doi:10.1182/blood-2001-12-0353 Prepublished online April 17, 2002; Roy Martin Guimond, Antonia Balassy, Melanie Barrette, Sylvie Brochu, Claude Perreault and Denis Claude immunoreactive T cells P-glycoprotein targeting: a unique strategy to selectively eliminate (373 articles) Plenary Papers Articles on similar topics can be found in the following Blood collections http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requests Information about reproducing this article in parts or in its entirety may be found online at: http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprints Information about ordering reprints may be found online at: http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtml Information about subscriptions and ASH membership may be found online at: digital object identifier (DOIs) and date of initial publication. the indexed by PubMed from initial publication. Citations to Advance online articles must include final publication). Advance online articles are citable and establish publication priority; they are appeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet Copyright 2011 by The American Society of Hematology; all rights reserved. 20036. the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.org From

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

doi:10.1182/blood-2001-12-0353Prepublished online April 17, 2002;
RoyMartin Guimond, Antonia Balassy, Melanie Barrette, Sylvie Brochu, Claude Perreault and Denis Claude immunoreactive T cellsP-glycoprotein targeting: a unique strategy to selectively eliminate
(373 articles)Plenary Papers �Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

P-GLYCOPROTEIN TARGETING: A UNIQUE STRATEGY
TO SELECTIVELY ELIMINATE IMMUNOREACTIVE T CELLS
Martin Guimond,1 Antonia Balassy,1,2 Mélanie Barrette,1,2
Sylvie Brochu,1 Claude Perreault,1 and Denis Claude Roy1
1Division of Hematology-Immunology, Maisonneuve-Rosemont Hospital Research Center Department of Medicine, Université de Montréal, and
2Theratechnologies Inc. Montreal, Canada
Scientific heading: ImmunobiologyRunning title: Pgp-modulation to eliminate immunoreactive T cellsWord count - Total text: 4444 words; abstract: 197 words.
DCR is the recipient of a clinician-scientist award of the F.R.S.Q.Supported by a grant from Theratechnologies-F.R.S.Q. and the Cancer Research Society of Canada.
Correspondence to: Denis Claude Roy, MDDepartment of Hematology-ImmunologyMaisonneuve-Rosemont Hospital Research Center5415 L'Assomption Blvd.Montreal, QC, H1T 2M4, Canada Tel.: (514) 252-3404Fax: (514) 252-3430E-mail: [email protected]
Copyright 2002 American Society of Hematology
Blood First Edition Paper, prepublished online April 17, 2002; DOI 10.1182/blood-2001-12-0353 For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

ABSTRACT
T lymphocytes have been found to harbor P-glycoprotein (Pgp) and to demonstrate modulation
of its ion channel transporter function according to the state of activation of T lymphocytes. We
hypothesized that cytotoxic chemicals that are extruded by Pgp could be used to specifically
eliminate immunoreactive T cell populations. In this study, we evaluated the capacity of 4,5-
dibromorhodamine methyl ester (TH9402), a photosensitizer structurally similar to rhodamine, a
dye transported by Pgp, and which becomes highly cytotoxic upon activation with visible light to
selectively deplete alloreactive T lymphocytes. Stimulation of T cells with mitogens or
allogeneic major histocompatibility complex mismatched cells resulted in the preferential
retention of the TH9402 rhodamine-derivative in activated T cells, both CD4+ and CD8+.
Photodynamic cell therapy of TH9402-exposed T cells lead to the selective elimination of
immunoreactive T cell populations. In addition, this treatment preserved resting T cells and their
capacity to respond to third party cells. Inhibition of Pgp enhanced cellular trapping of the dye in
non-activated T cells and resulted in their depletion after exposure to light. Targeting of Pgp-
deficient cells may therefore represent an appealing strategy for the prevention and treatment of
graft-versus-host disease, and other allo- or auto-immune disorders.
E-mail: [email protected]
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

INTRODUCTION
Graft-versus-host disease (GVHD) is the principal cause of mortality and the primary limitation
to the early and widespread use of allogeneic stem cell transplantation (SCT), a treatment that
often represents the only curative option for numerous patients with malignant diseases and
hereditary metabolic disorders. Depletion of T cells capable of recognizing and mounting an
immune response toward host cells from stem cell grafts abrogates GVHD.1-4 However, the
elimination of T cells also results in delayed T cell reconstitution, and thus, an increased rate of
infection, particularly with viral agents such as cytomegalovirus, herpes zoster, and Epstein-Barr
virus.5-7 In addition, the eradication of mature T cells is associated with an increased risk of graft
rejection and an increased incidence of relapse of malignant disease.1,5,8-10 Thus, T cells are
required early after allogeneic transplantation and depleting the graft of its T cell content is not
an ideal approach to prevention of post-transplant complications. Although new
immunosuppressive agents offer options to decrease the incidence and severity of GVHD, most
of the time these strategies are only partially effective and may also increase the incidence of
viral and fungal infections, and other adverse effects of profound immunosuppression. To
provide a solution to this conundrum, selective inactivation or elimination of alloreactive donor
T lymphocytes could allow early immune recovery and response toward infectious agents, and
potentially preserve graft-versus-leukemia (GVL) activity.11-13 In addition, a strategy to
selectively eliminate immunoreactive T cells could represent an important advance for the
treatment of a large number of patients with autoimmune disorders.
Recently there have been significant efforts to try to identify and eliminate T cell subsets capable
of mounting an immune response toward host cells and mediating GVHD. Strategies targeting
CD6+ or CD8+ T cells demonstrated convincing potential for the prevention of GVHD in HLA-
matched transplants using related and even unrelated donors.14-16 Interestingly, the elimination of
donor T cell subsets did not translate into a greater incidence of graft rejection.17 Moreover, the
partial loss of GVL activity that occurs with T cell depletion was able to be restored through the
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

administration of donor lymphocyte infusions at a time when patients are at lower risk for
GVHD.18 Delaying the infusion of donor T cells until the early post-transplant surge in pro-
inflammatory cytokines has abated may contribute to limiting the development of GVHD.19
However, in the context of HLA-mismatched transplantation, antigenic differences between
donor and host cells are particularly immunogenic. In addition, it is extremely difficult to
identify a priori T cell antigens that will be unique markers for cells capable of targeting both
major histocompatibility complex (MHC)-disparate antigens and the numerous distinguishing
peptide sequences expressed by host tissues.20 In contrast, when donor cells are exposed to host
cells ex vivo, cell subsets capable of recognizing host MHC antigens become activated and thus
display the peculiar antigenic and biochemical properties rendering these cells “visible”.
Importantly, a few monoclonal antibody-based strategies have been developed to specifically
eliminate such alloreactive cells.21-25
Rhodamine enters all cells and is extruded from the intracellular milieu through P-glycoprotein
(Pgp) active transport.26 Pgp, the product of the multidrug-resistance-1 (MDR1) gene, is a protein
expressed not only in normal stem cells, but also in T lymphocytes.27-30 Investigators have
proposed that T cell activation may actually lead to the inactivation of Pgp.29 Thus, activated T
cells should fail to extrude rhodamine. While rhodamine is not cytotoxic, 4,5-dibromorhodamine
methyl ester (TH9402), a rhodamine derivative, was found to harbor important photosensitizing
potential.31-33 Its phototoxicity is mediated primarily by singlet oxygen production, with
oxydative damage concentrated to mitochondria by the virtue of drug localization.31,34 The
structural similarity between rhodamine and TH9402 prompted us to evaluate the capacity of the
latter photosensitizing agent to be preferentially retained in Pgp-deficient activated T cells and
thus, lead to their selective elimination after exposure to visible light (514 nm).35
In the present study, we found that photodynamic cell therapy (PDCT) with TH9402 was highly
toxic against CD4+ and CD8+ T cells activated in response to mitogens or MHC-mismatched
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

