Novel antibodies targeting immune regulatory checkpoints for cancer therapy Chern Siang Lee, Mark Cragg, Martin Glennie & Peter Johnson Cancer Sciences Unit, Faculty of Medicine, University of Southampton, Southampton General Hospital, Southampton SO16 6YD, UK Correspondence Professor Martin Glennie, Cancer Sciences Unit, Faculty of Medicine, University of Southampton, Southampton General Hospital, Southampton SO16 6YD, UK. Tel.: +02 3 8079 6593 Fax: +02 3 8070 4061 E-mail: [email protected] ----------------------------------------------------------------------- Keywords cancer, immunomodulatory antibodies, immunotherapy, monoclonal antibodies ----------------------------------------------------------------------- Received 2 October 2012 Accepted 17 April 2013 Accepted Article Published Online 23 May 2013 Cancers must evade or suppress the immune system in order to develop. Better understanding of the molecular regulation governing tumour detection and effective activation of the immune system (so called immune regulatory checkpoints) has provided new targets for cancer immunotherapy.Therapeutic monoclonal antibodies against these targets are currently undergoing clinical evaluation with more in pre-clinical development; buoyed by the recent licence approval of the anti-CTLA-4 antibody, ipilumumab, for use in melanoma.This article will review the current status of the various antibodies and target molecules being investigated. Introduction The use of monoclonal antibodies (mAbs) as therapeutics has increased rapidly over the last 30 years. The first gen- eration of therapeutic mAbs were of mouse origin. This often resulted in a rapid induction of human anti-mouse antibody (HAMA) responses when infused into humans; limiting their half-life and therapeutic efficacy [1–4]. Sub- stitution of the murine Fc domain with human sequences through recombinant DNA technology resulted in chi- meric mAbs with improved pharmacokinetic and thera- peutic efficacy [5–7]. Progressive improvements in recombinant DNA and cloning techniques eventually led to the development of humanized mAbs carrying mouse hypervariable loops in the human V-region framework [8, 9]. Fully human mAbs are now available through the use of phage display libraries or genetically modified mice carry- ing human rather than mouse immunoglobulin loci [10– 13]. Current mAb nomenclature attaches suffixes to discriminate these various mouse, chimeric, humanized and human mAbs as well as mAb-derivatives such as Fc fusions and bispecific reagents (Table 1). Alongside small molecule target inhibitors, immune therapies and therapeutic mAbs, in particular, have become an important treatment strategy in oncology with numerous mAbs approved by the United States Food and Drug Administration (FDA) and the European Medicines Agency (EMA), and with many more undergoing clinical evaluation and pre-clinical development. Those such as rituximab, which directly target molecules expressed on the tumour (i.e. CD20 on lymphoma) can achieve their therapeutic effects through Fc-dependent cytotoxic effects, interacting with Fc receptor-expressing cells of the innate immune effector system to trigger antibody- dependent cell mediated cytotoxicity (ADCC) or serum proteins to elicit complement dependent cytotoxicity (CDC) [14, 15]. Others, such as trastuzumab and cetuximab, can also function by blocking key survival signals in the tumour cell provoking cell cycle arrest and cell death in addition to ADCC [16–19]. A detailed discussion of the activity and modes of action of these so-called direct tar- geting mAbs is beyond the scope of this article. Table 2 provides a brief overview of these agents, their targets and mechanisms of action with Table 3 summarising a selec- tion of promising agents currently undergoing clinical evaluation. In this review we will examine a different and fast-developing class of mAbs, those that do not target the tumour directly but instead target receptors of the immune system to promote anti-cancer immunity, so-called immunomodulatory mAb. British Journal of Clinical Pharmacology DOI:10.1111/bcp.12164 Br J Clin Pharmacol / 76:2 / 233–247 / 233 © 2013 The British Pharmacological Society

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Novel antibodies targetingimmune regulatorycheckpoints forcancer therapyChern Siang Lee, Mark Cragg, Martin Glennie & Peter Johnson
Cancer Sciences Unit, Faculty of Medicine, University of Southampton, Southampton General Hospital,
Southampton SO16 6YD, UK
CorrespondenceProfessor Martin Glennie, Cancer SciencesUnit, Faculty of Medicine, University ofSouthampton, Southampton GeneralHospital, Southampton SO16 6YD, UK.Tel.: +02 3 8079 6593Fax: +02 3 8070 4061E-mail: M.J.Glennie@soton.ac.uk-----------------------------------------------------------------------
Keywordscancer, immunomodulatory antibodies,immunotherapy, monoclonal antibodies-----------------------------------------------------------------------
Received2 October 2012
Accepted17 April 2013
Accepted ArticlePublished Online23 May 2013
Cancers must evade or suppress the immune system in order to develop. Better understanding of the molecular regulation governingtumour detection and effective activation of the immune system (so called immune regulatory checkpoints) has provided new targetsfor cancer immunotherapy. Therapeutic monoclonal antibodies against these targets are currently undergoing clinical evaluation withmore in pre-clinical development; buoyed by the recent licence approval of the anti-CTLA-4 antibody, ipilumumab, for use inmelanoma. This article will review the current status of the various antibodies and target molecules being investigated.
Introduction
The use of monoclonal antibodies (mAbs) as therapeuticshas increased rapidly over the last 30 years. The first gen-eration of therapeutic mAbs were of mouse origin. Thisoften resulted in a rapid induction of human anti-mouseantibody (HAMA) responses when infused into humans;limiting their half-life and therapeutic efficacy [1–4]. Sub-stitution of the murine Fc domain with human sequencesthrough recombinant DNA technology resulted in chi-meric mAbs with improved pharmacokinetic and thera-peutic efficacy [5–7]. Progressive improvements inrecombinant DNA and cloning techniques eventually ledto the development of humanized mAbs carrying mousehypervariable loops in the human V-region framework [8,9]. Fully human mAbs are now available through the use ofphage display libraries or genetically modified mice carry-ing human rather than mouse immunoglobulin loci [10–13]. Current mAb nomenclature attaches suffixes todiscriminate these various mouse, chimeric, humanizedand human mAbs as well as mAb-derivatives such as Fcfusions and bispecific reagents (Table 1).
Alongside small molecule target inhibitors, immunetherapies and therapeutic mAbs, in particular, havebecome an important treatment strategy in oncology with
numerous mAbs approved by the United States Food andDrug Administration (FDA) and the European MedicinesAgency (EMA), and with many more undergoing clinicalevaluation and pre-clinical development. Those such asrituximab, which directly target molecules expressed onthe tumour (i.e. CD20 on lymphoma) can achieve theirtherapeutic effects through Fc-dependent cytotoxiceffects, interacting with Fc receptor-expressing cells ofthe innate immune effector system to trigger antibody-dependent cell mediated cytotoxicity (ADCC) or serumproteins to elicit complement dependent cytotoxicity(CDC) [14, 15]. Others, such as trastuzumab and cetuximab,can also function by blocking key survival signals in thetumour cell provoking cell cycle arrest and cell death inaddition to ADCC [16–19]. A detailed discussion of theactivity and modes of action of these so-called direct tar-geting mAbs is beyond the scope of this article. Table 2provides a brief overview of these agents, their targets andmechanisms of action with Table 3 summarising a selec-tion of promising agents currently undergoing clinicalevaluation. In this review we will examine a different andfast-developing class of mAbs, those that do not targetthe tumour directly but instead target receptors of theimmune system to promote anti-cancer immunity,so-called immunomodulatory mAb.
British Journal of ClinicalPharmacology
DOI:10.1111/bcp.12164
Br J Clin Pharmacol / 76:2 / 233–247 / 233© 2013 The British Pharmacological Society

Immunomodulatory mAbs forcancer immunotherapy
Immunomodulatory mAbs can provoke immune rejectionof the tumour without having to identify directly tumourantigens that may be unique to an individual cancer. Theadvantage of such an approach, in comparison with con-ventional therapies and even direct targeting mAbs, isthe potential for developing immunological memory tothe tumour, preventing its recurrence, the main issue ofmodern cancer treatment.
Regulation of T-cell responsesThe recognition of antigenic peptides (e.g. tumour-associated antigens) bound to the major histocompatibil-ity complex (MHC) by the T-cell receptor (TCR) alone isinsufficient for T- cell activation to effect immune destruc-tion of the cancer cell. Co-regulatory (inhibitory orstimulatory) signals are also required, transmitted bya series of receptor-ligand pairs (see Figure 1). Theseco-regulatory molecules can be constitutively (e.g. CD28on naive T-cells) or variably expressed on the cell surface atdifferent stages of an immune response (early activatoryphase or late executory phase) and interact in immuno-logical synapses formed between the different cell types ofthe adaptive immune system and tumour cells. Ultimately,they determine the fate of the T-cell response; activationand differentiation into effector T-cells, deletion or anergy.Such regulatory checkpoints are crucial for the mainte-nance of self-tolerance under physiological conditions butthere is an increasing recognition of the important role theimmune system plays in cancer formation [20]. Cancer cellscan co-opt these mechanisms or release immunosuppres-sive cytokines such as transforming growth factor β(TGF-β) to evade and suppress the immune system inorder to develop into a tumour [21]. Tumour infiltratinglymphocytes (TILs) that fail to eliminate the tumour but yet
possess cytolytic functions in vitro suggest an endogenousanti-tumour effect that can be harnessed if immune toler-ance can be broken [22]. Such immune tolerance of thecancer by the host can be disrupted by using mAbs totarget immune checkpoint receptors in order to boostweak, ineffectual endogenous anti-tumour immuneresponses to therapeutic levels. For example, immunecheckpoint receptors that inhibit effector T-cell functionand boost regulatory T-cell functions (regulatory T-cellsplay an immunosuppressive role in the tumourmicroenvironment by inhibiting anti-tumour immuneresponses) are called checkpoint blockers and serve aspromising therapeutic targets.
Immunomodulatory mAbs can be either activatory(agonists) towards a stimulatory receptor by mimickingligand engagement (immunostimulatory mAb) or inhibi-tory (antagonistic) against a checkpoint blocker by block-ing ligand engagement. Figure 2 illustrates the multipletherapeutic targets that may be manipulated with theseimmunomodulatory mAbs. Examples of these mAbs aredetailed below and a selection of agents undergoing clini-cal evaluation are summarized in Table 4.
Antagonists of checkpoint blockersLigation of the T-cell activator CD28 by a member of the B7family of co-stimulatory molecules, B7-1 (also known asCD80) or B7-2 (also known as CD86), greatly enhances TCR-induced survival, proliferation, differentiation and eventualactivation of the naive T-cell. Concurrently as one of theimmunological counterbalances to attenuate unwanted orexcessive T-cell responses, cytotoxic T-lymphocyte associ-ated antigen 4 (CTLA4), is transported from intracellularstores to the cell surface where it serves to down-regulatethe T-cell activation signal by avid competitive binding toB7 molecules. For this reason, CTLA4 has been referred toas a brake on the immune system and was proposed as aviable target for blocking with mAbs to enhance anti-cancer responses.
