Prolonged exposure of dendritic cells to maturation stimuli favors the induction of type-2 cytotoxic T lymphocytes Andrea Boni*, Giandomenica Iezzi**, Elena Degl'Innocenti, Matteo Grioni, Elena Jachetti, Annalisa Camporeale and Matteo Bellone Cancer Immunotherapy and Gene Therapy Program, Istituto Scientifico H San Raffaele, Milan, Italy Dendritic cell (DC) maturation influences the priming and polarization of T lymphocytes. We recently found that early activated DC (i.e. DC exposed to pro- maturation stimuli for 8 h) were more prone to prime in vivo a type-1 cytotoxic T cell (Tc1) response than DC exposed to pro-maturation stimuli for 48 h (48h-DC). We investigated whether 48h-DC, conversely, allowed the induction of Tc2 cells. Antigen- pulsed mouse bone-marrow-derived DC at any maturation stage, in the presence of exogenous IL-12, skewed in vitro naive CD8 + T cells towards Tc1 cells, but 48h-DC most potently, in the presence of exogenous IL-4, favored the induction of Tc2 cells. In vivo, full maturation of DC promoted expansion of Tc2 and fall of Tc1 cells. Tc2 cells maintained a high cytolytic activity and produced significant amounts of IL-4, IL-5, IL-10 and TGF-b. Our results indicate that polarization of naive CD8 + T cells to Tc2 cells is dependent on the amount of time DC have been exposed to maturation stimuli, and might be favored in late and/or chronic phases of an immune response. Supporting information for this article is available at http://www.wiley-vch.de/contents/jc_2040/2006/35597_s.pdf Introduction Upon antigenic stimulation, T lymphocytes proliferate and acquire a variety of effector functions. Indeed, activation of naive CD4 + T cells in the presence of IL-12 results in the development of Th1 cells, which mainly secrete interferon (IFN)-c, interleukin (IL)-2, tumor necrosis factor (TNF)-a and lymphotoxin, and are commonly associated with a cell-mediated immune response. Conversely, the presence of IL-4 skews the Th response towards Th2 cells producing IL-4, IL-10 and IL- 13, relevant cytokines against helminthiasis and allergic diseases [1]. More recently, a third subset of CD4 + T cells has been identified, which express CD25, produce IL-10 and TGF-b, and display regulatory functions [2]. The polarization of CD8 + CTL has been less characterized [3]. IL-12 also appears to be critical for the generation of Tc1 cells, characterized by a high production level of IFN-c and cytolytic activity, CC chemokine receptor (CCR)5 expression and the ability to migrate to the inflamed tissues where they are Correspondence: Matteo Bellone, Cancer Immunotherapy and Gene Therapy Program, Dibit, Istituto Scientifico H San Raffaele, Via Olgettina 58, 20132 Milan, Italy Fax: +39-02-2643-4786 e-mail: [email protected] Received 13/10/05 Revised 28/6/06 Accepted 17/10/06 [DOI 10.1002/eji.200535597] Key words: Cellular differentiation CTL Dendritic cells Rodent Th1/Th2 cells Abbreviations: 0h-DC: DC not exposed to maturation stimuli 8h-DC: DC exposed to maturation stimuli for 8 h 48h-DC: DC exposed to maturation stimuli for 48 h Tc1 cells: CTL of type 1 Tc2 cells: CTL of type 2 Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3157 f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu * Current address: NIH/NCI Building 10, Clinical Research Center, 9000 Rockville Pike, Bethesda, MD 20892-1201, USA ** Current address: Molecular Biomedicine, Swiss Federal Institute of Technology, Wagistrasse 25/27, CH-8952 Schlieren, Switzerland

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Prolonged exposure of dendritic cells to maturationstimuli favors the induction of type-2 cytotoxicT lymphocytes
Andrea Boni*, Giandomenica Iezzi**, Elena Degl'Innocenti,Matteo Grioni, Elena Jachetti, Annalisa Camporeale and Matteo Bellone
Cancer Immunotherapy and Gene Therapy Program, Istituto Scientifico H San Raffaele,Milan, Italy
Dendritic cell (DC) maturation influences the priming and polarization ofT lymphocytes. We recently found that early activated DC (i.e. DC exposed to pro-maturation stimuli for 8 h) were more prone to prime in vivo a type-1 cytotoxic T cell(Tc1) response than DC exposed to pro-maturation stimuli for 48 h (48h-DC). Weinvestigated whether 48h-DC, conversely, allowed the induction of Tc2 cells. Antigen-pulsed mouse bone-marrow-derived DC at any maturation stage, in the presence ofexogenous IL-12, skewed in vitro naive CD8+ Tcells towards Tc1 cells, but 48h-DC mostpotently, in the presence of exogenous IL-4, favored the induction of Tc2 cells. In vivo,full maturation of DC promoted expansion of Tc2 and fall of Tc1 cells. Tc2 cellsmaintained a high cytolytic activity and produced significant amounts of IL-4, IL-5,IL-10 and TGF-b. Our results indicate that polarization of naive CD8+ Tcells toTc2 cellsis dependent on the amount of time DC have been exposed to maturation stimuli, andmight be favored in late and/or chronic phases of an immune response.
Supporting information for this article is available athttp://www.wiley-vch.de/contents/jc_2040/2006/35597_s.pdf
Introduction
Upon antigenic stimulation, T lymphocytes proliferateand acquire a variety of effector functions. Indeed,activation of naive CD4+ T cells in the presence of IL-12results in the development of Th1 cells, which mainlysecrete interferon (IFN)-c, interleukin (IL)-2, tumornecrosis factor (TNF)-a and lymphotoxin, and arecommonly associated with a cell-mediated immune
response. Conversely, the presence of IL-4 skews the Thresponse towards Th2 cells producing IL-4, IL-10 and IL-13, relevant cytokines against helminthiasis and allergicdiseases [1]. More recently, a third subset of CD4+
Tcells has been identified, which express CD25, produceIL-10 and TGF-b, and display regulatory functions [2].
The polarization of CD8+ CTL has been lesscharacterized [3]. IL-12 also appears to be critical forthe generation of Tc1 cells, characterized by a highproduction level of IFN-c and cytolytic activity,CC chemokine receptor (CCR)5 expression and theability to migrate to the inflamed tissues where they areCorrespondence: Matteo Bellone, Cancer Immunotherapy and
Gene Therapy Program, Dibit, Istituto Scientifico H SanRaffaele, Via Olgettina 58, 20132 Milan, ItalyFax: +39-02-2643-4786e-mail: [email protected]
Received 13/10/05Revised 28/6/06
Accepted 17/10/06
[DOI 10.1002/eji.200535597]
Key words:Cellular
differentiation �CTL � Dendriticcells � Rodent �Th1/Th2 cells
Abbreviations: 0h-DC: DC not exposed to maturation stimuli �8h-DC: DC exposed to maturation stimuli for 8 h � 48h-DC: DCexposed to maturation stimuli for 48 h � Tc1 cells: CTL oftype 1 � Tc2 cells: CTL of type 2
Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3157
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
* Current address: NIH/NCI Building 10, Clinical ResearchCenter, 9000 Rockville Pike, Bethesda, MD 20892-1201, USA
** Current address: Molecular Biomedicine, Swiss FederalInstitute of Technology, Wagistrasse 25/27,CH-8952 Schlieren, Switzerland

highly efficient in eliminating viral infections and tumorcells. IL-4 [4–6] and TGF-b [7, 8] are critical for thepolarization of Tc2 cells, mainly characterized by IL-4,IL-5, IL-10 and TGF-b production [4, 6, 9, 10], CCR4expression, and a reduced capacity to migrate to theinflamed tissues [11].
