Clustered and non-clustered GABA A receptors in cultured hippocampal neurons S.B. Christie, R.-W. Li, C.P. Miralles, B.-Y. Yang, and A.L. De Blas * Department of Physiology and Neurobiology, University of Connecticut, 3107 Horsebarn Hill Rd., U-4156, Storrs, CT 06269, USA Received 7 July 2005; revised 17 August 2005; accepted 23 August 2005 Available online 21 September 2005 In cultured hippocampal neurons, ; 2 subunit-containing GABA A Rs form large postsynaptic clusters at GABAergic synapses and small clusters outside GABAergic synapses. We now show that a pool of non- clustered ; 2 subunit-containing GABA A Rs are also present at the cell surface. We also demonstrate that myc - or EGFP-tagged ; 2 , A 2 , B 3 or A 1 subunits expressed in these neurons assemble with endogenous subunits, forming GABA A Rs that target large postsynaptic clusters, small clusters outside GABAergic synapses or a pool of non-clustered surface GABA A Rs. In contrast, myc - or EGFP-tagged D subunits only form non-clustered GABA A Rs, which can be induced to form clusters by antibody capping. A myc -tagged chimeric ; 2 subunit possessing the large intracellular loop (IL) of the y-subunit IL ( myc ; 2 S/D-IL) assembled into GABA A Rs, but it did not form clusters, therefore behaving like the D subunit. Thus, the large intracellular loops of ; 2 and D play an important role in determining the synaptic clustering/ non-clustering capacity of the GABA A Rs. D 2005 Elsevier Inc. All rights reserved. Introduction The g 2 GABA A receptor (GABA A R) subunit knockout and conditional knockout mouse models demonstrate that the g 2 subunit is essential for the postsynaptic clustering of GABA A Rs (Gu ¨ nther et al., 1995; Essrich et al., 1998; Schweizer et al., 2003). This interpretation is also supported by the results obtained after knocking down the g 2 subunit by RNA interference (Li et al., in press). The g 2 knockout mice show a loss of most postsynaptically clustered GABA A Rs accompanied by severe deficits in GABAer- gic synaptic transmission, sensorimotor deficits and neonatal lethality. In these animals, the expression of the GABA A R a and h subunits was not affected, and they formed functional GABA A Rs just containing ah subunits that were translocated to the surface (Gu ¨ nther et al., 1995; Essrich et al., 1998). Thus, the postsynaptic accumulation of GABA A Rs apposed to GABA-releasing presy- naptic terminals is thought to be central to the phasic inhibitory GABAergic synaptic transmission in the brain. The g 2 and y subunits do not co-assemble within the same GABA A R (Quirk et al., 1994, 1995; Araujo et al., 1998; Jechlinger et al., 1998). Contrary to the postsynaptic clustering of the g 2 subunit-containing GABA A Rs, y subunit-containing GABA A Rs are non-synaptic or perisynaptic as shown by immunoelectron microscopy in granule cells of the cerebellum (Nusser et al., 1998) and dentate gyrus of the hippocampus (Wei et al., 2003). These extrasynaptic GABA A R receptors mediate the tonic GABAergic inhibition by sensing the ambient levels and/or spillover of GABA released from synapses (Brickley et al., 1996; De Schutter, 2002; Nusser and Mody, 2002; Stell et al., 2003; Yeung et al., 2003). Gephyrin, a glycine receptor (GlyR) interacting protein present in glycinergic and GABAergic synapses, has also been suggested to be required for the clustering of GABA A Rs (Essrich et al., 1998; Kneussel et al., 1999). However, not all GABA A Rs require gephyrin for clustering as shown in a gephyrin knockout mouse (Fischer et al., 2000; Kneussel et al., 2001; Levi et al., 2004). There is no biochemical evidence for a direct interaction of gephyrin with any of the GABA A R subunits (Meyer et al., 1995). Nevertheless, gephyrin plays an important role in normal synaptic transmission because deficits in clustering of GABA A Rs and GABAergic synaptic trans- mission are apparent in gephyrin / mice (Levi et al., 2004). Furthermore, a missense mutation in collybistin, a GDP – GTP exchange factor that is important for gephyrin clustering, leads to impaired inhibitory synapse formation (Harvey et al., 2004). There still exists an incomplete understanding of the key molecular mechanisms by which g 2 subunit-containing GABA A Rs (g 2 -GABA A Rs) are selectively retained and/or concentrated at GABAergic synapses, while y-GABA A Rs do not. However, it is likely that an interaction of the GABA A R with scaffolding proteins at the GABAergic synapses occurs via the IL of some of the GABA A R subunits. Studies using the yeast two-hybrid system have identified several proteins that interact with the IL domain of GABA A R subunits (recently reviewed or discussed by Bedford et al., 2001; Fritschy and Brunig, 2003; Herring et al., 2003; Charych et al., 2004a; Keller et al., 2004; Luscher and Keller, 2004). 1044-7431/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.mcn.2005.08.014 * Corresponding author. Fax: +1 860 486 5439. E-mail address: [email protected] (A.L. De Blas). Available online on ScienceDirect (www.sciencedirect.com). www.elsevier.com/locate/ymcne Mol. Cell. Neurosci. 31 (2006) 1 – 14

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 31 (2006) 1 – 14
Clustered and non-clustered GABAA receptors in cultured
hippocampal neurons
S.B. Christie, R.-W. Li, C.P. Miralles, B.-Y. Yang, and A.L. De Blas*
Department of Physiology and Neurobiology, University of Connecticut, 3107 Horsebarn Hill Rd., U-4156, Storrs, CT 06269, USA
Received 7 July 2005; revised 17 August 2005; accepted 23 August 2005
Available online 21 September 2005
In cultured hippocampal neurons, ;2 subunit-containing GABAARs
form large postsynaptic clusters at GABAergic synapses and small
clusters outside GABAergic synapses. We now show that a pool of non-
clustered ;2 subunit-containing GABAARs are also present at the cell
surface. We also demonstrate that myc- or EGFP-tagged ;2, A2, B3 or
A1 subunits expressed in these neurons assemble with endogenous
subunits, forming GABAARs that target large postsynaptic clusters,
small clusters outside GABAergic synapses or a pool of non-clustered
surface GABAARs. In contrast, myc- or EGFP-tagged D subunits only
form non-clustered GABAARs, which can be induced to form clusters
by antibody capping. A myc-tagged chimeric ;2 subunit possessing the
large intracellular loop (IL) of the y-subunit IL (myc;2S/D-IL)
assembled into GABAARs, but it did not form clusters, therefore
behaving like the D subunit. Thus, the large intracellular loops of ;2
and D play an important role in determining the synaptic clustering/
non-clustering capacity of the GABAARs.
D 2005 Elsevier Inc. All rights reserved.
Introduction
The g2 GABAA receptor (GABAAR) subunit knockout and
conditional knockout mouse models demonstrate that the g2subunit is essential for the postsynaptic clustering of GABAARs
(Gunther et al., 1995; Essrich et al., 1998; Schweizer et al., 2003).
This interpretation is also supported by the results obtained after
knocking down the g2 subunit by RNA interference (Li et al., in
press). The g2 knockout mice show a loss of most postsynaptically
clustered GABAARs accompanied by severe deficits in GABAer-
gic synaptic transmission, sensorimotor deficits and neonatal
lethality. In these animals, the expression of the GABAAR a and
h subunits was not affected, and they formed functional GABAARs
just containing ah subunits that were translocated to the surface
(Gunther et al., 1995; Essrich et al., 1998). Thus, the postsynaptic
accumulation of GABAARs apposed to GABA-releasing presy-
1044-7431/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2005.08.014
* Corresponding author. Fax: +1 860 486 5439.
E-mail address: [email protected] (A.L. De Blas).
Available online on ScienceDirect (www.sciencedirect.com).
naptic terminals is thought to be central to the phasic inhibitory
GABAergic synaptic transmission in the brain.
The g2 and y subunits do not co-assemble within the same
GABAAR (Quirk et al., 1994, 1995; Araujo et al., 1998;
Jechlinger et al., 1998). Contrary to the postsynaptic clustering
of the g2 subunit-containing GABAARs, y subunit-containing
GABAARs are non-synaptic or perisynaptic as shown by
immunoelectron microscopy in granule cells of the cerebellum
(Nusser et al., 1998) and dentate gyrus of the hippocampus (Wei
et al., 2003). These extrasynaptic GABAAR receptors mediate the
tonic GABAergic inhibition by sensing the ambient levels and/or
spillover of GABA released from synapses (Brickley et al., 1996;
De Schutter, 2002; Nusser and Mody, 2002; Stell et al., 2003;
Yeung et al., 2003).
Gephyrin, a glycine receptor (GlyR) interacting protein
present in glycinergic and GABAergic synapses, has also been
suggested to be required for the clustering of GABAARs
(Essrich et al., 1998; Kneussel et al., 1999). However, not all
GABAARs require gephyrin for clustering as shown in a
gephyrin knockout mouse (Fischer et al., 2000; Kneussel et
al., 2001; Levi et al., 2004). There is no biochemical evidence
for a direct interaction of gephyrin with any of the GABAAR
subunits (Meyer et al., 1995). Nevertheless, gephyrin plays an
important role in normal synaptic transmission because deficits
in clustering of GABAARs and GABAergic synaptic trans-
mission are apparent in gephyrin�/� mice (Levi et al., 2004).
Furthermore, a missense mutation in collybistin, a GDP–GTP
exchange factor that is important for gephyrin clustering, leads
to impaired inhibitory synapse formation (Harvey et al., 2004).
There still exists an incomplete understanding of the key
molecular mechanisms by which g2 subunit-containing GABAARs
(g2-GABAARs) are selectively retained and/or concentrated at
GABAergic synapses, while y-GABAARs do not. However, it is
likely that an interaction of the GABAAR with scaffolding proteins
at the GABAergic synapses occurs via the IL of some of the
GABAAR subunits. Studies using the yeast two-hybrid system
have identified several proteins that interact with the IL domain of
GABAAR subunits (recently reviewed or discussed by Bedford et
al., 2001; Fritschy and Brunig, 2003; Herring et al., 2003; Charych
et al., 2004a; Keller et al., 2004; Luscher and Keller, 2004).

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–142
Nevertheless, the identified proteins seem to be involved in
GABAAR trafficking rather than in synaptic clustering.
In this communication, we have tested the hypothesis that the
IL domains of g2 and y GABAAR subunits play a role in the
synaptic clustering capacity of the GABAARs.
Results
Before studying the expression in hippocampal neurons shown
below, all the tagged subunits and the chimera used in this study
were coexpressed in HEK293 cells with non-tagged h2 and a6
subunits. Expression, assembly and translocation to the cell surface
were demonstrated by immunofluorescence after life cell incubation
with anti-myc or anti-EGFP antibodies. These experiments showed
myc or EGFP immunofluorescence at the cell surface of the HEK293
cells that colocalized with h2 and a6 cell surface immunofluor-
escence (as determined with anti-h2/3 and anti-a6 antibodies
respectively) in triple-label experiments. Live cell incubation with
the anti-EGFP antibody of the cotransfected HEK293 cells induced
the capping of the EGFP-tagged subunit (g2 or y) and the colocalizedcapping of the h2 and a6 subunits (not shown).
Exogenous mycc2 or c2-EGFP form clusters at GABAergic synapses
that colocalize with endogenous GABAAR clusters
This is shown in Fig. 1. Hippocampal neurons transfected
with only mycg2S DNA express the exogenous subunit, which
forms large clusters (arrows, Fig. 1A) localized postsynaptically
to GAD+ (GABAergic) terminals (arrows, Fig. 1C). The mycg2S
also forms small clusters that are present in dendritic areas not
receiving GABAergic innervation (arrowheads, Figs. 1A and C).
