ORIGINAL ARTICLE Plasminogen activator inhibitor-1 as regulator of tumor-initiating cell properties in head and neck cancers Yueh-Chun Lee, MD 1,2 Cheng-Chia Yu, PhD 3,4,5 Chih Lan, MS 6 Che-Hsin Lee, PhD 7,8 Hsueh-Te Lee, PhD 9 Yu-Liang Kuo, PhD 10,11 Po-Hui Wang, MD, PhD 2,12,13 * Wen-Wei Chang, PhD 6,14 * 1 Radiation Oncology Department, Chung Shan Medical University Hospital, Taichung City, Taiwan, 2 Institute of Medicine, Chung Shan Medical University, Taichung, Taiwan, 3 School of Dentistry, Chung Shan Medical University, Taichung, Taiwan, 4 Department of Dentistry, Chung Shan Medical University Hospital, Taichung, Taiwan, 5 Institute of Oral Sciences, Chung Shan Medical University, Taichung, Taiwan, 6 School of Biomedical Sciences, Chung Shan Medical University, Taichung City, Taiwan, 7 Graduate Institute of Basic Medical Science, School of Medicine, China Medical University, Taichung, Taiwan, 8 Department of Microbiology, School of Medicine, China Medical University, Taichung, Taiwan, 9 Institute of Anatomy and Cell Biology, School of Medicine, National Yang Ming University, Taipei, Taiwan, 10 Department of Medical Imaging, Chung Shan Medical University Hos- pital, Taichung, Taiwan, 11 School of Medical Imaging and Radiological Sciences, Chung Shan Medical University, Taichung, Taiwan, 12 Department of Obstetrics and Gynecology, Chung Shan Medical University Hospital, Taichung, Taiwan, 13 School of Medicine, Chung Shan Medical University, Taichung, Taiwan, 14 Department of Medical Research, Chung Shan Medical University Hospital, Taichung City, Taiwan. Accepted 8 May 2015 Published online 00 Month 2015 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/hed.24124 ABSTRACT: Background. The existence of tumor-initiating cells (TICs) has been described in head and neck cancers. Plasminogen activator inhibitor-1 (PAI-1) has been demonstrated to act as a prognostic factor in head and neck cancers. Methods. Tiplaxtinin (PAI-039), a specific inhibitor of PAI-1, and PAI-1- specific siRNA were used to examine the role of PAI-1 in the self- renewal property of head and neck cancer-TICs by tumorsphere forma- tion. Western blot, real-time polymerase chain reaction, and luciferase- based reporter assay were used to study the effect of PAI-039 in the sex-determining region Y-box 2 (Sox2) expression. Results. PAI-039 suppressed the self-renewal capability of head and neck cancer-TICs derived from head and neck cancer cell lines through the inhibition of Sox2 expression. PAI-039 decreased the activity of the core promoter and the enhancer of the Sox2 gene in head and neck cancer-TICs. Knockdown of PAI-1 expression also inhibited self-renewal and radioresistance properties of head and neck cancer-TICs. Conclusion. The inhibition of PAI-1 by PAI-039 or siRNA could suppress head and neck cancer-TICs within head and neck cancer cell lines through the downregulation of Sox2. V C 2015 Wiley Periodicals, Inc. Head Neck 00: 000–000, 2015 KEY WORDS: plasminogen activator inhibitor-1 (PAI-1), tiplaxtinin (PAI-039), tumor-initiating cells, head and neck cancer, Sox2 INTRODUCTION Head and neck cancers are a group of cancers that occur in the paranasal sinuses, nasopharynx, oral cavities, oro- pharynx, hypopharynx, larynx, and salivary glands. 1 Head and neck squamous cell carcinoma (HNSCC) accounts for more than 90% cases of head and neck cancers. 2 Head and neck cancers account for 3% of all cancers in the United States 3 and they are ranked sixth of the most com- mon cancers worldwide. 4 The survival rate of head and neck cancers is near 50%, and the relatively high local recurrence rate, the metastatic spread, and development of second primary cancer in the head and neck region are thought to be associated with the poor 5-year survival rate of head and neck cancers. 1 Tumor-initiating cells (TICs), or cancer stem cells, are a subpopulation of cancer cells that have been shown to participate in tumor initiation, resistance to therapy, and metastasis. 5 In head and neck cancers, the reported markers of TICs include CD44, 2,6 CD133, 7 aldehyde dehydrogenase (ALDH), 8 Grp78, 9 side population, 10 and c-Met. 11 Head and neck cancer-TICs could also be enriched by tumorsphere cultivation. 12 Transcriptional factors in the maintenance of embryonic stem cells have been frequently found to be overexpressed in cancers and TICs. It has been reported that the phosphorylated Nanog promotes the growth of head and neck cancers through direct transactivation of Bmi1. 13 The ectopic expression of the sex-determining region Y-box 2 (Sox2) in HNSCC cell lines increased their self-renewal and chemoresistance properties. 14 Sox2 expression in head and neck cancers is associated with the recurrence and poor prognosis of patients. 15,16 The urokinase plasminogen activator (uPA)-plasmin system has been described to be associated with cancer progression. 17 Plasminogen activator inhibitor (PAI)21 *Corresponding author: P.-H. Wang, School of Medicine and Institute of Medicine, Chung Shan Medical University, 110, Section 1, Chien-Kuo North Road, Taichung 40201, Taiwan. E-mail: [email protected]. W.-W. Chang, School of Biomedical Sciences, Chung Shan Medical University, 110, Section 1, Chien-Kuo North Road, Taichung 40201, Taiwan. E-mail: [email protected] Contract grant sponsor: This work was supported by Chung Shan Medical University Hospital (grant no. CSH-2013-A-030) and Ministry of Science and Technology in Taiwan (grant no. MOST-103-2314-B-040-015-MY3). HEAD & NECK—DOI 10.1002/HED MONTH 2015 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Plasminogen activator inhibitor-1 as regulator of tumor-initiating cell properties inhead and neck cancers
Yueh-Chun Lee, MD1,2 Cheng-Chia Yu, PhD3,4,5 Chih Lan, MS6 Che-Hsin Lee, PhD7,8 Hsueh-Te Lee, PhD9 Yu-Liang Kuo, PhD10,11
Po-Hui Wang, MD, PhD2,12,13* Wen-Wei Chang, PhD6,14*
1Radiation Oncology Department, Chung Shan Medical University Hospital, Taichung City, Taiwan, 2Institute of Medicine, Chung Shan Medical University, Taichung, Taiwan,3School of Dentistry, Chung Shan Medical University, Taichung, Taiwan, 4Department of Dentistry, Chung Shan Medical University Hospital, Taichung, Taiwan, 5Institute of OralSciences, Chung Shan Medical University, Taichung, Taiwan, 6School of Biomedical Sciences, Chung Shan Medical University, Taichung City, Taiwan, 7Graduate Institute of BasicMedical Science, School of Medicine, China Medical University, Taichung, Taiwan, 8Department of Microbiology, School of Medicine, China Medical University, Taichung, Taiwan,9Institute of Anatomy and Cell Biology, School of Medicine, National Yang Ming University, Taipei, Taiwan, 10Department of Medical Imaging, Chung Shan Medical University Hos-pital, Taichung, Taiwan, 11School of Medical Imaging and Radiological Sciences, Chung Shan Medical University, Taichung, Taiwan, 12Department of Obstetrics and Gynecology,Chung Shan Medical University Hospital, Taichung, Taiwan, 13School of Medicine, Chung Shan Medical University, Taichung, Taiwan, 14Department of Medical Research, ChungShan Medical University Hospital, Taichung City, Taiwan.
