and Mohamed,2001) dengan menggunakan kombinasi empatmetode yaitu metode free collection (koleksibebas dengan tangan), leaf litter sampling(pengumpulan dan penyaringan serasah), honeybait (umpan madu), soil core sampling
(pengumpulan dan penyaringan tanah) padalevel permukaan tanah (ground level) danpengambilan sampel hanya dilakukan padapagi-siang hari.
Alat-alat yang digunakan dalam penelitianini adalah pinset, vial/botol koleksi, sekop,saringan dan wadah datar berwarna putih,kapas, kertas A4, kertas label, alat tulis, kuaskecil, mikroskop binokuler, plastik ukuran 1dan 2 kg, karet gelang, kamera, GPS, jarum,sterofoam, kertas lancip (card point), lem,kotak spesimen sedangkan bahan yangdibutuhkan adalah umpan madu, alkohol 96 %dan kapur barus.Koleksi Semut di Lapangan
Pengambilan sampel semut dilakukan padalevel permukaan (ground level) tanah padaperkebunan pisang dengan membuat transeksepanjang 180 m yang dibagi menjadi tigasubtransek, masing-masing subtransek berjarak60 m. Pada setiap subtransek semut dikoleksidengan metoda quadra protocol tersebut,meliputi sebagai berikut:
Metode free collection (koleksi bebasdengan tangan) dengan pengambilan sampeldilakukan di sepanjang subtransek untukmencari semut sebanyak mungkin terutamadicari pada tempat-tempat semut bersarang dantempat yang disukai semut antara lain padakayu-kayu lapuk atau busuk, bagian bawahpohon-pohon, ranting pohon perdu, dibawahbatu dilakukan selama 30 menit. Semut yangditemukan dikoleksi kedalam botol koleksiyang telah berisi alkohol.
Metode leaf litter sampling (pengumpulandan penyaringan serasah) dilakukan disepanjang subtransek dengan mengumpulkansarasah lalu disaring di bawah dan ditampungdengan wadah, semut yang ditemukandikoleksi, dilakukan selama 30 menit padamasing-masing subtransek.
Metode honey bait (umpan madu), disetiapsubtransek ditempatkan 15 umpan dengan jarakantar umpan 4 m, madu diteteskan pada kertasukuran A4 (yang telah dibagi menjadi empatbagian) yang diletakkan didasar hutan, semut

Serli Afri Susanti, Henny Herwina*dan Dahelmi 235
yang ditemukan pada umpan dikoleksi kedalambotol koleksi yang telah berisi alkohol.
Metode soil core sampling (pengumpulandan penyaringan tanah) pada level permukaantanah (ground level) yang diambil lima titiksampel tanah dengan ukuran 20 x 20 x 15 cmyang berjarak ± 50 cm dari umpan madu, jadisatu sampel tanah diambil pada tiga titik umpanmadu, tanah diambil menggunakan sekop laludisaring dengan ayakan tangan yangdibawahnya di tampung oleh wadah berwarnaputih datar, semut yang ditemukan danberjatuhan ke wadah di ambil dan dimasukkanke botol koleksi yang telah berisi alkohol.Di Laboratorium
Semut yang didapatkan di bawa kelaboratorium untuk disortir, dipisahkan sampaimorfospesies dan di pinning lalu identifikasimenggunakan buku acuan Bolton (1994),Hashimoto (2003), Bolton (2012), spesimenyang ada di laboratorium dan bantuan ahli Prof.Seiki Yamane dari Kagoshima University.Analisa DataData ditampilkan dalam bentuk tabel dandianalisis untuk menghitung indeks diversitasmenggunakan rumus sebagai berikut:
Rumus indeks diversitas Shannon-WienerH = −(Magurran,2004)
HASIL DAN PEMBAHASAN
Telah ditemukan total sebanyak 20 jenis semutyang tergolong ke dalam empat subfamili, 12tribe, 14 genera, dan 901 individu (Tabel 1).Subfamili Formicinae memiliki jenis yangpaling banyak yaitu 9 jenis dan 5 genera diikutioleh subfamili Myrmicinae (6 jenis dan 4genera), Dolichoderinae (3 jenis dan 3 genera)dan subfamili paling sedikit adalah Ponerinaeyang terdiri dari 2 jenis dan 2 genera. Genusyang paling banyak ditemukan pada penelitianini adalah Camponotus dan Polyrhachis darisubfamili Formicinae serta Pheidole dari
subfamili Myrmicinae dan dengan masing-masing 3 jenis. Subfamili Formicinaemerupakan subfamili kedua terbesar setelahMyrmicinae. Formicinae dapat ditemukan diseluruh dunia, sebagian besar dari merekaumumnya pemakan bangkai (scavenger),mencari makan di tanah atau di vegetasi danumumnya aktif bergerak cepat (Suriyapong,2003). Genus yang paling sedikit adalahDolichoderus, Iridomyrmex, Tapinoma,
Anoplolepis, Plagiolepis, Oecophylla,Crematogaster, Monomorium, Myrmicaria,
Hypoponera, Odontoponera yang masing-masingnya hanya ditemukan satu jenis.
Jenis yang paling banyak ditemukanindividunya adalah Tapinoma melanocephalumyaitu sebanyak 367 individu. Jenis iniditemukan dengan menggunakan metode free
collection, honey bait dan soil core sampling.Genus ini umumnya generalized foragers,scavenger dan penyuka gula sehingga jumlahindividu ini banyak didapatkan di metodehoney bait. Genus ini biasanya bersarang ditanah, bebatuan, kayu dan batang pohon. Semutini merupakan salah satu semut hama yangtersebar luas di kawasan tropis oleh aktivitasmanusia (Shattuck, 2000). Dari keempatmetode pengambilan sampel, metode yangefektif untuk mengoleksi Tapinomamelanocephalum adalah menggunakan metodehoney bait. Namun melihat dari frekuensinya(Tabel 1), jenis yang sering ditemukan adalahAnoplolepis gracilipes dan Oecophylla
smaragdina (12%) dan diikuti Iridomyrmexanceps dan Odontoponera denticulata (10%).Mungkin ini dikarenakan disetiap subtranseksemut ini sering ditemukan, walaupunindividunya tidak sebanyak Tapinoma tetapijenis ini disetiap metode dapat ditemukandibandingkan dengan Tapinoma yangindividunya banyak tetapi hanya denganmetode honey bait karena semut ini penyukagula sehingga lebih banyak ditemukan denganmetode tersebut dan ada yang tidak ditemukandengan metode lainnya seperti leaf litter
sampling.
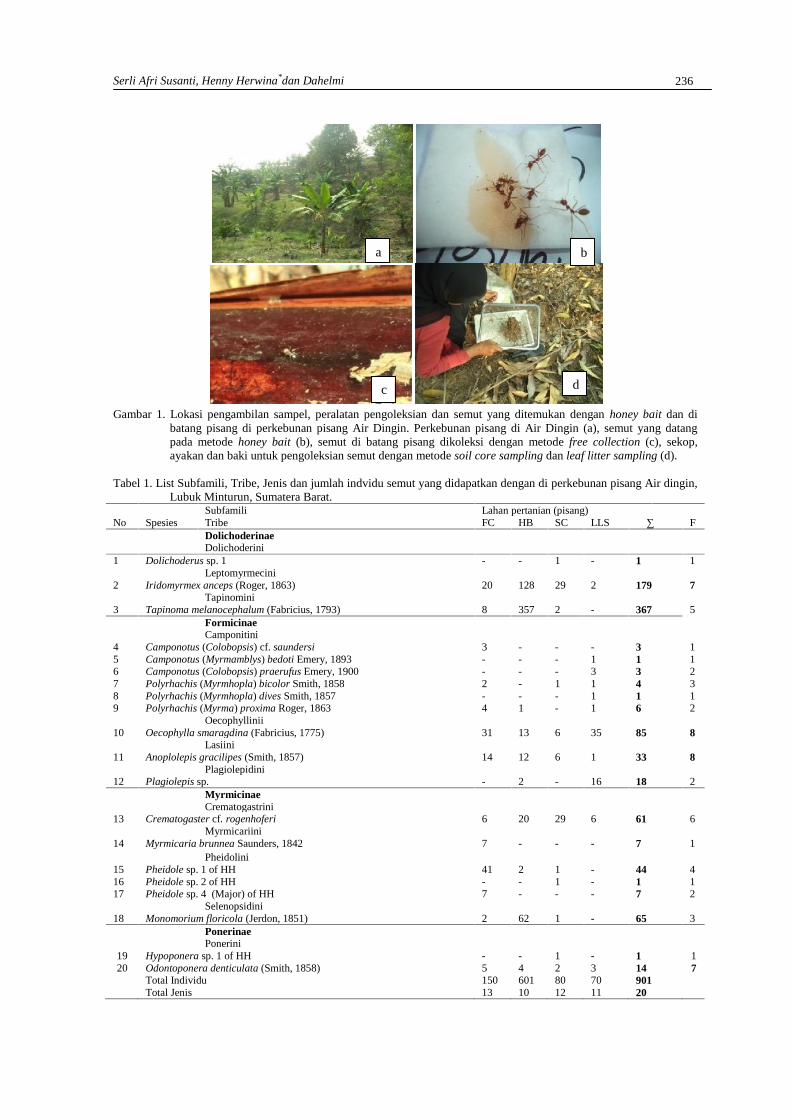
Serli Afri Susanti, Henny Herwina*dan Dahelmi 236
Gambar 1. Lokasi pengambilan sampel, peralatan pengoleksian dan semut yang ditemukan dengan honey bait dan dibatang pisang di perkebunan pisang Air Dingin. Perkebunan pisang di Air Dingin (a), semut yang datangpada metode honey bait (b), semut di batang pisang dikoleksi dengan metode free collection (c), sekop,ayakan dan baki untuk pengoleksian semut dengan metode soil core sampling dan leaf litter sampling (d).
Tabel 1. List Subfamili, Tribe, Jenis dan jumlah indvidu semut yang didapatkan dengan di perkebunan pisang Air dingin,Lubuk Minturun, Sumatera Barat.
No SpesiesSubfamiliTribe
Lahan pertanian (pisang)FC HB SC LLS ∑ F
DolichoderinaeDolichoderini
1 Dolichoderus sp. 1 - - 1 - 1 1Leptomyrmecini
2 Iridomyrmex anceps (Roger, 1863) 20 128 29 2 179 7Tapinomini
3 Tapinoma melanocephalum (Fabricius, 1793) 8 357 2 - 367 5FormicinaeCamponitini
456789
Camponotus (Colobopsis) cf. saundersiCamponotus (Myrmamblys) bedoti Emery, 1893Camponotus (Colobopsis) praerufus Emery, 1900Polyrhachis (Myrmhopla) bicolor Smith, 1858Polyrhachis (Myrmhopla) dives Smith, 1857Polyrhachis (Myrma) proxima Roger, 1863
3--2-4
-----1
---1--
-13111
313416
112312
Oecophyllinii10 Oecophylla smaragdina (Fabricius, 1775) 31 13 6 35 85 8
Lasiini11 Anoplolepis gracilipes (Smith, 1857) 14 12 6 1 33 8
Plagiolepidini12 Plagiolepis sp. - 2 - 16 18 2
MyrmicinaeCrematogastrini
13 Crematogaster cf. rogenhoferi 6 20 29 6 61 6Myrmicariini
14 Myrmicaria brunnea Saunders, 1842 7 - - - 7 1Pheidolini
151617
Pheidole sp. 1 of HHPheidole sp. 2 of HHPheidole sp. 4 (Major) of HH
41-7
2--
11-
---
4417
412
Selenopsidini18 Monomorium floricola (Jerdon, 1851) 2 62 1 - 65 3
PonerinaePonerini
1920
Hypoponera sp. 1 of HHOdontoponera denticulata (Smith, 1858)Total IndividuTotal Jenis
-515013
-460110
128012
-37011
11490120
17
dc
ba
Serli Afri Susanti, Henny Herwina*dan Dahelmi 237
Tabel 2. Perbandingan data semut yang ditemukan di perkebunan pisang Air Dingin dengan semut dipisang dengan gejala BBTV (Herwina et al., 2013) dan semut di dua perkebunan pisang(Pasar usang dan Kampung Pisang Kab. Agam) (Herwina et al., 2014)
No Spesies BBTV
Ps. Usang&K. Pisang
Jenis yangsamaditemukan
Jenis yang tidakditemukan penelitiansebelumnya
1 Anoplolepis gracilipes (Smith, 1857) √ √ √2 Pheidole sp. 4 (Major) of HH √3 Pheidole sp. 1 of HH √ √ √4 Pheidole sp. 2 of HH √ √ √5 Tapinoma melanocephalum (Fabricius, 1793) √ √ √6 Camponotus (Colobopsis) cf. saundersi √7 Oecophylla smaragdina (Fabricius, 1775) √ √ √8 Polyrhachis (Myrma) proxima Roger, 1863 √9 Myrmicaria brunnea Saunders, 1842 √10 Hypoponera sp. 1 of HH √11 Odontoponera denticulata (Smith, 1858) √12 Polyrhachis (Myrmhopla) dives Smith, 1857 √13 Camponotus (Myrmamblys) bedoti Emery, 1893 √14 Plagiolepis sp. √15 Camponotus (Colobopsis) praerufus Emery, 1900 √16 Polyrhachis (Myrmhopla) bicolor Smith, 1858 √17 Dolichoderus sp. 2 √18 Iridomyrmex anceps (Roger, 1863) √19 Monomorium floricola (Jerdon, 1851) √ √ √20 Crematogaster cf. rogenhoferi √
Total 6 9 6 11
Tabel 3. Indeks diversitas Shannon-Wiener yang ditemukan di perkebunan pisang Air Dingin,Lubuk Minturun, Sumatera Barat.Metode H’ H’ totalFC 2,14 1,90HB 1,23SS 1,57LLS 1,64
Jenis yang paling sedikit jumlah individunya(hanya 1 individu) ditemukan pada 5 jenis yaituCamponotus (Myrmamblys) bedoti,Polyrhachis (Mrymhopla) dives, dan Pheidolesp. 2 of HH, Hypoponera sp. 1 of HH danDolichoderus sp. 2. Jumlah jenis lebih banyakditemukan di metode free collection (13 jenisdan 150 individu) diikuti metode soil coresampling (12 jenis dan 80 individu), metodeleaf litter sampling (11 jenis dan 70 individu)dan metode honey bait (10 jenis dan 601individu) (Tabel 1). Pada penelitian ini metodefree collection yang sangat baik dalampengoleksian semut di lahan pertanian hal inidikarenakan semut-semut yang didapatkan
banyak ditemukan di bagian pohon sepertidibagian batang, akar dan ranting pohon.
Jumlah jenis pada penelitian ini lebihrendah dibandingkan dengan beberapapenelitian sebelumnya pada pisang sepertitentang jenis-jenis semut pada pisang dengangejala BBTV di Sumatera Barat oleh Herwinaet al., (2013) yang ditemukan 24 jenis danHerwina et al., (2014) tentang jenis-jenis semutpermukaan pada dua perkebunan pisang diSumatera Barat ditemukan 33 jenis. Walaupundemikian, total jenis semut yang sama denganpenelitian ini dengan Herwina et al., (2013) danHerwina et al., (2014) masing-masingnyahanya 6 jenis dan semut yang ditemukan dipenelitian ini dan tidak ditemukan pada

Serli Afri Susanti, Henny Herwina*dan Dahelmi 238
penelitian sebelumnya terdapat sebanyak 11jenis (Tabel 2). Pada penelitian Herwina et al.,(2013), jenis semut yang banyak didominasioleh Technomyrmrex dan Tetramorium
(masing-masing 4 jenis) dan Herwina et al.,(2014), yang didominasi oleh Pheidole (5 jenis)dan Monomorium (3 jenis).
Perbedaan jumlah dan jenis semut yangdidapatkan kemungkinan karena dipengaruhioleh metode yang digunakan, lokasipengambilan sampel dan faktor lingkungan.Dimana pada penelitian semut di pisang dengangejala BBTV (Herwina et al., 2013), metodeyang digunakan hanya menggunakan koleksilangsung pada lima lokasi dan semutpermukaan pada dua perkebunan pisang(Herwina et al., 2014) walaupun penelitian inimenggunakan metode yang sama denganpenelitian ini tetapi lokasi dan faktorlingkungannya juga sudah berbeda yangpastinya akan mempengaruhi keragaman semutyang ditemukan.
Indeks diversitas Shannon-Wiener diperkebunan pisang Air Dingin adalah 1,90 yangtergolong rendah (Tabel 3). Dilihat dari nilaiindeks diversitas permetode, indeks diversitasdengan metode free collection lebih tinggi(2,14) diikuti oleh metode leaf litter sampling(1,64), soil sampling (1,57) dan paling rendahmetode honey bait (1,23). Hal ini disebabkankarena jumlah jenis yang ditemukan padametode free collection lebih banyak dan dalamjumlah individu tidak ada yang terlalumendominan sedangkan dengan metode honeybait dengan adanya jenis semut yang dominandalam jumlah individunya seperti Tapinomamelanocephalum dibanding dengan jenislainnya yang ada satu jenis hanya ditemukansatu individu.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Telah ditemukan total 20 jenis semut yangtergolong ke dalam empat subfamili, 12 tribe,
14 genera dan 901 individu. Formicinaeditemukan (9 jenis dan 5 genera) diikutiMyrmicinae (6 jenis dan 4 genera),Dolichoderinae (3 jenis dan 3 genera) danPonerinae (2 jenis dan 2 genera). Camponotus,Polyrhachis dan Pheidole memiliki jenis yangbanyak masing-masing tiga jenis. Nilai IndeksDiversitas (H’) semut di Pisang Air Dingintermasuk kategori rendah yaitu 1,90.
UCAPAN TERIMAKASIHUcapan terimakasih kepada Prof. Seiki Yamane,Kagoshima University dan Rijal Satria M.Sc,Tokyo Metropolitan University yang telahmembantu dalam pengidentifikasian. PadaNindy Ladyfandela, Fadli, Larissa hilmi, FitriRoza Wiranata, Deffi Surya Ningsih, RiniOktavia dan Nila Suryayulni yang telahmembantu pengoleksian sampel di lapangan.
DAFTAR PUSTAKA
Bestia, A. 2011. Diversitas Semut(Hymenoptera: Formicidae) Pada EmpatHabitat di Hutan Lindung Sungai PulaiPulau Bintan Kepulauan Riau. Tesis,Universitas Andalas. Padang.
Bolton, B. 1994. Identification Guide to the AntGenera of the World. Harvard UniversityPress London. England.
Bolton, B. 2012. New General Catalogue of theants of the World. Harvard UniversityPress London. England.[not a publication].
Borror, D. J., C. A. Triplehorn, and N. F.Johnson, 1992. Pengenalan PelajaranSerangga. Edisi Keenam. Terjemahanoleh Soetiyon Partosoedjono. Gajah MadaUniversity Press. Yogyakarta.
Departemen pertanian. 2007. Prospek dan ArahPengembangan Agribisis Pisang. Edisikedua. Badan Penelitian danPengembangan. Depertemen Pertanian.Jakarta.
Hashimoto, Y., S. Yamane, and M. Mohamed.2001. How to Design an InventoryMethod for Ground Level Ants in TropicalForest. Nature and Human Activities 6:25-30.
Hashimoto, Y. 2003. Identification Guide to theAnt Subfamily of Borneo. Tools for

Serli Afri Susanti, Henny Herwina*dan Dahelmi 239
Monitoring Soil Biodiversity in theASEAN Region. Darwin Initiative
Herwina, H., N. Nasir, Jumjunidang andYaherwandi. 2013. The composition ofant species on banana plants with BananaBunchy-top Virus (BBTV) symptoms inWest Sumatra, Indonesia. AsianMyrmecology 5: 151-161.
Herwina, H., N. Nasir., Mairawita and Erniwati.2014. Ground ant species (Formicidae) intwo banana plantations of West Sumatra.Prosiding Semirata 2014. Bogor. [in print].
Magurran, A. E. 2004. Measuring BiologicalDiversity. Blackwell Sciene Ltd. UnitedKingdom.
Mohamed, M. 2003. Manual for Bornean Ant(Formicidae) Identification. Tools forMonitoring Soil Biodiversity in TheASEAN Region. Darwin Initiative.
Primack, R. B., J. Supriatna, M. Indrawan danP. Kramadibrata. 1988. Biologi
Konservasi. Yayasan Obor Indonesia.Jakarta.
Shattuck, S. O. 2000. Australian Ants: TheirBiology and Identification. CSIRO Public.Collingwood.
Suriyapong, Y. 2003. Study of GroundDwelling Ant Populations and TheirRelationship to Some Ecological Factorsin Sakaerat Environmental ResearchStation Nakhon Ratchasima. Thesis,Suranaree University of Technology.Thailand.
Wilson, E. O. 1979. The Insect Societies.Harvard University Press. Cambridge.Massachusetts. London.

ISSN 978-602-14989-0-3 BioETI
Daya hambat formulasi minyak daun Kayu Manis denganpenambahan minyak Serai Wangi sebagai biopestisida dalammenghambat Fusarium pada batang buah Naga secara InvitroSHYNTIA HARSARI1) NASRIL NASIR1)*) FUJI ASTUTI FEBRIA1) JUMJUNIDANG 2) DANNURMANSYAH3)
1)Labor Mikrobiologi, Jurusan Biologi, FMIPA, Universitas Andalas, Padang, Sumatera Barat 251632)Balai Penelitian Tanaman Buah Tropika, Jl. Raya Solok-Aripan Km. 7 Solok3) Balai Penelitian Tanaman Rempah dan Obat. KP. Laing. Solok. Sumatera Barat.E-mail: [email protected]
ABSTRAKPenelitian tentang daya hambat formulasi minyak daun kayu manis dengan penambahan minyak serai wangi sebagaibiopestisida dalam menghambat Fusarium pada batang buah naga secara invitro telah dilakukan di Balai Penelitiantanaman buah tropika (Balitbu) di Aripan Solok, dari bulan Juni sampai Agustus 2014. Penelitian dilakukan denganmetode eksperimen menggunakan Rancangan Acak Lengkap (RAL) dengan 6 ulangan untuk masing-masing perlakuan.Perlakuan terdiri dari kontrol (tanpa pemberian biopestisida), minyak daun kayu manis dengan penambahan minyak seraiwangi dengan konsentrasi 500 ppm, 1000 ppm, dan 1500 ppm. Hasil efektif didapat pada perlakuan pemberian formulasibiopestisida minyak daun kayu manis dengan penambahan minyak serai wangi dalam menghambat Fusarium sp. padakonsentrasi 1500 ppm dengan daya hambat 65,88 %.
Key words: buah naga, Fusarium sp., biopestisida, kayu manis, dan serai wangi
Pendahuluan
Buah naga (Hylocereus polyrhizus) merupakansalah satu tanaman buah yang kini mulaibanyak dibudidayakan di Indonesia. Budidayabuah naga semakin berkembang seiring denganpermintaan pasar yang terus meningkat. Upayauntuk memenuhi permintaan pasar domestikyang semakin tinggi, hingga dilakukanperluasan budidaya buah naga (Masyahit et al.,
2009). Namun, semenjak 5 tahun terakhir,serangan patogen mampu menghancurkanpertanian buah naga di Sumatera Barat danRiau lebih dari 50% (Nasir, 2013). MenurutJumjunidang (2012), ada beberapa penyebabyang berbahaya yang mengancam produksibuah naga di Indonesia diantaranya yaitupenyakit busuk batang yang disebabkan olehmikroba salah satunya cendawan dari genusFusarium sp., Gejala serangan Fusarium padabuah naga ditandai dengan perubahan warnadaun yang paling tua menjadi kekuninganterjadi pada satu sisi tanaman atau pada daunyang sejajar dengan petiol tanaman. Daun yangterinfeksi akan layu dan mengering, tetapi tetap
menempel pada tanaman. Kelayuan akanberlanjut ke bagian daun yang lebih muda dantanaman akan segera mati. Sejauh ini, belumada laporan keberhasilan pengendalian patogenpada buah naga, termasuk serangan Fusariumsp.
Pada beberapa tanaman yang diserang olehFusarium sp., pengendalian dilakukan hanyamenggunakan pestisida kimia. MenurutQuijano (2001), penggunaan pestisida kimiadapat memberikan dampak negatif terhadaplingkungan dan manusia diantaranya, residupestisida di dalam tanah dapat meracuniorganisme non target, dan meracuni lingkungansekitar. Bahkan, residu pestisida pada tanamanmasuk ke rantai makanan, sehingga dapatmeracuni konsumen. Akibat lain diantaranyaresistensi, ancaman bagi predator, dan lain-lain.Pemanfaatan biopestisida merupakan alternatifpengendalian yang tepat. Mengingatbiopestisida mudah didapat dan harganya relatifmurah, biopestisida yang telah diujikandiantaranya minyak daun kayu manis dan seraiwangi.

Shyntia Harsari, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 241
Daun dari tanaman kayu manis merupakanserasah yang belum dimanfaatkan dandibiarkan berserakan. Pemanfaatan daun kayumanis sebagai biopestisida sudah diteliti danterbukti mampu mengendalikan patogenFusarium oxysporum penyebab penyakit layutomat dan Sclerotium rofsii penyebab penyakitbusuk pangkal batang tanaman cabai(Nurmansyah, 2001; Chrisnawati, 2001).Minyak daun kayu manis mengandungsenyawa sinamaldehid 55-65%, eugenol 4-8%.Kandungan lain adalah methyl ketene, furfutal,benzaldehyde, nonylaldehyd, hydrocinnamicaldehyde, cuminaldehyde dan cumarine(Rismunandar dan Farry, 2001). MenurutNurmansyah, Syamsu dan Nasrun (1997)minyak daun kayu manis pada konsentrasi 500ppm dapat menghambat patogen Sclerotium
rofsii penyebab penyakit busuk pangkal batangtanaman kacang tanah dan cabai.
Chrisnawati (2001), melaporkan bahwaminyak serai wangi mampu mengendalikanpertumbuhan vegetatif Fusarium oxysporum fsp lycopersici dengan daya hambat 40,75 –52,68 % pada konsentrasi 750 ppm. Komponenutama minyak serai wangi adalah citronellaldan geraniol. Nurmansyah (2001) mencobakanminyak serai wangi dengan penambahan sirih-sirih dan kayu manis hasilnya menunjukkanbahwa kombinasi tersebut dapat meningkatkandaya antifungal pestisida nabati yang dapatmenghambat Fusarium oxysporum var Vanillaedan Fusarium oxysporum var Zynggiberi100%, dengan konsentrasi 1000 ppm.
Sejauh ini, belum ada penelitian tentangkombinasi penggunaan pestisida nabati minyakdaun kayu manis dengan penambahan minyakserai wangi untuk menghambat Fusarium
secara invitro. Dipilihnya serai wangi sebagaisalah satu penambahan dalam komposisibiopestisida, karena pada minyak serai wangiterdapat kandungan sitronellal dan geraniolyang dapat bersifat antifungal terhadapbeberapa jamur patogen tanaman (Sait, 1991).
Pada penelitian ini, akan di uji kemampuandaya hambat minyak daun kayu manis dengan
penambahan minyak serai wangi sebagaibiopestisida untuk menghambat Fusarium sp.,pada batang buah naga secara invitro.
BAHAN DAN METODE
Penelitian ini dilaksanakan pada bulan Juni –Agustus 2014 di laboratorium Balai Penelitiantanaman buah tropika (Balitbu) di Aripan Solok.Penelitian ini dilakukan dengan metodeeksperimen menggunakan Rancangan AcakLengkap (RAL) 4 perlakuan dan 6 ulanganuntuk masing-masing perlakuan. Perlakuanterdiri dari kontrol (tanpa permberianbiopestisida), formulasi minyak kayu manis +serai wangi dengan konsentrasi 500 ppm, 1000ppm dan 1500 ppm. Koleksi isolat jamurFusarium sp. diperoleh dari Balai PenelitianTanaman Buah (Balitbu) Tropika, di Aripan,Solok dan formulasi minyak daun kayu manis +serai wangi diperoleh dari Balai TanamanRempah dan Obat (Balitro) di Laing, Solok.
Parameter yang diamati berupapertumbuhan koloni jamur pada media PDApada masing-masing perlakuan yangdibandingkan dengan perlakuan kontrol.Pengamatan dilakukan sampai diameter jamurFusarium sp., pada perlakuan kontrolmemenuhi cawan petri. Daya hambat pengaruhpemberian formulasi minyak daun kayu manis+ serai wangi terhadap pertumbuhan jamurFusarium sp., dihitung dengan rumus sebagaiberikut:
Dari hasil uji daya hambat formulasi minyakdaun kayu manis + serai wangi terhadappertumbuhan jamur Fusarium sp., dilakukan ujiresistensi patogen pada perlakuan yangmemiliki daya hambat 100% (jamur padacawan petri tidak tumbuh). Uji resistensidilakukan dengan cara memindahkan kembaliinokulasi jamur patogen yang pertumbuhannyaterhambat karena pengaruh pemberianformulasi minyak daun kayu manis + seraiwangi ke medium PDA tanpa penambahanformulasi minyak. Selanjutnya diinkubasi pada

Shyntia Harsari, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 242
suhu ruang. Pertumbuhan koloni diketahuidengan mengukur diameter koloni jamur yangtumbuh pada setiap perlakuan. Jika kolonijamur tersebut tidak tumbuh, berarti formulasiminyak yang diberikan dapat membunuh kolonijamur, namun jika koloni jamur tersebuttumbuh berarti formulasi minyak yangdiberikan hanya dapat menghambatpertumbuhan koloni jamur.
HASIL DAN PEMBAHASAN
Pada pengamatan yang telah dilakukan, dayahambat formulasi minyak daun kayu manisdengan penambahan minyak serai wangi dalammenghambat pertumbuhan jamur Fusarium sppada hari ke-7 seperti yang dapat dilihat padaTabel 1.
Tabel 1. Persentase Daya hambat minyak daunkayu manis dengan penambahanminyak serai wangi dalammenghambat jamur Fusarium sp.,
Perlakuan Diameterkoloni (cm)
Persentasedayahambat (%)
Kontrol 9 a 0KM + SW (500 ppm) 4,72 b 47,55KM + SW (1000 ppm) 4,35 c 51,66KM + SW (1500 ppm) 3,07 d 65,88Keterangan:angka-angka pada tabel yang diikuti huruf kecilyang sama menunjukkan nilai yang tidakberbeda nyata pada DNMRT 5%. KM = Kayumanis, SW = Serai wangi
Pada Tabel 1 Dapat dilihat dari hasil analisisstatistik menunjukkan bahwa pemberiankonsentrasi yang berbeda menunjukkanpengaruh yang signifikan (berbeda nyata)terhadap jamur Fusarium sp., Semakin tinggikonsentrasi, semakin tinggi pula daya hambatterhadap jamur Fusarium sp., Pada konsentrasi500 ppm minyak daun kayu manis denganpenambahan minyak serai wangi mampumenghambat jamur Fusarium. sp., sebesar47,55 %, pada konsentrasi 1000 ppm JamurFusarium sp., mampu menghambat 51,66 %namun, pada konsentrasi 1500 mampumenghambat > 65 % yaitu sebesar 65,88 %.
konsentrasi terbaik untuk menghambat jamurFusarium sp., adalah pada konsentrasi 1500ppm dengan persentase daya hambat > 65 %yaitu sebesar 65,88 %. Maka, formulasiminyak daun kayu manis dengan penambahanminyak serai wangi dapat digunakan sebagaibiopestisida dalam menghambat jamurFusarium sp.
Bailey, Boyetchko dan Lange (2010)menyatakan bahwa konsentrasi suatu bahanyang berfungsi sebagai antimikroba merupakansalah satu faktor penentu besar kecilnyakemampuan dalam menghambat pertumbuhanmikroba yang diuji. Kerusakan yangditimbulkan komponen antimikroba dapatbersifat fungisidal (membunuh jamur) danfungistatik (menghentikan sementarapertumbuhan jamur). Suatu komponen akanbersifat fungisidal atau fungistatik tergantungpada sifat senyawa aktifnya, konsentrasi, danmedia yang digunakan.
Terhambatnya pertumbuhan jamurFusarium sp., disebabkan adanya komponensenyawa minyak atsiri yang terdapat dalamdaun kayu manis yang mempunyai aktifitasfungisidal. Rendemen minyak yang terdapatdalam daun kayu manis berkisar antara 0,24 –0,56% dengan komponen utamanya adalaheugenol berkisar antara 90,20 – 91,02% dansinamaldehid 23,80 – 42,00%. Sinamaldehidyang merupakan komponen utama minyakatsiri daun kayu manis secara alami didalamkulit batang dan daun dari genusCinnamommum, telah terbukti mempunyaiaktifitas antifungal yang kuat dan merupakanfungisida terbaik terhadap jamur pembusukkayu (Yen dan Chang, 2008). Senyawa volatile
yang dihasilkan dari minyak daun kayu manisjuga mampu menghambat jamur patogentanaman secara invitro (Nurmansyah, 2001).Selain itu, kandungan utama minyak seraiwangi seperti sitronellal dan graniol jugabersifat antifungal dan efektif untukpengendalian jamur Fusarium sp., keduanyatermasuk kelompok terpenoid yang tergolongmonoterpen yang mampu menekan
Shyntia Harsari, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 243
pertumbuhan jamur patogen. Senyawa-senyawaini dapat menghambat proses metabolismejamur sehingga akan mengganggupertumbuhan jamur (Nurmansyah, 2001).
Menurut Knoblock, Pauli, Iberl, Weiganddan Weis (1989), mengatakan bahwa,komponen minyak atsiri yang bersifatantifungal mampu menembus dinding sel jamur.Akibatnya, akan mengganggu prosesmetabolisme dalam sel, mekanisme kerja darikelompok terpenoid dapat mereduksi miselium.Akibatnya terjadi pemendekan pada ujung hifa,mampu menghambat proses metabolismedengan cara mengakumulasi globula lemak didalam sitoplasma sel, mengurangi jumlahmitokondria dan merusak membran nukleus.
Formulasi minyak daun kayu manis denganpenambahan minyak serai wangi dapatmenghambat jamur Fusarium sp., Hal inidisebabkan daya kerja komponen yang bersifatantifungal dikedua minyak tersebut sinergis,karena bahan aktifnya sama-sama mempunyaisifat anti jamur.
Kontrol (a) 500 ppm (b)
1000 ppm (c) 1500 ppm (d)
Gambar 1. Pertumbuhan diameter koloni jamurFusarium sp., dengan perlakuan (a)kontrol, (b) formulasi minyak daunkayu manis + serai wangi 500 ppm, (c)formulasi minyak daun kayu manis +serai wangi 1000 ppm, (d) formulasiminyak daun kayu manis + serai wangi1500 ppm
KESIMPULAN
Berdasarkan hasil penelitian yang telahdilakukan tentang daya hambat formulasiminyak daun kayu manis dengan penambahanminyak serai wangi sebagai pestisida nabatidalam menghambat Fusarium buah naga secarainvitro diperoleh kesimpulan sebagai berikut:Kombinasi biopestisida minyak daun kayumanis + serai wangi menunjukkan kerja yangsinergis, sehingga mampu menghambat jamurFusarium sp., dan konsentrasi efektif minyakdaun kayu manis + minyak serai wangi dalammenghambat Fusarium sp. secara invitroadalah pada konsentrasi 1500 ppm denganpersentase daya hambat sebesar 65,88 %.
DAFTAR PUSTAKA
Bailey, K.L., S.M. Boyetchko, and T. Lange.2010. Social and economic driversshaping the future of biological control:A Canadian perspective on the factorsaffecting the development and use ofmicrobial biopesticides BiologicalControl 52 (2010) 221–229
Chrisnawati. 2001. Uji daya kendali Pestisidanabati minyak serai wangi dan fraksinyaterhadap
Fusarium oxysporum f sp lycopersici penyebabpenyakit layu tanaman tomat. Journalstigma An Agricultural Science Journal.Faperta Unand. Padang. P 350-353.
Jumjunidang, Riska dan I. Muas. 2012.Outbreak Penyakit Busuk BatangTanaman Buah Naga di Sumatera Barat.Laporan Hasil survey OPT di SentaProduksi Buah Naga Sumatera Barat.Balitbu Tropika Solok. 6 hal.
Knoblock, K.,A, Pauli.,B, Iberl, H, Weigandand N, Weis. 1989. Antibacterial andAntifungal propertis of Essential oilComponds. J. Ess, oil. Res 1p; 119-128
Masyahit, M., K. Sijam, Y. Awang and M. G.Satar . 2009. The First Report of theOccurreance of Anthracnose DiseaseCaused by Colletotrichumgloeosporioides (Penz.) Penz. &acc. onDragon Fruit (Hylocereus spp.) inPeninsular Malaysia. American Journal ofApplied Scienses. 6 (5): 902-912.

Shyntia Harsari, Nasril Nasir, Fuji Astuti Febria, Jumjunidang dan Nurmansyah 244
Nasir. N. 2013. Serangan Penayakit Pada BuahNaga Hylocereus polyrhizus DiKepulauan Riau. Laporan Penelitian.Jurusan Biologi FMIPA UniversitasAndalas. 5 hal.
Nurmansyah, H. Syamsu dan Nasrun (1997).Kajian kemungkinan Pemanfaatan limbahproduksi kayu manis (C. Burmanii)sebagai fungisida nabati. ProsidingSeminar Kongres Nasional dan SeminarIlmiah PFI IVX 27-29 oktober 1995.Perhimpunan Fitopatologi Indonesia.Palembang, hal. 258-261.
Nurmansyah. 2001. Uji efikasi minyak kayumanis (Cinnamommum burmanii)terhadap jamur Fusarium oxysporum.Prosiding KSN. PFI XVI. 21-23 Agustus2001. Perhimpunan Fitopatologi Indonesia.Bagian Hama dan Penyakit InstitutPertanian Bogor. P. 260-264.
Quijano. Sarojeni V. Rengam. 2001. PestisidaBerbahaya Bagi Kesehatan. Solo:Yayasan Duta
Awam Pesticide Action Network Asia and thePacific
Rismunandar dan F.B. Paimin. 2001. KayuManis Budidaya dan Pengolahannya.Penebar Swadaya. Jakarta. 120 hlm.
Sait, S. 1991. Potensi minyak atsiri daunIndonesia sebagai sumber bahan obat.Prosiding Forum Komunikasi IlmiahPengembangan atsiri di Sumatera.Bukittinggi.Balitro. Bogor.
Yen, T,B dan S,T, Chang. 2008. Synergisticeffects of cinnamaldehyde in combinationwith Eugenol against wood decay fungi.Short Communication. BioresourceTechnology 99,232-236.

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman jenis tumbuhan asing invasif di vegetasisemak belukar di Hutan Pendidikan dan Penelitian Biologi(HPPB) Universitas AndalasSOLFIYENI, SYAMSUARDI, CHAIRUL, WELLA YURANTI DAN AFRIDA YULIA
Labor Ekologi Tumbuhan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKTelah dilakukan penelitian tentang keanekaragaman tumbuhan asing invasif pada vegetasi semak belukar di HutanPendidikan dan Penelitian Biologi (HPPB) Universitas Andalas yang bertujuan untuk mengetahui komposisi dan strukturtumbuhan asing invasif pada vegetasi semak di kawasan hutan tesebut. Penelitian ini menggunakan metode transekdengan peletakan plot secara sistematik sampling. Jumlah plot pengamatan sebanyak 40 plot dengan ukuran 2 x 2 meter.Hasil penelitian menunjukkan pada vegetasi semak belukar HPPB ditemukan 28 jenis tumbuhan asing invasif. Familidominan pada vegetasi ini adalah famili Leguminosae dan Compositae. Jenis tumbuhan asing invasif yang mendominasiadalah Mimosa pudica, Borreria laevis dan Melastoma malabathricum dengan Nilai Penting (NP) masing-masingnya24,16%, 23,58% dan 22,62%. Indeks keanekaragaman jenis tumbuhan asing invasif pada kawasan ini tergolong sedangyaitu 2,89.
Key words: tumbuhan asing invasif, komposisi, struktur, HPPB
Pendahuluan
Keanekaragaman hayati merupakan sumberdaya penting yang perlu dijaga kelestariannyakarena memiliki fungsi yang beranekaragamdiantaranya fungsi yang bersifat ekologis danekonomis. Indonesia adalah salah satu negarayang kaya dengan keanekaragaman jenis flora.Keanekaragaman hayati di Indonesia termasukdalam golongan tertinggi di dunia, jauh lebihtinggi dari pada keanekaragaman sumber dayahayati di Amerika maupun Afrika tropis,apalagi bila dibandingkan dengan daerahberiklim sedang dan dingin. Jenis tumbuh-tumbuhan di Indonesia secara keseluruhanditaksir sebanyak 25.000 jenis atau lebih dari10 % dari flora didunia (Soemarwoto, 1983).
Salah satu pulau di Indonesia yangmemiliki keanekaragaman hayati yang tinggidan endemisitas yang luar biasa yaitu pulauSumatera. Kekayaan tersebut terdapat dalamberbagai tipe ekosistem, dan habitat mulai daridataran rendah sampai pegunungan. Kawasandataran rendah Sumatera diantaranya adalahHPPB.
HPPB merupakan Hutan Pendidikan danPenelitian Biologi (HPPB) Universitas Andalasterletak di kawasan kampus UniversitasAndalas, Limau Manis Padang. Kawasan iniberbatasan dengan kawasan hutan lindung yangmerupakan cadangan air untuk kotamadyaPadang. HPPB termasuk hutan tropis dataranrendah, dengan ketinggian 200–460 m dpldengan luas 150 Ha (Nasir, 2010). Di dalamhutan HPPB terdapat berbagai macam flora danfauna yang beberapa diantaranya termasukbiota yang dilindungi. HPPB telah dijadikansebagai salah satu daerah kunci biodiversitasyang penting di Sumatera (ConservationInternasional, 2006). Selain itu, HPPB jugatelah digunakan sebagai salah satu lokasi dalamriset biodiversity sejak tahun 1982 hingga saatini (Rahman, 1994).
Menurut Rahman dkk., (1994), ditinjau darisudut ekologi HPPB merupakan gabungan daritiga tipe vegetasi yaitu: vegetasi semak belukar,vegetasi bekas perladangan serta vegetasi hutanprimer dan skunder. Secara umum hutan initergolong hutan skunder, ditandai denganditemukannya banyak daerah terbuka denganpohon-pohon bekas perladangan dan spesies

Solfiyeni, Syamsuardi, Chairul, Wella Yuranti dan Afrida Yulia 246
pionir. Jenis tumbuhan yang terdapat di HPPBterdiri dari 165 jenis pohon, 27 jenis vegetasidasar, 29 jenis paku-pakuan dan 32 jenis gulma.
Tipe vegetasi semak belukar di kawasanHPPB terletak di pinggiran kawasan hutan.Kawasan ini merupakan areal terbuka dansering dilalui dengan pejalan kaki ataupunmenggunakan kendraan. Berdasarkanpengamatan sepintas, pada vegetasi semakbelukar ini terdapat beberapa jenis tumbuhanasing invasif. Oleh sebab itu, terjadikekhawatiran akan terjadinya degradasikeanekaragaman tumbuhan spesies asli diHPPB karena keanekaragaman hayati yang adadiseluruh dunia saat ini mengalami berbagaiancaman diantaranya adalah keberadaan jenis-jenis asing invasif (Invasive Alien Spesies /IAS). Adanya jenis-jenis asing invasif sangatbesar pengaruhnya terhadap suatu ekosistem.Invasi tumbuhan invasif (IAS) ke dalam hutandapat menurunkan keanekaragaman tumbuhanhutan karena tumbuhan invasif dapatmenguasai bahkan menggantikan tumbuhanasli di hutan tersebut.
Spesies asing invasif didefinisikan sebagaispesies yang bukan spesies lokal dalam suatuekosistem yang dapat mendominasi suatuhabitat baru. Spesies asing invasif dapatmenyebabkan gangguan terhadap pertumbuhanekonomi dan lingkungan, serta berdampakburuk bagi kesehatan manusia. Spesiestumbuhan asing invasif dilaporkan telahmenjadi permasalahan ekologi di beberapakawasan konservasi di Indonesia, sepertiAcacia nilotica di Taman Nasional Baluran,Passiflora suberosa di TN Gunung GedePangrango, Chromolaena odorata di TNlUjung Kulon, Lantana camara di TN MeruBetiri, Merremia peltata di TN Bukit BarisanSelatan, dan Eichhornia crassipes di TamanNasional Wasur (BLK 2010; Purwono dkk.,
2002).Tumbuhan invasif mempunyai
karakteristik sebagai berikut: produksi biji yangberlimpah dalam setahun, koloni yang stabil,kemampuan menyebarkan melebihi akar dalam
tanah, cepat pulih kembali setelah dipotong,hampir tidak mempunyai predator, bijidormansinya lama, akan pecah apabila kondisilingkungan sesuai, perkecambahan tidakserentak, biji berkecambah bila ada cahaya,tidak dapat berkecambah dalam gelap,kecambah teradaptasi dengan tempat terbukadalam berbagai variasi suhu dan kelembapan,tidak tergantung pada jenis tanah tertentu,populasi tinggi dan mampu memproduksi bijisangat banyak dan berkesinambungan. Dapatmengendalikan pertumbuhan populasitumbuhan asli, bahkan sifat ini sangat menonjolpada tumbuhan asing invasif seperti:Chromolaena odorata, Mimosa pigra, danMikania micrantha (Credit ValleyConservation, 2004).
Saat ini di Indonesia ada 113 IAS (InvasifAlien Spesies) atau jenis invasif, 40diantaranya asli dari Indonesia, 59 dari negaraluar, dan sisanya belum diketahui apakahberasal dari Indonesia atau dari negara luar.Dari 113 Invasif Alien Spesies tersebut, 27diantaranya termasuk dalam kategori yangsangat berbahaya dan dapat menjadi salah satupenyebab merosotnya keanekaragaman hayati.Reproduksi tumbuhan invasif alien spesies,memiliki pertumbuhan yang sangat cepat,sehingga menjadi salah satu gulma terganas diIndonesia. Sekarang tumbuhan invasif mulaimerambah ke hutan alami, yang dikhawatirkankehadirannya dapat merusak keanekaragamantumbuhan asli di Indonesia (Binggeli, 1997).Sehubungan dengan permasalahan yangditimbulkan oleh jenis-jenis tumbuhan invasifdan sejauh ini penelitian mengenai spesies inidi HPPB belum banyak diungkap. Oleh karenaitu perlu dilakukan penelitian mengenai jenis-jenis tumbuhan invasif di HPPB sebagai salahsatu upaya untuk melindungi keanekaragamanhayati, mengingat kawasan ini merupakankunci biodiversitas yang penting di SumateraBarat. Tujuan dari penelitian ini adalah untukmengetahui komposisi dan struktur jenis-jenistumbuhan invasif di HPPB

Solfiyeni, Syamsuardi, Chairul, Wella Yuranti dan Afrida Yulia 247
BAHAN DAN METODE
Penelitian ini dilaksanakan dari bulan Junisampai Oktober 2014. Penelitian dilakukanpada vegetasi semak belukar (HPPB) LimauManis Padang, kemudian dilanjutkan diLaboratorium Ekologi Tumbuhan danHerbarium Universitas Andalas (ANDA)Jurusan Biologi, FMIPA, Universitas Andalas.
Metoda yang digunakan adalah metodatransek dengan menggunakan plot yangberukuran 2 x 2 meter, dan jumlah plotsebanyak 40 plot. Plot diletakkan berselangseling disepanjang transek dengan jarak 5meter.
Alat dan bahan yang digunakan dalampenelitian ini adalah: meteran, pancang, talirafia, GPS, altimeter, kamera digital, guntingtanaman, parang, label gantung, karung plastik,karet gelang, koran bekas, plastik ukuran 20 kg,alkohol 70%, papan pres untuk spesimen, oven,buku identifikasi, label herbarium, lakban,benang jagung, jarum jahit, alat-alat tulis.
Teknik pengumpulan data di lapangandilakukan dengan cara pengamatan langsungpada setiap plot dengan mengamati jenis-jenistumbuhan invasif, jumlah individu masing-masing jenis serta habitus dari setiap jenistumbuhan invasif yang ditemukan. Dilakukanpemotretan, pengambilan sampel jenis-jenistumbuhan yang belum diketahui namanya dilapangan. Selanjutnya jenis yang dikoleksi dilapangan diidentifikasi di laboratorium denganmenggunakan buku-buku identifikasi dan list
species invasive.Analisis Data
Komposisi:1. Jenis-jenis tumbuhan invasif serta familidominan dan co-dominanPersentase famili = Jumlah individu suatu famili x 100 %
Jumlah semua individu
Famili dikatakan dominan pada suatu kawasanjika memiliki persentase > 20 % dan co-dominan jika persentasenya 10% - 20%(Johnston and Gillman, 1995).Struktur
Untuk analisis data yang didapatkan darilapangan digunakan parameter – parameteryang digunakan oleh Mueller-Dombois danEllenberg (1974), yaitu: Kerapatan Relatif,Frekuensi Relatif dan Nilai Penting.Kerapatan = Jumlah individu
Luas areal contoh
Kerapatan Relatif (KR) = Kerapatan suatu jenis x 100%Kerapatan seluruh jenis
Frekuensi = Jumlah unit contoh terdapatnya suatu jenisJumlah seluruh unit contoh
Frekuensi Relatif = Frekuensi suatu jenisx 100%Frekuensi Seluruh jenis
Nilai Penting (NP) =KR + FR
Untuk melihat keanekaragaman spesiesdigunakan Indeks Shanon atau Shanon Index ofGeneral diversity (H) dimana :
H = - ∑ {(ni/N) log (ni/N)}H = Indeks keanekaragaman Shanonni = Nilai penting dari tiap spesiesN = Total Nilai penting.(Sumber = Odum, 1994; dalam Indriyanto,2008)
HASIL DAN PEMBAHASAN
Berdasarkan Tabel 1 dapat dilihat pada vegetasisemak belukar HPPB ditemukan 28 jenistumbuhan asing invasif yang tergolongkedalam 15 famili. Jumlah individu adalahsebanyak 1161 individu. Jumlah jenistumbuhan asing invasif yang ditemukan jauhlebih banyak dibandingkan dengan tumbuhaninvasif di kawasan taman hutan Kenali Jambi,dimana hanya ditemukan 6 jenis tumbuhanasing invasif yang tergolong kedalam 4 famili(Susanti dkk., 2013). Akan tetapi jumlah jenistumbuhan asing invasif pada vegetasi semakbelukar HPPB ini lebih sedikit dibandingkandengan hasil penelitian Sunaryo dkk., (2012)yang menemukan tumbuhan asing invasifsebanyak 74 jenis di taman nasional gunungGede Pangrango, Jawa Barat.