The role of Tc2 cells in the immune response isdebatable. Tc2 cells have been found in vivo in patientsaffected by allergic [12], autoimmune [13], viral [10,14], neurologic [15] and neoplastic diseases [16], and inmany cases their presence was associated with diseaseseverity and progression [13–16]. In mouse tumormodels, IL-4-transduced tumor cells elicited potent anti-tumor activities [17] and induced Tc2 cells able to curelung metastases [5, 17]. IL-4 produced by CTL wasrequired for the generation of protective tumor-asso-ciated CD8+ T cells [5, 18]. In other models, Tc2 cellswere far less effective in curing subcutaneous tumors[19, 20] and lung metastases [21]. Finally, CD8+ T cellsubpopulations exhibiting regulatory function have alsobeen described [14, 22].
Dendritic cells (DC), professional antigen presentingcells (APC) with a unique ability in priming T cellresponses [23], play a key role in T cell polarization. Inparticular, it has been reported that subsets of DC,derived from distinct precursors, induce differentiallypolarized CD4+ and CD8+ T cells [24]. More recently, ithas been reported that even DC comprised within thesame subset can differentially drive Th1 or Th2expansion in vitro, depending on their activation stageand antigen load [25, 26]. Of relevance, immature DC[27] and plasmacytoid DC [28] induced IL-10-secretingregulatory CD8+ T cells.
We recently observed that antigen-loaded bone-marrow-derived DC exposed to a single pro-maturationstimulus (i.e. LPS, CD40L or TNF) for 48 h (48h-DC)were less efficient than recently activated DC (i.e. DCexposed to maturation stimuli for 8 h; 8h-DC) ininducing a protective and therapeutic immune responseagainst B16 melanoma. This inefficiency correlated withreduced IL-12 production by 48h-DC and a lower IFN-cproduction by the primed CTL [29]. We hypothesizedthat, as has been reported for human CD4+ T cells [26],48h-DC might favor the induction of Tc2 cells. Herein,we show that the activation stage of DC directly impactson the polarization of naive CD8+ T cells toward Tc2cells.
Results
The polarization of naive CD8+ T lymphocytes toTc2 cells in vitro is favored by 48h-DC and IL-4
In order to assess the capacity of DC to polarize naiveCD8+ T cells towards Tc1 or Tc2 at different activationstages, DC induced from bone marrow precursors [29,30] were exposed to either LPS or TNF-a for differenttime periods. As previously reported [29], untreated DC(not exposed to maturation stimuli; 0h-DC) showed aheterogeneous expression of MHC class I, MHC class II,CD40, CD80 and CD86 (Fig. 1 and data not shown),suggesting that 0h-DC comprised a mixed population ofimmature and mature DC. This is a well-expected[31–33] and unavoidable phenomenon, which has beenattributed to the in vitro manipulation of DC (i.e.pipetting and transfer of cultured DC into wells).Nevertheless, the expression of the indicated markersincreased with exposure time to the pro-maturationstimulus, showing no substantial differences betweenLPS- and TNF-a-treated DC (Fig. 1A, B).
The 0h-, 8 h- and 48h-DC were loaded with thepeptide SIINFEKL [34] at a concentration (100 nM) thatin preliminary experiments (peptide range 1–1000 nM)favored the induction of the highest percentage of Tc2cells, and cultured together with naive OT-1 cells, ahomogeneous population of TCR-transgenic CD8+ cells[35], in the presence or absence of optimal amounts ofIL-12 or IL-4. Seven days later, cells were collected andassessed for cytokine production by flow cytometry.Similarly to what we found in vivo [29], both LPS- andTNF-a-treated 8h-DC, in the absence of added cytokines,induced efficient Tc1 differentiation, whereas 0 h- and48h-DC failed to induce Tc1 development (data notshown).
The ability to skew the OT-1 immune responsetowards Tc1 cells was associated with IL-12 productionby the DC subpopulations, with 8h-DC being far higherproducers of IL-12 than 0 h- or 48h-DC [29]. However,48h-DC did not appear to be exhausted, because in thatDC population a consistent percentage of CD11c+ cellswas still IL-12+ at flow cytometry after intracellularstaining [29]. Addition of exogenous IL-12 rescued thecapacity of 0 h- and 48h-DC to promote Tc1 polariza-tion, leading to the generation of a very large fraction ofIFN-c-producing Tcells (approximately 75%; Fig. 1C, D)under any condition.
Conversely, exogenous IL-4 promoted Tc2 polariza-tion primarily when 48h-DC were used as APC,irrespective of the pro-maturation stimulus used(Fig. 1C, D). Indeed, statistically significant numbersof IL-4-producing (10.3�6.1%; n=6; 0h-DC vs. 48h-DC:p<0.02;8h-DC vs.48h-DC:p<0.04) andespecially IL-10-producing CD8+ T cells (11.5�4.6%; n=5; 0h-DC vs.
Andrea Boni et al. Eur. J. Immunol. 2006. 36: 3157–31663158
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

48h-DC: p<0.007; 8h-DC vs. 48h-DC: p <0.03) werefoundonly in co-cultures ofOT-1 cells and48h-DC.WhenIL-13 was used, a prototypic type-2 cytokine, bona fideTc2 cells accounted for >40% of the cultured cells(Fig.1E). Both0 h- and8h-DC inducedonlymarginal Tc2development (Fig. 1C, D). In the presence of exogenousIL-4 together with IL-12, the percentage of IFN-c+ cellswas reduced, and no Tc2 switch was obtained (seeSupplementary Fig. 1). Contaminating Tc1 cells areinvariably found in Tc2 preparations [6, 7, 11, 19]. In allour test conditions, approximately 25% of IFN-c-producing cells were present in the Tc2 cultures(Fig. 1C, D), whereas CD8+ T cells producing bothIFN-c and a Tc2 cytokine were <5% (data not shown).
The role of TCR/MHC interactions in T cell polariza-tion is well recognized [36]. Hence, we assessed thecapacity of DC at various activation stages to induce
polarization of a transgenic T cell population with adifferent specificity, i.e. cells from TCR7mice expressinga TCR specific for the epitope 33–41 of lymphocyticchoriomeningitis virus (LCMV) glycoprotein 1 (gp33–41)[37, 38]. In this set of experiments we also tested CpG aspro-maturation stimulus (see Supplementary Fig. 2).
Similarly to the results obtained with OT-1 cells, inthe presence of IL-4, 48h-DC efficiently drove Tc2
Figure 2. Polarization of TCR7 cells. TCR7 T cell blasts, obtainedby in vitro co-cultures with gp33–41-loaded DC in the absence orin the presence of exogenous IL-12 and IL-4, were analyzed forcytokine production by intracellular staining. Antigen-pulsedand unpulsed RMA cells were used as stimulators. A gate wasperformed on live CD8+ cells. The panels depict IFN-c+ (y-axes)and IL-4+ (x-axes) cells within the gated population. Numbersin the panels refer to positive cells. Data shown arerepresentative of at least two independent experiments.