Accepted 8 May 2015
Published online 00 Month 2015 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/hed.24124
ABSTRACT: Background. The existence of tumor-initiating cells (TICs)has been described in head and neck cancers. Plasminogen activatorinhibitor-1 (PAI-1) has been demonstrated to act as a prognostic factorin head and neck cancers.Methods. Tiplaxtinin (PAI-039), a specific inhibitor of PAI-1, and PAI-1-specific siRNA were used to examine the role of PAI-1 in the self-renewal property of head and neck cancer-TICs by tumorsphere forma-tion. Western blot, real-time polymerase chain reaction, and luciferase-based reporter assay were used to study the effect of PAI-039 in thesex-determining region Y-box 2 (Sox2) expression.Results. PAI-039 suppressed the self-renewal capability of head andneck cancer-TICs derived from head and neck cancer cell lines through
the inhibition of Sox2 expression. PAI-039 decreased the activity of thecore promoter and the enhancer of the Sox2 gene in head and neckcancer-TICs. Knockdown of PAI-1 expression also inhibited self-renewaland radioresistance properties of head and neck cancer-TICs.Conclusion. The inhibition of PAI-1 by PAI-039 or siRNA could suppresshead and neck cancer-TICs within head and neck cancer cell linesthrough the downregulation of Sox2. VC 2015 Wiley Periodicals, Inc. HeadNeck 00: 000–000, 2015
KEY WORDS: plasminogen activator inhibitor-1 (PAI-1), tiplaxtinin(PAI-039), tumor-initiating cells, head and neck cancer, Sox2
INTRODUCTIONHead and neck cancers are a group of cancers that occurin the paranasal sinuses, nasopharynx, oral cavities, oro-pharynx, hypopharynx, larynx, and salivary glands.1 Headand neck squamous cell carcinoma (HNSCC) accounts formore than 90% cases of head and neck cancers.2 Headand neck cancers account for 3% of all cancers in theUnited States3 and they are ranked sixth of the most com-mon cancers worldwide.4 The survival rate of head andneck cancers is near 50%, and the relatively high localrecurrence rate, the metastatic spread, and development ofsecond primary cancer in the head and neck region are
thought to be associated with the poor 5-year survivalrate of head and neck cancers.1
Tumor-initiating cells (TICs), or cancer stem cells, area subpopulation of cancer cells that have been shown toparticipate in tumor initiation, resistance to therapy, andmetastasis.5 In head and neck cancers, the reportedmarkers of TICs include CD44,2,6 CD133,7 aldehydedehydrogenase (ALDH),8 Grp78,9 side population,10 andc-Met.11 Head and neck cancer-TICs could also beenriched by tumorsphere cultivation.12 Transcriptionalfactors in the maintenance of embryonic stem cells havebeen frequently found to be overexpressed in cancers andTICs. It has been reported that the phosphorylated Nanogpromotes the growth of head and neck cancers throughdirect transactivation of Bmi1.13 The ectopic expressionof the sex-determining region Y-box 2 (Sox2) in HNSCCcell lines increased their self-renewal and chemoresistanceproperties.14 Sox2 expression in head and neck cancers isassociated with the recurrence and poor prognosis ofpatients.15,16
The urokinase plasminogen activator (uPA)-plasminsystem has been described to be associated with cancerprogression.17 Plasminogen activator inhibitor (PAI)21
*Corresponding author: P.-H. Wang, School of Medicine and Institute ofMedicine, Chung Shan Medical University, 110, Section 1, Chien-Kuo NorthRoad, Taichung 40201, Taiwan. E-mail: [email protected]. Chang, School of Biomedical Sciences, Chung Shan MedicalUniversity, 110, Section 1, Chien-Kuo North Road, Taichung 40201, Taiwan.E-mail: [email protected]
Contract grant sponsor: This work was supported by Chung Shan MedicalUniversity Hospital (grant no. CSH-2013-A-030) and Ministry of Science andTechnology in Taiwan (grant no. MOST-103-2314-B-040-015-MY3).
HEAD & NECK—DOI 10.1002/HED MONTH 2015 1

inhibits the proteolytic activity of uPA through directbinding to uPA to promote the endocytosis of the trimole-cules uPA/PAI-1/uPA receptor (uPAR) complex.18 PAI-1has been demonstrated to cause lower proliferation andhigher apoptosis in subcutaneous implanted tumors inPAI-1 deficiency mice when compared with the wild-typemice.19 In patients with HNSCC, the high level of PAI-1expression has been associated with a shorter disease-freesurvival as well as with tumors with perineural inva-sion.20 The concentration of PAI-1 was higher in tumors
of oral squamous cell carcinoma (OSCC) than in normaloral tissues.21 In patients with the early-stage OSCC, thelower PAI-1 expression was correlated with a lowdisease-specific death.22 PAI-1 has also been suggested toserve as a novel prognostic factor for patients withHNSCC.20 In addition to the clinical association of PAI-1and tumor progression, PAI-1 has been reported to playroles in tumor behaviors, such as inhibition of apoptosis,and promoting invasiveness and angiogenesis17; however,the role of PAI-1 in the maintenance of TICs remains
FIGURE 1. The cytotoxic effect of tiplaxtinin (PAI-039) in plasminogen activator inhibitor-1 (PAI-1) expressed oral cancer cell lines. (A) PAI-1 expres-sion of 3 oral cancer cell lines (SAS, GNM, or OC3) and primary human oral keratinocytes (HOKs) was determined by Western blot. (B, C) The cyto-toxic effect of PAI-039 in 3 PAI-1 expressed head and neck squamous cell carcinoma (HNSCC) cell lines (B) or HOK (C) was determined by WST-1reagent. Plots and IC50 values were created and calculated by GraFit software. GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
LEE ET AL.
