Mutation Research 770 (2014) 105–111 Contents lists available at ScienceDirect Mutation Research/Genetic Toxicology and Environmental Mutagenesis jo ur nal home page: www.elsevier.com/locate/gentox Comm unit y ad dress: www.elsevier.com/locate/mutres Molecular instability induced by aluminum stress in Plantago species Sofia Correia a,∗ , Manuela Matos a , Vanessa Ferreira a , Neusa Martins b , Sandra Gonc ¸ alves b , Anabela Romano b , Olinda Pinto-Carnide a a Institute of Biotechnology and Bioengineering, Centre of Genomics and Biotechnology, University of Trás-os-Montes and Alto Douro (IBB/CGB-UTAD), P.O. Box 1013, 5000-801 Vila Real, Portugal b Institute of Biotechnology and Bioengineering, Centre of Genomics and Biotechnology (IBB/CGB), Faculty of Sciences and Technology, University of Algarve, Gambelas Campus, Ed. 8, 8005-139 Faro, Portugal a r t i c l e i n f o Article history: Received 7 March 2014 Received in revised form 20 May 2014 Accepted 9 June 2014 Available online 16 June 2014 Key-words: Aluminum Plantago Genotoxic effects Genomic template stability (GTS) Inter Simple Sequence Repeat (ISSR) a b s t r a c t Aluminum (Al) is one of the most abundant metals on earth’s crust and Al toxicity represents one of the major factors that limit plant growth and productivity in acid soils (with a pH ≤ 5.0). In this study the mutagenic/genotoxic effects of Al were evaluated in roots and leaves of two Plantago, species, Plantago almogravensis and Plantago lagopus, using ISSRs markers. Both species were exposed to 400 M Al during 7 and 21 days. Ten ISSR primers produced polymorphic bands. In P. almogravensis, a total of 257 and 258 bands in roots and 255 and 265 bands in leaves were produced in the presence and absence of Al, respectively. In P. lagopus were produced 279 and 278 a total bands in roots and 275 and 274 bands in leaves, under the same conditions. The changes in ISSR profiles after Al treatment were considered as gain and/or loss of bands compared with the controls. The results suggest that changes in genomic template stability (GTS) could be detected with ISSR profiles. This molecular marker proved to be a good tool to detect the effects of Al on DNA profiles. It seems that Al did not interfere significantly with DNA integrity in both species but generated less ISSR stability in P. almogravensis than in P. lagopus. The results confirm the tolerance of P. almogravensis and suggest the same behavior of P. lagopus. Although further studies are required for confirmation the Al tolerance behavior of P. lagopus, a potential application for phytoremediation can be also considered due its wide distribution. © 2014 Elsevier B.V. All rights reserved. 1. Introduction Soil contains an average of 7% total aluminum (Al), which repre- sents the third most abundant element in the earth’s crust [1,2]. Al constitutes a phytotoxic barrier to crop production and its toxicity is one of the major factors that limit plant growth and productivity in acid soils [1,3,4]. Acid soils account for 30–40% of the Word’s arable land and are spreading due to natural, agricultural and industrial processes [5]. Although Al naturally exists as an insoluble silicate and oxide at neutral or weakly acid pH, the problem becomes par- ticularly severe when the soil pH drops below 5.0 because Al is solubilized into the most phytotoxic form Al 3+ . Al rapidly affects a number of cellular processes in the root apex, which is considered to be the primary target of Al stress, caus- ing an inhibition of elongation and lateral root formation as the first symptoms of Al toxicity in plants [6,7]. This response indi- cates that Al quickly disrupts root cell elongation, which leads ∗ Corresponding author. Tel.: +351 933496732. E-mail addresses: sofi[email protected], sofi[email protected] (S. Correia). to the decrease in water and nutrient uptake, and eventually to limited growth of the whole plant due to cell division inhibition [8,9]. Al also affects a wide range of physiological processes. At cellular level, the strong binding affinity of Al with oxygen donor ligands, such as proteins, nucleic acids and phospholipids, results in the inhibition of cell division, cell extension and transport, interfering with cytoskeleton structure and function, phosphorus metabolism and disruption of calcium homeostasis [10]. At the molecular level, Al stress causes major changes in the expression patterns of impor- tant oxidative stress responding genes, which may cause damage to cellular components if antioxidant defenses are overwhelmed [10]. To prevent or avoid Al toxicity, some plant species evolved and developed the ability to exclude Al from plant tissue (Al excluders), or to tolerate high internal Al concentrations (Al accumulators), allowing them to grow in acid soils with a high concentration of Al 3+ . The first mechanism involves the secretion through the root apices of Al-chelating substances, such as organic acid anions, that can form stable complexes with Al which are less phytotoxic than the ionic form, Al 3+ (Al resistance). The other one involves an http://dx.doi.org/10.1016/j.mrgentox.2014.06.002 1383-5718/© 2014 Elsevier B.V. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

M
SAa
Bb
A
a
ARRAA
KAPGGI
1
scoalpats
wific
h1
Mutation Research 770 (2014) 105–111
Contents lists available at ScienceDirect
Mutation Research/Genetic Toxicology andEnvironmental Mutagenesis
jo ur nal home page: www.elsev ier .com/ locate /gentoxComm uni t y ad dress : www.elsev ier .com/ locate /mutres
olecular instability induced by aluminum stress in Plantago species
ofia Correiaa,∗, Manuela Matosa, Vanessa Ferreiraa, Neusa Martinsb, Sandra Gonc alvesb,nabela Romanob, Olinda Pinto-Carnidea
Institute of Biotechnology and Bioengineering, Centre of Genomics and Biotechnology, University of Trás-os-Montes and Alto Douro (IBB/CGB-UTAD), P.O.ox 1013, 5000-801 Vila Real, PortugalInstitute of Biotechnology and Bioengineering, Centre of Genomics and Biotechnology (IBB/CGB), Faculty of Sciences and Technology, University oflgarve, Gambelas Campus, Ed. 8, 8005-139 Faro, Portugal
r t i c l e i n f o
rticle history:eceived 7 March 2014eceived in revised form 20 May 2014ccepted 9 June 2014vailable online 16 June 2014
ey-words:luminumlantagoenotoxic effectsenomic template stability (GTS)
a b s t r a c t
Aluminum (Al) is one of the most abundant metals on earth’s crust and Al toxicity represents one of themajor factors that limit plant growth and productivity in acid soils (with a pH ≤ 5.0). In this study themutagenic/genotoxic effects of Al were evaluated in roots and leaves of two Plantago, species, Plantagoalmogravensis and Plantago lagopus, using ISSRs markers. Both species were exposed to 400 �M Al during7 and 21 days. Ten ISSR primers produced polymorphic bands. In P. almogravensis, a total of 257 and258 bands in roots and 255 and 265 bands in leaves were produced in the presence and absence of Al,respectively. In P. lagopus were produced 279 and 278 a total bands in roots and 275 and 274 bandsin leaves, under the same conditions. The changes in ISSR profiles after Al treatment were consideredas gain and/or loss of bands compared with the controls. The results suggest that changes in genomictemplate stability (GTS) could be detected with ISSR profiles. This molecular marker proved to be a good
nter Simple Sequence Repeat (ISSR) tool to detect the effects of Al on DNA profiles. It seems that Al did not interfere significantly with DNAintegrity in both species but generated less ISSR stability in P. almogravensis than in P. lagopus. The resultsconfirm the tolerance of P. almogravensis and suggest the same behavior of P. lagopus. Although furtherstudies are required for confirmation the Al tolerance behavior of P. lagopus, a potential application forphytoremediation can be also considered due its wide distribution.