Ipilimumab is a fully human mAb that blocks CTLA4function. It is the first immunomodulatory mAb to belicensed after demonstrating a statistically and clinicallysignificant survival benefit in a pivotal phase III clinical trial[23]. Patients with metastatic melanoma who had receivedat least one previous line of treatment were randomizedto receive ipilimumab with a glycoprotein 100 (gp100)peptide vaccine, ipilimumab alone or gp100 alone as thecontrol arm. The median overall survival (OS) was 10months [95% confidence interval (CI) 8.5, 11.5] in patientsreceiving both ipilimumab and gp100 vs. 6.4 months (95%CI 5.5, 8.7) in the gp100 alone group [hazard ratio (HR) fordeath 0.68, P < 0.001]. The median OS in the ipilumumabonly group was 10.1 months (95% CI 8.0, 13.8) and thehazard ratio of death was 0.66 (P = 0.003) when comparedwith gp100 alone. There was no difference in OS betweenthe two ipilimumab containing groups. Further clinicaltrials are on-going in multiple tumour types as either
Table 1Antibody nomenclature
Substem-suffix Type of molecule
-a-mab Rat-o-mab Mouse
-e-mab Hamster-i-mab Primate
-xi-mab Chimeric-xizu-mab Chimeric-humanized
-zu-mab Humanized-u-mab Fully human antibody from phage display or transgenic mice
technology
-stim Fc fusion peptide-cept Fc fusion protein
-axo-mab Bispecific mouse-rat hybrid mAb
C. S. Lee et al.
234 / 76:2 / Br J Clin Pharmacol
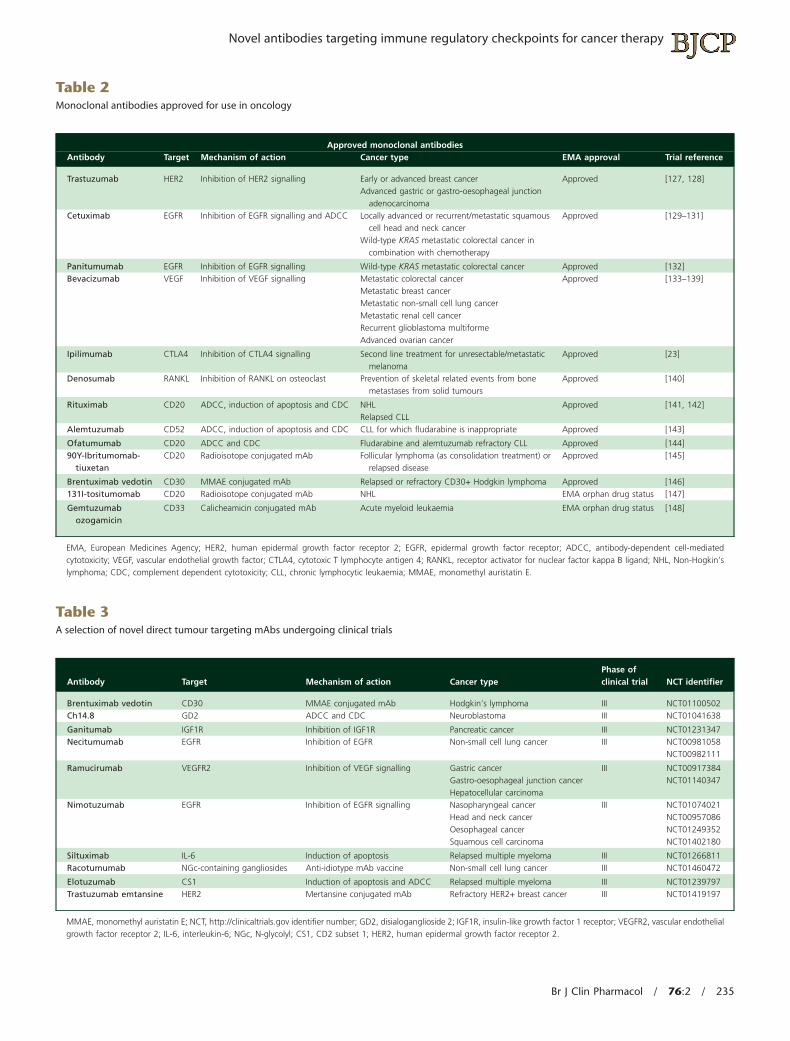
Table 2Monoclonal antibodies approved for use in oncology
Approved monoclonal antibodiesAntibody Target Mechanism of action Cancer type EMA approval Trial reference
Trastuzumab HER2 Inhibition of HER2 signalling Early or advanced breast cancerAdvanced gastric or gastro-oesophageal junction
adenocarcinoma
Approved [127, 128]
Cetuximab EGFR Inhibition of EGFR signalling and ADCC Locally advanced or recurrent/metastatic squamouscell head and neck cancer
Wild-type KRAS metastatic colorectal cancer incombination with chemotherapy
Approved [129–131]
Panitumumab EGFR Inhibition of EGFR signalling Wild-type KRAS metastatic colorectal cancer Approved [132]Bevacizumab VEGF Inhibition of VEGF signalling Metastatic colorectal cancer
Metastatic breast cancerMetastatic non-small cell lung cancerMetastatic renal cell cancerRecurrent glioblastoma multiformeAdvanced ovarian cancer
Approved [133–139]
Ipilimumab CTLA4 Inhibition of CTLA4 signalling Second line treatment for unresectable/metastaticmelanoma
Approved [23]
Denosumab RANKL Inhibition of RANKL on osteoclast Prevention of skeletal related events from bonemetastases from solid tumours
Approved [140]
Rituximab CD20 ADCC, induction of apoptosis and CDC NHLRelapsed CLL
Approved [141, 142]
Alemtuzumab CD52 ADCC, induction of apoptosis and CDC CLL for which fludarabine is inappropriate Approved [143]
Ofatumumab CD20 ADCC and CDC Fludarabine and alemtuzumab refractory CLL Approved [144]90Y-Ibritumomab-
tiuxetanCD20 Radioisotope conjugated mAb Follicular lymphoma (as consolidation treatment) or
relapsed diseaseApproved [145]
Brentuximab vedotin CD30 MMAE conjugated mAb Relapsed or refractory CD30+ Hodgkin lymphoma Approved [146]131I-tositumomab CD20 Radioisotope conjugated mAb NHL EMA orphan drug status [147]
Gemtuzumabozogamicin
CD33 Calicheamicin conjugated mAb Acute myeloid leukaemia EMA orphan drug status [148]
EMA, European Medicines Agency; HER2, human epidermal growth factor receptor 2; EGFR, epidermal growth factor receptor; ADCC, antibody-dependent cell-mediatedcytotoxicity; VEGF, vascular endothelial growth factor; CTLA4, cytotoxic T lymphocyte antigen 4; RANKL, receptor activator for nuclear factor kappa B ligand; NHL, Non-Hogkin’slymphoma; CDC, complement dependent cytotoxicity; CLL, chronic lymphocytic leukaemia; MMAE, monomethyl auristatin E.
Table 3A selection of novel direct tumour targeting mAbs undergoing clinical trials
Antibody Target Mechanism of action Cancer typePhase ofclinical trial NCT identifier
Brentuximab vedotin CD30 MMAE conjugated mAb Hodgkin’s lymphoma III NCT01100502Ch14.8 GD2 ADCC and CDC Neuroblastoma III NCT01041638
Ganitumab IGF1R Inhibition of IGF1R Pancreatic cancer III NCT01231347Necitumumab EGFR Inhibition of EGFR Non-small cell lung cancer III NCT00981058
NCT00982111
Ramucirumab VEGFR2 Inhibition of VEGF signalling Gastric cancerGastro-oesophageal junction cancerHepatocellular carcinoma
III NCT00917384NCT01140347
Nimotuzumab EGFR Inhibition of EGFR signalling Nasopharyngeal cancerHead and neck cancerOesophageal cancerSquamous cell carcinoma
III NCT01074021NCT00957086NCT01249352NCT01402180
Siltuximab IL-6 Induction of apoptosis Relapsed multiple myeloma III NCT01266811Racotumumab NGc-containing gangliosides Anti-idiotype mAb vaccine Non-small cell lung cancer III NCT01460472
Elotuzumab CS1 Induction of apoptosis and ADCC Relapsed multiple myeloma III NCT01239797Trastuzumab emtansine HER2 Mertansine conjugated mAb Refractory HER2+ breast cancer III NCT01419197
MMAE, monomethyl auristatin E; NCT, http://clinicaltrials.gov identifier number; GD2, disialoganglioside 2; IGF1R, insulin-like growth factor 1 receptor; VEGFR2, vascular endothelialgrowth factor receptor 2; IL-6, interleukin-6; NGc, N-glycolyl; CS1, CD2 subset 1; HER2, human epidermal growth factor receptor 2.
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 235

monotherapy or in combination with other modalities(see Table 4). The question of scheduling ipilimumab withcytotoxic chemotherapy is of particular importancebecause although chemotherapy may increase antigendelivery it may also delete key immune cells. A recentphase II, three-armed, placebo-controlled study (CA184-041) randomized 204 treatment naive patients withadvanced non-small cell lung cancer to chemotherapy
only, chemotherapy and concurrent ipilimumab or twocycles of chemotherapy before ipilimumab was phased in.Non-progressive patients who tolerated treatment thenreceived ipilimumab or placebo maintenance every 12weeks. The study met its primary end-point of immune-related progression free survival (irPFS) for the phasedtreatment group (HR 0.72, P = 0.05) but not for the concur-rent treatment group (HR 0.81, P = 0.13) compared with the
Antigen presenting cellor tumour cell
(depending on context) T-cell
BTLAHVEM
CTLA4B7 1/2
TCR
CD28
PD-1PD-L1
MHC
B7 1/2
T-cellResponse?
Immunoglobulinsuperfamily members
CD40
4-1BB
CD27
OX40
CD40L
4-1BBL
CD70
OX40L
TNFR superfamilymembers
TIM3GAL-9
LAG3MHC2
CD40
Signal 2: Co-stimulatory signals
Signal 2: Co-Inhibitory signals
Outcome
4-1BB 4-1BBL
CD40L
Signal 1:Antigenic
stimulation
Signal 3:Cytokines
AnergyDeletion
Effector T-cell
Activation
Proliferation
Differentiation
Abortiveactivation
VISTAUnidentified
receptor
Figure 1Schematic representation of an immune synapse and downstream events. Signal 1 is generated by T-cell receptor (TCR) recognition of peptide antigenpresented on major histocompatibility complex (MHC) by antigen-presenting cells (APCs, e.g. dentritic cells) or tumour cells. Signal 2 is provided byco-regulatory (co-stimulatory or inhibitory) molecules interacting with their cognate ligands expressed on APCs. These co-regulatory molecules can beexpressed constitutively in naive T-cells (e.g. CD28 and CD27) or induced by TCR engagement leading to expression at different time points of T-cellactivation. Signal 3 is mostly provided by cytokines secreted by APCs or other immune cells including CD4 T-helper cells to help regulate the effectordifferentiation pathway. Successful activation of a naive CD8 T-cell allows survival and proliferation of the reactive clone, allowing their progeny todifferentiate into cytotoxic T lymphocytes that kill tumour cells expressing the tumour associated antigen. Unsuccessful activation leads to anergy ordeletion of the T-cell. Transient, abortive activation of T-cells leading to deletion or anergy may occur in the context of antigen presentation by immature(or insufficiently activated) dendritic cells. CD40/CD40L and 4-1BB/4-1BBL are known as bi-directional signalling molecules and ligand engagementactivates intra-cellular signalling transduction pathways. Furthermore the receptors can also be expressed on the target cell and may play a part in therapywith mAb. LAG3, lymphocyte activation gene 3; TIM3, T-cell immunoglobulin and mucin domain 3; GAL-9, galectin-9; PD-1, programmed death-1; PDL-1,programmed death ligand-1; HVEM, herpes virus entry mediator; BTLA, B and T lymphocyte activator; CTLA4, cytotoxic T-lymphocyte associated antigen 4;VISTA, V-domain Ig suppressor of T-cell activation; TNFR, tumour necrosis factor receptor
C. S. Lee et al.
236 / 76:2 / Br J Clin Pharmacol

control group. There was also an improvement in WHO-criteria progression free survival (HR 0.69, P = 0.02) [24]. Aphase III trial (NCT01285609) examining this regime insquamous cell lung cancer is now underway as a subsetanalysis suggested greater efficacy in favour of squamoushistology. The parallel study examining extensive stagesmall cell lung cancer patients (n = 130) also arrived at asimilar conclusion favouring the phased ipilimumabregime for irPFS (HR 0.63, P = 0.03) over the concurrentregime (HR 0.75, P = 0.11) compared with the control group[25]. Although small, these clinical studies highlight theneed for more well-designed phase II trials in other cancersto test the effect of scheduling for these novel mAbs incombination with other modalities.