2 HEAD & NECK—DOI 10.1002/HED MONTH 2015

unknown. Because of the pathological role of PAI-1 invascular diseases, small molecule inhibitors of PAI-1have been developed for preclinical characterization.Tiplaxtinin (also named as PAI-039) is an indole oxoace-tic acid derivative and exhibited oral efficacy in the ani-mal models of acute arterial thrombosis in vivo.23
Recently, PAI-039 also displayed anti-angiogenesis andantitumor growth activities in the human bladder cancerxenograft model in vivo.24
For the present study, we investigated the effect ofPAI-039 on head and neck cancer-TICs derived from
PAI-1 expressing human HNSCC cell lines. During treat-ment of PAI-039 at the concentration below IC50 value,the self-renewal capability and radioresistance propertiesof head and neck cancer-TICs were inhibited. We alsofound that PAI-039 could transcriptionally downregulatethe expression of Sox2 and Nanog. The activity of thecore promoter and the SRR2 enhancer of the Sox2 genecould be suppressed by PAI-039. Knockdown of PAI-1by siRNA also inhibited the expression of Sox2 andNanog as well as the self-renewal and radioresistanceproperties of head and neck cancer-TICs. Our results
FIGURE 2. Tiplaxtinin (PAI-039) suppressed self-renewal capability of head and neck cancer-tumor-initiating cells (TICs). Head and neck cancer-TICs were enriched by tumorsphere cultivation from SAS (A), GNM (B), or OC3 (C) cells and self-renewal capability was determined by the formationof secondary sphere under the treatment with 0.1% dimethyl sulfoxide (DMSO) or indicated concentration of PAI-039 for 7 days. *p< .05;**p< .01. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
PLASMINOGEN ACTIVATOR INHIBITOR–1 IN TUMOR–INITIATING CELLS OF HEAD AND NECK CANCER
HEAD & NECK—DOI 10.1002/HED MONTH 2015 3

suggest that targeting PAI-1 may be a novel strategyagainst head and neck cancer-TICs.
MATERIALS AND METHODS
Cell culture and regents SAS or GNM cells weremaintained in Dulbecco modified Eagle’s medium
The cells were maintained in Dulbecco modified Eagle’smedium (DMEM; Gibco, Invitrogen Corporation, Carls-bad, CA) containing 10% fetal bovine serum (FBS; Gibco).OC3 cells were kindly provided by Dr. Ying-Ray Lee(Department of Medical Research, Chiayi Christian Hospi-tal, Chiayi City, Taiwan) and maintained as previouslyreported.25 Human oral keratinocytes were purchased fromScienCell Research Laboratories (Carlsbad, CA) and main-tained in oral keratinocyte medium (ScienCell ResearchLaboratories). Cells were maintained in a standard humidi-fied incubator at 378C in 5% CO2. PAI-039 was purchasedfrom Axon MedChem (Groningen, Netherlands) and dis-
solved in dimethyl sulfoxide (DMSO) at a stock concentra-tion of 100 mM and stored at 2208C.
Cell proliferation/survival determination. Cells were platedin wells of 96-well plates as 1 3 104 cells/well in 0.1%DMSO or different concentration of PAI-039-containingmedium and cultured at 378C for 48 hours. Cell prolifera-tion/survival was determined by metabolic proliferationreagent WST-1 (Roche Applied Science, Mannheim, Ger-many). The 440 nm absorbance of the DMSO-treatedgroup was set as 100% and data were presented as per-centage of DMSO control. IC50 value was calculated byGraFitsoftware (Erithacus Software, Surrey, UK).
Enrichment of head and neck cancer-tumor-initiating cells fromhead and neck cancer cells and determination of self-renewalcapability by tumorsphere cultivation. SAS, GNM, or OC3cells were plated at a density of 104 live cells/10-mmultralow attachment dishes (Corning, Tewksbury, MA)
FIGURE 3. Aldehyde dehydrogenase (ALDH)-positive SAS cells display tumor-initiating cell (TIC) characteristics. SAS cells were stained with ALDE-FLUOR substrate to differentiate into ALDH-negative or ALDH-positive population of cells and sorted by fluorescence activated cell sorter (FACS).(A) It showed the purity of sorted ALDH-negative or ALDH-positive cells. (B) The sorted ALDH-positive cells were cultured in tumorsphere conditionand the expression of ALDH1A1 (green fluorescence) was determined in the formed tumorspheres. Red fluorescence indicated the cell nuclei. (C)The abilities of colony formation, (D) cell migration, (E) or cell invasion were compared between sorted ALDH-negative and ALDH-positive cells. (F)ALDH-negative or ALDH-positive cells were subcutaneously injected into nude mice as indicated by the cell numbers for tumorigenicity determina-tion and recorded for 2 months. (G) The tumor volumes in 1 3 105 groups were shown. DEAB, diethylaminobenzaldehyde. *p< .05. [Color figurecan be viewed in the online issue, which is available at wileyonlinelibrary.com.]
LEE ET AL.
4 HEAD & NECK—DOI 10.1002/HED MONTH 2015

with serum-free DMEM/F12 medium (Gibco), N2 supple-ment (Gibco), 10 ng/mL human recombinant basic fibro-blast growth factor-basic (Novus Biologicals, Littleton,CO), and 10 ng/mL epidermal growth factor (PeproTech,Rocky Hill, NJ) and the medium was changed every 3days until the tumorsphere formation was observed inabout 4 weeks.26 For determination of self-renewal capa-bility of head and neck cancer-TICs, primary tumor-spheres derived from HNSCC cell lines were dissociatedinto single cell suspension by HyQTase solution (GEHealthcare HyClone, Logan, UT) at 378C for 5 minutes,and plated at a density of 103 live cells/well of ultralowattachment 6-well plates (Corning) with the mediumdescribed above. The number of secondary tumorsphereswas counted by inverted microscopy at day 7.
ALDEFLUOR assay, cell sorting and determination of radiosensi-tivity. ALDH-negative or ALDH-positive SAS cells werestained with ALDEFLUOR assay kit (StemCell Technolo-gies, Vancouver, British Columbia, Canada) after our previ-ous report27 and were sorted by FACSAria II cell sorter (BDBiosciences, San Jose, CA). For determination of radiosensi-tivity of ALDH-negative or ALDH-positive SAS cells withor without PAI-039 treatment, 5 3 104 sorted cells/tubewere suspended in DMEM/10% FBS medium in 1.5 mLmicrotubes and irradiated by Elekta Axesse linear accelera-tor (Elekta AB, Stockholm, Sweden) at a dose rate of 6 Gymin21. Irradiated cells were then seeded into wells of 96-well plates at a density of 1 3 104/well and cultured for 72hours. Cell viability was determined by Thiazolyl Blue Tet-razolium Blue (MTT; Sigma–Aldrich, St. Louis, MO).
Tumorigenicity determination. All the animal experimentswere approved by the Institutional Animal Care and UseCommittee of Chung Shan Medical University, Taichung,Taiwan (Institutional Animal Care and Use Committeeapproval no. 1303). Tumorigenicity of different cell num-bers of sorted ALDH-negative or ALDH-positive SAS cellswas determined by xenograftment assay in BALB/c nudemice, as previously reported.28 Tumor volume was meas-ured twice per week until 2 months and calculated accord-ing to the formula: tumor volume 5 (length 3 width2)/2.
