Morphology and molecular phylogeny of Amphidiniopsis rotundata sp. nov. (Peridiniales, Dinophyceae), a benthic marine dinoflagellate MONA HOPPENRATH 1,3 *, MARINA SELINA 2 ,AIKA YAMAGUCHI 3 AND BRIAN LEANDER 3 1 Senckenberg Research Institute, German Centre for Marine Biodiversity Research, Su ¨dstrand 44, D-26382 Wilhelmshaven, Germany 2 A. V. Zhirmunsky Institute of Marine Biology FEB RAS, Far Eastern Federal University,Vladivostok 690041, Russia 3 University of British Columbia, Departments of Botany and Zoology, #3529-6270 University Boulevard, Vancouver, BC V6T 1Z4, Canada HOPPENRATH M., SELINA M., YAMAGUCHI A. AND LEANDER B. 2012. Morphology and molecular phylogeny of Amphidiniopsis rotundata sp. nov. (Peridiniales, Dinophyceae), a benthic marine dinoflagellate. Phycologia 51: 157–167. DOI: 10.2216/11-35.1 A new dinoflagellate species within the benthic, heterotrophic, and thecate genus Amphidiniopsis was discovered, independently, in sediment samples taken on opposite sides of the Pacific Ocean: (1) the Vancouver area, Canada, and (2) Vostok Bay, the Sea of Japan, Russia. The cell morphology was characterized using light and scanning electron microscopy, and the phylogenetic position of this species was inferred from small-subunit ribosomal DNA sequences. The thecal plate pattern [formula: apical pore complex 49 3a 70 5c 5(6)s 5- 2-9] and ornamentation, as well as the general cell shape without an apical hook or posterior spines, demonstrated that this taxon is different from all other described species within the genus. Amphidiniopsis rotundata sp. nov. was dorsoventrally flattened, 24.5–38.5 mm long, 22.6– 32.5 mm wide. The sulcus was characteristically curved and shifted to the left of the ventral side of the cell. This work presents the first molecular study including a representative of the genus Amphidiniopsis, and led us to propose a new combination, Amphidiniopsis dragescoi comb. nov. (formerly Thecadinium dragescoi), and also suggests a close relationship between Amphidiniopsis, Herdmania (another benthic genus), and Archaeperidinium minutum (a planktonic species). KEY WORDS: Amphidiniopsis, Archaeperidinium, Herdmania, Northeast Pacific, Sand-dwelling, Taxonomy, Sea of Japan, SSU rDNA, Thecadinium dragescoi INTRODUCTION Amphidiniopsis was introduced by Woloszyn ´ska (1928) with the type species A. kofoidii. The history of records, nomenclatural changes and classification schemes of Amphidiniopsis species were summarized in Hoppenrath (2000a) and Hoppenrath et al. (2009a). The genus includes 14 validly described species: A. aculeata Hoppenrath, Koeman & Leander, A. arenaria Hoppenrath, A. cristata Hoppenrath, A. dentata Hoppenrath, A. galericulata Hoppenrath, A. hexagona Yoshimatsu, Toriumi & Dodge, A. hirsutum (Balech) Dodge, A. kofoidii Woloszynska, A. korewalensis Murray & Patterson, A. pectinaria Toriumi, Yoshimatu & Dodge, A. sibbaldii Nicholls, A. swedmarkii (Balech) Dodge, A. urnaeformis Gail 1950, and A. uroensis Toriumi, Yoshimatu & Dodge (Woloszyn ´ska 1928; Gail 1950; Balech 1956; Dodge 1982; Nicholls 1998, 1999; Hoppenrath 2000a; Yoshimatsu et al. 2000; Murray & Patterson 2002; Toriumi et al. 2002; Hoppenrath et al. 2009a). All of these species are heterotrophic sand dwellers with diverse cell morphologies, including morphological variability within the same species (Bursa 1963; Selina & Hoppenrath 2008). Species are laterally or dorsoventrally flattened, with a complete or incomplete cingulum, and with or without an apical hook (e.g. Balech 1956; Dodge 1982; Hoppenrath 2000a; Yoshimatsu et al. 2000; Murray & Patterson 2002; Toriumi et al. 2002). Currently, Amphidiniopsis is characterized by an ascending cingulum, a distinctive curved sulcus and hypothecal plate pattern (Hoppenrath et al. 2009a). Because the genus contains a great deal of morphological heterogeneity, it is possible that Amphidiniopsis contains a pattern of subclades that represent several different ‘genus-level’ taxa. This hypoth- esis can be tested with molecular phylogenetic data, which were unavailable before this study. A revision of the genus is also needed, but to do so, the type species (A. kofoidii) needs to be defined unambiguously and reinvestigated, especially specimens from the type locality. Similarities between Amphidiniopsis and other genera and the system- atic placement of this genus within the order Peridiniales have been discussed in Hoppenrath (2000a). Comparative morphology and molecular phylogenetic analyses suggest that Thecadinium dragescoi Balech is distinct from, and only distantly related to, Thecadinium Kofoid & Skogsberg and may instead be a member of Amphidiniopsis (Hoppenrath et al. 2004). It has been shown repeatedly that patterns of thecal plates can be interpreted differently, and that this affects the plate formula and of course the discussion about species similarities (e.g. Murray & Patterson 2002; Hoppenrath et al. 2004; Hoppenrath & Selina 2006). The heterogeneity within the genus and the absence of molecular phylogenetic information made the * Corresponding author ([email protected]). Phycologia (2012) Volume 51 (2), 157–167 Published 12 March 2012 157

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Morphology and molecular phylogeny of Amphidiniopsis rotundata sp. nov.
