Research Report Neural evidence for impaired action selection in right hemiparetic cerebral palsy M. van Elk a, ⁎ , C. Crajé a, b , M.E.G.V. Beeren a , B. Steenbergen b , H.T. van Schie b , H. Bekkering a a Donders Institute for Brain, Cognition and Behaviour, Radboud University Nijmegen, Nijmegen, The Netherlands b Behavioural Science Institute, Radboud University Nijmegen, Nijmegen, The Netherlands ARTICLE INFO ABSTRACT Article history: Accepted 22 June 2010 Available online 28 June 2010 Recent studies suggest that in addition to low-level motor impairments, individuals with hemiparetic cerebral palsy (HCP) are characterized by anticipatory action planning deficits as well. In the present EEG study we investigated the neural and temporal dynamics of action planning in participants with right-sided HCP (n = 10) and in left-handed control subjects (n = 10). An anticipatory planning task was used in which participants were required to grasp and rotate a hexagonal knob over different angles (60°, 120° or 180°). At a behavioral level, participants with HCP were slower in their movements and often selected an inappropriate grip when grasping the object. At a neural level, individuals with HCP showed a strong reduction in the amplitude of the P2 component, likely reflecting an impaired process of action selection. In addition, a strong correlation was observed between the P2 amplitude and grasping and rotation times. The P2 component was localized to sources in the dorsal posterior cingulate cortex (dPCC), an area that is known to be involved in orienting visual body parts in space. Together these findings suggest that anticipatory planning deficits in cerebral palsy arise mainly due to an impaired process of action selection. © 2010 Elsevier B.V. All rights reserved. Keywords: Hemiparetic cerebral palsy Motor planning Action selection Event-related potentials P2 1. Introduction Individuals with cerebral palsy (CP) are characterized by non- progressive disorders of movement and posture that are attributed to disturbances in the fetal or infant brain (Bax et al., 2005). Clinical studies have shown large individual variability in terms of the brain areas affected, ranging from lesions in both gray and white matter, brain malformations to no detectable brain abnormalities at all (Korzeniewski et al., 2008; Wu et al., 2006). About 20–33% of the individuals with CP are categorized with hemiparetic CP (HCP), a form of CP affecting the left or right lateral side of the body (Koman et al., 2004; Wuet al., 2006). In addition to low-level problems with motor execution, one recurring finding is that individuals with HCP are characterized by deficits in anticipatory action planning as well (for review, see Steenbergen & Gordon, 2006). These action planning deficits are occurring in both the affected and the (relatively) unaffected arm, and as such these problems have a major impact on activities in daily life. Previous studies have indicated that action planning deficits are especially apparent when the right body side is affected (Craje et al., 2009; Mutsaarts et al., 2007), in line with the proposed role of the left hemisphere in action planning (Haaland & Harrington, 1996; Vingerhoets, 2008). Support for the notion that action planning deficits are a BRAIN RESEARCH 1349 (2010) 56 – 67 ⁎ Corresponding author. Donders Institute for Brain, Cognition and Behaviour, P.O. Box 9104, 6500 HE Nijmegen, The Netherlands. Fax: + 31 24 3616066. E-mail address: [email protected] (M. van Elk). 0006-8993/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2010.06.055 available at www.sciencedirect.com www.elsevier.com/locate/brainres

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Research Report
Neural evidence for impaired action selection in righthemiparetic cerebral palsy
M. van Elka,⁎, C. Crajéa,b, M.E.G.V. Beerena, B. Steenbergenb, H.T. van Schieb, H. Bekkeringa
aDonders Institute for Brain, Cognition and Behaviour, Radboud University Nijmegen, Nijmegen, The NetherlandsbBehavioural Science Institute, Radboud University Nijmegen, Nijmegen, The Netherlands
A R T I C L E I N F O
⁎ Corresponding author. Donders Institute for24 3616066.
E-mail address: [email protected]
0006-8993/$ – see front matter © 2010 Elsevidoi:10.1016/j.brainres.2010.06.055
A B S T R A C T
Article history:Accepted 22 June 2010Available online 28 June 2010
Recent studies suggest that in addition to low-level motor impairments, individuals withhemiparetic cerebral palsy (HCP) are characterized by anticipatory action planning deficitsas well. In the present EEG study we investigated the neural and temporal dynamics ofaction planning in participants with right-sided HCP (n=10) and in left-handed controlsubjects (n=10). An anticipatory planning task was used inwhich participants were requiredto grasp and rotate a hexagonal knob over different angles (60°, 120° or 180°). At a behaviorallevel, participants with HCP were slower in their movements and often selected aninappropriate grip when grasping the object. At a neural level, individuals with HCP showeda strong reduction in the amplitude of the P2 component, likely reflecting an impairedprocess of action selection. In addition, a strong correlation was observed between the P2amplitude and grasping and rotation times. The P2 component was localized to sources inthe dorsal posterior cingulate cortex (dPCC), an area that is known to be involved in orientingvisual body parts in space. Together these findings suggest that anticipatory planningdeficits in cerebral palsy arise mainly due to an impaired process of action selection.
© 2010 Elsevier B.V. All rights reserved.
Keywords:Hemiparetic cerebral palsyMotor planningAction selectionEvent-related potentialsP2
1. Introduction
Individuals with cerebral palsy (CP) are characterized by non-progressive disorders of movement and posture that areattributed to disturbances in the fetal or infant brain (Bax etal., 2005).Clinical studieshaveshownlarge individualvariabilityin terms of thebrain areas affected, ranging from lesions in bothgray and white matter, brain malformations to no detectablebrain abnormalities at all (Korzeniewski et al., 2008; Wu et al.,2006). About 20–33% of the individuals with CP are categorizedwith hemiparetic CP (HCP), a formofCP affecting the left or rightlateral side of the body (Koman et al., 2004; Wuet al., 2006).
Brain, Cognition and Beha
(M. van Elk).
er B.V. All rights reserved
In addition to low-level problems withmotor execution, onerecurring finding is that individuals with HCP are characterizedbydeficits inanticipatoryactionplanningaswell (for review, seeSteenbergen&Gordon, 2006). These action planning deficits areoccurring in both the affected and the (relatively) unaffectedarm, and as such these problems have a major impact onactivities in daily life. Previous studies have indicated thataction planning deficits are especially apparent when the rightbody side is affected (Craje et al., 2009; Mutsaarts et al., 2007), inline with the proposed role of the left hemisphere in actionplanning (Haaland & Harrington, 1996; Vingerhoets, 2008).Support for the notion that action planning deficits are a
viour, P.O. Box 9104, 6500 HE Nijmegen, The Netherlands. Fax: +31
.

57B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
characteristic feature of HCP comes mainly from behavioralstudies. Whereas healthy participants typically grasped objectswith a grip that allows a comfortable posture at the end of amovement (Rosenbaum Vaughan et al., 1992), individuals withHCP often grasped objects with an initial comfortable grip, evenif that resulted in an awkward end posture or task failures(Mutsaarts et al., 2005, 2006; Steenbergen et al., 2004). Inaddition, it has been found that HCP participants failed toanticipate the fingertip forces required for smoothly grasping anobject (Duff & Gordon, 2003). Other studies indicate that peoplewith HCP show less efficient grasping kinematics that – incontrast to healthy controls – are not influenced by later taskdemands (Chen & Yang, 2007; Steenbergen & van der Kamp,2004). Although these studies provide tentative support for thenotion that participants with HCP are characterized by antici-patory planningdeficits, theneural and functionalmechanismsunderlying these impairments remain poorly understood.
An important aspect of action planning involves theselection of the correct motor program (Andersen & Cui,2009). If you are grasping a pen, for instance, depending onyour action intention you need to select a specific grip thatallows you to use the object in a proper way (e.g. writing with apen requires a different grip than moving a pen; cf. Daprati &Sirigu, 2006). Accordingly, in the present study we hypothe-sized that action planning problems in individuals with HCPmay be related to an impaired process of action selection.
To investigate this hypothesis we used an action planningtask in which subjects were required to make a rotatingmovement with their unaffected hand (cf. Mutsaarts et al.,2006). We measured subjects’ EEG while they were preparingactions of varying difficulty (i.e. preparing to rotate a disk over60°, 120° or 180°) and effects of task difficulty were explored bymeasuring event-related potentials (ERPs). Due to their hightemporal resolution and the limited restrictions on subjects’hand and arm movements, ERPs provide the opportunity tocapture the time-course of visuo-motor processing.