·
Figure 1. The polarization of CD8+ T lymphocytes to Tc2 cells invitro is favored by 48h-DC and IL-4. Bone-marrow-derived DC,after exposure to LPS (A, C) or TNF-a (B, D) for the last 8 or 48 hof culture, were double-stained with mAb against CD11c andthe indicated specificity, and analyzed by flow cytometry. Agate was performed on live CD11c+ cells. (A, B) Histogramsillustrate the expression of specific markers on CD11+ cells(black profiles). Isotype controls are represented by openprofiles. In the upper right corner of each panel theMFI of eachcurve is reported. (C, D) OT-1 T cell blasts, obtained by in vitroco-cultures with SIINFEKL-loaded DC, were analyzed forcytokine production by intracellular staining. Antigen-pulsedand unpulsed RMA cells were used as stimulators. A gate wasperformed on live CD8+ cells. Percentages of double-positiveCD8 and IFN-c-, IL-4- or IL-10-producing cells are reported.Background staining (i.e. blasts in the presence of unpulsedRMA) for all cytokineswas<0.1%. (E) OT-1 blasts, obtained from48h-DC co-cultures, were analyzed for IL-4 and IL-13 produc-tion as described above. The panel depicts IL-13+ (y-axes) andIL-4+ (x-axes) cellswithin the gated population. Numbers in thepanels refer to positive cells. In each panel, representativeexamples of at least three independent experiments areshown.
Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3159
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

polarization of TCR7 cells, leading to the development ofup to 10�0.4 % of IL-4 single producers and 18�0.9% ofIFN-c/IL-4 double-producing cells (Fig. 2, lower rightpanel). Conversely, 0 h- and 8h-DC induced onlymarginal frequencies of Tc2 cells. (Fig. 2, lower panels).In the presence of IL-12, all DC populations drove astrong Tc1 polarization (Fig. 2, middle panels). At oddswith OT-1 cells, in the absence of polarizing cytokines,TCR7 cells acquired IFN-c production capacity whenprimed also by 0 h- or 48h-DC (Fig. 2, upper panels).This may be related to the fact that Tc1 cell induction inTCR7 mice appears to be less dependent on IL-12 [38].However, 8 h- and 48h-DC led to a much greaterexpansion of primed cells than 0h-DC (approximately8.2-fold vs. 1.9-fold, data not shown).
Phenotype and functional characterizationof Tc1 and Tc2 populations
Irrespective of the polarizing cytokine added to theculture and the time the DC were exposed to thematuration stimulus, activated DC induced a rapid (peakat day 5) and comparable*8-fold expansion of Tc1 andTc2 cells (data not shown). To directly measure thenumber of cell divisions required to display the Tc1 andTc2 phenotype, naive TCR7 cells were stained withcarboxyfluorescein diacetate succinimidyl ester (CFSE)and co-cultured with gp33–41-pulsed 48h-DC and thepolarizing cytokines for 3 days. As shown in Fig. 3, IFN-cproduction was readily acquired by activated cells evenbefore completing the first division. Conversely, IL-4appeared to be produced only by cells that hadcompleted at least four cycles, indicating that more celldivisions are required in order to acquire a Tc2phenotype.
Flow cytometry analysis (Fig. 4) did not detect anysubstantial difference between the two populations of
Tc1 and Tc2 cells induced by either 8 h- or 48h-DC. Onthe other hand, a few remarkable differences were foundbetween Tc1 and Tc2 cells. In all populations from the7-day co-cultures, �90% were CD8+ cells, all of whichwere, in the case of OT-1 cells, Va–2+Vb5+(data notshown). Both Tc1 and Tc2 cells similarly exhibited aphenotypic profile typical of activated effector cells,including high levels of CD44 (% of positive cells withinthe populations induced by 48h-DC: Tc1: 96�2.8 andTc2: 95�3.8; n=4; not shown), CD25 and CD28, andlow levels of CD62L expression. However, as previouslydescribed by others [21], a difference was foundbetween Tc1 and Tc2 cells in the expression of CD25and CD62L. Indeed, Tc2 cells homogeneously expressedCD25 at lower intensity than Tc1 cells, and a higherpercentage of Tc2 cells down-regulated CD62L (% ofpositive cells within the populations induced by 48h-DC:Tc1: 44�14 and Tc2: 20�7; p<0.03; n=6).
OX40 has been described as a co-stimulatorymolecule involved in Th2 polarization [39]. Its expres-sion on CTL was marginal and lower than that found inTh2 cells (Fig. 4). However, OX40 appeared slightly butconsistently up-regulated in Tc2 cells (% of positive cellswithin the populations induced by 48h-DC: Tc1: 3.2�1.6and Tc2: 23�9.4; p<0.005; n=4; Fig 3).
Most, if not all, Tc1 and Tc2 resulted negative forCD137 (% of positive cells within the populationsinduced by 48h-DC: Tc1: 0.9�0.2 and Tc2: 1�0.8; n=3;data not shown) and CD154 (CD40L; Fig. 4). At least forCD154, this expected result [40] may be due to theabsence of brefeldin A in the culture medium [41].
When tested in standard cytotoxicity assays, bothOT-1 Tc1 and Tc2 populations efficiently killed bothRMA cells expressing OVA [30] and RMA cells loadedwith the SIINFEKL peptide, but not unpulsed RMA cells(Fig. 5). We also found no difference in cytolytic activitybetween Tc2 cells obtained by 8 h- and 48h-DC co-cultures (Fig. 5). Finally, a peptide titration assayconfirmed a very similar sensitivity for SIINFEKL by Tc1and Tc2 cells (Fig. 5E), strongly supporting theirequivalent lytic potential.
We also tested by ELISA the cytokines produced by48h-DC-induced Tc1 and Tc2 cells upon antigen-specificstimulation. As expected, Tc1 cells produced highamounts of IFN-c (13�1.5 ng/mL; n=3), minimalamounts of IL-10 and no IL-5 or TGF-b (Fig. 5F).Conversely, Tc2 cells produced significant amounts ofIL-5, IL-10 and TGF-b (Fig. 5F), and approximately 1 logless IFN-c (1.8�0.2 ng/mL; n=3) than Tc1 cells.
The 48h-DC favor the polarization of naiveCD8+ T lymphocytes to Tc2 cells in vivo
In order to verify whether fully matured DC were able toskew an endogenous CD8+ Tcell response towards a Tc2
Figure 3. Cell divisions of Tc1 and Tc2 populations. Naive TCR7cells were stained with CFSE and co-cultured with gp33–41-loaded 48h-DC in the presence of IL-12 (left panel) or IL-4 (rightpanel). At day 3, cellswere analyzed by flow cytometry for CFSEexpression and cytokine production by intracellular staining.Dot plots illustrate the expression of diluted CFSE (x-axes) andcytokine production (y-axes). Data are representative of threeindependent experiments.
