Review Oxidative stress in Perna perna and other bivalves as indicators of environmental stress in the Brazilian marine environment: Antioxidants, lipid peroxidation and DNA damage ☆ Eduardo Alves de Almeida a , Afonso Celso Dias Bainy b , Ana Paula de Melo Loureiro c , Glaucia Regina Martinez d , Sayuri Miyamoto a , Janice Onuki a , Lívea Fujita Barbosa a , Camila Carrião Machado Garcia a , Fernanda Manso Prado a , Graziella Eliza Ronsein a , Carlos Alexandre Sigolo a , Cláudia Barbosa Brochini a , Ana Maria Gracioso Martins a , Marisa Helena Gennari de Medeiros a , Paolo Di Mascio a, ⁎ a Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo, CP 26.077, 05513-970, São Paulo, Brazil b Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, Brazil c Departamento de Análises Clínicas e Toxicológicas, Faculdade de Ciências Farmacêuticas, Universidade de São Paulo, 05508-900, São Paulo, Brazil d Departamento de Bioquímica e Biologia Molecular, Setor de Ciências Biológicas, Universidade Federal do Paraná, CP 19046, 81531-990, Curitiba-PR, Brazil Received 25 October 2005; received in revised form 25 February 2006; accepted 25 February 2006 Available online 19 April 2006 Abstract Oxidative stress can take place in marine bivalves under a series of environmental adverse conditions. The study of different systems related to oxidative stress in these organisms can give important information about their physiological status and also about environmental health. Bivalves have been proposed as good sentinel organisms in pollution monitoring studies through the analysis of biochemical biomarkers, and most of the biomarkers analyzed are those related to oxidative stress. However, it is very important to know how other environmental factors not associated to the presence of pollutants might affect these parameters. We have studied a series of mechanisms related to oxidative stress in mussels which inhabit the Brazilian coast, especially in Perna perna species, subjected to different stress conditions, such as the exposure to different contaminants in the laboratory and in the field, the exposure of mussels to air and re-submersion, simulating the tidal oscillations, and in mussels collected at different seasons. Both oxidative damage levels and antioxidant defense systems were strongly affected by the different environmental stress. This review summarizes the data obtained in some studies carried out in bivalves from the Brazilian coast. © 2006 Elsevier Inc. All rights reserved. Keywords: Oxidative stress; Mussel; Antioxidant; DNA damage; Lipid peroxidation; Environmental stress; Bivalve; Pollution Contents 1. Introduction .............................................................. 589 2. Sources of ROS/RNS in mussels ................................................... 589 3. Antioxidant defense systems ..................................................... 590 4. Lipid peroxidation ........................................................... 592 Comparative Biochemistry and Physiology, Part A 146 (2007) 588 – 600 www.elsevier.com/locate/cbpa ☆ This paper is part of a special issue of CBP dedicated to “The Face of Latin American Comparative Biochemistry and Physiology” organized by Marcelo Hermes- Lima (Brazil) and co-edited by Carlos Navas (Brazil), Rene Beleboni (Brazil), Tania Zenteno-Savín (Mexico) and the editors of CBP. This issue is in honour of Cicero Lima and the late Peter W. Hochachka, teacher, friend and devoted supporter of Latin American science. ⁎ Corresponding author. Tel.: +55 11 3091 3815x224, 223; fax: +55 11 3815 5579. E-mail address: [email protected] (P. Di Mascio). 1095-6433/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.cbpa.2006.02.040

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600www.elsevier.com/locate/cbpa
Review
Oxidative stress in Perna perna and other bivalves as indicators ofenvironmental stress in the Brazilian marine environment: Antioxidants,
lipid peroxidation and DNA damage☆
Eduardo Alves de Almeida a, Afonso Celso Dias Bainy b, Ana Paula de Melo Loureiro c,Glaucia Regina Martinez d, Sayuri Miyamoto a, Janice Onuki a, Lívea Fujita Barbosa a,Camila Carrião Machado Garcia a, Fernanda Manso Prado a, Graziella Eliza Ronsein a,Carlos Alexandre Sigolo a, Cláudia Barbosa Brochini a, Ana Maria Gracioso Martins a,
Marisa Helena Gennari de Medeiros a, Paolo Di Mascio a,⁎
a Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo, CP 26.077, 05513-970, São Paulo, Brazilb Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, 88040-900, Florianópolis, Brazil
c Departamento de Análises Clínicas e Toxicológicas, Faculdade de Ciências Farmacêuticas, Universidade de São Paulo, 05508-900, São Paulo, Brazild Departamento de Bioquímica e Biologia Molecular, Setor de Ciências Biológicas, Universidade Federal do Paraná, CP 19046, 81531-990, Curitiba-PR, Brazil
Received 25 October 2005; received in revised form 25 February 2006; accepted 25 February 2006Available online 19 April 2006
Abstract
Oxidative stress can take place in marine bivalves under a series of environmental adverse conditions. The study of different systems related tooxidative stress in these organisms can give important information about their physiological status and also about environmental health. Bivalveshave been proposed as good sentinel organisms in pollution monitoring studies through the analysis of biochemical biomarkers, and most of thebiomarkers analyzed are those related to oxidative stress. However, it is very important to know how other environmental factors not associated tothe presence of pollutants might affect these parameters. We have studied a series of mechanisms related to oxidative stress in mussels whichinhabit the Brazilian coast, especially in Perna perna species, subjected to different stress conditions, such as the exposure to differentcontaminants in the laboratory and in the field, the exposure of mussels to air and re-submersion, simulating the tidal oscillations, and in musselscollected at different seasons. Both oxidative damage levels and antioxidant defense systems were strongly affected by the different environmentalstress. This review summarizes the data obtained in some studies carried out in bivalves from the Brazilian coast.© 2006 Elsevier Inc. All rights reserved.
Keywords: Oxidative stress; Mussel; Antioxidant; DNA damage; Lipid peroxidation; Environmental stress; Bivalve; Pollution
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5892. Sources of ROS/RNS in mussels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5893. Antioxidant defense systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5904. Lipid peroxidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 592
☆ This paper is part of a special issue of CBP dedicated to “The Face of Latin American Comparative Biochemistry and Physiology” organized by Marcelo Hermes-Lima (Brazil) and co-edited by Carlos Navas (Brazil), Rene Beleboni (Brazil), Tania Zenteno-Savín (Mexico) and the editors of CBP. This issue is in honour of CiceroLima and the late Peter W. Hochachka, teacher, friend and devoted supporter of Latin American science.⁎ Corresponding author. Tel.: +55 11 3091 3815x224, 223; fax: +55 11 3815 5579.E-mail address: [email protected] (P. Di Mascio).
1095-6433/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.cbpa.2006.02.040

589E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
5. DNA damage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5935.1. Oxidative DNA damage . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 593
5.1.1. Strand breaks . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5935.1.2. 8-Oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodGuo) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5935.1.3. 8-OxodGuo formation in DNA by 1O2: mechanistic studies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5945.1.4. 8-OxodGuo levels in mussels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 594
5.2. Etheno DNA adducts . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5976. Conclusions. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 597Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 597References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 597
1. Introduction
Marine bivalves are constantly subjected to different types ofenvironmental stress, due to their sessile condition and filterfeeding habits (Goldberg, 1975). Examples of these includevariations in water temperature due to seasonal changes, oxygenand salinity fluctuations related to tidal oscillations, predation,desiccation during air exposure at low tides, changes in foodavailability, and the presence of contaminants in the environ-ment (Kennedy, 1976; Widdows et al., 1979; Guderley et al.,1994). To deal with this, mussels have developed a series ofadaptations that allows them to survive under these adverseconditions, including changes in respiratory and overallmetabolic rate, activation of alternative pathways for energyproduction, and the induction of a series of biochemical defenseand repair mechanisms.
Many studies have focused on the evaluation of differentbiochemical aspects of stress responses in marine bivalves. Asthese organisms represent important economic resources formany coastal populations in several countries, such studiescould furnish important information about the physiologicalstatus of the animals, helping to diagnose the environmentalquality of areas for bivalve farming. Most studies regarding theeffects of environmental stress in mussels are centered on thoseresponses that can potentially lead these organisms to anoxidative stress condition, since there is a close relationshipbetween environmental stress and the rate of cellular reactiveoxygen and nitrogen species (ROS/RNS) generation in theorganism (Storey, 1996). ROS/RNS can be produced as by-products of cellular metabolism, and if they are not immediatelyintercepted by antioxidant defenses they could oxidize differentcell components (Halliwell, 1993; Lemaire and Livingstone,1993). When the rate of ROS/RNS production exceeds the rateof its decomposition by antioxidant defenses and repairsystems, oxidative stress can be established, leading to theoxidation of key cell components like proteins, DNA and fattyacids (Sies, 1993).
Exposure of aquatic organisms to pollutants can promote anincrease in the rate of ROS/RNS production, thus theassessment of oxidative stress-related parameters in specificsentinel organisms could be included in environmentalpollution monitoring studies to predict the impact of pollutantspresent in the environment (Pellerin-Massicote, 1994; Walker etal., 1996; Livingstone, 2001). However, other environmental
parameters not related to pollution, such as temperature,salinity, and others can cause important changes in somebiochemical systems that have been proposed as biomarker.Therefore, basic information about the influence of these factorson the biomarker to be used in different species is required to getaccurate measurements.
Bivalve mollusc farming along the Brazilian coast, espe-cially in Santa Catarina State, has increased significantly in thelast ten years, with a production surpassing 8000tons in 2001(personal communication). The main species cultivated are thebrown mussel Perna perna and Crassostrea gigas, and anincreasing interest has developed in the cultivation of otherbivalves, such as the oysters Crassostrea rhizophorae and theclam Nodipecten nodosus.
In Brazil few studies have dealt with the evaluation of themarine environmental quality, either in farming zones or in wildareas, and these are mostly related to the measurement of fecalcoliforms in water (balneability studies), or the analysis of somespecific chemicals in water. In collaboration with bivalvecultivators from southern Brazil, we started a series of studieson biochemical mechanisms related to oxidative stress inbivalve species from the Brazilian coast, especially in P. pernamussels, one of the most abundant species in Brazil. Bysubmitting mussels to different types of environmental stressingagents we obtained data sets on the responses of this mollusc toadverse conditions, providing information to help diagnose thephysiological status of these organisms in the field, as well as topredict the quality of the marine environment. The data obtainedin these studies are promising to be used on more extensiveprograms of marine environmental monitoring along theBrazilian coast.
