Photosynthesis Research 67: 185–197, 2001. © 2001 Kluwer Academic Publishers. Printed in the Netherlands. 185 Regular paper Non-photochemical chlorophyll fluorescence quenching and structural rearrangements induced by low pH in intact cells of Chlorella fusca (Chlorophyceae) and Mantoniella squamata (Prasinophyceae) Reimund Goss 1,∗ & Gyözö Garab 2 1 Institut für Botanik, Universität Leipzig, Johannisallee 21-23, D-04103 Leipzig, Germany; 2 Institute of Plant Biology, Biological Research Center, Hungarian Academy of Sciences, Szeged, P.O. Box 521, H-6701 Szeged, Hungary; ∗ Author for correspondence Received 1 May 2000; accepted in revised form 30 November 2000 Key words: chiral macrodomains, circular dichroism, light adaptation, low-pH-induced fluorescence quenching, non-photochemical quenching of chlorophyll fluorescence, structural flexibility, xanthophyll cycle Abstract We have used circular dichroism (CD) spectroscopy and chlorophyll fluorescence induction measurements in order to examine low-pH-induced changes in the chiral macro-organization of the chromophores and in the efficiency of non-photochemical quenching of the chlorophyll a fluorescence (NPQ) in intact, dark-adapted cells of Chlorella fusca (Chlorophyceae) and Mantoniella squamata (Prasinophyceae). We found that: (i) high proton concentrations enhanced the formation of chiral macrodomains of the complexes, i.e. the formation of large aggregates with long-range chiral order of pigment dipoles; this was largely independent of the low-pH-induced accumulation of de-epoxidized xanthophylls; (ii) lowering the pH led to NPQ; however, efficient energy dissipation, in the absence of excess light, could only be achieved if a substantial part of violaxanthin was converted to zeaxanthin and antheraxanthin in Chlorella and Mantoniella, respectively; (iii) the low-pH-induced changes in the chiral macro-organization of pigments were fully reversed by titrating the cells to neutral pH; (iv) at neutral pH, the presence of antheraxanthin or zeaxanthin did not bring about a sizeable NPQ. Hence, low-pH-induced NPQ in dark adapted algal cells appears to be associated both with the presence of de-epoxidized xanthophylls and struc- tural changes in the chiral macrodomains. It is proposed that the macrodomains, by providing a suitable structure for long-distance migration of the excitation energy, in the presence of quenchers associated with de-epoxidized xanthophylls, facilitate significantly the dissipation of unused excitation energy. Abbreviations: CD – circular dichroism; Chl – chlorophyll; Fm – maximum fluorescence with all PS II reaction centres closed; Fo – minimum fluorescence with all PS II reaction centres in the open state; Fv – variable part of the Chl a fluorescence; LHC – light-harvesting complex; LHC II – the main Chl a/b light-harvesting complex of PS II; NPQ – non-photochemical quenching of the Chl a fluorescence; PAR – photosynthetically active radiation; PFD – photon flux density; PS II – Photosystem II; psi – polymerization or salt induced Introduction Higher plants and algae have developed short-term protection mechanisms against over-excitation of the photosynthetic apparatus in high light (for reviews see Horton et al. 1996 and Demmig-Adams and Adams 1996). Changes in the pigmentation of light- harvesting proteins in combination with conforma- tional changes, that are induced by the build-up of a proton gradient, have been proposed to switch the antenna system from a light-harvesting state to a state that effectively dissipates excitation energy to heat (Horton et al. 1996). In this process of en- hanced thermal dissipation in the antenna system,

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Photosynthesis Research 67: 185–197, 2001.© 2001 Kluwer Academic Publishers. Printed in the Netherlands.
185
Regular paper
Non-photochemical chlorophyll fluorescence quenching and structuralrearrangements induced by low pH in intact cells of Chlorella fusca(Chlorophyceae) and Mantoniella squamata (Prasinophyceae)
Reimund Goss1,∗ & Gyözö Garab2
1Institut für Botanik, Universität Leipzig, Johannisallee 21-23, D-04103 Leipzig, Germany; 2Institute of PlantBiology, Biological Research Center, Hungarian Academy of Sciences, Szeged, P.O. Box 521, H-6701 Szeged,Hungary; ∗Author for correspondence
Received 1 May 2000; accepted in revised form 30 November 2000
Key words: chiral macrodomains, circular dichroism, light adaptation, low-pH-induced fluorescence quenching,non-photochemical quenching of chlorophyll fluorescence, structural flexibility, xanthophyll cycle
Abstract
We have used circular dichroism (CD) spectroscopy and chlorophyll fluorescence induction measurements in orderto examine low-pH-induced changes in the chiral macro-organization of the chromophores and in the efficiency ofnon-photochemical quenching of the chlorophyll a fluorescence (NPQ) in intact, dark-adapted cells of Chlorellafusca (Chlorophyceae) and Mantoniella squamata (Prasinophyceae). We found that: (i) high proton concentrationsenhanced the formation of chiral macrodomains of the complexes, i.e. the formation of large aggregates withlong-range chiral order of pigment dipoles; this was largely independent of the low-pH-induced accumulationof de-epoxidized xanthophylls; (ii) lowering the pH led to NPQ; however, efficient energy dissipation, in theabsence of excess light, could only be achieved if a substantial part of violaxanthin was converted to zeaxanthinand antheraxanthin in Chlorella and Mantoniella, respectively; (iii) the low-pH-induced changes in the chiralmacro-organization of pigments were fully reversed by titrating the cells to neutral pH; (iv) at neutral pH, thepresence of antheraxanthin or zeaxanthin did not bring about a sizeable NPQ. Hence, low-pH-induced NPQ indark adapted algal cells appears to be associated both with the presence of de-epoxidized xanthophylls and struc-tural changes in the chiral macrodomains. It is proposed that the macrodomains, by providing a suitable structurefor long-distance migration of the excitation energy, in the presence of quenchers associated with de-epoxidizedxanthophylls, facilitate significantly the dissipation of unused excitation energy.
Abbreviations: CD – circular dichroism; Chl – chlorophyll; Fm – maximum fluorescence with all PS II reactioncentres closed; Fo – minimum fluorescence with all PS II reaction centres in the open state; Fv – variable part ofthe Chl a fluorescence; LHC – light-harvesting complex; LHC II – the main Chl a/b light-harvesting complex ofPS II; NPQ – non-photochemical quenching of the Chl a fluorescence; PAR – photosynthetically active radiation;PFD – photon flux density; PS II – Photosystem II; psi – polymerization or salt induced
Introduction
Higher plants and algae have developed short-termprotection mechanisms against over-excitation of thephotosynthetic apparatus in high light (for reviewssee Horton et al. 1996 and Demmig-Adams andAdams 1996). Changes in the pigmentation of light-
harvesting proteins in combination with conforma-tional changes, that are induced by the build-up ofa proton gradient, have been proposed to switch theantenna system from a light-harvesting state to astate that effectively dissipates excitation energy toheat (Horton et al. 1996). In this process of en-hanced thermal dissipation in the antenna system,

186
de-epoxidation of epoxidized xanthophylls via the op-eration of xanthophyll cycles has been shown to playan important role (for reviews see Demmig-Adams1990; Pfündel and Bilger 1994; Gilmore 1997). Thishas been thoroughly documented mainly by the aidof chlorophyll (Chl) fluorescence induction measure-ments (Demmig-Adams et al. 1990; Gilmore andYamamoto 1993; Goss et al. 1995) and analysis ofultrafast fluorescence decay kinetics (Gilmore et al.1995; Wagner et al. 1996; Richter et al. 1999). Re-cently, by using mutants with substantially reducedquenching capability, it has been clearly demonstratedthat the �pH- and zeaxanthin-dependent quenchingof the excitation energy is associated with a con-formational change of the thylakoid membranes (Liet al. 2000). This, and earlier data, underscorethe significance of structural rearrangements in non-photochemical quenching of the Chl a fluorescence(NPQ). However, at present, relatively little is knownabout the structural basis of the thermal dissipation,and our knowledge concerning the nature and mech-anism(s) of the structural rearrangements in the chro-mophoric system, that evidently accompany the al-ternations in the photophysical functions (i.e. energyutilization vs energy dissipation), is limited. In order toelucidate these questions and ultimately the physicalmechanism(s) of NPQ, investigations that provide in-formation on the molecular organization and structuralflexibility of the pigment system may be of particularimportance.
Structure and structural flexibility of the pigmentsystem in thylakoids can be investigated with the aidof circular dichroism (CD) spectroscopy (reviewed byGarab 1996). For photosynthetic pigments, CD canoriginate (i) from asymmetry in the molecular struc-ture; the so-called intrinsic CD of most photosyntheticpigments is very weak; (ii) short-range interactionsbetween different pigment molecules usually give riseto excitonic CD bands; these have provided importantinformation on the molecular architecture of differ-ent pigment–protein complexes (see e.g. Van Metter1977; Gülen et al. 1997); (iii) in large, densely packed,chirally organized aggregates an additional CD sig-nal arises, which carries physical information on thesize and long-range chiral order of extended arrays ofchromophores (Keller and Bustamante 1986a,b; Garab1996).
