Special issue: Cephalopod Biology Polarization vision can improve object detection in turbid waters by cuttlefish ☆ Lelia Cartron a , Noam Josef b , Amit Lerner b , Steven D. McCusker b , Anne-Sophie Darmaillacq a , Ludovic Dickel a , Nadav Shashar b, ⁎ a Université de Caen Basse-Normandie, Groupe Mémoire et Plasticité comportementale EA 4259, F-14032 Caen, France b Department of Life Sciences, Eilat Campus, Ben-Gurion University of the Negev, P.O. Box 653, Beer-Sheva 84105, Israel abstract article info Article history: Received 2 January 2012 Received in revised form 2 May 2012 Accepted 25 January 2013 Available online 5 March 2013 Keywords: Contrast enhancement Optical depth Polarized-light Sepia Target detection The use of polarization vision to improve visibility through turbid water has been proposed by different re- searchers, but has rarely been tested behaviorally. Cephalopods perceive the polarization of light but the function of this sensitivity is not fully understood yet. In this two-phase study, we examined the ability of cut- tlefish to detect an artificial stimulus through water of different turbidity, with or without polarization infor- mation. We tested responses of Sepia prashadi and Sepia pharaonis to computer generated looming stimuli, presented through different levels of water turbidity. Stimuli were presented with a modified LCD monitor delivering polarized pattern that could not be detected without a polarization analyzer, and via a CRT monitor delivering an unpolarized, intensity contrasted image. The results showed that both cuttlefish can use their polarization vision to detect objects, such as a predator. Both species detected the pattern when it included a polarization component, through higher turbidity than when it contained only intensity information. Cuttlefish are nectobenthic organisms which probably often experience low visibility conditions. Therefore the ability to see further into turbid water and to better detect prey and predator would be beneficial for their survival. © 2013 Elsevier B.V. All rights reserved. 1. Introduction Visibility and small object detection in seawater is limited to distances of tens of meters and often less. This limitation is created by a combina- tion of processes that degrade the image including scattering, absorption and the masking of objects by veiling light (Lythgoe, 1979). In addition to the illumination conditions, and the properties and capabilities of the vi- sual and processing systems of the observer, the properties of the waters, and especially their particle content or turbidity, are keys in determining the actual distance of detection. Polarization has been found to increase contrast and detection range in scattering media both above and under water (Flamarique and Browman, 2001; Schechner and Karpel, 2004, 2005; Schechner et al., 2003; Shashar et al., 1995, 1998, 2004; for review see also Shashar et al., 2011). Moreover, marine organisms may affect and even generate localized polarization via scattering, reflection and bire- fringence of their tissues. Polarization may therefore provide a more reli- able channel for the detection of predator and prey in conditions where luminance or chromatic signals do not contain sufficient or reliable visual contrast (see Marshall and Cronin, 2011 for a discussion on polarization- sensitivity vs. -vision). Many cephalopod species are sensitive to the linear polarization of light (Gleadall and Shashar, 2004; for review see also Hanlon and Shashar, 2003). This sensitivity arises from the mostly orthogonal arrangement of the microvilli in the retina (Moody, 1962; Shashar et al., 2002). For each photoreceptor cell, the microvilli protrude mostly later- ally on opposite sides of the receptor's outer segment, so that all the microvilli of a given cell are aligned along a dominant axis. The alignment of the retinal molecules of visual pigment within the microvillar mem- brane (Laughlin et al., 1975) along with the alignment for the microvilli in the cell is such that the photoreceptor cell absorbs light most efficiently in one orientation of polarization (for review see: Goldsmith, 1977, 1991). While cephalopods are known to use their polarization vision for prey detection and recognition (Shashar et al., 1998, 2000), it is not yet clear whether they use it for detection of predators or other objects or for improvement of visibility through turbid waters. One limitation in testing polarization functions so far was that creating dynamic polarized stimuli free of confounding luminance and chromatic cues has been difficult, though not impossible. To overcome this limitation, Pignatelli et al. (2011) developed an experimental set-up based on the use of a modified liquid crystal display (LCD) monitor to deliver computer-generated controlled polarized stimuli. In this study, we used the same design and tested cuttlefish responses to large, rapidly approaching, stimuli through different levels of water turbidity. We compared cuttlefish's behavioral responses to intensity based stimuli vs. polarization based stimuli, to examine the animal ability to detect objects through turbid waters in the intensity and polarization domains. Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85 ☆ This article is part of a special issue on Cephalopod Biology published under the auspices of CephRes-ONLUS ( www.cephalopodresearch.org). ⁎ Corresponding author. Tel.: +972 8 6304520/531; fax: +972 8 6304538. E-mail address: [email protected] (N. Shashar). 0022-0981/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.jembe.2013.02.013 Contents lists available at SciVerse ScienceDirect Journal of Experimental Marine Biology and Ecology journal homepage: www.elsevier.com/locate/jembe

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
Contents lists available at SciVerse ScienceDirect
Journal of Experimental Marine Biology and Ecology
j ourna l homepage: www.e lsev ie r .com/ locate / jembe
Special issue: Cephalopod Biology
Polarization vision can improve object detection in turbid waters by cuttlefish☆
Lelia Cartron a, Noam Josef b, Amit Lerner b, Steven D. McCusker b, Anne-Sophie Darmaillacq a,Ludovic Dickel a, Nadav Shashar b,⁎a Université de Caen Basse-Normandie, Groupe Mémoire et Plasticité comportementale EA 4259, F-14032 Caen, Franceb Department of Life Sciences, Eilat Campus, Ben-Gurion University of the Negev, P.O. Box 653, Beer-Sheva 84105, Israel
☆ This article is part of a special issue on Cephalopodauspices of CephRes-ONLUS (www.cephalopodresearch⁎ Corresponding author. Tel.: +972 8 6304520/531;
E-mail address: [email protected] (N. Shashar).
0022-0981/$ – see front matter © 2013 Elsevier B.V. Alhttp://dx.doi.org/10.1016/j.jembe.2013.02.013
a b s t r a c t
a r t i c l e i n f oArticle history:Received 2 January 2012Received in revised form 2 May 2012Accepted 25 January 2013Available online 5 March 2013
Keywords:Contrast enhancementOptical depthPolarized-lightSepiaTarget detection
The use of polarization vision to improve visibility through turbid water has been proposed by different re-searchers, but has rarely been tested behaviorally. Cephalopods perceive the polarization of light but thefunction of this sensitivity is not fully understood yet. In this two-phase study, we examined the ability of cut-tlefish to detect an artificial stimulus through water of different turbidity, with or without polarization infor-mation. We tested responses of Sepia prashadi and Sepia pharaonis to computer generated looming stimuli,presented through different levels of water turbidity. Stimuli were presented with a modified LCD monitordelivering polarized pattern that could not be detected without a polarization analyzer, and via a CRT monitordelivering an unpolarized, intensity contrasted image. The results showed that both cuttlefish can use theirpolarization vision to detect objects, such as a predator. Both species detected the pattern when it includeda polarization component, through higher turbidity than when it contained only intensity information.Cuttlefish are nectobenthic organisms which probably often experience low visibility conditions. Thereforethe ability to see further into turbid water and to better detect prey and predator would be beneficial fortheir survival.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Visibility and small object detection in seawater is limited to distancesof tens of meters and often less. This limitation is created by a combina-tion of processes that degrade the image including scattering, absorptionand themasking of objects by veiling light (Lythgoe, 1979). In addition tothe illumination conditions, and the properties and capabilities of the vi-sual and processing systems of the observer, the properties of the waters,and especially their particle content or turbidity, are keys in determiningthe actual distance of detection. Polarization has been found to increasecontrast and detection range in scattering media both above and underwater (Flamarique and Browman, 2001; Schechner and Karpel, 2004,2005; Schechner et al., 2003; Shashar et al., 1995, 1998, 2004; for reviewsee also Shashar et al., 2011).Moreover, marine organismsmay affect andeven generate localized polarization via scattering, reflection and bire-fringence of their tissues. Polarization may therefore provide a more reli-able channel for the detection of predator and prey in conditions whereluminance or chromatic signals do not contain sufficient or reliable visualcontrast (see Marshall and Cronin, 2011 for a discussion on polarization-sensitivity vs. -vision).
Biology published under the.org).fax: +972 8 6304538.
l rights reserved.
