Phenotypic profiling and gene expression analyses for aromatic and volatile compounds in Chamoes (Cucumis melo) Jeongyeo Lee • Min Keun Kim • Seung Hwan Hwang • Jungeun Kim • Jong Moon Ahn • Sung Ran Min • Sang Un Park • Soon Sung Lim • HyeRan Kim Received: 21 May 2013 / Accepted: 27 January 2014 / Published online: 11 February 2014 Ó Springer Science+Business Media Dordrecht 2014 Abstract Gotgam chamoe (GgC), a native oriental melon in Korea, is known to possess the aroma of a dried per- simmon, an agronomic relevance for melon breeding pro- gram. The volatile compounds and the transcript levels of aromatic compound genes in cultivar (Ohbokggul chamoe [OC]) and GgC were profiled. A total of 62 volatile com- pounds were identified and quantified. Twenty-eight vola- tile compounds were specific to either the OC or the GgC. The amounts of volatile alcohol, saturated hydrocarbon, and unsaturated hydrocarbon compounds were 2.2, 2.7, and 1.1 times higher in OC, respectively. The amounts of ketone volatiles were 1.2 times higher in GgC, whereas the total amounts of esters were similar. In the shikimate pathway, transcriptional patterns with the fruit parts were different between the two chamoes for CmDAHPS, CmDHD/SDH, and CmEPSPS. The expression levels of all six genes investigated, especially CmCS, were highest in the peel of both chamoes compared to the other parts. The transcript levels of the aromatic amino acid biosynthesis genes demonstrate that phenylalanine and tyrosine are present more in edible parts of the chamoe, while trypto- phan may be accumulated low in the chamoe. In addition, phenylalanine and tryptophan are synthesized more in GgC than the OC. Keywords Cucumis melo Chamoe Volatile aromatic compounds Shikimate pathway Aromatic amino acids Abbreviations Cm Cucumis melo PE Peel PU Pulp S Stalk OC Ohbokggul chamoe GgC Gotgam chamoe GC Gas chromatography E4P Erythrose 4-phosphate PEP Phosphoenolpyruvate Jeongyeo Lee and Min Keun Kim have contributed equally to this work. Electronic supplementary material The online version of this article (doi:10.1007/s11033-014-3211-9) contains supplementary material, which is available to authorized users. J. Lee J. Kim S. R. Min H. Kim (&) Plant Systems Engineering Research Center, Korea Research Institute of Bioscience and Biotechnology (KRIBB), 125 Gwahangno, Yuseong-gu, Daejeon 305-806, Korea e-mail: [email protected] M. K. Kim Division of Environment-Friendly Research, Gyeongsangnam-do Agricultural Research and Extension Service, Jinju 660-360, Korea S. H. Hwang S. S. Lim (&) Department of Food Science and Nutrition, Hallym University, 1 Hallymdaehak-gil, Chuncheon 200-702, Gangwon-do, Korea e-mail: [email protected] J. M. Ahn Breeding & Research Institute, Nongwoo Bio Co., LTD, Yeoju, Kyonggi-do 469-885, Korea S. U. Park Department of Crop Science, College of Agriculture & Life Sciences, Chungnam National University, 79 Daehangno, Yuseong-gu, Daejeon 305-764, Korea 123 Mol Biol Rep (2014) 41:3487–3497 DOI 10.1007/s11033-014-3211-9

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Phenotypic profiling and gene expression analyses for aromaticand volatile compounds in Chamoes (Cucumis melo)
Jeongyeo Lee • Min Keun Kim • Seung Hwan Hwang •
Jungeun Kim • Jong Moon Ahn • Sung Ran Min •
Sang Un Park • Soon Sung Lim • HyeRan Kim
Received: 21 May 2013 / Accepted: 27 January 2014 / Published online: 11 February 2014
� Springer Science+Business Media Dordrecht 2014
Abstract Gotgam chamoe (GgC), a native oriental melon
in Korea, is known to possess the aroma of a dried per-
simmon, an agronomic relevance for melon breeding pro-
gram. The volatile compounds and the transcript levels of
aromatic compound genes in cultivar (Ohbokggul chamoe
[OC]) and GgC were profiled. A total of 62 volatile com-
pounds were identified and quantified. Twenty-eight vola-
tile compounds were specific to either the OC or the GgC.
The amounts of volatile alcohol, saturated hydrocarbon,
and unsaturated hydrocarbon compounds were 2.2, 2.7, and
1.1 times higher in OC, respectively. The amounts of
ketone volatiles were 1.2 times higher in GgC, whereas the
total amounts of esters were similar. In the shikimate
pathway, transcriptional patterns with the fruit parts were
different between the two chamoes for CmDAHPS,
CmDHD/SDH, and CmEPSPS. The expression levels of all
six genes investigated, especially CmCS, were highest in
the peel of both chamoes compared to the other parts. The
transcript levels of the aromatic amino acid biosynthesis
genes demonstrate that phenylalanine and tyrosine are
present more in edible parts of the chamoe, while trypto-
phan may be accumulated low in the chamoe. In addition,
phenylalanine and tryptophan are synthesized more in GgC
than the OC.
Keywords Cucumis melo � Chamoe � Volatile aromatic
compounds � Shikimate pathway � Aromatic amino acids
Abbreviations
Cm Cucumis melo
PE Peel
PU Pulp
S Stalk
OC Ohbokggul chamoe
GgC Gotgam chamoe
GC Gas chromatography
E4P Erythrose 4-phosphate
PEP Phosphoenolpyruvate
Jeongyeo Lee and Min Keun Kim have contributed equally to this
work.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s11033-014-3211-9) contains supplementarymaterial, which is available to authorized users.
J. Lee � J. Kim � S. R. Min � H. Kim (&)
Plant Systems Engineering Research Center, Korea Research
Institute of Bioscience and Biotechnology (KRIBB),
125 Gwahangno, Yuseong-gu, Daejeon 305-806, Korea
e-mail: [email protected]
M. K. Kim
Division of Environment-Friendly Research,
Gyeongsangnam-do Agricultural Research and Extension
Service, Jinju 660-360, Korea
S. H. Hwang � S. S. Lim (&)
Department of Food Science and Nutrition, Hallym University,
1 Hallymdaehak-gil, Chuncheon 200-702, Gangwon-do, Korea
e-mail: [email protected]
J. M. Ahn
Breeding & Research Institute, Nongwoo Bio Co., LTD,
Yeoju, Kyonggi-do 469-885, Korea
S. U. Park
Department of Crop Science, College of Agriculture & Life
Sciences, Chungnam National University, 79 Daehangno,
Yuseong-gu, Daejeon 305-764, Korea
123
Mol Biol Rep (2014) 41:3487–3497
DOI 10.1007/s11033-014-3211-9

CM Chorismatemutase
AS Anthranilate synthase
DAHPS 3-deoxy-D-arabino-heptulosonate 7-phosphate
synthase
CS Chorismate synthase
EPSP 5-enolpyruvylshikimate 3-phosphate synthase
AAT Alcohol acetyltransferase
Introduction
Melons (Cucumis melo) are one of the major horticultural
crops in the world, and include cantaloupe, honeydew, and
various mixed specialty types. It is a diploid species
(2n = 2X = 24) belonging to the Cucurbitaceae family,
which includes the cucumber, watermelon, and squash.
With its various phenotypes, the melon has been suggested
as a model for studying fruit ripening, sex determination,
and phloem physiology [1–4]. The oriental melon (C. melo
var. makuwa) called the ‘chamoe’ in Korean, has been used
traditionally for the treatment of various disorders as a liver
tonic, cardio-protective, anti-diabetic, anti-obesity, and
anti-cancer medicine [5–7]. The chamoe is a popular fruit
crop cultivated mainly in Asia and a high market value
crop in Korea. It is thought that melon and cucumber
originate in Asia. There are two major landrace of chamoe
in Korea: Sunghwan chamoe and Gotgam chamoe. The
Sunghwan chamoe has a green strain basis and a blue skin.
The Sunghwan chamoe contains more nutrients (such as
water, calcium, carbohydrates, and vitamins) than the
cultivar of chamoes. Moreover, it has greater disease
resistance compared to cultivar of chamoes [7]. With these
advantages, the whole genome of the melon was sequenced
using Sunghwan chamoe-derived double haploid lines [8].
Gotgam chamoe has green skin with distinctive green
stripes running from end to end, and a very thick, light
green flesh. The Gotgam chamoe has the aroma of a dried
persimmon, from which its name is derived. Breeders have
attempted to introduce the aroma of the Gotgam chamoe to
melons as well as chamoes because it adds more flavor and
savor. However, the constituents of this aroma as well as
the genotyping of this trait have not been studied in detail
up to now.
Aroma is an important trait of fresh and processed fruits
and vegetables. The aromas of fruits are determined by
distinctive combinations as well as the presence or absence
of volatile components [9–12]. Volatiles are naturally
produced by plants (flowers, fruits, vegetables, and herbs)
and animals. More than 300 compounds, including alco-
hols, aldehydes, esters, acids, ketones, and hydrocarbons
have been reported in fruits and vegetables [13–15]. The
volatile components of muskmelon have been identified
and include volatile aldehydes, alcohols, and an especially
large number of esters [16–18]. Volatile ester compounds
are key contributors to unique aromas, are produced during
ripening, and play an important role in determining the
final sensory quality of fruits [19, 20].
