Cell Tissue Res (2004) 316:45–54 DOI 10.1007/s00441-004-0856-z REGULAR ARTICLE M. Sundqvist · S. Holmgren Neurotrophin receptors and enteric neuronal development during metamorphosis in the amphibian Xenopus laevis Received: 19 August 2003 / Accepted: 12 January 2004 / Published online: 20 February 2004 # Springer-Verlag 2004 Abstract During metamorphosis, the frog intestine goes through a dramatic shortening with extensive apoptosis and regeneration in the epithelial layer and connective tissue. Our aim was to study changes in the enteric nervous system represented by one inhibitory (vasoactive intestinal polypeptide; VIP) and one excitatory (sub- stance P, neurokinin A; SP/NKA) nerve population and concomitant changes in neurotrophin receptor occurrence during this development in the gut of Xenopus laevis adults and tadpoles at different stages of metamorphosis (NF stages 57–66). Sections were incubated with anti- bodies against the neurotrophin Trk receptors and p75 NTR , and the neurotransmitters VIP and SP/NKA. Trk-immu- noreactive nerves increased dramatically but transiently in number during early metamorphic climax. Nerves immu- noreactive for p75 NTR were present throughout the gut, decreased in number in the middle intestine during climax, and increased in the large intestine during late metamorphosis. The percentage of VIP-immunoreactive nerves did not change during metamorphosis. SP/NKA- immunoreactive nerves were first apparent at NF stag- es 61–62 in the middle intestine and increased in the stomach and large intestine during metamorphosis. En- docrine cells expressing SP/NKA increased in number in stomach, proximal, and middle intestine during metamor- phic climax. Thus, neurotrophin receptors are expressed transiently in neurons of the enteric nervous system during metamorphosis in Xenopus laevis and SP/NKA innervation is more abundant in the intestine of the postmetamorphic frog than in the tadpole. Keywords Enteric nervous system · p75 · Trk receptors · Tachykinins · Xenopus laevis (Anura) Introduction The amphibian intestine goes through extensive remod- elling during metamorphosis. The larval intestine is long and coiled and is well suited for a herbivorous diet. However, during metamorphosis, the intestine shortens dramatically and the mucosa becomes more folded to suit the more carnivorous diet of the adult frog (Fig. 1). In the African clawed frog, Xenopus laevis, the intestine short- ens to approximately 1/10 of its larval length (Ishizuya- Oka and Shimozawa 1987). The histological changes that occur in both the epithelial layer and the extraepithelial layers of the gut have been studied extensively (Kordylewski 1983; Dauca This study was supported by grants from the Swedish Research Council to S. Holmgren M. Sundqvist ( ) ) · S. Holmgren Department of Zoophysiology, University of Göteborg, Box 463, 405 30 Göteborg, Sweden e-mail: [email protected] Tel.: +46-31-7733694 Fax: +46-31-7733807 Fig. 1 Morphological changes of the tadpole and the tadpole gut during prometamorphosis (NF stages 56–59) and metamorphic climax (NF stages 60–66) in Xenopus laevis (bars relative length of the intestine in the different stages). At NF stage 57–58, the approximate length of the tadpole is 50–60 mm and that of the tadpole gut 150 mm. At NF stage 61–62, the tadpole is approx- imately the same size but the gut is 2/3 shorter. At NF stage 65–66, the froglet is only approximately 25–30 mm having lost its tail and the gut is reduced to 15–20 mm

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cell Tissue Res (2004) 316:45–54DOI 10.1007/s00441-004-0856-z
R E G U L A R A R T I C L E
M. Sundqvist · S. Holmgren
Neurotrophin receptors and enteric neuronal developmentduring metamorphosis in the amphibian Xenopus laevis
Received: 19 August 2003 / Accepted: 12 January 2004 / Published online: 20 February 2004� Springer-Verlag 2004
Abstract During metamorphosis, the frog intestine goesthrough a dramatic shortening with extensive apoptosisand regeneration in the epithelial layer and connectivetissue. Our aim was to study changes in the entericnervous system represented by one inhibitory (vasoactiveintestinal polypeptide; VIP) and one excitatory (sub-stance P, neurokinin A; SP/NKA) nerve population andconcomitant changes in neurotrophin receptor occurrenceduring this development in the gut of Xenopus laevisadults and tadpoles at different stages of metamorphosis(NF stages 57–66). Sections were incubated with anti-bodies against the neurotrophin Trk receptors and p75NTR,and the neurotransmitters VIP and SP/NKA. Trk-immu-noreactive nerves increased dramatically but transiently innumber during early metamorphic climax. Nerves immu-noreactive for p75NTR were present throughout the gut,decreased in number in the middle intestine duringclimax, and increased in the large intestine during latemetamorphosis. The percentage of VIP-immunoreactivenerves did not change during metamorphosis. SP/NKA-immunoreactive nerves were first apparent at NF stag-es 61–62 in the middle intestine and increased in thestomach and large intestine during metamorphosis. En-docrine cells expressing SP/NKA increased in number instomach, proximal, and middle intestine during metamor-phic climax. Thus, neurotrophin receptors are expressedtransiently in neurons of the enteric nervous systemduring metamorphosis in Xenopus laevis and SP/NKAinnervation is more abundant in the intestine of thepostmetamorphic frog than in the tadpole.
Keywords Enteric nervous system · p75 · Trk receptors ·Tachykinins · Xenopus laevis (Anura)
Introduction
The amphibian intestine goes through extensive remod-elling during metamorphosis. The larval intestine is longand coiled and is well suited for a herbivorous diet.However, during metamorphosis, the intestine shortensdramatically and the mucosa becomes more folded to suitthe more carnivorous diet of the adult frog (Fig. 1). In theAfrican clawed frog, Xenopus laevis, the intestine short-ens to approximately 1/10 of its larval length (Ishizuya-Oka and Shimozawa 1987).
The histological changes that occur in both theepithelial layer and the extraepithelial layers of the guthave been studied extensively (Kordylewski 1983; Dauca
This study was supported by grants from the Swedish ResearchCouncil to S. Holmgren
M. Sundqvist ()) · S. HolmgrenDepartment of Zoophysiology,University of G�teborg,Box 463, 405 30 G�teborg, Swedene-mail: [email protected].: +46-31-7733694Fax: +46-31-7733807
Fig. 1 Morphological changes of the tadpole and the tadpole gutduring prometamorphosis (NF stages 56–59) and metamorphicclimax (NF stages 60–66) in Xenopus laevis (bars relative length ofthe intestine in the different stages). At NF stage 57–58, theapproximate length of the tadpole is 50–60 mm and that of thetadpole gut 150 mm. At NF stage 61–62, the tadpole is approx-imately the same size but the gut is 2/3 shorter. At NF stage 65–66,the froglet is only approximately 25–30 mm having lost its tail andthe gut is reduced to 15–20 mm

and Hourdry 1985; Ishizuya-Oka and Shimozawa 1987;Ishizuya-Oka and Ueda 1996). The larval epithelium ofthe intestinal mucosa undergoes apoptosis during meta-morphic climax (NF stages 60–66: Niewkoop and Faber1967) according to Etkin and Gilbert (1968) and isreplaced by adult epithelium in an anterior to posteriorgradient (Ishizuya-Oka et al. 1997). In the connectivetissue, some cells undergo apoptosis but the tissue most-ly proliferates to form a thick layer of submucosa(Kordylewski 1983; Ishizuya-Oka and Shimozawa 1987;Ishizuya-Oka and Ueda 1996). The muscle layers thicken,mainly because of the contraction of the intestine, butadditional cell division and proliferation has been sug-gested to occur in some species (Janes 1934; Kordylewski1983; Dauca and Hourdry 1985). The amphibian stomachepithelium and connective tissue go through the samebasic development as that in the intestine (Ishizuya-Okaand Shimozawa 1987; Rovira et al. 1995).
