Polymorphism and regulation of the spxB (pyruvate oxidase) virulence factor gene by a CBS-HotDog domain protein (SpxR) in serotype 2 Streptococcus pneumoniae Smirla Ramos-Montañez, 1† Ho-Ching Tiffany Tsui, 1† Kyle J. Wayne, 1 Jordan L. Morris, 1 Lindsey E. Peters, 1 Faming Zhang, 2 Krystyna M. Kazmierczak, 1 Lok-To Sham 1 and Malcolm E. Winkler 1 * Departments of 1 Biology and 2 Chemistry, Indiana University Bloomington, Bloomington, IN 47405, USA. Summary spxB-encoded pyruvate oxidase is a major virulence factor of Streptococcus pneumoniae. During aerobic growth, SpxB synthesizes H2O2 and acetyl phosphate, which play roles in metabolism, signalling, and oxi- dative stress. We report here the first cis- and trans- acting regulatory elements for spxB transcription. These elements were identified in a genetic screen for spontaneous mutations that caused colonies of strain D39 to change from a semitransparent to an opaque appearance. Six of the seven opaque colonies recov- ered (frequency ª 3 ¥ 10 -5 ) were impaired for SpxB function or expression. Two mutations changed amino acids in SpxB likely required for cofactor or subunit binding. One mutation defined a cis-acting adjacent direct repeat required for optimal spxB transcription. The other three spontaneous mutations created the same frameshift near the start of the trans-acting spxR regulatory gene. The SpxR protein contains helix–turn–helix, CBS and HotDog domains implicated in binding DNA, adenosyl compounds, and CoA-containing compounds respectively, and suggest that SpxR positively regulates spxB tran- scription in response to energy and metabolic state. Microarray analyses unexpectedly demonstrated that SpxR also positively regulates the strH exoglycosi- dase gene, which, like spxB, has been implicated in colonization. Finally, SpxR is required for full viru- lence in a murine model of infection. Introduction Streptococcus pneumoniae (pneumococcus) is an aerotolerant anaerobe that causes a variety of serious mucosal and invasive diseases in humans, including pneumonia, otitis media (ear infection), sinusitis, menin- gitis and septicaemia (reviewed in Musher, 2000; Tomasz, 2000; Tuomanen et al., 2004; Orihuela and Toumanen, 2006). Invasive pneumococcal disease results in high annual mortality and morbidity of well over one million people worldwide, especially among young, elderly, debili- tated and immunosuppressed individuals (Janoff and Rubins, 1997; 2004; Orihuela et al., 2004a; Briles et al., 2006). Resistance to a range of antibiotics is increasing at an alarming rate among clinical isolates of S. pneumoniae (Appelbaum, 2002; McCormick et al., 2003; Klugman, 2004), and multiresistant drug strains are emerging among serotypes not targeted by current vaccines (Pichichero and Casey, 2007). As part of its life cycle, pneumococcus exists as a commensal bacterium that inhabits and colonizes the nasopharynx of up to 10% and 40% of healthy adults and children, respectively, at any time (Orihuela and Toumanen, 2006). The transition from commensal bacterium to opportunistic pathogen often occurs following another respiratory tract infection, such as influenza (Barry, 2005). Knowledge of many aspects of pneumococcal colonization, persistence and invasion remains incomplete (Orihuela et al., 2004b; LeMessurier et al., 2006; Orihuela and Toumanen, 2006). Several virulence factors have been identified among the different capsular serotypes of S. pneumoniae (see Hava et al., 2003; Jedrzejas, 2004; Lopez and Garcia, 2004; Mitchell, 2004; Orihuela et al., 2004b; LeMessurier et al., 2006). Among these, pyruvate oxidase, encoded by spxB, is required for most stages of infection (Spellerberg et al., 1996; Pericone et al., 2000; Orihuela et al., 2004b; Hoffmann et al., 2006; Regev-Yochay et al., 2007). Pyruvate oxidase is a decarboxylase that catalyses the conversion of pyruvate, inorganic phosphate (P i), and molecular oxygen (O2) to hydrogen peroxide (H2O2), carbon dioxide (CO2) and acetyl phosphate, which acts as a high-energy phosphoryl group donor (Fig. 1). This reac- tion involves the two bound cofactors thiamin diphosphate Accepted 1 December, 2007. *For correspondence. E-mail [email protected]; Tel. (+1) 812 856 1318; Fax (+1) 812 855 6705. † Contributed equally to this work. Molecular Microbiology (2008) 67(4), 729–746 doi:10.1111/j.1365-2958.2007.06082.x First published online 2 January 2008 © 2008 The Authors Journal compilation © 2008 Blackwell Publishing Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Polymorphism and regulation of the spxB (pyruvate oxidase)virulence factor gene by a CBS-HotDog domain protein (SpxR)in serotype 2 Streptococcus pneumoniae
Smirla Ramos-Montañez,1† Ho-Ching Tiffany Tsui,1†
Kyle J. Wayne,1 Jordan L. Morris,1
Lindsey E. Peters,1 Faming Zhang,2
Krystyna M. Kazmierczak,1 Lok-To Sham1 andMalcolm E. Winkler1*Departments of 1Biology and 2Chemistry, IndianaUniversity Bloomington, Bloomington, IN 47405, USA.
Summary
spxB-encoded pyruvate oxidase is a major virulencefactor of Streptococcus pneumoniae. During aerobicgrowth, SpxB synthesizes H2O2 and acetyl phosphate,which play roles in metabolism, signalling, and oxi-dative stress. We report here the first cis- and trans-acting regulatory elements for spxB transcription.These elements were identified in a genetic screen forspontaneous mutations that caused colonies of strainD39 to change from a semitransparent to an opaqueappearance. Six of the seven opaque colonies recov-ered (frequency ª 3 ¥ 10-5) were impaired for SpxBfunction or expression. Two mutations changedamino acids in SpxB likely required for cofactor orsubunit binding. One mutation defined a cis-actingadjacent direct repeat required for optimal spxBtranscription. The other three spontaneous mutationscreated the same frameshift near the start of thetrans-acting spxR regulatory gene. The SpxR proteincontains helix–turn–helix, CBS and HotDog domainsimplicated in binding DNA, adenosyl compounds,and CoA-containing compounds respectively, andsuggest that SpxR positively regulates spxB tran-scription in response to energy and metabolic state.Microarray analyses unexpectedly demonstrated thatSpxR also positively regulates the strH exoglycosi-dase gene, which, like spxB, has been implicatedin colonization. Finally, SpxR is required for full viru-lence in a murine model of infection.
Introduction
Streptococcus pneumoniae (pneumococcus) is anaerotolerant anaerobe that causes a variety of seriousmucosal and invasive diseases in humans, includingpneumonia, otitis media (ear infection), sinusitis, menin-gitis and septicaemia (reviewed in Musher, 2000; Tomasz,2000; Tuomanen et al., 2004; Orihuela and Toumanen,2006). Invasive pneumococcal disease results in highannual mortality and morbidity of well over one millionpeople worldwide, especially among young, elderly, debili-tated and immunosuppressed individuals (Janoff andRubins, 1997; 2004; Orihuela et al., 2004a; Briles et al.,2006). Resistance to a range of antibiotics is increasing atan alarming rate among clinical isolates of S. pneumoniae(Appelbaum, 2002; McCormick et al., 2003; Klugman,2004), and multiresistant drug strains are emergingamong serotypes not targeted by current vaccines(Pichichero and Casey, 2007). As part of its life cycle,pneumococcus exists as a commensal bacterium thatinhabits and colonizes the nasopharynx of up to 10% and40% of healthy adults and children, respectively, at anytime (Orihuela and Toumanen, 2006). The transition fromcommensal bacterium to opportunistic pathogen oftenoccurs following another respiratory tract infection, suchas influenza (Barry, 2005). Knowledge of many aspectsof pneumococcal colonization, persistence and invasionremains incomplete (Orihuela et al., 2004b; LeMessurieret al., 2006; Orihuela and Toumanen, 2006).
Several virulence factors have been identified amongthe different capsular serotypes of S. pneumoniae (seeHava et al., 2003; Jedrzejas, 2004; Lopez and Garcia,2004; Mitchell, 2004; Orihuela et al., 2004b; LeMessurieret al., 2006). Among these, pyruvate oxidase, encoded byspxB, is required for most stages of infection (Spellerberget al., 1996; Pericone et al., 2000; Orihuela et al., 2004b;Hoffmann et al., 2006; Regev-Yochay et al., 2007).Pyruvate oxidase is a decarboxylase that catalyses theconversion of pyruvate, inorganic phosphate (Pi), andmolecular oxygen (O2) to hydrogen peroxide (H2O2),carbon dioxide (CO2) and acetyl phosphate, which acts asa high-energy phosphoryl group donor (Fig. 1). This reac-tion involves the two bound cofactors thiamin diphosphate
Accepted 1 December, 2007. *For correspondence. [email protected]; Tel. (+1) 812 856 1318; Fax (+1) 812855 6705. †Contributed equally to this work.
Molecular Microbiology (2008) 67(4), 729–746 � doi:10.1111/j.1365-2958.2007.06082.xFirst published online 2 January 2008
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd

(ThDP) and flavin adenine dinucleotide (FAD) (Sedewitzet al., 1984; Muller et al., 1994; Tittmann et al., 1998;2005). The crystal structure of the pyruvate oxidase tet-ramer from Lactobacillus plantarum shows that the ThDPand FAD cofactors are closely situated in active sitescomprised of segments from adjacent subunits (Mulleret al., 1994; Tittmann et al., 2005). While aspects of themechanism of the two-electron reduction of O2 catalysedby SpxB are typical of other ThDP enzymes, recentresults indicate that this mechanism involves novel kineticcoupling via a flavin-mediated radical (Tittmann et al.,2005).
Pyruvate oxidase plays several critical roles in pneumo-coccal metabolism and virulence. Many bacteria usephosphotransacetylase (Pta) to convert acetyl-CoA intoacetyl phosphate (reviewed in Wolfe, 2005). In pneumo-coccus, the additional acetyl phosphate synthesized bythe SpxB pyruvate oxidase accounts for as much as 85%of the pool synthesized during some aerobic growth con-ditions (Pericone et al., 2003). This acetyl phosphateserves as a precursor for ATP biosynthesis (Fig. 1) (Peri-cone et al., 2003), which is important in a bacterium thatlacks a tricarboxylic acid cycle and electron transport(Hoskins et al., 2001; Tettelin et al., 2001; Lanie et al.,2007). In addition, there has been speculation that thispool of acetyl phosphate acts in the phosphorylation ofresponse regulators and thereby affects global regulationpatterns (Spellerberg et al., 1996; Kuboniwa et al., 2006).
Besides synthesizing acetyl phosphate, SpxB pyruvateoxidase produces relatively high concentrations (~1 mM)
of extracellular H2O2 (Pericone et al., 2000; 2003;Belanger et al., 2004; Regev-Yochay et al., 2007). Para-doxically, pneumococcus lacks a catalase, and themechanisms used to cope with this endogenous oxidativestress are largely unknown (Pericone et al., 2003). Long-term incubation of pneumococcal strains in laboratorymedia at stationary phase results in cell death that ispartly prevented by deletion of spxB (Belanger et al.,2004; Regev-Yochay et al., 2007). On the other hand,spxB mutants are hypersensitive to sublethal concentra-tions of added H2O2 (Pericone et al., 2003; Regev-Yochayet al., 2007), presumably because of decreased ATP pro-duction (Pericone et al., 2003). The copious amount ofH2O2 produced by pneumococcus inhibits the growth ofcompeting bacteria in the nasopharynx and in vitro culture(Pericone et al., 2000; Regev-Yochay et al., 2006),damages host cells (reviewed in Mitchell, 2004), and likelyalters their production of cytokines (DeYulia et al., 2005;Xue et al., 2005). Notably, bacterial-produced H2O2 reactswith host-produced nitric oxide to produce toxic com-pounds that cause cellular damage in pneumococcalmeningitis (Hoffmann et al., 2006). Unencapsulated spxBmutants are less adherent to mammalian cells than theirisogenic spxB + parent strain (Spellerberg et al., 1996),and encapsulated spxB knock-out mutants are stronglyattenuated in murine infection models following intranasalor intratracheal, but not intravenous, inoculation (Orihuelaet al., 2004b; Orihuela and Toumanen, 2006).
Changes in SpxB expression seem to be correlatedwith phase variation in S. pneumoniae (Overweg et al.,
Fig. 1. Proposed pathway for acetyl-CoA and acetyl phosphate biosynthesis in S. pneumoniae. The reaction catalysed by SpxB pyruvateoxidase is highlighted. The top line shows an abbreviated version of the Embden–Meyerhof–Parnas pathway for glycolysis to pyruvate and thelactate dehydrogenase (Ldh) reaction from pyruvate to lactate. The pyruvate → acetoin → acetyl-CoA pathway is proposed based on thepresence of genes encoding a putative acetoin dehydrogenase in S. pneumoniae genomes (Hoskins et al., 2001; Tettelin et al., 2001; Lanieet al., 2007) and characterization of these steps in Bacillus subtilis metabolism (see Renna et al., 1993; Huang et al., 1999; Shirk et al., 2002).Additional enzymes and coenzymes shown: GapA, glyceraldehyde-3-phosphate dehydrogenase; CoA-SH, free coenzyme A, Pta,phosphotransacetylase; AckA, acetate kinase. The strong positive regulation of spxB and strH transcription by the SpxR protein reported hereis indicated along with the hypothesis that SpxR senses metabolic state by binding adenosyl and CoA-containing compounds. See text foradditional details.