Immunofluorescence analysis. Tumorspheres derived fromsorted ALDH-positive SAS cells were transferred ontocollagen-coated coverslips for adhesion overnight and thenfixed/permeabilized with cold acetone/methanol for 5minutes. After blocking with 1% bovine serum albumin/phosphate-buffered saline, coverslips were incubated withmouse anti-human ALDH1A1 antibody (BD Biosciences)at 48C overnight followed by incubation of Alexa-488 con-jugated anti-mouse immunoglobulin G antibody (JacksonImmunoResearch, West Grove, PA). Tumorspheres werecounterstained with Nuclear-ID Red DNA stain dye (EnzoLife Sciences, Farmingdale, NY) and the fluorescence sig-nals were recorded by inverted fluorescence microcopy(Carl Zeiss Microscopy GmbH, Oberkochen, Germany).
Soft agar colony formation assay
The 2 3 104 cells were suspended in 0.3% agar/DMEM containing 10% FBS, loaded onto 0.6% agar/
DMEM/10% FBS-coated wells of 6-well plates and cul-tured at 378C for 4 weeks. Wells were stained with crys-tal violet and counted colonies with a diameter �100 mm.
In vitro cell migration/invasion assay. Cell migration/invasionassay was performed by transwell chambers (Corning), asour previous report.29 Briefly, 1 3 105 cells were suspendedin 0.5% FBS DMEM medium, loaded onto the upper cham-ber with a porous membrane (8.0 mm pore size), inserted intothe lower chamber containing 10% FBS DMEM medium,and cultured at 378C for 12 hours. After removing the cellsdid not migrate across the membrane with a cotton swab, themigrated cells were stained with crystal violet and countedunder an inverted microscopy. For invasion assay, the mem-brane was coated with 0.5 mg/mL matrigel (BD Biosciences)before cell loading and then followed the protocol of migra-tion described above.
Western blot analysis. Cells were lysed with NP-40 lysisbuffer and 25 mg of total protein were separated bysodium dodecyl sulfate-polyacrylamide gel electrophoresisand transferred to polyvinylidene fluoride membrane. Pro-tein detection was conducted as in our previous report.30
Nestin antibody was purchased from Santa Cruz Biotech-nology (Dallas, TX), Nanog antibody was purchased fromCell Signaling Technology (Danvers, MA), Sox2 antibodywas purchased from Novus Biologicals and glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) antibody waspurchased from GeneTex International Corporation (Hsin-chu, Taiwan).
FIGURE 4. Tiplaxtinin (PAI-039) potentiated the radiosensitivity ofhead and neck cancer-tumor-initiating cells (TICs). Aldehydedehydrogenase (ALDH)-negative or ALDH-positive cells weresorted from SAS cells and irradiated at a dose rate of 6 Gy min21
in the presence of 0.1% dimethyl sulfoxide (DMSO) or 70 mM PAI-039. Irradiated cells were then plated into 96-well plates at adensity of 1 3 104 and cultured for 72 hours. Cell viability wasdetermined with MTT assay and presented as survival fraction tononirradiated/0.1% DMSO group. *p< .01 when the ALDH-positive group was compared with the ALDH-negative group;¶p< .01 when the ALDH-positive/PAI-039 group was comparedwith the ALDH-positive group; ‡p< .01 when the ALDH-negative/PAI-039 group was compared with the ALDH-negative group.
PLASMINOGEN ACTIVATOR INHIBITOR–1 IN TUMOR–INITIATING CELLS OF HEAD AND NECK CANCER
HEAD & NECK—DOI 10.1002/HED MONTH 2015 5
Quantitative real-time reverse transcriptase-polymerase chainreaction. Total RNA was extracted using a Quick RNAMiniPrep kit (Zymo Research, Irvine, CA) and reversetranscribed to cDNA using oligo(dT) primer (RevertAidFirst Strand cDNA Synthesis Kit, Fermentas). Reversetranscriptase-polymerase chain reaction (RT-PCR) forsimultaneous detection and quantification of the cDNAsamples was performed on an ABI StepOnePlus Real-Time PCR System and analyzed with the StepOne soft-ware (Applied Biosystems, Life Technologies, Carlsbad,CA). Fifty nanograms of cDNA sample were used in anSYBR Green-based quantitative polymerase chain reac-tion reaction after the cycling condition and calculation,as previously reported.30 The primer sequences used inthis study were as follows:
PAI-1: 50-GGAATGACCGACATGTTCAG-30 and 50-ACTCTCGTTCACCTCGATCT-30;
Nanog: 50-CATGAGTGTGGATCCAGCTTG-30 and 50-CCTGAATAAGCAGATCCATGG-30;
Sox2: 50-AACCCCAAGATGCACAACTC-30 and 50-CGGGGCCGGTATTTATAATC-30;
MRPL19 (internal control): 50-GGGATTTGCATTCAGAGATCAG-30 and 50- GGAAGGGCATCTCGTAAG-30.
Luciferase-based reporter assay. The core promoter, SRR1or SRR2 enhancer of Sox2 gene, was cloned into pGL3-basic luciferase reporter vector (Promega, Madison, WI)using the following primers:
Core promoter: 50-CAAGTGGCTAGCCGCTTAGCCTCGTCGATGAAC-30 and 50-CAAGTGGCTAGCGCGGGCGCTGTGCGCGGGCCC-30;
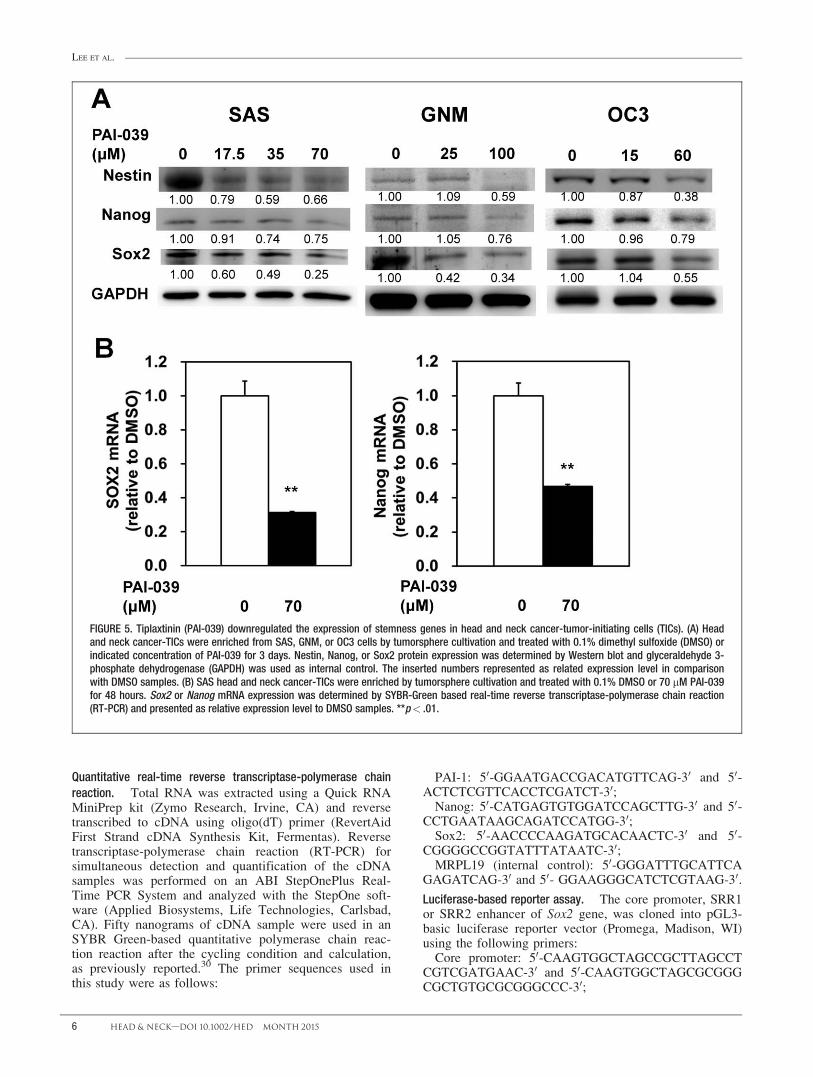
FIGURE 5. Tiplaxtinin (PAI-039) downregulated the expression of stemness genes in head and neck cancer-tumor-initiating cells (TICs). (A) Headand neck cancer-TICs were enriched from SAS, GNM, or OC3 cells by tumorsphere cultivation and treated with 0.1% dimethyl sulfoxide (DMSO) orindicated concentration of PAI-039 for 3 days. Nestin, Nanog, or Sox2 protein expression was determined by Western blot and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as internal control. The inserted numbers represented as related expression level in comparisonwith DMSO samples. (B) SAS head and neck cancer-TICs were enriched by tumorsphere cultivation and treated with 0.1% DMSO or 70 mM PAI-039for 48 hours. Sox2 or Nanog mRNA expression was determined by SYBR-Green based real-time reverse transcriptase-polymerase chain reaction(RT-PCR) and presented as relative expression level to DMSO samples. **p< .01.