The larger (synaptic) and smaller clusters of the endogenous
GABAAR associated protein, gephyrin, were highly colocalized
with both large (arrows, Fig. 1B) and small (arrowheads, Fig.
1B) mycg2S clusters respectively. Moreover, the size of the largemycg2S clusters at GABAergic synapses and small clusters
outside GABAergic synapses was similar to that of the
endogenous GABAAR and gephyrin clusters in non-transfected
cells (Figs. 1D–F). These larger (synaptic) and smaller clusters
have been characterized in detail elsewhere (Christie et al.,
2002a).
Table 1 shows that there is no statistically significant
difference (P = 0.58, Student’s t test) between the density ofmycg2S clusters in transfected cells (22 T 2 clusters per 100 Am2
dendrite area; n = 193 clusters in 15 dendrites from 9 cells) and
the density of endogenous g2 clusters (20 T 1 clusters per 100
Am2 dendrite area; n = 386 clusters in 18 dendrites from 9
cells) in untransfected cells. Neither is there significant differ-
ence (P = 0.74) between the density of endogenous gephyrin
clusters in transfected cells (19 T 2 clusters/100 Am2; n = 175
in 15 dendrites from 9 cells) and the density of endogenous
gephyrin clusters in untransfected cells (19 T 1 clusters/100
Am2; n = 357 in 18 dendrites from 9 cells). In transfected
neurons (Table 1) 82 T 3% of mycg2S clusters were colocalized
with gephyrin, while 91 T 3% of gephyrin clusters were
colocalized with mycg2S clusters. These values were not
significantly different than the 85 T 2% colocalization of
endogenous g2 clusters with gephyrin clusters (P = 0.19) or
the 93 T 2% colocalization of gephyrin clusters with endoge-
nous g2 clusters (P = 0.32) observed in untransfected neurons.
Thus, we find that the exogenous expression of the mycg2S subunit
in the transfected neurons integrates with the endogenous
GABAAR assembly without significantly altering the targeting of
GABAARs, density of the clusters or their colocalization with
gephyrin.
Transfection of hippocampal neurons with only g2S-EGFP
(Figs. 1G–I) or g2L-EGFP (Figs. 1J–L) also formed large and
small clusters as shown by EGFP fluorescence (Figs. 1G and J)
and with an anti-g2 antibody (Fig. 1H). These clusters
colocalized with endogenous GABAAR subunits, as shown by
immunofluorescence with anti-h2/3 GABAAR subunit (arrows,
Figs. 1I and K), at GABAergic (vGAT+) synapses (arrows, Fig
1L). These experiments strongly suggest that the mycg2S, g2S-
EGFP or g2L-EGFP co-assemble with endogenous subunits to
form GABAARs. This notion is further supported because others
have shown that some myc- and EGFP-tagged GABAAR
subunits co-assemble with endogenous subunits forming func-
tional GABAARs (Connolly et al., 1996; Kittler et al., 2000;
Alldred et al., 2005). Moreover, g2 GABAAR subunits that do
not assemble with a or h GABAAR subunits are retained within
the ER, do not translocate to the cell surface and are rapidly
degraded (Connolly et al., 1999).
It is also worth mentioning that it has been proposed (Kittler
et al., 2000) that: (I) a g2S when tagged at the N-terminus with
EGFP (EGFP-g2S), but not when tagged at the C-terminus
(g2S-EGFP), can form clusters that colocalize with other
GABAAR subunits, (II) exogenous a and h subunits need to
be cotransfected together with the EGFP-g2S for the latter to
assemble into GABAARs and to form clusters. Our results
instead show that (I) the g2S and g2L GABAAR subunits
tagged at the C-terminus with EGFP can also form synaptic
GABAAR clusters and (II) cotransfection of exogenous a and hsubunits is not required since g2S-EGFP and g2L-EGFP can co-
assemble with endogenous GABAAR subunits and form clusters
of similar density and size to the endogenous GABAAR
clusters, probably because the endogenous subunits are rate-
limiting for receptor assembly.
Various GABAAR subunits tagged at the N-terminus or the
C-terminus form clusters that colocalize with endogenous GABAAR
clusters
We also tagged other GABAAR subunits with EGFP (a1-EGFP,
a2-EGFP and h3-EGFP) and made a double-tagged mycg2S-EGFP
subunit. Hippocampal neurons transfected with each of these
tagged subunits displayed large EGFP clusters (arrows, Figs. 2A, D
and G, respectively) that were colocalized with endogenous
gephyrin clusters (arrows, Figs. 2B, E and H) in areas postsynaptic
to GAD+ boutons (arrows, Figs. 2C, F and I). Similar results were
obtained using an a2-EGFP made by us (not shown) or another a1-
EGFP construct produced in Dr. Stefano Vicini’s laboratory (not
shown). Smaller clusters of EGFP-tagged subunit that colocalized
with smaller gephyrin clusters were observed in dendritic areas not
receiving GABAergic innervation (arrowheads, Fig. 2).
These experiments strongly suggest that the tagged a1, a2 or h3
subunits co-assemble with the endogenous subunit partners (e.g.
exogenous-tagged a assembles with endogenous h and g subunits
or exogenous-tagged h assembles with endogenous a or g
subunits) since only GABAAR pentamers that contain the g
subunit (plus have a and h) target to GABAergic synapses (Essrichet al., 1998; Schweizer et al., 2003; Li et al., in press).
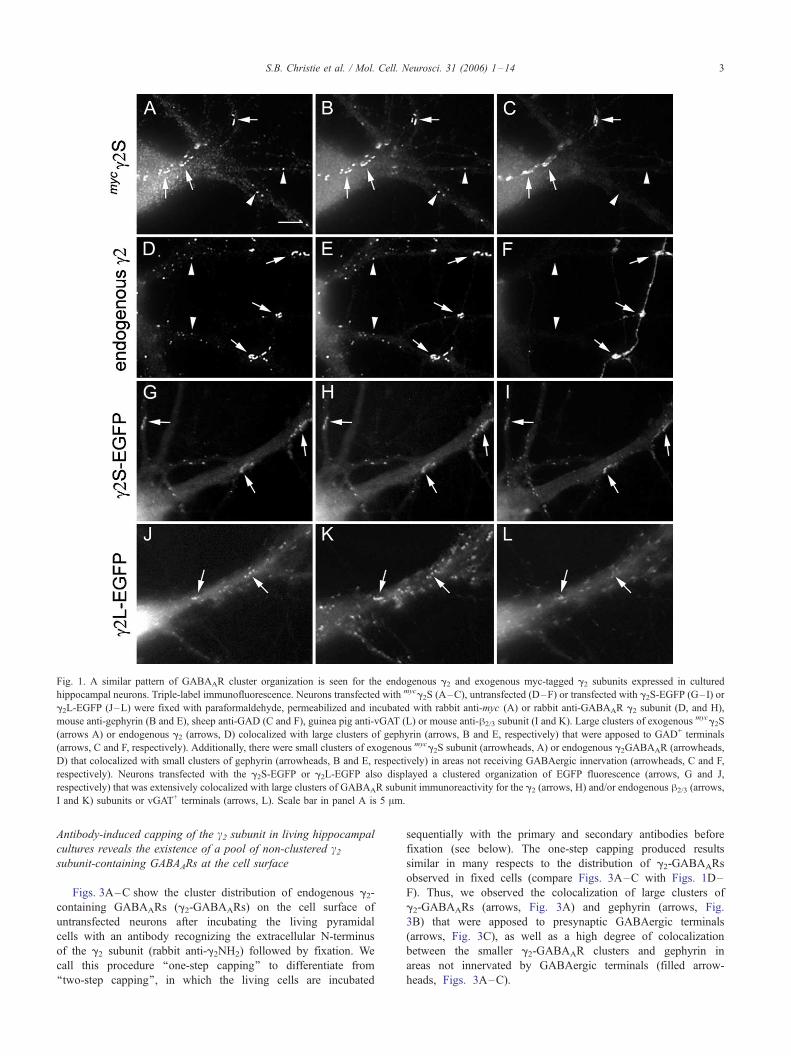
Fig. 1. A similar pattern of GABAAR cluster organization is seen for the endogenous g2 and exogenous myc-tagged g2 subunits expressed in cultured
hippocampal neurons. Triple-label immunofluorescence. Neurons transfected with mycg2S (A–C), untransfected (D–F) or transfected with g2S-EGFP (G–I) or
g2L-EGFP (J–L) were fixed with paraformaldehyde, permeabilized and incubated with rabbit anti-myc (A) or rabbit anti-GABAAR g2 subunit (D, and H),
mouse anti-gephyrin (B and E), sheep anti-GAD (C and F), guinea pig anti-vGAT (L) or mouse anti-h2/3 subunit (I and K). Large clusters of exogenousmycg2S
(arrows A) or endogenous g2 (arrows, D) colocalized with large clusters of gephyrin (arrows, B and E, respectively) that were apposed to GAD+ terminals
(arrows, C and F, respectively). Additionally, there were small clusters of exogenous mycg2S subunit (arrowheads, A) or endogenous g2GABAAR (arrowheads,
D) that colocalized with small clusters of gephyrin (arrowheads, B and E, respectively) in areas not receiving GABAergic innervation (arrowheads, C and F,
respectively). Neurons transfected with the g2S-EGFP or g2L-EGFP also displayed a clustered organization of EGFP fluorescence (arrows, G and J,
respectively) that was extensively colocalized with large clusters of GABAAR subunit immunoreactivity for the g2 (arrows, H) and/or endogenous h2/3 (arrows,
I and K) subunits or vGAT+ terminals (arrows, L). Scale bar in panel A is 5 Am.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 3
Antibody-induced capping of the c2 subunit in living hippocampal
cultures reveals the existence of a pool of non-clustered c2subunit-containing GABAARs at the cell surface
Figs. 3A–C show the cluster distribution of endogenous g2-
containing GABAARs (g2-GABAARs) on the cell surface of
untransfected neurons after incubating the living pyramidal
cells with an antibody recognizing the extracellular N-terminus
of the g2 subunit (rabbit anti-g2NH2) followed by fixation. We
call this procedure ‘‘one-step capping’’ to differentiate from
‘‘two-step capping’’, in which the living cells are incubated
sequentially with the primary and secondary antibodies before
fixation (see below). The one-step capping produced results
similar in many respects to the distribution of g2-GABAARs
observed in fixed cells (compare Figs. 3A–C with Figs. 1D–
F). Thus, we observed the colocalization of large clusters of
g2-GABAARs (arrows, Fig. 3A) and gephyrin (arrows, Fig.
3B) that were apposed to presynaptic GABAergic terminals
(arrows, Fig. 3C), as well as a high degree of colocalization
between the smaller g2-GABAAR clusters and gephyrin in
areas not innervated by GABAergic terminals (filled arrow-
heads, Figs. 3A–C).