Based on our interest in early action planning, ERP analysisfocused on the action preparation interval and more specif-ically on the anterior P2 (also labeled P2a or P3f; Makeig et al.,1999; Potts, 2004), which is a positive deflection over fronto-central sites with a peak latency of about 200 ms after theonset of a stimulus. Typically a larger P2 amplitude is observedfor stimuli with a to-be-attended feature (Kenemans et al.,1993; Potts, 2004; Smid et al., 1999) and accordingly the P2 hasbeen associated with the evaluation of task-relevant stimuli.In addition the P2 amplitude is larger for trials that require anovert response (Gajewski et al., 2008; Makeig et al., 1999; Pottset al., 1996) and the P2 has been associated with theanticipation of action consequences as well (Nikolaev et al.,2008). Interestingly, in a recent study we found a stronger P2component (as reflected in a frontal selection positivity; FSP)when subjects were required to grasp compared to when theyhad to point towards a 3D target object (van Elk et al., in press).This task-specific modulation of the P2 was only found whensubjects were required to actually perform themovement, butnot when they withheld from a response. In sum, thesestudies suggest that the P2 reflects an action selectionmechanism, enabling the coupling of relevant visual infor-mation to specific responses (Kuhn et al., 2009; Smid et al.,1999).
In the present study, it was hypothesized that motorplanning deficits in participants with HCP should becomeapparent in slower reaction and movement times, lessaccurate rotations and more awkward grips compared tocontrol subjects. Based on previous studies (Smid et al., 1999;van Elk et al., in press), we expected that the planning of morecomplex actions (i.e. rotating a disc over a larger angle) mightbe accompanied by a stronger anterior P2 component. Inaddition, differences in action planning between individualswith HCP and control participants may be reflected in asmaller amplitude of the anterior P2 for HCP compared tocontrol participants.
2. Results
2.1. Behavioral results
TheRTanalyses revealedamaineffect of rotation,F(2, 17)=6.5,p<.01, η2 =.43, reflecting that RTs increased with increasedrotation angles (see Fig. 2). There was a marginal trend fordirection, F(1, 18)=3.7, p =.07, η2 =.17, suggesting slower RTs forcounter-clockwise rotations (777 ms, SE=119 ms) than forclockwise rotations (738 ms, SE=104 ms; see Fig. 1). No effectof group was found (F<1).
The analyses of grasping times showed a main effect ofrotation, F(2,17)=9.1, p <.005, η2 =.52, reflecting slower graspingtimeswith increased rotation angles (see Fig. 1). Amain effect ofdirection, F(1,18)=4.7, p <.05, η2 =.21, reflected slower graspingtimes for counter-clockwise (2077ms, SE=190 ms) compared toclockwise rotations (1925 ms, SE=188 ms; see Fig. 1). A margin-ally significant effect of group, F(1,18)=4.2, p =.056, η2 =.19,indicated slower grasping times for HCP subjects (2381 ms,SE=263 ms) than for control subjects (1621 ms, SE=263 ms; seeFig. 1).
As expected, the analyses of rotation times showed a maineffect of rotation, F(2, 17)=9.8, p <.001, η2 =.54, reflecting longerrotation timeswith increased rotations (see Fig. 1). Amaineffectof direction, F(1,18)=12.2, p <.005, η2 =.40, reflected slowerrotation times for counter-clockwise (3471ms, SE=196) com-pared to clockwise rotations (3198ms, SE=176 ms; see Fig. 1). Amain effect of group, F(1,18)=6.6, p <.05, η2 =.27, reflected slowerrotation times for HCP subjects (3801 ms, SE=258 ms) than forcontrol subjects (2869ms, SE=257 ms; see Fig. 1).
With respect to rotation errors (difference betweeninstructed and realized rotation angle), a main effect ofrotation, F(2,17)=4.4, p <.05, η2 =.34, reflected larger rotationerrors with increased rotations (see Fig. 1). No effect of groupwas found, F(1,18)=2.2, p =.15.
In the analysis of the selection of initial grip types forgrasping the hexagon it was found that HCP subjects moreoften grasped the hexagon with an initial comfortable handgrip (average number of selection of Grip 3 per category=6.1)compared to control subjects (average number of selection ofGrip 3 per category=2.5), F(1,18)=4.1, p =.059. In an additionalanalysis based on the coding of anticipatory grips, a margin-ally significant interaction was found between direction androtation, F(2,17)=2.9, p =.08. This interaction reflected that –overall – subjects showed anticipatory grip selection for 180°rotations. Importantly, a significant interaction between
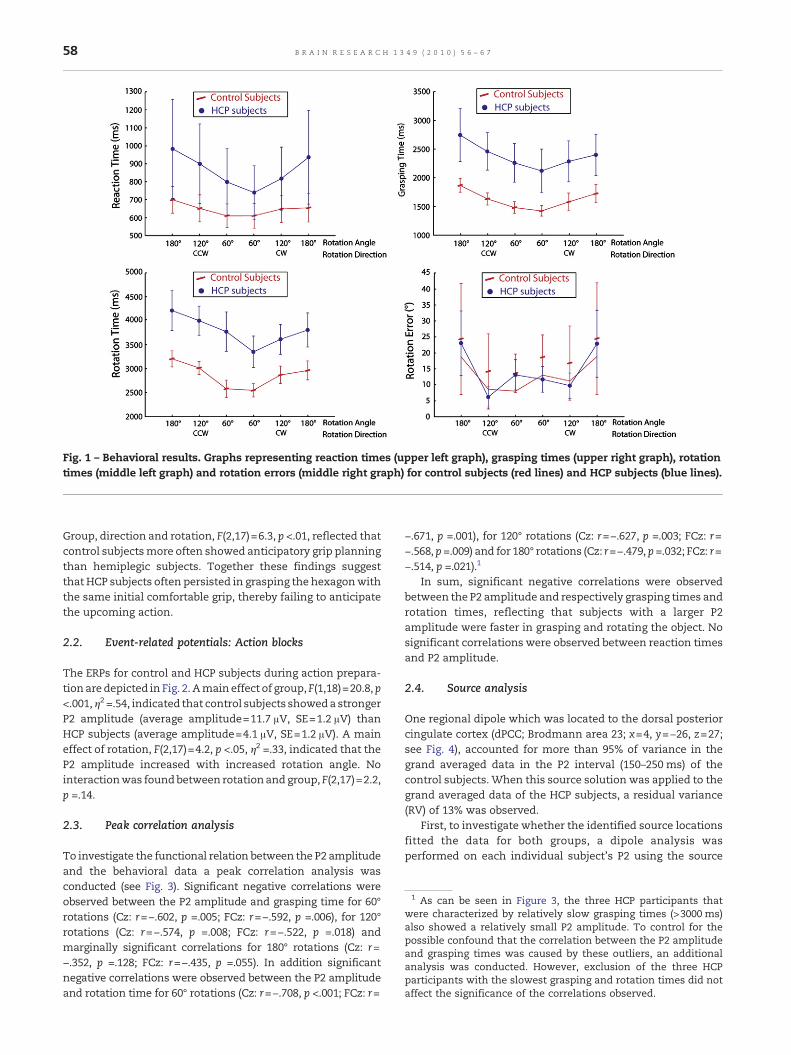
Fig. 1 – Behavioral results. Graphs representing reaction times (upper left graph), grasping times (upper right graph), rotationtimes (middle left graph) and rotation errors (middle right graph) for control subjects (red lines) and HCP subjects (blue lines).
1 As can be seen in Figure 3, the three HCP participants thatwere characterized by relatively slow grasping times (>3000 ms)also showed a relatively small P2 amplitude. To control for thepossible confound that the correlation between the P2 amplitudeand grasping times was caused by these outliers, an additionalanalysis was conducted. However, exclusion of the three HCPparticipants with the slowest grasping and rotation times did notaffect the significance of the correlations observed.
