The International Journal of Biochemistry & Cell Biology 39 (2007) 2083–2092 Overexpression of phospholipase C-1 inhibits NGF-induced neuronal differentiation by proliferative activity of SH3 domain Truong Le Xuan Nguyen a , Keqiang Ye b , Sung-Woo Cho c , Jee-Yin Ahn a,∗ a Department of Molecular Cell Biology, Center for Molecular Medicine, Samsung Biomedical Research Institute, Sungkyunkwan University School of Medicine, Suwon 440-746, Republic of Korea b Department of Pathology and Laboratory Medicine, Emory University, School of Medicine, Atlanta, GA 30322, USA c Department of Biochemistry and Molecular Biology, University of Ulsan, College of Medicine, Seoul 138-736, Republic of Korea Received 3 May 2007; accepted 30 May 2007 Available online 7 June 2007 Abstract Since the biological role of phospholipase C (PLC) 1 in neuronal differentiation still barely understood, here, we report that overexpression of PLC 1 inhibits neurite outgrowth and prolonged proliferation ability of PLC 1 contribute to the alteration of cell cycle regulatory proteins, subsequently exiting from cell growth arrest. Deletion of the SH3 or the entire SH223 domains, but not deletion of the N-SH2 or both the N-SH2 and C-SH2 domains expressing cells abolishes the differentiation-inhibitory effects of PLC 1, displaying depression of PCNA and elevation of cyclin D1. Moreover, these cells declined CDK1 and CDK2 expression and increased p21 WAF -1, accompanying with G 2 /M accumulation. Some antiproliferative reagents are able to restore neurite outgrowth in PLC 1 cells, showing G 2 /M arrest. Our findings suggest that the proliferation activity of PLC 1 via its SH3 domain may be coupled with the flight from growth arrest by NGF, thereby inhibiting neuronal differentiation. © 2007 Elsevier Ltd. All rights reserved. Keywords: Phospholipase C (PLC) 1; Differentiation; Proliferation; PC12 cells; SH domain 1. Introduction Phospholipase C (PLC) elicits the hydrolysis of phosphatidylinositol 4,5-bisphosphate, resulting in the generation of inositol 1,4,5-triphsophate (IP3) and dia- cylglycerol (DAG), which in turn release intracellular Ca2+ and the activation of protein kinase C (PKC), respectively. One PLC isoform, PLC 1, has been iden- tified as an excellent substrate for nerve growth factor receptor, is crucial for the regulation of cell proliferation and differentiation (Pawson, 1995). The SH3 domain ∗ Corresponding author. Tel.: +82 31 299 6134; fax: +82 31 299 6139. E-mail address: [email protected] (J.-Y. Ahn). has been implicated in the mitogenesis of PLC 1, but not lipase activity of this protein (Huang et al., 1995; Smith et al., 1994; Ye et al., 2002). It was also recently reported that the SH3 domain of PLC 1 binds to the Ras nucleotide exchange factor, Sos, thus augmenting Ras activity (Kim et al., 2000), and also elicits the acti- vation of mitogen-activated protein kinase (MAPK) via an interaction with the SH2 domain containing leukocyte protein, SLP-76 (Yablonski, Kadlecek, & Weiss, 2001). Quite recently, we demonstrated that the SH3 domain is also required for the activation of nuclear PI3K and cell proliferation (Ye et al., 2002). Recently, there has been increased interest in the role of PLC 1 in cell differentiation. The results of a number of studies have indicated that PLC 1 per- forms a function in certain cell differentiation events, 1357-2725/$ – see front matter © 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2007.05.020

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
ocnPiic©
K
1
pgcCrtra
f
1
The International Journal of Biochemistry & Cell Biology 39 (2007) 2083–2092
Overexpression of phospholipase C-�1 inhibits NGF-inducedneuronal differentiation by proliferative activity of SH3 domain
Truong Le Xuan Nguyen a, Keqiang Ye b, Sung-Woo Cho c, Jee-Yin Ahn a,∗a Department of Molecular Cell Biology, Center for Molecular Medicine, Samsung Biomedical Research Institute,
Sungkyunkwan University School of Medicine, Suwon 440-746, Republic of Koreab Department of Pathology and Laboratory Medicine, Emory University, School of Medicine, Atlanta, GA 30322, USA
c Department of Biochemistry and Molecular Biology, University of Ulsan, College of Medicine, Seoul 138-736, Republic of Korea
Received 3 May 2007; accepted 30 May 2007Available online 7 June 2007
bstract
Since the biological role of phospholipase C (PLC) �1 in neuronal differentiation still barely understood, here, we report thatverexpression of PLC �1 inhibits neurite outgrowth and prolonged proliferation ability of PLC �1 contribute to the alteration ofell cycle regulatory proteins, subsequently exiting from cell growth arrest. Deletion of the SH3 or the entire SH223 domains, butot deletion of the N-SH2 or both the N-SH2 and C-SH2 domains expressing cells abolishes the differentiation-inhibitory effects of
LC �1, displaying depression of PCNA and elevation of cyclin D1. Moreover, these cells declined CDK1 and CDK2 expression andncreased p21WAF-1, accompanying with G2/M accumulation. Some antiproliferative reagents are able to restore neurite outgrowthn PLC �1 cells, showing G2/M arrest. Our findings suggest that the proliferation activity of PLC �1 via its SH3 domain may beoupled with the flight from growth arrest by NGF, thereby inhibiting neuronal differentiation.
2007 Elsevier Ltd. All rights reserved.
PC12 c
eywords: Phospholipase C (PLC) �1; Differentiation; Proliferation;. Introduction
Phospholipase C (PLC) elicits the hydrolysis ofhosphatidylinositol 4,5-bisphosphate, resulting in theeneration of inositol 1,4,5-triphsophate (IP3) and dia-ylglycerol (DAG), which in turn release intracellulara2+ and the activation of protein kinase C (PKC),
espectively. One PLC isoform, PLC �1, has been iden-
ified as an excellent substrate for nerve growth factoreceptor, is crucial for the regulation of cell proliferationnd differentiation (Pawson, 1995). The SH3 domain∗ Corresponding author. Tel.: +82 31 299 6134;ax: +82 31 299 6139.
E-mail address: [email protected] (J.-Y. Ahn).
357-2725/$ – see front matter © 2007 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocel.2007.05.020
ells; SH domain
has been implicated in the mitogenesis of PLC �1, butnot lipase activity of this protein (Huang et al., 1995;Smith et al., 1994; Ye et al., 2002). It was also recentlyreported that the SH3 domain of PLC �1 binds to theRas nucleotide exchange factor, Sos, thus augmentingRas activity (Kim et al., 2000), and also elicits the acti-vation of mitogen-activated protein kinase (MAPK) viaan interaction with the SH2 domain containing leukocyteprotein, SLP-76 (Yablonski, Kadlecek, & Weiss, 2001).Quite recently, we demonstrated that the SH3 domain isalso required for the activation of nuclear PI3K and cellproliferation (Ye et al., 2002).
