IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007 1309 Neuronal Network Morphology and Electrophysiologyof Hippocampal Neurons Cultured on Surface-Treated Multielectrode Arrays Walid V. Soussou*, Geoffrey J. Yoon, Roberta Diaz Brinton, and Theodore W. Berger, Senior Member, IEEE Abstract—Toward the development of biocompatible surfaces for implantable electrode arrays and the creation of patterned neuronal networks, the impact of select biochemical substrates [poly-D-lysine (PDL), polyornithine (PO), polyethylenimine (PEI), and a basement membrane extract (BM)] on network morphology and spontaneous electrophysiological activity of dissociated hippocampal neurons was investigated. Cultured in serum-free Neurobasal medium at 100 000 , neurons attached to each substrate. PDL, PO, and PEI induced little or no neuronal clustering and process fasciculation, whereas the addition of BM promoted these features. The ratios of somas to processes, and axons to dendrites, as determined by immunohistochemical staining and image analysis were comparable across all substrates. Spontaneous firing was recorded using planar multielectrode ar- rays (MEAs) at the third week in vitro for the two most divergent morphologies according to Euclidian cluster analysis, namely those induced by PO BM and PEI. Mean spike amplitude, mean firing rate, median interspike interval (ISI), mean burst rate, and correlation index were analyzed and compared to morphological features. Synchronized bursting was highly correlated with neu- ronal clustering and process fasciculation. Spike amplitude was negatively correlated with thin branching which was most evident in neurons grown on PEI. These data indicate that factors, which influence adherence of neurons to surfaces, can profoundly impact both neuronal network morphology and electrophysiological activity in vitro. Index Terms—Basement membrane, multielectrode array, neuronal network morphology, spontaneous electrophysiological activity. Manuscript received December 16, 2005; revised November 1, 2006. Asterisk indicates corresponding author. *W. V. Soussou is with the Neuroscience Program, University of Southern California, Los Angeles, CA 90089-2520 USA (e-mail: [email protected]. edu). G. J. Yoon is at 501 St Paul St #1407, Baltimore, MD 21202 USA (e-mail: [email protected]). R. D. Brinton is with the Neuroscience Program, John Stauffer Pharmaceu- tical Sciences Center 504, University of Southern California, Los Angeles, CA 90089-9121 USA (e-mail: [email protected]). T. W. Berger is with the Department of Biomedical Engineering, and the Center for Neural Engineering, Neuroscience Program, University of Southern California, Los Angeles, CA 90089-1451 USA (e-mail: [email protected]). This paper contains supplementary multimedia material available at http:// ieeexplore.ieee.org, provided by the author. This material consists of six videos. Each file is a time-lapse photomicrograph sequence of dissociated hippocampal neurons growing on MEAs coated with one of six biochemical substrate combi- nations (PDL, PDL BM, PEI, PEI BM, PO, PO BM). Pictures were taken at 2, 4, 6, 11, 14, and 17 days in vitro and joined into time-lapse movies. Process fasciculation and cell clustering is most readily observable on PDL BM and PO BM, while PEI holds somas fixed and promotes extensive fine branching. This material is 31.3 MB in size. Digital Object Identifier 10.1109/TBME.2006.889195 I. INTRODUCTION C ONTROL over the interface between neurons and Multi- Electrode Arrays (MEAs) is crucial for the development of viable electrode-based neural prostheses [1], [2], cell-based biosensors [3], and the creation of patterned neuronal circuits for the study of network dynamics [4], [5]. For these purposes, a diverse set of MEAs has been developed for recording the ac- tivity of neuronal populations, including microwire arrays [6], [7], silicon- or ceramic-based implantable arrays [8]–[11], and planar MEAs [12]–[18]. These neuron-silicon interface devices have to overcome several mechanical challenges from material disintegration in the saline oxidative environment, to biocom- patibility and tissue toxicity, to cellular attachment and encap- sulation. Various surface modifications are being investigated to reduce reactive gliosis and increase neuronal interaction with implanted devices [19]–[21]. Most of these approaches involve selective patterning of biologically active molecules on surfaces to spatially confine cellular attachment [4], [22], [23]. Physio- logical and morphological effects of substrates have been re- ported [24]–[34], as well as an increase in recorded activity by neuronal patterning [16]. This report investigates the rela- tionship between substrate induced morphological properties of neuronal networks and their spontaneous electrophysiological activity. Dissociated neurons cultured on planar MEAs (pMEAs) provide an ideal system for the investigation of the neuron-elec- trode interface. First, pMEAs can stimulate and record elec- trophysiological activity of many neurons simultaneously, while still enabling microscopic morphological observations and biochemical studies [12], [23], [35]. Second, dissociated neuronal cultures are simplified neural tissue preparations that can be grown in a highly controlled environment and still form synapses and develop into interconnected networks retaining physiological activity and plasticity [38]–[41]. Neurons must attach to, survive on, and form tight contacts with electrodes in order to stimulate and record their activity. At the neuron-elec- trode interface, it is the properties of the substrate covering the pMEAs that determine the degree of neural attachment, motility, growth, and survival [22], [32]. This paper, for the first time, describes the relationship between substrate coatings, network morphology, and elec- trophysiological activity on pMEAs. To this end, dissociated hippocampal neurons are cultured on the surface of pMEAs coated with three biochemical substrates: Poly-D-Lysine (PDL), PolyOrnithine (PO), and PolyEthylenImine (PEI) (Fig. 1). PDL 0018-9294/$25.00 © 2007 IEEE

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007 1309
Neuronal Network Morphology andElectrophysiologyof Hippocampal Neurons
Cultured on Surface-Treated Multielectrode ArraysWalid V. Soussou*, Geoffrey J. Yoon, Roberta Diaz Brinton, and Theodore W. Berger, Senior Member, IEEE
Abstract—Toward the development of biocompatible surfacesfor implantable electrode arrays and the creation of patternedneuronal networks, the impact of select biochemical substrates[poly-D-lysine (PDL), polyornithine (PO), polyethylenimine (PEI),and a basement membrane extract (BM)] on network morphologyand spontaneous electrophysiological activity of dissociatedhippocampal neurons was investigated. Cultured in serum-freeNeurobasal medium at 100 000 cells cm2, neurons attached toeach substrate. PDL, PO, and PEI induced little or no neuronalclustering and process fasciculation, whereas the addition ofBM promoted these features. The ratios of somas to processes,and axons to dendrites, as determined by immunohistochemicalstaining and image analysis were comparable across all substrates.Spontaneous firing was recorded using planar multielectrode ar-rays (MEAs) at the third week in vitro for the two most divergentmorphologies according to Euclidian cluster analysis, namelythose induced by PO+ BM and PEI. Mean spike amplitude, meanfiring rate, median interspike interval (ISI), mean burst rate, andcorrelation index were analyzed and compared to morphologicalfeatures. Synchronized bursting was highly correlated with neu-ronal clustering and process fasciculation. Spike amplitude wasnegatively correlated with thin branching which was most evidentin neurons grown on PEI. These data indicate that factors, whichinfluence adherence of neurons to surfaces, can profoundly impactboth neuronal network morphology and electrophysiologicalactivity in vitro.
Index Terms—Basement membrane, multielectrode array,neuronal network morphology, spontaneous electrophysiologicalactivity.
Manuscript received December 16, 2005; revised November 1, 2006. Asteriskindicates corresponding author.
*W. V. Soussou is with the Neuroscience Program, University of SouthernCalifornia, Los Angeles, CA 90089-2520 USA (e-mail: [email protected]).
G. J. Yoon is at 501 St Paul St #1407, Baltimore, MD 21202 USA (e-mail:[email protected]).
R. D. Brinton is with the Neuroscience Program, John Stauffer Pharmaceu-tical Sciences Center 504, University of Southern California, Los Angeles, CA90089-9121 USA (e-mail: [email protected]).
T. W. Berger is with the Department of Biomedical Engineering, and theCenter for Neural Engineering, Neuroscience Program, University of SouthernCalifornia, Los Angeles, CA 90089-1451 USA (e-mail: [email protected]).
This paper contains supplementary multimedia material available at http://ieeexplore.ieee.org, provided by the author. This material consists of six videos.Each file is a time-lapse photomicrograph sequence of dissociated hippocampalneurons growing on MEAs coated with one of six biochemical substrate combi-nations (PDL, PDL+BM, PEI, PEI+BM, PO, PO+BM). Pictures were taken at2, 4, 6, 11, 14, and 17 days in vitro and joined into time-lapse movies. Processfasciculation and cell clustering is most readily observable on PDL+BM andPO+BM, while PEI holds somas fixed and promotes extensive fine branching.This material is 31.3 MB in size.
Digital Object Identifier 10.1109/TBME.2006.889195
I. INTRODUCTION
CONTROL over the interface between neurons and Multi-Electrode Arrays (MEAs) is crucial for the development
of viable electrode-based neural prostheses [1], [2], cell-basedbiosensors [3], and the creation of patterned neuronal circuitsfor the study of network dynamics [4], [5]. For these purposes,a diverse set of MEAs has been developed for recording the ac-tivity of neuronal populations, including microwire arrays [6],[7], silicon- or ceramic-based implantable arrays [8]–[11], andplanar MEAs [12]–[18]. These neuron-silicon interface deviceshave to overcome several mechanical challenges from materialdisintegration in the saline oxidative environment, to biocom-patibility and tissue toxicity, to cellular attachment and encap-sulation. Various surface modifications are being investigatedto reduce reactive gliosis and increase neuronal interaction withimplanted devices [19]–[21]. Most of these approaches involveselective patterning of biologically active molecules on surfacesto spatially confine cellular attachment [4], [22], [23]. Physio-logical and morphological effects of substrates have been re-ported [24]–[34], as well as an increase in recorded activityby neuronal patterning [16]. This report investigates the rela-tionship between substrate induced morphological properties ofneuronal networks and their spontaneous electrophysiologicalactivity.