730 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

2000; Pericone et al., 2000; 2002). Pneumococcal phasevariation was detected by changes between transparentand opaque colony morphologies on clear tryptic soy agar(TSA) plates lacking blood, but seeded with catalase(Weiser et al., 1994). A general model has emerged thattransparent variants are more proficient in colonization,produce more teichoic acid and H2O2, but less capsulethan opaque variants, which are more proficient in sepsis(reviewed in Weiser, 2006). However, the genetic mecha-nisms underlying phase variation have remained elusive,and may in fact, vary among different serotypes ofS. pneumoniae (Weiser, 2006). The proposed roles ofspxB function and regulation in pneumococcal phasevariation have been somewhat contradictory. On onehand, it was stated that SpxB expression level wasunlikely to determine colony morphology (Overweg et al.,2000). On the other hand, some opaque variants werelater found to be defective in SpxB function (Periconeet al., 2002), and there have been recent suggestions thatSpxB does determine colony morphology and may play arole in phase variation (Belanger et al., 2004; Weiser,2006).
During a recent genomics project, we noticed that twodistantly separated isolates of encapsulated serotypestrain D39 showed different colony morphologies thatwere correlated with H2O2 production (Lanie et al., 2007).We also noted a polymorphism between the amino acidsequence of SpxB of serotype 2 strains and those of otherpneumococcal serotypes. To understand these morpho-logical and mutational differences and their possible rela-tionship to phase variation, we performed a systematicgenetic screen for opaque colony formation in strain D39.We report here that spontaneous opaque variants containseveral types of mutations that affect the function andregulation of spxB, including replication-slippage muta-tions in a gene, designated spxR (streptococcus pyruvateoxidase regulator) (see Spellerberg et al., 1996), thatencodes a novel regulator. We also report on the long-standing issue of whether lack of spxB function influencesglobal transcription patterns and present the first charac-terization of the spxR regulon.
Results
Polymorphism of spxB in serotype 2 strains and theirderivatives
A previous study compared spxB function in an isolate ofvirulent, encapsulated serotype 2 strain D39 with that ofunencapsulated laboratory strain R6, which was originallyderived from strain D39 (Belanger et al., 2004). A majorconclusion of this study was that R6 acquired a mutation inspxB that increased H2O2 production and thereby contrib-uted to the ‘rough’ colony morphology of strain R6
compared with D39. Recently, we compared the correctedsequence of R6 with those of two isolates of strain D39(Lanie et al., 2007), including the one studied previously[D39 (IU1680); Table S1; Belanger et al., 2004]. Compari-son of these three related sequences allowed us to deducea likely progenitor D39 sequence (Lanie et al., 2007).
The deduced progenitor sequence of spxB is stillpresent in strain D39 (IU1690) obtained from NCTC(Table S1) and encodes amino acids Asn95, Ile152 andAla282 (Fig. 2A; Table 1). SpxB of R6 or D39 (IU1680)contains an Asn95Asp or Ala282Thr change, respectively,which likely arose by spontaneous mutation in these iso-lates (Table 1; Lanie et al., 2007). In addition, SpxB ofD39 (IU1690) and R6 contains Ala282 instead of Pro282,which is present in SpxB of all other pneumococcal strainssequenced to date (Table 1) and at the equivalent positionin the pyruvate oxidases of other bacterial species (datanot shown; see Discussion). We determined the rates ofH2O2 production normalized to initial cell densities ofvarious strains freshly inoculated into diluted brain heartinfusion (BHI) broth (Experimental procedures). The dou-bling times of these strains were comparable in BHI broth(data not shown). D39 (IU1690), R6 and strain WU2(serotype 3) produced H2O2 at similar rates, and TIGR4(serotype 4) produced H2O2 at a slightly higher rate thanD39 (IU1690) (bars 8, 1, 22 and 23; Fig. 3), despite thepolymorphism in SpxB (Table 1). In comparison, D39(IU1680) produced about 2.9-fold less H2O2 per min thanD39 (IU1690) (bars 4 and 8; Fig. 3). We confirmedthat this decrease in H2O2 production was due to theAla282Thr mutation by swapping the corresponding allelefrom D39 (IU1680) into D39 (IU1690) rpsL1 (bars 9 and11; Fig. 3). Conversely, we increased H2O2 production inD39 (IU1680) rpsL1 by swapping in the allele encodingAla282 from D39 (IU1690) (bars 5 and 7; Fig. 3).
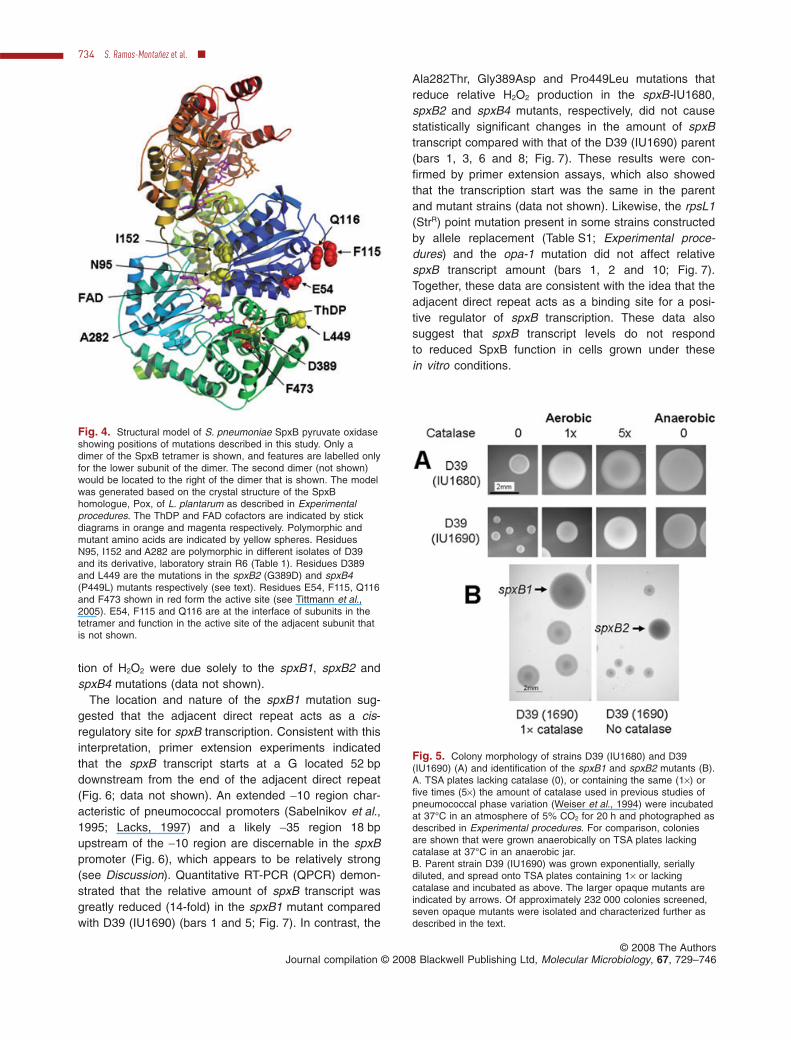
We constructed a model of pneumococcal SpxBbased on the crystal structure of its Pox homologue inL. plantarum (Fig. 4) (Experimental procedures; Mulleret al., 1994). Ile152 is located away from the ThDP andFAD cofactor binding sites and active site residues(Fig. 4). The Asn95Asp mutation in strain R6 is locatednear the FAD binding site, but the Asn95Asp change wasnot deleterious to SpxB function (Fig. 3). Likewise, thePro282 to Ala282 polymorphism in serotype 2 SpxB(Table 1) did not significantly reduce SpxB activity (Fig. 3),even though Ala282 is located in the FAD binding site. Bycontrast, the Ala282Thr mutation in strain D39 (IU1680)did reduce SpxB activity (Fig. 3), probably by reducingFAD binding (Fig. 4). Finally, one implication of theseresults is that R6 and its progenitor D39 strain likely pro-duced similar amounts of H2O2, which does not support acentral assumption underlying the conclusions in theprevious work comparing spxB function in R6 and D39(IU1680) (Belanger et al., 2004).
cis- and trans-regulation of pneumococcal spxB 731
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

Genetic screen for opaque colonies in D39 (IU1690)
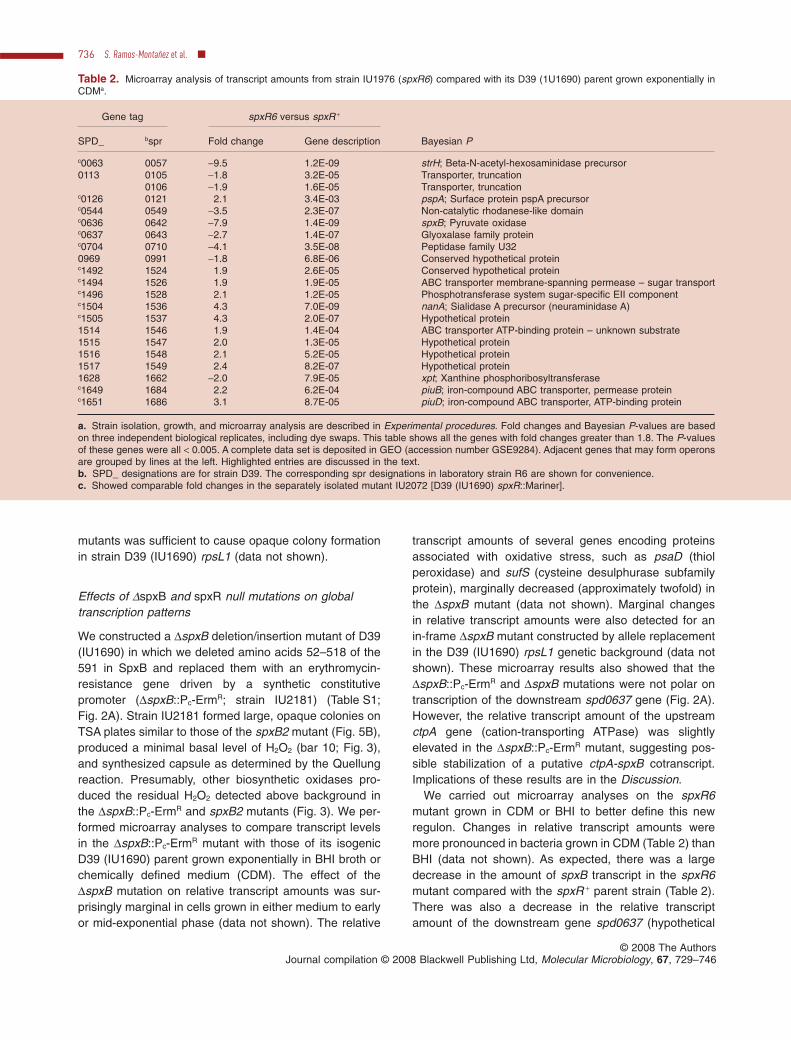
In previous studies of pneumococcal phase variation,opaque colonies appeared larger and more dome-shapedthan umbilicated transparent colonies on clear TSA plates(Weiser et al., 1994). We noticed that D39 (IU1690)formed smaller, more transparent colonies than those ofD39 (IU1680) on TSA plates lacking or containing thesame amount (1¥) of catalase used in previous studies ofphase variation (Fig. 5A; Weiser et al., 1994). At a higherconcentration of added catalase (5¥) or under anaerobicgrowth conditions, both strains formed very opaque colo-nies, although D39 (IU1680) colonies remained some-what larger than those of D39 (IU1690) (Fig. 5A).
Strains producing higher levels of H2O2 were reported toappear smaller, more transparent, and umbilicated com-pared with strains producing less H2O2, even on bloodagar plates (Belanger et al., 2004). The dependence ofthe colony morphology of D39 (IU1690) on catalase and
the presence of oxygen (Fig. 5A) suggested that thedifference in colony morphology between D39 (IU1690)and D39 (IU1680) was due to reduced H2O2 productionby D39 (IU1680) (Figs 3–5; Table 1). Consistent withthis interpretation, allele swap experiments showed thatthe semitransparent or opaque colony morphologiesdepended on whether Ala282 or Thr282, respectively, waspresent in SpxB in the D39 (IU1690) and D39 (IU1680)genetic backgrounds (data not shown).
Phase variants were reported to appear in serotype2-derived strains, such as R6 and D39, relatively infre-quently compared with other serotypes (Weiser et al.,1994; Saluja and Weiser, 1995). In fact, the frequency ofphase variation in strain D39 was not determineddirectly, and a frequency of < 10-6-10-7 was deducedfrom experiments going back to work by Avery andcoworkers (Weiser et al., 1994). We wanted to deter-mine the spontaneous frequency of opaque colonyformation by the semitransparent strain D39 (IU1690) for
Fig. 2. Structures of the spxB (A) and spxR(B) genes and flanking regions in theS. pneumoniae D39 and R6 genomes. Thefigures are drawn to scale. Also indicated arethe positions of PCR primers used to amplifysegments of genomic DNA for sequencedeterminations, PCR primers used in QPCRdeterminations of transcript amounts, 50-meroligomers spotted onto microarrays used todetermine relative transcript amounts, andprimers used in primer extension assays (seeExperimental procedures). The diagramsindicate the locations of the polymorphisms(AA95, AA152 and AA282) and mutations(DspxB, spxB1, spxB2 and spxB4) in spxB (A)and the spxR::Mariner insertion and spxR3,spxR6 and spxR7 Dag frameshift mutations inspxR (B). The HTH, DRTGG-CBS andHotDog domains in the SpxR protein, whichare hypothesized to bind to DNA, adenosylcompounds, and CoA-containing compounds,respectively, are indicated above the spxRreading frame (B). See text for additionaldetails.
Table 1. Polymorphism of SpxB in different serotypes of Streptococcus pneumoniae.