antigens. This PDCT selectively preserved resting T lymphocytes and their ability to proliferate
and to demonstrate cytotoxicity toward third-party antigens. PDCT may therefore have clinical
utility for the selection of non-MHC reactive T cells to prevent GVHD and accelerate immune
reconstitution post-transplantation, or for the treatment of immunoreactive disorders. Moreover,
our findings identify targeting of MDR1 inhibition as a unique physiologic approach to
specifically eliminate activated T cells.
MATERIALS AND METHODS
Human cells. Blood samples were obtained with the informed consent of healthy donors under
clinical protocols approved by the Human Subjects Protection Committee of the Maisonneuve-
Rosemont Hospital. Peripheral blood (PB) samples were collected in preservative-free heparin
and mononuclear cells separated by ficoll-hypaque density gradient centrifugation (Ficoll-Paque;
Pharmacia, Piscataway, NJ). The T lymphoblastic cell line CEM and the Pgp-expressing KG1a
cells were obtained from the American Type Culture Collection (ATCC, Rockville, MD).
PHA stimulation. Peripheral blood mononuclear cells (PBMC) were cultured for 72 hours at a
concentration of 3 X 106cells/mL in flasks (Nunclon, Nunc, Denmark) with 2 µg/mL
phytohemagglutinin (PHA; Sigma, St Louis, MO) in X-Vivo 15 medium (Bio-Whittaker,
Walkersville, MD) supplemented with 15% human AB serum (HAB)(Sigma), 2mM L-
glutamine, 1 mM sodium pyruvate, 100 U/mL penicillin, and 100 µg/mL streptomycin (all from
Gibco, Grand Island, NY).
Allogeneic T cell activation. Activation of responder (A) T lymphocytes against stimulator (B)
cells was conducted in a one-way mixed lymphocyte reaction (MLR).36 A and B individuals
presented three major HLA mismatches. Briefly, responder cells from subject A were cultured
for 4 days at 37°C with the same number of irradiated PBMC from subject B (50 Gy)(137Cs;
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Gamma Cell, Atomic Energy of Canada, Ottawa, ON, Canada) in medium supplemented with 50
U/mL rhIL-2 (R&D Systems, Minneapolis, MN).
Photodynamic cell therapy with TH9402. After in vitro activation, cells were harvested, washed,
resuspended at a final concentration of 1 X 106 cells/mL, and incubated at 37°C with 10 µM
TH9402 (Theratechnologies Inc., Montreal, QC, Canada) in X-vivo 15 medium with 2.5% HAB.
After a 40-minute incubation, cells were centrifuged and dye efflux favored by resuspending
cells in TH9402-free medium for 90 minutes. At the end of the latter dye efflux period, cells
were exposed to a fluorescent light-scanning device (PDCT-Xerox Series 4, Theratechnologies
Inc.) delivering 5 joules/cm2 at a wavelength of 514 nm.
T cell proliferation assay. Proliferative activity of responder cells exposed to photodynamic
therapy and non-treated controls was assessed on day 5 of a MLR in a standard 3H-thymidine
labeling assay. The total number of cells (responder (A) and irradiated stimulator (B) cells)
present before PDCT was not adjusted after PDCT. These cells were restimulated with a fixed
number (1 x 105) of irradiated stimulator (B) or third-party (C) cells at different
responder/stimulator cell ratios (2:1, 1:1, 1:2, and 1:4) in 96-well U-bottomed microtiter plates
(Nunc). Cultures in triplicate were labeled with 1 µCi (0.037 MBq) 3H-thymidine (Perkin Elmer,
Woodbridge, Canada) per well for 18 hours, harvested onto glass fibre filter mats, and 3H-
thymidine incorporation was measured using a liquid scintillation counter (Wallac, Gaithersburg,
MD).
CTL-precursor and limiting dilution assays. Limiting dilution analyses (LDA) were used to
calculate the frequencies of responding cytotoxic T lymphocyte precursor (CTLp) cells and
clonogenic CEM and KG1a cells in treated and untreated conditions using previously described
methods.37,38 Briefly, to determine CTLp frequency: 24 replicates of graded numbers of treated
or untreated responder (A) cells (3 X 106 to 450 cells/well) were seeded in 96-well microtiter
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

plates in the presence of 1 X 105 irradiated (50 Gy) fresh PBMC stimulator cells (B or C).
Control wells consisted of stimulator cells only. After 9 days of culture in medium supplemented
with 50 U/mL of rhIL-2, each well was tested for cytolytic activity against 5 X 103 initial
stimulator (B) and third party (C) cells using a standard 51Cr- release assay. The supernatant (100
µL) was harvested from each well and counted in a gamma counter. Spontaneous release was
less than 15%. Results for individual wells were expressed as a percentage of specific lysis
calculated as follows: % specific lysis = 100 X (experimental release – spontaneous release
[medium only]) / (maximum release [1% Triton X-100] – spontaneous release). To measure
CEM and KG1a, clonogenic cell frequencies, cells were grown in a similar LDA (from 5 X 105
to 0.5 cells per well) in RPMI-1640 medium (Life Technologies, Inc., Gaithersburg, MD)
supplemented with 10% fetal bovine serum, fed every 4 days and scored for growth under an
inverted phase microscope.
Immunophenotypic analysis. Expression of cell surface T cell antigens was evaluated by direct
immunofluorescence using standard techniques.39 MAbs used in this study were anti-CD3-FITC
(UCHT1), anti-CD25-PE (B1.49.9), anti-CD3-APC (UCHT1)(Coulter Immunology, Hialeah,
FL), anti-CD4-APC (RPA-T4), anti-CD8-APC (RPA-T8) (Pharmigen, San Diego, CA).
Nonspecific binding was determined using appropriate isotypic controls. Immunofluorescence
reactivity was determined by automated multi-parameter flow cytometry analyzing at least 104
cells in each sample (FACSCalibur; Becton Dickinson, Mountain View, CA) and processed
using Cell Quest software (Becton Dickinson).
Hematopoietic progenitor cell assay. PB cells from healthy donors mobilized with G-CSF
(Amgen, Thousand Oaks, CA) underwent PDCT and were plated in methylcellulose medium
(MethoCult H4434; StemCell Technologies Inc, Vancouver, BC, Canada) on 35-mm plastic
culture dishes, according to the manufacturer's instructions. Colony-forming units-granulocyte-
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