Recently, there has been increased interest in therole of PLC �1 in cell differentiation. The results ofa number of studies have indicated that PLC �1 per-forms a function in certain cell differentiation events,

of Bioc
2084 T.L.X. Nguyen et al. / The International Journalincluding insulin-like growth factor (IGF)-induced mus-cle differentiation and vascular endothelial growth factor(VEGF)-induced endothelial cell differentiation (Meyer,Latz, & Rahimi, 2003). Moreover, Bae et al. (1998)reported PLC �1 inhibits NGF-induced differentiationof PC12 cells. These studies implied an important rolefor PLC �1 in response to specific growth factor and/ordifferent cell types.
To better understand the role of PLC �1 functionin neuronal differentiation, we used stable PC12 ratpheochromocytoma cell line, which is comparable toadrenal chromaffin cells of neural crest origin (Greene& Tishler, 1976), overexpressing wild-type (WT) ofthis gene and its deletion mutants. Nerve growth factor(NGF) promotes PC12 cells differentiation into a sympa-thetic neuron-like phenotype, and this effect is regulatedby the sustained activation of mitogen-activated pro-tein kinase (MAPK) (Gomez, Tonks, Morrison, Harmar,& Cohen, 1990; Traverse, Gomez, Paterson, Marshell,& Cohen, 1992), coincident with cell cycle arrest(Anderson & Axel, 1986; Unsicjker, Krisch, Otten, &Yhoenen, 1978). Although previous studies showed pos-itive effect of PLC �1 in H9c2 cardiac myoblasts (Honget al., 2001) or endothelial cell differentiation (Meyer etal., 2003), we found that PLC �1 overexpression ren-dered PC12 cells completely resistant to differentiationby NGF. Now, we report that differentiation inhibitionphenomenon is clearly regulated by SH3 domain of PLC�1. Further exploration of this phenomenon revealed thatPLC �1 can induce profound change in the expressionof cell cycle regulatory proteins and exit from cell cycleattest by NGF. Pharmacological inhibition of cell cycleprogression, arresting specifically at G2/M were recapit-ulate neuronal phenotype in WT-PLC �1 cells and cellswith deletion mutants of SH2 domain, suggesting themechanism of PLC �1 inhibitory action on the NGF-induced neuronal differentiation may involve cell cycleregulatory machinery and the escape from growth arrestby NGF through its SH3 domain.
2. Materials and methods
2.1. Cell cultures
PC12 cells were maintained in medium A (Dul-becco’s modified Eagle’s medium with 10% fetal bovineserum, 5% horse serum, and 100 units penicillin-streptomycin) at 37 ◦C, under a 5% CO atmosphere in
2a humidified incubator. The Flag-tagged PLC �1 stablytransfected PC12 cells (Tet-off cell line) were culturedin medium B (Dulbecco’s modified Eagle’s medium,10% horse serum, 5% fetal bovine serum, 100 �g/mlhemistry & Cell Biology 39 (2007) 2083–2092
G418, 100 �g/ml hygromycin B, 2 �g/ml tetracyclineand 100 units of penicillin-streptomycin). The PLC �1was induced in medium B without tetracycline for 24 h.
2.2. Reagents and antibodies
The mouse monoclonal anti-HA utilized in thisstudy was purchased from Sigma. Anti-Akt, anti-P44/42MAPK, anti-Phospho-Akt (Ser473), anti-Phospho-p44/42 MAPK and anti-CDK1 antibodies were obtainedfrom Cell Signaling. The anti-CDK2 and the anti-PCNAwere purchased from Abcam and Oncogene. Anti-cyclinD1 (DCS-6) and anti-GAP43 were acquired from SantaCruz. The anti-p21WAF1 and anti-tubulin were providedby CalBiochem and Molecular Probes. The cell pro-liferation assay kit was purchased from BoehringerMannheim. Chemicals used were 0.5 mM sodiumnitropruside (Sigma), 10 �M forskolin (CalBiochem),200 nM rapamycin (CalBiochem). All chemicals not ref-erenced above were obtained from Sigma.
2.3. Immunoblotting
The cells were rinsed and harvested with lysisbuffer (Tris 50 mM, pH 7.4, NaCl 40 mM, EDTA1 mM, Triton X-100 0.5%, Na3VO4 1.5 mM, NaF50 mM, sodium pyrophosphate 10 mM, glycerolphos-phate 10 mM, PMSF 1 mM, Protease inhibitor cocktail10 mM), vortexed, and centrifuged for 10 min at13,000 rpm at 4 ◦C. Aliquots (70 �g) of the pro-teins were analyzed via Western blotting using a1:1000 dilution of monoclonal anti-Akt, anti-Phospho-Akt, anti-p44/42 MAPK, anti-Phospho-p44/42 MAPK,anti-PCNA, anti-GAP43, anti-p21WAF1, anti-CDK1,anti-CDK2 and anti-Cyclin D1. The immunocomplexeswere visualized with enhanced chemiluminescencereagent (Amersham), in accordance with the manufac-turer’s instructions.
2.4. Assay for phospholipase activity
The induced and uninduced PLC �1 cells werelabeled with myo-[2-3H]inositol (2 �l Ci/ml; Perkin-Elmer Life Sciences, Boston, MA, USA) in inositol-freemedium for 24 h. The labeled cells were pretreated for15 min with 20 mM LiCl in serum-free DMEM contain-ing 1 mg/ml bovine serum albumin. The cells were thenstimulated with NGF for 10 min. The reaction was ter-
minated via the addition of 0.6 ml ice-cold 5% HClO4.After 30 min on ice, the extract was eluted through aBio-Rad Dowex AG 1-X8 anion exchange column. Totalinositol phosphate was eluted with a solution contain-
of Bioc
iq
2o
pwwimumifilV
2
wfil(ro(aw
2
pwptlatA
2
wsofa
To address how PLC �1 participates in the neuronaldifferentiation, we examined neurite outgrowth with orwithout tetracycline in WT-PLC �1 and its mutant cells,as well as control PC12 cells (Fig. 2A and Supplementary
Fig. 1. Schematic outline of the domain organization of PLC �1 con-structs and immunoblotting analysis of the expression of Flag-PLC
T.L.X. Nguyen et al. / The International Journal
ng 1 M ammonium formate and 0.1 M formic acid, anduantified via liquid scintillation counting.