Another CTLA4 mAb, tremelimumab (previouslyticilimumab; Pfizer-MedImmune), suffered initial disap-
pointment after a phase III clinical trial in metastatic mela-noma was stopped early after failing to demonstratesuperiority over chemotherapy as first line treatment [26,27]. It recently demonstrated interesting activity in a smallphase II study (n = 20) of advanced hepatocellular carci-noma in chronic hepatitis C infection with a disease controlrate of 76.4% and 33% of patients experiencing stabledisease of more than 12 months duration. The median OSwas 7.5 months (95% CI 4.6, 18.0) [28]. While the exactmechanism of action of anti-CTLA4 mAbs remains a matterof discussion and research, their therapeutic activity andpotential for patient benefit is now available in the clinic.
The programmed death 1 (PD-1) receptor expressed byactivated T-cells is another key immune checkpoint recep-tor with a negative regulatory role when engaged by itsligands PD-L1 (also known as B7-H1) and PD-L2 (also
Tumour cell
PD-1
CTLA4
CD28
B7-1/2
TCR
MHC
CD40
CD40
CD27
OX40
4-1BB
BTLATIM3LAG3
LAG3
PD-1
CTLA4
PD-L1
TCR
MHC
T-cellactivation
Anti-CTLA-4
Anti-4-1BB
Anti-PD1
Anti-OX40
Anti-CD40
APC
Anti-CD40
Anti-CD27
Anti-LAG3Anti-TIM3 Anti-BTLA
Effector CTL
RegulatoryT-cell Cognate ligand binding induces
secretion of imunosuppressivecytokines, e.g. TGF-b
Anti-PD-L1
Legend:• Stimulatory receptor• Inhibitory receptor• Peptide• Agonist mAb• Antagonist mAb
Figure 2Schematic representation of the multiple immunomodulatory receptors that are potential therapeutic targets as monotherapy or combination therapy withimmunomodulatory mAbs. T-cell activation refers to the eventual activity of a complex network of different T-cell lineages (e.g. Th1, Th2, Th17, Treg etc)which can have differential expression of checkpoint regulators. APC, antigen presenting cells; CTL, cytotoxic T lymphocytes
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 237
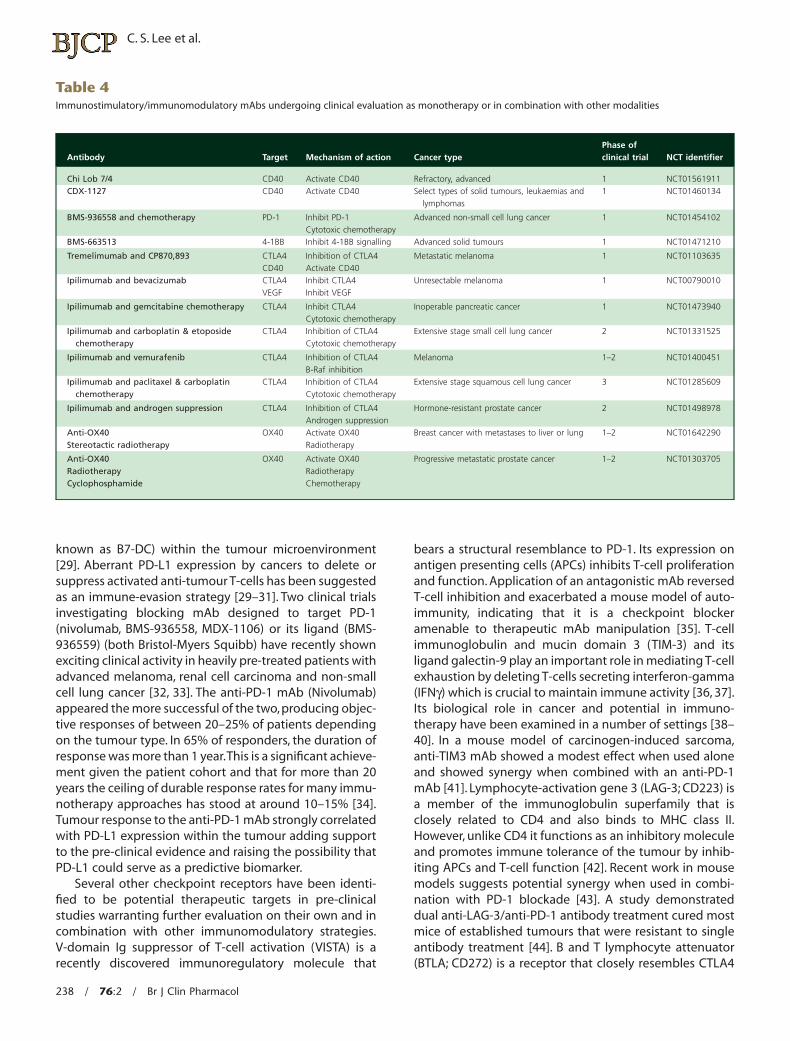
known as B7-DC) within the tumour microenvironment[29]. Aberrant PD-L1 expression by cancers to delete orsuppress activated anti-tumour T-cells has been suggestedas an immune-evasion strategy [29–31]. Two clinical trialsinvestigating blocking mAb designed to target PD-1(nivolumab, BMS-936558, MDX-1106) or its ligand (BMS-936559) (both Bristol-Myers Squibb) have recently shownexciting clinical activity in heavily pre-treated patients withadvanced melanoma, renal cell carcinoma and non-smallcell lung cancer [32, 33]. The anti-PD-1 mAb (Nivolumab)appeared the more successful of the two, producing objec-tive responses of between 20–25% of patients dependingon the tumour type. In 65% of responders, the duration ofresponse was more than 1 year.This is a significant achieve-ment given the patient cohort and that for more than 20years the ceiling of durable response rates for many immu-notherapy approaches has stood at around 10–15% [34].Tumour response to the anti-PD-1 mAb strongly correlatedwith PD-L1 expression within the tumour adding supportto the pre-clinical evidence and raising the possibility thatPD-L1 could serve as a predictive biomarker.
Several other checkpoint receptors have been identi-fied to be potential therapeutic targets in pre-clinicalstudies warranting further evaluation on their own and incombination with other immunomodulatory strategies.V-domain Ig suppressor of T-cell activation (VISTA) is arecently discovered immunoregulatory molecule that
bears a structural resemblance to PD-1. Its expression onantigen presenting cells (APCs) inhibits T-cell proliferationand function. Application of an antagonistic mAb reversedT-cell inhibition and exacerbated a mouse model of auto-immunity, indicating that it is a checkpoint blockeramenable to therapeutic mAb manipulation [35]. T-cellimmunoglobulin and mucin domain 3 (TIM-3) and itsligand galectin-9 play an important role in mediating T-cellexhaustion by deleting T-cells secreting interferon-gamma(IFNγ) which is crucial to maintain immune activity [36, 37].Its biological role in cancer and potential in immuno-therapy have been examined in a number of settings [38–40]. In a mouse model of carcinogen-induced sarcoma,anti-TIM3 mAb showed a modest effect when used aloneand showed synergy when combined with an anti-PD-1mAb [41]. Lymphocyte-activation gene 3 (LAG-3; CD223) isa member of the immunoglobulin superfamily that isclosely related to CD4 and also binds to MHC class II.However, unlike CD4 it functions as an inhibitory moleculeand promotes immune tolerance of the tumour by inhib-iting APCs and T-cell function [42]. Recent work in mousemodels suggests potential synergy when used in combi-nation with PD-1 blockade [43]. A study demonstrateddual anti-LAG-3/anti-PD-1 antibody treatment cured mostmice of established tumours that were resistant to singleantibody treatment [44]. B and T lymphocyte attenuator(BTLA; CD272) is a receptor that closely resembles CTLA4
Table 4Immunostimulatory/immunomodulatory mAbs undergoing clinical evaluation as monotherapy or in combination with other modalities
Antibody Target Mechanism of action Cancer typePhase ofclinical trial NCT identifier
Chi Lob 7/4 CD40 Activate CD40 Refractory, advanced 1 NCT01561911CDX-1127 CD40 Activate CD40 Select types of solid tumours, leukaemias and
lymphomas1 NCT01460134
BMS-936558 and chemotherapy PD-1 Inhibit PD-1Cytotoxic chemotherapy
Advanced non-small cell lung cancer 1 NCT01454102
BMS-663513 4-1BB Inhibit 4-1BB signalling Advanced solid tumours 1 NCT01471210
Tremelimumab and CP870,893 CTLA4CD40
Inhibition of CTLA4Activate CD40
Metastatic melanoma 1 NCT01103635
Ipilimumab and bevacizumab CTLA4VEGF
Inhibit CTLA4Inhibit VEGF
Unresectable melanoma 1 NCT00790010
Ipilimumab and gemcitabine chemotherapy CTLA4 Inhibit CTLA4Cytotoxic chemotherapy
Inoperable pancreatic cancer 1 NCT01473940
Ipilimumab and carboplatin & etoposidechemotherapy
CTLA4 Inhibition of CTLA4Cytotoxic chemotherapy
Extensive stage small cell lung cancer 2 NCT01331525
Ipilimumab and vemurafenib CTLA4 Inhibition of CTLA4B-Raf inhibition
Melanoma 1–2 NCT01400451
Ipilimumab and paclitaxel & carboplatinchemotherapy
CTLA4 Inhibition of CTLA4Cytotoxic chemotherapy
Extensive stage squamous cell lung cancer 3 NCT01285609
Ipilimumab and androgen suppression CTLA4 Inhibition of CTLA4Androgen suppression
Hormone-resistant prostate cancer 2 NCT01498978
Anti-OX40Stereotactic radiotherapy
OX40 Activate OX40Radiotherapy
Breast cancer with metastases to liver or lung 1–2 NCT01642290
Anti-OX40RadiotherapyCyclophosphamide
OX40 Activate OX40RadiotherapyChemotherapy
Progressive metastatic prostate cancer 1–2 NCT01303705
C. S. Lee et al.
238 / 76:2 / Br J Clin Pharmacol

and PD-1 [45]. It binds to the herpes virus entry mediator(HVEM). The interaction between BTLA and HVEM canfunction either as a stimulatory or inhibitory signaldepending on the context of other binding partners orcell-type expression [46, 47]. Recent work demonstrates itspotential role in maintaining immune-tolerance to self [45,48] and tumour, making it a potential target for therapy[49–51].
Agonists of positive regulators(immunostimulators)Immunostimulatory mAbs have agonistic properties onstimulatory immune receptors. Although not yet as clini-cally successful as ipilumumab, preclinical models showthat they also hold great promise, equal to that of thecheckpoint blockers. As discussed earlier, physiologicalligation of the TCR without CD28 engagement is insuffi-cient for full T-cell activation. However, a class of ‘super-agonist’ anti-CD28 mAbs has been developed which bindto a particular epitope in CD28 and can activate T-cellswithout concomitant TCR engagement. The initial clinicaltrial with superagonist anti-CD28 antibodies resulted inextremely rapid and dysregulated immune activation,leading to severe inflammatory reactions promoted by aso-called ‘cytokine storm’ (for further details see Eastwoodet al. [52]).