58 B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
Group, direction and rotation, F(2,17)=6.3, p <.01, reflected thatcontrol subjectsmore often showed anticipatory grip planningthan hemiplegic subjects. Together these findings suggestthat HCP subjects often persisted in grasping the hexagonwiththe same initial comfortable grip, thereby failing to anticipatethe upcoming action.
2.2. Event-related potentials: Action blocks
The ERPs for control and HCP subjects during action prepara-tion are depicted in Fig. 2. Amain effect of group, F(1,18)=20.8, p<.001, η2 =.54, indicated that control subjects showeda strongerP2 amplitude (average amplitude=11.7 μV, SE=1.2 μV) thanHCP subjects (average amplitude=4.1 μV, SE=1.2 μV). A maineffect of rotation, F(2,17)=4.2, p <.05, η2 =.33, indicated that theP2 amplitude increased with increased rotation angle. Nointeractionwas foundbetween rotationand group, F(2,17)=2.2,p =.14.
2.3. Peak correlation analysis
To investigate the functional relation between the P2 amplitudeand the behavioral data a peak correlation analysis wasconducted (see Fig. 3). Significant negative correlations wereobserved between the P2 amplitude and grasping time for 60°rotations (Cz: r=−.602, p =.005; FCz: r=−.592, p =.006), for 120°rotations (Cz: r=−.574, p =.008; FCz: r=−.522, p =.018) andmarginally significant correlations for 180° rotations (Cz: r=−.352, p =.128; FCz: r=−.435, p =.055). In addition significantnegative correlations were observed between the P2 amplitudeand rotation time for 60° rotations (Cz: r=−.708, p <.001; FCz: r=
−.671, p =.001), for 120° rotations (Cz: r=−.627, p =.003; FCz: r=−.568, p =.009) and for 180° rotations (Cz: r=−.479, p =.032; FCz: r=−.514, p =.021).1
In sum, significant negative correlations were observedbetween the P2 amplitude and respectively grasping times androtation times, reflecting that subjects with a larger P2amplitude were faster in grasping and rotating the object. Nosignificant correlations were observed between reaction timesand P2 amplitude.
2.4. Source analysis
One regional dipole which was located to the dorsal posteriorcingulate cortex (dPCC; Brodmann area 23; x=4, y=−26, z=27;see Fig. 4), accounted for more than 95% of variance in thegrand averaged data in the P2 interval (150–250 ms) of thecontrol subjects. When this source solution was applied to thegrand averaged data of the HCP subjects, a residual variance(RV) of 13% was observed.
First, to investigate whether the identified source locationsfitted the data for both groups, a dipole analysis wasperformed on each individual subject's P2 using the source
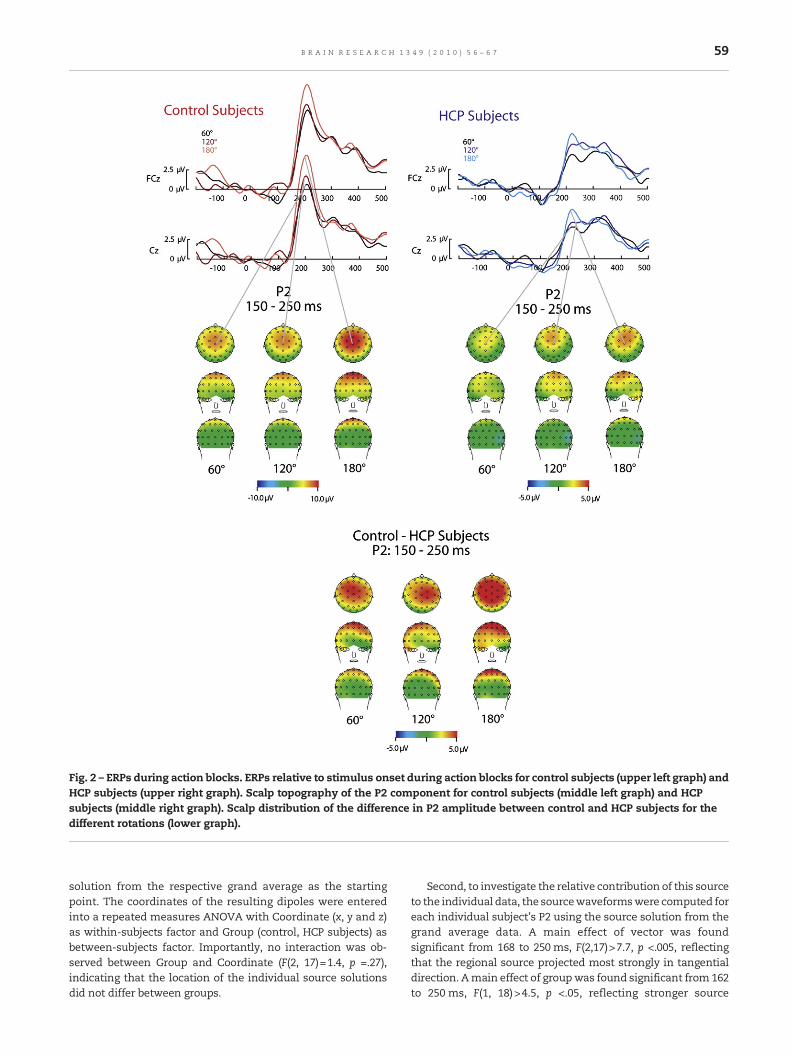
Fig. 2 – ERPs during action blocks. ERPs relative to stimulus onset during action blocks for control subjects (upper left graph) andHCP subjects (upper right graph). Scalp topography of the P2 component for control subjects (middle left graph) and HCPsubjects (middle right graph). Scalp distribution of the difference in P2 amplitude between control and HCP subjects for thedifferent rotations (lower graph).
59B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
solution from the respective grand average as the startingpoint. The coordinates of the resulting dipoles were enteredinto a repeated measures ANOVA with Coordinate (x, y and z)as within-subjects factor and Group (control, HCP subjects) asbetween-subjects factor. Importantly, no interaction was ob-served between Group and Coordinate (F(2, 17)=1.4, p =.27),indicating that the location of the individual source solutionsdid not differ between groups.
Second, to investigate the relative contribution of this sourceto the individual data, the sourcewaveformswere computed foreach individual subject's P2 using the source solution from thegrand average data. A main effect of vector was foundsignificant from 168 to 250 ms, F(2,17)>7.7, p <.005, reflectingthat the regional source projected most strongly in tangentialdirection. Amain effect of groupwas found significant from 162to 250 ms, F(1, 18)>4.5, p <.05, reflecting stronger source
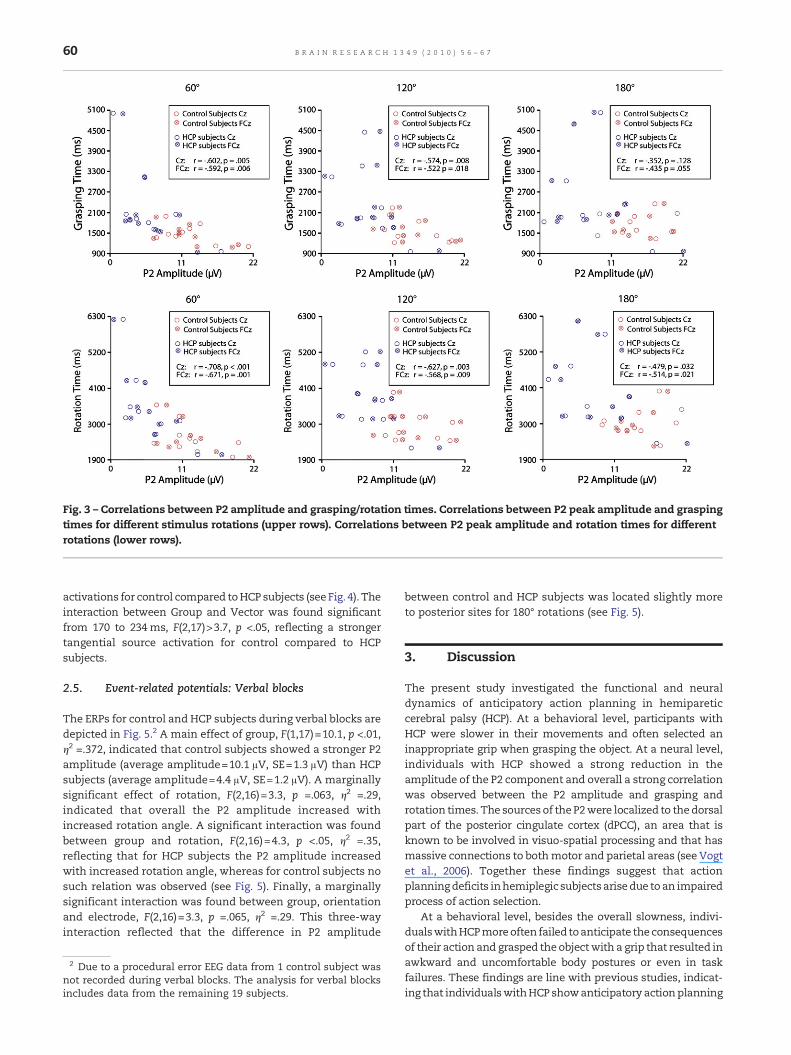
Fig. 3 – Correlations between P2 amplitude and grasping/rotation times. Correlations between P2 peak amplitude and graspingtimes for different stimulus rotations (upper rows). Correlations between P2 peak amplitude and rotation times for differentrotations (lower rows).