Solfiyeni, Syamsuardi, Chairul, Wella Yuranti dan Afrida Yulia 248
Tabel 1. Jenis-Jenis Tumbuhan Asing Invasif di Hutan Pendidikan dan Penelitian Biologi (HPPB)
No Famili Spesies Asal Jumlahindividu
1 Acanthaceae Asystasia gangetica (L.)T. Anders. Afrika 61
2 Compositae Ageratum conyzoides L. Am. Utara 17Austroeupatorium inulifolium (Kunt.) R.M. King & H. Rob. Am. Selatan 7Clibadium surinamense L. Am. Tropik 48Mikania micrantha Kunt. Am. Selatan 73Spagneticola trilobata(L.C.Rich) Pruski 91
3 Costaceae Cheilocostus speciosus(J.Koenig) Sm. Asia Tenggara 24 Lamiaceae Hyptis capitata Jaq. Am. Tropik 85 Malvaceae Sida acuta Burm. Asia 46 Melastomataceae Clidemia hirta (L.) Don. L. Am. Selatan 2
Melastoma malabathricum L. Asia 1337 Leguminoceae Acacia auriculiformis Benth. Australia 1
Calliandra calothrysus Meisn. Am. Tengah 2Mimosa pudica L Am. Tropik 189Mimosa pigra L. Am. Tropik 8Leucana leucocephala (Lam.) De Wit
Centrosemavirginianum (L). Benth.Camaecristanictitans(L). Moench
Mexico& Amerika TengahAsia Tenggara
6
243
8 Myrtaceae Rhodomyrtus tomentosa (Aiton). Hassk. Asia Tenggara 49 Oxalidaceae Oxalis barrelieri L. Am. Tropik 1610 Piperaceae Piper aduncum L. Am. Tropik 311 Poaceae Imperata cylindrica (L.) Raeusch. Asia Tropik 113
Themeda gigantea (Cav.) Hac - 5412 Rubiaceae Borreria leavis (Lamk.) Griseb. Am. Tropik 19413 Rosaceae Rubus moluccanus Anet. Asia Timur & Pasifik 314 Verbenaceae Lantana camara L. Amerika 13
Stachytarpheta jamaicensis (L.) (Vah). Am. Tropik 6215 Vitaceae Cissus hastata Miq. Asia Selatan & Asia
Tenggara2
Total 1161
Tabel 2. Famili Dominan dan Co-dominan Tumbuhan Asing Invasif di HPPB
No. Famili Persentase (%)
12345
LeguminosaeCompositaeRubiaceaePoaceaeMelastomataceae
21,6220,3216,7014,3811,62
Tabel 3. Jenis-jenis Tumbuhan Asing Invasif yang Utama di HPPB
No. Nama Jenis Famili NP (%)1. Mimosa pudica Leguminoceae 24,16
2. Borreria laevis Rubiaceae 23,583. Melastoma malabathricum Melastomataceae 22,624. Imperata cylindica Poaceae 15,325. Lantana camara Verbenaceae 14,736. Mikania micrantha Compositae 14,487. Clibadium surinamense Compositae 11,058. Themeda gigantea Poaceae 8,639. Piper aduncum Piperaceae 8,1810. Stachytarpheta jamaicensis Verbenaceae 7,81

Solfiyeni, Syamsuardi, Chairul, Wella Yuranti dan Afrida Yulia 249
Pada Tabel 3 dapat dilihat jenis yang banyakditemukan dan mendominasi areal penelitianadalah Borreria laevis yaitu sebanyak 194individu dengan INP 23,58 %. Jenis Borreria
laevis ini merupakan gulma semusim yangberasal dari Amerika tropik, tumbuh pada tanahterbuka atau sedikit ternaungi terutama padatanah keras, daerah penyebarannya cukup luasmeliputi ketinggian 1 – 1000 meter di ataspermukaan laut serta tumbuhan ini berbungasepanjang tahun (Nasution, 1986). Selain itujenis yang banyak dijumpai adalah Mimosa
pudica, Melastoma malabathricum danImperata cylindrica.
Mimosa pudica juga banyak ditemukan dilokasi penelitian yaitu 189 individu dengan INP24,16 %. Jenis ini ditemukan tumbuh tersebar(jarang-jarang) dan juga banyak yang tumbuhmengelompok. Mimosa pudica berasal dariAmerika tropik, sering ditemukan di tanah yangtidak diusahakan, di tepi jalan, di tepi sungaidan di pekarangan. Daerah penyebarannyameliputi ketinggian 1 – 1200 meter di ataspermukaan laut (Nasution, 1986).
Melastoma malabathricum juga termasukjenis yang umum ditemukan di areal penelitiandengan jumlah 133 individu dan INP 22,62 %.Syamsuardi, Mansurdin dan Suryani (2007)menyatakan berdasarkan hasil percobaanpolinasi dan pemeriksaan ratio-pollenmenunjukkan adanya strategi reproduktiftumbuhan ini sehingga mampu menguasaihabitatnya. Sifat tumbuhan ini yang mampumenghasilkan bunga dengan waktu mekar yangcukup panjang, dan system reproduksi yangtidak saja mampu menghasilkan biji melaluiperkawinan silang (out-crossing) tetapi jugasecara selfing. Sehingga keterbatasan jumlahindividu pada awal kolonisasi tidak menjadihambatan dalam keberhasilan polinasi.Disamping itu jumlah biji yang dihasilkansangat banyak yang berkisar antara 1410-3265perbuah memfasilitasi keberhasilan penguasaansuatu habitat bahkan dapat menggantikanvegetasi asli. Melastoma malabathricum inijuga merupakan salah satu jenis tumbuhan
invasif yang ditemukan di Hutan Kenali Jambi,dimana jenis ini menempati urutan keduamendominasi komunitas tumbuhan invasif dikawasan hutan tersebut (Susanti dkk., 2013).
Famili dominan berdasarkan besarnyapersentase adalah famili Leguminosae danCompositae dengan persentase masingmasingnya 21,62 % dan 20, 32 %. Sedangkanfamili yang tergolong co-dominan adalahRubiaceae, Poaceae dan Melastomataceaedengan persentase masing-masingnya adalah16,70 %, 14,38% dan 11,62 %. FamiliLeguminosae merupakan salah satu familitumbuhan dikotil yang terpenting dan terbesar.Sebanyak 1800 jenis tumbuhan berbunga yangtergolong kedalam lebih kurang 650 genustergabung kedalam famili Leguminosae (Polhilland Raven, 1981). Famili Asteraceae adalahsalah satu famili tumbuhan yang menjadipenyusun vegetasi penutup lantai hutan di wanawisata Nglimut (Kumolo dan Utami, 2011).Asteraceae merupakan takson tumbuhandengan keanekaragaman jenis yang cukuptinggi. Famili ini terdiri dari 1.100 marga yangmeliputi 20.000 spesies (Cronquist, 1981).
Indeks keanekaragaman jenis tumbuhanasing invasif pada vegetasi semak belukar dikawasan HPPB ini tergolong sedang yaitu 2,89.Beberapa jenis dari tumbuhan asing invasifyang ditemukan pada penelitian ini mempunyaikerapatan yang tinggi dan sebagian jenis jugamempunyai kerapatan yang rendah. Kalausemua jenis yang didapatkan mempunyaikerapatan yang hampir sama besar, makakeanekaragaman jenisnya akan menjadi tinggi(Indriyanto, 2008).
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Pada vegetasi semak belukar kawasan HPPB
ditemukan 1161 individu tumbuhan asinginvasif dari 28 jenis yang tergolong kedalam15 famili, dengan famili dominan adalahLeguminosae dan Compositae.

Solfiyeni, Syamsuardi, Chairul, Wella Yuranti dan Afrida Yulia 250
2. Jenis yang paling dominan adalah Mimosapudica dengan INP 24,16 %. Nilai indekskeanekaragaman tumbuhan asing invasiftergolong sedang yaitu 2,89.
DAFTAR PUSTAKA
Binggeli, P. 1997. An Overview of InvasiveWoody Plants in The Tropic. http://www.agric.wa.gov.au/progserv/plants/weeds.
Cronquist, A. 1981. 1981. An IntegratedSystem of Classification of FloweringPlants. Columbia University Press. NewYork.
Conservation Internasional. 2006. ProsidingLokakarya Penentuan Daerah KunciBiodiversitas Di Sumatra dan DiskusiPemanfaatan Data Bersama, Jejaring,Monitoring Serta Identifikasi KebutuhanKonservasi Pada Masa Mendatang.
Credit Valley Conservation. 2004. A Qunickreference quide to Invasive Plansp.Toronto & Regin Conservation.Diakses tgl 4 Nopember 2013.
Dombois, D. M. and Ellenberg, H. 1974. Aimsand Methods of Vegetation Ecology. JohnWiley & Sons. Toronto.
Indriyanto, 2008. Ekologi Hutan. PT. BumiAksara. Jakarta.
Johnston dan Gillman. 1995. Tree PopulationStudy in Ow Diversity Forest. Gunaya.i.Floristic Composition and Stand Structure.Biodiversity and Conversation 4: 339-362.
Kumolo, F.B dan Utami, S. 2011. Jenis-jenisTumbuhan Anggota Asteraceae di WanaWisata Nglimut Gonoharjo KabupatenKendal Jawa Tengah. Jurnal Bioma.Univesitas Diponegoro. Semarang.
Nasir, N. 2010. Eksplorasi Kelompok Jahe-jahean Hutan Pendidikan dan PenelitianBiologi Universitas Andalas. ProsidingSemirata Bidang MIPA ke 23 tahun 2010Universitas Riau. Pekan Baru.
Nasution, U. 1986. Gulma danPengendaliannya di Perkebunan KaretSumatera Utara dan Aceh. PT. Gramedia.Jakarta.
Odum, 1994. Dasar-dasar Ekologi. GadjahMada University Press. Yogyakarta.
Polhill, R.M. & Raven, P.H. 1981. Advances inLegume Systematics. Royal BotanicGardens, Kew.
Purwono,B., Wardhana ,B.S. Wijanako,K.,Setiowati,E., Kurniawati,D.S. 2002.Keanekaragaman Hayati danPengendalian Jenis Asing Invasif. KantorMentri Lingkungan Hidup RI dan NatureConservasi. Jakarta.
Rahman, M., dkk.1994. Inventarisasi SumberDaya Flora di Hutan Pendidikan danPenelitian Biologi (HPPB) UniversitasAndalas. Padang.
Soemarwoto, O. 1983. Ekologi LingkunganHidup dan Pembangunan. PenerbitDjambatan. Jakarta.
Sunaryo, Tahan, U., Eka, F.T. 2012. JenisTumbuhan Asing Invasif yangMengancam Ekosistem di TamanNasional Gunung Gede Pangrango, ResortBodogol, Jawa Barat. Berk. Penel. Hayati:17 (147 – 152).
Susanti,T., Suraida, Harlis, F. 2013.Keanekaragaman Tumbuhan Invasif diKawasan Taman Hutan Kenali Kota Jambi.Prosiding Semirata FMIPA Unila.Lampung.

ISSN 978-602-14989-0-3 BioETI
Diversitas Ikan Gabus (Channa spp.) di Sumatera Barat danvariasi morfologinyaSYAIFULLAH, ANAS SALSABILA DAN DENNY PUTRI
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian mengenai Diversitas dan Variasi Morfologi Ikan Gabus (Channa spp) di Sumatera Barat yang dikoleksi daribeberapa pasar tradisional di Kabupaten 50 Kota, Agam, Tanah Datar, Solok, dan Padang Pariaman didapatkan sebanyak83 individu ikan yang terdiri dari 81 individu Channa striatus Bloch dan 2 individu Channa lucius Cuvier. Variasimorfologi dari karakter morfometrik dan meristik C. striata dari seluruh individu berdasarkan analisa PCA dan UPGMAmenunjukkan populasi dari Kabupaten Solok dan 50 Kota lebih tinggi variasinya dibandingkan populasi dari Agam,Tanah Datar dan Padang Pariaman. Selain variasi morfologi juga ditemukan adanya dua macam variasi pola warna tubuhyaitu, bagian dorsal polos dan bagian dorsal dengan garis-garis tegak dan berwarna gelap.
Key words: morfologi, ikan gabus, Channa
Pendahuluan
Keanekaragaman ikan yang hidup di perairantropis air tawar dan laut Indonesia merupakanaset nasional yang perlu diinventarisir jenis dankeberadaanya, distribusinya, sifat-sifat hidupnyaserta dinamika populasinya. Hal ini pentingdalam penentuan kebijakan dibidang eksplorasikelautan dan perikananan.
Dalam menjaga kelestarian dankeseimbangan populasi ikan gabus, sertapengenalan biologi dan habitatnya, maka upayaini akan memberikan manfaat bagi masyarakatitu sendiri dalam melestarikan sumber daya ikandan seiring dengan upaya peningkatanpenangkapan secara berkelanjutan.
Morfologi ikan gabus memiliki bentuk tubuhpanjang dengan dasar sirip dorsal dan sirip analpanjang. Beberapa jenis memiliki sirip pelvikdan ada pula yang tidak, bila ada sirip pelvikmemiliki 6 duri sirip lunak dan tidak memilikiduri sirip keras. Sisik ktenoid atau sikloid(Nelson, 1994). Sirip ekor membulat. Punyalubang hidung anterior dan berbentuk tabung.Mulut terminal dan besar. Rahang bawahberkembang, terdapat gigi-gigi berbentuk taring.Pada provomer dan langit-langit ada jenis yangmemiliki gigi dan ada yang tidak (Courtenaydan Williams, 2004).
Kelompok ikan ini diidentifikasikan pertamakali sebagai Channa tahun 1763 oleh Cronovius.Pada literatur tahun 1777, kelompok ikan inijuga ditulis dengan nama yang sama olehScopolli. Pada tahun 1766, Bloch menamakansebagai Ophicephalus. Berdasarkan buktispesimen yang ada kebanyakan ahli iktiologisepakat bahwa kedua nama ini adalah sinomin(Chou dan Ng, 2004). Terdapat 26 jenis Channayang telah teridentifikasi, dengan ukuranpanjang tubuh bervariasi mulai dari 17 cmsampai 1,8 meter (Courtenay dan Williams,2004). Ikan gabus adalah ikan lokal yangmemiliki nilai ekonomis yang tinggi sebagaisalah satu sumber protein, baik dalam bentuksegar atau telah diasinkan (Djajadiredja dkk.,1977).
Daerah sebaran utama jenis-jenis Channaadalah di Asia Tenggara meliputi Indonesiakhususnya Sumatra dan Kalimantan. Di Sumatra,dilaporkan terdapat delapan jenis ikan gabus, diantaranya; Channa bankanensis, C. lucius, C.cyanospilos, C. marulioides, C. melasoma, C.micropeltes, C. striata dan C. pleurophthalama.Semua ikan tersebut dilaporkan diperoleh dariSumatra bagian timur dan selatan. Hanya satujenis, yaitu C. striata yang dilaporkan terdapathampir seluruh tempat di Sumatra. Selain itu adatambahan satu jenis lagi diduga terdapat di

Syaifullah, Anas Salsabila dan Denny Putri 252
Sumatra, yaitu C. melanoptera (Courtenay danWilliams, 2004).
Secara morfologi, sulit untuk mengetahuijenis kelamin ikan gabus, karena tidak adakarakter morfologi eksternal yang dapatdijadikan patokan. Mengetahui jenis kelamindapat dilakukan dengan mengamati strukturanatomi internal, dan melalui tingkah laku ikanini. Dua ekor ikan ditempatkan pada wadahyang sama dan dibatasi kaca pembatas yangdapat dibuka tutup. Bila tiap kali kaca pembatasdibuka ikan menunjukkan perilaku mengancam,berarti jenis kelamin kedua ikan tersebut dapatdipastikan sama. Ikan yang jenis kelaminnyasama canderung bersikap agresif terhadap satusama lain (Chou dan Ng, 2004). SementaraAxelrod dan Vorderwinkler (1983) menulisbahwa pada C. asiatica umumnya jantanberukuran lebih besar dan berwarna lebih cerahdari pada yang betina. Kedewasaan seksual padaC. striata setelah beumur 2 tahun, denganukuran tubuh sekitar 25-30 cm atau lebih(Wijeyaratne, 1989).
Kondisi habitat ikan gabus bervariasi,tergantung jenisnya. Kebanyakan hidup disungai kecil, rawa, sawah, kolam, dan selokan.pH tempat hidup bervariasi, C. bankanensis
lingkungan yang sangat asam sekitar pH 2,8-3,8(Lee dan Ng, 1991 cit. Courtenay dan Williams,2004). Sedangkan tiga spesies, yaitu C. gachua,C. punctata, dan C. striata, toleran terhadaprentang pH yang luas, yaitu pH 4,25 sampai 9,4(Varma, 1979 cit. Courtenay dan Williams,2004). Meskipun tidak begitu signifikan,pertumbuhan ikan ini baik pada kondisi yangsedikit asam, karena efisiensi pencernaannyameningkat pada suasana lingkungan asam(Wijeyaratne, 1989).
Anak ikan gabus sebagian besar memakanplankton, khususnya zooplankton. Saat juvenil,makanan berganti menjadi larva serangga,udang-udangan kecil, serta anak ikan jenis lain.Ikan dewasa memangsa ikan lain, udang-udangan, katak, kepiting, anak burung, bahkanmamalia kecil. Saat kekurangan makanan, ikanini bersifat kanibal (Courtenay dan Williams,
2004). Makan yang dimakan biasanya sebesarbukaan mulut, yaitu 16-20% dari panjangstandar (Manzano, 1989).
Ikan gabus sebagai sumber makanan jugamempunyai nilai kesehatan, karena dalamdagingnya terkandung albumin yang tinggi,yang tidak terdapat pada ikan konsumsi lainseperti; ikan lele, nila, mas, gurami dansebagainya. Serum albumin dapat digunakansebagai obat mempercepat penyembuhan lukabekas operasi (Anonimous, 2003b). Penggunaanekstrak dari ikan gabus sangat baik, karenaselain lebih cepat sembuh juga lebih murah (Lia,2003).
Anak ikan gabus, ikut berperan secara tidaklangsung pada kehidupan alamiah, karenamemakan jentik-jentik nyamuk yang merupakanmusuh alami manusia. Namun akhir-akhir inibanyak anak ikan gabus ditangkap untukdijadikan makanan ikan gurami jenis Lou Han.Penangkapan anak ikan dikhawatirkan akanmenurunkan populasi ikan gabus dimasa depan,juga meningkatkan populasi nyamuk(Anonimous, 2003a).
Penelitian ini bertujuan untuk mengetahuidiversitas ikan gabus dan variasi morfologinyadi beberapa tempat di Sumatera Barat.Selanjutnya diharapkan penelitian ini dapatbermanfaat menambah informasikeanekaragaman ikan yang ada di Sumatra Barat,serta dapat menunjang program budidaya ikanini sebagai salah satu komoditas ekonomiSumatera Barat.
BAHAN DAN METODE
Penelitian ini menggunakan metoda deskriptifdengan observasi dan koleksi dari hasiltangkapan masyarakat, yang dijual di pasartradisional setempat. Penentuan lokasiberdasarkan informasi masyarakat mengenaikeberadaan ikan gabus dan nama daerah ikangabus yang berbeda. Dari hal di atas makaditentukan sebagai berikut; Kabupaten 50 Kota,Kabupaten Agam, Kabupaten Tanah Datar,Kabupaten Solok, dan Kabupaten PadangPariaman. Pengidentifikasian berdasarkan

Syaifullah, Anas Salsabila dan Denny Putri 253
deskripsi Courtenay dan Williams (2004) danVierke (2004). Selain itu juga dilengkapi dengankajian morfometrik pengukuran terhadapbeberapa karakter kuantitatif dan kualitatifmenurut Caillet, Love, dan Ebeling (1986),seperti Gambar 1 dibawah ini.
Gambar 1. Skema pengukuran morfometrik ikangabus
Keterangan:a. sisik sepanjang gurat sisi, b. sisik di atas gurat sisi, c.sisik di bawah gurat sisi, d. dan sisik di sekeliling batangekor. 1. panjang standar, 2. tinggi tubuh, 3. tinggi batangekor, 4. panjang batang ekor, 5. panjang nape, 6.panjang dasar sirip dorsal, 7. panjang dasar sirip anal, 8.tinggi sirip dorsal, 9. tinggi sirip anal, 10. panjang sirippektoral, 11. panjang sirip pelvik, 12. panjang duri siripdorsal yang terpanjang, 13. panjang kepala, 14. lebarkepala, 15. panjang moncong, 16. lebar suborbital, 17.diameter mata, 18. panjang rahang atas, dan 19. lebar‘gape’.
Data hasil pengamatan karakter morfologisemua populasi Channa spp dari berbagai lokasidianalisis dengan menggunakan analisiskomponen prinsip (PCA) untuk mengetahui polapenyebaran Channa spp dengan menggunakanprogram MVSP 3.1 dan Analisis UPGMA untukmengetahui jarak euclidian dilakukan denganprogram NTSYS ver.2.0.2i dan (Rohlf, 2001).
HASIL DAN PEMBAHASAN
Ikan gabus merupakan salah satu plasma nutfahsumber daya ikan di Sumatra Barat. Dari hasilkoleksi dan identifikasi yang telah dilakukan,didapatkan bahwa terdapat dua jenis ikan gabus
yang diperdagangkan, yaitu C. striata Bloch.dan C. lucius Cuvier.
C. striata yang ditemukan pada sebagianpasar-pasar tradisional di Sumatra Barat berasaldari berbagai lokasi. Sedangkan C. lucius hanyaditemukan di pasar tradisional Kabupaten Solok.C. striata pada daerah yang berbeda memilikinama daerah yang berbeda-beda, sementaradiketahui hanya satu nama daerah untuk Clucius.
Jumlah C. striata dan C. lucius yangditemukan merupakan hasil tangkapan yangdiperdagangkan dalam jumlah yang terbatas,karena ikan ini belum dibudidayakan secarakhusus. Harga jual ikan gabus jauh lebih tinggidaripada harga ikan air tawar lain yangdiperdagangkan. Ikan ini tidak tersedia di pasarsepanjang waktu. Jenis C. striata kebanyakanditangkap dari kolam budidaya ikan mas,sedangkan C. lucius ditangkap dari sungaisekitar danau Singkarak. Ikan gabus dewasayang berukuran sekitar 25 cm atau lebih,kebanyakan ditemukan berpasangan. Hal inisesuai dengan laporan Courtenay dan Williams(2004) yang menyatakan bahwa ikan gabusbersifat monogami dan berpasangan sepanjangtahun.
Masyarakat menganggap C. striata sebagaihama karena memakan anak ikan mas di kolambudidaya, sehingga banyak ikan ini yanglangsung dibunuh bila ditemukan. Hal ini amatdisayangkan karena berdasarkan penelitianManzaro (1988), meski C striata memakan anakikan budidaya, tapi secara keseluruhanmeningkatkan populasi ikan dewasa, yangberarti pula meningkatkan produksi ikanbudidaya. Philipina bahkan telah menerapkansistem pemeliharaan ikan budidaya secarapolikultur, di mana ikan tilapia (Oreochromis
mossambicus) yang dibudidayakan dipeliharabersama C striata.
Data kuantitatif hasil pengukuran masing-masing variabel karakter pada C. striata
menunjukkan sedikit perbedaan antara hasilyang diperoleh dengan yang telahdideskripsikan oleh Bloch, seperti yang dikutip

Syaifullah, Anas Salsabila dan Denny Putri 254
dari Courteney dan Wiliam (2004). Perbedaantersebut yaitu pada jumlah duri lunak siripdorsal (deskipsi Bloch 37-46, hasil koleksi 35-44), duri sirip pektoral (deskripsi Bloch 15-17,hasil koleksi 13-17), serta jumlah sisiksepanjang gurat sisi (deskripsi Bloch 50-57,hasil koleksi 48-58). Cailliet et. al. (1986)menyatakan karena tidak mungkin mengkoleksiseluruh individu untuk dijadikan variabel, makadata yang ada dikatakan sebagai populasisampel bukan populasi biologi. Populasi sampeltersebut diharapkan mendekati populasi biologiyang ada. Caillit et. al. (1986) juga menyatakandengan adanya perbedaan hasil observasidengan deskripsi yang telah ada bisa sajamengacu pada jenis baru atau jenis yang samanamun belum terobservasi sebelumnya. Biladilihat dari data kuantitatif sampel yangbersangkutan keseluruhan, tidak menunjukkanperbedaan maupun pengelompokan. Semuasampel menunjukkan bahwa individu-individuyang diperoleh tersebut adalah C. striata.
Berdasarkan dari hasil pengukuran spesimenyang diperoleh, deskripsi dari ikan C striataadalah sebagai berikut: duri lunak dorsal 35-44;duri lunak anal 24-27; total duri pektoral 13-17.Sisik sepanjang gurat sisi 48-58; sisik di atasgurat sisi 4-5; sisik di bawah gurat sisi 8-13;sisik di sekeliling batang ekor 8-13. Panjangstandar 113-414 mm. Tinggi tubuh 12,1-20,3%SL. Tinggi batang ekor 8,3-11,5% SL. Panjangbatang ekor 5,1-10,7% SL. Panjang nape 8,1-15,4% SL. Panjang dasar sirip dorsal 57,1-64,3 % SL. Panjang dasar sirip anal 33,8-41,7%SL. Tinggi sirip dorsal 5,1-9,9% SL. Tinggisirip anal 3,5-7,8% SL. Panjang sirip pektoral15,6-21,9% SL. Panjang sirip pelvik 9,9-14,8%SL. Panjang duri sirip dorsal terpanjang 9,3-14,4% SL. Panjang kepala 26,6-35,0% SL.Lebar kepala 9,6-13,2% SL. Panjang moncong3.0-7,5% SL. Lebar suborbit 1,1-2,5% SL.Panjang. orbit ke sudut preoperkulum 4,3-8,7%SL. Diameter mata 2,8-5,7% SL. Panjangrahang atas 10,2-15,8% SL. Lebar gape 15,2-19,5% SL. Pola warna tubuh belang dan adapula yang polos. Warna dasar bagian dorsal
kehitaman, kecoklatan atau kelabu. Warna dasarventral umummya putih, dengan atau tanpabercak. Pada prevomer dan palatin tidakterdapat taring, hanya rangkaian gigi-gigi kecilyang runcing, pada mandibula terdapat beberapagigi taring.
Variasi individu pada bentuk tubuh danmorfologi sirip sering berhubungan denganhabitat yang ditempati populasi ikan air tawar.Meski banyak korelasi antara karakter morfologidengan habitat yang ditempati, belum ada buktiempiris langsung yang mendukung adanyakorelasi tersebut secara fungsional (Boily danMagnan, 2002). Variasi morfologi merupakaninteraksi plastisitas fenotip dengan keragamangenetik. Pada karakter tertentu, variasimorfologi bisa saja muncul cenderung akibatplastisitas fenotip, sementara pada karakterlainnya variasi morfologi muncul cemderungakibat keragaman genetik (Mittlebach, Osenberg,dan Wainwright, 1999).
Gambar 2. Foto C. striata dengan pola warna belang danfoto C. striata dengan pola warna poloskehitaman
Sedangkan morfologi ikan C. lucius,berdasarkan dari hasil spesimen yang diperoleh,deskripsi C. licius di Sumatra Barat adalahsebagai berikut: duri lunak dorsal 39-40; durilunak anal 28-29; total duri pektoral 16. Sisiksepanjang gurat sisi 60; sisik di atas gurat sisi5,5; sisik di bawah gurat sisi 14; sisik disekeliling batang ekor 11. Panjang standar 178-251 mm. Tinggi tubuh 18,6-19,7% SL. Tinggibatang ekor 12,3-12,4% SL. Panjang batangekor 6,2-6,7% SL. Panjang nape 10,0-11,5% SL.Panjang dasar sirip dorsal 60,7-62,9 % SL.Panjang dasar sirip anal 43,8-44,9% SL. Tinggi
Syaifullah, Anas Salsabila dan Denny Putri 254
dari Courteney dan Wiliam (2004). Perbedaantersebut yaitu pada jumlah duri lunak siripdorsal (deskipsi Bloch 37-46, hasil koleksi 35-44), duri sirip pektoral (deskripsi Bloch 15-17,hasil koleksi 13-17), serta jumlah sisiksepanjang gurat sisi (deskripsi Bloch 50-57,hasil koleksi 48-58). Cailliet et. al. (1986)menyatakan karena tidak mungkin mengkoleksiseluruh individu untuk dijadikan variabel, makadata yang ada dikatakan sebagai populasisampel bukan populasi biologi. Populasi sampeltersebut diharapkan mendekati populasi biologiyang ada. Caillit et. al. (1986) juga menyatakandengan adanya perbedaan hasil observasidengan deskripsi yang telah ada bisa sajamengacu pada jenis baru atau jenis yang samanamun belum terobservasi sebelumnya. Biladilihat dari data kuantitatif sampel yangbersangkutan keseluruhan, tidak menunjukkanperbedaan maupun pengelompokan. Semuasampel menunjukkan bahwa individu-individuyang diperoleh tersebut adalah C. striata.
Berdasarkan dari hasil pengukuran spesimenyang diperoleh, deskripsi dari ikan C striataadalah sebagai berikut: duri lunak dorsal 35-44;duri lunak anal 24-27; total duri pektoral 13-17.Sisik sepanjang gurat sisi 48-58; sisik di atasgurat sisi 4-5; sisik di bawah gurat sisi 8-13;sisik di sekeliling batang ekor 8-13. Panjangstandar 113-414 mm. Tinggi tubuh 12,1-20,3%SL. Tinggi batang ekor 8,3-11,5% SL. Panjangbatang ekor 5,1-10,7% SL. Panjang nape 8,1-15,4% SL. Panjang dasar sirip dorsal 57,1-64,3 % SL. Panjang dasar sirip anal 33,8-41,7%SL. Tinggi sirip dorsal 5,1-9,9% SL. Tinggisirip anal 3,5-7,8% SL. Panjang sirip pektoral15,6-21,9% SL. Panjang sirip pelvik 9,9-14,8%SL. Panjang duri sirip dorsal terpanjang 9,3-14,4% SL. Panjang kepala 26,6-35,0% SL.Lebar kepala 9,6-13,2% SL. Panjang moncong3.0-7,5% SL. Lebar suborbit 1,1-2,5% SL.Panjang. orbit ke sudut preoperkulum 4,3-8,7%SL. Diameter mata 2,8-5,7% SL. Panjangrahang atas 10,2-15,8% SL. Lebar gape 15,2-19,5% SL. Pola warna tubuh belang dan adapula yang polos. Warna dasar bagian dorsal
kehitaman, kecoklatan atau kelabu. Warna dasarventral umummya putih, dengan atau tanpabercak. Pada prevomer dan palatin tidakterdapat taring, hanya rangkaian gigi-gigi kecilyang runcing, pada mandibula terdapat beberapagigi taring.
Variasi individu pada bentuk tubuh danmorfologi sirip sering berhubungan denganhabitat yang ditempati populasi ikan air tawar.Meski banyak korelasi antara karakter morfologidengan habitat yang ditempati, belum ada buktiempiris langsung yang mendukung adanyakorelasi tersebut secara fungsional (Boily danMagnan, 2002). Variasi morfologi merupakaninteraksi plastisitas fenotip dengan keragamangenetik. Pada karakter tertentu, variasimorfologi bisa saja muncul cenderung akibatplastisitas fenotip, sementara pada karakterlainnya variasi morfologi muncul cemderungakibat keragaman genetik (Mittlebach, Osenberg,dan Wainwright, 1999).
Gambar 2. Foto C. striata dengan pola warna belang danfoto C. striata dengan pola warna poloskehitaman
Sedangkan morfologi ikan C. lucius,berdasarkan dari hasil spesimen yang diperoleh,deskripsi C. licius di Sumatra Barat adalahsebagai berikut: duri lunak dorsal 39-40; durilunak anal 28-29; total duri pektoral 16. Sisiksepanjang gurat sisi 60; sisik di atas gurat sisi5,5; sisik di bawah gurat sisi 14; sisik disekeliling batang ekor 11. Panjang standar 178-251 mm. Tinggi tubuh 18,6-19,7% SL. Tinggibatang ekor 12,3-12,4% SL. Panjang batangekor 6,2-6,7% SL. Panjang nape 10,0-11,5% SL.Panjang dasar sirip dorsal 60,7-62,9 % SL.Panjang dasar sirip anal 43,8-44,9% SL. Tinggi
Syaifullah, Anas Salsabila dan Denny Putri 254
dari Courteney dan Wiliam (2004). Perbedaantersebut yaitu pada jumlah duri lunak siripdorsal (deskipsi Bloch 37-46, hasil koleksi 35-44), duri sirip pektoral (deskripsi Bloch 15-17,hasil koleksi 13-17), serta jumlah sisiksepanjang gurat sisi (deskripsi Bloch 50-57,hasil koleksi 48-58). Cailliet et. al. (1986)menyatakan karena tidak mungkin mengkoleksiseluruh individu untuk dijadikan variabel, makadata yang ada dikatakan sebagai populasisampel bukan populasi biologi. Populasi sampeltersebut diharapkan mendekati populasi biologiyang ada. Caillit et. al. (1986) juga menyatakandengan adanya perbedaan hasil observasidengan deskripsi yang telah ada bisa sajamengacu pada jenis baru atau jenis yang samanamun belum terobservasi sebelumnya. Biladilihat dari data kuantitatif sampel yangbersangkutan keseluruhan, tidak menunjukkanperbedaan maupun pengelompokan. Semuasampel menunjukkan bahwa individu-individuyang diperoleh tersebut adalah C. striata.
Berdasarkan dari hasil pengukuran spesimenyang diperoleh, deskripsi dari ikan C striataadalah sebagai berikut: duri lunak dorsal 35-44;duri lunak anal 24-27; total duri pektoral 13-17.Sisik sepanjang gurat sisi 48-58; sisik di atasgurat sisi 4-5; sisik di bawah gurat sisi 8-13;sisik di sekeliling batang ekor 8-13. Panjangstandar 113-414 mm. Tinggi tubuh 12,1-20,3%SL. Tinggi batang ekor 8,3-11,5% SL. Panjangbatang ekor 5,1-10,7% SL. Panjang nape 8,1-15,4% SL. Panjang dasar sirip dorsal 57,1-64,3 % SL. Panjang dasar sirip anal 33,8-41,7%SL. Tinggi sirip dorsal 5,1-9,9% SL. Tinggisirip anal 3,5-7,8% SL. Panjang sirip pektoral15,6-21,9% SL. Panjang sirip pelvik 9,9-14,8%SL. Panjang duri sirip dorsal terpanjang 9,3-14,4% SL. Panjang kepala 26,6-35,0% SL.Lebar kepala 9,6-13,2% SL. Panjang moncong3.0-7,5% SL. Lebar suborbit 1,1-2,5% SL.Panjang. orbit ke sudut preoperkulum 4,3-8,7%SL. Diameter mata 2,8-5,7% SL. Panjangrahang atas 10,2-15,8% SL. Lebar gape 15,2-19,5% SL. Pola warna tubuh belang dan adapula yang polos. Warna dasar bagian dorsal
kehitaman, kecoklatan atau kelabu. Warna dasarventral umummya putih, dengan atau tanpabercak. Pada prevomer dan palatin tidakterdapat taring, hanya rangkaian gigi-gigi kecilyang runcing, pada mandibula terdapat beberapagigi taring.
Variasi individu pada bentuk tubuh danmorfologi sirip sering berhubungan denganhabitat yang ditempati populasi ikan air tawar.Meski banyak korelasi antara karakter morfologidengan habitat yang ditempati, belum ada buktiempiris langsung yang mendukung adanyakorelasi tersebut secara fungsional (Boily danMagnan, 2002). Variasi morfologi merupakaninteraksi plastisitas fenotip dengan keragamangenetik. Pada karakter tertentu, variasimorfologi bisa saja muncul cenderung akibatplastisitas fenotip, sementara pada karakterlainnya variasi morfologi muncul cemderungakibat keragaman genetik (Mittlebach, Osenberg,dan Wainwright, 1999).
Gambar 2. Foto C. striata dengan pola warna belang danfoto C. striata dengan pola warna poloskehitaman
Sedangkan morfologi ikan C. lucius,berdasarkan dari hasil spesimen yang diperoleh,deskripsi C. licius di Sumatra Barat adalahsebagai berikut: duri lunak dorsal 39-40; durilunak anal 28-29; total duri pektoral 16. Sisiksepanjang gurat sisi 60; sisik di atas gurat sisi5,5; sisik di bawah gurat sisi 14; sisik disekeliling batang ekor 11. Panjang standar 178-251 mm. Tinggi tubuh 18,6-19,7% SL. Tinggibatang ekor 12,3-12,4% SL. Panjang batangekor 6,2-6,7% SL. Panjang nape 10,0-11,5% SL.Panjang dasar sirip dorsal 60,7-62,9 % SL.Panjang dasar sirip anal 43,8-44,9% SL. Tinggi

Syaifullah, Anas Salsabila dan Denny Putri 255
sirip dorsal 5,8-7,0% SL. Tinggi sirip anal 4,8-6,4% SL. Panjang sirip pektoral 16,0-17,0 % SL.Panjang sirip pelvik 10,9-11,1% SL. Panjangduri sirip dorsal terpanjang 12,0-12,9% SL.Panjang kepala 31,6-31,7% SL. Lebar kepala11,5-11,7% SL. Panjang moncong 3.5-4,8% SL.Lebar suborbit 1,2-1,6% SL. Panjang. orbit kesudut preoperkulum 4,5-4,6% SL. Diametermata 3,3-3,6% SL. Panjang rahang atas 7,7-9,6% SL. Lebar gape 16,4-16,7% SL. Polawarna tubuh belang dengan warna dasar kuning.Terdapat 6 bulatan hitam memanjang tersusundari operkulum hingga pangkal sirip ekor. Padapalatin dan mandibula terdapat gigi-gigi runcing.
Gambar 3. Foto C. lucius Cuvier. dari Sumani (Sungai)
Dari analisa morfologi ikan gabus C. striatadari beberapa lokasi dengan menggunakanPrinsipal Component Analisis (PCA) didapatkanadanya individu-individu ikan gabus C. striatayang berasal dari Solok dan sebagian dari Kab.50 Kota mempunyai variasi morfologi lebih luasdibandingkan dengan individu dari lokasilainnya (lihat Gambar 4). Sedangkan populasiikan yang berasal dari lokasi lainnya salingtumpang tindih, hal ini menunjukkan persamaankarakter morfologi dari individu-individu ikangabus yang tersebar di Sumatera Barat. Olehkarenanya diversitas morfologi yang berasal dariKabupaten Solok dan 50 Kota inilah yang perludiamati lebih jauh lagi, sehingga pemisahanbeberapa individu dari populasi lainnyamerupakan indikasi variasi morfologi yangmenunjukkan adanya plastisitas morfologi darikarakter-karakter yang diuji. Apabila ditemukanadanya variasi morfologi dari ikan gabus C.striata, maka perlu dilakukan penyelamatansumber plasma nutfah untuk dikembangkansebagai ikan budidaya.
Gambar 4. Analisa PCA yang menunjukkanadanya variasi morfologi pd ikangabus (C. striata) yang dikoleksidari beberapa lokasi di SumateraBarat.
Karakter morfologi yang berperan dalammemisahkan individu dalam analsis PCA adalahukuran panjang standard, panjang batang ekor,tinggi sirip dorsal, tinggi sirip anal, panjangsirip pektoral, lebar kepala, diameter mata, dankarakter meristik lainnya sepeti: sirip lemahanal, sisik diatas gurat sisi, sisik di bawah guratsisi, sisik sebelum dorsal, sisik batang ekor.
Selanjutnya analisa UPGMA terhadapkarakter morfologi ikan gabus C. striata dapatdilihat pada dendogram (Gambar 5), bahwa ikangabus yang berasal dari Kabupaten Pasaman,Pesisir Selatan dan Agam mempunyaikedekatan morfologi. Sedangkan individu dariKabupaten 50 Kota dan Tanah Datar, kemudianpopulasi Padang dan Solok mempunyaikemiripan morfologi yang lebih jauh.
Gambar 5. Dendogram dari keseluruhan populasiChanna striata di beberapa lokasi diSumatera Barat.
Syaifullah, Anas Salsabila dan Denny Putri 255
sirip dorsal 5,8-7,0% SL. Tinggi sirip anal 4,8-6,4% SL. Panjang sirip pektoral 16,0-17,0 % SL.Panjang sirip pelvik 10,9-11,1% SL. Panjangduri sirip dorsal terpanjang 12,0-12,9% SL.Panjang kepala 31,6-31,7% SL. Lebar kepala11,5-11,7% SL. Panjang moncong 3.5-4,8% SL.Lebar suborbit 1,2-1,6% SL. Panjang. orbit kesudut preoperkulum 4,5-4,6% SL. Diametermata 3,3-3,6% SL. Panjang rahang atas 7,7-9,6% SL. Lebar gape 16,4-16,7% SL. Polawarna tubuh belang dengan warna dasar kuning.Terdapat 6 bulatan hitam memanjang tersusundari operkulum hingga pangkal sirip ekor. Padapalatin dan mandibula terdapat gigi-gigi runcing.
Gambar 3. Foto C. lucius Cuvier. dari Sumani (Sungai)
Dari analisa morfologi ikan gabus C. striatadari beberapa lokasi dengan menggunakanPrinsipal Component Analisis (PCA) didapatkanadanya individu-individu ikan gabus C. striatayang berasal dari Solok dan sebagian dari Kab.50 Kota mempunyai variasi morfologi lebih luasdibandingkan dengan individu dari lokasilainnya (lihat Gambar 4). Sedangkan populasiikan yang berasal dari lokasi lainnya salingtumpang tindih, hal ini menunjukkan persamaankarakter morfologi dari individu-individu ikangabus yang tersebar di Sumatera Barat. Olehkarenanya diversitas morfologi yang berasal dariKabupaten Solok dan 50 Kota inilah yang perludiamati lebih jauh lagi, sehingga pemisahanbeberapa individu dari populasi lainnyamerupakan indikasi variasi morfologi yangmenunjukkan adanya plastisitas morfologi darikarakter-karakter yang diuji. Apabila ditemukanadanya variasi morfologi dari ikan gabus C.striata, maka perlu dilakukan penyelamatansumber plasma nutfah untuk dikembangkansebagai ikan budidaya.
Gambar 4. Analisa PCA yang menunjukkanadanya variasi morfologi pd ikangabus (C. striata) yang dikoleksidari beberapa lokasi di SumateraBarat.
Karakter morfologi yang berperan dalammemisahkan individu dalam analsis PCA adalahukuran panjang standard, panjang batang ekor,tinggi sirip dorsal, tinggi sirip anal, panjangsirip pektoral, lebar kepala, diameter mata, dankarakter meristik lainnya sepeti: sirip lemahanal, sisik diatas gurat sisi, sisik di bawah guratsisi, sisik sebelum dorsal, sisik batang ekor.
Selanjutnya analisa UPGMA terhadapkarakter morfologi ikan gabus C. striata dapatdilihat pada dendogram (Gambar 5), bahwa ikangabus yang berasal dari Kabupaten Pasaman,Pesisir Selatan dan Agam mempunyaikedekatan morfologi. Sedangkan individu dariKabupaten 50 Kota dan Tanah Datar, kemudianpopulasi Padang dan Solok mempunyaikemiripan morfologi yang lebih jauh.
Gambar 5. Dendogram dari keseluruhan populasiChanna striata di beberapa lokasi diSumatera Barat.
Syaifullah, Anas Salsabila dan Denny Putri 255
sirip dorsal 5,8-7,0% SL. Tinggi sirip anal 4,8-6,4% SL. Panjang sirip pektoral 16,0-17,0 % SL.Panjang sirip pelvik 10,9-11,1% SL. Panjangduri sirip dorsal terpanjang 12,0-12,9% SL.Panjang kepala 31,6-31,7% SL. Lebar kepala11,5-11,7% SL. Panjang moncong 3.5-4,8% SL.Lebar suborbit 1,2-1,6% SL. Panjang. orbit kesudut preoperkulum 4,5-4,6% SL. Diametermata 3,3-3,6% SL. Panjang rahang atas 7,7-9,6% SL. Lebar gape 16,4-16,7% SL. Polawarna tubuh belang dengan warna dasar kuning.Terdapat 6 bulatan hitam memanjang tersusundari operkulum hingga pangkal sirip ekor. Padapalatin dan mandibula terdapat gigi-gigi runcing.
Gambar 3. Foto C. lucius Cuvier. dari Sumani (Sungai)
Dari analisa morfologi ikan gabus C. striatadari beberapa lokasi dengan menggunakanPrinsipal Component Analisis (PCA) didapatkanadanya individu-individu ikan gabus C. striatayang berasal dari Solok dan sebagian dari Kab.50 Kota mempunyai variasi morfologi lebih luasdibandingkan dengan individu dari lokasilainnya (lihat Gambar 4). Sedangkan populasiikan yang berasal dari lokasi lainnya salingtumpang tindih, hal ini menunjukkan persamaankarakter morfologi dari individu-individu ikangabus yang tersebar di Sumatera Barat. Olehkarenanya diversitas morfologi yang berasal dariKabupaten Solok dan 50 Kota inilah yang perludiamati lebih jauh lagi, sehingga pemisahanbeberapa individu dari populasi lainnyamerupakan indikasi variasi morfologi yangmenunjukkan adanya plastisitas morfologi darikarakter-karakter yang diuji. Apabila ditemukanadanya variasi morfologi dari ikan gabus C.striata, maka perlu dilakukan penyelamatansumber plasma nutfah untuk dikembangkansebagai ikan budidaya.
Gambar 4. Analisa PCA yang menunjukkanadanya variasi morfologi pd ikangabus (C. striata) yang dikoleksidari beberapa lokasi di SumateraBarat.
Karakter morfologi yang berperan dalammemisahkan individu dalam analsis PCA adalahukuran panjang standard, panjang batang ekor,tinggi sirip dorsal, tinggi sirip anal, panjangsirip pektoral, lebar kepala, diameter mata, dankarakter meristik lainnya sepeti: sirip lemahanal, sisik diatas gurat sisi, sisik di bawah guratsisi, sisik sebelum dorsal, sisik batang ekor.
Selanjutnya analisa UPGMA terhadapkarakter morfologi ikan gabus C. striata dapatdilihat pada dendogram (Gambar 5), bahwa ikangabus yang berasal dari Kabupaten Pasaman,Pesisir Selatan dan Agam mempunyaikedekatan morfologi. Sedangkan individu dariKabupaten 50 Kota dan Tanah Datar, kemudianpopulasi Padang dan Solok mempunyaikemiripan morfologi yang lebih jauh.
Gambar 5. Dendogram dari keseluruhan populasiChanna striata di beberapa lokasi diSumatera Barat.

Syaifullah, Anas Salsabila dan Denny Putri 256
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Diversitas ikan gabus yang ditemukan pada
beberapa lokasi di Sumatera Barat adalahChanna striata Bloch dan C. lucius Cuvier.
2. Karakter morfologi yang bevariasi pada ikangabus ini, karakter morfometrik (tinggi tubuh,panjang batang ekor, panjang dasar siripanal , lebar “gape”) dan karakter meristik(sirip lunak ekor, warna tubuh, sisiksepanjang garis, sisik atas gurat sisi, sisikbawah gurat sisi, sisik sekitar batang ekor).
Ucapan terima kasihPenelitian ini merupakan upaya untukmenambah informasi diversitas spesies ikan diSumatera Barat, terima kasih kepada DennyPutri yang telah mengkoleksi ikan gabus dariberbagai pasar tradisional untuk penelitianskripsinya.
DAFTAR PUSTAKA
Anonimous. 2003a. Kompas Cyber Media: IkanLou Han atau Nyamuk Demam Berdarah.http://www.kompas.com/kompas-cetak/0305/28/daerah/336316. htm.
Anonimous. 2003b. Kompas Cyber Media:Potensi Serum Albumin dari Ikan Gabus.http://www.kompas.com/kompas-cetak/0301/04/jatim/70587.htm.
Anonimous. 2005. Channa/Frankenfish.http://www.channa.info/. 23 April 2006.
Axelrod, H. R. dan W. Vorderwinkler. 1983.Encyclopedia of Tropical Fish. T. F. H.Publication, Inc. Ltd. US.
Boesch, D. F. 2002a. Snakehead scientificadvisory panel report recommendscommon herbicides ans a pesticide toerredicate fish from crofton pond.http://www.dnr.state.md.us/dnrnews/pressrelease2002/072602a.html. 27 Ap-ril 2006.
Boesch, D. F. 2002b. Northern Snakeheads Fish.http://www.dnr.state.md.us/fishe ries/snakeheadinfosheet.html. 27 April 2006.
Boily, P. dan P. Magnan. 2002. RelationshipBetween Individual Variation InMorphological Characters and SwimmingCosts In Brook Charr (Salvelinusfontinalis) and Yellow Perch (Perca
flavescens). The Journal of ExperimentalBiology 205: 1031-1036.
Chou, L.M. dan P.K.L. Ng. 2004. Snakeheads(Pisces: Channidae): Natural History,Biology and Economic Importance.http://www.snakeheads.org. 23 April 2006.
Courtenay,W.R.Jr. dan J.D.Williams. 2004.Snakehead (Pisces, Channidae) A Bio-logical Synopsis and Risk Assessment.http://www.usgs .gov. 23 April 2006.
Caillet, G. M., M. S. Love, dan A.W. Ebeling.1986. Fishes A Field and LaboratoryManual on Their Structure, Identification,and Natural History. Waveland Press.
Djajadiredja, R., S. Hatimah, dan Z. Arifin.1977. Buku pengenalan sumber perikanandarat bagian 1 (jenis-jenis ikan ekonomispenting). Direktorat Jendral PerikananDepartemen Pertanian.
Lagler, K. F., J. E. Bardach, dan R. R. Miller.1962. Ichthyology. John Wiley and Sons,
Moyle, P. B. dan J. J. Cech, Jr. 2000. Fishes AnIntroduction to Ichthyology 4th edition.Prentice Hall. USA.
Manzano, V. B. 1989. Polyculture Grouper(Ephinephelus tauvina) and Tilapia(Oreochromis mossambicus) In Brackishwater Pond. In: Huisman, E. A., N.Zonneveld, dan A. H. M. Bouwmans (Eds.).Aquaculture Research In Asia ManagementTechniques And Nutrition. PudocWageninger. Netherland.
Mittlebach, G. G., C. W. Osenberg, dan P. C.Wainwright. 1999. Variation in feedingmorphology between pumpkinseedpopulations: phenotypic or evolution?Evolutionary Ecology Reseach 1: 111-128.
Nelson, J.S. 1994 Fishes of the World. 3rdedition. John Wiley & Sons. New York.
Rohlf, F.J., 2001. NTSyst. NumericalTaxonomy and Multivariate AnalysisSystem Version 2.0.2. Applied Biostatistic
Tan, H. H. dan P. K. L. Ng. 2005. The LabyrinthFishes (Teleostei: Anabantoidei,Channoidei) of Sumatra, Indonesia. TheRaffles Buletin of Zoology 13: 115-138.
Wijeyaratne, M. J. S. 1989. Food Intake andFood Conversion Efficiency of TheSnakehead Ophichepalus striatus Bloch InA Peaty Swamp In Sri Langka In: Huisman,E. A., N. Zonneveld, dan A. H. M.Bouwmans (Eds.). Aquaculture ResearchIn Asia Management Techniques AndNutrition. Pudoc Wageninger. Netherland.