CD40 is expressed by APCs [B cells, dentritic cells(DCs), monocytes and macrophages], as well as non-haemopoietic tissues such as vascular endothelium andmultiple epithelia [53]. It is also often found on the surfaceof carcinoma and lymphoma cells [54]. The ligand, CD40L(CD154) is primarily expressed on activated CD4 T-cells andligand binding activates APCs [53, 54]. An intact CD40/CD40L system appears crucial for a normal B-cell humoralimmune response and DC activation. It fulfils critical func-tions in stimulating antigen presentation leading to immu-nity and priming of T-cells to drive full effector cytotoxic Tlymphocyte (CTL) responses [55, 56]. Incompletely acti-vated DCs cause T-cell anergy or depletion after an abor-tive transient activation [57]. As such, CD40 and DCactivation is a key regulator of many facets of immunestimulation (see Figure 2).
Agonist mAbs targeting CD40 have been shown tosubstitute for CD4 T-cell help by mimicking CD40L engage-ment [55, 56, 58] and can overcome T-cell tolerancein tumour-bearing mice, evoking highly effective CTLresponses, and augment the efficacy of anti-tumour vac-cines in pre-clinical models [59–62]. There are currentlyfour anti-CD40 mAbs undergoing early clinical trials;CP-870,893 (Pfizer), Chi Lob 7/4 (Cancer Research UKBiotherapeutics Development Unit), HCD122 (Novartis)and dacetuzumab (SGN-40, Seattle Genetics). All of theseappear relatively well-tolerated with some evidence ofgenerating objective or biochemical responses [63–66].However, their relative potency seems extremely wide-ranging with the maximum tolerated dose for CP-870,893
far lower than that for SGN-40 or Chi Lob 7/4. We [67] andothers [68] recently showed that interaction with theinhibitory Fc gamma receptor (FcγRIIB, CD32b) is critical forthe immunostimulatory and therapeutic activity of anti-CD40 mAb in mice. This new finding might explain thedifferences in mAb potency as immunoglobulin subclass(IgG 1, 2a, 2b, 3) have varying binding affinities to differentFc gamma receptors (FcγRI, FcγRIIB, FcγRIII and FcγRIV) [69].Currently it remains unclear how this will translate in thehuman system owing to our differences in IgG subclassand Fc receptor interactions from mice [69, 70]. This infor-mation will have significant implications for immuno-globulin selection in the design of the next generation ofimmunostimulatory mAbs if found to affect clinicalsuccess.
Interestingly, although much of the work in pre-clinicalmodels has indicated that anti-CD40 mAbs function byproducing effective CTL responses, a phase I pancreaticcancer trial of CP-870,893 in combination withgemcitabine showed therapeutic efficacy but suggestedan alternative mode of action. In this study,CD40 activationwas insufficient by itself to invoke an anti-tumour T-cellimmunity and instead the investigators concluded that themAb was re-educating macrophages to kill tumour andstromal cells [71].This highlights the role that innate immu-nity cells play in the tumour microenvironment and thepotential collateral effects these immunomodulatorymAbs might have on other cell types. For example, anti-CD40 mAbs could also directly target CD40 expressingtumour cells (e.g. lymphoma) causing apoptosis or deathvia ADCC or CDC and at the same time activate DCs whichmay be presenting tumour antigens to primed CD8 T-cells[54] (see Figure 2).
Other members of the TNFR superfamily are alsotargets with clinical potential for immunostimulatorymAbs, such as 4-1BB, OX40 and CD27. 4-1BB (CD137) is aco-stimulatory molecule that can function independentlyof CD28 [72]. It is stably expressed on activated T-cells aswell as other myeloid cells [73–76]. The cognate ligand(4-1BBL) is expressed on APCs like activated DCs, Bcells and macrophages. Signalling through 4-1BB/4-1BBLcauses T-cell expansion, cytokine induction and up-regulation of anti-apoptotic genes in order to preventactivation-induced cell death (AICD) of the expandingT-cells [77–79]. This permits downstream differentiation ofCTL as well as the establishment of long-lived memory[80]. Much remains to be understood of the biology andfunction of the 4-1BB/4-1BBL pathway as the result oftreatment with agonist anti-4-1BB mAbs is dependent onthe context. Anti-4-1BB mAbs have been shown to reduceB-cell numbers and function in an immunosuppressiverole against autoantibody production [81]. This paradoxi-cal effect has now been examined in a number of autoim-mune settings [82–85]. An early phase clinical trialexamining anti-cancer properties of an agonistic anti-4-1BB mAbs is on-going but early results testing a fully
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 239

humanized mAb, urelumab (BMS-663513, Bristol-MyersSquibb) in 83 patients with advanced cancers (54 mela-noma, 15 renal cell carcinoma, 13 ovarian cancer and oneprostate cancer) suggest activity at various dose rangeswith partial responses in three melanoma patients andstable disease in four patients in the expansion cohort. It iswell-tolerated but may cause reversible hepatic toxicity insome patients [86].
OX40 (CD134) is a TNF receptor expressed primarily onactivated CD4 and CD8 T-cells that transmits a potentco-stimulatory signal upon engagement of its ligandOX40L [87]. OX40L, the only known ligand for OX40, ismainly found on activated APCs [88–91]. In vivo, blockadeof OX40/OX40L interaction mitigates autoimmunity [92–94] and preclinical experiments confirmed enhancementof anti-tumour T-cell responses with OX40 agonist treat-ment [95]. An agonistic mouse mAb targeting OX40 hasnow undergone a first-in-man phase I dosing study in 30patients (NCT01644968). Limited data published so farsuggest the mouse mAb was tolerated in the study sub-jects (the maximum tolerated dose was not reached) withserological evidence of activation/proliferation of T-cells[96]. Clinical data published to date showed that twopatients (ovarian and melanoma) displayed stable diseasein the lowest dose cohort [97]. This approach will now betested in combination with radiotherapy in metastaticbreast cancer (NCT01642290) as well as in metastatic pros-tate cancer in combination with chemoradiotherapy(NCT01303705). As this is a mouse mAb, repeated dosingwill require a non-immunogenic humanized product ofwhich two are currently under development [96, 98].
CD27 is another TNF receptor that is expressed onT-cells, memory B cells, plasma cells and some natural killercells and engagement by its ligand CD70 leads to prolif-eration, survival and functional activation [99–102]. A fullyhuman agonistic mAb targeting CD27 (CDX-1127) hasbeen developed and has now entered a phase I study(NCT01460134).
It has become clear that the kinetics of clinicalresponses achieved with immunomodulatory mAbs areappreciably slower than those observed following chemo-therapy. Apparent initial tumour progression may beobserved, and it may be 2 to 3 months before regression isobserved. Responses, if seen, tend to be sustained. Earlyclinical trials using conventional early tumour responsetime points may underestimate the therapeutic effects ofthese agents and specific Immune Related Response Crite-ria have been proposed [103].
Toxicity: the dark side ofimmune stimulation
In comparison with most cytotoxic agents, mAbs are well-tolerated.Toxicities tend to be either acute hypersensitivityreactions at the time the mAb is infused, or delayed
immune-mediated toxicities, related to the mechanismof action of the mAb. Infusion reactions range from mildskin reactions, pyrexia and an influenza-like response, to, inrare instances, frank anaphylaxis, systemic inflammatoryresponse syndrome and life-threatening cytokine releasesyndrome. Close monitoring during infusions is required,but most such acute reactions are readily controlled withslowing or interruption of the infusion and appropriatesymptom management (see reference [104] for a clinicalmanagement review).
Immune related adverse events (IRAEs) as seen bypatients receiving ipilimumab are a new class of sideeffects particular to treatment using these novelimmunostimulatory or immunomodulatory mAbs. Osten-sibly this is due to a breakdown of immune self-tolerancealthough initial animal toxicology studies failed to predictsuch adverse effects. CTLA4 knock-out mice also didnot suffer from the common IRAEs in humans [105,106]. The most common serious manifestations includeenterocolitis with diarrhoea, hepatitis, dermatitis andendocrinopathies (hypophysitis, hypopituitarism, adrenalinsufficiency, hypothyroidism or hypogonadism). IRAEs arereported in up to 60% of patients treated with ipilimumab,with severe toxcities (grade 3 or 4) in about 10–15% ofpatients [23]. The clinical experience in the managementof IRAEs gained by investigators of ipilimumab in clinicaltrials have been published [107, 108] and managementalgorithms are also available at the FDA Risk Eliminationand Management System (REMS) website [109] or the UKElectronic Medicines Compendium website. The manage-ment strategy typically involves early detection of toxicity,interruption of mAb therapy, close clinical monitoring,early symptomatic relief and if appropriate, the timely useof corticosteroids to prevent rapid deterioration. Moderatetoxicities (grade 2) can usually be reversed successfullywith treatment interruption or with additional low dosecorticosteroids (prednisolone 0.5 mg kg−1 day−1 or equiva-lent) if symptoms do not resolve within a week. Severeor life-threatening (grade 3 or 4) toxicities require perma-nent cessation of mAb therapy and prompt treatmentwith high dose corticosteroid (prednisolone 1 to2 mg kg−1 day−1 or equivalent). The steroid should betapered gradually at least over 1 month as prematurewithdrawal may lead to a rebound inflammatory response[107]. Persistent inflammatory IRAEs may require addi-tional immune-suppression with infliximab, myco-phenolate or tacrolimus [107].
Perhaps not surprisingly, the occurrence of IRAEs hasbeen correlated with tumour reponse, with one studyreporting clinical responses in 33% of patients with grade3 or 4 IRAEs, and only 5% of patients with no evidence ofimmune toxicity [110]. Interestingly, clinical responseshave been maintained even when systemic immunosup-pression with corticosteroids has been required to treatIRAEs [111, 112]. The reason for this remains unclear and isthe subject of further investigation.
C. S. Lee et al.
240 / 76:2 / Br J Clin Pharmacol

Emerging effects of other immunecell types
Our discussion has so far focused on the effects on thesenovel mAbs on T-cells. However, other cell types also play arole in immune escape by interacting directly with thetumour cells or acting in tandem with other immunosup-pressive cells; the so-called immunosuppressive network[113]. These additional cell types include regulatory T-cells(Tregs), myeloid-derived suppressor cells (a heterogene-ous population of bone-marrow derived cells of myeloidlineage characterized by their potential to suppress T-cellresponses) and tumour associated macrophages. Theeffects of novel immunomodulatory mAbs on these otherimmune cell types would depend on their receptor-ligandexpression profile.CTLA4 is constitutively expressed on thecell surface of Tregs and functions as a potent mediator ofself-tolerance and immune homeostasis [114], which mayhelp explain the immune-related toxicity profile ofipilimumab. The potential activity of antagonistic CTLA4mAbs in depletion/inhibition of Tregs to remove Treg sup-pression and enhance antitumor immunity has been pro-posed. Murine models and early clinical studies proposingthis mechanism of action [115, 116] have not been borneout by more recent and larger clinical series [117–119]. Ithas also been proposed that OX-40 stimulation is requiredfor regulatory T-cell mediated control of colitis [120], whichhighlights the intriguing possibility of differential activa-tion by the agonistic OX-40 mAb on T-cell effector functionand T-cell regulation depending on organ/tumour loca-tion. As mentioned above CP-870,893 (anti CD40) acti-vated macrophages can lead to tumour cytolysis. Theexpression, distribution and function of the targets ofthese novel mAbs are still the subject of research whichwill shed further light on potential toxicities and currentlyunidentified mechanisms of actions. For example, a posi-tive co-stimulatory role for PD-L2 has been reportedwhereby PD-L2 deficient mice have impaired CTL functionwith increased tumour growth [121]. Another recentreport surprisingly demonstrated a favourable role in PD-1expressing T-cell infiltrates in human papilloma virus asso-ciated head and neck cancer [122].