macrophage (CFU-GM) and colony-forming units-mix (CFU-mix) were scored after 14 days of
culture at 37ºC in a fully humidified 5% CO2 atmosphere.38
Functional evaluation of Pgp-170. Pgp substrate efflux modulation by cyclosporin-A and
verapamil was determined in an accumulation assay using TH9402. Cells were stained with 10
µM TH9402 for 40 minutes, washed and resuspended in either medium alone, with 1 µg/mL
cyclosporin A (Novartis Pharma, Dorval, QC, Canada) or 5 µg/mL verapamil (Sabex,
Boucherville, QC, Canada). Cellular retention of the dye was assessed by flow cytometry
(FACSCalibur, Becton Dickinson).40,41 Calibration beads were used in all experiments to ensure
stable energy delivery (Calibrate3 and -APC, Becton-Dickinson). Positive controls for functional
Pgp-expression consisted of KG1a cells.
Statistical analysis. To determine CTLp frequency, experimental wells were scored positive if
the percent specific lysis of a well exceeded the mean + 3 SD of the wells where only stimulator
cells were present. Cytotoxicity and clonogenicity at each serial cell concentration were assessed
in an "all-or-nothing" (positive or negative) fashion, and frequency within the test population
was estimated by use of chi-square minimization.38,42
RESULTS
Photodynamic elimination of T cells. In order to assess the sensitivity of proliferating T cells to
TH9402 PDCT, the lymphoblastic T cell line CEM was used as target. Cells were incubated for
40 minutes with 10 µM TH9402, washed and incubated with TH9402-free medium for 90
minutes and then exposed to light (5 joules/cm2). These parameters were previously found to
eliminate more than 99.9% of malignant K562 cells, while preserving more than 50% of
hematopoietic progenitor cells.32 TH9402 PDCT when compared to untreated controls depleted
more than 99.97% of CEM T cells measured by LDA (Figure 1A).
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 1.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

The potential of this TH9402-based PDCT method to eliminate activated T lymphocytes was
evaluated by comparing proliferative responses of treated (PDCT) versus untreated PHA-
activated normal PBMCs toward MHC-mismatched stimulator cells. In untreated controls, PHA-
stimulated cells were able to proliferate when subsequently exposed to MHC-disparate
stimulatory cells in an MLR (Figure 1B). In contrast, TH9402 PDCT completely abrogated the
response of PHA-stimulated cells to MHC-mismatched cells. The specificity of PDCT for
activated cells was evaluated by treating resting PBMC, incubated in IL-2 containing medium
only, and then measuring proliferative response in allogeneic mismatch MLR (Figure 1C).
Interestingly, PDCT did not affect the response of these resting cells, a finding that indicates a
higher level of sensitivity to PDCT for primed versus resting T cells.
Depletion of alloreactive T lymphocyte subsets. The clinical application of PDCT in the context
of allogeneic transplantation must rely on both specific elimination of T cells that are reactive
toward host cells and preservation of T cells capable of subsequent response to infectious or
other foreign antigens. To clarify this issue, PBMCs (individual A) were first exposed for 4 days
to allogeneic stimulator (individual B) cells mismatched at 3 MHC loci (A, B and DR). After this
activation process, cells were exposed to PDCT and then presented with either the same
stimulator (B) cells for a second time, or with third-party (C) cells in a conventional 3H-
thymidine incorporation assay (Figure 2). Increasing concentrations of TH9402 and light
intensity induced gradually decreasing proliferation toward stimulator B cells. In contrast, the
capacity of residual cells to proliferate when exposed to third party C cells was preserved except
at the highest treatment intensity. Moreover, to discriminate between the effect of TH9402, light
and PDCT, A cells were treated with either TH9402 alone, light alone or TH9402 PDCT.
Proliferative responses were preserved after exposure to either TH9402 without light or to light
alone (data not shown) (p = NS). The highest TH9402 concentration (10 µΜ) and light intensity
(5 J/cm2) were selected for all subsequent experiments because these achieved maximum
elimination of specific alloreactivity and only slightly affected response to third party C cells.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 2.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 3.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Immunophenotypic analysis. In order to evaluate the specificity of PDCT for activated T
lymphocytes, the expression of the inducible α chain of the IL-2 receptor (IL-2R: CD25) was
measured on CD4+ and CD8+ T cell populations from treated samples and untreated controls
(Figure 3). At the end of 4-day MLR, cells were exposed to TH9402 PDCT or medium, and
immunophenotypic analysis performed after 3-day culture in IL-2 containing medium. At least
98% of CD8+CD25+ cells and 96% of CD4+CD25+ cells were eliminated by PDCT when
measured by flow cytometry (Table 1). In contrast, most unactivated (CD25-) T cells were
spared; their increased proportion after PDCT confirms the selectivity of PDCT for activated
lymphocytes. In addition, CD25- T cells also stained negatively for propidium iodide, a finding
that indicates preservation of T cell integrity (data not shown).
CTL-precursor frequency after photodynamic therapy. To confirm the specificity of PDCT for
anti-host T cells, cytotoxic T lymphocyte precursors (CTLp) were enumerated following
treatment with TH9402 or medium using limiting dilution analysis (LDA). The number of CTL-
p active against B and C cells was determined after TH9402 PDCT of (AXB) MLR primed cells
(Figure 4A). In untreated samples, more anti-B than anti-C CTL-p were detected, a finding
probably reflecting the primary nature of the immune response against C cells versus the
secondary immune reaction against B cells. TH9402 PDCT eliminated anti-B cell CTL-p by a
1000-fold, but anti-C cell CTLp frequency was decreased by only 30-fold, confirming the
preferential targeting of previously activated CTLp.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 4.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Progenitor recovery after photodynamic therapy. The effect of PDCT on mobilized PB cells
from normal individuals (n = 4) was utilized to assess the toxicity of the procedure for other
spontaneously proliferating cells. These cells were exposed to the same PDCT conditions as for
T cell depletion. Survival of hematopoietic progenitor cells was evaluated using a semi-solid
culture assay (Figure 4B). These conditions, which induced 100- to 1000-fold decreases in
activated T cells, did not cause a significant decrease in the growth of CFU-GM (p = 0.09) nor
CFU-mix colonies (p = 0.1).
Kinetics of dye retention. The specificity of TH9402-mediated killing for activated T cells could
be due to a differential accumulation or retention of the dye in resting versus activated T cells.
To test this hypothesis, TH9402 influx/efflux kinetics were evaluated in PHA-stimulated and
resting lymphocytes. At the end of the incubation period, retention of the dye was higher in
PHA-stimulated CD3+ cells than in resting lymphocytes (Figure 5A). In addition, activated
lymphocytes continued to sequester more TH9402 over time than resting lymphocytes, even
after reaching the plateau phase (p<.05). Moreover, after a MLR, CD25 expressing T
lymphocytes, whether CD4+ or CD8+, retained more TH9402 compared to CD25- T cells
(p<.05)(Figure 5B, C). These data indicate that both TH9402 accumulation and TH9402
retention are increased in the proliferating and activated T cells.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom
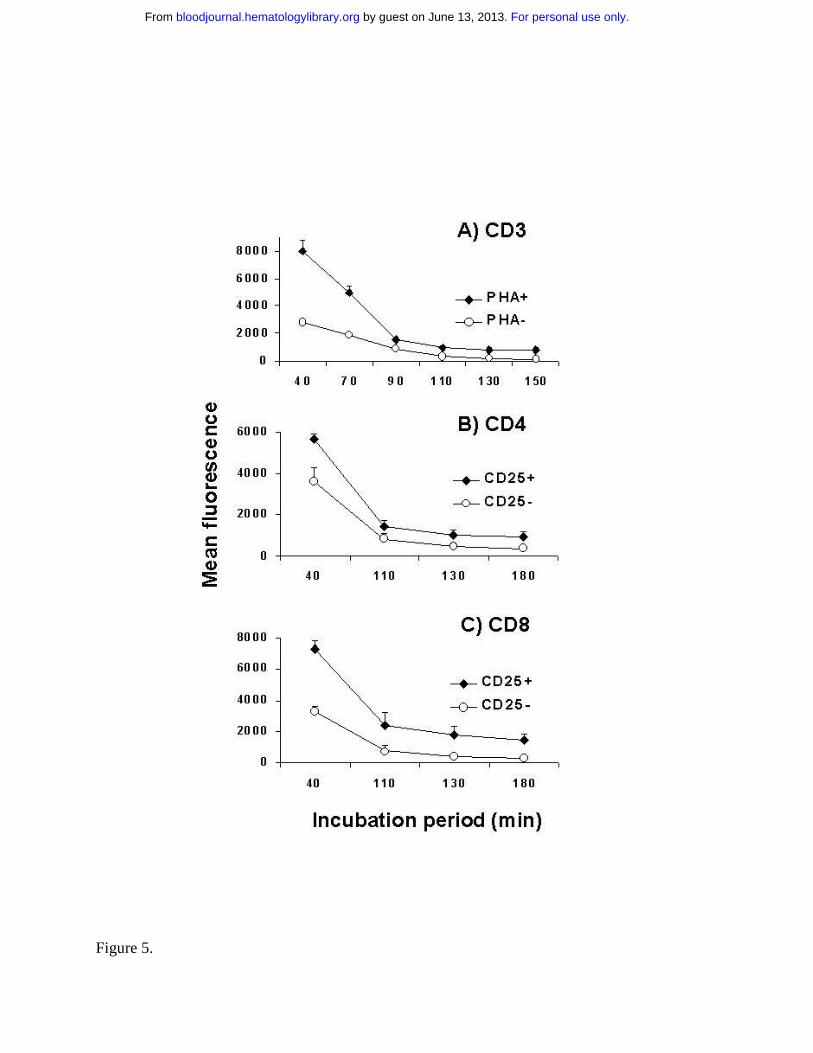
Figure 5.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Interestingly, resting T cells not only incorporated lower levels of TH9402 than PHA activated
cells, but also a large proportion of the former cells demonstrated a second peak of lower
fluorescence intensity (Figure 6A). This bimodal distribution indicates the existence among
resting cells of two populations with a different propensity to eliminate the dye. Similar results
obtained using MLR-activated T cells (data not shown) indicate that both TH9402 accumulation
and TH9402 retention are increased in the proliferating and activated T cells.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 6
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