Andrea Boni et al. Eur. J. Immunol. 2006. 36: 3157–31663160
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

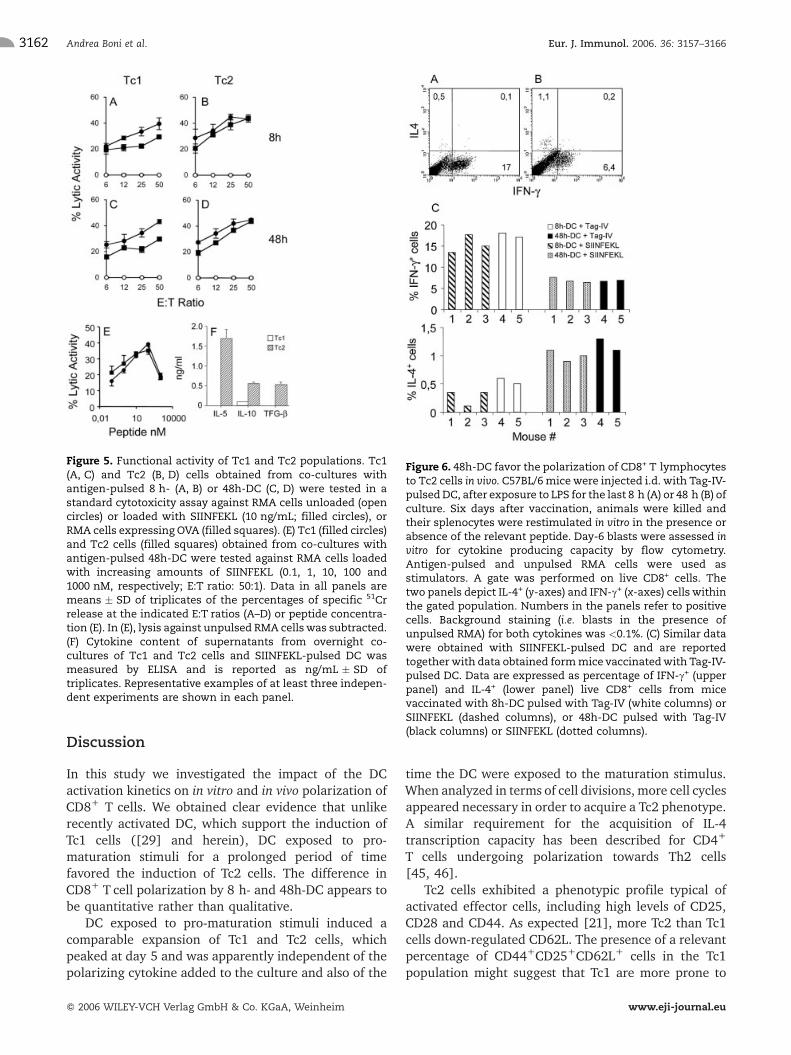
phenotype in vivo, C57BL/6 mice were immunized oncewith 8 h- or 48h-DC pulsed with either SIINFEKL or Tag-IV, the immunodominant epitope of the large T antigenof SV-40 [42]. On day 6, animals were killed and theirsplenocytes were cultured in vitro for an additional6 days in the presence of the relevant peptide. Finally,blasts were assessed for their cytokine releasingpotential by intracellular staining and flow cytometry.
As previously reported [43], Tag-IV-pulsed 8h-DCfavored the induction of Tc1 cells (Fig. 6A). In theabsence of an adoptively transferred TCR-transgenicpopulation, intracellular staining with IL-4 is generallypoor [44]; yet in those animals we found a population ofCD8+ cells producing IL-4+ (� 0.5%; Fig. 6A) which wasconsistently above the background (�0.1%; not shown).On the other hand, mice vaccinated with antigen-pulsed
48h-DC developed more than twice the amount of Tag-IV-specific and IL-4-producing bona fide Tc2 cells, andgenerated less than half of the Tag-IV-specific CTLproducing IFN-c found in mice vaccinated with antigen-pulsed 8h-DC (Fig. 6B). Similar data were obtained withSIINFEKL-pulsed DC. Indeed, when 48h-DC were usedto induce polarization of CTL, the percentage of CD8+
cells producing IFN-c dropped from 15.4�2.1% to6.9�0.6% (p<0.03; n=3). Conversely, IL-4+ cells rosefrom 0.3�0.1% to 1.0�0.1% (p<0.003; n=3). Resultsare reported together with data obtained from micevaccinated with Tag-IV-pulsed DC in Fig. 6C. Alltogether, these findings suggest that also in vivo 48h-DC have lost at least part of their Tc1-polarizingpotential and favor the induction of Tc2 cells.
Figure 4. Phenotype of Tc1 and Tc2 populations. Flow cytometry analysis of naive, Tc1 and Tc2 OT-1 cells induced by co-culturewith 8 h- and 48-DC pulsed with SIINFEKL. Cells were triple-stained with isotype controls or the indicated mAb. Histogramsillustrate the expression of OX40 (solid lines). Isotype controls are indicated by dashed lines. A gatewas performedon live CD8/Va2double-positive T cells. As positive control, activated Th2 cells were obtained fromDO11.10 splenocytes. The second column fromthe left reports dot plots of live CTL populations stained with CD8. Columns 3–5 from the left show dot plots on gated live CD8+
cells. In the upper quadrants of each panel the percentage of positive cells is reported. Data are representative of at least threeindependent experiments.
Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3161
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Discussion
In this study we investigated the impact of the DCactivation kinetics on in vitro and in vivo polarization ofCD8+ T cells. We obtained clear evidence that unlikerecently activated DC, which support the induction ofTc1 cells ([29] and herein), DC exposed to pro-maturation stimuli for a prolonged period of timefavored the induction of Tc2 cells. The difference inCD8+ T cell polarization by 8 h- and 48h-DC appears tobe quantitative rather than qualitative.
DC exposed to pro-maturation stimuli induced acomparable expansion of Tc1 and Tc2 cells, whichpeaked at day 5 and was apparently independent of thepolarizing cytokine added to the culture and also of the
time the DC were exposed to the maturation stimulus.When analyzed in terms of cell divisions, more cell cyclesappeared necessary in order to acquire a Tc2 phenotype.A similar requirement for the acquisition of IL-4transcription capacity has been described for CD4+
T cells undergoing polarization towards Th2 cells[45, 46].
Tc2 cells exhibited a phenotypic profile typical ofactivated effector cells, including high levels of CD25,CD28 and CD44. As expected [21], more Tc2 than Tc1cells down-regulated CD62L. The presence of a relevantpercentage of CD44+CD25+CD62L+ cells in the Tc1population might suggest that Tc1 are more prone to
Figure 6. 48h-DC favor the polarization of CD8+ T lymphocytesto Tc2 cells in vivo. C57BL/6mice were injected i.d. with Tag-IV-pulsedDC, after exposure to LPS for the last 8 h (A) or 48 h (B) ofculture. Six days after vaccination, animals were killed andtheir splenocytes were restimulated in vitro in the presence orabsence of the relevant peptide. Day-6 blasts were assessed invitro for cytokine producing capacity by flow cytometry.Antigen-pulsed and unpulsed RMA cells were used asstimulators. A gate was performed on live CD8+ cells. Thetwo panels depict IL-4+ (y-axes) and IFN-c+ (x-axes) cells withinthe gated population. Numbers in the panels refer to positivecells. Background staining (i.e. blasts in the presence ofunpulsed RMA) for both cytokines was <0.1%. (C) Similar datawere obtained with SIINFEKL-pulsed DC and are reportedtogether with data obtained formmice vaccinatedwith Tag-IV-pulsed DC. Data are expressed as percentage of IFN-c+ (upperpanel) and IL-4+ (lower panel) live CD8+ cells from micevaccinated with 8h-DC pulsed with Tag-IV (white columns) orSIINFEKL (dashed columns), or 48h-DC pulsed with Tag-IV(black columns) or SIINFEKL (dotted columns).
