Moult in the Mediterranean monk seal from Cap Blanc, western Sahara Enric Badosa, Teresa Pastor*, Manel Gazo & Alex Aguilar Departament de Biologia Animal, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain Received 28 March 2005. Accepted 15 December 2005 At birth, the Mediterranean monk seal (Monachus monachus) is covered with dark lanugo. This is shed earlier and faster in females, so they complete the moult when they are significantly younger (64 days) than males (82 days). Moulted pups present a greyish dorsal and a white ventral pelage. Only the hair is shed in the first moult. In subsequent moults, the hairs are shed along with large sheets of cornified epidermis. A series of sequential stages were identified, which included a long pre-moult phase (brown pelage) and a post-moult phase (dull appear- ance). The shedding process lasted 15 days. It is different to that of the Hawaiian monk seal and elephant seals, as Mediterranean monk seals partly moult in the water. The intermoult period was close to one year except in females nursing a pup. Such females have longer intermoult periods and can even moult whilst still lactating. In males, the process of develop- ing the mature pelage pattern of bulls is gradual. It involves at least two annual moults and can be completed by the age of 4 years. Key words: Mediterranean monk seal, Monachus, moult, lanugo, shedding, pelage. INTRODUCTION The Mediterranean monk seal (Monachus monachus) is one of the most endangered mammals in the world (IUCN 1996), with probably fewer than 350 individuals surviving today (Aguilar 1999). Its current distribution is restricted to the southern and western Mediterranean, and to two locations in the eastern Atlantic: the Desertas Islands (Madeira) and the Cap Blanc Peninsula (Western Sahara – Mauritania). The only sizeable extant colony inhabits this latter location. Like all other pinnipeds that evolved outside of frozen habitats (Bowen 1991), the Mediterranean monk seal is covered with a dark lanugo at birth. In addition, it has a white, spotted ventral patch that is sexually dimorphic and individually unique (Badosa et al. 1998). After shedding the lanugo, moulted pups have a greyish dorsal and white ventral pelage. In females, the grey pelage is main- tained in subsequent moults and throughout their lifespan. Juvenile males are also grey. However, when they become bulls, they acquire a black pelage with a white, spotted ventral patch, that resembles their pup lanugo (Samaranch & González 2000). The only aspects of the moult that have been studied are the relation between its seasonality and the female reproductive cycle (Pastor & Aguilar 2003). Available information is limited to observations collected sporadically in the Greco-Turkish area (Yediler et al. 1993; Güçlüsoy & Savas 2003) and a description of an individual from the Cap Blanc Peninsula showing a strange pelage condition (Marchessaux 1989). Current information indicates that this condition is associ- ated with a moulting individual. By contrast, the moult process – including its progression, timing, seasonality, histology and energetic cost – is well known in other pinnipeds, and particularly in the congeneric Hawaiian monk seal (Monachus schauislandi) (Kenyon & Rice 1959; Johanos et al. 1994). The colony of Mediterranean monk seals located on the Cap Blanc Peninsula provided an excellent opportunity to study the moult in this species. The population is small enough to allow the iden- tification of most individuals, but large enough to permit studies at the population level and to assess individual variability. In this study, we describe the complete moult process from pups to adults. We focus on: timing, duration and seasonality, and relate this process to the biological and reproductive traits of individuals. The study is based on data collected over a seven-year period (1993–1999) in the Cap Blanc colony. MATERIALS & METHODS The seal colony is distributed along the west coast of the Cap Blanc Peninsula (Western Sahara – Mauritania), in the southern part of the Western Sahara, between 20°40’ and 21°20’N, and 17°00’ and 17°20’W. The largest concentration of the African Zoology 41(2): 183–192 (October 2006) *Author for correspondence. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Moult in the Mediterranean monk seal fromCap Blanc, western Sahara
Enric Badosa, Teresa Pastor*, Manel Gazo & Alex AguilarDepartament de Biologia Animal, Universitat de Barcelona, Diagonal 645, 08028 Barcelona, Spain
Received 28 March 2005. Accepted 15 December 2005
At birth, the Mediterranean monk seal (Monachus monachus) is covered with dark lanugo. Thisis shed earlier and faster in females, so they complete the moult when they are significantlyyounger (64 days) than males (82 days). Moulted pups present a greyish dorsal and a whiteventral pelage. Only the hair is shed in the first moult. In subsequent moults, the hairs are shedalong with large sheets of cornified epidermis. A series of sequential stages were identified,which included a long pre-moult phase (brown pelage) and a post-moult phase (dull appear-ance). The shedding process lasted 15 days. It is different to that of the Hawaiian monk sealand elephant seals, as Mediterranean monk seals partly moult in the water. The intermoultperiod was close to one year except in females nursing a pup. Such females have longerintermoult periods and can even moult whilst still lactating. In males, the process of develop-ing the mature pelage pattern of bulls is gradual. It involves at least two annual moults and canbe completed by the age of 4 years.
Key words: Mediterranean monk seal, Monachus, moult, lanugo, shedding, pelage.