(Peridiniales, Dinophyceae), a benthic marine dinoflagellate
MONA HOPPENRATH1,3*, MARINA SELINA
2, AIKA YAMAGUCHI3
AND BRIAN LEANDER3
1Senckenberg Research Institute, German Centre for Marine Biodiversity Research, Sudstrand 44,
D-26382 Wilhelmshaven, Germany2A. V. Zhirmunsky Institute of Marine Biology FEB RAS, Far Eastern Federal University,Vladivostok 690041, Russia3University of British Columbia, Departments of Botany and Zoology, #3529-6270 University Boulevard, Vancouver,
BC V6T 1Z4, Canada
HOPPENRATH M., SELINA M., YAMAGUCHI A. AND LEANDER B. 2012. Morphology and molecular phylogeny ofAmphidiniopsis rotundata sp. nov. (Peridiniales, Dinophyceae), a benthic marine dinoflagellate. Phycologia 51: 157–167.DOI: 10.2216/11-35.1
A new dinoflagellate species within the benthic, heterotrophic, and thecate genus Amphidiniopsis was discovered,independently, in sediment samples taken on opposite sides of the Pacific Ocean: (1) the Vancouver area, Canada, and(2) Vostok Bay, the Sea of Japan, Russia. The cell morphology was characterized using light and scanning electronmicroscopy, and the phylogenetic position of this species was inferred from small-subunit ribosomal DNA sequences.The thecal plate pattern [formula: apical pore complex 49 3a 70 5c 5(6)s 5- 2-9] and ornamentation, as well as the generalcell shape without an apical hook or posterior spines, demonstrated that this taxon is different from all other describedspecies within the genus. Amphidiniopsis rotundata sp. nov. was dorsoventrally flattened, 24.5–38.5 mm long, 22.6–32.5 mm wide. The sulcus was characteristically curved and shifted to the left of the ventral side of the cell. This workpresents the first molecular study including a representative of the genus Amphidiniopsis, and led us to propose a newcombination, Amphidiniopsis dragescoi comb. nov. (formerly Thecadinium dragescoi), and also suggests a closerelationship between Amphidiniopsis, Herdmania (another benthic genus), and Archaeperidinium minutum (a planktonicspecies).
KEY WORDS: Amphidiniopsis, Archaeperidinium, Herdmania, Northeast Pacific, Sand-dwelling, Taxonomy, Sea ofJapan, SSU rDNA, Thecadinium dragescoi
INTRODUCTION
Amphidiniopsis was introduced by Wołoszynska (1928) with
the type species A. kofoidii. The history of records,
nomenclatural changes and classification schemes of
Amphidiniopsis species were summarized in Hoppenrath
(2000a) and Hoppenrath et al. (2009a). The genus includes
14 validly described species: A. aculeata Hoppenrath,
Koeman & Leander, A. arenaria Hoppenrath, A. cristata
Hoppenrath, A. dentata Hoppenrath, A. galericulata
Hoppenrath, A. hexagona Yoshimatsu, Toriumi & Dodge,
A. hirsutum (Balech) Dodge, A. kofoidii Woloszynska, A.
korewalensis Murray & Patterson, A. pectinaria Toriumi,
Yoshimatu & Dodge, A. sibbaldii Nicholls, A. swedmarkii
(Balech) Dodge, A. urnaeformis Gail 1950, and A. uroensis
Toriumi, Yoshimatu & Dodge (Wołoszynska 1928; Gail
1950; Balech 1956; Dodge 1982; Nicholls 1998, 1999;
Hoppenrath 2000a; Yoshimatsu et al. 2000; Murray &
Patterson 2002; Toriumi et al. 2002; Hoppenrath et al.
2009a). All of these species are heterotrophic sand dwellers
with diverse cell morphologies, including morphological
variability within the same species (Bursa 1963; Selina &
Hoppenrath 2008). Species are laterally or dorsoventrally
flattened, with a complete or incomplete cingulum, and
with or without an apical hook (e.g. Balech 1956; Dodge
1982; Hoppenrath 2000a; Yoshimatsu et al. 2000; Murray
& Patterson 2002; Toriumi et al. 2002). Currently,
Amphidiniopsis is characterized by an ascending cingulum,
a distinctive curved sulcus and hypothecal plate pattern
(Hoppenrath et al. 2009a). Because the genus contains a
great deal of morphological heterogeneity, it is possible that
Amphidiniopsis contains a pattern of subclades that
represent several different ‘genus-level’ taxa. This hypoth-
esis can be tested with molecular phylogenetic data, which
were unavailable before this study. A revision of the genus
is also needed, but to do so, the type species (A. kofoidii)
needs to be defined unambiguously and reinvestigated,
especially specimens from the type locality. Similarities
between Amphidiniopsis and other genera and the system-
atic placement of this genus within the order Peridiniales
have been discussed in Hoppenrath (2000a).
Comparative morphology and molecular phylogenetic
analyses suggest that Thecadinium dragescoi Balech is
distinct from, and only distantly related to, Thecadinium
Kofoid & Skogsberg and may instead be a member of
Amphidiniopsis (Hoppenrath et al. 2004). It has been shown
repeatedly that patterns of thecal plates can be interpreted
differently, and that this affects the plate formula and of
course the discussion about species similarities (e.g. Murray
& Patterson 2002; Hoppenrath et al. 2004; Hoppenrath &
Selina 2006). The heterogeneity within the genus and the
absence of molecular phylogenetic information made the* Corresponding author ([email protected]).
Phycologia (2012) Volume 51 (2), 157–167 Published 12 March 2012
157

transfer of T. dragescoi into Amphidiniopsis tenuous at best
(e.g. Hoppenrath et al. 2004; Yamaguchi et al. 2011).
However, recently reported molecular phylogenetic data
demonstrated a novel clade comprised of T. dragescoi,
Herdmania litoralis Dodge emend. Hoppenrath, and
Archaeperidinium minutum (Kofoid) Jorgensen (Yamaguchi
et al. 2011). This is consistent with the suggestion that H.
litoralis should be placed in the order Peridiniales, despite
the unusual tabulation pattern that is difficult to categorize
(Hoppenrath 2000b).
In the present study, an unidentifiable species was
recognized as a member of Amphidiniopsis, as currently
circumscribed, and was described as a new species from two
localities on opposite sides of the Pacific Ocean: Canada
and Russia. The new species is characterized with light and
scanning electron microscopy and with molecular phyloge-
netic analyses of the first small-subunit (SSU) ribosomal
(r)DNA sequence for any Amphidiniopsis species.
MATERIAL AND METHODS
Sampling
In Canada, sand samples were collected with a spoon
during low tide at Centennial Beach, Boundary Bay, BC,
Canada from April 2005 to August 2007. The sand samples
were transported directly to the laboratory, and the
flagellates were separated from the sand by extraction
through a fine filter (mesh size 45 mm) using melting
seawater ice (Uhlig 1964). The flagellates that accumulated
in a Petri dish beneath the filter were identified at the light
microscope level. This sample was used as a source of
isolated cells of this new species (see below).
In Russia, sampling was carried out in Vostok Bay in the
Sea of Japan (42u539130N, 132u439360E). The samples were
collected from May 2006 to December 2006 and from May
2007 to January 2008. The upper 5-cm sand layer was
collected under water using a square reservoir at 0.5–1.0-m
depth. Filtered seawater was added to the sand, which was
then thoroughly agitated. The resultant suspension was
subjected to two stages of filtration through fine gauze
(mesh size of 150 and 80 mm, respectively). The sample was
then concentrated by filtration through 20-mm mesh-size
gauze. A 20–80-mm fraction was collected in this manner.
