Hydrobiologia 381: 77–103, 1998. Zheng Mianping, S. H. Hurlbert & W. D. Williams (eds), Saline Lakes VI. © 1998 Kluwer Academic Publishers. Printed in Belgium. 77 Molecular indicators of microbial contributions to recent and Tertiary hypersaline lacustrine sediments in China Ruiliang Wang 1 , Simon C. Brassell 1 , Jiamo Fu 2 & Guoying Sheng 2 1 Biogeochemical Laboratories, Department of Geological Sciences, Indiana University, Bloomington, IN 47405-1403, U.S.A. 2 Institute of Geochemistry, Chinese Academy of Sciences, Guangzhou, 510640, P.R.C. Key words: Archaea, bacteria, biomarkers, carboxyl acids, hypersalinity Abstract In this paper, we investigate the molecular characteristics of recent sediment from the Ejinur salt lake (northern China) and eight Tertiary (Eocene) core samples from Qianjiang Formation (Eq 1-4 ) of Jianghan basin, central eastern China, focussing on the distributions and abundances of carboxylic acid isolated from extractable organic matter. n-Alkanoic acids in sediments from both study areas (Ejinur and Jianghan) show a pronounced even over odd predominance (EOP) and a bimodel distribution. In the lower molecular weight range C 16 and C 18 components are prominent with the former dominant. For higher homologues ( ≥ C 20 ), docosanoic (C 22 ) and tetracosanoic (C 24 ) acids dominant the n-alkanoic acid homologues for the Jianghan and Ejinur samples, respectively. Alkanoic acids with an isoprenoid skeleton are more abundant in Jianghan, including C 20 ,C 21 ,C 24 ,C 25 and C 30 homologues, with a C 25 component (3, 7, 11, 15, 19-pentamethyleicosanoic acid) most pronounced in the lower part of the Qianjiang Fm. The carbon skeletons of these isoprenoid acids are identical to those of isoprenoid hydrocarbons previously recognized in source rocks and petroleums from Jianghan, and attributed to Archaea (formally called archaebacteria; e.g. halophiles, methanogens). The similarity in the distributions of isoprenoid alkanoic acids and alkanes suggests that these isoprenoid acids must derive from comparable biological sources, although they appear to be formed geochemically from biosynthetic precursors (e.g. alkanes, alkenes, alkanols or alkenols). The possibility that these acids are formed as oxidation artifacts from alkanes can be discounted because of the dissimilarity between the distributions of n-alkanoic acids and n-alkanes. Iso and anteiso branched carboxylic acids are prevalent in both the Ejinur sample and the upper portion of the Qiangiang Fm (Eq 1-3 ). They derive from bacteria, perhaps sulfate reducing bacteria, and their abundances may aid assessment of the importance of bacterial contributions of organic matter in different sedimentary environments. The presence of hopanoid acids and a 3-carboxy steroidal acid further attest to contributions from bacterial and eukaryotic sources, respectively. The occurrence of carboxylic acids in the Jianghan samples confirms the potential for these compounds to survive in ancient sediments and source rocks, notably in hypersaline settings. Introduction Organic molecules synthesized by organisms survive beyond the demise of their biological sources and can be preserved in the geological record as biomarkers. They can help to identify and distinguish the sources of sedimentary organic matter, and they permit recog- nition of microorganisms that leave no morphological fossil remains (Brassell & Eglinton, 1986). The distri- butions and abundances of the major groups of organic components can also reflect depositional conditions, including characteristics like plankton productivity, salinity, water temperatures and oxicity levels (ten Haven et al., 1987a; de Leeuw & Sinninghe Damsté, 1990; Li et al., 1992; Brassell et al., 1986a; Didyk et al., 1978). Evaluation of such compounds in sed- hysh-6.tex; 18/11/1998; 13:21; p.1 article: hysh-6; pips: 180263 (hydrkap:bio2fam) v.1.1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hydrobiologia 381: 77–103, 1998.Zheng Mianping, S. H. Hurlbert & W. D. Williams (eds), Saline Lakes VI.© 1998Kluwer Academic Publishers. Printed in Belgium.
77
Molecular indicators of microbial contributions to recent and Tertiaryhypersaline lacustrine sediments in China
Ruiliang Wang1, Simon C. Brassell1, Jiamo Fu2 & Guoying Sheng21Biogeochemical Laboratories, Department of Geological Sciences, Indiana University, Bloomington,IN 47405-1403, U.S.A.2Institute of Geochemistry, Chinese Academy of Sciences, Guangzhou, 510640, P.R.C.