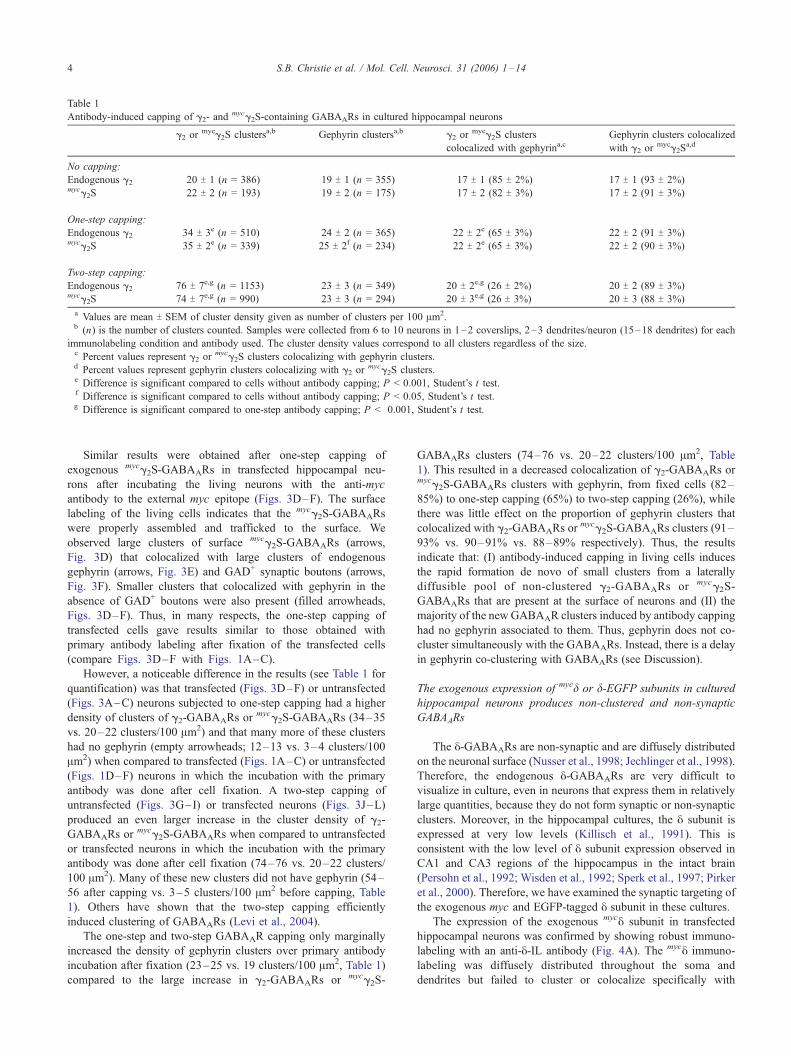
Table 1
Antibody-induced capping of g2- andmycg2S-containing GABAARs in cultured hippocampal neurons
g2 ormycg2S clustersa,b Gephyrin clustersa,b g2 or
mycg2S clusters
colocalized with gephyrina,cGephyrin clusters colocalized
with g2 ormycg2S
a,d
No capping:
Endogenous g2 20 T 1 (n = 386) 19 T 1 (n = 355) 17 T 1 (85 T 2%) 17 T 1 (93 T 2%)mycg2S 22 T 2 (n = 193) 19 T 2 (n = 175) 17 T 2 (82 T 3%) 17 T 2 (91 T 3%)
One-step capping:
Endogenous g2 34 T 3e (n = 510) 24 T 2 (n = 365) 22 T 2e (65 T 3%) 22 T 2 (91 T 3%)mycg2S 35 T 2e (n = 339) 25 T 2f (n = 234) 22 T 2e (65 T 3%) 22 T 2 (90 T 3%)
Two-step capping:
Endogenous g2 76 T 7e,g (n = 1153) 23 T 3 (n = 349) 20 T 2e,g (26 T 2%) 20 T 2 (89 T 3%)mycg2S 74 T 7e,g (n = 990) 23 T 3 (n = 294) 20 T 3e,g (26 T 3%) 20 T 3 (88 T 3%)
a Values are mean T SEM of cluster density given as number of clusters per 100 Am2.b (n) is the number of clusters counted. Samples were collected from 6 to 10 neurons in 1–2 coverslips, 2–3 dendrites/neuron (15–18 dendrites) for each
immunolabeling condition and antibody used. The cluster density values correspond to all clusters regardless of the size.c Percent values represent g2 or
mycg2S clusters colocalizing with gephyrin clusters.d Percent values represent gephyrin clusters colocalizing with g2 or
mycg2S clusters.e Difference is significant compared to cells without antibody capping; P < 0.001, Student’s t test.f Difference is significant compared to cells without antibody capping; P < 0.05, Student’s t test.g Difference is significant compared to one-step antibody capping; P < 0.001, Student’s t test.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–144
Similar results were obtained after one-step capping of
exogenous mycg2S-GABAARs in transfected hippocampal neu-
rons after incubating the living neurons with the anti-myc
antibody to the external myc epitope (Figs. 3D–F). The surface
labeling of the living cells indicates that the mycg2S-GABAARs
were properly assembled and trafficked to the surface. We
observed large clusters of surface mycg2S-GABAARs (arrows,
Fig. 3D) that colocalized with large clusters of endogenous
gephyrin (arrows, Fig. 3E) and GAD+ synaptic boutons (arrows,
Fig. 3F). Smaller clusters that colocalized with gephyrin in the
absence of GAD+ boutons were also present (filled arrowheads,
Figs. 3D–F). Thus, in many respects, the one-step capping of
transfected cells gave results similar to those obtained with
primary antibody labeling after fixation of the transfected cells
(compare Figs. 3D–F with Figs. 1A–C).
However, a noticeable difference in the results (see Table 1 for
quantification) was that transfected (Figs. 3D–F) or untransfected
(Figs. 3A–C) neurons subjected to one-step capping had a higher
density of clusters of g2-GABAARs ormycg2S-GABAARs (34–35
vs. 20–22 clusters/100 Am2) and that many more of these clusters
had no gephyrin (empty arrowheads; 12–13 vs. 3–4 clusters/100
Am2) when compared to transfected (Figs. 1A–C) or untransfected
(Figs. 1D–F) neurons in which the incubation with the primary
antibody was done after cell fixation. A two-step capping of
untransfected (Figs. 3G–I) or transfected neurons (Figs. 3J–L)
produced an even larger increase in the cluster density of g2-
GABAARs ormycg2S-GABAARs when compared to untransfected
or transfected neurons in which the incubation with the primary
antibody was done after cell fixation (74–76 vs. 20–22 clusters/
100 Am2). Many of these new clusters did not have gephyrin (54–
56 after capping vs. 3–5 clusters/100 Am2 before capping, Table
1). Others have shown that the two-step capping efficiently
induced clustering of GABAARs (Levi et al., 2004).
The one-step and two-step GABAAR capping only marginally
increased the density of gephyrin clusters over primary antibody
incubation after fixation (23–25 vs. 19 clusters/100 Am2, Table 1)
compared to the large increase in g2-GABAARs or mycg2S-
GABAARs clusters (74–76 vs. 20–22 clusters/100 Am2, Table
1). This resulted in a decreased colocalization of g2-GABAARs ormycg2S-GABAARs clusters with gephyrin, from fixed cells (82–
85%) to one-step capping (65%) to two-step capping (26%), while
there was little effect on the proportion of gephyrin clusters that
colocalized with g2-GABAARs ormycg2S-GABAARs clusters (91–
93% vs. 90–91% vs. 88–89% respectively). Thus, the results
indicate that: (I) antibody-induced capping in living cells induces
the rapid formation de novo of small clusters from a laterally
diffusible pool of non-clustered g2-GABAARs or mycg2S-
GABAARs that are present at the surface of neurons and (II) the
majority of the new GABAAR clusters induced by antibody capping
had no gephyrin associated to them. Thus, gephyrin does not co-
cluster simultaneously with the GABAARs. Instead, there is a delay
in gephyrin co-clustering with GABAARs (see Discussion).
The exogenous expression of mycd or d-EGFP subunits in cultured
hippocampal neurons produces non-clustered and non-synaptic
GABAARs
The y-GABAARs are non-synaptic and are diffusely distributed
on the neuronal surface (Nusser et al., 1998; Jechlinger et al., 1998).
Therefore, the endogenous y-GABAARs are very difficult to
visualize in culture, even in neurons that express them in relatively
large quantities, because they do not form synaptic or non-synaptic
clusters. Moreover, in the hippocampal cultures, the y subunit is
expressed at very low levels (Killisch et al., 1991). This is
consistent with the low level of y subunit expression observed in
CA1 and CA3 regions of the hippocampus in the intact brain
(Persohn et al., 1992; Wisden et al., 1992; Sperk et al., 1997; Pirker
et al., 2000). Therefore, we have examined the synaptic targeting of
the exogenous myc and EGFP-tagged y subunit in these cultures.
The expression of the exogenous mycy subunit in transfected
hippocampal neurons was confirmed by showing robust immuno-
labeling with an anti-y-IL antibody (Fig. 4A). The mycy immuno-
labeling was diffusely distributed throughout the soma and
dendrites but failed to cluster or colocalize specifically with

Fig. 2. Various exogenous GABAAR-EGFP subunits form GABAAR clusters that colocalize with gephyrin clusters at GABAergic synapses. Triple-label
immunofluorescence. Hippocampal neurons transfected with either a1-EGFP (A), h3-EGFP (D) or mycg2S-EGFP (G) were fixed, permeabilized and labeled
with mouse anti-gephyrin (B, E and H) and sheep anti-GAD (C, F and I). For all EGFP fusion subunits examined, the larger clusters of EGFP fluorescence
(arrows, A, D and G) colocalized with the larger clusters of gephyrin (arrows, B, E and H) that were apposed to GAD+ synapses (arrows, C, F and I). In
addition, extensive colocalization was observed between the small EGFP clusters (arrowheads, A, D and G) and gephyrin (arrowheads, B, E and H) in the
absence of GABAergic terminals (arrowheads, C, F and I). Scale bar in panel A is 5 Am.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 5
gephyrin clusters or GAD at GABAergic synapses (arrows, Figs.
4B and C, respectively) or with any of the small gephyrin clusters
outside of GABAergic synapses (arrowheads, Fig. 4B). Similar
subcellular distribution of anti-y-IL immunolabeling was observed
in fixed neurons that had been transfected with either an untagged
y subunit (not shown) or a novel y-EGFP subunit generated in our
laboratory (Figs. 4D–F).
In the cerebellar granule cells, the a6 subunit co-assembles with
the y subunit (and h) and forms extrasynaptic y-GABAARs or with
the g2 subunit (and h) and forms synaptic g2-GABAARs (Nusser et
al., 1998; Jechlinger et al., 1998). When we transfected hippo-
campal neurons with a plasmid encoding the exogenous non-tagged
a6 subunit (a subunit not expressed in the hippocampus or in these
cultures), they produced a6-GABAAR clusters that colocalized with
gephyrin (arrows, Figs. 5A and B, respectively) at GABAergic
GAD+ synapses (not shown). The observed clustering and synaptic
targeting were presumably mediated by the assembly of exogenous
a6 with endogenous g2 and h subunits since the synaptic targeting
of the a subunits is severely impaired unless it co-assembles with g2(Gunther et al., 1995; Essrich et al., 1998; Li et al., in press).
When we cotransfected hippocampal neurons with a combina-
tion of y-EGFP and a6, we found a diffuse distribution of the y-EGFP fluorescence (Fig. 5C), similar to that observed in cells
expressing only the y-EGFP (Fig. 4D), thereby discounting a role
for a6 in significantly recruiting y-EGFP to postsynaptic clusters
since there was no colocalization with gephyrin clusters (Fig. 5D).
These results are consistent with the notion that in the brain a6 co-
assembles with either g2 or y (or y-EGFP) but not with both
subunits in the same receptor pentamer (Quirk et al., 1994, 1995;
Araujo et al., 1998; Jechlinger et al., 1998).
Further evidence for the non-synaptic distribution of the y-EGFP containing GABAARs that are present at the cell surface was
obtained by antibody-induced capping. One-step capping of living
neurons with polyclonal anti-GFP antibody revealed numerous
small and disorganized EGFP clusters that decorate the processes
(arrowheads, Fig. 5E) of cells transfected with the y-EGFP subunit.
These small EGFP clusters are not readily observed in areas of
contact by GAD+ terminals (arrows, Fig. 5F).
In order to confirm that in cotransfected neurons the exogenous
a6 co-assembled with y-EGFP to form surface expressed but non-
clustered GABAARs, we performed a two-step capping of living
neurons using a polyclonal anti-GFP and a secondary antibody
(arrowheads, Fig. 5G). We observed that the y-EGFP clusters
induced by two-step capping frequently colocalized with a subset of
small a6-GABAAR clusters (arrowheads, Figs. 5G and H,
respectively). However, none of the induced y-EGFP clusters
colocalized with the larger and synaptic clusters of a6 (arrows, Figs.