The shikimate pathway is considered as the common
route for the production of aromatic amino acids such as
phenylalanine, tyrosine, and tryptophan, which are found
only in microorganisms and plants, and never found in
animals [21, 22]. The shikimate pathway consists of seven
enzymes that catalyze the sequential conversion of ery-
throse 4-phosphate (E4P) and phosphoenolpyruvate (PEP)
to chorismate [23]. Chorismate, the end product of the
shikimate pathway, is the precursor of three aromatic
amino acids and several other aromatic compounds of
primary metabolism [24]. Chorismate is converted by
chorismatemutase (CM) and anthranilate synthase (AS) to
prephenate and anthranilate, respectively. The aromatic
amino acids are then biosynthesized from prephenate and
anthranilate. Phenylalanine is a general precursor of vari-
ous phenolic compounds [25]. Tryptophan is a precursor of
alkaloids, phytoalexins, indoleglucosinolates, and auxin.
Tyrosine is a precursor of isoquinoline alkaloids, pigment
betalains, and quinines [26, 27]. The key genes in the
shikimate and aromatic amino acid biosynthesis pathways
that have been identified in melons [28], apples [29],
grapes [30], oranges [31], pears [32], and strawberries [33]
enable us to investigate the expression levels of the genes
encoding the key enzymes for biosynthesis of aromatic
compounds.
In this study, we profiled and compared volatile com-
pounds in the different fruit parts of Gotgam (Korean
landrace) and Ohbokggul (cultivar) chamoes to elucidate
aroma of GgC, one of the breeding target traits. We also
analyzed the expression patterns of volatile biosynthesis
genes in both chamoes to investigate the correlations
between the metabolic pathway and transcription.
Materials and methods
Plant materials
Two chamoes (C. melo L. var. makuwa), Ohbokggul cha-
moe (Nongwoo Bio Co. F1 cultivar) and Gotgam chamoe
(C. melo var. makuwa, Nongwoo Bio Co. Accession No.
1638), were grown in a greenhouse at an experimental
farm, and obtained from Nongwoo Bio Co. (Korea) during
the fruiting season in October 2012. Ohbokggul and Got-
gam chamoes are differentiated by shape, size, color, and
flavor (Fig. 1). Three fruits of each chamoe were collected
and their peels, pulps, and stalks were separated. The
samples were frozen in liquid nitrogen, and stored at
-80 �C until analyzed.
3488 Mol Biol Rep (2014) 41:3487–3497
123

Extraction of volatile compounds
A 1 cm long 50/30 lm polydimethylsiloxane/divinylbenzene/
carboxen-coated fiber was used for this analysis. The SPME
fiber was conditioned in a gas chromatography (GC) injection
port at 220 �C for 3 h and at 250 �C for 5 min prior to use.
The fresh fruit of each chamoe (2.0 g) were placed in a
15 mL thermo stated vial. During the SPME extraction pro-
cedure the SPME fiber was introduced for 30 min into the
thermostated vial (25 �C) with a rubber septum containing
2.0 g of the fresh fruit of each chamoe. The absorbed com-
ponent was injected into a GC by desorption at 220 �C for
1 min in the injector (splitless mode). The SPME procedure
was repeated three times and the results were presented as the
mean ± standard deviation.
Analysis of GC and GC-mass spectrometry (MS)
GC analysis was performed using an Agilent 6890 N GC
mainframe equipped with an HP-5 (30 m 9 0.32 mm ID,
film thickness 0.25 lm) fused-silica capillary column (Ag-
ilent, USA) and a flame ionization detector. The injector and
detector temperatures for each analysis were 220 and
280 �C, respectively. The carrier gas was nitrogen at a flow
rate of 1.0 ml/min. The column temperature was maintained
at 50 �C for 5 min and programmed as follows: rise from 50
to 85 �C at a rate of 2.5 �C/min, rise from 85 to 250 �C at a
rate of 5 �C/min, and hold at 250 �C for 5 min. GC–MS
analysis was performed on a GC/MSD Polaris Q (Thermo
Finnigan, USA) with an HP-5 (30 m 9 0.32 mm ID, film
thickness 0.25 lm) fused-silica capillary column (Agilent,
USA). Helium was used as the carrier gas at a flow rate of
1.0 ml/min. For GC–MS detection, an electron ionization
system with system energy 70 eV, trap current 250 lA, and
ion source temperature 200 �C was used. The oven tem-
perature program was the same as described for GC, and
injections were used in the splitless mode.
Identification of samples
The identification of the chemical constituents was based
on comparisons of their relative retention times. The
retention indices (RIs) were calculated for all compounds
using a homologous series of n-alkanes (C5-C20) under
same GC condition and by comparison of mass spectra
with NIST and WILEY library data of the GC–MS system
and with literature data. Total ion current chromatograms
were recorded in a mass range of 40–400 amu.
Analysis of reverse transcription-polymerase chain
reaction (RT-PCR)
Genbank accession numbers AB472682, AB472683,
AB472375, FJ604859, AB472685, AB472686, L47355,
AK226909, EU431334, NM_202520, M92353, U18968,
U18770, and NM_116495 were used as queries to identify
the homologous genes of the chamoe using our internal
transcriptome database of the chamoe (unpublished data).
To confirm the transcript level of selected genes, we per-
formed RT-PCR using primers listed in Table 1. For RT-
PCR analysis, total RNA was extracted with the RNeasy
Plant Mini Kit (QIAGEN, Germany), and cDNA was
synthesized using the iScript Select cDNA Synthesis Kit
(Bio-Rad, UK) according to the manufacturers’ instruc-
tions. The level of the C. melo actin gene (CmACT2) was
used as a loading control. RT-PCR reaction conditions
included 45 �C for 30 min, followed by 94 �C for 2 min;
28 cycles at 94 �C for 30 s; 52 �C for 30 s; 72 �C for
1 min; and a final extension step of 72 �C for 5 min. The
amplified gene products were sequenced and compared by
multiple alignments using ClustalW2 program (http://
www.ebi.ac.uk/Tools/msa/clustalw2/). The sequence vari-
ations were illustrated in Supplementary Fig. 1.
Analysis of quantitative real-time PCR
Quantitative real-time RT-PCR (qRT-PCR) was performed for
precise analysis of transcript levels. Primers that targeted each
gene and actin gene produced fragments of 80–90 bp were
designed using the Primer Quest computer program (http://eu.
idtdna.com/Scitools/Applications/Primerquest/). After cDNA
synthesis, qRT-PCR was performed using a SYBR� Green
Ohbokggul Chamoe Gotgam Chamoe
PEPU
S
Fig. 1 Photographs of Ohbokggul and Gotgam chamoes. PE peel,
PU pulp, S stalk
Mol Biol Rep (2014) 41:3487–3497 3489
123
RT-PCR kit (IQ Sybr Green Super Mix) on a MiniOption
detection system (Bio-Rad, UK). Results were analyzed using
Bio-Rad software (GeneXpression Macro Chromo4) and the
comparative threshold cycle (Ct) method according to the
manufacturer’s instructions for data normalization.
Results and discussion
Profiling of volatile compounds in cultivar and landrace
chamoes
Two chamoes, Ohbokggul (cultivar; OC) and Gotgam
chamoe (Korean landrace; GgC), were harvested in Octo-
ber 2012, and their volatile compounds were profiled from
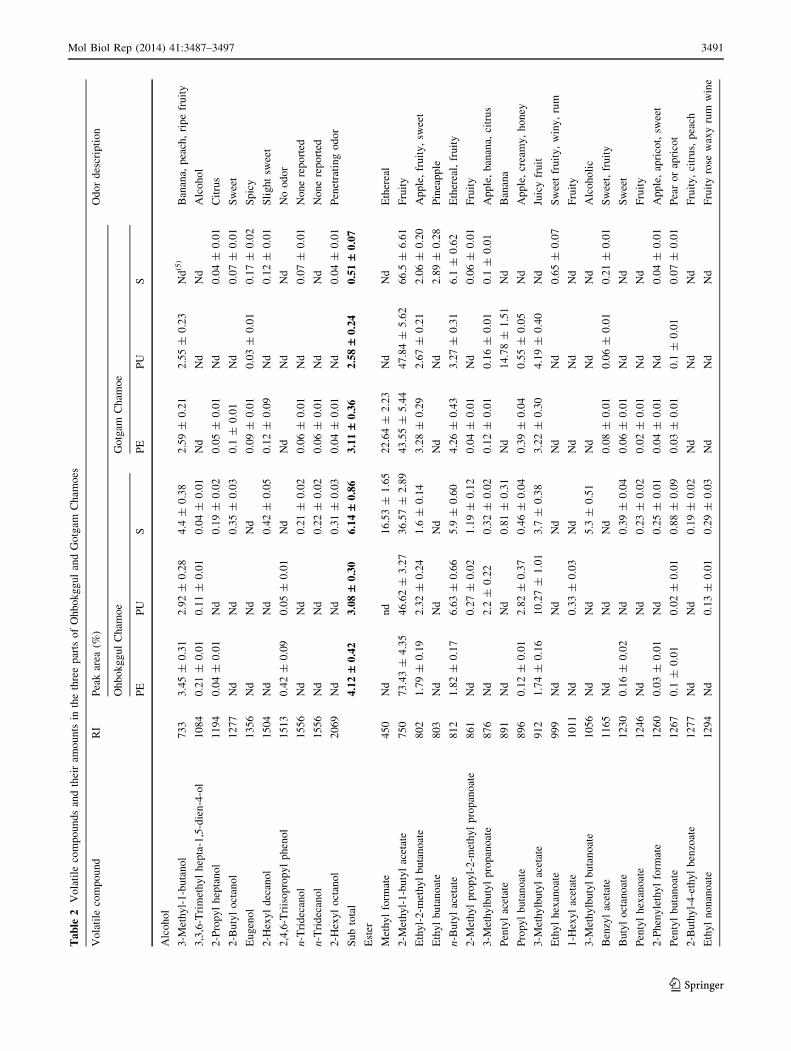
three parts: peels, pulps, and stalks (Fig. 1). A total of 62
volatile compounds were identified and quantified,
including 10 alcohols, 35 esters, 5 saturated hydrocarbons,
10 unsaturated hydrocarbons, and 2 ketones from both
chamoes. A total of 55 volatile compounds were identified
from the OC, and 43 volatile compounds from the GgC.