In the bullfrog, Rana catesbeiana, changes have alsobeen reported in the enteric nervous system (ENS). Anelectron-microscopic study has shown that larval neuronsin the myenteric plexus in the duodenum degenerateduring metamorphosis and are replaced by adult neuronsin this species. The time-course of these developmentalalterations corresponds to the apoptosis and proliferation/differentiation that occurs in the epithelium (Torihashi1990). However, the question arises as to whether thesealterations can be correlated with changes in the expres-sion of neurotransmitters in enteric neurons duringmetamorphosis. This may reflect functional adaptationsof the ENS to the post-metamorphic gastrointestinal tract.
Neuronal development is regulated by neurotrophicfactors in the ENS and in the peripheral and centralnervous systems (for reviews, see Barbacid 1994; Kaplanand Miller 2000; Huang and Reichardt 2001; Patapoutianand Reichardt 2001). The mammalian neurotrophin fam-ily of growth factors includes nerve growth factor (NGF),brain-derived neurotrophic factor (BDNF), neurotrophin 3(NT-3) and neurotrophin 4/5 (NT-4/5). These factors bindto the trk (tropomyosin-related kinase) family of tyrosinekinase receptors comprising Trk A, Trk B and Trk C.NGF binds Trk A, whereas BDNF and NT-4/5 both bindto Trk B, and NT-3 binds to Trk C. NT-3 can also bindTrk A and Trk B with lower affinity (Kaplan et al. 1991a,1991b; Klein et al. 1991a, 1991b; Lamballe et al. 1991).In addition to these high-affinity receptors, the neu-rotrophins activate a low-affinity receptor, p75NTR, whichis a member of the tumour necrosis factor receptorsuperfamily (Johnson et al. 1986; Radeke et al. 1987).The high-affinity receptors support neuronal differentia-tion, growth and survival. For example, BDNF, theforemost ligand for Trk B, is thought to play a role innetwork formation and activity-dependent plasticity inneurons in many parts of the central nervous system (forreviews, see McAllister et al. 1999; Cohen-Cory 2002).Such developmental processes might also be active in theremodelling of the ENS during metamorphosis.
Trk receptors are present on enteric ganglia and glialcells in a number of mammalian species (Hoehner et al.
1996; Esteban et al. 1998) and in the reptilian and teleostgut (Lucini et al. 1999, 2001). In Xenopus laevis, Trk-likereceptors have been detected in the stomach and intestineas early as stage 41 by using immunohistochemistry.Immunoreactivity disappears around stages 48–49 but isonce again observed in adult specimens (Holmberg et al.2001). Similarly, p75NTR has been found in Xenopus inenteric ganglia of the early developing intestine (Hutsonand Bothwell 2001). The role of the low-affinity receptorp75NTR is still controversial; it has been suggested to workas a co-receptor, enhancing and specifying responses ofneurotrophins on Trk receptors but growing evidence alsosuggests a role for the receptor in neuronal apoptosis(Barrett 2000; Bibel and Barde 2000; Miller and Kaplan2001; Roux and Barker 2002).
The neurons of the ENS can contain a number ofdifferent neurotransmitters, for example acetylcholine,adrenaline/noradrenaline, serotonin, gamma-aminobutyr-ic acid, glutamate or nitric oxide. They can also containseveral neuropeptides, including tachykinins, such assubstance P (SP) and neurokinin A (NKA), and vasoac-tive intestinal peptide (VIP), pituitary adenylyl cyclase-activating polypeptide (PACAP), neuropeptide Y, motilinand galanin. Some of these peptides (e.g. SP, NKA) areconsidered to have mostly excitatory effects on otherneurons or on muscle cells, whereas some are consideredpredominantly to generate inhibitory responses (VIP,PACAP; Shuttleworth and Keef 1995; Furness 2000;Olsson and Holmgren 2001).
In this study, we have investigated the expression ofthe neurotrophin high-affinity receptors Trk A, Trk B andTrk C and the low-affinity receptor p75 NTR in the ENSduring metamorphosis in Xenopus laevis. We correlatethis to changes in the excitatory and inhibitory neuronalpopulations represented by NKA/SP and VIP, respective-ly, during this time, with the aim to further our under-standing of enteric neuronal development in the amphib-ian gut. The amount of SP/NKA- or VIP-containingendocrine cells has also been considered, since thisreflects the functionality of these transmitters.
Materials and methods
Animals
Xenopus laevis tadpoles and froglets (n=58) were staged accordingto Niewkoop and Faber (1967). Animals in stages 57–66 werekilled by using an overdose of anaesthetic, i.e. 0.05% MS222(3-aminobenzoic acid ethyl ester; Sigma). Adult frogs wereanaesthetised with MS222 and decapitated. All experiments wereapproved by the animal ethics committee of G�teborg.
Tissues
The gastrointestinal tract was removed and fixed in Zamboni’sfixative, consisting of 15% picric acid, 2% formaldehyde inphosphate-buffered saline (PBS), 0.9% NaCl (pH 7.2), overnight.Subsequently, the tissue was washed repeatedly in 80% ethanol toremove the picric acid and dehydrated by washes in increasinglyconcentrated ethanol, followed by treatment in xylene for 30 min
46

and rehydration in progressively more diluted ethanol and finallyPBS. The tissue was stored (at least overnight) in PBS-sucrosesolution (0.9% NaCl, 30% sucrose in phosphate buffer). Smallsamples were then taken from stomach, proximal intestine (imme-diately after the pyloric sphincter), middle intestine (at approxi-mately 40%–50% of the length of the total small intestine) andlarge intestine. The samples were embedded in OCT (Sakura),frozen in isopentane cooled by liquid nitrogen and later cut intosections (10 mm thick) on a cryostat (Zeiss). The sections werepicked up on gelatine-coated slides and left to air-dry overnight.