5G and H, respectively), which are postsynaptic to GAD+ terminals

Fig. 3. Antibody-induced capping of endogenous g2- ormycg2-GABAARs that are present on the cell surface. Triple-label immunofluorescence. One-step
antibody capping (A–F) of live neurons with an affinity-purified rabbit antibody to the N-terminus of the g2 (A) or a rabbit anti-myc (D) induced the formation
of GABAAR clusters of g2-GABAARs in untransfected cells (A) or mycg2S-GABAARs in transfected cells (D). Two-step antibody-induced capping (G–L)
induced the formation of additional clusters of endogenous g2-GABAARs in untransfected cells (G) ormycg2S-GABAARs in transfected cells (J). After capping,
cells were fixed, permeabilized and incubated with a mixture of mouse anti-gephyrin (B, E, H and K) and sheep anti-GAD (C, F, I and L) followed by a mixture
of the secondary antibodies. Larger clusters of the endogenous g2-GABAARs (arrows, A and G) and the exogenous mycg2S-GABAARs (arrows, D and J)
colocalized with large gephyrin clusters (arrows, B, E, H and K) at GAD+ synapses (arrows, C, F, I and L). Smaller clusters of the endogenous g2-GABAARs
(filled arrowheads, A and G) and the exogenous mycg2S-GABAARs (filled arrowheads, D and J) were also colocalized with gephyrin (filled arrowheads, B, E,
H and K) in areas not innervated by GABAergic terminals (filled arrowheads, C, F, I and L). However, aggregates of endogenous g2-GABAARs (empty
arrowheads, A and G) or exogenous mycg2S-GABAARs (empty arrowheads, D and J) that did not colocalize with gephyrin (empty arrowheads, B, E, H and K)
or GAD+ terminals (empty arrowheads, C, F, I and L) were abundant throughout all processes. These clusters were greater in number after two-step capping
than one-step capping. Scale bar in panel A is 5 Am.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–146
(not shown). These and the aforementioned results indicate that a6
co-assembled with either the endogenous g2 or exogenous y-EGFPsubunits (but not with both), forming GABAARs that were
differentially targeted to synaptic or non-synaptic areas, respec-
tively. This is consistent with the behavior of endogenous a6 in
cerebellar granule cells, where it assembles with either y or g2(Quirk et al., 1995; Jechlinger et al., 1998) to form extrasynaptic or
synaptic receptors, respectively, in the same cell (Nusser et al.,
1998).
The large intracellular loop of the c2 and d subunits plays a role in
the synaptic clustering of the GABAA receptors
Studies with other receptors of the same family have shown that
the large IL domain of the GlyR h subunit (Kirsch et al., 1995) and
the nAChR a3 subunit (Williams et al., 1998) is critical to the
synaptic targeting and clustering of these receptors. We have now
tested the hypothesis that the IL of the g2 and y subunits plays an
important role in determining the clustering of the GABAARs. For
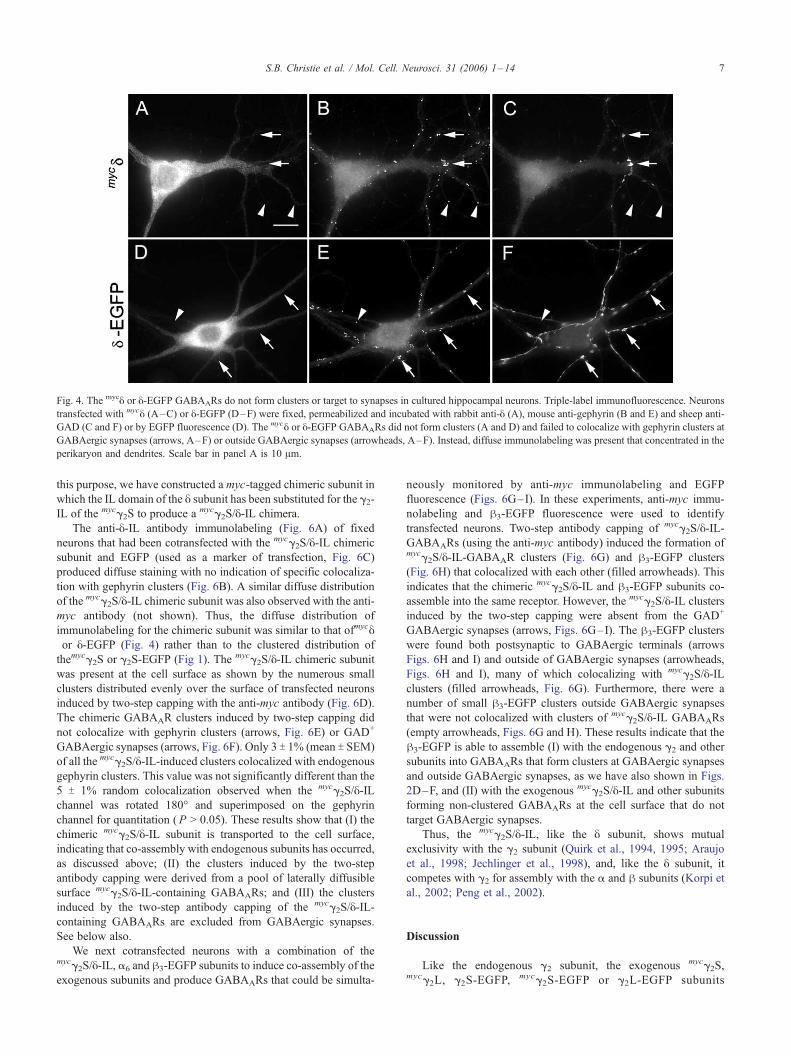
Fig. 4. The mycy or y-EGFP GABAARs do not form clusters or target to synapses in cultured hippocampal neurons. Triple-label immunofluorescence. Neurons
transfected with mycy (A–C) or y-EGFP (D–F) were fixed, permeabilized and incubated with rabbit anti-y (A), mouse anti-gephyrin (B and E) and sheep anti-
GAD (C and F) or by EGFP fluorescence (D). The mycy or y-EGFP GABAARs did not form clusters (A and D) and failed to colocalize with gephyrin clusters at
GABAergic synapses (arrows, A–F) or outside GABAergic synapses (arrowheads, A–F). Instead, diffuse immunolabeling was present that concentrated in the
perikaryon and dendrites. Scale bar in panel A is 10 Am.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 7
this purpose, we have constructed a myc-tagged chimeric subunit in
which the IL domain of the y subunit has been substituted for the g2-IL of the mycg2S to produce a mycg2S/y-IL chimera.
The anti-y-IL antibody immunolabeling (Fig. 6A) of fixed
neurons that had been cotransfected with the mycg2S/y-IL chimeric
subunit and EGFP (used as a marker of transfection, Fig. 6C)
produced diffuse staining with no indication of specific colocaliza-
tion with gephyrin clusters (Fig. 6B). A similar diffuse distribution
of the mycg2S/y-IL chimeric subunit was also observed with the anti-
myc antibody (not shown). Thus, the diffuse distribution of
immunolabeling for the chimeric subunit was similar to that ofmycyor y-EGFP (Fig. 4) rather than to the clustered distribution of
themycg2S or g2S-EGFP (Fig 1). The mycg2S/y-IL chimeric subunit
was present at the cell surface as shown by the numerous small
clusters distributed evenly over the surface of transfected neurons
induced by two-step capping with the anti-myc antibody (Fig. 6D).
The chimeric GABAAR clusters induced by two-step capping did
not colocalize with gephyrin clusters (arrows, Fig. 6E) or GAD+
GABAergic synapses (arrows, Fig. 6F). Only 3 T 1% (mean T SEM)
of all the mycg2S/y-IL-induced clusters colocalized with endogenousgephyrin clusters. This value was not significantly different than the
5 T 1% random colocalization observed when the mycg2S/y-ILchannel was rotated 180- and superimposed on the gephyrin
channel for quantitation (P > 0.05). These results show that (I) the
chimeric mycg2S/y-IL subunit is transported to the cell surface,
indicating that co-assembly with endogenous subunits has occurred,
as discussed above; (II) the clusters induced by the two-step
antibody capping were derived from a pool of laterally diffusible
surface mycg2S/y-IL-containing GABAARs; and (III) the clusters
induced by the two-step antibody capping of the mycg2S/y-IL-containing GABAARs are excluded from GABAergic synapses.
See below also.
We next cotransfected neurons with a combination of themycg2S/y-IL, a6 and h3-EGFP subunits to induce co-assembly of the
exogenous subunits and produce GABAARs that could be simulta-
neously monitored by anti-myc immunolabeling and EGFP
fluorescence (Figs. 6G–I). In these experiments, anti-myc immu-
nolabeling and h3-EGFP fluorescence were used to identify
transfected neurons. Two-step antibody capping of mycg2S/y-IL-GABAARs (using the anti-myc antibody) induced the formation ofmycg2S/y-IL-GABAAR clusters (Fig. 6G) and h3-EGFP clusters
(Fig. 6H) that colocalized with each other (filled arrowheads). This
indicates that the chimeric mycg2S/y-IL and h3-EGFP subunits co-
assemble into the same receptor. However, the mycg2S/y-IL clusters
induced by the two-step capping were absent from the GAD+
GABAergic synapses (arrows, Figs. 6G–I). The h3-EGFP clusters
were found both postsynaptic to GABAergic terminals (arrows
Figs. 6H and I) and outside of GABAergic synapses (arrowheads,
Figs. 6H and I), many of which colocalizing with mycg2S/y-ILclusters (filled arrowheads, Fig. 6G). Furthermore, there were a
number of small h3-EGFP clusters outside GABAergic synapses
that were not colocalized with clusters of mycg2S/y-IL GABAARs
(empty arrowheads, Figs. 6G and H). These results indicate that the
h3-EGFP is able to assemble (I) with the endogenous g2 and other
subunits into GABAARs that form clusters at GABAergic synapses
and outside GABAergic synapses, as we have also shown in Figs.
2D–F, and (II) with the exogenous mycg2S/y-IL and other subunits
forming non-clustered GABAARs at the cell surface that do not
target GABAergic synapses.
Thus, the mycg2S/y-IL, like the y subunit, shows mutual
exclusivity with the g2 subunit (Quirk et al., 1994, 1995; Araujo
et al., 1998; Jechlinger et al., 1998), and, like the y subunit, it
competes with g2 for assembly with the a and h subunits (Korpi et
al., 2002; Peng et al., 2002).