INTRODUCTIONThe Mediterranean monk seal (Monachus monachus)is one of the most endangered mammals in theworld (IUCN 1996), with probably fewer than 350individuals surviving today (Aguilar 1999). Itscurrent distribution is restricted to the southernand western Mediterranean, and to two locationsin the eastern Atlantic: the Desertas Islands(Madeira) and the Cap Blanc Peninsula (WesternSahara – Mauritania). The only sizeable extantcolony inhabits this latter location.
Like all other pinnipeds that evolved outside offrozen habitats (Bowen 1991), the Mediterraneanmonk seal is covered with a dark lanugo at birth. Inaddition, it has a white, spotted ventral patch thatis sexually dimorphic and individually unique(Badosa et al. 1998). After shedding the lanugo,moulted pups have a greyish dorsal and whiteventral pelage. In females, the grey pelage is main-tained in subsequent moults and throughout theirlifespan. Juvenile males are also grey. However,when they become bulls, they acquire a blackpelage with a white, spotted ventral patch, thatresembles their pup lanugo (Samaranch &González 2000). The only aspects of the moult thathave been studied are the relation between itsseasonality and the female reproductive cycle(Pastor & Aguilar 2003). Available information islimited to observations collected sporadically inthe Greco-Turkish area (Yediler et al. 1993; Güçlüsoy
& Savas 2003) and a description of an individualfrom the Cap Blanc Peninsula showing a strangepelage condition (Marchessaux 1989). Currentinformation indicates that this condition is associ-ated with a moulting individual. By contrast, themoult process – including its progression, timing,seasonality, histology and energetic cost – is wellknown in other pinnipeds, and particularly inthe congeneric Hawaiian monk seal (Monachusschauislandi) (Kenyon & Rice 1959; Johanos et al.1994).
The colony of Mediterranean monk seals locatedon the Cap Blanc Peninsula provided an excellentopportunity to study the moult in this species.The population is small enough to allow the iden-tification of most individuals, but large enough topermit studies at the population level and to assessindividual variability. In this study, we describethe complete moult process from pups to adults.We focus on: timing, duration and seasonality, andrelate this process to the biological and reproductivetraits of individuals. The study is based on datacollected over a seven-year period (1993–1999) inthe Cap Blanc colony.
MATERIALS & METHODSThe seal colony is distributed along the west coastof the Cap Blanc Peninsula (Western Sahara –Mauritania), in the southern part of the WesternSahara, between 20°40’ and 21°20’N, and 17°00’and 17°20’W. The largest concentration of the
African Zoology 41(2): 183–192 (October 2006)
*Author for correspondence. E-mail: [email protected]

colony is at Costa de las Focas, where seals arefound all year round. In this area seals haul outand breed on narrow beaches inside two maincaves. Twice a day the tide washes out the caves,forcing the seals to leave the haul-out sites.
A series of surveys were conducted on the studyarea from May 1993 – December 1999. The obser-vation effort, timing and duration of these surveysvaried between years. All the months were coveredduring the observation period (for further detailssee Pastor & Aguilar 2003). Information on moultingwas collected by direct observation of identifiedseals swimming near the caves, using binocularsand photo-identification techniques. Periodicphoto-identification surveys were conductedannually, each comprising 16–34 effective fielddays (for further details see Forcada & Aguilar2000).
Animals were individually identified by a com-bination of natural (scars, wounds and disruptivecoloration patterns) and artificial marks. Non-moulted pups were sexed and identified by thedesign of their ventral patch (Badosa et al. 1998).Moulted pups were identified by their flankpigmentation pattern and by marking with twotypes of plastic tag: Temple tags (Temple Tags Inc.,Temple, TX, U.S.A.) or Supertags (Dalton Supplies,
Oxon, U.K.). Juveniles and adults were cata-logued in age classes according to their morpho-logical characteristics (Samaranch & González2000). Adults were recognized individually by thescarring pattern on their heads and backs (Forcada& Aguilar 2000).
Although moulting is a continuous process, aseries of sequential stages were identified. Thesewere termed ‘phases A to F’, and were used tostandardize observation criteria for moultingindividuals other than pups (Table 1). The identifi-cation, description and duration of these differentphases were carried out by analysing chronologicalsequences of photos of known individuals andsightings. The actual moult was considered to startduring phase B and finish during phase E. Phase Acorresponded to a pre-moult stage. This was iden-tified as a clearly observable alteration in colourand a degradation of the pelage that always oc-curred before the moult. This degradation maystart long before the actual moult, especially in ju-veniles. However, it is a process that clearly an-nounces a moult event. Phase F corresponded to apost-moult stage.
The intermoult was calculated as the periodbetween two consecutive moults during phase B.If a seal was not observed moulting during phase
184 African Zoology Vol. 41, No. 2, October 2006
Table 1.Sequential stages of the moulting process including:pre-moult, actual moult and post-moult for juveniles andadults. Data are given as mean ± standard deviation followed by range and sample size (n). ND: not determined
Phase Duration (days)
Juveniles Adults
Pre-moultA: the phase before shedding the skin and hair. The pelage acquires 192 ± 108 75 ± 29a brown coloration. In juveniles, this phase is characterized by the 35–352 26–153presence of black spots all around the body, and around the throat in n = 8 n = 7particular; a black coloration appears around eyes and nostrils.