ISSN 978-602-14989-0-3 BioETI
Ekstrak limbah jus Jeruk sebagai feed additive alamiFakultas Peternakan Universitas JambiUCOP HAROEN
Fakultas Peternakan, Universitas JambiE-mail:
ABSTRAKPenggunaan antibiotik sintetik dalam dunia peternakan dapat menimbulkan dampak negatif bagi kesehatan ternak danmanusia. Antibiotik sintetik dapat mengakibatkan residu bahan kimia berbahaya dalam produk yang dihasilkan danmenyebabkan resistensi bakteri-bakteri berbahaya yang terdapat di dalam tubuh ternak. Residu dari feed additive tersebutakan terbawa dalam produk-produk ternak seperti daging dan telur dan ini akan berbahaya bagi konsumen yangmengkonsumsinya. Oleh karena itu akhir-akhir ini permintaan konsumen telah mengarah pada penggunaan produk ternakyang aman dan berkualitas melalui penggunaan bahan-bahan alam serta menghindari penggunaan feed additive darisintetis (Patra 2011). Ekstrak limbah jus merupakan salah satu bahan yang dapat digunakan sebagai feed additive alamisebagai pengurangan pemakaian feed additive sintetis. Limbah jus jeruk mengandung senyawa-senyawa aktif yangberguna untuk kesehatan ternak antara lain limonoid (dominan), kumarin relative tinggi (+++), sementara alkaloid,flavonoid, steroid, triterpenoid, fenolik relative sedang (++) (Haroen., 2013). Potensi limbah jus jeruk sebagai sumberfeed additive alami sangat besar. Berdasarkan produksi jeruk tiap tahun di Provinsi Jambi cukup tinggi yaitu denganproduksi sebesar 3047 ton dan meningkat mencapai 35.702 ton. Hal ini ketersediaan limbah jus jeruk di Provinsi Jambicukup besar (Litbang Pertanian, 2010). Penelitian bertujuan untuk memurnikan senyawa aktif limonoid yang dominanpada ekstrak limbah jus jeruk, menentukan pelarut yang cocok untuk mendapatkan ekstrak jus jeruk yang mengandungsenyawa limonoid terbanyak, mengukur aktivitas antibakteri senyawa limonoid dari ekstrak limbah jus.
Key words: Purifikasi, limonoid, etilasetat, methanol, n-heksana, limbah jus jeruk
Pendahuluan
Pada sistim produksi ternak, penggunaan feedadditive sintetis seperti antibiotik telahdicobakan terutama untuk tujuan peningkatanpertumbuhan untuk perbaikan konsumsi danefisiensi pakan serta pencegahan penyakit(Patra, 2011). Penggunaan feed additive untuktujuan tersebut, telah menjadi fokus perhatianoleh masyarakat yang peduli terhadap asfekkesehatan, seperti adanya residu kimia yangterdapat pada daging dan telur serta resistensiterhadap antibiotik (Barton., 2000). Bahkanmasyarakat Ekonomi Eropa mengeluarkansemacam aturan pembatasan dan pelarang-anpenggunaan beberapa feed additive antibiotikdan pemacu pertumbuhan (growth promoter)dalam ransum ternak, dimulai sejak 1 Januari2006 dan pada akhir 2006 tidak diperkenankanlagi adanya feed additive baru dalam campuranpakan ternak di kawasan negara-negara UniEropa untuk melindungi konsumennya dan
mencegah efek negatif bagi manusia (TheEauropean Parliament and the Council of theEuropean Union, 2003). Oleh karena itu akhir-akhir ini permintaan konsumen telah mengarahpada pengguna-an produk ternak yang amandan berkualitas melalui penggunaan bahan-bahan alam serta menghindari penggunaan feedadditive dari sintetis (Patra., 2011).
Berbagai upaya telah dilakukan untukmencari bahan tambahan dalam ransum ternaksebagai pengganti feed additive sintetis yangberbahaya tersebut. Oleh karena itu peng-gunaan feed additive alami merupakanalternative untuk mengurangi residu feed
additive dalam daging ayam. Salah satu feedadditive alami yang dapat digunakan adalahsenyawa bioaktif limonoid yang bersal darilimbah pembuatan jus jeruk.
Limbah jus jeruk merupakan hasilsampingan dari pembuatan minuman jus jeruk.Berdasarkan hasil uji profil fitokimia terutamakandungan bioaktif atau senyawa metabolik

Ucop Haroen 258
sekunder dalam limbah jus jeruk menunjukkankadar kumarin relatif tinggi (+++), untukflavonoid, alkaloid, steroid, triterpenoid,fenolik, saponin menunjukkan kadar relativesedang (++) (Haroen., 2013). Selain senyawabioaktif tersebut diatas limbah jus jeruk inimasih mengandung senyawa-senyawa aktifdominan yang berguna untuk kesehatan ternak.Senyawa bioaktif yang dominan dalam limbahjus jeruk adalah senyawa limonoid (Miller et
al., 2004) dan ( Miller et al., 2008).. Beberapapeneliti mengevaluasi aktifitas biologis darisenyawa aktif limonoid jeruk dimana senyawaini berpotensi sebagai feed additive, (Wing.2000; Roy. et al., 2006; Miller. et al., 1989).Disamping itu juga Roza et al., (2007)melaporkan bahwa senyawa limonoid yangtelah diekstraksi dapat berperan sebagaiantibakteri. Selain itu juga penggunaan limbahjus jeruk ini merupakan salah satu upaya untukmenekan pemakaian feed additive sintetis.
Berdasarkan latar belakang yang dikemukakan di atas maka permasalahan pokokdalam penelitian ini adalah bagaimana senyawaaktif limonoid yang dominan dalam limbahjus jeruk di duga dapat menggantikan fungsidari antibiotic sintetis yang biasa diberikankepada ternak. Sehingga efek buruk daripenggunaan antibiotik sintetis ini bisa kitahindari, kesehatan ternak terjaga dan produkyang dihasilkan oleh ternak juga aman untukdikonsumsi oleh masyarakat.
BAHAN DAN METODE
Ruang lingkup penelitian ini adalahmenggunakan pelarut yang cocok untukmenghasilkan ekstrak limbah jus jeruk denganmetode maserasi serta uji aktifitas anti bakteri,sehingga limbah jus jeruk dapat digunakansebagai sumber feed addtive alami pada ayambroiler. Penelitian dibagi menjadi beberapaTahap percobaan yaitu:
Tahap 1. Pemurnian senyawa limonoid yangmerupakan senyawa dominan terdapatdalam limbah jus jeruk.
Penelitian ini dilaksanakan di LaboratoriumKimia Organik Bahan Alam FakultasMatematika dan Ilmu Pengetahuan AlamUniversitas Andalas. Penelitian dilakukanmulai bulan April 2012 – September 2012.Peubah yang DiukurData yang di ukur jumlah ekstrak limonoid daripelarut etilasetat , methanol, n-heksana seratpemurnian (purifikasi) senyawa limonoidlimbah jus jeruk.
Tahap 2. Uji aktifitas antibakteri daripelarut etilasetat, methanol dan n-heksanadalam menghambat perkembangan bakteripathogen.Penelitian ini dilaksanakan di LaboratoriumKimia Organik Bahan Alam FakultasMatematika dan Ilmu Pengetahuan AlamUniversitas Andalas. Penelitian dilakukanmulai bulan Oktober 2012 – Desember 2012.Rancangan Percobaan.Rancangan yang dipergunakan adalah RancanganAcak Lengkap terdiri 6 perlakuan dan 4 ulangan.P0 = Larutan antibiotik tanpa limonoid (kontrol)
P1 = Larutan 0 ppm limonoid (kontrol)P2 = Larutan 250 ppm limonoidP3 = Larutan 500 ppm limonoidP4 = Larutan 750 ppm limonoidP5 = Larutan 1000 ppm limonoid
Peubah yang di ukur1. Kandungan senyawa murni limonoid
(senyawa dominan) dari limbah jus jeruk2. Diameter zona hambat yaitu dengan
mengukur zona bening yang terbentuk padaperlakuan ekstrak limonoid (mm)
3. Kosentrasi hambat minimum dari ekstraklimonoid yang mempunyai aktivitasantibakteri yang terbesar
HASIL DAN PEMBAHASAN
Isolasi konstituen metabolit sekunder darilimbah jus jeruk pada penelitian ini dilakukanpemisahan menggunakan metoda kromatografi.Hal ini disebabkan kromato-grafi merupakanteknik pemisahan yang didasarkan padaperbedaan migrasi masing-masing komponen

Ucop Haroen 259
melalui fasa diam dan di bawah pengaruh eluensebagai fasa gerak (Still, 1978). Hasilpemurnian pada penelitian ini dari 1 kg serbukkering limbah jus jeruk di dapat sebanyak17,85 mg senyawa aktif limonoid murni (Tabel1).Tabel 1. Hasil pemurnian senyawa aktif limonoid
limbah jus jeruk
Fraksi Kristalisasi Hasil BeratWarna Bentuk
Vial4-8
Aseton,Heksan
PutihKekuningan
Jarum17,85mg
Hasil penelitian tahap pertama menunjukkanbahwa limbah jus jeruk mengandung semuametabolic sekunder yang umumnya terdapatpada tanaman dan mengandung senyawalimonoid murni sebanyak 17,85 mg/kg. Isolasisenyawa limonoid murni penting dilakukanguna mengetahui seberapa besar jumlahsenyawa aktif yang dominan (limonoid murni)yang terdapat dalam limbah jus jeruk.
Setelah didapat senyawa aktif limonoidmurni dari limbah jus jeruk maka hasil isolasidiuji kemurnianya dengan kromatografilapisan tipis (KLT) dengan berbagai komposisieluen dan juga dilakukan pengelusian berulang-ulang. Hasil yang diperoleh memperlihatkannoda tunggal berwarna ungu dengan penampaknoda lampu UV λ 254 nm (Arbain, 1995).Berdasarkan pengamatan tersebut dapatdisimpulkan bahwa senyawa aktif limonoidhasil isolasi relatif murni. Selanjutnyadilakukan pengukuran titik leleh terhadapsenyawa aktif hasil isolasi dan didapatkanlahjarak titik leleh 145,2 - 147o C. Berdasarkanjarak titik leleh, yang memberikan jarak titikleleh yang cukup pendek, mengindi-kasikansenyawa hasil isolasi telah murni. Suatusenyawa dapat dikatakan murni apabila nodayang dihasilkan selalu tunggal denganmenggunakan bermacam eluen (Manjang,1985).Dari Tabel 2 di atas terlihat bahwa zonahambat dari ektrak limbah jus jeruk denganbeberapa pelarut terhadap bakteri E. coli danSalmonela yang diperoleh berkisar antara 8.00– 11.750. Hasil analisis ragam menunjukkanbahwa perlakuan pemberian ekstrak limbah jus
jeruk (methanol, etilasetat dan n-heksana) nyata(P<0.05) berpengaruh terhadap zona hambatbakteri E. coli dan Salmonella.
Dari uji Duncan menunjukkan zona hambatterhadap E. coli maupun Salmonella yangmendapat perlakuan pemberian ekstrak limbahjus jeruk-etilasetat berbeda nyata (P<0.05)lebih tinggi dibandingkan dengan zona hambatekstrak limbah jus jeruk-metanol dan ekstraklimbah jus jeruk-n-heksana. Sementara ituantara pemberian ekstrak limbah jus jeruk-metanol dan ekstrak limbah jus jeruk-n-heksanaberbeda tidak nyata berbeda (P>0.05) dalammenghambat perkembangan bakteri E. coli danSalmonela. Gambaran zona hambat yangterbentuk dari pengujian berdasarkan jenispelarut terlihat pada Gambar 1.
Hasil analisis ragam menunjukan bahwaperlakuan pemberian ekstrak limbah jus jeruk(metanol, etilasetat dan n-heksana) nyata(P<0.05) berpengaruh terhadap daya hambatminimum (MIC) pada bakteri E. coli danSalmonella. Hasil uji MIC terhadap bakteri E.coli dan Salmonella yang diperoleh dalampenelitian ini berkisar antara 27.427 – 44.54%dan 4.72 – 12.645%. Dari uji MIC terlihatbahwa ekstrak limbah jus jeruk ternyata lebihefektif dalam menghambat perkembanganbakteri E. coli dari pada Salmonella.
Dari uji Duncan hasil uji MIC dari ekstraklimbah jus jeruk-etilasetat berbeda nyata(P<0.05) lebih tinggi dibandingkan denganekstrak limbah jus jeruk-metanol dan ekstraklimbah jus jeruk-n-heksana terhadap bakteri E.coli dan Salmonella.Sementara itu hasil ujiMIC antara ekstrak limbah jus jeruk-methanoldan ekstrak limbah jus jeruk-n-heksana berbedatidak nyata (P>0.05) terhadap E. coli kecualidengan Salmonella berbeda nyata (P<0.05).Hasil uji MIC ini sejalan dengan yangdiperoleh pada pengamatan zona hambat,dimana ekstrak limbah jus jeruk-etilasetatmengasilkan zona hambat yang lebih baikterhadap bakteri E. coli dan Salmonela. Lebihefektifnya ekstrak limbah jus jeruk-etilasetatdibanding ekstrak limbah jus jeruk

Ucop Haroen 260
Tabel 2. Pengaruh Pemberian ekstrak dari limbah jus jeruk-metanol, ekstrak limbah jus jeruk-etilasetat dan ekstrak limbah jus jeruk-n-heksana terhadap zona hambat (mm) aktivitasantibakteri E. coli dan Salmonella
B a k t e r iP e r l a k u a n
Ekstrak limbah jusjeruk-Metanol
Ekstrak limbah jusjeruk-Etilasetat
Ekstrak limbah jusjeruk-n-heksana
E. Coli 9,125±0.144 b 11,500±0.00 a 9,312±0.125 b
Salmonella 8,062±0.125 b 11,750 ±0.00 a 8,000±0.000 b
Keterangan : Konsentrasi Disc 30 mg/ml.Superskrip huruf kecil yang berbeda pada baris yang sama menunjukkan berbedanyata pada taraf P<0.05
Gambar 1. Efek beberapa jenis pelarut ekstrak limbah jus jeruk terhadap zona hambat bakteri E. colidan Salmonella
Zona hambat yang terbentuk dari perlakuan pemberian ekstrak (limbah jus jeruk-etilasetat)terhadap bakteri E. coli dan Salmonela sebesar 11,50 mm dan 11,750 mm.
Tabel 3. Pengaruh pemberian ekstrak limbah jus jeruk-metanol, ekstrak limbah jus jeruk-etilasetatdan ekstrak limbah jus jeruk-n-heksana terhadap daya hambat (minimum inhibitoryconcentration) (%) antibakteri E. Coli dan Salmonella
B a k t e r iP e r l a k u a n
Ekstrak limbah jusjeruk-Metanol
Ekstrak limbah jusjeruk-Etilasetat
Ekstrak limbah jusjeruk-n-heksana
E. coli 27,427±0.482 b 44,540±0.00 a 27,662±3,534 b
Salmonella 4,720±0.00 c 12,645±0,273 a 9,300±0,00 b
Keterangan : Konsentrasi MIC 30 mg/ml.Superskrip huruf kecil yang berbeda pada baris yang sama menunjukkan berbeda nyata padataraf P<0.05
Tabel 4. Uji MIC konsentrasi ekstrak limbah jus jeruk-etilasetat terhadap bakteri E. coli danSalmonella
KonsentrasiLimbah jus jeruk-etilasetat
(ppm)
Daya hambat (%)
E. coli Salmonella
250 44,54±0.000 c 43,11±0.000 b
500 57,72±1.150 b 58,855±0.578 a
750 60.255±0,601 a 57,465±1,861 a
1000 39,0404±0.615 d 38,625±0.190 c
Keterangan : Konsentrasi MIC 30 mg/ml.Superskrip huruf kecil yang berbeda pada kolom yang sama menunjukkan berbedanyata pada taraf P<0.05
OAc
tOAc

Ucop Haroen 261
-methanol dan ekstrak limbah jus jeruk-n-heksana dalam menghasilkan nilai MICkemungkinan dise-babkan karena kemampuanetilasetat sebagai pelarut mampu menghasilkankonsen-trasi senyawa-senyawa fitokimia yanglebih tinggi dibandingkan dengan pelarutmethanol dan n-heksana.
Hasil analisis ragam menunjukkan bahwakonsentrasi ekstrak limbah jus jeruk-etilasetatberpengaruh nyata (P<0.05) terhadap dayahambat bakteri E. coli dan Salmonella. Dari ujiDuncan daya hambat ekstrak limbah jus jeruk-etilasetat pada konsentrasi 750 ppm terhadapbakteri E. coli dan Salmonella nyata (P<0.05)lebih tinggi dibandingkan dengan taraf 1000,500 dan 250 ppm, kecuali pada bakteriSalmonella antara konsentrasi 500 ppm dan 750ppm berbeda tidak nyata (P>0.05). Hasil inimenunjukkan bahwa ekstrak limbah jus jeruk-etilasetat pada konsentrasi 750 ppm efektifdalam menghambat perkembangan bakteri E.coli, sementara itu pada konsentrasi 500 ppmcukup efektif dalam menghambat perkembanganbakteri Salmonella. Rendahnya daya hambatterhadap pertumbuhan bakteri pada konsentrasi1000 ppm disebabkan reaksi antara ekstraklimbah jus jeruk dengan pertumbuhan bakterihanya berlangsung sampai pada konsentrasi 750ppm. Hal ini disebabkan aktivitas ekstrak yangcukup tinggi dalam menghambat pertumbuhanbakteri terjadi pada konsentrasi yang tidakterlalu tinggi, sehingga terhambatnyapertumbuhan bakteri, dengan kata lain reaksinyatelah berlangsung pada saat konsentrasi sebelummencapai 1000 ppm sehingga terjadi penurunandaya hambat terhadap bakteri.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Hasil isolasi dan purifikasi dari limbah jus
jeruk didapat senyawa murni limonoidsebanyak 17,85 mg/ kg limbah jus jeruk.
2. Berdasarkan uji aktifitas antibakteri pelarutetilasetat lebih efektif dalam menghambat
perkembangan bakteri E.coli maupunsalmonella yaitu 11,500 dan 11,750 mm.
DAFTAR PUSTAKA
Arbain. D. 1995. “Phytochemical Survey OneWay Approach”, HEDS-USAID projectUnand.
Haroen, U. Y. Marlida., Mirzah dan A.Budianyah. 2013. Extraction and isolasiphytochemical and antimicrobial activity oflimonoid compounds from orange wastejuice. Pak. J. Nutr, 12 (8): 730 -735.
Manjang Y. Kimia Analisa Organik, ProyekPeningkatan Pengembangan PerguruanTinggi. Univ. Andalas, Padang 1985. 5 -20.
Miller, E.G., Fanous, R., Rivera-Hidalgo, F.,Binnie, W.H., and Hasegawa, S. 1989. Theeffect of citrus limonoids on hamsterbuccal pouch carcinogenesis.Carcinogenesis 10:1535-1537.
Oluremi, O. I. A., V.O., Ojighen and E. H.Ejembi. 2006. The nutritive potentials ofSweet orange (Citrus sinensis) rid inbroiler production. Int. J. Poult. Sci., 5:613-617.
Roy. A., and Saraf Shailendra. 2006. LimonoidsOverview Of Significant BioactiveTriterpenes Distributed in Plants Kingdom.Biol. Pharm. Bull 29(2)191-201.
Roza. J.M. Liu.Z.X and Guthrie. N. 2007.Effetc Of Citrus Flavonoids and TocotrieNols on Serum Cholesterol Level inHypercholes-terolemic Subjects.Alternative Therapies. Vol 13.
Still, Clark., Kahn, M., and Mitra, A., 1978.Rapid Chromatographic Technique forPreparatives Separations with ModerateResolution. Journal of Organic Chemistry.Vol. 43. No. 14.
The European Parliament and the Council of theEuropean Union. 2003. Regulation (EC)1831/2003 of The European Parliament andof the Council of 22 September 2003 onadditives for use in animal nutrition.Brussels, Belgium.
Van den Bogaard, A.E., N. London, C. Driesen,dan E.E. Stobberingh. 2001. Antibioticresistance of faecal Eschericia coli inpoultry, poultry farmers and poultryslaughterers. J. Antimicrobial Chemotherapy 47 : 763-771.
Wing. J. M. 2000. Citrus Feed Stuffs for DairyCattle. Animal Science DepartementFlorida. University of Florida. Gainesvella.

ISSN 978-602-14989-0-3 BioETI
Analisis mikroba pada Kerang Air Tawar (Contradenscontradens) di Danau Singkarak, Kabupaten Solok, SumatraBaratVIVI FITRIANI DAN ARMEIN LUSI ZESWITA
Program Studi Pendidikan Biologi Sekolah Tinggi Keguruan dan Ilmu Pendidikan(STKIP) PGRI Sumatera BaratE-mail:
ABSTRACTContradens contradens is typical of freshwater mussels or brackish water economically valuable. Contradens contradens
a shellfish known people with the name Lokan. Lokan consumed by people as a source of animal protein and animal feedingredients. Singkarak Lake used by people in daily life from the source of drinking water, toilets, fisheries, irrigation,PLTA and Tourism. The many activities of the community could be expected to cause pollution and affect theContradens contradens become vectors of biotoksin because his diet filter-feeder. Lokan meat is an excellent medium forbacterial growth. This study aims to determine the presence of Escherichia coli and Salmonella sp. on fresh meatLokan(Contradens contradens) originating from Singkarak Lake. The research was conducted in Januari–Mei 2014. Thisresearch is a descriptive method, by looking at and analyzing the presence of microorganisms are bacteria E. coli andSalmonella sp. The method used to determine the bacteriological quality of the meat is Lokan by MPN method andexamination of Salmonella. Data were analyzed by calculating the number of bacteria E. coli by observing the number ofpositive results of the estimation of presumptive test, confirmative test and completed test. The next number of bacteriafrom each positive results are matched with MPN table. Test Salmonella sp. done by looking at the colony grows. Resultsof bacteriological tests on meat samples were examined in BAPELKES Padang showed that of the four samples tested,three of which were negative for the bacteria E. coli samples take in Nagari Singkarak samples contained negativeSalmonella sp. Of this study is suggested to consumers that cooking Lokan perfectly, to avoid diseases that may occurbecause it is caused by bacteria.
Key words: bacterium, meat, Contradens contradens
Pendahuluan
Pelecypoda merupakan kelas kedua terbesarFilum Mollusca yang banyak dimanfaatkanoleh masyarakat sebagai sumber protein hewaniataupun sebagai bahan baku industri (Kastoro,1992). Di perairan Indonesia hidupberanekaragam jenis Pelecypoda. ada yanghidup di air tawar (sungai dan danau) dimanabiasanya disebut dengan pensi atau lokan dankerang yang hidup di laut. Pemanfaatan sumberprotein hewani ini telah mulai diminati olehsebagian masyarakat Indonesia terutama jenis-jenis Pelecypoda yang memiliki arti ekonomi(Kastoro, 1992; Suin dan lswandi, 1994).
Salah satu hewan yang mendiami dasarperairan adalah Contradens contradens. Hasilpenelitian Priantna dkk. (2013) Contradens
contradens merupakan spesies kijing air tawar
Unionidae terbanyak yang didapati di SungaiBrantas dengan jumlah individu sebanyak 41individu. Jenis kerang ini dipanen o1ehmasyarakat dalam jumlah yang banyak. Karenamerupakan salah satu jenis makanan yang enakrasanya. Penduduk di sekitar Danau Singkarakmengenal kerang dengan istilah alo-alo namunpemanenannya tidak sebanyak kerang air tawar(Corbicula sumatrana) yang banyak ditemukandi danau ini. Kerang ini dijual dalam bentuktanpa cangkang. Kerang ini sama halnyadengan Corbicula sumatrana merupakan satusumber makanan yang praktis dan enak sebagaipengganti protein hewani lain. Kerang dipanenmasyarakat dengan ukuran yang bervariasi.
Kerang-kerangan menjadi vektor daribiotoksin karena pola makannya yang bersifatfilter-feeder yaitu dengan cara menyaringmakanan yang terbawa arus atau aliran air,

Vivi Fitriani dan Armein Lusi Zeswita 263
melalui insang dan meloloskan bahan-bahanyang diperlukan. Proses ini menyebabkanterkumpulnya plankton, senyawa kimia danpartikel-partikel kecil lainnya di dalam saluranpencernaan kerang-kerangan (Hikmah, 2011).Umumnya di Indonesia Contradens contradens
merupakan kerang yang khas air tawar, tetapibeberapa spesies dapat hidup pada air payau.Contradens contradens juga dilaporkanterdapat di Danau Maninjau, Danau Singkarak,Danau Diatas dan Danau Dibawah, yaituContradens contradens (Taher, 1986).
Danau Singkarak merupakan salah satudanau terluas yang terdapat di Sumatera Barat.Di Danau Singkarak banyak terdapatContradens contradens atau Lokan yangdimanfaatkan oleh masyarakat, selain tempathabitat Lokan danau juga dimanfaatkan dalamkehidupan sehari-hari, mulai dari sumber airminum, MCK, perikanan, irigasi, PembangkitListrik Tenaga Air (PLTA) dan tempatpariwisata (Nofriza, 2005). Banyaknya aktifitasmasyarakat di Danau Singkarak maka dapatterjadinya pencemaran pada perairan. MenurutAlcamo (1995) dalam Marlina (2009) perairanmerupakan tempat hidup berbagai mikroorganisme dan makroorganisme. Antaramikroorganisme dan makroorganisme akanterjadi interaksi, seperti bakteri akanbersimbiosis dengan organisme yang hidup diperairan seperti plankton, zooplankton, ikan,udang dan kerang.
Sebagian besar Lokan dipasarkan dalamkeadaan segar (tidak mendapatkan perlakuan)tanpa kemasan, sehingga meningkatkanperkembangan bakteri aerob karena kontakdengan udara. Bakteri aerob yang dapat tumbuhadalah Salmonella dan lain sebagainya.Salmonella merupakan salah satu penyebabpenyakit infeksi. Faktor yang mempengaruhiantara lain kebersihan sanitasi yang burukmemegang peranan penting dalam penyebaranpenyakit. Salmonella dapat tumbuh pada susudan hasil olahannya, kerang-kerangan, teluryang dibekukan, daging dan hasil olahannya.Untuk itu perlu diteliti uji bakteriologis daging
segar Lokan Contradens contradens yangberasal dari Danau Singkarak
BAHAN DAN METODE
Pengambilan sampel Contradens contradens diDanau Singkarak telah dilaksanakan pada bulanJanuari-Mei 2014. Uji Bakteriologis dilakukandi Labor Mikrobiologi BAPELKES Padang.
Alat yang digunakan adalah autoklaf,inkubator, timbangan, labu erlemeyer, tabungreaksi, tabung durham, lampu spiritus, cawanpetri, pipet steril 5 ml dan 10 ml, jarum ose,gelas ukur, batang pengaduk, rak tabung reaksi,spray alkohol, gunting, kompor, saringan,ember, pinset, mortil, kulkas, kamera digitaldan alat tulis. Bahan-bahan yang digunakanadalah daging Lokan , selenith broth, mediumLB, BGLB, EMB, SSA, kantong plastik,aquades, kertas label, korek api, selotip dantissue.
Penelitian ini dilakukan melalui pendekatandeskriptif. Sampel lokan diambil pada danausingkarak dengan 2 kali pengambilan.
Analisis data dilakukan dengan caramenghitung jumlah bakteri E. coli denganmengamati jumlah hasil yang positif dari hasilpengujian pendugaan, penegasan danpenyempurna. Selanjutnya jumlah bakteri darimasing-masing hasil yang positif dicocokkandengan tabel MPN. Uji Salmonella sp.dilakukan dengan melihat koloni yang tumbuh.Hasil analisis ini selanjutnya dibandingkandengan Keputusan Menteri Kelautan danPerikanan Nomor : KEP.17/MEN/2004.
HASIL DAN PEMBAHASAN
Hasil uji bakteriologis terhadap daging segarLokan (Contradens contradens) yang berasaldari Danau Singkarak di LaboratoriumMikrobiologi BAPELKES Padang. Hasilpemeriksaan bakteri Escherchia coli padadaging Pensi dapat dilihat pada Tabel 1 dibawah ini.
1

Vivi Fitriani dan Armein Lusi Zeswita 264
Tabel 1. Keberadaan Esherichia coli pada ujipenyempurna dengan tabel MPN/100 mldengan Ragam : 5 x 10 ml, 1 x 1 ml dan1 x 0,1 ml.
Sampel 10ml
1ml
0,1ml
Index MPN/100ml
L1 0 0 0 2L2 0 0 0 5
Keterangan :L1 = Diambil pada tanggal 6 Mei 2014,L2 = Diambil pada tanggal 12 Mei 2014
Selanjutnya hasil pemeriksaan bakteriSalmonella sp pada daging Pensi menunjukkanhasil yang negatif. Uraian yang lebih jelasdapat dilihat pada Tabel 2.
Tabel 2. Hasil Pemeriksaan Bakteri Salmonellasp. pada Daging Segar Pensiyang berasal dari Danau Singkarak
Sampel Salmonella sp.
L1 NegatifL2 Negatif
Keterangan :L1 = Diambil pada tanggal 6 Mei 2014,L2 = Diambil pada tanggal 12 Mei 2014
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Dari keempat sampel yang diuji Salmonella
dinyatakan positif karena terdapat pertumbuhanbakteri Salmonella sp.
DAFTAR PUSTAKA
Bloom Balia, R.L, E. Harlina. dan Denny S.2011. Deteksi Coliform pada Daging SapiGiling Spesial yang dijual di HipermarketBandung. Fakultas Peternakan UniversitasPadjadjaran.http://pustaka.unpad.ac.id/archives/81123/diakses 26 Desember 2012.
Jawetz, M. A. 2005. Mikrobiologi Kedokteran(Medical Microbiology), SalembaMedika : Jakarta
Kastoro, W. W. 1992. Beberapa Aspek Biologidan Ekologi dari Jenis-jenis Molusca LautKomersial yang diperlukan untukmenunjang Usaha Budidaya. ProsedingTemu Karya Ilmiah Potensi Sumber DayaKerang-Kerang Sulawesi Selatan danSulewesi Tenggara. Balai PenelitianBudidaya Pantai Maros
Peraturan Menteri Kelautan dan Perikanan.KEP.17/MEN/2004. Sistem SanitasiKekerangan Indonesia.
Rachmawaty. 2008. Kajian Biofilter Mikrobiadari Remis (Corbicula sp) Jurnal BiologiFMIPA Universitas Negeri Makassar. Vol.9 (1) : Hlm 10-13, April 2008. ISSN :1411-4720.

ISSN 978-602-14989-0-3 BioETI
Analisis tingkat kematangan gonad ikan mungkuih Sicyopterusmacrostetholepis(Bleeker) hidup di Sungai Batang Kuranji KotaPadang berdasarkan umur, panjang dan berat tubuhWarnety Munir, Indra Junaidi Zakaria dan Nelmi Fitria
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRACTSicyopterus macrostetholepis is one of the freshwater gobiid fish that live on the Batang Kuranji river, it is known withcommon name as Mungkuih. This fish has a delicious taste and potential as ornamental fish, because it has a beautifulshape and attractive body colors. The condition of the fish will cause more arrests and not controlled, so it will endangerher existence. As conservation efforts, research on aspects of fish reproduction, including the gonads Maturity LevelAnalysis based on age, length and body weight had been done on January until June 2009, using the descriptive method.Based on age determination using scales, it were found the age of the fish ranged from 7 months to 46 months with abody length ranging from 36.7 to 100 mm and a weight of 0.73 to 12.32 g. There were found 4 fish gonad developmentstages some of which are on the stage: young gonads (GMI II), gonads mature stage (GMI III), post-ovulation (GMI V)and the spent stage (GMI VI) of female fish Mungkuih. Furthermore, in male fish, it were obtained at the stage of "non-reproductive, pre-nesting, nesting and post-nesting". Reproduce occurred over the year and a period to top reproducehappened in April.
Key words: development phase, gonad, age, length, weight
Pendahuluan
Salah satu jenis ikan Gobi (famili Gobiidae)yang banyak ditemukan di perairan tawar KotaPadang yaitu di Sungai Batang Kuranji adalahSicyopterus macrostetholepis (Bleeker, 1853),yang dikenal dengan Mungkuih. Mungkuihadalah kelompok ikan yang berwarna ungu tuadengan 5-6 pita warna yang samar-samar, sirippunggung dan sirip dubur berwarna ungusuram. Pada ekor terdapat sebuah pita warnagelap yang memanjang pada pinggiran ekordan antara kedua pita berwarna orange padajantan. Pada betina terdapat garis hitammemanjang ditengah ekor. Terdapat belahanpada bibir atas dan memiliki 49-54 deret sisiksepanjang sisi badan serta mempunyai 12-16sisik di depan sirip punggung (Kottelat, 1993).
Seperti keluarga Gobiidae lainnya, ikan inimempunyai ciri spesifik yaitu sirip pevikbersatu membentuk piringan penghisap,digunakan untuk menempel pada permukaanyang licin dan bertahan pada kondisi air berarusderas dan berbatu-batu. Tipe sisik cycloid dan
ctenoid, sirip dorsal terpisah. Keluarga ikaniniberukuran kecil dengan warna yang bagusdan tingkah laku yang sangat menarik sehinggaberpotensi besar dikembangkan sebagai ikanhias.
Ikan Mungkuih dijadikan santapan karenamemiliki cita rasa yang lezat dangurih,sehingga selalu ditangkap oleh penduduksekitar sebagai sumber mata pencarianuntukmenunjang kehidupan keluarga. Aktifitaspenangkapan berkelanjutan dan tidak terkontroldikhawatirkan akanmenyebabkan ikan ini suatusaat akan punah. Agar kelestarian ikanMungkuih tetap terjaga maka diperlukanpengelolaan. Aspek yang sangat penting untukkelestarian populasi ikan adalah aspek biologiterutama aspek reproduksi. Oleh karenaituperlu dipelajari aspek reproduksi yaitu tingkatkematangan gonad yang merupakan aspekpenting yang menentukan keberhasilan dalampemijahan.
Pengamatan kematangan gonad dapatdilakukan dengancara morfologidan histologi.Dari penelitian secara histologi akan diketahui

Warnety Munir, Indra Junaidi Zakaria dan Nelmi 266
anatomi perkembangan gonad secara jelas danmendetail (Effendie, 1997).Hasil penelitian inisangat penting sebagai imformasi dasar untukkebijakan berkaitan dengan pemanfaatan secaraberkelanjutan, termasuk aktivitas pembibitandan pembenihan. Pada penelitian ini telahdilakukan Analisis Tingkat Kematangan GonadIkan Mungkuih yang hidup di Sungai BatangKuranji cara dengan histologi dihubungkandengan umur, panjang total dan berat tubuhikan. Tingkat kematangan ditentukan menurutkriteria Sadovy dan Domeier (2005)
BAHAN DAN METODE
Metoda yang dipakai pada penelitian ini adalahmetode deskriptif. Sampel ikan yang digunakandiambil setiap bulan berturut-turut, mulaiJanuari-Juni 2009 dari Sungai Batang Kuranjiyang terletak 48’–0.56’ LS, 100021’– 100033’BT menggunakan kejutan listrik. Sampeldibawa kelaboratorium meng-gunakan kantungplastik yang telah diisi air. Di labor ikandimatikan, panjang ikan ditentukanmengunakan kaliper, berat digital. Umurditentukan dengan menghitung lingkaran sisikyang diambil dari bagian depan antara gurat sisi(linealateralis) sebelah kiri dan pangkal sirippunggung. Penghitungan di lakukan dibawahmikroskop. Setelah selesai dilakukan penentuannilai dimuka, gonad ikan dikeluarkan, difiksasidengan larutan Bouin. Selanjutnya dibuatsedian melalui prosedur standar, diwarnaidengan pewarna H&E. Pengamatan terhadapsediaan dilakukan mengunakan mikroskopcahaya (Nikon) ditujukan untuk penentuantahapan per kembangan gonad dan tingkatkematangan gonad berdasarkan kriteria Sadovyand Domeier (2005). Proses berikutya adalahmengelompokan ikan berdasarkan umur,panjang total dan berat tubuh
HASIL DAN PEMBAHASAN
Dari hasil penghitungan lingkaran sisik ikandiketahui umur ikan berkisar 7 hingga 46bulandengan kisar panjang 36,7 – 100, mm danberat 0,73 – 12,32 g.
Morfologi Gonad S. macrostetholepisGonad S. Macrostetholepis menempel
memanjang di kedua sisi ventro-lateral vertebra,pada bagian dorsal dinding rongga tubuh. Padaikan jantan, gonad menyerupai sepasangbenang, berwarna bening hingga putih. Padaikan betina, gonad seperti gergaji, bergerigi,warna tergantung pada status reproduksi ikan,mulai dari putih kekuningan hingga kuningkemerah-merahan. Hasil ini sesuaidengan pernyataan Uyeno (1986), gonad betinadan gonad jantan umumnya mempunyaiperbedaan baik dari struktur morfologi maupunstruktur histologinya. Ujung posterior bersatudengan saluran kemih, bermuara di kloaka,sedangkan bagian anterior menempel padabagian hati.Histologi S. macrostetholepis betina
Secara histologi antara lapisan korteks danmedula gonad S. macrostetholepis tidak dapatdibedakan karena jaringan yang membangunsangat tipis. Disebelah luar terdapat tunikaalbugenia dan kearah dalam dijumpai jaringanepitel germinal dan jaringan ikat fibrosa, daerahini membentuk lipatan-lipatan yang disebutlamella yang dibangun oleh oosit dalamberbagai tingkatan.
Berdasarkan hasil pengamatan yang telahdilakukan pada sedian histologis ovariumdidapatkan hampir semua ikan telah dewasa.Oosit yang terdapat dalam lamela berada padaberbagai tingkat perkembangan yangmenandakan ovarium dalam keadaan aktif,oosit tidak matang secara bersamaan (asinkron).
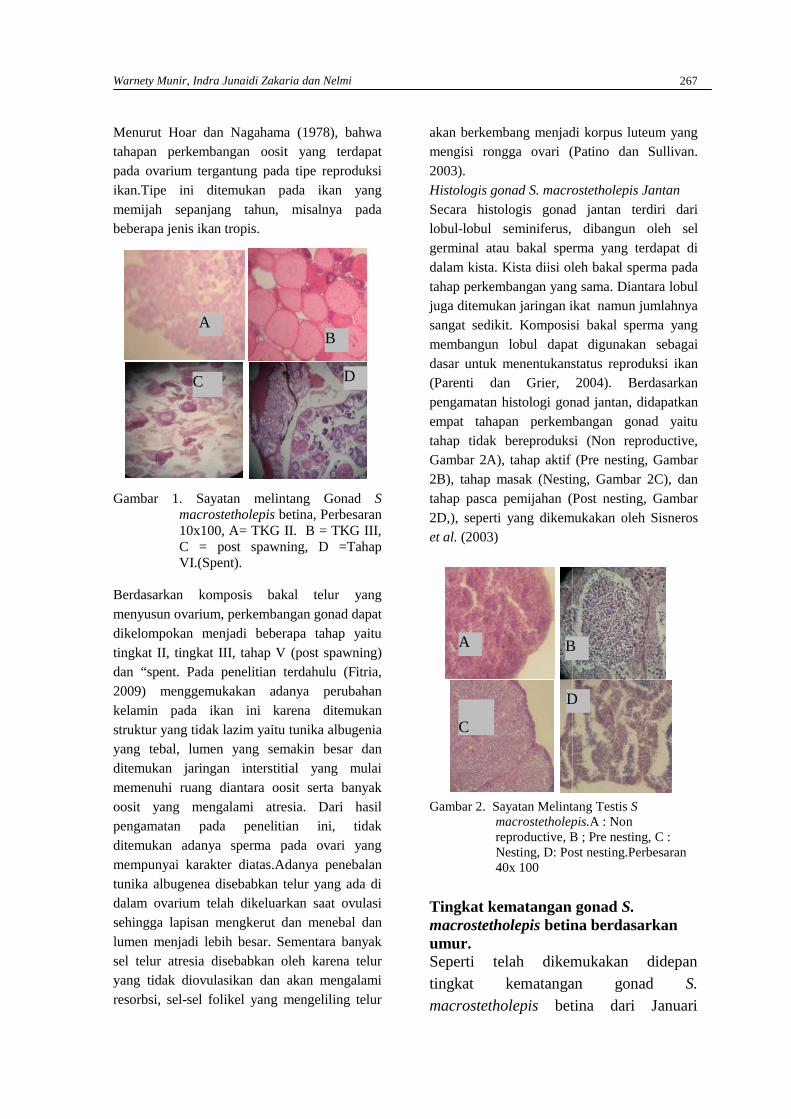
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 267
Menurut Hoar dan Nagahama (1978), bahwatahapan perkembangan oosit yang terdapatpada ovarium tergantung pada tipe reproduksiikan.Tipe ini ditemukan pada ikan yangmemijah sepanjang tahun, misalnya padabeberapa jenis ikan tropis.
Gambar 1. Sayatan melintang Gonad Smacrostetholepis betina, Perbesaran10x100, A= TKG II. B = TKG III,C = post spawning, D =TahapVI.(Spent).
Berdasarkan komposis bakal telur yangmenyusun ovarium, perkembangan gonad dapatdikelompokan menjadi beberapa tahap yaitutingkat II, tingkat III, tahap V (post spawning)dan “spent. Pada penelitian terdahulu (Fitria,2009) menggemukakan adanya perubahankelamin pada ikan ini karena ditemukanstruktur yang tidak lazim yaitu tunika albugeniayang tebal, lumen yang semakin besar danditemukan jaringan interstitial yang mulaimemenuhi ruang diantara oosit serta banyakoosit yang mengalami atresia. Dari hasilpengamatan pada penelitian ini, tidakditemukan adanya sperma pada ovari yangmempunyai karakter diatas.Adanya penebalantunika albugenea disebabkan telur yang ada didalam ovarium telah dikeluarkan saat ovulasisehingga lapisan mengkerut dan menebal danlumen menjadi lebih besar. Sementara banyaksel telur atresia disebabkan oleh karena teluryang tidak diovulasikan dan akan mengalamiresorbsi, sel-sel folikel yang mengeliling telur
akan berkembang menjadi korpus luteum yangmengisi rongga ovari (Patino dan Sullivan.2003).Histologis gonad S. macrostetholepis Jantan
Secara histologis gonad jantan terdiri darilobul-lobul seminiferus, dibangun oleh selgerminal atau bakal sperma yang terdapat didalam kista. Kista diisi oleh bakal sperma padatahap perkembangan yang sama. Diantara lobuljuga ditemukan jaringan ikat namun jumlahnyasangat sedikit. Komposisi bakal sperma yangmembangun lobul dapat digunakan sebagaidasar untuk menentukanstatus reproduksi ikan(Parenti dan Grier, 2004). Berdasarkanpengamatan histologi gonad jantan, didapatkanempat tahapan perkembangan gonad yaitutahap tidak bereproduksi (Non reproductive,Gambar 2A), tahap aktif (Pre nesting, Gambar2B), tahap masak (Nesting, Gambar 2C), dantahap pasca pemijahan (Post nesting, Gambar2D,), seperti yang dikemukakan oleh Sisneroset al. (2003)
Gambar 2. Sayatan Melintang Testis Smacrostetholepis.A : Nonreproductive, B ; Pre nesting, C :Nesting, D: Post nesting.Perbesaran40x 100
Tingkat kematangan gonad S.macrostetholepis betina berdasarkanumur.Seperti telah dikemukakan didepantingkat kematangan gonad S.
macrostetholepis betina dari Januari
C D
AB
A
C
D
B
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 267
Menurut Hoar dan Nagahama (1978), bahwatahapan perkembangan oosit yang terdapatpada ovarium tergantung pada tipe reproduksiikan.Tipe ini ditemukan pada ikan yangmemijah sepanjang tahun, misalnya padabeberapa jenis ikan tropis.
Gambar 1. Sayatan melintang Gonad Smacrostetholepis betina, Perbesaran10x100, A= TKG II. B = TKG III,C = post spawning, D =TahapVI.(Spent).
Berdasarkan komposis bakal telur yangmenyusun ovarium, perkembangan gonad dapatdikelompokan menjadi beberapa tahap yaitutingkat II, tingkat III, tahap V (post spawning)dan “spent. Pada penelitian terdahulu (Fitria,2009) menggemukakan adanya perubahankelamin pada ikan ini karena ditemukanstruktur yang tidak lazim yaitu tunika albugeniayang tebal, lumen yang semakin besar danditemukan jaringan interstitial yang mulaimemenuhi ruang diantara oosit serta banyakoosit yang mengalami atresia. Dari hasilpengamatan pada penelitian ini, tidakditemukan adanya sperma pada ovari yangmempunyai karakter diatas.Adanya penebalantunika albugenea disebabkan telur yang ada didalam ovarium telah dikeluarkan saat ovulasisehingga lapisan mengkerut dan menebal danlumen menjadi lebih besar. Sementara banyaksel telur atresia disebabkan oleh karena teluryang tidak diovulasikan dan akan mengalamiresorbsi, sel-sel folikel yang mengeliling telur
akan berkembang menjadi korpus luteum yangmengisi rongga ovari (Patino dan Sullivan.2003).Histologis gonad S. macrostetholepis Jantan
Secara histologis gonad jantan terdiri darilobul-lobul seminiferus, dibangun oleh selgerminal atau bakal sperma yang terdapat didalam kista. Kista diisi oleh bakal sperma padatahap perkembangan yang sama. Diantara lobuljuga ditemukan jaringan ikat namun jumlahnyasangat sedikit. Komposisi bakal sperma yangmembangun lobul dapat digunakan sebagaidasar untuk menentukanstatus reproduksi ikan(Parenti dan Grier, 2004). Berdasarkanpengamatan histologi gonad jantan, didapatkanempat tahapan perkembangan gonad yaitutahap tidak bereproduksi (Non reproductive,Gambar 2A), tahap aktif (Pre nesting, Gambar2B), tahap masak (Nesting, Gambar 2C), dantahap pasca pemijahan (Post nesting, Gambar2D,), seperti yang dikemukakan oleh Sisneroset al. (2003)
Gambar 2. Sayatan Melintang Testis Smacrostetholepis.A : Nonreproductive, B ; Pre nesting, C :Nesting, D: Post nesting.Perbesaran40x 100
Tingkat kematangan gonad S.macrostetholepis betina berdasarkanumur.Seperti telah dikemukakan didepantingkat kematangan gonad S.
macrostetholepis betina dari Januari
C D
AB
A
C
D
B
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 267
Menurut Hoar dan Nagahama (1978), bahwatahapan perkembangan oosit yang terdapatpada ovarium tergantung pada tipe reproduksiikan.Tipe ini ditemukan pada ikan yangmemijah sepanjang tahun, misalnya padabeberapa jenis ikan tropis.
Gambar 1. Sayatan melintang Gonad Smacrostetholepis betina, Perbesaran10x100, A= TKG II. B = TKG III,C = post spawning, D =TahapVI.(Spent).
Berdasarkan komposis bakal telur yangmenyusun ovarium, perkembangan gonad dapatdikelompokan menjadi beberapa tahap yaitutingkat II, tingkat III, tahap V (post spawning)dan “spent. Pada penelitian terdahulu (Fitria,2009) menggemukakan adanya perubahankelamin pada ikan ini karena ditemukanstruktur yang tidak lazim yaitu tunika albugeniayang tebal, lumen yang semakin besar danditemukan jaringan interstitial yang mulaimemenuhi ruang diantara oosit serta banyakoosit yang mengalami atresia. Dari hasilpengamatan pada penelitian ini, tidakditemukan adanya sperma pada ovari yangmempunyai karakter diatas.Adanya penebalantunika albugenea disebabkan telur yang ada didalam ovarium telah dikeluarkan saat ovulasisehingga lapisan mengkerut dan menebal danlumen menjadi lebih besar. Sementara banyaksel telur atresia disebabkan oleh karena teluryang tidak diovulasikan dan akan mengalamiresorbsi, sel-sel folikel yang mengeliling telur
akan berkembang menjadi korpus luteum yangmengisi rongga ovari (Patino dan Sullivan.2003).Histologis gonad S. macrostetholepis Jantan
Secara histologis gonad jantan terdiri darilobul-lobul seminiferus, dibangun oleh selgerminal atau bakal sperma yang terdapat didalam kista. Kista diisi oleh bakal sperma padatahap perkembangan yang sama. Diantara lobuljuga ditemukan jaringan ikat namun jumlahnyasangat sedikit. Komposisi bakal sperma yangmembangun lobul dapat digunakan sebagaidasar untuk menentukanstatus reproduksi ikan(Parenti dan Grier, 2004). Berdasarkanpengamatan histologi gonad jantan, didapatkanempat tahapan perkembangan gonad yaitutahap tidak bereproduksi (Non reproductive,Gambar 2A), tahap aktif (Pre nesting, Gambar2B), tahap masak (Nesting, Gambar 2C), dantahap pasca pemijahan (Post nesting, Gambar2D,), seperti yang dikemukakan oleh Sisneroset al. (2003)
Gambar 2. Sayatan Melintang Testis Smacrostetholepis.A : Nonreproductive, B ; Pre nesting, C :Nesting, D: Post nesting.Perbesaran40x 100
Tingkat kematangan gonad S.macrostetholepis betina berdasarkanumur.Seperti telah dikemukakan didepantingkat kematangan gonad S.
macrostetholepis betina dari Januari
C D
AB
A
C
D
B