Key words:Archaea, bacteria, biomarkers, carboxyl acids, hypersalinity
Abstract
In this paper, we investigate the molecular characteristics of recent sediment from the Ejinur salt lake (northernChina) and eight Tertiary (Eocene) core samples from Qianjiang Formation (Eq1−4) of Jianghan basin, centraleastern China, focussing on the distributions and abundances of carboxylic acid isolated from extractable organicmatter.
n-Alkanoic acids in sediments from both study areas (Ejinur and Jianghan) show a pronounced even over oddpredominance (EOP) and a bimodel distribution. In the lower molecular weight range C16 and C18 components areprominent with the former dominant. For higher homologues (≥C20), docosanoic (C22) and tetracosanoic (C24)acids dominant then-alkanoic acid homologues for the Jianghan and Ejinur samples, respectively. Alkanoic acidswith an isoprenoid skeleton are more abundant in Jianghan, including C20, C21, C24, C25 and C30 homologues,with a C25 component (3, 7, 11, 15, 19-pentamethyleicosanoic acid) most pronounced in the lower part of theQianjiang Fm. The carbon skeletons of these isoprenoid acids are identical to those of isoprenoid hydrocarbonspreviously recognized in source rocks and petroleums from Jianghan, and attributed to Archaea (formally calledarchaebacteria; e.g. halophiles, methanogens). The similarity in the distributions of isoprenoid alkanoic acidsand alkanes suggests that these isoprenoid acids must derive from comparable biological sources, although theyappear to be formed geochemically from biosynthetic precursors (e.g. alkanes, alkenes, alkanols or alkenols).The possibility that these acids are formed as oxidation artifacts from alkanes can be discounted because of thedissimilarity between the distributions ofn-alkanoic acids andn-alkanes.Iso andanteisobranched carboxylicacids are prevalent in both the Ejinur sample and the upper portion of the Qiangiang Fm (Eq1−3). They derivefrom bacteria, perhaps sulfate reducing bacteria, and their abundances may aid assessment of the importance ofbacterial contributions of organic matter in different sedimentary environments. The presence of hopanoid acidsand a 3-carboxy steroidal acid further attest to contributions from bacterial and eukaryotic sources, respectively.
The occurrence of carboxylic acids in the Jianghan samples confirms the potential for these compounds tosurvive in ancient sediments and source rocks, notably in hypersaline settings.
Introduction
Organic molecules synthesized by organisms survivebeyond the demise of their biological sources and canbe preserved in the geological record as biomarkers.They can help to identify and distinguish the sourcesof sedimentary organic matter, and they permit recog-nition of microorganisms that leave no morphologicalfossil remains (Brassell & Eglinton, 1986). The distri-
butions and abundances of the major groups of organiccomponents can also reflect depositional conditions,including characteristics like plankton productivity,salinity, water temperatures and oxicity levels (tenHaven et al., 1987a; de Leeuw & Sinninghe Damsté,1990; Li et al., 1992; Brassell et al., 1986a; Didyket al., 1978). Evaluation of such compounds in sed-
hysh-6.tex; 18/11/1998; 13:21; p.1
article: hysh-6; pips: 180263 (hydrkap:bio2fam) v.1.1

78
iment sequences provides a measure of variations inenvironmental conditions over geological time.
Carboxylic (or fatty) acids constitute an impor-tant group of biomarkers because they are ubquitousconstituents of extractable organic matter (EOM) andare informative about the microbial sources of or-ganic matter. Some carboxylic acids exhibit a markedspecies dependence (Gillan & Johns, 1986). For in-stance, iso and anteisocarboxylic acids are estab-lished indicators for bacterial contributions (Cran-well, 1973) induding specific components reported insulfate-reducing bacteria (Boon et al., 1977a, 1978a).In contrast, common species of algae tend to produceonly n-carboxylic acids (Smith et al., 1983a; Har-wood & Russell, 1984; Lechevalier & Lechevalier,1988). Other differences between algal and bacterialcarboxylic acids include the positions of unsaturationin their alkenoic acids; algae primarily contain19Z
isomers, with C16 and C18 homologues dominant (Er-win, 1973), whereas bacteria commonly synthesize16:119Z and 18:1111Z (Gillan & Johns, 1986). Theoccurrence of carboxylic acids in microbes (Ratledge& Wilkinson, 1988), however, is not universal becausethey are absent in Archaea (formerly termed archae-bacteria). Prokaryotes within the domain Archaea arecharacterized by ether-linked, rather than ester-linked,lipids, and therefore yield virtually no carboxylic acidson saponification of biolipids (Kates, 1978).
