Accepted Manuscript Mycobacterium bovis Bacillus Calmette-Guérin killed by extended freeze-drying reduces colitis in mice Micheline Lagranderie, Christoph Kluge, Helene Kiefer-Biasizzo, Mohammad Abolhassani, Marie-Anne Nahori, Catherine Fitting, Michel Huerre, Antonio Bandeira, Herve Bercovier, Gilles Marchal PII: S0016-5085(11)00619-6 DOI: 10.1053/j.gastro.2011.05.002 Reference: YGAST 57078 To appear in: Gastroenterology Received date: 22 July 2010 Revised date: 28 April 2011 Accepted date: 5 May 2011 Please cite this article as: Lagranderie, M., Kluge, C., Kiefer-Biasizzo, H., Abolhassani, M., Nahori, M.A., Fitting, C., Huerre, M., Bandeira, A., Bercovier, H., Marchal, G., Mycobacterium bovis Bacillus Calmette-Guérin killed by extended freeze-drying reduces colitis in mice, Gastroenterology (2011), doi: 10.1053/j.gastro.2011.05.002. This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. All studies published in Gastroenterology are embargoed until 3PM ET of the day they are published as corrected proofs on-line. Studies cannot be publicized as accepted manuscripts or uncorrected proofs.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Accepted Manuscript
Mycobacterium bovis Bacillus Calmette-Guérin killed by extended freeze-drying reduces colitis in mice
Micheline Lagranderie, Christoph Kluge, Helene Kiefer-Biasizzo, Mohammad Abolhassani, Marie-Anne Nahori, Catherine Fitting, Michel Huerre, Antonio Bandeira, Herve Bercovier, Gilles Marchal
PII: S0016-5085(11)00619-6 DOI: 10.1053/j.gastro.2011.05.002
Reference: YGAST 57078
To appear in: Gastroenterology
Received date: 22 July 2010 Revised date: 28 April 2011 Accepted date: 5 May 2011 Please cite this article as: Lagranderie, M., Kluge, C., Kiefer-Biasizzo, H., Abolhassani, M., Nahori, M.A., Fitting, C., Huerre, M., Bandeira, A., Bercovier, H., Marchal, G., Mycobacterium bovis Bacillus Calmette-Guérin killed by extended freeze-drying reduces colitis in mice, Gastroenterology (2011), doi: 10.1053/j.gastro.2011.05.002. This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. All studies published in Gastroenterology are embargoed until 3PM ET of the day they are published as corrected proofs on-line. Studies cannot be publicized as accepted manuscripts or uncorrected proofs.

1
Mycobacterium bovis Bacillus Calmette-Guérin killed by extended freeze-
drying reduces colitis in mice
Short title: Dehydrated BCG immunoregulates colitis in mice
MICHELINE LAGRANDERIE,1,2 CHRISTOPH KLUGE,2 HELENE KIEFER-BIASIZZO,3
MOHAMMAD ABOLHASSANI,1 MARIE-ANNE NAHORI,4 CATHERINE FITTING,5
MICHEL HUERRE, 6 ANTONIO BANDEIRA,7,* HERVE BERCOVIER,8 and GILLES
MARCHAL1,2
1 Laboratoire d’Immunothérapie, 2 Immunotherapix, 3 Plate forme de cytométrie, 4 Unité des
interactions bactéries-cellules, 5 Unité de Recherche Cytokines et inflammation, 6 Unité de
Recherche et d’Expertise Histotechnologie et Pathologie, 7 Unité du développement des
lymphocytes. 1, 4, 5, 6,7,* Institut Pasteur Paris, France. 2 BioTop Institut Pasteur Paris, France. 8 Hebrew University of Jerusalem, Department of Microbiology and Molecular Genetics,
Faculty of Medicine, Jerusalem, Israel. * present address : Unité de Biologie des Populations
Lymphocytaires
Corresponding author
Gilles Marchal, MD
Immunotherapix, BioTop Institut Pasteur, 28 rue du Dr Roux 75015 Paris, France.
Tel: 33 1 45 68 86 68 Fax: 33 1 40 61 33 32
e-mail: [email protected]
This work was supported by the Institut Pasteur and Immunotherapix
C.K., H.K‐B., M.A., M‐A.N., C.F., M.H., A.B. declare no conflict of interest
M.L., H.B. and G.M. are shareholders of Immunotherapix, a start‐up of the Institut Pasteur
edicated to EFD BCG development. d
C.K., H.K‐B., M.A., M‐A.N., C.F., M.H., experimental works, M.L., A.B., H.B. , G.M. conception
of new experiments, data analysis and writing.

2
ABSTRACT
BACKGROUND & AIMS: Mycobacterium bovis Bacillus Calmette-Guérin (BCG), killed by
extended freeze-drying (EFD BCG), induces secretion of interleukin (IL)-10 and reduces lung
inflammation in a mouse model of asthma. We investigated the effects of EFD BCG in mouse models
of inflammatory bowel disease (IBD).
METHODS: EFD BCG was administered subcutaneously to mice with colitis, induced by dextran
sodium sulfate (DSS), oxazolone (OXA), or adoptive transfer of CD4+CD45RBhighFoxp3- T cells from
C57Bl/6 Foxp3GFP mice to RAG2-/- mice.
RESULTS: EFD BCG, administered either before induction of DSS- and oxazolone- induced colitis
or after development of acute or chronic DSS-induced colitis, reduced symptom scores, loss of body
weight, and inflammation. Although transfer of CD4+CD45RBhighFoxp3- cells induced colitis in
RAG2-/- mice, administration of EFD BCG at the time of the transfer converted Foxp3- T cells to
Foxp3+ T cells and the mice did not develop colitis. EFD BCG protected mice from colitis via a
mechanism that required expansion of T regulatory (Treg) cells and production of IL-10 and
transforming growth factor-β (TGF-β). EFD BCG activated the RXRα–PPAR-γ heterodimer, blocked
translocation of NF-κB to the nucleus, and reduced colonic inflammation; it did not increase the
number of colon tumors that formed in mice with chronic, DSS-induced colitis.
CONCLUSION: EFD BCG controls severe colitis in mice by expanding Treg cell populations and
PPAR-γ and might be developed to treat patients with IBD.
Keywords: Killed BCG; IL-10; therapeutic vaccine; immune regulation