The concentrated sample was fixed with Lugol’s solution.
Light microscopy
For Canadian material, living cells were observed using a
Leica DMIL inverted microscope (Wetzlar, Germany) and
isolated by micropipetting. These were further character-
ized and images captured at the light microscope level using
differential intereference contrast optics on a Zeiss Ax-
ioplan 2 imaging microscope (Carl-Zeiss, Oberkochen,
Germany) connected to a Leica DC500 color digital camera
(Wetzlar, Germany).
Isolated fixed cells of the Russian material were
transferred onto a glass slide and observed directly with
an Olympus BX41 light microscope (Tokyo, Japan). Images
were captured using an Olympus C2020Z digital camera
Figs 1–8. Light micrographs of Amphidiniopsis rotundata sp. nov.Figs 1–4, 8. Living cells from Canada.Figs 5–7. Lugol-fixed cells from Russia.
Figs 1–4. Through focus of a single cell. Note ventral sulcus (Fig. 1), single pusule (p) (Figs 2, 3), and nucleus (Figs 3, 4).Figs 5, 6. Through focus of sulcus.Fig. 7. Large central nucleus (n).Fig. 8. A cell with two pusules (p). Scale bars 5 10 mm.
158 Phycologia, Vol. 51 (2), 2012

(Tokyo, Japan). Cell dimensions were measured for 10 cells
at 3640 magnification using an ocular micrometer.
Scanning electron microscopy
Aliquots of the agitated sand samples were fixed overnight
with two drops of acidic Lugol’s solution. Cells were
transferred onto a 5-mm polycarbonate membrane filter
(Corning Separations Div., Acton, MA), washed with
distilled water, dehydrated with a graded series of ethanol
and critical-point-dried with CO2. Filters were mounted on
stubs, sputter-coated with gold and viewed using an Hitachi
S4700 scanning electron microscope (SEM). Some SEM
images were presented on a black background using Adobe
Photoshop 6.0 (Adobe Systems, San Jose, CA). Cell sizes
were calculated from calibrated SEM images.
The SEM stubs with the natural samples used in this
study have been deposited at the Senckenberg Research
Institute and Natural History Museum in the Centre of
Excellence for Dinophyte Taxonomy, Wilhelmshaven,
Germany, with the designations CEDiT2011H11 and
CEDiT2011RM12.
Figs 9–16. Scanning electron micrographs of Amphidiniopsis rotundata sp. nov. (Figs 9–11, 14–16 stub CEDiT2011H11 and Figs 12, 13 stubCEDiT2011RM12, stored at the Centre of Excellence for Dinophyte Taxonomy, Wilhelmshaven, Germany).
Figs 9–11. Ventral views. Note the marginal listlike structure (arrow) of the Sp-plate and its generally prominent, spinelike marginal listat the border to the Ss-plate (arrowhead).Figs 12, 13. Dorsal views. Note the variable lengths of the plate border between 2a and 3a (arrow) and the beginning and end of thefourth cingular plate (4c) (arrowheads).Fig. 14. Apical view. Note the border between 2a and 3a plates (arrow).Fig. 15. Antapical view.Fig. 16. Detail of the sulcal plates. Scale bars 5 10 mm.
Hoppenrath et al.: Amphidiniopsis rotundata sp. nov. 159

DNA extraction, polymerase chain reaction (PCR) amplifi-
cation, sequencing, molecular phylogenetic analyses
Thirty-four living cells were isolated by micropipetting for
DNA extraction (Canadian sample from 18 June 2007).
The cells were manually isolated and washed three times in
sterile f/2-medium. Genomic DNA was extracted using the
MasterPure Complete DNA and RNA purification kit
(EPICENTRE, Madison, WI, USA). The SSU rDNA
sequence was PCR amplified using puReTaq Ready-to-go
PCR beads (GE Healthcare, Quebec, Canada), with an
error rate of 1 per 20,000–40,000 bases, and universal
eukaryotic primers (PF1-R4, as reported previously in
Hoppenrath et al. 2009b). The PCR product of the expected
size was gel isolated and cloned into pCR2.1 vector using a
TOPO TA cloning kit (Invitrogen Corporation, CA, USA).
One clone was completely sequenced with ABI big-dye
reaction mix using both vector primers and two internal
primers oriented in both directions (acc. no. AB639343).
The sequence identity was evaluated initially by BLAST
and then by phylogenetic analyses.
The new SSU rDNA sequence was aligned with other
dinoflagellate/alveolate sequences using MacClade 4 (Mad-
dison & Maddison 2000), forming a 63-taxon alignment.
The alignment is available on request. To find the model of
evolution that fits the data set for the maximum likelihood
Figs 17–20. Scanning electron micrographs of Amphidiniopsis rotundata sp. nov. (stub CEDiT2011H11 stored at the Centre of Excellence forDinophyte Taxonomy, Wilhelmshaven, Germany).
Fig. 17. Apical view of all epithecal plates. Scale bar 5 10 mm.Fig. 18. Dorsal view of the epitheca. Scale bar 5 10 mm.Fig. 19. Outside view of the apical pore complex with ventral platelet (asterisk). Scale bar 5 1 mm.Fig. 20. Inside view of the apical pore complex with ventral platelet (asterisk) and opening of the apical pore. Scale bar 5 1 mm.
160 Phycologia, Vol. 51 (2), 2012

(ML) analysis, we used the FindModel server (http://www.
hiv.lanl.gov/content/sequence/findmodel/findmodel.html).
The general time reversible (GTR) plus gamma model of
nucleotide substitution was chosen as the best-fitting model.
ML analyses were performed using PhyML (Guindon &
Gascuel 2003). The starting tree was generated by BIONJ
with optimization of topology, branch lengths, and rate
parameters selected. The GTR model of nucleotide
substitution was chosen and the gamma distribution
parameter and proportion of variable rates was estimated
from the original data set. Eight categories of substitution
rates were selected. PhyML bootstrap analysis was
conducted with the same parameters described above (100
pseudoreplicates; one heuristic search per pseudoreplicate).
The alignment was also analyzed with Bayesian methods
using the MrBayes 3.1.2 (Huelsenbeck & Ronquist 2001).