Many cephalopod species are sensitive to the linear polarization oflight (Gleadall and Shashar, 2004; for review see also Hanlon andShashar, 2003). This sensitivity arises from the mostly orthogonalarrangement of the microvilli in the retina (Moody, 1962; Shashar et al.,2002). For each photoreceptor cell, the microvilli protrude mostly later-ally on opposite sides of the receptor's outer segment, so that all themicrovilli of a given cell are aligned along a dominant axis. The alignmentof the retinal molecules of visual pigment within the microvillar mem-brane (Laughlin et al., 1975) along with the alignment for the microvilliin the cell is such that the photoreceptor cell absorbs lightmost efficientlyin one orientation of polarization (for review see: Goldsmith, 1977, 1991).
While cephalopods are known to use their polarization vision for preydetection and recognition (Shashar et al., 1998, 2000), it is not yet clearwhether they use it for detection of predators or other objects or forimprovement of visibility through turbidwaters. One limitation in testingpolarization functions so far was that creating dynamic polarized stimulifree of confounding luminance and chromatic cues has been difficult,though not impossible. To overcome this limitation, Pignatelli et al.(2011) developed an experimental set-up based on the use of a modifiedliquid crystal display (LCD) monitor to deliver computer-generatedcontrolled polarized stimuli. In this study, we used the same design andtested cuttlefish responses to large, rapidly approaching, stimuli throughdifferent levels of water turbidity. We compared cuttlefish's behavioralresponses to intensity based stimuli vs. polarization based stimuli, toexamine the animal ability to detect objects through turbid waters inthe intensity and polarization domains.

81L. Cartron et al. / Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
2. Methods
This study used a behavioral experiment to examine the responseof cuttlefish to stimuli presented at different levels of turbidity.
2.1. Animals
Two cuttlefish individuals, Sepia prashadi (dorsal mantle length=6 cm) and Sepia pharaonis (dorsal mantle length=7 cm), were usedin this experiment. They were caught, under a special permit fromthe Israeli Nature and Parks Authority, at night in the Gulf of Aqaba(Eilat, Israel) in shallow waters. Since cuttlefish are a protected andpoorly studied species in this region, we were limited to one individ-ual of each species. The cuttlefish were kept separately in individualtanks (50×50×50 cm) with running sea water, under shaded naturalillumination, and fed daily with juvenile fish (total length 4 to 5 cm;obtained from Ardag Ltd., Eilat). Cuttlefish were released exactlywhere they were caught at the end of the experiment.
2.2. Apparatus
The housing tank (50×50×50 cm) was a glass aquarium with 3sides blocked by opaque screens and one left open. In front of thislast side, an experimental compartment (15×28×50 cm) was in-stalled in the housing tank (Fig. 1). A smaller aquarium (14×28×20 cm) was placed just next to it, in which the water turbidity wasvaried. The short sides (14×20 cm), the bottom and the topwere cov-ered with opaque screens to block incoming light from the outside.A computer screen linked to a laptop was placed in front of the exper-imental compartment, on the far side of this turbidity tank. Two typesof computer screen were used in this experiment. A CRT monitor wasused to deliver contrasted stimuli without polarization. A modifiedLCD monitor where the external polarizing filter had been removed(following Pignatelli et al., 2011; Temple et al., 2012), was used to de-liver a polarized stimulus which was unseen by polarization insensi-tive animals (Fig. 2B).
2.3. Stimuli
Five types of white stimuli on a black background were presentedto each cuttlefish through both types of screens. They consisted infour geometrical shapes (circle, rectangle, lozenge and star) and a
Fig. 1. A scheme of the apparatus used in behavioral experiments. The laptop was used to gewas either a CRT or a LCD monitor. Dimensions in cm.
grouper's image (Fig. 2). Each shape suddenly appeared and expand-ed (loomed) to 300% of its original size in 0.5 s to occupy half the sizeof the screen. One should note that in the CRT screen (Fig. 2A) the fishshowed an intensity contrast from 28% of brightness in the back-ground (70 on a grayscale where 255 represents full white and 0full black) vs. up to 100% brightness (255) in the fish's bright areas;the LCD (polarization) screen had a constant intensity of 83% (212)brightness throughout the screen (Fig. 2B). The partial (percent) po-larization contrast of the LCD image ranged from 94% polarization inthe background to 60% polarization in the least polarized parts ofthe fish (Fig. 2C). The orientation of polarization ranged from 50° inthe background to up to 85° in the fish (Fig. 2D). This difference ine-vector orientation should be large enough to be detected by cuttle-fish (Temple et al., 2012).