MoultB: the skin begins to shed on the nostrils, big scars, slit of the penis, ND 7.25 ± 6.13anus, snout, and around the eyes. Skin is also shed on the hind and 1–13rear flippers. n = 4
C: the skin on the head and flippers is totally shed. ND 1.5 ± 0.71–2
n = 2
D: the skin starts to shed on the back and belly. ND ND
E: old skin only remains on the sides of the seal’s trunk. ND 5.5 ± 3.53–8
n = 2
Post-moultF: shedding is finished, the animal has a dark grey, in some cases almost ND 58 ± 2.83black, dull appearance, and the seal seems to be dry even in the water. 13–109
n = 5
B, but at a later phase, the onset of the moult wasestimated. This estimate took into account theduration of each of the phases (B to E) estimated inthis paper.
The duration of the intermoults in breeding andnon-breeding adult females were compared, todetect any effects of reproduction on the occur-rence of the moult. The onset of the moult was alsotaken as phase B for these calculations.
Statistical analyses were carried out using theSPSS 9.0.1 statistical software package. When in-sufficient data were available for each sex, resultswere pooled. Only one observation per individualwas included in all statistical calculations to assuredata independence. Prior to any analyses, datadistributions were tested for normality using theKolmogorov–Smirnov test (K–S test). The t-testwas used to compare means. Standard deviation isgiven as a measure of variability about the mean.
RESULTSData on moulting were obtained from 103 pups, 38juveniles, 34 females and 13 bulls. After prelimi-nary observations, we distinguished between thefirst moult after birth (moult of the foetal pelage orlanugo) and the subsequent annual moults.
Moult of the lanugoNineteen pups were specifically monitored to
estimate the moult timing (11 females and eight
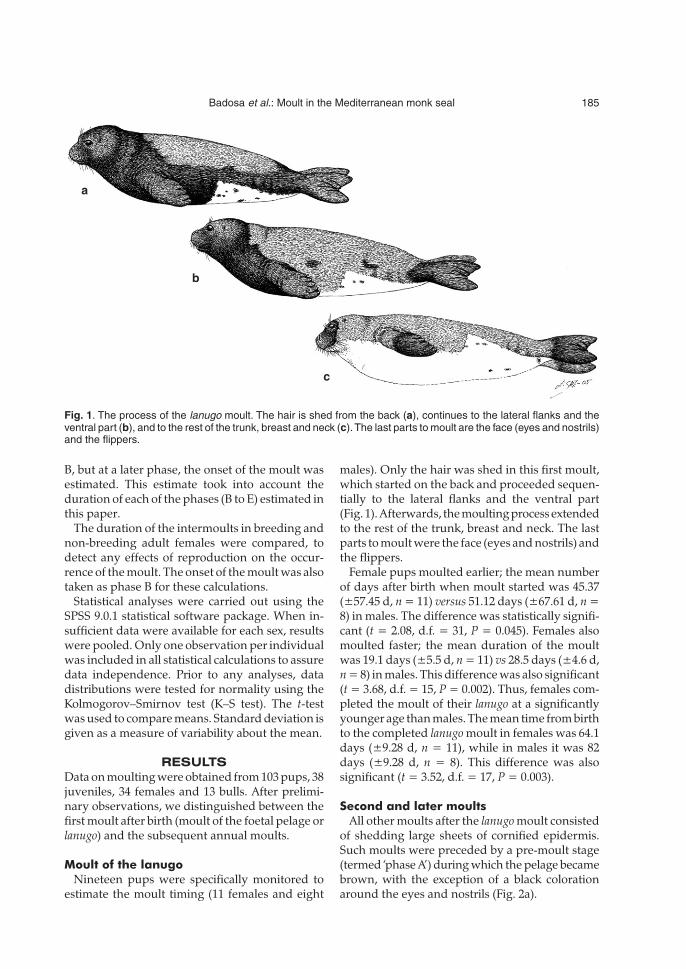
males). Only the hair was shed in this first moult,which started on the back and proceeded sequen-tially to the lateral flanks and the ventral part(Fig. 1). Afterwards, the moulting process extendedto the rest of the trunk, breast and neck. The lastparts to moult were the face (eyes and nostrils) andthe flippers.
Female pups moulted earlier; the mean numberof days after birth when moult started was 45.37(±57.45 d, n = 11) versus 51.12 days (±67.61 d, n =8) in males. The difference was statistically signifi-cant (t = 2.08, d.f. = 31, P = 0.045). Females alsomoulted faster; the mean duration of the moultwas 19.1 days (±5.5 d, n = 11) vs 28.5 days (±4.6 d,n = 8) in males. This difference was also significant(t = 3.68, d.f. = 15, P = 0.002). Thus, females com-pleted the moult of their lanugo at a significantlyyounger age than males. The mean time from birthto the completed lanugo moult in females was 64.1days (±9.28 d, n = 11), while in males it was 82days (±9.28 d, n = 8). This difference was alsosignificant (t = 3.52, d.f. = 17, P = 0.003).
Second and later moultsAll other moults after the lanugo moult consisted
of shedding large sheets of cornified epidermis.Such moults were preceded by a pre-moult stage(termed ‘phase A’) during which the pelage becamebrown, with the exception of a black colorationaround the eyes and nostrils (Fig. 2a).