2. Sources of ROS/RNS in mussels
ROS/RNS are produced in organisms under a series of phy-siological conditions, being related both to the basal metabolismand to the influence of environmental factors. ROS/RNS in-cludes the hydroxyl radical (•OH), the superoxide anion radical(O2
•−), hydrogen peroxide (H2O2), singlet oxygen (1O2), nitricoxide (NO), peroxinitrite (ONOO−), alkoxyl (RO•), andperoxyl (ROO•) radicals, among others. They are particularlytransient species due to their high chemical reactivity and canreact with DNA, proteins, carbohydrates and lipids in adestructive manner (Storey, 1996). Under physiological

590 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
conditions, the mitochondrion is the major site of O2•− and H2O2
generation in organisms, due to univalent reduction of O2. Thepropensity for electron transport chain to generate such reactivespecies through uncoupled electron transfer to O2 depends onfactors that include the availability of molecular oxygen, thereduction state of the electron carriers, and the mitochondriamembrane potential (Turrens and Boveris, 1980; Turrens et al.,1985; Dawson et al., 1993; Korshunov et al., 1997). Indeed,cellular generation of •OH, the most reactive ROS could berelated to the reduction of H2O2 by O2
•− (Haber–Weiss reaction)(Kehrer, 2000), but this has been demonstrated to be nearly zeroin biological systems. On the other hand, in the presence oftransition metals, like iron and copper, O2
•− and H2O2 cangenerate •OH through Fenton reaction (Halliwell, 1992). Othernon-transition metal ions can also be implicated in ROS/RNSgeneration in mitochondria. Cadmium, for example, is known togenerate ROS due to an inhibitory effect on mitochondriaelectron transport (Stohs et al., 2000). As a result, the respiratorychain becomes highly reduced and the electrons are transferreddirectly to available oxygen, leading to an enhancement in theROS formation (Ochi et al., 1987; Tatrai et al., 2001).
Cellular oxygen availability is also related to the generationof ROS/RNS and oxidative stress. It seems clear that duringhyperoxia the increase in molecular oxygen concentration isdirectly proportional to the production of ROS/RNS, and thatduring hypoxia, at low oxygen availability, the rate of ROS/RNS should be decreased (Michiels et al., 2002). However,recently was proposed that ROS/RNS generation could increasein mitochondria under hypoxic conditions, and that thesereactive species would serve as signaling molecules whichwould activate factors involved in hypoxic responses. Thedecrease in cytochrome oxidase Vmax during hypoxia isresponsible for an increase in mitochondria redox state(Chandel et al., 2000), which in turns, accelerates ROS/RNSgeneration, triggering the activation of different transcriptionalfactors involved in numerous cellular hypoxia responses.Although these mechanisms are mostly studied in mammalsdue to pathological dysfunctions, these studies become relevantin mussels, since these organisms face daily oscillations inoxygen availability associated with the tide cycle.
Increased production of ROS/RNS in organisms can beassociated also to the biotransformation of organic xenobiotics,since O2
•− and H2O2 can be generated at substantial amountsduring biotransformation reactions, due to uncoupled transfer ofelectrons from cytochrome P450 through NADPH P450reductase to the substrate (Cesaratto et al., 2004; Zangar etal., 2004). Nevertheless, it is believed that the major organicxenobiotic-stimulated ROS/RNS production in biologicalsystems is the redox cycling reactions, which involve theunivalent reduction of a suitable xenobiotic (e.g. quinones,nitroaromatics) catalyzed by the microsomal NAD(P)H-depen-dent flavoprotein reductases (Kappus, 1986). Under aerobicconditions, the resultant reduced xenobiotic reacts withmolecular oxygen to produce O2
•−, and is covered back to theparent compound, so completing the redox cycle (Lemaire et al.,1994). O2
•− may in turns form H2O2 and, subsequently, thehighly reactive •OH radical.
Other systems can be associated to cellular ROS/RNSgeneration, such as those involved on the inflammatoryresponse or the activity of several oxidases (Jones, 1986).Also, the interaction between different radicals can generateother physiologically important ROS/RNS, like the formationof peroxynitrite (ONOO−) by the reaction of O2
•− with NO(Huie and Padmaja, 1993), or the Russel's generation of 1O2 bythe reaction of two adjacent lipoperoxyl radicals during lipidperoxidation (Russell, 1957; Miyamoto et al., 2003a,b).
Despite the existence of numerous evidences of ROS/RNSgeneration in many organisms, there are few studies directlyevidencing the generation of these reactive species in bivalves,and most of them are related to the immune response of theseorganisms. Bivalve haemocytes are able to generate ROS(Cajaraville et al., 1996), and rate of ROS generation can beaffected by exposure to xenobiotics like copper, benzo[a]pyrene,and paraquat (Gómez-Mendikute and Cajaraville, 2003). Someother direct evidences of ROS generation in mussels came fromstudies on xenobiotic response, as revised by Livingstone(2001). ROS production inmicrosomal extracts ofMytilus edulisis NAD(P)H-dependent, indicating redox cycling reactions(Garcia Martinez et al., 1995; Garcia Martinez and Livingstone,1995). Cancio and Cajaraville (1997) have histochemicallyidentified a series of oxidases capable of generating ROS/RNSin different tissues of mussels Mytilus galloprovincialis.
To our knowledge there are no direct evidences of cellularROS/RNS generation in bivalves from South America, whichremains to be shown. However, several studies have indirectlyindicated the formation of ROS/RNS, through the measurementof cellular oxidized components, which includes DNA strandbreaks, DNAoxidative damage (especially the oxidative product8-oxo-7,8-dihydro-2′-deoxyguanosine, 8-oxodGuo), lipid per-oxidation products, and the protein carbonyls (Walker et al.,2000; Labieniec and Gabryelac, 2004; McDonagh et al., 2005).
3. Antioxidant defense systems
Organisms have evolved antioxidant defense mechanismsthat prevent and intercept ROS/RNS, as well as repairmechanisms for oxidized components. In the former areincluded metal chelators (ferritin, transferrin, coeruplasmin,metallothionein and others), which prevent ROS/RNS produc-tion by intercepting reagents in the Fenton reaction. Also, cellscontain antioxidant enzymes that can intercept ROS/RNS,protecting molecular targets against oxidative injury. The threemajor antioxidant enzymes are the superoxide dismutases(SOD), which decomposes O2
•− to H2O2, catalase (CAT) thatdecomposes H2O2 to molecular oxygen and water, andglutathione peroxidases (GPx) which reduce both H2O2 andlipid hydroperoxides, associated to glutathione oxidation (GSH)(Hebbel, 1986; Sies, 1993). Oxidized glutathione (GSSG)should be reduced to GSH by the ancillary flavoenzymeglutathione reductase (GR), which uses NADPH as electrondonor (Hasspieler et al., 1994). This catalytic cycle lowers theNADPH/NADP+ ratio that in turn should be maintained high inorder to prevent oxidative damage. The activities of twoenzymes of the oxidative pentose phosphate pathway glucose-

591E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
6-phosphate dehydrogenase and 6-phosphogluconate dehydro-genase as well as other enzymes from the intermediarymetabolism, such as the malic enzyme, isocitrate dehydroge-nase, recycle NADPH. Enzymes involved in the transport andelimination of reactive compounds carry other indirectantioxidant functions, e.g. the glutathione-S-transferases andthe transport systems for the glutathione-S-conjugates (Sies,1993). Non-enzymatic defenses include the fat-soluble vita-mins, α-tocopherol and β-carotene, as well as several lowmolecular weight compounds like ascorbic acid and GSH.
The values we have measured for antioxidant defenses in P.perna (Table 1) agree with those reported for other bivalvespecies (Viarengo et al., 1991; Solé et al., 1995; Power andSheehan, 1996; Manduzio et al., 2004; Frenzilli et al., 2004),and also to those exhibited by fishes and mammals, in general(Filho et al., 2001). Some differences observed betweenmussels, vertebrate and other invertebrate species can beobserved, and this can be rationalized in terms of differentenvironmental conditions (i.e. water temperature or oxygenavailability), as well as due to inter-specific differences, such asthe biological rhythms and the reproductive cycles.
Some differences can also be observed in antioxidantdefenses between different tissues of mussels (Table 1). Forexample, the activity of GPx, GR and SOD, as well as the levelsof total GSH, are similar in both digestive glands and gills of P.perna, contrasting with results presented by Manduzio et al.(2004) for the musselM. edulis, which showed higher GR, GPxand SOD in gills. Comparatively to digestive gland, gills have∼3-fold lower CAT activity, 2-fold higher G6PDH and 4-foldGST activities (Almeida et al., 2005).
Antioxidant defenses can change significantly in Brazilianmussel species in response to different stress conditions. In P.perna, these defenses can change seasonally, mostly accordingto reproduction cycles, water temperature and food availability(Filho et al., 2001), in accordance with the profiles observedfrom mussel species from the Northern Hemisphere (Viarengoet al., 1991; Solé et al., 1995; Power and Sheehan, 1995, 1996;Cancio et al., 1999). Mussels sampled in summer showedincreased activity of SOD, CAT and GPx, and increased levelsof GSH and GSSG (Filho et al., 2001). Elevated temperatureand higher food availability would induce oxygen consumption
Table 1Level of antioxidant and complementary enzymes and of total GSH in thedigestive gland and gill of mussels P. perna caught in winter (Almeida et al.,2005)
Digestive gland Gill
SOD 178.1±47.9 212.7±57.1CAT 11.4±4.4 3.1±0.9GPx 8.2±3.3 9.3±7.6GST 129.6±33.1 630.7±173.5G6PDH 50.8±13.7 116.4±34.6GR 84.1±15.2 84.5±21.0Total GSH 2.6±0.5 1.9±0.5
U/mg protein for SOD and CAT.mU/mg protein for GPx, GR, G6PDH and GST.GSH concentrations in μmol/g tissue.
and cellular oxyradical generation, which could be compensatedby the increase of antioxidant defenses. Hence, the decreasedactivity of the antioxidant defense system and GSH/GSSGcontent occurring during winter and spring (Filho et al., 2001)could be directly responsible for an enhanced susceptibility ofmussels to oxidative stress in these periods, despite no increasein lipid peroxidation levels has been observed (Filho et al.,2001). P. perna also showed increased SOD, CAT and GSTactivities in autumn, which was associated with an intensivereproductive activity observed in P. perna at this season(Magalhães, 1998; Filho et al., 2001).