60 B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
activations for control compared toHCP subjects (see Fig. 4). Theinteraction between Group and Vector was found significantfrom 170 to 234ms, F(2,17)>3.7, p <.05, reflecting a strongertangential source activation for control compared to HCPsubjects.
2.5. Event-related potentials: Verbal blocks
The ERPs for control and HCP subjects during verbal blocks aredepicted in Fig. 5.2 A main effect of group, F(1,17)=10.1, p <.01,η2 =.372, indicated that control subjects showed a stronger P2amplitude (average amplitude=10.1 μV, SE=1.3 μV) than HCPsubjects (average amplitude=4.4 μV, SE=1.2 μV). A marginallysignificant effect of rotation, F(2,16)=3.3, p =.063, η2 =.29,indicated that overall the P2 amplitude increased withincreased rotation angle. A significant interaction was foundbetween group and rotation, F(2,16)=4.3, p <.05, η2 =.35,reflecting that for HCP subjects the P2 amplitude increasedwith increased rotation angle, whereas for control subjects nosuch relation was observed (see Fig. 5). Finally, a marginallysignificant interaction was found between group, orientationand electrode, F(2,16)=3.3, p =.065, η2 =.29. This three-wayinteraction reflected that the difference in P2 amplitude
2 Due to a procedural error EEG data from 1 control subject wasnot recorded during verbal blocks. The analysis for verbal blocksincludes data from the remaining 19 subjects.
between control and HCP subjects was located slightly moreto posterior sites for 180° rotations (see Fig. 5).
3. Discussion
The present study investigated the functional and neuraldynamics of anticipatory action planning in hemipareticcerebral palsy (HCP). At a behavioral level, participants withHCP were slower in their movements and often selected aninappropriate grip when grasping the object. At a neural level,individuals with HCP showed a strong reduction in theamplitude of the P2 component and overall a strong correlationwas observed between the P2 amplitude and grasping androtation times. The sourcesof the P2were localized to thedorsalpart of the posterior cingulate cortex (dPCC), an area that isknown to be involved in visuo-spatial processing and that hasmassive connections to bothmotor and parietal areas (see Vogtet al., 2006). Together these findings suggest that actionplanningdeficits inhemiplegic subjectsarisedue to an impairedprocess of action selection.
At a behavioral level, besides the overall slowness, indivi-dualswithHCPmoreoften failed toanticipate theconsequencesof their action and grasped the objectwith a grip that resulted inawkward and uncomfortable body postures or even in taskfailures. These findings are line with previous studies, indicat-ing that individualswithHCP showanticipatory actionplanning
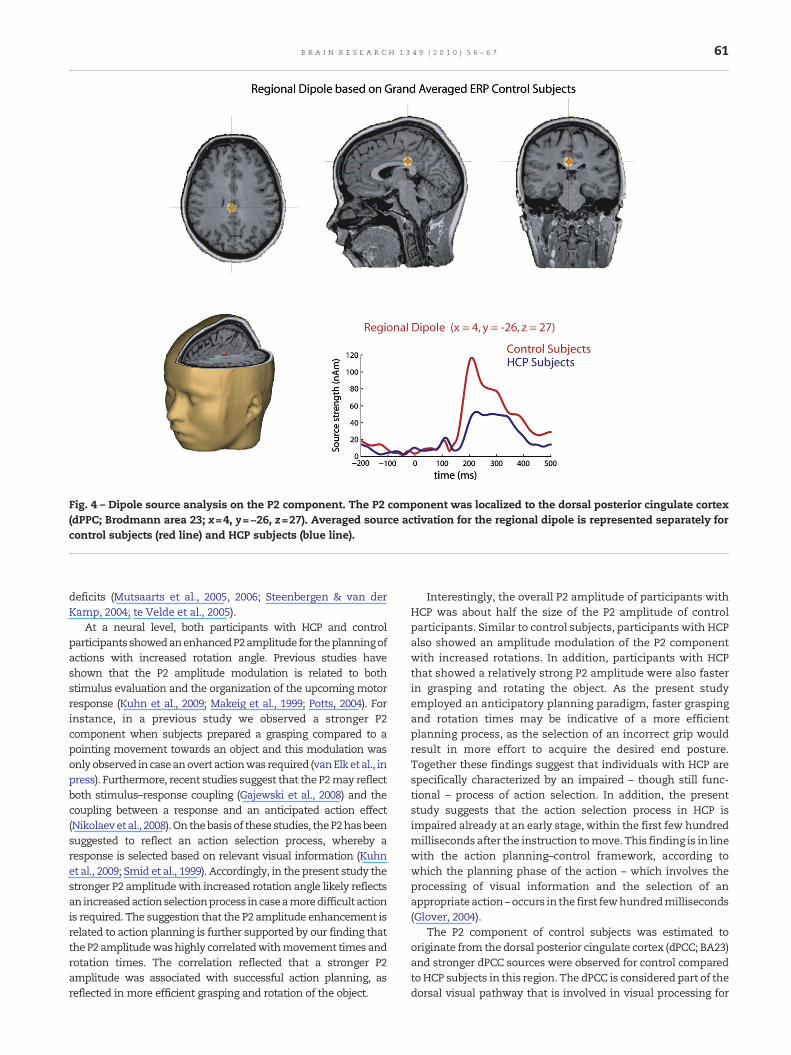
Fig. 4 – Dipole source analysis on the P2 component. The P2 component was localized to the dorsal posterior cingulate cortex(dPPC; Brodmann area 23; x=4, y=−26, z=27). Averaged source activation for the regional dipole is represented separately forcontrol subjects (red line) and HCP subjects (blue line).
61B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
deficits (Mutsaarts et al., 2005, 2006; Steenbergen & van derKamp, 2004; te Velde et al., 2005).
At a neural level, both participants with HCP and controlparticipantsshowedanenhancedP2amplitude for theplanningofactions with increased rotation angle. Previous studies haveshown that the P2 amplitude modulation is related to bothstimulus evaluation and the organization of the upcoming motorresponse (Kuhn et al., 2009; Makeig et al., 1999; Potts, 2004). Forinstance, in a previous study we observed a stronger P2component when subjects prepared a grasping compared to apointing movement towards an object and this modulation wasonlyobserved incaseanovertactionwasrequired (vanElketal., inpress). Furthermore, recent studies suggest that the P2may reflectboth stimulus–response coupling (Gajewski et al., 2008) and thecoupling between a response and an anticipated action effect(Nikolaevetal., 2008).On thebasisof thesestudies, theP2hasbeensuggested to reflect an action selection process, whereby aresponse is selected based on relevant visual information (Kuhnet al., 2009; Smid et al., 1999). Accordingly, in the present study thestronger P2 amplitudewith increased rotation angle likely reflectsan increasedactionselectionprocess incaseamoredifficultactionis required. The suggestion that the P2 amplitude enhancement isrelated to action planning is further supported by our finding thattheP2 amplitudewashighly correlatedwithmovement timesandrotation times. The correlation reflected that a stronger P2amplitude was associated with successful action planning, asreflected in more efficient grasping and rotation of the object.