Dissociated neurons cultured on planar MEAs (pMEAs)provide an ideal system for the investigation of the neuron-elec-trode interface. First, pMEAs can stimulate and record elec-trophysiological activity of many neurons simultaneously,while still enabling microscopic morphological observationsand biochemical studies [12], [23], [35]. Second, dissociatedneuronal cultures are simplified neural tissue preparations thatcan be grown in a highly controlled environment and still formsynapses and develop into interconnected networks retainingphysiological activity and plasticity [38]–[41]. Neurons mustattach to, survive on, and form tight contacts with electrodes inorder to stimulate and record their activity. At the neuron-elec-trode interface, it is the properties of the substrate coveringthe pMEAs that determine the degree of neural attachment,motility, growth, and survival [22], [32].
This paper, for the first time, describes the relationshipbetween substrate coatings, network morphology, and elec-trophysiological activity on pMEAs. To this end, dissociatedhippocampal neurons are cultured on the surface of pMEAscoated with three biochemical substrates: Poly-D-Lysine (PDL),PolyOrnithine (PO), and PolyEthylenImine (PEI) (Fig. 1). PDL
0018-9294/$25.00 © 2007 IEEE

1310 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
Fig. 1. Chemical structure of polycationic substrates. (A) PDL, (B) PO, and (C)PEI. All three polymers have amine groups (NH )which are positively chargedat physiological pH, however, both PDL and PO have additional carbonyl groups(C = O) which contribute partial negative charges.
and PO are polycationic molecules first used to allow cells,which have an overall negative surface charge, to attach to sim-ilarly charged glass surfaces [24]–[26]. Their synthetic natureallows them not to interfere with cell physiology, while PDL’sD-enantiomeric configuration enables it to withstand prote-olytic cleavage. PO is similar to PDL in structure and charge,but has the amine group closer to the backbone. PEI, lackingthe slightly negatively charged carbonyl moieties of PDL andPO, is therefore a more positively charged polycationic organicmolecule on which neurons were reported to mature faster [29].In addition to these three synthetic molecules, a solubilizedbasement membrane (BM) extract was also used as a culturesubstrate. BM is a reconstituted extracellular matrix derivedfrom a mouse sarcoma, and consists of laminin (56%), collagenIV (31%), entactin (8%), heparan sulfate proteoglycan, matrixmetalloproteinases and various growth factors, including EGF,bFGF, NGF, PDGF, IGF-1, and TFG-B. The structure andactivity of reconstituted BM is reported to be similar to in vivoextracellular matrix [27], and its laminin is known to promoteaxonal growth [28], [30], [33]. These four substrates werecombined to yield a total of eight different substrate conditions(Fig. 2), onto which hippocampal neurons were cultured. Theeffects of substrates on neuron physiology was assessed firstmorphologically, by comparing cell attachment, and branchingpatterns, then immunohistochemically, by labeling axons anddendrites, and finally electrophysiologically, by analyzingspontaneous activity patterns of neurons cultured on thesedifferent substrate combinations. Euclidian cluster analysis wasconducted to group the substrates conditions by similarity ofmorphological features. These network morphological featureswere then correlated with the electrophysiological activityparameters. The observed relations between surface modifica-tions and network morphological and physiological activity are
useful for controlling patterned neuronal networks on MEAsand may hold in vivo implications for the design of neuralimplant surface treatments.
II. MATERIALS AND METHODS
A. Substrate Preparation
Glass coverslips and pMEAs were cleaned with a milddetergent (MEA cleaning solution, ALA scientific, Westbury,NY) thoroughly rinsed with distilled water, and sterilized with70% ethanol and UV light overnight. The following substrateswere dissolved in 1 mM sodium borate buffer pH 8.5: 100
PDL, 100 PEI, 100 PO. The coverslipsor MEAs were coated with 0.5 ml of these substrates per .After an overnight incubation, surfaces were rinsed with boratebuffer several times and either coated with 250 BM(Matrigel, Becton Dickinson, Franklin Lakes, NJ), or kept inbuffer overnight. Surfaces were rinsed again after this secondcoating and stored in buffer or culture medium until shortlybefore culture, at which time they were dried until cell seeding.Eight surface conditions were thus created (Fig. 2).
B. Cell Culture
Neurons were cultured according to a slightly modified pro-tocol from the one described in [41]. Pregnant Sprague-Dawleyrats were sacrificed by CO2 inhalation 18 days after plugging.Following decapitation, the embryonic brains were removedfrom the skull and placed in cold calcium and magnesium-freeHank’s salt solution (HSS) buffered with HEPES pH 7.3.Hippocampi then were dissected under a stereo microscopeand kept in cold HSS. Meningeal membranes were removedduring three HSS rinses, and then the hippocampi were me-chanically dissociated by repeated pipetting through a seriesof fire-polished Pasteur pipettes with decreasing tip diam-eters. Dissociated cells were then passed through a 40cell strainer to remove large clusters. The cells were mixedin Neurobasal medium (NBM, InVitrogen, Carlsbad, CA)supplemented with 0.5 mM glutamine, B-27 supplement (In-Vitrogen), Penicillin-Streptavidin, and 25 glutamate. Cellswere then plated at a density of oncoated coverslips or MEAs. Culture medium was replaced bi-weekly with NBM without additional glutamate. Cultures werekept in a humidified incubator at 37 in 10% , whichyielded a pH of 7.18 and no noticeable difference from culturesmaintained at 5% and hence pH 7.28. Cells survived for3–4 weeks under these conditions. Cell survival was monitoredby conducting LDH assays (Boehringer Mannheim, Germany)to obtain a measure of cell death.
C. Image Analysis
Cell cultures were taken out of the incubator periodically, andphotographed through a 10x phase contrast objective (DMIRBmicroscope, Leica, Germany; SPOT RT camera, Diagnostic In-struments, MI) before feeding or recording. Pictures were re-named to blind the conditions and visually scored according tothe following morphological features on scales from 1 to 5, withhalf point increments.
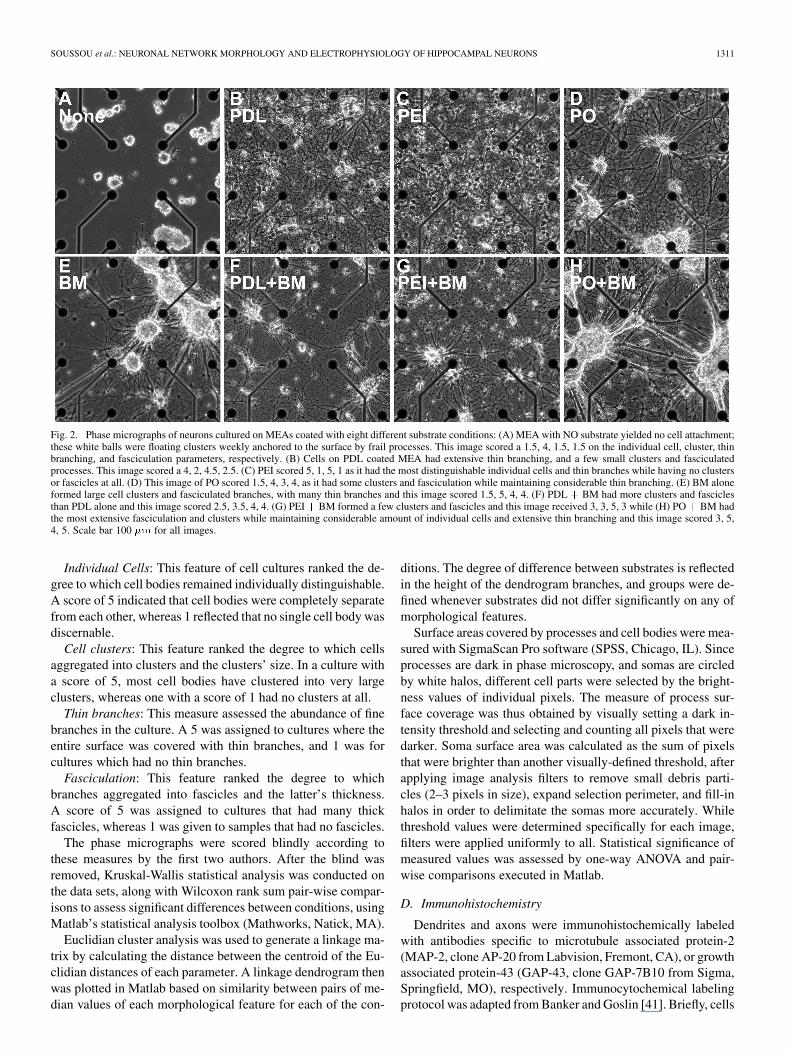
SOUSSOU et al.: NEURONAL NETWORK MORPHOLOGY AND ELECTROPHYSIOLOGY OF HIPPOCAMPAL NEURONS 1311
Fig. 2. Phase micrographs of neurons cultured on MEAs coated with eight different substrate conditions: (A) MEA with NO substrate yielded no cell attachment;these white balls were floating clusters weekly anchored to the surface by frail processes. This image scored a 1.5, 4, 1.5, 1.5 on the individual cell, cluster, thinbranching, and fasciculation parameters, respectively. (B) Cells on PDL coated MEA had extensive thin branching, and a few small clusters and fasciculatedprocesses. This image scored a 4, 2, 4.5, 2.5. (C) PEI scored 5, 1, 5, 1 as it had the most distinguishable individual cells and thin branches while having no clustersor fascicles at all. (D) This image of PO scored 1.5, 4, 3, 4, as it had some clusters and fasciculation while maintaining considerable thin branching. (E) BM aloneformed large cell clusters and fasciculated branches, with many thin branches and this image scored 1.5, 5, 4, 4. (F) PDL + BM had more clusters and fasciclesthan PDL alone and this image scored 2.5, 3.5, 4, 4. (G) PEI+ BM formed a few clusters and fascicles and this image received 3, 3, 5, 3 while (H) PO+ BM hadthe most extensive fasciculation and clusters while maintaining considerable amount of individual cells and extensive thin branching and this image scored 3, 5,4, 5. Scale bar 100 �m for all images.