The most volatile compounds were detected in the stalk (46
compounds) of the OC and in the peel (32 compounds) of
the GgC. Ester compounds were the most different between
the two chamoes in terms of the number of detected vol-
atile compounds. Five saturated hydrocarbon compounds
were detected in common amongst both chamoes, but in
different amounts and patterns (Table 2).
Alcohols are known to contribute to the flavor and
aroma of fruit and serve as precursors for ester synthesis.
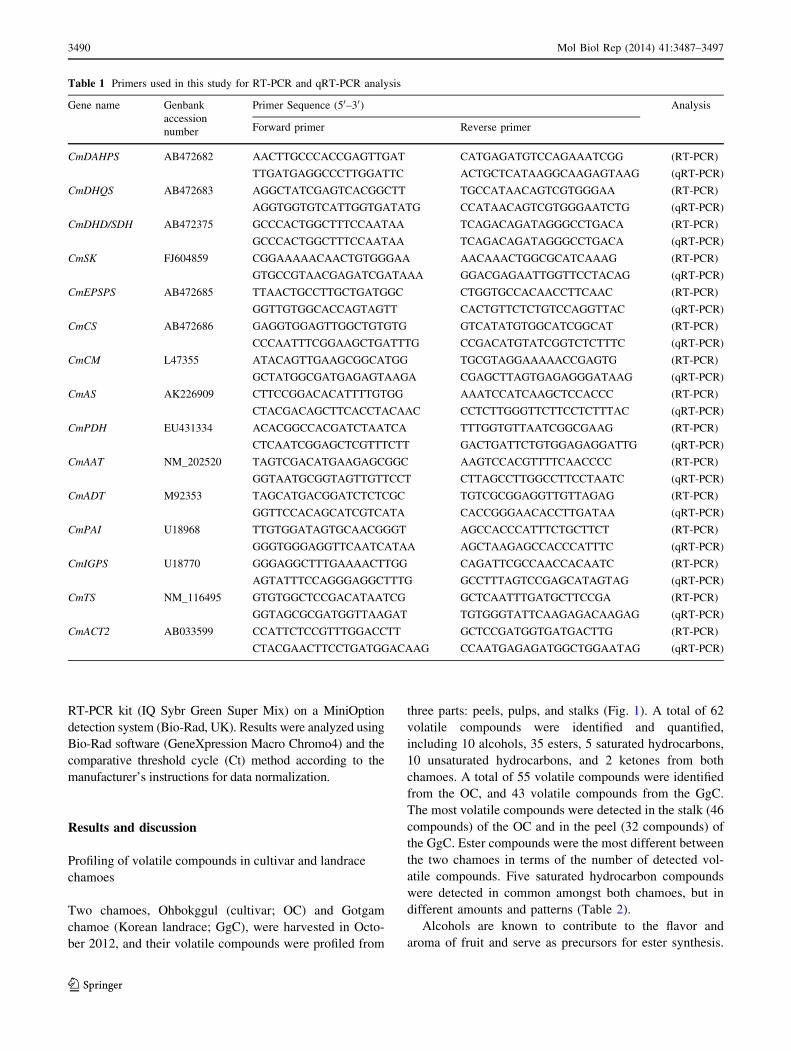
Table 1 Primers used in this study for RT-PCR and qRT-PCR analysis
Gene name Genbank
accession
number
Primer Sequence (50–30) Analysis
Forward primer Reverse primer
CmDAHPS AB472682 AACTTGCCCACCGAGTTGAT CATGAGATGTCCAGAAATCGG (RT-PCR)
TTGATGAGGCCCTTGGATTC ACTGCTCATAAGGCAAGAGTAAG (qRT-PCR)
CmDHQS AB472683 AGGCTATCGAGTCACGGCTT TGCCATAACAGTCGTGGGAA (RT-PCR)
AGGTGGTGTCATTGGTGATATG CCATAACAGTCGTGGGAATCTG (qRT-PCR)
CmDHD/SDH AB472375 GCCCACTGGCTTTCCAATAA TCAGACAGATAGGGCCTGACA (RT-PCR)
GCCCACTGGCTTTCCAATAA TCAGACAGATAGGGCCTGACA (qRT-PCR)
CmSK FJ604859 CGGAAAAACAACTGTGGGAA AACAAACTGGCGCATCAAAG (RT-PCR)
GTGCCGTAACGAGATCGATAAA GGACGAGAATTGGTTCCTACAG (qRT-PCR)
CmEPSPS AB472685 TTAACTGCCTTGCTGATGGC CTGGTGCCACAACCTTCAAC (RT-PCR)
GGTTGTGGCACCAGTAGTT CACTGTTCTCTGTCCAGGTTAC (qRT-PCR)
CmCS AB472686 GAGGTGGAGTTGGCTGTGTG GTCATATGTGGCATCGGCAT (RT-PCR)
CCCAATTTCGGAAGCTGATTTG CCGACATGTATCGGTCTCTTTC (qRT-PCR)
CmCM L47355 ATACAGTTGAAGCGGCATGG TGCGTAGGAAAAACCGAGTG (RT-PCR)
GCTATGGCGATGAGAGTAAGA CGAGCTTAGTGAGAGGGATAAG (qRT-PCR)
CmAS AK226909 CTTCCGGACACATTTTGTGG AAATCCATCAAGCTCCACCC (RT-PCR)
CTACGACAGCTTCACCTACAAC CCTCTTGGGTTCTTCCTCTTTAC (qRT-PCR)
CmPDH EU431334 ACACGGCCACGATCTAATCA TTTGGTGTTAATCGGCGAAG (RT-PCR)
CTCAATCGGAGCTCGTTTCTT GACTGATTCTGTGGAGAGGATTG (qRT-PCR)
CmAAT NM_202520 TAGTCGACATGAAGAGCGGC AAGTCCACGTTTTCAACCCC (RT-PCR)
GGTAATGCGGTAGTTGTTCCT CTTAGCCTTGGCCTTCCTAATC (qRT-PCR)
CmADT M92353 TAGCATGACGGATCTCTCGC TGTCGCGGAGGTTGTTAGAG (RT-PCR)
GGTTCCACAGCATCGTCATA CACCGGGAACACCTTGATAA (qRT-PCR)
CmPAI U18968 TTGTGGATAGTGCAACGGGT AGCCACCCATTTCTGCTTCT (RT-PCR)
GGGTGGGAGGTTCAATCATAA AGCTAAGAGCCACCCATTTC (qRT-PCR)
CmIGPS U18770 GGGAGGCTTTGAAAACTTGG CAGATTCGCCAACCACAATC (RT-PCR)
AGTATTTCCAGGGAGGCTTTG GCCTTTAGTCCGAGCATAGTAG (qRT-PCR)
CmTS NM_116495 GTGTGGCTCCGACATAATCG GCTCAATTTGATGCTTCCGA (RT-PCR)
GGTAGCGCGATGGTTAAGAT TGTGGGTATTCAAGAGACAAGAG (qRT-PCR)
CmACT2 AB033599 CCATTCTCCGTTTGGACCTT GCTCCGATGGTGATGACTTG (RT-PCR)
CTACGAACTTCCTGATGGACAAG CCAATGAGAGATGGCTGGAATAG (qRT-PCR)
3490 Mol Biol Rep (2014) 41:3487–3497
123
Ta
ble
2V
ola
tile
com
po
un
ds
and
thei
ram
ou
nts
inth
eth
ree
par
tso
fO
hb
ok
gg
ul
and
Go
tgam
Ch
amo
es
Vo
lati
leco
mp
ou
nd
RI
Pea
kar
ea(%
)O
do
rd
escr
ipti
on
Oh
bo
kg
gu
lC
ham
oe
Go
tgam
Ch
amo
e
PE
PU
SP
EP
US
Alc
oh
ol
3-M
eth
yl-
1-b
uta
no
l7
33
3.4
5±
0.3
12
.92
±0
.28
4.4
±0
.38
2.5
9±
0.2
12
.55
±0
.23
Nd
(5)
Ban
ana,
pea
ch,
rip
efr
uit
y
3,3
,6-T
rim
eth
yl
hep
ta-1
,5-d
ien
-4-o
l1
08
40
.21
±0
.01
0.1
1±
0.0
10
.04
±0
.01
Nd
Nd
Nd
Alc
oh
ol
2-P
rop
yl
hep
tan
ol
11
94
0.0
4±
0.0
1N
d0
.19
±0
.02
0.0
5±
0.0
1N
d0
.04
±0
.01
Cit
rus
2-B
uty
lo
ctan
ol
12
77
Nd
Nd
0.3
5±
0.0
30
.1±
0.0
1N
d0
.07
±0
.01
Sw
eet
Eu
gen
ol
13
56
Nd
Nd
Nd
0.0
9±
0.0
10
.03
±0
.01
0.1
7±
0.0
2S
pic
y
2-H
exy
ld
ecan
ol
15
04
Nd
Nd
0.4
2±
0.0
50
.12
±0
.09
Nd
0.1
2±
0.0
1S
lig
ht
swee
t
2,4
,6-T
riis
op
rop
yl
ph
eno
l1
51
30
.42
±0
.09
0.0
5±
0.0
1N
dN
dN
dN
dN
oo
do
r
n-T
rid
ecan
ol
15
56
Nd
Nd
0.2
1±
0.0
20
.06
±0
.01
Nd
0.0
7±
0.0
1N
on
ere
po
rted
n-T
rid
ecan
ol
15
56
Nd
Nd
0.2
2±
0.0
20
.06
±0
.01
Nd
Nd
No
ne
rep
ort
ed
2-H
exy
lo
ctan
ol
20
69
Nd
Nd
0.3
1±
0.0
30
.04
±0
.01
Nd
0.0
4±
0.0
1P
enet
rati
ng
od
or
Su
bto
tal
4.1
2–
0.4
23
.08
–0
.30
6.1
4–
0.8
63
.11
–0
.36
2.5
8–
0.2
40
.51
–0
.07
Est
er
Met
hy
lfo
rmat
e4
50
Nd
nd
16
.53
±1
.65
22
.64
±2
.23
Nd
Nd
Eth
erea
l
2-M
eth
yl-
1-b
uty
lac
etat
e7
50
73
.43
±4
.35
46