.5. Phase-contrast microscopy and measurementsf neurite outgrowth
The cells (1 × 104) were seeded in 6-well culturelates. After 1 day of culture, the medium was exchangedith the induced medium. After 24 h, 50 ng/ml NGFas administered to the samples. The cells were then
ncubated for an additional 5 days. For phase-contrasticroscopy, the cells were photographed and scored
nder an Axiovert 100 microscope (Carl Zeiss, Ger-any). Neuritic processes longer than two cell bodies
n length were counted as neuritis and a minimum ofve random fields were photographed in each case. The
ength of the neurites was measured using the Axioision program.
.6. Flow cytometry assay
The cells were collected via trypsinization. Afterashing with PBS 1× (two times), 2 × 106 cells werexed in 70% ethanol at −20 ◦C, for at least 24 h. The pel-
ets were resuspended in 50 �l phosphate/citrate buffer0.2 M Na2HPO4, 0.1 M citrate, pH 7.5) for 30 min atoom temperature. The cells were then washed in 5 mlf PBS, and incubated with propidium iodide solution20 �g/ml pI, 20 �g/ml RNase A in 1× PBS) for 30 mint room temperature. The samples were then analyzedith the FacsCalibur system (Becton Dickinson, USA).
.7. Cell proliferation assay
The cells (1.0 × 103) were seeded in 96-well culturelates coated with 10 �g/ml poly-l-lysine, and treatedith 50 ng/ml of NGF in complete medium. The cellroliferation assay was conducted in accordance withhe manufacturer’s instructions. In brief, the cells wereabeled with bromodeoxyuridine (BrdU) for 4 h at 37 ◦C,nd then incubated with anti-BrdU-peroxidase. The reac-ions were quantified via detection of absorbance at450–A690, using an ELISA reader.
.8. Treatments with antiproliferative agents
WT-PLC �1, �SH2N and �SH22 cells were culturedith 100 ng/ml NGF or 100 ng/ml NGF with 0.5 mM
odium nitropruside (SNP), 10 �M forskolin (FOR),r 200 nM rapamycin (RAP) in the induced mediumor 7 days. The phase-contrast photographs and FSACnalysis were performed as mentioned above. For cell
hemistry & Cell Biology 39 (2007) 2083–2092 2085
proliferation assay, 2 × 104 cells were seeded in 6-wellculture plates. After treatments with chemicals, cellswere trypsinized and triplicate counted from trypan bluestaining.
3. Results
3.1. Response of PC12 cells overexpressing PLC γ1and its variant mutants to NGF exposure
To characterize the profound role of PLC �1 inNGF-mediated PC12 cell differentiation, we employeda Tet-off PC12 cell line that had been stably transfectedwith the inducible form of Flag-tagged PLC �1, whichharbored a variety of SH domain truncations. The struc-ture of PLC �1 and its mutants are depicted in Fig. 1A.The expression of the PLC �1 constructs was confirmedvia immunoblotting against anti-Flag antibody after 24 hof induction in culture medium with or without tetracy-cline. The endogenous PLC �1 was also detected againstanti-PLC �1 antibody (Fig. 1B).
�1 and its deletion mutants. (A) Constructs of various PLC �1. (B)Stably transfected PC12 cells were cultured in medium with or with-out tetracycline for 24 h, and 70 �g of protein was employed in 10%sodium dodecyl sulfate-polyacrylamide gel electrophoresis, followedby immunoblotting with anti-Flag antibody and anti-PLC �1 antibody.
2086 T.L.X. Nguyen et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 2083–2092
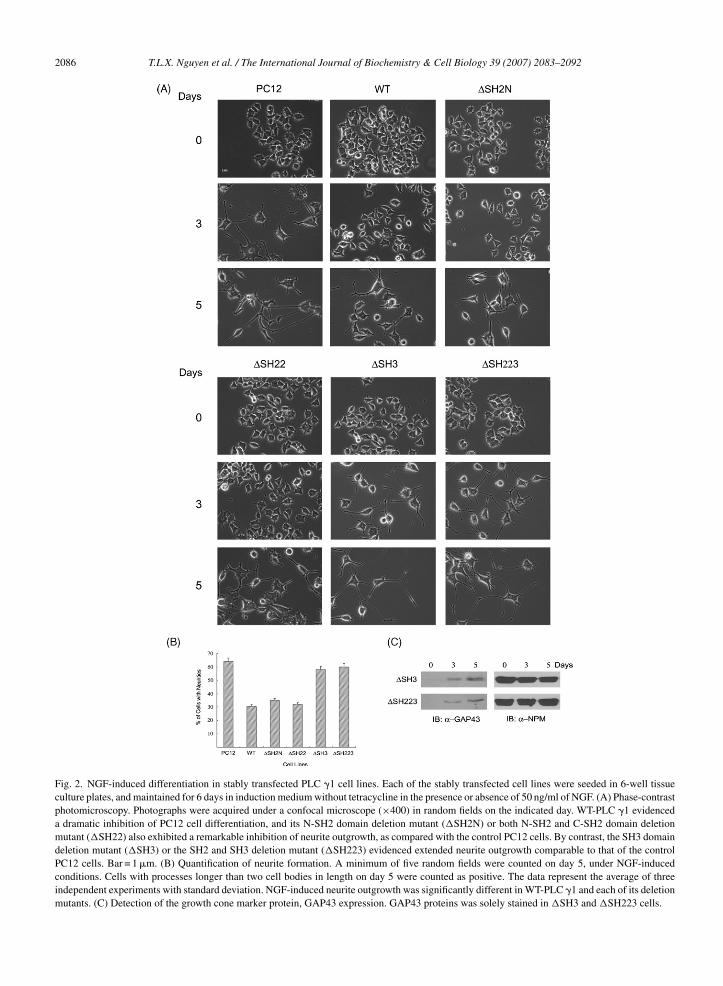
Fig. 2. NGF-induced differentiation in stably transfected PLC �1 cell lines. Each of the stably transfected cell lines were seeded in 6-well tissueculture plates, and maintained for 6 days in induction medium without tetracycline in the presence or absence of 50 ng/ml of NGF. (A) Phase-contrastphotomicroscopy. Photographs were acquired under a confocal microscope (×400) in random fields on the indicated day. WT-PLC �1 evidenceda dramatic inhibition of PC12 cell differentiation, and its N-SH2 domain deletion mutant (�SH2N) or both N-SH2 and C-SH2 domain deletionmutant (�SH22) also exhibited a remarkable inhibition of neurite outgrowth, as compared with the control PC12 cells. By contrast, the SH3 domaindeletion mutant (�SH3) or the SH2 and SH3 deletion mutant (�SH223) evidenced extended neurite outgrowth comparable to that of the controlPC12 cells. Bar = 1 �m. (B) Quantification of neurite formation. A minimum of five random fields were counted on day 5, under NGF-inducedconditions. Cells with processes longer than two cell bodies in length on day 5 were counted as positive. The data represent the average of threeindependent experiments with standard deviation. NGF-induced neurite outgrowth was significantly different in WT-PLC �1 and each of its deletionmutants. (C) Detection of the growth cone marker protein, GAP43 expression. GAP43 proteins was solely stained in �SH3 and �SH223 cells.