Individual Cells: This feature of cell cultures ranked the de-gree to which cell bodies remained individually distinguishable.A score of 5 indicated that cell bodies were completely separatefrom each other, whereas 1 reflected that no single cell body wasdiscernable.
Cell clusters: This feature ranked the degree to which cellsaggregated into clusters and the clusters’ size. In a culture witha score of 5, most cell bodies have clustered into very largeclusters, whereas one with a score of 1 had no clusters at all.
Thin branches: This measure assessed the abundance of finebranches in the culture. A 5 was assigned to cultures where theentire surface was covered with thin branches, and 1 was forcultures which had no thin branches.
Fasciculation: This feature ranked the degree to whichbranches aggregated into fascicles and the latter’s thickness.A score of 5 was assigned to cultures that had many thickfascicles, whereas 1 was given to samples that had no fascicles.
The phase micrographs were scored blindly according tothese measures by the first two authors. After the blind wasremoved, Kruskal-Wallis statistical analysis was conducted onthe data sets, along with Wilcoxon rank sum pair-wise compar-isons to assess significant differences between conditions, usingMatlab’s statistical analysis toolbox (Mathworks, Natick, MA).
Euclidian cluster analysis was used to generate a linkage ma-trix by calculating the distance between the centroid of the Eu-clidian distances of each parameter. A linkage dendrogram thenwas plotted in Matlab based on similarity between pairs of me-dian values of each morphological feature for each of the con-
ditions. The degree of difference between substrates is reflectedin the height of the dendrogram branches, and groups were de-fined whenever substrates did not differ significantly on any ofmorphological features.
Surface areas covered by processes and cell bodies were mea-sured with SigmaScan Pro software (SPSS, Chicago, IL). Sinceprocesses are dark in phase microscopy, and somas are circledby white halos, different cell parts were selected by the bright-ness values of individual pixels. The measure of process sur-face coverage was thus obtained by visually setting a dark in-tensity threshold and selecting and counting all pixels that weredarker. Soma surface area was calculated as the sum of pixelsthat were brighter than another visually-defined threshold, afterapplying image analysis filters to remove small debris parti-cles (2–3 pixels in size), expand selection perimeter, and fill-inhalos in order to delimitate the somas more accurately. Whilethreshold values were determined specifically for each image,filters were applied uniformly to all. Statistical significance ofmeasured values was assessed by one-way ANOVA and pair-wise comparisons executed in Matlab.
D. Immunohistochemistry
Dendrites and axons were immunohistochemically labeledwith antibodies specific to microtubule associated protein-2(MAP-2, clone AP-20 from Labvision, Fremont, CA), or growthassociated protein-43 (GAP-43, clone GAP-7B10 from Sigma,Springfield, MO), respectively. Immunocytochemical labelingprotocol was adapted from Banker and Goslin [41]. Briefly, cells

1312 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
were fixed with 4% paraformaldehyde for 10–15 min, and per-meabilized with 0.25% Triton-X for 5 min. Nonspecific bindingwas blocked with 10% Bovine Serum Albumin (BSA, RocheApplied Science, Indianapolis, IN), in PBS for 30 min. Fixedcells were then incubated with primary antibodies diluted in a 3%BSA in PBS solution for 2 h at 37 . After rinsing, secondaryantibodies (FITC labeled anti-IgG1, and Texas Red labeledanti-IgG2a from Rockland Immunochemicals, Gilbertsville,PA) were applied to the cells for 1 h in the dark at 37 .Following another rinsing, stained cells were finally mountedwith a polyvinyl alcohol and glycerol mountant containing1% DABCO (Sigma). Fluorescent photomicrographs weretaken with a 40x objective on a DMIRB inverted microscopeequipped with a Spot RT camera. This labeling protocol yieldedgreen fluorescent dendrites and red axons.
Surface coverage of fluorescent dendrites and axons wasquantified using SigmaScan Pro image analysis software. Foreach image, color channels were first separated into red andgreen images. Then, pixels brighter than a visually-set back-ground threshold were counted as dendrites or axons dependingon their color channel. Lastly, one-way ANOVA was run inMatlab on measured surface area values to determine statisticalsignificance.
E. Electrophysiological Analysis
MEAs and their MEA-60 recording system, which includespreamplifiers, amplifiers, A/D converters, and software torecord from 60 electrodes simultaneously, are commerciallyavailable from MultiChannelSystems (Reutlingen, Germany).MEAs had 30 electrode tip diameter and 200 interelec-trode spacing. Two weeks after cell seeding, MEAs were takenout of the incubator, placed in the preamplifier box, and pho-tographed. Arrays were covered with a small cap traversed bya perforated tube that provided a slow flow of humidified 10%
and 90% gas mixture. The chamber was maintainedat 37 . The first 5 min of recorded spontaneous activity wereanalyzed for each pMEA cell culture.
Extracellular spike trains were detected by conservatively set-ting a threshold value just above a visually determined back-ground noise for each recording channel. Whenever an actionpotential crossed that threshold, a timestamp was recorded alongwith a 3–ms spike waveform sampled at 20 or 25 kHz (1 ms pre-trigger and 2 ms posttrigger). Recorded waveforms were usedfor spike sorting using OfflineSorter (PlexonInc, TX). Wave-forms were also used to discard spikes that were less than fourtimes the signal-to-noise ratio. This re-thresholding was accom-plished in a custom Matlab script. Noise was calculated as thesmaller of the root-mean-square value of all 1 ms pretrigger orall 1 ms posttrigger segments for a unit. The contrasted signalwas the peak-to-peak spike amplitude. This re-threshholdingscript also allowed manual assignment of overlapping spikes tomultiple units. The last preprocessing step was elimination ofall units that had less than 10 spikes over the entire recordedperiod. This filtering reduced the effect of the large number ofsilent channels on parameters of interest. Sorted spike times-tamps were then imported into NeuroExplorer (Nex Technolo-gies, MA) for analysis. This program calculated spontaneous
firing and bursting parameters such as mean firing rates, the me-dian of interspike interval (ISI) distributions, mean burst rates(where bursts are calculated according to a surprise method de-scribed in [42]), and unit cross-correlations. A normalized indexof this last measure was used as a measure of firing synchrony.Correlation indices were calculated in Matlab as the mean of thecross-correlation over a window minus the mean ofthe entire cross-correlation multiplied by the number of bins anddivided by the number of reference events. This normalizationproduces a nonsymmetrical correlation matrix that is howevermore appropriate for large spike trains. Distributions of thesefive electrophysiological parameters between cultures on PEIand PO BM were compared with a Kolmogorov-Smirnov testin Matlab. Finally, the relation between morphological featuresand electrophysiological measures were statistically comparedwith Spearman rank correlation coefficients that were calculatedalso using Matlab’s statistical toolbox.
III. RESULTS
Fig. 2 shows photomicrographs of hippocampal neuroncultures seeded onto different substrates after 17 daysin culture. These cultures were plated with a density of
. At this density, approximately 20% ofthe surface area should be covered with somas. This densitywas selected to strike a balance between increasing the likeli-hood of neurons settling on top of electrodes, without creatinga confluent monolayer of cells, thus enabling distinction of cellbodies and processes. In addition, the lifespan of dissociatedcultures is decreased at lower plating densities [41].
Neurons did not attach at all, or very poorly at best, whenglass or MEA surfaces were not coated with any substrate [Fig.2(A)]. Any remaining cell bodies clustered and floated, whilethe few processes that extended out of them were frail, thinbut fasciculated and elongated. Hippocampal neurons did how-ever attach to coated surfaces and develop networks varying inmorphological features. Whether cultured on glass coverslips
or MEAs , hippocampal neurons attached and de-veloped morphologies that were reproducibly characteristic ofthe coating substrate on which they were cultured.
Networks gradually developed and accentuated their morpho-logical characteristics over weeks in culture. Branching patternsin the first week were not clearly distinguishable between condi-tions, while cells bodies had already started to form clusters onmost substrates except PEI. To track neuron morphological dif-ferentiation and motility over time, images of neurons culturedon MEAs were captured across a span of 17 days in vitro (DIV).Aligning and linking these pictures into time-lapse movies re-vealed that cells on PEI did not move once settled on the surface,while those on other conditions were more motile, and movedslowly along fasciculating processes to form clusters. Develop-ment of neuronal cultures on different substrates can be tracedin supplementary videos.