Straina AA95 (codon) AA152 (codon) AA282 (codon)
R6 (unencapsulated) Asp (GAC) Ile (ATT) Ala (GCA)D39 (IU1680) (2) Asn (AAC) Ile (ATT) Thr (ACA)D39 (IU1690) (2) Asn (ACC) Ile (ATT) Ala (GCA)INV104B (1), 23F (23), Sp11BS70 (11), Sp23BS72 (23) and Sp14BS69 (14) Asn (AAC) Ile (ATT) Pro (CCA)WU2 (3), TIGR4 (4), TIGR670 (6B), OXC141 (3), INV200 (14), Sp6BS73 (6),
Sp9BS68 (9), Sp3BS71 (3), Sp18BS74 (6), and Sp19BS75 (19)Asn (AAC) Val (GTT) Pro (CCA)
a. R6 is an unencapsulated derivative of serotype 2 strain D39. Serotypes of other strains are indicated in parenthesis (see Hiller et al., 2007).The Asn95Asp and Ala282Thr mutations in R6 and D39 (IU1680) are in boldface.
732 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746
two reasons. First, we would learn whether defectivespxB expression plays a major role in the appearance ofopaque colony variants, similar to those observed forD39 (IU1680) compared with D39 (IU1690) (Fig. 5).Second, this genetic screen would turn up regulatorymutations of spxB.
We screened approximately 232 000 colonies of D39(IU1690) by plating serial dilutions of cultures grown expo-nentially in tryptic soy broth (TSB) onto TSA plates,lacking (~165 000 colonies) or containing 1¥ catalase(~67 000 colonies) (Fig. 5B; Experimental procedures).Opaque colonies were found in both the presence andabsence of catalase, although the semitransparent D39(IU1690) parent and opaque variant colonies were largeron plates containing catalase (Fig. 5B). We recoveredseven spontaneous opaque variants from this screen.Thus, the frequency of opaque colony appearance of D39was approximately 3.0 ¥ 10-5, which is comparable withthat for other pneumococcal serotypes (Weiser et al.,1994). In the Discussion, this frequency is compared withthat of spontaneous resistance to the antibiotics rifampicinand optochin.
Of the seven opaque variants recovered in this screen,six showed increased colony size compared with thesemitransparent D39 (IU1690) parent, whereas one, con-taining a mutation designated opa-1, formed the samesized colony as the parent. We assayed the sevenopaque variants and a DspxB mutant for rates of H2O2
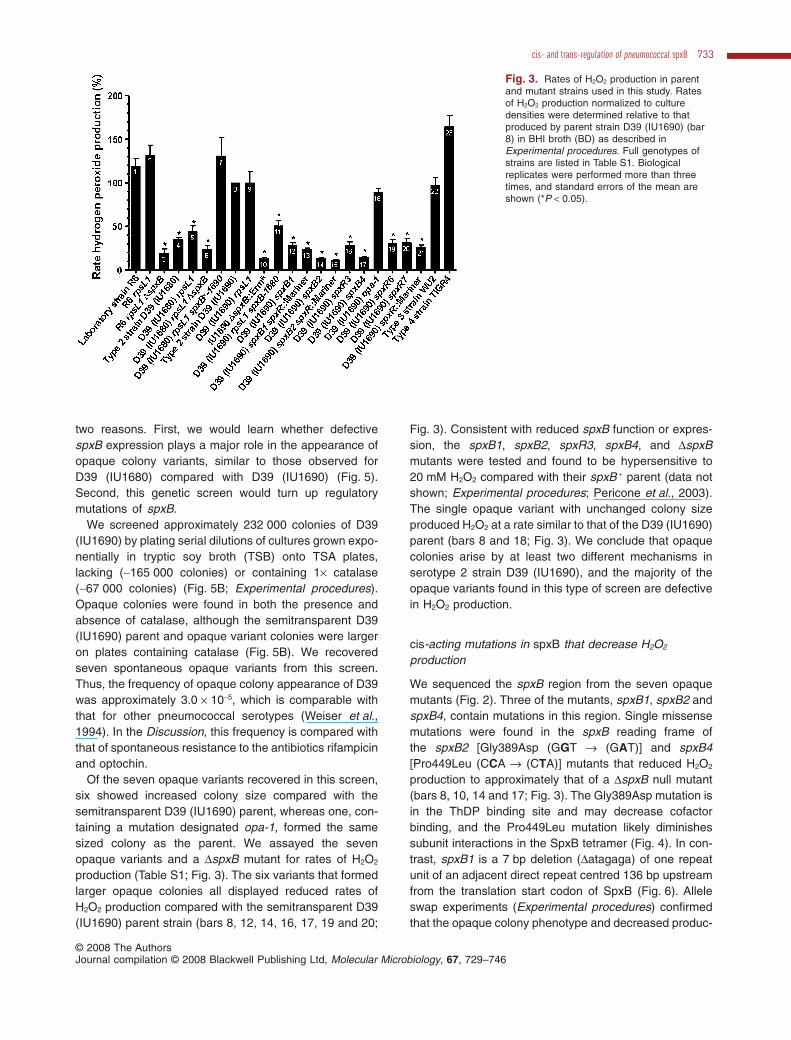
production (Table S1; Fig. 3). The six variants that formedlarger opaque colonies all displayed reduced rates ofH2O2 production compared with the semitransparent D39(IU1690) parent strain (bars 8, 12, 14, 16, 17, 19 and 20;
Fig. 3). Consistent with reduced spxB function or expres-sion, the spxB1, spxB2, spxR3, spxB4, and DspxBmutants were tested and found to be hypersensitive to20 mM H2O2 compared with their spxB + parent (data notshown; Experimental procedures; Pericone et al., 2003).The single opaque variant with unchanged colony sizeproduced H2O2 at a rate similar to that of the D39 (IU1690)parent (bars 8 and 18; Fig. 3). We conclude that opaquecolonies arise by at least two different mechanisms inserotype 2 strain D39 (IU1690), and the majority of theopaque variants found in this type of screen are defectivein H2O2 production.
cis-acting mutations in spxB that decrease H2O2
production
We sequenced the spxB region from the seven opaquemutants (Fig. 2). Three of the mutants, spxB1, spxB2 andspxB4, contain mutations in this region. Single missensemutations were found in the spxB reading frame ofthe spxB2 [Gly389Asp (GGT → (GAT)] and spxB4[Pro449Leu (CCA → (CTA)] mutants that reduced H2O2
production to approximately that of a DspxB null mutant(bars 8, 10, 14 and 17; Fig. 3). The Gly389Asp mutation isin the ThDP binding site and may decrease cofactorbinding, and the Pro449Leu mutation likely diminishessubunit interactions in the SpxB tetramer (Fig. 4). In con-trast, spxB1 is a 7 bp deletion (Datagaga) of one repeatunit of an adjacent direct repeat centred 136 bp upstreamfrom the translation start codon of SpxB (Fig. 6). Alleleswap experiments (Experimental procedures) confirmedthat the opaque colony phenotype and decreased produc-
Fig. 3. Rates of H2O2 production in parentand mutant strains used in this study. Ratesof H2O2 production normalized to culturedensities were determined relative to thatproduced by parent strain D39 (IU1690) (bar8) in BHI broth (BD) as described inExperimental procedures. Full genotypes ofstrains are listed in Table S1. Biologicalreplicates were performed more than threetimes, and standard errors of the mean areshown (*P < 0.05).
cis- and trans-regulation of pneumococcal spxB 733
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746
tion of H2O2 were due solely to the spxB1, spxB2 andspxB4 mutations (data not shown).
The location and nature of the spxB1 mutation sug-gested that the adjacent direct repeat acts as a cis-regulatory site for spxB transcription. Consistent with thisinterpretation, primer extension experiments indicatedthat the spxB transcript starts at a G located 52 bpdownstream from the end of the adjacent direct repeat(Fig. 6; data not shown). An extended -10 region char-acteristic of pneumococcal promoters (Sabelnikov et al.,1995; Lacks, 1997) and a likely -35 region 18 bpupstream of the -10 region are discernable in the spxBpromoter (Fig. 6), which appears to be relatively strong(see Discussion). Quantitative RT-PCR (QPCR) demon-strated that the relative amount of spxB transcript wasgreatly reduced (14-fold) in the spxB1 mutant comparedwith D39 (IU1690) (bars 1 and 5; Fig. 7). In contrast, the
Ala282Thr, Gly389Asp and Pro449Leu mutations thatreduce relative H2O2 production in the spxB-IU1680,spxB2 and spxB4 mutants, respectively, did not causestatistically significant changes in the amount of spxBtranscript compared with that of the D39 (IU1690) parent(bars 1, 3, 6 and 8; Fig. 7). These results were con-firmed by primer extension assays, which also showedthat the transcription start was the same in the parentand mutant strains (data not shown). Likewise, the rpsL1(StrR) point mutation present in some strains constructedby allele replacement (Table S1; Experimental proce-dures) and the opa-1 mutation did not affect relativespxB transcript amount (bars 1, 2 and 10; Fig. 7).Together, these data are consistent with the idea that theadjacent direct repeat acts as a binding site for a posi-tive regulator of spxB transcription. These data alsosuggest that spxB transcript levels do not respondto reduced SpxB function in cells grown under thesein vitro conditions.
Fig. 4. Structural model of S. pneumoniae SpxB pyruvate oxidaseshowing positions of mutations described in this study. Only adimer of the SpxB tetramer is shown, and features are labelled onlyfor the lower subunit of the dimer. The second dimer (not shown)would be located to the right of the dimer that is shown. The modelwas generated based on the crystal structure of the SpxBhomologue, Pox, of L. plantarum as described in Experimentalprocedures. The ThDP and FAD cofactors are indicated by stickdiagrams in orange and magenta respectively. Polymorphic andmutant amino acids are indicated by yellow spheres. ResiduesN95, I152 and A282 are polymorphic in different isolates of D39and its derivative, laboratory strain R6 (Table 1). Residues D389and L449 are the mutations in the spxB2 (G389D) and spxB4(P449L) mutants respectively (see text). Residues E54, F115, Q116and F473 shown in red form the active site (see Tittmann et al.,2005). E54, F115 and Q116 are at the interface of subunits in thetetramer and function in the active site of the adjacent subunit thatis not shown.
Fig. 5. Colony morphology of strains D39 (IU1680) and D39(IU1690) (A) and identification of the spxB1 and spxB2 mutants (B).A. TSA plates lacking catalase (0), or containing the same (1¥) orfive times (5¥) the amount of catalase used in previous studies ofpneumococcal phase variation (Weiser et al., 1994) were incubatedat 37°C in an atmosphere of 5% CO2 for 20 h and photographed asdescribed in Experimental procedures. For comparison, coloniesare shown that were grown anaerobically on TSA plates lackingcatalase at 37°C in an anaerobic jar.B. Parent strain D39 (IU1690) was grown exponentially, seriallydiluted, and spread onto TSA plates containing 1¥ or lackingcatalase and incubated as above. The larger opaque mutants areindicated by arrows. Of approximately 232 000 colonies screened,seven opaque mutants were isolated and characterized further asdescribed in the text.
734 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

trans-acting mutations in spxR that decrease H2O2
production
DNA sequencing demonstrated that the three remainingmutations spxR3, spxR6 and spxR7 were not located inthe spxB region. The relative amount of spxB transcriptamount dropped significantly (13-fold) in the spxR3 andspxR6 mutants (bars 1, 7 and 9; Fig. 7), suggesting thatspxR3 and spxR6 could inactivate a positive regulatorof spxB transcription. Similar results were obtained in pri-mer extension assays (data not shown). To locate thespxR3, spxR6 and spxR7 mutations, we performed trans-poson mutagenesis and screened for insertions that led toopaque colony formation. We performed in vitro Marinertransposon jumps and subsequent screening in the R6genetic background because of its higher transformationfrequency compared with D39 (see Belanger et al., 2004;Lanie et al., 2007). As a preliminary experiment, weswapped the spxB1 deletion mutation from D39 (IU1690)(Fig. 6) into strain R6 and confirmed opaque colony for-mation and reduced H2O2 production in R6. This resultsuggested that spxB transcription is likely regulated by thesame mechanism in the related R6 and D39 strains(Belanger et al., 2004; Lanie et al., 2007). We screened4224 spectinomycin-resistant R6 colonies containingtransposons before finding a mutant that formed largeropaque colonies on TSA plates and had reduced H2O2
production (data not shown). This number of transposoninsertions represented approximately 80% to 90% cover-age of the chromosome, but was not saturating, becausewe did not recover insertions in spxB.
Polymerase chain reaction indicated that the insertion,designated spxR::Mariner, was not located in the spxBregion of the chromosome (Fig. 2A). Inverse PCR (Experi-
mental procedures) located the Mariner insertion in themiddle of gene spr0991 in R6 (spd0969 in D39; Fig. 2B).The spxR::Mariner insertion was moved into strain D39(IU1690) by transformation with purified genomic DNAfrom the R6 spxR::Mariner mutant strain IU2002 or bytransformation with an amplicon containing the spxR::Ma-riner insertion (Table S1). The resulting strains (IU2013and IU2072) formed opaque colonies on TSA plates, andIU2072 showed reduced H2O2 production similar to that ofthe spxR3, spxR6 and spxR7 mutants (bars 8, 16, 19, 20and 21; Fig. 3). We sequenced the spxR region in D39(IU1690) and the spxR3, spxR6 and spxR7 mutants. Allthree spontaneous mutants, which arose in two indepen-dent cultures (spxR3 in one and spxR6 and spxR7 fromanother), contained a Dag deletion in a run of five agdinucleotide repeats near the beginning of spxR (codon90 of 495; Fig. 2B). This Dag deletion results in a frame-shift mutation that truncates the SpxR protein after 93amino acids (Fig. 2B) and likely arose by a slippagereplication error of the kind that mediates phase variationin a variety of bacteria (Moxon et al., 1998; Periconeet al., 2002; Fox et al., 2005). The SpxR regulatory proteincontains several domains, including putative helix–turn–helix (HTH) DNA-binding and CBS adenosyl compound-binding domains (Fig. 2B), which are described in theDiscussion. Finally, we used allele exchange to confirmthat the Dag mutation from the spxR3, spxR6 and spxR7
Fig. 6. Promoter region of the spxB gene in S. pneumoniae R6and D39. The predicted translation start codon, transcription startsite, extended -10 region, and likely -35 region are indicated. Thetranscription start site was mapped by primer extension(Experimental procedures). The spxB and spxR mutationsdescribed herein did not alter this transcription start site. Onerepeat of the adjacent direct repeat is deleted in the spxB1 mutant.A putative cre element that could bind the catabolite repressorCcpA is indicated. See text for details.
