OELANDIELLA , THE EARLIEST CAMBRIAN HELCIONELLOID MOLLUSC FROM SIBERIA by ALEXANDER P. GUBANOV and JOHN S. PEEL ABSTRACT. The helcionelloid molluscs Oelandiella korobkovi and O. sibirica Vostokova are redescribed on the basis of type material from the lowest Cambrian of northern Siberia. Oelandiella is reinstated as a recognizable genus separated from Latouchella Cobbold, of which it has been considered generally to be a junior synonym, by differences in the prominent comarginal plication. The bilaterally symmetrical shells of both of these genera are readily delimited from Oelandia Westerga ˚rd, originally described from the Middle Cambrian of Sweden, which has a characteristic asymmetrical alternation of plicae. H ELCIONELLOIDS are univalved molluscs which are conspicuous in Lower and Middle Cambrian fossil assemblages as part of the first major diversification of the Mollusca. They were interpreted as members of the untorted exogastric class Monoplacophora by Runnegar and Pojeta (1974; see Runnegar 1996 for summary of later references) but Peel (1991a, 1991b) proposed a new class Helcionelloida, members of which were considered to be untorted but endogastric (see also Geyer 1994). In the latter class, the apex lies posteriorly, whilst it is anterior in the exogastric Monoplacophora (partially redefined as class Tergomya by Peel 1991a, 1991b). Helcionelloids are present in the earliest shelly faunas of northern Siberia (Text-fig. 1) where they are represented by small, planispirally coiled shells of similar proportions to Latouchella Cobbold, 1921 (see also Gubanov and Peel 1998). Indeed, these tiny shells (length 1–3 mm) are often referred to Cobbold’s genus which was originally described from the Lower Cambrian of Shropshire (e.g. Rozanov et al. 1969; Brasier 1989). The Siberian fossils were first described, however, under the names Oelandiella korobkovi and O. sibirica by Vostokova (1962) who noted the great similarity between her new genus and Oelandia Westerga ˚rd, 1936, originally described from the Middle Cambrian of Sweden (see also Knight 1941; Peel and Yochelson 1987). Missarzhevsky (in Rozanov et al. 1969; translated into English as Raaben 1981) changed the generic assignment of Vostokova’s species to Latouchella, without explanation, and this synonymy has been accepted by subsequent authors (although only tentatively by Rozanov and Zhuravlev 1992). Moreover, Knight et al. (1960) regarded Oelandia as a junior synonym of Latouchella, illustrating the type species of the former as a representative example of the latter. Again, this practice as been followed by most subsequent authors (see Peel and Yochelson 1987). Nevertheless, the differences between the genera are clear and Latouchella, Oelandia and Oelandiella are readily separated from each other by differences in the pattern of their prominent transverse plications (Text-fig. 2). In this paper we redescribe Oelandiella on the basis of Vostokova’s (1962) original material from the Kotuj River of north Siberia, clarifying the previously uncertain relationship to Oelandia and Latouchella, as exemplified by recent redescriptions of their type materials by Peel and Yochelson (1987) and Gubanov and Peel (1998). Whilst Vostokova’s (1962) species have often been reported by later authors (e.g. Brasier 1989; Esakova and Zhegallo 1996), the type specimens do not appear to have been described or illustrated since her original publication. STRATIGRAPHICAL SETTING In the original description, Vostokova (1962) indicated that all molluscs assigned to her new genus Oelandiella occurred in the Aldanian Stage of the Lower Cambrian of the Anabar anticline (basins of [Palaeontology, Vol. 42, Part 2, 1999, pp. 211–222] q The Palaeontological Association

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

O E L A N D I E L L A, T H E E A R L I E S T C A M B R I A N
H E L C I O N E L L O I D M O L L U S C F R O M S I B E R I A
by A L E X A N D E R P . G U B A N O V and J O H N S . P E E L
ABSTRACT. The helcionelloid molluscsOelandiella korobkoviandO. sibirica Vostokova are redescribed on the basisof type material from the lowest Cambrian of northern Siberia.Oelandiella is reinstated as a recognizable genusseparated fromLatouchellaCobbold, of which it has been considered generally to be a junior synonym, by differencesin the prominent comarginal plication. The bilaterally symmetrical shells of both of these genera are readily delimitedfrom OelandiaWesterga˚rd, originally described from the Middle Cambrian of Sweden, which has a characteristicasymmetrical alternation of plicae.
H E L C I O N E L L O I D S are univalved molluscs which are conspicuous in Lower and Middle Cambrianfossil assemblages as part of the first major diversification of the Mollusca. They were interpreted asmembers of the untorted exogastric class Monoplacophora by Runnegar and Pojeta (1974; see Runnegar1996 for summary of later references) but Peel (1991a, 1991b) proposed a new class Helcionelloida,members of which were considered to be untorted but endogastric (see also Geyer 1994). In the latter class,the apex lies posteriorly, whilst it is anterior in the exogastric Monoplacophora (partially redefined as classTergomya by Peel 1991a, 1991b).