Warnety Munir, Indra Junaidi Zakaria dan Nelmi 268
sampai Juni dapat dikelompokan menjadiempat tahap berdasarkan komposisi bakaltelur yang menyusunnya yaitu tingkat II,tingkat III, tahap V (post spawaning) dantahap VI (“spent”). Keempat tahapanperkembangan gonad tersebut tidakpernah dijumpai secara bersamaan padakisaran kelompok umur, maupun bulanyang sama. Pada hampir semua bulanpengamatan ditemukan 3 tahapperkembangan kecuali pada Januariditemukan 2 tahap perkembangan(Gambar 3). Tahap III ditemukan padasemua umur dan bulan pengamatan .
Semua ikan yang diamati sudahdewasa karena gonad berada pada faseaktif. Pada ikan usia terendah (< 11,pengamatan bulan Maret April) sudahmempunyai gonad pada Tahapperkembangpan III. Tahap ini ditemukandengan jumlah terbanyak setiap bulanpengamatan, sehingga timbul asumsibahwa ikan ikan ini berproduksisepanjang bulan selama pengamatan.Asumsi ini didukung oleh kenyataanditemukan ikan pada tahap spentpadasemuabulanpengamatan walaupun pascaovulasi hanya ditemukan pada bulan Juni,tidak ditemukan pada Januari, Februari,April, Maret dan Mei (Gambar 3a),Tahap spemt mulai ditemukan pada umur16 bulan. Jumlah individu tahap spentpaling tinggi ditemukan Maret pada umur21 – 25 bulan. Dari kenyataan ini didugapuncak reproduksi terjadi pada Maret.
Tingkat Kematangan Gonad S.macrostetholepis Betina BerdasarkanPanjang.Dari hasil pengamatan yang telahdilakukan (Gb.4) pada sedian histologisgonad ikankematangan gonad ditemukan
ada yang berbeda pada kisaran panjangyang sama danpada bulan pengamatanberbeda. TKG II tidak hanya ditemukanpada ikan yang pendek tetapi pada ikanterpanjang (lebih dari 96mm).TKG IIIsudah ditemukan pada ikan terpendek (<35 mm), namun jumlah tertinggiditemukan pada kisaran panjang 56.00-65.99 dan 66.00–75.99 mm hampirsemua bulan pengamatn. Spent sudahditemukan pada ikan panjang 46 mm.
Kematangan Gonad S.macrostetholepis Betina BerdasarkanBerat.Hasil pengamatan tingkat kematangangonad berdasar berat tubuh total (Gb.5)menunjukan pada berat kurang dari 1 ggonad telah berada pada tahapperkembangantiga (TKG III), walaupunjumlah tertinggi ditemukan pada berat 1-3,99 g kecuali pangamatan pada Februari.Spent juga terjadi pada ikan mempunyaiberat lebih dari 1 g yaitu padapengamatan bulan April, tahap ini jugaditemukan pada ukuran yang lebih besaryaitu pada bulan Maret, April dan bulanJuni. Dari hasil ini dapat dikatakan bahwaspent yang menunjukan masa berbiak .tidak berkaitan dengan berat tubuh ikan.
Tingkat Kematangan Gonad S.macrostetholepis Jantan berdasarkanumur.
Dari hasilpengamatan terhadap gonadikan betina, timbul asumsi bahwa ikan inimelakukan reproduksi sepanjangpengamatan. untuk memdukung asumsiini digunakan hasil pengamatan terhadapperkembangan gonad jantan. Seperti telahdikemukakan diatas ditemukan 4 tingkatkematangan gonad, Ke-empat tahapangonad tersebut dijumpai pada kisaran
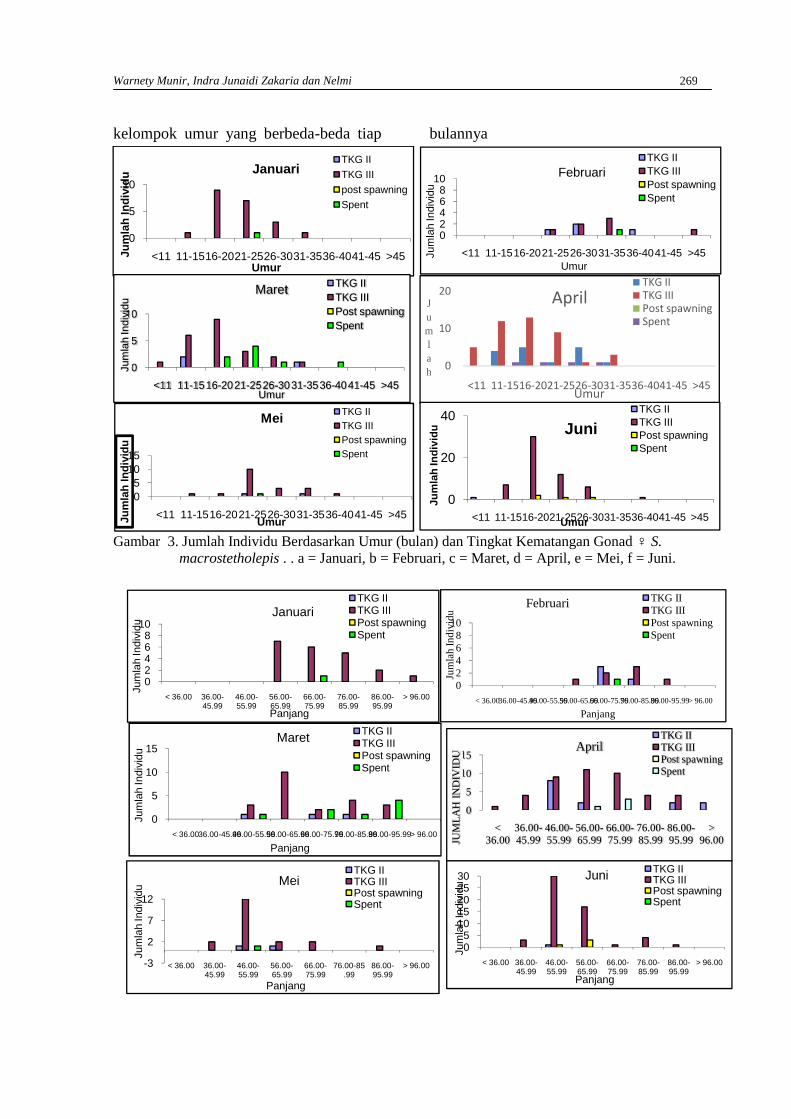
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 269
kelompok umur yang berbeda-beda tiap bulannya
Gambar 3. Jumlah Individu Berdasarkan Umur (bulan) dan Tingkat Kematangan Gonad ♀ S.macrostetholepis . . a = Januari, b = Februari, c = Maret, d = April, e = Mei, f = Juni.
0
5
10
<11 11-1516-2021-2526-30Jum
lah
Indi
vidu
Umur
Januari
0
5
10
15
< 36.0036.00-45.9946.00-55.9956.00-65.99
Jum
lah
Indi
vidu
Panjang
Maret
0
5
10
<11 11-1516-2021-2526-3031-35
Jum
lah
Indi
vidu
Umur
Maret
02468
10
< 36.00 36.00-45.99
46.00-55.99
56.00-65.99
Jum
lah
Indi
vidu
Panjang
Januari
05
1015
<11 11-1516-2021-2526-30Jum
lah
Indi
vidu
Umur
Mei
-3
2
7
12
< 36.00 36.00-45.99
46.00-55.99
56.00-65.99
Jum
lah
Indi
vidu
Panjang
Mei
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 269
kelompok umur yang berbeda-beda tiap bulannya
Gambar 3. Jumlah Individu Berdasarkan Umur (bulan) dan Tingkat Kematangan Gonad ♀ S.macrostetholepis . . a = Januari, b = Februari, c = Maret, d = April, e = Mei, f = Juni.
26-3031-3536-4041-45 >45Umur
JanuariTKG IITKG IIIpost spawningSpent
02468
10
<11 11-1516-2021-2526-30Jum
lah
Indi
vidu
Umur
Februari
56.00-65.9966.00-75.9976.00-85.9986.00-95.99> 96.00
Panjang
Maret TKG IITKG IIIPost spawningSpent
26-3031-3536-4041-45 >45Umur
Maret TKG IITKG IIIPost spawningSpent
0
5
10
15
<36.00
36.00-45.99
46.00-55.99
56.00-65.99JU
ML
AH
IN
DIV
IDU
April
56.00-65.99
66.00-75.99
76.00-85.99
86.00-95.99
> 96.00
Panjang
JanuariTKG IITKG IIIPost spawningSpent
02468
10
< 36.0036.00-45.9946.00-55.9956.00-65.99
Jum
lah
Indi
vidu
Februari
26-3031-3536-4041-45 >45Umur
Mei TKG IITKG IIIPost spawningSpent
05
1015202530
< 36.00 36.00-45.99
46.00-55.99
56.00-65.99
Jum
lah
Indi
vidu
Panjang56.00-65.99
66.00-75.99
76.00-85.99
86.00-95.99
> 96.00
Panjang
MeiTKG IITKG IIIPost spawningSpent
0
10
20
<11 11-1516-2021-2526-30
Jumlah
Umur
April
0
20
40
<11 11-1516-2021-2526-30
Jum
lah
Indi
vidu
Umur
Juni
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 269
kelompok umur yang berbeda-beda tiap bulannya
Gambar 3. Jumlah Individu Berdasarkan Umur (bulan) dan Tingkat Kematangan Gonad ♀ S.macrostetholepis . . a = Januari, b = Februari, c = Maret, d = April, e = Mei, f = Juni.
21-2526-3031-3536-4041-45 >45Umur
FebruariTKG IITKG IIIPost spawningSpent
46.00-55.99
56.00-65.99
66.00-75.99
76.00-85.99
86.00-95.99
>96.00
AprilTKG IITKG IIIPost spawningSpent
56.00-65.9966.00-75.9976.00-85.9986.00-95.99> 96.00
Panjang
Februari TKG IITKG IIIPost spawningSpent
46.00-55.99
56.00-65.99
66.00-75.99
76.00-85.99
86.00-95.99
> 96.00
Panjang
Juni TKG IITKG IIIPost spawningSpent
21-2526-3031-3536-4041-45 >45Umur
AprilTKG IITKG IIIPost spawningSpent
21-2526-3031-3536-4041-45 >45Umur
JuniTKG IITKG IIIPost spawningSpent
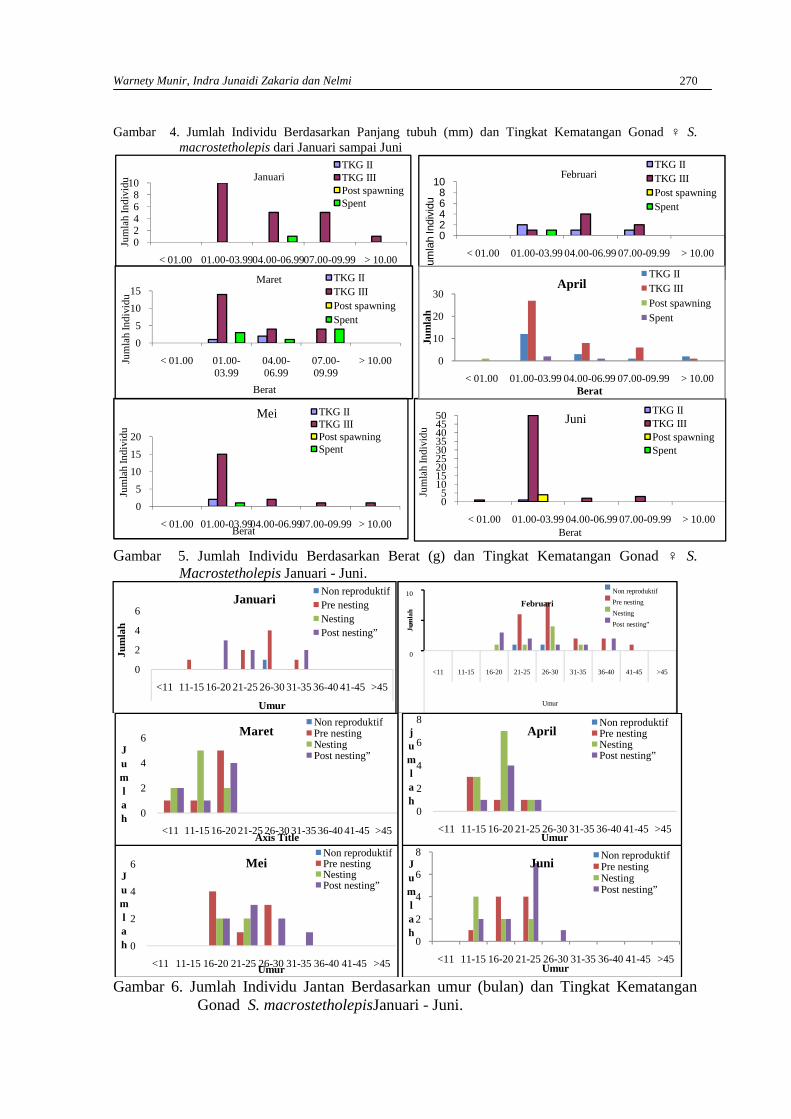
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 270
05
101520253035404550
< 01.00 01.00-03.99 04.00-06.99 07.00-09.99 > 10.00
Jum
lah
Indi
vidu
Berat
JuniTKG IITKG IIIPost spawningSpent
Gambar 4. Jumlah Individu Berdasarkan Panjang tubuh (mm) dan Tingkat Kematangan Gonad ♀ S.macrostetholepis dari Januari sampai Juni
Gambar 5. Jumlah Individu Berdasarkan Berat (g) dan Tingkat Kematangan Gonad ♀ S.Macrostetholepis Januari - Juni.
Gambar 6. Jumlah Individu Jantan Berdasarkan umur (bulan) dan Tingkat KematanganGonad S. macrostetholepisJanuari - Juni.
0
2
4
6
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
Jum
lah
Umur
JanuariNon reproduktifPre nestingNestingPost nesting”
0
5
10
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
Umur
Jum
lah
Februari
Non reproduktif
Pre nesting
Nesting
Post nesting”
0
2
4
6
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
Jumlah
Axis Title
MaretNon reproduktifPre nestingNestingPost nesting”
0
2
4
6
8
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
jumlah
Umur
AprilNon reproduktifPre nestingNestingPost nesting”
0
2
4
6
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
Jumlah
Umur
MeiNon reproduktifPre nestingNestingPost nesting”
0
2
4
6
8
<11 11-15 16-20 21-25 26-30 31-35 36-40 41-45 >45
Jumlah
Umur
JuniNon reproduktifPre nestingNestingPost nesting”
02468
10
< 01.00 01.00-03.9904.00-06.9907.00-09.99 > 10.00
Jum
lah
Indi
vidu
Berat
JanuariTKG IITKG IIIPost spawningSpent
02468
10
< 01.00 01.00-03.99 04.00-06.99 07.00-09.99 > 10.00
Jum
lah
Indi
vidu
Berat
FebruariTKG IITKG IIIPost spawningSpent
0
10
20
30
< 01.00 01.00-03.99 04.00-06.99 07.00-09.99 > 10.00
Jum
lah
Berat
AprilTKG IITKG IIIPost spawningSpent
0
5
10
15
< 01.00 01.00-03.99
04.00-06.99
07.00-09.99
> 10.00Jum
lah
Indi
vidu
Berat
Maret TKG IITKG IIIPost spawningSpent
0
5
10
15
20
< 01.00 01.00-03.9904.00-06.9907.00-09.99 > 10.00
Jum
lah
Indi
vidu
Berat
Mei TKG IITKG IIIPost spawningSpent

Warnety Munir, Indra Junaidi Zakaria dan Nelmi 271
Gambar 7. Jumlah Individu Jantan Berdasarkan panjang (cm) dan Tingkat KematanganGonad S. macrostetholepis mulai Januari hingga Juni
Gambar 8. Jumlah Individu Jantan Berdasarkan berat (g) dan Tingkat Kematangan Gonad S.macrostetholepis mulai Januari hingga Juni
0
2
4
6
<3.6 3.6-4.5 4.6-5.5 5.5-6.5 6.6-7.5
Jumlah
Panjang
Januari
0
1
2
3
4
<3.6 3.6-4.5 4.6-5.5 5.5-6.5 6.6-7.5
Jumlah
Panjang
Mei
0
2
4
6
<3.6 3.6-4.54.6-5.55.5-6.56.6-7.5
Jumlah
Panjang
April
0
2
4
6
.>1.5 1.6 -3.0 3.1-4.5 4.6-60
Jum
lah
Berat
Januari
012345
.<1.5 1.6 -3.0 3.1-4.5 4.6-60 6.1-75
Jum
lah
Berat
Maret
0
1
2
3
4
.<1.5 1.6 -3.0 3.1-4.5 4.6-60
Jum
lah
Berat
MEI
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 271
Gambar 7. Jumlah Individu Jantan Berdasarkan panjang (cm) dan Tingkat KematanganGonad S. macrostetholepis mulai Januari hingga Juni
Gambar 8. Jumlah Individu Jantan Berdasarkan berat (g) dan Tingkat Kematangan Gonad S.macrostetholepis mulai Januari hingga Juni
6.6-7.5 7.6-8.5 8.6-9/5 > 9.6Panjang
JanuariNon reproduktifPre nestingNestingPost nesting”
6.6-7.5 7.6-8.5 8.6-9/5 > 9.6Panjang
Non reproduktifPre nestingNestingPost nesting”
6.6-7.57.6-8.58.6-9/5 > 9.6Panjang
Non reproduktifPre nestingNestingPost nesting”
0
2
4
6
Jumlah
Panjang
June
4.6-60 6.1-75 7.6-8.0 8.1-9.5Berat
JanuariNon reproduktifPre nestingNestingPost nesting”
0
5
10
15
.<1.5 1.6 -3.0 3.1-4.5 4.6-60
Jum
lah
Berat
Februari
4.6-60 6.1-75 76-80 8.1-95Berat
MaretNon reproduktifPre nestingNestingPost nesting”
01234
.<1.5 1.6 -3.0 3.1-4.5 4.6-60
Jum
lah
Berat
April
4.6-60 6.1-75 76-80 8.1-95Berat
Non reproduktifPre nestingNestingPost nesting”
0123456
.<1.5 1.6 -3.0 3.1-4.5 4.6-60 6.1-75
Jum
lah
Berat
Juni
Warnety Munir, Indra Junaidi Zakaria dan Nelmi 271
Gambar 7. Jumlah Individu Jantan Berdasarkan panjang (cm) dan Tingkat KematanganGonad S. macrostetholepis mulai Januari hingga Juni
Gambar 8. Jumlah Individu Jantan Berdasarkan berat (g) dan Tingkat Kematangan Gonad S.macrostetholepis mulai Januari hingga Juni
Panjang
Non reproduktifPre nestingNestingPost nesting”
4.6-60 6.1-75 76-80 8.1-95Berat
FebruariNon reproduktifPre nestingNestingPost nesting”
4.6-60 6.1-75 76-80 8.1-95Berat
AprilNon reproduktifPre nestingNestingPost nesting”
4.6-60 6.1-75 76-80 8.1-95Berat
Non reproduktifPre nestingNestingPost nesting”

Warnety Munir, Indra Junaidi Zakaria dan Nelmi 272
272
Pada beberapa bulan pengamatan ditemukan tigatahapan perkembangan dan bahkan 4 tahapanperkembangan pada bulan yang sama dan bahkanpada kisaran umur yang sama yaitu pada Februari,pada kisaran umur 21 -25 dan 26 -29 bulan.Untukhasil pengamatan bulan Maret, April, Mai dan Juniditemukan tiga tahapan perkembangan yaitu pre-nesting, nesting, dan post nesting secara bersamaanpada selang yang sama, sedangkan tahap nonreproductive tidak ditemukan. Ke-tiga tahapperkembangan dijumpai pada kisaran umur ikan<11 bulan sampai 25 bulan. Tahap perkembanganpre-nesting terjadi selama pengamatan mulai bulanJanuari hingga bulan Juni,sedangkan nestingditemukan mulai bulan Februari hingga bulan Juninamum puncak nesting terjadi pada Maret danbulan April. dan post nesting di temukan hadirmendominasi.pada dua bulan pengamatan ini.Dari hasil pengamatan ini dapat disimpulkan bahwareproduksi terjadi sepanjang bulan, namun puncakreproduksi terjadi pada bulan Maret dan Aprilkarenakan tahap nesting
Tingkat Kematangan Gonad S. macrostetholepisJantan berdasarkan panjang
Dari hasil pengamatan perkembangan testisberdasarkan panjang ikan yang mempuyai kisaran3,6 – 4,5sudah mengalami nesting (memijah)kecuali pada Februari. Pada bulan pengamatanlainnya pada setiap bulan pengamatan ditemukanketiga tahap perkembangan testis lainya walaupunpada kisaran yang berbeda. Dari hasil pengamatanini untuk sementara dapat disimpulkan bahwapanjang ikan tidak menentukan tingkatkematanngan testis ikan ini.Tingkat Kematangan Gonad S. macrostetholepisJantan berdasarkan beratHasil pengamatan pada tahap perkembangan testisikan pada Januari memperlihatkan ikan yangmempunyai panjang kurang dari 1,5 cm sudahmelakukan reproduksi yaitu pada April ikan postnestting, Mei nesting dan post nesting sedangkandan Juni prenesting, nesting dan post nesting. Padabulan yang sama tidak ditemukan ke empat tahapperkembangan secara serentak dan berat tidakmenentukan tingkat perkembangan reproduksi ikanini, ini mungkin disebabkann ikan yang tertangkapsemuanya sudah dewasa.
Dari hasil pengamatan terlihat bahwa masareproduksi tidak berkaitan langsung dengan denganumur, panjang dan berat. Menurut Uyeno (1986)reproduksi dipengaruhi oeleh berbagai faktor, yaitufaktor eksternal seperti kadar hormon streroid, MIHmaupaun GTHl dan faktor eksternal sepertiintensitas cahaya, kekuatan arus, makanan daninteraksi sosial.
KESIMPULAN
Dari Penelitian yang telah dilakukan tentangperkembangan gonad ikan gobi S. macrostetholepis
bulan Januari hingga Juni 2009, maka diperolehkesimpulan yaitu;1. S. macrostetholepis yang tertangkap berumur
antara 7 bulan hingga 46 bulan, panjang 36,7 –100, mm dan berat 0,73 – 12,32 , dimana semuaindividu sudah berada dalam tahap matanggonad (dewasa).
2. Gonad S. macrostetholepis betinaberada padatahap TKG II, TKG III, post spawning danSpent. Sedangkan pada gonad S.
macrostetholepis jantan, didapatkan dalam tahapNon reproduksi, Pre nesting, Nesting dan Postnesting.
3. Reproduksi terjadi sepanjang bulanpengamatandengan puncak reproduksi terjadipada bulan Maret dan April.
4. Tahap perkembangan gonad tidak berhubunganlansung dengan umur, panjang dan umur ikan.
DAFTAR PUSTAKA
Burhanuddin, S, A, Nontji, dan Djamali.Pengamatan terhadap Ikan Gelodok
(Periopthalmus koelreutari) (PALLAS) di PulauPerid alam Press Seminar Ekosistem HutanMangrove.
Effendie, MI. 1997. Biologi Peikanan. YayasanPustaka Nusatama, Bogor. Hoar, W.S. andNagahama. 1978. The Selluler Resource of SexSteroid in Teleostei Gonad. Ann. Biology.Biophis.
Hutomo, M, S. Naamin, A. Nontji dan Djamali.1978. Pengamatan terhadap Ikan Gelodok(Periopthalmus koelreutari) (PALLAS) di PulauPeri dalam Press Seminar Ekosistem HutanMangrove.

Warnety Munir, Indra Junaidi Zakaria dan Nelmi 273
273
Kottelat, M, AJ. Whitten, S.N. Kartikasari dan S.Wirjoadmodjo. 1993. Fresh Water Fishes of
Western Indonesia and Sulawesi. PeriplusEdition Limited, Jakarta.
Lagler, K.F, J.E Bordach, dan R.R. Miller. 1962.Ichtiology. John Wiley and Sons, Inc. New York,London
Mc Manus, JF and WR. Mowry. 1960. StainingMethods Histologic and Histochemical. Paul. B.Hoeber.
Mohsin, M dan A, Ambak. 1996. Marine Fishesand FisherieesOf Malaysia and Neighbouring
Countries.Universitas Pertanian Malaysia Press.Serdang.
Moyle, PB and J.J Cech, Jr. 2000. Fishes AnIntroduction To Ichtyologi 4th Edition. PrenticeHall. USA.
Donald B. 2007. Fish Histology, Female
Reproductive System. Departemen of Biology
The University of Western Ontario LondonOntario. Canada
Sadovy, Y and M.L. Domeier. 2005. PerpelxingProblems Of Sexual Pattern In The Fish Genus
Paralabrax (Serranidae, Serraninae). J. Zool.Lond. 267; 121-133.
Sisneros, J.A,. Forlano, P.M, Knapp, R, and. Bass,A H. 2003. Seasonal variation of steroid
hormone levels in an intertidal-nesting fish, thevocal plainfin midshipman. General andComparative Endocrinology 136 (2004) 101–116.USA
Stacey, N. E. 1984. Control of Timing ofOvulation by Exogenous and EndogenousFactors dalam Fish Reproduction. Potts, G. W.dan Wootton, R. J. (Eds), Academic Press,London
Uyeno, T. 1986. Indo-Pasific Fish Biology. TheIchtiological Society of Japan. Tokyo.

ISSN 978-602-14989-0-3 BioETI
Jenis-jenis tumbuhan Invasif di Hutan Pendidikan danPenelitian Biologi (HPPB)WELLA YURANTI1, SYAMSUARDI1 DAN SOLFIYENI2
1) Herbarium Universitas Andalas (ANDA), Padang, Sumatera Barat 251632) Laboratorium Ekologi Tumbuhan, Jurusan Biologi, FMIPA, Universitas Andalas,
Padang, Sumatera Barat 25163E-mail: [email protected]
ABSTRAKPenelitian tentang jenis-jenis tumbuhan asing invasif yang terdapat di Hutan Pendidikan dan Penelitian Biologi (HPPB)telah dilakukan dari bulan Februari sampai Juni 2014 Limau Manis Padang, kemudian dilanjutkan di LaboratoriumTaksonomi Tumbuhan dan Herbarium Universitas Andalas (ANDA) Universitas Andalas, Padang. Metode surveydigunakan dalam penelitian ini dengan mengoleksi sampel tumbuhan invasif di sepanjang dua rute penjelajahan di HPPB.Identifikasi jenis menggunakan referensi taksnomi khususnya yang terkait dengan gulma dan jenis invasif. Hasilpenelitian menunjukkan bahwa 28 jenis tumbuhan asing (Allien spesies) invasif telah teridentifikasi yang termasuk kedalam 15 famili. Famili Leguminosae memiliki jumlah jenis terbanyak (tujuh jenis), Compositae (lima jenis) dan diikutioleh famili Melastomataceae, Poaceae dan Verbenaceae (masing-masing dua jenis) serta Acanthaceae, Costaceae,Lamiaceae, Malvaceae, Myrtaceae, Oxalidaceae, Piperaceae, Rubiaceae, Rosaceae, dan Vitaceae (masing-masing satujenis).
Key words: hutan tropis, keanekaragaman jenis, pengendalian, tumbuhan invasif
Pendahuluan
Indonesia adalah salah satu negara yang kayadengan keanekaragaman jenis flora.Keanekaragaman hayati di Indonesia termasukdalam golongan tertinggi didunia, jauh lebihtinggi dari pada keanekaragaman sumber dayahayati di Amerika maupun Afrika tropis,apalagi bila dibandingkan dengan daerahberiklim sedang dan dingin. Jenis tumbuh-tumbuhan di Indonesia secara keseluruhanditaksir sebanyak 25.000 jenis atau lebih dari10 % dari flora didunia (Soemarwoto, 1983).Dari sekian banyak jenis-jenis tumbuhan yangada sebagian besar terdapat di kawasan hutantropika basah, terutama hutan primer, yangmenutup sebagian besar daratan Indonesia.Hutan ini mempunyai struktur yang kompleksyang menciptakan lingkungan sedemikian rupasehingga memungkinkan beranekaragam jenisdapat tumbuh di dalamnya. Dari sekian banyakjenis tumbuhan yang ada banyak terdapat didalamnya jenis-jenis yang kisaran ekologinyasama tetapi banyak pula yang berbeda. Jenis-jenis tertentu mempunyai kisaran penyebaran
yang luas dan menduduki berbagai macamhabitat dan seirama dengan itu pula jenissemacam ini biasanya mempunyai variabilitasgenetika yang tinggi (Setiadi, 1983).
Hutan di Sumatera Barat memilikikeanekaragaman tumbuhan tertinggi di Asia(Yahara et al., 2013). HPPB (Hutan Pendidikandan Penelitian Biologi) merupakan salah satukomunitas hutan penting di Sumatera Baratyang terletak di Kota Padang dengan luasnya148 hektar yang di dalamnya terdapat berbagaimacam flora dan fauna yang beberapadiantaranya termasuk biota yang dilindungi.HPPB telah dijadikan sebagai salah satu daerahkunci biodiversitas yang penting di SumateraBarat (Conservation Internasional, 2006).Sehingga hutan ini menjadi penting untuk studidan konservasi keanekaragaman tumbuhan(plants diversity) di hutan hujan tropis dataranrendah. Akhir-akhir ini, terjadi kekawatiranakan merusak keanekaragaman tumbuhanHutan Pendidikan dan Penilitian Biologi(HPPB) dataran rendah Limau Manis akibatgangguan manusia maupun karena bencanaalam. Serbuan ‘alien plants species’ (tumbuhan

Wella Yuranti, Syamsuardi dan Solfiyeni 279
invasif) ke dalam hutan dapat menurunkankeanekaragaman tumbuhan hutan, karenatumbuhan invasif dapat menguasai bahkanmenggantikan tumbuhan asli di hutan tersebut.Sekarang tumbuhan invasif mulai merambahkehutan-hutan alami, yang dikhawatirkankehadirannya dapat merusak keanekaragamantumbuhan di Indonesia.
BAHAN DAN METODE
Penelitian ini dilaksanakan dari bulan Februarisampai Juni 2014 di Hutan Pendidikan danPenelitian Biologi (HPPB), Limau ManisPadang. Kemudian dilanjutkan di LaboratoriumTaksonomi Tumbuhan dan HerbariumUniversitas Andalas (ANDA) Jurusan Biologi,Fakultas Matematika dan Ilmu PengetahuanAlam, Universitas Andalas, Padang.
Selanjutnya dilakukan pengamatan,pemotretan gambar, pengambilan sampel(koleksi) jenis-jenis tumbuhan berbunga yangditemukan dilapangan. Cara koleksi dilakukanberdasarkan sifat hidup/habit (pohon, perdu,herba, calamus & calmus) tumbuhan asinginvasif. Sebagai contoh untuk pohon, dikoleksidengan panjang sampel kira-kira 30 cm dariujung tumbuhan dan setiap koleksi diberi labellapangan dan dilakukan pencatatan data atauinformasi yang akan hilang sebelum prosespengawetan nantinya seperti: Karaktermorfologi organ generatif yaitu warna bunga,dan organ lainnya yang mudah hilang jikadiawetkan antara lain warna batang, habitat danbentuk hidup. Kemudian dilakukan pengawetanlapangan terhadap semua jenis tumbuhan asingyang telah dikoleksi dan selanjutnya pembuatanspesimen di Herbarium Universitas Andalas(ANDA) seperti pengeringan, penyortiran,penempelan. Identifikasi spesies tumbuhanasing invasif dilakukan dengan menggunakanbuku panduan lapangan tentang IAS, dan listspecies invasive.
HASIL DAN PEMBAHASAN
Dari hasil pengamatan dan pengambilan sampeldi Hutan Pendidikan dan Penelitian Biologi(HPPB), Padang telah didapatkan dandiidentifikasi 28 jenis tumbuhan asing (Allienspesies) invasif yang termasuk ke dalam 15famili (Tabel 1). Famili Leguminosae memilikijumlah jenis terbanyak (tujuh jenis), FamiliCompositae (lima jenis) dan diikuti oleh familiMelastomataceae, Poaceae dan Verbenaceae(masing-masing dua jenis) serta Acanthaceae,Costaceae, Lamiaceae, Myrtaceae, Oxalidaceae,Piperaceae, Rubiaceae, Rosaceae, dan Vitaceae(masing-masing satu jenis).
Hasil penelitian Tjitrosoedirdjo (2005)berdasarkan spesimen herbarium menunjukkanbahwa telah ditemukan 1936 jenis tumbuhanasing yang tergabung dalam 187 famili denganfamili Poaceae, Compositae dan Cyperaceaememiliki jumlah jenis yang paling banyak.Pada penelitian di HPPB hanya ditemukan duajenis dari Poaceae yaitu Imperata cylindrica
(ilalang) dan Themeda gigantea (Pimpiang) danlima jenis Compositae (Ageratum conyzoides,Clibadium Surinamense, Austroeupatoriuminulifolium, Mikania mikrantha dansphaegneticola trilobata) dan tidak dijumpaijenis dari Cyperaceae. Kemudian jikadibandingkan dengan penelitian Susanti, Suraiddan Febriana (2013), jumlah jenis tumbuhanasing yang ditemukan di HPPB (28 jenis) lebihbanyak dibandingkan dengan yang ditemukandi Kawasan Hutan Taman Kenali Kota Jambi(6 jenis).
Selanjutnya sebahagian besar dari 28 jenistumbuhan asing yang ditemukan di HPPBmerupakan gulma yang sering dijumpai padaekosistem pertanian dan beberapa diantaranyatidak dijumpai di daerah pertanian seperti:Clidemia hyrta, Melastoma malabathricum,Acasia auriculiformis, Calliandra calothyrsus,
Mimosa pigra, Rhodomystus tomentosa, Piperaduncum, Rubus moluccanus, Lantana camara,Asystasia gangetica dan Cheilocostusspeciosus yang tumbuhan tersebut hidup liar di

Wella Yuranti, Syamsuardi dan Solfiyeni 280
Tabel 1. Jenis-Jenis Tumbuhan Invasif di Hutan Pendidikan dan Penelitian Biologi (HPPB)No Famili Spesies Asal Habitus1 Acanthaceae Asystasia gangetica (L.)T. Anders. Afrika Herba2 Compositae Ageratum conyzoides L. Am. Utara Herba
Austroeupatorium inulifolium (Kunt.) R.M. King & H. Am. Selatan PerduClibadium surinamense L. Am. Tropik PerduMikania micrantha Kunt. Am. Selatan HerbaSpagneticola trilobata (L.C.Rich) Pruski HerbaHerba
3 Costaceae Cheilocostus speciosus(J.Koenig) Sm. Asia Tenggara Perdu4 Lamiaceae Hyptis capitata Jaq. Am. Tropik Perdu5 Malvaceae Sida acuta Burm. Asia Perdu6 Melastomataceae Clidemia hirta (L.) Don. L. Am. selatan Perdu
Melastoma malabathricum L. Asia Perdu7 Leguminoceae Acacia auriculiformis Benth.
Calliandra calothrysus Meisn.Mimosa pudica LMimosa pigra L.Leucana leucocephala (Lam.) De WitCentrosema virginianum (L). Benth.Camaecrista nictitans (L). Moenc
AustraliaAm. TengahAm. TropikAm. TropikMexico & Am. tengahAsia TenggaraAfrikatimur
PohonPohonPerdu merayapPerduPohonPerduPerdu
8 Myrtaceae Rhodomyrtus tomentosa (Aiton). Hassk. Asia Tenggara Perdu9 Oxalidaceae Oxalis barrelieri L. Am. Tropik Herba10 Piperaceae Piper aduncum L. Am. Tropik Pohon kecil11 Poaceae Imperata cylindrica (L.) Raeusch. Asia Tropik Calmus
Themeda gigantea (Cav.) Hac - Calmus12 Rubiaceae Borreria leavis (Lamk.) Griseb. Am. Tropik Herba13 Rosaceae Rubus moluccanus Anet. Asia Timur & Pasifik Perdu14 Verbenaceae Lantana camara L. Amerika Perdu
Stachytarpheta jamaicensis (L.) (Vah). Am. Tropik Perdu15 Vitaceae Cissus hastata Miq. Asia Selatan & Tenggara Herba memanjat
Keterangan: Am. Amerika.* Nama ilmiah mengacu pada The Plant List (2013)
daerah terbuka. Sebahagian besar dari jenistumbuhan asing yang ditemukan merupakanperdu (54%) yang lainnya berupa herba (18%),pohon (14%) dan calamus (7%). Sebahagianbesar jenis yang ditemukan hidup tegak (82%)dan empat jenis (18%) memiliki salah satu sifatyang berpotensi menjadi lebih invasif yaitu:Mikania mikrantha, Centrosema virginianum,
Rubus moluccanus dan Cissus hastata sifatnyayang memanjat (climbing), sedangkanSphaegneticola trilobata dan Mimosa pudica
hidup merayap (creeping) di atas permukaantanah. Menurut Tjitrosoedirdjo, (2005),kebanyakan tumbuhan asing di Indonesiamerupakan gulma pertanian yang bersifat herbadan hidup pada agroekosistem buatan, denganlingkungan dan habitat yang seragam,sedangkan jenis tumbuhan asing invasif terdiridari tumbuhan yang bervariasi habitusnya danhidup pada habitat alami dengan lingkunganyang lebih heterogen.
Selanjutnya 19 jenis yang ditemukan diHPPB telah tercatat sebagai jenis tumbuhan
invasif dalam Global Invasive Species Database(GISD, 2014), sedangkan sembilan jenislainnya (Cassia mimosaides, Hyptis capitata,Clibadium surinamense, sphaegneticolatrilobata, Acasia auriculiformis, Oxalisbarrelieri, Themeda gigantea dan Cissushastata) walaupun tidak dijumpai dalamdatabase namun potensinya sebagai jenis invasifperlu mendapat perhatian karena kepadatanpopulasinya yang tinggi di lapangan. Dari jenis-jenis tumbuhan yang didapatkan dari lapanganmerupakan tumbuhan yang berpotensi menjadiinvasif spesies (Henderson, 2001). Hasilidentifikasi terhadap tumbuhan yang dikoleksijuga menunjukkan bahwa lima jenis diantaranya(Austroeupatorium inulifolium, Lantana camara,Mikania micrantha, Piper aduncum, dan
Stachytarpeta jamaicencis), termasuk kedalamtumbuhan asing invasif terrestrial yang pentingdi Indonesia karena adaptasinya yang tinggi,tidak mempunyai predator alami dan memilikireproduksi vegetatif dan generatif yang cepat(Tjitrosedirdjo, 2005). Hasil penelitian juga

Wella Yuranti, Syamsuardi dan Solfiyeni 281
mengungkapkan bahwa lima jenis yangditemukan (Mimosa pigra, Austroeupatorium
inulifolium, Sida acuta, Lantana camara, Piperaduncum, Mikania micrantha) termasuk jenistumbuhan asing invasif yang berada di hutan-hutan Indonesia yang perlu mendapat perhatian.Berkaitan dengan hal ini, lima jenis tumbuhanasing yang ditemukan (Imperata cylindrica,Clydemia hirta, Lantana camara, Leucaena
leucocephala, Mikania micrantha dan Mimosapigra), termasuk kedalam 100 jenis asing invasifdunia (32 jenis diantaranya berupa tumbuhan)yang paling merusak karena dapat menurunkanbiodiversitas di kawasan konservasi (Lowe et al.,2004; GISD, 2014). Berdasarkan hal ini, 28jenis tumbuhan asing yang ditemukan di HPPBperlu mendapat perhatian untukpengendaliannya, terutama enam jenistumbuhan yang termasuk diantara 32 jenistumbuhan invasif yang paling merusak diduniayaitu: Imperata cylindrica, Clydemia hirta,Lantana camara, Leucaena leucocephala,Mikania micrantha dan Mimosa pigra (ISSG,2004). Pada umumnya tumbuhan invasif yangdijumpai di HPPB merupakan tumbuhan asingatau pendatang yang berasal dari Amerika,namun ada juga yang berasal dari Afrika,Australia dan Asia timur & pasifik. TumbuhanRhodomyrtus tomentosa, Centrosemavirginianum dan Cissus hastata merupakantumbuhan asli Asia Tenggara yang dikenalsebagai jenis invasif di wilayah lain dantermasuk dalam Global Invasif Species
Database.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Ditemukan 28 jenis tumbuhan asing (Allienspesies) invasif yang termasuk ke dalam 15Famili. Leguminosae memiliki jumlah jenisterbanyak (tujuh jenis), diikuti Compositae (limajenis), Melastomataceae, Poaceae danVerbenaceae (masing-masing dua jenis) sertaAcanthaceae, Costaceae, Lamiaceae, Myrtaceae,
Oxalidaceae, Piperaceae, Rubiaceae, Rosaceaedan Vitaceae (masing-masing satu jenis).
DAFTAR PUSTAKA
Bloom Anonim, 2004. IUCN/SSC InvasiveSpesies Specialist Group (ISSG) (http://www.issg.org/gisd).
Conservation Internasional, 2006. ProsidingLokakarya penentuan daerah kuncibiodiversitas di sumatra dan diskusipemanfaatan data bersama, jejaring,monitoring serta identifikasi kebutuhankonservasi pada masa mendatang.
Global Invasive Species Database, 2014.Available from: http://www.issg.org/database/species/ecology.asp [Accessed10Juli 2014].
Henderson, L. 2001. 100 of the Worlds WorstInvasive Alien Species. Fondation D’Enterprise. http://www. Sabonet.org.za/aliens part 3 asteraceae.htm. 30 mei 2014.
Lowe S., Browne M., Boudjelas S.,De PoorterM. 2004. 100 of theWorld’s Worst InvasiveAlienSpecies A selection from the GlobalInvasive Species Database. The InvasiveSpecies Specialist Group (ISSG) aspecialist group of the Species SurvivalCommission (SSC) of the WorldConservation Union (IUCN).
Setiadi, Y. 1983. Pengertian Dasar TentangKonsep Ekosistem. IPB. Bogor.
Soemarwoto, O. 1983. Ekologi LingkunganHidup dan Pembangunan. Djambatan.Jakarta.
Susanti T, Suraida, dan Febriana F. 2013.Prosiding Keanekaragaman TumbuhanInvasif Di Kawasan Taman Hutan KenaliKota Jambi.
The Plant List. 2013. Version 1.1. Published onthe Internet; http://www. Theplantlist.org/(accessed 11thJuly).
Tjitrosoedirdjo, S.S., dan Veldkamp, F. 2005.Bartlettina sordid (Eupatorium sordidum)(Compositae), an invasive alien plantspecies in the G. Gede Pangrango NationalPark, West Java, Indonesia. Fl. Mal. Bull.14(3): 172.
Yahara, Fujii dan Tagane. 2013. Floristik Studyat altitudinal In Gunung Gadut. progressreport. Kyushu University.