LEE ET AL.
6 HEAD & NECK—DOI 10.1002/HED MONTH 2015

SRR1: 50-CACTTGGAGCTCGTTACACTTAACAGTTAAGGC-30 and 50-CAAGTGGCTAGCGGTTAAAGGGGTTGGCTAGG-30;
SRR2: 50-CGGAGCTCCTTTTTTAATCCTCATTTTTA-30
and 50-CGGCTAGCTTTTAAAAAATGAGAGGTGAG-30.The assay was conducted using a dual reporter assay
system according to our previous report.30
RNA interference. The specific siRNA oligos of PAI-1were purchased from Qiagen Sciences (Frederick, MD)and the sequences were listed as follows: 50-GGAGGACCUUUAGGUCAAATT-30 (sense) and 50-UUUGACCUAAAGGUCCUCCAA-30 (anti-sense). A negative controlsiRNA oligos were purchased from MDBio (Taipei,Taiwan) and the sequences were listed as follows:
50-GCGACGAUCUGCCUAAGAUTT-30 (sense) and50-AUCUUAGGCAGAUCGUCGCTT-30 (anti-sense).RNA oligos were transfected by Turbofect transfectionreagent (Fermentas, Hanover, MD), in accord with themanufacturer’s recommendation.
RESULTS
Cytotoxic effect of PAI-039 in oral cancer cell lines
We first examined the cytotoxic activity of PAI-039 inoral cancer cell lines. Three oral cancer cell lines (SAS,GNM, and OC3) used in this study were confirmed toexpress PAI-1 (Figure 1A). During the WST-1 assay,PAI-039 displayed cytotoxic effects to these PAI-1expressing oral cancer cell lines. The IC50 values of PAI-039 in SAS, GNM, or OC3 cells were (81.0 6 1.7) mM,(145.7 6 22.9) mM, or (75.7 6 1.7) mM, respectively (Fig-ure 1B). We also examined the cytotoxic effect of PAI-039 to human primary normal oral keratinocytes andfound that PAI-039 had no obvious cytotoxicity when theconcentration was tested up to 200 mM (Figure 1C).
PAI-039 impaired self-renewal capability and theincreased radiosensitivity of head and neckcancer-tumor-initiating cells
We next investigated the effect of PAI-039 on the self-renewal capability of head and neck cancer-TICs bytumorsphere cultivation, a common method used to enrichTICs and quantify their self-renewal capability.31 Thehead and neck cancer-TICs were first enriched from dis-sociated primary tumorspheres derived from SAS, GNM,or OC3 cells to examine the effect of PAI-039 on theself-renewal capability of head and neck cancer-TICs. Toavoid a cytotoxic effect, the PAI-039 concentrations usedin this study were below the IC50 values of each parentalcell line. With the dose-dependent treatment of PAI-039,the self-renewal capability was significantly suppressed inSAS (Figure 2A), GNM (Figure 2B), and OC3 (Figure2C) cells. It has been reported that TICs display a radio-resistance property32 and that the PAI-1 level in HNSCCcell lines is correlated with the responses of in vivo irra-diation.33 We next investigated the effect of PAI-039 onthe radiosensitivity of head and neck cancer-TICs. Headand neck cancer-TICs could also be identified as cancercells with high intracellular ALDH activity (ALDH-posi-tive)8 and ALDH expression have been recently demon-
strated to be associated with the worth prognosis inpatients with HNSCC.34 During ALDEFLUOR assay, theSAS cells were separated into ALDH-negative (generalHNSCC cells) or ALDH-positive (head and neck cancer-TICs), and the properties of TICs were determined. Thepurity of the ALDH-negative or ALDH-positive SAScells was 98.8% or 95.8%, respectively (Figure 3A). TheALDH-positive SAS cells could form tumorspheres withthe expression of ALDH1A1 (Figure 3B). The ALDH-positive SAS cells displayed a greater activity of colonyformation (Figure 3C), cell migration (Figure 3D), andcell invasion (Figure 3E) than the ALDH-negative cells.As few as 1 3 103 ALDH-positive SAS cells formedtumors when injected into nude mice (Figure 3F).Although 1 3 105 ALDH-negative SAS cells also formedtumors in nude mice, the tumor volumes were signifi-cantly smaller than the tumors derived from the samenumber of ALDH-positive cells (Figure 3G). Theseresults demonstrated that ALDH-positive SAS cells dis-play TIC characteristics. We next examined the cell sur-vival of ALDH-negative and ALDH-positive SAS cellsafter irradiation at dosages of 2, 4, 8, 16, or 32 Gy. Asshown in Figure 4, ALDH-positive SAS indeed displayeda radioresistant phenotype, and the treatment of 70 mMPAI-039 enhanced the efficacy of radiation in bothALDH-negative and ALDH-positive SAS cells (Figure 4).
PAI-039 inhibited the expression of stemness genes inhead and neck cancer-tumor-initiating cells
We next examined the effect of PAI-039 on the expres-sion of stemness genes in tumorspheres derived fromSAS, GNM, or OC3 cells. During the Western blot analy-sis, the expression of nestin, Nanog, or Sox2 was dose-dependently inhibited by PAI-039 in tumorspheresderived from all 3 HNSCC cell lines (Figure 5A). It hasbeen demonstrated that Sox2 transcriptionally controls
FIGURE 6. Tiplaxtinin (PAI-039) inhibited the activity of the corepromoter and the SRR2 enhancer of THE Sox2 gene in head andneck cancer-tumor-initiating cells (TICs). Head and neck cancer-TICs were enriched from SAS cells by tumorsphere cultivationand transfected with indicated luciferase reported vectors for 8hours. Transfected cells were further treated with 0.1% dimethylsulfoxide (DMSO) or 70 mM PAI-039 for 48 hours and the celllysate was collected for luciferase activity determination.**p< .01.