© 2014 Elsevier B.V. All rights reserved.
. Introduction
Soil contains an average of 7% total aluminum (Al), which repre-ents the third most abundant element in the earth’s crust [1,2]. Alonstitutes a phytotoxic barrier to crop production and its toxicity isne of the major factors that limit plant growth and productivity incid soils [1,3,4]. Acid soils account for 30–40% of the Word’s arableand and are spreading due to natural, agricultural and industrialrocesses [5]. Although Al naturally exists as an insoluble silicatend oxide at neutral or weakly acid pH, the problem becomes par-icularly severe when the soil pH drops below 5.0 because Al isolubilized into the most phytotoxic form Al3+.
Al rapidly affects a number of cellular processes in the root apex,hich is considered to be the primary target of Al stress, caus-
ng an inhibition of elongation and lateral root formation as therst symptoms of Al toxicity in plants [6,7]. This response indi-ates that Al quickly disrupts root cell elongation, which leads
∗ Corresponding author. Tel.: +351 933496732.E-mail addresses: [email protected], [email protected] (S. Correia).
ttp://dx.doi.org/10.1016/j.mrgentox.2014.06.002383-5718/© 2014 Elsevier B.V. All rights reserved.
to the decrease in water and nutrient uptake, and eventually tolimited growth of the whole plant due to cell division inhibition[8,9].
Al also affects a wide range of physiological processes. At cellularlevel, the strong binding affinity of Al with oxygen donor ligands,such as proteins, nucleic acids and phospholipids, results in theinhibition of cell division, cell extension and transport, interferingwith cytoskeleton structure and function, phosphorus metabolismand disruption of calcium homeostasis [10]. At the molecular level,Al stress causes major changes in the expression patterns of impor-tant oxidative stress responding genes, which may cause damageto cellular components if antioxidant defenses are overwhelmed[10].
To prevent or avoid Al toxicity, some plant species evolved anddeveloped the ability to exclude Al from plant tissue (Al excluders),or to tolerate high internal Al concentrations (Al accumulators),allowing them to grow in acid soils with a high concentration
of Al3+. The first mechanism involves the secretion through theroot apices of Al-chelating substances, such as organic acid anions,that can form stable complexes with Al which are less phytotoxicthan the ionic form, Al3+ (Al resistance). The other one involves an
1 Research 770 (2014) 105–111
oaft
ttu
cettttmp[
daselt
osevg(o[(agtt
cFIitasathipwnaaAtfsrDpTim
Table 1ISSR primers sequence.
Primer Sequence (5′–3′)
UBC 807 (AG)8TUBC 811 (GA)8CUBC 825 (AC)8TUBC 835 (AG)8YCUBC 836 (AG)8YAUBC 844 (CT)8RCUBC 853 (TC)8RTUBC 873 (GACA)4
UBC 880 (GGAGA)3
UBC y889 DBD(AC)7
06 S. Correia et al. / Mutation
rganic-mediated mechanism to detoxify Al internally through Al3+
ccumulation at the simplest either by Al cheating in the cytosol toorm harmless complexes on by sequestering it into organelles (Alolerance) [11,12].
Despite the considerable progress made in decipheringhe physiological and genetic basis of Al toxicity and resis-ance/tolerance, the understanding of the molecular mechanismsnderlying, these processes still remain limited.
The assessment of genotoxic effects of metals in plants, such asadmium, copper, lead, Al and other metals is an important topic innvironmental research and has been paid increasing attention tohis field in the last years [13–16]. Plants are ideal systems for geno-oxicity assays and have been used as a bioindicator to evaluate theoxic effects of pollutants, once they could overcome environmen-al stress by developing efficient and specific physio-biochemical
echanisms, thus providing vital information from the stand-oint of safeguarding biodiversity and ecological resources17,18].
Several studies have been using the comet assay to detect DNAegradation and micronucleus assay or chromosome aberrationssay to measure the genotoxic effect of metals on different plantspecies, namely in lupin [19], tobacco [20] and onion [21]. How-ver, these assays are limited by their relatively low sensitivity andow ability to provide information on the effects of toxic metals athe DNA level [22].
Advances in molecular biology have led to the developmentf selective and sensitive DNA analysis methods [14], making thetudy of the mechanisms involving metal-induced genotoxicityasier and more accessible [16]. Despite the existence of a largeariety of DNA molecular markers that can be used to assess theenotoxicity, the analysis of Random Amplified Polymorphic DNARAPD) has been successfully used to evaluate the mutagenic effectsf several metals, namely in rice [22], barley [14], common bean23] and eggplant [15]. However, Inter Simple Sequence RepeatISSR) assay could be more advantageous since it is described as
more reproducible technique. This molecular marker becomes ofreat importance since the detection of genotoxic effects involveshe comparison of profiles generated from control (unexposed) andreated (exposed) plants.
The current study was designed to detect the impact of Alontamination on two Plantago species, Plantago almogravensisranco and Plantago lagopus L. in terms of DNA damage, usingSSR molecular markers. The genus Plantago belongs to the fam-ly Plantaginaceae and has approximately 483 species [24]. Athe moment, there are no DNA sequences databases availablend are scarce molecular studies in both species. P. almograven-is is endemic from Portugal where only one population is knownnd was described as the only Al-hyperaccumulating species ofhe Plantaginaceae family [25]. On the other hand, P. lagopusas a wide distribution and its usage as a traditional medicine
s mainly due to the antioxidant activity of its extract [26]. Theresence of an apparent B chromosome in P. lagopus (2n = 12)as reported by Dhar et al. [27]. Later on, in 2014, this super-umerary chromosome was described as genetically inert, with
heterochromatic structure, rich in repetitive DNA and transpos-ble elements [28]. However, this species is not known to possessl-hyperaccumulation activity despite showing tolerance to cer-
ain herbicides [29]. Both species can have a potential applicationor phytoremediation. However, this application of P. almograven-is can be limited due to the fact that this species is presently inisk of global extinction and protected under the European Habitatsirective and the Portuguese law. For this reason, currently, these
lant species are being conserved through micropropagation [30].he main goal of this study was to evaluate the genetic instabilitynduced by Al stress in roots and leaves of both species using ISSRarkers.