When exposed to air for 4h, mussels P. perna showed nochanges in CAT and GPx activities but increased SODactivity, probably due to a preparative mechanism to deal withthe posterior re-oxygenation period, as proposed by Hermes-Lima et al. (1998) for anoxia tolerant animals (Almeida andBainy, 2006). After 1h of re-oxygenation, SOD activitydecreased to basal values. On the other hand, when thisspecies was exposed to air for a longer period (18h), nochanges were observed in SOD, CAT, GPx, GR, and G6PDHactivities, as well as in total GSH levels in both digestivegland and gills (Almeida et al., 2005).
The increase in the levels of 8-oxodGuo and lipidperoxidation observed after air exposure in P. perna (Almeidaet al., 2005) indicates the production of ROS under thiscondition. Increase in the GST activity was the only antioxidantdefense activated during prolonged hypoxic periods, althoughsuch increase would not prevent the oxidative injury during theexposure period. An increase in GST activity in gills of M.edulis was proposed as a response to oxidative stress when theactivities of other antioxidant enzymes are lowered (Power andSheehan, 1996; Sheehan and Power, 1999; Manduzio et al.,2004). The maintenance of antioxidant and complementaryenzymes at similar levels during air exposure and re-submersioncould contribute to the return of lipid and DNA damage levels tobasal values after ventilation.
Antioxidant defenses in mussels from the Brazilian coast canalso change in response to pollutant exposure. Metals such asCd, Fe, Cu and Pb at LC50 levels and under acute exposure, areable to decrease GPx activity and GSH levels, and to producelipid peroxidation in P. perna (Almeida et al., 2004b; Dafre etal., 2004). GSH depletion was also observed in the bivalvesUnio tumidus and M. galloprovincialis exposed to Cu (Doyotteet al., 1997; Canesi et al., 1998). Interestingly, we found that theenzyme phospholipid hydroperoxide glutathione peroxidase(PHGPx) had an inverse relationship with the levels of lipidperoxidation in mussels exposed to Fe and Cu, suggesting thisenzyme has a role in the protection against lipid peroxidationelicited by these metals (Almeida et al., 2004b).
In another study, P. pernamussels were exposed to paraquat,showing significant increases in GR and G6PDH activities, anddecreased GST activity (Dafre et al., 2004). Indeed, increasedCAT activity was observed in oyster C. rhizophorae exposed tofuradan, a carbamate pesticide. When exposed to differentdiesel oil concentrations, this oyster also presented a dose-dependent increase in GST activity, at salinities of 25 and 15ppt(Silva et al., 2005).

592 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
In field studies, we observed that mussel P. pernatransplanted to polluted sites presented a significant increasein GST activity after 150days, probably related to a great inputof pollutants into the marine environment due to the highrainfall index. Also, mussels Mytella guyanensis collected atpolluted mangroves showed substantial changes in the antiox-idant status, presenting increased GPx, GR, CAT and GSTactivities and GSSG levels, and decreased GSH, compared tocontrol mussels (Torres et al., 2002).
4. Lipid peroxidation
Cell membranes are potential targets of attack by ROS/RNS.The attack of membrane lipids by these species initiates anautocatalytic oxidation process known as lipid peroxidation.During this process, lipids are oxidized generating lipidhydroperoxides (LOOH) as primary products (Fig. 1). LOOHare also generated in 1O2 mediated oxidations (Terao andMatsushita, 1977; Frankel et al., 1979) and by the action ofenzymes such as lipoxygenases (Brash, 1999) and cycloox-ygenases (Hamberg and Samuelsson, 1980).
The formation of LOOH in membranes disrupts the normalcellular metabolism, triggering adaptive responses and/orcausing cell death (Girotti, 1998). Eukaryotic cells haveenzymes that can eliminate these hydroperoxides. Phospho-lipid hydroperoxides can be either detoxified directly by theaction PHGPx or by the consecutive action of phospholipase
R2R1 R1
OO
O
H O
H
R, O2
Acr-dGuo / Cro-dGuo adducts
DNA
D
4-hydro
acrolein (Acr)
2,4-d
crotonaldehyde (Cro)
Linoleic acid: R1= -(CH2)4-CH3, R2 = -(CH2)7-COOH
lipid (LH) lipid hydroperoxide
. .
Fig. 1. During lipid peroxidation a series of reactive aldehydes are formed as by-proand malonaldehyde. These aldehydes can react with DNA bases forming a series of
A2 and classical glutathione peroxidase (cGPx) (Ursini et al.,1991). Glutathione peroxidases are generally believed to beup-regulated by oxidants (Flohé et al., 1997). Indeed,increased activity of cGPx has been detected in tissuessubmitted to oxidative stress (Miyamoto et al., 2003c; Flohéet al., 1997). Increased activity of PHGPx has also beenfound in mussels P. perna exposed to different metals(Almeida et al., 2004b).
The LOOH not detoxified can participate in reactions thatenhance its toxicity. For example, LOOH are known todecompose in the presence of metal ions generating peroxyl(LOO•) (Eq. (1)) and/or alkoxyl radicals (LO•) (Eq. (2))(Girotti, 1998).
LOOH þ Fe3þ→LOO• þ Hþ þ Fe2þ ð1ÞLOOH þ Fe2þ→LO• þ OH− þ Fe3þ ð2Þ
These radicals can propagate the oxidation process to othercellular compartments; undergo fragmentation generatingketones, epoxides, and aldehydes (Esterbauer et al., 1991;Esterbauer, 1993) (Fig. 1); or generate 1O2 through themechanism proposed by Russell (1957). According to thismechanism, the reaction of LOO• occurs through the formationof a cyclic mechanism from a linear tetraoxide intermediate thatdecomposes yielding 1O2 (Howard and Ingold, 1968). Thegeneration of 1O2 by this mechanism has been recently con-firmed by our group in a study in which [18O]-isotopically-
R2
R O
OH
H
H
O
OO
OH
H
O
H
LOO.
[O]
HNE-dGuo Etheno / Ethano adducts
MDA-adducts
NA DNA
xy-2-nonenal (HNE)
malonaldehyde (MDA)
ecadienal (DDE)
ducts, including acrolein, crotonaldehyde, 2,4-decadienal, 4-hydroxy-2-nonenalDNA adducts.

Table 2Levels of 8-oxodGuo (8-oxodGuo residues/106 dGuo), etheno adducts (1,N2-εdGuo/107 dGuo, lipid peroxidation (nmol TBARs/g tissue) in different tissuesof P. perna collected in summer and in autumn
Oxidative injury Tissue Summer Autumn
Lipid peroxidation Digestive gland 54.9±18.3 41.5±11.0Mantle 44.6±18.8 90.5±23.1 ⁎
8-OxodGuo Digestive gland 12.8±5.4 30.8±8.6 ⁎
Mantle 18.2±7.2 51.4±19.9 ⁎
1,N2-εdGuo Digestive gland 16.7±9.5 ⁎ 5.3±1.0Mantle 12.2±5.6 15.5±7.2
⁎ Indicates statistical differences, p<0.05.
593E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
oxygen labeled linoleic acid hydroperoxides were reacted withmetal ions or peroxynitrite (Miyamoto et al., 2003a,b).
We have measured the levels of lipid peroxidation (asmalondialdehyde levels) in mussel species submitted todifferent stresses (Almeida et al., 2003b, 2004b, 2005). Thelevels of lipid peroxidation found in bivalves from the Braziliancoast are in accordance with those found in other aquatic speciesand mammal models, despite the significant differencesobserved between the tissues. Thus, the levels of lipidperoxidation in gills of C. rhizophorae (unpublished) and P.perna, for example, ranged from 4 to 15nmol/g of tissue, but inthe digestive gland or mantle tissues of P. perna and M.guyanensis it ranged from 40 to 120nmol/g of tissue−1, underphysiological conditions (Almeida et al., 2003b, 2005; Torres etal., 2002). Digestive gland and mantle tissues possess higherlevels of polyunsaturated fatty acids (PUFA) than gills (Riberaet al., 1991), and this could partially explain the higher levels oflipid peroxidation observed between different tissues.
The level of lipid peroxidation in mussel species collected inBrazil was also affected by environmental pollutants. Forexample, P. perna exposed to LC50 levels of Cu and Fe hadincreased levels of lipid peroxidation in the digestive gland after120h of exposure (Almeida et al., 2004b), which agrees withprevious findings in mussels from the Northern hemisphere(Viarengo et al., 1999). Mussels P. perna transplanted tocontaminated sites in Santa Catarina Island, Brazil, showed alsohigher levels of lipid peroxidation in the mantle tissue than inthe reference animals (Almeida et al., 2003a). Similarly,mangrove mussels M. guyanensis from polluted sites presentedhigher lipid peroxidation levels in the digestive gland than thereference mussels (Torres et al., 2002). These results indicatethat the measurement of this oxidative injury also serves as agood indication of environmental pollution.
Nevertheless, as in the case of antioxidants, lipid peroxida-tion levels in Brazilian bivalves species can be also influencedby other environmental factors like the air exposure/re-submersion cycles and seasonal changes P. perna exposed toair for 24h had increased levels of lipid peroxidation in both thedigestive gland and gills, another indication that ROS/RNS canbe produced in mussels even when they are not ventilating atlow tides (Almeida et al., 2005). When re-submersed for 3h,levels of lipid peroxidation turned back to control values, whichsuggests that mussels are able to handle the oxidative stresscaused by aerial exposure.
Seasonally, the levels of lipid peroxidation can also changein P. perna, mostly due to changes in the reproductive cycles.This species usually display a maximal annual gameteemission in later autumn, corresponding with the higher levelsof lipid peroxidation observed in the digestive gland at thisperiod (Filho et al., 2001). It has been reported that gonads ofP. perna in autumn have higher lipid and carbohydratemobilization besides protein synthesis due to the reproductiveactivity, and need a longer period to recover from this highmetabolic activity (Magalhães, 1998), justifying the increasedlipoperoxidation levels at this season. During the other seasonsthe levels of lipid peroxidation in this species were similar(Filho et al., 2001).
In our studies with P. perna collected in autumn, nodifferences were observed between the levels of lipidperoxidation in the digestive gland of mussels collected insummer and autumn (Table 2, unpublished data). On thecontrary, lipid peroxidation levels increased significantly inmantle tissue. Mussels studied by Filho et al. (2001) werecollected in May, corresponding to later autumn in Brazil, whileour studies were carried out in mussels collected in thebeginning of April, corresponding to the early autumn. Possibly,mussels collected on later autumn would present increased lipidperoxidation levels in digestive gland due to higher reproduc-tion activity than mussels from early autumn. Nevertheless, theincreased levels of lipid peroxidation in mantle tissue would bea consequence of the increased gametogenesis activity even atearly autumn. As the gonads of mussel are located in the mantletissue, a possible oxidative stress in consequence of increasedreproductive activity should be more evident in this tissue thanin the digestive gland.