PgP involvement in TH9402 efflux. In order to study mechanisms of retention of the TH9402
rhodamine derivative, we focussed on Pgp, which has been previously described as the main
channel involved in rhodamine efflux.43,44 In resting T lymphocytes, inactivation of Pgp by
cyclosporin-A lead to a disappearance of the peak of lower fluorescence intensity (M2: MFI =
32), and gave rise to a single peak of TH9402 fluorescence (M1) demonstrating slightly higher
retention of the dye (MFI = 600) than the M1 peak of the CSA-unexposed sample (MFI = 400)
(Figure 6A). In contrast, inactivation of the MDR1 channel had no major impact on retention of
the dye in PHA-stimulated lymphocytes. Moreover, the effect of cyclosporin-A on TH9402
efflux was durable and prevented the appearance of CD3+ T cells with low concentrations of dye
for more than 2 hours (Figure 6B).
To determine if the higher retention of the dye caused by Pgp inhibition translated into higher
cytotoxicity, we incubated resting T cells with verapamil and exposed them to PDCT. This
MDR1 inhibitor significantly enhanced the PDCT elimination of resting T cells (p<0.01) (Figure
7A). To investigate the extent of MDR1 involvement in TH9402-mediated effects, we repeated
the same experiment using the KG1a cell line, which demonstrates high levels of constitutive
expression of Pgp.45 Phototherapy with TH9402 was not cytotoxic to KG1a cells, but the
addition of verapamil led not only to increased retention of the dye (data not shown), but also to
depletion of 99.99% of clonogenic cells (Figure 7B). When used without PDCT, verapamil did
not deplete T or KG1a cells. These findings clearly identify Pgp as the principal modulator of
TH9402 cellular concentration and photodynamic cytotoxicity.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Figure 7.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

DISCUSSION
Selective elimination of donor T cell subsets recognizing host histocompatibility antigens
represents an appealing strategy to eradicate GVHD. However, in order to limit complications
with viral and fungal infections, graft rejection and relapse, which occur after physical or
functional T cell depletion, T cells capable of generating an immune response toward foreign
antigens must be preserved.5,11,46 Nevertheless, the specific elimination of such host-reactive T
cells represents a difficult task. In our present study, we uncovered a unique cytotoxic pathway
that takes advantage of the intrinsic modulation of the Pgp channel transporter to eradicate
immunoreactive T cells. Activated T lymphocytes demonstrated preferential accumulation and
retention of the TH9402 rhodamine derivative over resting T cells. Indeed, we found that resting
T lymphocytes, which express MDR1,29 extruded TH9402 through this channel transporter,
while the cellular activation process lead to an impairment in MDR1-mediated TH9402 efflux.
These kinetics of accumulation of TH9402 have resulted in the photodynamic eradication of
responder cells immunized ex vivo against stimulator cells, in conditions simulating MHC-
mismatched transplantation. In addition, this PDCT achieved drastic elimination of IL-2 receptor
expressing CD4+ and CD8+ cell populations. Notably, these findings translated into highly
efficient depletion of host-reactive cytotoxic T lymphocyte precursors. Moreover, the efficiency
of Pgp spared resting T cells and preserved their ability to generate proliferative and cytotoxic
responses against antigens other than host MHC.
Immunological tolerance may result from a variety of mechanisms, including deletion, anergy,
ignorance, and suppression.47- 51 PDCT using the TH9402 photosensitizer abrogated anti-host
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