Fig. 7. Relative spxB transcript amounts determined by QPCR inparent and spxB and spxR mutant strains. RNA was prepared frombacteria growing exponentially in BHI broth at 37°C in anatmosphere of 5% CO2, and QPCR was performed as described inExperimental procedures. The amount of spxB transcript wasnormalized to that of 16S rRNA in each preparation. Data from twoor more biological replicates were averaged and are expressedrelative to the normalized amount of spxB transcript in the D39(IU1690) parent strain. (*P < 0.05).
cis- and trans-regulation of pneumococcal spxB 735
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746
mutants was sufficient to cause opaque colony formationin strain D39 (IU1690) rpsL1 (data not shown).
Effects of DspxB and spxR null mutations on globaltranscription patterns
We constructed a DspxB deletion/insertion mutant of D39(IU1690) in which we deleted amino acids 52–518 of the591 in SpxB and replaced them with an erythromycin-resistance gene driven by a synthetic constitutivepromoter (DspxB::Pc-ErmR; strain IU2181) (Table S1;Fig. 2A). Strain IU2181 formed large, opaque colonies onTSA plates similar to those of the spxB2 mutant (Fig. 5B),produced a minimal basal level of H2O2 (bar 10; Fig. 3),and synthesized capsule as determined by the Quellungreaction. Presumably, other biosynthetic oxidases pro-duced the residual H2O2 detected above background inthe DspxB::Pc-ErmR and spxB2 mutants (Fig. 3). We per-formed microarray analyses to compare transcript levelsin the DspxB::Pc-ErmR mutant with those of its isogenicD39 (IU1690) parent grown exponentially in BHI broth orchemically defined medium (CDM). The effect of theDspxB mutation on relative transcript amounts was sur-prisingly marginal in cells grown in either medium to earlyor mid-exponential phase (data not shown). The relative
transcript amounts of several genes encoding proteinsassociated with oxidative stress, such as psaD (thiolperoxidase) and sufS (cysteine desulphurase subfamilyprotein), marginally decreased (approximately twofold) inthe DspxB mutant (data not shown). Marginal changesin relative transcript amounts were also detected for anin-frame DspxB mutant constructed by allele replacementin the D39 (IU1690) rpsL1 genetic background (data notshown). These microarray results also showed that theDspxB::Pc-ErmR and DspxB mutations were not polar ontranscription of the downstream spd0637 gene (Fig. 2A).However, the relative transcript amount of the upstreamctpA gene (cation-transporting ATPase) was slightlyelevated in the DspxB::Pc-ErmR mutant, suggesting pos-sible stabilization of a putative ctpA-spxB cotranscript.Implications of these results are in the Discussion.
We carried out microarray analyses on the spxR6mutant grown in CDM or BHI to better define this newregulon. Changes in relative transcript amounts weremore pronounced in bacteria grown in CDM (Table 2) thanBHI (data not shown). As expected, there was a largedecrease in the amount of spxB transcript in the spxR6mutant compared with the spxR + parent strain (Table 2).There was also a decrease in the relative transcriptamount of the downstream gene spd0637 (hypothetical
Table 2. Microarray analysis of transcript amounts from strain IU1976 (spxR6) compared with its D39 (1U1690) parent grown exponentially inCDMa.
Gene tag spxR6 versus spxR +
Bayesian PSPD_ bspr Fold change Gene description
c0063 0057 -9.5 1.2E-09 strH; Beta-N-acetyl-hexosaminidase precursor0113 0105 -1.8 3.2E-05 Transporter, truncation
0106 -1.9 1.6E-05 Transporter, truncationc0126 0121 2.1 3.4E-03 pspA; Surface protein pspA precursorc0544 0549 -3.5 2.3E-07 Non-catalytic rhodanese-like domainc0636 0642 -7.9 1.4E-09 spxB; Pyruvate oxidasec0637 0643 -2.7 1.4E-07 Glyoxalase family proteinc0704 0710 -4.1 3.5E-08 Peptidase family U320969 0991 -1.8 6.8E-06 Conserved hypothetical proteinc1492 1524 1.9 2.6E-05 Conserved hypothetical proteinc1494 1526 1.9 1.9E-05 ABC transporter membrane-spanning permease – sugar transportc1496 1528 2.1 1.2E-05 Phosphotransferase system sugar-specific EII componentc1504 1536 4.3 7.0E-09 nanA; Sialidase A precursor (neuraminidase A)c1505 1537 4.3 2.0E-07 Hypothetical protein1514 1546 1.9 1.4E-04 ABC transporter ATP-binding protein – unknown substrate1515 1547 2.0 1.3E-05 Hypothetical protein1516 1548 2.1 5.2E-05 Hypothetical protein1517 1549 2.4 8.2E-07 Hypothetical protein1628 1662 -2.0 7.9E-05 xpt; Xanthine phosphoribosyltransferasec1649 1684 2.2 6.2E-04 piuB; iron-compound ABC transporter, permease proteinc1651 1686 3.1 8.7E-05 piuD; iron-compound ABC transporter, ATP-binding protein
a. Strain isolation, growth, and microarray analysis are described in Experimental procedures. Fold changes and Bayesian P-values are basedon three independent biological replicates, including dye swaps. This table shows all the genes with fold changes greater than 1.8. The P-valuesof these genes were all < 0.005. A complete data set is deposited in GEO (accession number GSE9284). Adjacent genes that may form operonsare grouped by lines at the left. Highlighted entries are discussed in the text.b. SPD_ designations are for strain D39. The corresponding spr designations in laboratory strain R6 are shown for convenience.c. Showed comparable fold changes in the separately isolated mutant IU2072 [D39 (IU1690) spxR::Mariner].
736 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746
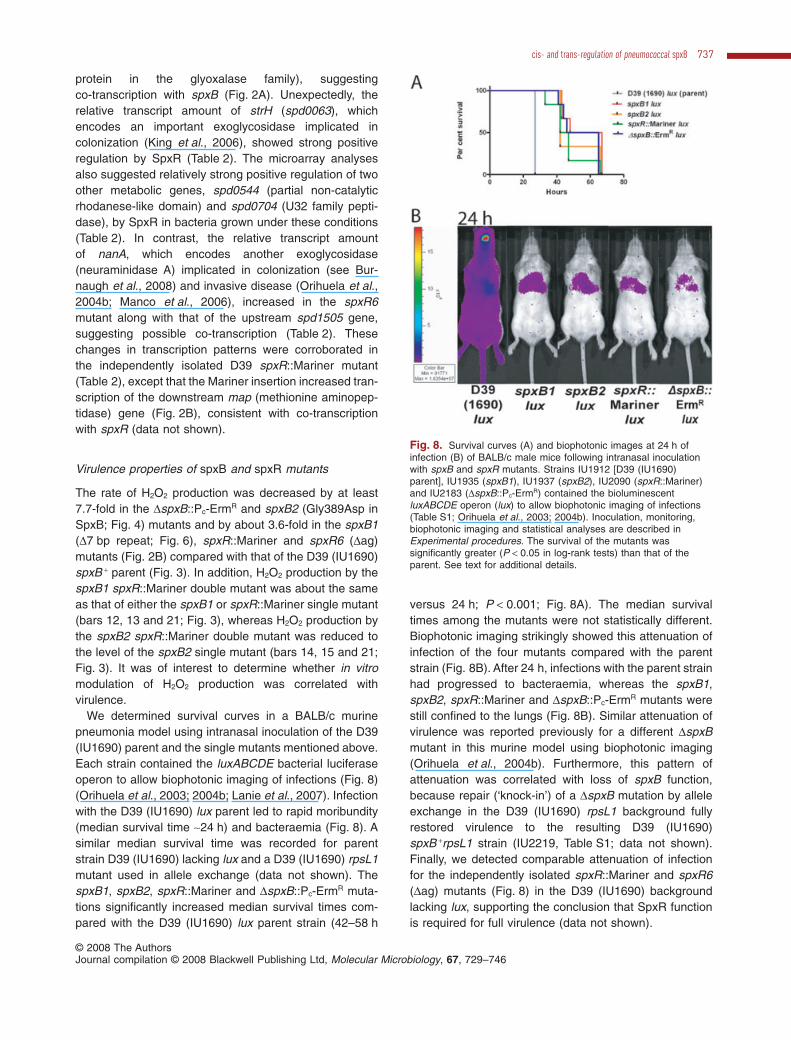
protein in the glyoxalase family), suggestingco-transcription with spxB (Fig. 2A). Unexpectedly, therelative transcript amount of strH (spd0063), whichencodes an important exoglycosidase implicated incolonization (King et al., 2006), showed strong positiveregulation by SpxR (Table 2). The microarray analysesalso suggested relatively strong positive regulation of twoother metabolic genes, spd0544 (partial non-catalyticrhodanese-like domain) and spd0704 (U32 family pepti-dase), by SpxR in bacteria grown under these conditions(Table 2). In contrast, the relative transcript amountof nanA, which encodes another exoglycosidase(neuraminidase A) implicated in colonization (see Bur-naugh et al., 2008) and invasive disease (Orihuela et al.,2004b; Manco et al., 2006), increased in the spxR6mutant along with that of the upstream spd1505 gene,suggesting possible co-transcription (Table 2). Thesechanges in transcription patterns were corroborated inthe independently isolated D39 spxR::Mariner mutant(Table 2), except that the Mariner insertion increased tran-scription of the downstream map (methionine aminopep-tidase) gene (Fig. 2B), consistent with co-transcriptionwith spxR (data not shown).
Virulence properties of spxB and spxR mutants
The rate of H2O2 production was decreased by at least7.7-fold in the DspxB::Pc-ErmR and spxB2 (Gly389Asp inSpxB; Fig. 4) mutants and by about 3.6-fold in the spxB1(D7 bp repeat; Fig. 6), spxR::Mariner and spxR6 (Dag)mutants (Fig. 2B) compared with that of the D39 (IU1690)spxB + parent (Fig. 3). In addition, H2O2 production by thespxB1 spxR::Mariner double mutant was about the sameas that of either the spxB1 or spxR::Mariner single mutant(bars 12, 13 and 21; Fig. 3), whereas H2O2 production bythe spxB2 spxR::Mariner double mutant was reduced tothe level of the spxB2 single mutant (bars 14, 15 and 21;Fig. 3). It was of interest to determine whether in vitromodulation of H2O2 production was correlated withvirulence.
We determined survival curves in a BALB/c murinepneumonia model using intranasal inoculation of the D39(IU1690) parent and the single mutants mentioned above.Each strain contained the luxABCDE bacterial luciferaseoperon to allow biophotonic imaging of infections (Fig. 8)(Orihuela et al., 2003; 2004b; Lanie et al., 2007). Infectionwith the D39 (IU1690) lux parent led to rapid moribundity(median survival time ~24 h) and bacteraemia (Fig. 8). Asimilar median survival time was recorded for parentstrain D39 (IU1690) lacking lux and a D39 (IU1690) rpsL1mutant used in allele exchange (data not shown). ThespxB1, spxB2, spxR::Mariner and DspxB::Pc-ErmR muta-tions significantly increased median survival times com-pared with the D39 (IU1690) lux parent strain (42–58 h
versus 24 h; P < 0.001; Fig. 8A). The median survivaltimes among the mutants were not statistically different.Biophotonic imaging strikingly showed this attenuation ofinfection of the four mutants compared with the parentstrain (Fig. 8B). After 24 h, infections with the parent strainhad progressed to bacteraemia, whereas the spxB1,spxB2, spxR::Mariner and DspxB::Pc-ErmR mutants werestill confined to the lungs (Fig. 8B). Similar attenuation ofvirulence was reported previously for a different DspxBmutant in this murine model using biophotonic imaging(Orihuela et al., 2004b). Furthermore, this pattern ofattenuation was correlated with loss of spxB function,because repair (‘knock-in’) of a DspxB mutation by alleleexchange in the D39 (IU1690) rpsL1 background fullyrestored virulence to the resulting D39 (IU1690)spxB +rpsL1 strain (IU2219, Table S1; data not shown).Finally, we detected comparable attenuation of infectionfor the independently isolated spxR::Mariner and spxR6(Dag) mutants (Fig. 8) in the D39 (IU1690) backgroundlacking lux, supporting the conclusion that SpxR functionis required for full virulence (data not shown).