Badosa et al.: Moult in the Mediterranean monk seal 185
Fig. 1. The process of the lanugo moult. The hair is shed from the back (a), continues to the lateral flanks and theventral part (b), and to the rest of the trunk, breast and neck (c).The last parts to moult are the face (eyes and nostrils)and the flippers.
a
b
c

In juveniles, this pre-moult stage (mean dura-tion = 192 ± 108 d, range 35–352, n = 8) was signifi-cantly longer (t = –2.96, d.f. = 9, P = 0.007) than inother individuals (mean duration = 75 ± 41 d,range 26–153, n = 7). Moreover, small black spotscovered the entire body surface, and the throat inparticular (Fig. 2a’). Data were only available froma single well-monitored individual to establish theonset of the juvenile moult. This was a female(no. 408), in which the moult occurred 675 days,i.e. about two years, after having shed the lanugo.During this period of time, the female spent aminimum of 236 days with a brownish pelage cor-responding to the pre-moult stage describedabove. No data were available to determine theduration of the actual moult in juveniles.
In adults, the moult started with shedding theskin from around the nostrils, larger scars, the slitof the penis, anus, snout, eyes and also from thehind and rear flippers (Fig. 2b). This stage, termedB, lasted one week (mean duration = 7.25 ± 6.13 d,range, 1–13, n = 4). During the phase termed C,the skin on the head and flippers was shedcompletely (Fig. 2c). The duration of this stage,which could only be estimated for two females,was approximately two days (mean duration =1.5 ± 0.7 d, range 1–2, n = 2). The skin continued toshed on the back and belly (phase D). The lengthof this phase could not be estimated due to its lim-ited duration (Fig. 2d). In phase E, the last pieces ofold skin remained on the sides of the trunk only(Fig. 2e). This phase lasted around five days (meanduration = 5.5 ± 3.5 d, range 3–8, n = 2). The shed-ding duration was estimated by adding the meanintervals between phases B to E, which averaged14.25 days. Moreover, the complete process alsolasted 14 days in two females (nos 83 and 48)tracked from phase B to phase E. When sheddingwas completed, the animal had a dark grey, and insome cases almost black, dull appearance and itseemed to be dry even in the water (Fig. 2f). Thispost-moult phase, termed F, could be identified foralmost two months (mean duration = 53.8 ± 2.8 d,range 13–109, n = 5).
Moult in femalesMoult ontogeny was investigated in two females
born in the same season (April and May 1994),whose moulting and reproductive events coin-cided. Both females had their juvenile moult inlate spring/early summer after a long pre-moultphase (5.5 and 7 months) and both had their firstadult moult in February. Approximately eight
months later, at least one of the two females gavebirth. However, she lost her pup four days later. Itwas not possible to confirm whether the second fe-male gave birth. The following year, both femalesmoulted in February and gave birth in October.
Lactation was not a constraint on moulting:three females (nos 55, 74 and 179) were observedto be nursing their pups when they moulted.Intermoult periods varied, depending on the re-productive status of the female. The intermoultperiod for non-parturient females (nos 21, 32, 43,49, 83, 119 and 408) was approximately one year(mean duration = 355 ± 18 d, range, 329–377, n =7), while that in parturients (nos 10, 73, 77 and 403)was more variable (mean duration = 399 ± 33 d,range 356–429, n = 4) and significantly longer (t =2.85, d.f. = 9, P = 0.01).
The duration of the intermoult period wasrelated to the duration of lactation. In femalesnos 73 and 77, lactation lasted 104 and 121 days,and their intermoult periods were 429 and421 days, respectively. By contrast, in femaleno. 403, whose lactation period was abruptly inter-rupted by the death of the pup, the intermoultperiod appeared to be much shorter (356 d). Infact, a female (no. 10) that was observed moultingin four consecutive years (1993–96) was found toalternately reduce and increase her intermoultperiods (351 d, 391 d, 303 d), with the longest inter-moult period corresponding to the year when sheraised a pup.
Moult in malesData from a single tagged male (no. 412) were
available to determine the exact age at the onset ofthe moult that led to acquiring the bull pelagepattern. This pup was monitored from the moultof its lanugo to the time when it became a bull,which occurred 1507 days, i.e. about four years,later. The process of developing the adult malepelage pattern appeared to occur gradually, andinvolved at least two annual moults. On 4 April1994, a male (no. 66) was reported as a large-sizedgrey seal. One year later, on 8 April 1995, it hadrecently completed a moult. However his overallaspect was neither that of a full bull nor that of alarge-sized grey seal, but intermediate. The dorsalpart and the top of the head were greyish-black incolour, while the entire ventral part (starting fromthe anus to the tip of the snout) was white (unlikebulls, which only have white on a clearly distinctventral patch). One year later, on 2 April 1996, hewas reported to moult and to have an overall bull
186 African Zoology Vol. 41, No. 2, October 2006

aspect, as described by Samaranch & González2000.
The intermoult period in two monitored bulls(nos 66 and 211) averaged one year (mean period= 363 ± 5.6 d). In fact, two consecutive intermoultperiods were estimated in bull no. 66. These lasted359 and 361 days, respectively.