Immunohistochemistry
Prior to incubation with the primary antibody, the sections werepre-incubated for 30 min with normal donkey serum. The sectionswere then incubated at room temperature overnight with primaryantibody (see Table 1). They were rinsed (3� 5 minutes) in high-salt PBS (2% NaCl) and subsequently incubated for 1 h with asecondary antibody conjugated to a fluorescent molecule (seeTable 1). After incubation, the sections were rinsed again, mountedin Vectashield mounting medium (Vector Laboratories, Burlin-game, Calif.) and studied by fluorescence microscopy (NikonEclipse E1000). Micrographs were taken with a digital camera(Nikon DXM1200) and further examined by using the Nikonsoftware image analysis programs Easy Image measurements andEasy Image analysis 2000. Double-staining was performed by usingthe antibody combinations SP-GP/Trk B and VIP-GP/Trk B. Sincethe p75NTR antibody and the Trk B antibody had been raised in thesame animal species, consecutive sections were stained to obtainthe extent of the co-expression of the receptors. Double-staining forenterochromaffin (EC) cells was performed by using NKA-2809/serotonin (5-HT) and VIP-MI/5-HT.
Absorption tests
Absorption tests for Trk A, Trk B and Trk C antisera, with thematching peptide and with the other Trk peptides, have beenperformed previously in our laboratory. The Trk A and Trk Cantibodies do not cross-react with the other peptides, whereas theantibody for the Trk B receptor cross-reacts with the Trk A andTrk C peptides in amphibian tissue (Holmberg et al. 2001). Thisimmunoreactivity will thus be referred to as Trk immunoreactivity.
No blocking peptide was available for the p75NTR antibody.However, the antibody used in the experiments was directed againstamino acids 266-357 of the p75NTR receptor, which includes thetransmembrane region of the receptor. This region is highly
conserved through evolution (Hutson and Bothwell 2001). Anabsorption test was also performed with the Trk A, Trk B and Trk Cpeptides; no cross-reaction with the p75NTR antibody in Xenopuslaevis tissue was found.
The NKA and SP antibodies used react with both SP and NKAand this immunoreactivity will be referred to as SP/NKA immu-noreactivity (Holmberg et al. 2001).
Analysis and statistics
Analyses were performed in the software Easy Image analysis 2000by measuring the area marked with the fluorescent antibody. Thearea represented by the muscle tissue in the micrographs was alsomeasured and the two areas were related to each other to adjust forthe sections being of different sizes depending on the develop-mental stage. Since the amount of neurons as a whole changedduring metamorphosis, the immunoreactivities to Trk, p75NTR,NKA/SP and VIP were related to acetylated tubulin (AcT), whichhas previously been used as a general neuronal marker (see Chitnisand Kuwada 1990; LeDizet and Piperno 1991; Hartenstein 1993;Eagleson et al. 1998; Schlosser 2003).
Endocrine cells were counted and related to the total number ofepithelial cells in the same image. The nucleus of the cells wasvisualised by using Vectashield mounting medium with 4,6diamino-2-phenyl-indole (Vector Laboratories). Endocrine cellsthat were labelled with 5-HT were referred to as EC cells.
Data from consecutive stages were grouped two by two. Alldata are shown as the mean € SEM. The data were statisticallycompared by using a one-way analysis of variance (ANOVA)followed by a Dunnett’s post hoc test comparing all stages with thestate at the beginning of metamorphosis (stages 57–58, alsoreferred to as prometamorphosis; Etkin and Gilbert 1968). Statis-tical comparisons were performed by using GraphPad Prism 3.0(GraphPad Software, San Diego, USA).
Results
General morphological changes
In the stomach, the muscular layer increased in thicknessthroughout the metamorphic climax and was significantlythicker at stages 61–66 than during prometamorphosis(stages 57–58; Fig. 2A). In the intestine, the muscularlayer followed the same general pattern (Fig. 3A, B),
Table 1 Summary of primaryand secondary antibodies used(DaR donkey anti-rabbit, DaMdonkey anti-mouse, DaGPdonkey anti-guinea-pig, DaGdonkey anti-goat)
Antiserum Code Host Dilution Source
Primary antiserum againstAcT(sea-urchin) acetylated tubulin T-6793 Mouse 1:1000 SigmaTrk A (human) SC-118-G Rabbit 1:200 Santa CruzTrk B (human) SC-12 Rabbit 1:200 Santa CruzTrk C (human) SC-117 Rabbit 1:200 Santa CruzNGFR p75 (human) SC-5634 Rabbit 1:40 Santa CruzNKA 2809 (rainbow trout, cod) NKA2809 Rabbit 1:6400 J�rgen Jensen,
G�teborg UniversitySP-GP (human) B-GP 450-1 Guinea-pig 1:400 Euro-DiagnosticaVIP-GP (synthetic, human) B-GP 340-X Guinea-pig 1:800 Euro-DiagnosticaVIP-MI (porcine) B-340 Rabbit 1:1000 Milab5-HT (human) 20079 Goat 1:200 DiasorinSecondary antiserum againstDaR-FITC (rabbit IgG) 711-095-151 Donkey 1:100 JacksonDaM-FITC (rabbit IgG) 715-095-150 Donkey 1:100 JacksonDaR-Cy3 (rabbit IgG) 711-165-152 Donkey 1:800 JacksonDaGP-DTAF (guinea-pig IgG) 706-015-148 Donkey 1:100 JacksonDaG-DTAF (goat IgG) 705-016-147 Donkey 1:100 JacksonDaG-Cy3 (goat IgG) 705-165-147 Donkey 1:800 Jackson
47
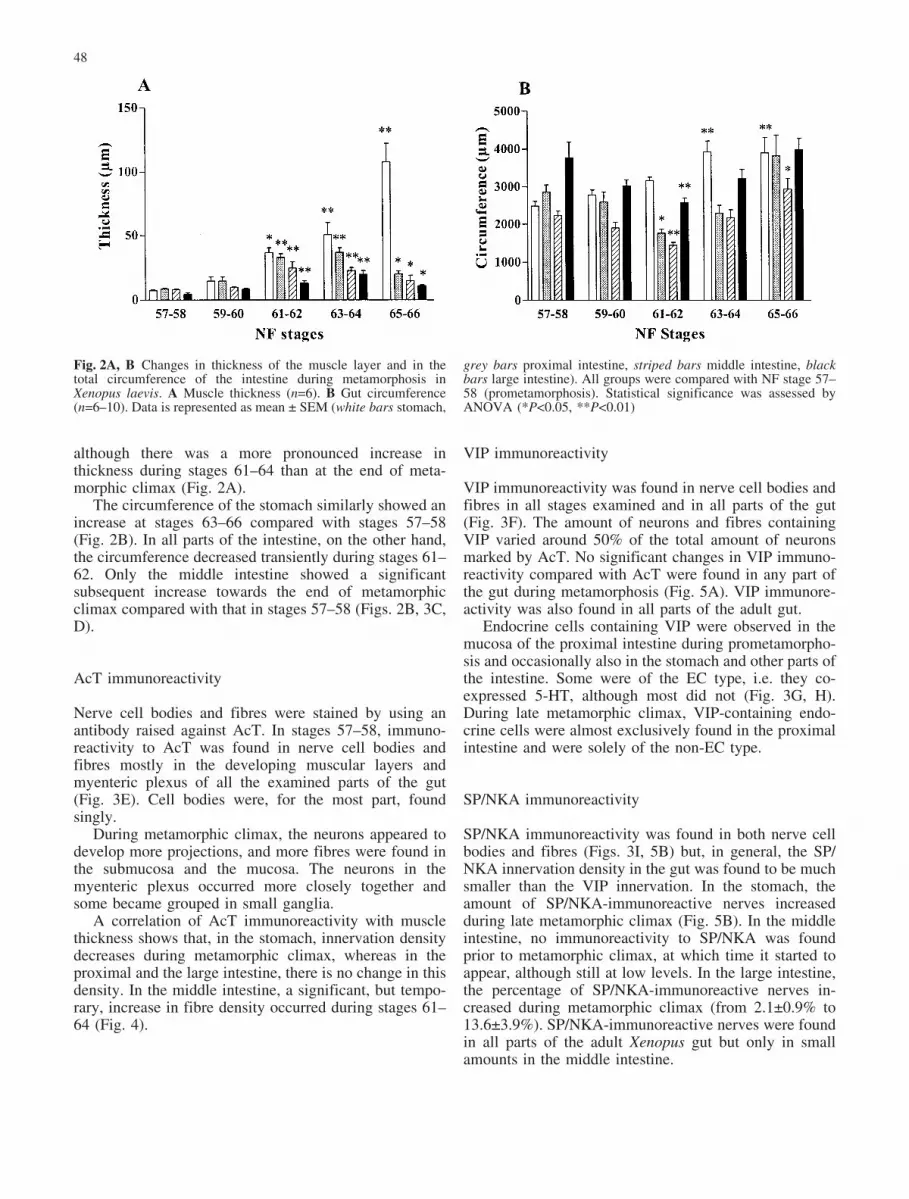
although there was a more pronounced increase inthickness during stages 61–64 than at the end of meta-morphic climax (Fig. 2A).