Fig. 8. Survival curves (A) and biophotonic images at 24 h ofinfection (B) of BALB/c male mice following intranasal inoculationwith spxB and spxR mutants. Strains IU1912 [D39 (IU1690)parent], IU1935 (spxB1), IU1937 (spxB2), IU2090 (spxR::Mariner)and IU2183 (DspxB::Pc-ErmR) contained the bioluminescentluxABCDE operon (lux) to allow biophotonic imaging of infections(Table S1; Orihuela et al., 2003; 2004b). Inoculation, monitoring,biophotonic imaging and statistical analyses are described inExperimental procedures. The survival of the mutants wassignificantly greater (P < 0.05 in log-rank tests) than that of theparent. See text for additional details.
cis- and trans-regulation of pneumococcal spxB 737
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

Discussion
Previous work suggested that spxB transcription isstrongly regulated in vitro and in vivo (Hoffmann et al.,2006; LeMessurier et al., 2006; Weiser, 2006). Normal-ized QPCR assays showed that the relative amount ofspxB transcript is significantly lower in D39 pneumococ-cus sampled from the lungs and blood of mice comparedwith the nasopharynx (LeMessurier et al., 2006). spxBregulation was also implied by the finding that SpxBpyruvate oxidase seems to be expressed at greaterlevels in transparent than opaque phase variants ofS. pneumoniae (Weiser, 2006). A further metabolic link tospxB regulation is suggested by the discovery that H2O2
production by pneumococcus is greatly decreased in aDcarB mutant deficient in carbamoyl phosphate syn-thetase (Hoffmann et al., 2006). However, no mechanismfor spxB regulation has yet been reported. We report herethat spxB transcription is regulated by both cis- andtrans-acting regulatory elements (Figs 2, 6 and 7). Thecis-acting element consists of an adjacent direct repeatlocated immediately upstream of the spxB promoter(Fig. 6). A spontaneous deletion of one repeat unit of thisdirect repeat (spxB1) caused a significant decrease inrelative spxB transcript amount (Fig. 7) and a concomitantdrop in H2O2 production (Fig. 3). Expansion and deletionof adjacent direct repeats were noted previously in acomparison of the complete genome sequences of strainsD39 (IU1690), D39 (IU1680) and R6 (Lanie et al., 2007)and likely occur by a slippage mechanism during replica-tion (Lovett, 2004). A likely hypothesis is that this adjacentdirect repeat binds a trans-acting positive regulatoryprotein that stimulates spxB transcription.
A candidate for this positive regulator is SpxR, whichwas identified in the same genetic screen (Fig. 5) usedto discover the cis-acting element upstream of spxB(Fig. 6). Spontaneous mutations in spxR were mappedby performing parallel transposon mutagenesis to locatespxR and then confirming that the spontaneous muta-tions were indeed located in this new regulatory gene.Three spontaneous spxR mutants were isolated fromscreens on TSA plates lacking (spxR6 and spxR7) orcontaining 1¥ added catalase (spxR3) (Fig. 5; Results);yet, each contains the same Dag deletion in a run of fiveag dinucleotide repeats that causes early truncation ofthe SpxR protein (Fig. 2B; Results). A frameshift muta-tion in a run of repeated bases early in a regulatory orstructural genes is a hallmark of phase variation thatmodulates virulence factor expression in numerous bac-terial pathogens (Moxon et al., 1998; Pericone et al.,2002; Fox et al., 2005). The recovery of at least twoindependent isolates of the same spxR spontaneousmutation (frequency ª 1 ¥ 10-5; Results) is consistentwith a phase variation mechanism. However, this inter-
pretation must be qualified, because pneumococcus pro-duces H2O2 during this screen (see below).
The SpxR protein, which hitherto has been classified asa conserved hypothetical protein, contains several evoca-tive structural features (Fig. 2B). A winged HTH domain inthe GntR family of DNA binding proteins (Rigali et al.,2002) is located at the amino terminus of SpxR. TandemDRTGG-CBS domains follow the HTH domain. TheDRTGG domain is often found associated with CBSdomains, which have been implicated in binding adenosylcompounds, such as AMP and ATP (Kemp, 2004; Scottet al., 2004). Unlike some CBS-domain proteins, SpxRcontains a single copy of the CBS domain instead of twoor four copies. The last domain in SpxR is a member ofthe ‘HotDog’ family, sometimes found associated with theCBS domain (Dillon and Bateman, 2004). The HotDogfold is found in thioesterase enzymes that catalysevarious interconversions of CoA-containing compounds,especially in fatty acid metabolism (Dillon and Bateman,2004). This combination of domains suggests that SpxRregulates spxB transcript amount in response to energyand metabolic state (Fig. 1), rather than oxidative stress.
Consistent with this interpretation, SpxR lacks cysteineresidues that could become oxidized by H2O2 (reviewed inImlay, 2003; Kiley and Storz, 2004). SpxR may detectenergy and metabolic state by binding ATP or AMP to itsCBS domain and acetyl-CoA or another CoA-containingcompound to its HotDog domain (Fig. 1). DspxB mutantsof D39 were reported to contain considerably less acetylphosphate and ATP than their spxB + parent duringaerobic growth in TH+Y medium (Pericone et al., 2003).Yet, in bacteria grown aerobically in BHI broth, the relativeamount of spxB transcript did not change significantly inthe spxB2 or spxB4 point mutants, which produced muchless H2O2 than their parent strain (Figs 2, 3, and 7). We donot know whether ATP pools dropped in these mutants orwere compensated by ATP produced by glycolysis or thePta/AckA pathway under these growth conditions (Fig. 1).Besides extending these experiments to additional growthstates, future experiments will determine whether purifiedSpxR acts directly by binding to the adjacent direct repeatupstream of spxB in response to ATP and other ligands.
NCBI BLAST analyses show that SpxB-like pyruvateoxidases are found in a restricted number of bac-terial species. In the phylum Firmicutes, order Lactobacil-lales, and family Streptococcaceae, spxB is presentin S. pneumoniae, S. oralis (96%), S. mitis (97%),S. sanguinis (97%) and Lactococcus lactis (41%), wherenumbers in parentheses refer to amino acid identity withS. pneumoniae SpxB. The spxB gene is conspicuouslyabsent from some other major pathogens in this family,including S. mutans, S. agalactiae (Group B Strep), andS. pyogenes (Group A Strep). However, S. pyogenes con-tains a homologue of SpxR containing all four of the
738 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

domains found in S. pneumoniae SpxR (Fig. 2B). Pre-sumably, SpxR regulates genes other than spxB inS. pyogenes. Likewise, S. agalactiae and S. mutans,which also lack spxB, contain homologues of SpxRlacking the HotDog domain. Oddly, the SpxR homologuesof L. lactis and S. mitis contain DRTGG-CBS or CBSdomains respectively, but lack HTH DNA bindingdomains. Evidently, the SpxR homologues in these otherspecies of streptococcus do not regulate spxB transcrip-tion directly. Taken together, these phylogenetic compari-sons suggest that the role of SpxR in regulating spxBtranscription, whether directly or indirectly, may be con-fined to S. pneumoniae. Of these full and partial homo-logues, only S. pneumoniae spxR contains a agagagagagslippage site early in the gene. The fragment of SpxRpotentially expressed in spontaneous frameshift mutants,such as spxR3, spxR6 and spxR7, would contain an intactGntR-HTH domain (Fig. 2B). Whether this fragment ofSpxR is expressed awaits further experiments.
Sequence and expression comparisons suggestthat pneumococcal spxB is regulated by fundamentallydifferent mechanisms from its poxB homologue inL. plantarum (Lorquet et al., 2004). Transcription ofL. plantarum poxB is decreased by catabolite repressionin the presence of glucose and increased strongly byaeration (Lorquet et al., 2004). The catabolite repressionis mediated by CcpA protein binding to a cre sitebetween the -35 and -10 regions of the poxB promoter(Hueck and Hillen, 1995), but the mechanism of aerobicinduction is unknown (Lorquet et al., 2004). A putativebinding site for the OhrR repressor, which responds tooxidative stress (Fuangthong and Helmann, 2002), islocated immediately downstream from the poxB -10region; but it is not known whether OhrR plays a directrole in poxB regulation (Lorquet et al., 2004). By com-parison, there is a putative cre site upstream of the adja-cent direct repeat upstream of the spxB promoter(Fig. 6). However, to date, we have not detected strongcatabolite repression of pneumococcal spxB transcrip-tion. BHI broth used in these experiments contains11 mM glucose, which is sufficient to elicit cataboliterepression in S. pneumoniae (Iyer et al., 2005). Basedon signal strengths in the microarray (see GEO entryGSE9284), QPCR, and primer extension experimentsreported here, as well as a previous report (LeMessurieret al., 2006), spxB mRNA is one of the more abundanttranscripts in D39 (IU1690) growing in BHI broth in anatmosphere of 5% CO2. Moreover, S. pneumoniae lackshomologues of the oxidative stress regulators PerR andOhrR found in Bacillus subtilis and OxyR found in Gram-negative bacteria (Pericone et al., 2003).
Microarray analyses led to several conclusions aboutthe effects of spxB and spxR null mutations on globaltranscription patterns (Results; Table 2). Unexpectedly, a
large drop (approximately eightfold; Fig. 3) in relativeH2O2 production in the DspxB::Pc-ErmR mutant was notaccompanied by strong relief of an oxidative stressresponse (Results). At most, there was a marginal(approximately twofold) response in relative transcriptamounts from some genes, including recurringdecreases in the relative transcript amounts of severalgenes associated with oxidative stress, such as psaD[thiol peroxidase; (Spd1464)], and sufS (cysteine desul-phurase subfamily protein (Spd0765), possibly involvedin maintaining iron-sulphur centres). Our samples weretaken in early and mid-exponential phase, and it is pos-sible that stronger effects might be detected instationary-phase cells. The marginal changes in relativetranscript amounts also did not support the idea thatacetyl phosphate synthesized by SpxB broadly modu-lates regulation by two-component regulatory systems.Deletion of spxB has been reported to reduce cellularacetyl phosphate amount by as much as 85% in sero-type 2 strains related to those used in this study (Peri-cone et al., 2003). Yet, this presumed large drop inacetyl phosphate level was not accompanied by analtered transcription pattern indicative of a generalchange in the phosphorylation states of those responseregulators whose regulons have been characterized inprevious microarray studies (reviewed in Paterson et al.,2006). One qualification to this interpretation is that theamount of acetyl phosphate remaining in a DspxBmutant may still be relatively high. A simple calculationbased on the data in Pericone et al. (2003) and anassumed cell volume of 0.5 ¥ 10-15 l per cell in a diplo-coccus pair suggests that the concentration of acetylphosphate remaining in a DspxB mutant (~9 mM) maybe comparable to that (~10 mM) which causes full phos-phorylation of the purified VicR response regulator (Nget al., 2005).
Our microarray analyses showed that the SpxR regulonincludes comparatively few genes (Table 2). As expectedfrom QPCR analyses (Fig. 7), the relative amount of thespxB transcript dropped greatly in the spxR6 or spxR::Ma-riner mutant (Table 2). Unexpectedly, the relative tran-script amount of the strH gene showed a large drop in thespxR mutants. Regulation by SpxR suggests that expres-sion of the StrH exoglycosidase is coupled to pneumo-coccal energy and metabolic state (Fig. 1). StrH releasessugars from glycoproteins on the surfaces of human cells(see Clarke et al., 1995; King et al., 2006), and a recentreport shows that these sugars can be used as a carbonsource for S. pneumoniae growing in culture and possiblyduring persistence in the airway (Burnaugh et al., 2008).An adjacent direct repeat sequence (atagaaatagaa)related to the one that precedes spxB (Fig. 6) is located172 bp upstream of the strH reading frame and may beanother site for SpxR binding.
cis- and trans-regulation of pneumococcal spxB 739
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

Besides spxB and strH, SpxR may directly or indirectlyregulate several other metabolic genes. The relative tran-script amounts of nanA (spd1504; neuraminidase A) andthe gene upstream from nanA (spd1505; hypotheticalprotein) increased in the spxR mutants, suggesting nega-tive coregulation. This result seems at odds with thefinding that NanA acts sequentially before StrH in thedegradation of glycoproteins (King et al., 2006; Burnaughet al., 2008). In addition, in serotype 6 strains, the relativetranscript amount from nanA was increased in a transpar-ent phase variant (King et al., 2004), whereas the spxRmutants are opaque variants of the serotype 2 strain usedin this study (Results). Besides possible serotype differ-ences, the opposite regulation of strH and nanA mayreflect the fact that there is a redundant neuraminidase(NanB) in S. pneumoniae (Manco et al., 2006; Burnaughet al., 2008), which was not detected in the SpxR regulon(Table 2). Of the remaining genes that are positively regu-lated by SpxR under this growth condition (Table 2),spd0704 was noteworthy, because it encodes a putativeU32 family peptidase related to collagenase.
The genetic screen used in this study (Fig. 5) suggeststhat opaque colonies of serotype 2 strain D39 arise by atleast two different mechanisms. The predominant mecha-nism (six of seven mutants) involved changes in the func-tion or regulation of the SpxB pyruvate oxidase, which hasbeen repeatedly linked to pneumococcal phase variation(Overweg et al., 2000; Pericone et al., 2002; Belangeret al., 2004; Weiser, 2006). The frequency of opaquecolony formation (~3 ¥ 10-5) was surprisingly high andmuch greater than the spontaneous mutation frequency atother genetic loci. We determined that the mutation fre-quency for the development of rifampicin or optochinresistance was 3.8 � 0.8 ¥ 10-8 and 5.7 � 0.8 ¥ 10-8,respectively, for strain D39 (IU1690) grown in TSB andspread onto blood agar plates containing these com-pounds (Experimental procedures). In contrast to anotherreport (Pericone et al., 2002), we did not detect largereductions in the mutation frequency of isogenicDspxB::Pc-ErmR mutant IU2181 (4.0 � 0.6 ¥ 10-8 and7.6 � 0.5 ¥ 10-8 for rifampicin and optochin resistancerespectively) compared with the spxB + parent. However,there were differences in some experimental details; forexample, we sampled exponentially growing instead ofstationary cultures (Pericone et al., 2002). We also did notdetect a significant difference in the number of viable cellsof the D39 (IU1690) spxB + parent and its DspxB::Pc-ErmR
derivative (IU2181) grown exponentially in TSB andplated onto blood agar (data not shown). This result isconsistent with reports showing that viability of spxBmutants is primarily lost during prolonged stationaryphase (Regev-Yochay et al., 2007).