ISSN 978-602-14989-0-3 BioETI
Inventarisasi jenis Kodok (Ranidae) sebagai komoditi ekspor diSumatera BaratWINCE HENDRI1) DAN NAWIR MUHAR2
1)Program Pacasarjana Universitas Andalas Padang2)Pendidikan Biologi FKIP Universitas Bung Hatta PadangE-mail:
ABSTRAKPenelitian ini betujuan untuk menginventarisasi dan menganalisis keanekaragaman jenis kodok (Ranidae) yang ditangkapuntuk diperjualbelikan penduduk di daerah Sumatera Barat sebagai komoditi ekspor. Penelitian dilakukan di daerahSijunjung, Ujung Gading (Pasaman), Mentawai (Siberut Tengah) dan Tarusan (Painan) dengan metode penangkapansecara langsung di lapangan. Hasil penelitian menunjukkan jenis kodok yang sering ditangkap dan diperjualbelikanadalah jenis Fejervarya limnocharis ditemukan pada lokasi Ujung Gading (Pasaman), Sijunjung dan Painan;Limnonectes blythii ditemukan di Sijunjung, Pasaman, Mentawai, Rana erythraea ditemukaan di Pasaman dan Painan,Rana chalconata dan Limnonectes sp. Dari penelitian yang telah dilakukan terlihat beberapa jenis kodok berukuran besarsepeti Limnonectes blythii dan. Limnonectes sp jarang ditemukan, tetapi kodok berukuran kecil Fejervarya limnocharisditemukan dalam jumlah yang banyak. Dari penelitian ini dapat disimpulkan bahwa keanekaragaman jenis kodokRanidae yang ditangkap di Sumatera Barat mempunyai gambaran spesies yang berbeda pada kawasana daerah yangberbeda. Penelitian ini perlu dilanjutkan dengan penelitian lebih mendalam kepada keanekaragaman genetik mengingatbeberapa jenis sangat jarang ditemukan serta menganalisis hubungan filogenetik di antara dan di dalam spseies ituditemukan.
Key words: inventarisasi, kenekaragaman, filogenetik
Pendahuluan
Amfibia adalah vertebrata pertama yang beralihdari kehidupan di air ke kehidupan di daratkarena tidak dapat beradaptasi secara penuhdengan lingkungan daratan maka hewan inihidup di antara lingkungan berair dan daratan,mempunyai kulit basah, berkelenjar, lembut,tanpa rambut, bulu dan sisik. Ampihibiatergolong hewan berdarah dingin dengan suhuyang bervariasi tergantung pada keadaanlingkungan (poikilotermal-ektotermal) (vanKampen, 1923; Storer dan Usinger, 1968).
Pada umumnya Amfibia bersifat tidakkhusus dalam pemilihan makanannya. Hal inimerupakan salah satu penunjang kesuksesandalam hal melangsungkan kehidupannya dialam bebas. Dari tingkah laku makan dankemampuan memangsa hewan apa saja yangdapat ditelannya sesuai dengan habitatnya dipinggir-pinggir sungai pada umumnya,memungkinkan kodok sebagai pengontrolhama tanaman secara biologis.
Ukuran bobot badan kodok jantan dewasa yangpernah ditemukan mencapai 900 g sedangkankodok betina mencaai 1100 g. panjang badang(dari mulut ke dubur) kodok jantan dewasa dpatmencapai 200 mm dan betina 220 mm (Sugiridan Soenarjo, 1980). Aktifitas mencari makankodok umumnya berlangsung pada malam hariapabila kelembaban tinggi. Perpindahan danperkawinan biasanya dilakukan apabila hujanmulai turun.
Beberapa penduduk di Sumatera Baratmenangkap beberapa jenis kodok dari alamdiperjualbelikan dan untuk diekspor sebagaisumber pendapatan.. Akhir-akhir ini jeniskodok tersebut sudah jarang ditemukan padahabitatnya karena populasinya menurun.Turunnya populasi juga disebabkan olehpenggunaan pestisida yang dapat mematikankecebong dan anak-anak kodok yang sedangtumbuh serta karena kerusakan habitat.
Pada masa yang akan datang, tekananterhadap populasi kodok akan terus berlanjutdan bukan tidak mungkin pada suatu saat

Wince Hendri dan Nawir Muhar 283
spesies yang ditangkap ini akan punah. Kajiankonservasi sebagai upaya dan saranaperlindungan di negara lain sudah banyakdilakukan, karena dari segi kepadatannyaterlihat penurunan populasi, namun diIndonesia dengan keanekaragaman yang tinggibelum ada amphibia yang dilindungi. Hal initentu memerlukan perhatian besar, terutamapada spesies endemik, langka dan mempunyainilai ekonomi tinggi bagi penduduk.
Pandangan umum menyatakan keaneka-ragaman fauna yang paling tinggi ada di hutantropik. Demikian pula halnya mengenaikeanekaragaman hewan Amphibia yangtertinggi ada di daerah tropik. Di hutan hujantropik terdapat hubungan komunitas yangkompleks. Hubungan tersebut akan lebihmenjadi khusus (terspesialisasi) dengan adanyadukungan masalah relung (niche) dan stenofagiinterspesies yang tersegrasi. Sehingga haltersebut menyebabkan keanekaragaman(kekayaan) spesies komunitas meningkat.Iskandar dan Erdelen (2004), melaporkanbahwa Indonesia, sebagai salah satu pusatkeanekaragaman yang terbesar di dunia, baikdari segi kekayaan alam jenisnya maupun darisegi tingkat endemisitasnya. 16% dari amfibidan reptil dunia terdapat di Indonesia, denganjumlah lebih dari 1100 jenis
Keanekaragaman hayati merupakan cirikhas suatu daerah yang menyangkut keragamandi dalam dan di antara oganisme hidup,kumpulan organisme, komunitas biotik danproses biotik yang masih bersifat alamiahmaupun yang sudah diubah oleh manusia.Keanekaragaman hayati dapat diukur dari levelgenetik beserta identitasnya, jumlah spesies,kumpulan spesies, komunitas biotik, prosesbiotik dan jumlah (seperti kelimpahan,biomassa, penutupan dan laju) serta strukturdari level-level tersebut.
Keberadaan spesies-spesies dan kelimpahanpopulasi amphibia lebih banyak terdapat padadaerah-daerah yang kelembaban udaranyacukup tinggi seperti, misalnya, dekat suatuperairan. Selain itu, ketinggian suatu daerah
juga berpengaruh terhadap keanekaragamanspesies dari amphibia. Hubungan perkerabatanevolusioner atau filogenetik di antara familiAmphibia merupakan suatu yang sangatmendasar untuk menginterpretasikanbiogeografinya dan sangat berarti dalammenyusun klasifikasinya.
Penelitian terhadap jenis katak padaumumnya di Sumatera Barat sudah banyakdilakukan baik aspek taksonomi, evolusi,ekologi maupun genetika terhadap jenistertentu. Seperti yang telah dilakukan olehDjong, 2003; Djong, et al (2007), Djong(2010); Inger and Iskandar (2009), namunjenis-jenis yang ditangkap, diekspor danmempunyai nilai ekonomis serta analisisiskeanekaragaman jenis kodok belum diketahuidengan jelas.
Berdasarkan hal tersebut di atas untukmeningkatkan ketersediaan kodok, sertamenjamin kebutuhan penduduk yangberkesinambungan, maka diperlukan informasijenis kodok apa saja yang ditangkap untukdiperjualbelikan penduduk di daerah SumateraBarat dengan judul penelitian “InventarisasiJenis Kodok (Ranidae) Sebagai KomoditiEkspor Di Sumatera Barat” dengan tujuanuntuk:1. Menginventarisasi jenis kodok yang
ditangkap dan diperjualbelikan penduduk didaerah Sumatera Barat.
2. Menganalisis keanekaragaman jenis dankeberadaan kodok yang ditangkap untukdiperjualbelikan penduduk di daerahSumatera Barat.
Dari penelitian ini diharapkan informasimengenai jenis kodok yang ditangkap di daerahSumatera Barat dalam upaya meningkatkanperdagangan kodok dan sebagai strategipelestarian ataupun usaha usaha domestifikasikodok dimasa yang akan datang.
BAHAN DAN METODE
Penelitian ini dilakukan di daerah SumateraBarat pada lokasi di daerah Sijunjung, UjungGading (Pasaman), Siberut Tengah (Mentawai)

Wince Hendri dan Nawir Muhar 284
dan Tarusan (Painan) dengan metode jelajah.Kodok yang didapatkan diidentifikasi dandikoleksi di laboratorium Biologi UniversitasBung Hatta. Kodok-kodok family Ranidaeyang didapatkan dipelihara dan diidentifikasiuntuk menetapkan karakteristik dan jenis untukkeperluan taksonomi. Alat-alat yang digunakanadalah lampu senter, ember plastik, karungplastik, kantung plastik, alat seksi, spuit 5 ml,spuit 2 ml, spuit 1 ml, cawan petri, gelas ukur10 ml, gelas ukur 100 ml, pipet ukur, tabungreaksi, pipet kecil, termos es, sarung tangan,loup, dan alat potret.
Sampel kodok ditangkap dari pinggir sungaidan semak sekitarnya pada malam hari, kodokyang didapatkan dimasukkan dalam kantongplastik, diberi label dan di bawa kelaboratorium untuk diidentifikasi.
HASIL DAN PEMBAHASAN
Dari penelitian yang telah dilakukan didapatkan5 jenis amfibia famili Ranidae, yaituFejervarya limnocharis, Rana calconata, Ranaerythraea, Limnonectes blythii dan,Limnonectes sp. Jenis-jenis kodok yangdijumpai kategori melimpah adalah Fejervarya
limnocharis, Rana erythraea, Rana chalconata.Sedangkan Rana bylthi dijumpai dalam jumlahyang sangat sedikit. Kerapatan jenis yangditemukan dapat dilihat pada Tabel 1.
Tabel 1. Keragaman jenis dan jumlah spesieskodok yang ditemukan pada tiaplokasi penangkapan
No Jenis
Lokasi
Sij
unju
ng
Pas
aman
Men
taw
ai
Pai
nan
12345
Fejervarya limnocharisLimnonectes blythiiRana erythraeaRana chalconataLimnonectes sp
62---
18253-
-2--2
4-25-
1. Fejervarya limnocharisKingdom : AnimaliaFilum : Chordata
Kelas : AmphibiaOrdo : AnuraFamili : RanidaeGenus : FejervaryaSpecies : Fejervarya limnocharis
Jenis kodok ini ditemukan di lokasiPasaman dan Painan. Dijumpai di sawahpenduduk dan pinggiran perairan yang banyakterdapat tanaman air, duduk di atas daun-daunrumputan sekitar di atas permukaan Kadang-kadang jenis ini dijumpai pada areal daratansekitar 1 meter dari tepi perairan. Jenis inibersifat semi-akuatik. Jenis ini ditangkap dandijual oleh penduduk. Kodok ini dikenaldengan Kodok Sawah dan dagingnya umumdikonsumsi manusia. Jari kaki belakangsetengah selaput pada ruas terakhir, memilikisepasang bintil metatarsal, tekstur kulitberkerut tertutup oleh bintil-bintil panjang yangtipis. Tympanum terlihat jelas. Punggungberwarna coklat dengan bercak gelap dengancorak bentuk huruf W atau U di antara bahu.seperti lumpur dengan bercak-bercak yanglebih gelap yang kurang jelas tetapi simetris.Perut berwarna putih, pada bagian bagianpunggung terdapat garis vertebral keputihan.Panjang tubuh 30 - 50 mm.(Gambar 1)
Tekstur kulit berkerut, tertutup oleh bintil-bintil tipis yang biasanya memanjang, parareldengan sumbu tubuh. Warna kulit kotor sepertilumpur dengan bercak-bercak yang lebih gelapyang kurang jelas tetapi simetris.
2. Limnonectes BlythiiKingdom : AnimaliaPhylum : ChordataClass : AmphibiaOrdo : AnuraFamili : RanidaeGenus : LimnonectesSpesies : Limnonectes blythii
Jenis kodok ini ditemukan di Sijunjung,Pasaman dan Mentawai, tetapi tidak ditemukandi Painan. Kerap dijumpai pada bagian tepianak air perairan, duduk di atas daun-daun

Wince Hendri dan Nawir Muhar 285
rumputan sekitar 10 cm di atas permukaan,walaupun dalam jumlah yang sedikit. Kadang-kadang jenis ini dijumpai pada areal daratansekitar 1 meter dari tepi perairan. Ukuran tubuhlebih besar. Kepala pipih, moncong halus danbentuk triangular. Ujung moncong dapatrincing atau tumpul, cantus rostalis tak jelas.Lore agak cekung, Lore dan pipi miringkebawah, antara mata dan tulang rahang atasterdapat alur sempit. Lipatan supra timpani
yang membujur dari bagian belakang matamelalui bagian atas gendang telinga menuju keaksila dalam bentuk lengkungan. Di antarabagian belakang kedua matanya terdapat lekukyang tak jelas. Membran timpani terlihat jelas.Pada kelopak mata atas bagian belakang bintil-bintil. Pada rahang bagian depan terdapat satupasang penonjolan tulang (apofisis), denganlidah oval yang tebal dan bercabang dua(Gambar 5).
Pada tungkai depan, ujung jari berbentukbulatan kecil, secara berurutan jari ketiga relatiflebih panjang dari jari pertama, keempat danjari kedua. Pada sisi medial jari kedua danketiga terdapat rigi dermal. rigi dermal sisilateral jari kedua tidak tumbuh dengan subur,sedangkan pada sisi lateral jari ketigamempunyai rigi tersebut. Tuberkel metakarpalbesar dan oval. Tungkai belakang berselaputrenang penuh, ujung jari-jari berakhir dengancakram kecil. Selaput renang antara jari kakikeempat dan kelima berpadanan tinggi dengansubartikular ketiga dan agak cekung. Secaraberurutan jari kaki keempat lebih panjang daripada jari ketiga, kelima, kedua dan jari pertama.Sisi lateral jari kelima dan sisi medial tarsallipatan kulit (lipatan tarsal) yang tidak jelasatau tidak ada. Tuberkel tarsal sebelah medialbesar dan memanjang, sedangkan tuberkel
subatikular berbentuk bulat atau lonjong. Kulithalus, warna kulit pada bagian atas kepala danpunggung coklat abu-abu terang, kelabu hitamsampai hitam dengan bercak hitam maupuncoklat. garis loreal yang membujur dari hidungsampai mata tidak pernah dijumpai. Garis diantara kedua mata tidak jelas. Tanda huruf W
pada bagian dorsal yang terdapat diantarakedua tungkai depan kadang-kadang terlihatdengan jelas, kadang-kadang tidak jelas. garispunggung yang membujur dari bagian antaratelinga sampai dubur tidak pernah dijumpai.Permukaan ventral kepala berpigmen. Bibirsering dengan bercak gelap atau aris vertikal.
Pada bagian lengan bagian atas kaki depanterdapat lorek tak teratur. Pada bagian dorsalpaha terdapat lorek yang tak teratur, sedangkandi bagian belakang terdapat totol yang takteratur. Pada betis dan tapak tungkai belakangterdapat lorek. Permukaan bawah tungkaibelakang berwarna kuning pucat atau orangepucat seharusnya atau dengan bercak coklat.hewan jantan tidak mempunyai kantung suara.
3. Rana erythraea
Kingdom : AnimaliaPhylum : ChordataClass : AmphibiaOrdo : AnuraFamili : RanidaeGenus : RanaSpesies : R. erythraea
Jenis kodok ini dijumpai pada tipe habitatkolam dengan bagian tepi berdekatan denganpersawahan. Kebiasaan pada habitat yangdisukainya adalah berdiam diri denganmemendamkan badannya ke dalam air, hanyamatanya berada di permukaan air. Warna kulitbagian dorsal kehijauan dan lipatan dorso-lateral berwarna kekuningan dengan pitaberwarna kehitaman. hijau lumut atau hijaumuda di punggungnya. Sepasang lipatandorsolateral yang jelas, besar, berwarnakekuningan dan kadang-kadang disertai dengangaris hitam, terdapat di kiri kanan punggung.Tangan dan kaki berwarna kuning coklat muda,dengan loreng-loreng terutama pada paha. Sisibawah tubuh berwarna putih. Kulit licin danhalus. Ukuran sekitar 45-75 mm. Tangandengan ujung jari melebar serupa piringan yangmeruncing, yang terbesar sekitar setengahdiameter timpanum (gendang telinga). Piringanpada jari kaki lebih kecil. Selaput renang

Wince Hendri dan Nawir Muhar 286
mencapai pangkal piringan di jari-jari kaki,kecuali pada jari keempat yang memiliki duaruas bebas dari selaput. Terdapat sekurangnyasatu bintil metatarsal di kaki, yakni di sisidalam (Gambar 2).
4. Rana chalconata
Kingdom : AnimaliaPhylum : ChordataClass : AmphibiaOrdo : AnuraFamili : RanidaeGenus : RanaSpesies : R.chalconota
Warna tubuh coklat muda berbintik hitam.Garis coklat tua memanjang pada bagiansamrping tubuh, mulai moncong sampaipangkal kaki belakang. Kulit halus dan licin.Panjang tubuh kodok katak 38 mm nnn- 60 mm.Putaran gelap tempat di belakang (kecuali didua), bintik dari melanophores pada permukaanventral kaki, tenggorokan keputihan, denganbintik gelap. Vomerine gigi berjarak satu samalain. Jari kaki berjumlah empat sepenuhnyaberselaput untuk distal subarticulartuberkulum; tuberkulum metatarsal luarditinggikan. Jantan dengan bantalanperkawinan keputihan pada jari pertama; vokalkantung bukaan di sudut-sudut lantai mulut;spinules keputihan pada butiran dari dorsum.
Pada jantan memiliki spinules dikelopakmata bagian atas. Jantan dalam sampel ini 30-40 mm, tetapi betina jauh lebih kecil (45-65mm). Kodok ini dengan diameter lebih keciltimpanum relatif pada jantan (Gambar 3).
5. Limnonectes sp
Kingdom : AnimaliaPhylum : ChordataClass : AmphibiaOrdo : AnuraFamili : RanidaeGenus : LimnonectesSpesies : Limnonectes sp1
Jenis kodok ini ditemukan di Pasaman danPainan. Dijumpai pada bagian tepi anak airperairan dengan cirri-iri hampir mirip denganLimnonectes blythii. Kepala pipih, monconghalus dan bentuk melebar. Ujung moncongruncing atau tumpul, cantus rostalis tak jelas.Lore dan pipi miring kebawah, antara mata dantulang rahang atas terdapat alur sempit. Lipatansupra timpani yang membujur dari bagianbelakang mata melalui bagian atas gendangtelinga menuju ke aksila dalam bentuklengkungan. Di antara bagian belakang keduamatanya terdapat lekuk yang tak jelas.Membran timpani sebagian tersembunyi dibawah lipatan kulit dari belakang mata sampaimemotong bagian tympanum. Pada kelopakmata atas bagian belakang terdapat bintil-bintil,sedangkan daerah okspital bersifat halus. Padatungkai depan, ujung jari berbentuk bulatankecil, secara berurutan jari ketiga relatif lebihpanjang dari jari pertama, keempat dan jarikedua.. Tungkai belakang berselaput renangpenuh, ujung jari-jari berakhir dengan cakramkecil. Selaput renang antara jari kaki keempatdan kelima berpadanan tinggi dengansubartikular ketiga dan agak cekung. Secaraberurutan jari kaki keempat lebih panjang daripada jari ketiga, kelima, kedua dan jari pertama.Tuberkel tarsal sebelah medial besar danmemanjang, sedangkan tuberkel subatikularberbentuk bulat atau lonjong. Kulit halus,warna kulit pada bagian atas kepala danpunggung coklat terang, kelabu hitam sampaihitam dengan bercak hitam maupun coklat.garis loreal yang membujur dari hidung sampaimata tidak pernah dijumpai. Garis di antarakedua mata tidak jelas. Tanda huruf W padabagian dorsal yang terdapat diantara keduatungkai depan kadang-kadang terlihat denganjelas, kadang-kadang tidak jelas. garispunggung yang membujur dari bagian antaratelinga sampai dubur tidak pernah dijumpai.Permukaan ventral kepala berpigmen. Bibirsering dengan bercak gelap atau aris vertikal(Gambar 4).

Wince Hendri dan Nawir Muhar 287
Gambar 1 Gambar 2
Gambar 3 Gambar 4
Gambar 5
Keterangan Gambar:1. Fajervarya limnocharis2. Rana erytharaea3. Rana chalconata4. Limnoneces sp5. Limnonectes blythii

Wince Hendri dan Nawir Muhar 288
Pada bagian lengan bagian atas kaki depanterdapat lorek tak teratur. Pada bagian dorsalpaha terdapat lorek yang tak teratur, sedangkandi bagian belakang terdapat totol yang takteratur. Pada betis dan tapak tungkai belakangterdapat lorek. Permukaan bawah tungkaibelakang berwarna kuning pucat atau orangepucat seharusnya atau dengan bercak coklat.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :
Jenis kodok yang biasa ditangkap untukdiperjual belikan penduduk di Sumatera Baratadalah jenis kodok Fajervarya limnocharis,Limnonectes blythii, Ranae erytharaea, danRana chalconata (Schlegel) dan Limnoneces sp
Jenis kodok yang paling melimpah ditemukanadalah jenis Fajervarya limnocharis di daerahPasaman dan Painan, Rana erytharaea, danRana chalconata di daerah Pasaman dan Painan.Sedangkan yang paling sedikit ditemukanadalah jenis Limnonectes blythii di Sijunjung,Pasaman dan Mentawai serta Limnonectes sp diPasaman dan Painan
DAFTAR PUSTAKA
Campbell, N.A., J.B. Reece, L.G. Matchell.2003. Biology jilid III. Jakarta: Erlangga.
Djong, T. H., D. I. Iskandar dan D. Gusman.2010. Hubungan Filogenetik SpesiesLimnonectes (Ranidae: Amphibia) asalSumatera Barat dan asal Asia TenggaraBerdasarkan gen 16S Ribosomal RNA.Makara Sains. 14 (1): 79-87
_____________., M. Matsui., M. Kuramoto., D.M. Belabut., Y. H. Sen., M. Nishioka andM. Sumida. 2007. Morphologicaldivergencce, reproduktive isolatingmechanisme, and moleculer phylogeneticrelationship among Indonesia, Malaysia,and Japan Population of Fejervaryalimnocharis oplex (Anura, Ranidae)Zoological Science 24: 1197-1212
Duellman, W. E. and L. Trueb. 1994. Biology ofamphibian. The Johns Hopkins UniversityPress. Baltimore 670. pp.
____________. 1993. Amphibian species of theworld: Additions and Corrections. UnivKansas Publ. Mus. Nat. Hist 21:1-372.
Hellen, Kurniati. 2006. The amphibians speciesin Gunung Halimun National Park, WestJava, Indonesia: 107 – 120. ResearchCenter for Biology, LIPI, Widya satwalokaBuilding, Jalan Raya Cibinong, Cibinong16911, West Java, Indonesia.
Inger, R. F., and D. T. Iskandar, 2005. ACollection of Amphibian from WESTSumatera, “Whith Description of NewSpecies of Megophrys (Amphibia: Anura.The Rafles of Zoology 53(1): 133-142
Iskandar, D. T and W.R. Erdelen. 2006.Conservation of Amphibians and Reptilesin Indonesia: Issues and Problems.Amphib. Reptile Conserv. 4(1):60-93
______________ 1998. Amfibi Jawa dan Bali. 1.Puslitbang Biologi-LIPI. Bogor. 132 hal.
Islam, M. M., Khan, M. M., Tjong, D. H., Alam,M. S., and Sumida, M. (2008). Geneticdifferentiation of the Fejervaryalimnocharis complex from Bangladesh andother Asian countries elucidated byallozyme analyses. Zoological science 25(3), 261.
IUCN (2004), Conservation International, andNatureServe. Global AmphibianAssessment. www.globalamphibians.org
Kusrini, M. D. and R. A. Alford. 2006.Indonesia’s exports of frogs’ legs. TrafficBull. 21(1): 13-24.
_______, M. D., A. Fitri, H. Utama, D. M. Nasir,D. Ardiansyah, V. Lestari and R. Rachmadi.2005. Project 202404: Ecology andconservation of frogs of Mount GedePangrango National Park. Bogor, InstitutPertanian Bogor: 23 hal.
______, M. D., A. Mardiastuti and A. Fitri.2003. Promoting frog conservation throughenvironmental education and researchexperience: Pilot project in west java,indonesia. Dalam: M. D. Kusrini, A.Mardiastuti and T. Harvey (eds) Prosidingseminar hasil penelitian konservasi amfibidan reptil di Indonesia. Bogor, 8 Mei 2003.Departemen Konservasi Sumber dayaHutan. Institut Pertanian 45-51.
Sugiri, N. 1979. Studi beberapa aspek biologikodok batu di beberapa wilayah dankedudukan taksanya. Program Pascasarjana IPB Bogor
Van Kampen, P. N. 1923. The Amphibia of theIndo-Australian Archipelago. E. J. Brill,Ltd. Leiden

ISSN 978-602-14989-0-3 BioETI
Jenis-jenis Hymenoptera sebagai serangga pengunjung padatanaman Mentimun (Cucumis sativus L. Cucurbitaceae) diLubuk Minturun, Kota Padang dan Sungai Pua, KabupatenAgamWITA PUSPITA SARI, HENNY HERWINA DAN DAHELMI
Labor Riset Taksonomi Hewan, Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian mengenai jenis-jenis Hymenoptera sebagai serangga pengunjung pada tanaman mentimun (Cucumis sativus L.Cucurbitaceae) telah dilakukan pada dua lokasi (Lubuk Minturun, Padang dan Sungai Pua, Kabupaten Agam) pada bulanJuni 2013 sampai dengan Januari 2014. Metode yang digunakan adalah pengkoleksian secara langsung menggunakan jalaserangga. Telah didapatkan 11 jenis serangga yang tergolong kedalam 5 famili, 8 genera, dan 93 individu. Familiterbanyak yang ditemukan adalah famili Vespidae (empat jenis) dan Anthoporidae (tiga jenis). Untuk famili Formicidaeditemukan dua jenis, sedangkan Apidae dan Colletidae masing-masing hanya ditemukan satu jenis. Genera yang palingbanyak ditemukan adalah Xylocopa (tiga jenis) dan Delta (dua jenis), sedangkan untuk genus yang lainnya hanyaditemukan masing-masing satu jenis. Jenis dengan individu yang paling banyak ditemukan adalah Xylocopa confusa
Linnaeus, 1902 (60 individu).
Key words: Cucumis sativus, serangga, pengunjung, bunga, jenis
Pendahuluan
Serangga merupakan agen penyerbuk yangsangat penting. Asosiasi antara seranggapenyerbuk (insect pollinators) dengan tanamanmerupakan bentuk asosisasi mutualisme.Asosiasi ini diduga telah terjadi sejak awalCretaceous (sekitar 130-90 juta tahun lalu)melalui proses koevolusi yang menghasilkankeanekaragaman tanaman dan serangga sepertisaat ini. Dominasi tanaman sangat bergantungpada hubungan mutualistik dengan seranggapenyerbuk. Asosisasi mutualisme antaraserangga dengan tanaman bervariasi antarspesies dan terjadi dalam spektrum yang luas.Bagi serangga, asosiasi dengan tanamanmemberi keuntungan, yaitu sebagai sumberpakan berupa serbuk sari (pollen) dan nektar.Serbuk sari mengandung 15-30% protein dannektar mengandung sekitar 50% gula dansenyawa lain, seperti lipid, asam amino,mineral, dan senyawa aromatik (Schoonhoven,Jermy dan Van Loon, 1998).
Menurut Atmowidi (2008), seranggapengunjung pada tanaman jarak pagar dapat
meningkatkan jumlah buah, dengan seranggapengunjung dari ordo Lepidoptera, ColeopteraThysanoptera, Diptera dan Hymenoptera,misalnya pada lebah. Hadirnya lebah sebagaiserangga penyerbuk bagi masyarakat dapatmeningkatkan hasil pertanian baik dari jumlahbuah dan kualitas buah yang dihasilkanterutama tanaman yang tidak dapatmengadakan penyerbukan sendiri. Sebagaicontoh pada tanaman dari Famili Cucurbitaceaeyaitu pare (Momordica charantia) danmentimun (C. sativus) yang memerlukanserangga penyerbuk karena bunga jantan danbetina terpisah (dioceus) (Delaplane dan Mayer,2000).Masing-masing serangga pengunjungbunga juga memiliki daerah sebaran secaravertikal maupun horizontal. Misalnya, Bombusspp. di Indonesia biasanya berada pada daerahpegunungan dengan ketinggian 800-2000 mdpl(Kahono, Erniwati dan Amir, 2005) sehinggaperbedaan ketinggian daerah tanam menjadipenting untuk diperhatikan. Keanekaragamanserangga pengunjung pada ketinggian yangberbeda diperkirakan juga beragam.

Wita Puspita Sari, Henny Herwina dan Dahelmi 283
Pada tanaman mentimun (Cucumis sativus),jumlah kunjungan lebah berpengaruh terhadapbuah yang dihasilkan. Tanaman yangdikunjungi lebah menghasilkan buah tiga kalilebih banyak dibandingkan dari tanaman yangtidak dikunjungi lebah. Kunjungan lebah 6 kalimeningkatkan lebih dari 50% buah, sedangkankunjungan kurang dari 1 kali menyebabkantanaman tidak atau sedikit menghasilkan buah(Gingras et al. 1999 di dalam Atmowidi, 2008).Hymenoptera diperkirakan ada sekitar 300.000jenis yang menempati berbagai tipe ekosistemdi dunia dan yang sudah diberi nama sekitar115.000 jenis. Suatu perbandinganmenyebutkan bahwa jenis Hymenopterasebanding dengan seluruh jumlah vertebratadarat dan air. Salah satu kelompok terbesardalam Hymenoptera adalah Formicidaememiliki jumlah jenis dari kelompok lainnya(Kahono dan Amir, 2003).
Sebagian besar serangga yangmengunjungi bunga adalah dari ordoHymenoptera. Ordo Hymenoptera terbagimenjadi dua subordo (Symphyta danApocrita). Hampir semua subordo Symphytapemakan tumbuhan dan kebanyakan adalahpemakan daun-daunan. Sedangkan subordoApocrita beberapa diantaranya termasukkedalam kelompok pemakan nektar (Borroret al., 1992). Dari ordo ini yang palingbanyak mengujungi bunga dan membantupenyerbukan adalah lebah. Diseluruh duniadiketahui jenis lebah (Apoidea) mencapai16.000 jenis (Michener, 2000).
Kelurahan Lubuk Minturun merupakansalah satu daerah Prima Tani di Sumatera Barat.Luas lahan sawah di Kelurahan LubukMinturun Sungai Lareh adalah 211 ha(BBSDLP, 2007). Tanaman mentimun telahlama diusahakan oleh masyarakat di daerah ini.Luas tanam mentimun 58 ha dengan rata-ratahasil 47,24 kwt/ha (BPS dan Bappeda Padang,2006). Kecamatan Sungai Pua merupakan salahsatu Kecamatan di Kabupaten Agam denganluas wilayah 3.650 Ha. Kecamatan Sungai Puamempunyai beberapa nagari, salah satunya
adalah Nagari Sungai Pua dengan luas wilayah1.213 Ha. Luas lahan yang dimanfaatkansebagai sawah ladang adalah 1061 Ha.Tanaman mentimun merupakan salah satu hasildari perladangan (Pemerintah kabupaten Agam,2013). Informasi mengenai jenis-jenishymenoptera sebagai serangga pengunjungpada tanaman mentimun (C. sativus) sangatsedikit sehingga perlu dilakukan penelitianuntuk melihat jenis-jenis hymenoptera yangmengunjungi tanaman mentimun (C. sativus)pada kedua lokasi diatas.
BAHAN DAN METODE
Penelitian ini dilakukan pada dua lokasi yaitu,Lubuk Minturun, Kota Padang dan Sungai Pua,Kabupaten Agam. Metode yang digunakanpada penelitian ini adalah pengkoleksian secaralangsung dengan menggunakan jala serangga.Pengkoleksian dilakukan mulai pukul 08.00–17.00 WIB saat cuaca cerah pada tanamanmentimun (C. sativus). Setiap pengkoleksiantersebut terbagi atas tiga yaitu pagi (08.00-11.00) WIB, siang (11.00-14.00) WIB, dan sore(14.00-17.00) WIB, selama 4 hari secaraberturut-turut pada masing-masing lokasi.
Serangga yang hinggap pada tanamanmentimun (C. sativus) ditangkap denganmenggunakan jala serangga. Sedangkanserangga-serangga kecil seperti semut langsungdikoleksi secara langsung denganmenggunakan pinset. Selama pengkoleksiandilakukan pencatatan suhu dan kelembapan.Jenis dan jumlah individu serangga pengunjungpada tanaman mentimun (C. sativus)ditampilkan dalam tabel dan grafik, kemudianditampilkan foto dan dibuat deskripsi masing-masingnya.
HASIL DAN PEMBAHASAN
Berdasarkan hasil identifikasi ditemukan 11jenis serangga yang tergolong kedalam 5 famili,8 genera, dan 93 individu. Famili terbanyakyang ditemukan adalah famili Vespidae (empatjenis) dan Anthoporidae (tiga jenis). Untuk

Wita Puspita Sari, Henny Herwina dan Dahelmi 284
famili Formicidae ditemukan dua jenis,sedangkan Apidae dan Colletidae masing-masing hanya ditemukan satu jenis. Generayang paling banyak ditemukan adalah Xylocopa
(tiga jenis) dan Delta (dua jenis), sedangkanuntuk genus yang lainnya hanya ditemukanmasing-masing satu jenis.
Dari Tabel 1 dapat dilihat bahwaHymenoptera didapatkan dengan jumlah jenisyang cukup tinggi (11 Jenis). Genus yangpaling banyak ditemukan jumlah jenisnya yangmengunjungi tanaman mentimun (C. sativus)adalah Xylocopa, dimana selama masapengamatan didapatkan tiga jenis yang berbedayaitu Xylocopa caerulea, Xylocopa confusa danXylocopa latipes. Menurut Delaplane danMayer (2000), Xylocopa merupakan seranggayang mengunjungi semua tanaman yangmemiliki bunga dan salah satunya adalahtanaman mentimun (C. sativus).
Jumlah individu untuk Xylocopa confusa(60 Individu), yang didapatkan pada setiapwaktu pengamatan di kedua lokasi pengamatan.Banyak jumlah individu yang didapatkan,diperkirakan karena berlimpahnya jumlahindividu disekitar lokasi pengamatan sehinggadidapatkan setiap periode waktu pengamatan.Rianti (2009) melaporkan bahwa Xylocopaconfusa kurang dipengaruhi oleh parameterlingkungan yang terjadi. Beberapa penelitianmelaporkan Xylocopa confusa juga merupakanpenyerbuk yang penting pada beberapatanaman. Pada penelitian yang dilakukan olehPateel (2007), Xylocopa merupakan salah satuserangga penyerbuk pada tanaman mentimun(C. sativus) di Dharward, India. Xylocopaconfusa dilaporkan sebagai seranggapengunjung pada bunga caisin (Brassica rapa;Brassicaceae) namun dengan kelimpahanrendah di Taman Nasional Gunung Halimun-Salak, Kab.Sukabumi, Jawa Barat (Atmowidiet al., 2007). Rianti (2009) menyatakan bahwa,Xylocopa confusa merupakan salah satuserangga yang menyerbuki jarak pagar (Jatropacurcas L; Euphobiaceae) di Gunto Indocement
Mining Quarryd, Kec. Kelapa Tunggal, JawaBarat.
Famili Formicidae merupakan Familiserangga kedua dengan jumlah individuterbanyak (dua jenis dan 12 individu). Duajenis yang di dapatkan yaitu Dolichocerus
thoracicus (8 Individu) dan Technomyrmex
albipes (4 Individu). Banyaknya kedua jenis iniditemukan, karena semut memiliki penyebaranyang luas. Sesuai dengan Holldobler danWilson (1990) yang mengatakan bahwa, semutbisa ditemukan dimana-mana pada kawasanterrestrial, akan tetapi mengalami penurunankeanekaragaman jenis sampai pada ketinggian2500 mdpl. Famili ini berkemungkinan dapatmembantu dalam penyerbukan karena adanyarambut tipis pada tubuh walaupun jarakperpindahannya tidak jauh. Menurut Rianti(2009) struktur tubuh semut memiliki rambutyang tipis sehingga memungkinkan adanyaserbuk sari yang melekat saat semut berpindahdari satu bunga ke bunga lainnya.
Selanjutnya masih dari Ordo Hymenoptera,Famili Apidae, di dapatkan satu jenis yaituApis dorsata. Jenis ini pada lokasi 1 didapatkan2 individu dan lokasi 2 di dapatkan 5 individu.Sedikitnya jumlah individu lebah yangdidapatkan pada penelitian ini, dimungkinkankarena sarang A. dorsata yang jauh dari lokasipenelitian. Sola et al (2005) mengatakan bahwa,A. dorsata membuat sarang di pohon tinggi didalam hutan dan Roubik (1989) melaporkanjarak pencarian pakan A. dorsata adalah 6,7-10km dari sarang, sehingga kemungkinanpencarian pakan juga mencakup pada jarak 400m dari tepi hutan. Walaupun jumlah individuyang ditemukan pada penelitian ini sedikit,namun A. dorsata merupakan salah satuserangga penyerbuk yang cukup penting untuktanaman mentimun (C.sativus). Pateel (2007)melaporkan, kelimpahan A. dorsata sebagaiserangga penyerbuk pada tanaman mentimun(C.sativus) dengan persentase 14%. Lebahmadu Apis sudah banyak dikenal sebagaipenyerbuk pada berbagai tanamanAngiospermae (Momose K, 1998) dan berbagai
Wita Puspita Sari, Henny Herwina dan Dahelmi 285
Tabel 1. Daftar jenis, famili dan jumlah individu hymenoptera sebagai serangga pengunjung pada tanaman C. sativuspada masing-masing lokasi penelitian pada setiap waktu pengkoleksian
NoFamili
GenusJenis
Jumlah Individu Lubuk Minturun Sungai PuaA B C A B C
123
AnthoporidaeXylocopaXylocopa caerulea Fabricius, 1804Xylocopa confusa Linnaeus, 1902Xylocopa latipes Drury, 1773
-13-
-10-
-1-
-24-
1192
-12-
-5-
1362
1602
4
ApidaeApisApis dorsata Fabricius, 1793 - 2 - 2 1 3 1 5 7
5Colletidae
Sp. 1 - 1 - 1 1 - - 1 2
6
7
FormicidaeDolichoderus
Dolichoderus thoracicus (F. Smith, 1860)Technomyrmex
Technomyrmex albipes (F. Smith, 1861)
-
-
-
-
4
-
4
-
-
-
-
-
4
4
4
4
8
4
89
10
11
VespidaeDelta
Delta sp. 1Delta sp. 2
EumenesEumenes sp.
VespaVespa affinis Linnaeus, 1764
--
-
-
11
-
1
--
-
-
11
-
1
1-
-
1
--
1
-
1-
1
1
2-
2
2
31
2
3Total Individu 13 16 5 34 26 16 17 59 93
Ket: A (Pagi : 08.00-11.00), B (Siang: 11.00-14.00), C (Sore: 14.00-17.00)- (Tidak ditemukan), (total individu)
Tabel 2. Daftar jenis, family dan jumlah individu hymenoptera sebagai serangga pengunjung pada tanaman C. sativusdibandingkan dengan penelitian serangga pengunjung pada Impatien balsamina dan Nerium oliander
NoFamili
GenusJenis
Impatien balsamina(Khairian, 2012)
Nerium oleander(Yuliani, 2013)
123
AnthoporidaeXylocopaXylocopa caerulea Fabricius, 1804Xylocopa confusa Linnaeus, 1902Xylocopa latipes Drury, 1773
--
-
4
ApidaeApisApis dorsata Fabricius, 1793 - -
5Colletidae
Sp. 1 - -
6
7
FormicidaeDolichoderus
Dolichoderus thoracicus (F. Smith, 1860)Technomyrmex
Technomyrmex albipes (F. Smith, 1861)
-
-
89
10
11
VespidaeDelta
Delta sp. 1Delta sp. 2
EumenesEumenes sp.
VespaVespa affinis Linnaeus, 1764
--
-
-
--
-
-Total Individu 2 3
Ket: A (Pagi : 08.00-11.00), B (Siang: 11.00-14.00), C (Sore: 14.00-17.00)- (Tidak ditemukan)

Wita Puspita Sari, Henny Herwina dan Dahelmi 286
Gambar 1. Jenis-jenis Hymenoptera sebagai serangga pengunjung pada tanaman mentimun (C.sativus) diLubuk Minturun, Kota Padang dan Sungai Pua, Kabupaten Agama. Xylocopa caerulea, b. Xylocopa confusa, c. Xylocopa latipes, d. Apis dorsata,e. Colletidae Sp.1, f. Dolichoderus thoracicus, g. Technomyrmex albipes, h. Eumenes sp., i.Vespa affinis, j. Delta sp. 1, k. Delta sp. 2
a b c
d e f
g h i
j k

Wita Puspita Sari, Henny Herwina dan Dahelmi 287
tanaman di lahan pertanian (Kreman, Williamdan Thorp, 2002). Apis dorsata merupakanpenyerbuk utama pada pertanaman Brassica
rapa (caisin), ditemukan sebanyak 498 individudengan persentase 8,36% (Atmowidi et al.,2007).
Famili Colletidae, di dapatkan satu jenisyaitu Sp.1 (Colletidae), dimana di dapatkan satudi Lubuk Minturun dan satu di Sungai Pua. Jenisini hanya di dapatkan 2 individu selama waktupengamatan. Sedikitinya jumlah individu yangdidapatkan, diperkirakan karena bentuk bungamentimun yang tidak sesuai dengan kelompokini. Famili Colletidae merupakan salah satukelompok lebah berlidah pendek (Short-
tounged), sehingga susah untuk mencapai nektaryang terletak didasar bunga. Michener (2007),berdasarkan analisis morfologinya lebah dibagimenjadi dua kelompok, yaitu lebah berlidahpanjang (Apidae, Megachilidae) dan yangberlidah pendek (Meganomidae, Mellitidae,Andrenidae, Halictidae, Stenotritidae danColletidae). Namun Famili ini merupakan salahsatu serangga penyerbuk pada tanaman Ourisiaglandulosa dan Wahlenbergia albomarginatadengan persentase 90% dan 95%. Famili inidikatakan efektif sebagai serangga penyerbukkarena dapat membawa 10 serbuk sari padasetiap kunjungan ke Ourisia glandulosa danmembawa 3 serbuk sari pada setiap kunjunganke Wahlenbergia albomarginata (Bischoff,2012).
Dari Tabel 2. Dapat dilihat jumlah jenis yangditemukan pada penelitian ini lebih banyakdibandingkan dengan penelitian yangsebelumnya pernah dilakukan mengenaiserangga pengunjung, Khairiah (2012) padatanaman Impatiens balsamina yangmendapatkan dua jenis, begitupun jikadibandingkan dengan Yuliani (2013) padatanaman Nerium oleander menemukan tigajenis. Menurut Gilman (1999), adanyaperbedaan jenis serangga yang berkunjungdipengaruhi oleh perbedaan karakter bunga.Keanekaragaman serangga berkaitan denganbanyaknya bunga yang dihasilkan oleh
tumbuhan. Banyaknya jenis yang didapatkanpada penelitian ini, diperkirakan karena adanyaperbedaan jenis tanaman, vegetasi dan kondisilingkungan, serta lokasi penelitian yang beradadi dekat area persawahan dan adanya aliran airdi sekitarnya.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Jenis Hymenoptera sebagai seranggapengunjung pada tanaman mentimun (Cucumissativus L.) telah didapatkan sebanyak 11 jenisyang tergolong kedalam 5 famili, 8 genera, dan93 individu. Famili terbanyak yang ditemukanadalah famili Vespidae (empat jenis) diikuti olehAnthoporidae (tiga jenis). Pada Formicidaeditemukan dua jenis, sedangkan Apidae danColletidae masing-masing hanya ditemukan satujenis serangga pengunjung.
Ucapan Terima kasihTerima kasih ditujukan kepada Dra. IzmiartiMS, Dr. Rizaldi dan Dr. Resti Rahayu yangtelah memberikan saran dan masukan padapenelitian ini. Terima kasih kepada Prof. Jun-Ichi Kojima dari Ibaraki University, Jepang danRijal Satria M.Sc dari Tokyo MetropolitanUniversity, Jepang atas bantuannya untukpengidentifikasian dan penulisan artikel ini.
DAFTAR PUSTAKA
Atmowidi T., Buchori, D., Manuwoto, S.,Suryobroto, B. dan Hidayat, P. 2007.Diversity of pollinator insects in relation ofseed set of Mustard (Brassica rappa L:Crusiferae). Hayati Journal Bioscience14:155-161.
Atmowidi, T. 2008. Keanekaragaman danPerilaku Kunjungan Serangga Penyerbukserta Pengaruhnya dalam PembentukanBiji Tanaman Caisin (Brassica rapaL.:Brassicaceae) [Disertasi]. Bogor:Institut Pertanian Bogor. (unpublish)
Bischoff, M., Campbell, D.R., Lord, J.M. danRobertson, A.W. 2012. The relativeimportance of solitary bees and shyrphid

Wita Puspita Sari, Henny Herwina dan Dahelmi 288
flies as pollinators of two outcrossing plantspecies in the New Zealand alpine. AustralEcology 20: 1-9.
Borror, D. J, Johnson, N.F. dan Triplehorn, C.A.1992. Pengenalan Pelajaran Serangga.Diterjemahkan oleh Suryobroto, M.Gadjah Mada University Press. Jogjakarta
Delaplane, K. S., D.F. Mayer. 2000. Croppollination by bees. CABI Publishing.Oxon.
Gilman, E.F. 1999. Hoya carnosa. CooperativeExtention Servis Institute of food andagriculture science. University of FloridaPr. Florida.
Hölldobler, B. dan Wilson, E.O. 1990. The Ants.Harvard University Press. Cambridge.USA
Kahono, S., Erniwati. dan Amir, M. 2005.Evaluasi Serangga Penyerbuk danPenyerbukan di Jawa: Pemilihan JenisPotensial sebagai Dasar PengembanganJenis dan Konservasinya. Laporan Teknik.Pusat Penelitian Biologi-LIPI: 789-797.
Kreman C., William M. N. dan Thorp R. W.2002. Crop pollination from native bees atrisk from agricultural intensification. PNAS99: 16812-16819.
Michener, C. D. 2003. The Bees of the World.The John Hopkins Univ. Press. Baltimore.
Michener, C. D. 2007. The Bees of the World.The John Hopkins Univ. Press. Baltimore.
Momose, K. 1998. Pollination biology on adipterocarp forest in Sarawak Malaysia.Characteristic of the plant pollinatorcommunity in the lowland dipterocarpforest. American Journal of Botany 10:1477-1501.
Pateel, M. C. 2007. Impact of honey beepollination on qualitative and quantitativeparameters of cucumber (Cucumis sativusL). Thesis of Graduate Master of ScienceAgricultural Entomologi, Dharwarduniversity. Dharward. (unpublish)
Rianti, P. 2009. Keanekaragaman, Efektifitas,dan Frekuensi kunjungan SeranggaPenyerbuk pada Tanaman Jarak Pagar(Jatropha curcas L: Euphorbiaceae).Thesis Pasca Sarjana Institut PertanianBogor. Bogor (unpublish)
Roubik, D.W. 1989. Ecology and NaturalHistory of Tropical bees. Cambridge Univ.Press. New York
Schoonhoven, L. M., Jermy, T. dan Van Loon,J.J.A. 1998. Physiology to Evolution:Insect-Plant Biology. Chapman and Hall.London.
Sola E., Widyaningrum, I.K. dan Mulyati, S.2005. A Photographic Guide to theCommon Insect of Gunung Halimun-SalakNational Park. Taman Nasional GunungHalimun Salak. Bogor

ISSN 978-602-14989-0-3 BioETI
Studi eksplorasi bakteri dari saluran pencernaan Ikan Nila(Oreochromis niloticus) yang dibudidayakan di Karamba JaringApung Danau Maninjau, Sumatera BaratYEMPITA EFENDI DAN YUSRA
Fakultas Perikanan dan Ilmu Kelautan, Universitas Bung Hatta, PadangE-mail:
ABSTRAKIkan Nila (Oreochromis niloticus) merupakan salah satu jenis ikan yang dibudidayakan di Karamba Jaring Apung (KJA)Danau Maninjau, Sumatera Barat. Penelitian ini bertujuan mengetahui jenis pakan dan bakteri yang terdapat pada saluranpencernaan ikan Nila yang diharapkan dapat dijadikan sebagai kandidat probiotik. Materi yang digunakan dalampenelitian ini adalah ikan Nila yang berasal dari petani ikan yang berada di sekitar Danau Maninjau. Metode penelitianyang digunakan adalah eksplorasi dan eksperiment. Pakan yang biasa digunakan oleh petani adalah Almabar, Bintang,Cargil, Comfeed dan Sinta. Dari identifikasi ditemukan 55 koloni bakteri yang secara morfologi dan biokimiadikelompokkan ke dalam tiga genus yakni Bacillus, Achromobacter dan Enterobacter.
Key words: eksplorasi, bakteri, Nila (Oreochromis niloticus), karamba jaring apung
Pendahuluan
Danau Maninjau merupakan salah satu perairanumum yang terletak di Kecamatan TanjungRaya, Kabupaten Agam, Propinsi SumateraBarat. Kegiatan perikanan yang berlangsungterdiri dari perikanan budidaya di karambajaring apung (KJA) dan perikanan tangkap.Syandri (2013) kegiatan budidaya ikan di KJAdimulai semenjak tahun 1992 dengan jumlahKJA 12 unit, pada tahun 1997 meningkatmenjadi 2854 unit, sampai sekarang sudahberkembang menjadi 13.000 unit
Usaha perikanan KJA memang berdampakpada peningkatan kesejahteraan penduduk,namun seiring denganpeningkatan jumlah KJAtersebut juga berdampak pada meningkatnyalimbah yang pada akhirnya memberikandampak negatif terhadap lingkungan perairan.Terjadinya eutrofikasi yang lebih cepat denganfrekuensi yang sering, sehingga menyebabkanmutu perairan menjadi menurun (Krismono danKrismono, 1998). Demikian juga halnyadengan limbah sisa pakan dan kotoran ikanyang menumpuk di dasar perairan danau.Beveridge (1996) limbah dari budidaya KJAadalah berupa makanan yang tidak dikonsumsi,feses, dan urin termasuk mikroorganisme,
parasit, dan organisme lainnya yang terdapat didalamnya. Peningkatan unsur hara daripenguraian sisa pakan akan mempercepatterjadinya eutrofikasi .
Upaya untuk menurunkan potensieutrofikasi perairan danau adalah denganmengurangi beban N dan P dari pakan yangdigunakan dalam budidaya. Tangko et al.,(2007), upaya yang dapat dilakukan untuk lebihmeningkatkan kualitas pakan adalah denganmenambahkan bahan aditif berupa probiotik.Prinsip dasar kerja probiotik adalah denganmemanfaatkan kemampuan mikroba untukmempermudah penyerapan oleh saluranpencernaan ikan (Feliatra dan Suryadi, 2004).
Probiotik tergolong dalam makananfungsional, dimana bahan makanan inimengandung komponen–komponen yang dapatmeningkatkan kesehatan ternak danmengefesiensikan pakan dengan caramanipulasi komposisi bakteri yang ada dalampakan udang. Namun tidak semua bakteri yangberpotensi sebagai probiotik. Perlu adanyaidentifikasi secara genetik yang menunjukanhasil identifikasi secara spesifik. Untuk ituperlu diketahui karakteristik bakteri probiotikyang terdapat pada ikan Nila (Tilapia nilotica),terutama dalam penemuan spesies baru atau