Extensive suites of carboxylic acids are widelyreported in lacustrine (Cranwell, 1973, 1974, 1978,1981, 1983, 1984; Brooks et al., 1976, 1977; Mat-suda & Koyama, 1977a, b; Matsuda, 1978; Meyers& Takeuchi, 1979; Quirk et al., 1980; Kawamura &Ishiwatari, 1981, 1984, 1985; Cardoso et al., 1983;Leenheer & Meyers, 1983; Leenheer et al., 1984;Goossens et al., 1989; Meyers & Ishiwatari, 1993,1994), coastal, estuarine and intertidal (Johnson &Calder, 1973; Volkman & Johns, 1977; Van Vleet &Quinn, 1976, 1979; Volkman et al., 1980), and marinecontemporary environments (Cooper & Blumer, 1968;Gaskell et al., 1975; Boon et al., 1975a, b, 1978a, b;Simoneit et al., 1977; Perry et al., 1979; Van Vleet &Quinn, 1979; Bobbie & White, 1980; Meyers et al.,1980; Leenheer & Meyers, 1983; Leenheer et al.,1984; Smith et al., 1983a, b; Parkes & Taylor, 1983;DeBaar et al., 1983; Wakeham & Lee, 1989). Theyinclude components that belong to a variety of struc-tural classes, notably straight-chain, branched-chain,isoprenoid, diterpenoid, steroid and hopanoid acids.The great majority of these compounds are productsof biosynthesis, inherited by sediments directly from
their biological source. A similarly diverse, yet differ-ent, range of carboxylic acids occurs in petroleums,including numerous components that are biodegrada-tion products formed by oxidation of thermally maturehydrocarbons (e.g. Seifert & Teeter, 1970a, b; Seifert,1975; Schmitter et al., 1981; Mackenzie et al., 1983;Behar & Albrecht, 1984; Cyr & Strausz, 1984; Jaffé& Gallardo, 1993). Reports of free carboxylic acidsin ancient sediments were initially restricted to ther-mally immature shales, notably the Green River oilshale (e.g. Burlingame et al., 1969; Cox, 1970; Dou-glas et al., 1970; Haug et al., 1971), in part reflectingthe perspective that they are unstable compounds thatreadily undergo diagenetic alteration and degradation(e.g. Gaskell et al., 1976). However, the range of theirgeological occurrences has continued to grow (Kven-volden, 1970; Van Dorsselaer, 1975; Barnes et al.,1979; McEvoy, 1983; Chicarelli et al., 1984; Suzuki& Taguchi, 1984; Schaeffer et al., 1993; Barakat &Rullkötter, 1994a, b) largely refuting this presumption.In addition, some carboxylic acids are source-specific.For example, the diterpenoid acids which occur widelyin both Recent marine and lacustrine sediments thatcontain terrestrial organic matter are derived fromconifers (e.g. Simoneit, 1977), and can persist asabundant consistuents in resinous coals and lignites(e.g. Shaw et al., 1980). For hypersaline environments,the characteristics of hydrocarbons and organo-sulfurcomponents are well documented, but poorly matchedby similarly detailed investigations of other compoundclasses. Recent reports, however, document the oc-currence of several structural classes of carboxylicacids in sulfur-rich lacustrine sediments (Barakat &Rullkötter, 1994a, b). Hence, we here investigate thedistributions and abundances of extractable carboxylicacids in modern and ancient hypersaline lake settingsto ascertain their value and utility in the geochemicalcharacterization of these environments. In particular,we seek molecular evidence that may attest to thesources of sedimentary organic matter, whether bacte-rial, algal, and/or Archaeal, and provide an indicationof microbial activity.