reactivity of T lymphocytes activated either with mitogens or with an allogeneic MLR. Flow-
cytometric evaluations demonstrated that most of the effect of TH9402 treatment was
attributable to depletion of alloreactive T cells expressing CD25, the inducible high affinity IL-2
receptor. Both activated CD4+ and CD8+ cells were sensitive to the PDCT shown by the
detection of less than 1% of the total pool of lymphocytes expressing CD25 after treatment.
While the scant number of CD25+ cells detected could represent activated T cells escaping
photodynamic eradication, it is also possible that they correspond to cells bound to die from
lethal damage of PDCT-mediated oxidative damage. Alternatively, these lymphocytes could
represent non-activated T lymphocytes, such as regulatory T cells, which have been found to
constitutively express CD25.52-54 The preservation of a regulatory T cell population would be
particularly useful since it has been shown to play an important role in induction of tolerance to
alloantigen via costimulatory blockade.55 The latter scenarios would explain why the evaluation
of the impact of PDCT on CTLp demonstrated greater elimination of anti-host clonogenic
cytotoxic precursors (Figure 4A) than of CD25+ T cell populations (Figure 3). Moreover, the 3
logs of depletion of CTLs observed with the LDA is of the same order as the threshold of 2 to 3
logs of T cell depletion thought to be required for the prevention of GVHD.56
Because T cell receptor diversity post-transplant is decreased according to the number of T cells
present in the graft, it is crucial that as many T cells as possible be spared.57 Our results
demonstrate that while phototherapy using TH9402 is highly toxic for activated T cells, it
remains selective and preserves a large proportion of the CD4+ and CD8+ cells that do not
express the IL-2 high affinity receptor and other cell lineages such as myeloid and erythroid
progenitors. Interestingly, the administration of such T cells has the potential to restore T cell
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

receptor diversity through expansion in response to homeostatic signals of the host and to
reconstitute the peripheral T cell pool.58,59 Indeed, non-activated T cells remain
immunocompetent and able to proliferate in response to new antigenic stimuli, whether
previously cultured in IL-2 only, mitogens, or stimulated with allogeneic cells. This is
corroborated by the ability of TH9402 exposed cells to generate CTLp against third-party
antigens, a finding that also confirms the selectivity of PDCT. Future studies will challenge us to
determine if the small decrease in reactivity toward third party cells observed after PDCT could
reflect the elimination of T cell clones with dual specificity and demonstrating the capacity to
react toward both host and third-party cells.60,61
Since only T cells recognizing an antigen expressed by stimulator cells will be eliminated, PDCT
should spare T cells recognizing tumor antigens (developmentally regulated antigens or
leukemia-specific antigens) provided care is taken to exclude neoplastic cells from the stimulator
cell population.62,63 NK cells also express high levels of Pgp and should be protected from PDCT
toxic effects.28,64 Though we demonstrate here that PDCT is effective at eliminating T cells
reactive to stimulator cell MHC antigens, the effect of PDCT on minor histocompatibility
antigen (MiHA) stimulated T cells has yet to be addressed.36,65 In addition, the observation that
TH9402 treated T cells respond to third party MHC antigens indicates preservation of T cell
signaling and effector pathways that should translate into elimination of viral and fungal
invaders.22 Moreover, the addition of donor T cells, although non-reactive toward host MHC
antigens, could help lower the incidence of graft rejection associated with T cell depletion.3,5,8,20
Indeed, T cells present after PDCT have the potential to act as veto cells to block anti-donor
reactivity of host T cells without requiring recognition of host alloantigens.66,67 In future studies,
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

it will be important to delineate the nature of the various T and NK cell populations that escape
elimination by TH9402 PDCT and their contribution to the prevention of immunologic and
infectious post-transplant complications.
Our findings indicate that TH9402 PDCT does not exhibit a broad antiproliferative effect, but
rather acts specifically against activated T cells according to intrinsic physiologic properties of
target cells. Modulation of Pgp activity, which results in differential retention and cytotoxicity
from TH9402, could reflect biomechanical modifications of such channel transporters with the
activation process.68,69 Interestingly, Pgp could also be inactivated by PDCT itself,70 an
inhibitory mechanism that would augment retention of the photosensitizer in those activated T
cells with partial inhibition of MDR1 function, without affecting resting T cells that have already
extruded most of the dye at the time of light application. The increased retention of TH9402, and
potentially of its photoproducts, in activated cells could enhance the efficacy and specificity of
the treatment. While we cannot exclude a contribution of metabolic changes induced by T cell
activation to altered mitochondrial targeting by this rhodamine-derivative,71 our findings clearly
indicate that Pgp plays an important role in the intracellular handling of TH9402, and identify a
novel approach that takes advantage of the functional inhibition of this pathway of resistance to
selectively eradicate activated T cells.
The current photodynamic approach could be applied directly for the ex vivo treatment of stem
cell grafts or donor lymphocyte infusions in order to prevent GVHD in the context of MHC-
mismatched allogeneic transplantation.72,73 Moreover, recent identification and sequencing of
several MiHAs and improvements in immunization strategies using dendritic cells should
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

facilitate the activation of effector cells directed at host MiHAs.74 This may broaden the
applicability of this PDCT to allogeneic HLA-matched transplant strategies. Finally, the
selectivity of TH9402 PDCT for activated T lymphocytes could be exploited for the targeted
elimination of both alloreactive T cell clones that develop after solid organ transplants and
autoreactive clones responsible for diseases such as lupus erythematosus, rheumatoid arthritis
and systemic sclerosis.75-78
ACKNOWLEDGEMENTS
The authors thank Drs B. Leonard, N. Beauger and G. Krosl for insightful scientific advice; Dr
M.A. Caligiuri for critical review of the manuscript; and C. LeHouillier and the members of the
Cell Therapy Laboratory and Apheresis Unit for their excellent technical assistance. We thank all
scientists at Theratechnologies Inc. for their close collaboration and technical support.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

LEGENDS TO FIGURES
Figure 1. TH9402 eliminates proliferating T cells. A) CEM T cells were incubated with
10 µM TH 9402 for 40 minutes, washed and exposed to 5 J/cm2 of light after a dye efflux period
of 90 minutes. Survival of clonogenic cells for treated (white) and untreated (black) cells was
evaluated using a limiting dilution assay. B) PBMC exposed to PHA or C) medium only for 72
hours underwent PDCT. Proliferation to MHC-incompatible cells was evaluated in an MLR at
different responder:stimulator (R:S) ratios. Results expressed as mean ± SEM of experiments
performed in triplicate.
Figure 2. Effect of TH9402 concentration and light intensity on the depletion of host- and
third party-reactive T cells. Donor cells were first primed against host cells in a one-way, 4-
day MLR, and then treated with increasing concentrations of TH9402 and light intensity. A)
After treatment, cells were co-cultured with irradiated stimulator cells from the same host or B)
third party cells for 5 days and proliferation was measured after addition of 3H-thymidine.
(*p<.05; **p<.01) Results expressed as mean ± SEM of the percentage of proliferation of the
untreated control at a R:S ratio of 2:1; experiments performed in triplicate.
Figure 3. TH9402 PDCT eliminates activated CD4+ and CD8+ cells. MLR-activated cells
underwent TH9402 PDCT and after 72 hours, T cell populations were assessed for CD25
expression using flow cytometry. Numbers indicate the percentage of cells, and dot plots are
representative of 3 experiments.
Figure 4. PDCT eliminates anti-stimulator and preserves anti-third party CTLp, and
normal hematopoietic progenitor cells. A) Effect of TH9402 and medium treatment of MLR-
activated cells on the frequency of CTLp directed against stimulator B and third party C cells.
Results are expressed as mean ± SEM of 3 experiments. B) Mobilized peripheral blood
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