CD measurements have revealed that light-harvesting proteins can exist in different aggregationstates in their native membranes (Garab et al. 1988a,c).In hypotonic medium, at low ionic strength, Photosys-
tem II (PS II) particles exists in a non-aggregated form.This is reflected in the CD spectrum by a conservativeband structure which contains CD bands given riseby excitonic interactions between Chl a, Chl b andcarotenoid molecules (Garab et al. 1991; Barzda etal. 1994). In intact granal thylakoids, PS II particles,because of the high self-aggregation of LHC II, havebeen shown to form densely packed aggregates withintensely interacting chromophores that are arrangedin a long-range chiral order, and possess dimensions(d = 200–400 nm) commensurate with the wavelengthof the visible light. These chiral macrodomains ex-hibit very intense, anomalously shaped bands withlong tails outside the principal absorbance (Garab etal. 1988a; Finzi et al. 1989; Barzda et al. 1994). Theresulting psi-type CD is superimposed on the excitonicCD signal of individual complexes and particles. Thistype of macro-organization of the complexes has beenshown to be associated with the granal ultrastructureof chloroplasts, and to play a key role in the lateral se-gregation (‘sorting’) of the two photosystems betweenthe stacked and unstacked regions (Garab et al. 1991;Garab and Mustárdy 1999). In isolated lamellar ag-gregates of LHC II, it has also been shown that inthe macroarrays, the excitation energy can migrate forlong distances (r > 64 nm) (Barzda et al. 1996b).
CD spectroscopy also has revealed dynamicchanges in the chiral macro-organization of thelight-harvesting system: Intact thylakoid membraneshave been shown to exhibit light-induced, reversiblechanges in all major psi-type CD bands (�CD) (Garabet al. 1988b). Macroaggregation of LHC II in vivoand its modulation by light has been proposed to playa regulatory role in the energy-dependent NPQ, qE(Horton et al. 1991; Horton 1999). As for the moredirect correlation between the light-induced changesin the chiral macro-organization and NPQ, it has beenshown that �CD is associated with a transient, non-photochemical quenching of the Chl a fluorescence(Barzda et al. 1996a). In addition, �CD has beenshown to be blocked by inhibitors of qE (Istokovicset al. 1997). These observations, despite some differ-ences in other features, strongly suggest that the struc-tural rearrangements in the chiral macro-organizationof the light-harvesting complexes play a significantrole in the regulation of energy utilization in the an-tenna system. Concerning the role of de-epoxidizedxanthophylls in the structural changes, the correlationis less evident. �CD has been observed in the ab-sence of zeaxanthin (cf. the experimental conditions inGarab et al. 1988b). However, experiments conducted

187
on thylakoids exposed to photoinhibitory treatmentssuggested that zeaxanthin increased significantly thestructural flexibility of membranes: In the presence ofzeaxanthin, the irreversible disassembly of the macro-domains was much smaller than in the absence of thesexanthophylls (Gussakovsky et al. 1997). Recently,by analysing negative stained electron micrographsof granal membranes that were exposed to photoin-hibitory conditions, it has been shown that structuralintegrity of the macroarray of complexes also dependson the phosphorylation of the membranes (Kitmitto etal. 1999). It is important to point out that the macroar-rays detected by this technique exhibited dimensionssimilar to the sizes of the chiral macrodomains. In ad-dition, in perfect agreement with earlier data from CDmeasurements (Gussakovsky et al. 1997), they wereshown to be desintegrated into smaller domains uponprolonged illumination with intense light.
As concerns the possible mechanism(s) of NPQand its correlation with light-induced structural re-arrangements in the chiral macrodomains, it is import-ant to point out that �CD has been observed not onlyin thylakoid membranes, but also in lamellar aggreg-ates of purified LHC II. These changes in the chiralmacro-organization of the chromophores have alsobeen shown to be associated with reversible quench-ing of the Chl a fluorescence (Barzda et al. 1996,1999; Gruszecki et al. 1999b). This type of revers-ible quenching in the fluorescence of LHC II, thatresembled to some extent the NPQ in thylakoids, wasfirst observed by Jennings et al. (1991). This quench-ing has also been shown to be largely facilitated bythe organization of LHC II in chiral macrodomains(Barzda et al. 1999). Recent data suggest that the vi-olaxanthin cycle pigments affect the supramolecularorganization of LHC II (Gruszecki et al. 1999a).
In thylakoid membranes, NPQ is normally in-duced by the light-driven proton gradient. However,it can also be mimicked by an artificial increase ofthe proton concentration. When incubated in a reac-tion medium buffered to pH 5, thylakoid membranesof higher plants exhibited a pronounced quenchingof chlorophyll fluorescence in the presence of zeax-anthin (Rees et al. 1992; Goss et al. 1995). Titrationof algal cells to pH 5 or incubation of intact cellsin a reaction medium pH 5 also led to an accumula-tion of de-epoxidized xanthophylls (Goss et al. 1998),and a strong NPQ. Although incubation at low pHmay not completely represent the light-driven protongradient, the main characteristics of the zeaxanthin-dependent NPQ induced by light and that provoked
by low pH appear to be similar. This suggests thatthe basic mechanisms underlying the two types ofquenching are at least very similar. On this basis, in-volvement of structural changes can be expected. Infact, structural changes that accompany the quenchingof fluorescence have been detected in isolated LHCII (Ruban et al. 1997b). LHC II is capable of under-going low-pH-induced quenching that depends on theaggregation state of the complexes and is influencedby the presence of zeaxanthin (Ruban et al. 1996,1997a). However, to our knowledge no evidence hasbeen presented for the occurrence of low-pH-inducedstructural changes in intact cells.
In this work, we induced non-photochemical fluor-escence quenching by low pH in dark-adapted wholecells of Chlorella fusca (Chlorophyceae) and Mantoni-ella squamata (Prasinophyceae) that contained dif-ferent amounts of de-epoxidized violaxanthin cyclepigments, and by means of CD investigated the ac-companying structural changes in the chiral macro-organization of the complexes. Our data strongly sug-gest that efficient quenching in intact cells at low pHrequires both structural rearrangements in the chiralmacrodomains and the accumulation of de-epoxidizedxanthophylls of the violaxanthin cycle.
Materials and methods
Plant material
Chlorella fusca cells (Shihirira et Krauss, strain C1.1.10) were grown in liquid medium according toHase and Morimura (1971) at a photon flux density(PFD) of 40 µmol m−2 s−1 (PAR) with a light:dark re-gime of 14:10 h. The temperature of the batch cultureswas held constant at 20 ◦C. Mantoniella squamatacells (Manton et Parke, strain LB 1965/1) were cul-tivated under the same conditions, but with a PFDof 20 µmol m−2 s−1 in artificial seawater mediumaccording to Müller (1962). Chlorella cells were har-vested directly from the logarithmically growing cul-ture at a chlorophyll content of 5 mg Chl (a+b) l−1,Mantoniella cells with a chlorophyll content of 2 mgChl (a+b+c) l−1. Before the incubation in reactionmedium pH 5, Chlorella and Mantoniella cells weredark-adapted for 2 h.

188
Incubation of intact cells in reaction media ofdifferent pH in the dark
All measurements were performed with intact cellsof Chlorella fusca and Mantoniella squamata. De-epoxidation of violaxanthin to zeaxanthin in Chlorellaand to antheraxanthin in Mantoniella and non-photochemical quenching of chlorophyll fluorescencewere induced by incubation of intact Chlorella andMantoniella cells in a reaction medium pH 5 in thedark.
Dark-adapted Chlorella and Mantoniella cellswere harvested by mild centrifugation (1000 × g for2 min) and resuspended in reaction media in whichthe pH was adjusted to pH 5 and pH 7.5. The reactionmedia consisted of 0.33 M sorbitol, 10 mM KCl, 5mM MgCl2, 30 mM ascorbate and 40 mM MES (pH5) or 40 mM HEPES (pH 7.5). In Chlorella, CCCP(100 µM) had to be added to ensure the penetrationof protons through the cell wall and the thylakoidmembrane. In order to block low-pH-induced anther-axanthin and zeaxanthin synthesis in the dark, 1 mMDTT was added. After a 10 min incubation in therespective reaction media the cells were used for fluor-escence measurements and for pigment determination.