of Biochemistry & Cell Biology 39 (2007) 2083–2092 2087
FP7tIPr&HiiNincismtmd
3S
PfcfaidtsitSPBwtiTdt�
ic
pw
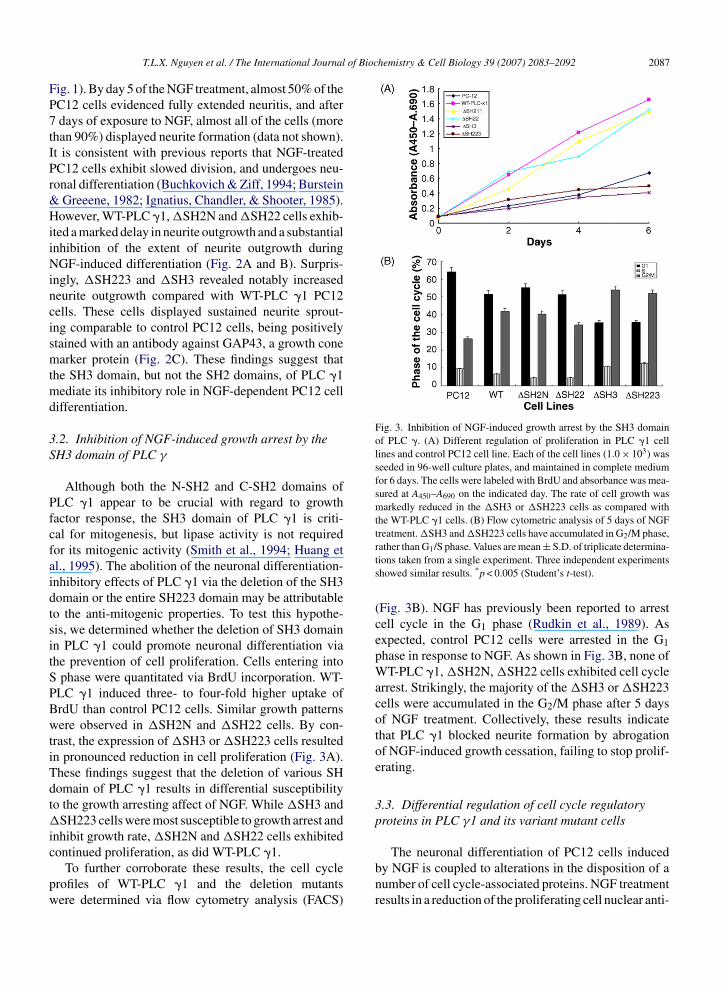
Fig. 3. Inhibition of NGF-induced growth arrest by the SH3 domainof PLC �. (A) Different regulation of proliferation in PLC �1 celllines and control PC12 cell line. Each of the cell lines (1.0 × 103) wasseeded in 96-well culture plates, and maintained in complete mediumfor 6 days. The cells were labeled with BrdU and absorbance was mea-sured at A450–A690 on the indicated day. The rate of cell growth wasmarkedly reduced in the �SH3 or �SH223 cells as compared withthe WT-PLC �1 cells. (B) Flow cytometric analysis of 5 days of NGFtreatment. �SH3 and �SH223 cells have accumulated in G2/M phase,
T.L.X. Nguyen et al. / The International Journal
ig. 1). By day 5 of the NGF treatment, almost 50% of theC12 cells evidenced fully extended neuritis, and afterdays of exposure to NGF, almost all of the cells (more
han 90%) displayed neurite formation (data not shown).t is consistent with previous reports that NGF-treatedC12 cells exhibit slowed division, and undergoes neu-onal differentiation (Buchkovich & Ziff, 1994; Burstein
Greeene, 1982; Ignatius, Chandler, & Shooter, 1985).owever, WT-PLC �1, �SH2N and �SH22 cells exhib-
ted a marked delay in neurite outgrowth and a substantialnhibition of the extent of neurite outgrowth duringGF-induced differentiation (Fig. 2A and B). Surpris-
ngly, �SH223 and �SH3 revealed notably increasedeurite outgrowth compared with WT-PLC �1 PC12ells. These cells displayed sustained neurite sprout-ng comparable to control PC12 cells, being positivelytained with an antibody against GAP43, a growth conearker protein (Fig. 2C). These findings suggest that
he SH3 domain, but not the SH2 domains, of PLC �1ediate its inhibitory role in NGF-dependent PC12 cell
ifferentiation.
.2. Inhibition of NGF-induced growth arrest by theH3 domain of PLC γ
Although both the N-SH2 and C-SH2 domains ofLC �1 appear to be crucial with regard to growthactor response, the SH3 domain of PLC �1 is criti-al for mitogenesis, but lipase activity is not requiredor its mitogenic activity (Smith et al., 1994; Huang etl., 1995). The abolition of the neuronal differentiation-nhibitory effects of PLC �1 via the deletion of the SH3omain or the entire SH223 domain may be attributableo the anti-mitogenic properties. To test this hypothe-is, we determined whether the deletion of SH3 domainn PLC �1 could promote neuronal differentiation viahe prevention of cell proliferation. Cells entering into
phase were quantitated via BrdU incorporation. WT-LC �1 induced three- to four-fold higher uptake ofrdU than control PC12 cells. Similar growth patternsere observed in �SH2N and �SH22 cells. By con-
rast, the expression of �SH3 or �SH223 cells resultedn pronounced reduction in cell proliferation (Fig. 3A).hese findings suggest that the deletion of various SHomain of PLC �1 results in differential susceptibilityo the growth arresting affect of NGF. While �SH3 and
SH223 cells were most susceptible to growth arrest andnhibit growth rate, �SH2N and �SH22 cells exhibited
ontinued proliferation, as did WT-PLC �1.To further corroborate these results, the cell cyclerofiles of WT-PLC �1 and the deletion mutantsere determined via flow cytometry analysis (FACS)
rather than G1/S phase. Values are mean ± S.D. of triplicate determina-tions taken from a single experiment. Three independent experimentsshowed similar results. *p < 0.005 (Student’s t-test).
(Fig. 3B). NGF has previously been reported to arrestcell cycle in the G1 phase (Rudkin et al., 1989). Asexpected, control PC12 cells were arrested in the G1phase in response to NGF. As shown in Fig. 3B, none ofWT-PLC �1, �SH2N, �SH22 cells exhibited cell cyclearrest. Strikingly, the majority of the �SH3 or �SH223cells were accumulated in the G2/M phase after 5 daysof NGF treatment. Collectively, these results indicatethat PLC �1 blocked neurite formation by abrogationof NGF-induced growth cessation, failing to stop prolif-erating.