progenitors from 5 normal donors were treated with TH9402 or medium. Results are expressed
as mean ± SEM of CFU-GM and CFU-Mix colonies.
Figure 5: Kinetics of incorporation of TH9402 in resting and activated lymphocytes. A)
TH9402 dye retention was analyzed in the CD3+ cells from samples incubated (diamonds) or not
(circles) with PHA. B) Dye retention was also measured in activated (CD25+) and non-activated
(CD25-) CD4+ and C) CD8+ cells within the same MLR-activated sample. MFI ± SEM of 3 to 6
experiments and p < 0.05 for all evaluations.
Figure 6. Impact of PgP inhibition on TH9402 content in resting and activated
lymphocytes. A) PHA-stimulated and resting lymphocytes were stained with TH9402 for 40
minutes and resuspended in medium alone or with cyclosporin-A. Flow cytometric assessment of
TH9402 content in CD3+ cells was performed 90 minutes after the end of the incubation period.
Numbers in parentheses indicate the MFI of corresponding cell populations. B) The impact of
cyclosporin-A exposure on the proportion of PHA-stimulated lymphocytes capable of
eliminating TH9402 (MFI of less than 100 units) was measured over time, starting after
completion of the 40-minute incubation period. The results are representative of 3 experiments.
Figure 7. Down-modulation of MDR1 function enhances TH9402-mediated cytotoxicity. A)
Resting PBMC were exposed to TH9402 in medium supplemented or not with verapamil.
Elimination of CD4+ and CD8+ cell populations was measured 3 days after PDCT using flow
cytometry, and compared with untreated controls. Inhibition of MDR1 function increased the
photodynamic elimination of T cells. B) Cytotoxicity of PDCT on KG1a cells, an MDR1
expressing cell line, was measured using a limiting dilution assay. Verapamil alone or PDCT
with TH9402 had no effect on KG1a cells but combining the inhibition of MDR1 with verapamil
to TH9402 PDCT resulted in the elimination of more than 3 logs of clonogenic cells. Results are
expressed as mean ± SEM of at least 2 experiments.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

REFERENCES
1. Goldman JM, Gale RP, Horowitz MM, et al. Bone marrow transplantation for chronic
myelogenous leukemia in chronic phase. Increased risk for relapse associated with T-cell
depletion. Ann Intern Med. 1988;108:806-814.
2. Champlin R. T-cell depletion to prevent graft-versus-host disease after bone marrow
transplantation. Hematol Oncol Clin North Am. 1990;4:687-698.
3. Aversa F, Tabilio A, Velardi A, et al. Treatment of high-risk acute leukemia with T-cell-
depleted stem cells from related donors with one fully mismatched HLA haplotype. N Engl
J Med. 1998;339:1186-1193.
4. Storek J, Storb R. T-cell reconstitution after stem-cell transplantation-by which organ.
Lancet. 2000;355:1843-1844.
5. Appelbaum FR. Haematopoietic cell transplantation as immunotherapy. Nature.
2001;411:385-389.
6. Gratama JW, van Esser JW, Lamers CH, et al. Tetramer-based quantification of
cytomegalovirus (CMV)-specific CD8+ T lymphocytes in T-cell-depleted stem cell grafts
and after transplantation may identify patients at risk for progressive CMV infection. Blood.
2001;98:1358-1364.
7. van Esser JW, van der Holt B, Meijer E, et al. Epstein-Barr virus (EBV) reactivation is a
frequent event after allogeneic stem cell transplantation (SCT) and quantitatively predicts
EBV-lymphoproliferative disease following T-cell-depleted SCT. Blood. 2001;98:972-978.
8. Urbano-Ispizua A, Rozman C, Pimentel P, et al. The number of donor CD3(+) cells is the
most important factor for graft failure after allogeneic transplantation of CD34(+) selected
cells from peripheral blood from HLA-identical siblings. Blood. 2001;97:383-387.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

9. Pollard CM, Powles RL, Millar JL, et al. Leukaemic relapse after Campath 1 treated bone
marrow transplantation for leukaemia. Lancet. 1986;2:1404.
10. Marmont AM, Horowitz MM, Gale RP, et al. T-cell depletion of HLA-identical transplants
in leukemia. Blood. 1991;78:2120-2130.
11. Yu XZ, Bidwell SJ, Martin PJ, Anasetti C. Anti-CD3 epsilon F(ab')2 prevents graft-versus-
host disease by selectively depleting donor T cells activated by recipient alloantigens. J
Immunol. 2001;166:5835-5839.
12. Guinan EC, Boussiotis VA, Neuberg D, et al. Transplantation of anergic histoincompatible
bone marrow allografts. N Engl J Med. 1999;340:1704-1714.
13. Greenberg PD, Riddell SR. Deficient cellular immunity-finding and fixing the defects.
Science. 1999;285:546-551.
14. Champlin R, Ho W, Gajewski J, et al. Selective depletion of CD8+ T lymphocytes for
prevention of graft-versus-host disease after allogeneic bone marrow transplantation. Blood.
1990;76:418-423.
15. Soiffer RJ, Fairclough D, Robertson M, et al. CD6-depleted allogeneic bone marrow
transplantation for acute leukemia in first complete remission. Blood. 1997;89:3039-3047.
16. Soiffer RJ, Weller E, Alyea EP, et al. CD6+ donor marrow T-cell depletion as the sole form
of graft-versus-host disease prophylaxis in patients undergoing allogeneic bone marrow
transplant from unrelated donors. J Clin Oncol. 2001;19:1152-1159.
17. Roy DC, Tantravahi R, Murray C, et al. Natural history of mixed chimerism after bone
marrow transplantation with CD6-depleted allogeneic marrow: a stable equilibrium. Blood.
1990;75:296-304.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