Seasonality of eventsIn general, the seasonality of the moult was
extended in all morphological categories. Moultingpups (n = 103) were observed throughout thewhole year, with a slight peak in October/Novem-ber, that is, one month after the peak in births(Fig. 3a). No sexual differences were observed.
Badosa et al.: Moult in the Mediterranean monk seal 187
Fig. 2. The moult process. a, Pre-moult stage during which the pelage becomes brown, with the exception of a blackcoloration around the eyes and nostrils; a’, in juveniles, small black spots cover the entire body surface. b, The actualmoult starts around orifices and on the hind and rear flippers; c, the skin on the head and flippers is totally shed; d, theskin starts to shed on the back and belly; e, old skin remains on the sides of the trunk. f, Shedding is finished, theanimal has a dark grey, in some cases almost black, dull appearance.
a
b
c
d
a’
e
f
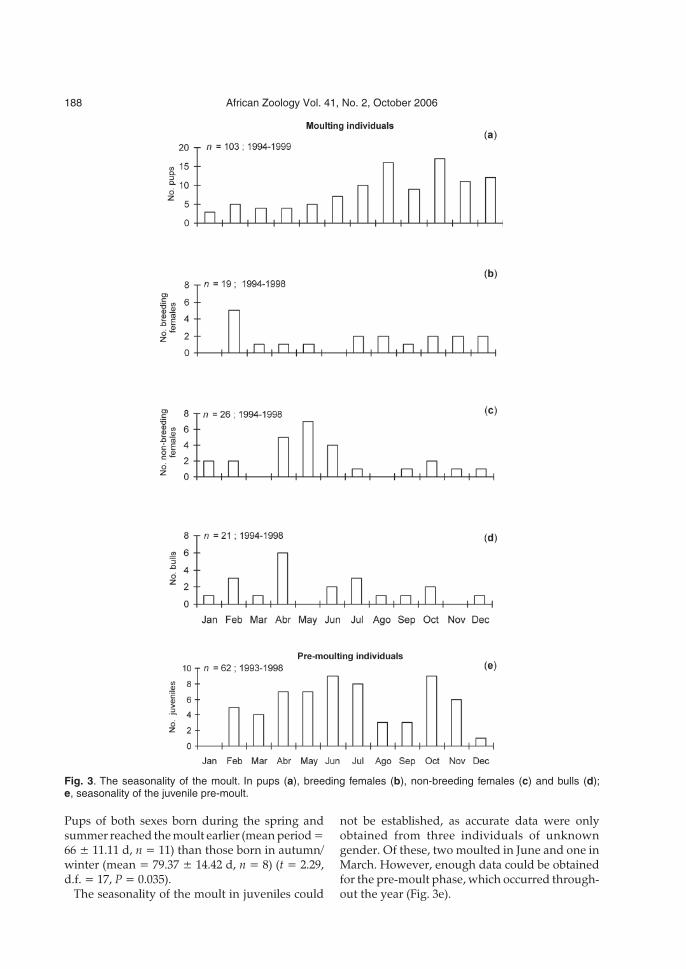
Pups of both sexes born during the spring andsummer reached the moult earlier (mean period =66 ± 11.11 d, n = 11) than those born in autumn/winter (mean = 79.37 ± 14.42 d, n = 8) (t = 2.29,d.f. = 17, P = 0.035).
The seasonality of the moult in juveniles could
not be established, as accurate data were onlyobtained from three individuals of unknowngender. Of these, two moulted in June and one inMarch. However, enough data could be obtainedfor the pre-moult phase, which occurred through-out the year (Fig. 3e).
188 African Zoology Vol. 41, No. 2, October 2006
Fig. 3. The seasonality of the moult. In pups (a), breeding females (b), non-breeding females (c) and bulls (d);e, seasonality of the juvenile pre-moult.

By contrast, sufficient data were obtained tostudy seasonality in adults. Moults were normallydistributed over time (1 sample K–S test: non-parturients, P = 0.22; parturients, P = 0.81; bulls,P = 0.92). Moult in females occurred throughoutthe year, but peaked in different periods depend-ing on their breeding status (Fig. 3b,c). The peakfor non-parturient females was recorded in May,while the peak for parturients took place in Febru-ary. The moult in bulls occurred throughout theyear, with a peak in April (Fig. 3d). However, thedifferences observed between the various groupswere not statistically significant.
DISCUSSIONMediterranean monk seal pups, like those of mostother Phocids, are born with their foetal pelage(lanugo). This pelage is dark and begins to bemoulted to a shorter, greyish pelage approxi-mately 50 d after birth. The moult is not related toweaning, which is protracted and occurs 120 dafter birth (Aguilar et al. 2002). This contrasts withother species, including the congeneric Hawaiianmonk seal, in which both events are strictly associ-ated (Bowen 1991). Two other Phocid speciesmoult the lanugo before weaning: hooded seals,Cystophora cristata, and harbour seals (Phocavitulina). Harbour seals moult their foetal coatin utero, so pups are born with the new pelage. Asthe insulating layer of air trapped within thelanugo disappears when the pelage is wet, thepre-weaning moult and prenatal fat depositionenables pups to minimize heat loss when enteringthe water soon after birth (Oftedal et al. 1991).Owing to their cave-dwelling habits, Mediterra-nean monk seal pups are also forced to enter thesea soon after birth, particularly during high tidesor when the high swell reaches the bottom of thecaves (Gazo et al. 2000). However, pups do notappear to be physically prepared for this. Pupmortality rates are high, mainly during thepre-moult period (Gazo et al. 2000).