3.3. Differential regulation of cell cycle regulatoryproteins in PLC γ1 and its variant mutant cells
The neuronal differentiation of PC12 cells inducedby NGF is coupled to alterations in the disposition of anumber of cell cycle-associated proteins. NGF treatmentresults in a reduction of the proliferating cell nuclear anti-
of Bioc
2088 T.L.X. Nguyen et al. / The International Journalgen (PCNA), a molecular marker of S phase cells (Bravo& Macdonald-Bravo, 1987). Conversely, cyclin D1, G1phase specific cyclin protein is greatly increased (Yan& Ziff, 1995). To characterize the correlation betweenNGF-mediated differentiation and the effects of PLC�1 and its mutants on cell cycle regulatory proteins,the expressions of cyclin D1 and PCNA were evaluatedvia immunoblotting (Fig. 4A, left and middle panels).Similar quantities of proteins were employed in the West-
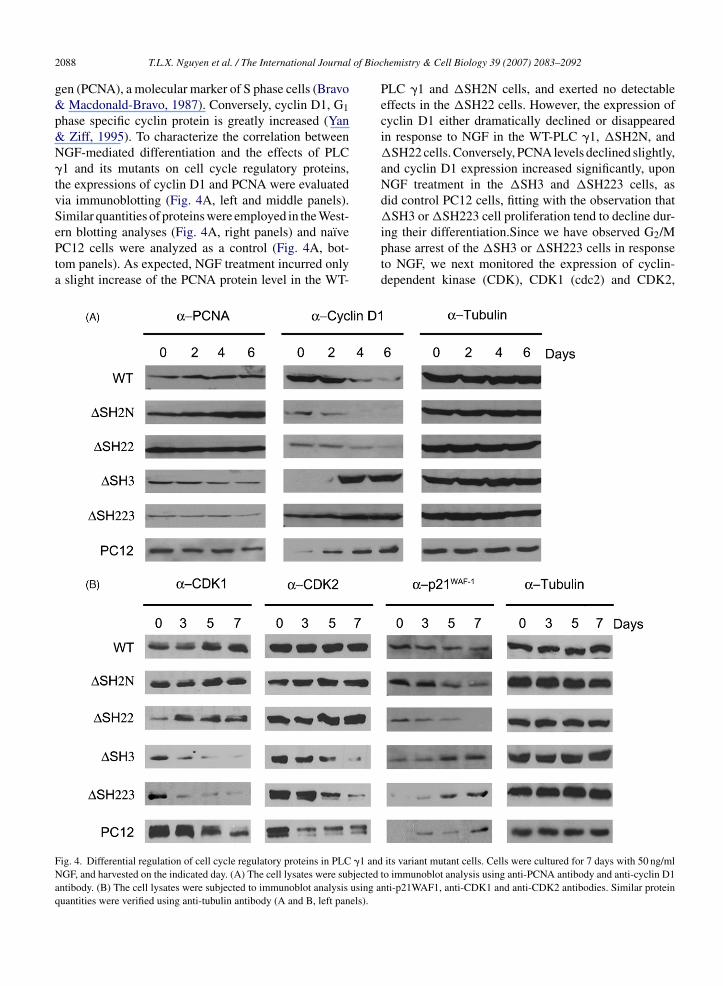
ern blotting analyses (Fig. 4A, right panels) and naı̈vePC12 cells were analyzed as a control (Fig. 4A, bot-tom panels). As expected, NGF treatment incurred onlya slight increase of the PCNA protein level in the WT-Fig. 4. Differential regulation of cell cycle regulatory proteins in PLC �1 andNGF, and harvested on the indicated day. (A) The cell lysates were subjectedantibody. (B) The cell lysates were subjected to immunoblot analysis using aquantities were verified using anti-tubulin antibody (A and B, left panels).
hemistry & Cell Biology 39 (2007) 2083–2092
PLC �1 and �SH2N cells, and exerted no detectableeffects in the �SH22 cells. However, the expression ofcyclin D1 either dramatically declined or disappearedin response to NGF in the WT-PLC �1, �SH2N, and�SH22 cells. Conversely, PCNA levels declined slightly,and cyclin D1 expression increased significantly, uponNGF treatment in the �SH3 and �SH223 cells, asdid control PC12 cells, fitting with the observation that�SH3 or �SH223 cell proliferation tend to decline dur-
ing their differentiation.Since we have observed G2/Mphase arrest of the �SH3 or �SH223 cells in responseto NGF, we next monitored the expression of cyclin-dependent kinase (CDK), CDK1 (cdc2) and CDK2,its variant mutant cells. Cells were cultured for 7 days with 50 ng/mlto immunoblot analysis using anti-PCNA antibody and anti-cyclin D1nti-p21WAF1, anti-CDK1 and anti-CDK2 antibodies. Similar protein

of Bioc
w&l(1ppAiPtiaoaciap
osp
3W
dfiaacNwpacEee1ctnmarWfn
T.L.X. Nguyen et al. / The International Journal
hich promotes cell cycle progression (Van den HeuvelHarlow, 1993), and p21WAF-1, which negatively regu-
ate CDK activity, arresting cell cycle at G1/S or G2/MBillon, van Grunsven, & Rudkin, 1996; Parker et al.,995). Previous studies showed that NGF represses therotein level of specific CDK, but increased the level of21WAF-1 (Buchkovich & Ziff, 1994; Yan & Ziff, 1995).s expected, NGF exposure caused a gradual decline
n the protein levels of CDK1 and CDK2 in controlC12 cells as well as �SH3 or �SH223 cells, correlating
heir neurite formation. In contrast, WT-PLC �1 exhib-ted a bit increase in CDK1 expression, but not muchffection in CDK2 expression level. Similar patternsf CDK1 and CDK2 levels were observed in �SH2Nnd �SH22 cells (Fig. 4B). NGF-treated control PC12ells and SH3 domain deleted mutant cells showed thencreased level of p21WAF-1, while WT-PLC �1, �SH2Nnd �SH22 cells showed a lower level expression of21WAF-1 (Fig. 4B).
Collectively, these results suggest that overexpressionf PLC �1 inhibits growth arrest and alters the expres-ion level of cell cycle proteins towards a less neuronalhenotype in PC12 cells through its SH3 domain.