5. DNA damage
5.1. Oxidative DNA damage
5.1.1. Strand breaksThe oxidation of DNA by ROS/RNS can produce strand
breaks and a number of different modified DNA bases. DNAstrand breaks represent a major class of oxidative damage toDNA under oxidative stress (Cadet et al., 1997). Hydroxylradicals are able to abstract hydrogen atoms from the sugarmoiety with a preference for carbon 4. The resulting radicals areconverted in most cases into strand breaks through mechanismsthat are well documented (Breen and Murphy, 1995; Cadet etal., 1997). The presence of DNA strand breaks is generallydetermined by the Comet assay, which measures the electro-phoretic migration of relaxed or fragmented DNA away fromthe nuclei of cells immobilized in agarose (Steinert, 1999). Inbivalve species, strand breaks have been extensively studied inthe Northern Hemisphere, mainly related to oxidative stressgenerated due to xenobiotic exposure. To our knowledge, thereare no reports on such studies in bivalve species from theBrazilian coast.
5.1.2. 8-Oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodGuo)Several products of DNA damage formed by the action of
various ROS have been identified (Dizdaroglu, 1993). Recently,

HN
N
N
NH2N H2N H2N
O
dR
HN
N
N
N
O
dR
OHHN
N
NH
N
O
dR
O
dR = 2-deoxyribose
dGuo 8-oxodGuo
ROS / RNS
Fig. 2. The oxidation of 2′-deoxyguanosine (dGuo) by reactive oxygen and nitrogen species (ROS/RNS) can lead to the formation of 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodGuo).
594 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
the evaluation of modified DNA bases, especially 8-oxodGuolevels (Fig. 2), has proved to be a good indicator of oxidativestress caused by xenobiotic exposure in marine organisms,including bivalves (Mallins and Haimanot, 1990; Canova et al.,1998; Rodríguez-Ariza et al., 1999; Livingstone, 2001). Thedetection of 8-oxodGuo has been considered a marker of DNAdamage and oxidative stress, as it represents the main oxidativeDNA product induced in different experimental models by •OHand 1O2, thus being considered as a fingerprint to •OH and 1O2
attack to DNA (Dizdaroglu et al., 1991; Halliwell, 1993).
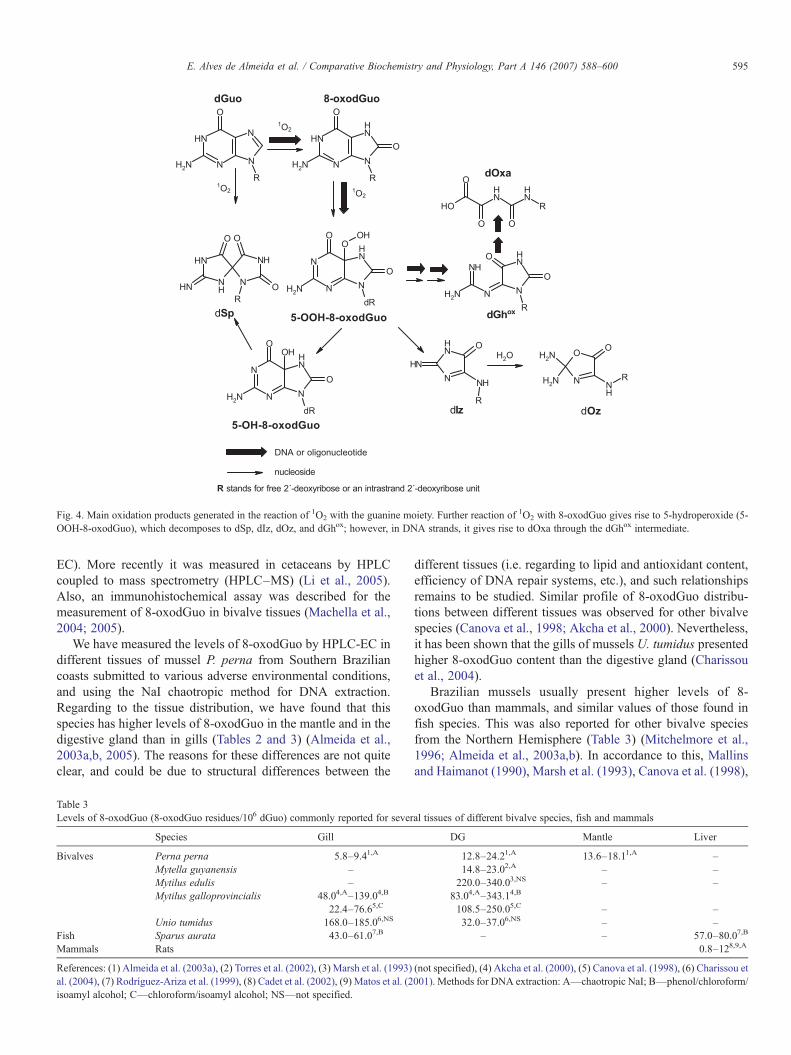
5.1.3. 8-OxodGuo formation in DNA by 1O2: mechanisticstudies
The reaction of 1O2 with DNA is highly specific, contrary tothe reactions with •OH. The reactivity of 1O2 toward isolatedDNA has been studied in detail using the hydrophilic non-ionicN,N′-di(2,3-dihydroxypropyl)-1,4-naphthalenedipropanamide(DHPN) as a water-soluble naphthalene 1O2 carrier (Fig. 3). Inparticular, the endoperoxide prepared with [18O]-isotopicallylabeled molecular oxygen (DHPN18O2) was employed to assessthe reactivity of 1O2 towards biological targets, since theoxidation labeled products formed could be detected andquantified using high performance liquid chromatography–electrospray ionization–tandem mass spectrometry (HPLC–ESI–MS/MS) assay (Martinez et al., 2000). Studies conductedon DNA showed that 8-oxo-7,8-dihydroguanine (8-oxoGua)was the exclusive 1O2 oxidation product within double-strandedisolated DNA (Ravanat et al., 2001). The potential ability for1O2 to oxidize cellular DNAwas also assessed by measurementof the level of [18O]-8-oxodGuo in DNA. This result clearly
O
O
O
NH
OH
OH O
[1
X
R
RX
R =
X
or
Fig. 3. Thermolysis of naphthalene endoperoxides with appropriate substituents actingdi(2,3-dihydroxypropyl)-1,4-naphthalenedipropanamide. NDPO2=endoperoxide of
demonstrated that singlet oxygen, when released within cells,was able to directly oxidize cellular DNA (Ravanat et al., 2000).
Using the accurate HPLC–ESI–MS/MS method and theendoperoxide DHPNO2 or DHPN18O2 acting as a cleanchemical source of 1O2 the predominant final 1O2-oxidationproducts of 8-oxodGuo were identified as the 2-amino-5-[(2′-deoxy-β-D-erythro-pentofuranosyl)amino]-4H-imidazol-4-one(dIz), the 2,2-diamino-4-[2′-deoxy-β-D-erythro-pentofurano-syl) amino]-5-(2H)-oxazolone (dOz) and the diastereomericspiroiminodihydantoin 2′-deoxyribonucleosides (dSp) (Fig. 4)(Martinez et al., 2002). Mass spectrometry measurementsprovide evidence for the formation of an oxidized nucleosidethat displays the same mass spectrometric characteristics thanoxidized guanidinohydantoin 2′-deoxyribonucleoside com-pound (dGhox). Overall, the formation of the currentlyreported 1O2-oxidation products of 8-oxodGuo may berationalized in term of the transient formation of a 5-hydroperoxide, also proposed for the one-electron oxidationof 8-oxoGuo (Luo et al., 2000, 2001). The 1O2-mediatedoxidation of 8-oxoGua in single stranded DNA was found togive rise to oxaluric acid as the overwhelming final stabledecomposition product (Duarte et al., 2000).
5.1.4. 8-OxodGuo levels in musselsThe presence of 8-oxodGuo in a DNA template contribute to
misreading and GC→TA transversions (Kuchino et al., 1987),thus being implicated in carcinogenic processes. Levels of 8-oxodGuo in marine organisms have been assessed throughdifferent methodologies, especially by high performance liquidchromatography coupled to electrochemical detection (HPLC–
ONa
O2]
R
R
+
X = 16O or 18O
as a pure chemical source of 18O-labeled 1O2. DHPNO2=endoperoxide of N,N′-disodium 1,4-naphthalenedipropanoate.
NN
NHN
HN
HN
O
H2N
H2N
H2N H2N
H2N
H2N
H2N
R
NN
NH
HN
O
O
R
N
NH
NH
NH
O
R
N
O
NH
O
R
NH
NNH
O O
O
R
HONH
NH
O
O O
R
NH
N
NH
N
O
O
R
NH
N
N
N
O
O
O
dR
OH
NH
N
N
N
O
O
OH
dR
dGhox
nucleoside
DNA or oligonucleotide
1O2
1O2
1O2
R stands for free 2´-deoxyribose or an intrastrand 2´-deoxyribose unit
dGuo 8-oxodGuo
dOxa
dOzdIz
dSp
H2O
5-OOH-8-oxodGuo
5-OH-8-oxodGuo
Fig. 4. Main oxidation products generated in the reaction of 1O2 with the guanine moiety. Further reaction of 1O2 with 8-oxodGuo gives rise to 5-hydroperoxide (5-OOH-8-oxodGuo), which decomposes to dSp, dIz, dOz, and dGhox; however, in DNA strands, it gives rise to dOxa through the dGhox intermediate.
595E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
EC). More recently it was measured in cetaceans by HPLCcoupled to mass spectrometry (HPLC–MS) (Li et al., 2005).Also, an immunohistochemical assay was described for themeasurement of 8-oxodGuo in bivalve tissues (Machella et al.,2004; 2005).