Figure 5. Functional activity of Tc1 and Tc2 populations. Tc1(A, C) and Tc2 (B, D) cells obtained from co-cultures withantigen-pulsed 8 h- (A, B) or 48h-DC (C, D) were tested in astandard cytotoxicity assay against RMA cells unloaded (opencircles) or loaded with SIINFEKL (10 ng/mL; filled circles), orRMA cells expressing OVA (filled squares). (E) Tc1 (filled circles)and Tc2 cells (filled squares) obtained from co-cultures withantigen-pulsed 48h-DC were tested against RMA cells loadedwith increasing amounts of SIINFEKL (0.1, 1, 10, 100 and1000 nM, respectively; E:T ratio: 50:1). Data in all panels aremeans � SD of triplicates of the percentages of specific 51Crrelease at the indicated E:T ratios (A–D) or peptide concentra-tion (E). In (E), lysis against unpulsed RMA cellswas subtracted.(F) Cytokine content of supernatants from overnight co-cultures of Tc1 and Tc2 cells and SIINFEKL-pulsed DC wasmeasured by ELISA and is reported as ng/mL � SD oftriplicates. Representative examples of at least three indepen-dent experiments are shown in each panel.
Andrea Boni et al. Eur. J. Immunol. 2006. 36: 3157–31663162
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

migration to the lymphoid organs than Tc2 and generatea central memory population [47, 48], although Tc2 cellstoomay efficiently migrate to lymph nodes [19] and giverise to central memory cells [49].
Conversely, OX40 was slightly but consistently up-regulated in Tc2 cells. Its ligand OX40L is marginallyexpressed on immature and recently activated DC,whereas it is strongly up-regulated on 48h-DC [50].Hence, it might be possible that OX40-OX40L interactionis actively involved in the generation and maintenanceof the Tc2 phenotype.
In presence of the relevant antigen, well-proliferatedTc2 cells produced IL-10, IL-4, IL-5 and TGF-b, andefficiently killed specific targets. The cytolytic activity ofTc2 cells is under discussion. Whereas some investiga-tors have reported on non-cytolytic Tc2 cells [51], othershave shown similar [6, 19, 52] and even slightly highercytotoxic potential for Tc2 cells [11]. The differentresults may be, at least in part, due to the purity of thedifferent Tc2 cell preparations. The Tc1 cells (approxi-mately 25%) contaminating our Tc2 population canaccount for part but not all of the cytolytic activityattributed to Tc2 cells. Hence, although a quantitativeevaluation cannot be performed due to the contaminat-ing Tc1 population, our Tc2 populations appear similarlycytolytic to Tc1 cells.
Since it has been described that IL-10-producingCD8+ T cells may have a regulatory function on theimmune response [14, 27, 28], and that regulatory CD8+
T cells may act directly on APC [53, 54], we verified theeffects of Tc2 cells on DC during a secondary immuneresponse. As reported elsewhere [55], Tc2, and not Tc1cells, modified, through a cell-to-cell contact mechan-ism, the APC function of DC. Indeed, Tc2-conditionedDC displayed a reduced expression of MHC class II andco-stimulatory molecules, produced IL-10 instead ofIL-12, and, in the absence of exogenous polarizingcytokines, favored the differentiation of both naiveCD4+ and CD8+ T cells towards type-2 cells, thereforesuggesting a modulating activity for Tc2 cells.
The data reported herein, together with those fromour and other laboratories [26, 29, 55], allow us tohypothesize the following scenario. In the early phasesof an immune response, tissue-resident DC are exposedto strong pro-inflammatory stimuli (e.g. LPS, bacterialCpG DNA and double-stranded viral DNA) and largeamounts of antigen. As a consequence, maturing DCrapidly up-regulate chemokine receptors, which favortheir migration to the draining lymph nodes at a time atwhich IL-12 is maximally produced [56]. Here, earlyactivated DC should rapidly induce CD4+ T cellactivation [57], possibly Th1 polarization [46], andeven faster priming of potent CTL [58, 59] with Tc1phenotype [29]. In the following phase of the inflam-matory reaction, DCmay be trapped within the inflamed
tissue, where scavenger cells are fully committed toeliminating the inflammation/antigen source [60]. Inthe later phases of the inflammatory reaction, DCexposed to maturation stimuli for a prolonged period oftime, although equally efficient to early activated DC inmigrating to the draining lymph nodes [29], have lostmost of their potential to induce powerful type-1responses, and bear reduced amounts of antigen. Thisscenario should favor the induction of Th2 and Tc2 cells.
Tc2 cells migrate less efficiently than Tc1 cells [11]and should, therefore, be less effective in the tissues. Ashas been described for Th2 cells [61], Tc2 cells mightalter the APC function of lymph-node-resident DC andfavor, together with IL-4 produced by both Th2 and Tc2cells, the induction of a self-sustained regulatory loop.The narrow time window for efficient activation of anantigen-specific immune response should avoid the riskof over-activation of the immune response, and/oractivation of bystander and possibly auto-reactiveT cells. The self-sustained loop induced in the laterphases of the inflammatory reaction should favor thephysiological shutdown of the immune response with-out excessive damage by Tc1 cells.
Conversely, in the presence of a chronic stimulationof the immune system, the regulatory loop describedabove may dominate, and explain at least in part the Tc2switch found in pathologies such as allergy [12],psoriasis [13], type-1 human immunodeficiency virusand chronic hepatitis C virus infections [10, 14], multi-ple sclerosis [15] and lung carcinoma [16]. A deepermolecular definition of the mechanisms underlyingpathological skews of the immune response would be ofhelp in developing more potent immunotherapeuticapproaches for those diseases.
Materials and methods
Mice and reagents
Female C57BL/6 (H-2b) and BALB/c (H-2d)mice, 8–10 wk old,were purchased from Charles River (Calco, Italy). OT-1 (H-2b)[35] and DO11.10 (H-2d) [62] mice were bred to thehomozygous condition. OT-1 mice express a Va2Vb5 Tg TCRspecific for OVA257–264 (SIINFEKL). DO11.10 mice express a Tgab TCR specific for OVA323–339 presented in the context of I-Ad
molecules. LCMV gp33 mice, which express a TCR specific forgp33–41 in association with H-2Db [37], were backcrossed ontoC57BL/6 background over six generations to obtain TCR7mice[38]. All mice were housed in a specific pathogen-free animalfacility, and treated in accordance with the European UnionGuidelines together with the approval of the InstitutionalEthics Committee. Unless specified, all chemical reagents werefrom Sigma-Aldrich, and mAb were from BD PharMingen (SanDiego, CA).
Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3163
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Generation and functional characterization of Tc1 andTc2 cells from OT-1 and TCR7 mice
Single-cell suspensions (1�106 cells/mL) of spleen and lymphnodes cells from OT-1 or TCR7 mice were seeded into 6-wellplates together with different populations of DC (ratio 5:1; seebelow) loaded with 100 nM SIINFEKL peptide [34] or 10 nMgp33–41 (Research Genetics, Huntsville, AL) in RPMI 1640supplemented with penicillin/streptomycin, 10 mM HEPES,10 mMNa-pyruvate, 50 lM 2-mercaptoethanol and 10% heat-inactivated FCS. In order to obtain highly polarized Tc1 or Tc2cells, IL-12 (3.5 ng/mL) or IL-4 (5 ng/mL; R&D Systems,Minneapolis, MN), respectively, was added to the cultures. Insome experiments IL-4 and IL-12 were added together to theculture medium.