2.4. Light attenuation measurements
The entire set upwas covered from the outsidewith a black opaquesheet to eliminate external illumination aswell as veiling illumination.A portable fiber-optic spectrophotometer (S2000, Ocean Optics Inc.,FL, USA) was used to measure light intensity (I) on a spectrum be-tween 400 and 700 nm. It was coupled with a rotatable linear polariz-ing filter to measure light intensity at three different orientation of thefilter (0°, 45° and 90°). Incoming light, intensity and polarization aswell as light passing through the turbidity tank were measured.Light measurements were taken in clear water (Cl) and four increas-ing levels of turbidity (ClbT1bT2bT3bT4; Table 1).
Partial (%) polarization and e-vector orientation were calculatedfrom the set of three readings (i0, i45, i90) following the equationsused by Sabbah et al. (2005). Light intensity, %polarization ande-vector orientation were averaged for wavelengths between 420and 550 nm which are around the pick of maximal absorption forthe cuttlefish's rhodopsin at 492 nm (Mathger et al., 2006).
2.5. Procedure
The cuttlefish was gently pushed from the housing tank into theexperimental compartment. It was given 5 to 10 min to settle downand familiarize to the apparatus. Each cuttlefish was tested in two tothree sessions per day. Each session was composed of 5 presentationsof a single stimulus through the same type ofmonitor spaced by 5-minintervals. Between sessions, the cuttlefish had no less than 15 min to
nerate the presented images, and was not seen by the animal. The presentation screen

Fig. 2. Grouper's image delivered by CRT (A) and by LCD monitors (B) both as seen by a human observer. Note that the fish pattern could not be seen on the LCD monitor. Imagepolarimeter reveals the partial (percent) polarization (C) and e-vector orientation (D) of the grouper's image when delivered by LCD monitor.
82 L. Cartron et al. / Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
calm down. The presentations were recorded from above with avideo camera (Sanyo Xacti VPC-CG10). Responses of the cuttlefishto the stimuli were quantified as the number of presentations (outof 5) in which the animal changed its body pattern (“pattern re-sponse”, Fig. 3) or moved away from the screen (“escape response”).In S. prashadi, the body pattern passed from a uniform to a disruptivecoloration when the stimulus appeared (Fig. 3B). In S. pharaonis, itchanged from a light uniform pattern to a dark pattern with blackspots (Fig. 3C, D). At the end of the daily sessions, the experimentalcompartment was opened and the cuttlefish was allowed to swimfreely into the housing tank.
In the first part of the experiment, with clear water in the turbiditychamber, three sessions were run using the CRT monitor. In session 1,we compared the cuttlefish's responses to the 4 geometrical shapesand a grouper image (Fig. 2) presented in a random order. In sessions2 and 3, we compared the response to five successive presentations ofthe circle and the grouper, which induced the strongest response insession 1. Then, those three sessionswere repeatedwith the LCDmon-itor. This allowed us to determine which stimulus was the most effec-tive to elicit a response from the cuttlefish when presented on eachtype of screens. Only the grouper's image stimulus was used in the fol-lowing part of the experiment.
Table 1Percent of the light's passing through the turbidity tank in the intensity (%I) and linearpolarization (%P — partial polarization) domains for each level of turbidity (from clearwater, CL to T4) tested with Sepia prashadi and Sepia pharaonis. There was no effect onthe orientation of polarization.
S. prashadi S. pharaonis
%I %P %I %P
CL 100 100 100 100T1 85 98 85 98T2 57 85 56 85T3 41 75 43 77T4 28 63 29 64
In the second part of the experiment, the turbidity level was in-creased with fine natural sand (diameterb1 mm) collected underwa-ter in the Gulf of Aqaba (Eilat, Israel). For each turbidity level and forclear seawater, the light intensity and %polarization passing throughthe tankweremeasuredwith the portable fiber-optic spectrophotom-eter. Then, for each turbidity level (ClbT1bT2bT3bT4), the cuttlefishwas given two sessions with five presentations of the grouper's image,one session with the CRT monitor and one session with the LCD mon-itor, in a random order.
2.6. Statistical analysis
Behavioral data were analyzed using StatXact 7 (Cytel Studio soft-ware). A significance threshold of α=0.05 was used. In the first partof the experiment, responses to the CRT and LCD monitors werepooled and McNemar tests were used to compare the number of be-havioral responses to the circle and the grouper image. In the secondpart, Cochran's Q tests were used to compare the variation in re-sponse to the CRT and the LCD monitor with turbidity level. If thenull hypothesis was rejected, Wilcoxon Signed Rank tests were usedfor post-hoc pairwise comparisons.