By 14 DIV, neural networks had developed characteristicmorphological features: cells formed small clusters and someprocesses fasciculated on PDL and PO [Fig. 2(B) and (D)],whereas PEI kept cell bodies distinct and prevented clusteringand process fasciculation [Fig. 2(C)]. Addition of a BM coat to
SOUSSOU et al.: NEURONAL NETWORK MORPHOLOGY AND ELECTROPHYSIOLOGY OF HIPPOCAMPAL NEURONS 1313
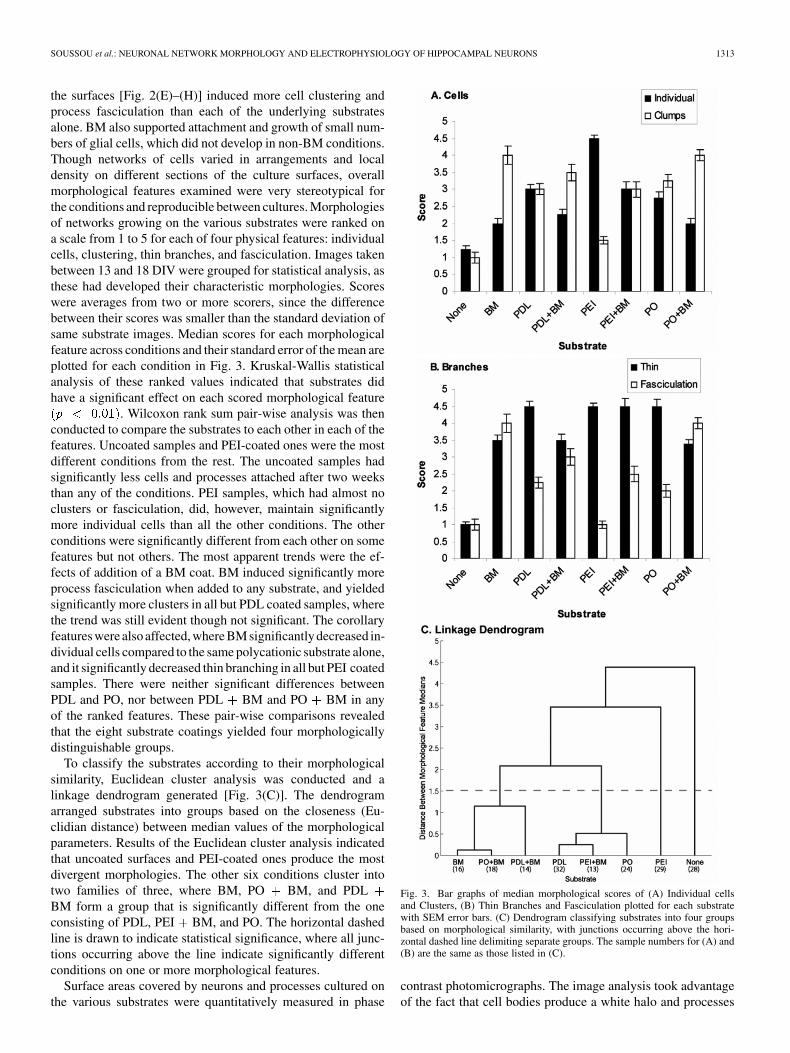
the surfaces [Fig. 2(E)–(H)] induced more cell clustering andprocess fasciculation than each of the underlying substratesalone. BM also supported attachment and growth of small num-bers of glial cells, which did not develop in non-BM conditions.Though networks of cells varied in arrangements and localdensity on different sections of the culture surfaces, overallmorphological features examined were very stereotypical forthe conditions and reproducible between cultures. Morphologiesof networks growing on the various substrates were ranked ona scale from 1 to 5 for each of four physical features: individualcells, clustering, thin branches, and fasciculation. Images takenbetween 13 and 18 DIV were grouped for statistical analysis, asthese had developed their characteristic morphologies. Scoreswere averages from two or more scorers, since the differencebetween their scores was smaller than the standard deviation ofsame substrate images. Median scores for each morphologicalfeature across conditions and their standard error of the mean areplotted for each condition in Fig. 3. Kruskal-Wallis statisticalanalysis of these ranked values indicated that substrates didhave a significant effect on each scored morphological feature
. Wilcoxon rank sum pair-wise analysis was thenconducted to compare the substrates to each other in each of thefeatures. Uncoated samples and PEI-coated ones were the mostdifferent conditions from the rest. The uncoated samples hadsignificantly less cells and processes attached after two weeksthan any of the conditions. PEI samples, which had almost noclusters or fasciculation, did, however, maintain significantlymore individual cells than all the other conditions. The otherconditions were significantly different from each other on somefeatures but not others. The most apparent trends were the ef-fects of addition of a BM coat. BM induced significantly moreprocess fasciculation when added to any substrate, and yieldedsignificantly more clusters in all but PDL coated samples, wherethe trend was still evident though not significant. The corollaryfeatures were also affected, where BM significantly decreased in-dividual cells compared to the same polycationic substrate alone,and it significantly decreased thin branching in all but PEI coatedsamples. There were neither significant differences betweenPDL and PO, nor between PDL BM and PO BM in anyof the ranked features. These pair-wise comparisons revealedthat the eight substrate coatings yielded four morphologicallydistinguishable groups.
To classify the substrates according to their morphologicalsimilarity, Euclidean cluster analysis was conducted and alinkage dendrogram generated [Fig. 3(C)]. The dendrogramarranged substrates into groups based on the closeness (Eu-clidian distance) between median values of the morphologicalparameters. Results of the Euclidean cluster analysis indicatedthat uncoated surfaces and PEI-coated ones produce the mostdivergent morphologies. The other six conditions cluster intotwo families of three, where BM, PO BM, and PDLBM form a group that is significantly different from the oneconsisting of PDL, PEI BM, and PO. The horizontal dashedline is drawn to indicate statistical significance, where all junc-tions occurring above the line indicate significantly differentconditions on one or more morphological features.
Surface areas covered by neurons and processes cultured onthe various substrates were quantitatively measured in phase
Fig. 3. Bar graphs of median morphological scores of (A) Individual cellsand Clusters, (B) Thin Branches and Fasciculation plotted for each substratewith SEM error bars. (C) Dendrogram classifying substrates into four groupsbased on morphological similarity, with junctions occurring above the hori-zontal dashed line delimiting separate groups. The sample numbers for (A) and(B) are the same as those listed in (C).
contrast photomicrographs. The image analysis took advantageof the fact that cell bodies produce a white halo and processes
1314 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
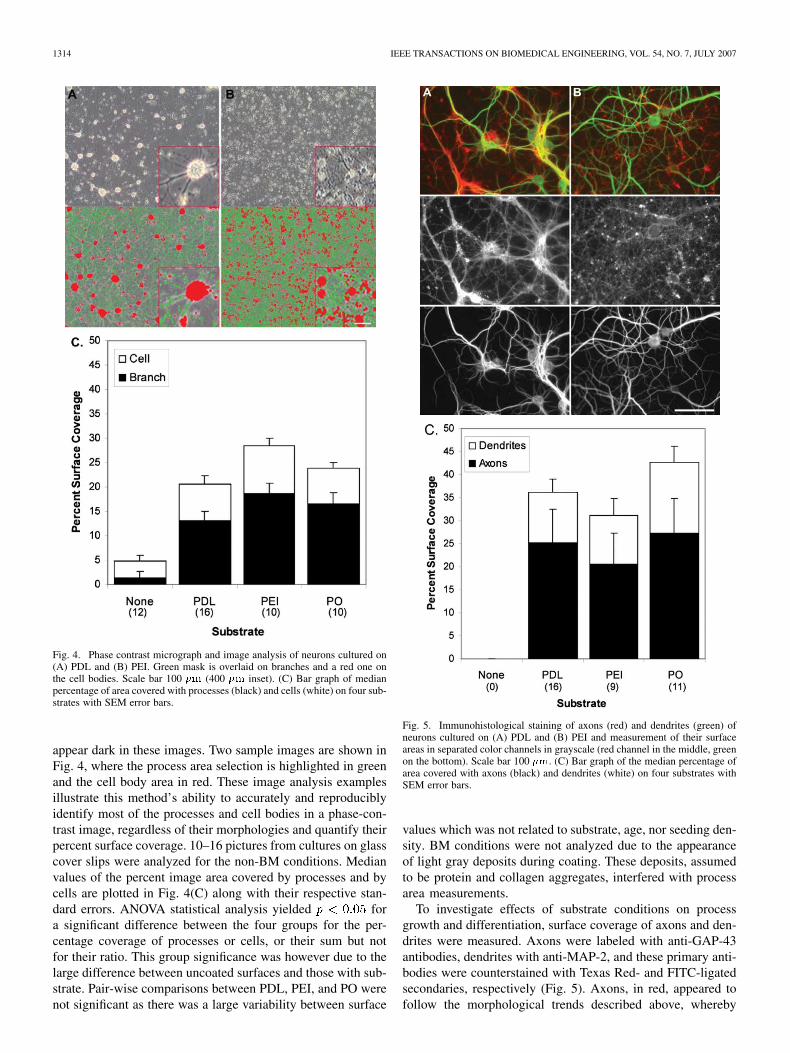
Fig. 4. Phase contrast micrograph and image analysis of neurons cultured on(A) PDL and (B) PEI. Green mask is overlaid on branches and a red one onthe cell bodies. Scale bar 100 �m (400 �m inset). (C) Bar graph of medianpercentage of area covered with processes (black) and cells (white) on four sub-strates with SEM error bars.
appear dark in these images. Two sample images are shown inFig. 4, where the process area selection is highlighted in greenand the cell body area in red. These image analysis examplesillustrate this method’s ability to accurately and reproduciblyidentify most of the processes and cell bodies in a phase-con-trast image, regardless of their morphologies and quantify theirpercent surface coverage. 10–16 pictures from cultures on glasscover slips were analyzed for the non-BM conditions. Medianvalues of the percent image area covered by processes and bycells are plotted in Fig. 4(C) along with their respective stan-dard errors. ANOVA statistical analysis yielded fora significant difference between the four groups for the per-centage coverage of processes or cells, or their sum but notfor their ratio. This group significance was however due to thelarge difference between uncoated surfaces and those with sub-strate. Pair-wise comparisons between PDL, PEI, and PO werenot significant as there was a large variability between surface
Fig. 5. Immunohistological staining of axons (red) and dendrites (green) ofneurons cultured on (A) PDL and (B) PEI and measurement of their surfaceareas in separated color channels in grayscale (red channel in the middle, greenon the bottom). Scale bar 100 �m. (C) Bar graph of the median percentage ofarea covered with axons (black) and dendrites (white) on four substrates withSEM error bars.
values which was not related to substrate, age, nor seeding den-sity. BM conditions were not analyzed due to the appearanceof light gray deposits during coating. These deposits, assumedto be protein and collagen aggregates, interfered with processarea measurements.
To investigate effects of substrate conditions on processgrowth and differentiation, surface coverage of axons and den-drites were measured. Axons were labeled with anti-GAP-43antibodies, dendrites with anti-MAP-2, and these primary anti-bodies were counterstained with Texas Red- and FITC-ligatedsecondaries, respectively (Fig. 5). Axons, in red, appeared tofollow the morphological trends described above, whereby

SOUSSOU et al.: NEURONAL NETWORK MORPHOLOGY AND ELECTROPHYSIOLOGY OF HIPPOCAMPAL NEURONS 1315
they were fasciculated on PO BM, very finely branched onPEI, and with intermediate fasciculation on PDL. Dendritesappeared less affected by the substrate in terms of thickness andfasciculation. The median area coverage of dendrites and axonson PDL, PEI, and PO are plotted in Fig. 5(E) with the respectivestandard errors. ANOVA statistical analysis of axonal or den-dritic areas, and their sum or ratios yielded , indicatingno significant difference between these groups, probably due toa large variability between surface values which was not relatedto substrate, age, nor seeding density. However, in all cases,axons occupied more surface than dendrites.