3
Introduction
Inflammatory bowel diseases (IBD) are chronic disorders of the gastrointestinal tract,
including Crohn’s disease and ulcerative colitis. The pathogenesis of IBD has not been
completely elucidated, but there is increasing evidence that inflammatory cytokines and
chemokines produced in the lesions with local neutrophil accumulation seem to play a major
role in acute flares of the disease (1-3). The pathogenesis of IBD involves gut epithelial barrier
defects, environmental, microbial and genetic factors (1, 4, 5). IBD has been linked to
deficiencies in the regulation of the immune response resulting in an excess of inflammatory
stimuli and mediators (6).
Analyses of immune responses in animal models have improved our understanding of
the physiopathology of IBD (7, 8). However, the treatment of IBD patients remains a major
challenge in clinical practice. Recently developed therapeutic approaches such as inhibitors of
pro-inflammatory cytokines, blockade of T cells or of lymphocyte migration, or PPAR-
γ agonists may be associated with side effects and treatment failure may occur over time (9-
11). PPAR-γ expression is impaired in the epithelial cells of the colon in patients with
ulcerative colitis, suggesting that PPAR-γ that inhibits NF-κBp65 translocation may have a
protective effect in IBD (12, 13). Accumulating data from clinical and experimental studies
have also highlighted the important role of regulatory T cells producing TGF-β or IL-10 in the
control of IBD (14, 15). We have shown that Mycobacterium bovis BCG killed by extended
freeze-drying (EFD BCG) reverses established inflammation in a murine model of asthma by
increasing the number of IL-10 producing Tregs and by enhancing PPAR-γ expression (16,
17). Therefore, we analyzed the effect of EFD BCG in three experimental models of IBD: 1)
DSS- induced colitis which is a useful murine model of IBD for studies of the cellular basis of

4
disease and the validation of therapeutic agents for human IBD (18, 19) 2) oxazolone-induced
colitis (20) and 3) spontaneous colitis developing after the adoptive transfer of naïve T cells
(CD4+CD45RBhigh) into RAG2-/- mice (21).
We found that EFD BCG prevented DSS- and oxazolone- induced colitis and cured mice
with established acute and chronic DSS-induced colitis as shown by normal colon length,
decreased of pro-inflammatory cytokines and myeloperoxidase (MPO) activity. EFD BCG
enhanced PPAR-γ expression and inhibited nuclear NF-κB translocation. Furthermore it
converted naïve T cells into Tregs expressing Foxp3 at the local (gut) and systemic (spleen)
levels in the model of adoptive transfer to RAG2-/- mice. Blockades of anti-inflammatory
cytokines (IL-10, TGF-β) and Treg depletion cancelled the protective effect of EFD BCG in
DSS-induced colitis.

5
Materials and Methods
Mice
Male BALB/c and C57Bl/6 mice were purchased from Charles River France or Centre
d’Elevage Janvier (Le Genest, St Isle, France) for the DSS- and oxazolone- induced colitis
models. Female Swiss mice from the Centre d’Elevage Janvier were used to study the
incidence of adenocarcinomas in DSS-induced long-term colitis (22). Female RAG2-/- mice
(from the central animal house of Institut Pasteur) and C57Bl/6-Foxp3GFP mice (kindly
provided by B. Malissen) were used as recipient and donor mice respectively in a model of
colitis induced by the transfer of naïve T cells. Mice were 6-7 week-old at the start of the study
and were kept in specific pathogen-free conditions in accordance with national guidelines for
animal welfare.
Acute and chronic chemically induced colitis
Acute DSS model: DSS (MW= 36000-50000), (MP Biomedicals, Illkirch, France) was
added to the drinking water of C57Bl/6 mice (2.5% DSS given for 5 days to mice from
Elevage Janvier, and 3% DSS given for 6 days to mice from Charles River France). Mice were
treated subcutaneously at the base of the tail with 100 μl of PBS or 100μg of EFD BCG
prepared as previously described (16): 1) preventively (21 days before DSS administration),
and mice were sacrificed 7 days after the end of DSS treatment, or 2) curatively
(administration 2 days after the end of the course of DSS treatment), and mice were sacrificed
20 days after EFD BCG treatment.
Chronic DSS model: C57Bl/6 mice were subjected to one cycle of treatment with 1.5% DSS in
drinking water for 7 days followed by 10 days of sterile tap water and then to 2 cycles of 5

6
days 1.5 % DSS followed by 10 days of sterile tap water. Mice were treated subcutaneously at
the base of the tail (2 days after the first DSS cycle) with 100 μl of PBS or 100 μg of EFD
BCG. All mice were sacrificed 18 days after the third DSS cycle.
Acute oxazolone model: BALB/c mice were treated subcutaneously, at the base of the
tail, with 100 μl of PBS or 100μg of EFD BCG. Three weeks later, mice were epicutaneously
sensitized with 3% oxazolone (4-ethoxymethylene-2-phenyl-2-oxazolin-5-one; Sigma-
Aldrich). Mice were anesthetized 8 days later and treated intrarectally with 1% oxazolone in
50% ethanol as previously described (23). Control mice received 50% ethanol alone. Mice
were sacrificed 4 days after the oxazolone challenge.
Adoptive transfer of naïve T cells
CD4+ spleen cells from female C57Bl/6 Foxp3 GFP naïve mice were purified on an
AutoMACS (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer’s
instructions and were then sorted into CD4+CD45RBhighFoxp3- cells (purity ≥ 90%)
(FACSAria, BD). The purified naïve cells (3x105) were intravenously transferred to female
RAG2-/- recipient mice receiving concomitant treatment with 100 μg of EFD BCG or 100 μl of
PBS (subcutaneously at the base of the tail).
In vivo treatment with anti-IL-10, anti-TGF-β or anti-CD25 antibodies
C57Bl/6 mice were EFD BCG-treated 21 days before the induction of acute colitis (3%
DSS). Two days after DSS exposure, groups of mice were treated intraperitoneally with 1)
0.5mg of anti-CD25 antibody (clone PC61) gift from A.Herbelin, 2) 1mg of anti-TGF-
β antibody (clone 2G7) provided by L. Chatenoud, 3) 0.5mg of anti-IL-10 antibody (clone

7
JES-2A5), followed by a second injection at the end of DSS treatment (day 6) as previously
described (16), 4) 0.5 mg of rat IgG1 isotypic antibody. All mice were killed 10 days after the
start of DSS treatment.
Statistical analysis
Data are presented as mean ± SD. The Instat package from Graph Pad Software (San
Diego, CA, USA) was used to analyze the data, using the t-test with Welch’s correction.