3. Results
The light intensity and partial (%) polarization decreased with in-creasing turbidity levels (Table 1). However, intensity decreasedfaster than partial polarization with increasing turbidity. E-vector ori-entation remained relatively unaffected in all turbidity levels. Similarconditions were presented to S. prashadi and S. pharaonis.
3.1. Sepia prashadi
In the first part of the experiment, the cuttlefish responded to allshapes for both types of screen but the responses were stronger forthe circle and the grouper. Responses to CRT and LCD monitors were

Fig. 3. Body patterns of Sepia prashadi when relaxed (A) and in response to a visual stimulus (B) and of Sepia pharaonis in relaxed (C) condition or in response to a stimulus (D).
83L. Cartron et al. / Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
not different (P=0.25) so results were pooled for this part of the ex-periment (Fig. 4A). In the10 presentations (5 CRT+5 LCD), the cuttle-fish showedmore pattern responses to the grouper image (9/10) thanto the circle (2/10; P=0.006). Moreover, the cuttlefish had a tendencyto showmore escape response to the grouper image (5/10) than to thecircle (1/10), though not significant (P=0.052). These results tend toshow that the grouper image was more efficient in inducing a re-sponse. Therefore, the grouper imagewas chosen for the following ex-periments to test the effects of different levels of water turbidity.
In the second part of the experiment, we compared responsesto the CRT and LCD monitors through increasing levels of turbidity(Fig. 5A). The number of pattern responses decreased with turbidityfor both CRT (P=0.006) and LCD (P=0.005) monitor whereas theoverall number of escape responses did not significantly vary withturbidity (CRT, P=0.400; LCD, P=0.440). The number of pattern re-sponses was stable (4 to 5/5 times) from the clear water through T2(P=0.250) and then dropped sharply to 0 at T3 for the CRT (T2vs.T3; P=0.062) monitor and at T4 for the LCD monitor (T3/T4 P=0.062). The number of escape responses reached 0 at T3 for the CRTmonitor and T4 for the LCD monitor. These results showed that thegrouper image was still detected by the cuttlefish at T3 based on
Fig. 4. Responses of Sepia prashadi (A) and Sepia pharaonis (B) to the circle and the grouper ianimal changed body pattern or moved away from the screen; CRT monitor; LCD moMacNemar test.
polarization vision but not on intensity vision alone (response CRT/LCD, P=0.031).
3.2. Sepia pharaonis
In the first part of the experiment, the cuttlefish responded to allshapes presented through both types of screens, but the responseswere stronger for the circle and the grouper image. Responses toCRT monitor did not differ from that to the LCD (P=0.25) so resultswere pooled for this part of the experiment (Fig. 4B). In 10 presenta-tions (5 CRT+5 LCD), the cuttlefish showed more pattern responsesto the grouper image (8/10) than to the circle, though this differencewas not significant (4/10; P=0.052). There was no significant differ-ences in the number of escape responses from the circle (1/10) andthe grouper images (4/10; P=0.125). Since the grouper image wasmore effective to induce responses by the cuttlefish it was chosenfor presentation in the following experiments.