Interestingly, the overall P2 amplitude of participants withHCP was about half the size of the P2 amplitude of controlparticipants. Similar to control subjects, participants with HCPalso showed an amplitude modulation of the P2 componentwith increased rotations. In addition, participants with HCPthat showed a relatively strong P2 amplitude were also fasterin grasping and rotating the object. As the present studyemployed an anticipatory planning paradigm, faster graspingand rotation times may be indicative of a more efficientplanning process, as the selection of an incorrect grip wouldresult in more effort to acquire the desired end posture.Together these findings suggest that individuals with HCP arespecifically characterized by an impaired – though still func-tional – process of action selection. In addition, the presentstudy suggests that the action selection process in HCP isimpaired already at an early stage, within the first few hundredmilliseconds after the instruction tomove. This finding is in linewith the action planning–control framework, according towhich the planning phase of the action – which involves theprocessing of visual information and the selection of anappropriateaction –occurs in the first fewhundredmilliseconds(Glover, 2004).
The P2 component of control subjects was estimated tooriginate from the dorsal posterior cingulate cortex (dPCC; BA23)and stronger dPCC sources were observed for control comparedtoHCP subjects in this region. The dPCC is considered part of thedorsal visual pathway that is involved in visual processing for
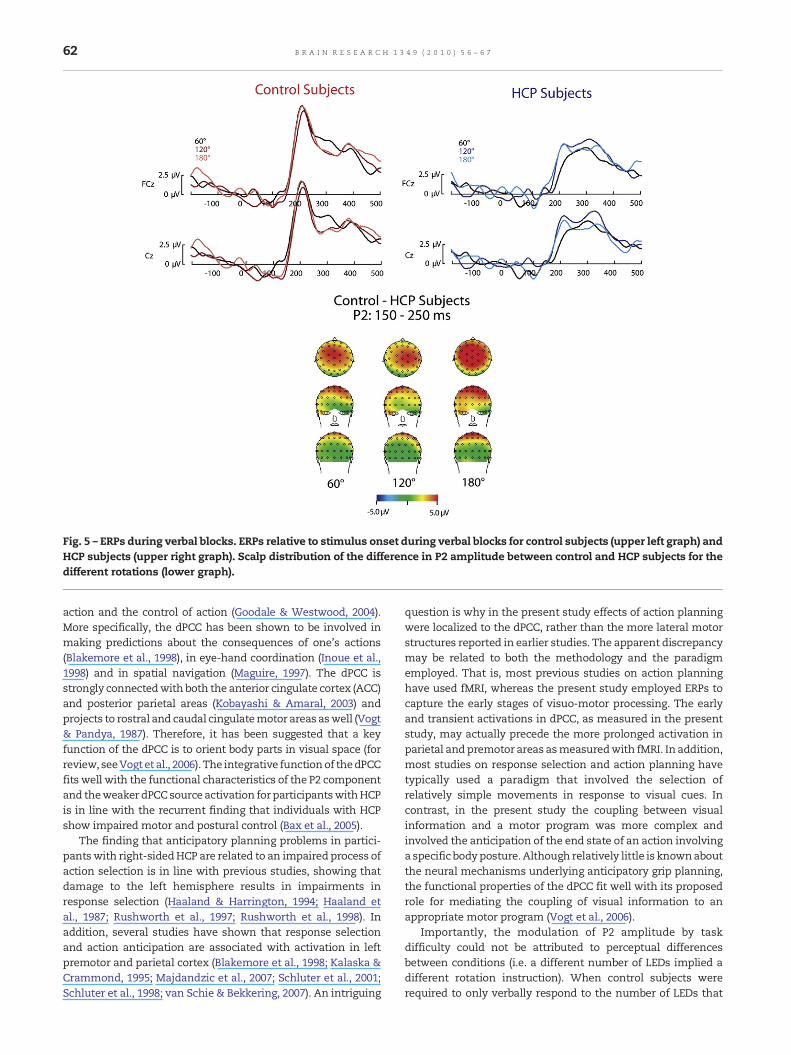
Fig. 5 – ERPs during verbal blocks. ERPs relative to stimulus onset during verbal blocks for control subjects (upper left graph) andHCP subjects (upper right graph). Scalp distribution of the difference in P2 amplitude between control and HCP subjects for thedifferent rotations (lower graph).
62 B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
action and the control of action (Goodale & Westwood, 2004).More specifically, the dPCC has been shown to be involved inmaking predictions about the consequences of one's actions(Blakemore et al., 1998), in eye-hand coordination (Inoue et al.,1998) and in spatial navigation (Maguire, 1997). The dPCC isstrongly connectedwith both the anterior cingulate cortex (ACC)and posterior parietal areas (Kobayashi & Amaral, 2003) andprojects to rostral and caudal cingulatemotor areas aswell (Vogt& Pandya, 1987). Therefore, it has been suggested that a keyfunction of the dPCC is to orient body parts in visual space (forreview, seeVogt et al., 2006).The integrative functionof thedPCCfits well with the functional characteristics of the P2 componentand theweaker dPCCsource activation forparticipantswithHCPis in line with the recurrent finding that individuals with HCPshow impaired motor and postural control (Bax et al., 2005).
The finding that anticipatory planning problems in partici-pantswith right-sidedHCP are related to an impaired process ofaction selection is in line with previous studies, showing thatdamage to the left hemisphere results in impairments inresponse selection (Haaland & Harrington, 1994; Haaland etal., 1987; Rushworth et al., 1997; Rushworth et al., 1998). Inaddition, several studies have shown that response selectionand action anticipation are associated with activation in leftpremotor and parietal cortex (Blakemore et al., 1998; Kalaska &Crammond, 1995; Majdandzic et al., 2007; Schluter et al., 2001;Schluter et al., 1998; van Schie & Bekkering, 2007). An intriguing
question is why in the present study effects of action planningwere localized to the dPCC, rather than the more lateral motorstructures reported in earlier studies. The apparent discrepancymay be related to both the methodology and the paradigmemployed. That is, most previous studies on action planninghave used fMRI, whereas the present study employed ERPs tocapture the early stages of visuo-motor processing. The earlyand transient activations in dPCC, as measured in the presentstudy, may actually precede the more prolonged activation inparietal and premotor areas asmeasuredwith fMRI. In addition,most studies on response selection and action planning havetypically used a paradigm that involved the selection ofrelatively simple movements in response to visual cues. Incontrast, in the present study the coupling between visualinformation and a motor program was more complex andinvolved the anticipation of the end state of an action involvinga specific bodyposture.Although relatively little is knownaboutthe neural mechanisms underlying anticipatory grip planning,the functional properties of the dPCC fit well with its proposedrole for mediating the coupling of visual information to anappropriate motor program (Vogt et al., 2006).
Importantly, the modulation of P2 amplitude by taskdifficulty could not be attributed to perceptual differencesbetween conditions (i.e. a different number of LEDs implied adifferent rotation instruction). When control subjects wererequired to only verbally respond to the number of LEDs that

63B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
was lighted, no modulation of the P2 amplitude was observed.This finding further supports the interpretation that the P2-effect reflects a selection-for-action mechanism, related toresponse selection on the basis of visual information (Smid etal., 1999; van Elk et al., in press). Although for participants withHCP a modulation of the P2 component was observed in theverbal task, the effect appeared to be weaker and did not showthe same consistent pattern as observed in the action planningtask. It could well be that the effects observed in the visual taskfor individuals with HCP reflected the persistent activation ofthe stimulus–response coupling required for the action plan-ning task. The notion that individuals with HCP may haveparticular difficulty with controlling and flexibly switchingbetween different stimulus–response couplings opens interest-ing avenues for future research.