PLASMINOGEN ACTIVATOR INHIBITOR–1 IN TUMOR–INITIATING CELLS OF HEAD AND NECK CANCER
HEAD & NECK—DOI 10.1002/HED MONTH 2015 7

Nanog expression.35 We further determined the mRNAexpression of Nanog or Sox2 in PAI-039-treated second-ary tumorspheres from the SAS cell line. During thequantitative RT-PCR analysis, PAI-039 displayed a dose-dependent inhibition of Nanog or Sox2 (Figure 5B)mRNA expression in SAS tumorsphere cells.
PAI-039 inhibited the activity of the core promoter andthe SRR2 enhancer of the Sox2 gene
The transcription of the Sox2 gene could be regulatedby a core promoter36 or 2 enhancers, SRR1 or SRR2.37
To investigate the mechanism of PAI-039 induced down-regulation of Sox2 transcription, we cloned the core pro-moter or enhancers (SRR1 or SRR2) in the Sox2 geneinto pGL3-basic luciferase reporter plasmid. After weenriched the head and neck cancer-TICs from the SAStumorspheres, the head and neck cancer-TICs were trans-fected with the core promoter, or an SRR1 or SRR2reporter plasmid, and treated with 70 mM PAI-039 for 48
hours. As shown in Figure 5, PAI-039 downregulated theactivity of the core promoter or the SRR2 enhancer, butnot the SRR1, of the Sox2 gene (Figure 6).
Knockdown of PAI-1 by the specific siRNA inhibited self-renewal of head and neck cancer-tumor-initiating cells
We used PAI-1-specific siRNA oligos to knockdownthe PAI-1 expression in the SAS tumorsphere cells andexamined their self-renewal property. The PAI-1-specificsiRNA oligos efficiently knockdown the expression ofPAI-1 after being transfected into the SAS tumorspherecells (Figure 7A). The PAI-1 knockdown SAS tumor-sphere cells formed small secondary tumorspheres andcaused a reduction in the sphere number in comparisonwith the control RNA oligos transfected group (Figure7B). We also found that the expression of Sox2 andNanog was downregulated in PAI-1 knockdown SAStumorspheres at the mRNA (Figure 7C) and protein level(Figure 7D). Furthermore, the knockdown of PAI-1 in theSAS tumorspheres enhanced their sensitivity to radiation
FIGURE 7. Knockdown of plasminogen activator inhibitor-1 (PAI-1) suppressed the self-renewal capability of head and neck cancer-tumor-initiating cells (TICs). Head and neck cancer-TICs were enriched from SAS cells by tumorsphere cultivation and transfected with negative control(Ctrl) or the PAI-1 specific (KD) siRNA oligos for 8 hours. (A) The PAI-1 protein expression was determined at day 2 post-transfection by Westernblot. (B) Transfected cells were performed by secondary sphere formation to determine the self-renewal capability. Secondary spheres werecounted at day 7 and were divided into 2 groups according to the diameter (>100 mm or 50–100 mm). (C) The mRNA expression of PAI-1, Sox2, orNanog in siRNA oligos transfected secondary spheres was determined by SYBR-Green based quantitative polymerase chain reaction at day 2 post-transfection. (D) The protein expression of nestin, Sox2, or Nanog in siRNA oligos transfected secondary spheres was determined by Western blotat day 3 post-transfection. (E) The siRNA oligos transfected secondary spheres were irradiated as indicated dose and cultured for 72 hours anddetermined the cell viability by MTT. GAPDH, glyceraldehyde 3-phosphate dehydrogenase. *p< .05; **p< .01.
LEE ET AL.
8 HEAD & NECK—DOI 10.1002/HED MONTH 2015

(Figure 7E). Based on these results, the knockdown ofPAI-1 with specific RNA oligos in head and neck cancer-TICs could inhibit their self-renewal and radioresistanceproperties.
DISCUSSIONPAI-1 has been known to be a metastasis-associated
gene in cancer. In transforming growth factor-b inducedepithelial-mesenchymal transition (EMT), PAI-1 wasinduced through Sp1 activation.38 EMT has been consid-ered to be an important induction mechanism in the gen-eration of TICs.39 The treatment of transforming growthfactor-b or the forced expression of EMT-related tran-scriptional factors (Snail1 or Twist1) in immortalizedhuman mammary epithelial cells caused cells to displayTIC behaviors.39 Although the function of PAI-1 in cellmotility or in the invasiveness of cancer cells is well-known,40 the involvement of PAI-1 in TIC biologyremains unknown. In this study, we provide evidence thatthe treatment of PAI-039, a PAI-1 inhibitor, or the knock-down of PAI-1 expression by siRNA could inhibit theself-renewal capability head and neck cancer-TICs (Fig-ures 2 and 7B). To our knowledge, this is the first reportto demonstrate that PAI-1 is involved in the maintenanceof TICs.
We found that the inhibition of PAI-1 by PAI-039 orsiRNA-mediated gene silence could increase the radiosen-sitivity of head and neck cancer-TICs derived from SAScells (Figures 4 and 7E). It has been reported that irradia-tion leads to a dose-dependent increase of PAI-1 expres-sion in several HNSCC cell lines.41,42 The correlationbetween PAI-1 and radiotherapy has also been reported inother types of cancer, such as rectal43 or liver44 cancer. Inaddition to head and neck cancer-TICs, the inhibition ofPAI-1 by PAI-039 also sensitized non-TICs of head andneck cancer cells to radiation (Figure 4). Recently, it hasbeen demonstrated that the non-TIC cancer cells coulddedifferentiate to TICs through microenvironment signals,signal transduction pathways, or transcriptional net-works.45 This indicates that a successful strategy of can-cer treatment will be to target both TICs and non-TICswith combination therapy. Based on our observation andothers studies, the inhibition of PAI-1 with PAI-1 inhibi-tors may be considered to be a combination therapy withcancer radiation treatment. During the development ofanticancer agents, it often requires optimization from theearly generation of lead compounds. For example, suber-oylanilide hydroxamic acid, a potent inhibitor of histonedeacetylases that works at a low mM range and is cur-rently being used in advanced clinical trials for the treat-ment of cancer,46 was developed from hexamethylenebisacetamide, which suppressed cell proliferation ofmurine erythroleukemia cells at 5 mM.46 Although theeffective concentration of PAI-039 in this study rangedfrom 25 to 100 mM, PAI-039 could serve as a lead com-pound for future development of more potent compoundsin the suppression of PAI-1. It is also beneficial to exam-ine the additive or synergistic effects when a combinationof PAI-039 and other anticancer agents are used in treat-ment of head and neck cancer cells.