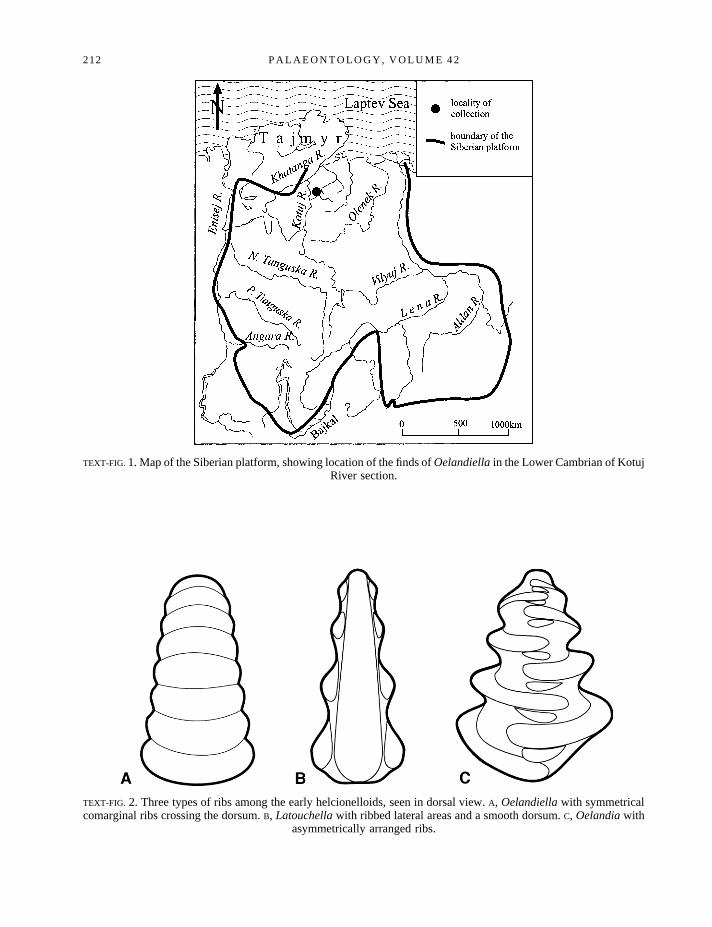
Helcionelloids are present in the earliest shelly faunas of northern Siberia (Text-fig. 1) where they arerepresented by small, planispirally coiled shells of similar proportions toLatouchellaCobbold, 1921 (seealso Gubanov and Peel 1998). Indeed, these tiny shells (length 1–3 mm) are often referred to Cobbold’sgenus which was originally described from the Lower Cambrian of Shropshire (e.g. Rozanovet al. 1969;Brasier 1989). The Siberian fossils were first described, however, under the namesOelandiella korobkoviandO. sibiricaby Vostokova (1962) who noted the great similarity between her new genus andOelandiaWesterga˚rd, 1936, originally described from the Middle Cambrian of Sweden (see also Knight 1941; Peeland Yochelson 1987). Missarzhevsky (in Rozanovet al. 1969; translated into English as Raaben 1981)changed the generic assignment of Vostokova’s species toLatouchella, without explanation, and thissynonymy has been accepted by subsequent authors (although only tentatively by Rozanov and Zhuravlev1992). Moreover, Knightet al. (1960) regardedOelandiaas a junior synonym ofLatouchella, illustratingthe type species of the former as a representative example of the latter. Again, this practice as beenfollowed by most subsequent authors (see Peel and Yochelson 1987). Nevertheless, the differencesbetween the genera are clear andLatouchella, OelandiaandOelandiellaare readily separated from eachother by differences in the pattern of their prominent transverse plications (Text-fig. 2).
In this paper we redescribeOelandiellaon the basis of Vostokova’s (1962) original material from theKotuj River of north Siberia, clarifying the previously uncertain relationship toOelandiaandLatouchella,as exemplified by recent redescriptions of their type materials by Peel and Yochelson (1987) and Gubanovand Peel (1998). Whilst Vostokova’s (1962) species have often been reported by later authors (e.g. Brasier1989; Esakova and Zhegallo 1996), the type specimens do not appear to have been described or illustratedsince her original publication.
S T R A T I G R A P H I C A L S E T T I N G
In the original description, Vostokova (1962) indicated that all molluscs assigned to her new genusOelandiellaoccurred in the Aldanian Stage of the Lower Cambrian of the Anabar anticline (basins of
[Palaeontology, Vol. 42, Part 2, 1999, pp. 211–222] q The Palaeontological Association
212 P A L A E O N T O L O G Y , V O L U M E 4 2
TEXT-FIG. 1. Map of the Siberian platform, showing location of the finds ofOelandiellain the Lower Cambrian of KotujRiver section.
TEXT-FIG. 2. Three types of ribs among the early helcionelloids, seen in dorsal view.A, Oelandiellawith symmetricalcomarginal ribs crossing the dorsum.B, Latouchellawith ribbed lateral areas and a smooth dorsum.C, Oelandiawith
asymmetrically arranged ribs.
G UB A N OV A N D P E E L : C AM B R I A N H E L C I O N E L L O I D 213
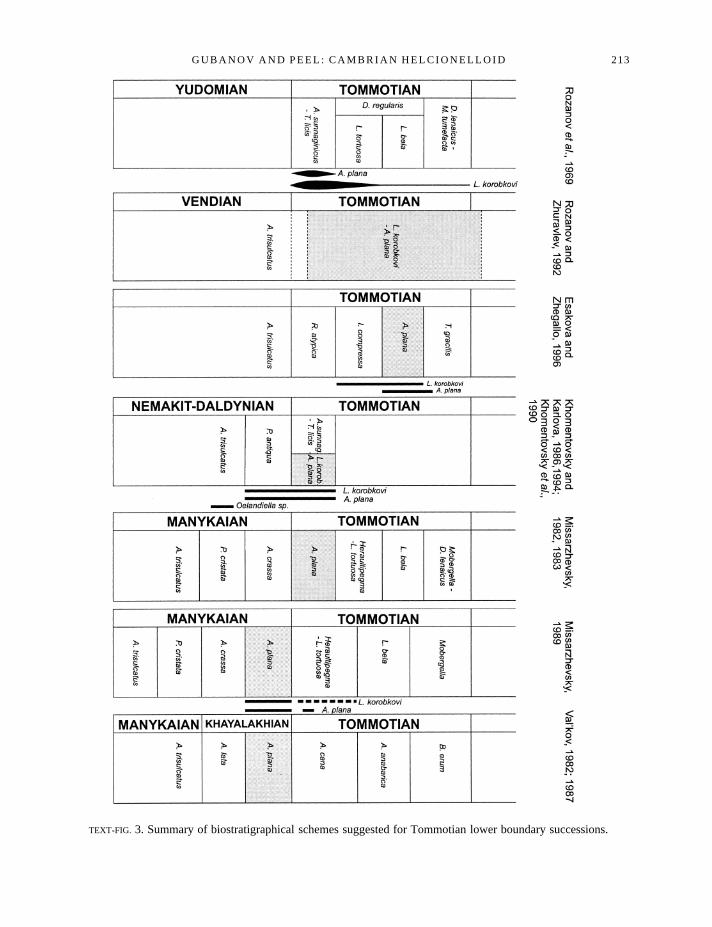
TEXT-FIG. 3. Summary of biostratigraphical schemes suggested for Tommotian lower boundary successions.