Conclusion
The use of mAbs is firmly established as a treatment forcancer. The emergence of immunomodulatory mAbs as anovel class of drugs will see cancer immunotherapy takeon an increasingly important strategic role in cancer man-agement. They can be used to produce powerful tumour-specific T-cell responses with the hope of eradicatingestablished tumour and generating long term tumourimmunity. Our increasing understanding in the regulationof the immune system will inform future strategies inimmunomodulatory mAb therapies, including identifica-
tion of new targets and rational drug combinations.Already in pre-clinical models it has been demonstratedthat synergistic activity can be achieved through combina-tions of immunostimulatory and immunomodulatorymAbs [123, 124]. Potentially the more exciting role of theseagents could be in combination with small molecularinhibitors and/or cytotoxic chemotherapy agents whereclinical trials are already ongoing or are being planned[125, 126]. The activity of such agents combined with thedurable effects observed with the checkpoint blockers hasthe potential to achieve objective responses in a greaterproportion of patients than is currently the case.
Competing Interests
All authors have completed the Unified Competing Inter-est form at http://www.icmje.org/coi_disclosure.pdf anddeclare CSL has not received support from any organiza-tion for the submitted work, has no financial relationshipswith any organizations that might have an interest in thesubmitted work in the previous 3 years or any other rela-tionships or activities that could appear to have influencedthe submitted work. MC has not received support from anyorganization for the submitted work. He acts as a consult-ant to Bioinvent and Roche and has received institutionalsupport from Bioinvent for grants and patents. MG has notreceived support from any organization for the submittedwork. He receives an institutional grant from CancerResearch UK. He acts as a consultant to a number ofbiotech companies to write general antibody expertreports and he receives institutional payments and royal-ties from antibody patents and licenses. PJ has notreceived support from any organization for the submittedwork. He acts as a consultant to Roche and Pfizer and hasreceived payments for lectures from Millennium Takedaand Pfizer.
We thank Dr Juliet Gray and Dr Ruth French for theirhelpful suggestions and proof-reading of the manuscript.
REFERENCES
1 Levy R, Miller RA. Biological and clinical implications oflymphocyte hybridomas – tumor-therapy withmonoclonal-antibodies. Annu Rev Med 1983; 34: 107–16.
2 Lowder JN, Levy R. Monoclonal antibodies – therapeuticand diagnostic uses in malignancy. West J Med 1985; 143:810–8.
3 Schroff RW, Foon KA, Beatty SM, Oldham RK, Morgan AC Jr.Human anti-murine immunoglobulin responses in patientsreceiving monoclonal antibody therapy. Cancer Res 1985;45: 879–85.
4 Shawler DL, Bartholomew RM, Smith LM, Dillman RO.Human immune response to multiple injections of murinemonoclonal IgG. J Immunol 1985; 135: 1530–5.
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 241

5 Boulianne GL, Hozumi N, Shulman MJ. Production offunctional chimaeric mouse/human antibody. Nature 1984;312: 643–6.
6 Morrison SL, Johnson MJ, Herzenberg LA, Oi VT. Chimerichuman antibody molecules: mouse antigen-bindingdomains with human constant region domains. Proc NatlAcad Sci U S A 1984; 81: 6851–5.
7 Kuus-Reichel K, Grauer LS, Karavodin LM, Knott C,Krusemeier M, Kay NE. Will immunogenicity limit the use,efficacy, and future development of therapeuticmonoclonal antibodies? Clin Diagn Lab Immunol 1994; 1:365–72.
8 Jones PT, Dear PH, Foote J, Neuberger MS, Winter G.Replacing the complementarity-determining regions in ahuman antibody with those from a mouse. Nature 1986;321: 522–5.
9 Queen C, Schneider WP, Selick HE, Payne PW, Landolfi NF,Duncan JF, Avdalovic NM, Levitt M, Junghans RP,Waldmann TA. A humanized antibody that binds to theinterleukin 2 receptor. Proc Natl Acad Sci U S A 1989; 86:10029–33.
10 Winter G, Griffiths AD, Hawkins RE, Hoogenboom HR.Making antibodies by phage display technology. Annu RevImmunol 1994; 12: 433–55.
11 Low NM, Holliger PH, Winter G. Mimicking somatichypermutation: affinity maturation of antibodies displayedon bacteriophage using a bacterial mutator strain. J MolBiol 1996; 260: 359–68.
12 Bruggemann M, Spicer C, Buluwela L, Rosewell I, Barton S,Surani MA, Rabbitts TH. Human antibody production intransgenic mice: expression from 100 kb of the human IgHlocus. Eur J Immunol 1991; 21: 1323–6.
13 Mendez MJ, Green LL, Corvalan JR, Jia XC, Maynard-CurrieCE, Yang XD, Gallo ML, Louie DM, Lee DV, Erickson KL, LunaJ, Roy CM, Abderrahim H, Kirschenbaum F, Noguchi M,Smith DH, Fukushima A, Hales JF, Klapholz S, Finer MH,Davis CG, Zsebo KM, Jakobovits A. Functional transplant ofmegabase human immunoglobulin loci recapitulateshuman antibody response in mice. Nat Genet 1997; 15:146–56.
14 Lim SH, Beers SA, French RR, Johnson PW, Glennie MJ,Cragg MS. Anti-CD20 monoclonal antibodies: historical andfuture perspectives. Haematologica 2010; 95: 135–43.
15 Beers SA, Chan CH, French RR, Cragg MS, Glennie MJ. CD20as a target for therapeutic type I and II monoclonalantibodies. Semin Hematol 2010; 47: 107–14.
16 Valabrega G, Montemurro F, Aglietta M. Trastuzumab:mechanism of action, resistance and future perspectives inHER2-overexpressing breast cancer. Ann Oncol 2007; 18:977–84.
17 Spector NL, Blackwell KL. Understanding the mechanismsbehind trastuzumab therapy for human epidermal growthfactor receptor 2-positive breast cancer. J Clin Oncol 2009;27: 5838–47.
18 Tejani MA, Cohen RB, Mehra R. The contribution ofcetuximab in the treatment of recurrent and/or metastatichead and neck cancer. Biologics 2010; 4: 173–85.
19 Kawaguchi Y, Kono K, Mimura K, Sugai H, Akaike H, Fujii H.Cetuximab induces antibody-dependent cellularcytotoxicity against EGFR-expressing esophagealsquamous cell carcinoma. Int J Cancer 2007; 120: 781–7.
20 Hanahan D, Weinberg RA. Hallmarks of cancer: the nextgeneration. Cell 2011; 144: 646–74.
21 Drake CG, Jaffee E, Pardoll DM. Mechanisms of immuneevasion by tumors. Adv Immunol 2006; 90: 51–81.
22 Rosenberg SA, Spiess P, Lafreniere R. A new approach tothe adoptive immunotherapy of cancer withtumor-infiltrating lymphocytes. Science 1986; 233: 1318–21.
23 Hodi FS, O’Day SJ, McDermott DF, Weber RW, Sosman JA,Haanen JB, Gonzalez R, Robert C, Schadendorf D, Hassel JC,Akerley W, van den Eertwegh AJ, Lutzky J, Lorigan P, VaubelJM, Linette GP, Hogg D, Ottensmeier CH, Lebbe C, PeschelC, Quirt I, Clark JI, Wolchok JD, Weber JS, Tian J, Yellin MJ,Nichol GM, Hoos A, Urba WJ. Improved survival withipilimumab in patients with metastatic melanoma. N Engl JMed 2010; 363: 711–23.
24 Lynch TJ, Bondarenko I, Luft A, Serwatowski P, Barlesi F,Chacko R, Sebastian M, Neal J, Lu HL, Cuillerot JM, Reck M.Ipilimumab in combination with paclitaxel and carboplatinas first-line treatment in stage IIIB/IV non-small-cell lungcancer: results from a randomized, double-blind,multicenter phase II study. J Clin Oncol 2012; 30: 2046–54.
25 Reck M, Bondarenko I, Luft A, Serwatowski P, Barlesi F,Chacko R, Sebastian M, Lu H, Cuillerot JM, Lynch TJ.Ipilimumab in combination with paclitaxel and carboplatinas first-line therapy in extensive-disease-small-cell lungcancer: results from a randomized, double-blind,multicenter phase 2 trial. Ann Oncol 2013; 24: 75–83.
26 Ribas A. Clinical development of the anti-CTLA-4 antibodytremelimumab. Semin Oncol 2010; 37: 450–4.
27 Ribas A, Hauschild A, Kefford R, Gomez-Navarro J, Pavlov D,Marshall MA. Phase III, open-label, randomized, comparativestudy of tremelimumab (CP-675,206) and chemotherapy(temozolomide [TMZ] or dacarbazine [DTIC]) in patientswith advanced melanoma. ASCO Meeting Abstracts 2008;26 (Suppl. 15): LBA9011.
28 Melero I, Sangro B, Riezu-Boj JI, Inarrairaegui M, Lasarte JJ,Sarobe P, Larrea E, Prieto J. Abstract 4387: antiviral andantitumoral effects of the anti-CTLA4 agent tremelimumabin patients with hepatocellular carcinoma (HCC) andchronic hepatitis C virus (HCV) infection: results from aphase II clinical trial. Cancer Research 2012; 72 (Suppl. 8):4387.
29 Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, FliesDB, Roche PC, Lu J, Zhu G, Tamada K, Lennon VA, Celis E,Chen L. Tumor-associated B7-H1 promotes T-cell apoptosis:a potential mechanism of immune evasion. Nat Med 2002;8: 793–800.
30 Hamanishi J, Mandai M, Iwasaki M, Okazaki T, Tanaka Y,Yamaguchi K, Higuchi T, Yagi H, Takakura K, Minato N,Honjo T, Fujii S. Programmed cell death 1 ligand 1 andtumor-infiltrating CD8+ T lymphocytes are prognosticfactors of human ovarian cancer. Proc Natl Acad Sci U S A2007; 104: 3360–5.
C. S. Lee et al.
242 / 76:2 / Br J Clin Pharmacol

31 Zou W, Chen L. Inhibitory B7-family molecules in thetumour microenvironment. Nat Rev Immunol 2008; 8:467–77.
32 Topalian SL, Hodi FS, Brahmer JR, Gettinger SN, Smith DC,McDermott DF, Powderly JD, Carvajal RD, Sosman JA, AtkinsMB, Leming PD, Spigel DR, Antonia SJ, Horn L, Drake CG,Pardoll DM, Chen L, Sharfman WH, Anders RA, Taube JM,McMiller TL, Xu H, Korman AJ, Jure-Kunkel M, Agrawal S,McDonald D, Kollia GD, Gupta A, Wigginton JM, Sznol M.Safety, activity, and immune correlates of anti-PD-1antibody in cancer. N Engl J Med 2012; 366: 2443–54.
33 Brahmer JR, Tykodi SS, Chow LQ, Hwu WJ, Topalian SL, HwuP, Drake CG, Camacho LH, Kauh J, Odunsi K, Pitot HC, HamidO, Bhatia S, Martins R, Eaton K, Chen S, Salay TM, AlaparthyS, Grosso JF, Korman AJ, Parker SM, Agrawal S, GoldbergSM, Pardoll DM, Gupta A, Wigginton JM. Safety and activityof anti-PD-L1 antibody in patients with advanced cancer.N Engl J Med 2012; 366: 2455–65.
34 Ribas A, Butterfield LH, Glaspy JA, Economou JS. Currentdevelopments in cancer vaccines and cellularimmunotherapy. J Clin Oncol 2003; 21: 2415–32.