We have measured the levels of 8-oxodGuo by HPLC-EC indifferent tissues of mussel P. perna from Southern Braziliancoasts submitted to various adverse environmental conditions,and using the NaI chaotropic method for DNA extraction.Regarding to the tissue distribution, we have found that thisspecies has higher levels of 8-oxodGuo in the mantle and in thedigestive gland than in gills (Tables 2 and 3) (Almeida et al.,2003a,b, 2005). The reasons for these differences are not quiteclear, and could be due to structural differences between the
Table 3Levels of 8-oxodGuo (8-oxodGuo residues/106 dGuo) commonly reported for sever
Species Gill
Bivalves Perna perna 5.8–9.41,A
Mytella guyanensis –Mytilus edulis –Mytilus galloprovincialis 48.04,A–139.04,B
22.4–76.65,C
Unio tumidus 168.0–185.06,NS
Fish Sparus aurata 43.0–61.07,B
Mammals Rats
References: (1) Almeida et al. (2003a), (2) Torres et al. (2002), (3) Marsh et al. (1993)al. (2004), (7) Rodríguez-Ariza et al. (1999), (8) Cadet et al. (2002), (9) Matos et al. (2isoamyl alcohol; C—chloroform/isoamyl alcohol; NS—not specified.
different tissues (i.e. regarding to lipid and antioxidant content,efficiency of DNA repair systems, etc.), and such relationshipsremains to be studied. Similar profile of 8-oxodGuo distribu-tions between different tissues was observed for other bivalvespecies (Canova et al., 1998; Akcha et al., 2000). Nevertheless,it has been shown that the gills of mussels U. tumidus presentedhigher 8-oxodGuo content than the digestive gland (Charissouet al., 2004).
Brazilian mussels usually present higher levels of 8-oxodGuo than mammals, and similar values of those found infish species. This was also reported for other bivalve speciesfrom the Northern Hemisphere (Table 3) (Mitchelmore et al.,1996; Almeida et al., 2003a,b). In accordance to this, Mallinsand Haimanot (1990), Marsh et al. (1993), Canova et al. (1998),
al tissues of different bivalve species, fish and mammals
DG Mantle Liver
12.8–24.21,A 13.6–18.11,A –14.8–23.02,A – –220.0–340.03,NS – –
83.04,A–343.14,B
108.5–250.05,C – –32.0–37.06,NS – –
– – 57.0–80.07,B
0.8–128,9,A
(not specified), (4) Akcha et al. (2000), (5) Canova et al. (1998), (6) Charissou et001). Methods for DNA extraction: A—chaotropic NaI; B—phenol/chloroform/

596 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
and Rodríguez-Ariza et al. (1999) also observed higher levels of8-oxodGuo in mussels and fishes as background levels or whenexposed to common pollutants than those observed inmammalian models. The reasons for the increased levels of 8-oxodGuo in aquatic species compared to mammals areunknown. Different factors can be listed here to explain thisincrement of 8-oxodGuo in aquatic species, like the rate ofbioaccumulation and elimination of transition metals thatcontributes to enhance 8-oxodGuo formation, differencesrelated to metabolic rate, or different efficiency of antioxidantdefenses and/or repair mechanisms, but this remains to beclarified.
The artifact oxidation of dGuo due to different methodolog-ical approaches for DNA extraction would contribute to theseobservations. Some authors have proposed that the classicphenol/chloroform method for DNA extraction should beavoided when measuring 8-oxodGuo, because dGuo oxidationmay occur. Most adequate method would be the NaI chaotropicmethod, as proposed by Helbock et al. (1998). However, wehave also studied the levels of 8-oxodGuo in DNA from rattissues and CV1-P monkey cells using the same procedures thanthose used for mussel samples, the NaI chaotropic method, andfound these same discrepancies between mammals and mussels(Cadet et al., 2002; Matos et al., 2000, 2001).
Interestingly, Brazilian mussels showed lower levels of 8-oxodGuo than mussels from Northern Hemisphere (Table 3),which could also be related to differences in the methodologiesfor DNA extraction and hydrolysis. Akcha et al. (2000) studiedthe effects of different DNA extraction methods on 8-oxodGuomeasurements in mussel tissues. When the classical phenol/chloroform method for DNA extraction was used, mussels M.galloprovincialis presented 73+24 and 256+48 residues of 8-oxodGuo/106 dGuo in gills and the digestive gland, respec-tively. In our studies with Brazilian mussels, we always used theChaotropic NaI method for DNA extraction (Wang et al., 1994;Helbock et al., 1998), and when Akcha et al. (2000) used thismethod in M. galloprovincialis the levels of 8-oxodGuo in gillsand the digestive gland were, respectively, 48+11 and 83+20.3residues of 8-oxodGuo/106 dGuo. This could indicate anartifactual oxidation of dGuo residues by using the classicalphenol/chloroform method, as already proposed (Cadet et al.,1998).
As mentioned, the exposure of organisms to pollutants canresult in an increase in ROS/RNS production, leading toincreases in the levels of oxidative DNA damage. We observedthat the mussel P. perna exposed to LC50 levels of Pb and Cd for24h showed higher levels of 8-oxodGuo than in the controlanimals (Almeida et al., 2004a). Also, P. perna musselstransplanted to a polluted site in Florianópolis city (SantaCatarina, Southern Brazil) presented increased levels of 8-oxodGuo in gills and digestive gland, but not in mantle tissue,one year after transplantation (Almeida et al., 2003a).Moreover, mussels M. guyanensis collected in pollutedmangroves of Florianópolis city also presented increased levelsof 8-oxodGuo than reference mussels (Torres et al., 2002).These results clearly indicate an association between contam-inant exposure and increased levels of oxidative DNA damage
in Brazilian mussel species, as observed in other mussel speciesfrom the Northern Hemisphere. Canova et al. (1998) observedhigher levels of 8-oxodGuo in gills and digestive gland of M.galloprovincialis mussels exposed to benzo[a]pyrene. Mallinsand Haimanot (1990) detected a significant increase in thelevels of 8-oxodGuo in fishes (English sole) collected at acontaminated area. In addition, Rodríguez-Ariza et al. (1999)observed elevated levels of 8-oxodGuo in the fish Sparusaurata exposed to dieldrin, paraquat and copper or in animalscaught at contaminated areas.
In response to other environmental stresses not related tocontamination we observed, for example, that mussels P. pernacaught at rocky coast had almost 6 times higher 8-oxodGuolevels than that caught at farming areas, which are permanentlysubmersed (Almeida et al., 2004a). Most mussels are sessileanimals from mesolitoral, thus subjected to air exposure andperiods of anoxia and re-oxygenation due to tidal oscillations.Indeed, when they are re-submersed at high tides an elevation inthe oxygen consumption may also increase the cellulargeneration of ROS/RNS due to the simultaneous enhancedflux of reducing equivalents and oxygen (Jones, 1986; Hermes-Lima et al., 1998). Farmed mussels, being constantly immersedin seawater are not subjected to these stressing conditions, thuspresenting lower levels of oxidative DNA damage.
In order to better understand the influence of these cycles ofair exposure/re-submersion on 8-oxodGuo formation inbivalves, we have measured the levels of this lesion in differenttissues of P. perna exposed to air for 24h, and in musselsexposed to air for 24h followed by re-submersion in seawaterfor 3h (Almeida et al., 2005). It has been shown that bivalvesreduces their oxygen uptake when exposed to air at low tides(Widdows et al., 1979; Griffiths and Buffestein, 1981; Marsdenand Weatherhead, 1998; Willson and Burnett, 2000). Despitethis, mussels P. perna exposed to air for 24h presented higherlevels of 8-oxodGuo in gills, but not in the digestive gland, thanthose mussels kept constantly immersed in seawater. Thisclearly indicates that ROS/RNS can be produced even athypoxic conditions in mussels. After the re-oxygenation period,the levels of 8-oxodGuo decreased to control values, indicatingthat mussels are able to keep basal levels of oxidative damagewhen they are ventilating, which may point to the need ofoxygen to support metabolic processes related to oxidativedamage repair.
Seasonally, the levels of 8-oxodGuo can also change in P.perna. We observed that mussels collected in autumnpresented higher levels of this lesion in both the digestivegland and the mantle tissue (Table 2, unpublished results).Contrary to SOD activity that was increased in summer, 8-oxodGuo increment was not correlated with temperatureincreases. Nevertheless, the increased SOD activity observedin summer would be protecting mussels against DNA damageat this season. In the digestive gland, the 8-oxodGuo levelsobserved in autumn were doubled in comparison to summervalues, while in the mantle tissue they were 3-fold increased.Contrariwise, Charissou et al. (2004) observed no seasonalchanges in the levels of 8-oxodGuo in the digestive gland andgills of the bivalve U. tumidus.

Stress biomarkers
Lesion
Environmental stress
ROS/RNS
Lipid peroxidation
aldehydes
defenses
ROS/RNS
Antioxidant
Fig. 5. Environmental stress is associated with the rate of ROS/RNS generationin organisms, producing lesions in cellular components like DNA and lipids, andthe induction of antioxidant defense systems. These systems can be assessed asstress biomarkers in living organisms.
597E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
5.2. Etheno DNA adducts
In vitro studies have shown that oxidized α,β-unsaturatedaldehydes from lipid peroxidation are able to react with DNAbases generating a series of etheno adducts in DNA (Fig. 1)(Sodum andChung, 1988; Loureiro et al., 2000). The role ofDNAetheno adducts in mutagenic and carcinogenic processes has beenwell investigated (Foiles et al., 1993; Bartsch et al., 1994). Someof these lesions accumulate in DNA after chronic carcinogenexposure, leading to miscoding upon replication or transcription(Swenberg et al., 1992). Numerous etheno adducts have also beendetected as background lesions in tissues of rodents and humansnot exposed to carcinogens (Chung et al., 1996; Loureiro et al.,2002). In vitro studies have shown that 1,N6-ethenoadenine (εA)and 3,N4-ethenocytosine (εC) are formed by iron mediated lipidperoxidation in rat liver microsome incubation mixtures contain-ing nucleosides or nucleotides (el Ghissassi et al., 1995). Elevatedlevels of εdAdo and εdCyd adducts, probably due to excessivegeneration of lipid peroxidation products, were observed in liverDNA of a Long–Evans–Cinnamon rat strain that accumulatescopper in the liver (Nair et al., 1996).
We have studied for the first time the levels of the etheno-DNA adduct 1,N2-etheno-2′-deoxyguanosine (1,N2-εdGuo) inthe digestive gland and mantle tissue of P. perna musselsthrough a recently improved methodology, coupling highperformance liquid chromatography to electrospray ionizationtandem mass spectrometry (Loureiro et al., 2002). This assayhas opened new perspectives for the use of 1,N2-εdGuo as anindicator of oxidative stress in marine bivalves. The levels of 1,N2-εdGuo measured in autumn in the digestive gland of P.perna (Table 2) agree with those found in mammalian tissues(∼5 residues of 1,N2-εdGuo/107 dGuo), but it was higher inmantle tissue (∼15 residues of 1,N2-εdGuo/107 dGuo)(Loureiro et al., 2002; Almeida et al., 2003b). In summer, thelevels of 1,N2-εdGuo in mantle tissue were similar to thosefound in autumn in this species, but in the digestive gland thevalues increased to ∼17 residues of 1,N2-εdGuo/107 dGuo,clearly indicating that level of this lesion in P. perna maychange seasonally (unpublished results).