Tomeasure cell division, naive TCR7 cells were stained withCFSE as described in [29], and cultured as described above.After 3 days, cells were analyzed by flow cytometry for CFSEexpression and intracellular cytokine production (uponstimulation by phorbol 12-myristate 13-acetate (PMA)/iono-mycin triggering) after staining with cytokine (IFN-c or IL-4)and CD8 antibodies. Alternatively, on day 6 of culture,unmarked Tc1 and Tc2 cells were analyzed for cell surfacemolecule expression and intracellular cytokine production(upon antigen-specific stimulation by RMA cells [63] pulsedwith 1 nM SIINFEKL, or PMA/ionomycin triggering) bystaining with FITC-labeled anti-CD44, anti-CD62L, anti-CD137, anti-Va2, anti-IFN-c, PE-labeled anti-CD8, anti-CD28, anti-CD25, anti-OX40, anti-Vb5, anti-IL-4, and biotin-labeled anti-CD40L, anti-IL-10 antibodies, followed by Cy-chrome-labeled streptavidin as described in [29, 43]. Deadcells were excluded by physical parameter and, when possible,by the addition of propidium iodide immediately beforesample acquisition. As positive control for OX40 expression,Th2 cells were induced by in vitro culture of DO11.10splenocytes and lymph nodes in the presence of IL-4, asdescribed in [64].
Cytokines were measured on day 5 in culture supernatantsfrom Tc1 or Tc2 cells by sandwich ELISA (BD PharMingen),following the manufacturer's instructions. Tc1 and Tc2 cellswere also evaluated for their cytolytic activity in a standard 4-h51Cr-release assay, using as targets RMA cells [63] unpulsed orpulsed with SIINFEKL (from 0.1 to 1000 nM).
Generation and functional characterization of DC
Bone-marrow-derived DC cells were obtained from C57BL/6mice as previously described [29, 30]. To induce maturation,DC were exposed to LPS (1 lg/mL), TNF-a (50 ng/mL; R&DSystems) or ODN-1826 (CpG; 5 lg/mL; InvivoGen, San Diego,CA) for 8 or 48 h of culture. On day 7 of the in vitro culture,non-adherent and loosely adherent cells were collected andevaluated for mycoplasma contamination by PCR (positivecultures were discarded). DC were also analyzed by flowcytometry after incubation with normal mouse serum for15 min at 4�C and double staining with the PE-conjugatedanti-CD11c mAb and one of the following FITC-conjugatedmAb: CD3, CD4, CD8, CD19, CD40, CD80, CD86, NK/2B4, Kb,Db or I-Ab. Propidium iodide was added immediately beforesample acquisition. Cytokine producing capacity was assessed
by flow cytometry after a 4-h stimulation with PMA (60 ng/mL) and ionomycin (1 lg/mL) and staining as previouslydescribed [29]. Biotin-labeled anti-CD11c followed by Cy-chrome-labeled streptavidin, PE-conjugated anti-IL12p70(clone C15.6) or FITC-conjugated anti-IL-10 mAb was usedfor staining.
In vivo experiments
DC were pulsed with 1 lM SIINFEKL or VVYDFLKC (Tag-IV;[42]) peptides for 1 h at 37�C, washed, and resuspended inPBS. Mice were injected i.d. with 5�105 DC. Animals werekilled 6 days later. Splenocytes were restimulated in vitro inthe presence or absence of 100 nM of the relevant peptide.Day-6 blasts were assessed in vitro for cytokine producingcapacity by flow cytometry as described above.
Statistical analysis
Statistical analyses were performed using the Student's t-test.Differences between values were considered statisticallysignificant for p<0.05.
Acknowledgements: We are grateful to Dr. M. Battaglia(San Raffaele Telethon Institute for Gene Therapy,Milan, Italy) for providing us with reagents, and Dr. C.Nembrini (Institute for Integrative Biology, MolecularBiomedicine, Swiss Federal Institute of Technology,Zurich-Schlieren, Switzerland) for helpful experimentalsuggestions. We also thank several colleagues at theCIGTP for critical reading of the manuscript. Thanks toMichael John of the Vita-Salute San Raffaele Universityfor his help with the English language revision. Thestudy was supported by Associazione Italiana per laRicerca sul Cancro (AIRC) and Ministero della Salute eMinistero dell'Istruzione dell'Universit� e della Ricerca.
References
1 Mosmann, T. R. and Coffman, R. L., Th1 and Th2 cells: Different patternsof lymphokine secretion lead to different functional properties. Annu. Rev.Immunol. 1989. 7: 145–173.
2 Bach, F. J., Regulatory T cells under scrutiny. Nat. Rev. Immunol. 2003. 3:189–198.
3 Woodland, D. L. and Dutton, R. W., Heterogeneity of CD4(+) and CD8(+)T cells. Curr. Opin. Immunol. 2003. 15: 336–342.
4 Croft, M., Carter, L., Swain, S. L. and Dutton, R. W., Generation ofpolarized antigen-specific CD8 effector populations: Reciprocal action ofinterleukin (IL)-4 and IL-12 in promoting type 2 versus type 1 cytokineprofiles. J. Exp. Med. 1994. 180: 1715–1728.
5 Rodolfo, M., Zilocchi, C., Accornero, P., Cappetti, B., Arioli, I. andColombo, M. P., IL-4-transduced tumor cell vaccine induces immunor-egulatory type 2 CD8 T lymphocytes that cure lung metastases uponadoptive transfer. J. Immunol. 1999. 163: 1923–1928.
6 Sad, S., Marcotte, R. and Mosmann, T. R., Cytokine-induced differentia-tion of precursor mouse CD8+ T cells into cytotoxic CD8+ T cells secretingTh1 or Th2 cytokines. Immunity 1995. 2: 271–279.
7 Erard, F., Garcia-Sanz, J. A., Moriggl, R. and Wild, M. T., Presence orabsence of TGF-beta determines IL-4-induced generation of type 1 or type 2CD8 T cell subsets. J. Immunol. 1999. 162: 209–214.
Andrea Boni et al. Eur. J. Immunol. 2006. 36: 3157–31663164
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

8 Sheu, B. C., Lin, R. H., Lien, H. C., Ho, H. N., Hsu, S. M. and Huang, S. C.,Predominant Th2/Tc2 polarity of tumor-infiltrating lymphocytes in humancervical cancer. J. Immunol. 2001. 167: 2972–2978.
9 Salgame, P., Abrams, J. S., Clayberger, C., Goldstein, H., Convit, J.,Modlin, R. L. and Bloom, B. R., Differing lymphokine profiles of functionalsubsets of human CD4 and CD8 T cell clones. Science 1991. 254: 279–282.
10 Maggi, E., Giudizi, M. G., Biagiotti, R., Annunziato, F., Manetti, R.,Piccinni, M. P., Parronchi, P. et al., Th2-like CD8+ T cells showing B cellhelper function and reduced cytolytic activity in human immunodeficiencyvirus type 1 infection. J. Exp. Med. 1994. 180: 489–495.