The circumference of the stomach similarly showed anincrease at stages 63–66 compared with stages 57–58(Fig. 2B). In all parts of the intestine, on the other hand,the circumference decreased transiently during stages 61–62. Only the middle intestine showed a significantsubsequent increase towards the end of metamorphicclimax compared with that in stages 57–58 (Figs. 2B, 3C,D).
AcT immunoreactivity
Nerve cell bodies and fibres were stained by using anantibody raised against AcT. In stages 57–58, immuno-reactivity to AcT was found in nerve cell bodies andfibres mostly in the developing muscular layers andmyenteric plexus of all the examined parts of the gut(Fig. 3E). Cell bodies were, for the most part, foundsingly.
During metamorphic climax, the neurons appeared todevelop more projections, and more fibres were found inthe submucosa and the mucosa. The neurons in themyenteric plexus occurred more closely together andsome became grouped in small ganglia.
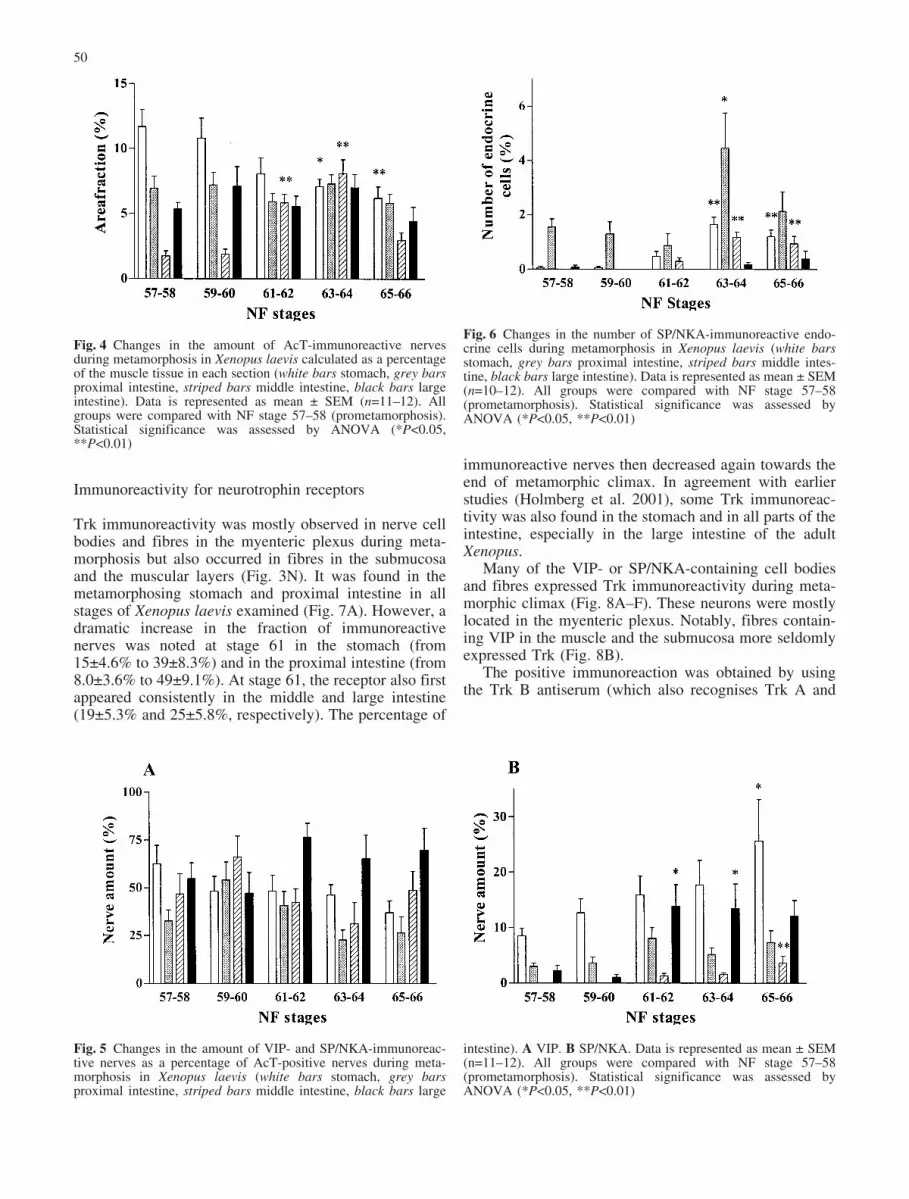
A correlation of AcT immunoreactivity with musclethickness shows that, in the stomach, innervation densitydecreases during metamorphic climax, whereas in theproximal and the large intestine, there is no change in thisdensity. In the middle intestine, a significant, but tempo-rary, increase in fibre density occurred during stages 61–64 (Fig. 4).
VIP immunoreactivity
VIP immunoreactivity was found in nerve cell bodies andfibres in all stages examined and in all parts of the gut(Fig. 3F). The amount of neurons and fibres containingVIP varied around 50% of the total amount of neuronsmarked by AcT. No significant changes in VIP immuno-reactivity compared with AcT were found in any part ofthe gut during metamorphosis (Fig. 5A). VIP immunore-activity was also found in all parts of the adult gut.