In preliminary studies with contaminant exposure, we foundthat mussel P. perna transplanted to a polluted site presentedhigher levels of 1,N2-εdGuo than reference mussels in summer,indicating that contaminant exposure can affect the levels of thislesion in bivalves. Interestingly, the levels of lipid peroxidationin these studies followed the same profile as for 1,N2-εdGuolevels, indicating a positive correlation between lipid peroxida-tion and 1,N2-εdGuo levels. However, more extensive studiesare needed to corroborate this.
6. Conclusions
The importance of the study of the effects of environmen-tal stress in aquatic organisms has increased during the lastdecade, especially in those species cultivated for humanconsumption. Indeed, the study of stress response, especiallythose related to oxidative stress (i.e. antioxidant defensesystems, DNA damage and lipid peroxidation) in aquatic
organisms can provide important information to be used astools for examining the environmental quality (Fig. 5). Theresults presented and discussed here with bivalve species fromthe Brazilian coast indicate that different environmentalfactors, like seasonality, air exposure and re-submersioncycles, and the exposure to different classes of contaminants,can cause substantial changes in their antioxidant defensesystems and oxidative injury levels. The assessment of thesesystems in Brazilian bivalves is promising for future studieson environmental quality in Brazil.
Acknowledgements
The authors acknowledge the Brazilian research fundinginstitutions FAPESP (Fundação de Amparo à Pesquisa doEstado de São Paulo), CNPq (Conselho Nacional para oDesenvolvimento Científico e Tecnológico), CNPq-Instituto doMilênio Redoxoma, PRONEX/FINEP (Programa de Apoio aosNúcleos de Excelência) and Fundo Bunka de Pesquisa BancoSumitomo Mitsui and John Simon Guggenheim MemorialFoundation (P.D.M. Fellowship). EAA and SM are post-doctoral recipients of CNPq and FAPESP fellowship, respec-tively. APML, GRM and JO were post-doctoral recipients ofFAPESP fellowships. LFB, CCMG, GER and CAS aredoctorate recipients of FAPESP fellowships.
References
Akcha, F., Ruiz, S., Zampieron, C., Venier, P., Burgeot, T., Cadet, J., Narbonne,J.F., 2000. Benzo[a]pyrene-induced DNA damage in Mytilus galloprovin-cialis: measurement of bulky DNA adducts and DNA oxidative damage interms of 8-oxo-7,8-dihydro-2′-deoxyguanosine formation. Biomarkers 5,355–367.
Almeida, E.A., Bainy, A.C.D., 2006. Effects of aerial exposure on antioxidantdefenses in the brown mussel Perna perna. Braz. J. Biol. Technol. 49 (2),225–229.
Almeida, E.A., Bainy, A.C.D., Loureiro, A.P.M., Medeiros, M.H.G., Di Mascio,P., 2003a. DNA and lipid damage in the brown mussel Perna perna from acontaminated site. Bull. Environ. Contam. Toxicol. 71, 270–275.

598 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
Almeida, E.A., Marques, S.A., Klitzke, C.F., Bainy, A.C.D., Medeiros, M.H.G.,Di Mascio, P., Loureiro, A.P.M., 2003b. DNA damage in digestive gland andmantle tissue of the mussel Perna perna. Comp. Biochem. Physiol., C 135,295–303.
Almeida, E.A., Gomes, O.F., Bainy, A.C.D., Medeiros, M.H.G., Di Mascio,P., 2004a. Lesões oxidativas de DNA de glândulas digestivas de mexilhõesPerna perna como indicadoras de estresse ambiental. Biotemas 17,163–178.
Almeida, E.A., Miyamoto, S., Bainy, A.C.D., Medeiros, M.H.G., Di Mascio, P.,2004b. Protective effects of phospholipid hydroperoxide glutathioneperoxidase (PHGPx) against lipid peroxidation in mussels Perna pernaexposed to different metals. Mar. Pollut. Bull. 49, 386–392.
Almeida, E.A., Bainy, A.C.D., Dafre, A.L., Gomes, O.F., Medeiros, M.H.G., DiMascio, P., 2005. Oxidative stress in digestive gland and gill of the brownmussel (Perna perna) exposed to air and re-submersed. J. Exp. Mar. Biol.Ecol. 318, 21–30.
Bartsch, H., Barbin, A., Marion, M.J., Nair, J., Guichard, Y., 1994. Formation,detection, and role in carcinogenesis of etheno bases in DNA. Drug Metab.Rev. 26, 349–371.
Brash, A.R., 1999. Lipoxygenases: occurrence, functions, catalysis, andacquisition of substrate. J. Biol. Chem. 274, 23679–23682.
Breen, A.P., Murphy, J.A., 1995. Reactions of oxyl radicals with DNA. FreeRadical Biol. Med. 18, 1033–1077.
Cadet, J., Berger, M., Douki, T., Ravanat, J.L., 1997. Oxidative damage to DNA:formation, measurement, and biological significance. Rev. Physiol.,Biochem. Pharmacol. 131, 1–87.
Cadet, J., D'Am, C., Douki, T., Pouget, J.P., Ravanat, J.L., Sauvaigo, S., 1998.Facts and artifacts in the measurement of oxidative base damage to DNA.Free Radical Res. 29, 541–550.
Cadet, J., Douki, T., Frelon, S., Gasparutto, D., Ravanat, J.-L., Sauvaigo, S.,Martinez, G.R., Medeiros, M.G.H., Di Mascio, P., 2002. Oxidative damageto DNA: formation and measurement. In: Pasquier, C. (Ed.), Proceedings ofthe 11th Conference of SFRRI, Paris, Monduzzi Editore. S.P.A. Bologna,Italy, pp. 129–135.
Cajaraville, M.P., Olabarrieta, I., Marigomez, I., 1996. In vitro activities inmussel hemocytes as biomarkers of environmental quality: a case study inthe Abra Estuary (Biscay Bay). Ecotoxicol. Environ. Saf. 35, 253–260.
Cancio, I., Cajaraville, M.P., 1997. Histochemistry of oxidases in several tissuesof bivalve molluscs. Cell Biol. Int. 21, 575–584.
Cancio, I., Ibabe, A., Cajaraville, M.P., 1999. Seasonal variation of peroxisomalenzyme activities and peroxisomal structure in mussels Mytilus gallopro-vincialis and its relationship with the lipid content. Comp. Biochem. Physiol.C 123, 135–144.
Canesi, L., Ciacci, C., Piccoli, G., Stocchi, V., Viarengo, A., Gallo, G., 1998.In vitro and in vivo effects of heavy metals on mussel digestive glandhexokinase activity: the role of glutathione. Comp. Biochem. Physiol. C120, 261–268.
Canova, S., Degan, P., Peters, L.D., Livingstone, D.R., Voltan, R., Venier, P.,1998. Tissue dose, DNA adducts, oxidative DNA damage and CYP1A-immunopositive proteins in mussels exposed to waterbone benzo[a]pyrene.Mutat. Res. 399, 17–30.
Cesaratto, L., Vascotto, C., Calligaris, S., Tell, G., 2004. The importance ofredox state in liver damage. Ann. Hepatol. 3, 86–92.
Chandel, N.S., McClintock, D.S., Feliciano, C.E., Wood, T.M., Melendez, J.A.,Rodriguez, A.M., Schumacker, P.T., 2000. Reactive oxygen speciesgenerated at mitochondrial complex III stabilize hypoxia-inducible factor-1α during hypoxia. J. Biol. Chem. 275, 25130–25138.
Charissou, A.M., Cossu-Leguille, C., Vasseur, P., 2004. Relationship betweentwo oxidative stress biomarkers, malondialdehyde and 8-oxo-7,8-dihydro-2′-deoxyguanosine, in the freshwater bivalve Unio tumidus. Sci. TotalEnviron. 322, 109–122.
Chung, F.L., Chen, H.C.J., Nath, R.G., 1996. Lipid peroxidation as a potentialendogenous source for the formation of exocyclic DNA adducts.Carcinogenesis 17, 2105–2111.
Dafre, A.L., Medeiros, I.D., Müller, I.C., Ventura, E.C., Bainy, A.C.D., 2004.Antioxidant enzymes and thiol/disulfide status in the digestive gland of thebrown mussel Perna perna exposed to lead and paraquat. Chem. Biol.Interact. 149, 97–105.
Dawson, T.L., Gores, G.J., Nieminen, A.L., Herman, B., Lemasters, J.J., 1993.Mitochondria as a source of reactive oxygen species during reductive stressin rat hepatocytes. Am. J. Physiol., Cell Physiol. 264, 961–967.
Dizdaroglu, M., 1993. Quantitative determination of oxidative base damage inDNA by stable isotope-dilution mass spectrometry. FEBS Lett. 315 (1), 1–6.
Dizdaroglu, M., Rao, G., Halliwell, B., Gajewski, E., 1991. Damage to the DNAbases in mammalian chromatin by hydrogen peroxide in the presence offerric and cupric ions. Arch. Biochem. Biophys. 285, 317–324.
Doyotte, A., Cossu, C., Jacquin, M.C., Babut, M., Vasseur, P., 1997. Antioxidantenzymes, glutathione and lipid peroxidation of experimental or fieldexposure in the gills and the digestive gland of the freshwater bivalve Uniotumidus. Aquat. Toxicol. 39, 93–110.
Duarte, V., Gasparutto, D., Jaquinod, M., Cadet, J., 2000. In vitro DNAsynthesis opposite oxazolone and repair of this DNA damage using modifiedoligonucleotides. Nucleic Acids Res. 28, 1555–1563.
el Ghissassi, F., Barbin, A., Nair, J., Bartsch, H., 1995. Formation of 1,N4-ethenocytosine by lipid peroxidation products and nucleic acid bases. Chem.Res. Toxicol. 8, 278–283.
Esterbauer, H., 1993. Cytotoxicity and genotoxicity of lipid-oxidation products.Am. J. Clin. Nutr. 57, 779S–786S.
Esterbauer, H., Shaur, J.S., Zollner, H., 1991. Chemistry and biochemistry of 4-hydroxynonenal, malondialdehyde and related aldehydes. Free Radical Biol.Med. 11, 81–128.
Filho, D.W., Tribess, T., Gáspari, C., Cláudio, F.D., Torres, M.A., Magalhães,A.R.M., 2001. Seasonal changes in antioxidant defenses of the digestivegland of the brown mussel (Perna perna). Aquaculture 203, 149–158.