18. Alyea E, Weller E, Schlossman R, et al. T-cell-depleted allogeneic bone marrow
transplantation followed by donor lymphocyte infusion in patients with multiple myeloma:
induction of graft-versus-myeloma effect. Blood. 2001;98:934-939.
19. Krenger W, Hill GR, Ferrara JL. Cytokine cascades in acute graft-versus-host disease.
Transplantation. 1997;64:553-558.
20. Martin PJ, Rowley SD, Anasetti C, et al. A phase I-II clinical trial to evaluate removal of
CD4 cells and partial depletion of CD8 cells from donor marrow for HLA-mismatched
unrelated recipients. Blood. 1999;94:2192-2199.
21. Koh MB, Prentice HG, Lowdell MW. Selective removal of alloreactive cells from
haematopoietic stem cell grafts: graft engineering for GVHD prophylaxis. Bone Marrow
Transplant. 1999;23:1071-1079.
22. Montagna D, Yvon E, Calcaterra V, et al. Depletion of alloreactive T cells by a specific
anti-interleukin-2 receptor p55 chain immunotoxin does not impair in vitro antileukemia and
antiviral activity. Blood. 1999;93:3550-3557.
23. Garderet L, Snell V, Przepiorka D, et al. Effective depletion of alloreactive lymphocytes
from peripheral blood mononuclear cell preparations. Transplantation. 1999;67:124-130.
24. Mavroudis DA, Jiang YZ, Hensel N, et al. Specific depletion of alloreactivity against
haplotype mismatched related individuals by a recombinant immunotoxin: a new approach
to graft-versus-host disease prophylaxis in haploidentical bone marrow transplantation.
Bone Marrow Transplant. 1996;17:793-799.
25. Fehse B, Frerk O, Goldmann M, Bulduk M, Zander AR. Efficient depletion of alloreactive
donor T lymphocytes based on expression of two activation-induced antigens (CD25 and
CD69). Br J Haematol. 2000;109:644-651.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

26. Weaver JL, Pine PS, Aszalos A, et al. Laser scanning and confocal microscopy of
daunorubicin, doxorubicin, and rhodamine 123 in multidrug-resistant cells. Exp Cell Res.
1991;196:323-329.
27. Chaudhary PM, Mechetner EB, Roninson IB. Expression and activity of the multidrug
resistance P-glycoprotein in human peripheral blood lymphocytes. Blood. 1992;80:2735-
2739.
28. Ludescher C, Pall G, Irschick EU, Gastl G. Differential activity of P-glycoprotein in normal
blood lymphocyte subsets. Br J Haematol. 1998;101:722-727.
29. Pilarski LM, Paine D, McElhaney JE, Cass CE, Belch AR. Multidrug transporter P-
glycoprotein 170 as a differentiation antigen on normal human lymphocytes and
thymocytes: modulation with differentiation stage and during aging. Am J Hematol.
1995;49:323-335.
30. Coon JS, Wang YZ, Bines SD, Markham PN, Chong AS, Gebel HM. Multidrug resistance
activity in human lymphocytes. Hum Immunol. 1991;32:134-140.
31. Villeneuve L. Ex vivo photodynamic purging in chronic myelogenous leukaemia and other
neoplasias with rhodamine derivatives. Biotechnol Appl Biochem. 1999;30 (Pt 1):1-17.
32. Roy DC, Paquette Y, Balassy A, et al. Elimination of chronic myeloid leukemia cells with a
novel photodynamic treatment. Blood. 1999;94:144a.
33. Roy DC, Boileau J, Laplante J, et al. Phase I study of autologous progenitor cell
transplantation purged with a photodynamic approach for patients with chronic myeloid
leukemia. Blood. 2000;96:583a.
34. Pal P, Zeng H, Durocher G, et al. Phototoxicity of some bromine-substituted rhodamine
dyes: synthesis, photophysical properties and application as photosensitizers. Photochem
Photobiol. 1996;63:161-168.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

35. Guimond M, Brochu S, Perreault C, Molfino N, Roy DC. Specific elimination of anti-host T
cell alloreactivity using a photodynamic approach [Abstract]. FASEB J. 2000;14:A1074.
36. Brochu S, Baron C, Hetu F, Roy DC, Perreault C. Oligoclonal expansion of CTLs directed
against a restricted number of dominant minor histocompatibility antigens in hemopoietic
chimeras. J Immunol. 1995;155:5104-5114.
37. Moretta A, Pantaleo G, Moretta L, Mingari MC, Cerottini JC. Quantitative assessment of
the pool size and subset distribution of cytolytic T lymphocytes within human resting or
alloactivated peripheral blood T cell populations. J Exp Med. 1983;158:571-585.
38. Roy DC, Ouellet S, Le Houillier C, Ariniello PD, Perreault C, Lambert JM. Elimination of
neuroblastoma and small-cell lung cancer cells with an anti-neural cell adhesion molecule
immunotoxin. J Natl Cancer Inst. 1996;88:1136-1145.
39. Guimond M, Busque L, Baron C, et al. Relapse after bone marrow transplantation: evidence
for distinct immunological mechanisms between adult and paediatric populations. Br J
Haematol. 2000;109:130-137.
40. Pallis M, Turzanski J, Harrison G, et al. Use of standardized flow cytometric determinants
of multidrug resistance to analyse response to remission induction chemotherapy in patients
with acute myeloblastic leukaemia. Br J Haematol. 1999;104:307-312.
41. Huet S, Marie JP, Gualde N, Robert J. Reference method for detection of Pgp mediated
multidrug resistance in human hematological malignancies: a method validated by the
laboratories of the French Drug Resistance Network. Cytometry. 1998;34:248-256.
42. Taswell C. A solution to the problems of cytolysis assays with additional applications to
other immunological and biochemical assays. J Immunol. 1987;138:333-341.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

43. Legrand O, Simonin G, Beauchamp-Nicoud A, Zittoun R, Marie JP. Simultaneous activity
of MRP1 and Pgp is correlated with in vitro resistance to daunorubicin and with in vivo
resistance in adult acute myeloid leukemia. Blood. 1999;94:1046-1056.
44. Leith CP, Kopecky KJ, Chen IM, et al. Frequency and clinical significance of the expression
of the multidrug resistance proteins MDR1/P-glycoprotein, MRP1, and LRP in acute
myeloid leukemia: a Southwest Oncology Group Study. Blood. 1999;94:1086-1099.
45. Lehne G, De Angelis P, den Boer M, Rugstad HE. Growth inhibition, cytokinesis failure
and apoptosis of multidrug-resistant leukemia cells after treatment with P-glycoprotein
inhibitory agents. Leukemia. 1999;13:768-778.
46. Champlin RE, Passweg JR, Zhang MJ, et al. T-cell depletion of bone marrow transplants for
leukemia from donors other than HLA-identical siblings: advantage of T-cell antibodies
with narrow specificities. Blood. 2000;95:3996-4003.
47. Mirshahidi S, Huang CT, Sadegh-Nasseri S. Anergy in peripheral memory CD4(+) T cells
induced by low avidity engagement of T cell receptor. J Exp Med. 2001;194:719-731.
48. Wells AD, Walsh MC, Bluestone JA, Turka LA. Signaling through CD28 and CTLA-4
controls two distinct forms of T cell anergy. J Clin Invest. 2001;108:895-903.
49. de St Groth BF. DCs and peripheral T cell tolerance. Semin Immunol. 2001;13:311-322.
50. Iwashiro M, Messer RJ, Peterson KE, Stromnes IM, Sugie T, Hasenkrug KJ.
Immunosuppression by CD4+ regulatory T cells induced by chronic retroviral infection.
Proc Natl Acad Sci U S A. 2001;98:9226-9230.
51. Yu X, Carpenter P, Anasetti C. Advances in transplantation tolerance. Lancet.
2001;357:1959-1963.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