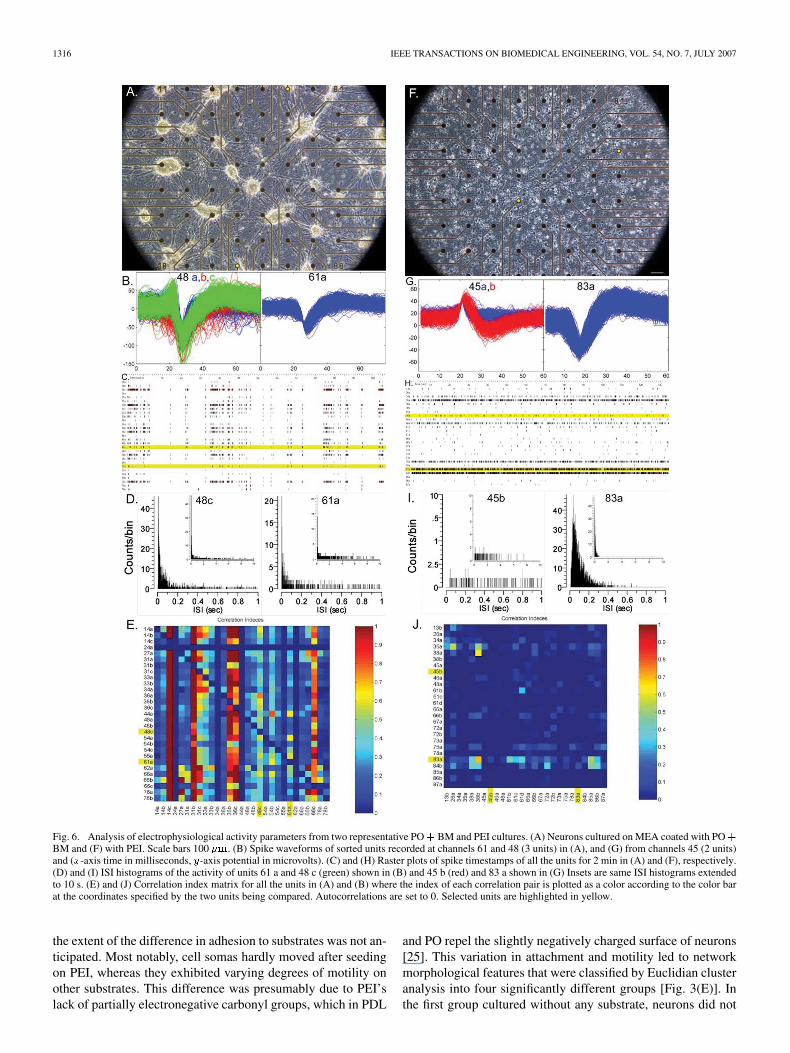
To determine the impact of morphology on electrophysi-ology, spontaneous activity of hippocampal neurons growingon substrate-coated pMEAs was recorded during the thirdweek in culture. The two conditions that produced the mostdifferent morphological features, namely PEI and PO BM,were compared. Inspection of a few overall activity measuresrevealed consistency with some of the above morphologicalobservations (Table I). Of all electrodes recorded, 21% exhib-ited spike activity on either substrate, which is consistent witha 100 000 plating density, and in agreement with thelack of difference in their measured cell surface coverage. Afternoise filtering and spike sorting, the PO BM condition hadmore units per channel, consistent with its large cell clusterson top of multiunit electrodes, whereas cell bodies were moredispersed on PEI [Fig. 6(A) and (F)]. Fig. 6 illustrates theseobserved differences with two units from each condition.Fig. 6(B) and (G) contrasts spike waveforms and amplitudes.PO BM had more units per channel and their amplitude wasoften larger. The overall mean firing rate of neurons growingon PEI was greater than that of neurons cultured on PO BM(see Fig. 7). Furthermore, the firing patterns of neurons in thesetwo conditions were qualitatively very different. Rastergrams[Fig. 6(C) and (H)], revealed the key difference in activity:neurons cultured on PO BM frequently fired in synchronousbursts, whereas those grown on PEI tended to fire individuallyin sporadic or continuous asynchronous patterns. Consistentwith the bursting observed in the rastergrams, ISI histogramsof the firing patterns [Fig. 6(D) and (I)] displayed bimodalcurves for PO BM consisting of a narrow distribution ofshort ISIs, accounted for by high intraburst firing rates, fol-lowed by a smaller and broadly spread peak of longer ISIscorresponding to interburst rates. In contrast, PEI’s histogramswere monophasic and represented sporadic or continuousfiring with spreads intermediate to PO BM’s dual peaks.Lastly, synchrony between active units on a particular MEAwas revealed with color plot of correlation indices [Fig. 6(E)and (J)]. A high index value indicates that two channel on anarray frequently fire within 200 ms of each other, whereas alow index reflects asynchronous firing. Negative correlationindices, which reflected the likelihood of units to fire outsidea 200 ms interval from the other (i.e., asynchronously), werealways small in magnitude indicating that there wereno delayed correlations. The color graph plots high correlationindices in red, and low valued ones in blue at the intersectionbetween their respective units on the - and -axes. The activityof networks exhibited more correlated firing on PO BM thanon PEI.
TABLE IELECTROPHYSIOLOGY SUMMARY
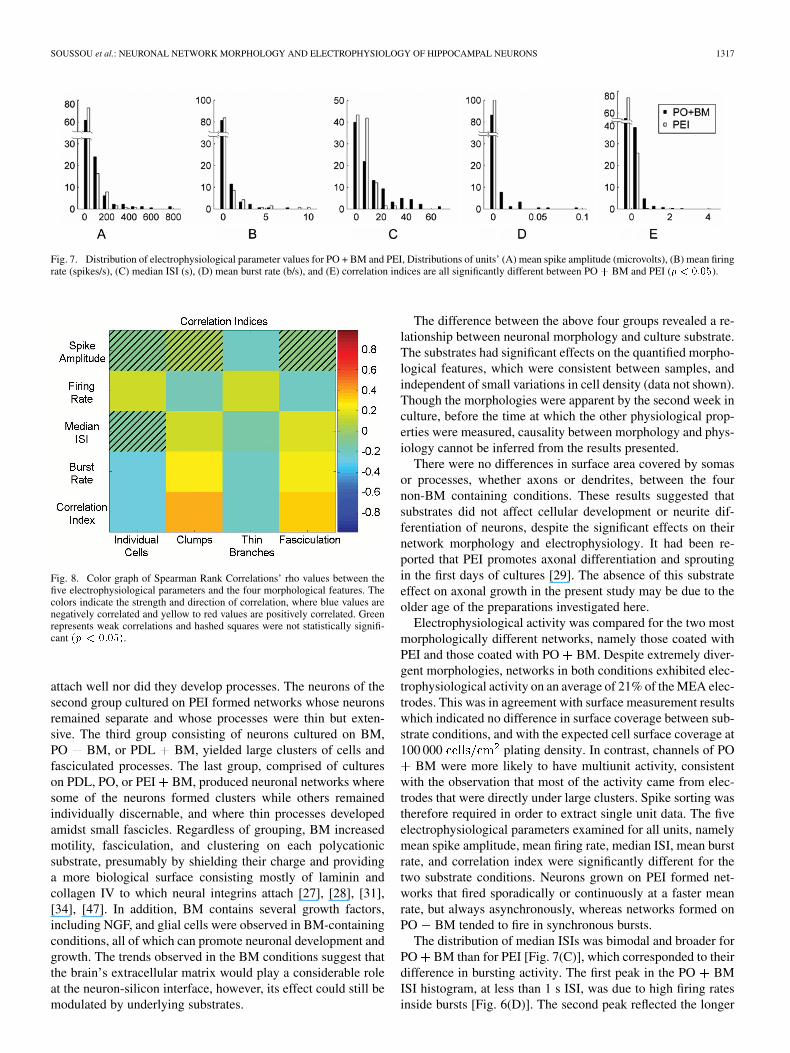
These electrophysiological parameters were not normallydistributed, and thus for stastical analysis, their distributionswere plotted and analyzed with Kolmogorov-Smirnov tests(Fig. 7). The distributions were significantly different for all ex-amined parameters. The mean spike amplitude distribution wasshifted towards larger amplitudes for the PO BM conditionFig. 7(A), while the mean firing rate distribution was broaderwith PEI as substrate Fig. 7(B). The median ISI distributionof the neurons was bimodal on PO BM Fig. 7(C) whichreflected their bursting activity, while neurons on PEI exhibitedno bursting at all Fig. 7(D). Finally, the correlation index distri-bution was shifted towards more correlation between neuronscultured on PO BM than on PEI Fig. 7(E).
Finally, the question of whether morphological features arerelated to electrophysiological parameters was addressed witha Spearman rank correlation matrix. Fig. 8 is a color plot ofSpearman’s correlation index (rho), where positive correlationsare red and negative correlations are blue, and statistically in-significant correlations are hashed. Results of this analysis in-dicated that neuronal networks with greater clustering and fas-ciculation had more synchronous firing and faster bursting, al-though its mean firing rate dropped. Neural networks character-ized by abundant individual cells and thin branching were nega-tively correlated with synchronous activity and bursting. Thesedata indicated that individual cells were less likely to burst insynchrony, despite having higher mean firing rates. In addition,thin branches were also negatively correlated with smaller spikeamplitudes.