11 Cerwenka, A., Morgan, T. M., Harmsen, A. G. and Dutton, R. W.,Migration kinetics and final destination of type 1 and type 2 CD8 effectorcells predict protection against pulmonary virus infection. J. Exp. Med.1999.189: 423–434.
12 Meissner, N., Kussebi, F., Jung, T., Ratti, H., Baumgarten, C., Werfel, T.,Heusser, C. and Renz, H., A subset of CD8+ T cells from allergic patientsproduce IL-4 and stimulate IgE production in vitro. Clin. Exp. Allergy 1997.27: 1402–1411.
13 Inaoki, M., Sato, S., Shirasaki, F., Mukaida, N. and Takehara, K., Thefrequency of type 2 CD8+ T cells is increased in peripheral blood frompatients with psoriasis vulgaris. J. Clin. Immunol. 2003. 23: 269–278.
14 Accapezzato, D., Francavilla, V., Paroli, M., Casciaro, M., Chircu, L. V.,Cividini, A., Abrignani, S. et al., Hepatic expansion of a virus-specificregulatory CD8(+) Tcell population in chronic hepatitis C virus infection. J.Clin. Invest. 2004. 113: 963–972.
15 Ochi, H.,Wu, X.M., Osoegawa,M., Horiuchi, I., Minohara, M., Murai, H.,Ohyagi, Y. et al., Tc1/Tc2 and Th1/Th2 balance in Asian and Western typesof multiple sclerosis, HTLV-I-associated myelopathy/tropical spastic para-paresis and hyperIgEaemic myelitis. J. Neuroimmunol. 2001. 119: 297–305.
16 Ito, N., Nakamura, H., Tanaka, Y. and Ohgi, S., Lung carcinoma: Analysisof T helper type 1 and 2 cells and T cytotoxic type 1 and 2 cells byintracellular cytokine detection with flow cytometry. Cancer 1999. 85:2359–2367.
17 Tepper, R. I., Pattengale, P. K. and Leder, P.,Murine interleukin-4 displayspotent anti-tumor activity in vivo. Cell 1989. 57: 503–512.
18 Schuler, T., Kammertoens, T., Preiss, S., Debs, P., Noben-Trauth, N. andBlankenstein, T., Generation of tumor-associated cytotoxic T lymphocytesrequires interleukin 4 from CD8(+) T cells. J. Exp. Med. 2001. 194:1767–1775.
19 Kemp, R. A. and Ronchese, F., Tumor-specific Tc1, but not Tc2, cells deliverprotective antitumor immunity. J. Immunol. 2001. 167: 6497–6502.
20 Helmich, B. K. and Dutton, R. W., The role of adoptively transferredCD8 T cells and host cells in the control of the growth of the EG7 thymoma:Factors that determine the relative effectiveness and homing properties ofTc1 and Tc2 effectors. J. Immunol. 2001. 166: 6500–6508.
21 Dobrzanski, M. J., Reome, J. B. and Dutton, R. W., Therapeutic effects oftumor-reactive type 1 and type 2 CD8+ T cell subpopulations in establishedpulmonary metastases. J. Immunol. 1999. 162: 6671–6680.
22 Filaci, G. and Suciu-Foca, N., CD8+ T suppressor cells are back to the game:Are they players in autoimmunity? Autoimmun. Rev. 2002. 1: 279–283.
23 Banchereau, J. and Steinman, R. M., Dendritic cells and the control ofimmunity. Nature 1998. 392: 245–252.
24 Moser, M. and Murphy, K. M., Dendritic cell regulation of Th1-Th2development. Nat. Immunol. 2000. 1: 199–205.
25 Boonstra, A., Asselin-Paturel, C., Gilliet, M., Crain, C., Trinchieri, G., Liu,Y. J. and O'Garra, A., Flexibility of mouse classical and plasmacytoid-derived dendritic cells in directing T helper type 1 and 2 cell development:Dependency on antigen dose and differential Toll-like receptor ligation. J.Exp. Med. 2003. 197: 101–109.
26 Langenkamp, A., Messi, M., Lanzavecchia, A. and Sallusto, F., Kinetics ofdendritic cell activation: Impact on priming of Th1, Th2 and nonpolarizedT cells. Nat. Immunol. 2000. 1: 311–316.
27 Dhodapkar, M. V., Steinman, R. M., Krasovsky, J., Munz, C. andBhardwaj, N., Antigen-specific inhibition of effector T cell function inhumans after injection of immature dendritic cells. J. Exp. Med. 2001. 193:233–238.
28 Gilliet, M. and Liu, Y. J., Generation of human CD8 T regulatory cells byCD40 ligand-activated plasmacytoid dendritic cells. J. Exp. Med. 2002. 195:695–704.
29 Camporeale, A., Boni, A., Iezzi, G., Degl'Innocenti, E., Grioni, M.,Mondino, A. and Bellone, M., Critical impact of the kinetics of dendriticcells activation on the in vivo induction of tumor-specific T lymphocytes.Cancer Res. 2003. 63: 3688–3694.
30 Bellone, M., Cantarella, D., Castiglioni, P., Crosti, M. C., Ronchetti, A.,Moro, M., Garancini, M. P. et al., Relevance of the tumor antigen in thevalidation of three vaccination strategies for melanoma. J. Immunol. 2000.165: 2651–2656.
31 Inaba, K., Inaba, M., Romani, N., Aya, H., Deguchi, M., Ikehara, S.,Muramatsu, S. and Steinman, R. M., Generation of large numbers ofdendritic cells from mouse bone marrow cultures supplemented withgranulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1992. 176:1693–1702.
32 Lutz, M. B., Kukutsch, N., Ogilvie, A. L., Rossner, S., Koch, F., Romani, N.and Schuler, G., An advanced culture method for generating largequantities of highly pure dendritic cells from mouse bone marrow. J.Immunol. Methods 1999. 223: 77–92.
33 Gatti, E., Velleca, M. A., Biedermann, B. C., Ma, W., Unternaehrer, J.,Ebersold, M. W., Medzhitov, R. et al., Large-scale culture and selectivematuration of human Langerhans cells from granulocyte colony-stimulatingfactor-mobilized CD34+ progenitors. J. Immunol. 2000. 164: 3600–3607.
34 Rotzschke, O., Falk, K., Stevanovic, S., Jung, G., Walden, P. andRammensee, H. G., Exact prediction of a natural T cell epitope. Eur. J.Immunol. 1991. 21: 2891–2894.
35 Hogquist, K. A., Jameson, S. C., Heath, W. R., Howard, J. L., Bevan, M. J.and Carbone, F. R., T cell receptor antagonist peptides induce positiveselection. Cell 1994. 76: 17–27.
36 Constant, S. L. and Bottomly, K., Induction of Th1 and Th2 CD4+ T cellresponses: The alternative approaches. Annu. Rev. Immunol. 1997. 15:297–322.
37 Pircher, H., Burki, K., Lang, R., Hengartner, H. and Zinkernagel, R. M.,Tolerance induction in double specific T-cell receptor transgenic mice varieswith antigen. Nature 1989. 342: 559–561.
38 Nembrini, C., Abel, B., Kopf, M. andMarsland, B. J., Strong TCR signaling,TLR ligands, and cytokine redundancies ensure robust development oftype 1 effector T cells. J. Immunol. 2006. 176: 7180–7188.