Flohé, L., Wingender, E., Brigelius-Flohé, R., 1997. The regulation ofglutathione peroxidases, p. 415–435. In: Forman, H., Cadenas, E. (Eds.),Oxidative Stress and Signal Transduction. Chapman & Hall, New York, NY.
Foiles, P.G., Miglietta, L.M., Nishikawa, A., Kusmierek, J.T., Singer, B., Chung,F.C., 1993. Development of monoclonal antibodies specific for 1,N2-ethenodeoxyguanosine and N2,3-ethenodeoxyguanosine and their use forquantitation of adducts in G12 cells exposed to chloroacetaldehyde.Carcinogenesis 14, 113–116.
Frankel, E.N., Neff, W.E., Bressler, T.R., 1979. Analysis of autooxidized fats bygas-chromatography mass spectrometry: V. Photosensitized oxidation.Lipids 14, 961–967.
Frenzilli, G., Bocchetti, R., Pagliarecci, M., Nigro, M., Annarumma, F.,Scarcelli, V., Fattorini, D., Regoli, F., 2004. Time-course evaluation of ROS-mediated toxicity in mussels, Mytilus galloprovincialis, during a fieldtranslocation experiment. Mar. Environ. Res. 58, 609–613.
Garcia Martinez, P., Livingstone, D.R., 1995. Benzo[a]pyrene-dione stimulatedoxyradical production by microsomes of digestive gland of the commonmussel Mytilus edulis L. Mar. Environ. Res. 39, 185–189.
Garcia Martinez, P., Winston, G.W., Metosh-Dickey, C., O'Hara, S.C.M.,Livingstone, D.R., 1995. Nitrofurantoin-stimulated reactive oxygen speciesproduction and genotoxicity in digestive gland microsomes and cytosol ofthe common mussel (Mytilus edulis, L.). Toxicol. Appl. Pharmacol. 131,332–341.
Girotti, A.W., 1998. Lipid hydroperoxide generation, turnover, and effectoraction in biological systems. J. Lipid Res. 39, 1529–1542.
Goldberg, E.D., 1975. The mussel watch: a first step in global marinemonitoring. Mar. Pollut. Bull. 6, 111–132.
Gómez-Mendikute, A., Cajaraville, M.P., 2003. Comparative effects ofcadmium, copper, paraquat and benzo[a]pyrene on the actin cytoskeletonand production of reactive oxygen species (ROS) in mussel haemocytes.Toxicol. In Vitro 17, 539–546.
Griffiths, R.J., Buffestein, R., 1981. Aerial exposure and energy input in thebivalve Choromytilus meridionalis (Kr.). J. Mar. Biol. Ecol. 52,219–229.
Guderley, H., Demers, A., Couture, P., 1994. Acclimatization of blue mussel(Mytilus edulis Linnaeus, 1758) to intertidal conditions: effects on mortalityand gaping during air exposure. J. Shellfish Res. 13, 379–385.
Halliwell, B., 1992. Reactive oxygen species and the central nervous system. J.Neurochem. 59, 1609–1623.
Halliwell, B., 1993. Oxidative DNA damage: meaning and measurement. In:Halliwell, B., Auroma, O.I. (Eds.), DNA and Free Radicals. Ellis Horwood,New York, pp. 67–79.

599E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
Hamberg, M., Samuelsson, B., 1980. Stereochemistry in the formation of 9-hydroxy-10,12-octadecadienoic acid and 13-hydroxy-9,11-octadecadienoicacid from linoleic acid by fatty acid cyclooxygenase. Biochim. Biophys.Acta 617, 545–547.
Hasspieler, B.M., Behar, J.V., Di Giulio, RT., 1994. Glutathione-dependentdefense in channel catfish (Ictalurus punctatus) and brown bullhead(Ameriurus nebulosus). Ecotoxicol. Environ. Saf. 28, 82–90.
Hebbel, R.P., 1986. Erythrocyte antioxidants and membrane vulnerability. J.Lab. Clin. Med. 107, 401–404.
Helbock, H.J., Beckman, K.B., Shigenaga, M.K., Walter, P.B., Woodal, A.A.,Yeo, H.C., Ames, B.N., 1998. DNA oxidation matters: the HPLC-electrochemical detection assay of 8-oxo-deoxyguanosine and 8-oxo-guanine. Proc. Natl. Acad. Sci. U. S. A. 95, 288–293.
Hermes-Lima, M., Storey, J.M., Storey, K.B., 1998. Antioxidant defences andmetabolic depression. The hypothesis of preparation for oxidative stress inland snail. Comp. Biochem. Physiol. B 120, 437–448.
Howard, J.A., Ingold, K.U., 1968. The self-reaction of sec-butylperoxyradicals. Confirmation of the Russell mechanism. J. Am. Chem. Soc. 90,1056–1058.
Huie, R.E., Padmaja, S., 1993. The reaction of NO and superoxide. Free Radic.Res. Commun. 18, 195–199.
Jones, D.P., 1986. Renal metabolism during normoxia, hypoxia, and ischemicinjury. Annu. Rev. Physiol. 48, 33–50.
Kappus, H., 1986. Overview of enzyme systems involved in bio-reduction ofdrugs and in redox cycling. Biochem. Pharmacol. 35, 1–6.
Kehrer, J.P., 2000. The Haber–Weiss reaction and mechanisms of toxicity.Toxicology 149, 43–50.
Kennedy, V.S., 1976. Desiccation, higher temperatures and upper intertidallimits of three species of sea mussels (Mollusca: Bivalvia) in New Zealand.Mar. Biol. 35, 127–137.
Korshunov, S.S., Skulachev, V., Starkov, A.A., 1997. High protonic potentialactuates a mechanism of production of reactive oxygen species inmitochondria. FEBS Lett. 416, 15–18.
Kuchino, Y., Mori, F., Kasai, H., Iwai, S., Miura, K., Ohtsuka, E., Nishimura, S.,1987. Misreading of DNA templates containing 8-hydroxydeoxyguanosineat the modified base and adjacent residues. Nature 327, 77–79.
Labieniec, M., Gabryelac, T., 2004. Response of DNA, proteins and membranebilayer in the digestive gland cells of freshwater mussel Unio tumidus totannins exposure. Toxicol. In Vitro 18, 773–781.
Lemaire, P., Livingstone, D.R., 1993. Pro-oxidant/antioxidant processes andorganic xenobiotic interactions in marine organisms, in particular theflounder Platichthys flesus and the mussel Mytilus edulis. Trends Comp.Biochem. Physiol. 1, 1119–1147.
Lemaire, P., Matthews, A., Forlin, L., Livingstone, D.R., 1994. Stimulation ofoxyradical production of hepatic microsomes of flounder (Platichthys flesus)and perch (Perca fluviatilis) by model and pollutant xenobiotics. Arch.Environ. Contam. Toxicol. 26, 191–200.
Li, C.S., Wu, K.Y., Chang-Chien, G.P., Chou, C.C., 2005. Analysis of oxidativeDNA damage 8-hydroxy-2′-deoxyguanosine as a biomarker of exposuresto persistent pollutants for marine mammals. Environ. Sci. Technol. 39,2455–2460.
Livingstone, D.R., 2001. Contaminant-stimulated reactive oxygen speciesproduction and oxidative damage in aquatic organisms. Mar. Pollut. Bull.42, 656–666.
Loureiro, A.P.M., Di Mascio, P., Gomes, O.F., Medeiros, M.H.G., 2000. trans,trans-2,4-Decadienal-induced 1,N2-etheno-2′-deoxyguanosine adduct for-mation. Chem. Res. Toxicol. 13, 601–609.
Loureiro, A.P.M., Marques, S.A., Garcia, C.C.M., Di Mascio, P., Medeiros,M.H.G., 2002. Development of an on-line liquid chromatography-electrospray tandem mass spectrometry system to quantitatively deter-mine 1,N2-etheno-2′-deoxyguanosine in DNA. Chem. Res. Toxicol. 15,1302–1308.
Luo, W., Muller, J.G., Rachlin, E.M., Burrows, C.J., 2000. Characterization ofspiroiminodihydantoin as a product of one-electron oxidation of 8-oxo-7,8-dihydroguanosine. Org. Lett. 2, 613.
Luo, W., Muller, J.G., Rachlin, E.M., Burrows, C.J., 2001. Characterization ofhydantoin products from one-electron of 8-oxo-7,8-dihydroguanosine in anucleoside model. Chem. Res. Toxicol. 14, 927.
Machella, N., Regoli, F., Cambria, A., Santella, R.M., 2004. Oxidativedamage to DNA: an immunohistochemical approach for detection of 7,8.dihydro-8-oxodeoxyguanosine in marine organisms. Mar. Environ. Res.58, 725–729.
Machella, N., Regoli, F., Santella, R.M., 2005. Immunofluorescent detection of8-oxo-dG and PAH bulky adducts in fish liver and mussel digestive gland.Aquat. Toxicol. 71, 335–343.
Magalhães, A.R.M., 1998. Efeito da parasitose por Trematoda bucephalidae nareprodução, composição bioquímica e índice de condição de mexilhões Pernaperna. Doctoral Thesis. Instituto de Biociências, USP, São Paulo, SP, Brazil.
Mallins, D.C., Haimanot, R., 1990. 4,6-Diamino-5-formamidopyrimidine, 9-hydroxyguanine and 8-hydroxyadenine in DNA from neoplastic liver ofEnglish sole exposed to carcinogens. Biochem. Biophys. Res. Commun.173, 614–619.
Manduzio, H., Monsinjon, T., Galap, C., Leboulenger, F., Rocher, B., 2004.Seasonal variations in antioxidant defences in the blue mussels Mytilusedulis collected from a polluted area: major contributions in gills of aninducible isoform of Cu/Zn-superoxide dismutase and of glutathione S-transferase. Aquat. Toxicol. 70, 83–93.
Marsden, I.D., Weatherhead, M.A., 1998. Effects of aerial exposure on oxygenconsumption by the New Zealand mussel Perna canaliculus (Gmelin, 1791)from an intertidal habitat. J. Exp. Mar. Biol. Ecol. 230, 15–29.
Marsh, J.W., Chipman, J.K., Livingstone, D.R., 1993. Formation of DNAadducts following laboratory exposure of the mussel, Mytilus edulis, toxenobiotics. Sci. Total Environ. 208, 567–572.
Martinez, G.R., Ravanat, J.L., Medeiros, M.G.H., Di Mascio, P., 2000.Synthesis of a naphthalene endoperoxyde as a source of 18O-labeled singletoxygen for mechanistic studies. J. Am. Chem. Soc. 122, 10212–10213.