Yempita Efendi dan Yusra 290
galur bakteri lokal sehingga dapat memperkayaisolat bakteri asli Indonesia. Tujuan penelitianini adalah untuk mengisolasi, identifikasi dankarakterisasi bakteri yang berasal dari saluranpencernaan ikan Nila (Oreochromis niloticus)sebagai kandidat probiotik yang diharapkansebagai salah satu alternatif untukmengantisipasi terjadinya kasus kematianmasal ikan di KJA Danau Maninjau.
BAHAN DAN METODE
Bahan yang dipergunakan dalam penelitian iniadalah ikan Nila (Oreochromis niloticus) yangdibeli dari petani ikan yang terletak di sekitarDanau Maninjau. Pengambilan sampel di KJAberdasarkan jenis pakan ditentukan secarapurpossive sampling.
Bahan-bahan kimia yang akan digunakandalam penelitian ini adalah: bacto agar (Oxoid),CaCO3, aquadest steril, alkohol, spiritus. Untukpewarnaan Gram dibutuhkan zat warna crystalviolet, lugol, alkohol 96%, 70%, safranin, H2O2
3%, pewarna malacyt green. Uji uji biokimiadigunakan KOH 1%, NaCl, potasium kromat,perak nitrat, fenoftalein, p-aminodimetilanilinoksalat 1%, NaOH 0,1N, minyak immersi,bromthymol blue, pereaksi kovacs dan asamsulfanilat.
Media yang digunakan adalah Glukosa
Tripton Agar (GTA) + CaCO3, TSA(trypticase soy agar), trypton broth, sulfit agar,nitrat broth, TSIA (triple sugar iron agar),Baird Parker Agar (BPA), brain heart infusion
(BHI) dan Simmons citrate.Isolasi bakteri saluran pencernaan dilakukan
dengan teknik pourplate pada media glukosetripton agar + CaCO3. Satu gram homogenatsaluran pencernaan disuspensikan ke dalam 9mL air laut streril kemudian dibuat seripengenceran hingga 10-8. Masing masing seripengenceran ditanam pada media nutrient agarkemudian diinkubasi selama 48 jam pada suhu37 oC.
Identifikasi isolat bakteri meliputikarakteristik morfologi dan biokimia bakteriyakni: pewarnaan Gram, pewarnaan spora,
motilitas, uji TSIA, pembentukan gas, katalase,oksidase, motilitas, indol, urea, sitrat, laktosa,glukosa, sukrosa, MR dan VP, OF test, reduksinitrat dan gelatin (Fardiaz, 1989; Hadioetomo,1985 dan Lay, 1994).Uji Aktivitas Produksi Enzim ProteolitikUji ini dilakukan dengan prosedur (Jacob danGerstein, 1960) dalam Bairagi et al., (2002).Isolat-isolat yang diperoleh dari hasil isolasi diinokulasikan dengan cara streak pada mediaagar yang diperkaya dengan skim milk (4%).Inkubasi pada 37°C selama 24 jam. Adanyaaktivitas produksi enzimproteolitik ditunjukanoleh terbentuknya zona bening.
HASIL DAN PEMBAHASAN
Berdasarkan penelitian diketahui lima jenispakan yang biasa digunakan oleh petanipembudidaya ikan Nila (Tilapia nilotica) yangberada di sekitar Danau Maninjau. Sebaranjenis pakan yang biasa digunakan dapat dilihatpada Tabel 1.
Tabel 1. Sebaran Jenis Pakan yang DigunakanPetani Ikan .
No Jenis Pakan12345
AlmabarBintangCargilComfeedSinta
Berdasarkan Tabel 1 terlihat bahwa jenispakan yang biasa digunakan oleh petanipembudidaya ikan di KJA yang terdapat disekitar Danau Maninjau adalah lima jenis yaknialmabar, bintang, cargil, comfeed dan sinta,namun pada waktu penelitian dilakukan ikanyang diambil sebagai sampel hanya berasal dari4 pakan yakni Almabar, Bintang, Comfeed danSinta. Hal ini karena petani yang menggunakanpakan Cargil baru melakukan pemanenanterhadap ikan yang mereka budidayakan.Untuk mengetahui jenis bakteri dari saluranpencernaan ikan Nila (Tilapia nilotica) yangakan dijadikan kandidat probiotik dilakukan
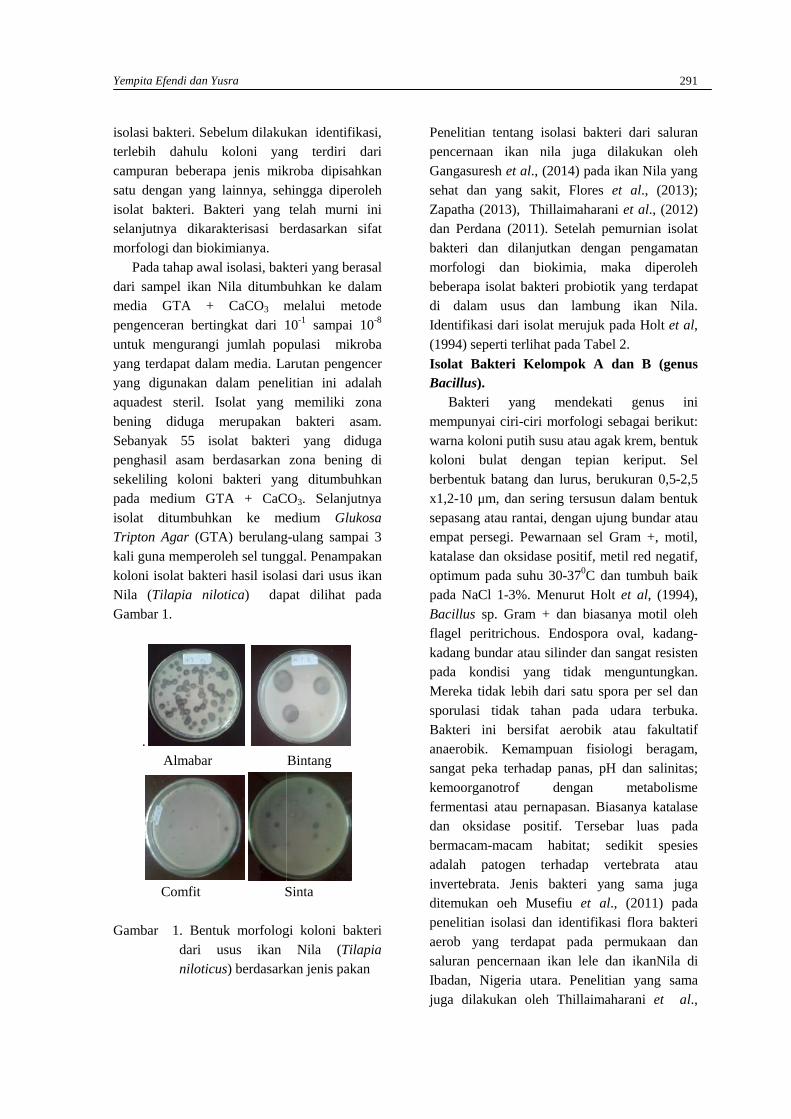
Yempita Efendi dan Yusra 291
isolasi bakteri. Sebelum dilakukan identifikasi,terlebih dahulu koloni yang terdiri daricampuran beberapa jenis mikroba dipisahkansatu dengan yang lainnya, sehingga diperolehisolat bakteri. Bakteri yang telah murni iniselanjutnya dikarakterisasi berdasarkan sifatmorfologi dan biokimianya.
Pada tahap awal isolasi, bakteri yang berasaldari sampel ikan Nila ditumbuhkan ke dalammedia GTA + CaCO3 melalui metodepengenceran bertingkat dari 10-1 sampai 10-8
untuk mengurangi jumlah populasi mikrobayang terdapat dalam media. Larutan pengenceryang digunakan dalam penelitian ini adalahaquadest steril. Isolat yang memiliki zonabening diduga merupakan bakteri asam.Sebanyak 55 isolat bakteri yang didugapenghasil asam berdasarkan zona bening disekeliling koloni bakteri yang ditumbuhkanpada medium GTA + CaCO3. Selanjutnyaisolat ditumbuhkan ke medium GlukosaTripton Agar (GTA) berulang-ulang sampai 3kali guna memperoleh sel tunggal. Penampakankoloni isolat bakteri hasil isolasi dari usus ikanNila (Tilapia nilotica) dapat dilihat padaGambar 1.
.Almabar Bintang
Comfit Sinta
Gambar 1. Bentuk morfologi koloni bakteridari usus ikan Nila (Tilapianiloticus) berdasarkan jenis pakan
Penelitian tentang isolasi bakteri dari saluranpencernaan ikan nila juga dilakukan olehGangasuresh et al., (2014) pada ikan Nila yangsehat dan yang sakit, Flores et al., (2013);Zapatha (2013), Thillaimaharani et al., (2012)dan Perdana (2011). Setelah pemurnian isolatbakteri dan dilanjutkan dengan pengamatanmorfologi dan biokimia, maka diperolehbeberapa isolat bakteri probiotik yang terdapatdi dalam usus dan lambung ikan Nila.Identifikasi dari isolat merujuk pada Holt et al,(1994) seperti terlihat pada Tabel 2.
Isolat Bakteri Kelompok A dan B (genusBacillus).
Bakteri yang mendekati genus inimempunyai ciri-ciri morfologi sebagai berikut:warna koloni putih susu atau agak krem, bentukkoloni bulat dengan tepian keriput. Selberbentuk batang dan lurus, berukuran 0,5-2,5x1,2-10 μm, dan sering tersusun dalam bentuksepasang atau rantai, dengan ujung bundar atauempat persegi. Pewarnaan sel Gram +, motil,katalase dan oksidase positif, metil red negatif,optimum pada suhu 30-370C dan tumbuh baikpada NaCl 1-3%. Menurut Holt et al, (1994),Bacillus sp. Gram + dan biasanya motil olehflagel peritrichous. Endospora oval, kadang-kadang bundar atau silinder dan sangat resistenpada kondisi yang tidak menguntungkan.Mereka tidak lebih dari satu spora per sel dansporulasi tidak tahan pada udara terbuka.Bakteri ini bersifat aerobik atau fakultatifanaerobik. Kemampuan fisiologi beragam,sangat peka terhadap panas, pH dan salinitas;kemoorganotrof dengan metabolismefermentasi atau pernapasan. Biasanya katalasedan oksidase positif. Tersebar luas padabermacam-macam habitat; sedikit spesiesadalah patogen terhadap vertebrata atauinvertebrata. Jenis bakteri yang sama jugaditemukan oeh Musefiu et al., (2011) padapenelitian isolasi dan identifikasi flora bakteriaerob yang terdapat pada permukaan dansaluran pencernaan ikan lele dan ikanNila diIbadan, Nigeria utara. Penelitian yang samajuga dilakukan oleh Thillaimaharani et al.,
Yempita Efendi dan Yusra 291
isolasi bakteri. Sebelum dilakukan identifikasi,terlebih dahulu koloni yang terdiri daricampuran beberapa jenis mikroba dipisahkansatu dengan yang lainnya, sehingga diperolehisolat bakteri. Bakteri yang telah murni iniselanjutnya dikarakterisasi berdasarkan sifatmorfologi dan biokimianya.
Pada tahap awal isolasi, bakteri yang berasaldari sampel ikan Nila ditumbuhkan ke dalammedia GTA + CaCO3 melalui metodepengenceran bertingkat dari 10-1 sampai 10-8
untuk mengurangi jumlah populasi mikrobayang terdapat dalam media. Larutan pengenceryang digunakan dalam penelitian ini adalahaquadest steril. Isolat yang memiliki zonabening diduga merupakan bakteri asam.Sebanyak 55 isolat bakteri yang didugapenghasil asam berdasarkan zona bening disekeliling koloni bakteri yang ditumbuhkanpada medium GTA + CaCO3. Selanjutnyaisolat ditumbuhkan ke medium GlukosaTripton Agar (GTA) berulang-ulang sampai 3kali guna memperoleh sel tunggal. Penampakankoloni isolat bakteri hasil isolasi dari usus ikanNila (Tilapia nilotica) dapat dilihat padaGambar 1.
.Almabar Bintang
Comfit Sinta
Gambar 1. Bentuk morfologi koloni bakteridari usus ikan Nila (Tilapianiloticus) berdasarkan jenis pakan
Penelitian tentang isolasi bakteri dari saluranpencernaan ikan nila juga dilakukan olehGangasuresh et al., (2014) pada ikan Nila yangsehat dan yang sakit, Flores et al., (2013);Zapatha (2013), Thillaimaharani et al., (2012)dan Perdana (2011). Setelah pemurnian isolatbakteri dan dilanjutkan dengan pengamatanmorfologi dan biokimia, maka diperolehbeberapa isolat bakteri probiotik yang terdapatdi dalam usus dan lambung ikan Nila.Identifikasi dari isolat merujuk pada Holt et al,(1994) seperti terlihat pada Tabel 2.
Isolat Bakteri Kelompok A dan B (genusBacillus).
Bakteri yang mendekati genus inimempunyai ciri-ciri morfologi sebagai berikut:warna koloni putih susu atau agak krem, bentukkoloni bulat dengan tepian keriput. Selberbentuk batang dan lurus, berukuran 0,5-2,5x1,2-10 μm, dan sering tersusun dalam bentuksepasang atau rantai, dengan ujung bundar atauempat persegi. Pewarnaan sel Gram +, motil,katalase dan oksidase positif, metil red negatif,optimum pada suhu 30-370C dan tumbuh baikpada NaCl 1-3%. Menurut Holt et al, (1994),Bacillus sp. Gram + dan biasanya motil olehflagel peritrichous. Endospora oval, kadang-kadang bundar atau silinder dan sangat resistenpada kondisi yang tidak menguntungkan.Mereka tidak lebih dari satu spora per sel dansporulasi tidak tahan pada udara terbuka.Bakteri ini bersifat aerobik atau fakultatifanaerobik. Kemampuan fisiologi beragam,sangat peka terhadap panas, pH dan salinitas;kemoorganotrof dengan metabolismefermentasi atau pernapasan. Biasanya katalasedan oksidase positif. Tersebar luas padabermacam-macam habitat; sedikit spesiesadalah patogen terhadap vertebrata atauinvertebrata. Jenis bakteri yang sama jugaditemukan oeh Musefiu et al., (2011) padapenelitian isolasi dan identifikasi flora bakteriaerob yang terdapat pada permukaan dansaluran pencernaan ikan lele dan ikanNila diIbadan, Nigeria utara. Penelitian yang samajuga dilakukan oleh Thillaimaharani et al.,
Yempita Efendi dan Yusra 291
isolasi bakteri. Sebelum dilakukan identifikasi,terlebih dahulu koloni yang terdiri daricampuran beberapa jenis mikroba dipisahkansatu dengan yang lainnya, sehingga diperolehisolat bakteri. Bakteri yang telah murni iniselanjutnya dikarakterisasi berdasarkan sifatmorfologi dan biokimianya.
Pada tahap awal isolasi, bakteri yang berasaldari sampel ikan Nila ditumbuhkan ke dalammedia GTA + CaCO3 melalui metodepengenceran bertingkat dari 10-1 sampai 10-8
untuk mengurangi jumlah populasi mikrobayang terdapat dalam media. Larutan pengenceryang digunakan dalam penelitian ini adalahaquadest steril. Isolat yang memiliki zonabening diduga merupakan bakteri asam.Sebanyak 55 isolat bakteri yang didugapenghasil asam berdasarkan zona bening disekeliling koloni bakteri yang ditumbuhkanpada medium GTA + CaCO3. Selanjutnyaisolat ditumbuhkan ke medium GlukosaTripton Agar (GTA) berulang-ulang sampai 3kali guna memperoleh sel tunggal. Penampakankoloni isolat bakteri hasil isolasi dari usus ikanNila (Tilapia nilotica) dapat dilihat padaGambar 1.
.Almabar Bintang
Comfit Sinta
Gambar 1. Bentuk morfologi koloni bakteridari usus ikan Nila (Tilapianiloticus) berdasarkan jenis pakan
Penelitian tentang isolasi bakteri dari saluranpencernaan ikan nila juga dilakukan olehGangasuresh et al., (2014) pada ikan Nila yangsehat dan yang sakit, Flores et al., (2013);Zapatha (2013), Thillaimaharani et al., (2012)dan Perdana (2011). Setelah pemurnian isolatbakteri dan dilanjutkan dengan pengamatanmorfologi dan biokimia, maka diperolehbeberapa isolat bakteri probiotik yang terdapatdi dalam usus dan lambung ikan Nila.Identifikasi dari isolat merujuk pada Holt et al,(1994) seperti terlihat pada Tabel 2.
Isolat Bakteri Kelompok A dan B (genusBacillus).
Bakteri yang mendekati genus inimempunyai ciri-ciri morfologi sebagai berikut:warna koloni putih susu atau agak krem, bentukkoloni bulat dengan tepian keriput. Selberbentuk batang dan lurus, berukuran 0,5-2,5x1,2-10 μm, dan sering tersusun dalam bentuksepasang atau rantai, dengan ujung bundar atauempat persegi. Pewarnaan sel Gram +, motil,katalase dan oksidase positif, metil red negatif,optimum pada suhu 30-370C dan tumbuh baikpada NaCl 1-3%. Menurut Holt et al, (1994),Bacillus sp. Gram + dan biasanya motil olehflagel peritrichous. Endospora oval, kadang-kadang bundar atau silinder dan sangat resistenpada kondisi yang tidak menguntungkan.Mereka tidak lebih dari satu spora per sel dansporulasi tidak tahan pada udara terbuka.Bakteri ini bersifat aerobik atau fakultatifanaerobik. Kemampuan fisiologi beragam,sangat peka terhadap panas, pH dan salinitas;kemoorganotrof dengan metabolismefermentasi atau pernapasan. Biasanya katalasedan oksidase positif. Tersebar luas padabermacam-macam habitat; sedikit spesiesadalah patogen terhadap vertebrata atauinvertebrata. Jenis bakteri yang sama jugaditemukan oeh Musefiu et al., (2011) padapenelitian isolasi dan identifikasi flora bakteriaerob yang terdapat pada permukaan dansaluran pencernaan ikan lele dan ikanNila diIbadan, Nigeria utara. Penelitian yang samajuga dilakukan oleh Thillaimaharani et al.,
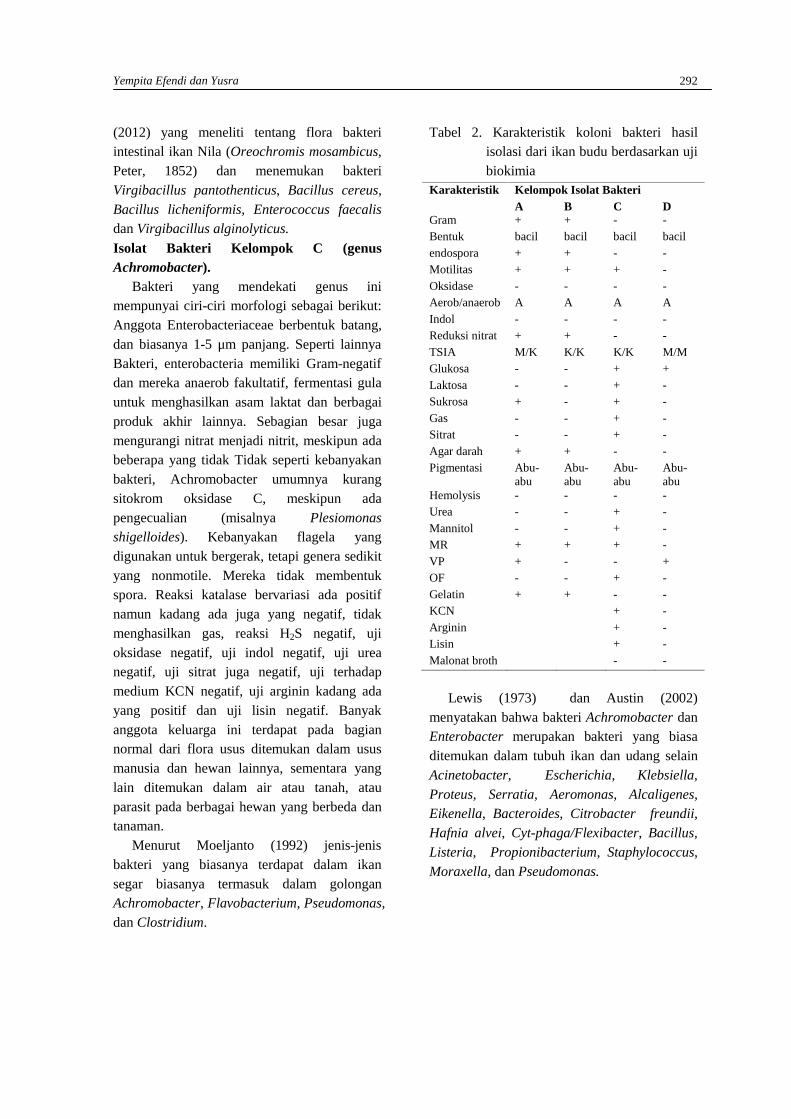
Yempita Efendi dan Yusra 292
(2012) yang meneliti tentang flora bakteriintestinal ikan Nila (Oreochromis mosambicus,Peter, 1852) dan menemukan bakteriVirgibacillus pantothenticus, Bacillus cereus,
Bacillus licheniformis, Enterococcus faecalisdan Virgibacillus alginolyticus.
Isolat Bakteri Kelompok C (genusAchromobacter).
Bakteri yang mendekati genus inimempunyai ciri-ciri morfologi sebagai berikut:Anggota Enterobacteriaceae berbentuk batang,dan biasanya 1-5 μm panjang. Seperti lainnyaBakteri, enterobacteria memiliki Gram-negatifdan mereka anaerob fakultatif, fermentasi gulauntuk menghasilkan asam laktat dan berbagaiproduk akhir lainnya. Sebagian besar jugamengurangi nitrat menjadi nitrit, meskipun adabeberapa yang tidak Tidak seperti kebanyakanbakteri, Achromobacter umumnya kurangsitokrom oksidase C, meskipun adapengecualian (misalnya Plesiomonasshigelloides). Kebanyakan flagela yangdigunakan untuk bergerak, tetapi genera sedikityang nonmotile. Mereka tidak membentukspora. Reaksi katalase bervariasi ada positifnamun kadang ada juga yang negatif, tidakmenghasilkan gas, reaksi H2S negatif, ujioksidase negatif, uji indol negatif, uji ureanegatif, uji sitrat juga negatif, uji terhadapmedium KCN negatif, uji arginin kadang adayang positif dan uji lisin negatif. Banyakanggota keluarga ini terdapat pada bagiannormal dari flora usus ditemukan dalam ususmanusia dan hewan lainnya, sementara yanglain ditemukan dalam air atau tanah, atauparasit pada berbagai hewan yang berbeda dantanaman.
Menurut Moeljanto (1992) jenis-jenisbakteri yang biasanya terdapat dalam ikansegar biasanya termasuk dalam golonganAchromobacter, Flavobacterium, Pseudomonas,
dan Clostridium.
Tabel 2. Karakteristik koloni bakteri hasilisolasi dari ikan budu berdasarkan ujibiokimia
Karakteristik Kelompok Isolat BakteriA B C D
Gram + + - -Bentuk bacil bacil bacil bacilendospora + + - -Motilitas + + + -Oksidase - - - -Aerob/anaerob A A A AIndol - - - -Reduksi nitrat + + - -TSIA M/K K/K K/K M/MGlukosa - - + +Laktosa - - + -Sukrosa + - + -Gas - - + -Sitrat - - + -Agar darah + + - -Pigmentasi Abu-
abuAbu-abu
Abu-abu
Abu-abu
Hemolysis - - - -Urea - - + -Mannitol - - + -MR + + + -VP + - - +OF - - + -Gelatin + + - -KCN + -Arginin + -Lisin + -Malonat broth - -
Lewis (1973) dan Austin (2002)menyatakan bahwa bakteri Achromobacter danEnterobacter merupakan bakteri yang biasaditemukan dalam tubuh ikan dan udang selainAcinetobacter, Escherichia, Klebsiella,
Proteus, Serratia, Aeromonas, Alcaligenes,Eikenella, Bacteroides, Citrobacter freundii,Hafnia alvei, Cyt-phaga/Flexibacter, Bacillus,
Listeria, Propionibacterium, Staphylococcus,Moraxella, dan Pseudomonas.

Yempita Efendi dan Yusra 293
Isolat Bakteri Kelompok D (genusEnterobacter)
Enterobacter merupakan bakteri gramnegatif yang berbentuk basil, dengan ukuran0,6 – 1,0 µm x 1,2 – 3,0 µm, motil, tidakmembentuk spora, berkapsul, bersifat aerob,menghasilkan gas dan memiliki flagel. Bakteriini sering ditemukan bersama Escherichiacoli hidup bebas di alam seperti di air, tanahdan juga di saluran pencernaan manusia danhewan. Oksidase negatif, katalase ada yangpositif dan kadang negatif, motil, uji sitratpositif, indol negatif, uji urease positif, sitratpositif, uji gula (laktosa, glukosa dan sukrosa)positif, uji terhadap manitol positif, uji VPnegatif dan uji OF positif.
Sifat pertumbuhan dari Enterobacter yaitudapat tumbuh baik hampir di semua mediabuatan pada laboratorium mikrobiologi. Bentukkoloni Enterobacter (Aerobacter aeroge nes)besar, berwarna putih-merah, keruh, cembung,bulat dan halus. Selain itu A. aerogenes jugamengurai karbohidrat seperti glukosa danlaktosa menjadi asam dan gas sepertihalnya Escherichia coli. Enterobacteraerogenes dapat hidup sebagai saprobe disaluran pencernaan hewan danmanusia. Enterobacter aerogenes adalah salahsatu jenis bakteri coliform, yang merupakankelompok bakteri yang digunakan sebagaiindikator kondisi sanitasi yang tidak baikterhadap makanan dan minuman.
Bakteri ini juga ditemukan pada penelitianHarbi et al., (2012) yang meneliti bakteri yangterdapat di dalam saluran pencernaan ikan Mas(Cyprinus carpio) yang di bekukan, Takroo danRitu (2011) yang melakukan isolasi bakteri dariikan Rohu (Labeo rohita) di India.
Deteksi produksi enzim proteolitikekstraseluler
Kemampuan memproduksi enzimproteolitik ekstraseluler dideteksi menggunakanmedium pengujian yaitu medium yangdiperkaya dengan substrat enzimnya (skimmilk). Pendeteksian didasarkan padaterbentuknya zona hidrolisis disekitarkoloni
bakteri yang diuji. Hasil uji produksi enzimproteolitik ekstraseluler menunjukan bahwasemua isolate uji memproduksi enzimproteolitik ekstraseluler.
Pada penelitian ini difokuskan salah satunyapada jenis enzim proteolitik karena komponenutama pakan ikan adalah protein. Berdasarkanhasil penelitian diperoleh seluruh isolatmempunyai kemampuan menghasilkan enzimproteolitik ekstraseluler. Kemampuan bakteriprobiotik untuk memproduksi enzim proteolitikekstraseluler mempunyai peranan pentingdalam ikut serta mencerna senyawa-senyawayang bersifat protein. Hasil penelitianGeovanny dan Shen (2008) menunjukanadanya peningkatan yang nyata pada aktivitasenzim proteolitik pada udang yang diberiperlakuan probiotik dibandingkan kontrol.Penelitian yang sama mengenai pengaruhpemberian probiotik terhadap aktivitas enzimpencernaan juga dilakukan oleh Ziaei-Nejad etal. (2006). Zhou et al., (2009). Musikasang etal. (2009) menetapkan kemampuan untukmencerna protein sebagai salah satu kriteriaseleksi probiotik. Adanya enzim proteolitik iniselanjutnya akan meningkatkan jumlahsenyawa yang bersifat protein yang dicernasehingga menurunkan jumlah limbah yangmengandung Nitrogen yang berasal dari prosespencernaan. Hal ini menguntungkan karenaakan menekan jumlah amonia yang berasal dariproses mineralisasi N-organik yang diharapkandapat memecahkan masalah kematian masalikan yang sering terjadi di Danau Maninjau.Penelitian tentang uji aktivitas enzimproteolitik yang terdapat pada saluranpencernaan ikan sebagai kandidat probiotikjuga dilakukan oleh Subagyo dan Djunaedi(2011) menemukan seluruh isolat mempunyaikemampuan menghasilkan enzim proteolitikprotease. Hasil penelitian Mubarik et al.,(2006) menunjukkan bahwa isolat NU-2merupakan bakteri proteolitik dengan nilaiIndeks Proteolitik (IP) sebesar 1.89 danberpotensi untuk dijadikan probiotik karena

Yempita Efendi dan Yusra 294
mampu menghasilkan baik protease, α-amilase,dan glukoamilase ekstraseluler.
KESIMPULAN
Berdasarkan hasil seleksi diperoleh 55 isolatbakteri dari saluran pencernaan ikan Nila(Tilapia nilotica) yang secara morfologi danbiokimia dikelompokkan ke dalam tiga genusyakni Bacillus, Achromobacter danEnterobacter. Berdasarkan produksi enzimproteolitik ekstra-selluler diketahui bahwabakteri dari genus Bacillus dapat dijadikansebagai kandidat probiotik.
UCAPAN TERIMA KASIH
Penelitian ini dibiayai oleh Direktorat JenderalPendidikan Tinggi, Kementerian PendidikanNasional, melalui DIPA Kopertis Wilayah XTahun 2014 Nomor SP DIPA-023.04.2.532476/2014 tanggal 5 Desember 2013,sesuai dengan Surat Perjanjian Hibah PenelitianNomor: 01/Kontrak/010/KM/2014
DAFTAR PUSTAKA
Austin, B. 2002. The bacterial microflora offish. Mini-Review. The Scientific WorldJournal. 2: 558.572
Bairagi, A., K. Ghosh, S. Kumarsen dan A. K.Ray, 2002. Enzyme producing bacterialflora isolated from fish digestive tracts.Aquaculture International, 10: 109–121.
Beveridge M.C.M., 1996, Cage aquaculture,fishing news books, Oxford, 346p.
Fardiaz, S., 1989. Mikrobiologi panganpenuntun praktek laboratorium. Bogor:Jurusan Teknologi Pangan dan Gizi,Fakultas Teknologi Pertanian, InstitutPertanian Bogor, Bogor.
Feliatra., I. Efendi dan E. Suryadi. 2004. Isolasidan identifikasi bakteri probiotik dari ikanKerapu Macan (Ephinephelus fuscogatus)dalam upaya efisiensi pakan ikan. JurnalNatur Indonesia, 6(2): 75-80.
Flores. M. L dan M. A.O. Novoa, 2013. Theuse of lactic acid bacteria isolated fromintestinal tract of Nile tilapia(Oreochromis niloticus), as growthpromoters in fish fed low protein diets.
Latin America Journal Aquatic Resources.41(3): 490-497
Gangasuresh, P dan M. R. Bharathi, 2014.Comparative diversity profiles ofgastrointestinal micro flora or normal andsick “Oreochromis mossambicus (Peters).Asian Journal of Pharmaceutical Science& Technology, 4(2): 91-96
Geovanny , D. G. R dan M. A. Shen, 2008.Influence of probiotics on the growth anddigestive enzyme activity of White PacificShrimp (Litopenaeus vannamei). JournalOcean Univ. Chin. 7: 215-218.
Hadioetomo, R. ,. 1985. Mikrobiologi dasardalam praktek teknik dan prosedur dasarlaboratorium. Gramedia. Jakarta.
Holt, J. G; N. R, Krieg; P. H. A. Sneath; , J. T.Staley dan S. T. Williams, 1994. Bergey’smanual of determinative bacteriology.Ninth Edition. Williams and Wilkins.United Stated of America..
Lay B. W., 1994. Analisis mikroba dilaboratorium. Jakarta: PT. Raja Persada.
Lewis, H. D, 1973. Predominant aerobicbacteriaof fish and shellfish. Departmentof Veterinary Microbiology. Texas A &M University. College Station. Texas.
Moeljanto, 1992. Pengawetan dan pengolahanhasil perikanan, Jakarta , PenebarSwadaya
Musikasang, H., A. Tani, A. H. Kittikun dan S.Maneerat, 2009. Probiotic potential oflactic acid bacteria isolated from chickengastrointestinal digestive tract. WorldJournal Microbiology and Biotechnology,25: 1337–1345.
Subagiyo dan A. Djunaedi. 2011. Skriningkandidat bakteri probiotik dari saluranpencernaan ikan Kerapu berdasarkanaktivitas anti bakteri dan produksi enzimproteolitik ekstraseluler. Jurnal IlmuKelautan, 16(1): 41-48
Syandri, H. 2013. Penggunaan ikan Nilem(Osteochilus haselti CV) dan ikan Tawes(Puntius javanicus CV) sebagai agenhayati pembersih perairan DanauManinjau, Sumatera Barat. Jurnal NaturIndonesia, 6(2): 87-90.
Tangko A. M., A. Mansyur dan Reski, 2007.Penggunaan probiotik pada pakanpembesaran ikan Bandeng dalam kerambajaring apung di laut, Jurnal RisetAkuakultur II (1): 33-40.
Thillaimaharani, K. A., A. R. Logesh, K.Sharmila, B. Kaja Magdoom dan M.Kalaiselvam, 2012. Studies on the

Yempita Efendi dan Yusra 295
intestinal bacterial flora of tilapiaOreochromis mossam bicus (Peters, 1852)and optimization of alkaline protease byVirgibacillus pantothenticus. Journal ofMicrobiology and Antimicrobials, 4(5):79-87.
Trakroo, M. D dan R. Agarwal, 2011.Qualitative and quantitative study onbacterial flora of farm raised Rohu, Labeorohita (Ham.) in India. J.Recent Trends inBiosci.,1(2): 66-71.
Zapata, A. A. 2013. Antimicrobial activities oflactic acid bacteria strains isolated fromNile Tilapia intestine (Oreochromis
niloticus). Journal of Biology and LifeScience 4(1). 164-171.
Ziaei-Nejad S., M. H. Rezaei, G. A. Takami, D.L. Lovett, A.R. Mirvaghefi dan M.Shakouri, 2006. The effect of Bacillus spp.bacteria used as probiotics on digestiveenzyme activity, survival and growth intheIndian white shrimp Fenneropenaeusindicus. Aquaculture, 252: 516-524.
Zhou.X., Y. Wang dan W. Li, 2009 . Effect ofprobiotic on larvae shrimp (Penaeusvannamei) based on water quality,survival rate and digestive enzymeactivities. Aquaculture, 287: 349–353.

ISSN 978-602-14989-0-3 BioETI
Efektivitas beberapa insektisida aerosol dengan metode GlassJar dan semprot terhadap Kecoak Jerman (Blattella germanicaL.) Strain PLZ-SMRDYOSI RAHMAN1), RESTI RAHAYU1 DAN DAHELMI2)
1)Labor Riset Fisiologi Hewan, Jurusan Biologi, FMIPA Universitas Andalas, Limau Manis Padang – 251632)Labor Riset Taksonomi Hewan, Jurusan Biologi, FMIPA FMIPA Universitas Andalas, Limau Manis Padang – 25163E-mail: [email protected]
ABSTRAKEfektivitas beberapa insektisida aerosol terhadap B. germanica strain PLZ-SMRD dapat dilihat dengan menggunakanmetode glass jar dan semprot menggunakan insektisida ByYR, HtYR, MtYR, RdYR dan VpYR. Penelitian dilakukan diLaboratorium Riset Fisiologi Hewan, Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA),Universitas Andalas, Padang pada bulan November sampai Desember 2013. Kriteria efektivitas digolongkanberdasarkan knockdown time 90% (waktu kelumpuhan). Hasil penelitian memperlihatkan bahwa efek kelumpuhantercepat pada strain PLZ-SMRD pada insektisida ByYR dan RdYR menggunakan metode glass jar.
Key words: B. germanica, insektisida aerosol, metode glass jar, metode semprot, knockdown
Pendahuluan
B. germanica atau kecoak jerman tergolongdalam filum Arthropoda, kelas Dictyoptera, danfamili Blattidae. Blattidae merupakan familikecoak yang terdistribusi luas, terdiri dari duaspesies terbesar yaitu B. germanica dan Supellalongipalpa (Gillot, 2005). Kecoak mempunyaibentuk tubuh bulat pipih dorsoventral, kepalaagak memendek dan dilengkapi sepasangantena filiform. Pada dada terdapat 3 pasangkaki, 2 pasang sayap berbentuk membran(Robinson, 2005).
Kecoak jerman dapat ditemukan di seluruhdunia, lebih menyukai tempat yang didiamimanusia (IFAS, 2006). Kecoak mampu hidupdi luar maupun di dalam ruangan. Serangga inidapat bertindak sebagai agen penularanpenyakit dan bisa menyebabkan alergi terhadaporang yang sensitif atas keberadaan seranggaini (Lesmana, 2003).
Kecoak jerman adalah salah satu hama yangmeninggalkan sisa makanan yang keluar darimulutnya serta membuang kotoran di tempatyang dilewatinya sehingga menimbulkan aromabusuk (Rozendaal, 1997). Kecoak ini
merupakan perusak kayu dengan membentukterowongan akibat gigitannya (Bell, 2007).Metode pengendalian kecoak jerman telahbanyak dikenal dan digunakan oleh manusia.Metode yang sering digunakan adalah secarakimiawi menggunakan insektisida. Penggunaaninsektisida kimiawi atau sintetik pada dasarnyasangat efektif untuk membunuh kecoak atauserangga lain (Baskoro et al., 2010).Penggunaan insektisida dapat membunuhkecoak secara cepat, tetapi dosis yangdigunakan oleh masyarakat cenderungberlebihan dari yang dianjurkan dan padaakhirnya dalam waktu lama akanmengakibatkan resistensi (Untung, 2008).
Efektivitas insektisida terhadap kecoakjerman ditentukan oleh cara pengaplikasiannya.Aplikasi semprot dan glass jar adalah metodeyang paling sering digunakan. Makadilakukanlah penelitian dengan dua metodetersebut, metode glass jar mengacu kepadapenelitian Schraf et al., (1995) dan metodesemprot mengacu kepada penelitian Pratama(2011). Berdasarkan penelitian yang telahdilakukan diperoleh knockdown time (KT) yangberbeda antara kedua metode tersebut.

Yosi Rahman, Resti Rahayu dan Dahelmi 297
BAHAN DAN METODE
Tanah Penelitian ini dilakukan dengan metodeeksperimental dengan pengulangan sebanyaktiga kali. Subjek penelitian yang digunakanadalah kecoak jerman strain PLZ-SMRD.Insektisida aerosol yang digunakan yaitu ByYR,HtYR, MtYR, RdYR, dan VpYR.Metode Pengujian EfektivitasMetode Glass Jar
Beberapa insektisida aerosol disemprotkanke gelas ukur untuk mendapatkan 1 mlinsektisida. Insektisida tersebut dimasukkan kedalam petridish dan digoyang-goyangkansampai merata, kemudian dikeringkan anginkanselama ±1-2 jam. Petridish yang telah kering,pada pinggirnya dioleskan campuran vaselinedan baby oil. Sepuluh ekor kecoak dimasukkanke dalam petridish, selanjutnya diamati setiapmenit sampai 10 menit, setiap 10 menit sampai50 menit dan setiap satu jam sampai 96 jam.Setiap pengamatan dilakukan pencatatanjumlah knockdown (kelumpuhan) kecoak.Metode Semprot
Sepuluh ekor kecoak masing-masing strainuji dimasukkan ke dalam petridish yang telahdiolesi campuran vaseline dan baby oil.Petridish dimasukkan ke dalam kotakberukuran 30 cm x 30 cm x 60 cm, kemudiandari mulut kotak disemprotkan insektisida kearah petridish selama satu detik. Kotakdiangkat dan diamati kondisi (kelumpuhan)kecoak dalam petridish setiap menit sampai 10menit, setiap 10 menit sampai 50 menit dansetiap satu jam sampai 96 jam. Setiappengamatan dilakukan pencatatan jumlahkejatuhan kecoak.
Kriteria Efektivitas InsektisidaMetode Pengujian Efikasi Hygene
Lingkungan oleh Direktorat Pupuk danPestisida (2004) merupakan dasar penentuankriteria efektivitas insektisida yang digunakandalam penelitian ini. Kriteria efektivitasinsektisida tersebut berdasarkan waktukelumpuhan (knockdown time 90%/KT90)dalam periode tertentu. KT90 merupakan waktuyang diperlukan untuk melumpuhkan 90%
hewan uji. Suatu insektisida dikatakan efektifuntuk kecoak jerman apabila KT90 mampudicapai paling lama 20 menit setelahpemaparan.
HASIL DAN PEMBAHASAN
Pengujian tentang uji efektivitas beberapainsektisida aerosol terhadap strain PLZ-SMRDdengan metode glass jar dan semprotmenunjukkan perbedaan rata-rata kelumpuhanseperti strain KRSA-BDG. Rata-ratakelumpuhan kecoak strain PLZ-SMRDterhadap insektisida ByYR dengan metodeglass jar pada menit awal sebanyak 73,3%(Gambar 2.a). Pada menit ke-2 terjadipertambahan kelumpuhan kecoak sebanyak80,0% dan selanjutnya sebanyak 90,0% padamenit ke-10. Pada menit ke-20 semua kecoakmengalami kelumpuhan (100%).
Insektisida ByYR yang dipaparkan denganmetode semprot menyebabkan kelumpuhanpada kecoak sebanyak 90,0% pada menit ke-1sampai menit ke-9. Pada menit ke-10 terjadikelumpuhan kecoak sebesar 96,7%. Semuakecoak mengalami kelumpuhan 100% pada jamke-24 (Gambar 2.a).
Pada Gambar 2.b terlihat rata-ratakelumpuhan kecoak pada menit ke-1 sebanyak26,7% terhadap insektisida HtYRmenggunakan metode glass jar. Pada menit ke-2 kelumpuhan kecoak bertambah menjadi36,7% dan berlanjut hingga menit ke-50sebanyak 96,7%. Setelah satu jam, kecoakmengalami kelumpuhan 100%.
Kecoak yang terkena insektisida HtYRdengan menggunakan metode semprotmenyebabkan rata-rata kelumpuhan padahewan uji sebanyak 53,3% pada menit ke-1.Selanjutnya, pada menit ke-3 rata-ratakelumpuhan kecoak menjadi 63,3% danseterusnya hingga mencapai 96,7% setelah 1jam. Pada jam ke-2 terjadi kelumpuhan 100%kecoak.Gambar 2.c menujukkan rata-rata kelumpuhankecoak jerman terhadap insektisida MtYR

Yosi Rahman, Resti Rahayu dan Dahelmi 298
dengan metode glass jar sebanyak 50,0% padamenit awal dan pada menit selanjutnyasebanyak 56,7%. Pada menit ke-3 sampai menitke-90 kecoak mengalami kelumpuhan hingga90,0%. Semua kecoak mengalami kelumpuhansetelah 12 jam.
Pemaparan kecoak dengan metode semprotmengakibatkan kelumpuhan kecoak terjadipada menit pertama sebanyak 76,7% dan menitke-2 hingga menit ke-5 masih sebanyak 76,7%.Pada menit ke-6 rata-rata kelumpuhanmeningkat menjadi 83,3%. Selanjutnya menitke-20 terjadi kelumpuhan kecoak sebanyak96,7%. Kecoak mengalami kelumpuhan 100%pada menit ke-30.
Rata-rata kelumpuhan kecoak jerman padaGambar 2.d dengan metode glass jar terhadapinsektisida RdYR mencapai 63,3% pada menitke-1, 76,7% pada menit ke-2 dan 80,0% padamenit ke-3. Pada menit ke-6 rata-ratakelumpuhan kecoak bertambah sebanyak80,0% dan akhirnya pada menit ke-20, semuakecoak mengalami kelumpuhan.Kecoak yang terkena insektisida uji di atasdengan metode semprot menyebabkankelumpuhan sebanyak 76,7% pada menit ke-1dan menit ke-2 sebanyak 80,0%. Kelumpuhankecoak meningkat rata-ratanya mencapai96,7% pada menit ke-10. Kelumpuhan kecoak100% terjadi pada jam ke-3.
Pada Gambar 2.e terlihat bahwa rata-ratakelumpuhan kecoak jerman terhadapinsektisida VpYR menggunakan metode glass
jar pada menit ke-1 sebanyak 90,0%.Kelumpuhan kecoak meningkat pada menit ke-4 sebanyak 96,7% dan seterusnya hingga padamenit ke-30, kecoak mengalami kelumpuhan100%.
Berbeda halnya dengan metode semprot,pada metode ini kelumpuhan kecoak padamenit pertama sampai menit ke-3 sebanyak73,3% dan menit ke-4 mengalami pertambahanrata-rata kelumpuhan sebanyak 96,7%. Padajam ke-1, kecoak mengalami kelumpuhan100%.
Berdasarkan beberapa pernyataan di atasdapat diketahui bahwa efek kelumpuhantercepat terhadap kecoak jerman strain PLZ-SMRD terdapat pada insektisida ByYR danRdYR menggunakan metode glass jar. Hal inidisebabkan oleh kandungan bahan aktif dalamByYR adalah imiprotrin. Imiprotrin merupakanpiretroid sintetik dengan bentuk cair danberwarna kuning. Efek yang ditimbulkanapabila keracunan imiprothrin antara lainhipersensitivitas, fibrilasi otot, tremor, ataksia,pernapasan tidak teratur/cepat, excess salivasi,urinasi, nonkarsinogenik dan nonmutagenik.Toksisitas akut oral LD50 pada tikus betina 50sebesar 2400mg/kg dan jantan 4500mg/kg(Anonimous, 2008b)
Bahan aktif yang lainnya yaitu esbiothrinyang merupakan senyawa piretroid sintetisdengan efek kejatuhan (knockdown) cepatterhadap hama rumah tangga. Zat ini biasanyadigunakan masyarakat untuk pemberantasannyamuk, lalat dan kecoak (WHO, 2002).Esbiothrin memiliki mekanisme kerjanonsistemik dengan aplikasi melalui racunkontak, racun perut dan pernapasan sertasebagai modulator sodium channel 378-432mg/kg pada tikus. (Anonimous 2008a).
RdYR mengandung senyawa transflutrindan siflutrin. Siflutrin tergolong dalam piretroid,zat ini bersifat toksik terhadap kedua strain.Piretroid bersifat racun terhadap serabut saraf(Sigit et al., 2006) dan menunjukkan efekkejatuhan yang cepat pada serangga sehinggamenimbulkan kelumpuhan dan berakhir dengankematian (Dadang, 2007). Siflutrin bersifatsangat akut toksik pada lebah dan ikan. Padatingkat ekosistem menunjukkan efek toksikterhadap beberapa organisme, termasuk alga,zooplankton, nematoda, serangga dan ikan.Beberapa peneliti dari Australia melakukanpengamatan residu siflutrin pada gandum yangdisimpan selama bertahun-tahun denganberbagai kondisi suhu dan kelembaban. Merekamenemukan bahwa siflutrin mampu bertahan
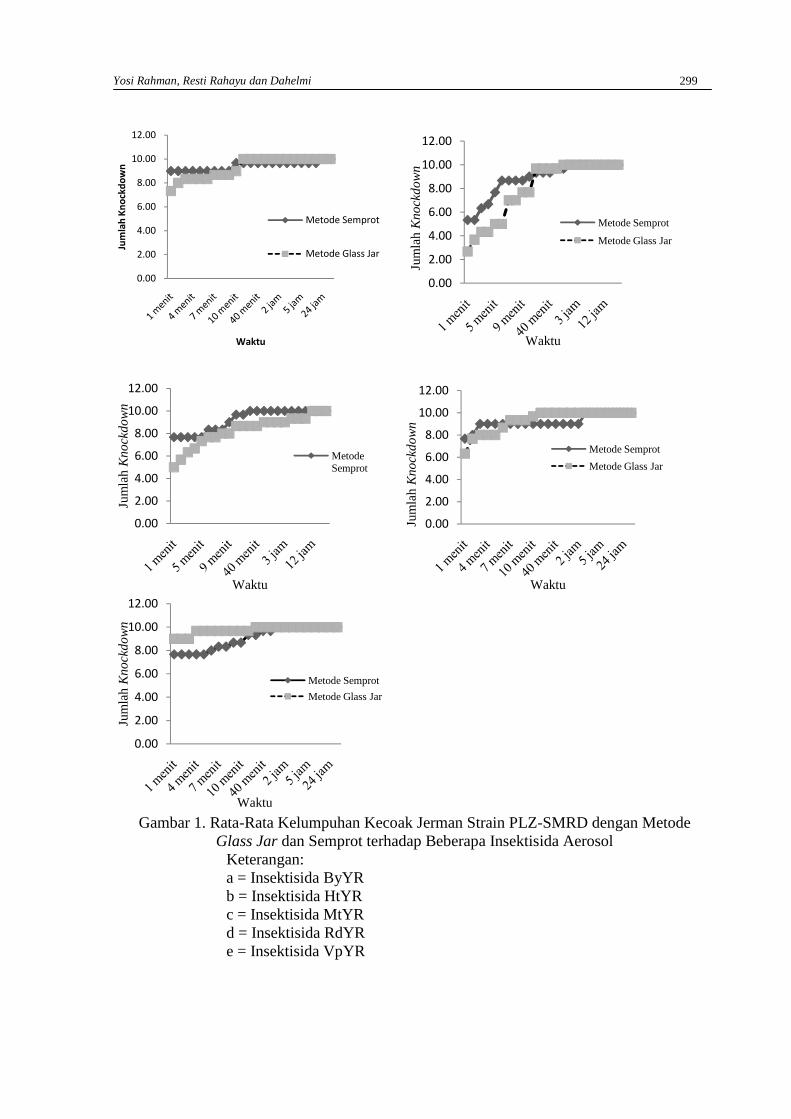
Yosi Rahman, Resti Rahayu dan Dahelmi 299
Gambar 1. Rata-Rata Kelumpuhan Kecoak Jerman Strain PLZ-SMRD dengan MetodeGlass Jar dan Semprot terhadap Beberapa Insektisida Aerosol
Keterangan:a = Insektisida ByYRb = Insektisida HtYRc = Insektisida MtYRd = Insektisida RdYRe = Insektisida VpYR
0.00
2.00
4.00
6.00
8.00
10.00
12.00Ju
mla
h Kn
ockd
own
Waktu
Metode Semprot
Metode Glass Jar
0.00
2.00
4.00
6.00
8.00
10.00
12.00
Jum
lah
Kno
ckdo
wn
Waktu
Metode Semprot
Metode Glass Jar
0.00
2.00
4.00
6.00
8.00
10.00
12.00
Jum
lah
Kno
ckdo
wn
Waktu
MetodeSemprot
0.00
2.00
4.00
6.00
8.00
10.00
12.00Ju
mla
hK
nock
dow
n
Waktu
Metode Semprot
Metode Glass Jar
0.00
2.00
4.00
6.00
8.00
10.00
12.00
Jum
lah
Kno
ckdo
wn
Waktu
Metode Semprot
Metode Glass Jar

Yosi Rahman, Resti Rahayu dan Dahelmi 300
selama masa penelitian (52 minggu).Diperkirakan, siflutrin memiliki ketahananpada gandum tersebut selama 23-114 mingguyang tergantung pada suhu, kelembaban, danisomernya (Cox, 1994).
Selain kandungan bahan aktif, efektivitasbeberapa insektisida aerosol dipengaruhi olehperbedaan metode (aplikasi) pemaparannya,faktor lainnya yaitu variasi ukuran nozel yangberfungsi mengeluarkan massa insektisida padasaat penyemprotan. Djojosumarto (2008)menyatakan penyemprotan dilakukan denganmenggunakan alat semprot (sprayer). Sprayermemiliki nozel (cerat, spuyer) yang berfungsimemecah cairan pestisida menjadi butiran-butiran cairan sangat halus, yang disebutdengan spray droplet. Butiran semprot inididistribusikan ke bidang sasaran. Johnson(1914) menyatakan variasi massa yangdikeluarkan masing-masing insektisidadisebabkan oleh perbedaan ukuran nozel.Pemilihan tipe dan ukuran nozel yang tepatsangat penting untuk menentukan keefektifanaplikasi pestisida. Nozel adalah faktor yangpaling utama dalam menentukan jumlah massapestisida yang keluar pada suatu area,sedangkan metode pengaplikasian, luaspermukaan area target dan Potential driftmerupakan faktor lainnya.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Efek kelumpuhan tercepat terlihat pada strain
PLZ-SMRD menggunakan ByYR, RdYRdengan metode glass jar.
DAFTAR PUSTAKA
Anonimous. 2008a. Mekanisme KerjaEsbiothrin. http://sitem.herts.ac.uk/aeru/footprint/en/Reports/80.htm. Diakses pada9 Januari 2014.
_______. 2008b. Pyrethroid.http://www.sumitomo-chem.com.au/msds/esbiothrin+d-
phenothrin + imiprothrin.pdf. Diaksespada 9 Januari 2014.
Baskoro, A. D. Umi Kalsum, MuhammadSuhail Bin Satri. 2010. Uji PotensiEkstrak Etanol Daun Menta (Menthaarvensis L.var.Javanica(BI.) Hook.)sebagai Insektisida terhadap kecoaPeriplaneta americana Dewasa. [Skripsi].Fakultas Kedokteran. UniversitasBrawijaya.
Bell, W. J., L. M. Roth and C. A. Nalepa. 2007.Cockroaches; Ecology, Behavior andNatural History. The Johns HopkinsUniversity Press. Baltimore.
Cox, C. 1994. Insecticide Fact Sheet.Cyfluthrin. Eugene. Oregon. Vol. 14, No.2. Danish Pest Infestation Laboratory,Skovbrynet 14, DK-2800 Lyngby,Denmark.
Dadang. 2007. Bahan Kuliah Pestisida danTeknik Aplikasi (Insektisida). IPB. Bogor.
Direktorat Pupuk dan Pestisida. 2004. MetodePengujian Efikasi Hygene LingkunganDepartemen Pertanian Republik Indonesia.Jakarta.
Djojosumarto, P. 2008. Pestisida &Aplikasinya. Agromedia. Jakarta.
Gillott, C. 2005. Entomology, Third Edition.Springer. Netherlands.
IFAS (Institute of Food and AgriculturalSciences). 2006. German Cockroach,Blattella germanica (Linnaeus) (Insecta:Blattodea: Blattellidae. Entomology andNematology Department, FloridaCooperative Extension Service. FloridaUniversity. Florida.http://entomology .ifas.ufl.edu/creatures.
Johnson, M.P and Larry D. Swetnam. 1914.Sprayer Nozzles Selection and Calibration.University of Kentucky. College ofAgriculture. U.S.
Lesmana, D. 2003. Aktivitas repelensi ekstraksepuluh spesies tanaman terhadapBlattella germanica L. [Skripsi]. FakultasPertanian. Institut Pertanian Bogor.
Pratama, F. 2011. Perbandingan efikasi limainsektisida aerosol komersial terhadap
kecoa jerman, Blattella germanica,(Dictyoptera: Blattellidae) strain VCRU,bandung, dan surabaya. Skripsi SarjanaSekolah Ilmu dan Teknologi Hayati, ITB.
Robinson, W.H. 2005. Handbook of UrbanInsect and Arachnids. CambridgeUniversity press. New York.