Experimental
Samples
The two sample settings are the Ejinur Salt Lake (ESL)and the Jianghan basin (JB; Figure 1). The modernevaporitic ESL lies in the semi-arid area of Inner
hysh-6.tex; 18/11/1998; 13:21; p.2
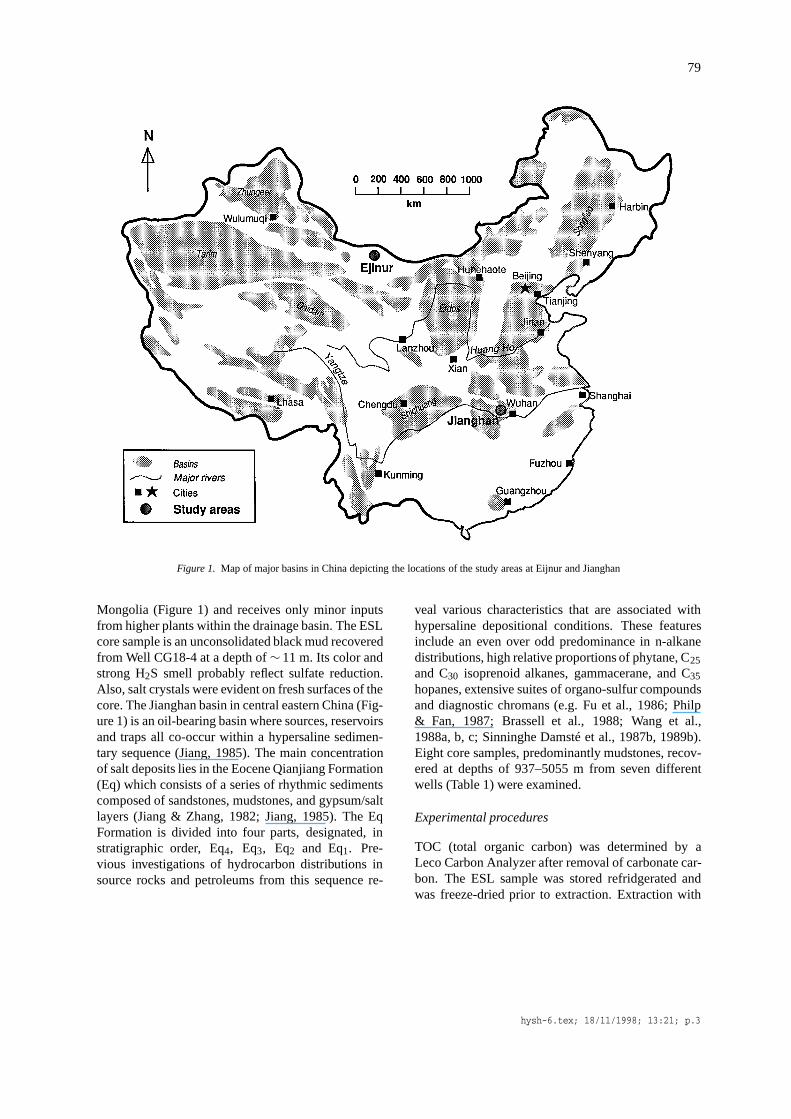
79
Figure 1. Map of major basins in China depicting the locations of the study areas at Eijnur and Jianghan
Mongolia (Figure 1) and receives only minor inputsfrom higher plants within the drainage basin. The ESLcore sample is an unconsolidated black mud recoveredfrom Well CG18-4 at a depth of∼ 11 m. Its color andstrong H2S smell probably reflect sulfate reduction.Also, salt crystals were evident on fresh surfaces of thecore. The Jianghan basin in central eastern China (Fig-ure 1) is an oil-bearing basin where sources, reservoirsand traps all co-occur within a hypersaline sedimen-tary sequence (Jiang, 1985). The main concentrationof salt deposits lies in the Eocene Qianjiang Formation(Eq) which consists of a series of rhythmic sedimentscomposed of sandstones, mudstones, and gypsum/saltlayers (Jiang & Zhang, 1982; Jiang, 1985). The EqFormation is divided into four parts, designated, instratigraphic order, Eq4, Eq3, Eq2 and Eq1. Pre-vious investigations of hydrocarbon distributions insource rocks and petroleums from this sequence re-
veal various characteristics that are associated withhypersaline depositional conditions. These featuresinclude an even over odd predominance in n-alkanedistributions, high relative proportions of phytane, C25and C30 isoprenoid alkanes, gammacerane, and C35hopanes, extensive suites of organo-sulfur compoundsand diagnostic chromans (e.g. Fu et al., 1986; Philp& Fan, 1987; Brassell et al., 1988; Wang et al.,1988a, b, c; Sinninghe Damsté et al., 1987b, 1989b).Eight core samples, predominantly mudstones, recov-ered at depths of 937–5055 m from seven differentwells (Table 1) were examined.