IV. DISCUSSION
There is an increased use of microfabricated devices withbrain tissue, in applications ranging from planar MEAs asbiosensors [3], [43], to implantable MEAs for Deep-BrainStimulation [44] or neuroprosthetic applications [2]. Issues ofcell attachment and biocompatibility are critical for the func-tioning of these devices [45]. Several biotic coatings have beeninvestigated to control neuronal attachment [22], [24], [30],[32], [46] or glial scarring [21], however, little is known aboutthe effects of these various surface treatments on neuronalnetwork morphology and electrophysiology. Here, dissociatedhippocampal neurons cultured on pMEAs were used to inves-tigate the relationship between various polycationic moleculesand BM coatings on neuronal attachment and physiology.
The various substrate coatings yielded neuronal networkswith different morphological features. Neurons did not attach touncoated glass surfaces, which are electronegatively charged,as neurons also have a weakly negative overall surface chargeand are therefore repelled [41]. As expected, the neurons didattach to surfaces coated with polycationic molecules; however,
1316 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
Fig. 6. Analysis of electrophysiological activity parameters from two representative PO+ BM and PEI cultures. (A) Neurons cultured on MEA coated with PO+BM and (F) with PEI. Scale bars 100 �m. (B) Spike waveforms of sorted units recorded at channels 61 and 48 (3 units) in (A), and (G) from channels 45 (2 units)and (x-axis time in milliseconds, y-axis potential in microvolts). (C) and (H) Raster plots of spike timestamps of all the units for 2 min in (A) and (F), respectively.(D) and (I) ISI histograms of the activity of units 61 a and 48 c (green) shown in (B) and 45 b (red) and 83 a shown in (G) Insets are same ISI histograms extendedto 10 s. (E) and (J) Correlation index matrix for all the units in (A) and (B) where the index of each correlation pair is plotted as a color according to the color barat the coordinates specified by the two units being compared. Autocorrelations are set to 0. Selected units are highlighted in yellow.
the extent of the difference in adhesion to substrates was not an-ticipated. Most notably, cell somas hardly moved after seedingon PEI, whereas they exhibited varying degrees of motility onother substrates. This difference was presumably due to PEI’slack of partially electronegative carbonyl groups, which in PDL
and PO repel the slightly negatively charged surface of neurons[25]. This variation in attachment and motility led to networkmorphological features that were classified by Euclidian clusteranalysis into four significantly different groups [Fig. 3(E)]. Inthe first group cultured without any substrate, neurons did not
SOUSSOU et al.: NEURONAL NETWORK MORPHOLOGY AND ELECTROPHYSIOLOGY OF HIPPOCAMPAL NEURONS 1317
Fig. 7. Distribution of electrophysiological parameter values for PO + BM and PEI, Distributions of units’ (A) mean spike amplitude (microvolts), (B) mean firingrate (spikes/s), (C) median ISI (s), (D) mean burst rate (b/s), and (E) correlation indices are all significantly different between PO+ BM and PEI (p < 0:05).
Fig. 8. Color graph of Spearman Rank Correlations’ rho values between thefive electrophysiological parameters and the four morphological features. Thecolors indicate the strength and direction of correlation, where blue values arenegatively correlated and yellow to red values are positively correlated. Greenrepresents weak correlations and hashed squares were not statistically signifi-cant (p < 0:05).
attach well nor did they develop processes. The neurons of thesecond group cultured on PEI formed networks whose neuronsremained separate and whose processes were thin but exten-sive. The third group consisting of neurons cultured on BM,PO BM, or PDL BM, yielded large clusters of cells andfasciculated processes. The last group, comprised of cultureson PDL, PO, or PEI BM, produced neuronal networks wheresome of the neurons formed clusters while others remainedindividually discernable, and where thin processes developedamidst small fascicles. Regardless of grouping, BM increasedmotility, fasciculation, and clustering on each polycationicsubstrate, presumably by shielding their charge and providinga more biological surface consisting mostly of laminin andcollagen IV to which neural integrins attach [27], [28], [31],[34], [47]. In addition, BM contains several growth factors,including NGF, and glial cells were observed in BM-containingconditions, all of which can promote neuronal development andgrowth. The trends observed in the BM conditions suggest thatthe brain’s extracellular matrix would play a considerable roleat the neuron-silicon interface, however, its effect could still bemodulated by underlying substrates.
The difference between the above four groups revealed a re-lationship between neuronal morphology and culture substrate.The substrates had significant effects on the quantified morpho-logical features, which were consistent between samples, andindependent of small variations in cell density (data not shown).Though the morphologies were apparent by the second week inculture, before the time at which the other physiological prop-erties were measured, causality between morphology and phys-iology cannot be inferred from the results presented.
There were no differences in surface area covered by somasor processes, whether axons or dendrites, between the fournon-BM containing conditions. These results suggested thatsubstrates did not affect cellular development or neurite dif-ferentiation of neurons, despite the significant effects on theirnetwork morphology and electrophysiology. It had been re-ported that PEI promotes axonal differentiation and sproutingin the first days of cultures [29]. The absence of this substrateeffect on axonal growth in the present study may be due to theolder age of the preparations investigated here.
Electrophysiological activity was compared for the two mostmorphologically different networks, namely those coated withPEI and those coated with PO BM. Despite extremely diver-gent morphologies, networks in both conditions exhibited elec-trophysiological activity on an average of 21% of the MEA elec-trodes. This was in agreement with surface measurement resultswhich indicated no difference in surface coverage between sub-strate conditions, and with the expected cell surface coverage at100 000 plating density. In contrast, channels of PO
BM were more likely to have multiunit activity, consistentwith the observation that most of the activity came from elec-trodes that were directly under large clusters. Spike sorting wastherefore required in order to extract single unit data. The fiveelectrophysiological parameters examined for all units, namelymean spike amplitude, mean firing rate, median ISI, mean burstrate, and correlation index were significantly different for thetwo substrate conditions. Neurons grown on PEI formed net-works that fired sporadically or continuously at a faster meanrate, but always asynchronously, whereas networks formed onPO BM tended to fire in synchronous bursts.
The distribution of median ISIs was bimodal and broader forPO BM than for PEI [Fig. 7(C)], which corresponded to theirdifference in bursting activity. The first peak in the PO BMISI histogram, at less than 1 s ISI, was due to high firing ratesinside bursts [Fig. 6(D)]. The second peak reflected the longer

1318 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
interburst intervals (mode 40s). The median ISI, therefore,was related to the mean bursting rate which was positivelyand significantly correlated with clusters and fasciculation, andnegatively correlated with thin branching, and individual cells.Similar correlations also were observed for correlation indices,which were measures of synchronous firing of units within a
window. Therefore, neuron clustering and fascicu-lation were correlated with networks that burst synchronously,whereas individual cells with extensive thin branching firedasynchronously.
This difference in bursting activity between the two groupsis in accord with a theoretical framework presented by Lathamet al.. for explaining bursting activity in isolated networks. [48],[49] According to their model, networks shift from steady firingrates to bursting when their fraction of endogenously active neu-rons drops below a certain threshold. This criterion could bemet by the morphological differences in the networks grownon the two conditions. PO BM had significantly less indi-vidual cells than PEI, these isolated neurons would be morelikely to fire independently at their endogenous rates than theneurons clumped on PO BM. Neurons in clumps can coupleephaptically and fire as one, yielding a network with fewer en-dogeneously firing units, which according to the model pro-duces bursting activity. In addition, branch fascicules increaseinterconnections between clumps and could hence promote syn-chrony across the network. Though the results are only cor-relative, this framework suggests that PO BM could shiftthe activity of cultured neuronal populations into synchronizedbursting by inducing cell clustering and process fasciculation.
The substrates also affected the size of the recorded signal, asunits on PO BM had significantly larger mean spike ampli-tudes than those on PEI, and the larger spikes were significantlynegatively correlated with thin branching. This might be due tocurrents carried by thinner processes being smaller, or to incom-plete electrode coupling on PEI substrates. In general, recordedextracellular spike amplitudes depend on the proximity of axonhillocks to recording electrodes and on the coupling betweensomata and electrode tips [50]. Large neuronal clusters wouldprovide a better surface seal with electrodes. Though there was apositive correlation between clusters and mean spike amplitude,the correlation was not significant perhaps due to the separationof active neurons in the cluster from the electrode surface byother nonactive cells.
There is considerable debate as to what constitutes “healthy”morphology and electrical activity in dissociated neuronal cul-tures. Some researchers favor cells that do not clump, branchesthat do not fasciculate, and activity that does not burst, and in-vestigate single neuron morphology or development of synapticactivity. [41], [51], [52] others point to the tendency of neu-rons to cluster in vivo, form fasciculated nerve bundles, and firein synchronous bursts at specific frequencies and have inves-tigated this bursting. [39], [49], [53] The major difference be-tween these two clans is cell density and recording age. Net-works above a certain cell density and two weeks of age tend toexhibit bursting activity not observed in younger or lower den-sity networks. The results of LDH assays performed on cultureson the various substrates did not indicate any increased mor-tality of neurons over the 3 week period investigated, except for
those on uncoated surfaces. However, there was a distinct differ-ence between the clumping and fasciculation observed on thoseuntreated condition and that on BM containing conditions. Theclumps on the uncoated surfaces contained mostly dead cells,and their branches were weakly attached to the surface, and mostof the cells floated away by the end of the first week. All coatedconditions are therefore assumed to be in similar health, albeitexpressing different morphologies and activity patterns.
In summary, while a variety of surface treatments havebeen investigated for attachment of neurons on silicon ormetal surfaces; little was known about differential effects ofthese substrates on neuronal network morphology and electro-physiological activity. Here, substrates were shown to have asignificant effect on neuronal network morphology. The elec-trophysiological activity of the networks was also altered bythe underlying substrates. In addition, morphological featureswere significantly correlated with electrophysiological param-eters. Understanding these correlations will enable control ofnetwork architectural types through the selection of underlyingsubstrates.