Martinez, G.R., Medeiros, M.H.G., Ravanat, J.L., Cadet, J., Di Mascio, P., 2002.[18O]-labeled singlet oxygen as a tool for mechanistic studies of 8-oxo-7,8-dihydroguanine oxidative damage: detection of spiroiminodihydantoin,imidazolone and oxazolone derivatives. J. Biol. Chem. 383, 607–617.
Matos, H.R., Di Mascio, P., Medeiros, M.H.G., 2000. Protective effect oflycopene on lipid peroxidation and oxidative DNA damage in cell culture.Arch. Biochem. Biophys. 383, 56–59.
Matos, H.R., Capelozzi, V.L., Gomes, O.F., Di Mascio, P., Medeiros, M.H.G.,2001. Lycopene inhibits DNA damage and liver necrosis in rats treated withferric nitrilotriacetate. Arch. Biochem. Biophys. 396, 171–177.
McDonagh, B., Tyther, R., Sheehan, D., 2005. Carbonylation and glutathionyla-tion of proteins in the blue mussel Mytilus edulis detected by proteomicanalysis and Western blotting: actin as a target for oxidative stress. Aquat.Toxicol. 73, 315–326.
Michiels, C., Minet, E., Mottet, D., Raes, E., 2002. Regulation of geneexpression by oxygen: NF-kappaB and HIF-1, two extremes. Free Radic.Biol. Med. 33, 1231–1242.
Mitchelmore, C.L., Chipman, J.K., Garcia-Martinez, P., Lemaire, P., Peters,L.D., Livingstone, D.R., 1996. Normal status of hepatic 7-ethoxyresorufinO-deethylase (EROD) activity, antioxidant enzymes and DNA oxidationin turbot (Scophthalmus maximus) and other flatfish species followingexposure to nitroaromatic compounds. Mar. Environ. Res. 42, 329–333.
Miyamoto, S., Martinez, G.R., Martins, A.P., Medeiros, M.H.G., Di Mascio, P.,2003a. Direct evidence of singlet molecular oxygen [O2(
1Δg)] production inthe reaction of linoleic acid hydroperoxide with peroxynitrite. J. Am. Chem.Soc. 125, 4510–4517.
Miyamoto, S., Martinez, G.R., Medeiros, M.H.G., Di Mascio, P., 2003b. Singletmolecular oxygen generated from lipid hydroperoxides by the Russellmechanism: studies using 18O-labeled linoleic acid hydroperoxide andmonomol light emission measurements. J. Am. Chem. Soc.. 125, 6172–6179.
Miyamoto, S., Dupas, C., Murota, K., Terao, J., 2003c. Phospholipidhydroperoxides are detoxified by phospholipase A2 and GSH peroxidasein rat gastric mucosa. Lipids 38, 641–649.
Nair, J., Sone, H., Nagao, M., Barbin, A., Bartsch, H., 1996. Copper-dependentformation of miscoding etheno-DNA adducts in the liver of Long–Evans–Cinnamon (LEC) rats developing hereditary hepatitis and hepatocellularcarcinoma. Cancer Res. 56, 1267–1271.
Ochi, T., Takahashi, K., Ohsawa, M., 1987. Indirect evidences for the inductionof a pro-oxidant state by cadmium chloride in cultured mammalian cells anda possible mechanism for the induction. Mutat. Res. 180, 257–266.

600 E. Alves de Almeida et al. / Comparative Biochemistry and Physiology, Part A 146 (2007) 588–600
Pellerin-Massicote, J., 1994. Oxidative processes as indicators of chemical stressin marine bivalves. J. Aquat. Ecosyst. Health 3, 101–111.
Power, A., Sheehan, D., 1995. Seasonal variations in the levels of antioxidantenzymes in Mytilus edulis. Biochem. Soc. Trans. 23, 354S–362S.
Power, A., Sheehan, D., 1996. Seasonal variations in the antioxidant defensesystems of gill and digestive gland of the blue mussel,Mytilus edulis. Comp.Biochem. Physiol. C 114, 99–103.
Ravanat, J.L., Di Mascio, P., Martinez, G.R., Medeiros, M.H.G., Cadet, J., 2000.Singlet oxygen induces oxidation of cellular DNA. J. Biol. Chem. 275,40601–40604.
Ravanat, J.L., Di Mascio, P., Martinez, G.R., Medeiros, M.H.G., 2001. Singletoxygen induces oxidation of cellular DNA. J. Biol. Chem. 276,40601–40604.
Ribera, D., Narbonne, J.F., Michel, X., Livingstone, D.R., O`hara, S., 1991.Responses of antioxidants and lipid peroxidation in mussels to oxidativedamage exposure. Comp. Biochem. Physiol. C 100, 177–181.
Rodríguez-Ariza, A., Alhama, J., Díaz-Méndez, F.M., López-Barea, J., 1999.Content of 8-oxodG in chromosomal DNA of Sparatus aurata fish asbiomarker of oxidative stress and environmental pollution. Mutat. Res. 438,97–107.
Russell, G.A., 1957. Deuterium-isotope effects in the autoxidation of aralkylhydrocarbons. Mechanism of the interaction of peroxyl radicals. J. Am.Chem. Soc. 79, 3871–3877.
Sheehan, D., Power, A., 1999. Effects of seasonality on xenobiotic andantioxidant defense mechanisms in bivalve molluscs. Comp. Biochem.Physiol. C 123, 193–199.
Sies, H., 1993. Strategies of antioxidant defenses. Eur. J. Biochem. 215,213–219.
Silva, A.Z., Zanette, J., Ferreira, J.F., Guzenski, J., Marques, M.R.F., Bainy,A.C.D., 2005. Effects of salinity on biomarker responses in Crassostrearhizophorae (Mollusca, Bivalvia) exposed to diesel oil. Ecotoxicol.Environ. Saf. 62 (3), 376–382.
Sodum, R.S., Chung, F.L., 1988. 1,N2-Ethenodeoxyguanosine as a potentialmarker for DNA adduct formation by trans-4-hydroxy-2-nonenal. CancerRes. 48, 320–323.
Solé, M., Porte, C., Albaigés, J., 1995. Seasonal variation in the mixed functionoxygenase system and antioxidant defenses of the mussel Mytilusgalloprovincialis. Environ. Toxicol. Chem. 14, 157–164.
Steinert, S.A., 1999. DNA damage as a bivalve biomarker. Biomarkers 4,492–496.
Stohs, S.J., Hagchi, D., Hassoun, E., Bagshi, M., 2000. Oxidative mechanismsin the toxicity of chromium and cadmium ions. J. Environ. Pathol. Toxicol.Oncol. 19, 201–213.
Storey, K.B., 1996. Oxidative stress: animal adaptations in nature. Braz. J. Med.Biol. Res. 29, 1715–1733.
Swenberg, J.A., Fedtke, N., Ciroussel, F., Barbin, A., Bartsch, H., 1992. Ethenoadducts formed in DNA of vinyl chloride-exposed rats are highly persistentin liver. Carcinogenesis 13, 727–729.
Tatrai, E., Kovacikova, Z., Hudak, A., Adamis, Z., Ungvary, G., 2001.Comparative in vitro toxicity of cadmium and lead on redox cycling in typeII pneumocytes. J. Appl. Toxicol. 21, 479–483.
Terao, J., Matsushita, S., 1977. Products formed by photosensitized oxidation ofunsaturated fatty acid esters. J. Am. Oil Chem. Soc. 54, 234–238.
Torres, M.A., Testa, C.P., Gáspari, C., Masutti, M.B., Panitz, C.M.N., Curi-Pedrosa, R., Almeida, E.A., Di Mascio, P., Whilhelm-Filho, D., 2002.Oxidative stress in the mussel Mytella guyanensis from polluted mangroveson Santa Catarina Island, Brazil. Mar. Pollut. Bull. 44, 923–932.
Turrens, J.F., Boveris, A., 1980. Generation of superoxide anion by the NADHdehydrogenase of bovine heart mitochondria. Biochem. J. 191, 421–427.
Turrens, J.F., Alexandre, A., Lehninger, A.L., 1985. Ubisemiquinone is theelectron donor for superoxide formation by complex III of heartmitochondria. Arch. Biochem. Biophys. 237, 408–414.
Ursini, F., Maiorino, M., Sevanian, A., 1991. Membrane hydroperoxides. In:Sies, H. (Ed.), Oxidative Stress: Oxidants and Antioxidants. AcademicPress, New York, pp. 319–336.
Viarengo, A., Canesi, L., Pertica, M., Livingstone, D.R., 1991. Seasonalvariations in the antioxidant defense enzymes and lipid peroxidation of thedigestive gland of mussels. Comp. Biochem. Physiol. C 100, 187–190.
Viarengo, A., Burlando, B., Cavaletto, M., Marchi, B., Ponzano, E., Blasco, J.,1999. Role of metallothionein against oxidative stress in the mussel Mytilusgalloprovincialis. Am. J. Physiol. 277, R1612–R1619.
Walker, C.H., Hopkin, S.P., Sibly, R.M., Peakall, D.B., 1996. Principles ofEcotoxicology. Taylor and Francis, London. 321 pp.
Walker, S.T., Mantle, D., Bythell, J.C., Thomason, J.C., 2000. Oxidative-stress:comparison of species specific and tissue specific effects in the marinebivalves Mytilus edulis (L.) and Dosinia lupinus (L.). Comp. Biochem.Physiol. B 127, 347–355.
Wang, L., Hirayasu, K., Ishizawa, M., Kobayashi, Y., 1994. Purification ofgenomic DNA from human whole blood by isopropanol-fractionation withconcentrated NaI and SDS. Nucleic Acids Res. 22, 1774–1775.
Widdows, J., Bayne, B.L., Livingstone, D.R., Newell, R.I.E., Donkin, P., 1979.Physiological and biochemical responses of bivalve molluscs to exposure toair. Comp. Biochem. Physiol. A 62, 301–308.
Willson, L.L., Burnett, L.E., 2000. Whole animal and gill tissue oxygen uptakein the Eastern oyster, Crassostrea virginica: effects of hypoxia, hypercapnia,air exposure, and infection with the protozoan parasite Perkinsus marinus. J.Exp. Mar. Biol. Ecol. 246, 233–240.
Zangar, R.C., Davydov, D.R., Verma, S., 2004. Mechanisms that regulateproduction of reactive oxygen species by cytochrome P450. Toxicol. Appl.Pharmacol. 199, 316–331.
Related Documents