Determination of xanthophyll cycle pigments
The concentration of violaxanthin cycle pigments wasdetermined using HPLC pigment analysis. Samplesfor HPLC were taken before and directly after incub-ating the cells for 10 min at a different pH. Algalcells were harvested by filtration and the pigmentswere immediately extracted with 90% methanol/0.2M ammonium acetate (90:10) and 10% ethyl acet-ate. The pigment extract was centrifuged for 90 s at13 000 × g (Hermle Z 231 M) and the supernatantwas directly loaded onto the HPLC column. HPLCseparation was performed on a Gynkotek HPLC sys-tem consisting of an HPLC pump (Gynkotek M 480)equipped with a photodiode array detector (GynkotekUVD 340 S). For the separation of Chlorella pigmentsa 250/8/4 Nucleosil 120-5, C-18 column (Macherey-Nagel, Düren, Germany) was used with the followingeluents and gradient program. Eluent A consisted ofacetonitril/25 mM Tris–HCl pH 7.5 (97:3), eluent B ofmethanol/ethyl acetate (75:25). The elution was per-formed consecutively with eluent A and eluent B for18 min and 15 min, respectively. The temperature ofthe HPLC column was adjusted to 20 ◦C, the flowrate was 1 ml min−1. The separation of Mantoniellapigments was performed as described by Goss et al.
(1998). Quantification of xanthophyll cycle pigmentsand Chl a was carried out using calibration curves cal-culated with purified and quantified authentic standardpigments.
Determination of non-photochemical quenching ofchlorophyll fluorescence
Room temperature fluorescence measurements wereperformed using a PAM fluorometer (Walz, Effeltrich,Germany) equipped with an ED 101 US cuvette. Thealgal cell cultures were used at a concentration of 5mg Chl (a+b) l−1 in the case of Chlorella cells, and2 mg Chl (a+b+c) l−1 for Mantoniella. The fluor-escence parameters Fo and Fm of dark-adapted cellswere recorded before and after 10 min of incubation atdifferent pH values. For the Chl fluorescence measure-ments Chlorella and Mantoniella cells were incubateddirectly in the ED 101 US cuvette. One hundred µMCCCP was added to Chlorella cells directly after thedetermination of Fo and Fm of the dark-adapted state.
To test if zeaxanthin and antheraxanthin were ableto induce NPQ in the absence of high proton con-centrations, algal cells with high concentrations ofzeaxanthin and antheraxanthin, that were induced bya preillumination (350 µmol photons m−2 s−1 for5 min), were incubated in the corresponding reac-tion medium at pH 7.5. (Note that incubation ofdark-adapted cells at pH 7.5 per se does not lead tode-epoxidation of violaxanthin.)
The PS II quantum yield of intact cells underthe various measurement conditions was calculatedas Fv/Fm with Fv = Fm − Fo. The minimum (Fo)and maximum (Fm) fluorescence yields were determ-ined directly before and after 10 min of the respectiveincubation in reaction media pH 7.5 or pH 5.
CD-spectroscopy
Since harvesting of the cells by centrifugation disrup-ted the main bands, that are characteristic of intactcells, for the CD measurements Chlorella and Man-toniella cells in the respective culture medium weretitrated to pH 7.5, 6.5, 5.5 and 5 by the addition ofsmall amounts of 1 N HCl. Before the addition ofHCl, 100 µM CCCP and 30 mM ascorbate were addedto the culture medium. In Chlorella, in the absenceof CCCP, the CD was insensitive to the lowering ofthe pH. For Mantoniella, addition of CCCP was notnecessary.
Accumulation of de-epoxidized xanthophylls wasprevented by the addition of 1 mM DTT. In order to
189
prove that the changes induced by low pH were revers-ible, the pH of the medium was titrated back to neutralpH by the addition of small volumes of 1 N KOH.
CD spectra were recorded at room temperature ina Jobin Yvon CD 6 dichrograph using a bandpass of2 nm. Chlorella and Mantoniella cells were obtaineddirectly from the culture medium at chlorophyll con-centrations of about 5 mg Chl (a+b) l−1 and 2 mgChl (a+b+c) l−1, respectively. The low chlorophyllconcentration of Mantoniella cells brought about dis-tortions in the baseline which, however, cannot be heldresponsible for the low-pH-induced variations in themain CD bands. As pointed out earlier (Goss et al.2000), Mantoniella cells must be harvested in the log-arithmic growth phase at relatively low Chl content,and this alga cannot be centrifuged without the loss ofsome characteristic, psi-type CD bands.
Results
Low-pH-induced de-epoxidation of the violaxanthincycle pigments
Table 1 shows the concentrations of the violaxanthincycle pigments in Chlorella and Mantoniella cells un-der the experimental conditions used in this study.It can be seen that the dark-adapted cells containedhigh amounts of violaxanthin and low concentrationsof antheraxanthin and zeaxanthin. The total violax-anthin cycle pigment pool size was slightly higherin Chlorella than in Mantoniella. Illumination of thecells led to a conversion of about 40 mol violax-anthin to antheraxanthin in Mantoniella, and 30 molviolaxanthin to zeaxanthin in Chlorella. Transferringdark adapted cells from the culture medium to thepH 7.5 medium had no significant effect on the pig-ment composition. However, a 10 min incubation atpH 5 led to a strong de-epoxidation of violaxanthin inboth algae. In Chlorella, it induced a strong accumula-tion of zeaxanthin with antheraxanthin concentrationsbeing rather low. In Mantoniella, the accumulationof zeaxanthin was suppressed compared to antherax-anthin. These data agree well with literature data onChlorella (Hager 1967, 1969) and our earlier data onMantoniella (Goss et al. 1998). Addition of the viol-axanthin de-epoxidase inhibitor DTT (1 mM), as ex-pected, blocked almost completely the de-epoxidationof violaxanthin in both algae.
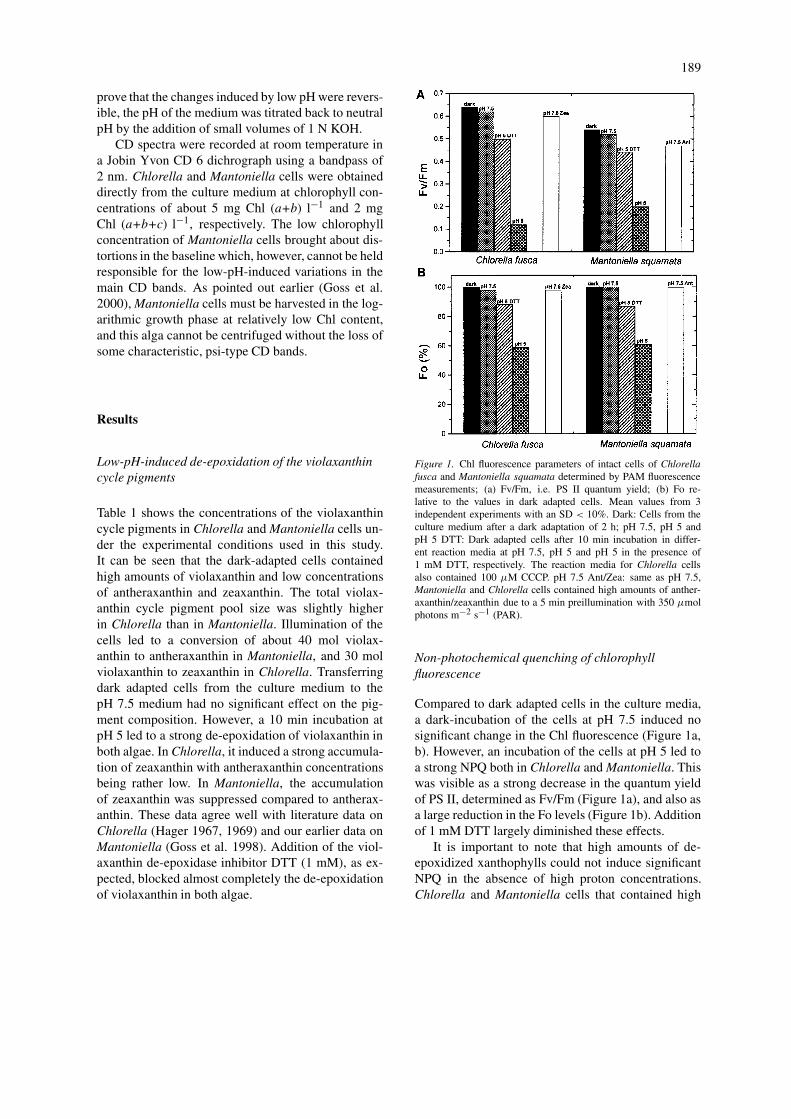
Figure 1. Chl fluorescence parameters of intact cells of Chlorellafusca and Mantoniella squamata determined by PAM fluorescencemeasurements; (a) Fv/Fm, i.e. PS II quantum yield; (b) Fo re-lative to the values in dark adapted cells. Mean values from 3independent experiments with an SD < 10%. Dark: Cells from theculture medium after a dark adaptation of 2 h; pH 7.5, pH 5 andpH 5 DTT: Dark adapted cells after 10 min incubation in differ-ent reaction media at pH 7.5, pH 5 and pH 5 in the presence of1 mM DTT, respectively. The reaction media for Chlorella cellsalso contained 100 µM CCCP. pH 7.5 Ant/Zea: same as pH 7.5,Mantoniella and Chlorella cells contained high amounts of anther-axanthin/zeaxanthin due to a 5 min preillumination with 350 µmolphotons m−2 s−1 (PAR).