8
Results
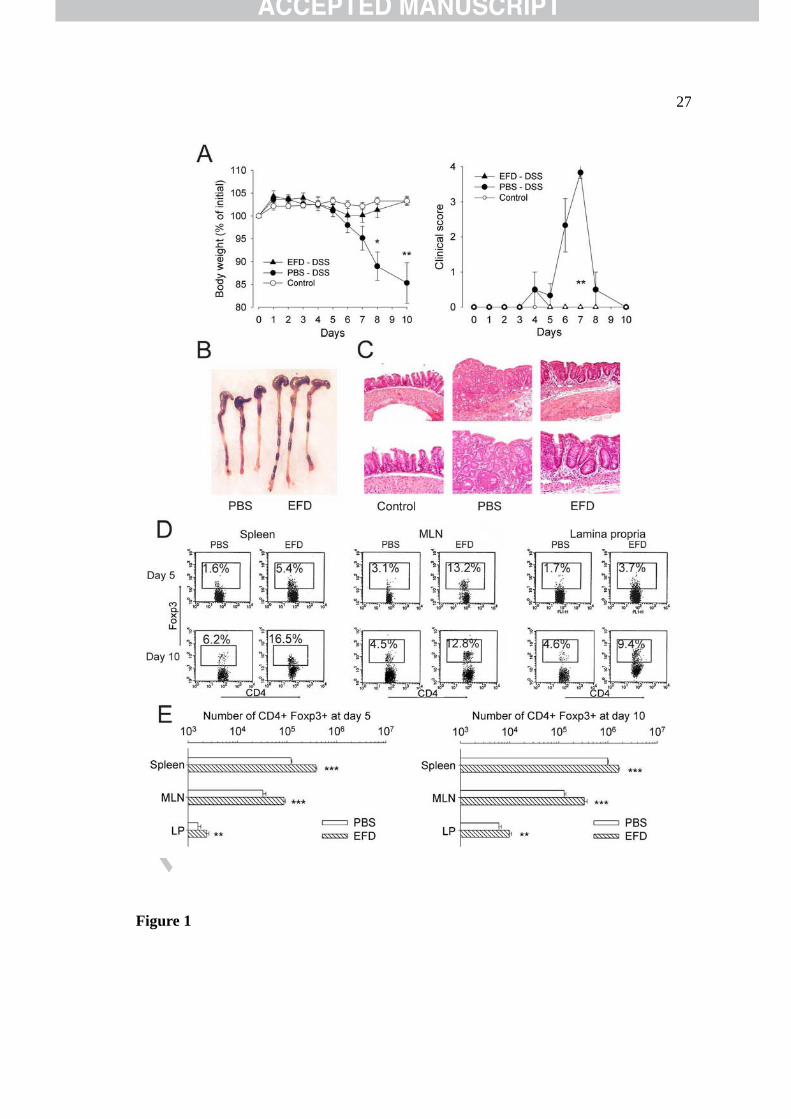
EFD BCG prevents acute DSS-induced colitis
Acute colitis resulted in substantial body weight loss in PBS-treated mice, whereas mice
treated preventively with EFD BCG displayed only slight weight loss (Figure 1A). Clinical
scores peaked at day 7 with diarrhea and wet blood present on the anus of PBS-treated mice
whereas only one mouse displayed diarrhea and none had wet blood on the anus in the EFD
BCG-treated group (Figure 1A; P <0.01). Colon length was significantly shorter in PBS- than
in EFD BCG-treated and naïve mice (4.7 ± 0.5 cm, 6.1 ± 0.2 cm and 6.4 ±0.5 cm respectively;
Figure 1B). Inflammatory infiltrate and a loss of normal colonic architecture were observed in
the colon of PBS-treated mice whereas EFD BCG-treated mice displayed only mild infiltration
(Figure 1C). Five and 10 days after DSS exposure, EFD BCG-treated mice had a higher
percentage and absolute number of CD4+Foxp3+ cells in the spleen, mesenteric lymph nodes
(MLNs) and lamina propria (LP) than PBS-treated mice (Figure 1D, E, Supplementary Figure
1). By contrast, total cell and CD4+ T cell numbers were higher after PBS- than after EFD
BCG-treatment (Supplementary Figure 2).
EFD BCG cures acute and chronic DSS-induced colitis
We then assessed the therapeutic effect of EFD BCG administered after the onset (day 6)
of acute IBD (2.5% DSS for 5 days), at a time at which clinical scores were high (3.4 ± 0.6)
and substantial weight loss was observed (Figure 2A). Mouse body weight began to increase 2
days after the EFD BCG treatment, reaching that of control mice by day 17 and remaining
significantly higher than that of PBS-treated mice from day 24 until the end of the experiment
(Figure 2A; P<0.05). Two days after EFD BCG treatment clinical scores fell to one third (2.7 ±

9
0.4 versus 0.8 ± 0.6) those of PBS-treated mice and, at the end of the experiment colon length
in EFD BCG-treated mice (7.8 ± 0.2 cm) was similar to that in naive mice (8.1 ± 0.4 cm)
whereas PBS-treated mice had significantly shorter colons (6.2 ± 0.3 cm, P<0.001). IBD are
chronic diseases in which flares are followed by remissions. In an experimental chronic model
(three cycles of treatment with 1.5% DSS interrupted by 10 days of tap water) (Figure 2B), we
tested the efficacy of EFD BCG administered 2 days after the end of the first DSS cycle.
Weight loss was observed after each DSS cycle in PBS-treated mice whereas EFD BCG-
treated mice rapidly gained weights to reach the same weight as control mice (day 16) even
after two further DSS cycles (Figure 2B). PBS-treated mice developed chronic features with
high clinical scores until the end of the experiment (1.8 ± 0.4) whereas EFD BCG-treated mice
had normal stools. The colon lengths of both PBS- and EFD BCG -treated mice (6.8 ± 0.5 cm
and 7.2 ± 0.2 cm respectively) were shorter than that of naïve mice (7.9 ± 0.3 cm). High levels
of inflammatory cytokines (IFN-γ, IL-6 and IL-17) were found in the spleen cell culture
supernatants of PBS-treated mice but EFD BCG treatment significantly decreased these levels
in acute and chronic colitis (Figure 2C; P<0.01).
The nuclear receptor PPAR-γ can inhibit NF-κB activation and inflammatory cytokine
expression in IBD (13). In acute and chronic DSS-induced models of IBD, EFD BCG
treatment significantly decreased NF-κB activation (Figure 2D; P<0.001) and increased
PPAR-γ expression (Figure 2E; P<0.001) in nuclear extracts of colon cells.
Antibodies against IL-10, TGF-β or CD25 blocked the protective effect of EFD BCG
Tregs producing IL-10 or TGF-β play a major role in the modulation of gut and lung
inflammation (14-16). Therefore, in the acute DSS model, mice treated preventively with EFD

10
BCG received, 2 days after the exposure to DSS, injections of antibodies against IL-10, TGF-
β, CD25 or rat IgG1 isotype. More efficient inhibition of IL-10 production was achieved by
administering a second injection of anti IL-10 at the end of DSS treatment. EFD BCG-treated
mice had a stable weight (Figure 3A), low clinical scores (0.3 ± 0.3), and significantly longer
colons than PBS-treated mice (7.9 ± 0.5 cm and 6.4 ± 0.2 cm; P<0.01). By contrast EFD BCG-
treated mice receiving in addition 2 injections of anti-IL-10 antibody or 1 injection of anti-
TGF-β or -CD25 antibodies displayed severe weight loss (Figure 3A), high clinical scores (3.8
± 0.4, 4.0 ± 0.1, and 3.5 ± 0.5 respectively) and a significantly shorter colons (6.0 ± 0.3 cm, 6.4
± 0.4 cm and 5.5 ± 0.4 cm respectively; P<0.001). Neutrophil accumulation in the colon was
evaluated by measuring the myeloperoxidase (MPO) activity in the colonic homogenates.
MPO activity in the colon increased significantly after DSS administration and was decreased
by EFD BCG -treatment unless the EFD BCG-treated mice received blocking antibodies
(Figure 3B; P<0.001). The injection of IgG1 isotype antibody into EFD BCG-treated mice did
not influence the curative effect of EFD BCG (data not shown).
For all the groups of mice described above, the levels of IL-17 involved in inflammation
(24) and of TGF-β and IL-10 that displayed immunoregulatory effect (25) were measured in
the serum and their transcripts were analyzed by semi-quantitative RT-PCR on colon extracts.
EFD BCG treatment resulted in a strong increase in TGF-β and IL-10 production in serum
(Figure 3C; P<0.001). Antibodies against TGF-β, IL-10 and CD25 strongly downregulated
TGF-β (Figure 3C; P<0.001). Serum IL-10 was decreased significantly by anti-IL-10 antibody
and to a less extent by antibodies against TGF-β and CD-25 (Figure 3C; P<0.01). In EFD
BCG-treated mice receiving antibodies against IL-10, TGF-β, or CD25, IL-17 levels were
higher than (serum) or similar (colon) to those of PBS-treated mice; EFD BCG-treated mice