An important issue in research with clinical populationsconcerns the question whether left-handed control partici-pants comprise a valid comparison for participants with right-sidedHCP. It could be thatmost HCP participants had a geneticpredisposition to become right-handed prior to their prenatalbrain injuries. As the relation between handedness and thestructural and functional organization of the brain is wellestablished (Kloppel et al., 2007; Knecht et al., 2000; Willems &Hagoort, 2009; Willems et al., 2009a; Willems et al., 2009b), thecomparison between HCP and left-handed control partici-pants may have been confounded. However, although hand-edness has a strong genetic component, hand preference isdetermined by environmental factors as well (for review, seeLlaurens et al., 2009). From their birth onwards participantswith HCP predominantly used their unaffected left hand,thereby providing a good match with left-handed controlparticipants in terms of action experience. In addition, in arecent study it was found that adult ‘converted’ left-handers –who had been forced as children to write with their righthand – showed a comparable left-hemispheric lateralizationof the central sulcus as right-handers, thereby underlining theplasticity of themotor system caused by forced use of the non-dominant hand (Kloppel et al., 2010). In sum, given theequivalence in action experience and the supposed effect ofboth forced and spontaneous left-hand use on hemisphericspecialization, the comparison between left-handed controlparticipants and subjects with right HCP seems warranted.
Table 1 – Participant information for participants withHCP. Gen=gender; IH=impaired hand; UH=unimpairedhand; Ratio=score (impaired hand)/score (unimpairedhand).
Part Gen Age Box and Blocks Purdue Pegboard
IH UH Ratio IH UH Ratio
1 M 18.8 69 20 .29 28 4 .142 F 19.3 76 11 .15 31 0 0
4. Conclusion
The main finding of the present study is that anticipatoryplanning deficits in subjects with right hemiparetic cerebralpalsy (HCP) arise due to impairments in action selection,occurring at an early stage during action preparation, asevidenced by a reduced P2 amplitude and a weaker sourceactivation of the dorsal posterior cingulate cortex.
3 M 15.7 47 56 1.19 22 21 .964 M 17.5 63 9 .14 20 0 .05 M 18.1 49 33 .67 23 2 .096 M 20.1 49 26 .53 30 0 07 F 18.4 47 18 .38 32 0 .08 M 21.1 57 60 1.05 22 23 1.059 M 17.8 56 60 1.07 27 20 .7410 F 19.3 49 16 .33 28 0 .0
5. Experimental procedures
5.1. Subjects
In total 20 subjects participated in the study. The groupconsisted of 10 participants diagnosed with right spastic
hemiparetic cerebral palsy (HCP; 3 females, mean age=18.3 -years, SD=1.2 years) and 10 neurologically healthy controlparticipants (2 males, mean age=19.7 years, SD=2.2 years).Participants with HCP were recruited via their school (a schoolfor special educationMariëndael in Arnhem, theNetherlands),or via the Dutch society of parents of physically disabledchildren (‘BOSK’). Hand function was tested using the Box andBlocks test (gross dexterity; Mathiowetz et al., 1985) and thePurdue Pegboard test (fine dexterity; Tiffin, 1985) for both theimpaired and unimpaired hand. The severity of the paresiswas estimated by calculating the ratio between the score ofthe impaired and the unimpaired hand. Accordingly, a scorenear 1 indicates that hand function among both hands wascomparable (i.e. mild paresis), whereas a score near 0 indicatesa strong difference between the impaired and unimpairedhand (i.e. severe paresis). Participant information is providedin Table 1.
Control subjects were students from the Radboud Univer-sity Nijmegen, who participated for course credits or anexperimental fee. All control participants were left-handed,as assessed by an online inventory for handedness. Allparticipants gave informed consent prior to the experiment.The study was approved by the local ethics committee, inaccordance with the declaration of Helsinki.
5.2. Experimental setup
The experiment was conducted in an electrically and sound-shielded room. The experimental setup is schematicallypresented in Fig. 6. Subjects were seated comfortably behinda table on which a custom-made rotatable disk (width 40 cm)was placed. A hexagonal knob (width 11 cm and depth 6 cm)was placed at the center of the disk to allow and detectgrasping actions of the subject. At each of the six sides of thehexagon an arrow was inserted (length 15 cm and width0.5 cm). At 0.5 cm from the edge of the disk six LEDs wereplaced at 0°, 60°, 120°, 180°, 240° and 300°. In front of the disk aresponse box was placed from which the subject started thegraspingmovements. Both the response box and the rotatabledisk were controlled by a PC running Presentation software(Neurobehavioral Systems Inc., Albany, CA) to switch on the

Fig. 6 – Experimental setup. Top: Subjects were seated behind a table on which a custom-made disk was placed that could begrasped at the middle hexagon. LEDs on the side of the disk instructed subjects to either rotate the disk in a particularorientation (action blocks) or to report the number of LEDs (verbal blocks). Bottom: Subjects were allowed to freely select theirinitial handgrip for grasping the hexagon (Grip 1–5; grip 6 was biomechanically impossible).
64 B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
LEDs, to detect release of the button box (reaction time), thegrasping of the hexagon (grasping time), the release of thehexagon (rotation time) and to measure the rotation angle.Markers for the different eventswere sent to the EEG computerand stored for offline analysis.
5.3. Experimental procedure
At the beginning of each trial a number of LEDs turned on anddepending on the task instructions subjects responded byrotating the disk (action blocks) or by verbally reporting thenumber of LEDs (verbal blocks). Verbal blocks were included tocontrol for perceptual differences between conditions (i.e.more visual information projected onto the retina when alarge rotation is required). During action blocks a combinationof LEDs instructed the subject to rotate the disk with a specificangle in a specific direction (see Fig. 6). For instance, if duringan action block LEDs 1, 2 and 3were lighted, this indicated thatthe subject was required to rotate the disk 120° in a clockwisedirection. During verbal blocks a combination of LEDsinstructed the subject to report the number of LEDs thatwere lighted, after the LEDs switched off. For instance, ifduring a verbal block LEDs 1, 2 and 3 were lighted, the subjectwas required to verbally report the number of LEDs (i.e. ‘three’)after the LEDs had turned off.
In line with previous studies using participants withunilateral brain damage (Kimura & Archibald, 1974; Mutsaartset al., 2005; Rushworth et al., 1998; Steenbergen et al., 2004;Steenbergen & van der Kamp, 2004), during action blocksparticipants always performed the movements with their non-
affected/left hand (i.e. the hand ipsilateral to the affectedhemisphere). The rationale for having participants performthe action with the left hand was that most hemiplegicparticipants could correctly perform the grasping and rotationactions only with their ipsilateral hand. This manipulationallowed us to directly assess effects of action planning, withoutthe possible confound of physical limitations in motor abilitiesbetween participants.
Subjects were instructed to grasp the disk with the thumband fingers placed on opposite sides of the hexagon. At thebeginning of the experiment it was explained that the objectcould be grasped with 5 different grip types (see Grips 1–5 inFig. 6; Grip 6 was biomechanically impossible). During theexperiment subjects were free to choose by which grip theygrasped the object in order to rotate it. After the LEDs switchedon, subjects released the starting position (i.e., they releasedthe response button on the button box), grasped the hexagonand rotated the disk according to the task instructionsprovided by the LEDs. The initial grip by which subjectsgrasped the object was coded online by an experimenter. Afterrotation, subjects returned to the starting position uponwhichthe next trial was initiated.
During verbal blocks, the LEDs remained on for 1000–1500 ms after which the subjects verbally reported thenumber of lighted LEDs to the experimenter. After verballyresponding, subjects pressed the response button uponwhich the next trial was initiated. Each block consisted of 30trials (2 Directions × 3 Rotation angles × 5 Replications) whichwere presented in random order. Each subject performed threeaction blocks and three verbal blocks and block order was

65B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
counterbalanced between subjects (block order wasAVAVAVorVAVAVA).
5.4. EEG measurements
The electroencephalogram (EEG) was recorded using 61active electrodes that were placed in an actiCAP (BrainPro-ducts, Munich, Germany). Electrode positions were based onthe M-10 Equidistant 61-Channel-Arrangement, with aninter-electrode distance of 37±3 mm (given a head circum-ference of 58 cm). All electrodes were referenced to the leftmastoid online and re-referenced offline to the linkedmastoids. The impedance of the electrodes was kept below10 kOhm. EEG and EOG signals were amplified using two 32-channel BrainAmp DC EEG amplifiers. The signal wassampled at 500 Hz and filtered online with an 80 Hz highcut-off filter and a 10 s time-constant.