Non-photochemical quenching of chlorophyllfluorescence
Compared to dark adapted cells in the culture media,a dark-incubation of the cells at pH 7.5 induced nosignificant change in the Chl fluorescence (Figure 1a,b). However, an incubation of the cells at pH 5 led toa strong NPQ both in Chlorella and Mantoniella. Thiswas visible as a strong decrease in the quantum yieldof PS II, determined as Fv/Fm (Figure 1a), and also asa large reduction in the Fo levels (Figure 1b). Additionof 1 mM DTT largely diminished these effects.
It is important to note that high amounts of de-epoxidized xanthophylls could not induce significantNPQ in the absence of high proton concentrations.Chlorella and Mantoniella cells that contained high
190
Table 1. Pigment contents (mol pigment) per 1000 mol Chl a of the xantho-phyll cycle pigments, violaxanthin, antheraxanthin and zeaxanthin, of intactcells of Chlorella fusca and Mantoniella squamata under different condi-tions; mean values from 5 independent experiments with an SD < 10%.Dark-adapted and illuminated: Cells in the culture medium at neutral pHafter 2 h in the dark, or 5 min illumination with light of 350 µmol m−2 s−1
(PAR) intensity, respectively; pH 7.5, pH 5 and pH5 DTT: Cells incubatedin the reaction media adjusted to pH 7.5 and pH 5 and pH 5 in the presenceof 1 mM DTT, respectively. The reaction media for Chlorella cells alsocontained 100 µM CCCP. For further details, see ‘Materials and methods’.
Treatment violaxanthin antheraxanthin zeaxanthin
Chlorella fusca
Dark-adapted 62.15 3.02 12.86
Illuminated 37.81 11.21 30.54
pH 7.5 61.64 3.44 11.97
pH 5 35.43 10.04 37.31
pH 5 DTT 57.98 7.39 12.5
Mantoniella squamata
Dark-adapted 59.13 5.1 4.78
Illuminated 29.48 40.12 3.15
pH 7.5 59.16 5.23 4.11
pH 5 11.9 37.61 15.6
pH 5 DTT 55.51 9.58 6.13
concentrations of zeaxanthin or antheraxanthin, thatwere formed during a 5 min preillumination, exhibitedhigh Fv/Fm and Fo values, which were comparable tothose found in dark adapted cells at pH 7.5.
It is also to be noted that the low Fv/Fm values,0.54 and 0.64 found in dark-adapted Mantoniella andChlorella cells, respectively, are due to chlorores-piration in these algal species (Wilhelm and Duval1989).
CD-spectroscopy
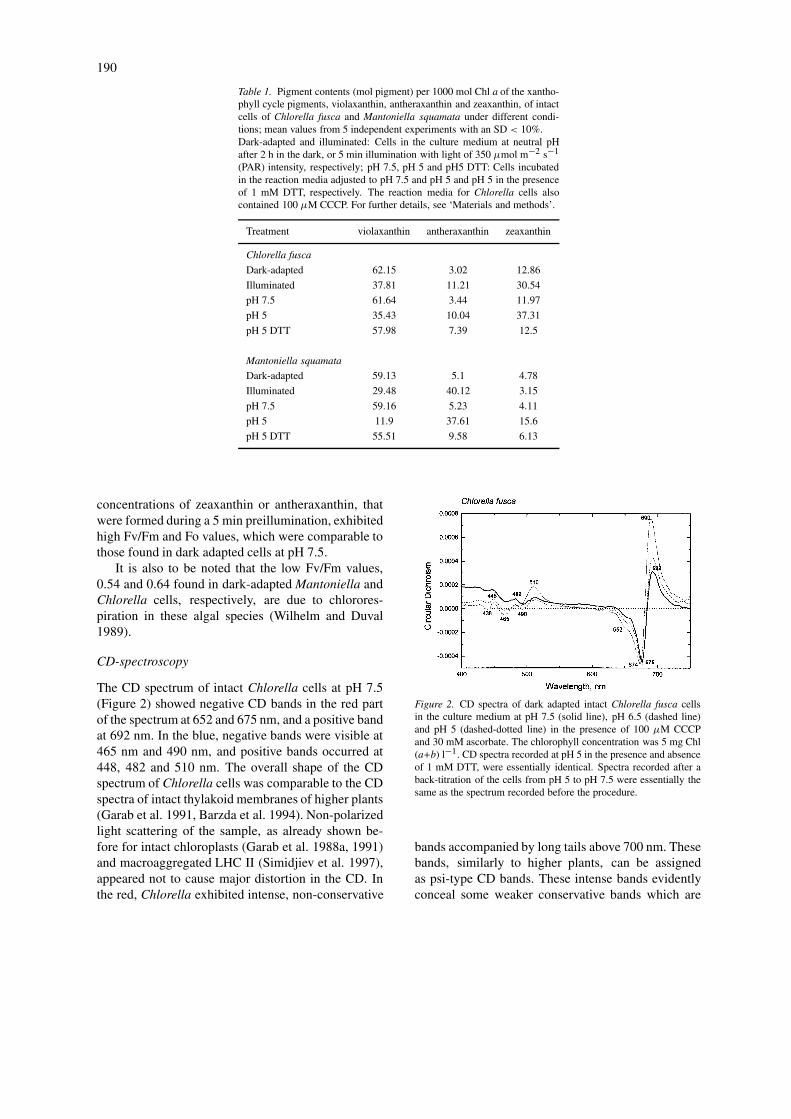
The CD spectrum of intact Chlorella cells at pH 7.5(Figure 2) showed negative CD bands in the red partof the spectrum at 652 and 675 nm, and a positive bandat 692 nm. In the blue, negative bands were visible at465 nm and 490 nm, and positive bands occurred at448, 482 and 510 nm. The overall shape of the CDspectrum of Chlorella cells was comparable to the CDspectra of intact thylakoid membranes of higher plants(Garab et al. 1991, Barzda et al. 1994). Non-polarizedlight scattering of the sample, as already shown be-fore for intact chloroplasts (Garab et al. 1988a, 1991)and macroaggregated LHC II (Simidjiev et al. 1997),appeared not to cause major distortion in the CD. Inthe red, Chlorella exhibited intense, non-conservative
Figure 2. CD spectra of dark adapted intact Chlorella fusca cellsin the culture medium at pH 7.5 (solid line), pH 6.5 (dashed line)and pH 5 (dashed-dotted line) in the presence of 100 µM CCCPand 30 mM ascorbate. The chlorophyll concentration was 5 mg Chl(a+b) l−1. CD spectra recorded at pH 5 in the presence and absenceof 1 mM DTT, were essentially identical. Spectra recorded after aback-titration of the cells from pH 5 to pH 7.5 were essentially thesame as the spectrum recorded before the procedure.
bands accompanied by long tails above 700 nm. Thesebands, similarly to higher plants, can be assignedas psi-type CD bands. These intense bands evidentlyconceal some weaker conservative bands which are
191
given rise by excitonic interactions of the pigment mo-lecules. Nonetheless, some of these split bands canclearly be identified, e.g. the shoulder at (−) 652 nmwhich is known to originate from LHC II (Van Metter1977), and which has been reported to remain invari-ant during the macroaggregation of isolated LHC II(Garab 1992).
Upon gradually lowering the pH of the culturemedium to pH 5 we observed an increase in the amp-litudes of the psi-type bands. In addition, both the (−)675 nm and the (+) 692 nm bands were shifted towardshorter wavelengths. A significant increase in the CDamplitude was also observed at (+) 510 nm. In higherplants, this latter band has also been shown to originatefrom a chiral macro-organization of the chromophores(Garab et al. 1988a; Barzda et al. 1994).
It was interesting to observe that addition of 1 mMDTT before titrating the cells to pH 5 did not preventthe increase of the psi-type CD bands in Chlorellacells exposed to low pH (data not shown). It must benoted, however, that minor alterations in the excitonicbands due to the presence of zeaxanthin could not beruled out. In an unreported experiment, we have alsoshown that back-titration of intact cells to neutral pHreversed fully the low-pH-induced changes in the CD.
Concerning the reversibility and the preferentialsensitivity of the psi-type CD, the low-pH-inducedchanges appear to be similar to the light-inducedchanges that have been reported earlier on higher plantthylakoids (Garab et al. 1988b; Istokovics et al. 1997).However, the two treatments induce opposite effects:While low pH increases, excess light decreases theamplitude of the main, psi-type bands. This clearlyindicates that at least as far as the macro-organizationof the chromophores is concerned, low pH and illu-mination lead to significantly different effects. Thisis not very surprising because the light-induced CD-changes in thylakoid membranes have been shown tobe largely independent of the transmembrane �pH,and amplified in excess light (see Barzda et al. 1996a;Istokovics et al., 1997).