.62
±3
.27
36
.57
±2
.89
43
.55
±5
.44
47
.84
±5
.62
66
.5±
6.6
1F
ruit
y
Eth
yl-
2-m
eth
yl
bu
tan
oat
e8
02
1.7
9±
0.1
92
.32
±0
.24
1.6
±0
.14
3.2
8±
0.2
92
.67
±0
.21
2.0
6±
0.2
0A
pp
le,
fru
ity
,sw
eet
Eth
yl
bu
tan
oat
e8
03
Nd
Nd
Nd
Nd
Nd
2.8
9±
0.2
8P
inea
pp
le
n-B
uty
lac
etat
e8
12
1.8
2±
0.1
76
.63
±0
.66
5.9
±0
.60
4.2
6±
0.4
33
.27
±0
.31
6.1
±0
.62
Eth
erea
l,fr
uit
y
2-M
eth
yl
pro
py
l-2
-met
hy
lp
rop
ano
ate
86
1N
d0
.27
±0
.02
1.1
9±
0.1
20
.04
±0
.01
Nd
0.0
6±
0.0
1F
ruit
y
3-M
eth
ylb
uty
lp
rop
ano
ate
87
6N
d2
.2±
0.2
20
.32
±0
.02
0.1
2±
0.0
10
.16
±0
.01
0.1
±0
.01
Ap
ple
,b
anan
a,ci
tru
s
Pen
tyl
acet
ate
89
1N
dN
d0
.81
±0
.31
Nd
14
.78
±1
.51
Nd
Ban
ana
Pro
py
lb
uta
no
ate
89
60
.12
±0
.01
2.8
2±
0.3
70
.46
±0
.04
0.3
9±
0.0
40
.55
±0
.05
Nd
Ap
ple
,cr
eam
y,
ho
ney
3-M
eth
ylb
uty
lac
etat
e9
12
1.7
4±
0.1
61
0.2
7±
1.0
13
.7±
0.3
83
.22
±0
.30
4.1
9±
0.4
0N
dJu
icy
fru
it
Eth
yl
hex
ano
ate
99
9N
dN
dN
dN
dN
d0
.65
±0
.07
Sw
eet
fru
ity
,w
iny
,ru
m
1-H
exy
lac
etat
e1
01
1N
d0
.33
±0
.03
Nd
Nd
Nd
Nd
Fru
ity
3-M
eth
ylb
uty
lb
uta
no
ate
10
56
Nd
Nd
5.3
±0
.51
Nd
Nd
Nd
Alc
oh
oli
c
Ben
zyl
acet
ate
11
65
Nd
Nd
Nd
0.0
8±
0.0
10
.06
±0
.01
0.2
1±
0.0
1S
wee
t,fr
uit
y
Bu
tyl
oct
ano
ate
12
30
0.1
6±
0.0
2N
d0
.39
±0
.04
0.0
6±
0.0
1N
dN
dS
wee
t
Pen
tyl
hex
ano
ate
12
46
Nd
Nd
0.2
3±
0.0
20
.02
±0
.01
Nd
Nd
Fru
ity
2-P
hen
yle
thy
lfo
rmat
e1
26
00
.03
±0
.01
Nd
0.2
5±
0.0
10
.04
±0
.01
Nd
0.0
4±
0.0
1A
pp
le,
apri
cot,
swee
t
Pen
tyl
bu
tan
oat
e1
26
70
.1±
0.0
10
.02
±0
.01
0.8
8±
0.0
90
.03
±0
.01
0.1
±0
.01
0.0
7±
0.0
1P
ear
or
apri
cot
2-B
uth
yl-
4-e
thy
lb
enzo
ate
12
77
Nd
Nd
0.1
9±
0.0
2N
dN
dN
dF
ruit
y,
citr
us,
pea
ch
Eth
yl
no
nan
oat
e1
29
4N
d0
.13
±0
.01
0.2
9±
0.0
3N
dN
dN
dF
ruit
yro
sew
axy
rum
win
e
Mol Biol Rep (2014) 41:3487–3497 3491
123
Ta
ble
2co
nti
nu
ed
Vo
lati
leco
mp
ou
nd
RI
Pea
kar
ea(%
)O
do
rd
escr
ipti
on
Oh
bo
kg
gu
lC
ham
oe
Go
tgam
Ch
amo
e
PE
PU
SP
EP
US
tra
ns-
2-H
exen
yl
acet
ate
13
08
0.1
9±
0.0
20
.22
±0
.02
0.3
1±
0.0
30
.03
±0
.01
0.0
5±
0.0
10
.06
±0
.01
Wax
y,
ban
ana-
lik
e
Oct
-1-e
n-3
-yl
acet
ate
14
78
0.1
1±
0.0
10
.58
±0
.06
Nd
Nd
Nd
Nd
Gre
en,
eart
hy
3-A
cety
lox
yb
uta
n-2
-yl
acet
ate
14
95
Nd
Nd
0.5
7±
0.0
6N
dN
dN
dS
wee
tet
her
-lik
e
Die
thy
l-2
-all
yl-
2-h
yd
rox
y-
pen
tan
edio
ate
15
52
Nd
Nd
0.2
4±
0.0
3N
dN
dN
dN
on
ere
po
rted
Eth
yl
do
dec
ano
ate
15
97
0.1
9±
0.0
20
.03
±0
.01
Nd
Nd
Nd
Nd
Pea
nu
t-li
ke
Lin
alo
ol
pro
pan
oat
e1
62
40
.06
±0
.01
0.0
1±
0.0
10
.16
±0
.02
Nd
Nd
Nd
Fre
shly
fru
ity
,fl
ora
l
a-T
erp
iny
lp
rop
ano
ate
17
03
0.0
8±
0.0
10
.07
±0
.01
0.3
±0
.03
0.0
4±
0.0
1N
d0
.52
±0
.05
Pin
ewo
od
y,
flo
ral
Iso
bo
rny
lp
rop
ano
ate
17
63
Nd
Nd
0.2
6±
0.0
30
.07
±0
.01
Nd
0.0
5±
0.0
1P
un
gen
tp
ine
cam
ph
or
wo
od
yla
ven
der
Ner
yl
pro
pan
oat
e1
79
4N
dN
d0
.23
±0
.02
0.0
4±
0.0
1N
d0
.04
±0
.01
Fru
ity
,ja
mm
y
Ger
any
lp
rop
ano
ate
18
34
0.0
±0
.01
Nd
0.1
5±
0.0
20
.05
±0
.01
0.0
3±
0.0
10
.14
±0
.01
Ro
se,
lem
on
aro
ma
[(Z
)-7
-met
hy
ltet
rad
ec-8
-en
yl]
acet
ate
18
92
Nd
Nd
0.1
5±
0.3
1N
dN
dN
dC
anta
lou
pe-
lik
e,fr
uit
y
Eth
yl
hex
adec
ano
ate
19
26
Nd
3.2
8±
0.3
1N
dN
dN
dN
dS
oft
,w
axy
Des
yl
ben
zoat
e2
03
0n
d0
.02
±0
.01
0.0
9±
0.0
10
.14
±0
.01
nd
0.1
6±
0.0
2N
on
ere
po
rted
Eth
yl
oct
adec
ano
ate
21
96
0.1
±0
.01
0.4
4±
0.0
50
.07
±0
.01
0.2
±0
.02
0.0
6±
0.0
10
.06
±0
.01
Wax
y
Eth
yl
iso
allo
cho
late
23
04
0.2
5±
0.0
3N
d0
.04
±0
.01
Nd
Nd
Nd
Pen
etra
tin
g,
Sic
ken
ing
Su
bto
tal
80
.21
–5
.04
76
.26
–6
.32
77
.24
–7
.45
78
.3–
8.8
67
3.7
6–
8.1
67
9.7
1–
7.8
1
Sat
ura
ted
hy
dro
carb
on
7-M
eth
yl
hep
tad
ecan
e1
28
8N
dN
d0
.21
±0
.02
0.0
8±
0.0
1N
d0
.06
±0
.01
No
ne
rep
ort
ed
n-N
on
adec
ane
17
93
0.0
7±
0.0
1N
d0
.26
±0
.03
0.0
5±
0.0
1N
d0
.04
±0
.01
No
od
or
6-M
eth
yl
oct
adec
ane
18
63
0.0
5±
0.0
1N
d0
.18
±0
.02
0.0
5±
0.0
1N
d0
.04
±0
.01
Eth
er
n-D
oco
san
e2
20
00
.05
±0