35 Wang L, Rubinstein R, Lines JL, Wasiuk A, Ahonen C, Guo Y,Lu LF, Gondek D, Wang Y, Fava RA, Fiser A, Almo S, NoelleRJ. VISTA, a novel mouse Ig superfamily ligand thatnegatively regulates T cell responses. J Exp Med 2011; 208:577–92.
36 Baitsch L, Baumgaertner P, Devevre E, Raghav SK, Legat A,Barba L, Wieckowski S, Bouzourene H, Deplancke B, RomeroP, Rufer N, Speiser DE. Exhaustion of tumor-specific CD8(+)T cells in metastases from melanoma patients. J Clin Invest2011; 121: 2350–60.
37 Zhu C, Anderson AC, Schubart A, Xiong H, Imitola J, KhourySJ, Zheng XX, Strom TB, Kuchroo VK. The Tim-3 ligandgalectin-9 negatively regulates T helper type 1 immunity.Nat Immunol 2005; 6: 1245–52.
38 Sakuishi K, Jayaraman P, Behar SM, Anderson AC, KuchrooVK. Emerging Tim-3 functions in antimicrobial and tumorimmunity. Trends Immunol 2011; 32: 345–9.
39 Anderson AC. Tim-3, a negative regulator of anti-tumorimmunity. Curr Opin Immunol 2012; 24: 213–6.
40 Ngiow SF, Teng MW, Smyth MJ. Prospects for TIM3-targetedantitumor immunotherapy. Cancer Res 2011; 71: 6567–71.
41 Ngiow SF, von Scheidt B, Akiba H, Yagita H, Teng MW,Smyth MJ. Anti-TIM3 antibody promotes T cellIFN-gamma-mediated antitumor immunity and suppressesestablished tumors. Cancer Res 2011; 71: 3540–51.
42 Sierro S, Romero P, Speiser DE. The CD4-like moleculeLAG-3, biology and therapeutic applications. Expert OpinTher Targets 2011; 15: 91–101.
43 Okazaki T, Okazaki IM, Wang J, Sugiura D, Nakaki F, YoshidaT, Kato Y, Fagarasan S, Muramatsu M, Eto T, Hioki K, Honjo T.PD-1 and LAG-3 inhibitory co-receptors act synergisticallyto prevent autoimmunity in mice. J Exp Med 2011; 208:395–407.
44 Woo SR, Turnis ME, Goldberg MV, Bankoti J, Selby M, NirschlCJ, Bettini ML, Gravano DM, Vogel P, Liu CL,
Tangsombatvisit S, Grosso JF, Netto G, Smeltzer MP, ChauxA, Utz PJ, Workman CJ, Pardoll DM, Korman AJ, Drake CG,Vignali DA. Immune inhibitory molecules LAG-3 and PD-1synergistically regulate T-cell function to promote tumoralimmune escape. Cancer Res 2012; 72: 917–27.
45 Watanabe N, Gavrieli M, Sedy JR, Yang J, Fallarino F, LoftinSK, Hurchla MA, Zimmerman N, Sim J, Zang X, Murphy TL,Russell JH, Allison JP, Murphy KM. BTLA is a lymphocyteinhibitory receptor with similarities to CTLA-4 and PD-1.Nat Immunol 2003; 4: 670–9.
46 Sedy JR, Gavrieli M, Potter KG, Hurchla MA, Lindsley RC,Hildner K, Scheu S, Pfeffer K, Ware CF, Murphy TL, MurphyKM. B and T lymphocyte attenuator regulates T cellactivation through interaction with herpesvirus entrymediator. Nat Immunol 2005; 6: 90–8.
47 Tamada K, Shimozaki K, Chapoval AI, Zhai Y, Su J, Chen SF,Hsieh SL, Nagata S, Ni J, Chen L. LIGHT, a TNF-like molecule,costimulates T cell proliferation and is required fordendritic cell-mediated allogeneic T cell response. JImmunol 2000; 164: 4105–10.
48 Oya Y, Watanabe N, Owada T, Oki M, Hirose K, Suto A,Kagami S, Nakajima H, Kishimoto T, Iwamoto I, Murphy TL,Murphy KM, Saito Y. Development of autoimmunehepatitis-like disease and production of autoantibodies tonuclear antigens in mice lacking B and T lymphocyteattenuator. Arthritis Rheum 2008; 58: 2498–510.
49 Derre L, Rivals JP, Jandus C, Pastor S, Rimoldi D, Romero P,Michielin O, Olive D, Speiser DE. BTLA mediates inhibitionof human tumor-specific CD8+ T cells that can be partiallyreversed by vaccination. J Clin Invest 2010; 120: 157–67.
50 Fourcade J, Sun Z, Pagliano O, Guillaume P, Luescher IF,Sander C, Kirkwood JM, Olive D, Kuchroo V, Zarour HM.CD8(+) T cells specific for tumor antigens can be rendereddysfunctional by the tumor microenvironment throughupregulation of the inhibitory receptors BTLA and PD-1.Cancer Res 2012; 72: 887–96.
51 Crawford A, Wherry EJ. Editorial: therapeutic potential oftargeting BTLA. J Leukoc Biol 2009; 86: 5–8.
52 Eastwood D, Bird C, Dilger P, Hockley J, Findlay L, Poole S,Thorpe SJ, Wadhwa M, Thorpe R, Stebbings R. Severity ofthe TGN1412 trial disaster cytokine storm correlated withIL-2 release. Br J Clin Pharmacol 2013; 76: 299–315.
53 Quezada SA, Jarvinen LZ, Lind EF, Noelle RJ. CD40/CD154interactions at the interface of tolerance and immunity.Annu Rev Immunol 2004; 22: 307–28.
54 Geldart T, Illidge T. Anti-CD 40 monoclonal antibody. LeukLymphoma 2005; 46: 1105–13.
55 Schoenberger SP, Toes RE, van der Voort EI, Offringa R,Melief CJ. T-cell help for cytotoxic T lymphocytes ismediated by CD40-CD40L interactions. Nature 1998; 393:480–3.
56 Ridge JP, Di Rosa F, Matzinger P. A conditioned dendriticcell can be a temporal bridge between a CD4+ T-helperand a T-killer cell. Nature 1998; 393: 474–8.
57 Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenicdendritic cells. Annu Rev Immunol 2003; 21: 685–711.
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 243

58 Bennett SR, Carbone FR, Karamalis F, Flavell RA, Miller JF,Heath WR. Help for cytotoxic-T-cell responses is mediatedby CD40 signalling. Nature 1998; 393: 478–80.
59 Khalil M, Vonderheide RH. Anti-CD40 agonist antibodies:preclinical and clinical experience. Update. Cancer Ther2007; 2: 61–5.
60 Sotomayor EM, Borrello I, Tubb E, Rattis FM, Bien H, Lu Z,Fein S, Schoenberger S, Levitsky HI. Conversion oftumor-specific CD4+ T-cell tolerance to T-cell primingthrough in vivo ligation of CD40. Nat Med 1999; 5: 780–7.
61 French RR, Chan HT, Tutt AL, Glennie MJ. CD40 antibodyevokes a cytotoxic T-cell response that eradicateslymphoma and bypasses T-cell help. Nat Med 1999; 5:548–53.
62 Diehl L, den Boer AT, Schoenberger SP, van der Voort EI,Schumacher TN, Melief CJ, Offringa R, Toes RE. CD40activation in vivo overcomes peptide-induced peripheralcytotoxic T-lymphocyte tolerance and augmentsanti-tumor vaccine efficacy. Nat Med 1999; 5: 774–9.
63 Vonderheide RH, Flaherty KT, Khalil M, Stumacher MS, BajorDL, Hutnick NA, Sullivan P, Mahany JJ, Gallagher M, KramerA, Green SJ, O’Dwyer PJ, Running KL, Huhn RD, Antonia SJ.Clinical activity and immune modulation in cancer patientstreated with CP-870,893, a novel CD40 agonist monoclonalantibody. J Clin Oncol 2007; 25: 876–83.
64 Advani R, Forero-Torres A, Furman RR, Rosenblatt JD,Younes A, Ren H, Harrop K, Whiting N, Drachman JG. Phase Istudy of the humanized anti-CD40 monoclonal antibodydacetuzumab in refractory or recurrent non-Hodgkin’slymphoma. J Clin Oncol 2009; 27: 4371–7.
65 Hussein M, Berenson JR, Niesvizky R, Munshi N, Matous J,Sobecks R, Harrop K, Drachman JG, Whiting N. A phase Imultidose study of dacetuzumab (SGN-40; humanizedanti-CD40 monoclonal antibody) in patients with multiplemyeloma. Haematologica 2010; 95: 845–8.
66 Byrd JC, Kipps TJ, Flinn IW, Cooper M, Odenike O, BendiskeJ, Rediske J, Bilic S, Dey J, Baeck J, O’Brien S. Phase I study ofthe anti-CD40 humanized monoclonal antibodylucatumumab (HCD122) in relapsed chronic lymphocyticleukemia. Leuk Lymphoma 2012; 53: 2136–42 [Epub2012/04/06].
67 White AL, Chan HT, Roghanian A, French RR, Mockridge CI,Tutt AL, Dixon SV, Ajona D, Verbeek JS, Al-Shamkhani A,Cragg MS, Beers SA, Glennie MJ. Interaction withFc{gamma}RIIB is critical for the agonistic activity ofanti-CD40 monoclonal antibody. J Immunol 2011; 187:1754–63.
68 Li F, Ravetch JV. Inhibitory Fcgamma receptor engagementdrives adjuvant and anti-tumor activities of agonistic CD40antibodies. Science 2011; 333: 1030–4.
69 Bruhns P. Properties of mouse and human IgG receptorsand their contribution to disease models. Blood 2012; 119:5640–9.
70 Bruhns P, Iannascoli B, England P, Mancardi DA, FernandezN, Jorieux S, Daeron M. Specificity and affinity of human
Fcgamma receptors and their polymorphic variants forhuman IgG subclasses. Blood 2009; 113: 3716–25.
71 Beatty GL, Chiorean EG, Fishman MP, Saboury B, TeitelbaumUR, Sun W, Huhn RD, Song W, Li D, Sharp LL, Torigian DA,O’Dwyer PJ, Vonderheide RH. CD40 agonists alter tumorstroma and show efficacy against pancreatic carcinoma inmice and humans. Science 2011; 331: 1612–6.
72 Saoulli K, Lee SY, Cannons JL, Yeh WC, Santana A, GoldsteinMD, Bangia N, DeBenedette MA, Mak TW, Choi Y, Watts TH.CD28-independent, TRAF2-dependent costimulation ofresting T cells by 4-1BB ligand. J Exp Med 1998; 187:1849–62.
73 Vinay DS, Kwon BS. 4-1BB signaling beyond T cells. Cell MolImmunol 2011; 8: 281–4.
74 Pollok KE, Kim YJ, Zhou Z, Hurtado J, Kim KK, Pickard RT,Kwon BS. Inducible T cell antigen 4-1BB. Analysis ofexpression and function. J Immunol 1993; 150: 771–81.
75 Melero I, Johnston JV, Shufford WW, Mittler RS, Chen L.NK1.1 cells express 4-1BB (CDw137) costimulatorymolecule and are required for tumor immunity elicited byanti-4-1BB monoclonal antibodies. Cell Immunol 1998; 190:167–72.
76 Wilcox RA, Chapoval AI, Gorski KS, Otsuji M, Shin T, Flies DB,Tamada K, Mittler RS, Tsuchiya H, Pardoll DM, Chen L.Cutting edge: expression of functional CD137 receptor bydendritic cells. J Immunol 2002; 168: 4262–7.