Kotuj and Popigaj rivers). All described specimens were collected during 1956–61 and, on the basis ofVostokova’s determinations, Savitsky (1962) established anOelandiella korobkovi-Anabarella planaBiozone as the basal part of the Aldanian Stage. Later, the Aldanian was divided largely into theTommotian and the Atdabanian stages, whilst the uppermost part was placed in the Botomian Stage (seeRepina and Rozanov 1984). Three different interpretations of the age of theOelandiella korobkovi-Anabarella planaBiozone exist. The first is that it is of early Tommotian age (Rozanovet al. 1969;Meshkovaet al. 1973; Zhuravlevaet al. 1979; Missarzhevsky 1980, 1983; Khomentovsky and Karlova1986). In contrast, Savitsky (1962), Val’kov (1975, 1982, 1987) and Missarzhevsky (1989;AnabarellaplanaBiozone in two last cases) suggested a pre-Tommotian age for this biozone. Voroninet al. (1982),Voronova et al. (1983) and Rozanov and Zhuravlev (1992) extended theOelandiella korobkovi-Anabarella planaBiozone to include the entire Tommotian, and even the lowermost Atdabanian. Thevariation in these age assignments has several causes, reflecting differences of interpretation: (1) incorrelation of fossil-bearing sections within Siberian platform succession, (2) in the range of theTommotian stage, and (3) in the range of this biozone (see Text-fig. 3).
At present it is difficult to estimate the stratigraphical position of the present samples more preciselythan Tommotian. However, Vostokova did not use acid processing as a method for obtaining small shellyfossils from her samples. When taking into account the fact that abundantOelandiellaoccur mostly nearthe base, the age of the redescribed specimens can probably be estimated as early Tommotian.
S Y S T E M A T I C P A L A E O N T O L O G Y
ClassHELCIONELLOIDA Peel, 1991aOrderHELCIONELLIDA Geyer, 1994
Family HELCIONELLIDAE Wenz, 1938
GenusOELANDIELLA Vostokova, 1962
Type species. Oelandiella korobkoviVostokova, 1962, from the Lower Cambrian of north Siberia.
Diagnosis. Small, univalve mollusc with tightly coiled, rapidly expanding shell of about one whorl, withprominent transverse ribs which cross the dorsum.
Oelandiella korobkoviVostokova, 1962
Text-figures 4–5, 6A–D, 7
1962 Oelandiella korobkoviVostokova, p. 52, pl. 1, figs 1–4.
Holotype. N8361–1, from the Lower Cambrian of the Kotuj River, north Siberia.
Material. Three specimens in addition to the holotype. A fourth, the best preserved specimen, was photographed(Vostokova 1962, pl. 1, fig. 1) and then destroyed during further preparation, as noted in the original description. Onlyone specimen (N8361–2; Vostokova 1962, pl. 1, fig. 4a-b) remains. Recently, from the same piece of rock, one morewell preserved specimen has been extracted.
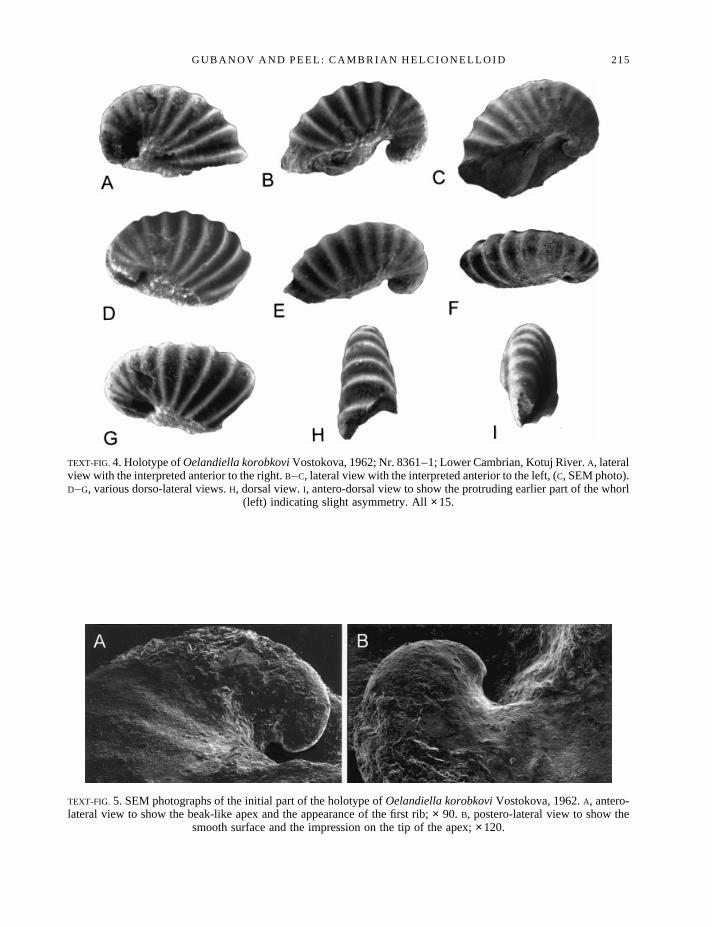
Description. The holotype (Text-fig. 4) is a small, laterally compressed univalve mollusc represented by an internalmould, with a maximum length ofc. 2·5 mm and width ofc. 1 mm; it is tightly coiled through approximately onewhorl. The initial part of the shell has a beak-like apex with a smooth surface (Text-fig. 5). The first rib appears afterone-third of the whorl. The aperture is oval and elongate with length:width of 2:1. The whorl has 13 prominent roundedribs which cross the dorsum and disappear towards the umbilici. The ribs are straight when viewed laterally andslightly curved within the sub-umbilical area towards the initial part of the mould.