11
without blocking antibodies had IL-17 levels similar to those of control naïve mice (Figure
3C). Interestingly, EFD BCG treatment resulted in significantly lower serum levels of IL-6
(278 ± 50 pg/ml versus 139 ± 29 pg/ml; P<0.01), TNF-α (340 ± 59 pg/ml versus 247 ± 56
pg/ml, P<0.05), and IL-13 (567 ± 64 pg/ml versus 290 ± 89 pg/ml; P<0.001) than PBS
treatment but had no significant effect on IFN-γ levels (139 ± 32 pg/ml versus 105 ± 96
pg/ml). The injection of Rat IgG1 isotype antibody into EFD BCG-treated mice had no effect
on cytokine levels (Figure 3C). The levels of IL-17, TGF-β and IL-10 transcripts in the colon
showed similar profiles to what was found in the serum (Figure 3D). T-bet, GATA-3, RORγt,
and Foxp3 are transcription factor signatures of theTh1, Th2, Th17, and Treg immune
responses, respectively. We therefore analyzed these transcription factors in spleen cell
extracts from the groups of mice described above. EFD BCG treatment increased Foxp3 and
T-bet expressions and strongly downregulated RORγt and GATA-3 whereas antibodies against
TGF-β, IL-10, or CD25 abolished the effect of EFD BCG on transcription (Figure 3E). The
same transcript profile, with an increase in Foxp3 and T-bet and decrease in GATA-3 and
RORγt transcripts, was obtained with colon cell extracts (data not shown).
CD4+Foxp3+ cells produce more IL-10 than TGF-β after EFD BCG treatment
Given the prominent role of IL-10- and TGF-β- producing cells in immune regulation,
we investigated the production of these cytokines by the Tregs induced by EFD BCG
treatment 10 days after exposure to DSS. After the in vitro restimulation of MLN and LP cells
with PMA-iono, the number of CD4+Foxp3+ cells producing IL-10 was higher in EFD BCG-
than PBS-treated mice, as shown by FACS analysis (Supplementary Figure 3A and B).

12
For Foxp3GFP mice, we quantified transcripts for IL-10 and TGF-β by RT-qPCR on
sorted CD4+Foxp3GFP+ MLN cells. After EFD BCG treatment we observed a 350% increase
in IL-10 transcript levels with no increase in TGF-β transcription when compared to PBS
treatment (Supplementary Figure 3C).
EFD BCG prevented acute oxazolone-induced colitis
We investigated whether EFD BCG also prevented the development of Th2-mediated
acute colitis, by treating BALB/c mice with EFD BCG three weeks before the induction of
oxazolone colitis. PBS-treated mice developed rapid-onset of colitis peaking 2 days after
intrarectal oxazolone challenge and marked by weight loss and diarrhea leading to 25% of
death (Figure 4A). Macroscopic examination of the colon 4 days after challenge showed a
shorter length and severe hemorrhagic colitis involving the distal colon (Figure 4B).
Histological examination of the distal colon of PBS-treated mice showed intense inflammation
characterized by epithelial cell loss, necrosis, patchy ulceration, inflammatory cells and
bleeding (Figure 4C). By contrast in mice EFD BCG-treated, weight loss, colon length and
inflammation were significantly reduced (Figure 4, A-C). In serum samples, EFD BCG
treatment significantly decreases levels of IL-5, IFN-γ and IL-17 inflammatory cytokines and it
also increased levels of IL-10 and TGF-β (Figure 4D; P<0.001).
EFD BCG converted CD4+FOXP3-T cells into CD4+FOXP3+Treg cells
We transferred, 3 x 105 naïve T cells (CD4+CD45highFoxp3-) from C57Bl/6-Foxp3GFP
mice, intravenously into RAG2-/- mice, which were simultaneously treated subcutaneously
with PBS or EFD BCG. As previously described by others (21), we observed that the transfer

13
of naïve CD4+ T cells to RAG2-/- mice induced colitis in recipient mice leading to weight loss
(Figure 5A) and diarrhea. RAG2-/- EFD BCG-treated mice had a clinical score of 0 and their
body weight remained higher than that of PBS-treated mice until the end of the experiment
(day 50; Figure 5A). The colon was significantly longer in EFD BCG-treated RAG2-/- mice and
the MPO content of the colon significantly was lower than that in PBS-treated mice (Figure
5B; P<0.01). Histological studies of the colon confirmed the anti-inflammatory effect of EFD
BCG (Figure 5C). After EFD BCG-treatment, levels of the inflammatory cytokines (IL-17,
TNF-α, IL-1β, IL-6) were significantly lower in the sera of RAG2-/- mice, whereas levels of
IL-10 and TGF-β were much higher (Figure 5D) and IFN-γ levels were only slightly lower
(185 pg/ml versus 127pg/ml). As observed in the DSS model, EFD BCG treatment
significantly reduced NF-κB translocation (P<0.01) and enhanced PPAR-γ expression
(P<0.05) in the colon of RAG2-/- mice (Figure 5E). The phosphorylated form of RXRα blocks
PPAR-γ activation (26). Consistent with increased PPAR-γ expression observed, EFD BCG-
treatment prevented the phosphorylation of RXRα (Figure 5E).
As determined by FACS analysis, 18 and 30 days after cell transfer, 9.3% and 11%
respectively of CD4+ spleen cells from recipient mice treated with EFD BCG displayed Foxp3
expression. By 50 days, the number of CD4+Foxp3+ cells slightly decreased (7.4%), and only
few CD4+cells of PBS-treated mice expressed Foxp3 (Figure 6A). Foxp3 was detected by
western blot analysis in the spleen and colon extracts only when mice were EFD BCG-treated
at the time of naïve T cells transfer (Figure 6B). In spleen cells extracts, T-bet was upregulated
whereas GATA-3 and RORγt were strongly downregulated when mice were EFD BCG-treated
(Figure 6C).

14
EFD BCG does not increase the risk of colon tumors development
As EFD BCG induces regulatory T cells, the effect of EFD BCG treatment on the
incidence of dysplasia and colon cancer was examined in a Swiss mouse model of long-term
colitis induced by several cycles of DSS (22). EFD BCG-treated mice had lower clinical scores
than PBS-treated mice during periods of 2 to 29 days after the third DSS cycle (Supplementary
Figure 4A; P<0.001). At the end of the experiment, 140 days after the last DSS cycle, NF-κB
translocation levels were lower and PPAR-γ levels were higher in the colonic nuclear extracts
of EFD BCG-treated mice than in those of PBS-treated (Supplementary Figure 4B; P<0.01).
Consistent with reduced inflammation, EFD BCG-treated mice had significantly longer colons
(Supplementary Figure 4C; P<0.01). EFD BCG treatment also significantly decreased the
frequency of ulcers (P<0.001) and neutrophil infiltration (P<0.05) (Supplementary Figure 4D).
Two cancers were observed in PBS-treated mice whereas none were found in the EFD BCG-
treated mice; the dysplasia index was not modified by the EFD BCG treatment.
(Supplementary Figure 4D).