77 Vinay DS, Kwon BS. Role of 4-1BB in immune responses.Semin Immunol 1998; 10: 481–9.
78 Croft M. Co-stimulatory members of the TNFR family: keysto effective T-cell immunity? Nat Rev Immunol 2003; 3:609–20.
79 Watts TH. TNF/TNFR family members in costimulation of Tcell responses. Annu Rev Immunol 2005; 23: 23–68.
80 Myers L, Lee SW, Rossi RJ, Lefrancois L, Kwon BS, Mittler RS,Croft M, Vella AT. Combined CD137 (4-1BB) and adjuvanttherapy generates a developing pool of peptide-specificCD8 memory T cells. Int Immunol 2006; 18: 325–33.
81 Mittler RS, Bailey TS, Klussman K, Trailsmith MD, HoffmannMK. Anti-4-1BB monoclonal antibodies abrogate Tcell-dependent humoral immune responses in vivo throughthe induction of helper T cell anergy. J Exp Med 1999; 190:1535–40.
82 Seo SK, Choi JH, Kim YH, Kang WJ, Park HY, Suh JH, Choi BK,Vinay DS, Kwon BS. 4-1BB-mediated immunotherapy ofrheumatoid arthritis. Nat Med 2004; 10: 1088–94.
83 Sun Y, Chen HM, Subudhi SK, Chen J, Koka R, Chen L, Fu YX.Costimulatory molecule-targeted antibody therapy of aspontaneous autoimmune disease. Nat Med 2002; 8:1405–13.
84 Sun Y, Lin X, Chen HM, Wu Q, Subudhi SK, Chen L, Fu YX.Administration of agonistic anti-4-1BB monoclonalantibody leads to the amelioration of experimentalautoimmune encephalomyelitis. J Immunol 2002; 168:1457–65.
C. S. Lee et al.
244 / 76:2 / Br J Clin Pharmacol

85 Foell J, Strahotin S, O’Neil SP, McCausland MM, Suwyn C,Haber M, Chander PN, Bapat AS, Yan XJ, Chiorazzi N,Hoffmann MK, Mittler RS. CD137 costimulatory T cellreceptor engagement reverses acute disease inlupus-prone NZB x NZW F1 mice. J Clin Invest 2003; 111:1505–18.
86 Sznol M, Hodi FS, Margolin K, McDermott DF, Ernstoff MS,Kirkwood JM, Wojtaszek C, Feltquate D, Logan T. Phase Istudy of BMS-663513, a fully human anti-CD137 agonistmonoclonal antibody, in patients (pts) with advancedcancer (CA). ASCO Meeting Abstracts 2008; 26 (Suppl. 15):3007.
87 Croft M. Control of immunity by the TNFR-related moleculeOX40 (CD134). Annu Rev Immunol 2010; 28: 57–78.
88 Weinberg AD, Wegmann KW, Funatake C, Whitham RH.Blocking OX-40/OX-40 ligand interaction in vitro and in vivoleads to decreased T cell function and amelioration ofexperimental allergic encephalomyelitis. J Immunol 1999;162: 1818–26.
89 Nohara C, Akiba H, Nakajima A, Inoue A, Koh CS, OhshimaH, Yagita H, Mizuno Y, Okumura K. Amelioration ofexperimental autoimmune encephalomyelitis withanti-OX40 ligand monoclonal antibody: a critical role forOX40 ligand in migration, but not development, ofpathogenic T cells. J Immunol 2001; 166: 2108–15.
90 Chen AI, McAdam AJ, Buhlmann JE, Scott S, Lupher ML Jr,Greenfield EA, Baum PR, Fanslow WC, Calderhead DM,Freeman GJ, Sharpe AH. Ox40-ligand has a criticalcostimulatory role in dendritic cell:T cell interactions.Immunity 1999; 11: 689–98.
91 Stuber E, Neurath M, Calderhead D, Fell HP, Strober W.Cross-linking of OX40 ligand, a member of the TNF/NGFcytokine family, induces proliferation and differentiation inmurine splenic B cells. Immunity 1995; 2: 507–21.
92 Yoshioka T, Nakajima A, Akiba H, Ishiwata T, Asano G,Yoshino S, Yagita H, Okumura K. Contribution ofOX40/OX40 ligand interaction to the pathogenesis ofrheumatoid arthritis. Eur J Immunol 2000; 30: 2815–23.
93 Murata K, Nose M, Ndhlovu LC, Sato T, Sugamura K, Ishii N.Constitutive OX40/OX40 ligand interaction inducesautoimmune-like diseases. J Immunol 2002; 169: 4628–36.
94 Redmond WL, Weinberg AD. Targeting OX40 and OX40L forthe treatment of autoimmunity and cancer. Crit RevImmunol 2007; 27: 415–36.
95 Weinberg AD, Rivera MM, Prell R, Morris A, Ramstad T, VettoJT, Urba WJ, Alvord G, Bunce C, Shields J. Engagement ofthe OX-40 receptor in vivo enhances antitumor immunity.J Immunol 2000; 164: 2160–9.
96 Weinberg AD, Morris NP, Kovacsovics-Bankowski M, UrbaWJ, Curti BD. Science gone translational: the OX40 agoniststory. Immunol Rev 2011; 244: 218–31.
97 Curti BD, Weinberg AD, Morris NP, Walker EB, Justice L, RossHJ, Crocenzi TS, Urba WJ. A phase I trial of a monoclonalantibody to OX40 in patients with advanced cancer.J Immunother 2006; 29: 645. doi: 10.1097/01.cji.0000211343.73588.59.
98 Morris NP, Peters C, Montler R, Hu HM, Curti BD, Urba WJ,Weinberg AD. Development and characterization ofrecombinant human Fc:OX40L fusion protein linked via acoiled-coil trimerization domain. Mol Immunol 2007; 44:3112–21.
99 Hendriks J, Gravestein LA, Tesselaar K, van Lier RA,Schumacher TN, Borst J. CD27 is required for generationand long-term maintenance of T cell immunity. NatImmunol 2000; 1: 433–40.
100 Hendriks J, Xiao Y, Borst J. CD27 promotes survival ofactivated T cells and complements CD28 in generation andestablishment of the effector T cell pool. J Exp Med 2003;198: 1369–80.
101 Xiao Y, Hendriks J, Langerak P, Jacobs H, Borst J. CD27 isacquired by primed B cells at the centroblast stage andpromotes germinal center formation. J Immunol 2004; 172:7432–41.
102 De Colvenaer V, Taveirne S, Delforche M, De Smedt M,Vandekerckhove B, Taghon T, Boon L, Plum J, Leclercq G.CD27-deficient mice show normal NK-cell differentiationbut impaired function upon stimulation. Immunol Cell Biol2011; 89: 803–11.
103 Wolchok JD, Hoos A, O’Day S, Weber JS, Hamid O, Lebbe C,Maio M, Binder M, Bohnsack O, Nichol G, Humphrey R, HodiFS. Guidelines for the evaluation of immune therapyactivity in solid tumors: immune-related response criteria.Clin Cancer Res 2009; 15: 7412–20.
104 Lenz HJ. Management and preparedness for infusion andhypersensitivity reactions. Oncologist 2007; 12: 601–9.
105 Waterhouse P, Penninger JM, Timms E, Wakeham A,Shahinian A, Lee KP, Thompson CB, Griesser H, Mak TW.Lymphoproliferative disorders with early lethality in micedeficient in Ctla-4. Science 1995; 270: 985–8.
106 Tivol EA, Borriello F, Schweitzer AN, Lynch WP, BluestoneJA, Sharpe AH. Loss of CTLA-4 leads to massivelymphoproliferation and fatal multiorgan tissuedestruction, revealing a critical negative regulatory role ofCTLA-4. Immunity 1995; 3: 541–7.
107 Weber J. Review: anti-CTLA-4 antibody ipilimumab: casestudies of clinical response and immune-related adverseevents. Oncologist 2007; 12: 864–72.
108 Weber JS, Kahler KC, Hauschild A. Management ofimmune-related adverse events and kinetics of responsewith ipilimumab. J Clin Oncol 2012; 30: 2691–7.
109 Bristol-MyersSquibb. Risk evaluation and mitigationstrategy for Yervoy (Ipilimumab) on the risks of andrecommended management for severe immune-mediatedadverse reaction [Website]: Bristol-Myers Squibb; 2011.Available at http://www.fda.gov/downloads/Drugs/DrugSafety/PostmarketDrugSafetyInformationforPatientsandProviders/UCM249435.pdf (last accessed 6 June 2012).
110 Attia P, Phan GQ, Maker AV, Robinson MR, Quezado MM,Yang JC, Sherry RM, Topalian SL, Kammula US, Royal RE,Restifo NP, Haworth LR, Levy C, Mavroukakis SA, Nichol G,Yellin MJ, Rosenberg SA. Autoimmunity correlates with
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 245

tumor regression in patients with metastatic melanomatreated with anti-cytotoxic T-lymphocyte antigen-4. J ClinOncol 2005; 23: 6043–53.
111 Amin A, DePril V, Hamid O, Wolchock J, Maio M, Neyns B,Chin K, Ibrahim R, Hoos A, O’Day S. Evaluation of the effectof systemic corticosteroids for the treatment ofimmune-related adverse events (irAEs) on thedevelopment or maintenance of ipilimumab clinicalactivity. J Clin Oncol 2009; 27 (15 Suppl.); abstr 9037.
112 Harmankaya K, Erasim C, Koelblinger C, Ibrahim R, Hoos A,Pehamberger H, Binder M. Continuous systemiccorticosteroids do not affect the ongoing regression ofmetastatic melanoma for more than two years followingipilimumab therapy. Med Oncol 2011; 28: 1140–4.
113 Zou W. Immunosuppressive networks in the tumourenvironment and their therapeutic relevance. Nat RevCancer 2005; 5: 263–74.
114 Friedline RH, Brown DS, Nguyen H, Kornfeld H, Lee J, ZhangY, Appleby M, Der SD, Kang J, Chambers CA. CD4(+)regulatory T cells require CTLA-4 for the maintenance ofsystemic tolerance. J Exp Med 2009; 206: 421–34.
115 O’Mahony D, Morris JC, Quinn C, Gao W, Wilson WH, GauseB, Pittaluga S, Neelapu S, Brown M, Fleisher TA, Gulley JL,Schlom J, Nussenblatt R, Albert P, Davis TA, Lowy I, PetrusM, Waldmann TA, Janik JE. A pilot study of CTLA-4 blockadeafter cancer vaccine failure in patients with advancedmalignancy. Clin Cancer Res 2007; 13: 958–64.
116 Hodi FS, Mihm MC, Soiffer RJ, Haluska FG, Butler M, SeidenMV, Davis T, Henry-Spires R, MacRae S, Willman A, Padera R,Jaklitsch MT, Shankar S, Chen TC, Korman A, Allison JP,Dranoff G. Biologic activity of cytotoxic Tlymphocyte-associated antigen 4 antibody blockade inpreviously vaccinated metastatic melanoma and ovariancarcinoma patients. Proc Natl Acad Sci U S A 2003; 100:4712–7.
117 Weber JS, Hamid O, Chasalow SD, Wu DY, Parker SM,Galbraith S, Gnjatic S, Berman D. Ipilimumab increasesactivated T cells and enhances humoral immunity inpatients with advanced melanoma. J Immunother 2012; 35:89–97.
118 Hamid O, Schmidt H, Nissan A, Ridolfi L, Aamdal S, HanssonJ, Guida M, Hyams D, Gomez H, Bastholt L, Chasalow S,Berman D. A prospective phase II trial exploring theassociation between tumor microenvironment biomarkersand clinical activity of ipilimumab in advanced melanoma.J Transl Med 2011; 9: 204.