Refined control of neurite pathways and connections allowsthe creation of defined circuits for the study of the computationalabilities of biological neuronal networks [4], [33]. This controlcan be achieved by cell patterning, which enables the align-ment of neurons on planar electrode arrays, thereby simplifiyinglong-term electrophysiology, microscopic observation, and dataanalysis [54]. Patterned networks are designed by creating con-trasts between repulsive and attractive surfaces for neurons toattach and grow on. The results presented above extend this con-trol by providing a more diverse substrate toolset to affect neu-ronal morphology and electrophysiological properties. For ex-ample, patterning electrode tips with PEI and the surroundingsurfaces with PDL or PO BM would allow neurons to attachto MEAs whether planar or implantable, and somas would mi-grate on the surface until they reached PEI, where they wouldadhere tightly and stop moving. This strong coupling of severalsomas at the electrode would also increase recorded spike am-plitude. In addition, process fasciculation would promote syn-chronization of bursting activity which might allow electricallystimulated responses to propagate further into the network. Pat-terning of substrates on MEAs therefore could be used to en-hance both recorded and stimulated signals in vitro. Coatingneuroprosthetic implants with biological molecules and growthfactors has also been shown to enhance biocompatibility andrecorded signal quality [55]. The results presented here shouldbe explored in vivo to elucidate the implications of patterningsurfaces for brain-implantable devices.
V. CONCLUSION
This report describes for the first time the effects of surfacecoating materials on neuronal network morphology and their re-lationships to electrophysiological activity. Neurons cultured ondifferent substrates develop networks with distinct morpholo-gies and activity patterns. Networks with neural clusters andaxonal fascicles fire in synchronized bursts whereas those withmore individual neurons and thin processes fire sporadically orsteadily. These relations between morphological and electro-physiological properties and neuron-interface attachment can be

SOUSSOU et al.: NEURONAL NETWORK MORPHOLOGY AND ELECTROPHYSIOLOGY OF HIPPOCAMPAL NEURONS 1319
harnessed for the control of neuronal network activity on pat-terned MEAs. In the future, the causality between these corre-lations should be investigated, as well as their direct relationto electrical excitability and applicability to brain implantabledevices.
ACKNOWLEDGMENT
The authors would like thank Dr. S. Chen and Dr. H. Guo andall the work-study students (E. Cheung, J. Toledo, J. Mara, andI-m. Shiou) for their help with cell culture. Additional thanks toDr. M. Kim for conducting LDH assays and Dr. H. Jourdi forhelpful comments on the manuscript.
REFERENCES
[1] W. L. Rutten, J. P. Smit, T. A. Frieswijk, J. A. Bielen, A. L. Brouwer, J.R. Buitenweg, and C. Heida, “Neuro-electronic interfacing with multi-electrode arrays,” IEEE Eng. Med. Biol. Mag., vol. 18, no. 3, pp. 47–55,May-Jun. 1999.
[2] T. W. Berger, M. Baudry, R. D. Brinton, J. S. Liaw, V. Z. Marmarelis,A. Y. Park, B. J. Sheu, and A. R. J. Tanguay, “Brain-Implantablebiomimetic electronics as the next era in neural prosthetics,” ProcIEEE, vol. 89, pp. 993–1012, 2001.
[3] G. W. Gross, B. K. Rhoades, H. M. Azzazy, and M. C. Wu, “The useof neuronal networks on multielectrode arrays as biosensors,” Biosens.Bioelectron., vol. 10, pp. 553–567, 1995.
[4] D. A. Stenger, J. J. Hickman, K. E. Bateman, M. S. Ravenscroft, W.Ma, J. J. Pancrazio, K. Shaffer, A. E. Schaffner, D. H. Cribbs, and C.W. Cotman, “Microlithographic determination of axonal/dendritic po-larity in cultured hippocampal neurons,” J. Neurosci. Meth., vol. 82,pp. 167–173, 1998.
[5] M. P. Maher, J. Pine, J. Wright, and Y. C. Tai, “The neurochip: A newmultielectrode device for stimulating and recording from cultured neu-rons,” J. Neurosci. Meth., vol. 87, pp. 45–56, 1999.
[6] S. A. Deadwyler, T. Bunn, and R. E. Hampson, “Hippocampalensemble activity during spatial delayed-nonmatch-to-sample perfor-mance in rats,” J. Neurosci., vol. 16, pp. 354–372, 1996.
[7] J. C. Williams, R. L. Rennaker, and D. R. Kipke, “Long-term neuralrecording characteristics of wire microelectrode arrays implanted incerebral cortex,” Brain Res. Brain Res. Protoc., vol. 4, pp. 303–313,1999.
[8] J. F. Hetke, J. L. Lund, K. Najafi, K. D. Wise, and D. J. Anderson, “Sil-icon ribbon cables for chronically implantable microelectrode arrays,”IEEE Trans. Biomed. Eng., vol. 41, no. 4, pp. 314–321, Apr. 1994.
[9] C. T. Nordhausen, P. J. Rousche, and R. A. Normann, “Optimizingrecording capabilities of the utah intracortical electrode array,” BrainRes., vol. 637, pp. 27–36, 1994.
[10] A. Bragin, J. Hetke, C. L. Wilson, D. J. Anderson, J. Engel, Jr, and G.Buzsaki, “Multiple site silicon-based probes for chronic recordings infreely moving rats: Implantation, recording and histological verifica-tion,” J. Neurosci. Meth., vol. 98, pp. 77–82, 2000.
[11] J. J. Burmeister, K. Moxon, and G. A. Gerhardt, “Ceramic-basedmultisite microelectrodes for electrochemical recordings,” AnalyticalChem., vol. 72, pp. 187–192, 2000.
[12] J. Pine, “Recording action potentials from cultured neurons with extra-cellular microcircuit electrodes,” J. Neurosci. Meth., vol. 2, pp. 19–31,1980.
[13] G. W. Gross, A. N. Williams, and J. H. Lucas, “Recording of sponta-neous activity with photoetched microelectrode surfaces from mousespinal neurons in culture,” J. Neurosci. Meth., vol. 5, pp. 13–22, 1982.
[14] U. Egert, B. Schlosshauer, S. Fennrich, W. Nisch, M. Fejtl, T. Knott,T. Müller, and H. Hämmerle, “A novel organotypic long-term cultureof the rat hippocampus on substrate-integrated multielectrode arrays,”Brain Res. Brain Res. Protoc., vol. 2, pp. 229–242, 1998.
[15] H. Oka, K. Shimono, R. Ogawa, H. Sugihara, and M. Taketani, “A newplanar multielectrode array for extracellular recording: Application tohippocampal acute slice,” J. Neurosci. Meth., vol. 93, pp. 61–67, 1999.
[16] J. C. Chang, G. J. Brewer, and B. C. Wheeler, “Modulation of neuralnetwork activity by patterning,” Biosens. Bioelectron., vol. 16, pp.527–33, 2001.
[17] Y. Jimbo, N. Kasai, K. Torimitsu, T. Tateno, and H. P. Robinson, “Asystem for MEA-based multisite stimulation,” IEEE Trans. Biomed.Eng., vol. 50, no. 2, pp. 241–248, Feb. 2003.
[18] F. O. Morin, Y. Takamura, and E. Tamiya, “Investigating neuronal ac-tivity with planar microelectrode arrays: Achievements and new per-spectives,” J. Biosci. Bioeng., vol. 100, pp. 131–143, 2005.
[19] L. Kam, W. Shain, J. N. Turner, and R. Bizios, “Correlation of as-troglial cell function on micro-patterned surfaces with specific geo-metric parameters,” Biomaterials, vol. 20, pp. 2343–2350, 1999.
[20] L. Kam, W. Shain, J. N. Turner, and R. Bizios, “Selective adhesion ofastrocytes to surfaces modified with immobilized peptides,” Biomate-rials, vol. 23, pp. 511–315, 2002.
[21] W. Shain, L. Spataro, J. Dilgen, K. Haverstick, S. Retterer, M. Isaacson,M. Saltzman, and J. N. Turner, “Controlling cellular reactive responsesaround neural prosthetic devices using peripheral and local interventionstrategies,” IEEE Trans. Neural Syst. Rehabil. Eng., vol. 11, no. 2, pp.186–188, Jun. 2003.
[22] D. Kleinfeld, K. H. Kahler, and P. E. Hockberger, “Controlled out-growth of dissociated neurons on patterned substrates,” J. Neurosci.,vol. 8, pp. 4098–4120, 1988.
[23] C. D. James, R. Davis, M. Meyer, A. Turner, S. Turner, G. Withers,L. Kam, G. Banker, H. Craighead, M. Isaacson, J. Turner, and W.Shain, “Aligned microcontact printing of micrometer-scale poly-L-ly-sine structures for controlled growth of cultured neurons on planar mi-croelectrode arrays,” IEEE Trans. Biomed. Eng., vol. 47, no. 1, pp.17–21, Jan. 2000.
[24] E. Yavin and Z. Yavin, “Attachment and culture of dissociated cellsfrom rat embryo cerebral hemispheres on polylysine-coated surface,”J. Cell. Biol., vol. 62, pp. 540–546, 1974.
[25] P. C. Letourneau, “Possible roles for cell-to-substratum adhesion inneuronal morphogenesis,” Dev. Biol., vol. 44, pp. 77–91, 1975.
[26] W. L. McKeehan and R. G. Ham, “Stimulation of clonal growth ofnormal fibroblasts with substrata coated with basic polymers,” J. Cell.Biol., vol. 71, pp. 727–734, 1976.
[27] H. K. Kleinman, M. L. McGarvey, J. R. Hassell, V. L. Star, F. B.Cannon, G. W. Laurie, and G. R. Martin, “Basement membrane com-plexes with biological activity,” Biochemistry, vol. 25, pp. 312–8, 1986.
[28] J. R. Sanes, “Extracellular matrix molecules that influence neural de-velopment,” Annu. Rev. Neurosci., vol. 12, pp. 491–516, 1989.
[29] I. H. Lelong, V. Petegnief, and G. Rebel, “Neuronal cells mature fasteron polyethyleneimine coated plates than on polylysine coated plates,”J. Neurosci. Res., vol. 32, pp. 562–8, 1992.