52. Sakaguchi S, Sakaguchi N, Shimizu J, et al. Immunologic tolerance maintained by CD25+
CD4+ regulatory T cells: their common role in controlling autoimmunity, tumor immunity,
and transplantation tolerance. Immunol Rev. 2001;182:18-32.
53. Ng WF, Duggan PJ, Ponchel F, et al. Human CD4(+)CD25(+) cells: a naturally occurring
population of regulatory T cells. Blood. 2001;98:2736-2744.
54. Baecher-Allan C, Brown JA, Freeman GJ, Hafler DA. CD4+CD25high regulatory cells in
human peripheral blood. J Immunol. 2001;167:1245-1253.
55. Taylor PA, Noelle RJ, Blazar BR. CD4(+)CD25(+) immune regulatory cells are required for
induction of tolerance to alloantigen via costimulatory blockade. J Exp Med.
2001;193:1311-1318.
56. Lowenberg B, Wagemaker G, van Bekkum DW, et al. Graft-versus-host disease following
transplantation of 'one log' versus 'two log' T-lymphocyte-depleted bone marrow from HLA-
identical donors. Bone Marrow Transplant. 1986;1:133-140.
57. Roux E, Dumont-Girard F, Starobinski M, et al. Recovery of immune reactivity after T-cell-
depleted bone marrow transplantation depends on thymic activity. Blood. 2000;96:2299-
2303.
58. Maury S, Salomon B, Klatzmann D, Cohen JL. Division rate and phenotypic differences
discriminate alloreactive and nonalloreactive T cells transferred in lethally irradiated mice.
Blood. 2001;98:3156-3158.
59. Hochberg EP, Chillemi AC, Wu CJ, et al. Quantitation of T-cell neogenesis in vivo after
allogeneic bone marrow transplantation in adults. Blood. 2001;98:1116-1121.
60. Heemskerk MH, de Paus RA, Lurvink EG, et al. Dual HLA class I and class II restricted
recognition of alloreactive T lymphocytes mediated by a single T cell receptor complex.
Proc Natl Acad Sci U S A. 2001;98:6806-6811.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

61. Basu D, Horvath S, Matsumoto I, Fremont DH, Allen PM. Molecular basis for recognition
of an arthritic peptide and a foreign epitope on distinct MHC molecules by a single TCR. J
Immunol. 2000;164:5788-5796.
62. Mutis T, Schrama E, van Luxemburg-Heijs SA, Falkenburg JH, Melief CJ, Goulmy E. HLA
class II restricted T-cell reactivity to a developmentally regulated antigen shared by
leukemic cells and CD34+ early progenitor cells. Blood. 1997;90:1083-1090.
63. Mavroudis DA, Dermime S, Molldrem J, et al. Specific depletion of alloreactive T cells in
HLA-identical siblings: a method for separating graft-versus-host and graft-versus-
leukaemia reactions. Br J Haematol. 1998;101:565-570.
64. Ruggeri L, Capanni M, Casucci M, et al. Role of natural killer cell alloreactivity in HLA-
mismatched hematopoietic stem cell transplantation. Blood. 1999;94:333-339.
65. Perreault C, Roy DC, Fortin C. Immunodominant minor histocompatibility antigens: the
major ones. Immunol Today. 1998;19:69-74.
66. Martin PJ. Prevention of allogeneic marrow graft rejection by donor T cells that do not
recognize recipient alloantigens: potential role of a veto mechanism. Blood. 1996;88:962-
969.
67. Fowler DH, Gress RE. Th2 and Tc2 cells in the regulation of GVHD, GVL, and graft
rejection: considerations for the allogeneic transplantation therapy of leukemia and
lymphoma. Leuk Lymphoma. 2000;38:221-234.
68. Anel A, Naval J, Gonzalez B, et al. Fatty acid metabolism in human lymphocytes. I. Time-
course changes in fatty acid composition and membrane fluidity during blastic
transformation of peripheral blood lymphocytes. Biochim Biophys Acta. 1990;1044:323-
331.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

69. Utsunomiya N, Tsuboi M, Nakanishi M. Early transmembrane events in alloimmune
cytotoxic T-lymphocyte activation as revealed by stopped-flow fluorometry. Proc Natl Acad
Sci U S A. 1986;83:1877-1880.
70. Kessel D, Woodburn K. Selective photodynamic inactivation of a multidrug transporter by a
cationic photosensitising agent. Br J Cancer. 1995;71:306-310.
71. Kim M, Cooper DD, Hayes SF, Spangrude GJ. Rhodamine-123 staining in hematopoietic
stem cells of young mice indicates mitochondrial activation rather than dye efflux. Blood.
1998;91:4106-4117.
72. Slavin S, Naparstek E, Nagler A, et al. Allogeneic cell therapy with donor peripheral blood
cells and recombinant human interleukin-2 to treat leukemia relapse after allogeneic bone
marrow transplantation. Blood. 1996;87:2195-2204.
73. van Rhee F, Kolb HJ. Donor leukocyte transfusions for leukemic relapse. Curr Opin
Hematol. 1995;2:423-430.
74. Vogt MH, Goulmy E, Kloosterboer FM, et al. UTY gene codes for an HLA-B60-restricted
human male-specific minor histocompatibility antigen involved in stem cell graft rejection:
characterization of the critical polymorphic amino acid residues for T-cell recognition.
Blood. 2000;96:3126-3132.
75. Verburg RJ, Kruize AA, van den Hoogen FH, et al. High-dose chemotherapy and
autologous hematopoietic stem cell transplantation in patients with rheumatoid arthritis:
results of an open study to assess feasibility, safety, and efficacy. Arthritis Rheum.
2001;44:754-760.
76. French LE, Lessin SR, Addya K, et al. Identification of clonal T cells in the blood of
patients with systemic sclerosis: positive correlation with response to photopheresis. Arch
Dermatol. 2001;137:1309-1313.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

77. Greinix HT, Volc-Platzer B, Kalhs P, et al. Extracorporeal photochemotherapy in the
treatment of severe steroid-refractory acute graft-versus-host disease: a pilot study. Blood.
2000;96:2426-2431.
78. Barr ML, Meiser BM, Eisen HJ, et al. Photopheresis for the prevention of rejection in
cardiac transplantation. Photopheresis Transplantation Study Group. N Engl J Med.
1998;339:1744-1751.
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom

Table 1. Impact of TH9402 PDCT on activated and non-activated CD4+ and CD8+ cells
evaluated using flow-cytometry.
CD4+ cells
(x106 cells)
CD8+ cells
(x106 cells)
Treatment† CD25+ CD25- CD25+ CD25-
Medium 1.68±0.61 0.96±0.69 1.11±0.40 0.50±0.34
TH9402 0.060±0.011 0.21±0.09 0.026±0.012 0.26±0.09†After exposure to TH9402 PDCT or medium, AXB* cells were cultured for 3 days in IL-2 supplemented medium. Absolute cell numbers are expressed in million cells (mean ± SEM of 3 experiments).
For personal use only. by guest on June 13, 2013. bloodjournal.hematologylibrary.orgFrom
Related Documents