5.5. Data analysis
Analysis of behavioral responses focused on reaction times,grasping times (time between release of the starting buttonand detection of grasping the hexagon) and rotation times(time between detection of grasping the hexagon and releaseof the hexagon). Response latencies exceeding the subject'smean bymore than 3 standard deviations were excluded fromanalysis. In addition the rotation error was calculated bysubtracting the actually rotated angle from the instructedrotation. Behavioral data were analyzed using a 2×3 repeatedmeasures general linear model (GLM) with Direction (counter-clockwise, clockwise) and Rotation (60°, 120°, 180°) as within-subject variables and Group (control subjects, HCP subjects) asbetween-subjects factor.
For the analysis of the selection of initial hand grip, thenumber of initial comfortable handgripswas analyzed, as thisis indicative of a deficit in anticipatory planning. In anadditional analysis, first each grip type was assigned a score(Grip 1=1; Grip 2=2 etc.). Based on the number of gripsselected, for each subject an average score was calculated perstimulus category. Accordingly, a low score is indicative ofanticipatory planning for clockwise movements (CW) and ahigh score is indicative of anticipatory planning for counter-clockwise movements. The individual scores were analyzedusing a 2×3 repeated measures general linear model (GLM)with Direction (counter-clockwise, clockwise) and Rotation(60°, 120°, 180°) aswithin-subject variables andGroup (controlsubjects, hemiparetic subjects) as between-subjects factor.Detailed analysis of the selection of the initial grip type isreported elsewhere (Crajé et al., submitted).
Analysis of event-related potentials (ERPs) focused on themovement preparation interval and ERPs were calculatedrelative to the onset of the LEDs from −200 to 500 ms using a100 ms pre-stimulus baseline. Trialswithmovement artifactswere excluded from analysis on the basis of careful visualinspection of the raw data. Ocular artifacts were correctedusing a semi-automatic correction procedure based on thealgorithm of Gratton and Coles (1983). The analyses wereconducted on the averaged P2 amplitude at Cz and FCz from180to 220 ms after stimulus onset. ERPs were tested forstatistical significance using a 3×2 repeated measures
general linear model (GLM) with Rotation (60°, 120°, 180°)and Electrode (Cz, FCz) as within-subject variables and Group(control subjects, HCP subjects) as between-subjects factor.
In a subsequent correlation analysis the functional relationbetween the ERPs and the behavioral measures was investigat-ed. First, for each individual subject thepeak amplitudeof theP2component was identified, by selecting the most positive peakbetween 150 ms and 250 ms at electrodes Cz and FCz. NextPearson's R was calculated between the individual peakamplitudes of the P2 component and grasping times androtation times. Correlations were calculated separately fordifferent rotations (60°, 120° and 180°).
5.6. Source analysis
Brain electromagnetic source analysis (BESA 5.0, MEGIS Soft-ware) was used for EEG dipole source analysis, by using a four-shell ellipsoid head model. For the source analysis of theaveraged P2 component in the action condition a single regionaldipolewasused, in linewithpreviousstudies thathavemodeledsources of fronto-central components (van Schie et al., 2004).The source model was computed from 150 to 250 ms afterstimulusonseton thegrandaveragewaveform, averagedacrossdifferent stimulus conditions to increase the signal-to-noiseratio. For the control subjects, the best fitting source model onthegrandaveragepotential distributionwascomputed (residualvariance (RV) was below 5%). The source solution for the controlsubjects was applied to the grand average potential distributionof the hemiparetic subjects.
First, to investigate whether the identified source locationdiffered between groups, a dipole analysis was performed oneach individual subject's P2 using the source solution from therespective grand average as the starting point. The x, y and zlocation coordinates for the individual dipole solutions wereentered in a repeatedmeasures ANOVAwith Coordinate (x, y, z)as within-subjects factor and Group (control, hemipareticsubjects) as between-subjects factor. Source locations werespecified in Talaraich coordinates and were projected on anindividual MRI.
Subsequently, to investigate whether the identified sourcediffered in strength betweenboth groups, the sourcewaveformswere computed for each individual subject's P2,using the sourcesolution from the grand average. To investigate group differ-ences between the source waveforms a sample-wise repeatedmeasures ANOVA was conducted on the 150–250ms interval,with Vector field (x, y and z) as within-subject factor and Group(control, HCP subjects) as between-subjects factor. The signif-icance criterion for each individual time-bin was set at 0.05. Tocontrol for multiple comparisons, a criterion of three consecu-tive significant intervals was used, thereby reducing p to50×0.053 =.00625.
Acknowledgments
This work was supported by the Netherlands Organization forScientific Research (grants 453-05-001 awarded to H.B. andgrant 400-04-046 awarded to C.C.).

66 B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
R E F E R E N C E S
Andersen, R.A., Cui, H., 2009. Intention, action planning, anddecision making in parietal-frontal circuits. Neuron 63 (5),568–583.
Bax, M., Goldstein, M., Rosenbaum, P., Leviton, A., Paneth, N., Dan,B., et al., 2005. Proposed definition and classification of cerebralpalsy, April 2005. Dev. Med. Child Neurol. 47 (8), 571–576.
Blakemore, S.J., Rees, G., Frith, C.D., 1998. How do we predict theconsequences of our actions? A functional imaging study.Neuropsychologia 36 (6), 521–529.
Chen, Y.P., Yang, T.F., 2007. Effect of task goals on the reachingpatterns of children with cerebral palsy. J. Mot Behav. 39 (4),317–324.
Craje, C., van der Kamp, J., Steenbergen, B., 2009. Visualinformation for action planning in left and right congenitalhemiparesis. Brain Res. 1261, 54–64.
Daprati, E., Sirigu, A., 2006. How we interact with objects: learningfrom brain lesions. Trends Cogn. Sci. 10 (6), 265–270.
Duff, S.V., Gordon, A.M., 2003. Learning of grasp control in childrenwith hemiplegic cerebral palsy. Dev. Med. Child Neurol. 45 (11),746–757.
Gajewski, P.D., Stoerig, P., Falkenstein, M., 2008. ERP–correlates ofresponse selection in a response conflict paradigm. Brain Res.1189, 127–134.
Glover, S., 2004. Separate visual representations in the planningand control of action. Behav. Brain Sci. 27 (1), 3–24 discussion24-78.
Goodale, M.A., Westwood, D.A., 2004. An evolving view of duplexvision: separate but interacting cortical pathways forperception and action. Curr. Opin. Neurobiol. 14 (2), 203–211.
Gratton, G., Coles, M.G., Donchin, E., 1983. A new method foroff-line removal of ocular artifact. Electroen Clin. Neuro 55 (4),468–484.
Haaland, K.Y., Harrington, D.L., 1994. Limb-sequencing deficits afterleft butnot righthemispheredamage. BrainCogn. 24 (1), 104–122.
Haaland, K.Y., Harrington, D.L., 1996. Hemispheric asymmetry ofmovement. Curr. Opin. Neurobiol. 6 (6), 796–800.
Haaland, K.Y., Harrington, D.L., Yeo, R., 1987. The effects of taskcomplexity on motor performance in left and right CVApatients. Neuropsychologia 25 (5), 783–794.
Inoue, K., Kawashima, R., Satoh, K., Kinomura, S., Goto, R.,Koyama, M., et al., 1998. PET study of pointing with visualfeedback of moving hands. J. Neurophysiol. 79 (1), 117–125.
Kalaska, J.F., Crammond, D.J., 1995. Deciding not to GO: neuronalcorrelates of response selection in a GO/NOGO task in primatepremotor and parietal cortex. Cereb. Cortex 5 (5), 410–428.
Kenemans, J.L., Kok, A., Smulders, F.T., 1993. Event-relatedpotentials to conjunctions of spatial frequency and orientationas a function of stimulus parameters and responserequirements. Electroencephalogr. Clin. Neurophysiol. 88 (1),51–63.
Kimura, D., Archibald, Y., 1974. Motor functions of the lefthemisphere. Brain 97 (2), 337–350.
Kloppel, S., van Eimeren, T., Glauche, V., Vongerichten, A.,Munchau, A., Frackowiak, R.S., et al., 2007. The effect ofhandedness on cortical motor activation during simplebilateral movements. Neuroimage 34 (1), 274–280.
Kloppel, S., Mangin, J.F., Vongerichten, A., Frackowiak, R.S.,Siebner, H.R., 2010. Nurture versus nature: long-term impact offorced right-handedness on structure of pericentral cortex andbasal ganglia. J. Neurosci. 30 (9), 3271–3275.