Experimental procedures
TOC (total organic carbon) was determined by aLeco Carbon Analyzer after removal of carbonate car-bon. The ESL sample was stored refridgerated andwas freeze-dried prior to extraction. Extraction with
hysh-6.tex; 18/11/1998; 13:21; p.3
80
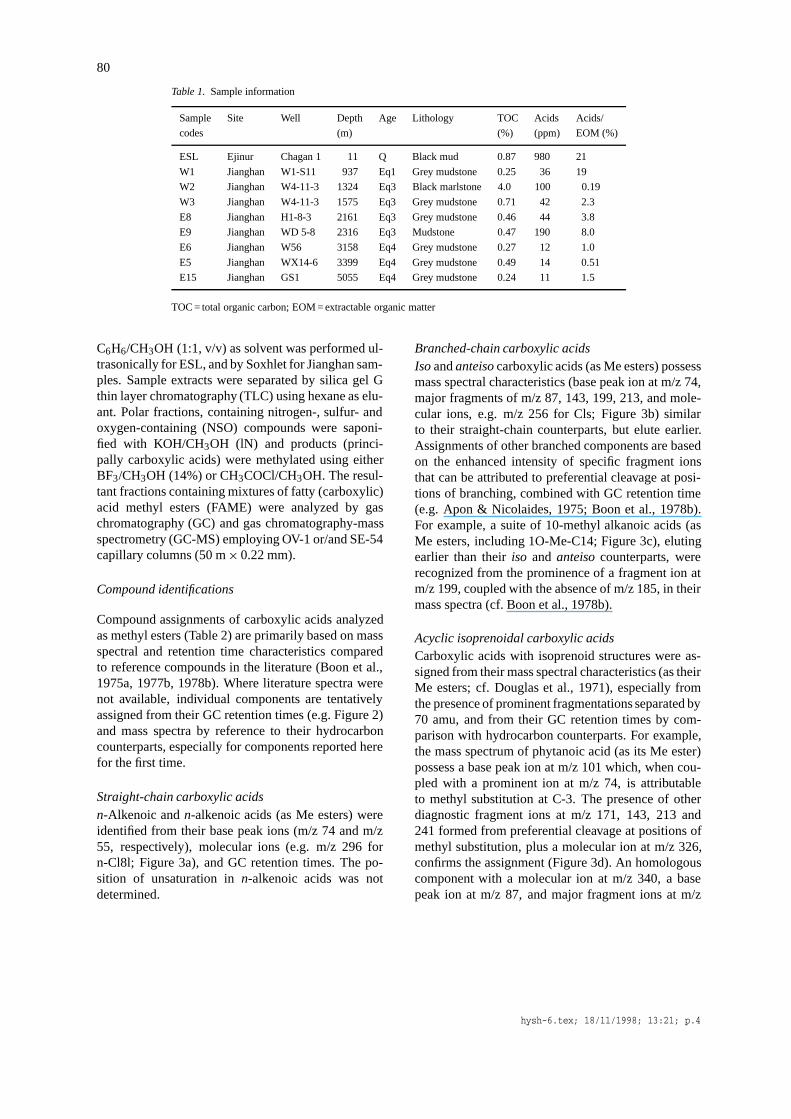
Table 1. Sample information
Sample Site Well Depth Age Lithology TOC Acids Acids/
codes (m) (%) (ppm) EOM (%)
ESL Ejinur Chagan 1 11 Q Black mud 0.87 980 21
W1 Jianghan W1-S11 937 Eq1 Grey mudstone 0.25 36 19
W2 Jianghan W4-11-3 1324 Eq3 Black marlstone 4.0 100 0.19
W3 Jianghan W4-11-3 1575 Eq3 Grey mudstone 0.71 42 2.3
E8 Jianghan H1-8-3 2161 Eq3 Grey mudstone 0.46 44 3.8
E9 Jianghan WD 5-8 2316 Eq3 Mudstone 0.47 190 8.0
E6 Jianghan W56 3158 Eq4 Grey mudstone 0.27 12 1.0
E5 Jianghan WX14-6 3399 Eq4 Grey mudstone 0.49 14 0.51
E15 Jianghan GS1 5055 Eq4 Grey mudstone 0.24 11 1.5
TOC = total organic carbon; EOM = extractable organic matter
C6H6/CH3OH (1:1, v/v) as solvent was performed ul-trasonically for ESL, and by Soxhlet for Jianghan sam-ples. Sample extracts were separated by silica gel Gthin layer chromatography (TLC) using hexane as elu-ant. Polar fractions, containing nitrogen-, sulfur- andoxygen-containing (NSO) compounds were saponi-fied with KOH/CH3OH (lN) and products (princi-pally carboxylic acids) were methylated using eitherBF3/CH3OH (14%) or CH3COCl/CH3OH. The resul-tant fractions containing mixtures of fatty (carboxylic)acid methyl esters (FAME) were analyzed by gaschromatography (GC) and gas chromatography-massspectrometry (GC-MS) employing OV-1 or/and SE-54capillary columns (50 m×0.22 mm).