The main difference between our experiments to deter-mine mutation frequency or identify colony morphology
variants was the plating medium. In the former, blood agar,which lessens effects of H2O2 production by pneumococ-cus (Weiser et al., 1994; Belanger et al., 2004), was used,whereas in the latter, TSA was used lacking or containing asuboptimal (1¥) concentration of added catalase (Fig. 5).Apossible explanation for the high frequency of opaquecolony formation in the morphology screen is that most ofthese mutants arose on the TSA plates and not in the liquidTSB cultures before plating. Pneumococcal colonies thatgrow to stationary phase and beyond on TSA plates mayaccumulate considerable H2O2 that can damage DNA,increase mutation rate, and induce global stress. Despitetheir sensitivity to H2O2 in some growth conditions (Peri-cone et al., 2003; Regev-Yochay et al., 2007), mutantswith decreased spxB expression survived and producedcolonies in this selection. Consistent with these ideas, weattempted to screen in the opposite direction for smallersemitransparent variants of starting opaque mutantsspxB1 (D7 direct repeat), spxR3 (Dag), spxR6 (Dag) andspxR7 (Dag). We screened more than 100 000 colonies ofeach strain (~600 000 colonies totally) and failed to find asingle transparent variant (data not shown). Thus, themutation frequency in the opaque → transparent directionwas less than 1 ¥ 10-6, possibly due to reduced H2O2
production by these mutants on the TSA plates.The cis-acting spxB1 and trans-acting spxR::Mariner
and spxR6 mutations reduced relative H2O2 production byabout 3.6-fold compared with the 7.7-fold drop caused bythe spxB2 and DspxB::Pc-ErmR mutations (Fig. 3), but allfive mutants showed a comparable reduction in virulencein a murine infection model (Results; Fig. 8). Thus, spxRacts as a virulence factor through its strong positive regu-lation of spxB, which had already been established as animportant virulence factor in single-strain murine infectionmodels (Orihuela et al., 2004b; Orihuela and Toumanen,2006).
Finally, the findings presented here do not fully supporta recent conjecture that pneumococcal spxB is strictly a‘suicide’ gene maintained by selective advantage in colo-nization of the nasopharynx (Regev-Yochay et al., 2007).This conjecture was partly based on the observation thata DspxB mutant did not exhibit reduced colonization in aninfant rat model (Regev-Yochay et al., 2007). However, inother single-strain infection models, DspxB mutants areclearly attenuated (Fig. 8; Orihuela et al., 2004b; Orihuelaand Toumanen, 2006). In addition, our discovery thatspxB transcription is strongly regulated by the SpxRprotein, which likely senses energy and metabolic state,argues that SpxB plays a prominent role in the metabo-lism of S. pneumoniae, probably through production ofATP (Fig. 1; Pericone et al., 2003). On the other hand, ourresults do support the previous observations that spxBmutants are hypersensitive to H2O2 (Results) and wouldnot likely accumulate in dense, stationary-phase cultures
740 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

of spxB + bacteria (Pericone et al., 2003; Regev-Yochayet al., 2007).
Experimental procedures
Bacterial strains and growth conditions
Strains used in this study are listed in Table S1. Strains weregrown on plates containing Trypticase Soy Agar II (Modified)(Becton-Dickinson; BD) and 5% (v/v) defibrinated sheepblood (TSAII BA) or TSB (BD) and 1% (wt/vol) Bacto Agar(Difco) (TSA). Plates were incubated at 37°C for the timesindicated in an atmosphere of 5% CO2, unless otherwiseindicated. Bacteria were cultured statically in BHI broth (BD),a CDM (formulated by JRH Biosciences; Talkington et al.,1992), or TSB at 37°C in an atmosphere of 5% CO2. Growthwas monitored by optical density (OD620) using a Spectronic20 spectrophotometer fitted for measurement of cappedtubes with an outer diameter of 16 mm. Bacteria were inocu-lated into BHI or TSB, serially diluted into the same medium,and propagated overnight. Overnight cultures that were still inexponential phase (OD620 ª 0.2–0.4) were diluted back to adensity of OD620 = 0.005 to start final cultures, which did notcontain antibiotics. For growth in CDM, overnight cultureswere first propagated in BHI and collected by microcentrifu-gation for 2 min at room temperature. Pellets were washed,resuspended and diluted in CDM to a density of OD620 = 0.01to start final cultures.
Construction and verification of S. pneumoniae mutants
Strains containing antibiotic markers were constructed bytransforming linear DNA amplicons synthesized by overlap-ping fusion PCR into competent pneumococcal cells asdescribed previously (Robertson et al., 2002; Ng et al.,2003). For antibiotic selections, TSAII BA plates were supple-mented with 250 mg kanamycin per ml, 100 mg spectinomycinper ml, 150 mg streptomycin per ml, or 0.3 mg erythromycinper ml. Primers synthesized for this study are listedin Table S2. The exchange of spxB and spxR alleles lackingantibiotic markers was carried out using the Janus method ofallelic replacement (Sung et al., 2001) in rpsL1 (StrR) mutantsEL908, IU1215 and IU1781 (Table S1). DspxB mutantscontain an in-frame deletion of amino acids 52–518 of SpxB(out of 591 amino acids totally) or the same deletion with aPc-ErmR cassette inserted (Fig. 2; Table S1). All constructswere confirmed and mutations located by DNA sequencing ofthe spxB and spxR regions (Fig. 2). DNA was prepared fromcell lysates and amplified using Pfu Turbo Polymerase (Strat-agene) and the primers indicated in Fig. 2. Amplicons weregel purified and sequenced in reaction mixtures thatcontained 1 ml of Big Dye Terminator reagent (AppliedBiosystems) as described previously (Lanie et al., 2007).Sequences were aligned and analysed using the Vector NTIprogram (Invitrogen).
Observation of colony morphology on TSA plates withor without catalase
Catalase (Worthington Biochemical Corporation) wasfreshly added in a volume of 20–30 ml to wells (21 mm
diameter) of a microtiter plate containing TSA and allowedto just absorb into the agar. 1¥ catalase (88.2 U per cm2
well surface) refers to the concentration used previously instudies of phase variation (Weiser et al., 1994). 5¥ catalasewas 441 U per cm2 well surface. In control experiments,catalase was denatured by heating at 95°C to confirm theactivity of the native enzyme. R6 and D39 strains weregrown exponentially in TSB to OD620 ª 0.1 and seriallydiluted such that 20 ml was added to each well. After 20 h ofincubation at 37°C, colonies were examined and photo-graphed on a dissecting microscope with oblique light and acalibrated scale. To examine colony morphology duringanaerobic growth, plates were incubated at 37°C in a GasPak Plus anaerobic system (BBL) containing envelopes withpalladium catalyst which generates 4.6–6.5% carbondioxide and < 0.2% oxygen in 100 min, according to infor-mation supplied by the manufacturer.
Isolation of spontaneous opaque mutants
Spontaneous opaque mutants were isolated in strain D39(IU1690) based on Weiser et al. (1994). Cells were grownexponentially in TSB to OD620 = 0.1–0.2 and serially dilutedso that ~400 cfu in 100 ml were spread onto each TSA plate(100 mm diameter) lacking or containing 1¥ catalase (5000 Uper plate). After 18–21 h, colonies were observed under adissecting microscope. Opaque colonies were single colonyisolated and stored for further study. Typically, 20 plates werescreened in each experiment, and the average number of cfuplated was based on counting and averaging the cfu on threeplates.
H2O2 release assays
The rate of H2O2 production was determined by a modificationof the protocol provided in the Amplex Red HydrogenPeroxide/Peroxidase Assay kit (Invitrogen) and Belangeret al. (2004). One hundred microlitres of reaction mixture(100 mM Amplex Red reagent, 0.2 U horseradish peroxidaseper ml in 0.05 M sodium phosphate buffer, pH 7.4) was dis-pensed into wells of a 96-well microtiter plate and warmed to37°C for 10 min. Exponential cultures were grown in BHI toOD620 = 0.1–0.15, centrifuged briefly in a microfuge, andresuspended in fresh BHI to OD620 ª 0.06. Twenty microlitresof freshly resuspended cells was added in duplicate or tripli-cate to 100 ml of pre-warmed reaction mixture. Control wellscontained 20 ml of BHI as a blank or a series of H2O2 con-centration standards diluted in BHI. Microtiter plates wereincubated at 37°C in a VERSAmax tunable reader driven bySoftMax Pro software (Molecular Devices) and absorbancewas read at 563 nm every 3 min for 1 h. Rates of H2O2
production in pmole per min were calculated and normalizedto the OD620 of the cell suspensions. Final values are shownrelative to that of strain D39 (IU1690), which was included inevery experiment as an internal standard (Fig. 3). The rateof H2O2 production for D39 (IU1690) (n = 25) was240 � 15 pmol per min per 20 ml of 0.1 OD620 cell suspen-sion, and the basal rate of H2O2 production in D39 (1690)DspxB::Pc-ErmR (IU2181) was 13 � 2% that of the D39(IU1690) parent strain (Fig. 3). Comparable results were
cis- and trans-regulation of pneumococcal spxB 741
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

obtained for the D39 (IU1690) rpsL1 parent and D39(IU1690) rpsL1 DspxB mutant (data not shown).
Hydrogen peroxide sensitivity assays
Hydrogen peroxide sensitivity assays were performed onbacterial cells grown exponentially in BHI to OD620 = 0.25 asdescribed in Pericone et al. (2003), except that samples wereserially diluted in physiological saline solution (0.9% (wt/vol)NaCl) at room temperature, instead of cold phosphate-buffered saline (PBS).
Molecular modelling
The amino acid sequences of the Pox and SpxB pyruvateoxidases from L. plantarum and S. pneumoniae D39 (1690),respectively, were aligned by using ExPASy MolecularBiology software. The crystal structure of L. plantarum Pox(PDB code, 1POX) (Muller et al., 1994) was used as thetemplate for homology modelling of pneumococcal SpxB.Amino acids that were different in the two sequences wereexchanged using the Coot molecular graphics program. Themost frequent side-chain conformations were selected andlocal energy minimization was applied for each replacementin Coot.
Rifampicin and optochin resistance assays
The frequencies of spontaneous mutation to rifampicin andoptochin resistance were determined based on Periconeet al. (2002) and Meier et al. (2003). Strains D39 (IU1690)and IU2181 (DspxB::Pc-ErmR) were grown exponentially inTSB to OD620 = 0.2 and 0.3. One hundred microlitres of cul-tures was spread directly in triplicate onto TSAII BA platescontaining 0.05 mg rifampicin per ml or 4 mg optochin per ml(both from Sigma). Total cfu were determined at each OD620
by serially diluting cultures into PBS at room temperature andspreading samples onto TSAII BA plates. Plates were scoredfor optochin or rifampicin-resistant colonies after 24 or 48 hrespectively.
Isolation of pneumococcal genomic DNA
Streptococcus pneumoniae DNA for transposon mutagenesisand inverse PCR was isolated from 5 ml of cells grown in BHIbroth to OD620 = 0.5–0.6 using a Master Pure Gram PositiveDNA Purification Kit (Epicentre) with the followingmodifications. Two microlitres of Ready Lyse™ Lysozymewas added to the resuspended pellet, and the mixture wasincubated for 45 min at 37°C. The yield of DNA was ~1 mg perml of starting culture.
In vitro mariner transposon mutagenesis
Marc9 transposase was purified and transposition reactionswere performed as described previously (Martin et al., 2000;Hava and Camilli, 2002) with the following modifications.Transposition reaction mixtures contained 21 ng of purifiedMarc9 transposase (Lampe et al., 1996), 500 ng of target
genomic DNA from strain D39 (IU1690), and 40 ng of purifiedpR412 plasmid DNA containing the Magellan5 (SpecR)mini-transposon (Martin et al., 2000). Gaps in transpositionproducts were repaired using T4 DNA polymerase andEscherichia coli DNA ligase (New England Biolabs) asdescribed previously (Hava and Camilli, 2002). Reaction mix-tures were mixed with 250 ml of strain R6 exponentially grownto OD620 = 0.3 and 750 ml of BHI containing 10% (v/v) heat-inactivated horse serum (Sigma), 10 mM glucose and 100 ngper ml of synthetic competence stimulating peptide-1(CSP-1) [generously provided by D.A. Morrison (UIC)] (Rob-ertson et al., 2002; Ng et al., 2003). Strain R6 was used asthe recipient for initial transformations because of its highertransformation frequency compared with strain D39 (IU1690)(see Belanger et al., 2004; Lanie et al., 2007). Transforma-tion mixtures were incubated for 2 h at 37°C, mixed with 3 mlnutrient soft agar, and poured onto TSAII BA plates contain-ing 100 mg spectinomycin per ml. Transformants werepatched onto TSAII BA + spectinomycin plates in the gridpattern created by a 48-prong transfer device (Sigma) andthen patched from the grid onto TSA plates. Patches wereincubated for 18–24 h before being examined under a dis-section microscope and scored for larger size and increasedopacity.