B = (C, G, T); D = (A, G, T); H = (A, C, T); R = (A, G); V = (A, C, G); Y = (C, T); N = (A, G,C, T).
2. Materials and methods
2.1. Plant material and Al treatment
At least 1000 seeds were randomly collected from 100 individuals of P. almo-gravensis and P. lagopus (50 of each species) in wild populations growing in Odemira(Portuguese Southwest coast) and in the center of Algarve, respectively. These werecombined by species and a total of 40 seeds, 20 from each species, were ran-domly selected and germinated in vitro, and the shoots obtained multiplied in MSmedium [31] containing 0.2 mg L−1 6-benzyladenine [30]. Plantlets were obtainedby cultivating the shoots for 3 weeks in 1/2MS medium [31] containing 0.5 mg L−1
indole-3-acetic acid at 25 ± 2 ◦C, under a light intensity of 69 �mol m−2 s−1, pho-toperiod of 16/8 h (light/dark).
Al treatment was aseptically applied. For that, the plantlets were individuallytransferred to test tubes (32 mm × 200 mm) using filter paper bridges as support.Each tube contained 20 ml autoclaved 1/4 MS liquid medium, with the pH adjustedto 4.0, and 400 �M AlCl3. This concentration of Al was selected based on prelimi-nary assays with a range of Al concentrations (0, 100, 200 and 400 �M) [32]. Thetreatment was applied during 7 days [33,34] and 21 days [29] (Fig. 1). Cultureswere incubated under the conditions described above without changing the culturemedium.
2.2. DNA extraction and ISSR procedures
Were collected after each Al treatment (7 and 21 days) five untreated (con-trol) and five Al-exposed plantlets of each Plantago species. Total DNA wasextracted from 100 mg of leaf and root tissue using the NucleoSpin® Plant II Kit(Macherey-Nagel) according to the manufacturer’s instructions. Concentrations ofthe extracted DNA samples were measured at 260 nm and the purity was estimatedby measuring the 260/280 m absorbance ratio by nanodrop (NanoDrop® ND-1000Spectrophotometer) followed by quality checking in a 1.0% (w/v) agarose gelelectrophoresis.
PCR conditions were optimized according to [35] with some minor modificationsdescribed in Ferreira et al. [36]. Amplification of genomic DNA used standard poly-merase chain reaction (PCR) protocols in 20.0 �L consisting of: 1.0 �L of genomicDNA (5 ng/�L), 2.0 �L of primer 0.5 �M, 1.2 �L of dNTPs (10 mM), 2.0 �L of 10 × PCRbuffer with (NH4)2SO4 (Fermentas), 1.6 �L of MgCl2 (25 mM) and 0.4 �L of Taq DNAPolymerase (5.0 U/�L) (Fermentas). Ten ISSR primers from the UBC#100/9 set (Uni-versity of British Columbia) previously selected by Ferreira et al. [36] were used andtheir sequences are presented in Table 1.
For amplification, the reaction mixtures were denatured 5 min at 94 ◦C followedby 45 cycles 30 s at 94 ◦C, 45 s at 52 ◦C, 2 min at 72 ◦C and a final 10 min extensionat 72 ◦C. All amplifications were conducted twice in order to evaluate the repro-ducibility of the polymorphic bands. PCR products were separated in 1.8% (w/v)agarose gels using TBE buffer, running at 100 V for about 3 h, stained with ethidiumbromide, and the molecular weights were estimated using the GeneRulerTM 100 bpDNA Ladder Plus (Fermentas).
Images were analyzed using the Molecular Image Gel-DocTM XR+ with ImageLabTM Software (BIO RAD), which assigns a fragment size to each band using analgorithm based on the 100 bp DNA ladder.
Three PCR technical replicates were included in the analysis, whichcorrespond to the three replicates developed for each treatment on bothspecies.
2.3. Estimation of genomic template stability
The arbitrary score of +1 was given to each obvious modification observed inthe ISSR patterns (disappearance of bands and/or appearance of new bands), andthe average was calculated for each experimental group (control, 7 and 21 days) ofleaves and roots of each species with the 10 ISSR primers analyzed.

S. Correia et al. / Mutation Research 770 (2014) 105–111 107
F antletr
T
G
w‘pamw[
Gfiua
3
alpod
aaAMsabOd
ig. 1. (A) Plantlets in control conditions: 1 – P. almogravensis; 2 – P. lagopus. (B) Plespectively; 4 and 6 – P. lagopus exposed to Al for 7 and 21 days, respectively.
Changes in the ISSR patterns were expressed as a decrease in GTS (Genomicemplate Stability) values. The GTS was calculated by the formula:
TS (%) =(
1 − a
n
)× 100
here ‘a’ is the average number of polymorphic bands in each treated sample andn’ the number of all bands in control. Polymorphism in ISSR profiles included disap-earance and appearance of new bands in comparison with the control profile. Theverage was calculated for each experimental group exposed to different Al treat-ents The average of the presence/absence bands detected by the ten primers usedere calculated for each experimental group exposed to different Al treatments
15].Dendrograms were constructed by cluster analysis using the Unweighted Pair
roup Method of the Arithmetic Averages (UPGMA) and the Simple Matching Coef-cient (SM) as implemented in the software package NTSYS-pc, version 2.02 g forntreated and treated plantlets of each time of exposition (7 and 21 days) on leavesnd roots of both species [37].
. Results and discussion
Metal toxicity in plants is described as a cause of various dam-ges in growth and development processes such as photosynthesis,ipid metabolism and nucleic acid biosynthesis. However, somelant species have developed the ability to tolerate metals in theirrgans. These plants have attracted much attention for phytoreme-iation purposes [9,38,39].
In the present study we report the application of ISSR markersnalysis for the assessment of Al genotoxicity in P. almogravensisnd P. lagopus. P. almogravensis was previously described as the onlyl-hyperaccumulating species of the Plantaginaceae family [25].oreover, recent physiological and biochemical studies demon-
trated the tolerance of this species to Al stress [32–34]. However, P.
lmogravensis has a limited application to phytoremediation once,esides being a rare species, it has a very narrow distribution.n the other hand, any hyper accumulative behavior was so farescribed in P. lagopus. On this way and taking into account its wides exposed to 400 �M Al: 3 and 5 – P. almogravenis exposed to Al for 7 and 21 days,
distribution throughout the Portuguese territory it was used forcomparison with a known Al-hyperacumulator Plantago species.
The effects of metals are dependent on the age of the plant, timeof exposure, and are accumulated at metabolically inactive sitessuch as cell walls and vacuoles as described by Fodor [40]. In orderto avoid the influence of plant age, all plants were exposed to Alafter 3 weeks in rooting medium to ensure that all plants had thesame age when exposed to Al treatment. One set of control plantsfor each treatment (7 and 21 days of exposure) of each species wasused to evaluate the effect of Al on both species.