Compound identifications
Compound assignments of carboxylic acids analyzedas methyl esters (Table 2) are primarily based on massspectral and retention time characteristics comparedto reference compounds in the literature (Boon et al.,1975a, 1977b, 1978b). Where literature spectra werenot available, individual components are tentativelyassigned from their GC retention times (e.g. Figure 2)and mass spectra by reference to their hydrocarboncounterparts, especially for components reported herefor the first time.
Straight-chain carboxylic acidsn-Alkenoic andn-alkenoic acids (as Me esters) wereidentified from their base peak ions (m/z 74 and m/z55, respectively), molecular ions (e.g. m/z 296 forn-Cl8l; Figure 3a), and GC retention times. The po-sition of unsaturation inn-alkenoic acids was notdetermined.
Branched-chain carboxylic acidsIsoandanteisocarboxylic acids (as Me esters) possessmass spectral characteristics (base peak ion at m/z 74,major fragments of m/z 87, 143, 199, 213, and mole-cular ions, e.g. m/z 256 for Cls; Figure 3b) similarto their straight-chain counterparts, but elute earlier.Assignments of other branched components are basedon the enhanced intensity of specific fragment ionsthat can be attributed to preferential cleavage at posi-tions of branching, combined with GC retention time(e.g. Apon & Nicolaides, 1975; Boon et al., 1978b).For example, a suite of 10-methyl alkanoic acids (asMe esters, including 1O-Me-C14; Figure 3c), elutingearlier than theiriso and anteisocounterparts, wererecognized from the prominence of a fragment ion atm/z 199, coupled with the absence of m/z 185, in theirmass spectra (cf. Boon et al., 1978b).
Acyclic isoprenoidal carboxylic acidsCarboxylic acids with isoprenoid structures were as-signed from their mass spectral characteristics (as theirMe esters; cf. Douglas et al., 1971), especially fromthe presence of prominent fragmentations separated by70 amu, and from their GC retention times by com-parison with hydrocarbon