15
Discussion
We have shown that EFD BCG treatment reduces lung inflammation in an asthma model
by recruiting plasmacytoid dendritic cells (pDCs) that promoted the differentiation of Tregs in
the draining lymph nodes and their migration to the spleen and lungs (17). We showed here
that EFD BCG efficiently reduced gut inflammation in DSS- and oxazolone-induced colitis
and in the model of RAG2-/- mice transferred with CD4+CD45high naïve T cells. In all the
models studied EFD BCG treatment led to significant generation of induced Tregs
CD4+Foxp3+ T that correlated with impaired development of colitis. EFD BCG 5 and 10 days
after DSS exposure increased the number of CD4+Foxp3+ cells in the spleen, MLNs and LP in
correlation with a decrease in the number of inflammatory cells in the gut. The curative effect
of EFD BCG depends on Tregs, IL-10 and TGF-β cytokines. Indeed, after EFD BCG
treatment, Foxp3 expression is increased in spleen cells extracts, whereas after the injection of
antibodies against TGF-β, IL-10 or CD25 to these mice, the expression of Foxp3 is strongly
decreased and colitis exacerbated. The CD4+Foxp3+ cells producing IL-10 were more
abundant in MLN of EFD BCG- than in PBS-treated mice. In addition, IL-10 transcript
expression was enhanced in CD4+Foxp3GFP+ cells from EFD BCG- treated mice when
compared to PBS-treated mice. By contrast TGF-β transcripts were not significantly different
in CD4+Foxp3+ cells isolated from PBS- and EFD BCG-treated mice. In accordance with these
results, it has been shown that Tregs do not need to produce TGF-β to regulate inflammation,
as TGF-β-/- Tregs can prevent colitis in a similar way to TGF-β-sufficient Tregs (27). The
higher levels of TGF-β observed in the sera of EFD BCG-treated mice can be provided by
cellular sources other than the Tregs. TGF-β has been clearly implicated in the conversion of
naïve CD4+ CD25- T cells into CD4+ CD25+ T cells through the induction of Foxp3 (28), and

16
the abolition of TGF-β production increases the levels of Th1 type cytokines in the human gut
mucosa (29). IL-10 also plays an essential role in inhibiting intestinal inflammation (15) and it
has been shown that TGF-β and IL-10 do not act independently in the prevention of colitis (30,
31). We found that antibodies against CD25, TGF-β and IL-10 abolished the protection
conferred by EFD BCG. The administration of anti-antibodies against IL-10 decreased IL-10
and TGF-β production in the sera and transcription levels in the colon, whereas antibodies
against TGF-β did not inhibit the production or transcription of IL-10. CD4+Foxp3+ cells that
preferentially produced IL-10 and not TGF-β after EFD BCG treatment may play an essential
role in the control of inflammatory responses in the gut tissues.
PBS treated mice exposed to DSS produced inflammatory Th1,Th2 and Th17
cytokines in the sera whereas EFD BCG treatment strongly downregulated all inflammatory
cytokines other than IFN-γ. However, at the transcriptional level only the expression of
GATA-3 and RORγt were reduced after EFD BCG treatment, whereas T-bet increased
significantly. The antibodies against IL-10, TGF-β, CD25 abolished the protection and
decreased T-bet expression whereas inflammatory Th1 cytokines (IFN-γ, TNF-α, IL-6,)
remained at high levels, the expression of Foxp3 and T-bet being enhanced only after EFD
BCG treatment without blocking antibodies. The function of T-bet in the coordination of Th1
inflammation in vivo is probably more complex than previously thought (32). A recent study
showed that T-bet is important not only for the differentiation of Th1 cells but also for the
control and regulation of Th1 inflammatory immune responses by T-bet+ Treg cells (33).
Garrett et al. showed that the restoration of T-bet expression in dendritic cells from T-bet-/-
RAG-/- mice ameliorated colitis and prevented neoplasia and, that T-bet plays the role of
“peacekeeper” in host-commensal relationships in ulcerative colitis (34, 35).

17
Murine IL-17 cells originate from naïve CD4+ T cells in the presence of IL-6 and TGF-β
and their development is then amplified by IL-23 (36) with RORγt acting as the master
transcription factor guiding Th17 differentiation (37). In the DSS-induced colitis model, EFD
BCG-treated mice had low levels of IL-17 and RORγt transcript whereas both transcript levels
were enhanced in PBS-treated mice and in EFD BCG-treated mice receiving blocking
antibodies. It has recently shown, in a murine model of asthma that a PPAR-γ agonist
downregulates NF-κB and IL-17 (17, 38) and that PPAR-γ is a potential new target for the
treatment of IBD (39). Indeed, EFD BCG may attenuate the colon inflammation by activating
RXRα/PPAR-γ heterodimer, blocking the NF-κB translocation from the cytoplasm to the
nucleus.
Therapies for IBD generally target events downstream from the inflammatory cascade
and many are highly specific (40) acting on a single factor of the disease. They therefore have
a limited clinical impact, and some treatments have induced serious side effects (10,12). By
contrast, EFD BCG treatment before or after the onset of disease induced persistent Tregs
exerting an immunoregulatory effect on Th1, Th2 and Th17 inflammatory cells, with an
increase in PPAR-γ expression and a decrease in the translocation of NF-κB to the nucleus.
Moreover, EFD BCG has no known side effects (16) and, despite its immunoregulatory effect,
it had no impact on cancer incidence after long-term multiple DSS treatments and, it did not
exacerbate microbial, parasite or virus infections (manuscript in preparation). EFD BCG
therefore appears to be a promising new treatment for IBD.
Acknowledgements
We would like to thank Delphine Guy-Grand for advice concerning the isolation of

18
lamina propria cells and helpful discussions.