Specimen N8361–2 (Text fig. 6A–D) is an internal mould tightly coiled through slightly more than one whorl(maximum length 3 mm and width of 1·2 mm). The oval aperture has a length:width ratio of 1·9:1. The well preservedlast quarter whorl bears about 14 tightly grouped ribs. The ribs cross the dorsum where they have a faintU-shaped
214 P A L A E O N T O L O G Y , V O L U M E 4 2
G UB A N OV A N D P E E L : C AM B R I A N H E L C I O N E L L O I D 215
TEXT-FIG. 4. Holotype ofOelandiella korobkoviVostokova, 1962; Nr. 8361–1; Lower Cambrian, Kotuj River.A, lateralview with the interpreted anterior to the right.B–C, lateral view with the interpreted anterior to the left, (C, SEM photo).D–G, various dorso-lateral views.H, dorsal view.I, antero-dorsal view to show the protruding earlier part of the whorl
(left) indicating slight asymmetry. All×15.
TEXT-FIG. 5. SEM photographs of the initial part of the holotype ofOelandiella korobkoviVostokova, 1962.A, antero-lateral view to show the beak-like apex and the appearance of the first rib;× 90. B, postero-lateral view to show the
smooth surface and the impression on the tip of the apex;×120.

bend opening outward toward the aperture (Text-fig. 6D). Most of the ribs disappear on the middle part of the lateralsurfaces towards the umbilici and only few almost reach the umbilical part of the whorl. When viewed laterally, theribs are gently convex towards the aperture.
The third, and newly obtained, specimen (Text-fig. 7) is the largest and the best preserved in the collection. It is aninternal mould tightly coiled through approximately one-and-one-third whorls and has a maximum length of 3·8 mmand a width of 1·2 mm. The dorsum of the initial part of the whorl touches the sub-apical surface of the last part of thewhorl after one complete revolution. Umbilici are not perforated. The initial whorl is slightly lifted up above the planeof the aperture and touches the arched sub-apical margin of the aperture. The aperture is elongate in plan view, with alength : width ratio ofc. 1·8:1 and gently convex margins when viewed laterally. The whorl has about 25 ribs, thespacing of which changes during ontogeny. The first part of the shell, which occupies about one revolution of thewhorl, bears rounded regularly repeated ribs with the same amplitude. Then repetition becomes irregular and theamplitude of some ribs increases whilst others disappear. Also, the distance between the ribs increases significantlyand the profile of individual ribs becomes asymmetrical, gently sloping outward away from the aperture. The early partof the whorl appears symmetrical but in the later stage, the shell becomes slightly asymmetrical, as clearly seen indorsal view (Text-fig. 4F, I).
Remarks. All three specimens have similar proportions. The main difference between them concerns thenumber of ribs. The holotype has seven ribs in one-quarter of a revolution of the whorl against 18 for thesecond specimen. The largest specimen has 12 ribs in the early stage of the shell and 10 in the later stage.
216 P A L A E O N T O L O G Y , V O L U M E 4 2
TEXT-FIG. 6A–D, Oelandiella korobkoviVostokova, 1962.A, lateral view, with the interpreted anterior to the right.B, postero-lateral view.C, antero-dorsal view to show a faintU-shaped bend of the ribs.D, dorsal view.E–H, Oelandiella sibiricaVostokova, 1962.E, antero-dorsal view.F, lateral view with the interpreted anterior to the
left. G, postero-lateral view.H, posterior view. All×15.

Oelandiella sibiricaVostokova, 1962
Text-figure 6E–H
1962 Oelandiella sibiricaVostokova, p. 53, pl. 1, figs 5–7.
Holotype. N8361–4 from the Lower Cambrian of the Kotuj River, north Siberia.
Description. The holotype is a small, coiled, laterally compressed internal mould of a helcionelloid mollusc with abroken initial part of the whorl (maximum lengthc. 3·85 mm and widthc. 1·35 mm). The comparatively well preservedfinal part of the whorl has an irregular alternation of about 20 (18 in one-quarter revolution of the whorl) differentlyexpressed, tightly gathered ribs. The ribs are straight when viewed laterally. The aperture is elongate, but its sub-apicalpart is not preserved.
Remarks. The rather poor preservation hinders detailed comparison of this species withO. korobkovi. Theproportions of the shell are similar to those ofO. korobkovi,but it is slightly more flattened laterally(length:width ratio of 2·8 against 2·2–2·6 forO. korobkovi). The ribs are less pronounced than inO. korobkovi and they are variable in width. Vostokova (1962) noticed three main differences betweenO. sibiricaandO. korobkovi. O. sibiricawas claimed to have a higher shell, with an irregular alternation ofwide and narrow ribs, and the initial part of the whorl appeared to be higher above the plane of the aperture.Our measurements show that the holotype ofO. sibirica has the same length:height ratio as the holotypeand the second specimen ofO. korobkovi,but slightly lower than the third specimen ofO. korobkovi. Thethird specimen ofO. korobkovihas a regular repetition of the ribs in the early stage of the whorl (which issimilar to the second specimen) and then the pattern becomes irregular and reminiscent ofO. sibirica.Differences in the length and emphasis of the ribs ofO. sibiricaare distinct in lateral aspect (Text-fig. 6A,F). However, when viewed dorsally, the ribs look almost the same in size, and dorsal views of the secondspecimen ofO. korobkoviand the holotype ofO. sibirica are rather similar (Text-fig. 7D–E). Moreover,examination of the second specimen ofO. korobkovireveals that some ribs are more pronounced, slightlywider, and longer than the others (Text-fig. 7A–C). Thus, the difference between the two species ofOelandiellarecognized by Vostokova (1962) is rather uncertain. It seems to be that Vostokova separatedthese species on the basis of the regularity of the alternation of the ribs. Regular repetition of ribs of thesame size and spacing characterizesO. korobkoviand these parameters increase proportionally duringontogeny. Hence, the number of ribs in one-quarter of a whorl is changed from about four (Vostokova1962, pl. 1, fig. 3), or even three for specimens described asLatouchella korobkovi(Rozanovet al. 1969,pl. 3, fig. 12), through seven in the holotype, to 18 in specimen N 8361–2.O. sibiricahas an alternation ofribs of slightly different size and spacing but this difference appears to be insufficient to justify itsseparation fromO. korobkovi.It is unprofitable to try to estimate the taxonomic value of the smalldifferences which exist between the two species on the base of the four available specimens. Solution ofthis problem awaits analysis of a huge collection from the basin of the Aldan River, Siberia, now inprogress.