For Baysian analysis, the TIM2+I+G model was chosen as
the best-fitting substitution rate model using jModeltest
0.1.1 (Posada 2008). This model was not implemented in
MrBayes 3.1.2. Because the next more-complex model
available is recommended in such situations the GTR
model with a gamma distribution and four Monte Carlo–
Markov chains starting from a random tree was used. A
total of 5,500,000 generations was calculated with trees
sampled every 1000 generations. The first 1375 trees in each
run were discarded as burn-in. Posterior probabilities
correspond to the frequency at which a given node was
found in the postburn-in trees.
RESULTS
Amphidiniopsis rotundata Hoppenrath & Selina sp. nov.
Figs 1–21
Ambitus cellulae rotundatus cum margine laevi, 24.8–
38.5 mm longae; 22.6–32.5 mm latae; et 18.0–21.5 mm altae.
Fig. 21. Line drawings of Amphidiniopsis rotundata sp. nov. showing the tabulation pattern.
Table 1. Plate formulae of morphologically similar, dorsoventrally flattened Amphidiniopsis species, ‘Thecadinium’ dragescoi andHerdmania litoralis.
APC 9 a 0 x c s - p -9
Amphidiniopsis APC 3–4 1–3 6–8 3–8 3–5 5 2A. rotundata APC 4 3 7 5 5(6) 5 2A. korewalensis APC 4 3 6 x 4? 4 6 2
New interp.1 APC 4 3 6 4? 4 5 2
A. pectinaria APC 4 3 7 3 4 5 2
New interp. APC 4 3 6 3 4 5 2
A. uroensis2 APC 3 3 6 3 4 5 2‘T.’ dragescoi3 APC 5 2 8 x 6(7) ? 4 2 1
4 APC 4 3 7 x 6(7) ? 5 2This study APC 4 3 7 6(7) 4? 4 2
H. litoralis5 APC 3 2 6 x 7 3 6 1 16 APC 4 3 7 x ? 3? 6 26 APC 4 3 7 ? 5–6? 5 2This study APC 4 3 7 5? 4–5? 5 2
1 New interp., our new interpretation of the tabulation (of the images provided in the original description).2 Same tabulation as in the original description but with different interpretation in the sulcal area (see Fig. 26).3 Hoppenrath 2000c.4 Hoppenrath et al. 2004.5 Hoppenrath 2000b.6 Yamaguchi et al. 2011.
Hoppenrath et al.: Amphidiniopsis rotundata sp. nov. 161

162 Phycologia, Vol. 51 (2), 2012

Cellulae dorsoventraliter complanatae. Tabulatio thecarum
laminarium: (APC) 49, 3a, 70, 5c, 5(6)s, 5-, 2-9. Laminae
thecae pusticulatae. Cingulum circa dimidio latitudinem suam
ascendens. Sulcus sinistorsum partisventralis cellulae transla-
tus. Nucleus in centro cellulae positus.
Cells 24.8–38.5 mm long, 22.6–32.5 mm wide, and 18.0–
21.5 mm deep, rounded with smooth outline, dorsoventrally
flattened. Plate formula: apical pore complex (APC) 49 3a
70 5c 5(6)s 5- 2-9. Pusticulate thecal ornamentation.
Cingulum ascending about half a cingular width. Sulcus
shifted to the left of the ventral cell side. Nucleus in cell
centre.
HABITAT: Marine, benthic (sand-dwelling).
HOLOTYPE: Fig. 9, SEM stub CEDiT2011H11.
ICONOTYPE/ISOTYPE: Fig. 21.
TYPE LOCALITY: Centennial Beach, Boundary Bay, British
Columbia, Canada (49u0.09N, 123u8.09W).
ETYMOLOGY: From Latin rotundus 5 more or less round/
circular; referring to the cell shape in ventral or dorsal view.
Morphology
Cells were rounded, dorsoventrally flattened, 24.8–38.5 mm
long (Canada: 24.8–38.5 mm, n 5 14; Russia 25.2–32.5 mm,
n 5 14), 22.6–32.5 mm wide (Canada: 22.6–32.5 mm, n 5 14;
Russia 23.4–30.1 mm, n 5 14) and 18.0–21.5 mm deep
(Canada: 18.9–21.5 mm, n 5 2; Russia 18.0–20.0 mm, n 5 4)
(Figs 1–15). The smaller epitheca was slightly narrower
than the hypotheca and was caplike (Figs 2–8). The sulcus
was distinctively curved and shifted to the left of the ventral
side of the cell (Figs 1, 5, 9–11). The cingulum was slightly
ascending about half a cingular width (Figs 1, 2, 5, 6, 9–11,
16). The large nucleus (n) was oval, located in the cell centre
in the upper half of the hyposome (Figs 4, 7). One or two
pusules (p) were visible in living cells (Figs 2, 3, 8). Cells
were colourless, and no food particles were observed. The
cells were characteristically smooth and somewhat globular
in outline without any apical hook or posterior spines/
protrusions (Figs 1–8). A ventral spinelike structure was
not obvious in any view under the light microscope.
Eleutheroschisis – daughter cell formation inside the parent
theca with new theca formation – was observed only once
and could not be documented.
The epithecal and hypothecal plates were all pusticulate,
ornamented with many small dots (Figs 9–18). The density
of dots was variable. Many large pores with a slightly raised
rim were randomly distributed over the epithecal and
hypothecal plates (Figs 9–18). Rows of pores tended to be
arranged along the precingular and postcingular margins
of the precingular and postcingular plates respectively
(Figs 9–14, 17, 18). A pluglike structure was visible inside
these pores in some cells (Figs 17, 18). A second size class of
pores was not observed. Epithecal and hypothecal plates
with growth bands were irregular, with only a very slightly
raised marginal listlike structure at the start of the growth
zone (Figs 9–11, 14, 15, 17, 18). This structure was more
developed into a real list at the margin of the posterior
sulcal plate that contacted the second antapical plate
(Figs 9, 10). The right and posterior sulcal plates were
ornamented like the epi- and hypothecal plates (Figs 9–11,
15, 16). The other sulcal plates were smooth with pores
(Figs 9–11, 16). The cingular plates were smooth without,
or only with a few, scattered pores (Figs 9–13, 18). The
sutures were narrow lines (Figs 12, 13), smooth with
relatively regular outline (Figs 9, 10, 17, 18), or broad with
an irregular outline (Figs 11, 14, 15).