Endocrine cells containing VIP were observed in themucosa of the proximal intestine during prometamorpho-sis and occasionally also in the stomach and other parts ofthe intestine. Some were of the EC type, i.e. they co-expressed 5-HT, although most did not (Fig. 3G, H).During late metamorphic climax, VIP-containing endo-crine cells were almost exclusively found in the proximalintestine and were solely of the non-EC type.
SP/NKA immunoreactivity
SP/NKA immunoreactivity was found in both nerve cellbodies and fibres (Figs. 3I, 5B) but, in general, the SP/NKA innervation density in the gut was found to be muchsmaller than the VIP innervation. In the stomach, theamount of SP/NKA-immunoreactive nerves increasedduring late metamorphic climax (Fig. 5B). In the middleintestine, no immunoreactivity to SP/NKA was foundprior to metamorphic climax, at which time it started toappear, although still at low levels. In the large intestine,the percentage of SP/NKA-immunoreactive nerves in-creased during metamorphic climax (from 2.1€0.9% to13.6€3.9%). SP/NKA-immunoreactive nerves were foundin all parts of the adult Xenopus gut but only in smallamounts in the middle intestine.
Fig. 2A, B Changes in thickness of the muscle layer and in thetotal circumference of the intestine during metamorphosis inXenopus laevis. A Muscle thickness (n=6). B Gut circumference(n=6–10). Data is represented as mean € SEM (white bars stomach,
grey bars proximal intestine, striped bars middle intestine, blackbars large intestine). All groups were compared with NF stage 57–58 (prometamorphosis). Statistical significance was assessed byANOVA (*P<0.05, **P<0.01)
48

Endocrine cells containing SP/NKA were most com-mon in the proximal intestine during prometamorphosis(Figs. 6, 3L). The cells were both of the EC type,expressing 5-HT, and the non-EC type, without 5-HT(Fig. 3J, K). In the stomach and in the middle and largeintestine, these cells were scarce. At stages 63–64, thenumber of endocrine cells increased and, at this time, theyalso appeared more consistently in the stomach andmiddle intestine. The increase of endocrine cells in thestomach and middle intestine was permanent, whereas theincrease in the proximal intestine was only transient and,at stage 65–66, the levels returned to prometamorphicvalues. In some sections from stages 63–65, clusters of
cells intensely stained for SP/NKA could be found in thesubmucosa and/or in the mucosa (Fig. 3M). No suchclusters were found in stage 66 or in adult tissue. In thelarge intestine, endocrine cells expressing SP/NKA werefound sporadically from stage 63, whereas in the adultfrog, the population was more established. The populationof endocrine cells in the large intestine was always of theEC type.
Fig. 3A–O Gut sections from the amphibian Xenopus laevis duringvarious stages of metamorphosis. A, B Middle intestine at stages 57and 61, respectively (green actin in muscular layers, white bracketsthickness of muscular layers). C, D Proximal intestine at stages 57and 61, respectively. Notice the different sizes of the proximalintestine at different times during metamorphosis (green actin inthe muscular layers). E AcT-immunoreactive nerve fibres in theproximal intestine at stage 62. F VIP immunoreactivity in theproximal intestine at stage 62 (filled arrowhead nerve fibres, openarrowheads endocrine cells). Some unspecific immunostaining ofmucus is seen on the surface of the mucosa. G, H Two VIP-
immunoreactive endocrine cells in the proximal intestine atstage 57, one co-expressing 5-HT (H). I SP/NKA immunorectivityin the proximal intestine at stage 64 (filled arrowheads nerve fibres,open arrowhead endocrine cell). J, K Enterochromaffin cell co-expressing SP/NKA and 5-HT in the middle intestine, stage 64. LSP/NKA-immunoreactive endocrine cells (arrowheads) in theproximal intestine at stage 59. M SP/NKA-immunoreactive endo-crine cells in the proximal intestine at stage 64. N Trk-immuno-reactive cell body and fibre in the stomach at stage 61 (arrow-heads). O p75NTR-immunoreactive fibre and cell body (arrow-heads) in the stomach at stage 62. Bar 50 �m
49
Immunoreactivity for neurotrophin receptors
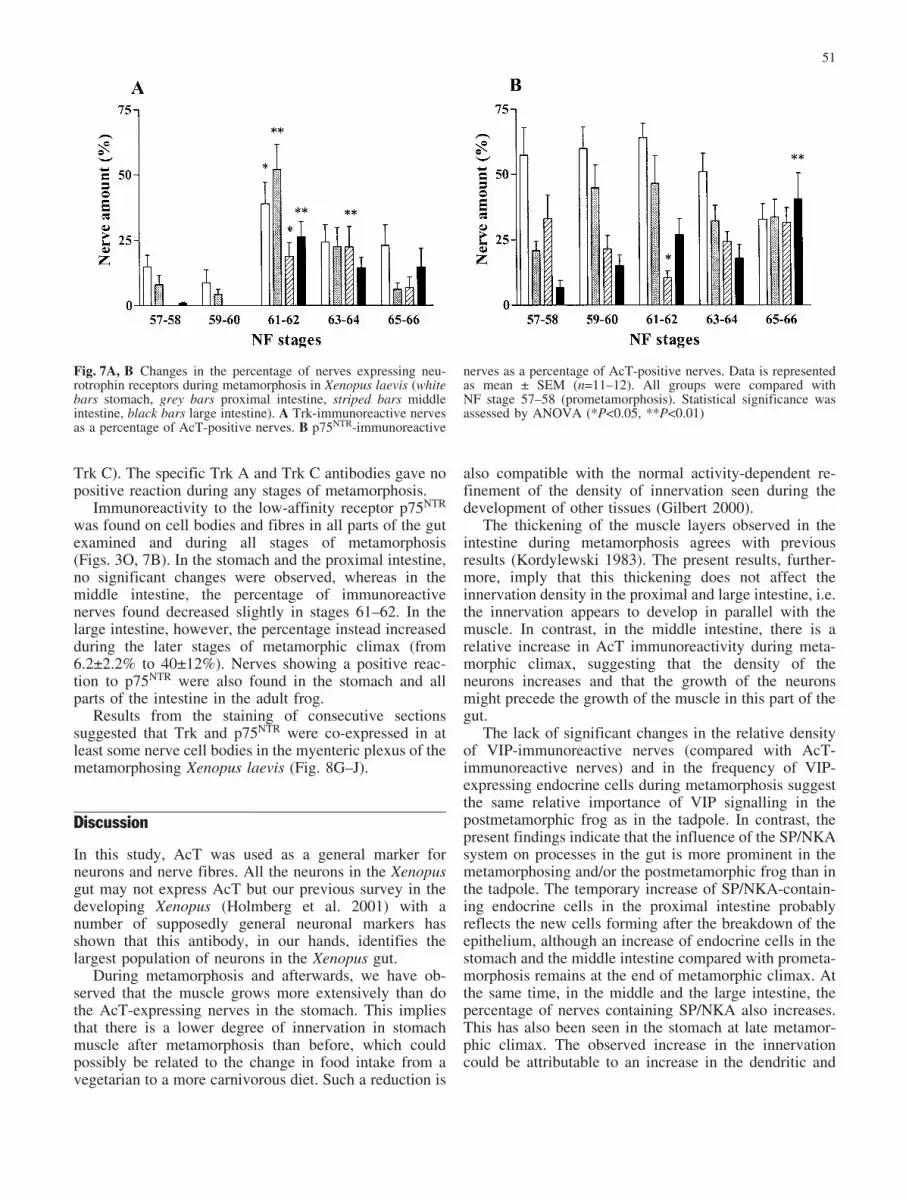
Trk immunoreactivity was mostly observed in nerve cellbodies and fibres in the myenteric plexus during meta-morphosis but also occurred in fibres in the submucosaand the muscular layers (Fig. 3N). It was found in themetamorphosing stomach and proximal intestine in allstages of Xenopus laevis examined (Fig. 7A). However, adramatic increase in the fraction of immunoreactivenerves was noted at stage 61 in the stomach (from15€4.6% to 39€8.3%) and in the proximal intestine (from8.0€3.6% to 49€9.1%). At stage 61, the receptor also firstappeared consistently in the middle and large intestine(19€5.3% and 25€5.8%, respectively). The percentage of
immunoreactive nerves then decreased again towards theend of metamorphic climax. In agreement with earlierstudies (Holmberg et al. 2001), some Trk immunoreac-tivity was also found in the stomach and in all parts of theintestine, especially in the large intestine of the adultXenopus.