The moult of the lanugo follows the same patternin both sexes, although the timing is quite different.Females start to moult earlier and the wholemoulting process is faster than in males. A delay inmale pups completing the moult has also beendescribed in northern elephant seals (Miroungaangustirostris), another Monachini species. In thiscase, it was viewed as one of male pups’ strategiesto avoid detection when they steal milk from nurs-ing females. This strategy would benefit maleweaners in terms of survival and future reproduc-
tive success (Reiter et al. 1978). However, thisexplanation cannot be applied to the Mediterra-nean monk seal as lanugo moulting is not associ-ated with weaning (see above). Besides, themoulting process is slower in pups of both sexesborn in the autumn/winter. This could be attrib-uted to a higher metabolic demand for tempera-ture regulation in winter that makes the moultprocess slower.
Unlike other pinnipeds and any other mammal,all the moults after the lanugo moult in members ofthe Monachini tribe [northern and southernelephant seals (Mirounga leonina), and monk seals]consist in shedding large sheets of cornified epi-dermis (Ling 1970). These moults are preceded bya pre-moult stage, which is commonly identifiedas the time between the last haul-out and the onsetof shedding. However, a clear-cut identification ofthis stage is unfeasible in the Mediterranean monkseal, as individuals continue to swim while moult-ing. The moult in Monachini is traumatic (theepidermis is shed). During the phase we callpre-moult, the physiological changes may be moredrastic than in other phocids and hence morepatent. In other Monachini, like the elephant seals,the pre-moult phase has not been seen to last aslong as in Mediterranean monk seals. This ispresumably because elephant seals, which aremigrant species, only haul-out when a moult isimminent. During this stage, the pelage acquires abrownish colour. In juveniles, it is covered withsmall black spots, mainly around the eyes andsnout, similar to descriptions of some northernelephant seal yearlings that exhibit a scabby moult(Le Boeuf & Laws 1994). This brownish colorationwas attributed to a deposition of iron oxide precip-itates in previous studies (Samaranch & González2000). However, our observations suggest that itcorresponds to a degradation of the pre-moultingskin. This has also been described in closely relatedspecies that are not exposed to iron deposits, likethe Hawaiian monk seal (Kenyon & Rice 1959;Ching 1994) and the southern elephant seal (Boydet al. 1993).
The Mediterranean monk seal’s shedding processis similar to that of the congeneric Hawaiian monkseal in that the old coat peels back and falls away inpatches (Kenyon & Rice 1959). However, in theHawaiian monk seal, the moult starts on the bellyand ends on the middle of the back (Ching 1994).The Mediterranean monk seal’s moult starts onthe face and flippers and ends on the sides of thetrunk, like the southern elephant seal (Ling &
Badosa et al.: Moult in the Mediterranean monk seal 189

Bryden 1981). The Mediterranean monk seal atCap Blanc spends more time in the water than itscongeneric species, due to the combination of cavedwelling and high tides. Cap Blanc seals spendsome hours in the water at least twice a day. Thefriction produced by the water may explain whythe skin of the head, back and hind flippers shedsearlier than that of the Hawaiian monk seal.
Phocid seals usually spend long periods on landto increase peripheral blood irrigation and thusaccelerate the moult (Boily 1995), and to enhancevitamin D synthesis through solar radiation(Mc Laren 1958; Moulton et al. 2000). However,moulting in the water does not seem to cause aproblem in the Mediterranean monk seal. Thus,the duration of the moult process (14 d) is notmuch longer than that of the Hawaiian monk seal(8–9 d) (Johnson & Johnson 1981). It is similar tothat of southern elephant seal females (7–14 d)(Boyd et al. 1993; Fedak et al. 1994), and evenshorter than that of the northern elephant seal(28 d) (Worthy et al. 1992).
Intermoult periods in non-parturient femalesand bulls appear to be regular and close toone year. By contrast, this period is variable inparturient females, ranging from 365–429 days.The intermoult period in females that haveabruptly lost their pups appears to be equivalentto that of non-parturient females, i.e. approxi-mately one year. If the pup is successfully weaned,the female’s intermoult period becomes longer.This delay in moulting in lactating females can beattributed to the high energy demand of lactation(Oftedal et al. 1987). It has also been observed inthe conspecific Hawaiian monk seal (Johanos et al.1994). However, it appears that moulting cannotbe postponed beyond a certain threshold, probablybecause the following reproductive cycle needs tostart. This is supported by the fact that at leastthree females with long lactation periods wereobserved moulting while still nursing their pups.Reproduction in phocid seals is characterized by aperiod of delayed implantation. The onset ofactive gestation is associated with the end of themoult (Ling 1970). The reproductive cycle in theMediterranean monk seal is annual and quiterigid, with individual females giving birth at ap-proximately same date (±15 d) year after year(Pastor & Aguilar 2003). Therefore, it is not surpris-ing that there is a limit to the length of time themoult can be delayed.