This study was performed with fully developed in vitro plantletsand not with seedlings, as the most of the assays described in theliterature [41,42]. During all the treatments, plantlets maintainedtheir morphological characteristics, revealing to be vigorous, with-out showing visible toxic effects, such as chlorotic leaves, brittleor brown roots (Fig. 1). Similar behavior was observed in previousstudies with P. almogravensis plantlets treated with 400 �M AlCl3,for 7 [33,34] and for 21 days [32].
ISSR markers were chosen for this study because they have thesame advantages of RAPDs, that are widely applied to metal eco-toxicological studies [14,15,17,22], and are described as a morereproducible technique than other molecular markers such asRAPDs. PCR in the analysis were included technical replicates toconfirm the reproducibility of each ISSR primer.
Ten ISSR primers produced polymorphic bands. In P. almo-gravensis, a total of 257 and 258 bands in roots and 255 and 265bands in leaves were produced in the presence and absence of Al,respectively. In P. lagopus, 279 and 278 bands in roots and 275and 274 bands in leaves, were produced in the same conditions
(Table 2).Previous studies showed that changes in DNA fingerprint reflectDNA alterations in genome from single base changes (point muta-tions) to complex chromosomal rearrangements [43–45], offering
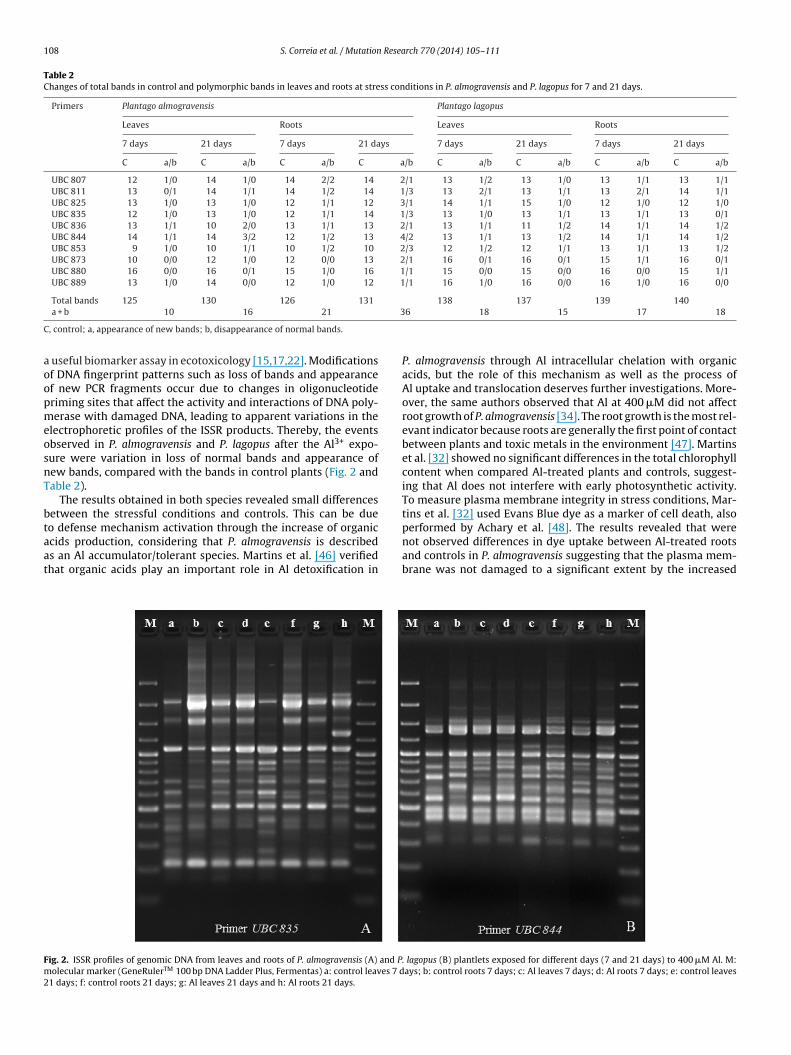
108 S. Correia et al. / Mutation Research 770 (2014) 105–111
Table 2Changes of total bands in control and polymorphic bands in leaves and roots at stress conditions in P. almogravensis and P. lagopus for 7 and 21 days.
Primers Plantago almogravensis Plantago lagopus
Leaves Roots Leaves Roots
7 days 21 days 7 days 21 days 7 days 21 days 7 days 21 days
C a/b C a/b C a/b C a/b C a/b C a/b C a/b C a/b
UBC 807 12 1/0 14 1/0 14 2/2 14 2/1 13 1/2 13 1/0 13 1/1 13 1/1UBC 811 13 0/1 14 1/1 14 1/2 14 1/3 13 2/1 13 1/1 13 2/1 14 1/1UBC 825 13 1/0 13 1/0 12 1/1 12 3/1 14 1/1 15 1/0 12 1/0 12 1/0UBC 835 12 1/0 13 1/0 12 1/1 14 1/3 13 1/0 13 1/1 13 1/1 13 0/1UBC 836 13 1/1 10 2/0 13 1/1 13 2/1 13 1/1 11 1/2 14 1/1 14 1/2UBC 844 14 1/1 14 3/2 12 1/2 13 4/2 13 1/1 13 1/2 14 1/1 14 1/2UBC 853 9 1/0 10 1/1 10 1/2 10 2/3 12 1/2 12 1/1 13 1/1 13 1/2UBC 873 10 0/0 12 1/0 12 0/0 13 2/1 16 0/1 16 0/1 15 1/1 16 0/1UBC 880 16 0/0 16 0/1 15 1/0 16 1/1 15 0/0 15 0/0 16 0/0 15 1/1UBC 889 13 1/0 14 0/0 12 1/0 12 1/1 16 1/0 16 0/0 16 1/0 16 0/0
Total bands 125 130 126 131 138 137 139 1403
C
aoopmeosnT
btaat
Fm2
a + b 10 16 21
, control; a, appearance of new bands; b, disappearance of normal bands.
useful biomarker assay in ecotoxicology [15,17,22]. Modificationsf DNA fingerprint patterns such as loss of bands and appearancef new PCR fragments occur due to changes in oligonucleotideriming sites that affect the activity and interactions of DNA poly-erase with damaged DNA, leading to apparent variations in the
lectrophoretic profiles of the ISSR products. Thereby, the eventsbserved in P. almogravensis and P. lagopus after the Al3+ expo-ure were variation in loss of normal bands and appearance ofew bands, compared with the bands in control plants (Fig. 2 andable 2).