The plate formula for this species was APC 49 3a 70 5c
5(6)s 5- 2-9 (Figs 9–18, 21; Table 1). The epitheca consisted
of 15 plates (Figs 9–14, 17, 18, 21). The oval to droplike
APC was centrally located and surrounded by a flange/list
(Figs 14, 17–20). The apical pore was a tiny hole in the
centre of a slightly raised, nearly circular central area
(Figs 17, 18), and was hidden behind a cover layer (Figs 14,
19). The apical pore was open to the inside of the apical
pore plate (Fig. 20). A ventrally located darker round area
reminiscent of a huge pore at low magnification (Fig. 14)
transpired to be a deeper-set platelet (Figs 17, 19, 20). Thus,
as described in other dinoflagellate species, the APC was
composed of two platelets, the pore plate (Po) and a small
canal plate (X). The APC was encircled by four apical
plates (Figs 17, 21), with plate 19 in contact with the sulcus
(Figs 9–11, 21) and sometimes disconnected from the canal
plate (ventral platelet) of the APC (Fig. 19). There were
three anterior intercalary plates (syn. apical intercalary
plate, epithecal intercalary plate) in a series (Figs 14, 17, 18,
21). The plate border between 2a and 3a was variable in
length (Figs 12–14, 17, 18). There were seven precingular
plates (Figs 9–14, 17, 18, 21). The slightly ascending
cingulum consisted of five smooth plates (Figs 9–13, 18,
21), but the sutures were difficult to discern. The deepened
part of the sulcus was shifted markedly to the left cell side
(Figs 9–11), and the complete sulcus was covering most of
the left ventral hypotheca with its right margin in about
mid-ventral cell position. Five sulcal plates with ornamen-
tation, as described above, were identified (Figs 9–11, 15,
16, 21). The right anterior sulcal plate (Sad) was seven
sided, relatively small and slightly elongated (Figs 10, 16).
The left anterior sulcal plate (Sas) was difficult to recognize
in most cells because it was partly hidden behind a list
formed by Sad; however, Sas can be clearly seen in cells
with rudimentarily developed sulcal lists (Fig. 16). The
broad right sulcal plate (Sd) was positioned in the middle of
the sulcus, and the left margin of this plate was
characteristically curved with a large smooth list (Figs 9–
11, 21). The left sulcal plate (Ss) was elongated and
characteristically curved (Figs 9–11, 16, 21) with a list along
part of its left margin formed by the first postcingular plate.
r
Fig. 22. Maximum-likelihood tree of dinoflagellates inferred from SSU rDNA sequences. The grey box indicates the Amphidiniopsis clade.ML bootstrap values (ML) over 50 and Bayesian posterior probabilities (PP) over 0.90 are shown at the nodes (ML/PP). The scale barrepresents inferred evolutionary distance in changes/site. The DNA sequence generated in the study is indicated by a black box.
Hoppenrath et al.: Amphidiniopsis rotundata sp. nov. 163

The posterior sulcal plate (Sp) reached the antapex (Figs 9–
11, 15) and formed a marginal list at the border to the Ss
plate that could reach a prominent size and spinelike shape
(Figs 9–11, 16). A mid-sulcal plate (Sm) could not be
identified unambiguously because this plate was positioned
near the centre of the sulcus behind the Sd plate; however, a
tiny left part of Sm was probably recorded in Fig. 16. The
flagella pore(s) were obscured from view. The hypotheca
consisted of seven plates: five postcingular and two
antapical plates (Figs 9–13, 16, 21). The fifth (5-)
postcingular plate was large and covered most of the right
ventral side of the hypotheca (Figs 9–11, 21). The medium-
sized, asymmetrical, third postcingular plate (3-) was
pentagonal and only slightly pointed into a right posterior
direction (Figs 12, 13, 21). The two antapical plates were
asymmetrical (Figs 12, 13, 15, 21).
Occurrence
The new species was recorded at Centennial Beach,
Boundary Bay, BC, Canada: 16 March, 26 April, 9 May,
6 June, 12 July, 4 August, 14 October 2005; 31 May 2006;
17 and 31 May, 18 June, 31 July, and 28 August 2007. It has
been seen in a sample from Parksville, Vancouver Island,
Canada on 18 September 2005. In most samples A.
rotundata occurred in very low cell densities. No sampling
took place during the winter months in Canada.
The new species was also recorded in Vostok Bay, Sea of
Japan, Russia: 20 August 2006; 28 May, 27 June, 12 July,
21 August, 4 and 22 September and 12 December 2007.
Cells in a sand sample from the island of Sylt (List
Harbor), German Bight, North Sea taken on 27 March
2009 and in a sand sample from the island of Wangerooge,
German Bight, North Sea taken on 22 April 2010 were also
identified at the light microscope level as A. rotundata.
Molecular phylogeny
The phylogenetic analyses of the 63-taxon alignment of
SSU rDNA sequence placed A. rotundata within a clade of
the Peridiniales together with species of Protoperidinium
and Diplopsalis group. Subclade IV was strongly supported
and consisted of A. rotundata, T. dragescoi, Archaeperidi-
nium minutum, and H. litoralis (Fig. 22). The closest sister
lineage to Amphidiniopsis rotundata was T. dragescoi with
moderate to high support values (ML bootstrap: 73%;
Bayesian posterior probability: 0.99) (Fig. 22). Archaeper-
idinium minutum formed the nearest sister lineage to the
Amphidiniopsis rotundata/T. dragescoi clade, but this
relationship lacked statistical support (Fig. 22). Thecadi-
nium dragescoi was only distantly related to the clade
comprised of T. kofoidii (Herdman) Larsen (the type) and
T. yashimaense Yoshimatsu, Toriumi & Dodge (Fig. 22).
DISCUSSION
Molecular data for the genus Amphidiniopsis are largely
unavailable and so the current considerations are restricted
to morphological data. The genus is characterized by an
ascending cingulum and a distinctive sulcus morphology
(curved) and hypothecal plate pattern (Hoppenrath et al.
2009a). The new species described here has these features
and is therefore identified as a member of this genus. Three
major morphological subgroups can be recognized within
the genus: (1) laterally flattened species with a complete
cingulum; (2) dorsoventrally flattened species with a
complete cingulum, a sulcus positioned in the middle of
the cell, no apical hook, and one or two anterior intercalary
plates; and (3) dorsoventrally flattened species with a
complete or incomplete cingulum, sulcus positioned in the
middle; the deepened sulcal part can be shifted to the cell’s
left side, with an apical hook pointing to the left, and with
three anterior intercalary plates. The new species was also
dorsoventrally flattened, which limits it to being a member
of either subgroup 2 or 3 above.
The first dorsoventrally flattened subgroup as described above
comprises species with one (A. swedmarkii) or two [A. aculeata,
A. hirsutum, A. hexagona, A. sp. (‘swedmarkii’)] anterior
intercalary plates (Hoppenrath et al. 2009a). Amphidiniopsis
aculeata is ornamented with spines (Hoppenrath et al.