In addition to the pericellular proteolysis role of theuPA-uPAR-PAI-1 system, PAI-1 could function as a sig-
nal molecule to regulate cell behavior. Through engage-ment with the low-density lipoprotein receptor-relatedprotein-1, PAI-1 could increase cell motility through theJak/Stat pathway.47,48 Recently, it has been reported thatthe activation of Jak/Stat by vitamin C could enhanceNanog expression in mouse embryonic stem cells orembryonal carcinoma cells.49 It would be interesting toinvestigate whether or not the Jak/Stat pathway isinvolved in the PAI-1 mediated self-renewal capability ofhead and neck cancer-TICs. In a study of mouse macro-phages, PAI-1 could induce the phosphorylation of focaladhesion kinase (FAK) to enhance the migration of mac-rophages.50 Treatment of FAK inhibitors or knockdownFAK expression by siRNA could reduce the self-renewalcapability and increase the radiosensitivity of ductal carci-noma in situ of breast cancer.51 The FAK expression inOSCC was significantly correlated with tumor size, necknode metastasis, and local recurrence.52 In a gliomastudy, the silence of Sox2 expression led to the reducedphosphorylation of FAKY397 and caused the inhibition ofcell invasiveness.53 It would also be interesting to studythe regulation of PAI-1, FAK, and Sox2 in head and neckcancer-TICs.
In conclusion, our results demonstrate that the inhibi-tion of PAI-1 by PAI-039 or siRNA mediated genesilencing reduces the self-renewal capability and increasesthe radiosensitivity of head and neck cancer-TICs. TheTIC targeting effect of PAI-039 is suggested to be associ-ated with the transcriptional downregulation of Sox2expression. The inhibition of PAI-1 may be considered tobe a therapeutic strategy in targeting TICs in HNSCC.
REFERENCES1. Machiels JP, Lambrecht M, Hanin FX, et al. Advances in the management
of squamous cell carcinoma of the head and neck. F1000Prime Rep 2014;6:44.
2. Kokko LL, Hurme S, Maula SM, et al. Significance of site-specific progno-sis of cancer stem cell marker CD44 in head and neck squamous-cell carci-noma. Oral Oncol 2011;47:510–516.
3. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer JClin 2013;63:11–30.
4. Haddad RI, Shin DM. Recent advances in head and neck cancer. N Engl JMed 2008;359:1143–1154.
5. Hermann PC, Bhaskar S, Cioffi M, Heeschen C. Cancer stem cells in solidtumors. Semin Cancer Biol 2010;20:77–84.
6. Prince ME, Sivanandan R, Kaczorowski A, et al. Identification of a subpo-pulation of cells with cancer stem cell properties in head and neck squa-mous cell carcinoma. Proc Natl Acad Sci U S A 2007;104:973–978.
7. Chen YS, Wu MJ, Huang CY, et al. CD133/Src axis mediates tumor initiat-ing property and epithelial-mesenchymal transition of head and neck can-cer. PLoS One 2011;6:e28053.
8. Clay MR, Tabor M, Owen JH, et al. Single-marker identification of headand neck squamous cell carcinoma cancer stem cells with aldehyde dehy-drogenase. Head Neck 2010;32:1195–1201.
9. Wu MJ, Jan CI, Tsay YG, et al. Elimination of head and neck cancer ini-tiating cells through targeting glucose regulated protein78 signaling. MolCancer 2010;9:283.
10. Tabor MH, Clay MR, Owen JH, et al. Head and neck cancer stem cells: theside population. Laryngoscope 2011;121:527–533.
11. Sun S, Wang Z. Head neck squamous cell carcinoma c-Met1 cells displaycancer stem cell properties and are responsible for cisplatin-resistance andmetastasis. Int J Cancer 2011;129:2337–2348.
12. Lim YC, Oh SY, Cha YY, Kim SH, Jin X, Kim H. Cancer stem cell traitsin squamospheres derived from primary head and neck squamous cell car-cinomas. Oral Oncol 2011;47:83–91.
13. Xie X, Piao L, Cavey GS, et al. Phosphorylation of Nanog is essential toregulate Bmi1 and promote tumorigenesis. Oncogene 2014;33:2040–2052.
14. Lee SH, Oh SY, Do SI, et al. SOX2 regulates self-renewal and tumorige-nicity of stem-like cells of head and neck squamous cell carcinoma. Br JCancer 2014;111:2122–2130.
15. Li W, Li B, Wang R, Huang D, Jin W, Yang S. SOX2 as prognostic factorin head and neck cancer: a systematic review and meta-analysis. Acta Oto-laryngol 2014;134:1101–1108.
PLASMINOGEN ACTIVATOR INHIBITOR–1 IN TUMOR–INITIATING CELLS OF HEAD AND NECK CANCER
HEAD & NECK—DOI 10.1002/HED MONTH 2015 9

16. Schr€ock A, Bode M, G€oke FJ, et al. Expression and role of the embryonicprotein SOX2 in head and neck squamous cell carcinoma. Carcinogenesis2014;35:1636–1642.
17. Van De Craen B, Declerck PJ, Gils A. The biochemistry, physiology andpathological roles of PAI-1 and the requirements for PAI-1 inhibition invivo. Thromb Res 2012;130:576–585.
18. Binder BR, Christ G, Gruber F, et al. Plasminogen activator inhibitor 1: physi-ological and pathophysiological roles. News Physiol Sci 2002;17:56–61.
19. Gutierrez LS, Schulman A, Brito–Robinson T, Noria F, Ploplis VA,Castellino FJ. Tumor development is retarded in mice lacking the gene forurokinase-type plasminogen activator or its inhibitor, plasminogen activa-tor inhibitor-1. Cancer Res 2000;60:5839–5847.
20. Speleman L, Kerrebijn JD, Look MP, Meeuwis CA, Foekens JA, BernsEM. Prognostic value of plasminogen activator inhibitor-1 in head andneck squamous cell carcinoma. Head Neck 2007;29:341–350.
21. Baker EA, Leaper DJ, Hayter JP, Dickenson AJ. Plasminogen activator sys-tem in oral squamous cell carcinoma. Br J Oral Maxillofac Surg 2007;45:623–627.
22. Magnussen S, Rikardsen OG, Hadler–Olsen E, Uhlin–Hansen L, SteigenSE, Svineng G. Urokinase plasminogen activator receptor (uPAR) and plas-minogen activator inhibitor-1 (PAI-1) are potential predictive biomarkersin early stage oral squamous cell carcinomas (OSCC). PLoS One 2014;9:e101895.
23. Elokdah H, Abou–Gharbia M, Hennan JK, et al. Tiplaxtinin, a novel, orallyefficacious inhibitor of plasminogen activator inhibitor-1: design, synthesis,and preclinical characterization. J Med Chem 2004;47:3491–3494.
24. Gomes–Giacoia E, Miyake M, Goodison S, Rosser CJ. Targeting plasmino-gen activator inhibitor-1 inhibits angiogenesis and tumor growth in ahuman cancer xenograft model. Mol Cancer Ther 2013;12:2697–2708.