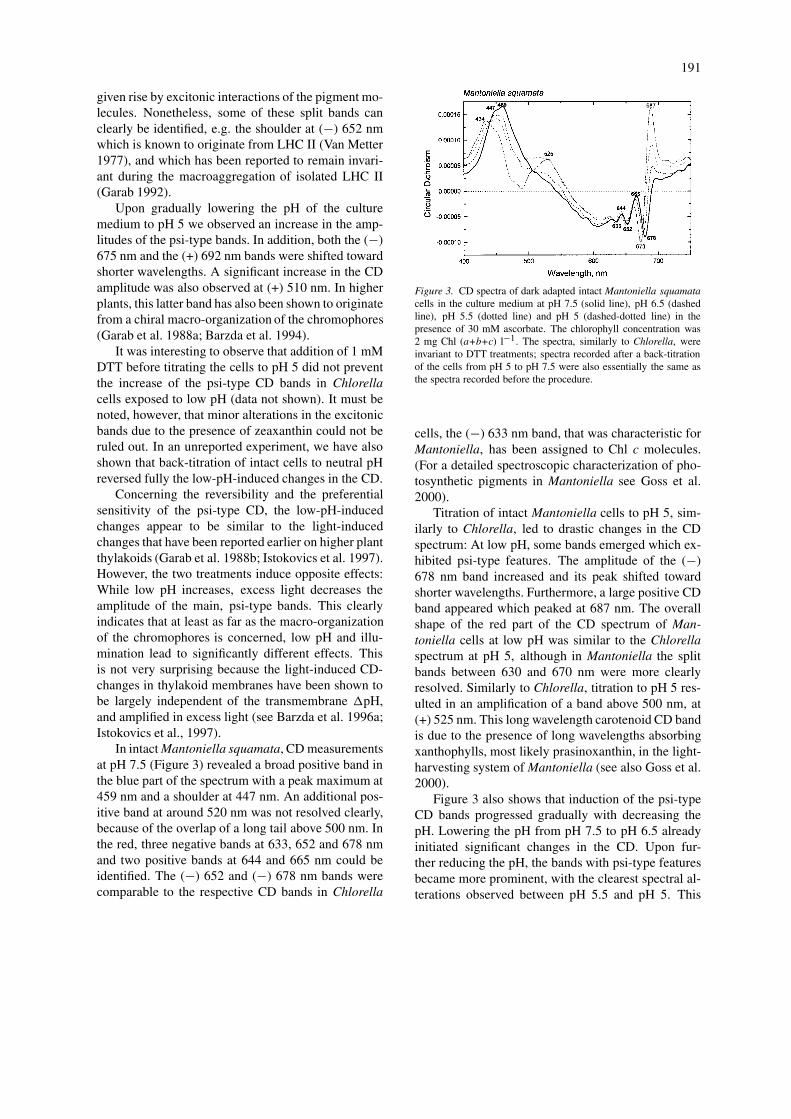
In intact Mantoniella squamata, CD measurementsat pH 7.5 (Figure 3) revealed a broad positive band inthe blue part of the spectrum with a peak maximum at459 nm and a shoulder at 447 nm. An additional pos-itive band at around 520 nm was not resolved clearly,because of the overlap of a long tail above 500 nm. Inthe red, three negative bands at 633, 652 and 678 nmand two positive bands at 644 and 665 nm could beidentified. The (−) 652 and (−) 678 nm bands werecomparable to the respective CD bands in Chlorella
Figure 3. CD spectra of dark adapted intact Mantoniella squamatacells in the culture medium at pH 7.5 (solid line), pH 6.5 (dashedline), pH 5.5 (dotted line) and pH 5 (dashed-dotted line) in thepresence of 30 mM ascorbate. The chlorophyll concentration was2 mg Chl (a+b+c) l−1. The spectra, similarly to Chlorella, wereinvariant to DTT treatments; spectra recorded after a back-titrationof the cells from pH 5 to pH 7.5 were also essentially the same asthe spectra recorded before the procedure.
cells, the (−) 633 nm band, that was characteristic forMantoniella, has been assigned to Chl c molecules.(For a detailed spectroscopic characterization of pho-tosynthetic pigments in Mantoniella see Goss et al.2000).
Titration of intact Mantoniella cells to pH 5, sim-ilarly to Chlorella, led to drastic changes in the CDspectrum: At low pH, some bands emerged which ex-hibited psi-type features. The amplitude of the (−)678 nm band increased and its peak shifted towardshorter wavelengths. Furthermore, a large positive CDband appeared which peaked at 687 nm. The overallshape of the red part of the CD spectrum of Man-toniella cells at low pH was similar to the Chlorellaspectrum at pH 5, although in Mantoniella the splitbands between 630 and 670 nm were more clearlyresolved. Similarly to Chlorella, titration to pH 5 res-ulted in an amplification of a band above 500 nm, at(+) 525 nm. This long wavelength carotenoid CD bandis due to the presence of long wavelengths absorbingxanthophylls, most likely prasinoxanthin, in the light-harvesting system of Mantoniella (see also Goss et al.2000).
Figure 3 also shows that induction of the psi-typeCD bands progressed gradually with decreasing thepH. Lowering the pH from pH 7.5 to pH 6.5 alreadyinitiated significant changes in the CD. Upon fur-ther reducing the pH, the bands with psi-type featuresbecame more prominent, with the clearest spectral al-terations observed between pH 5.5 and pH 5. This

192
Figure 4. Difference spectra, �CD (pH 5 – pH 7.5) of dark adaptedintact cells of Chlorella fusca (dashed-dotted line) and Mantoni-ella squamata (solid line). Chlorophyll concentration was 5 mgChl (a+b) l−1 in the case of Chlorella and 2 mg Chl (a+b+c) l−1
for Mantoniella. The difference spectrum of Mantoniella cells hasbeen multiplied by a factor of 2.5 to compensate for the lower Chlconcentration.
shows that the alterations clearly occur in a physiolo-gical range. Furthermore, even the changes inducedat pH 5 are fully reversible. It is also noteworthy thatin Mantoniella, the induction of the big CD did notrequire the addition of CCCP.
Similarly to Chlorella, 1 mM DTT had no in-fluence on the psi-type CD at pH 5. However, itmust be stressed again that minor alterations in somecarotenoid associated excitonic bands could be ob-scured by the psi-type CD and the poor quality ofthe spectra, which was the consequence of the lowchlorophyll concentration and high light-scattering inthe suspension.
Figure 4 shows the difference spectra of CD meas-ured at pH 5 minus pH 7.5 of intact Chlorella andMantoniella cells. These difference spectra are evid-ently dominated by psi-type bands, i.e. by the bandsat around (+) 685 nm and (−) 670 nm, and by thelong-wavelength carotenoid bands at (+) 510 nm inChlorella and (+) 530 nm in Mantoniella. Apart fromthe overall resemblance of these two difference spec-tra, significant differences were also detected. The redbands were broader in Chlorella than in Mantoniella,which is probably the consequence of a larger contri-bution of differential scattering in Chlorella. In bothalgae, high proton concentrations evidently inducedfurther changes in the organisation of carotenoid mo-lecules as can be seen from significant changes below500 nm. Some of these evidently originate from ex-citonic interactions, as indicated by the split characterof some overlapping bands centred around 493 and472 nm in Chlorella and 497, 474 and 457 nm in Man-
toniella (vertical bars in Figure 4). Investigations areunderway on isolated thylakoids and LHCs to identifythese bands more precisely and to establish their rolein the structural rearrangements as well as in NPQ.
Discussion
In the present study, we have provided experimentalevidence for low-pH-induced structural rearrange-ments in the pigment systems of dark adapted intactalgal cells. We have shown that macroaggregates withlong range chiral order can be induced in vivo bylowering the pH both in Mantoniella squamata andChlorella fusca. In Chlorella, as indicated by thepresence of psi-type CD at pH 7.5, the chiral macro-domains were already present at neutral pH but theywere strongly enhanced by lowering the pH to 5. InMantoniella, the psi-type features were hardly dis-cernible at neutral pH but clearly appeared at low pH.This difference between the CD spectra of Chlorellaand Mantoniella at pH 7.5 can be explained by thedifferences in the ultrastructures of their chloroplasts.