counterparts. For example,the mass spectrum of phytanoic acid (as its Me ester)possess a base peak ion at m/z 101 which, when cou-pled with a prominent ion at m/z 74, is attributableto methyl substitution at C-3. The presence of otherdiagnostic fragment ions at m/z 171, 143, 213 and241 formed from preferential cleavage at positions ofmethyl substitution, plus a molecular ion at m/z 326,confirms the assignment (Figure 3d). An homologouscomponent with a molecular ion at m/z 340, a basepeak ion at m/z 87, and major fragment ions at m/z
hysh-6.tex; 18/11/1998; 13:21; p.4

81
Table 2. Carboxylic acids tentatively identified as methyl esters (peak numbers from Figure 2; structures given in Appendix)
hysh-6.tex;18/11/1998;
13:21;p.5
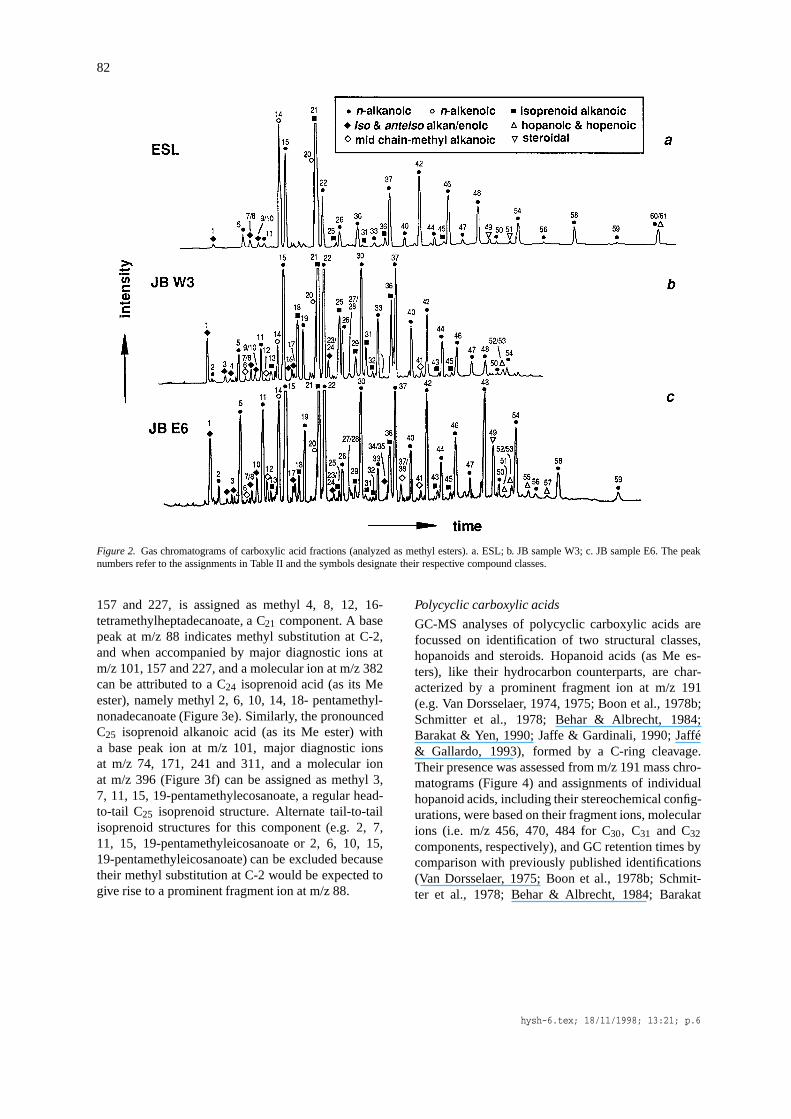
82
Figure 2. Gas chromatograms of carboxylic acid fractions (analyzed as methyl esters). a. ESL; b. JB sample W3; c. JB sample E6. The peaknumbers refer to the assignments in Table II and the symbols designate their respective compound classes.
157 and 227, is assigned as methyl 4, 8, 12, 16-tetramethylheptadecanoate, a C21 component. A basepeak at m/z 88 indicates methyl substitution at C-2,and when accompanied by major diagnostic ions atm/z 101, 157 and 227, and a molecular ion at m/z 382can be attributed to a C24 isoprenoid acid (as its Meester), namely methyl 2, 6, 10, 14, 18- pentamethyl-nonadecanoate (Figure 3e). Similarly, the pronouncedC25 isoprenoid alkanoic acid (as its Me ester) witha base peak ion at m/z 101, major diagnostic ionsat m/z 74, 171, 241 and 311, and a molecular ionat m/z 396 (Figure 3f) can be assigned as methyl 3,7, 11, 15, 19-pentamethylecosanoate, a regular head-to-tail C25 isoprenoid structure. Alternate tail-to-tailisoprenoid structures for this component (e.g. 2, 7,11, 15, 19-pentamethyleicosanoate or 2, 6, 10, 15,19-pentamethyleicosanoate) can be excluded becausetheir methyl substitution at C-2 would be expected togive rise to a prominent fragment ion at m/z 88.