2009a), Amphidiniopsis hirsutum possesses a row of irregular
spines at its posterior end and A. swedmarkii has two
antapical spines (Balech 1956; Hoppenrath 2000a). Amphi-
diniopsis rotundata has no antapical spines of any kind, like
the two remaining species of this group A. hexagona and A.
sp. (‘swedmarkii’). In both of these last species, the deep part
of the sulcus is not markedly-shifted to the left side of the cell
as in A. rotundata (Figs 9–11), and they differ in details of the
epithecal plates; both have only two anterior intercalary
plates and the second apical plate of A. sp. (‘swedmarkii’) is in
contact with the sulcus (Yoshimatsu et al. 2000; Hoppenrath
et al. 2009a). Within this group, the shape and size of the
third postcingular plate (3-) is characteristic for the species
(Hoppenrath et al. 2009a). The 3- plate of A. rotundata is of
medium size (in relation to the other hypothecal plates),
asymmetrical, and only slightly pointed into a right posterior
direction (Fig. 21). The relative size of that plate in A.
rotundata is similar to that of A. sp. (‘swedmarkii’), but it is
more asymmetrical, and the shape of the plate is more similar
to the 3- plate of A. hirsutum. However, the shortest side of
this plate in A. rotundata is the right posterior side, whereas
in A. hirsutum, it is the left posterior side (side-reversed
morphology) (Hoppenrath et al. 2009a). The new species is
different from all these species by its relative sulcus location
and by having three anterior intercalary plates.
The second subgroup of dorsoventrally flattened species
as described above comprises taxa with a complete (A.
pectinaria, A. uroensis) or incomplete (A. korewalensis)
cingulum (Murray & Patterson 2002; Toriumi et al. 2002;
Figs 25–30). The deepened part of their sulcus can be
markedly shifted to the cell’s left side, as in A. korewalensis
(Murray & Patterson 2002), moderately shifted as in A.
pectinaria (Toriumi et al. 2002), or only slightly shifted
as in A. uroensis (Toriumi et al. 2002). Interpreting the
plates differently from the original description (Murray &
Patterson 2002), A. korewalensis seems to have an extremely
wide sulcus covering most of the ventral side of the cell
(Fig. 25, Table 1). The three species have very similar thecal
plate patterns; however, A. uroensis has only three apical
plates (39) instead of four (49). The 3- plate is similar in all
three species, and clearly points posteriorly (Figs 26, 28,
164 Phycologia, Vol. 51 (2), 2012
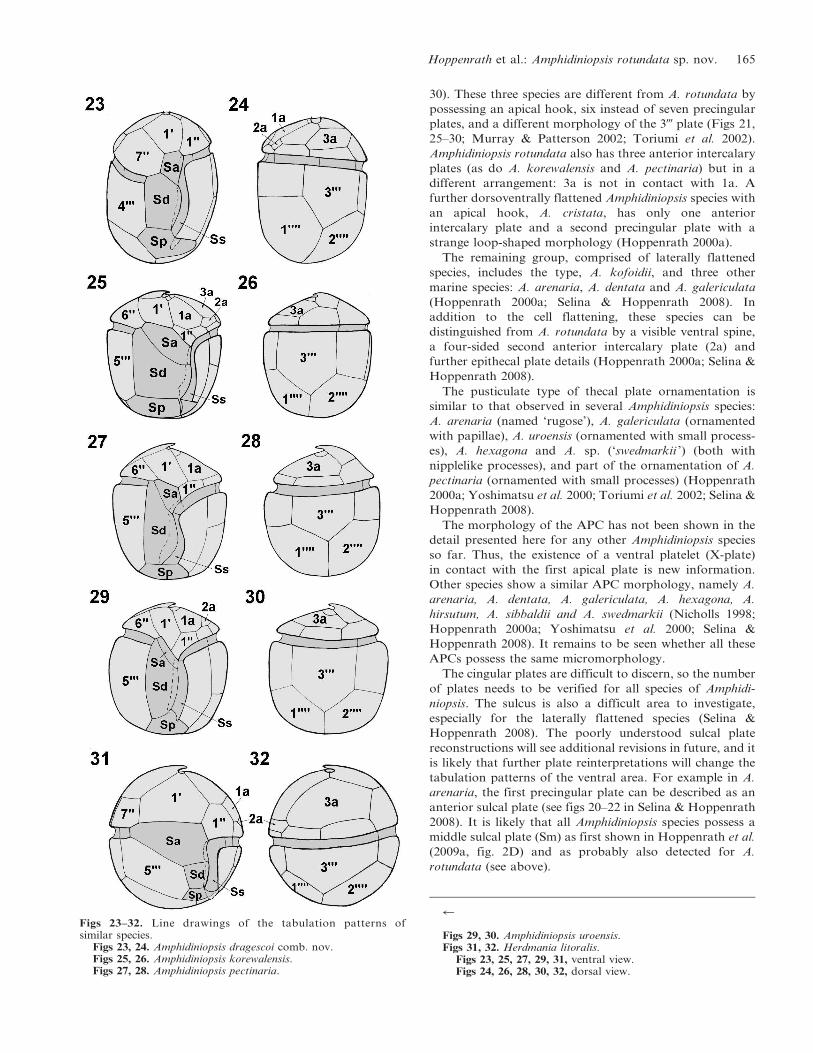
30). These three species are different from A. rotundata by
possessing an apical hook, six instead of seven precingular
plates, and a different morphology of the 3- plate (Figs 21,
25–30; Murray & Patterson 2002; Toriumi et al. 2002).
Amphidiniopsis rotundata also has three anterior intercalary
plates (as do A. korewalensis and A. pectinaria) but in a
different arrangement: 3a is not in contact with 1a. A
further dorsoventrally flattened Amphidiniopsis species with
an apical hook, A. cristata, has only one anterior
intercalary plate and a second precingular plate with a
strange loop-shaped morphology (Hoppenrath 2000a).
The remaining group, comprised of laterally flattened
species, includes the type, A. kofoidii, and three other
marine species: A. arenaria, A. dentata and A. galericulata
(Hoppenrath 2000a; Selina & Hoppenrath 2008). In
addition to the cell flattening, these species can be
distinguished from A. rotundata by a visible ventral spine,
a four-sided second anterior intercalary plate (2a) and
further epithecal plate details (Hoppenrath 2000a; Selina &
Hoppenrath 2008).