Many of the VIP- or SP/NKA-containing cell bodiesand fibres expressed Trk immunoreactivity during meta-morphic climax (Fig. 8A–F). These neurons were mostlylocated in the myenteric plexus. Notably, fibres contain-ing VIP in the muscle and the submucosa more seldomlyexpressed Trk (Fig. 8B).
The positive immunoreaction was obtained by usingthe Trk B antiserum (which also recognises Trk A and
Fig. 5 Changes in the amount of VIP- and SP/NKA-immunoreac-tive nerves as a percentage of AcT-positive nerves during meta-morphosis in Xenopus laevis (white bars stomach, grey barsproximal intestine, striped bars middle intestine, black bars large
intestine). A VIP. B SP/NKA. Data is represented as mean € SEM(n=11–12). All groups were compared with NF stage 57–58(prometamorphosis). Statistical significance was assessed byANOVA (*P<0.05, **P<0.01)
Fig. 4 Changes in the amount of AcT-immunoreactive nervesduring metamorphosis in Xenopus laevis calculated as a percentageof the muscle tissue in each section (white bars stomach, grey barsproximal intestine, striped bars middle intestine, black bars largeintestine). Data is represented as mean € SEM (n=11–12). Allgroups were compared with NF stage 57–58 (prometamorphosis).Statistical significance was assessed by ANOVA (*P<0.05,**P<0.01)
Fig. 6 Changes in the number of SP/NKA-immunoreactive endo-crine cells during metamorphosis in Xenopus laevis (white barsstomach, grey bars proximal intestine, striped bars middle intes-tine, black bars large intestine). Data is represented as mean € SEM(n=10–12). All groups were compared with NF stage 57–58(prometamorphosis). Statistical significance was assessed byANOVA (*P<0.05, **P<0.01)
50
Trk C). The specific Trk A and Trk C antibodies gave nopositive reaction during any stages of metamorphosis.
Immunoreactivity to the low-affinity receptor p75NTR
was found on cell bodies and fibres in all parts of the gutexamined and during all stages of metamorphosis(Figs. 3O, 7B). In the stomach and the proximal intestine,no significant changes were observed, whereas in themiddle intestine, the percentage of immunoreactivenerves found decreased slightly in stages 61–62. In thelarge intestine, however, the percentage instead increasedduring the later stages of metamorphic climax (from6.2€2.2% to 40€12%). Nerves showing a positive reac-tion to p75NTR were also found in the stomach and allparts of the intestine in the adult frog.
Results from the staining of consecutive sectionssuggested that Trk and p75NTR were co-expressed in atleast some nerve cell bodies in the myenteric plexus of themetamorphosing Xenopus laevis (Fig. 8G–J).
Discussion
In this study, AcT was used as a general marker forneurons and nerve fibres. All the neurons in the Xenopusgut may not express AcT but our previous survey in thedeveloping Xenopus (Holmberg et al. 2001) with anumber of supposedly general neuronal markers hasshown that this antibody, in our hands, identifies thelargest population of neurons in the Xenopus gut.
During metamorphosis and afterwards, we have ob-served that the muscle grows more extensively than dothe AcT-expressing nerves in the stomach. This impliesthat there is a lower degree of innervation in stomachmuscle after metamorphosis than before, which couldpossibly be related to the change in food intake from avegetarian to a more carnivorous diet. Such a reduction is
also compatible with the normal activity-dependent re-finement of the density of innervation seen during thedevelopment of other tissues (Gilbert 2000).
The thickening of the muscle layers observed in theintestine during metamorphosis agrees with previousresults (Kordylewski 1983). The present results, further-more, imply that this thickening does not affect theinnervation density in the proximal and large intestine, i.e.the innervation appears to develop in parallel with themuscle. In contrast, in the middle intestine, there is arelative increase in AcT immunoreactivity during meta-morphic climax, suggesting that the density of theneurons increases and that the growth of the neuronsmight precede the growth of the muscle in this part of thegut.
The lack of significant changes in the relative densityof VIP-immunoreactive nerves (compared with AcT-immunoreactive nerves) and in the frequency of VIP-expressing endocrine cells during metamorphosis suggestthe same relative importance of VIP signalling in thepostmetamorphic frog as in the tadpole. In contrast, thepresent findings indicate that the influence of the SP/NKAsystem on processes in the gut is more prominent in themetamorphosing and/or the postmetamorphic frog than inthe tadpole. The temporary increase of SP/NKA-contain-ing endocrine cells in the proximal intestine probablyreflects the new cells forming after the breakdown of theepithelium, although an increase of endocrine cells in thestomach and the middle intestine compared with prometa-morphosis remains at the end of metamorphic climax. Atthe same time, in the middle and the large intestine, thepercentage of nerves containing SP/NKA also increases.This has also been seen in the stomach at late metamor-phic climax. The observed increase in the innervationcould be attributable to an increase in the dendritic and
Fig. 7A, B Changes in the percentage of nerves expressing neu-rotrophin receptors during metamorphosis in Xenopus laevis (whitebars stomach, grey bars proximal intestine, striped bars middleintestine, black bars large intestine). A Trk-immunoreactive nervesas a percentage of AcT-positive nerves. B p75NTR-immunoreactive
nerves as a percentage of AcT-positive nerves. Data is representedas mean € SEM (n=11–12). All groups were compared withNF stage 57–58 (prometamorphosis). Statistical significance wasassessed by ANOVA (*P<0.05, **P<0.01)
51

axonal arborisation of intrinsic (enteric) nerves and alsoof extrinsic nerves in parts of the gut.