25. Lin SC, Liu CJ, Chiu CP, Chang SM, Lu SY, Chen YJ. Establishment ofOC3 oral carcinoma cell line and identification of NF-kappa B activationresponses to areca nut extract. J Oral Pathol Med 2004;33:79–86.
26. Chang WW, Hu FW, Yu CC, et al. Quercetin in elimination of tumor ini-tiating stem-like and mesenchymal transformation property in head andneck cancer. Head Neck 2013;35:413–419.
27. Wei L, Liu TT, Wang HH, et al. Hsp27 participates in the maintenance ofbreast cancer stem cells through regulation of epithelial-mesenchymal tran-sition and nuclear factor-jB. Breast Cancer Res 2011;13:R101.
28. Tsai LL, Hu FW, Lee SS, Yu CH, Yu CC, Chang YC. Oct4 mediates tumorinitiating properties in oral squamous cell carcinomas through the regula-tion of epithelial-mesenchymal transition. PLoS One 2014;9:e87207.
29. Chang WW, Lin RJ, Yu J, et al. The expression and significance of insulin-like growth factor-1 receptor and its pathway on breast cancer stem/progen-itors. Breast Cancer Res 2013;15:R39.
30. Chang YC, Tsai CH, Lai YL, et al. Arecoline-induced myofibroblast trans-differentiation from human buccal mucosal fibroblasts is mediated byZEB1. J Cell Mol Med 2014;18:698–708.
31. Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulat-ing evidence and unresolved questions. Nat Rev Cancer 2008;8:755–768.
32. Rycaj K, Tang DG. Cancer stem cells and radioresistance. Int J Radiat Biol2014;90:615–621.
33. Bayer C, Schilling D, Hoetzel J, et al. PAI-1 levels predict response to frac-tionated irradiation in 10 human squamous cell carcinoma lines of the headand neck. Radiother Oncol 2008;86:361–368.
34. Zhou C, Sun B. The prognostic role of the cancer stem cell marker alde-hyde dehydrogenase 1 in head and neck squamous cell carcinomas: a meta-analysis. Oral Oncol 2014;50:1144–1148.
35. Rodda DJ, Chew JL, Lim LH, et al. Transcriptional regulation of nanog byOCT4 and SOX2. J Biol Chem 2005;280:24731–24737.
36. Leis O, Eguiara A, Lopez–Arribillaga E, et al. Sox2 expression in breasttumours and activation in breast cancer stem cells. Oncogene 2012;31:1354–1365.
37. Miyagi S, Saito T, Mizutani K, et al. The Sox-2 regulatory regions displaytheir activities in two distinct types of multipotent stem cells. Mol Cell Biol2004;24:4207–4220.
38. Datta PK, Blake MC, Moses HL. Regulation of plasminogen activatorinhibitor-1 expression by transforming growth factor-beta -induced physi-cal and functional interactions between smads and Sp1. J Biol Chem 2000;275:40014–40019.
39. Mani SA, Guo W, Liao MJ, et al. The epithelial-mesenchymal transitiongenerates cells with properties of stem cells. Cell 2008;133:704–715.
40. Kwaan HC, Mazar AP, McMahon BJ. The apparent uPA/PAI-1 paradox incancer: more than meets the eye. Semin Thromb Hemost 2013;39:382–391.
41. Schilling D, Bayer C, Geurts–Moespot A, et al. Induction of plasminogenactivator inhibitor type-1 (PAI-1) by hypoxia and irradiation in humanhead and neck carcinoma cell lines. BMC Cancer 2007;7:143.
42. Artman T, Schilling D, Gnann J, Molls M, Multhoff G, Bayer C. Irradia-tion-induced regulation of plasminogen activator inhibitor type-1 and vas-cular endothelial growth factor in six human squamous cell carcinoma linesof the head and neck. Int J Radiat Oncol Biol Phys 2010;76:574–582.
43. Angenete E, Langenski€old M, Palmgren I, Falk P, Oresland T, IvarssonML. uPA and PAI-1 in rectal cancer–relationship to radiotherapy and clini-cal outcome. J Surg Res 2009;153:46–53.
44. Hageman J, Eggen BJ, Rozema T, Damman K, Kampinga HH, Coppes RP.Radiation and transforming growth factor-beta cooperate in transcriptionalactivation of the profibrotic plasminogen activator inhibitor-1 gene. ClinCancer Res 2005;11:5956–5964.
45. Li Y, Laterra J. Cancer stem cells: distinct entities or dynamically regulatedphenotypes? Cancer Res 2012;72:576–580.
46. Marks PA. Discovery and development of SAHA as an anticancer agent.Oncogene 2007;26:1351–1356.
47. Hou SX, Zheng Z, Chen X, Perrimon N. The Jak/STAT pathway in modelorganisms: emerging roles in cell movement. Dev Cell 2002;3:765–778.
48. Degryse B, Neels JG, Czekay RP, Aertgeerts K, Kamikubo Y, LoskutoffDJ. The low density lipoprotein receptor-related protein is a motogenicreceptor for plasminogen activator inhibitor-1. J Biol Chem 2004;279:22595–22604.
49. Wu H, Wu Y, Ai Z, et al. Vitamin C enhances Nanog expression via activa-tion of the JAK/STAT signaling pathway. Stem Cells 2014;32:166–176.
50. Thapa B, Koo BH, Kim YH, Kwon HJ, Kim DS. Plasminogen activatorinhibitor-1 regulates infiltration of macrophages into melanoma via phos-phorylation of FAK-Tyr925. Biochem Biophys Res Commun 2014;450:1696–1701.
51. Williams KE, Bundred NJ, Landberg G, Clarke RB, Farnie G. Focal adhe-sion kinase and Wnt signaling regulate human ductal carcinoma in situstem cell activity and response to radiotherapy. Stem Cells 2015;33:327–341.
52. de Vicente JC, Rosado P, Lequerica–Fern�andez P, Allonca E, Villalla�ın L,Hern�andez–Vallejo G. Focal adhesion kinase overexpression: correlationwith lymph node metastasis and shorter survival in oral squamous cell car-cinoma. Head Neck 2013;35:826–830.
53. Oppel F, M€uller N, Schackert G, et al. SOX2-RNAi attenuates S-phaseentry and induces RhoA-dependent switch to protease-independent amoe-boid migration in human glioma cells. Mol Cancer 2011;10:137.
LEE ET AL.
10 HEAD & NECK—DOI 10.1002/HED MONTH 2015
Related Documents



![Tissue-Type Plasminogen Activator-Mediated Activation of ... · TISSUE PLASMINOGEN ACTIVATOR IN STREPTOCOCCAL BINDING 197 sodium phosphate, 0.14 Msodium chloride [pH 7.4]) con- taining0.02%(wt/vol)](https://static.cupdf.com/doc/110x72/5f46a6d9df5f79688c496b2a/tissue-type-plasminogen-activator-mediated-activation-of-tissue-plasminogen.jpg)