The results obtained in both species revealed small differencesetween the stressful conditions and controls. This can be dueo defense mechanism activation through the increase of organic
cids production, considering that P. almogravensis is describeds an Al accumulator/tolerant species. Martins et al. [46] verifiedhat organic acids play an important role in Al detoxification inig. 2. ISSR profiles of genomic DNA from leaves and roots of P. almogravensis (A) and Polecular marker (GeneRulerTM 100 bp DNA Ladder Plus, Fermentas) a: control leaves 7 d
1 days; f: control roots 21 days; g: Al leaves 21 days and h: Al roots 21 days.
6 18 15 17 18
P. almogravensis through Al intracellular chelation with organicacids, but the role of this mechanism as well as the process ofAl uptake and translocation deserves further investigations. More-over, the same authors observed that Al at 400 �M did not affectroot growth of P. almogravensis [34]. The root growth is the most rel-evant indicator because roots are generally the first point of contactbetween plants and toxic metals in the environment [47]. Martinset al. [32] showed no significant differences in the total chlorophyllcontent when compared Al-treated plants and controls, suggest-ing that Al does not interfere with early photosynthetic activity.To measure plasma membrane integrity in stress conditions, Mar-tins et al. [32] used Evans Blue dye as a marker of cell death, alsoperformed by Achary et al. [48]. The results revealed that were
not observed differences in dye uptake between Al-treated rootsand controls in P. almogravensis suggesting that the plasma mem-brane was not damaged to a significant extent by the increased. lagopus (B) plantlets exposed for different days (7 and 21 days) to 400 �M Al. M:ays; b: control roots 7 days; c: Al leaves 7 days; d: Al roots 7 days; e: control leaves
S. Correia et al. / Mutation Research 770 (2014) 105–111 109
grave
ptt
afp1atCd
ibltt
Fc
Fig. 3. Genomic DNA template stability in leaves and roots of P. almo
roduction of ROS. These results suggest that the plantlets are ableo reduce the impact of oxidative damage under Al stress condi-ions, suggesting the tolerance of P. almogravensis [34].
The polymorphism values, for 7 days of treatment, were 8.00%nd 16.67% in P. almogravensis, and 13.04% and 12.23% in P. lagopusor leaves and roots, respectively. For the 21 days of treatment, theolymorphism observed was 12.31% and 27.48%, and 10.95% and2.86% for the same organs and species, respectively. These valuesre similar to the values obtained in a study using RAPDs to assesshe stress caused by the presence of heavy metals, in this case ofu2+, on Solanum melongena L. to which plants were exposed for 21ays to various concentrations of Cu2+ [15].
Changes in the ISSR patterns were also expressed as a decreasen GTS, a qualitative measure reflecting the changes in the num-er of ISSR profiles generated by Al-exposed P. almogravensis and P.
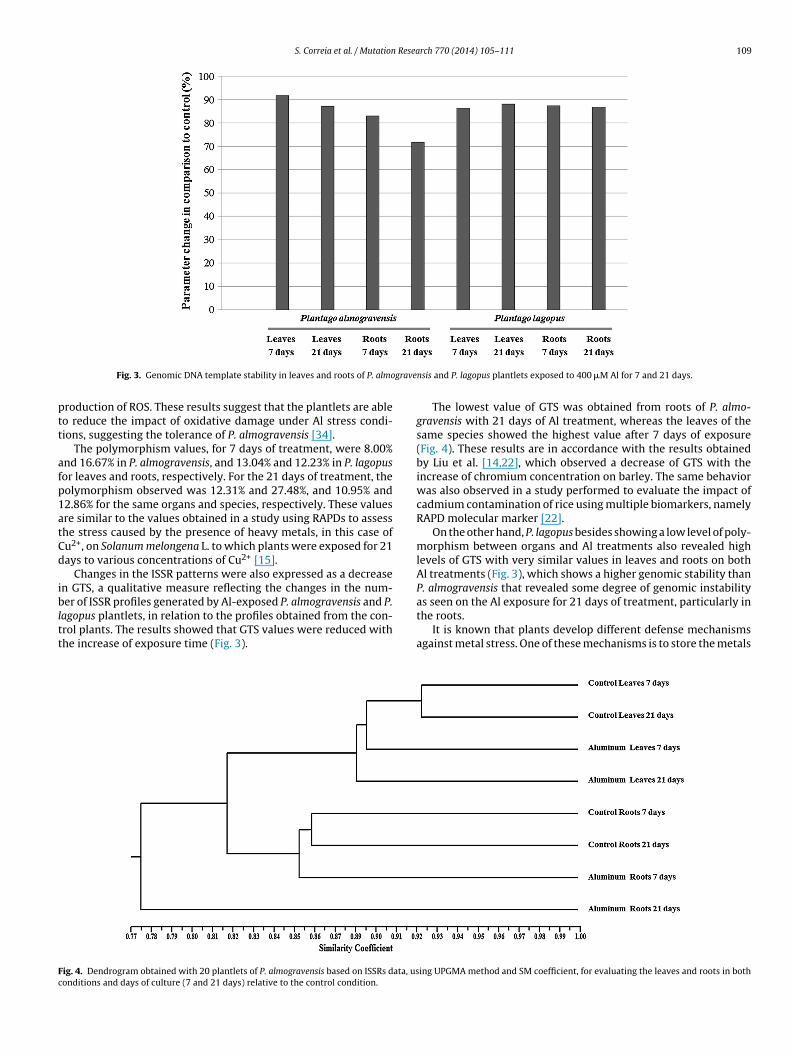
agopus plantlets, in relation to the profiles obtained from the con-rol plants. The results showed that GTS values were reduced withhe increase of exposure time (Fig. 3).
ig. 4. Dendrogram obtained with 20 plantlets of P. almogravensis based on ISSRs data, usonditions and days of culture (7 and 21 days) relative to the control condition.
nsis and P. lagopus plantlets exposed to 400 �M Al for 7 and 21 days.
The lowest value of GTS was obtained from roots of P. almo-gravensis with 21 days of Al treatment, whereas the leaves of thesame species showed the highest value after 7 days of exposure(Fig. 4). These results are in accordance with the results obtainedby Liu et al. [14,22], which observed a decrease of GTS with theincrease of chromium concentration on barley. The same behaviorwas also observed in a study performed to evaluate the impact ofcadmium contamination of rice using multiple biomarkers, namelyRAPD molecular marker [22].
On the other hand, P. lagopus besides showing a low level of poly-morphism between organs and Al treatments also revealed highlevels of GTS with very similar values in leaves and roots on bothAl treatments (Fig. 3), which shows a higher genomic stability thanP. almogravensis that revealed some degree of genomic instabilityas seen on the Al exposure for 21 days of treatment, particularly in
the roots.It is known that plants develop different defense mechanismsagainst metal stress. One of these mechanisms is to store the metals
ing UPGMA method and SM coefficient, for evaluating the leaves and roots in both
110 S. Correia et al. / Mutation Research 770 (2014) 105–111
F usingc
itatdcrAs
aprtgiestdrerdtswar
diaig
eAsosD
ig. 5. Dendrogram obtained with 20 plantlets of P. lagopus based on ISSRs data,onditions and days of culture (7 and 21 days) relative to the control condition.
n roots in order to prevent dispersal of the ions to other parts ofhe plant [49]. As a result, the accumulation of metals in the rootsffects its structure and growth more than it affects other parts ofhe plant [15]. Considering the number of bands that appear andisappear, polymorphism and GTS values (Table 2 and Fig. 3), It islear that the accumulation of Al in P. almogravensis occurs in theoots. Although P. almogravensis was previously described as anl-hyperaccumulator [25] this species revealed a lower genomictability than P. lagopus when exposed to Al.