Discussion
Like the endogenous g2 subunit, the exogenous mycg2S,mycg2L, g2S-EGFP, mycg2S-EGFP or g2L-EGFP subunits
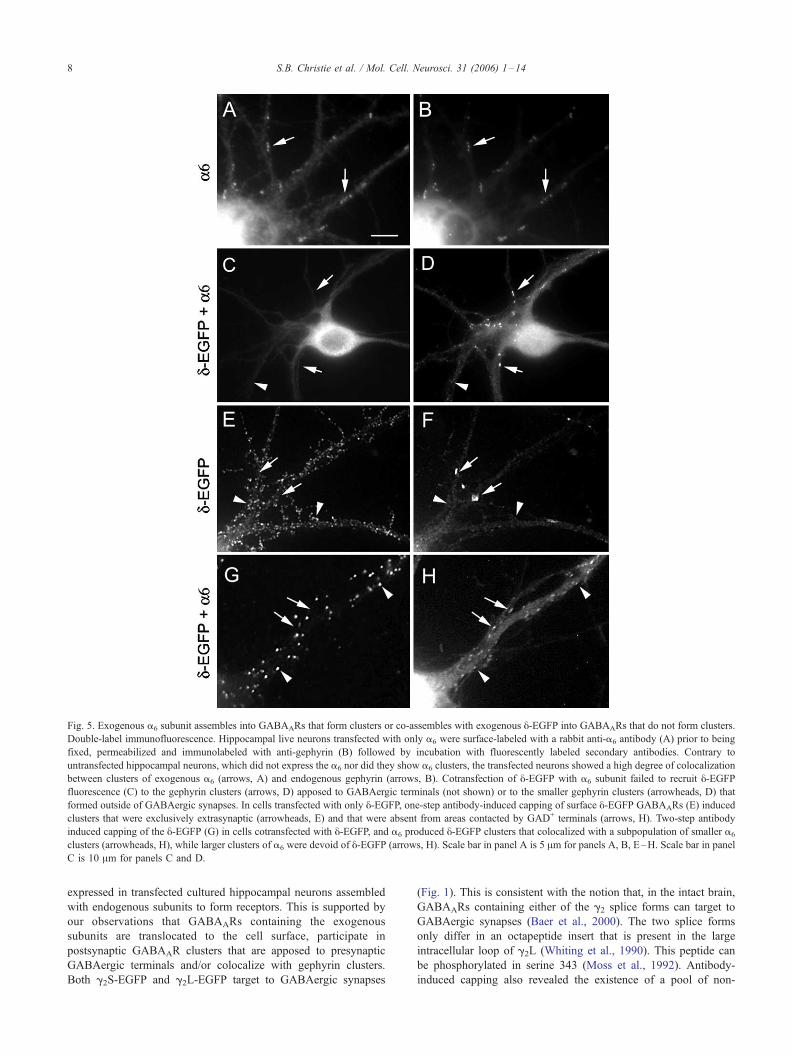
Fig. 5. Exogenous a6 subunit assembles into GABAARs that form clusters or co-assembles with exogenous y-EGFP into GABAARs that do not form clusters.
Double-label immunofluorescence. Hippocampal live neurons transfected with only a6 were surface-labeled with a rabbit anti-a6 antibody (A) prior to being
fixed, permeabilized and immunolabeled with anti-gephyrin (B) followed by incubation with fluorescently labeled secondary antibodies. Contrary to
untransfected hippocampal neurons, which did not express the a6 nor did they show a6 clusters, the transfected neurons showed a high degree of colocalization
between clusters of exogenous a6 (arrows, A) and endogenous gephyrin (arrows, B). Cotransfection of y-EGFP with a6 subunit failed to recruit y-EGFPfluorescence (C) to the gephyrin clusters (arrows, D) apposed to GABAergic terminals (not shown) or to the smaller gephyrin clusters (arrowheads, D) that
formed outside of GABAergic synapses. In cells transfected with only y-EGFP, one-step antibody-induced capping of surface y-EGFP GABAARs (E) induced
clusters that were exclusively extrasynaptic (arrowheads, E) and that were absent from areas contacted by GAD+ terminals (arrows, H). Two-step antibody
induced capping of the y-EGFP (G) in cells cotransfected with y-EGFP, and a6 produced y-EGFP clusters that colocalized with a subpopulation of smaller a6
clusters (arrowheads, H), while larger clusters of a6 were devoid of y-EGFP (arrows, H). Scale bar in panel A is 5 Am for panels A, B, E–H. Scale bar in panel
C is 10 Am for panels C and D.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–148
expressed in transfected cultured hippocampal neurons assembled
with endogenous subunits to form receptors. This is supported by
our observations that GABAARs containing the exogenous
subunits are translocated to the cell surface, participate in
postsynaptic GABAAR clusters that are apposed to presynaptic
GABAergic terminals and/or colocalize with gephyrin clusters.
Both g2S-EGFP and g2L-EGFP target to GABAergic synapses
(Fig. 1). This is consistent with the notion that, in the intact brain,
GABAARs containing either of the g2 splice forms can target to
GABAergic synapses (Baer et al., 2000). The two splice forms
only differ in an octapeptide insert that is present in the large
intracellular loop of g2L (Whiting et al., 1990). This peptide can
be phosphorylated in serine 343 (Moss et al., 1992). Antibody-
induced capping also revealed the existence of a pool of non-

Fig. 6. Surface GABAARs containing the chimeric mycg2S/y-IL subunit do not form clusters unless capping of the GABAARs in the living neuron is induced by
antibodies. Triple-label immunofluorescence. Hippocampal neurons cotransfected with the chimeric mycg2S/y-IL subunit and EGFP (A–C) were fixed,
permeabilized and incubated with rabbit anti-y-IL (A) and mouse anti-gephyrin (B). Transfected cells were identified by EGFP fluorescence (C). Most EGFP-
positive neurons also displayed y-IL immunoreactivity (used to indicate proper expression and monitor subcellular localization of the chimeric subunit) that
was diffusely distributed throughout the soma and proximal dendrites (A) and did not colocalize with clustered gephyrin (B). Two-step antibody-induced
capping of living cells with the rabbit anti-myc antibody and the FITC-conjugated donkey anti-rabbit IgG secondary antibody (D–I) of hippocampal neurons
transfected with only mycg2S/y-IL (D–F) or cotransfected with a mixture of mycg2S/y-IL, a6 and h3-EGFP (G–I) induces the formation of mycg2S/y-ILGABAAR clusters (D and G). The antibody-induced clusters (filled arrowheads, D and G) did not colocalized with gephyrin clusters (E) or were present at
GABAergic synapses containing GAD+ terminals (arrows, F and I) and postsynaptic gephyrin (arrows, E). In the triple-transfected neurons (G–I), antibody-
induced clustering of mycg2S/y-IL (filled arrowheads, G) induces the co-clustering of h3-EGFP (filled arrowheads, H). These neurons also have clusters of h3-
EGFP at GABAergic synapses (arrows, H) and outside GABAergic synapses (empty arrowheads, H) that do not colocalize with the antibody-induced mycg2S/y-IL clusters. Scale bar in panel A is 10 Am for panels A–C; scale bar in panel D is 5 Am for panels D–I.
S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 9
clustered GABAARs containing the endogenous g2 or the tagged
g2 subunit that is present at the cell surface of the non-transfected
and transfected hippocampal neurons, respectively. Quantification
of the cluster density indicates that the pool of non-clustered
endogenous or tagged g2-GABAARs at the cell surface is
significant.
A different result was obtained with the y subunit since the y-GABAARs were diffusely localized at the cell surface, not
forming any clusters at GABAergic synapses or outside
GABAergic synapses. We have also found that the chimericmycg2S/y-IL subunit, a g2 subunit with the IL of the y subunit, co-
assembles with other subunits to produce GABAARs that are
diffusely distributed on the cell surface and do not participate in
synaptic or non-synaptic GABAAR clusters, thus behaving like
the y subunit. These results support the notion that the large
intracellular loops of the native g2 and y subunits are domains
critical to the clustering of the GABAARs. The results also
suggest that the IL is more relevant to the clustering of GABAARs
than to their targeting to GABAergic synapses since y or mycg2S/
y-IL-containing GABAARs do not cluster at GABAergic synapses
or outside GABAergic synapses.
We and others have shown that clustered GABAARs are not
only localized to GABAergic synapses, forming large clusters, but
are capable of forming small clusters outside of GABAergic
synapses, often being mismatched to glutamatergic presynaptic
terminals (Rao et al., 2000; Christie et al., 2002a; Christie and de
Blas, 2003, Studler et al., 2005). Both large and small clusters in
these mature cultures have a high degree of colocalization with
gephyrin (Christie et al., 2002a). The present study, and that of
Levi et al. (2004), demonstrates that GABAAR clusters formed
rapidly after antibody-induced capping do not initially colocalize
with gephyrin. However, given enough time, gephyrin clusters
colocalize with the antibody-induced a2-GABAAR clusters,
suggesting that gephyrin is recruited to GABAAR clusters after
the latter are formed (Levi et al., 2004). Moreover, most
extrasynaptic clusters of a2 subunit-containing GABAARs in
immature cultures form independently of gephyrin (Studler et al.,
2005). Thus, gephyrin clustering follows GABAAR clustering.

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–1410
The antibody-induced capping supports a model of lateral g2-
GABAARs diffusion across the surface membrane of neurons
(Barnes, 2001; Triller and Choquet, 2005) that may also mediate
a dynamic exchange between surface clusters. This model is also
consistent with: (I) at the EM level, g2-GABAARs were not only
observed to be concentrated in postsynaptic membranes but were
also seen in non-synaptic membranes of granule cells of the intact
cerebellum (Nusser et al., 1998); (II) several reports describe the
presence of diffuse GABAAR subunit immunoreactivity that is
present outside of the synaptic clusters (Essrich et al., 1998;
Brunig et al., 2002b; Christie et al., 2002a,b; Christie and de
Blas, 2003; Levi et al., 2004; Studler et al., 2005); and (III) the
surface distribution of a5-GABAARs in cultured hippocampal
neurons is found in both GABAergic synapses (Christie and De
Blas, 2002) and extrasynaptically (Brunig et al., 2002a; Christie
and De Blas, 2002). Since the hippocampal a5-containing
GABAARs have a predominant a5h3g2 composition (Fritschy et
al., 1997; Sur et al., 1998), the enumerated data above suggest
that the presence of the g2 subunit within GABAARs does not
necessarily targets all the surface g2-GABAARs to the post-
synaptic region. Therefore, the g2 subunit may be essential for
the postsynaptic clustering of GABAARs (Gunther et al., 1995;
Essrich et al., 1998; Schweizer et al., 2003; Li et al., in press),
but it might not be sufficient since other subunits seem to also
play a role (i.e. a5 and a2).
For glycine receptors, it has been shown that their postsynaptic
accumulation in clusters is a highly dynamic process involving
lateral diffusion of the GlyRs (Meier et al., 2000, 2001; Rosenberg
et al., 2001; Meier, 2003; Triller and Choquet, 2003). The direct
interaction of the GlyR h-IL subunit with gephyrin seems to be
involved in the postsynaptic accumulation of the GlyR (Kirsch et
al., 1993; Meyer et al., 1995). The GlyR–gephyrin interaction is
important for the rate of surface accumulation, postsynaptic
accumulation, postsynaptic stabilization and internalization of
GlyRs from synapses to intracellular organelles (Kirsch et al.,
1993; Feng et al., 1998; Rasmussen et al., 2002; Hanus et al., 2004;
Levi et al., 2004). Furthermore, GlyR and Ca+ channel activity has
also been shown to be important for GlyR surface stability and the
postsynaptic accumulation of the glycine receptor (Kirsch and
Betz, 1998; Levi et al., 1998; Kneussel and Betz, 2000).
However, in the case of GABAARs clusters, the colocalizing
gephyrin has not been shown to directly interact with any of the
GABAAR subunit isoforms and appears to be dispensable for the
postsynaptic formation of some GABAergic postsynaptic receptor
clusters (Levi et al., 2004; Studler et al., 2005). Moreover, unlike
GlyR clustering, the clustering of GABAARs occurs independently
of receptor activation (Craig et al., 1994). Recent reports suggest
that the mechanism by which the GABAARs normally cluster and
are postsynaptically localized are dissociable events, with extra-
synaptic GABAAR clustering preceding colocalization with
gephyrin and/or synaptic targeting (Levi et al., 2004; Graf et al.,
2004; Studler et al., 2005). This is also supported by our antibody-
induced capping experiments as discussed above.
In the hippocampus, expression of the y subunit is primarily
localized to granule cells of the dentate gyrus (Sperk et al., 1997;
Pirker et al., 2000). Within these cells, electrophysiological and EM
immunocytochemical evidence supports exclusion of these sub-
types from GABAergic synapses (Stell and Mody, 2002; Wei et al.,
2003). The in situ hybridization studies (Persohn et al., 1992,
Wisden et al., 1992) also reveal low levels of y subunit expression
within the pyramidal cells of CA1 field. Our results showing that
the tagged y subunit distributed diffusely at the cell surface and did
not form synaptic or non-synaptic clusters were consistent with the
reported extrasynaptic distribution of the y subunit-containing
receptors in all cells expressing this subunit.