19
REFERENCES
1. Podolsky DK. Inflammatory bowel disease. N Engl J Med 2002;347:417-429.
2. Papadakis KA. Chemokines in inflammatory bowel disease. Curr Allergy Asthma Rep
2004;4:83-89
3. Chin AC, Parkos CA. Transepithelial migration and epithelial barrier function in IBD
potential targets for inhibiting neutrophil trafficking. Ann NY Acad Sci 2006;1072:276-287.
4. Loftus EV Jr. Clinical epidemiology of inflammatory bowel disease: incidence, prevalence,
and environmental influences. Gastroenterology 2004;126:1504-1517.
5. Van Limbergen J, Wilson DC, Satsangi J. The genetics of Crohn’s disease. Annu Rev
Genomics Hum Genet. 2009;10:89-116.
6. Neurath MF, Finotto S, Glimcher LH. The role of Th1/Th2 polarization in mucosal
immunity. Nat Med 2002;8:567-573.
7. Strober W, Fuss IJ, Blumberg RS. The immunology of mucosal models of inflammation.
Annu Rev Immunol 2002;20:495-549.
8. Kawada M, Arihiro A, Mizoguchi E. Insights from advances in research of chemically
induced experimental models of human inflammatory bowel disease. World J Gastroenterol
2007;13:5581-5593.
9. Peyrin-Biroulet L, Desreumaux P, Sandborn WJ, et al. Crohn’s disease: beyond
antagonists of tumor necrosis factor. Lancet 2008;372:67-81.
10. Stack WA, Mann SD, Roy AJ, et al. Randomised controlled trial of CDP571 antibody to
tumor necrosis factor-alpha in Crohn’s disease. Lancet 1997;349:521-524.
11. Bevan MJ, Fink PJ. Immunology: Surprising side effects. Nature 2008;454:837-838.
12. Desreumaux P, Dubuquoy L, Nutten S, et al. Attenuation of colon inflammation through
activators of the retinoid X receptor (RXR)/peroxisome proliferators-activated receptor gamma

20
(PPARgamma) heterodimer. A basis for new therapeutic strategies. J Exp Med 2001;193:827-
838.
13. Su CG, Wen X, Bailey ST, et al. A novel therapy for colitis utilizing PPARγ ligands to
inhibit the epithelial inflammatory response. J Clin Invest 1999;104:383-389.
14. Fantini MC, Becker C, Tubbe I, et al. Transforming growth factor beta induced Foxp3+
regulatory T cells suppress Th1 mediated experimental colitis. Gut 2006;55:671-680.
15. Asseman C, Mauze S, Leach MW, et al. An essential role for interleukin 10 in the function
of regulatory T cells that inhibit intestinal inflammation. J Exp Med 1999;190:995-1004.
16. Lagranderie M, Abolhassani M, Vanoirbeek J , et al. Mycobacterium bovis BCG killed by
extended freeze-drying reduces airway hyperresponsiveness in 2 animal models. J allergy Clin
Immunol. 2008; 121:471-478.
17. Lagranderie M, Abolhassani M, Vanoirbeek JAJ, et al. Mycobacterium bovis bacillus
Calmette-Guérin killed by extended freeze-drying target plasmacytoid dendritic cells to
regulate lung inflammation. J Immunol 2010;184;1062-1070.
18. Byrne FR, Viney JL. Mouse models of inflammatory bowel disease. Curr Opin Drug
Discov Devel 2006;9:207-217.
19. Melgar S, Karlsson L, Rehnström E, et al. Validation of murine dextran sulfate sodium-
induced colitis using four therapeutic agents for human inflammatory bowel disease. Int
Immunopharmacol 2008;8:836-844.
20. Boirivant M, Fuss IJ, Chu A, et al. Oxazolone colitis: a murine model of T helper cell type
2 colitis treatable with antibodies to interleukin 4. J Exp Med 1998;1929-1939.

21
21. Scheerens H, Hessel E, de Waal-Malefyt, et al. Characterization of chemokines and
chemokine receptors in two murine models of inflammatory bowel disease: IL-10-/- mice and
Rag2-/- mice reconstituted with CD4+CD45RBhigh T cells. Eur J Immunol 2001;31:1465-1474.
22. Cooper HS, Murthy S, Kido K, et al. Dysplasia and cancer in the dextran sulfate sodium
mouse colitis model. Relevance to colitis-associated neoplasia in the human: a study of
histopathology, B-catenin and p53 expression and the role of inflammation. Carcinogenesis
2000;21:757-768.
23. Wirtz S, Neufert C, Weigmann B, et al. Chemically induced mouse models of intestinal
inflammation. Nature protocols 2007;2:541-546.
24. Eastaff-Leung N, Mabarrack N, Barbour A, et al. Foxp3(+) regulatory T cells, Th17
effector cells, and cytokine environment in inflammatory bowel disease. J Clin Immunol
2010;30:80-89.
25. Liu H, Hu B, Xu D, et al. CD4+CD25+ regulatory T cells cure murine colitis: the role of
IL-10, TGF-β, and CTLA4. J Immunol 2003;171:5012-5017.
26. Yamazaki K, Shimizu M, Okuno M, et al. Synergistic effects of RXRα and PPARγ ligands
to inhibit growth in human colon cancer cells-phosphorylated RXRα is a critical target for
colon cancer management. Gut 2007;56:1557-1563.
27. Izcue A, Coombes JL, Powrie F. Regulatory T cells suppress systemic and mucosal
immune activation to control intestinal inflammation. Immunol Rev 2006;212:256-271.
28. Huber S, Schramm C, Lehr HA, et al. TGF-β signalling is required for the in vivo
expansion and immunosuppressive capacity of regulatory CD4+CD25+ T cells. J immunol
2004;173:6526-6531.

22
29. Di Sabatino A, Pickard KM, Rampton D, et al. Blockade of transforming growth factor
beta upregulates T-box transcription factor T-bet and increases T helper cell type 1 cytokine
and matrix metalloproteinase-3 production in the human gut mucosa. Gut 2008;57:605-612.
30. Fuss IJ, Boirivant M, Lacy B, et al. The interrelated roles of TGF-β and IL-10 in the
regulation of experimental colitis. J Immunol 2002;168:900-908.
31. Cottrez F, Groux H. Regulation of TGF-β response during T cell activation is modulated
by IL-10. J Immunol 2001;167:773-778.
32. Mosmann TR, Coffman RL. Th1 and Th2 cells: different patterns of lymphokine secretion
lead to different functional properties. Rev Immunol 1989;7:145-173.
33. Koch M, Tucker-Heard G, et al. The transcription factor T-bet controls regulatory T cell
homeostasis and function during type 1 inflammation. Nat Immunol 2009;10:595-602.
34. Garrett WS, Punit S, Gallini CA, et al. Colitis-associated colorectal cancer driven by T-bet
deficiency in dendritic cells. Cancer Cell 2009;16:208-219.
35. Garrett WS, Glimcher LH. T-bet-/- GAG2-/- ulcerative colitis: the role of T-bet as a
peacekeeper of host-commensal relationships. Cytokine 2009;48:144-147.
36. Aggarwal S, Ghilardi N, Xie MH, et al. Interleukin 23 promotes a distinct CD4 T cell
activation state characterized by the production of interleukin-17. J Biol Chem 2003;278:1010-
1914.
37. Ivanov II, McKenzie BS, Zhou L, et al. The orphan nuclear receptor ROR-gammat directs
the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006;126:1121-
1133.

23
38. Park SJ, Lee KS, Kim SR, et al. Peroxisome proliferators-activated receptor γ agonist
down-regulates IL-17 expression in a murine model of allergic airway inflammation. J
Immunol 2009;183:3259-3267.
39. Dubuquoy L, Rousseaux C, Thuru X, et al. PPARγ as a new therapeutic target in
inflammatory bowel diseases. Gut 2006;55:1341-1349.
40. Baumgart DC, Sandborn WJ. Inflammatory bowel disease: clinical aspects and established
and evolving therapies. Lancet 2007;369:1641-1657.
41. Arstila T, Arstila TP, Calbo S, et al. Identical T cell clones are located within the mouse
gut epothelium and lamina propria and circulate in the thoracic duct lymph. J Exp Med
2000;191:823-834.