Yosi Rahman, Resti Rahayu dan Dahelmi 301
Rozendaal, J. A. 1997. Handbook of urbanInsect and Arachnids. CambridgeUniversity Press. New York.
Scharf, M.E., Bennet G.W., Reid B.L., Qui C.1995. Comparison of Three InsecticideResistance Detection Methods for theGerman Cockroach (Dictyoptera:Blattellidae). J Econ Entomol. 88: 536-542.
Sigit, S.H., Koesharto F.X., Hadi U.K.,Gunandini D.J., Soviana S., Wirawan I.A.,Chalidaputra M., Rivai M., Priyambodo,Yusuf S. dan Utomo S. 2006. HamaPermukiman Indonesia (Pengenalan,
Biologi, dan Pengendalian. Unit KajianPengendalian Hama Permukiman FKHIPB. Bogor.
WHO. 2002. D-Allethrin. WHO Specificationsand Evaluations for Public HealthPesticides. Geneva. 1-24.
Untung, K. 2008. Manajemen ResistensiPestisida Sebagai PenerapanPengelolaan Hama Terpadu.http://kasumbogo.staff.ugm.ac.id/?satoewarna=index&winoto =base&ac.Diakses pada tanggal 17 Mei 2013

ISSN 978-602-14989-0-3 BioETI
Karakterisasi parsial senyawa antimikroba dari bakteri Bacilluscereus strain HVR22YUSRA DAN YEMPITA EFENDI
Fakultas Perikanan dan Ilmu Kelautan, Universitas Bung Hatta, PadangE-mail:
ABSTRAKPenelitian bertujuan untuk melakukan karakterisasi parsial senyawa antimikroba dari bakteri Bacillus cereus strainHVR22. Metode penelitian yang digunakan adalah metode eksperiment. Materi yang digunakan adalah bakteri Bacilluscereus strain HVR22. Karakterisasi meliputi aktivitas antimikroba pada suhu dingin (40C, -100C, -150C dan -200C),beberapa konsentrasi NaCl (2,5%, 5%, 7,5%, 10%, 12,5%, 15%, 17,5% dan 20%) dan lama penyimpanan (14 hari).Berdasarkan uji karakterisasi bakteri Bacillus cereus strain HVR22 masih memiliki aktivitas antimikroba pada suhurendah (40C sampai suhu -200C), masih dapat tumbuh pada konsentrasi garam sampai dengan 20% dan masih memilikiaktivitas setelah disimpan selama 14 hari terhadap lima bakteri patogen.
Key words: karakterisasi, senyawa antimikroba, Bacillus cereus strain HVR22
Pendahuluan
Masalah keamanan pangan (food safety)termasuk hasilperikanan masih merupakankendala utama. Mutu dan keamanan hasilperikanan perlu diperhatikan karena dapatmembahayakan kesehatan bagi konsumen.Mikroorganisme patogen yang sering terdapatdi dalam produk perikanan diantaranyaEscherichia coli, Staphylococcus aureusSalmonella thypi, Bacillus subtilis dan Listeria
monocytogenes. Bakteri patogen tersebutberesiko menimbulkan penyakit bahkankematian. Alternatif dalam mengatasimasalah tersebut adalah dengan pengolahandan pengawetan ikan.
Metode pengawetan yang telah banyakdiaplikasikan adalah penambahan bahanpengawet pada produk perikanan, baik bahanpengawet sintetis maupun alami. Penggunaanpengawet sintetis dapat menyebabkankemungkinan toksin akibat residu yang masihaktif, bahaya mikroorganisme yang resisten dandapat menimbulkan infeksi pada konsumen.Penggunaan bahan pengawet alami lebihberpotensi untuk diaplikasikan sebagaipengganti pengawet sintetis.
Bakteriosin merupakan salah satusubstansi antimikroba yang dihasilkan bakteriasam laktat dan memiliki aktivitas antagonistik,baik bakteriostatik maupun bakterisidal.Bakteriosin berpotensi digunakan sebagaibahan pengawet pangan alami yang amanuntuk dikonsumsi, karena zat aktif yangterdapat dalam bakteriosin adalah proteinyang dapat didegradasi oleh enzim proteolitik.Bakteriosin adalah peptida-peptida yangdiproduksi oleh sejumlah bakteri Gram positifdan Gram negative. Bakteriosin dapatdidefinisikan sebagai protein aktif ataukompleks protein yang menunjukkan aksibakterisidal melawan bakteri Gram positif,terutama spesies yang berkerabat dekat denganspesies penghasil (Jack et al., 1995; Ray;Parada et al., 2007). Penggunaan bakteriosinsebagai biopreservatif, perlu memperhatikandan menentukan jumlah konsentrasi bakteriosinyang harus ditambahkan dalam produk pangan,dan efisiensi bakteriosin dalam mengontrolbakteri-bakteri patogen (Ananou et al., 2005).
Bakteriosin selain dihasilkan oleh genusLactobacillaceae juga dapat dihasilkan olehbakteri gram positif lain, misalnya oleh bakteridari genus Bacillus sp. Hal ini didasarkan padapenelitian yang dilakukan oleh Sharma et al.,

Yusra dan Yempita Efendi 303
(2008); Hena dan Mamatha (2014) yangmeneliti tentang karakterisasi dan pemurnianbakteriosin dari Bacillus. sp MTCC 43 danBacillus subtilis BSF01 yang diaplikasikansebagai pengawet susu. Selanjutnya penelitianyang dilakukan oleh Bizani et al., (2005)tentang aktivitas antibakteri dari cerein 8A,bakteriosin yang dihasilkan oleh bakteriBacillus cereus. Berdasarkan penelitian Yusraet al., (2013) bakteri Bacillus cereus strainHVR22 merupakan bakteri potensial yangdiisolasi dari budu, produk fermentasi ikantradisional Sumatera Barat diketahuimenghasilkan suatu senyawa antimikroba yangdapat menghambat pertumbuhan lima bakteripatogen. Sebelum digunakan sebagai bahanpengawet harus dilakukan karakterisasi agarsenyawa antimikroba dari bakteri Bacilluscereus strain HVR22 dapat berfungsi maksimal.Tujuan penelitian ini adalah melakukankarakterisasi parsial bakteri terseleksi Bacilluscereus strain HVR22 yang berasal dari buduyang nantinya diharapkan dapat dijadikansebagai bahan pengawet alternatif yang amankhususnya untuk produk perikanan.
BAHAN DAN METODE
Bahan yang digunakan dalam penelitian iniadalah bakteri Bacillus Bacillus cereus strainHVR22 yang berasal dari ikan budu Tenggiri(Scomberomorus guttatus). Zat-zat yangdigunakan Rogosa Sharpe (MRS) Broth(Merck), Rogosa Sharpe (MRS) agar (Merck),nutrien agar (NA), larutan Mc Farland 0,5,NaOH, HCl, NaCl, alkohol, spiritus. Bakteriuji yang digunakan dalam penelititan ini adalahEscherichia coli ATCC 25922, Staphylococcus
aureus ATCC 25923, Salmonella thypi NBRC14237, Bacillus subtilis FNCC 056 danListeria monocytogenes FNCC 0734
1. Karakterisasi Isolat Bakteri Potensial danSenyawa Antimikroba yang Dihasilkannya1.1 Pengaruh Perlakuan Suhu Dinginterhadap Aktivitas Antimikroba
Untuk mengukur kemampuan antimikrobadari isolat bakteri terpilih dilakukan dengan
cara supernatan bebas sel dioptimasi padabeberapa variasi suhu dingin (-200C, -150C, -100C, 40C selama interval waktu 15, 30, 45 dan60 menit). Selanjutnya dilakukan ujiantimikroba menggunakan 5 bakteri patogendengan menggunakan metode cakram(Nofisulastri et al., 2006).1.2Optimasi Pertumbuhan Bakteri Potensial
Terpilih pada Beberapa KonsentrasiGaram
Sebanyak 1 ose kultur bakteri ditumbuhkanpada 10 ml MRS Broth, selanjutnyaditambahkan NaCl dengan konsentrasi yangberbeda (2,5%, 5%, 7,5%, 10%, 12,5%, 15%,17,5% dan 20%). Selanjutnya dilakukaninkubasi selama 24 jam pada suhu kamar(270C) dan diamati kekeruhannyamenggunakan alat spektrofotometer denganpanjang gelombang 600 nm (Axelsson, 2004.Modifikasi).
2. 3 Pengaruh Lama Penyimpanan terhadapAktivitas AntimikrobaUntuk mengukur kemampuan antimikroba dariisolat bakteri terpilih dilakukan dengan carasupernatan bebas sel dioptimasi pada suhu -100C, 40C dan 370C selama 14 hari.Selanjutnya dilakukan uji antimikrobamenggunakan 5 bakteri patogen denganmenggunakan metode cakram (Nofisulastri etal., 2006).
HASIL DAN PEMBAHASAN
1. Uji Kestabilan Supernatan Isolat BakteriBacillus cereus strain HVR22 pada SuhuRendah
Berdasarkan skreening uji antimikrobialterhadap lima bakteri patogen terpilih satuisolat Bacillus cereus strain HVR22 yangpaling potensial menghambat pertumbuhanlima bakteri patogen. Tujuan uji karakterisasisupernatan bakteri Bacillus cereus strainHVR22 pada suhu rendah adalah untuk melihatpotensi antimikroba dari isolat bakteri Bacillus
cereus strain HVR22 pada berbagai perlakuansuhu rendah. Aktifitas antimikrobial supernatanisolat bakteri Bacillus cereus strain HVR22

Yusra dan Yempita Efendi 304
pada berbagai suhu rendah dalam menghambatpertumbuhan lima bakteri patogen dapatdilihat pada Gambar 1.
Berdasarkan Gambar 1. dapat dilihat bahwaperlakuan suhu rendah masih berpengaruhterhadap aktivitas antimikroba isolat bakteriBacillus cereus strain HVR22 dalammenghambat bakteri patogen. Pada suhu 40Caktivitas antimikrobial yang paling tinggiadalah terhadap bakteri Salmonella thypidengan diameter zona beningnya 14 mm,diikuti oleh Bacillus subtilis 11 mm,Staphylococcus aureus, Escherichia coli danListeria monocytogenes. Pada suhu -100Caktivitas antimikrobial yang paling tinggiadalah terhadap bakteri Staphylococcus aureusdengan aktivitas zona hambatnya 11 mm,diikuti oleh Listeria monocytogenes, Bacillussubtilis, Salmonella thypi dan Escherichia coli.Pada suhu -150C, senyawa antimikroba yangdihasilkan oleh isolat bakteri Bacillus cereusstrain HVR22 memiliki aktivitas antimikrobialyang paling tinggi terhadap bakteri Bacillus
subtilis dengan diameter zona beningnya 13mm, diikuti oleh Listeria monocytogenes,Staphylococcus aureus, Escherichia coli danSalmonella thypi. Pada suhu -200C, aktivitasantimikrobial yang paling tinggi adalahterhadap bakteri Salmonella thypi dengandiameter zona beningnya 13 mm, diikuti olehbakteri Staphylococcus aureus, Listeria
monocytogenes, Escherichia coli dan Bacillussubtilis.
Pengaruh suhu terhadap pertumbuhanmikroba disebabkan karena suhumempengaruhi aktivitas enzim yangmengkatalisis sel-sel biokimia di dalam selmikroba. Di bawah suhu optimum, aktifitasenzim di dalam sel mikroba menurun,akibatnya pertumbuhan sel juga terhambat(Dwidjoseputo, 1978). Penggunaan suhurendah dalam pengawetan bahan pangandisebabkan karena aktivitas mikroorganismefood-borne melambat dan/atau berhenti padasuhu di atas pembekuan dan umumnya berhentipada suhu subfreezing karena seluruh reaksi
metabolik mikroorganisme dikatalisis olehenzim dan reaksi katalisis enzim tergantungpada suhu. Adanya kenaikan suhu akanmenyebabkan peningkatan laju reaksi (Jay,2005). Hasil penelitian ini sejalan denganpenelitian Gray et al., (2006) yang melakukanisolasi dan klasifikasi thuricin 17 yangdihasilkan oleh bakteri Bacillus thuringiensisNEB17, menemukan bahwa senyawaantimikrobanya memiliki stabilitas suhu yangluas mulai dari suhu -200C sampai 1000C danstabil pada suhu -200C dengan lamapenyimpanan 30 hari. Aktivitas senyawaantimikroba (Bacteriocin Like InhibitorySpectrum, BLIS) dari Bacillus cereus ATCC14579 100% aktif setelah diinkubasi pada suhu1000C selama 10 menit dan stabil pada suhu -200C dengan lama penyimpanan 30 hari(Risoen et al., 2004).2. Optimasi Pertumbuhan Bakteri Bacilluscereus strain HVR22 pada BeberapaKonsentrasi NaClTujuan dari uji optimasi isolat bakteri Bacillus
cereus strain HVR22 yang dikultur pada mediaMRSB, kemudian ditambahkan dengan NaCldengan konsentrasi 2,5%, 5%, 7,5%, 10%,12,5%, 15%, 17,5% dan 20% dan diinkubasiselama 24 jam, adalah untuk mengetahuikemampuan hidup bakteri ini pada beberapakonsentrasi NaCl sehingga diharapkan dapatdiaplikasikan sebagai bahan pengawet panganalami. Hal ini mengingat setiap bakterimemiliki toleransi tertentu terhadap konsentrasiNaCl. Optimasi pertumbuhan bakteri Bacilluscereus strain HVR22 pada berbagai konsentrasiNaCl dapat dilihat pada Gambar 2.
Berdasarkan Gambar 2 terlihat bahwa isolatbakteri Bacillus cereus strain HVR22 dapattumbuh pada medium MRSB yangditambahkan dengan NaCl dengan konsentrasiyang berbeda mulai 2.5% sampai dengan 20%.Hal ini dibuktikan dengan adanya kultur yangberwarna keruh pada pengamatan selama 24-72jam. Kultur bakteri yang semulanya beningberubah warna menjadi keruh sebagai tandaterjadinya pertumbuhan bakteri setelah masa
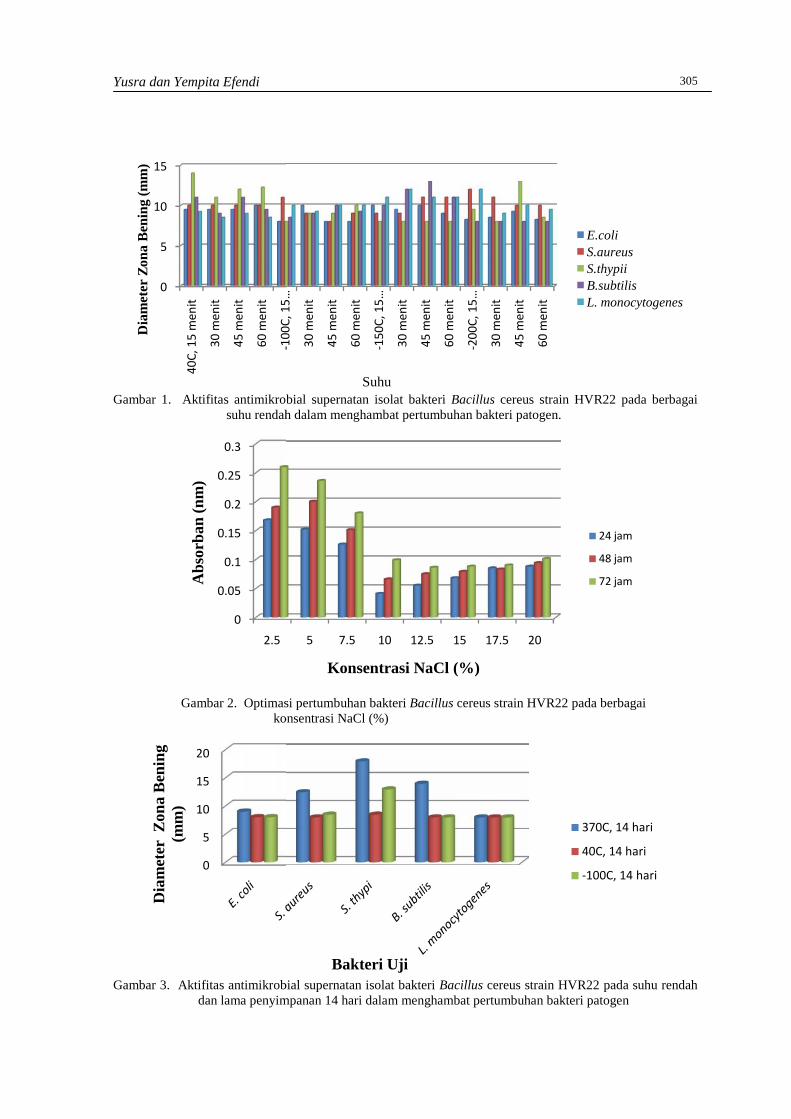
Yusra dan Yempita Efendi 305
Gambar 1. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada berbagaisuhu rendah dalam menghambat pertumbuhan bakteri patogen.
Gambar 2. Optimasi pertumbuhan bakteri Bacillus cereus strain HVR22 pada berbagaikonsentrasi NaCl (%)
Gambar 3. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada suhu rendahdan lama penyimpanan 14 hari dalam menghambat pertumbuhan bakteri patogen
0
5
10
15
40C,
15
men
it
30 m
enit
45 m
enit
60 m
enit
-100
C, 1
5…
Dia
met
er Z
ona
Ben
ing
(mm
)
0
0.05
0.1
0.15
0.2
0.25
0.3
2.5
Abs
orba
n (n
m)
0
5
10
15
20
Dia
met
er Z
ona
Ben
ing
(mm
)
Yusra dan Yempita Efendi 305
Gambar 1. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada berbagaisuhu rendah dalam menghambat pertumbuhan bakteri patogen.
Gambar 2. Optimasi pertumbuhan bakteri Bacillus cereus strain HVR22 pada berbagaikonsentrasi NaCl (%)
Gambar 3. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada suhu rendahdan lama penyimpanan 14 hari dalam menghambat pertumbuhan bakteri patogen
-100
C, 1
5…
30 m
enit
45 m
enit
60 m
enit
-150
C, 1
5…
30 m
enit
45 m
enit
60 m
enit
-200
C, 1
5…
30 m
enit
45 m
enit
60 m
enit
Suhu
2.5 5 7.5 10 12.5 15 17.5 20
Konsentrasi NaCl (%)
Bakteri Uji
Yusra dan Yempita Efendi 305
Gambar 1. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada berbagaisuhu rendah dalam menghambat pertumbuhan bakteri patogen.
Gambar 2. Optimasi pertumbuhan bakteri Bacillus cereus strain HVR22 pada berbagaikonsentrasi NaCl (%)
Gambar 3. Aktifitas antimikrobial supernatan isolat bakteri Bacillus cereus strain HVR22 pada suhu rendahdan lama penyimpanan 14 hari dalam menghambat pertumbuhan bakteri patogen
E.coliS.aureusS.thypiiB.subtilisL. monocytogenes
24 jam
48 jam
72 jam
370C, 14 hari
40C, 14 hari
-100C, 14 hari

Yusra dan Yempita Efendi 306
inkubasi yang disebabkan oleh prosesmetabolisme yang dilakukan oleh bakteritersebut. Sesuai dengan pendapat Lay (1994), didalam medium cair bakteri akan tumbuh dalamwaktu 24-48 jam. Pertumbuhan bakteri dalamsuatu media cair dapat terlihat dalam berbagaibentuk, yaitu: 1. Kekeruhan yang biasanyaterlihat pada seluruh bagian medium, 2.Pertumbuhan pada permukaan yang dapatberbentuk pelikel (pertumbuhan di atas bagianmedia cair), cincin (pertumbuhan berbentukcincin pada permukaan media cair), flokulenatau membran. 3. Sedimen atau endapan, yaitukumpulan sel-sel yang mengumpul pada dasartabung dan akan menyebar lagi jika tabungdigerakkan atau dikocok. Medium pada bagianatas tabung mungkin akan tetap bening jikainkubasi dilakukan lebih lama.
Terlihat bahwa secara umum terjadipeningkatan tingkat kekeruhan setelah inkubasi24 jam sampai dengan 72 jam. Tingkatkekeruhan yang tertinggi terjadi pada kulturMRSB yang ditambah dengan 2,5% NaCl yaknidengan nilai OD (0,26), diikuti denganpenambahan NaCl 5% dengan nilai OD (0,236)dan yang paling rendah pada penambahan NaCl10% dengan lama inkubasi 24 jam. Hal inimungkin disebabkan karena kondisi optimumuntuk pertumbuhan isolat bakteri Bacilluscereus strain HVR22 terdapat pada konsentrasigaram 2,5% dibandingkan dengan yang lain,mengingat ikan yang digunakan sebagai bahandasar pada pembuatan ikan budu ini merupakanikan air laut yang memiliki toleransi terhadapsalinitas atau kadar garam tertentu.
Garam merupakan bahan yang sangatpenting dalam pengawetan ikan, daging, danbahan pangan lainnya (Buckle et al., 1987).Garam berperan sebagai penghambat selektifpada mikroorganisme pencemar tertentu.Namun, masih tetap ada jenis mikroorganismeyang dapat tumbuh pada bahan pangan yangmengandung garam, baik garam dengankonsentrasi rendah, maupun garam dengankonsentrasi tinggi. Jenis ini disebut dengan
bakteri halofilik. Bakteri halofilik mem-butuhkan konsentrasi NaCl minimal tertentuuntuk pertumbuhannya (Fardiaz, 1989).
Kadar garam merupakan salah satu faktorpenting yang dapat mempengaruhi pertumbuh-an mikroorganisme. Bakteri asal laut, moderathalophilik dan halophilik sejati selalumembutuhkan NaCl yang tinggi untukpertumbuhannya (Stainer et al., 1976). Bakterihalophilik ekstrim membutuhkan NaCl yangtinggi untuk menjaga stabilitas dan aktivitaskatalitik enzim. Konsentrasi garam natriumyang sangat tinggi akan merusak fungsimembran karena dapat menyebabkan terjadinyaosmosis. Pada konsentrasi yang lebih tinggi,sebagian besar bakteri tidak mampu tumbuh lagi.Hal ini disebabkan karena garam mempengaruhipertumbuhan mikroba. Kandungan garam yangtinggi dapat menyebabkan tekanan osmotikyang tinggi sehingga mengakibatkan lisis darisel mikroba. Selain itu kandungan garam yangtinggi dapat menyerap air dari sel bakterisehingga menyebabkan sel menjadi kering.NaCl juga dapat terionisasi menjadi Cl- yangberbahaya bagi mikroba, garam dapatmengurangi kelarutan oksigen dalam air, yangmenyebabkan sel menjadi lebih sensitif terhadapCO2 dan mengganggu kerja enzim proteolitikdalam sel mikroba (Fardiaz, 1989).
Karakterisasi bakteri dari produk fermentasiikan berdasarkan konsentrasi NaCl jugadilakukan terhadap isolasi bakteri yang terdapatpada kecap ikan lemuru. Diketahui empatkelompok bakteri yang mampu tumbuh padakonsentrasi garam 18%, yaitu bakteriPediococcus halophilus dengan empat strainyang berbeda, sedangkan dua kelompok lagiyakni dari kelompok Aerococcus viridans tidakmampu tumbuh pada konsentrasi NaCl 18%(Sudiarta, 2011). Penelitian yang dilakukanterhadap produk fermentasi ikan jeot-galdiketahui bahwa isolat bakteri terpilih mamputumbuh pada konsentrasi NaCl 10-25% (Um danLee, 1996). Begitu juga pada penelitian produkfermentasi ikan teri (anchovy-jeot) yang berasal

Yusra dan Yempita Efendi 307
dari Korea diketahui bahwa bakteri hasil isolasimampu tumbuh pada konsentrasi garam 25%.Bakteri Bacillus sp. KYJ 968 mampu tumbuhhingga konsentrasi NaCl 15% (Ha et al., 2002).
3. Pengaruh Lama Penyimpanan dan Suhuterhadap Aktivitas Antimikroba bakteriBacillus cereus strain HVR22
Aktivitas penghambatan supernatan bakteriBacillus cereus strain HVR22 selama 14 haripada suhu -100C, 40C dan 370C diamati untukmengetahui stabilitas senyawa antimikrobaselama penyimpanan pada suhu dingin.Stabilitas aktivitas penghambatan ditentukanmelalui diameter zona hambat (berupa zonabening atau zona semu) yang dihasilkanterhadap kelima bakteri patogen indikator(Escherichia coli ATCC 25922, Staphylococcusaureus ATCC 25923, Salmonella thypi NBRC14237, Bacillus subtilis FNCC 056 dan Listeria
monocytogenes FNCC 0734). Stabilitas aktivitaspenghambatan supernatan bakteri Bacilluscereus strain HVR22 selama 14 hari pada suhu -100C, 40C dan 370C dapat dilihat pada Gambar 3.
Berdasarkan Gambar 3 dapat dilihat bahwaperlakuan suhu dan lama penyimpanan masihberpengaruh terhadap aktivitas antimikrobaisolat bakteri Bacillus cereus strain HVR22dalam menghambat bakteri patogen. Pada suhu370C dan lama penyimpanan 14 hari aktivitasantimikrobial yang paling tinggi adalah terhadapbakteri Salmonella thypi dengan diameter zonabeningnya 18 mm, diikuti oleh Bacillus subtilis14 mm, Staphylococcus aureus 12,5 mm,Escherichia coli dan Listeria monocytogenes.
Pada suhu 40C dan lama penyimpanan 14hari aktivitas antimikrobial yang paling tinggiadalah terhadap bakteri Salmonella thypi, diikutioleh Listeria monocytogenes, Bacillus subtilis,Staphylococcus aureus dan Escherichia coli.Pada suhu -100C dan lama penyimpanan 14 hari,senyawa antimikroba yang dihasilkan oleh isolatbakteri Bacillus cereus strain HVR22 memilikiaktivitas antimikrobial yang paling tinggiterhadap bakteri Salmonella thypi dengandiameter zona beningnya 13 mm, diikuti olehStaphylococcus aureus dengan diameter zona
beningnya 8,5 mm, Escherichia coli, Bacillus
subtilis dan Listeria monocytogenes.
Perbedaan lamanya umur simpan isolatbakteri Bacillus cereus strain HVR22mengalami penurunan aktivitas penghambatanpada penyimpanan selama pada suhu 40C,namun perpanjangan penyimpanan hingga 14hari mampu mengembalikan aktivitas peng-hambatan supernatan bakteri Bacillus cereusstrain HVR22 dengan menghasilkan diameterzona hambat terhadap bakteri Salmonella thypi.Nofisulastri et al., (2006), aktivitas antimikrobabakteriosin dari Pediococcus sp NWD 015 stabilpada suhu 40C dan lama penyimpanan 10 hari,tetapi menurun pada suhu 20oC padapenyimpanan 7 hari. Menurut Amanah (2011),penyimpanan 1 minggu pada suhu refrigerator
menyebabkan FBS (filtrat bebas sel) L.acidophilus Y-01 mengalami penurunanaktivitas penghambatan yang sangat nyata,namun perpanjangan penyimpanan hingga 2minggu mampu mengembalikan aktivitaspenghambatan FBS L. acidophilus Y-01 denganmenghasilkan diameter zona hambat yang tidakberbeda dengan kontrol. Aktivitas plantarisinselama penyimpanan suhu dingin bersifatfluktuatif. Penyimpanan plantarisin selama 10hari efektif digunakan untuk uji antagonistikterhadap bakteri Staphylococcus aureus ATCC25923 karena menghasilkan diameter zonahambat yang optimum.
Selanjutnya penelitian Bariyah (2012),Plantarisin dari empat galur L. plantarum 1A5,1B1, 2B2 dan 2C12 setelah mengalamipenyimpanan selama 15 hari pada suhu dingin(10ºC) masih mempunyai aktivitas antimikrobterhadap bakteri patogen indikator yaituSalmonella enteritidis ser. Typhimurium ATCC14028, E. coli ATCC 25922, S. aureus ATCC25923, B. cereus dan P. aeruginosa ATCC27853. Plantarisin 2C12 memiliki tingkatsensitivitas paling tinggi dibandingkan 1A5, B1dan 2C12 selama penyimpanan 15 hari padasuhu dingin.

Yusra dan Yempita Efendi 308
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :Setelah dilakukan karakterisasi diketahui bahwaisolat bakteri Bacillus cereus strain HVR22masih aktivitas senyawa antimikroba pada suhurendah (40C sampai suhu -200C), dapat tumbuhpada konsentrasi NaCl 2,5% sampai dengan20%, dan masih memiliki aktivitas setelahdisimpan selama 14 hari terhadap lima bakteripatogen.
DAFTAR PUSTAKA
Amanah, N. 2011. Identifikasi dan karakterisasisubstrat antimikroba dari bakteri asamlaktat kandidat probiotik yang diisolasi daridadih dan yogurt. Skripsi. FakultasPeternakan. Institut Pertanian Bogor,Bogor.
Ananou, S., A. Gálvez., M. Martínez-Bueno., M.Maqueda and E. Valdivia. 2005.Synergistic effect of enterocin AS-48 incombination with outer membranepermeabilizing treatments againstEscherichia coli O157:H7. Journal ofApplied Microbiology, 99: 1364–1372.
Axelsson, L., 2004. Lactic acid bacteria:classification and physiology. In: Salminen,S. von Wright, A. and Ouwehand A. (Eds.),lactic acid bacteria: microbiological andfunctional aspects. 3rd rev. and exp. ed.Marcel Dekker, Inc., New York, pp.1-66.
Bariyah, K. 2012. Aktivitas antimikrobbakteriosin asal Lactobacillus plantarumterhadap berbagai bakteri patogen selamapenyimpanan suhu dingin. Skripsi.Departemen Ilmu Produksi dan TeknologiPeternakan, Fakultas Peternakan, InstitutPertanian Bogor.
Bizani, D., A. S. Motta., J. A. C. Morrissy., R.M. S. Terra., A. A. Souto dan A. Brandelli.,2005. Antibacterial activity of cerein 8A, abacteriocin-like peptide produced byBacillus cereus. Int Micr, 8: 125-131.
Buckle, K. A., R. A. Edwards., G. H. Fleet danM. Wootton., 1987. Ilmu pangan. Purnomo,H dan Adiono, Penerjemah. Jakarta:Universitas Indonesia Press. Terjemahandari: Food Science.
Day, R. A. Jr. & A. L. Underwood. 2002.Analisis Kimia Kuantitatif. EdisiKeenam.Erlangga, Jakarta.
Dwidjoseputo, D., 1978. Dasar-dasarmikrobiologi. Penerbit Jambatan. Malang.
Fardiaz, S., 1989. Mikrobiologi panganpenuntun praktek laboratorium. Bogor:Jurusan Teknologi Pangan dan Gizi,Fakultas Teknologi Pertanian, InstitutPertanian Bogor, Bogor.
Gray, E. J., K. D. Lee., A. M. Souleimanov., M.R. Di Falco., X. Zhou., A. Ly., T. C.Charles., B. T. Driscoll dan D. L. Smith.,2006. A novel bacteriocin, turicin 17,produced by plant growth promotingRhizobacteria strain Bacillus thuringiensisNEB17: isolation and classification. J. ApplMicr, 100: 545-554.
Ha, Y. M., Y. H. Park dan Y. J. Kim., 2002. Ataxonomic study of Bacillus sp. isolatedfrom Korean salt fermented Anchovy. MolBio Today, 3(1): 25-29.
Hena, J. V dan C. Mamatha., 2014.Characterization of bacteriocin fromBacillus subtilis an Its application as milkpreservative. Asian Academic Research J.of Multidisciplinary, 1 (18):57-64.
Jack, R. W., J. R. Tagg dan R. Ray., 1995.Bacteriocins of gram positive bacteria.Micr Rev, 59(2): 171-200.
Jay, J. M., 2003. Modern food microbiology.Fourth Edition. Champman & Hall. NewYork. 254 p.
Lay B. W., 1994. Analisis mikroba dilaboratorium. Jakarta: PT. Raja Persada.
Nofisulastri, Z., Bachruddin dan E. Harmayanti,2006. Production and extraction ofantibacterial bacteriocin from Pediococcussp. NWD 015. Indon J Biotech, 11(2): 921-927.
Parada, J. L., C. R. Caron., A.B. P. Medeiros &C.R. Soccol. 2007. Bacteriocins from lacticacid bacteria: purification, properties anduse as biopreservatives. BrazilianArchive of Biology and Technology,50(3): 521-542.
Risoen, P. A., P. Ronning., I. K. Hegna dan A. B.Kolsto., 2004. Characterization of a broadrange antimicrobial substance fromBacillus cereus. J. Appl Micr, 96: 648-655.
Sharma, N., G. Kapoor., N. Gautam dan B.Neopaney, 2008. Characterization of apartially purified bacteriocin of Bacillus spMTCC 43 isolated from rhizosphere ofRadish (Raphanus sativus) and its

Yusra dan Yempita Efendi 309
application as a potential foodbiopreservative. J. Sci & Ind Research, 68:881-886.
Stanier, R. Y., M. Doudorrof dan E. A.Adelberg., 1976. The microbial world.Prantice-Hall, New York.
Sudiarta, I. W., 2011. Isolasi dan identfikasibakteri asam laktat indigenous dari kecapikan Lemuru (Sardinella longiceps) selamafermentasi. Tesis. Program PascasarjanaUniversitas Udayana, Bali.
Um, M. N. dan C.H. Lee., 1996. Isolation andidentification of Staphylococcus sp. fromKorean fermented fish products. J. MicrBiotech, 6: 340-346.
Yusra., F. Azima,. Novelina dan Periadnadi.2013. Antimicrobial activity of lactic acidbacteria isolated from budu of WestSumatera to food biopreservatives.Pakistan Journal of Nutrition, 12(7): 628-635.

ISSN 978-602-14989-0-3 BioETI
Penggunaan Air Kelapa, Air Cucian Beras dan Air RendamanJagung terhadap pertumbuhan Miselium Jamur Kuping Hitam(Auricularia polytricha (Mont.) Sacc) dalam media pembibitandan media poduksiZA’AZIZA RIDHA JULIA, NURMIATI DAN PERIADNADI
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: [email protected]
ABSTRAKPenelitian mengenai “Penggunaan Air Kelapa, Air Cucian Beras dan Air Rendaman Jagung Terhadap PertumbuhanMiselium Jamur Kuping Hitam (Auricularia polytricha (Mont.) Sacc) Dalam Media Pembibitan Dan Media Poduksi”telah dilaksanakan dari bulan April sampai September 2014 di Laboratorium Mikrobiologi, Jurusan Biologi, FakultasMatematika dan Ilmu Pengetahuan Alam, Universitas Andalas, Padang. Penelitian ini bertujuan untuk mengetahuipengaruh pencelupan air kelapa, air cucian beras dan air rendaman jagung terhadap kecepatan pertumbuhan miseliumjamur kuping hitam dalam media pembibitan dan media produksi. Serta untuk mengetahui perlakuan terbaikpertumbuhan miselium jamur kuping hitam pada media pembibitan dan media produksi. Penelitian ini menggunakanmetoda eksperimen Rancangan Acak Lengkap (RAL), yang terdiri dari 2 tahap, yaitu tahap media pembibitan dan tahapmedia produksi, dengan 4 perlakuan dan 6 kali ulangan. Tahap pertama, pada media pembibitan dengan perlakuanpenggunaan air kelapa, air rendaman beras, air rendaman jagung dan air kran (kontrol). Kemudian dilanjutkan tahapkedua pada media produksi, dengan perlakuan pencelupan air kelapa, air cucian beras, air rendaman jagung dan airkran (Kontrol). Hasil penelitian menunjukkan bahwa perlakuan air rendaman jagung dan air cucian beras pada media-pembibitan dan-produksi berpengaruh nyata terhadap kecepatan pertumbuhan miselium jamur kuping hitam. Perlakuanair rendaman jagung memberikan hasil terbaik pada media pembibitan dengan rata-rata kecepatan pertumbuhan miselium0,62 cm/hari,. Sedangkan perlakuan air cucian beras memberikan hasil terbaik pada media produksi dengan rata-ratakecepatan pertumbuhan miselium 0,63 cm/hari.
Key words: jamur kuping hitam, kecepatan pertumbuhan miselium, air kelapa, air cucian beras, air rendaman jagung.
Pendahuluan
Jamur kuping hitam (Auricularia polytricha(Mont.) Sacc) merupakan salah satu jamurkonsumsi jamur konsumsi yang bernilai gizitinggi dan sangat digemari oleh konsumen dibanyak negara, khususnya di Cina dan Jepang.Menurut Nurilla, Lilik dan Ellis (2013)kandungan gizi jamur ini diantaranya protein,lemak, karbohidrat, riboflavin, niacin, Ca, K, P,Na, dan Fe. Selain itu, jamur ini juga bisadimanfaatkan sebagai obat, sepertiantihiperlipidemia. Seiring denganmeningkatnya kesadaran masyarakat akanpentingnya konsumsi jamur kuping sebagaimakanan yang lezat dan bergizi, makapermintaan konsumen dan pasar terusmeningkat diberbagai daerah. Namun,
permintaan pasar tersebut tidak mudahdipenuhi karena terbatasnya bahan baku berupabibit dalam media tanam yang baik danmemenuhi syarat tumbuh jamur (Djarijah danSiregar,2001). Hal ini menjadi peluang untukmembuat usaha bibit dan budidaya jamurkuping.
Permasalahan yang terjadi dalampembibitan dan baglog adalah lambatnyapertumbuhan miselium yang berpotensikontaminasi, selain itu pertumbuhan miseliumberpacu dengan waktu pemesanan. Hal inimenyebabkan terjadinya kelangkaan bibitjamur, seperti yang terjadi di Sleman,Yogyakarta. Menurut Sigit, (2013), sulitnyamencari bibit jamur kuping membuat hasilproduksi petani jamur di PandansarenHarjobinangun Pakem Sleman turun tajam.

Za’aziza Ridha Julia, Nurmiati dan Periadnadi 311
Penurunan produktivitas jamur kuping inimencapai 75% dari hari biasanya.
Masyarakat di Sumatera Barat, sudah mulaimenekuni usaha budidaya jamur kuping.Namun usaha yang bergerak dibagianpembibitan jamur kuping masih langka,sehingga petani jamur harus memesan bibitjamur di luar pulau Sumatera, seperti di pulauJawa. Tempat pemesanan bibit jamur yang jauh,membutuhkan biaya pengiriman yang mahaldaripada harga bibit itu sendiri. Selain itu, tidakada jaminan kualitas bibit jamur yang baik.
Pada penelitian ini dilakukan penambahanbahan alami pada media bibit dan mediaproduksi jamur kuping, yang berpotensimerangsang pertumbuhan miselium jamur.Adapun bahan yang digunakan adalah airkelapa, air cucian beras dan air rendamanjagung. Berdasarkan uraian diatas, maka perludilakukan penelitian mengenai “PenggunaanAir Kelapa, Air Cucian Beras dan AirRendaman Jagung Terhadap PertumbuhanMiselium Jamur Kuping Hitam (Auriculariapolytricha (Mont.) Sacc) Dalam MediaPembibitan Dan Media Produksi” dalam usahameningkatkan pertumbuhan jamur kupinghitam.
BAHAN DAN METODE
Penelitian ini menggunakan metodaeksperimen Rancangan Acak Lengkap (RAL).Penelitian ini dilakukan pada bulan Aprilsampai September 2014 di LaboratoriumMikrobiologi, Jurusan Biologi, FakultasMatematika dan Ilmu Pengetahuan Alam,Universitas Andalas. Penelitian ini dilakukanmelalui 2 tahap, yaitu tahap media pembibitan,kemudian dilanjutkan dengan tahap mediaproduksi, yang terdiri 4 perlakuan dan 6 kaliulangan.Tahap I : Media Pembibitan
Tahap II : Media ProduksiA1: pencelupan air kelapa B1 :penambahan air kelapaA2: pencelupan air cucian beras B2 :pencelupan air cucian beras
A3: pencelupan air rendaman jagungB3 : pencelupan air rendaman jagungA4: pencelupan air kran (Kontrol)B4 : pencelupan air kran (Kontrol)
Parameter yang diamati ialah kecepatanpertumbuhan miselium pada media pembibitandan media produksi. Data yang diperoleh diujisecara statistik dengan menggunakanRancangan Acak Lengkap. Apabila dengan ujiF pada taraf 5 % terdapat perbedaan yang nyataantar perlakuan maka analisis ragamdilanjutkan dengan uji DNMRT (Duncan NewMultiple Range Test).
HASIL DAN PEMBAHASAN
I. CendawanKecepatan PertumbuhanMiselium Jamur Kuping Hitam pada MediaPembibitanKecepatan pertumbuhan miselium jamurkuping hitam pada media pembibitan, setelahdianalisa statistik dan uji DNMRT pada taraf5%, data tersebut menunjukkan perbedaannyata pada masing-masing perlakuan. Datatentang rata-rata kecepatan pertumbuhanmiselium masing-masing perlakuan dapatdilihat pada Tabel 1.
Tabel 1. Rata-rata kecepatan pertumbuhanmiselium jamur kuping hitam padamedia bibit setelah uji statistikdengan DNMRT 5%
No.
Jenis Perlakuan
Rata-RataKecepatanPertumbuhanMiselium(cm/hari)
Notasi
1. A1 (Air Kelapa) 0,48 d
2. A2 (Air Cucianberas)
0,58b
3. A3 (Air RendamanJagung)
0,62a
4. A4 (Kontrol) 0,52 c
Keterangan : Angka yang tidak diikuti huruf kecil yangsama berbeda nyata pada taraf = 0,05 pada ujiDNMRT
Dari Tabel 1 di atas, nilai kecepatanpertumbuhan miselium tertinggi terdapat padaperlakuan air rendaman jagung, dengan rata-
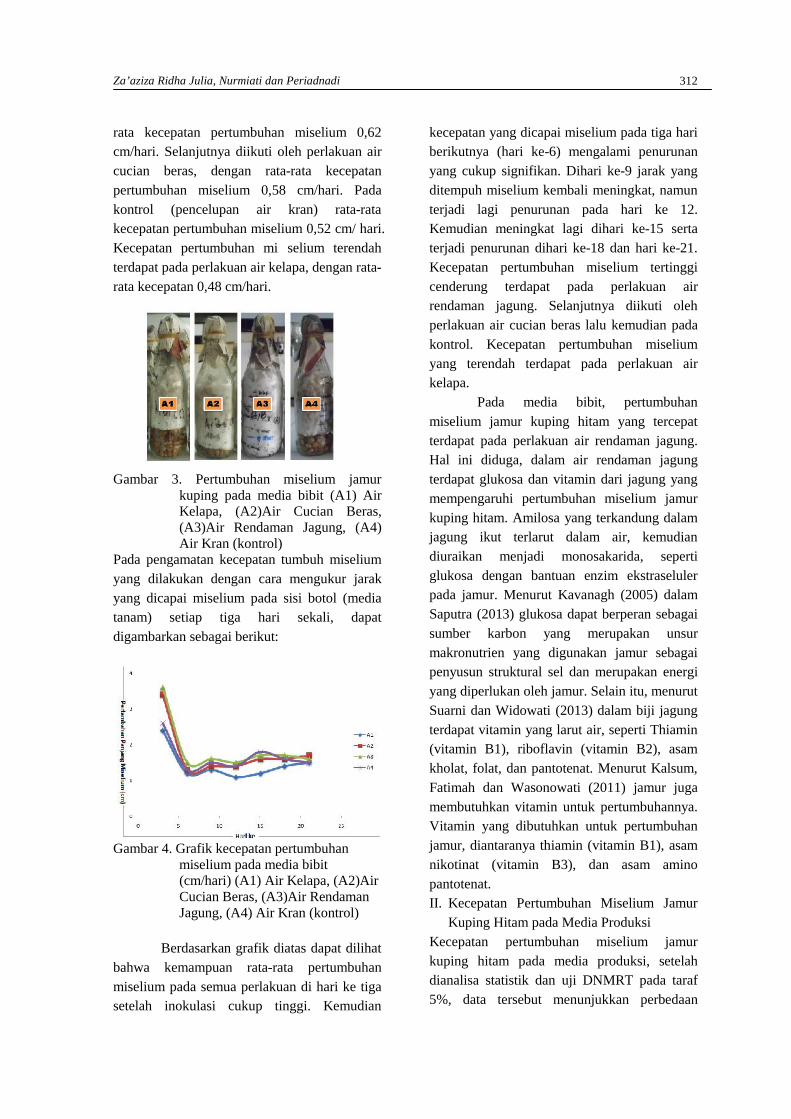
Za’aziza Ridha Julia, Nurmiati dan Periadnadi 312
rata kecepatan pertumbuhan miselium 0,62cm/hari. Selanjutnya diikuti oleh perlakuan aircucian beras, dengan rata-rata kecepatanpertumbuhan miselium 0,58 cm/hari. Padakontrol (pencelupan air kran) rata-ratakecepatan pertumbuhan miselium 0,52 cm/ hari.Kecepatan pertumbuhan mi selium terendahterdapat pada perlakuan air kelapa, dengan rata-rata kecepatan 0,48 cm/hari.
Gambar 3. Pertumbuhan miselium jamurkuping pada media bibit (A1) AirKelapa, (A2)Air Cucian Beras,(A3)Air Rendaman Jagung, (A4)Air Kran (kontrol)
Pada pengamatan kecepatan tumbuh miseliumyang dilakukan dengan cara mengukur jarakyang dicapai miselium pada sisi botol (mediatanam) setiap tiga hari sekali, dapatdigambarkan sebagai berikut:
Gambar 4. Grafik kecepatan pertumbuhanmiselium pada media bibit(cm/hari) (A1) Air Kelapa, (A2)AirCucian Beras, (A3)Air RendamanJagung, (A4) Air Kran (kontrol)
Berdasarkan grafik diatas dapat dilihatbahwa kemampuan rata-rata pertumbuhanmiselium pada semua perlakuan di hari ke tigasetelah inokulasi cukup tinggi. Kemudian
kecepatan yang dicapai miselium pada tiga hariberikutnya (hari ke-6) mengalami penurunanyang cukup signifikan. Dihari ke-9 jarak yangditempuh miselium kembali meningkat, namunterjadi lagi penurunan pada hari ke 12.Kemudian meningkat lagi dihari ke-15 sertaterjadi penurunan dihari ke-18 dan hari ke-21.Kecepatan pertumbuhan miselium tertinggicenderung terdapat pada perlakuan airrendaman jagung. Selanjutnya diikuti olehperlakuan air cucian beras lalu kemudian padakontrol. Kecepatan pertumbuhan miseliumyang terendah terdapat pada perlakuan airkelapa.
Pada media bibit, pertumbuhanmiselium jamur kuping hitam yang tercepatterdapat pada perlakuan air rendaman jagung.Hal ini diduga, dalam air rendaman jagungterdapat glukosa dan vitamin dari jagung yangmempengaruhi pertumbuhan miselium jamurkuping hitam. Amilosa yang terkandung dalamjagung ikut terlarut dalam air, kemudiandiuraikan menjadi monosakarida, sepertiglukosa dengan bantuan enzim ekstraselulerpada jamur. Menurut Kavanagh (2005) dalamSaputra (2013) glukosa dapat berperan sebagaisumber karbon yang merupakan unsurmakronutrien yang digunakan jamur sebagaipenyusun struktural sel dan merupakan energiyang diperlukan oleh jamur. Selain itu, menurutSuarni dan Widowati (2013) dalam biji jagungterdapat vitamin yang larut air, seperti Thiamin(vitamin B1), riboflavin (vitamin B2), asamkholat, folat, dan pantotenat. Menurut Kalsum,Fatimah dan Wasonowati (2011) jamur jugamembutuhkan vitamin untuk pertumbuhannya.Vitamin yang dibutuhkan untuk pertumbuhanjamur, diantaranya thiamin (vitamin B1), asamnikotinat (vitamin B3), dan asam aminopantotenat.II. Kecepatan Pertumbuhan Miselium Jamur
Kuping Hitam pada Media ProduksiKecepatan pertumbuhan miselium jamurkuping hitam pada media produksi, setelahdianalisa statistik dan uji DNMRT pada taraf5%, data tersebut menunjukkan perbedaan
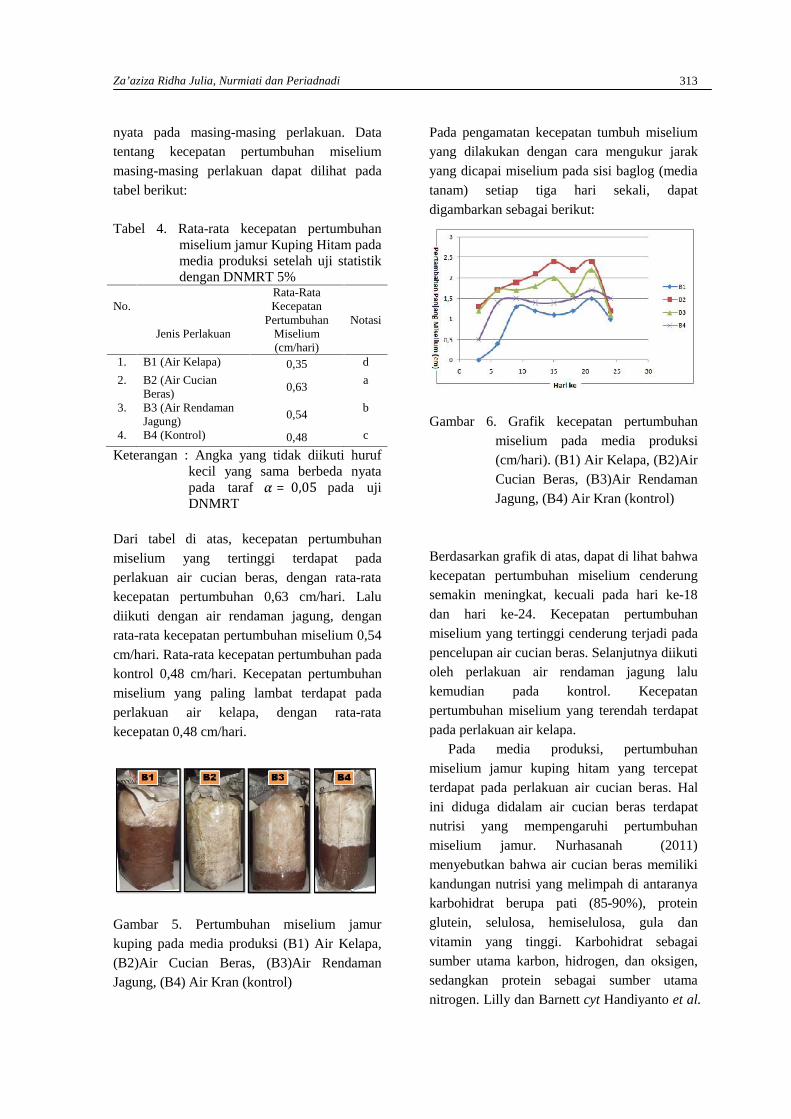
Za’aziza Ridha Julia, Nurmiati dan Periadnadi 313
nyata pada masing-masing perlakuan. Datatentang kecepatan pertumbuhan miseliummasing-masing perlakuan dapat dilihat padatabel berikut:
Tabel 4. Rata-rata kecepatan pertumbuhanmiselium jamur Kuping Hitam padamedia produksi setelah uji statistikdengan DNMRT 5%
No.
Jenis Perlakuan
Rata-RataKecepatan
PertumbuhanMiselium(cm/hari)
Notasi
1. B1 (Air Kelapa) 0,35 d
2. B2 (Air CucianBeras)
0,63a
3. B3 (Air RendamanJagung)
0,54b
4. B4 (Kontrol) 0,48 c
Keterangan : Angka yang tidak diikuti hurufkecil yang sama berbeda nyatapada taraf = 0,05 pada ujiDNMRT
Dari tabel di atas, kecepatan pertumbuhanmiselium yang tertinggi terdapat padaperlakuan air cucian beras, dengan rata-ratakecepatan pertumbuhan 0,63 cm/hari. Laludiikuti dengan air rendaman jagung, denganrata-rata kecepatan pertumbuhan miselium 0,54cm/hari. Rata-rata kecepatan pertumbuhan padakontrol 0,48 cm/hari. Kecepatan pertumbuhanmiselium yang paling lambat terdapat padaperlakuan air kelapa, dengan rata-ratakecepatan 0,48 cm/hari.
Gambar 5. Pertumbuhan miselium jamurkuping pada media produksi (B1) Air Kelapa,(B2)Air Cucian Beras, (B3)Air RendamanJagung, (B4) Air Kran (kontrol)
Pada pengamatan kecepatan tumbuh miseliumyang dilakukan dengan cara mengukur jarakyang dicapai miselium pada sisi baglog (mediatanam) setiap tiga hari sekali, dapatdigambarkan sebagai berikut:
Gambar 6. Grafik kecepatan pertumbuhanmiselium pada media produksi(cm/hari). (B1) Air Kelapa, (B2)AirCucian Beras, (B3)Air RendamanJagung, (B4) Air Kran (kontrol)
Berdasarkan grafik di atas, dapat di lihat bahwakecepatan pertumbuhan miselium cenderungsemakin meningkat, kecuali pada hari ke-18dan hari ke-24. Kecepatan pertumbuhanmiselium yang tertinggi cenderung terjadi padapencelupan air cucian beras. Selanjutnya diikutioleh perlakuan air rendaman jagung lalukemudian pada kontrol. Kecepatanpertumbuhan miselium yang terendah terdapatpada perlakuan air kelapa.
Pada media produksi, pertumbuhanmiselium jamur kuping hitam yang tercepatterdapat pada perlakuan air cucian beras. Halini diduga didalam air cucian beras terdapatnutrisi yang mempengaruhi pertumbuhanmiselium jamur. Nurhasanah (2011)menyebutkan bahwa air cucian beras memilikikandungan nutrisi yang melimpah di antaranyakarbohidrat berupa pati (85-90%), proteinglutein, selulosa, hemiselulosa, gula danvitamin yang tinggi. Karbohidrat sebagaisumber utama karbon, hidrogen, dan oksigen,sedangkan protein sebagai sumber utamanitrogen. Lilly dan Barnett cyt Handiyanto et al.