.01
0.0
2±
0.0
10
.36
±0
.04
0.0
3±
0.0
10
.07
±0
.01
0.0
7±
0.0
1N
on
ere
po
rted
4-M
eth
yl
do
cosa
ne
22
20
0.0
2±
0.0
10
.02
±0
.01
0.2
1±
0.0
10
.05
±0
.01
Nd
Nd
Wax
yty
pe
Su
bto
tal
0.1
9–
0.0
40
.04
–0
.02
1.2
2–
0.1
20
.26
–0
.05
0.0
7–
0.0
10
.21
–0
.04
Un
satu
rate
dh
yd
roca
rbo
n
Hep
ta-1
,2,4
,6-t
etra
ene
65
5N
dN
dn
dN
dN
d0
.85
±0
.08
Un
ple
asan
t
5-M
eth
yl-
3-o
cten
e8
38
0.2
6±
0.0
20
.48
±0
.05
nd
Nd
Nd
Nd
No
od
or
2-M
eth
yl-
2-d
ecen
e1
00
0N
dN
dn
dN
dN
d0
.32
±0
.03
Un
ple
asan
t
1,3
,5-H
epta
trie
ne
10
05
Nd
Nd
Nd
Nd
5.0
1±
0.8
0N
dS
wee
t,b
anan
a,fr
uit
y
1,3
,7-O
ctat
rien
e-5
-yn
e1
00
8N
d0
.3±
0.0
30
.07
±0
.01
Nd
Nd
Nd
Un
ple
asan
t
3-P
enty
l-1
-cy
clo
hex
ene
11
50
0.2
1±
0.0
2N
dN
dN
dN
dN
dF
lora
l
1,3
,5,7
-Cy
clo
oct
atet
raen
e1
19
60
.16
±0
.02
0.1
6±
0.0
20
.16
±0
.02
0.1
4±
0.0
10
.11
±0
.01
0.1
7±
0.0
2S
wee
to
lefi
n,
aro
ma
tra
ns-
Oci
men
e1
24
03
.04
±0
.81
5.9
2±
0.3
14
.79
±0
.70
4.1
8±
0.4
14
.29
±0
.71
Nd
Fre
sho
ran
ge
juic
e
3492 Mol Biol Rep (2014) 41:3487–3497
123
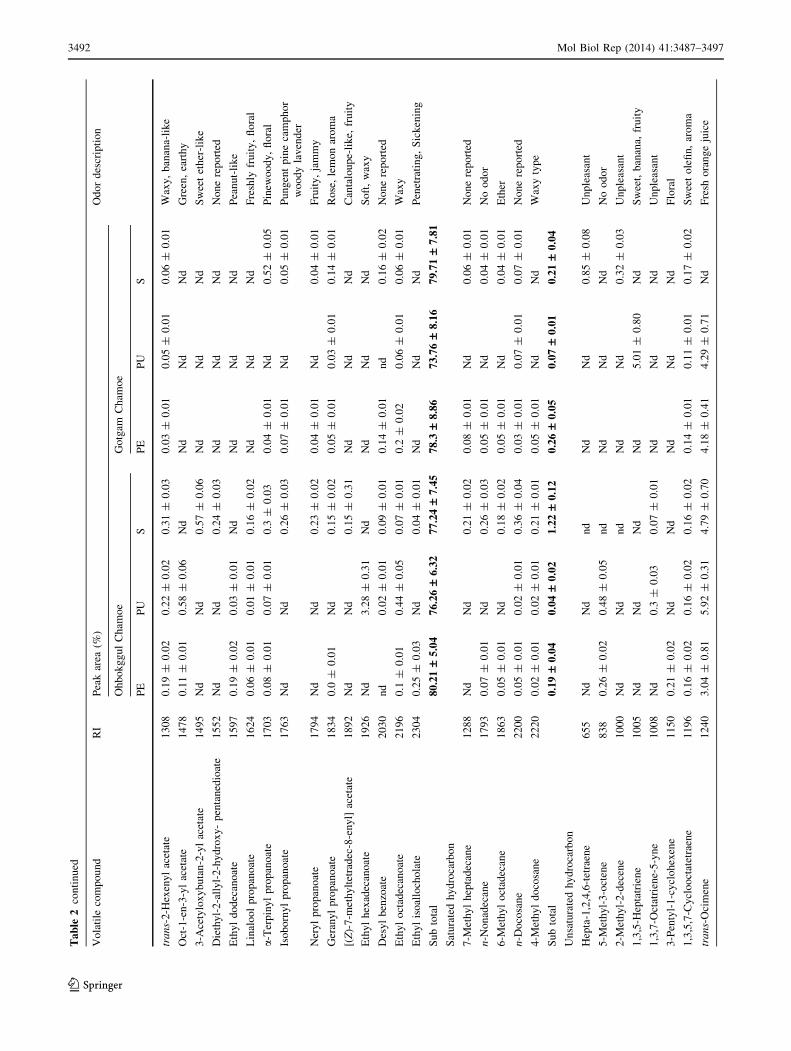
Consequently, their composition reflects the presence of
esters in the fruit [34]. Most of the alcohol compounds
were identified in the stalk part of the OC, and were 12.03
times higher there than in the stalk part of the GgC. Two
times more alcohol compounds were detected in peel part
of the GgC compared to that of the OC. The most abundant
alcohol compound detected in the peel, pulp, and stalk of
the OC was 3-methyl-1-butanol, but it was not detected in
stalk of the GgC. Among the detected alcohol compounds,
3,3,6-trimethyl hepta-1,5-dien-4-ol and 2,4,6-triisopropyl-
phenol were found only in the OC; however, eugenol was
detected only in the GgC (Table 2).
Esters are known to impart a fruity characteristic or
sweet odor in apples, strawberries, and pears [35, 36]. Our
results suggested that esters were the predominant volatile
compounds in chamoes. The number of ester compounds
was higher in the OC than the GgC, but total amounts of
esters were similar. The highest amount of esters was
detected from the peel part of the OC. The most dominant
ester in both chamoes was 2-methyl-1-butyl acetate, with a
peak range of 36.57–73.43 %. This compound was detec-
ted in different amounts depending on the part of the fruit
tested; however, it seemed to contribute to the majority of
the fruity odor of both chamoes. A total of 12 volatile
esters were identified only in the OC, and the total amount
of these specific compounds was higher in the pulp and
stalk parts which contribute fruity, citrus, and soft odors.
The GgC showed three specific ester compounds that were
detected mainly in the stalk, and these compounds provide
sweet fruity, winy, and pineapple odors (Table 2). Benzyl
acetate, which has been reported as the most important
synthetic aroma chemical in melon [37] and contributes
sweet and fruity flavors, was detected only in the GgC in
this study.
Beaulieu [29, 38] reported that methyl, ethyl, butyl, hexyl,
nonenyl, and benzyl acetates; butanoates; methylpropa-
notes; and hexanoates are the dominant esters in melon. In
apples, dominant esters are ethyl, butyl and hexyl acetates;
butanoates; and hexanoates [39]. Strawberries are comprised
primarily of methyl and ethyl acetates; butanoates; and
hexanoates [40, 41]. In our study, esters were dominated by
2-methyl-1-butyl, n-buthyl, and 3-methylbutyl acetate;
ethyl-2-methyl butanoate; and methyl formate in both cul-
tivar and landrace chamoes. Ester compounds such as
3-methylbutyl propanoate, ethyl hexadecanoate, 3-meth-
ylbutyl butanoate, and pentyl acetate were found only in
Ohbokggul chamoe, whereas ethyl butyrate and n-amyl
acetate were specific to Gotgam chamoes (Table 2). Esters
were the predominant compounds of our chamoes, but ester
compounds were different in each chamoe type, thus
reflecting their fruity flavor characters.