24
Figure Legends
Figure 1. Prophylactic EFD BCG treatment of C57Bl/6 mice reduced Th1-type
inflammation in acute DSS-induced colitis. (A) Body weight loss and clinical scores in PBS-
and EFD BCG-treated mice. (B) Colon length and (C) Representative H&E-stained colon
sections showing inflammatory infiltrate and loss of normal architecture in PBS-treated mice
and few inflammatory cells in EFD BCG-treated mice. Magnification x 4 (upper panel) and x
10 (lower panel). (D) Foxp3 expression on gated CD4+ T cells from the spleen, MLNs, and LP
on day 5 (upper panel) and day 10 (lower panel) after DSS exposure. (E) Absolute number of
CD4+Foxp3+ cells in the spleen, MLNs and LP on days 5 and 10. Data are representative of 2
independent experiments. n = 6 to 8 mice per group. * P<0.05, ** P<0.01.
Figure 2. EFD BCG treatment of C57Bl/6 mice reduced Th1-type inflammation. (A and
B) Body weight loss in acute and chronic model of DSS-induced colitis. (C) Inflammatory
cytokines in spleen cell supernatants. NF-κB (D) and PPAR-γ (E) transcription factors levels
determined in nuclear extracts of individual colon cells. n = 6 mice per group. *** P<0.001.
Figure 3. Antibodies against TGF-β, IL-10, CD25 blocked the protective effect of EFD
BCG. Prophylactic EFD BCG treatment was administered in an acute model of DSS-induced
colitis, and blocking antibodies and an isotype control were injected 2 days after the start of
DSS treatment. (A) Body weight loss. (B) MPO activity expressed as arbitrary units / mg of
colon. (C) IL-10, TGF-β, and IL-17 levels in individual serum and (D) transcripts from 2 pools
of 3 colons expressed as ratio versus GADPH. (E) Foxp3, T-bet, GATA-3, and ROR-γt
expression in the spleen after injection of blocking antibodies. Transcription factors were

25
analyzed by western blotting and expressed as ratios of the normalized intensities of the bands
(target proteins/ β-actin). For each transcription factor we analyzed 2 pools of spleen cell
extracts from 3 mice. Fine hatched bars represent the isotype control. Data are representative
of 2 independent experiments. n= 6 mice per group. ** P<0.01, *** P<0.001.
Figure 4. Prophylactic EFD BCG treatment reduced Th2-type inflammation. (A) Weight
loss measured from 8 days before oxazolone (OXA) sensitization to day 4 after intra-rectal
OXA challenge. (B) Colon length and distal lesions (inserts) in PBS- and EFD BCG- treated
mice. (C) Representative H&E-stained colon sections showing necrotic lesion with bleeding 4
days after OXA-challenge in PBS-treated mice and only mild inflammation in EFD BCG-
treated mice. Magnification x 10, white arrow showing haemorrhage. (D) Cytokines levels in
the individual serum.
Figure 5. EFD BCG treatment at the time of naïve CD4+ T cells transfer to RAG2-/- mice
reduced colitis features. Naïve CD4+CD45highFoxp3- (3x105) T cells were transferred to RAG2-
/- mice at the time of PBS or EFD BCG treatment. (A) Body weight loss, (B) colon length and
MPO activity in the colon (arbitrary units/mg) were measured on day 50. (C) Representative
H&E-stained colon sections showing inflammatory loss of normal architecture and ulceration
in PBS-treated mice and few inflammatory cells in EFD BCG-treated mice. Magnification x
10. (D) Cytokines levels in the individual serum. (E) NF-κB and PPAR-γ levels measured in
individual colon cell extracts and RXRα phosphorylation measured by western blotting in 2
pools of 3 colons. Data are representative of 2 independent experiments. n = 6 mice per group.
** P<0.01, *** P<0.001.

26
Figure 6. EFD BCG treatment converted CD4+Foxp3- cells into CD4+Foxp3+ cells in
RAG2-/- mice. (A) FACS analysis of CD4+Foxp3 GFP+ spleen cells (B) Foxp3 expression
analyzed by western blotting in spleen and colon extracts. (C) Transcription factors analyzed
by western blotting in 2 pools of 3 spleen cell extracts, and expressed as ratios of the
normalized intensities of the bands (target proteins/ β-actin). Data are representative of 2
independent experiments. n = 6 mice per group.
27
Figure 1

28
Figure 2

29
Figure 3

30
Figure 4

31
Figure 5

32
Figure 6

1
Supplementary Figures and methods
Supplementary Figure 1.
Isotype control (Rat IgG2a). No significant positive cells stained with Foxp3 isotype control
were found in the gate of CD4+ Foxp3+ Cells shown in Figure 1.
Supplementary Figure 2.
EFD BCG treatment reduced the number of inflammatory cells in the gut in DSS-induced
colitis. (A) Total number of cells and (B) absolute number of CD4+ T cells in the spleen, MLNs
and LP 5 and 10 days after DSS treatment. n = 8 mice per group.
Supplementary Figure 3.
CD4+Foxp3+ cells isolated from EFD BCG-treated mice produced large amounts of IL-10. (A)
FACS analysis of cells producing IL-10 isolated 10 days after DSS treatment from the spleen,
MLNs and LP and stimulated in vitro with PMA-iono. (B) Relative expression levels of IL-10
and TGF-β in Foxp3GFP+ cells isolated from MLNs of PBS- and EFD BCG-treated mice 10
days after DSS treatment. n= 5 to 8 mice per group.
Supplementary Figure 4.
EFD BCG treatment had no effect on the development of colon cancer. OF1 Swiss mice were
PBS- or EFD BCG-treated and were then submitted to three cycles of treatment with 5% DSS,
21 days later. (A) Clinical scores after the last DSS cycle. (B) NF-κB and PPAR-γ transcription
factor levels determined in nuclear extracts from individual colons. (C) Colon length and (D)

2
Indices of ulceration or erosion, polynuclear infiltration and dysplasia. n = 25 mice per group.
* P<0.05, ** P<0.01, *** P>0.001.