In Chlorella fusca, thylakoids possess a band-likestructure that spans the chloroplast. The inner bandscorrespond to grana membranes of higher plants,the outer bands show all features typical for stromathylakoids. In higher plants, via mediating the stackingof membranes (Staehelin and Arntzen 1983; Barber1986) and the lateral separation, i.e. the sorting ofthe photosystems in the membranes between granumand stroma regions (Garab et al. 1991; Garab andMustárdy 1999), LHC II plays a key role in the forma-tion of the granum. In the granum, PS II particles andthe chromophores are arranged in three-dimensionalarrays, which thus give rise to psi-type CD (Barzda etal. 1994). The low-pH-induced increase in the psi-typeCD in Chlorella is interpreted as a strengthening of thechiral macro-organization of the chromophores in thegranum-like regions of the inner bands, and indicatesan increase in the long-range order of the pigment di-poles. As inferred from the results obtained in LHC II(Barzda et al. 1996b), this facilitates the energy migra-tion in the membranes over long distances. However,long range order alone may facilitate both the utiliza-tion and the quenching of the excitation energy. Thus,the effect of this increased long range order on quench-ing becomes noticeable only upon the introduction ofquenchers. Our data show that in intact, dark adap-ted Chlorella cells, in accordance with most literature

193
data, the appearance of quenchers is associated withthe accumulation of zeaxanthin.
A similar explanation is offered for Mantoniella.In the chloroplasts of these cells, no differentiationin granum- and stroma-like regions can be observed.The thylakoid membranes are aligned parallel to eachother, with spacings of 2–8 nm (Krämer et al. 1988).This regular thylakoid arrangement gave rise only toweak anomalous CD (Goss et al. 2000). The in-duction of large, anomalously shaped CD bands atlow pH indicates the formation of three-dimensionalmacroaggregates with long-range chiral order. Theseare probably brought about by significant changes inthe membrane ultrastructure, e.g. via changes in thestacking interactions. The large amplitudes of the an-omalous, non-conservative CD bands, which resembleclosely those in Chlorella, strongly suggest a psi-typeorigin (Keller and Bustamante 1986a, b; Garab 1996).Hence, these chiral macrodomains also can provide asuitable structure for the migration of the excitationenergy for long distances. These macrodomains, inthe presence of quenchers which are evidently associ-ated with anteraxanthin, can contribute to the efficientenergy dissipation at low pH.
Low pH induced aggregation has been reported be-fore for isolated systems. It has earlier been shownthat high concentrations of metalic cations and highproton concentrations have similar effects on the ul-trastructure of granal thylakoids (Karukstis and Sauer1985). This, together with our earlier findings thatthe formation of chiral macrodomains is governed bymetalic cations (Garab et al. 1991), support the aboveinterpretation. Ruban et al. (1996, 1998) also usedhigh proton concentrations to induce the aggregationof LHC II. Further, they found that the addition ofexogenous zeaxanthin to purified LHC II stimulatedboth the aggregation and the fluorescence quenchingwhereas violaxanthin was found to be an inhibitor ofLHC II aggregation (Ruban et al. 1994; 1996; Phil-lip 1996). It is somewhat surprising that under ourexperimental conditions, i.e. in intact cells, aggrega-tion of light-harvesting complexes did not depend onthe presence of de-epoxidized xanthophyll cycle pig-ments. Titration of Chlorella and Mantoniella cells topH 5 in the presence of the violaxanthin de-epoxidaseinhibitor DTT prevented the de-epoxidation of violax-anthin but had no influence on the psi-type CD that isassociated with the chiral macroaggregates of the lightharvesting complexes.
The finding that low pH and excess light induceopposite changes in the chiral macro-organization of
the chromophores is unexpected because of the light-induced lumenal acidification of thylakoids. However,an asymmetric protonation due to photosynthesis evid-ently cannot result in the same strengthening of thecoupling between adjacent layers as an overall in-crease in the proton, or other, metallic cation con-centration. Furthermore, the light-induced reversiblestructural changes induced by excess light in thylakoidmembranes and in lamellar aggregates of LHC II arethought to be driven by release of Mg-ions, most prob-ably at the stromal side of the membrane (Garab etal. 1998; Cseh et al. 1998). Indeed, in LHC II, lighthas been shown to induce changes preferentially in theouter loop of the stromal side, around the site of phos-phorylation (Zer et al. 1999). It must also be pointedout that both CD and the light-induced CD-changes inalgal cells can depend on the pH, as it has been indic-ated in pilot experiments on higher plant thylakoids(Istokovics and Garab, unpublished). Thus, strictlyspeaking, the conclusions derived from the present ex-perimental findings are to be confined to dark adaptedmembranes.
High NPQ at low pH in the presence of de-epoxidized xanthophyll cycle pigments has earlierbeen observed in thylakoid membranes of higherplants where high concentrations of zeaxanthin wereinduced by actinic illumination (Rees et al. 1992) orby an artificial proton gradient from ATP-hydrolysis(Goss et al. 1995). In good agreement with theseresults and other literature data, we found that theenhanced long range order of the chromophores, in-duced by low pH, can lead to an enhanced quenchingonly in the presence of de-epoxidized xanthophylls(Demmig-Adams et al. 1990; Gilmore and Yamamoto1992, 1993). By employing a micellar system, Avitaland Malkin (1998) were able to show that zeaxanthinalso in a model system is a more efficient quencherof chlorophyll fluorescence than violaxanthin. Thiswould be easily explained with a mechanism of ‘re-verse’ energy transfer, i.e. from Chl a to zeaxanthin,as opposed to the preferential transfer from violax-anthin to Chl a (Frank et al. 1994). However, re-cent findings of Polivka et al. (1999) show that thezeaxanthin-dependent quenching cannot be accountedfor this type of mechanism. The role of zeaxanthin(or anteraxanthin) in NPQ can be explained better byinvoking the mechanism of enhanced internal conver-sion in carotenoid:porphyrin dyads. This mechanismfor controlling energy dissipation of Chls by caroten-oids has been proposed by Naqvi et al. (1997), whoalso provided some experimental evidence by using

194
LHC II preparations (Naqvi et al. 1999) and artificialsystems (Osuka et al. 1999). In terms of enhanced in-ternal conversion, the quenching due to an exchangeof an epoxidized xanthophyll (violaxanthin) with ade-epoxidized xanthophyll (antheraxanthin or zeax-anthin) can be explained by differences in the couplingbetween the Chl and its carotenoid partner. The valid-ity of this mechanism for the pigments affected bythe operation of the violaxanthin cycle remains tobe verified experimentally. Hence, other mechanisms(perhaps operating parallel with the enhanced internalconversion mechanism) cannot be discounted.
NPQ at low pH has also been reported in the com-plete absence of de-epoxidized xanthophylls and hasbeen ascribed to processes taking place at the reactioncentre of PS II. Krieger and Weis (1993) proposedthat acidification of the thylakoid lumen leads to aCa2+-release in the water splitting system, therebyinactivating PS II electron transport and inducing non-photochemical quenching of chlorophyll fluorescence.Furthermore, aggregation of isolated light-harvestingcomplexes by cations (Barzda et al. 1994) or by re-moval of detergent (Barzda et al. 1995) also leadsto a significant quenching of Chl a fluorescence ir-respective of the presence or absence of zeaxanthin.In addition, illumination of isolated LHC II can alsoinduce reversible fluorescence quenching (Jennings etal. 1991) that is independent of the presence of de-epoxidized xanthophylls (Barzda et al. 1999). Thephysiological significance of this type of structuralchanges in LHC II, beside their possible role in NPQ,is shown by the finding that phosphorylation of thesecomplexes can be regulated by light at the substratelevel (Zer et al. 1999).
A recent study, which revealed the key role ofthe PsbS protein in NPQ in higher plants (Li et al.2000), also demonstrated the importance of structuralchanges in the same process. This was indicated bythe absence of �A535 in the mutant with substantiallyreduced capability for NPQ. It is interesting to pointout that the light-induced reversible quenching in LHCII is also associated with structural rearrangements(Barzda et al. 1999; Gruszecki et al. 1999b), prob-ably due to a light-induced release of cations (Garabet al. 1998). Although the structural changes, in thechiral macro-organization of LHC II, similarly to thequenching, occur in the absence of zeaxanthin (Barzdaet al. 1996), it cannot be ruled out that they are modu-lated by the presence of these de-epoxidized pigments.The fact that different violaxanthin cycle pigments arealigned in a different manner in the supramolecular or-
ganization of LHC II (Gruszecki et al. 1999a) is in linewith this notion. It can also be speculated that differentmacro-organizations offer different conditions for theactivity of the violaxanthin cycle, which involves mo-bile enzymes and thus probably must rely substantiallyon the structural flexibilty of the membranes. Viceversa, as suggested by the findings of Gussakovsky etal. (1997), de-epoxidized xanthophylls may contributeto the overall structural flexibility of the membranes.