The pusticulate type of thecal plate ornamentation is
similar to that observed in several Amphidiniopsis species:
A. arenaria (named ‘rugose’), A. galericulata (ornamented
with papillae), A. uroensis (ornamented with small process-
es), A. hexagona and A. sp. (‘swedmarkii ’) (both with
nipplelike processes), and part of the ornamentation of A.
pectinaria (ornamented with small processes) (Hoppenrath
2000a; Yoshimatsu et al. 2000; Toriumi et al. 2002; Selina &
Hoppenrath 2008).
The morphology of the APC has not been shown in the
detail presented here for any other Amphidiniopsis species
so far. Thus, the existence of a ventral platelet (X-plate)
in contact with the first apical plate is new information.
Other species show a similar APC morphology, namely A.
arenaria, A. dentata, A. galericulata, A. hexagona, A.
hirsutum, A. sibbaldii and A. swedmarkii (Nicholls 1998;
Hoppenrath 2000a; Yoshimatsu et al. 2000; Selina &
Hoppenrath 2008). It remains to be seen whether all these
APCs possess the same micromorphology.
The cingular plates are difficult to discern, so the number
of plates needs to be verified for all species of Amphidi-
niopsis. The sulcus is also a difficult area to investigate,
especially for the laterally flattened species (Selina &
Hoppenrath 2008). The poorly understood sulcal plate
reconstructions will see additional revisions in future, and it
is likely that further plate reinterpretations will change the
tabulation patterns of the ventral area. For example in A.
arenaria, the first precingular plate can be described as an
anterior sulcal plate (see figs 20–22 in Selina & Hoppenrath
2008). It is likely that all Amphidiniopsis species possess a
middle sulcal plate (Sm) as first shown in Hoppenrath et al.
(2009a, fig. 2D) and as probably also detected for A.
rotundata (see above).
Figs 23–32. Line drawings of the tabulation patterns ofsimilar species.
Figs 23, 24. Amphidiniopsis dragescoi comb. nov.Figs 25, 26. Amphidiniopsis korewalensis.Figs 27, 28. Amphidiniopsis pectinaria.
r
Figs 29, 30. Amphidiniopsis uroensis.Figs 31, 32. Herdmania litoralis.
Figs 23, 25, 27, 29, 31, ventral view.Figs 24, 26, 28, 30, 32, dorsal view.
Hoppenrath et al.: Amphidiniopsis rotundata sp. nov. 165

In conclusion, according to all available morphological
data (see discussion above), the species described here
belongs to the genus Amphidiniopsis and is new to science.
The molecular phylogenetic data showed T. dragescoi as
the sister taxon to A. rotundata (Fig. 22). Hoppenrath
(2000c) reinvestigated T. dragescoi, amended the original
description of Balech (1956), and concluded that the species
does not belong to this genus. Molecular phylogenetic
analyses suggested later that T. dragescoi is distinct from,
and only distantly related to, other species of Thecadinium
and may be a member of Amphidiniopsis (Hoppenrath et al.
2004; Saldarriaga et al. 2004; Yamaguchi et al. 2011). The
sister relationship between T. dragescoi and A. rotundata is
now obvious from improved interpretations of the tabula-
tion (Figs 21, 23, 24, Table 1) and molecular phylogenetic
data. The deep part of the sulcus in T. dragescoi is shifted to
the cell’s left side as in A. rotundata (Hoppenrath 2000c, figs
10, 63), and the sulcal construction is Amphidiniopsis-like.
Thecadinium dragescoi is slightly dorsoventrally flattened
and has four apical plates, three anterior intercalary plates
and seven precingular plates like the first subgroup of
dorsoventrally flattened Amphidiniopsis species mentioned
above. The APC in T. dragescoi has the same morphology
as described for A. rotundata and many other Amphidi-
niopsis species (Hoppenrath 2000c). The third postcingular
plate of T. dragescoi is of the typical pentagonal posterior-
pointing shape (Fig. 24) and the 2a plate is square like that
described for laterally flattened Amphidiniopsis species
(Hoppenrath 2000c). There are also some differences
between T. dragescoi and all the other species, like a 3a
plate that is separated from 1a and 2a (Fig. 24), only four
postcingular plates (Fig. 24), and a descending cingulum –
which is the most problematic feature. In summary, the
data supporting the transfer of T. dragescoi into the genus
Amphidiniopsis outweigh the arguments for retaining it in
Thecadinium (see taxonomic appendix below). A revision of
the genus Amphidiniopsis will take time because additional
new species have been discovered and need to be formally
described, and the type species is in need of thorough
reinvestigation. Additional deliberation on the misclassifi-
cation of T. dragescoi at this juncture is inappropriate in
our opinion.
The molecular analysis placed Archaeperidinium minutum
as a close relative of the Amphidiniopsis clade, which is
difficult to interpret because morphological characters
could not be identified that unify these taxa (Yamaguchi
et al. 2011). Herdmania is also a close relative to the
Amphidiniopsis clade (Fig. 22). Herdmania litoralis has a
deepened sulcus part shifted strongly to the left ventral side
of the cell, as in A. rotundata and A. dragescoi comb. nov.
(Hoppenrath 2000b; Yamaguchi et al. 2011). Herdmania
also has an apical hook and three anterior intercalary
plates, as described for some Amphidiniopsis species. The
cingulum was described as being incomplete, but reinter-
preting the ventral plates brings the thecal morphology
closer to Amphidiniopsis species (Figs 31, 32, Table 1).
Herdmania also has an ascending cingulum (Hoppenrath
2000b; Yamaguchi et al. 2011). The hypothesis of an
evolutionary relationship between the genera Amphidiniop-
sis and Herdmania is strongly-supported by phylogenetic
analyses of both molecular and morphological data.
Taxonomic appendix
Amphidiniopsis dragescoi (Balech) Hoppenrath, Selina,
Yamaguchi & Leander comb. nov.
BASIONYM: Thecadinium dragescoi Balech 1956, Revue
Algologique 2: p. 37–40, figs 26–33.
FURTHER REFERENCE: Hoppenrath 2000, p. 101, figs 9, 10,
23–25, 61–69, as Thecadinium dragescoi.
EXCLUDED REFERENCE: Thecadinium dragescoi sensu Paul-
mier 1992, p. 60, pl. 70, figs 5, 6.