Za’aziza Ridha Julia, Nurmiati dan Periadnadi 314
(2014) menyatakan bahwa unsur-unsur C, H, O,N berperan sebagai unsur penyusun sel,fungsional sel (enzim), dan proses transferenergi, sehingga mempercepat pertumbuhanmiselium jamur. Air cucian beras jugamengandung vitamin dan mineral. Chetana, et
al. (2011) menyebutkan bahwa air cucian berasmengandung vitamin seperti niacin, riboflavindan thiamine, serta mineral seperti kalsium,magnesium dan besi. Vitamin dan mineraldalam air cucian beras berperan dalampertumbuhan miselium jamur kuping hitamkarena menurut Lilly dan Barnett (1951) cytHandiyanto et al. (2014) mineral sepertimagnesium, kalium, kalsium dan ferrumberperan dalam aktivasi enzim yang terlibatdalam reaksi enzimatik, sedangkan vitaminberperan sebagai katalisator.
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Perlakuan air rendaman jagung dan air
cucian beras pada media pembibitan danmedia produksi berpengaruh nyata terhadapkecepatan pertumbuhan miselium jamurkuping hitam.
2. Perlakuan air rendaman jagung memberikanhasil terbaik pada media pembibitanterhadap kecepatan pertumbuhan miseliumjamur kuping hitam. Adapun perlakuan aircucian beras memberikan hasil terbaik padamedia produksi terhadap kecepatanpertumbuhan miselium.
DAFTAR PUSTAKAChetana, S.H., B. Pratap, S. Roy, A. Jaiswal,
Shuruthi, dan Vedamurthy. 2011.Bioethanol Production From Rice WaterWaste : A Low Cost Motor Fuel.Pharmacologyonline 3 : 125-134
Djarijah, N.M dan A. Siregar. 2001. BudidayaJamur Kuping Pembibitan danPemeliharaan. Kanisius. Yogyakarta
Handiyanto, S., U. S. Hastuti, dan S.Prabaningtyas. 2014. Pengaruh MediumAir Cucian Beras terhadap KecepatanPertumbuhan Miselium Biakan MurniJamur Tiram Putih. Seminar Nasional XPendidikan Biologi FKIP UNS.Universitas Negeri Malang. Malang
Kalsum U., S. Fatimah dan C. Wasonowati.2011. Efektivitas Pemberian Air Leriterhadap Pertumbuhan dan Hasil JamurTiram Putih (Pleurotus osreatus).Agrogivor Vol. 4 No.2
Nurhasanah, Y.S. 2011. Air Cucian BerasDapat Suburkan Tanaman. InstitutPertanian Bogor. www.kabarkampus.comdiakses tanggal 17 September 2014
Nurilla, N., L. Setyobudi, dan E. Nihayati.2013. Studi Pertumbuhan dan ProduksiJamur Kuping (Auricularia auricula) padaSubstrat Serbuk Gergaji Kayu Dan SerbukSabut Kelapa. Universitas Brawijaya.Malang
Saputra, D. Y. 2013. Pengaruh PengaturanKeasaman Limbah Industri Teh TerhadapPelapukan Serta Ekspresinya PadaPertumbuhan Dan Produksi Jamur TiramCokelat (Pleurotus Cystidiosus O.K.Miller). Skripsi. Univ. Andalas. Padang
Sigit, A. 2013. Bibit Langka, Produksi JamurKuping Pakem Turun 75 Persen. http://KRjogja.com diakses tanggal 25 Oktober2013
Suarni dan S. Widowati. 2013. Struktur,Komposisi dan Nutrisi Jagung. BalaiPenelitian Tanaman Serealia. Bogor

ISSN 978-602-14989-0-3 BioETI
Keanekaragaman gulma pada kebun Kopi (Coffea arabica L.) dinagari Balingka, Kecamatan Ampek Koto, Kabupaten AgamZUHRI SYAM , CHAIRUL DAN INDAH PRAFITRI YUSA
Jurusan Biologi FMIPA, Universitas Andalas, Kampus Limau Manis Padang 25163E-mail: nurmiati @ fmipa.unand.ac.id
ABSTRAKKeanekaragaman gulma pada kebun kopi arabika Coffea arabica L. Di nagari Balingka, Kecamatan Ampek Koto,Kabupaten Agam telah dilakukan dari bulan Februari - Mei 2014. Tujuan penelitian ini adalah untuk mengetahuikomposisi dan struktur gulma pada kebun kopi arabika tersebut. Penelitian ini menggunakan metoda kuadrat secarasistematis dengan jumlah 32 plot ukuran 1x1 m, dengan jarak interval plot 10 m dengan intensitas sampel 1% dari luaskebun yang ada. Hasil penelitian menunjukkan komposisi gulma pada kebun kopi arabika adalah 11 famili, 21 genus dan25 spesies. Kerapatan gulma dengan jumlah total 3114 individu. Borreria laevis mendominasi pada kebun kopi arabikayaitu sebanyak 975 individu dengan SDR (Summed Dominance Ratio) tertinggi 22,809%. Indeks keanekaragaman padakebun kopi arabika adalah H’= 2,04 (sedang).
Key words: Gulma, kopi arabika, komposisi, struktur
Pendahuluan
Kopi merupakan jenis tumbuhan perkebunanyang sudah lama dibudidayakan dan memilikinilai ekonomi tinggi serta komoditi pentingdalam bidang perkebunan (Rahardjo, 2012).Sejauh ini produksi kopi di Indonesia telahmencapai 600 ribu ton pertahun dan lebih dari80% berasal dari perkebunan rakyat.Berdasarkan data produksi kopi di Indonesiatahun 2008-2012, pada tahun 2008 berkisar698.016 ton dan pada tahun 2012 turun menjadi657.138 ton (Departemen pertanian, 2012).Hasil survei lapangan yang telah dilakukan,salah satu kebun kopi arabika yang dikelolaoleh rakyat di Sumatera Barat terdapat didaerah Bancah, Nagari Balingka, KecamatanAmpek Koto, Kabupaten Agam. Luas kebunkopi arabika didaerah ini ±2 ha dengan jaraktanam antara pohon ke pohon berikutnya 2,5 x2,5 m dan masa penyiangan selama 2-3 bulan.Adapun umur kopi pada daerah ini yaitu 8tahun. Menurut Dinas Perkebunan PropinsiSumatera Barat (2011), Nagari BalingkaKecamatan Ampek Koto terletak padaketinggian 1000-1300 mdpl, dengan curahhujan >4500 mm/tahun, suhu udara 20oC –29oC dan kelembaban udara 88%.
Kopi arabika merupakan kopi yangmemiliki daya produksi yang lebih rendah,membutuhkan pemeliharaan yang rumit dansiklus pertumbuhan yang lebih lama (Prastowo,Karmawati, Rubijo, Siswanto, Indrawanto danMunarso, 2010). Kopi ini ditanam pada datarantinggi yang memiliki musim kering danpenghujan dan ketinggian sekitar 1000-1750mdpl (Najiyanti dan Danarti, 2004).Perkebunan kopi ini banyak mengalamigangguan yang sangat merugikan. Gangguan-gangguan tersebut kebanyakan disebabkan olehgulma.. Gulma merupakan tumbuhan yangberadaptasi dengan lingkungan dankeberadaannya dapat menimbulkan masalah.Salah satunya gulma dapat menjadi inang bagihama dan penyakit yang dapat menurunkanhasil produksi bagi tanaman budidaya. Untukmencegah serangan hama dan penyakit tersebutdiperlukan tindakan menjaga kebersihandisekitar tanaman dari adanya gulma.Banyaknya gulma yang tumbuh, maka semakinbanyak pula serangga yang berdatanganmerusak tanaman kopi (Utomo, Nuswandaridan Lontoh, 1989). Masalah keberadaan gulmapada pertanaman budidaya adalah persaingandalam pengambilan cahaya, air, unsur hara danruang tumbuh (Sastroutomo, 1990).

Zuhri Syam, Chairul dan Indah Prafitri Yusa 316
Persaingan gulma berpengaruh terhadapbaik buruknya pertumbuhan tanaman pokokdan tinggi rendahnya hasil produksi tanamanpokok. Persaingan antara gulma dan tanamanpokok yaitu memperebutkan air, hara, CO2 dancahaya. Sehingga persaingan antar gulmadengan tanaman adalah persaingan interspesifik karena terjadi antar spesies tumbuhanyang berbeda. Adapun tinggi rendahnyahambatan terhadap pertumbuhan atau hasiltanaman pokok dilihat dari segi gulma yangdipengaruhi oleh beberapa faktor yaitukerapatan gulma, jenis gulma, saat kemunculangulma, lama keberadaan gulma, kecepatantumbuh gulma, habitat gulma dan alelopati(Sukman dan Yakup, 1995).
Menurut penelitian sebelumnya Widiyanti(2013) menyatakan kondisi kebun sumberbenih kopi (Coffea sp.) di kebun Kasilat jampitBondowoso menunjukan hasil gulma yangbanyak adalah Imperata cylindrica, Paspalumconjugatum dan Cyperus rotundus. Sedangkanmenurut Moenandir (1990) mengatakan darilahan kopi di Desa Ampelgading Malang, pertahun terdapat gulma disekitar tanaman pokokkopi yaitu Setaria plicata, Paspalumconjugatum, Ageratum conyzoides, Cynodon
dactylon, Imperata cylindrica, Eleusine indica,Cyperus rotundus, Cyperus kilinga, Bidensbiternata, Erechtites valerianifolia danPanicum repens. Dengan adanya gulmadisekitar tanaman kopi dapat menurunkanproduksi biji 35% (dari 12,5 kw ha-1 menjadi 7kw ha-1).
Agar diperoleh tanaman kopi produksitinggi sangat diperlukan tindakan pemeliharaanseperti pemangkasan dan penyiangan gulma(Widiyanti, 2013). Pengendalian gulmaprinsipnya meningkatkan daya saing tanamanpokok dan melemahkan daya saing gulma.Sehingga pengendalian gulma merupakanpengelolaan organisme pengganggu dalamproses produksi pertanian (Sukman dan Yakup,1995). Pengendalian gulma dilakukan denganmengetahui jenis gulma dominan, tumbuhanbudidaya utama, alternatif pengendalian yang
tersedia, dampak ekonomi, ekologi dan parasit(Rambe, 2010).Berdasarkan uraian latar belakang diatas, makadirumuskan masalah sebagai berikut:Bagaimana komposisi gulma dan bagaimanastruktur gulma pada kebun kopi arabika (Coffea
arabica L.) di Nagari Balingka, KecamatanAmpek Koto, Kabupaten Agam. Adapun tujuandari penelitian ini adalah: Untuk mengetahuikomposisi gulma dan struktur gulma yangterdapat pada kebun kopi arabika (Coffeaarabica L.).
BAHAN DAN METODE
Penelitian ini telah dilaksanakan pada bulanFebruari - Mei 2014, dengan pengambilansampel di area kebun kopi di Balingka,Kecamatan Ampek Koto, Kabupaten Agam,kemudian dilanjutkan di Herbarium UniversitasAndalas (ANDA) dan Laboratorium EkologiJurusan Biologi Fakultas Matematika dan IlmuPengetahuan Alam Universitas Andalas Padang.Metoda penelitian yang digunakan dalampenelitian ini adalah Metoda Kuadrat denganpeletakan plot 1x1 secara sistematik, denganjarak interval plot 10 m dengan intensitassampel 1% dari luas kebun. Alat yangdigunakan dalam penelitian ini adalahtermometer, sling psycometer, meteran, soilmosture meter, pH meter, GPS, guntingtanaman, pancang, tali, label, kantong plastik,koran, karung, timbangan ohaus gram, oven,plastik packing, kamera digital, bukuidentifikasi tumbuhan dan alat-alat tulis.Sedangkan bahan yang digunakan adalahalkohol 70%.
Data yang didapatkan dilapangan dianalisisdengan menggunakan rumus berikut:
Kerapatan=
- Kerapatan Relatif (%) =100%- Frekuensi =
- Frekuensi Relatif (%) =100%

Zuhri Syam, Chairul dan Indah Prafitri Yusa 317
- Dominansi=
- Dominansi Relatif (%) =
x 100%
- Indeks Nilai Penting (INP) =KR + FR + DR
(Brower, Zar dan Von endle, 1990; Cox1992).
- Famili dominan dan co-dominanPersentase famili = 100%
Suatu famili dikatakan dominan pada suatukawasan jika memiliki persentase >20% daritotal individu dan co-dominan jikapersentasenya 10%-20% (Johnston danGillman, 1995).
- Summed Dominance Ratio (SDR) =
(Tjitrosoedirdjo, Utomo dan Wiroatmodjo,1984).
Hasil perhitungan nilai penting, selanjutnyadigunakan sebagai nilai untuk mengetahuibesarnya Indeks Keanekaragaman Jenis (H’)pada suatu komunitas dengan menggunakanrumus menurut (Johnston and Gillman, 1995).Odum (1996) mengatakan bahwakeanekaragaman jenis tumbuhan dapat dihitungmenggunakan Indeks KeanekaragamanShannon-Weinner yaitu:
H = -∑ ln ; pi = ni/N
Ket: H’ : Indeks KeanekaragamanJenis
pi : ni/Nni : Jumlah individu spesies ke-iN : Jumlah seluruh individu
Semakin besar nilai H’ maka akanmenunjukkan tingginya nilai keanekaragamanjenis. Besarnya nilai keanekaragaman jenisShannon didefinisikan sebagai berikut:H’>3,0 menunjukkan keanekaragaman tinggi1>H’≥3 menunjukkan keanekaragaman sedangH’<1 menunjukkan keanekaragaman renda
HASIL DAN PEMBAHASAN
Komposisi GulmaJenis gulma pada kebun kopi Arabika (C.
arabika L.) di Nagari Balingka, KecamatanAmpek Koto, Kabupaten Agam ditemukan 11famili, 21 genus, 25 spesies dan 3114 individu
(Tabel 1). Golongan teki-tekian 3 spesies,golongan rumput-rumputan 6 spesies, golonganberdaun lebar 13 spesies, dan golongan paku-pakuan 3 spesies. Gulma dengan jumlahindividu terbanyak adalah gulma jenis Borrerialaevis (975 individu) dan yang paling sedikitadalah jenis gulma Sonchus oleraceus (1individu), Tridax procumbens (1 individu) danCyperus cyperoides (1 individu). Berdasarkanhasil pengamatan , Borreria laevis merupakanjenis gulma dengan jumlah individu yangterbanyak (975 individu). Gulma ini didugamemiliki daya adaptasi yang tinggi dan dapatmengganggu pertumbuhan dan hasil tanamanbudidaya. Menurut Tjokrowardojo danDjauhariya (2013), mengatakan bahwa gulmaini merupakan salah satu jenis gulmakompetitif yang tinggi menghasilkan biji yangsangat banyak serta salah satu jenis gulmakompetitif yang tinggi menghasilkan biji yangsangat banyak serta mampu hidup di tempatyang terbuka dan agak terlindung sampaiketinggian hingga 1.100 m dpl. Gulma inibanyak hidup dan sering mengintervensi ladang,kebun, teh, karet, tebu dan lain-lain.
Selain itu pada kebun kopi Arabika jugaditemukan famili jumlah jenis terbanyak adalahfamili Asteraceae sebanyak tujuh jenis. JenisAsteraceae yang ditemukan adalah Ageratumconyzoides, Clibadium surinamensis, Emiliasonchifolia, Erigeron sumatrensis, Sonchus
oleraceus, Tridax procumbens dan Bidenspilosa. Adapun famili yang memiliki jumlahindividu terbanyak adalah famili Asteraceaeyaitu 1096 individu. Menurut Reader dan Buck(2000), gulma famili Asteraceae dapatberkembang biak melalui biji dan mempunyaikemampuan beradaptasi dengan lingkungan,misalnya sedikit banyaknya air dan tahanterhadap naungan. Kebutuhan akan cahaya,temperatur, air dan ruang terpenuhi sesuaidengan kebutuhannya, sehingga gulma inidapat berkembang cepat. Apabila lingkunganmenguntungkan baginya maka gulma ini akanterus berbunga sepanjang tahun.

Zuhri Syam, Chairul dan Indah Prafitri Yusa 318
Tjitrosoepomo, Soerjani dan Kostermans(1987) menyatakan bahwa gulma dari familiAsteraceae termasuk golongan gulma berdaunlebar dan semusim yang menyukai tanahsedikit lembab. Asteraceae dapat menghasilkanbiji sebanyak 40.000 pertanaman setiaptahunnya.
Sementara gulma yang sedikit ditemukanadalah jenis S.oleraceus, Tridax procumbensdan C. cyperoides. Menurut Robin (2011)menyatakan bahwa S. oleraceus ini merupakanjenis gulma berdaun lebar. Gulma inimerupakan tanaman musiman yang memilikibiji sedikit dan penyebarannya ketika suhutinggi. S. oleraceus umumnya tumbuh padahabitat yang terganggu serta tumbuh padaketinggian 400-700 mdpl, tetapi tidakditemukan pada ketinggian 1500 mdpl. Adanyatanaman kompetitif pada daerah tersebut makaakan sangat mengurangi biomassa dan produksidari gulma S. oleraceus ini.
T. procumbens atau gletang termasuk gulmadari famili Asteraceae yang merupakan sejenistumbuhan yang kebanyakan ditemukan liarpada daerah perkebunan. Gulma jenis ini(T.procumbens) termasuk kedalam golongangulma berdaun lebar. T. procumbens biasanyaditemukan pada tempat yang kering danmemiliki sinar matahari penuh (Susilo, 2013).
Dari tabel 1 diatas terlihat bahwa perbedaanjumlah individu gulma yang didapatkandipengaruhi oleh faktor lingkungan tempattumbuhnya yaitu suhu, temperatur, kelembaban,tanah, ruang tumbuh dan cahaya. Seperti yangdikatakan Moenandir (1993), menjelaskanbahwa yang mempengaruhi jumlah spesiesyang hidup pada suatu komunitas yaitu cahaya,dimana cahaya sangat berpengaruh terhadapjenis dan jumlah individu yang bisa tumbuh ditempat tersebut. Hal ini juga didukung olehpernyataan Lubis (1992), bahwa masalahgulma akan berbeda pada setiap tanaman, halini tergantung pada lokasi, iklim dan cahayayang diterima. Sastroutomo (1990), jugamenjelaskan bahwa komunitas gulma berbeda-beda pada satu tempat dengan tempat lainnya
baik pada jenis perkebunan yang sama maupunberbeda.
Struktur GulmaBerdasarkan hasil analisis data yang telah
dilakukan didapatkan struktur gulma padakebun kopi Arabika seperti terlihat pada Tabel2. Pada tabel 2 ini didapatkan nilai kerapatanrelatif, frekuensi relatif, dominansi relatif, nilaipenting, nilai SDR (Summed DominanceRatio) dan indeks keanekaragaman dari jenisgulma yang didapatkan. Masing-masingggulma bervariasi antara jenis yang satu denganjenis yang lainnya.
Pada lokasi ini nilai penting gulma tertinggiditemukan pada B. laevis (68,426%) denganSDR (22,809%). Sedangkan gulma yangmemiliki nilai penting terendah yaitu gulmajenis T. procumbens (0,689%) dengan SDR(0,230%) (Tabel 2). Hal ini menunjukkanbahwa gulma B.laevis paling dominan diantarajenis gulma lainnya pada kebun tanaman kopiarabika ini.
Tingginya nilai kerapatan relatif dandominansi relatif B. laevis yaitu (31,310%) dan(23,260), dibanding dengan gulma yang lainnyakarena mempunyai jumlah individu palingbanyak ditemukan disetiap plot pengamatandan penyebarannya yang luas pada kebun kopiArabika di Daerah Bancah, Nagari Balingka,Kecamatan Ampek Koto, Kabupaten Agam. B.laevis memiliki nilai penting dan nilai SDRpaling tinggi yaitu (68,426%) dan (22,809%).Kastanja (2012) juga menyatakan bahwa B.
laevis memiliki nilai penting tertinggi (5,64%)serta kemunculan terbanyak atau frekuensirelatif terbesar. Hal ini disebabkan bahwa jenisgulma B.laevis kebanyakan tumbuh pada lahanyang kering dan tergolong gulma penting padabeberapa lahan tanaman pangan. B.laevis
termasuk gulma penting tanaman pangan yangdijumpai pada pertanaman pagi gogo, jagung,kedelai, kacang tanah dan ketelapohon. GulmaB.laevis ini tergolong gulma berdaun lebar,mempunyai pertumbuhan yang cepat danmempunyai percabangan yang cukup banyakmengakibatkan untuk menghasilkan biji

Zuhri Syam, Chairul dan Indah Prafitri Yusa 319
menjadi banyak sehingga biomasanya menjaditinggi. Gulma ini tahan terhadap naungan,memiliki kerapatan yang tinggi dan penyebaranbiji yang merata, menyebabkan gulma inimenjadi salah satu gulma yang dominan(Suhardi, Sarbino dan Astina, 2011).
Borreria laevis salah satu jenis gulmadengan produksi biji yang sangat tinggi.Produksi bijinya dapat mencapai 9953 bijitanaman permusimnya dan tingkat dormansibiji yang cukup lama. Melalui produksi bijiyang banyak akan tumbuh dan berkembangindividu gulma yang banyak juga. Sehinggasemakin banyak produksi biji gulma pada suatulahan perkebunan maka kemungkinan gulmayang berkecambah akan semakin banyak pula(Sastroutomo, 1990).
Menurut Amperawati dan Basuki (1999)menyatakan bahwa faktor lain yangmempengaruhi keragaman gulma yaituketinggian daerah dari permukaan laut. Dalampenelitianya di kawasan Aek Nauli padaketinggian 950-1575 m dpl dijumpai lebihbanyak jenis tumbuhan berdaun lebar dansedikit jenis rerumputan. Ketinggian rata-ratalokasi penelitian 1200 m dpl dan suhu udararata-rata 24oC. Dengan ketinggian tempat daniklim tersebut, gulma yang dominan padategakan Eucalyptus spp. muda yaitu gulmaberdaun lebar seperti Borreria laevis (rumputkancing ungu).
B. laevis ditinjau dari segi kehadirannyayang tinggi pada suatu komunitas tumbuhandapat dikatakan bahwa semakin tinggi suatutempat biasanya berasosiasi denganpeningkatan keterbukaan, kecepatan angin,kelembaban udara dan penurunan suhu dapatmengakibatkan suatu komunitas yang tumbuhakan semakin homogen (Syafei, 1990). Hal inimenggambarkan bahwa tingkat penguasaanyang diberikan oleh spesies terhadap komunitas,semakin besar dan banyak (Soegianto, 1994)Selain B. laevis gulma yang memiliki kerapatanrelatif, frekuensi relatif, dan dominansi relatifyang tinggi adalah A. conyzoides yaitu(25,209%), (16,867%) dan (15,488%). Jenis
gulma A.conyzoides memiliki nilai penting(57,565%) dan SDR (19,188%), ini disebabkankarena jumlah individu hampir ditemukan padasetiap plot pengamatan.
Gulma A.conyzoides termasuk kedalamgolongan tumbuhan semusim yang banyaktumbuh pada di lahan pertanian, perkebunankaret, palawija, kopi, tembakau, cengkeh dankelapa sawit mulai dari pembibitan sampaiareal tanaman tua. Gulma ini dapat ditemukanhingga ketinggian 3.000 mdpl, meyukaiintensitas cahaya tinggi dan ternaungi. A.
conyzoides ini memiliki tekstur biji ringandengan jumlah biji yang banyak, dapat tersebardengan bantuan angin maka gulma ini cukupmengganggu perkebunan. Tumbuhan inimemiliki daya saing yang tinggi, sehinggadengan mudah tumbuh dimana-mana dansering menjadi gulma yang merugikan parapetani (Okunade, 2002).
A.conyzoides memiliki populasi yang lebihdominan dibandingkan tanaman liar lainyadalam suatu lahan. Tumbuhan ini diduga kuatmempunyai alelopati, suatu keadaan dimanatanaman mengeluarkan eksudat kimia yangdapat menekan pertumbuhan tanaman lainya. A.conyzoides menghasilkan alelopatidiidentifikasikan karena adanya 3 Phenolic acidyaitu Gallic acid, coumalic acid danprotocatechuic acid, yang dapat menghambatpertumbuhan beberapa gulma dan tanaman(Sukamto, 2007).
Berdasarkan hasil penelitian, nilai indekskeanekaragaman jenis tumbuhan pada kebunkopi arabika di Nagari Balingka, KecamatanAmpek Koto, Kabupaten Agam dikategorikankeanekaragaman jenis tingkat sedang karenajenis tumbuhan yang ada pada lokasi penelitianini tidak terlalu banyak. Nilai indeks darikeanekaragaman jenis gulma berada pada nilaiH’ = 2,04 (Tabel 2). Hal ini sesuai denganMargurran (2004) menyatakan bahwa nilaiindeks keanekaragaman Shannon yaitu 1>H≥3menunjukkan keanekaragaman sedang.Kemudian menurut Indriana (2009)menyatakan bahwa semakin besar nilai indeks
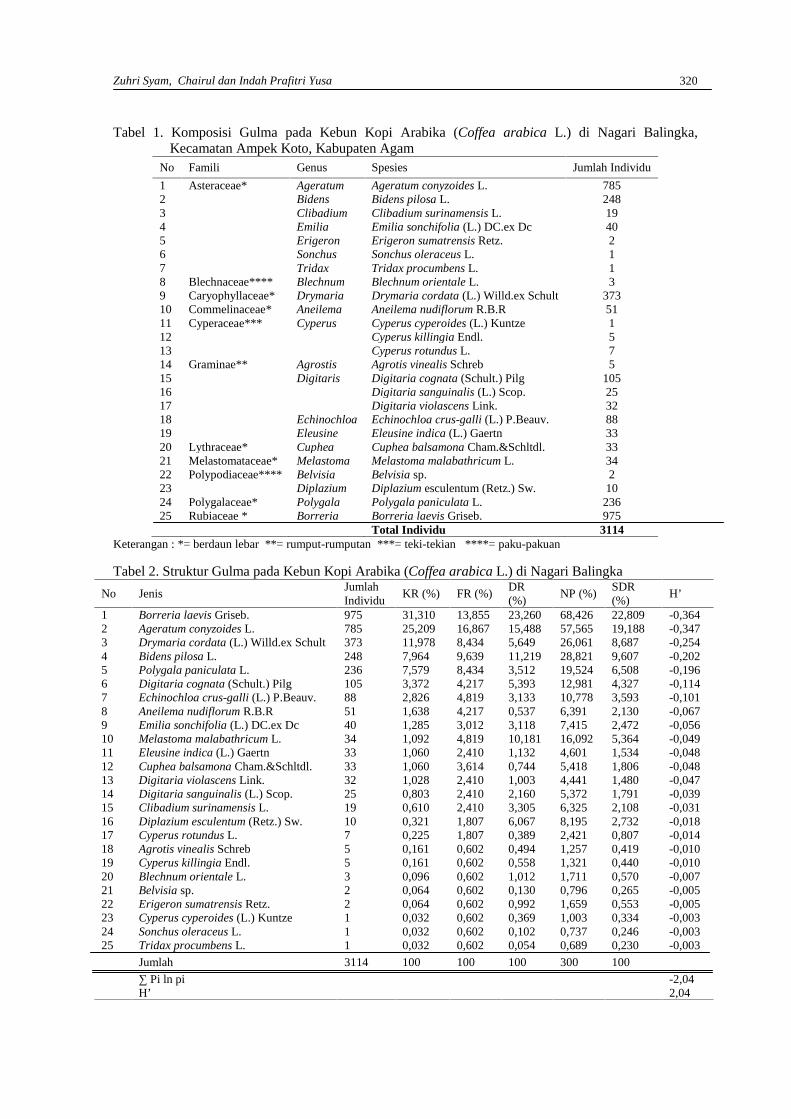
Zuhri Syam, Chairul dan Indah Prafitri Yusa 320
Tabel 1. Komposisi Gulma pada Kebun Kopi Arabika (Coffea arabica L.) di Nagari Balingka,Kecamatan Ampek Koto, Kabupaten Agam
No Famili Genus Spesies Jumlah Individu
1234567
Asteraceae* AgeratumBidensClibadiumEmiliaErigeronSonchusTridax
Ageratum conyzoides L.Bidens pilosa L.Clibadium surinamensis L.Emilia sonchifolia (L.) DC.ex DcErigeron sumatrensis Retz.Sonchus oleraceus L.Tridax procumbens L.
7852481940211
8910
Blechnaceae****Caryophyllaceae*Commelinaceae*
BlechnumDrymariaAneilema
Blechnum orientale L.Drymaria cordata (L.) Willd.ex SchultAneilema nudiflorum R.B.R
337351
11 Cyperaceae*** Cyperus Cyperus cyperoides (L.) Kuntze 112 Cyperus killingia Endl. 513 Cyperus rotundus L. 714 Graminae** Agrostis Agrotis vinealis Schreb 515 Digitaris Digitaria cognata (Schult.) Pilg 10516 Digitaria sanguinalis (L.) Scop. 2517 Digitaria violascens Link. 3218 Echinochloa Echinochloa crus-galli (L.) P.Beauv. 8819 Eleusine Eleusine indica (L.) Gaertn 3320 Lythraceae* Cuphea Cuphea balsamona Cham.&Schltdl. 3321 Melastomataceae* Melastoma Melastoma malabathricum L. 3422 Polypodiaceae**** Belvisia Belvisia sp. 223 Diplazium Diplazium esculentum (Retz.) Sw. 1024 Polygalaceae* Polygala Polygala paniculata L. 23625 Rubiaceae * Borreria Borreria laevis Griseb. 975
Total Individu 3114Keterangan : *= berdaun lebar **= rumput-rumputan ***= teki-tekian ****= paku-pakuan
Tabel 2. Struktur Gulma pada Kebun Kopi Arabika (Coffea arabica L.) di Nagari Balingka
No JenisJumlahIndividu
KR (%) FR (%)DR(%)
NP (%)SDR(%)
H’
12345678910111213141516171819202122232425
Borreria laevis Griseb.Ageratum conyzoides L.Drymaria cordata (L.) Willd.ex SchultBidens pilosa L.Polygala paniculata L.Digitaria cognata (Schult.) PilgEchinochloa crus-galli (L.) P.Beauv.Aneilema nudiflorum R.B.REmilia sonchifolia (L.) DC.ex DcMelastoma malabathricum L.Eleusine indica (L.) GaertnCuphea balsamona Cham.&Schltdl.Digitaria violascens Link.Digitaria sanguinalis (L.) Scop.Clibadium surinamensis L.Diplazium esculentum (Retz.) Sw.Cyperus rotundus L.Agrotis vinealis SchrebCyperus killingia Endl.Blechnum orientale L.Belvisia sp.Erigeron sumatrensis Retz.Cyperus cyperoides (L.) KuntzeSonchus oleraceus L.Tridax procumbens L.
97578537324823610588514034333332251910755322111
31,31025,20911,9787,9647,5793,3722,8261,6381,2851,0921,0601,0601,0280,8030,6100,3210,2250,1610,1610,0960,0640,0640,0320,0320,032
13,85516,8678,4349,6398,4344,2174,8194,2173,0124,8192,4103,6142,4102,4102,4101,8071,8070,6020,6020,6020,6020,6020,6020,6020,602
23,26015,4885,64911,2193,5125,3933,1330,5373,11810,1811,1320,7441,0032,1603,3056,0670,3890,4940,5581,0120,1300,9920,3690,1020,054
68,42657,56526,06128,82119,52412,98110,7786,3917,41516,0924,6015,4184,4415,3726,3258,1952,4211,2571,3211,7110,7961,6591,0030,7370,689
22,80919,1888,6879,6076,5084,3273,5932,1302,4725,3641,5341,8061,4801,7912,1082,7320,8070,4190,4400,5700,2650,5530,3340,2460,230
-0,364-0,347-0,254-0,202-0,196-0,114-0,101-0,067-0,056-0,049-0,048-0,048-0,047-0,039-0,031-0,018-0,014-0,010-0,010-0,007-0,005-0,005-0,003-0,003-0,003
Jumlah 3114 100 100 100 300 100
∑ Pi ln piH’
-2,042,04

Zuhri Syam, Chairul dan Indah Prafitri Yusa 321
Tabel 3. Famili Dominan dan Co-Dominan Gulma pada Kebun Kopi Arabika (Coffea arabica L.) diNagari Balingka, Kecamatan Ampek Koto, Kabupaten Agam
No Famili Jumlah IndividuDominan/Co-Dominan(%)
Golongan
1234567891011
AsteraceaeRubiaceaeCaryophyllaceaeGraminaePolygalaceaeCommelinaceaeMelastomaceaeLyhtraceaeCyperaceaePolypodiaceaeBlechnaceae
109697537328823651343313123
35,19631,31011,9789,2497,5791,6381,0921,0600,4170,3850,096
Berdaun lebarBerdaun lebarBerdaun lebarRumput-rumputanBerdaun lebarBerdaun lebarBerdaun lebarBerdaun lebarTeki-tekianPaku-pakuanPaku-pakuan
Jumlah 3114 100 %
diversitas atau keanekaragaman maka akansemakin tinggi keanekaragaman jenisnya. Halini dapat dipengaruhi oleh faktor lingkungansehingga habitat yang terbentuk hampir samaantar yang satu dengan yang lainnya.
Adanya keanekaragaman jenis gulma yangtumbuh pada perkebunan ini dipengaruhi olehlingkungan tempat tumbuhnya yaitu cahaya,suhu air, dan kelembaban. Kondisi lingkungansangat mempengaruhi keanekaragaman jenissuatu tumbuhan. Kondisi yang sangat ekstrimakan menyebabkan gangguan terhadapstabilitas kehidupan dan distribusi beragamtumbuhan (Ewusie, 1990). Selain itu jumlahindividu gulma yang banyak pada perkebunanini juga mempengaruhi nilai indekskeanekaragaman jenisnya (Mangoensoekarjo,1982). Odum (1996) juga menyatakan bahwatinggi rendahnya keanekaragaman jenis suatuorganisme didalam komunitasnya tergantungpada banyaknya (jumlah) individu yangterdapat pada komunitas tersebut.
Adapun keanekaragaman tumbuhandipengaruhi oleh pertumbuhan danperkembangbiakan dari masing-masing spesiestumbuhan. Pola pertumbuhan membentukrumpun dan cara perkembangbiakan yangberupa stolon menyebabkan tumbuhancenderung mempunyai sebaran yangberkembang. Tumbuhan yang memiliki teksturringan menyebab- kan sebaran cenderung acak
dikarenakan biji mudah tersebar melaluiperantara air, angin, binatang dan manusia.
Pada dataran tinggi, jenis tumbuhan akanditemukan lebih banyak jenis tetapi jumlahindividu yang ditemukan sedikit. Namunsebaliknya, di dataran rendah jenis tumbuhanyang ditemukan akan sedikit tetapi jumlahindividunya akan banyak ditemukan. Jika padasuatu daerah menjauhi garis pantai maka akansemakin bertambah dan beragam jenistumbuhan yang mampu tumbuh danberkembang pada daerah tersebut (Lovelles,1989).
Pada Tabel 3 dapat diketahui bahwa nilaipersentase famili gulma dominan yang palingbesar pada kebun kopi Arabika di NagariBalingka, Kecamatan Ampek Koto, KabupatenAgam adalah famili Asteraceae (35,196%).Famili Asteraceae ini memiliki nilai persentasepaling besar karena pada penelitian ini hampirditemukan pada setiap plot pengamatan.Hendrival, Wirda, dan Azis (2014) jugamendapatkan hasil bahwa famili Asteraceaejuga nerupakan famili yang dominan ditemukanpada pertanaman kedelai. Menurut Johnstondan Gillman (1995) menyatakan bahwa suatufamili dikatakan dominan pada suatu kawasanyaitu jika memiliki persentase >20% dari totalindividu dan persentase famili co-dominanyaitu 10% - 20%. Sedangkan famili yang co-

Zuhri Syam, Chairul dan Indah Prafitri Yusa 322
dominan diantaranya yaitu Caryophyllaceaedengan persentase 11,978%.
Asteraceae merupakan famili dengankeanekaragaman jenis yang cukup tinggi dandominan. Jenis-jenis tumbuhan familiAsteraceae yang dijumpai pada penelitian initerletak pada ketinggian 1325 mdpl. Hal inisesuai dengan Lawrence (1955) yangmenyatakan bahwa tumbuhan anggota darifamili Asteraceae dapat ditemui padaketinggian 0-1300 mdpl. Oleh karena itu, padadaerah Bancah, Nagari balingka, KecamatanAmpek Koto, Kabupaten Agam ini, jenis-jenistumbuhan famili Asteraceae yang tumbuh.
Famili Asteraceae termasuk kedalam gulmatahunan yang banyak tersebar. Jenis tumbuhanfamii Asteraceae ini merupakan tumbuhan yangtumbuh pada tanah yang telah diolah, ditempat-tempat terbuka, di pinggir-pinggir jalan,di timbunan sampah atau ladang yang telahditinggalkan. Gulma famili ini tergolongkedalam gulma yang ganas karena ituseringkali populasinya lebih dominandibanding tanaman liar lainnya dalam suatulahan (Sukamto, 2007).
KESIMPULAN
Dari hasil diatas dapat diambil beberapakesimpulan sebagai berikut :1. Komposisi gulma pada tanaman kopi
arabika terdiri dari 11 famili, 21 genus dan25 spesies dengan jumlah keseluruhan yaitu3114 individu. Terdiri dari golongan teki-tekian 3 spesies, golongan rumput-rumputan6 spesies, golongan berdaun lebar 13 spesiesdan golongan pakis-pakisan 3 spesies.
2. Gulma yang memiliki nilai penting danSDR (Summed Dominance Ratio) tertinggipada tanaman kopi arabika adalah Borrerialaevis (68,426%) dan (22,809%). Indekskeanekaragaman jenis gulma pada tanamankopi arabika adalah kategori sedang yaituH’= 2,04.
DAFTAR PUSTAKA
Amperawati, T. dan T.M. Basuki. 1999.Prosiding. Seminar Hasil-Hasil PenelitianBadan Penelitian Kehutanan PematangSiantar. Parapat. Hal.88-97.
Brower, J., J. Zar and C. Von Ende. 1990. Fieldand laboratory methods for generalecology. 3rd ed. Wm. C. Brown Publishers,Dubuque: xi + 237 hlm.
Dinas Perkebunan Propinsi Sumatera Barat.2011. Data Statistik Kabupaten Agam.Padang.
Ewusie, J.Y. 1990. Pengantar Ekologi Tropika.Diterjemahkan oleh U. Tanuwijaya. ITBPress. Bandung.
Departemen Pertanian, 2012. Produksi KopiMenurut Propinsi di Indonesia, 2008-2012. www.deptan.go.id. Diakses padatanggal 13 November 2013.
Hendrival, Z. Wirda dan A. Azis. 2014. PeriodeKritis Tanaman Kedelai TerhadapPersaingan Gulma. J. Floratek 9: 6 – 1.
Indriana, R. 2009. Keanekaragaman JenisTumbuhan Pada Area Bantaran KaliPembuangan di Kecamatan KarangTengah Kabupaten Demak. Skripsi IKIPPGRI. Semarang.
Johnston and Gillman. 1995. Tree PopulationStudy in ow Diversity ForestGuyana.i.Floristc Composition and StandStructure. Biodiversity and Conversation4;339-362.
Lawrence, G.H.M. 1955. An Introduction toPlant Taxonomy. Pp.15-17The MacmillanCompany. New York.
Lovelles, A.R. 1989. Prinsip-Prinsip BiologiTumbuhan Untuk Daerah Tropik.Gramedia. Jakarta.
Magurran, A.E. 2004. Measuring BiologicalDiversity. Blackwell Science Ltd.Australia.
Mangoensoekarjo, S. 1982. Ilmu Gulma danCara Pengendaliannya. LatihanPembekalan Keterampilan Teknik PetugasLapangan Proyek Terpadu PerkebunanLPP. Yogjakarta.
Moenandir, J. 1990. Pengantar Ilmu Gulma.PT. Raja Grafindo Persada. Jakarta.
Moenandir, J. 1993. Ilmu Gulma Dalam SistemPertanian. PT. Grafindo Persada. Jakarta.
Najiyati, S dan Danarti. 2004. Kopi Budidayadan Penanganan Lepas Panen. EdisiRevisi. Penebar Swadaya. Jakarta.

Zuhri Syam, Chairul dan Indah Prafitri Yusa 323
Odum, E.P. 1996. Dasar-Dasar Ekologi.Universitas Gadjah Mada. Yogyakarta.
Okunade, A.L. 2002. Ageratum conyzoides L.Asteraceae. Fitoterapia 73: 1-16.
Prastowo, B., E. Karmawati, Rubijo, Siswanto,C. Indrawanto dan S.J. Munarso. 2010.Budidaya dan Pasca Panen Kopi. PusatPenelitian dan Pengembangan Perkebunan.Bogor.
Rahardjo, P. 2012. Panduan Budidaya danPengelolaan Kopi Arabika dan Robusta.Penebar Swadaya. Jakarta.
Rambe, T.D, L.Pane, P.Sudharto dan Caliman.2010. Pengelolaan Gulma PadaPerkebunan Kelapa Sawit di PT. SmartTbk: Jakarta.
Reader dan Buck. 2000. Pertumbuhan GulmaPada Kondisi Lingkungan. PT. GramediaPres. Jakarta.
Robin, S. 2011. Dispersal and GeneticVariability of Sonchus oleraceus L. inRelation to its resistance to Als. Inhibitingherbicides. The University of Adelaide,Waite Research Institute. South Australia.
Ruswanto, I. 2011. Organisme PenggangguTumbuhan (Identifikasi dan AnalisisVegetasi Gulma). Agroekoteknologi.Fakultas Pertanian. Universitas JenderalSoedirman. Purwokerto.
Sastroutomo, S.S. 1990. Ekologi Gulma. PT.Gramedia Pustaka Utama. Jakarta.
Soegianto, A. 1994. Ekologi Kuatitatif: MetodeAnalisis Populasi dan Komunitas. UsahaNasional. Surabaya.
Suhardi, Sarbino dan Astina. 2011. StrukturKomunitas Gulma Pada PertanamanJagung (Zea mays L.) di Desa Suka MajuKecamatan Sungai Betung KabupatenBengkayang. Fakultas Pertanian,Universitas Tanjung Pura. Pontianak.
Sukamto. 2007. Babadotan (Ageratumconyzoides) Tanaman Multi Fungsi YangMenjadi Inang Potensial Virus Tanaman
http://www.ddbj.nig.ac.jp/. WartaPuslitbangbun.13 (3):2.
Sukman, Y. dan Yakup. 1995. Gulma danTeknik Pengendaliannya. Ed. 1, cet2. PT.Raya Grafindo. Jakarta.
Susilo, E. 2013. Tanggap Pertumbuhan AwalJarak Pagar (Jatropha curcas L.)Terhadap Bokkasi Gulma Gletang (Tridaxprocumbens) yang Diperkaya Kapur PadaTanah Ultisol. Agrovigor Vol 6 (1) : 63-72.
Syafei, E.S. 1990. Pengantar EkologiTumbuhan. Institusi Teknologi Bandung.Bandung.
Tjitrosoedirdjo, S., H. Utomo, dan J.Wiroatmodjo. 1984. Pengelolaan Gulmadi Perkebunan. PT. Gramedia. Jakarta.
Tjitrosoepomo, G., Soerjani, M danKostermans. 1987. Weeds of Rice inIndonesia. Balai Pustaka. Jakarta.
Tjokrowardojo, A.S. dan E. Djauhariya. 2013.Gulma Pada Budidaya Tanaman Jahe.Balai Penelitian Tanaman Obat danAromatik. Bogor.
Utomo, I.H.D,, Nuswandari, dan A,P Lontoh.1989. Periode Kritis Kacang HijauTerhadap Kompetisi Gulma. Prosiding keVII Himpunan Ilmu Gulma. Bogor.
Widiyanti, T. 2013. Kondisi Kebun SumberBenih Kopi (Coffea sp) di Kebun KalisatJampit Bondowoso. Balai BesarPerbenihan dan Proteksi TanamanPerkebun2an. Surabaya.
Related Documents