Knecht, S., Drager, B., Deppe, M., Bobe, L., Lohmann, H., Floel, A., etal., 2000. Handedness and hemispheric language dominance inhealthy humans. Brain 123 (Pt 12), 2512–2518.
Kobayashi, Y., Amaral, D.G., 2003. Macaque monkey retrosplenialcortex. II. Cortical afferents. J. Comp. Neurol. 466 (1), 48–79.
Koman, L.A., Smith, B.P., Shilt, J.S., 2004. Cerebral palsy. Lancet 363(9421), 1619–1631.
Korzeniewski, S.J., Birbeck, G., DeLano, M.C., Potchen, M.J., Paneth,N., 2008. A systematic review of neuroimaging for cerebralpalsy. J. Child Neurol. 23 (2), 216–227.
Kuhn, S., Gevers, W., Brass, M., 2009. The neural correlates ofintending not to do something. J. Neurophysiol. 101 (4),1913–1920.
Llaurens, V., Raymond, M., Faurie, C., 2009. Why are some peopleleft-handed? An evolutionary perspective. Philos. Trans. R. Soc.Lond. B Biol. Sci. 364 (1519), 881–894.
Maguire, E.A., 1997. Hippocampal involvement in humantopographical memory: evidence from functional imaging.Philos. Trans. R. Soc. Lond. B Biol. Sci. 352 (1360), 1475–1480.
Majdandzic, J., Grol, M.J., van Schie, H.T., Verhagen, L., Toni, I.,Bekkering, H., 2007. The role of immediate and final goals inaction planning: an fMRI study. Neuroimage 37 (2), 589–598.
Makeig, S., Westerfield, M., Jung, T.P., Covington, J., Townsend, J.,Sejnowski, T.J., et al., 1999. Functionally independentcomponents of the late positive event-related potential duringvisual spatial attention. J. Neurosci. 19 (7), 2665–2680.
Mathiowetz, V., Volland, G., Kashman, N., Weber, K., 1985. Adultnorms for the box and block test for manual dexterity. Amer. J.Occup. Ther. 157, 162–173.
Mutsaarts, M., Steenbergen, B., Bekkering, H., 2005. Anticipatoryplanning of movement sequences in hemiparetic cerebralpalsy. Mot. Control 9 (4), 439–458.
Mutsaarts, M., Steenbergen, B., Bekkering, H., 2006. Anticipatoryplanning deficits and task context effects in hemipareticcerebral palsy. Exp. Brain Res. 172 (2), 151–162.
Mutsaarts, M., Steenbergen, B., Bekkering, H., 2007. Impairedmotor imagery in right hemiparetic cerebral palsy.Neuropsychologia 45 (4), 853–859.
Nikolaev, A.R., Ziessler, M., Dimova, K., van Leeuwen, C., 2008.Anticipated action consequences as a nexus between actionand perception: evidence from event-related potentials. Biol.Psychol. 78 (1), 53–65.
Potts, G.F., 2004. An ERP index of task relevance evaluation ofvisual stimuli. Brain Cogn. 56 (1), 5–13.
Potts, G.F., Liotti, M., Tucker, D.M., Posner, M.I., 1996. Frontal andinferior temporal activity in visual target detection: evidencefrom high spatially sampled event-related potentials. BrainTopogr. 9 (1), 3–14.
Rosenbaum Vaughan, J., Barnes, H.J., Jorgensen, M.J., 1992. Timecourse ofmovement planning: selection of handgrips for objectmanipulation. J. Exp. Psychol LMC 18 (5), 1058–1073.
Rushworth, M.F., Nixon, P.D., Renowden, S., Wade, D.T.,Passingham, R.E., 1997. The left parietal cortex and motorattention. Neuropsychologia 35 (9), 1261–1273.
Rushworth, M.F., Nixon, P.D., Wade, D.T., Renowden, S.,Passingham, R.E., 1998. The left hemisphere and the selectionof learned actions. Neuropsychologia 36 (1), 11–24.
Schluter, N.D., Rushworth, M.F., Passingham, R.E., Mills, K.R., 1998.Temporary interference in human lateral premotor cortexsuggests dominance for the selection of movements A studyusing transcranialmagnetic stimulation.Brain 121 (Pt 5), 785–799.
Schluter, N.D., Krams, M., Rushworth, M.F., Passingham, R.E., 2001.Cerebral dominance for action in the human brain: theselection of actions. Neuropsychologia 39 (2), 105–113.
Smid, H.G., Jakob, A., Heinze, H.J., 1999. An event-related brainpotential study of visual selective attention to conjunctions ofcolor and shape. Psychophysiology 36 (2), 264–279.
Steenbergen, B., Gordon, A.M., 2006. Activity limitation inhemiplegic cerebral palsy: evidence for disorders in motorplanning. Dev. Med. Child Neurol. 48 (9), 780–783.
Steenbergen, B., van der Kamp, J., 2004. Control of prehension inhemiparetic cerebral palsy: similarities and differencesbetween the ipsi- and contra-lesional sides of the body. Dev.Med. Child Neurol. 46 (5), 325–332.

67B R A I N R E S E A R C H 1 3 4 9 ( 2 0 1 0 ) 5 6 – 6 7
Steenbergen, B., Meulenbroek, R.G., Rosenbaum, D.A., 2004.Constraints on grip selection in hemiparetic cerebral palsy:effects of lesional side, end-point accuracy, and context. BrainRes. Cogn. Brain Res. 19 (2), 145–159.
te Velde, A.F., van der Kamp, J., Becher, J.G., van Bennekom, C.,Savelsbergh, G.J., 2005. Planning and control in a manualcollision avoidance task by children with hemiparesis. Mot.Control 9 (4), 417–438.
Tiffin, J., 1985. Purdue-Pegboard Examiner Manual. ScienceResearch Associates, Chicago.
van Elk, M., van Schie, H. T., Neggers, S. F., & Bekkering, H. (inpress). Neural and temporal dynamics underlying visualselection for action. J Neurophysiol. doi:10.1152/jn.01079.2009.
van Schie, H.T., Bekkering, H., 2007. Neural mechanismsunderlying immediate and final action goals in object usereflected by slowwave brain potentials. Brain Res. 1148, 183–197.
van Schie, H.T., Mars, R.B., Coles, M.G., Bekkering, H., 2004.Modulation of activity in medial frontal and motor corticesduring error observation. Nat. Neurosci. 7 (5), 549–554.
Vingerhoets, G., 2008. Knowing about tools: neural correlates oftool familiarity and experience. Neuroimage 40 (3), 1380–1391.
Vogt, B.A., Pandya, D.N., 1987. Cingulate cortex of the rhesusmonkey. II. Cortical afferents. J Comp Neurol 262 (2), 271–289.
Vogt, B.A., Vogt, L., Laureys, S., 2006. Cytology and functionallycorrelated circuits of human posterior cingulate areas.Neuroimage 29 (2), 452–466.
Willems, R.M., Hagoort, P., 2009. Hand preference influencesneural correlates of action observation. Brain Res. 1269, 90–104.
Willems, R.M., Peelen, M.V., Hagoort, P., 2009a. Cerebrallateralization of face-selective and body-selective visual areasdepends on handedness. Cereb. Cortex.
Willems, R.M., Toni, I., Hagoort, P., Casasanto, D., 2009b.Body-specific motor imagery of hand actions: neural evidencefrom right- and left-handers. Front. Hum. Neurosci. 3, 39.
Wu, Y.W., Lindan, C.E., Henning, L.H., Yoshida, C.K., Fullerton, H.J.,Ferriero, D.M., et al., 2006. Neuroimaging abnormalities ininfants with congenital hemiparesis. Pediatr. Neurol. 35 (3),191–196.
Related Documents
![EfficacyofManipulativeAcupunctureTherapyMonitoredbyLSCI ...Bell’s palsy is an acute peripheral facial nerve palsy of un-knowncauseandaccountsfor50%ofallcasesoffacialnerve palsy [1].](https://static.cupdf.com/doc/110x72/60a4deb9e0003e748e568e41/efficacyofmanipulativeacupuncturetherapymonitoredbylsci-bellas-palsy-is-an.jpg)