ACKNOWLEDGEMENTS
We thank Drs N. Chomerat, IFREMER Concarneau,
and A. Coute, Museum National d’Histoire Naturelle
Paris, France, for the translation of the description into
Latin. This work was partly supported by a scholarship to
MH from the Deutsche Forschungsgemeinschaft (grant
Ho3267/1-1), postdoctoral research salary to MH and AY
from the Assembling the Tree of Life grant (NSF #EF-
0629624) and operating funds to BL from the National
Science and Engineering Research Council of Canada
(NSERC 283091-09) and the Canadian Institute for
Advanced Research, Programs in Evolutionary Biology
and Integrated Microbial Biodiversity. MS got grants
from Russian Government No. 11.G34.31.0010 and FEB
RAS 09-III-A-06-211.
REFERENCES
BALECH E. 1956. Etude des dinoflagelles du sable de Roscoff.Revue Algologique 2: 29–52.
BURSA A. 1963. Phytoplankton in coastal waters of the ArcticOcean at Point Barrow, Alaska. Arctic 4: 239–262.
DODGE J.D. 1982. Marine dinoflagellates of the British Isles. HerMajesty’s Stationery Office, London. 249 pp.
GAIL G.I. 1950. Manual for identification of the phytoplankton ofthe Sea of Japan. Transactions of the Pacific Research Institutefor Fisheries and Oceanography 33: 1–177. (In Russian).
GUINDON S. & GASCUEL O. 2003. PhyML – a simple, fast, andaccurate algorithm to estimate large phylogenies by maximumlikelihood. Systematic Biology 52: 696–704.
HOPPENRATH M. 2000a. Morphology and taxonomy of six marinesand-dwelling Amphidiniopsis species (Dinophyceae, Peridi-niales), four of them new, from the German Bight, North Sea.Phycologia 39: 482–497.
HOPPENRATH M. 2000b. An emended description of Herdmanialitoralis Dodge (Dinophyceae) including the plate formula. NovaHedwigia 71: 481–489.
HOPPENRATH M. 2000c. Morphology and taxonomy of the marinesand-dwelling genus Thecadinium (Dinophyceae), with thedescription of two new species from the North German WaddenSea. Phycologia 39: 96–108.
HOPPENRATH M. & SELINA M. 2006. Pseudothecadinium campbelliigen. nov. et sp. nov. (Dinophyceae), a phototrophic, thecate,marine planktonic species found in the Sea of Okhotsk, Russia.Phycologia 45: 260–269.
HOPPENRATH M., SALDARRIAGA J.F., SCHWEIKERT M., ELBRACHTER
M. & TAYLOR F.J.R. 2004. Description of Thecadinium mucosumsp. nov. (Dinophyceae), a new sand-dwelling marine dinoflagel-late, and an emended description of Thecadinium inclinatumBalech. Journal of Phycology 40: 946–961.
166 Phycologia, Vol. 51 (2), 2012

HOPPENRATH M., KOEMAN R.P.T. & LEANDER B.S. 2009a.Morphology and taxonomy of a new marine sand-dwellingAmphidiniopsis species (Dinophyceae, Peridiniales), A. aculeatasp. nov., from Cap Feret, France. Marine Biodiversity 39: 1–7.
HOPPENRATH M., BACHVAROFF T.R., HANDY S.M., DELWICHE C.F.& LEANDER B.S. 2009b. Molecular phylogeny of ocelloid-bearingdinoflagellates (Warnowiaceae) as inferred from SSU and LSUrDNA sequences. BMC Evolutionary Biology 9: 116 (15 pages).
HUELSENBECK J.P. & RONQUIST F. 2001. MRBAYES: Bayesianinference of phylogenetic trees. Bioinformatics 17: 754–755.
MADDISON D.R. & MADDISON W.P. 2000. MacClade 4: analysis ofphylogeny and character evolution version 4.0. Sinauer Associates,Sunderland, MA.
MURRAY S. & PATTERSON D.J. 2002. Amphidiniopsis korewalensissp. nov., a new heterotrophic benthic dinoflagellate. Phycologia41: 382–388.
NICHOLLS K.H. 1998. Amphidiniopsis sibbaldii sp. nov. (Thecadi-niaceae, Dinophyceae), a new freshwater sand-dwelling dinofla-gellate. Phycologia 37: 334–339.
NICHOLLS K.H. 1999. Validation of Amphidiniopsis sibbaldiiNicholls (Dinophyceae). Phycologia 38: 74.
PAULMIER G. 1992. Catalogue illustre des microphytes planctoniqueset benthiques des Cotes Normandes. Rapports internes de laDirection des Ressources Vivantes de l‘IFREMER, 108 pp.
POSADA D. 2008. jModelTest: phylogenetic model averaging.Molecular Biology and Evolution 25: 1253–1256.
SALDARRIAGA J.F., TAYLOR F.J.R., CAVALIER-SMITH T., MENDEN-DEUER S. & KEELING P.J. 2004. Molecular data and theevolutionary history of the dinoflagellates. European Journal ofProtistology 40: 85–111.
SELINA M. & HOPPENRATH M. 2008. An emended description ofAmphidiniopsis arenaria Hoppenrath 2000, based on materialfrom the Sea of Japan. European Journal of Protistology 44:71–79.
TORIUMI S., YOSHIMATSU S. & DODGE J.D. 2002. Amphidiniopsisuroensis sp. nov. and Amphidiniopsis pectinaria sp. nov.(Dinophyceae): two new benthic dinoflagellates from Japan.Phycological Research 50: 115–124.
UHLIG G. 1964. Eine einfach Methode zur Extraktion der vagilen,mesopsammalen Mikrofauna. Helgolander WissenschaftlicheMeeresuntersuchungen 11: 178–185.
WOŁOSZYNSKA J. 1928. Dinoflagellatae der polnischen Ostsee sowieder an Piasnica gelegenen Sumpfe. Archiv d’Hydrobiologied’Ichtyologie 3: 153–278.
YAMAGUCHI A., HOPPENRATH M., POSPELOVA V., HORIGUCHI T. &LEANDER B.S. 2011. Molecular phylogeny of the marinesand-dwelling dinoflagellate Herdmania litoralis and anemended description of the closely related planktonic genusArchaeperidinium Jorgensen. European Journal of Phycology 46:98–112.
YOSHIMATSU S., TORIUMI S. & DODGE J.D. 2000. Light andscanning microscopy of two benthic species of Amphidiniopsis(Dinophyceae), Amphidiniopsis hexagona sp. nov. and Amphidi-niopsis swedmarkii from Japan. Phycological Research 48:107–113.
Received 23 March 2011; accepted 25 July 2011
Associate editor: Stuart Sym
Hoppenrath et al.: Amphidiniopsis rotundata sp. nov. 167
Related Documents