Like the myc- or EGFP-tagged y subunit, the mycg2S/y-ILchimeric subunit formed GABAARs that translocated to the surface
but did not form or participate in synaptic or non-synaptic GABAAR
clusters. Non-synaptic clusters of these receptors were observed
only after antibody-induced capping. Thus, the loss of clustering
capacity observed for the mycg2S/y-IL-GABAARs, compared to themycg2S-GABAARs, is consistent with the hypothesis that the IL
domain of the g2 subunit is necessary for clustering to occur.
However, this interpretation contrasts a recent report in which
analysis of various chimeric g2–a2 subunits mapped the TM4
region of g2 as the domain that was ‘‘required and sufficient’’ for
synaptic clustering (Alldred et al., 2005). Because our mycg2S/y-ILchimeric subunit contains the TM4 domain of g2, we concluded that
the TM4 domain of g2 is not sufficient for clustering activity when
the y-IL is present, despite the ability of receptors containing these
chimeric subunits to access the surface. We were unable to test
whether the g2-IL was sufficient to induce the clustering of the mycy/g2S-IL-GABAARs. The chimeric mycy/g2S-IL subunit did not
express in HEK293 cells or in hippocampal neurons.
The apparent discrepancy between Alldred et al.’s (2005)
results and our results may be due to the fact that the various g2chimeras used by Alldred et al. (2005) were produced by
exchanging equivalent domains between g2 and a2, two subunits
that cluster at GABAergic synapses. In our case, we exchanged
equivalent domains between a subunit that clusters at GABAergic
synapses (g2) and a subunit that does not cluster at GABAergic
synapses (y). Thus, the a2 subunit may be neutral or weakly
permissive with regard to clustering and/or synaptic targeting,
while the y-IL may prevent the clustering of GABAARs.
Furthermore, post-translational palmitoylation of the g2-IL domain
has been shown to occur (Keller et al., 2004; Rathenberg et al.,
2004) and is essential in regulating the clustering of GABAARs at
postsynaptic sites (Rathenberg et al., 2004) which is also consistent
with our results. Perhaps in the mycg2S/y-IL GABAARs, the
presence of the y-IL disrupts or supersedes the g2 TM4 clustering
signal identified by Alldred et al. (2005). Therefore, GABAAR
clustering might depend on the presence of permissive domains
(e.g. possessing the g2-IL and the g2 TM4 domains but not the y-ILdomain) or restrictive (e.g. possessing the y-IL domain) signals that
are present within the pentameric receptor.
Experimental methods
Construction of c-myc tagged GABAAR subunits
Coding sequences for the GABAAR subunits rat g2S, rat y and
human g2L, contained within the pcDNA3.1 (+) mammalian
expression vector (Invitrogen), were tagged with the 10-amino-acid
epitope (EQKLISEEDL) from c-myc (Evan et al., 1985) between the
fourth and fifth amino acids of the mature subunit peptide sequence.
Briefly, a site-directed mutagenesis strategy described by Connolly
et al. (1996) was performed using the Gene Editor in vitro Site-
Directed Mutagenesis System (Promega) with PAGE purified 5Vphosphorylated antisense oligonucleotides (5V-GC ATA ATC TTC
ATA GTC ATC TAG GTC TTC TTC TGA TAT TAG TTT TTG
TTC ATC TGA CTT TTG GCTAGT G-3V, 5V-GCATAATCT TCA

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 11
TAG TCA TCT AGG TCT TCT TCT GAT ATT AGT TTT TGT
TCATCAGATTTCTGGCTAGTG 3Vand 5V-GTCATTCATTGC
TCT GGC GCC TAG GTC TTC TTC TGA TAT TAG TTT TTG
TTC ATG GTG CGG CTG CGT GCA CAG-3V) to create the
ratmycg2S, ratmycy and human mycg2L subunits, respectively. The
correct nucleotide sequences for the full-lengthmyc-tagged subunits
were confirmed by direct sequencing. Correct expression of themyc-
tagged subunit proteins was confirmed in transfected HEK293 cells
by immunolabeling with the monoclonal antibody (mAb) 9E10
(DSHB, University of Iowa) or a rabbit anti-myc (Upstate, Waltham
MA) antibody. A rabbit anti y-IL, produced in our laboratory to a
synthetic peptide (amino acids, a.a., 390–402) of y, was also used tostudy the expression of the mycy subunit and the mAb KC5-5E1,
raised in our laboratory against the g2-IL region (Fernando et al.,
1995), to study the expression of the mycg2S and mycg2L subunits.
Construction of chimeric subunits
To produce the chimeric mycg2S/y-IL subunit, BamHI and
EcoRI restriction sites flanking the large IL region were introduced
into the rat mycg2S and rat mycy subunits by site-directed
mutagenesis, as described above. Thus, the BamHI site spanning
the junction of the TM3/IL and the EcoRI site, six nucleotide bases
into the TM4 region, were created using the 5V phosphorylatedantisense oligonucleotides in the rat mycg2S (5V-TGG TTT CCG
GTT GCT CAC AAA ATA GGATTC GGTACC ATA CTC CAC
CAA AGC TGA-3V, 5V-GGC GGT AGG GAA GAA GAT GAA
TTC ATA GAA GTC CAT TTT GGC-3V) and in the mycy (5V-CCGTTT CTT CCT GTA GTC AGC ATT GAA GGATCC AAATGC
ATA CTC CAC CAG GGC AGC-3V, 5V-GGC TGC CGG GAA
CAC AGC GAA TTC ATA GAT GTC GAT GGT GTC-3V).The mutated clones were then double-digested with BamHI
and EcoRI enzymes, and the resultant fragments were gel-
extracted, purified and cross-ligated to produce the chimericmycg2S/y-IL subunit containing BamHI and EcoRI restriction sites.
The restriction sites flanking the IL region were then removed by
site-directed mutagenesis using the 5V phosphorylated oligos formycg2S/y-IL (5V-CCG TTT CTT CCT GTA GTC AGC ATT GAA
GTG CAG GGT ACC ATA CTC CAC CAA AGC TGA-3V, 5V-GCA GAA GGC GGT AGG GAA GAA GAT CCG AGC ATA
GAT GTC GAT GGT GTC TGC ATC-3V) to restore the original
nucleotide coding sequences appropriate to the respective subunit
isoform domains. The resulting chimeric subunit clones were then
sequenced to confirm that the mutagenesis reactions did not alter
the predicted primary amino acid sequence of the translated
chimeras. Correct expression of the mycg2S/y-IL chimera was
confirmed in transfected HEK293 cells, by separate immunolab-
eling experiments using rabbit anti-myc or rabbit anti y-ILantibodies.
Construction of EGFP-tagged GABAAR subunits
EGFP-tagged subunits (rat a1, a2, h3, g2S, g2L and human ytagged at the C-terminal) were fusion proteins constructed using
the pEGFP-N1 vector (Invitrogen, Carlsbad CA). Briefly, the
coding region for each subunit was amplified by PCR using
primers that created a restriction site before the 5V start codon
(NheI, for a1, h3, g2S, g2L and y; XhoI for a2) and replaced the 3Vstop codon with a restriction site (XhoI for a1, PstI for a2, h3, g2S,
g2L and y). The replacement of the 3V subunit insert’s stop codon
with either PstI or XhoI restriction sites ensured that the coding
region for the subunit was in frame with the start codon of the
EGFP coding region, thus creating a linker sequence between the
subunit and EGFP that was based on the sequence in the plasmid
vector (13 a.a. for PstI site and 20 a.a. for the XhoI site). EGFP-
tagged subunit clones were sequenced to confirm correctness and
expressed in HEK cells to confirm expression and detection by
both EGFP fluorescence and anti-subunit antibodies.
Transfection of primary cultures of hippocampal neurons
Hippocampal cultures were prepared according to Goslin et al.
(1998), as described elsewhere (Christie et al., 2002a; Christie and
de Blas, 2003). Briefly, dissociated cells were prepared from
embryonic day 18 Sprague–Dawley rat pup hippocampi and
plated at a density of 20,000–30,000 cells per 18 mm diameter
circular coverslip treated with poly-l-lysine (Sigma). The cover-
slips with cell suspension media were then placed in 5% CO2 at
37-C for 4–6 h to allow settling and attachment of cells. The
coverslips with attached cells were then placed upside down in 60
mm diameter petri dishes containing glial feeder layers (Goslin et
al., 1998) to maintain conditioning of the support medium (DMEM
containing 0.6% glucose, 26 mM NaHCO3, N-2 supplement and
0.1% ovalbumin) and placed in a 5% CO2 atmosphere at 37-C.After 2–3 days, a final concentration of 5 � 10�6 M cytosine
arabinoside was added for 24 h. Cultures were maintained by
replacement of one-third volume of fresh N2 supplemented
DMEM medium every 3–5 days.
Hippocampal neurons were maintained in culture for 12 days
prior to transfection by using the CalPhos Mammalian Transfection
Kit (BD Biosciences, Palo Alto, CA) following manufacturer’s
protocol using 1–2 Ag DNA of each plasmid for each coverslip
used in the transfection. Immunocytochemistry was done 7 days
after transfection, unless otherwise noted.
Antibodies
The primary antibodies, rabbit anti-g2 (1–15 a.a.) and rabbit
anti-y-IL (390–402 a.a.) were raised and affinity-purified in our
laboratory against synthetic peptides made to a unique extracellular
epitope (N-terminus for g2) or the large IL domain (for y) of ratGABAAR subunits. The specificity of the affinity-purified anti-
bodies has been determined by Western blots and immunopreci-
pitation of rat brain extracts, rat brain immunohistochemistry and
immunofluorescence after transfection of GABAAR subunits into
HEK293 cells. The rabbit anti-a6 and rabbit anti-g2 antibodies
have been used in previous studies (Gutierrez et al., 1996; Christie
et al., 2002a,b; Christie and De Blas, 2002, 2003; Charych et al.,
2004b). For antibody identification of myc- or EGFP-tagged
receptor subunits, rabbit affinity-purified anti-myc antibody from
Upstate (Charlottesville, VA) and either chicken anti-GFP or rabbit
anti-GFP from Chemicon (Temecula, CA) was used. The mono-
clonal mouse anti-gephyrin (mAb 7a) was purchased from
Cedarlane (Accurate Chemical and Scientific Corp., Westbury,
NY). Sheep anti-GAD (gift of I. Kopin, NINDS, Bethesda) and
guinea pig anti-synaptic vesicle GABA transporter (VGAT) from
Chemicon were used to identify interneurons and/or GABAergic
presynaptic processes. The secondary antibodies purchased from
Jackson Immunoreagents (West Grove, PA) were all raised in
donkey (crossreactivity eliminated by adsorption with IgG from
other species) and were conjugated with Texas Red, FITC or
AMCA fluorophores.

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–1412
Live cell fluorescence immunocytochemistry by incubation with
primary or combination of primary and secondary antibodies
Triple-label immunofluorescence detection of fixed and per-
meabilized cells was done as described elsewhere (Christie et al.,
2002a). For antibody-induced capping of surface receptors by
primary antibody (one-step capping), 19 DIV live neurons were
incubated in support medium containing anti-g2-NH2, rabbit anti-
myc or rabbit anti-GFP antibodies at 37-C for 30 min in a 5% CO2
atmosphere. Coverslips were then twice washed with pH- and
temperature-equilibrated DMEM medium at 37-C for 5 min. In the
two-step capping, there was a live cell incubation with the primary
antibody (done as above) followed by washing with temperature-
and pH-equilibrated DMEM and by incubation for 30 min at 37-Cwith DMEM containing FITC- or Texas-Red-conjugated donkey
anti-rabbit antibodies. Others have shown that antibody-induced
capping with this protocol induces the clustering of GABAARs at
the cell surface with little or no internalization (Levi et al., 2004).