Localization of transposon insertion
Inverse PCR was carried out as described in Sambrook andRussell (2001) with the following additions. Genomic DNAfrom R6 spxR::Mariner was digested overnight at 37°C withthe restriction enzyme NheI (NEB). Ligation reactions con-taining 10, 30, 60 or 100 ng of digested DNA and 1 ml of T4DNA ligase (NEB) in a total volume of 20 ml were incubated at16°C for 18 h. Ligation reactions were used as the DNAtemplate for PCR reactions using primers Mariner Fwd andMariner Rev (Table S2). All PCR reactions produced a promi-nent 5 kb product that was gel purified and used as thetemplate in DNA sequencing reactions. The exact location ofthe spxR::Mariner insertion was determined by PCR amplify-ing and sequencing the spxR locus of the mutant. The spxR-::Mariner insertion was PCR amplified from strain R6 andtransformed into strain D39 (IU1690) (Robertson et al., 2002;Ng et al., 2003). The resulting opaque mutant was character-ized further as described in Results.
RNA extraction and QPCR analyses
Total pneumococcal RNA was prepared from cells growingexponentially in BHI or CDM broth to OD620 = 0.2 by a rapidlysis procedure followed by purification using the RNAeasyminikit (Qiagen) as described previously (Robertson et al.,2002; Ng et al., 2005), with the inclusion of on-column treat-ment with DNase I (Qiagen). 5 mg of total RNA was furtherdigested using a DNA-Free kit (Ambion), diluted to about20 ng per ml, and quantified using a Quant-iT RNA assay kit(Molecular Probes). QPCR was performed with a two-stepPCR protocol. First-strand synthesis was performed in 20 mlcontaining 100 ng of RNA, 3 ml of random primers (0.1 mgper ml), 0.8 ml of dNTP mix (25 mM of each dNTP), and 1 mlof StrataScript reverse transcriptase (50 U per ml) in first-
742 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

strand buffer (Stratagene). Reactions lacking reverse tran-scriptase were prepared in parallel to control for possibleDNA contamination. cDNA from first-strand reactions wasdiluted eightfold and serially diluted in fourfold steps two orthree times more. Each PCR reaction contained 12.5 ml of2¥ Brilliant SYBR Green QPCR master mix (Stratagene),2 ml each of 2 mM PCR primers spxB6F and spxB6R(Table S2), 0.38 ml of a 1:500 dilution of the reference dyeprovided in the Brilliant SYBR kit, and 8 ml of diluted cDNAfrom the first strand reaction. 16S ribosomal RNA wasquantified using the same cDNA samples used for spxBtranscripts, except that rRNA cDNA samples were diluted100-fold further and primers KK387 and KK388 were used(Table S2). QPCR was performed using a StratageneMx3000P instrument and analysed with the SYBR Green(with dissociation curve) program. Four different dilutions ofcDNA from strain D39 (IU1690) were used to generate stan-dard curves. spxB transcript amounts were normalized to16S rRNA amounts and are the averages (�SEM) for atleast two independent RNA preparations and QPCR deter-minations (Fig. 7).
Microarray analysis
Microarray analyses were performed comparing relative tran-script amounts for strains D39 (IU1690) with those of strainsIU2181 (DspxB::Pc-ErmR), IU1976 (spxR6), and IU2072(spxR::Mariner) grown in BHI or CDM (Tables S1 and 2 anddata not shown). Total RNA was prepared from exponentiallygrowing cultures at OD620 = 0.2 as described previously (Nget al., 2005; Lanie et al., 2007). S. pneumoniae microarrayswere purchased from Ocimum Biosolutions. Synthesis, label-ling, hybridization, scanning and analysis using the Cyber-Tweb interface were performed as described previously (Nget al., 2005; Lanie et al., 2007) with the exception of thenormalization software used. Data were normalized withoutbackground subtraction by the global Lowess method usingBASE (BioArray Software Environment, http://iubase.cgb.indiana.edu), excluding empty wells and Arabidopsis controlspots. Intensity and expression ratio data for all transcriptsare deposited in the GEO database (accession numberGSE9284).
Primer extension assays
Primer extension mapped the spxB transcription startpoints for strains D39 (IU1690), IU1821 (spxB1), IU1823(spxB2), IU2143 (spxR3) and IU2072 (spxR::Mariner). TotalRNA was prepared as described above for the microarrayanalysis. Primer extension was performed using thePrimer Extension System-AMV Reverse Transcriptase kit(Promega Corp.) according to the manufacturer’s instruc-tions with the following modifications. Ten microlitres ofprimer labelling reactions containing 3.3 pmol of primerspxB7R (Fig. 2A; Table S2), 5 pmol of [g-32P]-ATP (6000 Ciper mmol, 10 mCi per ml), and 10 U of T4 polynucleotidekinase were incubated at 37°C for 30 min and then heatedto 90°C for 10 min to inactivate the enzyme. One microlitreof a 5-fold dilution of the labelled primer and 2 mg or6–10 mg of total RNA from strain D39 (IU1690) or the other
strains listed above, respectively, were used in extensionreactions. A sequencing ladder of the PCR amplicon syn-thesized with primers spxB7F and spxB7R was generatedby using the Sequenase PCR Product Sequencing kit (USBCorp.) and primer spxB7R (Fig. 2A; Table S2).
Murine pneumonia model of infection
All procedures were approved in advance by the InstitutionalAnimal Care and Use Committee and were performedaccording to recommendations of the National ResearchCouncil (National, 1996). Male BALB/c mice (4–5 weeks;Harlan) were maintained in a biosafety level 2 animal facility(70°F, 12 h light/dark). Mice were anaesthetized by inhaling4% isoflourane (Butler Animal Health Supply) delivered byan EZAnesthesia system (Euthanex Corp.) for 5 min. Inmost experiments, six mice were inoculated intranasally witheach bacterial strain to be tested. Six 1 ml aliquots of eachbacterial culture growing exponentially in BHI broth(OD620 = 0.45–0.48) were microcentrifuged for 2 min at13 500 g, and cell pellets were each resuspended in 60 ml ofPBS and combined for a total volume of 360 ml. A 50 mlaliquot of this suspension (~3.8 ¥ 108 cfu) was used as theinoculum. Anaesthetized mice were placed on their backs,and mouths were gently closed to allow inhalation of the 50 mlinoculum, which was delivered dropwise to the centre ofnoses. To ensure inhalation, mice were suspended verticallyafter inoculation for about 1 min until they awoke fromanaesthesia. CFUs used in inocula were confirmed by serialdilution and plating. Mice were monitored at 8 h intervals.Death was not used as an end-point. Moribund mice wereeuthanized by CO2 asphyxiation and that time-point was usedas ‘time of death’ in survival curves. For each strain, 10 ml ofblood was collected from the tail of one mouse and streakedfor single colonies to monitor colony morphology and checkfor revertants. No reversion was observed for any of themutants. Kaplan-Meir survival curves and log-rank testswere generated using GraphPad Prism software. Images ofmice infected with bioluminescent bacteria containing theluxABCDE operon (Table S1; Results) were obtained at 8 hintervals as described in Orihuela et al. (2003; 2004b). Inde-pendent confirmatory experiments were performed for mostof the bacterial mutants.
Acknowledgements
We thank the Indiana Institute for Molecular Biology foraccess to a DNA sequencing facility and the IUB Center forGenomics and Bioinformatics (CGB) for access to microarrayanalysis equipment. We thank Rebecca Lohmeyer for helpwith screening, Lilly Research Laboratories, Gregory Robert-son (Cumbre, Inc.), and Janet Yother (UAB) for bacterialstrains, Andrew Camilli (Tufts) for providing the Mariner trans-poson system, and Donald Morrison (UIC) for providing syn-thetic CSP and bacterial strains. We thank Dan Kearns andWai-Leung Ng for discussions. This work was supported byGrant AI060744 from the National Institutes of Health toM.E.W. and by funds from the Indiana METACyt Initiative ofIndiana University Bloomington, funded in part through amajor grant from the Lilly Endowment, Inc. to M.E.W.
cis- and trans-regulation of pneumococcal spxB 743
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

References
Appelbaum, P.C. (2002) Resistance among Streptococcuspneumoniae: implications for drug selection. Clin Infect Dis34: 1613–1620.
Barry, J.M. (2005) The Great Influenza: The Epic Story of theDeadliest Plague in History. New York: Penguin.
Belanger, A.E., Clague, M.J., Glass, J.I., and Leblanc, D.J.(2004) Pyruvate oxidase is a determinant of Avery’s roughmorphology. J Bacteriol 186: 8164–8171.
Briles, D.E., Paton, J.C., Swiatlo, E., and Crain, M.J. (2006)Pneumococcal vaccines. In Gram-Positive Pathogens. Fis-chetti, V.A., Novick, R.P., Ferretti, J.J., Portnoy D.A., andRood, J.I. (eds). Washington DC: American Society forMicrobiology Press, pp. 289–298.
Burnaugh, A.M., Frantz, L.J., and King, S.J. (2008) Growth ofStreptococcus pneumoniae on human glycoconjugates isdependent upon the sequential activity of bacterialexoglycosidases. J Bacteriol 190: 221–230.
Clarke, V.A., Platt, N., and Butters, T.D. (1995) Cloning andexpression of the beta-N-acetylglucosaminidase gene fromStreptococcus pneumoniae. Generation of truncatedenzymes with modified aglycon specificity. J Biol Chem270: 8805–8814.
DeYulia, G.J., Jr, Carcamo, J.M., Borquez-Ojeda, O., SheltonC.C., and Golde, D.W. (2005) Hydrogen peroxide gener-ated extracellularly by receptor–ligand interaction facilitatescell signaling. Proc Natl Acad Sci USA 102: 5044–5049.
Dillon, S.C., and Bateman, A. (2004) The Hotdog fold: wrap-ping up a superfamily of thioesterases and dehydratases.BMC Bioinformatics 5: 109.
Fox, K.L., Yildirim, H.H., Deadman, M.E., Schweda, E.K.,Moxon, E.R., and Hood, D.W. (2005) Novel lipopolysac-charide biosynthetic genes containing tetranucleotiderepeats in Haemophilus influenzae, identification of agene for adding O-acetyl groups. Mol Microbiol 58: 207–216.
Fuangthong, M., and Helmann, J.D. (2002) The OhrR repres-sor senses organic hydroperoxides by reversible formationof a cysteine-sulfenic acid derivative. Proc Natl Acad SciUSA 99: 6690–6695.
Hava, D.L., and Camilli, A. (2002) Large-scale identificationof serotype 4 Streptococcus pneumoniae virulence factors.Mol Microbiol 45: 1389–1406.
Hava, D.L., LeMieux, J., and Camilli, A. (2003) From nose tolung: the regulation behind Streptococcus pneumoniaevirulence factors. Mol Microbiol 50: 1103–1110.
Hiller, N.L., Janto, B., Hogg, J.S., Boissy, R., Yu, S., Powell,E., et al. (2007) Comparative genomic analyses of seven-teen Streptococcus pneumoniae strains: insights into thepneumococcal supragenome. J Bacteriol 189: 8186–8195.
Hoffmann, O., Zweigner, J., Smith, S.H., Freyer, D., Mahr-hofer, C., Dagand, E., et al. (2006) Interplay of pneumo-coccal hydrogen peroxide and host-derived nitric oxide.Infect Immun 74: 5058–5066.
Hoskins, J., Alborn, W.E., Arnold, J., Jr, Blaszczak, L.C.,Burgett, S., DeHoff, B.S., et al. (2001) Genome of thebacterium Streptococcus pneumoniae strain R6.J Bacteriol 183: 5709–5717.
Huang, M., Oppermann-Sanio, F.B., and Steinbuchel, A.(1999) Biochemical and molecular characterization of the
Bacillus subtilis acetoin catabolic pathway. J Bacteriol 181:3837–3841.
Hueck, C.J., and Hillen, W. (1995) Catabolite repression inBacillus subtilis: a global regulatory mechanism for thegram-positive bacteria? Mol Microbiol 15: 395–401.
Imlay, J.A. (2003) Pathways of oxidative damage. Annu RevMicrobiol 57: 395–418.
Iyer, R., Baliga, N.S., and Camilli, A. (2005) Catabolite controlprotein A (CcpA) contributes to virulence and regulationof sugar metabolism in Streptococcus pneumoniae.J Bacteriol 187: 8340–8349.
Janoff, E.N., and Rubins, J.B. (1997) Invasive pneumococcaldisease in the immunocompromised host. Microb DrugResist 3: 215–232.
Janoff, E.N., and Rubins, J.B. (2004) Immunodeficiency andinvasive pneumococcal disease. In The Pneumococcus.Tuomanen, E.I., Mitchell, T.J., Morrison, D.A., and Spratt,B.G. (eds). Washington, DC: American Society for Micro-biology Press, pp. 252–280.
Jedrzejas, M.J. (2004) Extracellular virulence factors ofStreptococcus pneumoniae. Front Biosci 9: 891–914.
Kemp, B.E. (2004) Bateman domains and adenosine deriva-tives form a binding contract. [comment]. J Clin Invest 113:182–184.
Kiley, P.J., and Storz, G. (2004) Exploiting thiol modifications.PLoS Biol 2: e400.
King, S.J., Hippe, K.R., Gould, J.M., Bae, D., Peterson, S.,Cline, R.T., et al. (2004) Phase variable desialylation ofhost proteins that bind to Streptococcus pneumoniae invivo and protect the airway. Mol Microbiol 54: 159–171.
King, S.J., Hippe, K.R., and Weiser, J.N. (2006) Deglycosy-lation of human glycoconjugates by the sequential activi-ties of exoglycosidases expressed by Streptococcuspneumoniae. Mol Microbiol 59: 961–974.
Klugman, K.P. (2004) Clinical relevance of antibiotic resis-tance in pneumococcal infections. In The Pneumococcus.Tuomanen, E.I., Mitchell, T.J., Morrison, D.A., and Spratt,B.G. (eds). Washington DC: American Society for Microbi-ology Press, pp. 331–338.