As shown in Table 2, all five saturated hydrocarbons
detected were identified mostly in the stalk of theTa
ble
2co
nti
nu
ed
Vo
lati
leco
mp
ou
nd
RI
Pea
kar
ea(%
)O
do
rd
escr
ipti
on
Oh
bo
kg
gu
lC
ham
oe
Go
tgam
Ch
amo
e
PE
PU
SP
EP
US
Bic
ycl
o[4
.4.0
]dec
a-1
,3,5
,7,9
-pen
ten
e1
24
20
.12
±0
.01
0.1
8±
0.0
2N
dN
dN
dN
dP
un
gen
t
1,(
E)-
8,(
Z)-
10
-Hex
adec
atri
ene
18
50
0.1
8±
0.0
2N
dN
dN
dN
dN
dU
np
leas
ant
Su
bto
tal
3.9
7–
0.9
7.0
4–
0.4
35
.02
–0
.73
4.3
2–
0.4
29
.41
–1
.52
1.3
4–
0.0
5
Ket
on
e
1-P
hen
yl
pro
pan
-2-o
ne
11
54
5.4
8±
0.8
17
.16
±0
.62
6.1
5±
0.4
27
.13
±0
.90
7.5
6±
0.7
62
.96
±0
.30
Ro
tten
egg
smel
l
4-H
yd
rox
yb
uta
no
icac
idla
cto
ne
16
18
0.5
4±
0.0
50
.8±
0.0
80
.33
±0
.02
2.0
5±
0.2
51
.45
±0
.35
3.4
9±
0.5
9C
ream
y
6.0
2–
0.8
67
.96
–0
.70
6.4
8–
0.4
49
.18
–1
.15
9.0
1–
1.1
16
.45
–0
.89
To
tal
94
.51
–7
.26
94
.38
–7
.70
96
.10
–9
.69
5.1
7–
10
.84
94
.83
–1
1.0
48
8.2
2–
8.8
6
Bo
ldv
alu
esin
dic
ate
tota
lam
ou
nt
of
vo
lati
leco
mp
ou
nd
s
RI
rete
nti
on
ind
ex,
PE
pee
l,P
Up
ulp
,S
stal
k,
Nd
no
td
etec
ted
Mol Biol Rep (2014) 41:3487–3497 3493
123
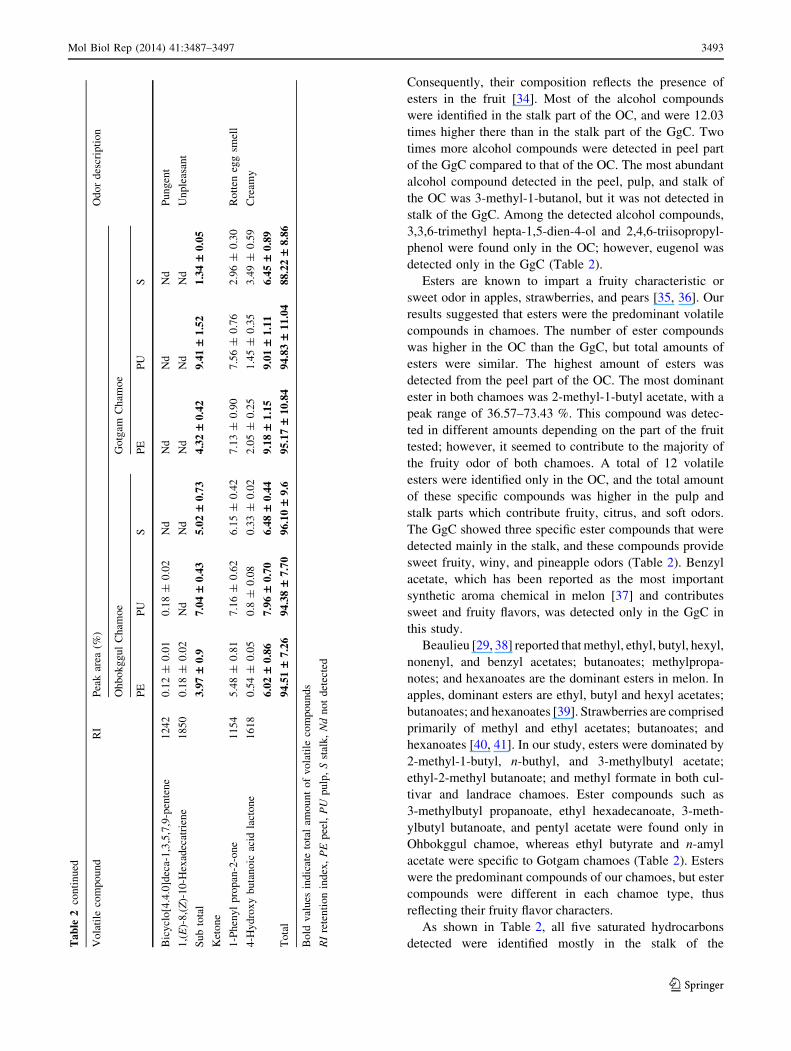
Ohbokggul chamoe (most abundant) and the peel of the
Gotgam chamoe (less abundant). The detected high
molecular weight compounds, 4-methyl docosane and
6-methyl octadecane, have been reported to donate waxy
and acetone-like odors. Five out of ten unsaturated hydro-
carbon compounds were detected only in the Ohbokggul
chamoe with a peak range of 0.07–0.48 %, and five com-
pounds were specific to the Gotgam chamoe with a peak
range of 0.32–5.01 %. 1,3,5-heptatriene, which gives sweet
and fruity odors was detected only in the pulp of the Gotgam
chamoe. Trans-ocimene, which provides a fresh orange
juice odor, was abundantly detected in both chamoes, but
not in the stalk of the Gotgam chamoe. Two ketones were
identified and the content of 4-hydroxy butanoic acid lac-
tone (which gives a creamy flavor) was higher in the Gotgam
chamoe than the Ohbokggul chamoe (Table 2). As a result,
volatile compounds were differently present within the
different parts of the two chamoes, and this contributed to
the aromatic differences between cultivar and native melon.
These data demonstrate the kinds and quantities of aroma
compounds present within different parts, and that the
aroma of chamoe was not related to any single compound,
but due to the combinations of volatiles.
Expression analysis of shikimate pathway genes
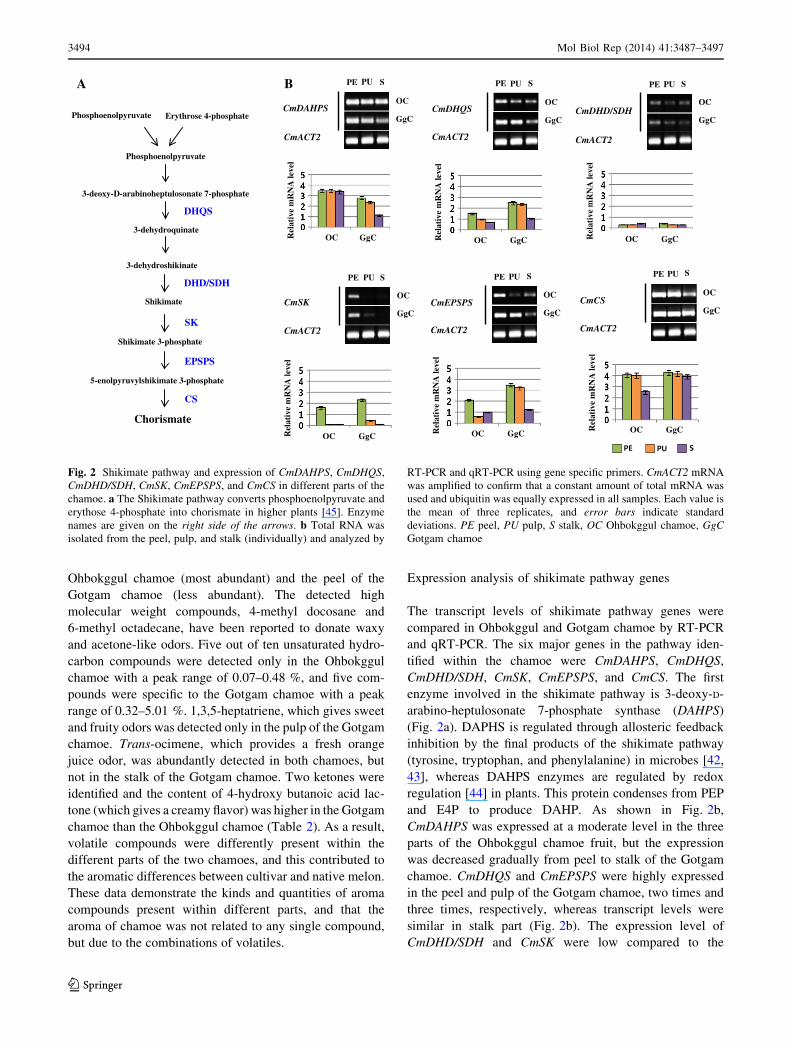
The transcript levels of shikimate pathway genes were
compared in Ohbokggul and Gotgam chamoe by RT-PCR
and qRT-PCR. The six major genes in the pathway iden-
tified within the chamoe were CmDAHPS, CmDHQS,
CmDHD/SDH, CmSK, CmEPSPS, and CmCS. The first
enzyme involved in the shikimate pathway is 3-deoxy-D-
arabino-heptulosonate 7-phosphate synthase (DAHPS)
(Fig. 2a). DAPHS is regulated through allosteric feedback
inhibition by the final products of the shikimate pathway
(tyrosine, tryptophan, and phenylalanine) in microbes [42,
43], whereas DAHPS enzymes are regulated by redox
regulation [44] in plants. This protein condenses from PEP
and E4P to produce DAHP. As shown in Fig. 2b,
CmDAHPS was expressed at a moderate level in the three
parts of the Ohbokggul chamoe fruit, but the expression
was decreased gradually from peel to stalk of the Gotgam
chamoe. CmDHQS and CmEPSPS were highly expressed
in the peel and pulp of the Gotgam chamoe, two times and
three times, respectively, whereas transcript levels were
similar in stalk part (Fig. 2b). The expression level of
CmDHD/SDH and CmSK were low compared to the
A
Phosphoenolpyruvate
3-deoxy-D-arabinoheptulosonate 7-phosphate
DHQS
3-dehydroquinate
DHD/SDH
3-dehydroshikinate
Shikimate
Shikimate 3-phosphate
SK
5-enolpyruvylshikimate 3-phosphate
Chorismate
EPSPS
CS
Phosphoenolpyruvate Erythrose 4-phosphate
BOC
CmDHD/SDH
CmACT2
GgC
PE PU S
OCCmDHQS
CmACT2
GgC
PE PU S
OCCmSK
CmACT2
GgC
PE PU S
OCCmEPSPS
CmACT2
GgC
PE PU S
OCCmCS
CmACT2
GgC
PE PU S
OCCmDAHPS
CmACT2
GgC
PE PU S
Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC
Fig. 2 Shikimate pathway and expression of CmDAHPS, CmDHQS,
CmDHD/SDH, CmSK, CmEPSPS, and CmCS in different parts of the
chamoe. a The Shikimate pathway converts phosphoenolpyruvate and
erythose 4-phosphate into chorismate in higher plants [45]. Enzyme
names are given on the right side of the arrows. b Total RNA was
isolated from the peel, pulp, and stalk (individually) and analyzed by
RT-PCR and qRT-PCR using gene specific primers. CmACT2 mRNA
was amplified to confirm that a constant amount of total mRNA was
used and ubiquitin was equally expressed in all samples. Each value is
the mean of three replicates, and error bars indicate standard
deviations. PE peel, PU pulp, S stalk, OC Ohbokggul chamoe, GgC
Gotgam chamoe
3494 Mol Biol Rep (2014) 41:3487–3497
123
A Shikimate pathway
Chorismate
Prephenate Anthranilate
ASCM
Arogenatep-hydroxyphenylpyruvate
PDH
Tyrosine
AATADT
Phenylalanine
Phosphoribosylanthranilate
IGPS
L-(o-carboxyphenylamino)-l-deoxyribulose-5-phosphate
Indole-3-glycerol phosphate
Indole
TS
TS
PAI
Tryptophan
BOC
GgC
PE PU S
CmAAT
CmACT2
CmPDH
CmACT2
OC
GgC
PE PU S
OC
CmCM
CmACT2
GgC
PE PU S
OC
GgC
PE PU S
CmAS
CmACT2
OC
GgC
PE PU S
CmPAI
CmACT2
OC
GgC
PE PU S
CmIGPS
CmACT2
OC
GgC
PE PU S
CmTS
CmACT2
OC
GgC
PE PU S
CmADT
CmACT2
Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC
Rel
ativ
e m
RN
A le
vel
OC GgC Rel
ativ
e m
RN
A le
vel
OC GgC
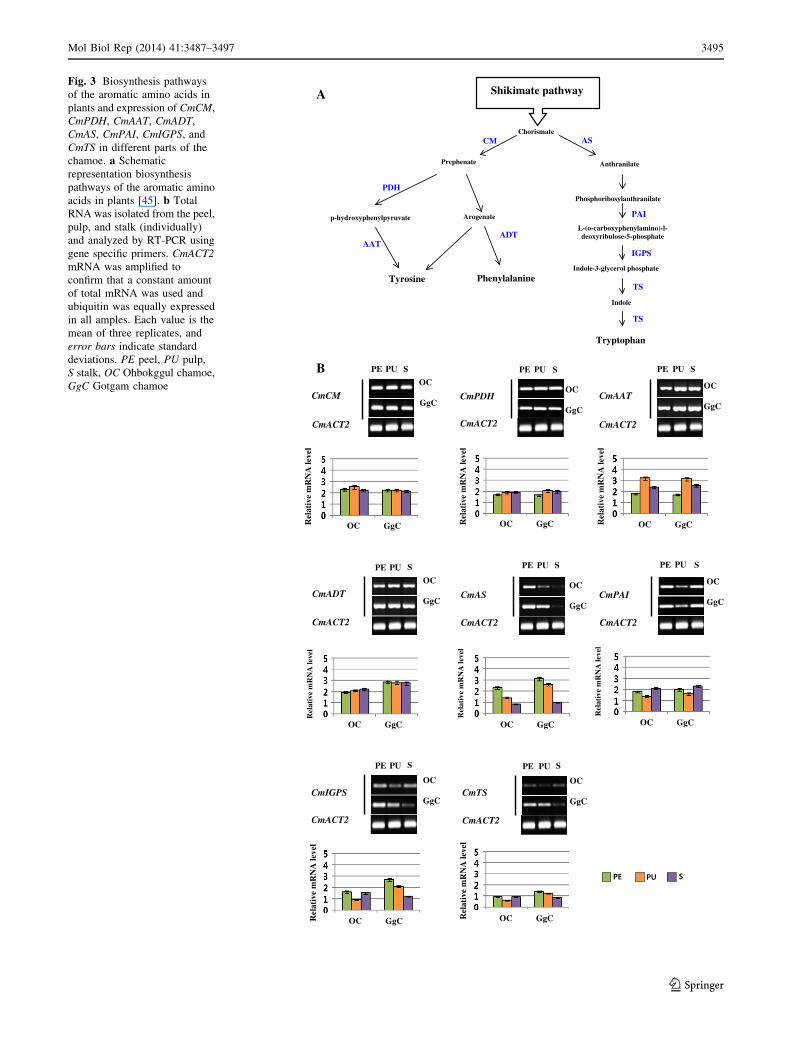
Fig. 3 Biosynthesis pathways
of the aromatic amino acids in
plants and expression of CmCM,