Concerning the possible role of the chiral macro-organization in the generation of quenchers, the ana-lysis of quenching in LHC II can be instructive. Ithas been shown unequivocally that the light-inducedreversible fluorescence quenching in LHC II, simil-arly to higher plant thylakoids (Barzda et al. 1996a),depends strongly on the presence of chiral macro-domains (Barzda et al. 1999). This, as emphasizedabove, does not mean, however, that the macrodo-mains directly quench the fluorescence. While it isclear that in the presence of low concentrations ofquenchers they facilitate quenching in the entire do-main, it is not clear what role their structural flexibiltymay play. This would be important to clarify becausesome observations suggest a correlation with the struc-tural flexibility of the artificial membranes in whichquenchers are generated by the light used for the ex-citation of Chl fluorescence. It has also been shownthat the light-induced reversible quenching in LHC IIoriginates from a very fast generation of quencherswhich remain at a very low concentration and whichare not identical with zeaxanthin (Barzda et al. 1999).However, it remains to be examined how the pres-ence of de-epoxidized xanthophylls can support thegeneration of quenching sites.
In summary, although the nature and role ofthe structural changes associated with fluorescencequenching remain to be clarified, it seems to be clearthat structural changes are involved in the quench-ing of the Chl a fluorescence in the light-harvestingantenna complexes both in intact and in isolated sys-tems. Thus, our results, showing significant, low-pH-induced structural rearrangements in the chiralmacro-organization of the chromophores in intact,dark-adapted algal cells substantiate the role of thestructural changes in the mechanisms that regulate theutilization of the excitation energy in the photosyn-thetic membranes.

195
Acknowledgements
The authors express their thanks to Prof. Dr. C. Wil-helm for fruitful discussions. Financial support fromthe Deutsche Forschungsgemeinschaft (GO 818/1-2) and a grant from the Hungarian Fund for BasicResearch, OTKA T 019226 are gratefully acknow-ledged. This work was also supported by an OM-OMFB/DAAD exchange program, TÉT D-44/96.
References
Avital S and Malkin S (1998) Quenching of chlorophyll fluores-cence by carotenoids in a micellar model system. In: Garab G(ed) Photosynthesis: Mechanisms and Effects, Vol I, pp 477–482.Kluwer Academic Publishers, Dordrecht, The Netherlands
Barber J (1986) Surface electric charges and protein phosphoryla-tion. In: Staehelin LA and Arntzen CJ (eds) Encyclopedia ofPlant Physiology, Vol 19, pp 653–664. Springer Verlag, Berlin
Barzda V, Mustárdy L and Garab G (1994) Size dependency of cir-cular dichroism in macroaggregates of photosynthetic pigment–protein complexes. Biochemistry 33: 10837–10841
Barzda V, Garab G, Gulbinas V and Valkunas L (1995) Long dis-tance migration of excitation energy and fluorescence quenchingmechanism in chiral macroaggregates of LHC II. In: Mathis P(ed) Photosynthesis: From Light to Biosphere, Vol I, pp 319–322. Kluwer Academic Publishers, Dordrecht, The Netherlands
Barzda V, Istokovics A, Simidjiev I and Garab G (1996a) Struc-tural flexibility of chiral macroaggregates of light-harvestingchlorophyll a/b pigment–protein complexes. Light-induced re-versible structural changes associated with energy dissipation.Biochemistry 35: 8981–8985
Barzda V, Garab G, Gulbinas V and Valkunas L (1996b) Evid-ence for long-range excitation migration in macroaggregates ofthe chlorophyll a/b light-harvesting antenna complexes. BiochimBiophys Acta 1273: 231–236
Barzda V, Jennings RC, Zucchelli G and Garab G (1999) Kin-etic analysis of the light-induced fluorescence quenching inlight-harvesting chlorophyll a/b pigment–protein complex ofPhotosystem II. Photochem Photobiol 70 (5): 751–759
Cseh Z, Papp E and Garab G (1998) Model calculations on themechanism of the light-induced structural changes in lamel-lar aggregates of LHC II. In: Garab G (ed) Photosynthesis:Mechanisms and Effects, Vol I, pp 345–348. Kluwer AcademicPublishers, Dordrecht, The Netherlands
Demmig-Adams B (1990) Carotenoids and photoprotection inplants: a role for the xanthophyll zeaxanthin. Biochim BiophysActa 1020: 1–24
Demmig-Adams B, Adams III WW, Heber U, Neimanis S, WinterK, Krüger A, Czygan FC, Bilger W and Björkman O (1990)Inhibition of zeaxanthin formation and of rapid changes in ra-diationless energy dissipation by dithiothreitol in spinach leavesand chloroplasts. Plant Physiol 92: 293–301
Demmig-Adams B and Adams III WW (1996) The role of xantho-phyll cycle carotenoids in the protection of photosynthesis.Trends Plant Sci 1: 21–26
Finzi L, Bustamante C, Garab G and Juang C-B (1989) Directobservation of large chiral domains in chloroplast thylakoidmembranes by differential polarization microscopy. Proc NatlAcad Sci USA 86: 8748–8752
Frank HA, Cua A, Chynwat V, Young AJ, Gosztola D andWasielewski MR (1994) Photophysics of the carotenoids asso-ciated with the xanthophyll cycle in photosynthesis. PhotosynthRes 41: 389–395
Garab G (1992) Macrodomain organization of complexes in thethylakoid membranes. In: Murata N (ed) Research in Photo-synthesis, Vol I, pp 171–178. Kluwer Academic Publishers,Dordrecht, The Netherlands
Garab G (1996) Linear and circular dichroism. In: Amesz J and HoffAJ (eds) Biophysical Techniques in Photosynthesis, pp 11–40.Kluwer Academic Publishers, Dordrecht, The Netherlands
Garab G and Mustárdy L (1999) Role of LHC II-containing macro-domains in the structure, function and dynamics of grana. Aust JPlant Physiol 26: 649–658
Garab G, Faludi-Daniel A, Sutherland JC and Hind G (1988a)Macro-organization of chlorophyll a/b light-harvesting com-plex in thylakoids and aggregates: Information from circulardifferential scattering. Biochemistry 27: 2425–2430
Garab G, Leegood RC, Walker DA, Sutherland JC and Hind G(1988b) Reversible changes in macro-organization of the light-harvesting chlorophyll a/b pigment–protein complex detected bycircular dichroism. Biochemistry 27: 2430–2434
Garab G, Wells KS, Finzi L and Bustamante C (1988c) Helic-ally organized macroaggregates of pigment–protein complexesin chloroplasts: Evidence from circular intensity differentialscattering. Biochemistry 27: 5839–5843
Garab G, Kieleczawa J, Sutherland JC, Bustamante C and Hind G(1991) Organization of pigment–protein complexes into macro-domains in the thylakoid membranes of wild-type and chloro-phyll b-less mutant of barley as revealed by circular dichroism.Photochem Photobiol 54: 273–281
Garab G, Istokovics A, Butiuc A, Simidjiev I and Dér A (1998)Light-induced ion movements in thylakoid membranes and isol-ated LHC II. In: Garab G (ed) Photosynthesis: Mechanismsand Effects, Vol I, pp 341–344. Kluwer Academic Publishers,Dordrecht, The Netherlands
Gilmore AM (1997) Mechanistic aspects of xanthophyll cycle-dependent photoprotection in higher plant chloroplasts andleaves. Physiol Plant 99: 197–209
Gilmore AM and Yamamoto HY (1992) Dark-induction ofzeaxanthin-dependent nonphotochemical fluorescence quench-ing mediated by ATP. Proc Natl Acad Sci USA 89: 1899–1903
Gilmore AM and Yamamoto HY (1993) Linear models re-lating xanthophylls and lumen acidity to non-photochemicalfluorescence quenching: Evidence that antheraxanthin explainszexanthin-independent quenching. Photosynth Res 35: 67–78
Gilmore AM, Hazlett T and Govindjee (1995) Xanthophyll cycle-dependent quenching of Photosystem II chlorophyll a fluores-cence: Formation of a quenching complex with a short fluores-cence lifetime. Proc Natl Acad Sci USA 92: 2273–2277
Goss R, Richter M and Wild A (1995) Role of �pH in the mechan-ism of zeaxanthin-dependent amplification of qE. J PhotochemPhotobiol B, Biol 27: 147–152
Goss R, Böhme K and Wilhelm C (1998) The xanthophyll cycleof Mantoniella squamata converts violaxanthin to antherax-anthin but not to zeaxanthin: Consequences for the mechanismof enhanced non-photochemical energy dissipation. Planta 205:613–621
Goss R, Wilhelm C and Garab G (2000) Organization of the pigmentmolecules in the chlorophyll a/b/c containing alga Mantoniellasquamata (Prasinophyceae) studied by means of absorbance, cir-cular and linear dichroism spectroscopy. Biochim Biophys Acta1457: 190–199