Kuboniwa, M., Tribble, G.D., James, C.E., Kilic, A.O., Tao, L.,Herzberg, M.C., et al. (2006) Streptococcus gordonii uti-lizes several distinct gene functions to recruit Porphyromo-nas gingivalis into a mixed community. Mol Microbiol 60:121–139.
Lacks, S.A. (1997) Cloning and expression of pneumococcalgenes in Streptococcus pneumoniae. Microb Drug Resist3: 327–337.
Lampe, D.J., Churchill, M.E., and Robertson, H.M. (1996) Apurified mariner transposase is sufficient to mediate trans-position in vitro. [erratum appears in EMBO J 1997 July 1;16 (13): 4153]. EMBO J 15: 5470–5479.
Lanie, J.A., Ng, W.L., Kazmierczak, K.M., Andrzejewski,T.M., Davidsen, T.M., Wayne, K.J., et al. (2007) Genomesequence of Avery’s virulent serotype 2 strain D39 of Strep-tococcus pneumoniae and comparison with that of unen-capsulated laboratory strain R6. J Bacteriol 189: 38–51.
LeMessurier, K.S., Ogunniyi, A.D., and Paton, J.C. (2006)Differential expression of key pneumococcal virulencegenes in vivo. Microbiology 152: 305–311.
Lopez, R., and Garcia, E. (2004) Recent trends on themolecular biology of pneumococcal capsules, lytic
744 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

enzymes, and bacteriophage. FEMS Microbiol Rev 28:553–580.
Lorquet, F., Goffin, P., Muscariello, L., Baudry, J.B., Ladero,V., Sacco, M., et al. (2004) Characterization and functionalanalysis of the poxB gene, which encodes pyruvateoxidase in Lactobacillus plantarum. J Bacteriol 186: 3749–3759.
Lovett, S.T. (2004) Encoded errors: mutations and rearrange-ments mediated by misalignment at repetitive DNAsequences. Mol Microbiol 52: 1243–1253.
McCormick, A.W., Whitney, C.G., Farley, M.M., Lynfield, R.,Harrison, L.H., Bennett, N.M., et al. (2003) Geographicdiversity and temporal trends of antimicrobial resistance inStreptococcus pneumoniae in the United States.[seecomment]. Nat Med 9: 424–430.
Manco, S., Hernon, F., Yesilkaya, H., Paton, J.C., Andrew,P.W., and Kadioglu, A. (2006) Pneumococcal neuramini-dases A and B both have essential roles during infection ofthe respiratory tract and sepsis. Infect Immun 74: 4014–4020.
Martin, B., Prudhomme, M., Alloing, G., Granadel, C., andClaverys, J.P. (2000) Cross-regulation of competencepheromone production and export in the early control oftransformation in Streptococcus pneumoniae. Mol Micro-biol 38: 867–878.
Meier, P.S., Utz, S., Aebi, S., and Muhlemann, K. (2003)Low-level resistance to rifampin in Streptococcuspneumoniae. Antimicrob Agents Chemother 47: 863–868.
Mitchell, T.J. (2004) Pneumolysin and other virulenceproteins. In The Pneumococcus. Tuomanen, E.I., Mitchell,T.J., Morrison, D.A., and Spratt, B.G. (eds). WashingtonDC: American Society for Microbiology Press, pp. 61–74.
Moxon, E.R., Lenski, R.E., and Rainey, P.B. (1998) Adaptiveevolution of highly mutable loci in pathogenic bacteria.Perspect Biol Med 42: 154–155.
Muller, Y.A., Schumacher, G., Rudolph, R., and Schulz, G.E.(1994) The refined structures of a stabilized mutant and ofwild-type pyruvate oxidase from Lactobacillus plantarum.J Mol Biol 237: 315–335.
Musher, D.M. (2000) Streptococcus pneumoniae. In Mandell,Douglas, and Bennett’s Principles and Practice of Infec-tious Diseases. Mandell, G.L., Bennett, J.E., and Dolin, R.(eds). Philadelphia, PA: Churchill Livingstone, pp. 2128–2146.
National, R.C. (1996) Guide for the Care and Use of Labo-ratory Animals. Washington, DC: National Academy Press.
Ng, W.L., Robertson, G.T., Kazmierczak, K.M., Zhao, J.,Gilmour R., and Winkler, M.E. (2003) Constitutive expres-sion of PcsB suppresses the requirement for the essentialVicR (YycF) response regulator in Streptococcus pneumo-niae R6. Mol Microbiol 50: 1647–1663.
Ng, W.L., Tsui, H.C., and Winkler, M.E. (2005) Regulationof the pspA virulence factor and essential pcsB mureinbiosynthetic genes by the phosphorylated VicR (YycF)response regulator in Streptococcus pneumoniae.J Bacteriol 187: 7444–7459.
Orihuela, C.J., and Toumanen, E. (2006) Streptococcuspneumoniae: invasion and inflammation. In Gram-PositivePathogens. Fischetti, V.A., Novick, R.P., Ferretti, J.J.,Portnoy, D.A., and Rood, J.I. (eds). Washington DC:American Society for Microbiology Press, pp. 253–267.
Orihuela, C.J., Gao, G., McGee, M., Yu, J., Francis, K.P., andTuomanen, E. (2003) Organ-specific models of Streptococ-cus pneumoniae disease. Scand J Infect Dis 35: 647–652.
Orihuela, C.J., Radin, J.N., Sublett, J.E., Gao, G., Kaushal,D., and Tuomanen, E.I. (2004a) Microarray analysis ofpneumococcal gene expression during invasive disease.Infect Immun 72: 5582–5596.
Orihuela, C.J., Gao, G., Francis, K.P., Yu, J., and Tuomanen,E.I. (2004b) Tissue-specific contributions of pneumococcalvirulence factors to pathogenesis. J Infect Dis 190: 1661–1669.
Overweg, K., Pericone, C.D., Verhoef, G.G., Weiser, J.N.,Meiring, H.D., De Jong, A.P., et al. (2000) Differentialprotein expression in phenotypic variants of Streptococcuspneumoniae. Infect Immun 68: 4604–4610.
Paterson, G.K., Blue, C.E., and Mitchell, T.J. (2006) Role oftwo-component systems in the virulence of Streptococcuspneumoniae. J Med Microbiol 55: 355–363.
Pericone, C.D., Overweg, K., Hermans, P.W., and Weiser,J.N. (2000) Inhibitory and bactericidal effects of hydrogenperoxide production by Streptococcus pneumoniae onother inhabitants of the upper respiratory tract. InfectImmun 68: 3990–3997.
Pericone, C.D., Bae, D., Shchepetov, M., McCool, T., andWeiser, J.N. (2002) Short-sequence tandem and nontan-dem DNA repeats and endogenous hydrogen peroxideproduction contribute to genetic instability of Streptococcuspneumoniae. J Bacteriol 184: 4392–4399.
Pericone, C.D., Park, S., Imlay, J.A., and Weiser, J.N. (2003)Factors contributing to hydrogen peroxide resistance inStreptococcus pneumoniae include pyruvate oxidase(SpxB) and avoidance of the toxic effects of the fentonreaction. J Bacteriol 185: 6815–6825.
Pichichero, M.E., and Casey, J.R. (2007) Emergence of amultiresistant serotype 19A pneumococcal strain notincluded in the 7-valent conjugate vaccine as an otopatho-gen in children. JAMA 298: 1772–1778.
Regev-Yochay, G., Trzcinski, K., Thompson, C.M., Malley,R., and Lipsitch, M. (2006) Interference between Strepto-coccus pneumoniae and Staphylococcus aureus: in vitrohydrogen peroxide-mediated killing by Streptococcuspneumoniae. J Bacteriol 188: 4996–5001.
Regev-Yochay, G., Trzcinski, K., Thompson, C.M., Lipsitch,M., and Malley, R. (2007) SpxB is a suicide gene of Strep-tococcus pneumoniae and confers a selective advantagein an in vivo competitive colonization model. J Bacteriol189: 6532–6539.
Renna, M.C., Najimudin, N., Winik, L.R., and Zahler, S.A.(1993) Regulation of the Bacillus subtilis alsS, alsD, andalsR genes involved in post-exponential-phase productionof acetoin. J Bacteriol 175: 3863–3875.
Rigali, S., Derouaux, A., Giannotta, F., and Dusart, J. (2002)Subdivision of the helix-turn-helix GntR family of bacterialregulators in the FadR, HutC, MocR, and YtrA subfamilies.J Biol Chem 277: 12507–12515.
Robertson, G.T., Ng, W.L., Foley, J., Gilmour R., andWinkler, M.E. (2002) Global transcriptional analysis of clpPmutations of type 2 Streptococcus pneumoniae and theireffects on physiology and virulence. J Bacteriol 184: 3508–3520.
Sabelnikov, A.G., Greenberg, B., and Lacks, S.A. (1995) An
cis- and trans-regulation of pneumococcal spxB 745
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746

extended -10 promoter alone directs transcription of theDpnII operon of Streptococcus pneumoniae. J Mol Biol250: 144–155.
Saluja, S.K., and Weiser, J.N. (1995) The genetic basis ofcolony opacity in Streptococcus pneumoniae: evidence forthe effect of box elements on the frequency of phenotypicvariation. Mol Microbiol 16: 215–227.
Sambrook, J., and Russell, D.W. (2001) Molecular cloning: alaboratory manual. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press.
Scott, J.W., Hawley, S.A., Green, K.A., Anis, M., Stewart, G.,Scullion, G.A., et al. (2004) CBS domains form energy-sensing modules whose binding of adenosine ligands isdisrupted by disease mutations. [see comment]. J ClinInvest 113: 274–284.
Sedewitz, B., Schleifer, K.H., and Gotz, F. (1984) Physiologi-cal role of pyruvate oxidase in the aerobic metabolism ofLactobacillus plantarum. J Bacteriol 160: 462–465.
Shirk, M.C., Wagner, W.P., and Fall, R. (2002) Isopreneformation in Bacillus subtilis: a barometer of central carbonassimilation in a bioreactor? Biotechnol Prog 18: 1109–1115.
Spellerberg, B., Cundell, D.R., Sandros, J., Pearce, B.J.,Idanpaan-Heikkila, I., Rosenow, C., and Masure, H.R.(1996) Pyruvate oxidase, as a determinant of virulence inStreptococcus pneumoniae. Mol Microbiol 19: 803–813.
Sung, C.K., Li, H., Claverys, J.P., and Morrison, D.A. (2001)An rpsL cassette, janus, for gene replacement throughnegative selection in Streptococcus pneumoniae. ApplEnviron Microbiol 67: 5190–5196.
Talkington, D.F., Voellinger, D.C., McDaniel, L.S., and Briles,D.E. (1992) Analysis of pneumococcal PspA microhetero-geneity in SDS polyacrylamide gels and the association ofPspA with the cell membrane. Microb Pathog 13: 343–355.
Tettelin, H., Nelson, K.E., Paulsen, I.T., Eisen, J.A., Read,T.D., Peterson, S., et al. (2001) Complete genomesequence of a virulent isolate of Streptococcuspneumoniae. [see comment]. Science 293: 498–506.
Tittmann, K., Proske, D., Spinka, M., Ghisla, S., Rudolph, R.,Hubner, G., and Kern, G. (1998) Activation of thiamindiphosphate and FAD in the phosphatedependent pyruvateoxidase from Lactobacillus plantarum. J Biol Chem 273:12929–12934.
Tittmann, K., Wille, G., Golbik, R., Weidner, A., Ghisla, S.,
and Hubner, G. (2005) Radical phosphate transfer mecha-nism for the thiamin diphosphate- and FAD-dependentpyruvate oxidase from Lactobacillus plantarum. Kineticcoupling of intercofactor electron transfer with phosphatetransfer to acetyl-thiamin diphosphate via a transient FADsemiquinone/hydroxyethyl-ThDP radical pair. Biochemistry44: 13291–13303.
Tomasz, A. (2000) Streptococcus pneumoniae: MolecularBiology and Mechanisms of Disease. Larchmont, NY: MaryAnn Liebert.
Tuomanen, E.I., Mitchell, T.J., Morrison, D.A., and Spratt,B.G. (2004) The Pneumococcus. Washington, D.C.: Ameri-can Society for Microbiology Press.
Weiser, J.N. (2006) Phase variation of Streptococcuspneumoniae. In Gram-Positive Pathogens. Fischetti, V.A.,Novick, R.P., Ferretti, J.J., Portnoy, D.A., and Rood, J.I.(eds). Washington DC: American Society for MicrobiologyPress, pp. 268–274.
Weiser, J.N., Austrian, R., Sreenivasan, P.K., and Masure,H.R. (1994) Phase variation in pneumococcal opacity: rela-tionship between colonial morphology and nasopharyngealcolonization. Infect Immun 62: 2582–2589.
Wolfe, A.J. (2005) The acetate switch. Microbiol Mol Biol Rev69: 12–50.
Xue, X., Piao, J.H., Nakajima, A., Sakon-Komazawa, S.,Kojima, Y., Mori, K., et al. (2005) Tumor necrosis factoralpha (TNFalpha) induces the unfolded protein response(UPR) in a reactive oxygen species (ROS)-dependentfashion, and the UPR counteracts ROS accumulation byTNFalpha. J Biol Chem 280: 33917–33925.
Supplementary material
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2958.2007.06082.x(This link will take you to the article abstract).
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
746 S. Ramos-Montañez et al. �
© 2008 The AuthorsJournal compilation © 2008 Blackwell Publishing Ltd, Molecular Microbiology, 67, 729–746
Related Documents