CmPDH, CmAAT, CmADT,
CmAS, CmPAI, CmIGPS, and
CmTS in different parts of the
chamoe. a Schematic
representation biosynthesis
pathways of the aromatic amino
acids in plants [45]. b Total
RNA was isolated from the peel,
pulp, and stalk (individually)
and analyzed by RT-PCR using
gene specific primers. CmACT2
mRNA was amplified to
confirm that a constant amount
of total mRNA was used and
ubiquitin was equally expressed
in all amples. Each value is the
mean of three replicates, and
error bars indicate standard
deviations. PE peel, PU pulp,
S stalk, OC Ohbokggul chamoe,
GgC Gotgam chamoe
Mol Biol Rep (2014) 41:3487–3497 3495
123

expression of other genes investigated. Those genes were
rarely expressed in the pulp and stalk, and shared a similar
expression pattern between Ohbokggul and Gotgam
chamoes. The last step of the shikimate pathway is cata-
lyzed by chorismate synthase (CS), which converts 5-e-
nolpyruvylshikimate 3-phosphate synthase (EPSP) to
chorismate (Fig. 2a). CmCS was the most expressed gene
among the six genes, and showed higher transcript levels in
the stalk of the Gotgam chamoe compared to that of the
Ohbokggul chamoe, while the other two parts showed
similar expression levels in both chamoes (Fig. 2b). Con-
sidering the expression patterns of CmEPSPS and CmCS,
some chorismate can accumulate in higher levels in the
pulp and stalk of the Gotgam chamoe than in those of the
Ohbokggul chamoe.
Differential expression of aromatic amino acid-related
genes
To compare the aroma-related gene expression in the both
chamoes, transcripts of eight aromatic amino acid biosyn-
thesis genes (Fig. 3a) [45] were identified from the chamoe
genome and monitored (Fig. 3). Identical expression pat-
terns were detected within the fruit parts for CmCM,
CmPDH, CmAAT, and CmADT from both chamoes. Their
expression was higher in all parts of the fruit compared to
the expression of the remaining four tryptophan biosyn-
thesis genes (Fig. 3b). The expression of CmAAT in the
pulp of the both chamoes was the highest among all of the
aromatic amino acid biosynthesis genes (Fig. 3b). Alcohol
acetyltransferase (AAT) is known to be a key enzyme in
aroma biochemistry since it catalyzes the last step in the
biosynthesis of volatile esters [46]. As previously described
by Lucchetta (2007) AAT gene is positively regulated by
ethylene during ripening of the melon [47].
Regulated enzyme CmADT was expressed higher in all
three parts of the Gotgam chamoe (Fig. 3b) than in the
Ohbokggul chamoe. This result suggests that the phenyl-
alanine accumulation level is higher in the Gotgam chamoe
than the Ohbokggul chamoe. In general, expression levels
of the tryptophan biosynthesis genes were lower in the both
chamoes than the expression levels of tyrosine and phen-
ylalanine biosynthesis genes. Those genes showed low
expression patterns in the stalk and higher expression
patterns the peel of the both chamoes. The first committed
step of tryptophan biosynthesis is generation of anthrani-
late from chorismate, catalyzed by AS (Fig. 3a). The
transcript level of CmAS was reduced from peel to stalk in
both chamoes. CmPAI expression was similar in peel and
stalk but reduced in the pulp of both chamoes. Expression
patterns of CmIGPS and CmPAI within the fruit parts were
different in the two chamoes. CmIGPS was expressed
higher in the peel and stalk versus the pulp of the
Ohbokggul chamoe while the expression was gradually
reduced from peel to stalk in the Gotgam chamoe. CmTS
exhibited a distinct Gotgam chamoe-specific expression
pattern, especially in the peel and pulp (Fig. 3b). These
collective results demonstrate that phenylalanine and
tyrosine present more in edible parts of the chamoes, and
that phenylalanine and tryptophan are synthesized more in
the Gotgam chamoe (Korean landrace) than the Ohbokggul
chamoe (cultivar). We suggest that in general the accu-
mulated amount of tryptophan may be low in chamoes.
This transcriptional difference in the aromatic amino acid
biosynthesis genes may be the cause of the variation in fruit
aromas among the chamoe varieties.
Acknowledgments This work was financially supported by Grants
from the Next-Generation Bio Green 21 Program (No. PJ008200),
Cabbage Genomics assisted breeding supporting Center (CGC)
research programs, and Hallym University Research Fund (HRF-
201302-006) funded by Rural Development Administration and
Ministry for Food, Agriculture, Forestry and Fisheries of the Korean
Government, and Hallym University, respectively.
References
1. Boualem A, Fergany M, Fernandez R, Troadec C, Martin A,
Morin H, Sari MA, Collin F, Flowers JM, Pitrat M, Purugganan
MD, Dogimont C, Bendahmane A (2008) A conserved mutation
in an ethylene biosynthesis enzyme leads to andromonoecy in
melons. Science 321(5890):836–838
2. Pech JC, Bouzayen M, Latche A (2008) Climacteric fruit ripen-
ing: ethylene-dependent and independent regulation of ripening
pathways in melon fruit. Plant Sci 175(1–2):114–120
3. Martin A, Troadec C, Boualem A, Rajab M, Fernandez R, Morin
H, Pitrat M, Dogimont C, Bendahmane A (2009) A transposon-
induced epigenetic change leads to sex determination in melon.
Nature 461(7267):1135–1138
4. Zhang B, Tolstikov V, Turnbull C, Hicks LM, Fiehn O (2010)
Divergent metabolome and proteome suggest functional inde-
pendence of dual phloem transport systems in cucurbits. Proc
Natl Acad Sci USA 107(30):13532–13537
5. Choi YJ, Chun H, Choi YH, Yum SH, Lee SY, Kim HJ, Shin YS,
Chung DS (2007) Nutritional components content of oriental
melon fruits cultivated under different greenhouse covering films.