In males, the process of developing the pelagepattern of bulls appears to occur gradually. It
involves at least two annual moults and culmi-nates in significant changes in morphology,pelage colour and behaviour. These changes areassociated with the acquisition of sexual maturity.According to a single observation, the process maybe completed by the age of four years, a figureconsistent with other phocids (Atkinson 1997).
Moulted Mediterranean monk seals show asilvery dark grey coloration, similar to the fullymoulted Hawaiian monk seals (Kenyon & Rice1959; Ching 1994), southern elephant seals (Boydet al. 1993) and northern elephant seals (Worthyet al. 1992). In northern elephant seals, this stage isonly detectable during one or two weeks as thenew hair grows fast (Worthy et al. 1992). By contrast,the post-moult phase in the Mediterranean monkseal can be identified for a longer period (up to54 d), presumably because the new pelage growsat a slower rate.
Moulting in the Mediterranean monk seal at CapBlanc was observed throughout the year, as wereother events in the biological cycle (Pastor &Aguilar 2003). The subtropical Hawaiian monkseal also presents a protracted moulting period,although it is shorter (from April–December)(Kenyon & Rice 1959; Johanos et al. 1994; Ching1994). The slight peak of moults in breedingfemales (in February) occurs four months laterthan the October peak in births (Gazo et al. 1999),as in the Hawaiian monk seal (Johanos et al. 1994)and the grey seal (Bonner 1981). Like other species(Bonner 1981; Thompson & Rothery 1987; Reeveset al. 1992; Green et al. 1995), this peak occurs earlierthan that of bulls (in April). Neither peak coincideswith that of pupping [October (Gazo et al. 1999)]nor with matings [December (Pastor & Aguilar2003)]. In Greece, the Mediterranean monk sealmoults from at least January–May (Yediler et al.1993), and, in Turkey, from October–January(Güçlüsoy & Savas 2003). This also indicates thatthe eastern Mediterranean population has anextremely protracted moulting season. It is likelythat the relatively mild variations in the environ-mental conditions throughout the year contributeto the overall lack of seasonality in the biologicalcycle, as previously suggested by Pastor & Aguilar2003.
ACKNOWLEDGEMENTSWe thank all colleagues and assisting personnel
who contributed to the fieldwork and made thisstudy possible. Special thanks are due to: EsteveGrau, Roser Samaranch, Gemma Cantos and
190 African Zoology Vol. 41, No. 2, October 2006

Sonia Rey. This study was funded by the Euro-pean Commission LIFE project B4-3200/94/0000/D2 and B4-3200/96/510, and by the FDS Foun-dation. T. Pastor also received a grant from theUniversity of Barcelona’s Vicerectorat de Recercaduring the study.
REFERENCESAGUILAR, A. 1999. Status of Mediterranean Monk Seal
(Monachus monachus) Populations. Aloès Editions,Tunis.
AGUILAR, A., CAPPOZZO, H.L., GAZO, M., PASTOR, T.& GRAU, E. 2002. Lactation and mother–pup behav-ior in the Mediterranean monk seal, Monachusmonachus: an unusual pattern for a phocid. Report tothe DG ENV European Commission. University ofBarcelona.
ATKINSON, S. 1997. Reproductive biology of seals. Jour-nal of Reproduction and Fertility 2: 175–194.
BADOSA, E., GRAU, E., APARICIO, F., LAYNA, J.F. &CEDENILLA, M.A. 1998. Individual variation andsexual dimorphism of colouration in Mediterraneanmonk seal pups (Monachus monachus). MarineMammal Science 14: 390–393.
BOILY, P. 1995. Theoretical heat flux in water and habitatselection of phocid seals and beluga whales duringthe annual molt. Journal of Theoretical Biology 172:235–244.
BONNER, W.N. 1981. Grey seal Halichoerus grypusFabricius, 1791. In: Handbook of Marine Mammals (eds)S.H. Ridgway & R.J. Harrison, pp. 111–144. AcademicPress, London.
BOWEN, W.D. 1991. Behavioural ecology of pinnipedneonates. In: The Behaviour of Pinnipeds, (ed.)D. Renouf, pp. 66–127. Chapman & Hall, London.
BOYD, I., ARNBOM, T. & FEDAK, M. 1993. Water flux,body composition, and metabolic rate during molt infemale southern elephant seals (Mirounga leonina).Physiological Zoology 66: 43–60.
CHING, P. 1994. The Hawaiian Monk Seal. University ofHawaii Press, Honolulu.
FEDAK, M.A., ARNBON, T.A., McCONELL, B.J., CHAM-BERS, C., BOYD, I.L., HARWOOD, J. & McCANN,T.S. 1994. Expenditure, investment, and acquisition ofenergy in southern elephant seals. In: Elephant Seals,(eds) B.J. Le Boeuf & R.M. Laws, pp. 354–373.University of California Press, Berkeley.
FORCADA, J. & AGUILAR, A. 2000. Use of photographicidentification in capture–recapture studies of Medi-terranean monk seals. Marine Mammal Science 16:767–793.