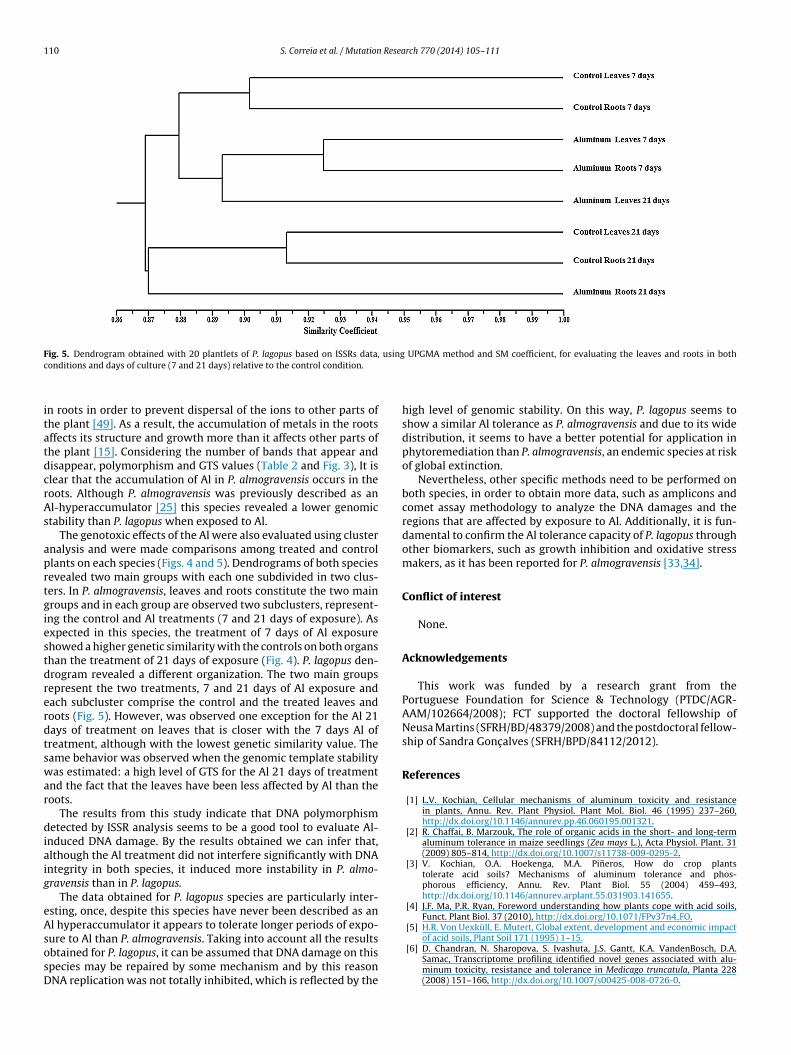
The genotoxic effects of the Al were also evaluated using clusternalysis and were made comparisons among treated and controllants on each species (Figs. 4 and 5). Dendrograms of both speciesevealed two main groups with each one subdivided in two clus-ers. In P. almogravensis, leaves and roots constitute the two mainroups and in each group are observed two subclusters, represent-ng the control and Al treatments (7 and 21 days of exposure). Asxpected in this species, the treatment of 7 days of Al exposurehowed a higher genetic similarity with the controls on both organshan the treatment of 21 days of exposure (Fig. 4). P. lagopus den-rogram revealed a different organization. The two main groupsepresent the two treatments, 7 and 21 days of Al exposure andach subcluster comprise the control and the treated leaves andoots (Fig. 5). However, was observed one exception for the Al 21ays of treatment on leaves that is closer with the 7 days Al ofreatment, although with the lowest genetic similarity value. Theame behavior was observed when the genomic template stabilityas estimated: a high level of GTS for the Al 21 days of treatment
nd the fact that the leaves have been less affected by Al than theoots.
The results from this study indicate that DNA polymorphismetected by ISSR analysis seems to be a good tool to evaluate Al-
nduced DNA damage. By the results obtained we can infer that,lthough the Al treatment did not interfere significantly with DNAntegrity in both species, it induced more instability in P. almo-ravensis than in P. lagopus.
The data obtained for P. lagopus species are particularly inter-sting, once, despite this species have never been described as anl hyperaccumulator it appears to tolerate longer periods of expo-
ure to Al than P. almogravensis. Taking into account all the resultsbtained for P. lagopus, it can be assumed that DNA damage on thispecies may be repaired by some mechanism and by this reasonNA replication was not totally inhibited, which is reflected by theUPGMA method and SM coefficient, for evaluating the leaves and roots in both
high level of genomic stability. On this way, P. lagopus seems toshow a similar Al tolerance as P. almogravensis and due to its widedistribution, it seems to have a better potential for application inphytoremediation than P. almogravensis, an endemic species at riskof global extinction.
Nevertheless, other specific methods need to be performed onboth species, in order to obtain more data, such as amplicons andcomet assay methodology to analyze the DNA damages and theregions that are affected by exposure to Al. Additionally, it is fun-damental to confirm the Al tolerance capacity of P. lagopus throughother biomarkers, such as growth inhibition and oxidative stressmakers, as it has been reported for P. almogravensis [33,34].
Conflict of interest
None.
Acknowledgements
This work was funded by a research grant from thePortuguese Foundation for Science & Technology (PTDC/AGR-AAM/102664/2008); FCT supported the doctoral fellowship ofNeusa Martins (SFRH/BD/48379/2008) and the postdoctoral fellow-ship of Sandra Gonc alves (SFRH/BPD/84112/2012).
References
[1] L.V. Kochian, Cellular mechanisms of aluminum toxicity and resistancein plants, Annu. Rev. Plant Physiol. Plant Mol. Biol. 46 (1995) 237–260,http://dx.doi.org/10.1146/annurev.pp.46.060195.001321.
[2] R. Chaffai, B. Marzouk, The role of organic acids in the short- and long-termaluminum tolerance in maize seedlings (Zea mays L.), Acta Physiol. Plant. 31(2009) 805–814, http://dx.doi.org/10.1007/s11738-009-0295-2.
[3] V. Kochian, O.A. Hoekenga, M.A. Pineros, How do crop plantstolerate acid soils? Mechanisms of aluminum tolerance and phos-phorous efficiency, Annu. Rev. Plant Biol. 55 (2004) 459–493,http://dx.doi.org/10.1146/annurev.arplant.55.031903.141655.