The transient, but prominent, relative increase in Trk-immunoreactive neurons, mainly in the myenteric plexus,which occurs during metamorphic climax, indicates aperiod of neuronal development. Such a developmentcould involve the growth and rearragement of existingneurons or the differentiation of precursor cells intopresumably more adult-type neurons, as in Rana cates-beiana (Torihashi 1990).
The Trk receptors in the metamorphosing Xenopus arepossibly of the Trk B-like type. We have found noimmunoreactivity in any of the sections examined when
using specific antibodies raised against the human Trk Aand Trk C receptors. This may indicate a true lack ofexpression of such receptors during the investigatedstages but it is also possible that the Trk A and Trk Cantibodies that we used were unable to recognize aXenopus counterpart of these receptors. Furthermore,there is an uncertainty as to the identity of the moleculereacting with the Trk B antiserum. This antiserum wasraised against human Trk B but was found, in absorptiontests, to react with all of the three Trk receptors.Nevertheless, we suggest that, as previously proposedfor the early developmental stages in Xenopus (NF 38-49;
Fig. 8A–J Gut sections from the amphibian Xenopus laevis duringvarious stages of metamorphosis. A VIP (green, open arrowheads),Trk (red, filled arrowhead) and co-expression (yellow, stripedarrowhead). Stomach at stage 61. B VIP (green, open arrowhead),Trk (red, filled arrowhead) and co-expression (yellow). Most VIP-containing fibres in the muscle layer do not express Trk. Proximalintestine at stage 62. C Co-expression of SP/NKA and Trk (yellow,arrowhead). Stomach, stage 61. D Same neuron as in C, showing
Trk immunorectivity (arrowhead). E Same neuron as in C, D,showing SP/NKA immunorectivity (arrowhead). F SP/NKA(green), Trk (red) and co-expression (yellow). Some SP/NKAfibres do not express Trk (arrowhead). Stomach, stage 63. G, HProximal intestine at stage 62. I, J Middle intestine at stage 64.Consecutive sections in G, H and in I, J show Trk and p75NTR
immunorectivity in the same neuronal cell bodies (arrowsheads).Bar 50 �m
52

Holmberg et al. 2001), the receptor is more Trk B-likethan Trk A-like or Trk C-like.
In humans, Trk C is expressed in neurons in the gut,and Trk A and Trk B in both neurons and glia (Hoehner etal. 1996; Esteban et al. 1998). On the other hand, inteleost and reptilian gut, Trk A-like and Trk C-likereceptors, but no Trk B-like receptors, have been found sofar (Lucini et al. 1999, 2001; Hannestad et al. 2000).However, these results do not preclude the possibility thatTrk B receptors are expressed in the amphibian gut, atleast at this specific time of development. Further studies,preferably involving genetic analyses, are needed in orderto be able to make conclusions about evolutionary trends.
Expression of the p75NTR receptor has been determinedduring the early development of the Xenopus gut (NFstage 45/46; Hutson and Bothwell 2001). The compara-tively high levels of the p75NTR receptor during all stagesof metamorphosis found in the present study suggest thecontinued physiological significance of p75NTR signallingduring this period. However, it is difficult to speculate ona specific function in Xenopus from the present results,with no regional or temporal common trends in therelative changes of p75NTR-expressing neurons in the gut.
In mammals, the p75NTR receptor has been suggestedto enhance responses to neurotrophins in low concentra-tions (Hantzopoulos et al. 1994; Roux and Barker 2002)and to make the Trk receptors more specific in theirbinding to the preferred ligand (Ip et al. 1993; Bibel et al.1999). In vitro, p75NTR stimulates axonal outgrowthand can, indeed, be essential for the differentiation ofsome neuronal precursor cells that also express Trk B(Yamashita et al. 1999; Hosomi et al. 2003). This couldbe one of the potential rationales for the simultaneousexpression of p75NTR receptors and Trk receptors onnerve cells in the developing amphibian gut. On the otherhand, a number of studies have demonstrated that, undercertain circumstances, p75NTR may have a role in apop-tosis (for reviews, see Barrett 2000; Bibel and Barde2000; Miller and Kaplan 2001; Roux and Barker 2002), aprocess that may indeed be involved in the rearragementof the ENS during metamorphosis in Xenopus (cf. Ranacatesbeiana; Torihashi 1990).
In summary, we have demonstrated that differentnerve populations undergo different developmentalchanges during metamorphosis. SP/NKA appears to bea more prominent transmitter in the postmetamorphicXenopus gut than in the tadpole, in contrast to VIP, whichdoes not change its relative innervation density duringmetamorphosis. The development is also region-specific.Finally, neuronal development during metamorphosismight be regulated, in part, by neurotrophins actingthrough Trk receptors and may also be influenced by thepresence of the p75NTR receptor.
Acknowledgements We thank Mrs. Christina Hagstr�m and Ms.Anna Holmberg for their valuable expert help with the immuno-histochemistry experiments.
References
Barbacid M (1994) The Trk family of neurotrophin receptors. JNeurobiol 25:1386–1403
Barrett GL (2000) The p75 neurotrophin receptor and neuronalapoptosis. Prog Neurobiol 61:205–229
Bibel M, Barde YA (2000) Neurotrophins: key regulators of cellfate and cell shape in the vertebrate nervous system. Genes Dev14:2919–2937
Bibel M, Hoppe E, Barde YA (1999) Biochemical and functionalinteractions between the neurotrophin receptors trk andp75NTR. Embo J 18:616–622
Chitnis AB, Kuwada JY (1990) Axonogenesis in the brain ofzebrafish embryos. J Neurosci 10:1892–1905
Cohen-Cory S (2002) The developing synapse: construction andmodulation of synaptic structures and circuits. Science298:770–776
Dauca M, Hourdry J (1985) Transformations in the intestinalepithelium during anuran metamorphosis. In: Metamorphosis:the Eighth Symposium of the British Society for Developmen-tal Biology. Oxford University Press, New York, pp 37–58
Eagleson GW, Ubink R, Jenks BG, Roubos EW (1998) Forebraindifferentiation and axonogenesis in amphibians. I. Differenti-ation of the suprachiasmatic nucleus in relation to backgroundadaptation behavior. Brain Behav Evol 52:23–36
Esteban I, Levanti B, Garcia-Suarez O, Germana G, Ciriaco E,Naves FJ, Vega JA (1998) A neuronal subpopulation in themammalian enteric nervous system expresses TrkA and TrkCneurotrophin receptor-like proteins. Anat Rec 251:360–370
Etkin W, Gilbert LI (1968) Metamorphosis. A problem in devel-opmental biology. Appleton-Century-Crofts, New York
Furness JB (2000) Types of neurons in the enteric nervous system. JAuton Nerv Syst 81:87–96
Gilbert SF (2000) Developmental biology. Sinauer, Sunderland,Mass.