GAZO, M., APARICIO, F., CEDENILLA, M.A., LAYNA,J.F. & GONZÁLEZ, L.M. 2000. Pup survival in theMediterranean monk seal (Monachus monachus)colony at Cabo Blanco Peninsula (Western Sahara –Mauritania). Marine Mammal Science 16: 158–168.
GAZO, M., APARICIO, F., CEDENILLA, M.A., LAYNA,J.F., GONZÁLEZ, L.M. & AGUILAR, A. 1999. Puppingseason, perinatal sex ratio and natality rates of theMediterranean monk seal (Monachus monachus) fromthe Cabo Blanco colony. Journal of Zoology (London)249: 393–401.
GREEN, K., BURTON, H.R., WONG, V., McFARLANE,R.A., FLAHERTY, A.A., PAHL, B.C. & HAIGH, S.A.1995. Difficulties in assessing population status of iceseals. Wildlife Research 22: 193–199.
GÜÇLÜSOY, H. & SAVAS, Y. 2003. Status of the Mediter-ranean monk seal, Monachus monachus, in the Foçapilot monk seal conservation area, Turkey. Zoology inthe Middle East 28: 5–16.
IUCN 1996. Red List of Threatened Animals. Gland,Switzerland.
JOHANOS, T.C., BECKER, B.L. & RAGEN, T.J. 1994.Annual reproductive cycle of the female Hawaiianmonk seal (Monachus schauislandi). Marine MammalScience 10: 13–30.
JOHNSON, B.W. & JOHNSON, P.A. 1981. Estimating theHawaiian monk seal on Laysan Island. Final Reportto U.S. Marine Mammal Commission. National Tech-nical Information Service, Bethesda, Maryland.
KENYON, K.W. & RICE, W.R. 1959. Life history of theHawaiian monk seal. Pacific Science 13: 243–252.
LE BOEUF, B.J. & LAWS, R.M. 1994. Elephant seals: anintroduction to the genus. In: Elephant Seals. Popula-tion Ecology, Behavior, and Physiology, (eds) B.J. LeBoeuf & R.M. Laws, pp. 1–26. University of CaliforniaPress, Berkeley.
LING, J.K. 1970. Pelage and molting in wild mammalswith special reference to aquatic forms. QuarterlyReview of Biology 45: 16–54.
LING, J.K. & BRYDEN, M.M. 1981. Southern elephantseal, Mirounga leonina Linnaeus, 1758. In: Handbook ofMarine Mammals, vol. 2: Seals, (eds) S.H. Ridgway &R.J. Harrison, pp. 297–327. Academic Press, London.
MARCHESSAUX, D. 1989. Recherches sur la biologie,l’écologie et le statut du phoque moine, Monachusmonachus. Ph.D. thesis, Université d’Aix-Marseille II,Marseille, France.
Mc LAREN, I.A. 1958. The biology of the ringed seal(Phoca hispida Schreber) in the eastern CanadianArctic. Bulletin of the Fisheries Research Board of Canada118: 1–97.
MOULTON, V.D., MILLER, E.H. & OCHOA-ACUÑA, H.2000. Haul out behaviour of captive harp seals(Pagophilus groenlandicus): incidence, seasonality andrelationships to weather. Applied Animal BehaviourScience 65: 367–378.
OFTEDAL, O.T., BONESS, D.J. & TEDMAN, R.A. 1987.The behavior, physiology, and anatomy of lactationin the pinnipedia. In: Current Mammalogy, (ed.)H.H. Genoways, pp. 175–245. Plenum PublishingCorporation, London.
OFTEDAL, O.T., BOWEN, W.D., WIDDOWSON, E.M. &BONESS, D.J. 1991. The prenatal molt and its ecologi-cal significance in hooded and harbor seals. CanadianJournal of Zoology 69: 2489–2493.
PASTOR, T. & AGUILAR, A. 2003. Reproductive cycle ofthe female Mediterranean monk seal in the westernSahara. Marine Mammal Science 19: 318–330.
REEVES, R.R., STEWART, B.S. & LEATHERWOOD, S.1992. Seals and Sirenians. Sierra Club Books, SanFrancisco.
REITER, J., STINSON, L. & LE BOEUF, B.J. 1978. North-ern elephant seal development: the transition fromweaning to nutritional independence. Behavioral
Badosa et al.: Moult in the Mediterranean monk seal 191

Ecology and Sociobiology 3: 337–367.SAMARANCH, R. & GONZÁLEZ, L.M. 2000. Changes
in morphology with age in Mediterranean monkseals (Monachus monachus). Marine Mammal Science16: 141–157.
THOMPSON, P. & ROTHERY, P. 1987. Age and sex differ-ences in the timing of moult in the common seal,Phoca vitulina. Journal of Zoology (London) 212: 597–603.
WORTHY, G.A., MORRIS, P.A., COSTA, D.P. & LEBOEUF, B.J. 1992. Molt energetics of northernelephant seal (Mirounga angustirotris). Journal ofZoology (London) 227: 257–265.
YEDILER, A., PANOU, A. & SCHRAMEL, P. 1993. Heavymetals in hair samples of the Mediterranean monkseal (Monachus monachus). Marine Pollution Bulletin 26:156–159.
192 African Zoology Vol. 41, No. 2, October 2006
Related Documents