.4. Regaining the ability to differentiation inT-PLC γ1 cells by antiproliferative agents
Because growth arrest is an essential for PC12 cellifferentiation, it is conceivable that the inability to dif-erentiate of WT-PLC �1, �SH2N, and �SH22 cellss responsible for escape from NGF-induced growthrrest. To address this issue whether forced growthrrest in these WT-PLC �1, �SH2N, and �SH22 cellsould regain the ability to differentiate in response toGF, we employed various antiproliferative reagents,hich induce cell growth arrest, such as sodium nitro-russide (SNP), forskolin, rapamycin. A nitric oxidegonist, SNP urges the expression of p21WAF-1 andause accumulation of cells in G2 phase (Peunova &nikolopov, 1995; Poluha et al., 1997). Forskolin, cAMPlevating agent, induce growth arrest inhibiting thexpression of CDK2 and CDK4 (Ammer & Schulz,997; Mark & Storm, 1997). Rapamycin causes growthessation in G1 phase by inhibiting p70S6 kinase. NGF-reated WT-PLC �1, �SH2N, and �SH22 cells showedo significant neurite outgrowth (Fig. 2A) and treat-ent of WT-PLC �1, �SH2N, and �SH22 cells with
ntiproloferative reagents alone did not stimulate neu-
ite formation (data not shown). However, treatment ofT-PLC �1, �SH2N, and �SH22 cells with SNP andorskolin during NGF-stimulation exhibited a markedeurite outgrowth of and obvious neurite extension of
hemistry & Cell Biology 39 (2007) 2083–2092 2089
PC12 cells (Fig. 5A and B). In contrast, rapamycin treat-ment in these cells was only able to force mere neuriteoutgrowth. Consistent with this observation, treatmentof SNP and forskolin with NGF, but not rapamycindramatically declined cell proliferation (Fig. 5C). Fur-thermore, FACS analysis revealed SNP and forskolingave rise to G2/M phase accumulation of these cells,whereas rapamycin elicited negligible change in thecell cycle distribution compared to control PC12 cells(Fig. 5D). Moreover, we also detected GAP43 expres-sion and inhibition of CDK1 expression in SNP andforskolin co-treated cells, implying the forced extendedneurite are structurally normal and SNP and forskolindrove cells into G2/M phase, as demonstrated by theFACS analysis (Fig. 5E). Different stable clones displaysimilar response upon NGF treatment or co-treatment ofSNP, forskolin, or rapamycin with NGF (data not sown).These observations suggest that WT-PLC �1, �SH2N,and �SH22 cells restore the ability to differentiate bymeans of which it can incur growth cessation, especiallyG2/M phase rather than G1/S phase. It also suggests thatthe inhibition of neurite outgrowth by overexpressionof WT-PLC �1, �SH2N-PLC �1, and �SH22-PLC �1might be caused to the exit from NGF-elicited growtharrest.
4. Discussion
Previous studies have demonstrated that SH domainsof PLC �1 inhibit NGF-induced differentiation of PC12cells. Here, we provide direct evidence that SH3 domain,but not SH2 domains, is essential for PLC �1’s inabilityto differentiate and this inability of neurite outgrowth ofPLC �1 is able to reconstitute if these cells are forced toarrest some stage of cell cycle. Although it is possible thatthere is the variation of our clones that stably transfectedwith WT-PLC �1 and its variant mutant constructs, thedegree of differentiation in each cell lines were similaramong different stable clones (data not shown).
We and others have previously revealed that SH3domain of PLC �1 is indispensable for cell proliferationand mediates a variety of signaling cascades includingMAPK and PI3K (Huang et al., 1995; Smith et al., 1994;Yablonski et al., 2001; Ye et al., 2002). Although com-pelling evidence exists to suggest that the differentiationof PC12 cells is regulated by sustained MAPK activation,we could not detect much difference of MAPK (Erk1 andErk2) activation among WT-PLC �1 and its mutant cells
(Supplementary Fig. 2A). Only lipase inactive mutantcell diminished MAPK activation (data not shown), aspreviously reported by us (Rong, Ahn, Chen, Suh, &Ye, 2003). Nevertheless Akt activation remained intact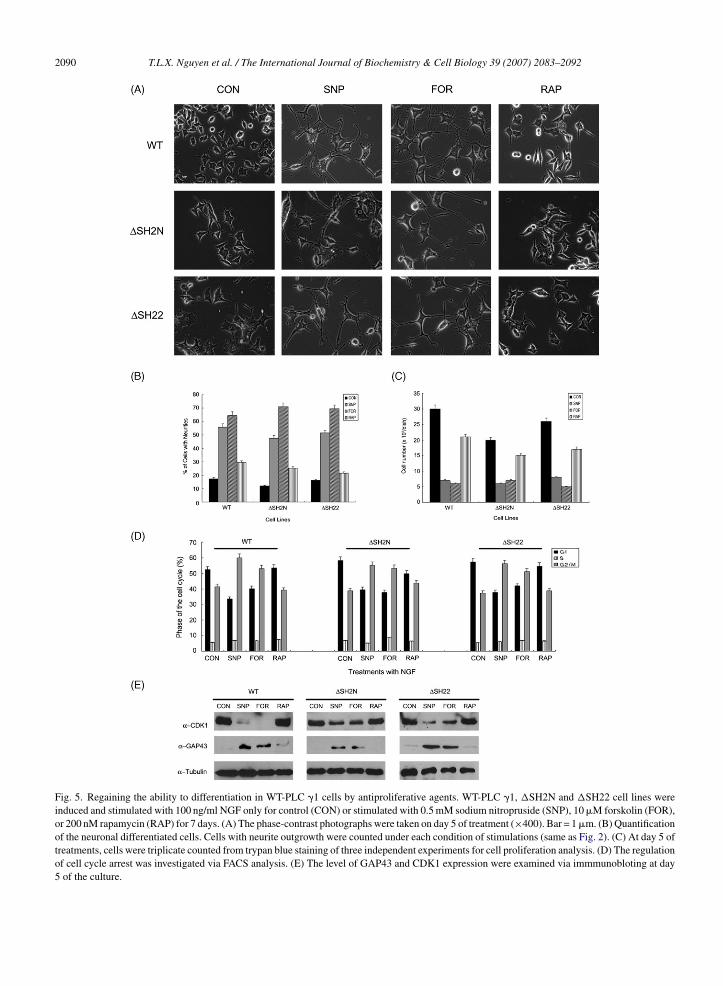
2090 T.L.X. Nguyen et al. / The International Journal of Biochemistry & Cell Biology 39 (2007) 2083–2092
Fig. 5. Regaining the ability to differentiation in WT-PLC �1 cells by antiproliferative agents. WT-PLC �1, �SH2N and �SH22 cell lines wereinduced and stimulated with 100 ng/ml NGF only for control (CON) or stimulated with 0.5 mM sodium nitropruside (SNP), 10 �M forskolin (FOR),or 200 nM rapamycin (RAP) for 7 days. (A) The phase-contrast photographs were taken on day 5 of treatment (×400). Bar = 1 �m. (B) Quantificationof the neuronal differentiated cells. Cells with neurite outgrowth were counted under each condition of stimulations (same as Fig. 2). (C) At day 5 oftreatments, cells were triplicate counted from trypan blue staining of three independent experiments for cell proliferation analysis. (D) The regulationof cell cycle arrest was investigated via FACS analysis. (E) The level of GAP43 and CDK1 expression were examined via immmunobloting at day5 of the culture.