O E L A N D I E L L A A N D O E L A N D I A
Oelandia pauciplicatawas described by Westerga˚rd (1936) as a new genus and species of gastropod fromthe Middle Cambrian of O¨ land, Sweden. The planispirally coiled shell ofOelandiashows a bilateralasymmetrical alternation of the sharpened ribs which cross the dorsum, but continue only a short distanceon the other side of the shell. In contrast, the comarginal rounded ribs ofOelandiellacross the dorsum andcontinue symmetrically on the other side of the shell (Text-fig. 8C–D). Available material ofOelandiais atleast twice as large asOelandiellaand has a significantly less tightly coiled and more expanded form. Thesub-apical part of the posteriorly extended aperture ‘trail’ ofOelandiais lifted up above the plane of theaperture and sometimes forms an upwardly pointing snorkel.
The redescription ofOelandia by Peel and Yochelson (1987) was based on the paratype andnew material (Text-fig. 8). These new specimens are crushed from the side, whereas the paratype
G UB A N OV A N D P E E L : C AM B R I A N H E L C I O N E L L O I D 217

(Text-fig. 8C–D) is preserved uncrushed, but with a broken apex. According to the retouched photo inWesterga˚rd’s paper, the holotype has a smooth dorsum, a rapidly expanded shell and its ribs are restrictedto the lateral surfaces of the shell, as inLatouchella costata.It is unclear from the illustration whether theholotype is bilaterally asymmetrical, as noted in the description, because only one side of the shell isexposed. The paratype of Westerga˚rd’s species and the additional two samples described by Peel andYochelson (1987) have a relatively slowly expanding shell with a bilaterally asymmetrical collabralplication and the last two have additionally a sub-apical snorkel.
218 P A L A E O N T O L O G Y , V O L U M E 4 2
TEXT-FIG. 7. Oelandiella korobkoviVostokova, 1962.A, lateral view with the anterior to the right.B, postero-lateralview. C, posterior view.D, oblique lateral view to show the changing from regularly repeated to irregularly repeatedribs.E, antero-lateral view.F, antero-dorsal view to show asymmetry of the shell.G, lateral view with the anterior to the
left. H, oblique lateral view.I, dorsal view to show asymmetry of the shell. All×20.

O E L A N D I E L L A A N D L A T O U C H E L L A
The holotype and paratype ofLatouchella costataCobbold, 1921 were recently redescribed (Gubanov andPeel 1998). The narrower, loosely coiled shell has comarginal ribs which are restricted to the lateralsurfaces and the initial part of the whorl is hemispherical, clearly distinguishingLatouchella fromOelandia. As we noted earlier (Gubanov and Peel 1998),L. costatais distinct from most other speciesassigned to the genus. A large number of helcionelloid molluscs, from the uppermost Vendian(Khomentovskyet al. 1990) to the Upper Cambrian (Weberset al. 1992), has been referred toLatouchella(Gubanov and Peel 1998). The morphology of these varies considerably, from tightly coiled, laterallycompressed and quickly expanded shells from the Lower Cambrian to conical and only slightly curvedHelcionella-like molluscs from the Middle and Upper Cambrian. With the exception of the type species,they have prominent comarginal ribs which cross the dorsum. The type species ofLatouchellais alsounusually laterally compressed when compared with the other species.
These molluscs which have been assigned toLatouchellacan be divided into three groups with differentage ranges: (1) an abundant group, dominated byOelandiella, in the Tommotian; (2) a few, including thetype species ofLatouchella, recorded from the upper Atdabanian–lower Botomian; (3) numeroushelcionelloid molluscs, includingOelandia, which appear in the lower Middle Cambrian. The fossilrecord between these three groups is poor and it is currently difficult to trace an evolutionary sequencewithin Latouchella-like molluscs. It is evident that taxonomic revision of the many species masqueradingin the literature under the nameLatouchella is a prerequisite to understanding this sequence. Severalgenera of helcionelloids have been proposed from Siberia (summarized in Missarzhevsky 1989; Esakovaand Zhegallo 1996), and the relationship of these to the redescribedLatouchellaandOelandiellaremainsto be clarified. Likewise, large faunas have been recorded from China (e.g. Yu 1979, 1987). Discussion ofthese helcionelloids lies outside the present context, but the basal Cambrian genusArchaeospiraYu, 1979,described in detail by Qian and Bengtson (1989), is of interest. As described by the latter authors,Archaeospira is a spirally coiled, sinistral shell with prominent comarginal plications which arecontinuous across the dorsum from one lateral area to the other. In terms of coiling and ornamentation,
G UB A N OV A N D P E E L : C AM B R I A N H E L C I O N E L L O I D 219
TEXT-FIG. 8. A–B, Latouchella costataCobbold, 1921; holotype, SM A 780, Sedgwick Museum, Cambridge; ComleyLimestone (Lower Cambrian), Shropshire, England,×15. C–F, Oelandia pauciplicataWesterga˚rd, 1936; MiddleCambrian, O¨ land, Sweden.C–D, paratype, SGU type 711, type collection, Geological Survey of Sweden (SGU),Uppsala, Sweden;×8. C, lateral view.D, antero-dorsal view to show the asymmetrically developed plications.E, LO
5757t; with anterior to the left;×4 . F, LO 5758t; with anterior to the right;×4.