Hannestad J, Marino F, Germana A, Catania S, Abbate F, CiriacoE, Vega JA (2000) Trk neurotrophin receptor-like proteins inthe teleost Dicentrarchus labrax. Cell Tissue Res 300:1–9
Hantzopoulos PA, Suri C, Glass DJ, Goldfarb MP, YancopoulosGD (1994) The low affinity NGF receptor, p75, can collaboratewith each of the Trks to potentiate functional responses to theneurotrophins. Neuron 13:187–201
Hartenstein V (1993) Early pattern of neuronal differentiation in theXenopus embryonic brainstem and spinal cord. J Comp Neurol328:213–231
Hoehner JC, Wester T, Pahlman S, Olsen L (1996) Localization ofneurotrophins and their high-affinity receptors during humanenteric nervous system development. Gastroenterology110:756–767
Holmberg A, Hagg U, Fritsche R, Holmgren S (2001) Occurrenceof neurotrophin receptors and transmitters in the developingXenopus gut. Cell Tissue Res 306:35–47
Hosomi S, Yamashita T, Aoki M, Tohyama M (2003) The p75receptor is required for BDNF-induced differentiation of neuralprecursor cells. Biochem Biophys Res Commun 301:1011–1015
Huang EJ, Reichardt LF (2001) Neurotrophins: roles in neuronaldevelopment and function. Annu Rev Neurosci 24:677–736
Hutson LD, Bothwell M (2001) Expression and function ofXenopus laevis p75(NTR) suggest evolution of developmentalregulatory mechanisms. J Neurobiol 49:79–98
Ip NY, Stitt TN, Tapley P, Klein R, Glass DJ, Fandl J, Greene LA,Barbacid M, Yancopoulos GD (1993) Similarities and differ-ences in the way neurotrophins interact with the Trk receptorsin neuronal and nonneuronal cells. Neuron 10:137–149
Ishizuya-Oka A, Shimozawa A (1987) Development of the con-nective tissue in the digestive tract of the larval and metamor-phosing Xenopus laevis. Anat Anz 164:81–93
Ishizuya-Oka A, Ueda S (1996) Apoptosis and cell proliferation inthe Xenopus small intestine during metamorphosis. Cell TissueRes 286:467–476
53

Ishizuya-Oka A, Ueda S, Damjanovski S, Li Q, Liang VC, Shi YB(1997) Anteroposterior gradient of epithelial transformationduring amphibian intestinal remodeling: immunohistochemicaldetection of intestinal fatty acid-binding protein. Dev Biol192:149–161
Janes RG (1934) Studies on the amphibian digestive system. I.Histological changes in the alimentary tract of anuran larvaeduring involution. J Exp Zool 67:73–91
Johnson D, Lanahan A, Buck CR, Sehgal A, Morgan C, Mercer E,Bothwell M, Chao M (1986) Expression and structure of thehuman NGF receptor. Cell 47:545–554
Kaplan DR, Miller FD (2000) Neurotrophin signal transduction inthe nervous system. Curr Opin Neurobiol 10:381–391
Kaplan DR, Hempstead BL, Martin-Zanca D, Chao MV, Parada LF(1991a) The trk proto-oncogene product: a signal transducingreceptor for nerve growth factor. Science 252:554–558
Kaplan DR, Martin-Zanca D, Parada LF (1991b) Tyrosine phos-phorylation and tyrosine kinase activity of the trk proto-oncogene product induced by NGF. Nature 350:158–160
Klein R, Jing SQ, Nanduri V, O’Rourke E, Barbacid M (1991a)The trk proto-oncogene encodes a receptor for nerve growthfactor. Cell 65:189–197
Klein R, Nanduri V, Jing SA, Lamballe F, Tapley P, Bryant S,Cordon-Cardo C, Jones KR, Reichardt LF, Barbacid M (1991b)The trkB tyrosine protein kinase is a receptor for brain-derivedneurotrophic factor and neurotrophin-3. Cell 66:395–403
Kordylewski L (1983) Light and electron microscopic observationsof the development of intestinal musculature in Xenopus. ZMikrosk Anat Forsch 97:719–734
Lamballe F, Klein R, Barbacid M (1991) TrkC, a new member ofthe trk family of tyrosine protein kinases, is a receptor forneurotrophin-3. Cell 66:967–979
LeDizet M, Piperno G (1991) Detection of acetylated alpha-tubulinby specific antibodies. Methods Enzymol 196:264–274
Lucini C, Girolamo P de, Maruccio L, Lamanna C, Castaldo L,Vega JA (1999) Trk-neurotrophin receptor-like immunoreac-tivity in the gut of teleost species. Cell Tissue Res 296:323–330
Lucini C, Girolamo P de, Lamanna C, Botte V, Vega JA, CastaldoL (2001) TrkA and TrkC neurotrophin receptor-like proteins inthe lizard gut. Cell Tissue Res 303:345–350
McAllister AK, Katz LC, Lo DC (1999) Neurotrophins andsynaptic plasticity. Annu Rev Neurosci 22:295–318
Miller FD, Kaplan DR (2001) Neurotrophin signalling pathwaysregulating neuronal apoptosis. Cell Mol Life Sci 58:1045–1053
Niewkoop PD, Faber J (1967) A normal table of Xenopus laevis(Daudin): a systematical and chronological survey of thedevelopment from the fertilized egg till the end of metamor-phosis. North-Holland, Amsterdam
Olsson C, Holmgren S (2001) The control of gut motility. CompBiochem Physiol A Mol Integr Physiol 128:481–503
Patapoutian A, Reichardt LF (2001) Trk receptors: mediators ofneurotrophin action. Curr Opin Neurobiol 11:272–280
Radeke MJ, Misko TP, Hsu C, Herzenberg LA, Shooter EM (1987)Gene transfer and molecular cloning of the rat nerve growthfactor receptor. Nature 325:593–597
Roux PP, Barker PA (2002) Neurotrophin signaling through thep75 neurotrophin receptor. Prog Neurobiol 67:203–233
Rovira J, Villaro AC, Bodegas ME, Valverde E, Sesma P (1995)Metamorphic changes in the stomach of the frog Ranatemporaria tadpoles. Tissue Cell 27:13–22
Schlosser G (2003) Hypobranchial placodes in Xenopus laevis giverise to hypobranchial ganglia, a novel type of cranial ganglia.Cell Tissue Res 312:21–29
Shuttleworth CW, Keef KD (1995) Roles of peptides in entericneuromuscular transmission. Regul Pept 56:101–120
Torihashi S (1990) Morphological changes of the myenteric plexusneurons in the bullfrog (Rana catesbeiana) duodenum duringmetamorphosis. J Comp Neurol 302:54–65
Yamashita T, Tucker KL, Barde YA (1999) Neurotrophin bindingto the p75 receptor modulates Rho activity and axonaloutgrowth. Neuron 24:585–593
54
Related Documents