of Bioc
ifiPdidptcatri1roNitaNdaacdcscptcc�
tssalrtawctmgIrwt
T.L.X. Nguyen et al. / The International Journal
n all PLC �1 cell lines (Supplementary Fig. 2B). Thisnding suggests that the inability to differentiate of WT-LC �1, �SH2N, and �SH22 cells are not caused by theefect of MAPK activation or Akt activation, the majorntracellular signaling pathways involved in neuronalifferentiation and proliferation.We hypothesize that therolonged proliferate activity of PLC �1 may contributeo this inhibitory role in neuronal differentiation becauseells must stop dividing before differentiation. Growthrrest during neuronal differentiation may be due tohe alteration of important proteins level for cell cycleegulation. NGF-induced differentiation of PC12 cellsncreased cyclin D1 inhibiting PCNA level (Yan & Ziff,995). �SH3 or �SH223 that showed normal neu-onal differentiation revealed similar expression patternf cyclin D1 and PCNA as control PC12 cells underGF-stimulation. Conversely, cyclin D1 was declined
n WT-PLC �1, �SH2N, and �SH22 cells, correlatinghe observation that these cells tend to continue prolifer-ting, thereby fail to format neurite outgrowth (Fig. 4A).GF also induces a host of other changes, including aecrease in CDK kinases and elevating p21WAF-1. Welso observed a down regulation of CDK1 and CDK2,nd gradual induction of p21WAF-1 in �SH3 or �SH223ells same as control PC12 cells. Although, we couldetect no obvious change of the level of CDK2, weonfirmed a gradual elevation of CDK1 and a depres-ion of p21WAF-1 in WT-PLC �1, �SH2N, and �SH22ells (Fig. 4B). Since our FACS analysis exhibited G2/Mhase arrest of differentiated �SH3 or �SH223 cells,his may explain why SH3 deletion mutants inhibit theell growth. Similarly, cyclin B1, which has been impli-ated in G2/M phase transition, was down regulated inSH3 or �SH223 cells (data not shown).Overexpression of WT-PLC �1 and its various dele-
ion mutants differentially modulates the expression ofeveral key regulatory proteins in cell cycle progres-ion due to their ability to differentiate. Once growthrrest is compelled following the treatment of antipro-iferative reagent, such as SNP and forskolin, but notapamycin in WT-PLC �1, �SH2N, and �SH22 cells,hese cells displayed reconstitution of neurite formationnd abrogation of cell proliferation (Fig. 5). Althoughe cannot rule out the possibility that these reagents
ould invoke diverse effect on cells, the ability thathey produced inhibition of cell growth under different
echanism argue that the recurrence of neurtie out-rowth caused by growth arrest, specifically G /M phase.
2t remains unclear why not all of the antiproliferativeeagents, but only those that caused G2/M phase arrestingere invoked WT-PLC �1, �SH2N, and �SH22 cellso reproduce neurite outgrowth under NGF treatment.
hemistry & Cell Biology 39 (2007) 2083–2092 2091
Interestingly, a lipase inactive mutant (LIM) of PLC�1 (His335 to Gln335) does not alter the differentiation-inhibitory effects of PLC �1, as the removal of SH3domain, although these mutants yield notable lipaseactivity; result in the significant neurite outgrowth(Supplementary Fig. 3). In contrast, �SH2N and �SH22cells, which show the loss of lipase activity, block neu-ronal differentiation comparable to WT-PLC �1, whichhas intact lipase activity, implying the enzymatic activ-ity of this protein has seemed not primarily relevant toneurite outgrowth in PC12 cells. Despite LIM-PLC �1contain whole SH domains and the enzymatic activity ofthis protein has not been implicated in the mitogenesis,somehow this lipase inactive point mutant bared normalneurite outgrowth.
In summary, we have determined that overexpres-sion of PLC �1 inhibits NGF-induced differentiation viaits SH3 domain, which is essential for mitogenic activ-ity of PLC �1, modulating the expression of cell cycleregulatory proteins and antiproliferative reagent recurneuronal differentiation in WT-PLC �1, �SH2N, and�SH22 cells. The temporal correlation between cessa-tion of proliferation occurred in SH3 domain deletionmutants, depression of regulatory protein, and neuronaldifferentiation suggest that the antiproliferative activityof SH3 deletion contributes to this phenomenon.
Acknowledgements
This work was supported by grants from the KoreaResearch Foundation (KRF-2005-204-E00015) and theKorea Science and Engineering Foundation (KOSEF)grant by the Korea government (MOST) (RO1-2006-000-10222-0). We thank Dr. Pann-Ghill Suh for hisgenerous gift of Flag-tagged WT-PLC �1 PC12 cells.
Appendix A. Supplementary data
Supplementary data associated with this arti-cle can be found, in the online version, atdoi:10.1016/j.biocel.2007.05.020.
References
Ammer, H., & Schulz, R. (1997). Regulation of stimulatory adeny-lyl cyclase signaling during forskolin-induced differentiation ofmouse neuroblastoma and rat glioma (NG108-15) cells. Neuro-science Letter, 230, 143–146.
Anderson, D. J., & Axel, R. (1986). A bipotential neuroendocrineprecursor whose choice of cell fate is determined by NGF andglucocorticoids. Cell, 47, 1079–1090.
Bae, S. S., Lee, Y. H., Chang, J. S., Galadari, S. H., Kim, Y. S.,Ryu, S. H., et al. (1998). Src homology domains of phospholipase

of Bioc
2092 T.L.X. Nguyen et al. / The International JournalC gamma1 inhibit nerve growth factor-induced differentiation ofPC12 cells. The Journal of Neurochemistry, 71, 178–185.
Billon, N., van Grunsven, L. A., & Rudkin, B. B. (1996). The CDKinhibitor p21WAF1/Cip1 is induced through a p300-dependentmechanism during NGF-mediated neuronal differentiation ofPC12 cells. Oncogene, 13, 2047–2054.
Bravo, R., & Macdonald-Bravo, H. (1987). Existence of two popu-lations of cyclin/proliferating cell nuclear antigen during the cellcycle: Association with DNA replication sites. The Journal of CellBiology, 105, 1549–1554.