its type speciesArchaeospira ornataYu, 1979 closely resemblesOelandiella korobkoviand Vostokova’sspecies name (ascribed toLatouchella) has been employed for material now placed withinA. ornatabyQian and Bengtson (1989, p. 112). Qian and Bengtson (1989) commented on the similarity of the twoforms, pointing out thatO. korobkoviwas not known to be anisometrically coiled, whilst a third species,Hubeispira nitidaYu, 1981, was apparently dextrally coiled. It is now known that at least some specimensof Oelandiella korobkovimay be slightly dextral (Text-fig. 4I). However, study of large samples isrequired before the suggestion of Qian and Bengtson (1989) that coiling differences of this kind may be afeature of different populations can be tested.
M O D E O F L I F E A N D A D A P T I V E S T R A T E G Y
The relatively wide aperture and pronounced, broadly rounded, comarginal plications suggest thatOelandiella, and many other helcionelloids, inhabited a bottom surface covered by an algal film, wherefood particles accumulated from the water column. The wideness of the aperture and the nature of theornamentation appear to rule out an infaunal mode of life, and suggest clamping against the substrate (cf.Linsley 1977). Moreover, studies by Stanley (1970) indicate that bivalves with prominent comarginalornamentation of the kind seen inOelandiella are slower burrowers than smooth forms. A benthicexistence was quickly mastered by the earliest helcionelloids and members of the class diversified intoother habitats, notably a semi-infaunal life style where they may have fed by filtration (Peel 1991a, 1991b).However, when considering filter feeding, it is relevant to recall that the earliest bivalves, contempora-neous with the helcionelloids, probably used the foot both for locomotion and feeding, and the gills onlyfor respiration (Morton 1996).
The success of helcionelloids as members of the epifauna is documented in the evolutionary lineagefrom OelandiellathroughBemellato Helcionellawhich may reflect adaptation from deposit feeding to agrazing habit on a somewhat harder sediment surface. In terms of morphology, this is reflected in achange from a coiled to a low conical shell form. The aperture widened and became sub-circular in planview, suggesting a decrease in mobility and increasing hardness of the substrate (Linsley 1977; Gubanov1984).
In contrast, an evolutionary lineage fromOelandiellavia AnabarellaVostokova, 1962 toWatsonellaGrabau, 1900 (=HeraultipegmaPojeta and Runnegar, 1976), through undescribed transitional forms fromthe Lower Cambrian (Tommotian) of Siberia and Spain, indicates adaptation to a softer substrate through asemi-infaunal mode of life. This trend is accompanied by a loss of the strong comarginal ornamentationand increased lateral compression. The same scenario was apparently repeated several times during theCambrian within the class Helcionelloida, for example withinEotebennaRunnegar and Jell, 1976 (Peel1991b).
The mid Early CambrianLatouchella, as exemplified by its type speciesL. costata, has a rather narrowshell and aperture, and a degree of lateral compression which is more similar to that ofAnabarellathanOelandiella. It is probable thatLatouchellacould partially burrow into the sediment in search of its foodand possibly also for increased protection. Loss of comarginal ribs on the median dorsal area might parallelthe reduction in ornamentation seen in theOelandiella–Anabarella–Watsonellalineage.
Oelandiahas an asymmetrical alternation of the prominent ribs on the lateral areas (Text-fig. 8C–F), thefunction of which is problematical. Peel and Yochelson (1987) suggested that the enigmatic ornamentationmight help anchor the apertural region of the laterally compressed shell or, in association with a gentlerocking movement, assist the process of embedding, but they found the dimensions of the individual ribs tobe excessive for this purpose alone. Whatever their function, asymmetrical alternating ribs occur rarely inthe Middle Cambrian (Yu and Ning 1985; Geyer 1986; Peel and Yochelson 1987) and were not widelyadopted by helcionelloids.
Acknowledgements. We thank Nina M. Kadlets (CNIGR Museum, St-Petersburg) for providing access to the typespecimen ofOelandiellaand the Swedish Natural Sciences Research Council (NFR) for financial support of the trip byAPG to St-Petersburg. Support from NFR and INTAS through grants to JSP is gratefully acknowledged.
220 P A L A E O N T O L O G Y , V O L U M E 4 2

R E F E R E N C E S
BRASIER, M. D.1989. Towards a biostratigraphy of the earliest skeletal biotas. 117–165.In COWIE, J. W.andBRASIER, M. D.
(eds).The Precambrian-Cambrian boundary. Oxford Science Publications, Oxford, 213 pp.COBBOLD, E. S.1921. The Cambrian horizons of Comley (Shropshire) and their Brachiopoda, Pteropoda, Gasteropoda
and etc.Quarterly Journal of the Geological Society, London, 76, 325–386.ESAKOVA, N. V. and ZHEGALLO, E. A. 1996. [Biostratigraphy and fauna of Lower Cambrian of Mongolia.] Nauka,
Moscow, 216 pp. [In Russian].GEYER, G.1986. Mittelkambrische Mollusken aus Marokko und Spanien.Senckenbergiana lethaea, 67, 55–118.—— 1994. Middle Cambrian mollusks from Idaho and early conchiferan evolution. 69–86.In LANDING, E. (ed.). Studies in
stratigraphy and paleontology in honor of Donald W. Fisher.Bulletin of the New York State Museum, 481, 1–380.GRABAU, A. W. 1900. Paleontology of the Cambrian terrains of the Boston basin.Occasional Papers of the Boston
Society for Natural History,4 (3), 601–694.GUBANOV, A. P. 1984. Forma rakoviny gastropod i ee svyaz’ so sredoj obitaniya i obrazom zhisni. 53–57.In: Geologo-
geofisicheskie issledovaniya v Sibiri. Institut Geologii i Geofiziki Sibirskogo Otdeleniya Akademii Nauk SSSR,Novosibirsk, 164 pp. [In Russian].