In the second part of the experiment, we compared responses tothe CRT and LCD monitors over increasing turbidity levels (Fig. 5B).The number of pattern responses decreased with turbidity for bothCRT (P=0.001) and LCD (P=0.024) monitors, whereas the number
mage. Values (score) are the number of presentations (out of a total of 10) in which thenitor. Significant difference (Pb0.05) or pairwise comparison is marked (* or ▲) after
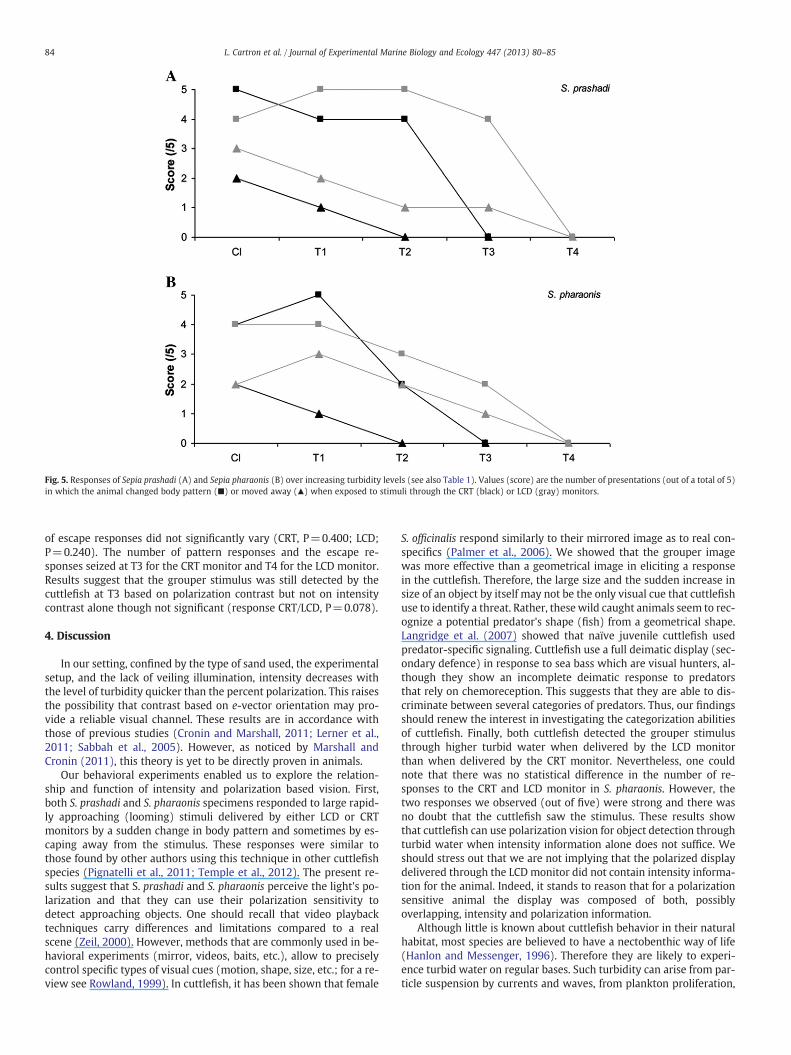
Fig. 5. Responses of Sepia prashadi (A) and Sepia pharaonis (B) over increasing turbidity levels (see also Table 1). Values (score) are the number of presentations (out of a total of 5)in which the animal changed body pattern (■) or moved away (▲) when exposed to stimuli through the CRT (black) or LCD (gray) monitors.
84 L. Cartron et al. / Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
of escape responses did not significantly vary (CRT, P=0.400; LCD;P=0.240). The number of pattern responses and the escape re-sponses seized at T3 for the CRT monitor and T4 for the LCD monitor.Results suggest that the grouper stimulus was still detected by thecuttlefish at T3 based on polarization contrast but not on intensitycontrast alone though not significant (response CRT/LCD, P=0.078).
4. Discussion
In our setting, confined by the type of sand used, the experimentalsetup, and the lack of veiling illumination, intensity decreases withthe level of turbidity quicker than the percent polarization. This raisesthe possibility that contrast based on e-vector orientation may pro-vide a reliable visual channel. These results are in accordance withthose of previous studies (Cronin and Marshall, 2011; Lerner et al.,2011; Sabbah et al., 2005). However, as noticed by Marshall andCronin (2011), this theory is yet to be directly proven in animals.