39 Linton, P. J., Bautista, B., Biederman, E., Bradley, E. S., Harbertson, J.,Kondrack, R. M., Padrick, R. C. and Bradley, L. M., Costimulation viaOX40L expressed by B cells is sufficient to determine the extent of primaryCD4 cell expansion and Th2 cytokine secretion in vivo. J. Exp. Med. 2003.197: 875–883.
40 Roy, M., Waldschmidt, T., Aruffo, A., Ledbetter, J. A. and Noelle, R. J.,The regulation of the expression of gp39, the CD40 ligand, on normal andcloned CD4+ T cells. J. Immunol. 1993. 151: 2497–2510.
41 Frentsch, M., Arbach, O., Kirchhoff, D., Moewes, B., Worm, M., Rothe,M., Scheffold, A. and Thiel, A., Direct access to CD4(+) T cells specific fordefined antigens according to CD154 expression. Nat. Med. 2005. 11:1118–1124.
42 Mylin, L. M., Bonneau, R. H., Lippolis, J. D. and Tevethia, S. S.,Hierarchyamong multiple H-2b-restricted cytotoxic T-lymphocyte epitopes withinsimian virus 40 T antigen. J. Virol. 1995. 69: 6665–6677.
43 Degl'Innocenti, E., Grioni, M., Boni, A., Camporeale, A., Bertilaccio, M.T., Freschi, M., Monno, A. et al., Peripheral T cell tolerance occurs earlyduring spontaneous prostate cancer development and can be rescued bydendritic cell immunization. Eur. J. Immunol. 2005. 35: 66–75.
44 Sander, B., Hoiden, I., Andersson, U., Moller, E. and Abrams, J. S.,Similar frequencies and kinetics of cytokine producing cells in murineperipheral blood and spleen. Cytokine detection by immunoassay andintracellular immunostaining. J. Immunol. Methods 1993. 166: 201–214.
45 Bird, J. J., Brown, D. R., Mullen, A. C., Moskowitz, N. H., Mahowald, M.A., Sider, J. R., Gajewski, T. F. et al., Helper T cell differentiation iscontrolled by the cell cycle. Immunity 1998. 9: 229–237.
46 Iezzi, G., Scotet, E., Scheidegger, D. and Lanzavecchia, A., The interplaybetween the duration of TCR and cytokine signaling determines T cellpolarization. Eur. J. Immunol. 1999. 29: 4092–4101.
Eur. J. Immunol. 2006. 36: 3157–3166 Cellular immune response 3165
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

47 Sallusto, F., Lenig, D., Forster, R., Lipp, M. and Lanzavecchia, A., Twosubsets of memory T lymphocytes with distinct homing potentials andeffector functions. Nature 1999. 401: 708–712.
48 Weninger, W., Crowley, M. A., Manjunath, N. and von Andrian, U. H.,Migratory properties of naive, effector, and memory CD8(+) T cells. J. Exp.Med. 2001. 194: 953–966.
49 Geginat, J., Lanzavecchia, A. and Sallusto, F., Proliferation anddifferentiation potential of human CD8+ memory T-cell subsets in responseto antigen or homeostatic cytokines. Blood 2003. 101: 4260–4266.
50 Granucci, F., Vizzardelli, C., Virzi, E., Rescigno, M. and Ricciardi-Castagnoli, P., Transcriptional reprogramming of dendritic cells bydifferentiation stimuli. Eur. J. Immunol. 2001. 31: 2539–2546.
51 Le Gros, G. and Erard, F., Non-cytotoxic, IL-4, IL-5, IL-10 producing CD8+
Tcells: Their activation and effector functions. Curr. Opin. Immunol.1994. 6:453–457.
52 Vukmanovic-Stejic, M., Vyas, B., Gorak-Stolinska, P., Noble, A. andKemeny, D. M., Human Tc1 and Tc2/Tc0 CD8 T-cell clones display distinctcell surface and functional phenotypes. Blood 2000. 95: 231–240.
53 Najafian, N., Chitnis, T., Salama, A. D., Zhu, B., Benou, C., Yuan, X.,Clarkson, M. R. et al., Regulatory functions of CD8+CD28– T cells in anautoimmune disease model. J. Clin. Invest. 2003. 112: 1037–1048.
54 Chang, C. C., Ciubotariu, R., Manavalan, J. S., Yuan, J., Colovai, A. I.,Piazza, F., Lederman, S. et al., Tolerization of dendritic cells by T(S) cells:The crucial role of inhibitory receptors ILT3 and ILT4. Nat. Immunol. 2002.3: 237–243.
55 Iezzi, G., Boni, A., Degl'Innocenti, E., Grioni, M., Bertilaccio, M. T. andBellone, M., Type-2 cytotoxic T lymphocytes modulate the activity ofdendritic cells toward type-2 immune responses. J. Immunol. 2006. 177:2131–2137.
56 Stoll, S., Delon, J., Brotz, T. M. and Germain, R. N., Dynamic imaging ofT cell-dendritic cell interactions in lymph nodes. Science 2002. 296:1873–1876.
57 Lanzavecchia, A., Immunology. Licence to kill. Nature 1998. 393: 413–414.
58 van Stipdonk, M. J., Lemmens, E. E. and Schoenberger, S. P., Naive CTLsrequire a single brief period of antigenic stimulation for clonal expansionand differentiation. Nat. Immunol. 2001. 2: 423–429.
59 Kaech, S. M. and Ahmed, R., Memory CD8+ T cell differentiation: Initialantigen encounter triggers a developmental program in naive cells. Nat.Immunol. 2001. 2: 415–422.
60 Rotta, G., Edwards, E. W., Sangaletti, S., Bennett, C., Ronzoni, S.,Colombo, M. P., Steinman, R. M. et al., Lipopolysaccharide or wholebacteria block the conversion of inflammatorymonocytes into dendritic cellsin vivo. J. Exp. Med. 2003. 198: 1253–1263.
61 Alpan, O., Bachelder, E., Isil, E., Arnheiter, H. and Matzinger, P.,'Educated' dendritic cells act as messengers from memory to naive T helpercells. Nat. Immunol. 2004. 5: 615–622.
62 Murphy, K. M., Heimberger, A. B. and Loh, D. Y., Induction by antigen ofintrathymic apoptosis of CD4+CD8+TCRlo thymocytes in vivo. Science 1990.250: 1720–1723.
63 Ljunggren, H. G. and Karre, K., Host resistance directed selectively againstH-2-deficient lymphoma variants. Analysis of the mechanism. J. Exp. Med.1985. 162: 1745–1759.
64 Casati, A., Zimmermann, V. S., Benigni, F., Bertilaccio, M. T., Bellone, M.and Mondino, A., The immunogenicity of dendritic cell-based vaccines isnot hampered by doxorubicin and melphalan administration. J. Immunol.2005. 174: 3317–3325.
Andrea Boni et al. Eur. J. Immunol. 2006. 36: 3157–31663166
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
Related Documents







![Tumour-infiltrating cytotoxic T lymphocytes in …...such as CD8+ and natural killer lymphocytes [17], indu-cing a cytotoxic cascade resulting in tumour cell death, while other TILs](https://static.cupdf.com/doc/110x72/5f4838f3212d137c1c54d55d/tumour-infiltrating-cytotoxic-t-lymphocytes-in-such-as-cd8-and-natural-killer.jpg)