—— andPEEL, J. S.1998. Redescription of the type species ofLatouchellaCobbold, 1921 (Mollusca) from the LowerCambrian of Comley (England).GFF, 120, 17–20.
KHOMENTOVSKY, V. V. andKARLOVA, G. A. 1986. O nizhnej granitse pestrotsvetnoj svity v bassejne r. Aldan. 3–32.InSOKOLOV, B. S. (ed.). Pozdnij dokembrij I rannij paleozoj Sibiri. Sibirskaya platforma I vneshnyaya zona Altae-Sayanskoj skladchatoj oblasti. Nauka, Novosibirsk, 220 pp. [In Russian].
—— —— 1994. [Ecological peculiarities of the Vendian–Cambrian small shelly fauna in the Siberian platform.]Stratigraphy and Geological Correlation,3, 206–215. [In Russian].
—— VAL’KOV, A. K. andKARLOVA, G. A. 1990. Novye dannye po biostratigrafii perekhodnykh vend-kembrijskikh sloev vbassejne srednego techeniya r. Aldan. 3–57.In KHOMENTOVSKY, V. V. andGIBSHER, A. S.(eds).Pozdnij dokembrij i rannijpaleozoj Sibiri. Voprosy regional’noj stratigrafii. Institut Geologii i Geofiziki Sibirskoe Otdelenie Akademii NaukSSSR, Novosibirsk, 168 pp. [In Russian].
KNIGHT, J. B. 1941. Paleozoic gastropod genotypes.Special Paper of the Geological Society of America, 32, 1–510.—— COX, L. R., KEEN, A. M., BATTEN, R. L., YOCHELSON, E. L.andROBERTSON, R.1960. Systematic descriptions. I169–
I1310. In MOORE, R. C.(ed.).Treatise on invertebrate paleontology. Part I. Mollusca 1. University of Kansas Pressand Geological Society of America, 351 pp.
LINSLEY, R. M. 1977. Some ’laws’ of gastropod shell form.Paleobiology,3, 196–206.MESHKOVA, N. P., ZHURAVLEVA, I. T. andLUCHININA, V. A. 1973. Nizhnij kembrij i nizhnyaya chast’ srednego kembriya
Olenekskogo podnyatiya. 194–214.In ZHURAVLEVA, I. T. (ed.).Problemy paleontologii i biostratigrafii nizhnegokembriya Sibiri i Dal’nego Vostoka.Nauka, Novosibirsk, 264 pp. [In Russian].
MISSARZHEVSKY, V. V. 1980. O pogranichnykh sloyakh kembriya i dokembriya zapadnogo sklona Olenekskogopodnyatiya (r. Olenek).Bulletin Moskovskogo Obschestva Ispytatelei Prirody,55, 23–34. [In Russian].
—— 1982. Raschlenenie i korrelatsiya pogranichnykh tolsch dokembriya i kembriya po nekotorym drevnejshimgruppam organizmov.Bulletin Moskovskogo Obschestva Ispytatelei Prirody,57, 52–67. [In Russian].
—— 1983. Stratigrafiya drevnejshikh tolshch fanerozoya Anabarskogo massiva.Sovetskaya Geologiya,9, 62–73. [InRussian].
—— 1989.Drevnejshie skeletnye okamenelosti i stratigrafiya pogranichnykh tolshch dokembriya i kembriya.Nauka,Moscow, 237 pp. [In Russian].
MORTON, B. 1996. The evolutionary history of the Bivalvia. 337–359.In TAYLOR, J. D. (ed.).Origin and evolutionaryradiation of the Mollusca. Oxford Science Publications, Oxford, 392 pp.
PEEL, J. S.1991a. Functional morphology of the Class Helcionelloida nov., and the early evolution of the Mollusca.157–177.In SIMONETTA, A. andCONWAY MORRIS, S.(eds).The early evolution of Metazoa and the significance ofproblematic taxa. Cambridge University Press, Cambridge, 296 pp.
—— 1991b. The classes Tergomya and Helcionelloida, and early molluscan evolution.Bulletin GrønlandsGeologiske Undersøgelse,161, 11–65.
—— andYOCHELSON, E. L.1987. New information onOelandia(Mollusca) from the Middle Cambrian of Sweden.Bulletin of the Geological Society of Denmark,36, 263–273.
QIAN YI and BENGTSON, S.1989. Palaeontology and biostratigraphy of the Early Cambrian Meishucunian Stage inYunnan Province, South China.Fossils and Strata, 24, 1–156.
RAABEN, M. E. (ed.) 1981.The Tommotian Stage and the Cambrian lower boundary problem. Amerind PublishingCompany, New Delhi, 359 pp.
REPINA, L. N. andROZANOV, A. Yu. 1984. Iz istorii stanovleniya yarusnoj shkaly nizhnego kembriya. 5–19.In ROZANOV,
G UB A N OV A N D P E E L : C AM B R I A N H E L C I O N E L L O I D 221

A. Yu. andSOKOLOV, B. S. (eds).Yarusnoe raschlenenie nizhnego kembriya. Stratigrafiya.Nauka, Moscow. [InRussian].
ROZANOV, A. Yu, MISSARZHEVSKY, V. V., VOLKOVA, N. A. , VORONOVA, L. G., KRYLOV, I. N., KELLER, B. M., KOROLYUK, I. K.,LENDZION, K., MICHNIAK, R., PYCHOVA, N. G. andSIDOROV, A. D. 1969.Tommotskij yarus i problema nizhnej granitsykembriya. Nauka, Moscow, 380 pp. [In Russian].