Following live cell incubations with primary or primary and
secondary antibodies, samples were briefly rinsed in phosphate-
buffered saline (PBS) and fixed by immersion in 4% paraformal-
dehyde, 4% sucrose in PBS for 15 min at RT followed by
permeabilization with 0.25% Triton X-100 in PBS for 5 min. Non-
specific antibody labeling was minimized by treatment with 5%
donkey serum in PBS for 30 min at room temperature. For labeling
cytoplasmic antigens (i.e. gephyrin and GAD), the fixed cultures
were incubated with a mixture of primary antibodies to the
intracellular antigens diluted in 0.25% Triton X-100 PBS for 2 h at
RT. Coverslips were washed with PBS and incubated for 1 h at
room temperature with a mixture of species-specific secondary
anti-IgG antibodies raised in donkey and conjugated to either
Texas Red, FITC or AMCA diluted in 0.25% Triton X-100 PBS.
The coverslips were washed with PBS and mounted using Prolong
anti-fade mounting solution (Molecular Probes; Eugene, Oregon).
Image acquisition and analysis
Images were collected using a 60� pan-fluor objective on a
Nikon Eclipse T200 microscope with a Spot 2e cooled CCD camera
(with a Kodak 1401E chip) driven by Spot image acquisition
software. Image files (1315 � 1035 pixel resolution) were then
sharpened using the unsharpen mask tool (settings: amount = 120%,
radius = 1.5 pixels, threshold = 0 levels) and merged for color
colocalization and figure preparation using PhotoShop 4.01
(Adobe). Control slides in which one or more primary antibodies
were omitted showed no spill over in the other two fluorescence
channels. Random drift of the sample’s fluorescence signal between
channels was controlled by alignment of all channels using triple-
labeled fluorescent microspheres (0.1 Am and 0.4 Am diameter;
Molecular Probes, Eugene, OR). Prior to processing images for
quantitation, two or three 50 Am dendritic lengths to be examined
were collected in 6–12 cells from each coverslip and condition.
Areas of the selected regions were measured directly for each
dendrite by outlining and dividing the total number of pixels
contained within the region by the constant value 81 pixels/Am2.
This value was determined by calibration of the Nikon 60�objective and CCD camera field (9 pixels/Am) using a Leica
micrometer. Quantification of signal was performed by normalizing
maximum intensity data between fluorophore channels followed by
the subtraction of the diffuse non-clustered fluorescence signal seen
in the dendrites. The two- or three-color channel images to be
compared were merged and clusters counted over 50 Am dendrite
lengths. A cluster in a fluorescence channel was considered
colocalizing with a cluster in the other fluorescence channel when
>25% of the surface of one of the two clusters overlapped with the
cluster in the other channel. Values were then normalized to 100 Am2
and used for statistical comparison by Student’s t test and reported as
mean T standard error of the mean (SEM).
Acknowledgments
This work was supported by National Institute of Neurological
Disorders and Stroke grants NS38752 and NS39287. We would
also like to thank Dr. Peter Seeburg for kindly providing the cDNA
clones for the GABAAR a1, a2, h3, g2S and g2L subunits, Dr.
Robert Macdonald for the GABAAR y subunit cDNA clone and
Dr. Stephano Vicini for one of the a1-EGFP plasmids.
References
Alldred, M.J., Mulder-Rosi, J., Lingenfelter, S.E., Chen, G., Luscher, B.,
2005. Distinct gamma2 subunit domains mediate clustering and
synaptic function of postsynaptic GABAA receptors and gephyrin.
J. Neurosci. 25, 594–603.
Araujo, F., Ruano, D., Vitorica, J., 1998. Absence of association between
delta and gamma2 subunits in native GABA(A) receptors from rat
brain. Eur. J. Pharmacol. 347, 347–353.
Baer, K., Essrich, C., Balsiger, S., Wick, M.J., Harris, R.A., Fritschy, J.M.,
Luscher, B., 2000. Rescue of gamma2 subunit-deficient mice by
transgenic overexpression of the GABAA receptor gamma2S or
gamma2L subunit isoforms. Eur. J. Neurosci. 12, 2639–2643.
Barnes, E.M., 2001. Assembly and intracellular trafficking of GABAA
receptors. Int. Rev. Neurobiol. 48, 1–29.
Bedford, F.K., Kittler, J.T., Muller, E., Thomas, P., Uren, J.M., Merlo, D.,
Wisden, W., Triller, A., Smart, T.G., Moss, S.J., 2001. GABA(A)
receptor cell surface number and subunit stability are regulated by the
ubiquitin-like protein Plic-1. Nat. Neurosci. 4, 908–916.
Brickley, S.G., Cull-Candy, S.G., Farrant, M., 1996. Development of a
tonic form of synaptic inhibition in rat cerebellar granule cells
resulting from persistent activation of GABAA receptors. J. Physiol.
497, 753–759.
Brunig, I., Scotti, E., Sidler, C., Fritschy, J.-M., 2002a. Intact
sorting, targeting, and clustering of gamma-aminobutyric acid A
receptor subtypes in hippocampal neurons in vitro. J. Comp.
Neurol. 443, 43–55.
Brunig, I., Suter, A., Knuesel, I., Luscher, B., Fritschy, J.-M., 2002b.
GABAergic terminals are required for postsynaptic clustering of
dystrophin but not of GABA(A) receptors and gephyrin. J. Neurosci.
22, 4805–4813.
Charych, E.I., Yu, W., Miralles, C.P., Serwanski, D.R., Li, X., Rubio,
M., de Blas, A.L., 2004a. The brefeldin A-inhibited GDP/GTP
exchange factor 2, a protein involved in vesicular trafficking, interacts
with the beta subunits of the GABA-A receptors. J. Neurochem. 90,
173–189.
Charych, E.I., Yu, W., Li, R., Serwanski, D.R., Miralles, C.P., Li, X., Yang,
B.Y., Pinal, N., Walikonis, R., De Blas, A.L., 2004b. A four PDZ
domain-containing splice variant form of GRIP1 is localized in
GABAergic and glutamatergic synapses in the brain. J. Biol. Chem.
279, 38978–38990.
Christie, S.B., De Blas, A.L., 2002. a5 subunit-containing GABAARs form
clusters at GABAergic synapses in hippocampal cultures. NeuroReport
13, 2355–2358.
Christie, S.B., de Blas, A.L., 2003. GABAergic and glutamatergic axons
innervate the axon initial segment and organize GABAA receptor

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–14 13
clusters of cultured hippocampal pyramidal cells. J. Comp. Neurol. 456,
361–374.
Christie, S.B., Miralles, C.P., de Blas, A.L., 2002a. GABAergic innervation
organizes synaptic and extrasynaptic GABAAR clustering in cultured
hippocampal neurons. J. Neurosci. 22, 684–697.
Christie, S.B., Li, R., Miralles, C.P., Riquelme, R., Yang, B.Y., Charych, E.,
Yu, W., Daniels, S.B., Cantino, M.E., De Blas, A.L., 2002b. Synaptic
and Extrasynaptic GABAA receptor and gephyrin clusters. In: Azmitia,
E.C., DeFelipe, J., Jones, E.G., Rakic, P., Ribak, C.E. (Eds.), Progress in
Brain Research, vol. 136. Elsevier, New York, pp. 157–179.
Connolly, C.N., Krishek, B.J., McDonald, B.J., Smart, T.G., Moss, S.J.,
1996. Assembly and cell surface expression of heteromeric and
homomeric gamma-aminobutyric acid type A receptors. J. Biol. Chem.
271, 89–96.
Connolly, C.N., Uren, J.M., Thomas, P., Gorrie, G.H., Gibson, A., Smart,
T.G., Moss, S.J., 1999. Subcellular localization and endocytosis of
homomeric gamma2 subunit splice variants of gamma-aminobutyric
acid type A receptors. Mol. Cell. Neurosci. 13, 259–271.
Craig, A.M., Blackstone, C.D., Huganir, R.L., Banker, G., 1994. Selective
clustering of glutamate and gamma-aminobutyric acid receptors
opposite terminals releasing the corresponding neurotransmitters. Proc.
Natl. Acad. Sci. U. S. A. 91, 12373–12377.
De Schutter, E., 2002. Cerebellar cortex: computation by extrasynaptic
inhibition? Curr. Biol. 12, R363–R365.
Essrich, C., Lorez, M., Benson, J.A., Fritschy, J.-M., Luscher, B., 1998.
Postsynaptic clustering of major GABAA receptor subtypes requires the
gamma 2 subunit and gephyrin. Nat. Neurosci. 1, 563–571.
Evan, G.I., Lewis, G.K., Ramsay, G., Bishop, J.M., 1985. Isolation of
monoclonal antibodies specific for human c-myc proto-oncogene
product. Mol. Cell. Biol. 12, 3610–3616.
Feng, G., Tintrup, H., Kirsch, J., Nichol, M.C., Kuhse, J., Betz, H., Sanes,
J.R., 1998. Dual requirement for gephyrin in glycine receptor clustering
and molybdoenzyme activity. Science 282, 1321–1324.
Fernando, L.P., Khan, Z.U., McKernan, R.M., De Blas, A.L., 1995.
Monoclonal antibodies to the human gamma 2 subunit of the
GABAA/benzodiazepine receptors. J. Neurochem. 64, 1305–1311.
Fischer, F., Kneussel, M., Tintrup, H., Haverkamp, S., Rauen, T., Betz, H.,
Wassle, H., 2000. Reduced synaptic clustering of GABA and glycine
receptors in the retina of the gephyrin null mutant mouse. J. Comp.
Neurol. 427, 634–648.
Fritschy, J.-M., Brunig, I., 2003. Formation and plasticity of GABAergic
synapses: physiological mechanisms and pathophysiological implica-
tions. Pharmacol. Ther. 98, 299–323.
Fritschy, J.-M., Benke, D., Johnson, D.K., Mohler, H., Rudolph, U., 1997.
GABAA-receptor alpha-subunit is an essential prerequisite for receptor
formation in vivo. Neuroscience 81, 1043–1053.
Goslin, K., Asmussen, H., Banker, G., 1998. Rat hippocampal neurons in
low-density culture. In: Banker, G., Goslin, K. (Eds.), Culturing Nerve
Cells, 2nd. edR MIT Press, Cambridge, MA, pp. 339–370.
Graf, E.R., Zhang, X., Jin. S, X., Linhoff, M.W., Craig, A.M., 2004.
Neurexins induce differentiation of GABA and glutamate postsynaptic
specializations via neuroligins. Cell 119, 1013–1026.
Gunther, U., Benson, J., Benke, D., Fritschy, J-M., Reyes, G.,
Knoflach, F., Crestani, F., Aguzzi, A., Arigoni, M., Lang, Y.,
Bluethmann, H., Mohler, H., Luscher, B., 1995. Benzodiazepine-
insensitive mice generated by targeted disruption of the gamma 2
subunit gene of gamma-aminobutyric acid type A receptors. Proc.
Natl. Acad. Sci. U. S. A. 92, 7749–7753.
Gutierrez, A., Khan, Z.U., De Blas, A.L., 1996. Immunocytochemical
localization of the alpha 6 subunit of the gamma-aminobutyric acidA
receptor in the rat nervous system. J. Comp. Neurol. 365, 504–510.
Hanus, C., Vannier, C., Triller, A., 2004. Intracellular association of glycine
receptor with gephyrin increases its plasma membrane accumulation
rate. J. Neurosci. 24, 1119–1128.
Harvey, K., Duguid, I.C., Alldred, M.J., Beatty, S.E., Ward, H., Keep, N.H.,
Lingenfelter, S.E., Pearce, B.R., Lundgren, J., Owen, M.J., Smart, T.G.,
Luscher, B., Rees, M.I., Harvey, R.J., 2004. The GDP–GTP exchange
factor collybistin: an essential determinant of neuronal gephyrin
clustering. J. Neurosci. 24, 5816–5826.