J Bio-Env Con 16(1):72–77
6. Shin Y-S, Lee J-E, Yeon I-K, Do H-W, Cheung J-D, Kang C-K,
Choi S-Y, Youn S-J, Cho J-G, Kwoen D-J (2008) Antioxidant
effects and tyrosinase inhibition activity of oriental melon
(Cucumis melo L. var. makuwa Makino) extracts. J Life Sci
18(7):963–967
7. Kim J-H, Suh J-K, Kang Y-H (2012) Anticancer effects of the
extracts of oriental melon (Cucumis melo L. var. makuwa Mak-
ino) seeds. Korean J Plant Res 25(5):647–651
8. Garcia-Masa J, Benjak A, Sanseverino W, Bourgeois M, Mir G,
Gonzalez VM, Henaff E, Camara F, Cozzuto L, Lowy E, Alioto
T, Capella-Gutierrez S, Blanca J, Canizares J, Ziarsolo P,
Gonzalez-Ibeas D, Rodrıguez-Moreno L, Droege M, Du L,
Alvarez-Tejado M, Lorente-Galdos B, Melec M, Yang L, Wengk
Y, Navarroj A, Marques-Bonet T, Aranda MA, Nuez F, Pico B,
Gabaldon T, Roma G, Guigo R, Casacuberta JM, Arus P, Pu-
igdomenech P (2012) The genome of melon (Cucumis melo L.).
Proc Natl Acad Sci USA 109(29):11872–11877
3496 Mol Biol Rep (2014) 41:3487–3497
123

9. Thomson DMH (1987) The meaning of flavor. Elsevier, London,
pp 1–21
10. Kuentzel H, Bahri D (1990) Synthetic ingredients of food fla-
vourings. Blackie & Son, London, pp 115–157
11. Tucker GA (1993) Biochemistry of fruit ripening. Chapman and
Hall, London, pp 1–51
12. Schwab W, Davidovich-Rikanati R, Lewinsohn E (2008) Biosyn-
thesis of plant-derived flavor compounds. Plant J 54(4):712–732
13. Dirinck P, Pooter HD, Schamp N (1989) Aroma development in
ripening fruits. American Chemical Society, Washington, DC,
pp 24–34
14. Latrasse A (1991) Volatile compounds in food and beverages.
Marcel Dekker, New York, pp 329–387
15. Rapparini F, Predieris S (2003) Pear fruit volatiles. Hort Rev
28:237–324
16. Beaulieu JC, Grimm CC (2001) Identification of volatile com-
pounds in cantaloupe at various developmental stages using solid
phase microextraction. J Agric Food Chem 49(3):1345–1352
17. Khanom MM, Ueda Y (2008) Bioconversion of aliphatic and
aromatic alcohols to their corresponding esters in melons (Cuc-
umis melo L. cv. Prince melon and cv. Earl’s favorite melon).
Postharvest Biol Technol 50(1):18–24
18. Obando-Ulloa JM, Ruiz J, Monforte AJ, Fernandez-Trujillo JP
(2010) Aroma profile of a collection of near-isogenic lines of
melon. Food Chem 118(3):815–822
19. Manrıquez D, El-Sharkawy I, Flores FB, Regad F, Bouzayen M,
Latche A, Pech JC (2006) Fruit-specific gene expression and
biochemical characteristics of two highly divergent alcohol
dehydrogenases of melon. Plant Mol Biol 61(4–5):675–685
20. D’Auria JC, Pichersky E, Schaub A, Hansel A, Gershenzon J
(2007) Characterization of a BAHD acyltransferase responsible
for producing the green leaf volatile (Z)-3-hexen-1-yl acetate in
Arabidopsis thaliana. Plant J 49(2):194–207
21. Kishore GM, Shah DM (1988) Amino acid biosynthesis inhibi-
tors as herbicides. Annu Rev Biochem 57:627–663
22. Herrmann KM, Weaver LM (1999) The shikimate pathway.
Annu Rev Plant Physiol Plant Mol Biol 50:473–503
23. Campbell SA, Richards TA, Mui EJ, Samuel BV, Coggins JR,
McLeod R, Roberts CW (2004) A complete shikimate pathway in
Toxoplasma gondii: an acident eukaryotic innovation. Int J
Parasitol 34(1):5–13
24. Dewick PM (1998) The biosynthesis of shikimate metabolites.
Nat Prod Rep 15(1):17–58
25. Vogt T (2010) Phenylpropanoid biosynthesis. Mol Plant 3(1):2–20
26. Kutchan TM (1995) Alkaloid biosynthesis: the basis for meta-
bolic engineering of medicinal plants. Plant Cell 7(7):1059–1070
27. Radwanski ER, Last RL (1995) Tryptophan biosynthesis and
metabolism: biochemical and molecular genetics. Plant Cell
7(7):921–934
28. Itay G, Bar E, Portnoy V, Lev S, Burger J, Schaffer AA, Tadmor
Y, Gepstein S, Giovannoni JJ, Katzir N, Lewinsohn E (2010)
Branched-chain and aromatic amino acid catabolism into aroma
volatiles in Cucumis melo L. fruit. J Exp Bot 61(4):1111–1123
29. Beaulieu JC (2006) Volatile changes in cantaloupe during growth,
maturation, and in stored fresh-cuts prepared from fruit harvested at
various maturities. J Am Soc Hort Sci 131(1):127–139
30. Pan Q-H, Chen F, Zhu B-Q, Ma L-Y, Li L, Li J-M (2012)
Molecular cloning and expression of gene encoding aromatic
amino acid decarboxylase in ‘Vidal blanc’ grape berries. Mol
Biol Rep 39(4):4319–4325
31. Qiao Y, Xie BJ, Zhang Y, Zhang Y, Fan G, Yao XL, Pan SY
(2008) Characterization of aroma active compounds in fruit juice
and peel oil of jinchen sweet orange fruit (Citrus sinensis (L.)
Osbeck) by GC-MS and GC-O. Molecules 13(6):1333–1344
32. Li G, Huijuan J, Ruiyuan W, Sayed H, Yuanwen T (2012)
Characterization of aromatic volatile constituents in 11 Asian
pear cultivars belonging to different species. Afr J Agric Res
7(34):4761–4770
33. Cumplido-Laso G, Medina-Puche L, Moyano E, Hoffmann T,
Sinz Q, Ring L, Studart-Wittkowski C, Caballero JL, Schwab W,
Munoz-Blanco J, Blanco-Portales R (2012) The fruit ripening-
related gene FaAAT2 encodes an acyl transferase involved in
strawberry aroma biogenesis. J Exp Bot 63(11):4275–4290
34. Song J, Forney CF (2007) Flavor volatile production and regu-
lation in fruit. Can J Plant Sci 88(3):537–550
35. Echeverria G, Graell J, Lara I, Lopez ML (2008) Physico
chemical measurements in ‘Mondial Gala’ apples stored at dif-
ferent at mospheres: influence on consumer acceptability. Post-
harvest Biol Technol 50(2):135–144
36. Villatoro C, Echeverria G, Graell J, Lopez ML, Lara I (2008)
Long-Term storage of Pink Lady apples modifies volatile-
involved enzyme activities: consequences on production of vol-
atile esters. J Agric Food Chem 56(19):9166–9174
37. Bauer K, Garbe D, Surburg H (1990) Common fragrance and
flavor materials. VCH, New York, pp 87–88
38. Beaulieu JC (2006) Effect of cutting and storage on acetate and
nonacetate esters in convenient, ready-to-eat fresh-cut melons
and apples. Hort Sci 41(1):65–73
39. Dixon J, Hewett EW (2000) Factors affecting apple aroma/fla-
vour volatile concentration: a review. NZ J Crop Hortic Sci
28:155–173
40. Larson M, Poll L (1992) Odour thresholds of some important
aroma compounds in strawberries. Z. Lebensm. Unters Forsch
195(2):120–123
41. Ulrich D, Hoberg E, Rapp A, Kecke S (1997) Analysis of
strawberry flavor description of aroma types by quantification of
volatile compounds. Z Lebensm. Unters Forsch 205:218–223
42. Salcher O, Lingens F (1980) Regulation of phospho-2-keto-3-deoxy-
heptonate aldolase (DAHP Synthase) and anthranilate synthase of
Pseudomonas aureofaciens. J Gen Microbiol 121(2):473–476
43. Hartmann M, Schneider TR, Pfeil A, Heinrich G, Lipscomb WN,
Braus GH (2003) Evolution of feedback-inhibited beta/alpha
barrel isoenzymes by gene duplication and a single mutation.
Proc Natl Acad Sci USA 100(3):862–867
44. Entus R, Poling M, Herrman KM (2002) Redox regulation of
Arabidopsis 3-Deoxy-D-arabino-Heptulosonate 7-Phosphate
Synthase. Plant Physiol 129(4):1866–1871
45. Tzin V, Galili G (2010) The Biosynthetic pathways for shikimate
and aromatic amino acids in Arabidopsis thaliana. The Arabi-
dopsis Book 8:e0132. doi:10.1199/tab.0132
46. Fellman JK, Miller TW, Mattison DS, Mattheis JP (2000) Factors
that influence biosynthesis of volatile flavor compounds in apple
fruits. Hort Sci 35:1026–1033
47. Lucchetta L, Manriquez D, El-Sharkawy I, Flores FB, Sanchez-
Bel P, Zouine M, Ginies C, Bouzayen M, Rombaldi C, Pech JC,
Latche A (2007) Biochemical and catalytic properties of three
recombinant alcohol acyltransferases of melon. Sulfur-containing
ester formation, regulatory role of CaSH in activity and sequence
elements conferring substrate preference. J Agr Food Chem
55(13):5213–5220
Mol Biol Rep (2014) 41:3487–3497 3497
123
Related Documents