Our behavioral experiments enabled us to explore the relation-ship and function of intensity and polarization based vision. First,both S. prashadi and S. pharaonis specimens responded to large rapid-ly approaching (looming) stimuli delivered by either LCD or CRTmonitors by a sudden change in body pattern and sometimes by es-caping away from the stimulus. These responses were similar tothose found by other authors using this technique in other cuttlefishspecies (Pignatelli et al., 2011; Temple et al., 2012). The present re-sults suggest that S. prashadi and S. pharaonis perceive the light's po-larization and that they can use their polarization sensitivity todetect approaching objects. One should recall that video playbacktechniques carry differences and limitations compared to a realscene (Zeil, 2000). However, methods that are commonly used in be-havioral experiments (mirror, videos, baits, etc.), allow to preciselycontrol specific types of visual cues (motion, shape, size, etc.; for a re-view see Rowland, 1999). In cuttlefish, it has been shown that female
S. officinalis respond similarly to their mirrored image as to real con-specifics (Palmer et al., 2006). We showed that the grouper imagewas more effective than a geometrical image in eliciting a responsein the cuttlefish. Therefore, the large size and the sudden increase insize of an object by itself may not be the only visual cue that cuttlefishuse to identify a threat. Rather, these wild caught animals seem to rec-ognize a potential predator's shape (fish) from a geometrical shape.Langridge et al. (2007) showed that naïve juvenile cuttlefish usedpredator-specific signaling. Cuttlefish use a full deimatic display (sec-ondary defence) in response to sea bass which are visual hunters, al-though they show an incomplete deimatic response to predatorsthat rely on chemoreception. This suggests that they are able to dis-criminate between several categories of predators. Thus, our findingsshould renew the interest in investigating the categorization abilitiesof cuttlefish. Finally, both cuttlefish detected the grouper stimulusthrough higher turbid water when delivered by the LCD monitorthan when delivered by the CRT monitor. Nevertheless, one couldnote that there was no statistical difference in the number of re-sponses to the CRT and LCD monitor in S. pharaonis. However, thetwo responses we observed (out of five) were strong and there wasno doubt that the cuttlefish saw the stimulus. These results showthat cuttlefish can use polarization vision for object detection throughturbid water when intensity information alone does not suffice. Weshould stress out that we are not implying that the polarized displaydelivered through the LCD monitor did not contain intensity informa-tion for the animal. Indeed, it stands to reason that for a polarizationsensitive animal the display was composed of both, possiblyoverlapping, intensity and polarization information.
Although little is known about cuttlefish behavior in their naturalhabitat, most species are believed to have a nectobenthic way of life(Hanlon and Messenger, 1996). Therefore they are likely to experi-ence turbid water on regular bases. Such turbidity can arise from par-ticle suspension by currents and waves, from plankton proliferation,

85L. Cartron et al. / Journal of Experimental Marine Biology and Ecology 447 (2013) 80–85
and from runoffs. Therefore, cuttlefish's use of their polarization visionto see further or sharper in turbid waters has a clear advantage foravoiding predators and hiding from prey.
Acknowledgments
We thank the support of Ben Gurion University's Marine Biologyand Biotechnology program. Comments by two anonymous reviewersgreatly improved this manuscript. We thank the Israeli National Cen-ter for Mariculture and the Ardag Company for the assistance in ani-mal maintenance. Special thanks to the Israeli Nature and ParksAuthority for monitoring the capture, maintenance, and release ofthe cuttlefish. This study was partly supported by an Israeli ScienceFoundation grant # 10/1081 to NS, by the Halperin and Shechter Foun-dations, and by a grant from the French Ministère de l'EnseignementSupérieur et de la Recherche to LC. [SS]
References
Cronin, T.W., Marshall, J., 2011. Patterns and properties of polarized light in air andwater. Philos. Trans. R. Soc. Lond. B 366, 619–626.
Flamarique, I.N., Browman, H.I., 2001. Foraging and prey-search behaviour of smalljuvenile rainbow trout (Oncorhynchus mykiss) under polarized light. J. Exp. Biol.204, 2415–2422.
Gleadall, I., Shashar, N., 2004. An Octopus' Garden: the visual world of Cephalopods. In:Prete, F.D. (Ed.), Complex Worlds from Simpler Nervous Systems. MIT Press,Cambridge, Mass, pp. 269–307.
Goldsmith, T.H., 1977. Membrane adaptations of visual photoreceptors for the analysisof plane-polarized light. In: Castellani, A. (Ed.), Research in Photobiology. PlenumPress, New York, NY, pp. 651–658.
Goldsmith, T.H., 1991. Photoreception and vision. In: Prosser, C.L. (Ed.), Neural and In-tegrative Animal Physiology. Wiley-Liss, New York, NY, pp. 171–245.
Hanlon, R.T., Messenger, J.B., 1996. Cephalopod Behaviour. Cambridge University Press,Cambridge, pp. 1–232.
Hanlon, R.T., Shashar, N., 2003. Aspects of the sensory ecology of cephalopods. In:Collin, S.P., Marshall, J. (Eds.), Sensory Processing of the Aquatic environment.Springer, New York, NY, pp. 226–282.
Langridge, K.V., Broom, M., Osorio, D., 2007. Selective signalling by cuttlefish to preda-tors. Curr. Biol. 17, R1044–R1045.
Laughlin, S.B., Menzel, R., Snyder, A.W., 1975. Membranes, dichroism and receptor sen-sitivity. In: Snyder, A.W., Menzel, R. (Eds.), Photoreceptor Optics. Springer Verlag,New York, NY, pp. 237–259.