Herring, D., Huang, R., Singh, M., Robinson, L.C., Dillon, G.H.,
Leidenheimer, N.J., 2003. Constitutive GABAA receptor endocytosis
is dynamin-mediated and dependent on a dileucine AP2 adaptin-binding
motif within the beta 2 subunit of the receptor. J. Biol. Chem. 278,
24046–24052.
Jechlinger, M., Pelz, R., Tretter, V., Klausberger, T., Sieghart, W., 1998.
Subunit composition and quantitative importance of hetero-oligomeric
receptors: GABAA receptors containing alpha6 subunits. J. Neurosci.
18, 2449–2457.
Keller, C.A., Yuan, X., Panzanelli, P., Martin, M.L., Alldred, M., Sassoe-
Pognetto, M., Luscher, B., 2004. The gamma2 subunit of GABA(A)
receptors is a substrate for palmitoylation by GODZ. J. Neurosci. 24,
5881–5891.
Killisch, I., Dotti, C.G., Laurie, D.J., Luddens, H., Seeburg, P.H., 1991.
Expression patterns of GABAA receptor subtypes in developing
hippocampal neurons. Neuron 7, 927–936.
Kirsch, J., Betz, H., 1998. Glycine-receptor activation is required for
receptor clustering in spinal neurons. Nature 392, 717–720.
Kirsch, J., Wolters, I., Triller, A., Betz, H., 1993. Gephyrin antisense
oligonucleotides prevent glycine receptor clustering in spinal neurons.
Nature 366, 745–748.
Kirsch, J., Kuhse, J., Betz, H., 1995. Targeting of glycine receptor subunits
to gephyrin-rich domains in transfected human embryonic kidney cells.
Mol. Cell. Neurosci. 6, 450–461.
Kittler, J.T., Wang, J., Connolly, C.N., Vicini, S., Smart, T.G., Moss, S.J.,
2000. Analysis of GABAA receptor assembly in mammalian cell lines
and hippocampal neurons using gamma 2 subunit green fluorescent
protein chimeras. Mol. Cell. Neurosci. 16, 440–452.
Kneussel, M., Betz, H., 2000. Clustering of inhibitory neurotransmitter
receptors at developing postsynaptic sites: the membrane activation
model. Trends Neurosci. 23, 429–435.
Kneussel, M., Bransdtatter, J.H., Laube, B., Stahl, S., Muller, U., Betz, H.,
1999. Loss of postsynaptic GABA(A) receptor clustering in gephyrin-
deficient mice. J. Neurosci. 19, 9289–9297.
Kneussel, M., Brandstatter, J.H., Gasnier, B., Feng, G., Sanes, J.R., Betz,
H., 2001. Gephyrin-independent clustering of postsynaptic gaba(a)
receptor subtypes. Mol. Cell. Neurosci. 17, 973–982.
Korpi, E.R., Mihalek, R.M., Sinkkonen, S.T., Hauer, B., Hevers, W.,
Homanics, G.E., Sieghart, W., Luddens, H., 2002. Altered receptor
subtypes in the forebrain of GABA(A) receptor delta subunit-
deficient mice: recruitment of gamma 2 subunits. Neuroscience 109,
733–743.
Levi, S., Vannier, C., Triller, A., 1998. Strychnine-sensitive stabilization of
postsynaptic glycine receptor clusters. J. Cell. Sci. 111, 335–345.
Levi, S., Logan, S.M., Tovar, K.R., Craig, A.M., 2004. Gephyrin is critical for
glycine receptor clustering but not for the formation of functional
GABAergic synapses in hippocampal neurons. J. Neurosci. 24, 207–217.
Li, R.-W., Yu, W., Christie, S., Miralles, C.P., Bai, J., LoTurco, J.J., De
Blas, A.L., in press. Disruption of postsynaptic GABAA receptor
clusters leads to decreased GABAergic innervation of pyramidal
neurons. J. Neurochem.
Luscher, B., Keller, C.A., 2004. Regulation of GABAA receptor trafficking,
channel activity, and functional plasticity of inhibitory synapses.
Pharmacol. Ther. 102, 195–221.
Meier, J., 2003. The enigma of transmitter-selective receptor accumulation
at developing inhibitory synapses. Cell Tissue Res. 311, 271–276.
Meier, J., Meunier-Durmort, C., Forest, C., Triller, A., Vannier, C., 2000.
Formation of glycine receptor clusters and their accumulation at
synapses. J. Cell. Sci. 113, 2783–2795.
Meier, J., Vannier, C., Serge, A., Triller, A., Choquet, D., 2001. Fast and
reversible trapping of surface glycine receptors by gephyrin. Nat.
Neurosci. 4, 253–260.
Meyer, G., Kirsch, J., Betz, H., Langosch, D., 1995. Identification of a
gephyrin binding motif on the glycine receptor beta subunit. Neuron 15,
563–572.

S.B. Christie et al. / Mol. Cell. Neurosci. 31 (2006) 1–1414
Moss, S.J., Doherty, C.A., Huganir, R.L., 1992. Identification of the cAMP-
dependent protein kinase and protein kinase C phosphorylation sites
within the major intracellular domains of the beta 1, gamma 2S, and
gamma 2L subunits of the gamma-aminobutyric acid type A receptor.
J. Biol. Chem. 267, 14470–14476.
Nusser, Z., Mody, I., 2002. Selective modulation of tonic and phasic inhibitions
in dentate gyrus granule cells. J. Neurophysiol. 87, 2624–2628.
Nusser, Z., Sieghart, W., Somogyi, P., 1998. Segregation of different
GABAA receptors to synaptic and extrasynaptic membranes of
cerebellar granule cells. J. Neurosci. 18, 1693–1703.
Peng, Z., Hauer, B., Mihalek, R.M., Homanics, G.E., Sieghart, W., Olsen,
R.W., Houser, C.R., 2002. GABA(A) receptor changes in delta subunit-
deficient mice: altered expression of alpha4 and gamma2 subunits in the
forebrain. J. Comp. Neurol. 446, 179–197.
Persohn, E., Malherbe, P., Richards, J.G., 1992. Comparative molecular
neuroatomy of cloned GABAA receptor subunits in the rat CNS.
J. Comp. Neurol. 326, 193–216.
Pirker, S., Schwarzer, C., Wieselthaler, A., Sieghart, W., Sperk, G., 2000.
GABA(A) receptors: immunocytochemical distribution of 13 subunits
in the adult rat. Neuroscience 101, 815–850.
Quirk, K., Gillard, N.P., Ragan, C.I., Whiting, P.J., McKernan, R.M., 1994.
Model of subunit composition of gamma-aminobutyric acid A receptor
subtypes expressed in rat cerebellum with respect to their alpha and
gamma/delta subunits. J. Biol. Chem. 269, 16020–16028.
Quirk, K., Whiting, P.J., Ragan, C.I., McKernan, R.M., 1995. Character-
isation of delta-subunit containing GABAA receptors from rat brain.
Eur. J. Pharmacol. 290, 175–181.
Rao, A., Cha, E.M., Craig, A.M., 2000. Mismatched appositions of
presynaptic and postsynaptic components in isolated hippocampal
neurons. J. Neurosci. 20, 8344–8353.
Rasmussen, H., Rasmussen, T., Triller, A., Vannier, C., 2002. Strychnine-
blocked glycine receptor is removed from synapses by a shift in
insertion/degradation equilibrium. Mol. Cell. Neurosci. 19, 201–215.
Rathenberg, J., Kittler, J.T., Moss, S.J., 2004. Palmitoylation regulates the
clustering and cell surface stability of GABAA receptors. Mol. Cell.
Neurosci. 26, 217–251.
Rosenberg, M., Meier, J., Triller, A., Vannier, C., 2001. Dynamics of glycine
receptor insertion in the neuronal plasma membrane. J. Neurosci. 21,
5036–5044.
Schweizer, C., Balsiger, S., Bluethmann, H., Mansuy, I.M., Fritschy, J.-M.,
Mohler, H., Luscher, B., 2003. The gamma 2 subunit of GABA(A)
receptors is required for maintenance of receptors at mature synapses.
Mol. Cell. Neurosci. 24, 442–450.
Sperk, G., Schwarzer, C., Tsunashima, K., Fuch, K., Sieghart, W., 1997.
GABAA receptor subunits in the rat hippocampus immunocytochemical
distribution of 13 subunits. Neuroscience 80, 987–1000.
Stell, B.M., Mody, I., 2002. Receptors with different affinities mediate
phasic and tonic GABA(A) conductances in hippocampal neurons.
J. Neurosci. 22, RC223.
Stell, B.M., Brickley, S.G., Tang, C.Y., Farrant, M., Mody, I., 2003.
Neuroactive steroids reduce neuronal excitability by selectively enhanc-
ing tonic inhibition mediated by delta subunit-containing GABAA
receptors. Proc. Natl. Acad. Sci. U. S. A. 100, 14439–14444.
Studler, B., Sidler, C., Fritschy, J.M., 2005. Differential regulation of
GABA(A) receptor and gephyrin postsynaptic clustering in
immature hippocampal neuronal cultures. J. Comp. Neurol. 484,
344–355.
Sur, C., Quirk, K., Dewar, D., Atack, J., McKernan, R., 1998. Rat and
human hippocampal alpha5 subunit-containing gamma-aminobutyric
AcidA receptors have alpha5 beta3 gamma2 pharmacological character-
istics. Mol. Pharmacol. 54, 928–933.
Triller, A., Choquet, D., 2003. Synaptic structure and diffusion dynamics of
synaptic receptors. Biol. Cell. 95, 465–476.
Triller, A., Choquet, D., 2005. Surface trafficking of receptors between
synaptic and extrasynaptic membranes: and yet they do move!. Trends
Neurosci. 28, 133–139.
Wei, W., Zhang, N., Peng, Z., Houser, C.R., Mody, I., 2003. Perisynaptic
localization of delta subunit-containing GABA(A) receptors and their
activation by GABA spillover in the mouse dentate gyrus. J. Neurosci.
23, 10650–10661.
Whiting, P., McKernan, R.M., Iversen, L.L., 1990. Another mechanism for
creating diversity in gamma-aminobutyrate type A receptors: RNA
splicing directs expression of two forms of gamma 2 phosphorylation
site. Proc. Natl. Acad. Sci. U. S. A. 87, 9966–9970.
Williams, B.M., Temburni, M.K., Levey, M.S., Bertrand, S., Bertrand, D.,
Jacob, M.H., 1998. The long internal loop of the alpha 3 subunit targets
nAChRs to subdomains within individual synapses on neurons in vivo.
Nat. Neurosci. 1, 557–562.
Wisden, W., Laurie, D.J., Monyer, H., Seeburg, P.H., 1992. The
distribution of 13 GABAA receptor subunit mRNAs in the rat
brain: I. Telencephalon, diencephalon, mesencephalon. J. Neurosci.
12, 1040–1062.
Yeung, J.Y., Canning, K.J., Zhu, G., Pennefather, P., MacDonald, J.F.,
Orser, B.A., 2003. Tonically activated GABAA receptors in hippo-
campal neurons are high-affinity, low-conductance sensors for extrac-
ellular GABA. Mol. Pharmacol. 63, 2–8.
Related Documents





![A - Benzodiazepine-Chloride Receptor-Targeted Therapy for ......nisms through GABAA and GABAB receptors [12]. GABA is classified into two main categories: GABAA and GABAB. GABAA and](https://static.cupdf.com/doc/110x72/60f82a0e0bab2d34196b5ccd/a-benzodiazepine-chloride-receptor-targeted-therapy-for-nisms-through.jpg)