[4] J.F. Ma, P.R. Ryan, Foreword understanding how plants cope with acid soils,Funct. Plant Biol. 37 (2010), http://dx.doi.org/10.1071/FPv37n4 FO.
[5] H.R. Von Uexküll, E. Mutert, Global extent, development and economic impact
of acid soils, Plant Soil 171 (1995) 1–15.[6] D. Chandran, N. Sharopova, S. Ivashuta, J.S. Gantt, K.A. VandenBosch, D.A.Samac, Transcriptome profiling identified novel genes associated with alu-minum toxicity, resistance and tolerance in Medicago truncatula, Planta 228(2008) 151–166, http://dx.doi.org/10.1007/s00425-008-0726-0.

Resea
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S. Correia et al. / Mutation
[7] K. Tahara, M. Norisada, T. Yamanoshita, K. Kojima, Role of aluminum-binding ligands in aluminum resistance of Eucalyptus camaldulensis andMelaleuca cajuputi, Plant Soil 302 (2008) 175–187, http://dx.doi.org/10.1007/s11104-007-9464-5.
[8] X. Dong, R. Shen, R. Chen, Z. Zhu, J. Ma, Secretion of malate and citrate fromroots is related to high Al-resistance in Lespedeza bicolor, Plant Soil 306 (2008)139–147, http://dx.doi.org/10.1007/s11104-008-9564-x.
[9] C. Poschenrieder, B. Gunsé, I. Corrales, J. Barceló, A glance into aluminumtoxicity and resistance in plants, Sci. Total Environ. 400 (2008) 356–368,http://dx.doi.org/10.1016/j.scitotenv.2008.06.003.
10] J. Navascués, C. Pérez-Rontomé, D.H. Sánchez, C. Staudinger, S. Wienkoop, R.Rellán-Alvarez, M. Becana, Oxidative stress is a consequence, not a cause, ofaluminum toxicity in the forage legume Lotus corniculatus, New Phytol. 193(2012) 625–636, doi:0.1111/j.1469-8137.2011.03978.x.
11] P.R. Ryan, E. Delhaize, The convergent evolution of aluminium resistancein plants exploits a convenient currency, Funct. Plant Biol. 37 (2010) 275,http://dx.doi.org/10.1071/fp09261.
12] I. Brunner, C. Sperisen, Alumium exclusion and aluminum tolerance in woodyplants, Front. Plant Sci. 4 (2013), http://dx.doi.org/10.3389/fpls.2013.00172.
13] M. Monteiro, C. Santos, R.M. Mann, A.M.V.M. Soares, T. Lopes,Evaluation of cadmium genotoxicity in Lactuca sativa L. usingnuclear microsatellites, Environ. Exp. Bot. 60 (2007) 421–427,http://dx.doi.org/10.1016/j.envexpbot.2006.12.018.
14] W. Liu, Y.S. Yang, P.J. Li, Q.X. Zhou, L.J. Xie, Y.P. Han, Risk assess-ment of cadmium-contaminated soil on plant DNA damage usingRAPD and physiological indices, J. Hazard. Mater. 161 (2009) 878–883,http://dx.doi.org/10.1016/j.jhazmat.2008.04.038.
15] D.A. Körpe, S. Aras, Evaluation of copper-induced stress on eggplant (Solanummelongena L.) seedlings at the molecular and population levels by use of variousbiomarkers, Mutat. Res. Genet. Toxicol. Environ. Mutagen 719 (2011) 29–34,http://dx.doi.org/10.1016/j.mrgentox.2010.10.003.
16] E. Rodriguez, R. Azevedo, H. Moreira, L. Souto, C. Santos, Pb2+ expo-sure induced microsatellite instability in Pisum sativum in a locus relatedwith glutamine metabolism, Plant Physiol. Biochem. 62 (2013) 19–22,http://dx.doi.org/10.1016/j.plaphy.2012.10.006.
17] W. Liu, P.J. Li, X.M. Qi, Q.X. Zhou, L. Zheng, T.H. Sun, Y.S. Yang,DNA changes in barley (Hordeum vulgare) seedlings induced by cad-mium pollution using RAPD analysis, Chemosphere 61 (2005) 158–167,http://dx.doi.org/10.1016/j.chemosphere.2005.02.078.
18] M.S. Monteiro, T. Lopes, R.M. Mann, C. Paiva, A.M.V.M. Soares, C. Santos,Microsatellite instability in Lactuca sativa chronically exposed to cad-mium, Mutat. Res. Genet. Toxicol. Environ. Mutagen 672 (2009) 90–94,http://dx.doi.org/10.1016/j.mrgentox.2008.10.012.
19] R. Rucinska, R. Sobkowiak, E.A. Gwózdz, Genotoxicity of lead in lupin root cellsas evaluated by the comet assay, Cell. Mol. Biol. Lett. 9 (2004) 519–528.
20] T. Gichner, I. Znidar, J. Száková, Evaluation of DNA damage and mutagenicityinduced by lead in tobacco plants, Mutat. Res. Genet. Toxicol. Environ. Mutagen652 (2008) 186–190, http://dx.doi.org/10.1016/j.mrgentox.2008.02.009.
21] I. Carruyo, Y. Fernández, L. Marcano, X. Montiel, Z. Torrealba, Correlation oftoxicity with lead content in root tip cells (Allium cepa L.), Biol. Trace Elem. Res.125 (2008) 276–285, http://dx.doi.org/10.1007/s12011-008-8175-y.
22] W. Liu, Y.S. Yang, Q. Zhou, L. Xie, P. Li, T. Sun, Impact assessment of cadmiumcontamination on rice (Oryza sativa L.) seedlings at molecular and popula-tion levels using multiple biomarkers, Chemosphere 67 (2007) 1155–1163,http://dx.doi.org/10.1016/j.chemosphere.2006.11.011.
23] F. Al-Qurainy, Toxicity of heavy metals and their molecular detection on Phase-olus vulgaris (L.), Aust. J. Basic Appl. Sci. 3 (2009) 3025–3035.
24] B. Tutel, I. Kandemir, S. Kus, A. Kence, Classification of Turkish Plantago L. speciesusing numerical taxonomy, Turk. J. Bot. 29 (2005) 51–61.
25] C. Branquinho, H.C. Serrano, M.J. Pinto, M.A. Martins-Louc ão, Revis-iting the plant hyperaccumulation criteria to rare plants andearth abundant elements, Environ. Pollut. 146 (2007) 437–443,http://dx.doi.org/10.1016/j.envpol.2006.06.034.
26] U.S. Harput, Y. Genc, I. Saracoglu, Cytotoxic and antioxidative activities of Plan-tago lagopus L. and characterization of its bioactive compounds, Food Chem.Toxicol. 50 (2012) 1554–1559, http://dx.doi.org/10.1016/j.fct.2012.01.019.