Buchkovich, K. J., & Ziff, E. B. (1994). Nerve growth factor regulatesthe expression and activity of p33cdk2 and p34cdc2 kinases inPC12 pheochromocytoma cells. Molecular Biology of the Cell, 5,1225–1241.
Burstein, D. E., & Greeene, L. A. (1982). Nerve growth factor has bothmitogenic and antimitogenic activity. Developmental Biology, 94,477–482.
Decker, S. J. (1995). Nerve growth factor-induced growth arrest andinduction of p21Cip1/WAF1 in NIH-3T3 cells expressing TrkA.The Journal of Biological Chemistry, 270, 30841–30844.
Gomez, N., Tonks, N. K., Morrison, C., Harmar, T., & Cohen, P.(1990). Evidence for communication between nerve growth factorand protein tyrosine phosphorylation. FEBS Letter, 271, 119–122.
Gotoh, Y., Nishida, E., Yamashita, T., Hoshi, M., Kawakami, M., &Sakai, H. (1990). Microtubule-associated-protein (MAP) kinaseactivated by nerve growth factor and epidermal growth factorin PC12 cells. Identity with the mitogen-activated MAP kinaseof fibroblastic cells. European Journal of Biochemistry, 193,661–669.
Greene, L. A., & Tischler, A. S. (1976). Establishment of a nora-drenergic clonal line of rat adrennal pheochromocytoma cellswhich respond to nerve growth factor. Proceedings of the NationalAcademy of Sciences, 73, 2424–2428.
Hong, F., Moon, K., Kim, S. S., Kim, Y. S., Choi, Y. K., Bae, Y.S., et al. (2001). Role of phospholipase C-gamma1 in insulin-likegrowth factor I-induced muscle differentiation of H9c2 cardiacmyoblasts. Biochemical and Biophysical Research Communica-tion, 282, 816–822.
Huang, P. S., Davis, L., Huber, H., Goodhart, P. J., Wegrzyn, R. E.,Oliff, A., et al. (1995). An SH3 domain is required for the mitogenicactivity of microinjected phospholipase C-gamma 1. FEBS Letter,358, 287–292.
Ignatius, M. J., Chandler, C. R., & Shooter, E. M. (1985). Nerve growthfactor-treated, neurite-bearing PC12 cells continue to synthesizeDNA. The Journal of Neuroscience, 5, 343–351.
Kim, B. Y., Kang, D. O., Oh, W. K., Kim, J. H., Choi, Y. K., Jang, J. S., etal. (2000). Involvement of SH2-SH2-SH3 domain of phospholipasec gamma 1 in NF-kappaB signaling. FEBS Letter, 472, 45–49.
Mark, M. D., & Storm, D. R. (1997). Coupling of epidermal growth
factor (EGF) with the antiproliferative activity of cAMP inducesneuronal differentiation. The Journal of Biological Chemistry, 272,17238–17244.Meyer, R. D., Latz, C., & Rahimi, N. (2003). Recruitment and acti-vation of phospholipase Cgamma1 by vascular endothelial growth
hemistry & Cell Biology 39 (2007) 2083–2092
factor receptor-2 are required for tubulogenesis and differentiationof endothelial cells. The Journal of Biological Chemistry, 278,16347–16355.
Parker, S. B., Eichele, G., Zhang, P., Rawls, A., Sands, A. T., Bradley,A., et al. (1995). p53-Independent expression of p21Cip1 in muscleand other terminally differentiating cells. Science, 267, 1024–1027.
Pawson, T. (1995). Protein-tyrosine kinases. Getting down to specifics.Nature, 373, 477–478.
Peunova, N., & Enikolopov, G. (1995). Nitric oxide triggers a switchto growth arrest during differentiation of neuronal cells. Nature,375, 68–73.
Poluha, W., Schonhoff, C. M., Harrington, K. S., Lachyankar, M. B.,Crosbie, N. E., Bulseco, D. A., et al. (1997). A novel, nerve growthfactor-activated pathway involving nitric oxide, p53, and p21WAF1regulates neuronal differentiation of PC12 cells. The Journal ofBiological Chemistry, 272, 24002–24007.
Rong, R., Ahn, J. Y., Chen, P., Suh, P. G., & Ye, K. (2003). Phos-pholipase activity of phospholipase C-gamma1 is required fornerve growth factor-regulated MAP kinase signaling cascade inPC12 cells. The Journal of Biological Chemistry, 278, 52497–52503.
Rudkin, B. B., Lazarovici, P., Levi, B. Z., Abe, Y., Fujita, K., & Guroff,G. (1989). Cell cycle-specific action of nerve growth factor in PC12cells: Differentiation without proliferation. The EMBO Journal, 8,3319–3325.
Sherr, C. J. (1994). G1 phase progression: Cycling on cue. Cell, 79,551–555.
Smith, M. R., Liu, Y. L., Matthews, N. T., Rhee, S. G., Sung, W.K., & Kung, H. F. (1994). Phospholipase C-gamma 1 can induceDNA synthesis by a mechanism independent of its lipase activity.Proceedings of the National Academy of Science, 91, 6554–6558.
Traverse, S., Gomez, N., Paterson, H., Marshell, C., & Cohen,P. (1992). Sustained activation of the mitogen-activated protein(MAP) kinase cascade may be required for differentiation of PC12cells. Comparison of the effects of nerve growth factor and epider-mal growth factor. The Biochemistry Journal, 288, 351–355.
Unsicjker, K., Krisch, B., Otten, U., & Yhoenen, H. (1978). Nervegrowth factor-induced fiber outgrowth from isolated rat adrenalchromaffin cells: Impairment by glucocorticoids. Proceedings ofthe National Academy of Sciences, 75, 3498–3502.
Van den Heuvel, S., & Harlow, E. (1993). Distinct roles for cyclin-dependent kinases in cell cycle control. Science, 262, 2050–2054.
Yablonski, D., Kadlecek, T., & Weiss, A. (2001). Identification of aphospholipase C-gamma1 (PLC-gamma1) SH3 domain-bindingsite in SLP-76 required for T-cell receptor-mediated activationof PLC-gamma1 and NFAT. Molecular and Cellular Biology, 21,4208–4218.
Yan, G. Z., & Ziff, E. B. (1995). NGF regulates the PC12 cell cyclemachinery through specific inhibition of the Cdk kinases and induc-
tion of cyclin D1. The Journal of Neuroscience, 15, 6200–6212.Ye, K., Aghdasi, B., Luo, H. R., Moriarity, J. L., Wu, F. Y., Hong, J. J.,et al. (2002). Phospholipase C gamma 1 is a physiological guaninenucleotide exchange factor for the nuclear GTPase PIKE. Nature,415, 541–544.
Related Documents