[30] M. Matsuzawa, F. F. Weight, R. S. Potember, and P. Liesi, “Direc-tional neurite outgrowth and axonal differentiation of embryonic hip-pocampal neurons are promoted by a neurite outgrowth domain of theB2-chain of laminin,” Int. J. Dev. Neurosci., vol. 14, pp. 283–295, 1996.
[31] A. Goldberger, P. Flaherty, R. Villalba, L. A. Riblet, and A. Cor-nell-Bell, Extracellular matrix proteins influence cell morphology andfunction in rat neural cultures, Becton Dickinson Labware, Tech. Bull.1997.
[32] M. J. Ignatius, N. Sawhney, A. Gupta, B. M. Thibadeau, O. R. Mon-teiro, and I. G. Brown, “Bioactive surface coatings for nanoscale in-struments: Effects on cns neurons,” J. Biomed. Mater. Res., vol. 40,pp. 264–274, 1998.
[33] T. Esch, V. Lemmon, and G. Banker, “Local presentation of substratemolecules directs axon specification by cultured hippocampal neu-rons,” J. Neurosci., vol. 19, pp. 6417–6426, 1999.
[34] A. Dityatev and M. Schachner, “Extracellular matrix molecules andsynaptic plasticity,” Nat. Rev. Neurosci., vol. 4, pp. 456–468, 2003.
[35] Y. Jimbo, H. P. Robinson, and A. Kawana, “Simultaneous measure-ment of intracellular calcium and electrical activity from patternedneural networks in culture,” IEEE Trans. Biomed. Eng., vol. 40, no. 8,pp. 804–810, Aug. 1993.
[36] R. O. Wong, “Calcium imaging and multielectrode recordings of globalpatterns of activity in the developing nervous system,” Histochem. J.,vol. 30, pp. 217–229, 1998.
[37] T. Tateno, Y. Jimbo, and H. P. Robinson, “Spatio-temporal cholinergicmodulation in cultured networks of rat cortical neurons: Spontaneousactivity,” Neuroscience, vol. 134, pp. 425–437, 2005.
[38] M. H. Droge, G. W. Gross, M. H. Hightower, and L. E. Czisny, “Mul-tielectrode analysis of coordinated, multisite, rhythmic bursting in cul-tured cns monolayer networks,” J. Neurosci., vol. 6, pp. 1583–1592,1986.
[39] H. Kamioka, E. Maeda, Y. Jimbo, H. P. Robinson, and A. Kawana,“Spontaneous periodic synchronized bursting during formation of ma-ture patterns of connections in cortical cultures,” Neurosci. Lett., vol.206, pp. 109–112, 1996.
[40] R. M. Fitzsimonds, H. J. Song, and M. M. Poo, “Propagation of ac-tivity-dependent synaptic depression in simple neural networks,” Na-ture, vol. 388, pp. 439–448, 1997.

1320 IEEE TRANSACTIONS ON BIOMEDICAL ENGINEERING, VOL. 54, NO. 7, JULY 2007
[41] G. Banker and K. Goslin, Culturing Nerve Cells, 2nd ed. Cambridge,MA: MIT Press, 1999.
[42] C. R. Legendy and M. Salcman, “Bursts and recurrences of bursts inthe spike trains of spontaneously active striate cortex neurons,” J. Neu-rophysiol., vol. 53, pp. 926–939, 1985.
[43] G. Gholmieh, W. Soussou, S. Courellis, V. Z. Marmarelis, T. W.Berger, and M. Baudry, “A biosensor for detecting changes in cog-nitive processing based on nonlinear systems analysis,” Biosens.Bioelectron., vol. 16, pp. 491–501, 2001.
[44] P. S. Motta and J. W. Judy, “Multielectrode microprobes for deep-brainstimulation fabricated with a customizable 3-D electroplating process,”IEEE Trans. Biomed. Eng., vol. 52, no. 5, pp. 923–933, May 2005.
[45] R. D. Brinton, W. Soussou, M. Baudry, M. Thompson, and T. W.Berger, “The biotic/abiotic interface: Achievements and foreseeablechallenges,” in Towards Replacement Parts for the Brain: ImplantableBiomimetic Electronics as Neural Prostheses, T. W. Berger and D. L.Glanzman, Eds. Cambridge, MA: MIT Press, 2005.
[46] D. W. Branch, B. C. Wheeler, G. J. Brewer, and D. E. Leckband,“Long-term maintenance of patterns of hippocampal pyramidal cellson substrates of polyethylene glycol and microstamped polylysine,”IEEE Trans. Biomed. Eng., vol. 47, no. 3, pp. 290–300, Mar. 2000.
[47] R. Milner and I. L. Campbell, “The integrin family of cell adhesionmolecules has multiple functions within the cns,” Neurosci. Res., vol.69, pp. 286–291, 2002.
[48] P. E. Latham, B. J. Richmond, P. G. Nelson, and S. Nirenberg, “In-trinsic dynamics in neuronal networks. I. Theory,” J. Neurophysiol.,vol. 83, pp. 808–827, 2000.
[49] P. E. Latham, B. J. Richmond, S. Nirenberg, and P. G. Nelson,“Intrinsic dynamics in neuronal networks. II. Experiment,” J. Neuro-physiol., vol. 83, pp. 828–835, 2000.
[50] E. Claverol-Tinture and J. Pine, “Extracellular potentials in low-densitydissociated neuronal cultures,” J. Neurosci. Meth., vol. 117, pp. 13–21,2002.
[51] Q. Liu, M. Coulombe, J. Dumm, K. M. Shaffer, A. E. Schaffner, J. L.Barker, J. J. Pancrazio, D. A. Stenger, and W. Ma, “Synaptic connec-tivity in hippocampal neuronal networks cultured on micropatternedsurfaces,” Brain Res. Dev. Brain Res., vol. 120, pp. 223–231, 2000.
[52] W. Ma, Q. Y. Liu, D. Jung, P. Manos, J. J. Pancrazio, A. E. Schaffner,J. L. Barker, and D. A. Stenger, “Central neuronal synapse formationon micropatterned surfaces,” Brain. Res. Dev. Brain Res., vol. 111, pp.231–243, 1998.
[53] M. Canepari, M. Bove, E. Maeda, M. Cappello, and A. Kawana, “Ex-perimental analysis of neuronal dynamics in cultured cortical networksand transitions between different patterns of activity,” Biol. Cybern.,vol. 77, pp. 153–162, 1997.
[54] M. Matsuzawa, T. Tabata, W. Knoll, and M. Kano, “Formation of hip-pocampal synapses on patterned substrates of a laminin- derived syn-thetic peptide,” Eur. J. Neurosci., vol. 12, pp. 903–910, 2000.
[55] P. R. Kennedy and R. A. Bakay, “Restoration of neural output froma paralyzed patient by a direct brain connection,” Neuroreport, vol. 9,pp. 1707–1711, 1998.
Walid V. Soussou was born in Beirut, Lebanon, in1975. He received the B.S. degree in biochemistryfrom Boston College, Chestnut Hill, MA, in 1995,and the Ph.D. in neuroscience from the University ofSouthern California, Los Angeles, in 2005.
He has interned at Harvard Medical Schooland worked as a Research Technician at BostonUniversity. He is currently a postdoctoral fellowat the Burnham Institute for Medical Research. Heis author on six research papers and three bookchapters and has presented his research at several
conferences.Dr. Soussou is a member of the Society for Neuroscience and is a recipient of
the 2005 MIT Arab Student Organization’s Science and Technology GraduateStudent Award.
Geoffrey J. Yoon graduated magna cum laude withthe B.A. degree in biology from the Universityof Southern California, Los Angeles, in 2002. Heis now a 5th-year medical student at The JohnsHopkins School of Medicine, Baltimore, MD.
Roberta Diaz Brinton received the Ph.D. degreein psychobiology and neuropharmacology from theUniversity of Arizona, Tucson, in 1984 as an NIHPredoctoral Fellow.
She continued her postdoctoral training in Neu-roendocrinology at Rockefeller University, NewYork, as an NIH Postdoctoral Fellow from 1984to 1988. She is Associate Professor of MolecularPharmacology and Toxicology and of BiologicalSciences (Neurobiology), and also serves as theDirector of the University of Southern California
(USC) Science, Technology and Research Program (STAR). She co-foundedthe biotechnology company CoCensys in 1988. She joined the faculty ofUniversity of Southern California, School of Pharmacy and the NeuroscienceProgram in 1988. Her laboratory was named the Kenneth T. and Eileen L.Norris Foundation laboratory for Neuroscience Research in 1999.
Dr. Brinton serves on the scientific advisory boards for the Institute for theStudy of Aging and the Diabetes Insipidus Foundation. She has served on sci-entific review boards for the National Institutes of Health and serves on the Ex-ecutive Committee of the USC Program in Neuroscience.
Theodore W. Berger (M’03–SM’04) received thePh.D. degree in physiological psychology from Har-vard University, Cambridge, MA, in 1976, receivingthe James McKeen Cattell Award from the New YorkAcademy of Sciences for his thesis research.is theDavid Packard Professor of Engineering at the Uni-versity of Southern California, Los Angeles.
He conducted postdoctoral research at the Univer-sity of California, Irvine and was an Alfred P. SloanFoundation Fellow at The Salk Institute.In 1979, hejoined the Departments of Neuroscience and Psychi-
atry at the University of Pittsburgh, and since 1992, he has been Professor ofBiomedical Engineering and Biological Sciences at the University of SouthernCalifornia, receiving the David Packard Chair in 2003. He became Director ofthe Center for Neural Engineering in 1997.
Dr. Berger has received a McKnight Foundation Scholar Award, twice re-ceived an NIMH Research Scientist Development Award, received an NIMHSenior Scientist Award, the Lockheed Senior Research Award, the Person ofthe Year “Impact Award” by the AARP for his work in neural prostheses, andhas been both a National Academy of Sciences International Scientist Lecturerand an IEEE Distinguished Lecturer. He was elected a Fellow of the AmericanAssociation for the Advancement of Science in 1986 and a Fellow of the Amer-ican Institute for Medical and Biological Engineering in 1998.
Related Documents