27] M.K. Dhar, B. Friebe, A.K. Koul, B.S. Gill, Origin of an apparent Bchromosome by mutation, chromosome fragmentation and spe-
cific DNA sequence amplification, Chromosoma 111 (2002) 332–340,http://dx.doi.org/10.1007/s00412-002-0214-4.28] G. Kour, S. Kaul, M.K. Dhar, Molecular characterization of repetitive DNAsequences from B chromosome in Plantago lagopus L., Cytogenet. Genome. Res.142 (2014) 121–128, http://dx.doi.org/10.1159/000356472.
[
rch 770 (2014) 105–111 111
29] E. Lior, M. Sibony, J. Kigel, B. Rubin, Polymorphism in the resis-tance of Plantago lagopus to herbicides, Weed Res. 40 (2000) 457,http://dx.doi.org/10.1046/j. 1365-3180.2000.00201.x.
30] S. Gonc alves, N. Martins, A. Romano, Micropropagation and conservation ofendangered species Plantago algarbiensis and P. almogravensis, Biol. Plant. 53(2009) 774–778, http://dx.doi.org/10.1007/s10535-009-0142-8.
31] T. Murashige, F. Shoog, A revised medium for rapid growth and bio-assays with tobacco tissue cultures, Physiol. Plant. 15 (1962) 473–497.
32] N. Martins, S. Gonc alves, A. Romano, Aluminum inhibits root growthand induces hydrogen peroxide accumulation in Plantago algarbien-sis and P. almogravensis seedlings, Protoplasma 250 (2013) 1295–1302,http://dx.doi.org/10.1007/s00709-013-0511-1.
33] N. Martins, S. Goncalves, A. Romano, Metabolism and aluminum accu-mulation in Plantago almogravensis and P. algarbiensis in responseto low pH and aluminum stress, Biol. Plant. 57 (2013) 325–331,http://dx.doi.org/10.1007/s10535-012-0271-3.
34] N. Martins, M.L. Osório, S. Goncalves, J. Osório, A. Romano, Differencesin Al tolerance between Plantago algarbiensis and P. almogravensis reflecttheir ability to respond to oxidative stress, Biometals 26 (2013) 427–437,http://dx.doi.org/10.1007/s10534-013-9625-3.
35] E. Zietkiewicz, A. Rafalski, D. Labuda, Genome fingerprinting by simplesequence repeat (SSR)-anchored polymerase chain reaction amplification,Genomics 20 (1994) 176–183, http://dx.doi.org/10.1006/geno.1994.1151.
36] V. Ferreira, M. Matos, S. Correia, N. Martins, S. Gonc alves, A. Romano,O. Pinto-Carnide, Genetic diversity of two endemic and endan-gered Plantago species, Biochem. Syst. Ecol. 51 (2013) 37–44,http://dx.doi.org/10.1016/j.bse.2013.08.003.
37] F.J. Rohlf, NTSYS-pc, Numerical Taxonomy and Multivariate Analysis Version 2.02j, Applied Biostatistics Inc., Setauket, New York, 1998.
38] M.N.V. Prasad, H.M.O. Freitas, Metal hyperaccumulation in plants – biodiversityprospecting for phytoremediation technology, Electron. J. Biotechnol. 6 (2003),http://dx.doi.org/10.2225/vol6-issue3-fulltext-6.
39] S.K. Panda, F. Baluska, H. Matsumoto, Aluminum stress signaling in plants, PlantSignal. Behav. 4 (2009) 592–597, http://dx.doi.org/10.4161/psb.4.7.8903.
40] F. Fodor, Physiological responses of vascular plants to heavy metals, in: M.N.V.Prasad, K. Strzałka (Eds.), Physiology and Biochemistry of Metal Toxicityand Tolerance in Plants, Kluwer Academic, Dordrecht/Boston, 2002, pp. 149–177.
41] I. Kranner, L. Colville, Metals and seeds: biochemical and molecular implica-tions and their significance for seed germination, Environ. Exp. Bot. 72 (2011)93–105, http://dx.doi.org/10.1016/j.envexpbot.2010.05.005.
42] A. Méndez-Hurtado, R. Rangel-Méndez, L. Yánez-Espinosa, J. Flores, Tol-erance to cadmium of Agave lechuguilla (Agavaceae) seeds and seedlingsfrom sites contaminated with heavy metals, Scientific World J. 1–11 (2013),http://dx.doi.org/10.1155/2013/167834.
43] F.A. Atienzar, M. Conradi, A.J. Evenden, A.N. Jha, M.H. Depledge, Qualita-tive assessment of genotoxicity using random amplified polymorphic DNA:Comparison of genomic template stability with key fitness parameters inDaphnia magna exposed to benzo[a]pyrene, Environ. Toxicol. Chem. 18 (1999)2275–2282, http://dx.doi.org/10.1002/etc.5620181023.
44] F.A. Atienzar, A.J. Evenden, A.N. Jha, M.H. Depledge, Use of the random ampli-fied polymorphic DNA (RAPD) assay for the detection of DNA damage andmutations: possible implications of confounding factors, Biomarkers 7 (2002)94–101, http://dx.doi.org/10.1080/13547500110057425.
45] F.A. Atienzar, P. Venier, A.N. Jha, M.H. Depledge, Evaluation of the randomamplified polymorphic DNA (RAPD) assay for the detection of DNA damage andmutations, Mutat. Res. Genet. Toxicol. Environ. Mutagen 521 (2002) 151–163,http://dx.doi.org/10.1016/S1383-5718(02)00216-4.
46] N. Martins, S. Gonc alves, J. Osório, A. Romano, Metabolism, aluminumaccumulation in Plantago almogravensis and P. algarbiensis in responseto low pH and aluminum stress, Biol. Plant. 57 (2013) 325–331,http://dx.doi.org/10.1007/s10535-012-0271-3.
47] M. Lamhamdi, A. Bakrim, A. Aarab, R. Lafont, F. Sayah, Lead phytotoxicity onwheat (Triticum aestivum L.) seed germination and seedlings growth, C R Biol.334 (2011) 118–126, http://dx.doi.org/10.1016/j.crvi.2010.12.006.
48] V.M.M. Acharya, N.L. Parinandi, B.B. Panda, Calcium channel blockers protectagainst aluminium-induced DNA damage and block adaptive response to geno-
toxic stress in plant cells, Mutat. Res. Genet. Toxicol. Environ. Mutagen 751(2013) 130–138, http://dx.doi.org/10.1016/j.mrgentox.2012.12.008.49] J.C. Fernandes, F.S. Henriques, Biochemical, physiological, and struc-tural effects of excess copper in plants, Bot. Rev. 57 (1991) 246–273,http://dx.doi.org/10.1007/bf02858564.
Related Documents