119 Graziani G, Tentori L, Navarra P. Ipilimumab: a novelimmunostimulatory monoclonal antibody for thetreatment of cancer. Pharmacol Res 2012; 65: 9–22.
120 Griseri T, Asquith M, Thompson C, Powrie F. OX40 isrequired for regulatory T cell-mediated control of colitis.J Exp Med 2010; 207: 699–709.
121 Shin T, Yoshimura K, Crafton EB, Tsuchiya H, Housseau F,Koseki H, Schulick RD, Chen L, Pardoll DM. In vivocostimulatory role of B7-DC in tuning T helper cell 1 andcytotoxic T lymphocyte responses. J Exp Med 2005; 201:1531–41.
122 Badoual C, Hans S, Merillon N, Van Ryswick C, Ravel P,Benhamouda N, Levionnois E, Nizard M, Si-Mohamed A,Besnier N, Gey A, Rotem-Yehudar R, Pere H, Tran T, GuerinCL, Chauvat A, Dransart E, Alanio C, Albert S, Barry B,Sandoval F, Quintin-Colonna F, Bruneval P, Fridman WH,Lemoine FM, Oudard S, Johannes L, Olive D, Brasnu D,Tartour E. PD-1-expressing tumor-infiltrating T cells are afavorable prognostic biomarker in HPV-associated headand neck cancer. Cancer Res 2013; 73: 128–38.
123 Gray JC, French RR, James S, Al-Shamkhani A, Johnson PW,Glennie MJ. Optimising anti-tumour CD8 T-cell responsesusing combinations of immunomodulatory antibodies. EurJ Immunol 2008; 38: 2499–511.
124 Curran MA, Kim M, Montalvo W, Al-Shamkhani A, Allison JP.Combination CTLA-4 blockade and 4-1BB activationenhances tumor rejection by increasing T-cell infiltration,proliferation, and cytokine production. PLoS One 2011; 6:e19499.
125 Mellman I, Coukos G, Dranoff G. Cancer immunotherapycomes of age. Nature 2011; 480: 480–9.
126 Vanneman M, Dranoff G. Combining immunotherapy andtargeted therapies in cancer treatment. Nat Rev Cancer2012; 12: 237–51.
127 Slamon DJ, Leyland-Jones B, Shak S, Fuchs H, Paton V,Bajamonde A, Fleming T, Eiermann W, Wolter J, Pegram M,Baselga J, Norton L. Use of chemotherapy plus amonoclonal antibody against HER2 for metastatic breastcancer that overexpresses HER2. N Engl J Med 2001; 344:783–92.
128 Bang YJ, Van Cutsem E, Feyereislova A, Chung HC, Shen L,Sawaki A, Lordick F, Ohtsu A, Omuro Y, Satoh T, Aprile G,Kulikov E, Hill J, Lehle M, Ruschoff J, Kang YK. Trastuzumabin combination with chemotherapy versus chemotherapyalone for treatment of HER2-positive advanced gastric orgastro-oesophageal junction cancer (ToGA): a phase 3,open-label, randomised controlled trial. Lancet 2010; 376:687–97.
129 Vermorken JB, Mesia R, Rivera F, Remenar E, Kawecki A,Rottey S, Erfan J, Zabolotnyy D, Kienzer HR, Cupissol D,Peyrade F, Benasso M, Vynnychenko I, De Raucourt D,Bokemeyer C, Schueler A, Amellal N, Hitt R. Platinum-basedchemotherapy plus cetuximab in head and neck cancer. NEngl J Med 2008; 359: 1116–27.
130 Van Cutsem E, Kohne CH, Hitre E, Zaluski J, Chang Chien CR,Makhson A, D’Haens G, Pinter T, Lim R, Bodoky G, Roh JK,Folprecht G, Ruff P, Stroh C, Tejpar S, Schlichting M,Nippgen J, Rougier P. Cetuximab and chemotherapy asinitial treatment for metastatic colorectal cancer. N Engl JMed 2009; 360: 1408–17.
131 Cunningham D, Humblet Y, Siena S, Khayat D, Bleiberg H,Santoro A, Bets D, Mueser M, Harstrick A, Verslype C, Chau I,Van Cutsem E. Cetuximab monotherapy and cetuximabplus irinotecan in irinotecan-refractory metastaticcolorectal cancer. N Engl J Med 2004; 351: 337–45.
132 Van Cutsem E, Peeters M, Siena S, Humblet Y, Hendlisz A,Neyns B, Canon JL, Van Laethem JL, Maurel J, Richardson G,Wolf M, Amado RG. Open-label phase III trial of
C. S. Lee et al.
246 / 76:2 / Br J Clin Pharmacol

panitumumab plus best supportive care compared withbest supportive care alone in patients withchemotherapy-refractory metastatic colorectal cancer.J Clin Oncol 2007; 25: 1658–64.
133 Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T,Hainsworth J, Heim W, Berlin J, Baron A, Griffing S,Holmgren E, Ferrara N, Fyfe G, Rogers B, Ross R, KabbinavarF. Bevacizumab plus irinotecan, fluorouracil, and leucovorinfor metastatic colorectal cancer. N Engl J Med 2004; 350:2335–42.
134 Sandler A, Gray R, Perry MC, Brahmer J, Schiller JH, DowlatiA, Lilenbaum R, Johnson DH. Paclitaxel-carboplatin alone orwith bevacizumab for non-small-cell lung cancer. N Engl JMed 2006; 355: 2542–50.
135 Miller K, Wang M, Gralow J, Dickler M, Cobleigh M, Perez EA,Shenkier T, Cella D, Davidson NE. Paclitaxel plusbevacizumab versus paclitaxel alone for metastatic breastcancer. N Engl J Med 2007; 357: 2666–76.
136 Escudier B, Pluzanska A, Koralewski P, Ravaud A, Bracarda S,Szczylik C, Chevreau C, Filipek M, Melichar B, Bajetta E,Gorbunova V, Bay JO, Bodrogi I, Jagiello-Gruszfeld A, MooreN. Bevacizumab plus interferon alfa-2a for treatment ofmetastatic renal cell carcinoma: a randomised, double-blindphase III trial. Lancet 2007; 370: 2103–11.
137 Kreisl TN, Kim L, Moore K, Duic P, Royce C, Stroud I, GarrenN, Mackey M, Butman JA, Camphausen K, Park J, Albert PS,Fine HA. Phase II trial of single-agent bevacizumabfollowed by bevacizumab plus irinotecan at tumorprogression in recurrent glioblastoma. J Clin Oncol 2009;27: 740–5.
138 Friedman HS, Prados MD, Wen PY, Mikkelsen T, Schiff D,Abrey LE, Yung WK, Paleologos N, Nicholas MK, Jensen R,Vredenburgh J, Huang J, Zheng M, Cloughesy T.Bevacizumab alone and in combination with irinotecan inrecurrent glioblastoma. J Clin Oncol 2009; 27: 4733–40.
139 Perren TJ, Swart AM, Pfisterer J, Ledermann JA,Pujade-Lauraine E, Kristensen G, Carey MS, Beale P,Cervantes A, Kurzeder C, du Bois A, Sehouli J, Kimmig R,Stahle A, Collinson F, Essapen S, Gourley C, Lortholary A,Selle F, Mirza MR, Leminen A, Plante M, Stark D, Qian W,Parmar MK, Oza AM. A phase 3 trial of bevacizumab inovarian cancer. N Engl J Med 2011; 365: 2484–96.
140 Stopeck AT, Lipton A, Body JJ, Steger GG, Tonkin K, de BoerRH, Lichinitser M, Fujiwara Y, Yardley DA, Viniegra M, Fan M,Jiang Q, Dansey R, Jun S, Braun A. Denosumab comparedwith zoledronic acid for the treatment of bone metastasesin patients with advanced breast cancer: a randomized,double-blind study. J Clin Oncol 2010; 28: 5132–9.
141 Coiffier B, Lepage E, Briere J, Herbrecht R, Tilly H,Bouabdallah R, Morel P, Van Den Neste E, Salles G, GaulardP, Reyes F, Lederlin P, Gisselbrecht C. CHOP chemotherapy
plus rituximab compared with CHOP alone in elderlypatients with diffuse large-B-cell lymphoma. N Engl J Med2002; 346: 235–42.
142 McLaughlin P, Grillo-Lopez AJ, Link BK, Levy R, CzuczmanMS, Williams ME, Heyman MR, Bence-Bruckler I, White CA,Cabanillas F, Jain V, Ho AD, Lister J, Wey K, Shen D, DallaireBK. Rituximab chimeric anti-CD20 monoclonal antibodytherapy for relapsed indolent lymphoma: half of patientsrespond to a four-dose treatment program. J Clin Oncol1998; 16: 2825–33.
143 Lundin J, Kimby E, Bjorkholm M, Broliden PA, Celsing F,Hjalmar V, Mollgard L, Rebello P, Hale G, Waldmann H,Mellstedt H, Osterborg A. Phase II trial of subcutaneousanti-CD52 monoclonal antibody alemtuzumab(Campath-1H) as first-line treatment for patients with B-cellchronic lymphocytic leukemia (B-CLL). Blood 2002; 100:768–73.
144 Wierda WG, Kipps TJ, Mayer J, Stilgenbauer S, Williams CD,Hellmann A, Robak T, Furman RR, Hillmen P, Trneny M, DyerMJ, Padmanabhan S, Piotrowska M, Kozak T, Chan G, DavisR, Losic N, Wilms J, Russell CA, Osterborg A. Ofatumumab assingle-agent CD20 immunotherapy infludarabine-refractory chronic lymphocytic leukemia. J ClinOncol 2010; 28: 1749–55.
145 Witzig TE, Gordon LI, Cabanillas F, Czuczman MS,Emmanouilides C, Joyce R, Pohlman BL, Bartlett NL,Wiseman GA, Padre N, Grillo-Lopez AJ, Multani P, White CA.Randomized controlled trial of yttrium-90-labeledibritumomab tiuxetan radioimmunotherapy versusrituximab immunotherapy for patients with relapsed orrefractory low-grade, follicular, or transformed B-cellnon-Hodgkin’s lymphoma. J Clin Oncol 2002; 20: 2453–63.
146 Younes A, Gopal AK, Smith SE, Ansell SM, Rosenblatt JD,Savage KJ, Ramchandren R, Bartlett NL, Cheson BD, de VosS, Forero-Torres A, Moskowitz CH, Connors JM, Engert A,Larsen EK, Kennedy DA, Sievers EL, Chen R. Results of apivotal phase II study of brentuximab vedotin for patientswith relapsed or refractory Hodgkin’s lymphoma. J ClinOncol 2012; 30: 2183–9.
147 Kaminski MS, Estes J, Zasadny KR, Francis IR, Ross CW, TuckM, Regan D, Fisher S, Gutierrez J, Kroll S, Stagg R, TidmarshG, Wahl RL. Radioimmunotherapy with iodine (131)Itositumomab for relapsed or refractory B-cell non-Hodgkinlymphoma: updated results and long-term follow-up of theUniversity of Michigan experience. Blood 2000; 96:1259–66.
148 Sievers EL, Appelbaum FR, Spielberger RT, Forman SJ,Flowers D, Smith FO, Shannon-Dorcy K, Berger MS,Bernstein ID. Selective ablation of acute myeloid leukemiausing antibody-targeted chemotherapy: a phase I study ofan anti-CD33 calicheamicin immunoconjugate. Blood 1999;93: 3678–84.
Novel antibodies targeting immune regulatory checkpoints for cancer therapy
Br J Clin Pharmacol / 76:2 / 247
Related Documents