3
Supplementary methods
Evaluation of the clinical and histological severity of colitis
From the day preceding DSS treatment until the end of the experiment, clinical
evaluation of the disease was carried out daily during flares and every 2-3 days during
remission phases. Each animal was weighted and clinical colitis was scored, score of 0
corresponding to a normal fecal pellet, 1 to anal inflammation with mucous secretion, 2 to a
loose fecal pellet, 3 to diarrhea and 4 to hemorrhage (wet blood on or around the anus). At the
end of the experiment, mice were sacrificed and colon lengths were measured and colons were
fixed in 10% formalin (Sigma-Aldrich) and paraffin embedded, sections were cut and H&E
stained.
We evaluated the incidence of colitis-associated dysplasia/cancers in the colon using
Swiss OF1 mice subjected to 3 cycles of treatment with 5% DSS in the drinking water each
lasting 7 days with an interval of 2 weeks between cycles. Twenty-one days before the first
DSS cycle, one group of mice was treated subcutaneously with EFD BCG (100 μg) and a
second group was treated with PBS. All groups of mice were sacrificed 140 days after the end
of the third DSS cycle (25 mice in each group). Colons were fixed in 10% formalin and
paraffin embedded, sections were cut and H&E stained and indices of ulceration or erosion,
polynuclear infiltration and dysplasia were calculated.
Determination of colon MPO activity
Each colon was weighed and homogenized for 30s in a Potter-Elvehjem glass
homogenizer (Thomas, Philadelphia, PA) at 4°C in 0.5 ml of PBS. The extracts were
centrifuged (10,000 g, 10 min, 4°C), and the supernatants were discarded. The pellets were

4
resuspended in 1 ml of PBS supplemented with HTAB (0.5%) and EDTA (5 mM) and
homogenized again. After centrifugation, 50μl of the supernatant was placed in a test tube with
200 μl PBS-HTAB-EDTA, 2 ml HBSS, 100 μl O-dianisidine dihydrochloride (1.25 mg/ml),
and 100 μl H2O2 (0.05% = 0.4 mM). After 30 min of incubation with shaking at 37°C, the
reaction was stopped by adding of 100 μl of NaN3 (1%). Absorbance was read at 460 nm and
the MPO activity was determined in arbitrary units / mg of colon.
Blood, spleen and colon collection and cell isolation
At the end of experiments, mice were bled and sera were frozen until cytokine
determination using Bio-Plex Cytokine Assay for all cytokines (Bio-Rad, Marnes La Coquette,
France) except TGF-β measured by ELISA (eBioscience, San Diego, CA). In some
experiments spleens cells were cultured for 96 hours in vitro (without further antigen
stimulation) in RPMI medium (Gibco) supplemented with 10% FCS, culture supernatants were
removed and frozen until cytokine determination (Bio-Plex). Spleen, MLN and LP cells were
isolated as previously described (17, 18, 41).
In vitro cell activation and intracellular IL-10 staining
For intracellular IL-10 staining, MLN and LP cells were stimulated by incubation for 5h
in the presence of 50ng/ml phorbol 12-myristate 13-acetate (PMA) and 500 ng/ml ionomycin
(both from Sigma-Aldrich). For the last 2 h, 1 μl of GolgiPlug (BD Biosciences) was added to
1 ml of cell culture. After the CD4 surface staining cells were fixed and intracellular staining
was then performed in permeabilization-solution (BD Biosciences) with anti-IL-10 and anti-
Foxp3 antibodies (eBioscience). Cells were analyzed on a FACScalibur (BD Biosciences).

5
Transcription factors in the colon and spleen
Nuclear proteins were extracted from colon homogenates after protease inhibitor
treatment. Nuclear extracts (10 μg) from all groups of mice were tested for NF-κB activation
or PPAR-γ expression with NF-κBp65 or PPAR-γ TransAMTM transcription factor assay kits
(Active Motif, Carlsbad, CA) according to the manufacturer’s recommendation.
The proteins extracted from colon and spleen cells were resolved by 7.5% SDS-PAGE.
Protein bands were probed with mouse monoclonal anti-Foxp3, -T-bet, -GATA-3, -RORγt
(Santa Cruz Biotechnology, Santa Cruz, CA), or β-actin mouse monoclonal antibody (Ac-15
Abcam, Cambridge, UK). As secondary antibodies, we used HRP-conjugated polyclonal goat
anti-rabbit (Dako Cytomation, Denmark) or goat anti-rabbit IgG (Santa Cruz Biotechnology).
The immune complex was visualized with an enhanced chemiluminescence detection system
(Amersham, France). For data acquisition and background normalization, we used Scan-
Analyze, as previously described (18). Data are expressed as ratios of the normalized
intensities of the bands (target proteins versus β-actin).
The Foxp3 expression was also analyzed by FACS in spleen, MLN and LP cells isolated
5 and 10 days after DSS administration and in spleen cells of RAG2-/- mice 18, 30 and 50 days
after the transfer of CD4+CD45RBhighFoxp3- T cells from C57Bl/6 Foxp3GFP donor mice.
Semi-quantitative RT-PCR and quantitative RT-qPCR
Total RNA was isolated from frozen colons with Trizol reagent
(Invitrogen,Carlsbad,CA). The first-strand cDNA was synthesized from 2 μg of total RNA
with 2,000 pmol Oligo dT, 1.0 mM dNTP, 200 U Moloney murine leukemia virus reverse

6
transcriptase, and 5x RT buffer (Promega, Madison, WI). The mouse-specific sense and
antisense primers used were as follows: IL-17A Sense, 5’- GCTCCAGAAGGCCCTCAGA-3’,
Antisense 5’-AGCTTTCCCTCCGCATTGA-3’, TGF-β Sense, 5’-
CAACGCCATCTATGAGAAAACC-3’, Antisense. 5’-AAGCCCTGTATTCCGTCTCC3’,
IL-10 Sense, 5’-AGAGACTTGCTCTTGCACTACCAA-3’, Antisense, 5’-
GTAAGAGCAGGCAGCATAGCAGT-3’, GAPDH Sense, 5’-
AATGGTGAAGGTCGGTGTGAAC-3’, Antisense, 5’-GAAGATGGTGATGGGCTTCC3’.
The PCR products were subjected to electrophoresis in a 2% agarose gel (Invitrogen,
Carlsbad, CA) and stained with a 0.5 mg/ml ethidium bromide in Tris/borate/EDTA buffer.
The intensity of the corresponding bands was normalized to that of GAPDH.
Foxp3GFP+ and Foxp3GFP- cells from the MLNs of PBS- and EFD BCG-treated mice
were sorted with a FACSAria (BD Biosciences). Sorted cells were suspended in lysis- buffer
(Pure Link RNA micro-kit (Invitrogen Carlsbad). RNA-isolation was performed as
recommended by the manufacturer (Pure Link RNA micro-kit (Invitrogen Carlsbad). The
cDNA synthesis was performed with the QuantiTect Reverse Transcription Kit (Qiagen,
Hilden). The qPCR reactions were run on the Roche Lightcycler 480. Each reaction was
performed in duplicate. The following primer sequences were used for the qPCR reactions:
tgf-beta 1-f: CTCCCGTGGCTTCTAGTGC tgf-beta 1-r:
GCCTTAGTTTGGACAGGATCTG; foxp3-f: CCCATCCCCAGGAGTCTTG, foxp3-r:
ACCATGACTAGGGGCACTG; il-10-f: GGTTGCCAAGCCTTATCGGA, il-10-r:
ACCTGCTCCAACTGCCTTGCT; tbp-f: CCTATGACCCCTATCACT, tbp-r:
GTCCGTGGCTCTCTTAT.

7
The raw Ct values were obtained with the LightCycler Software using the second
derivative maximum method. Changes in relative gene expression were calculated by the ∆∆Ct
method implemented in REST2009 Software with TBP used for normalization.




Related Documents