——andZHURAVLEV, A. Yu. 1992. The Lower Cambrian fossil record of the Soviet Union. 205–282.In LIPPS, J. H.andSIGNOR, P. W.(eds).Origin and early evolution of the Metazoa.Topics in Geobiology, 10. Plenum Press, New Yorkand London, 570 pp.
RUNNEGAR, B. 1996. Early evolution of the Mollusca: the fossil record. 77–87.In TAYLOR, J. D. (ed.). Origin andevolutionary radiation of the Mollusca.Oxford Science Publications, Oxford, 392 pp.
—— and JELL, P. A. 1976. Australian Middle Cambrian molluscs and their bearing on early molluscan evolution.Alcheringa, 1, 109–138.
—— andPOJETA, J. Jr 1974. Molluscan phylogeny: the paleontological viewpoint.Science, 186, 311–317.SAVITSKY, V. E. 1962. O sootnoshenii kembriya i verkhnego dokembriya Anabarskogo shchita. 53–54.In SOKOLOV, B. S.
(ed.).Soveshchanie po stratigrafii otlozhenij pozdnego dokembriya Sibiri i Dal’nego Vostoka. Sibirski NauchnoIssledovatel’ski Institut Geologii Geofiziki i Mineral’nogo Syr’ya, Novosibirsk, 124 pp. [In Russian].
STANLEY, S. 1970. Relation of shell form to life habits in the Bivalvia (Mollusca).Memoir of the Geological Society ofAmerica, 125, 1–206.
VAL’KOV, A. K. 1975.Biostratigrafiya i khiolity kembriya severo-vostoka Sibirskoj platformy. Nauka, Moscow, 140 pp.[In Russian].
—— 1982.Biostratigrafiya nizhnego kembriya Vostoka Sibirskoj platformy. Uchuro-Majskij rajon. Nauka, Moscow,92 pp. [In Russian].
—— 1987. Biostratigrafiya nizhnego kembriya Vostoka Sibirskoj platformy. Yudomo-Olenekskij rajon. Nauka,Moscow, 135 pp. [In Russian].
VORONIN, Yu. I., VORONOVA, L. G., GRIGOR’EVA, N. V., DROZDOVA, N. A., ZHEGALLO, E. A., ZHURAVLEV, A. Y u.,RAGOZINA, A. L.,ROZANOV, A. Yu., SAYUTINA, T. A., SYSOEVA, V. A. and FONIN, V. D. 1982. Granitsa dokembriya i kembriya vgeosinklinal’nykh oblastyakh (opornyj razrez Salany-Gol, M. N. R.).Nauka, Moscow, 164 pp. [In Russian].
VORONOVA, L. G., GRIGOR’EVA, N. V., ZHEGALLO, E. A., MISSARZHEVSKY, V. V. and SYSOEV, V. A. 1983. Vozrast sloev‘‘ Oelandiella’’ korobkovi - Anabarella planana Sibirskoj platforme.Izvestiya Akademii Nauk SSSR, Seriyageologicheskaya, 12, 80–84. [In Russian].
VOSTOKOVA, V. A. 1962. Kembrijskie gastropody Sibirskoj platformy i Tajmyra.Trudy Nauchno Issledovatel’skogoInstituta Geologii Arktiki,28, 51–74. [In Russian].
WEBERS, G. F., POJETA, J. Jr and YOCHELSON, E. L. 1992. Cambrian Mollusca from the Minaret Formation, EllsworthMountains, West Antarctica.Memoir of the Geological Society of America,170, 181–245.
WENZ, W. 1938. Gastropoda. Allgemeiner Teil und Prosobranchia. 1–720.In SCHINDEWOLF, O. H.(ed.).Handbuch derPalaozoologie.Band 6. Borntrager, Berlin, 1639 pp.
WESTERGARD, A. H. 1936.Paradoxides oelandicusbeds of Oland.Sveriges Geologiska Underso¨kning, Series C, 394, 1–66.YU WEN 1979. Earliest Cambrian monoplacophorans and gastropods from western Hubei and their biostratigraphical
significance.Acta Palaeontologica Sinica, 18, 233–270. [In Chinese, with English summary].—— 1981. New Earliest Cambrian monoplacophorans and gastropods from W. Hubei and E. Yunnan.Acta
Palaeontologica Sinica, 20, 552–556. [In Chinese, with English summary].—— 1987. Yangtze micromolluscan fauna in Yangtze region of China with notes on Precambrian-Cambrian
boundary. 19–344.In: Stratigraphy and palaeontology of systematic boundaries in China. Precambrian-Cambrianboundary (1). Nanjing University Publishing House, 344 pp. [In Chinese, with English summary].
—— andNING HUI 1985. Two Cambrian monoplacophorans from Borohore, Xinijang.Acta Palaeontologica Sinica,24, 47–50. [In Chinese, with English summary].
ZHURAVLEVA, I. T. , MESHKOVA, N. P., LUCHININA, V. A. , KASHINA, L. N., KORSHUNOV, V. I.andPEL’MAN. Yu. L. 1979. Osobennostiorganicheskogo mira na rubezhe kembriya i dokembriya (sever Sibirskoj platformy i sopredel’nye territorii). 193–200.In SOKOLOV, B. S.(ed.).Paleontologiya dokembriya i rannego kembriya.Nauka, Leningrad, 248 pp. [In Russian].
ALEXANDER P. GUBANOV
JOHN S. PEEL
Historical Geology and PalaeontologyDepartment of Earth Sciences, Uppsala University
Norbyvagen 22, S-752 36, Uppsala, Swedene-mail [email protected]
e-mail [email protected] received 2 April 1998Revised typescript received 10 September 1998
222 P A L A E O N T O L O G Y , V O L U M E 4 2
Related Documents