Lerner, A., Sabbah, S., Erlick, C., Shashar, N., 2011. Navigation by light polarization inclear and turbid waters. Philos. Trans. R. Soc. Lond. B 366, 671–679.
Lythgoe, J.N., 1979. The Ecology of Vision. Clarendon Press, Oxford, UK, pp. 1–224.Marshall, J., Cronin, T.W., 2011. Polarisation vision. Curr. Biol. 21, R101–R105.Mathger, L.M., Barbosa, A., Miner, S., Hanlon, R.T., 2006. Color blindness and contrast
perception in cuttlefish (Sepia officinalis) determined by a visual sensorimotorassay. Vision Res. 46, 1746–1753.
Moody, M.F., 1962. Evidence for intraocular discrimination of vertically and horizontal-ly polarized light by Octopus. J. Exp. Biol. 39, 21–30.
Palmer, M.E., Calve, M.R., Adamo, S.A., 2006. Response of female cuttlefish Sepiaofficinalis (Cephalopoda) to mirrors and conspecifics: evidence for signaling infemale cuttlefish. Anim. Cogn. 9, 151–155.
Pignatelli, V., Temple, S.E., Chiou, T.H., Roberts, N.W., Collin, S.P., Marshall, N.J., 2011.Behavioural relevance of polarization sensitivity as a target detection mechanismin cephalopods and fishes. Philos. Trans. R. Soc. Lond. B 366, 734–741.
Rowland, W.J., 1999. Studying visual cues in fish behavior: a review of ethological tech-niques. Environ. Biol. Fish. 56, 285–305.
Sabbah, S., Lerner, A., Erlick, C., Shashar, N., 2005. Under water polarization vision. Aphysical examination. In: Pandalai, S.G. (Ed.), Recent Research Developments inExperimental and Theoretical Biology. TRN Press, Kerala, India, pp. 123–176.
Schechner, Y.Y., Karpel, N., 2004. Clear underwater vision. Proceedings of the 2004IEEE Computer Society Conference on Computer Vision and Pattern Recognition,1, pp. 536–543.
Schechner, Y.Y., Karpel, N., 2005. Recovery of underwater visibility and structure by po-larization analysis. IEEE J. Ocean. Eng. 30, 570–587.
Schechner, Y.Y., Narasimhan, S.G., Nayar, S.K., 2003. Polarization-based vision throughhaze. Appl. Opt. 42, 511–525.
Shashar, N., Adessi, L., Cronin, T.W., 1995. Polarization vision as a mechanism for detec-tion of transparent objects. In: Gulko, D., Jokiel, P.L. (Eds.), Ultraviolet Radiationand Coral Reefs. Hawaii Institute of Marine Biology, University of Hawaii; SeagrantPublications UNIHI-SEAGRANT CR-95-03, Honolulu, HW, pp. 207–211.
Shashar, N., Hanlon, R.T., Petz, A.D., 1998. Polarization vision helps detect transparentprey. Nature 393, 222–223.
Shashar, N., Hagan, R., Boal, J.G., Hanlon, R.T., 2000. Cuttlefish use polarization sensitiv-ity in predation on silvery fish. Vision Res. 40, 71–75.
Shashar, N., Milbury, C.A., Hanlon, R.T., 2002. Polarization vision in cephalopods: neu-roanatomical and behavioral features that illustrate aspects of form and function.Mar. Freshw. Behav. Physiol. 35, 57–68.
Shashar, N., Sabbah, S., Cronin, T.W., 2004. Transmission of linearly polarized light inseawater: implications for polarization signaling. J. Exp. Biol. 207, 3619–3628.
Shashar, N., Johnsen, S., Lerner, A., Sabbah, S., Chiao, C.C., Mäthger, L.M., Hanlon, R.T.,2011. Underwater linear polarization: physical limitations to biological functions.Philos. Trans. R. Soc. Lond. B 366, 649–654.
Temple, S.E., Pignatelli, V., Cook, T., How, M.J., Chiou, T.H., Roberts, N.W., Marshall, N.J.,2012. High-resolution polarisation vision in a cuttlefish. Curr. Biol. 22, R121–R122.
Zeil, J., 2000. Depth cues, behavioural context, and natural illumination: some potentiallimitations of video playback techniques. Acta Ethol. 3, 39–48.
Related Documents