196
Gruszecki WI, Grudzinski W, Banaszek-Glos A, Matula M, KernenP, Krupa Z and Sielewiesiuk J (1999a) Xanthophyll pigmentsin light-harvesting complex II in monomolecular layers: Local-isation, energy transfer and orientation. Biochim Biophys Acta1412: 173–183
Gruszecki WI, Grudzinski W, Matula M, Kernen P and KrupaZ (1999b) Light-induced excitation quenching and structuraltransition in light-harvesting complex II. Photosynth Res 59:175–185
Gülen D, Van Grondelle G and Van Amerongen H (1997) Structuralinformation on the light harvesting complex II of green plantsthat can be deciphered from polarised absorption characteristics.J Phys Chem B 101: 7256–7261
Gussakovsky EE, Barzda V, Shahak Y and Garab G (1997) Irre-versible disassembly of chiral macrodomains in thylakoids dueto photoinhibition. Photosynth Res 51: 119–126
Hager A (1967) Untersuchungen über die lichtinduzierten revers-iblen Xanthophyllumwandlungen in Chlorella und Spinacia.Planta 74: 148–172
Hager A (1969) Lichtbedingte pH-Erniedrigung in einemChloroplasten-Kompartiment als Ursache der enzymatischenViolaxanthin- zu Zeaxanthin-Umwandlung. Beziehungen zurPhotophosphorylierung. Planta 89: 224–243
Hase E and Morimuro Y (1971) Synchronous and homocontinuouscultures of algae. In: Pietro S (ed) Methods in Enzymology 23,pp 81–106. Academic Press, New York
Horton P, Ruban AV, Rees D, Pascal AA, Noctor G and Young AJ(1991) Control of the light harvesting function of chloroplastmembranes by aggregation of the LHC II chlorophyll–proteincomplex. FEBS Lett 292: 1–4
Horton P, Ruban AV and Walters RG (1996) Regulation of lightharvesting in green plants. Annu Rev Plant Physiol Plant MolBiol 47: 655–684
Horton P (1999) Hypothesis: Are grana necessary for regulation oflight harvesting? Aust J Plant Physiol 26: 659–669
Istokovics A, Simidjiev I, Lajko F and Garab G (1997) Charac-terization of the light-induced reversible changes in the chiralmacro-organization of the chromophores in chloroplast thylakoidmembranes. Temperature dependence and effect of inhibitors.Photosynth Res 54: 45–53
Jennings RC, Garlaschi FM and Zucchelli G (1991) Light-inducedfluorescence quenching in the light-harvesting chlorophyll a/bprotein complex. Photosynth Res 34: 375–385
Karukstis KK and Sauer K (1985) The effects of cation-inducedand pH-induced membrane stacking on chlorophyll fluorescencedecay kinetics. Biochim Biophys Acta 806: 374–388
Keller D and Bustamante C (1986a) Theory of the interaction oflight with large inhomogeneous molecular aggregates. I. Absorp-tion. J Chem Phys 84: 2961–2971
Keller D and Bustamante C (1986b) Theory of the interactionof light with large inhomogeneous molecular aggregates. II.Psi-type circular dichroism. J Chem Phys 84: 2972–2979
Kitmitto A, Mustafa AO, Ford JW, Holzenburg A and Ford RC(1999) Does photoinhibition and/or phosphorylation of Pho-tosystem II influence its in vivo oligomeric state? BiochimBiophys Acta 1413: 21–30
Krämer P, Wilhelm C, Wild A, Mörschel E and Rhiel E (1988)Ultrastructure and freeze-fracture studies of thylakoids of Man-toniella squamata (Prasinophyceae). Protoplasma 147: 170–177
Krieger A and Weis E (1993) The role of calcium in the pH-dependent control of photosystem II. Photosynth Res 37: 117–130
Li X-P, Björkman O, Shih C, Rosenquist M, Jansson S and NiyogiKK (2000) A pigment–binding protein essential for the regula-tion of photosynthetic light harvesting. Nature 403: 391–395
Müller D (1962) Über jahres- und lunarperiodische Erscheinungenbei einigen Braunalgen. Bot Mar 4: 140–155
Naqvi KR, Melo TB, Raju BB, Jávorfi T, Simidjiev I and GarabG (1997) Quenching of chlorophyll a singlets and triplets bycarotenoids in the light-harvesting complex of Photosystem II:Comparison of aggregates with trimers. Spectrochim Acta PartA 53: 2659–2667
Naqvi KR, Jávorfi T, Melo TB and Garab G (1999) More on thecatalysis of internal conversion in chlorophyll a by an adja-cent carotenoid in light-harvesting complex (Chl a/b LHC II)of higher plants: Time-resolved triplet-minus-singlet spectra ofdetergent-perturbed complexes. Spectrochim Acta Part A 55:193–204
Osuka A, Kume T, Haggquist GW, Jávorfi T, Lima JC, Melo Eand Naqvi KR (1999) Photophysical characteristics of two modelantenna systems: A fucoxanthin-pyropheophorbide dyad and itsperidinin analogue. Chem Phys Let 313: 499–504
Pfündel E and Bilger W (1994) Regulation and possible function ofthe violaxanthin cycle. Photosynth Res 42: 89–109
Phillip D, Ruban AV, Horton P, Asato A and Young AJ (1996)Quenching of chlorophyll fluorescence in the major light har-vesting complex of Photosystem II. Proc Natl Acad Sci USA 93:1492–1497
Polivka T, Herek JL, Zigmantas D, Akerlund H-E and SundströmV (1999) Direct observation of the (forbidden) S1 state incarotenoids. Proc Natl Acad Sci USA 96: 4914–4917
Rees D, Noctor G, Ruban AV, Crofts J, Young AJ and HortonP (1992) pH dependent chlorophyll fluorescence quenching inspinach thylakoids from light treated or dark adapted leaves.Photosynth Res 31: 11–19
Richter M, Goss R, Wagner B and Holzwarth AR (1999) Char-acterization of the fast and slow reversible components ofnon-photochemical quenching in isolated pea thylakoids by pi-cosecond time resolved chlorophyll fluorescence analysis. Bio-chemistry 38: 12718–12726
Ruban AV, Young AJ and Horton P (1994) Modulation of chloro-phyll fluorescence quenching in isolated light harvesting com-plex of Photosystem II. Biochim Biophys Acta 1186: 123–127
Ruban AV, Young AJ and Horton P (1996) Dynamic propertiesof the minor chlorophyll a/b binding proteins of PhotosystemII, an in vitro model for photoprotective energy dissipation inthe photosynthetic membrane of green plants. Biochemistry 35:674–678
Ruban AV, Calkoen F, Kwa SLS, Van Grondelle R, Horton P andDekker J (1997a) Characterisation of LHC II in the aggregatedstate by linear and circular dichroism spectroscopy. BiochimBiophys Acta 1321: 61–70
Ruban AV, Phillip D, Young AJ and Horton P (1997b) Carotenoid-dependent oligomerization of the major chlorophyll a/b light-harvesting complex of Photosystem-II of plants. Biochemistry36: 7855–7859
Ruban AV, Phillip D, Young AJ and Horton P (1998) Excited-stateenergy level does not determine the differential effect of violax-anthin and zeaxanthin on chlorophyll fluorescence quenching inisolated light-harvesting complex of Photosystem II. PhotochemPhotobiol 68: 829–834
Simidjiev I, Barzda V, Mustárdy L and Garab G (1997) Isolation oflamellar aggregates of the light-harvesting chlorophyll a/b pro-tein complex of Photosystem II with long-range chiral order andstructural flexibility. Anal Biochem 250: 169–175

197
Staehelin LA and Arntzen CJ (1983) Regulation of chloroplastmembrane function: Protein phosphorylation changes the spa-tial organization of membrane components. J Cell Biol 97:1327–1337
Van Metter RL (1977) Excitation energy transfer in the light-harvesting chlorophyll a/b protein. Biochim Biophys Acta 462:642–658
Wagner B, Goss R, Richter M and Holzwarth AR (1996) Pico-second time-resolved study on the nature of high-energy statequenching in isolated pea thylakoids. Different localization ofzeaxanthin dependent and independent quenching mechanisms.J Photochem Photobiol B Biol 36: 339–350
Wilhelm C and Duval JC (1990) Fluorescence induction kinetics asa tool to detect a chlororespiratory activity in the prasinophyceanalga Mantoniella squamata. Biochim Biophys Acta 1016: 197–202
Zer H, Vink M, Keren N, Dilly-Hartwig HG, Paulsen H, HerrmannRG, Andersson B and Ohad I (1999) Regulation of thylakoidprotein phosphorylation at the substrate level: Reversible light-induced conformational changes expose the phosphorylation siteof the light-harvesting complex II. Proc Natl Acad Sci USA 96:8277–8282
Related Documents
![[3,3]-Sigmatropic rearrangements - Massey Universitygjrowlan/stereo2/lecture11.pdf · 123.702 Organic Chemistry Claisen rearrangements • One of the most useful sigmatropic rearrangements](https://static.cupdf.com/doc/110x72/5adcada77f8b9a213e8bd8b0/33-sigmatropic-rearrangements-massey-gjrowlanstereo2lecture11pdf123702.jpg)