Polycyclic carboxylic acids
GC-MS analyses of polycyclic carboxylic acids arefocussed on identification of two structural classes,hopanoids and steroids. Hopanoid acids (as Me es-ters), like their hydrocarbon counterparts, are char-acterized by a prominent fragment ion at m/z 191(e.g. Van Dorsselaer, 1974, 1975; Boon et al., 1978b;Schmitter et al., 1978; Behar & Albrecht, 1984;Barakat & Yen, 1990; Jaffe & Gardinali, 1990; Jaffé& Gallardo, 1993), formed by a C-ring cleavage.Their presence was assessed from m/z 191 mass chro-matograms (Figure 4) and assignments of individualhopanoid acids, including their stereochemical config-urations, were based on their fragment ions, molecularions (i.e. m/z 456, 470, 484 for C30, C31 and C32components, respectively), and GC retention times bycomparison with previously published identifications(Van Dorsselaer, 1975; Boon et al., 1978b; Schmit-ter et al., 1978; Behar & Albrecht, 1984; Barakat
hysh-6.tex; 18/11/1998; 13:21; p.6
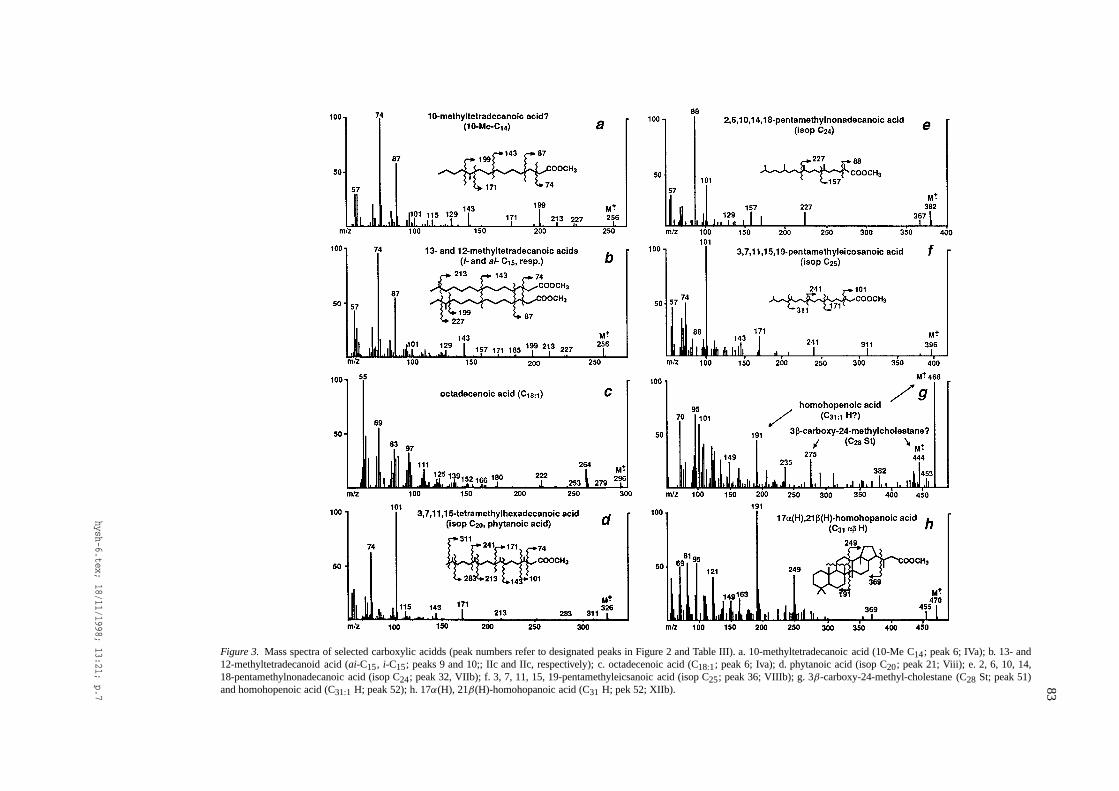
83
Figure 3. Mass spectra of selected carboxylic acidds (peak numbers refer to designated peaks in Figure 2 and Table III). a. 10-methyltetradecanoic acid (10-MeC14; peak 6; IVa); b. 13- and12-methyltetradecanoid acid (ai-C15, i-C15; peaks 9 and 10;; IIc and IIc, respectively); c. octadecenoic acid (C18:1; peak 6; Iva); d. phytanoic acid (isop C20; peak 21; Viii); e. 2, 6, 10, 14,18-pentamethylnonadecanoic acid (isop C24; peak 32, VIIb); f. 3, 7, 11, 15, 19-pentamethyleicsanoic acid (isop C25; peak 36; VIIIb); g. 3β-carboxy-24-methyl-cholestane (C28 St; peak 51)and homohopenoic acid (C31:1 H; peak 52); h. 17α(H), 21β(H)-homohopanoic acid (C31 H; pek 52; XIIb).
hysh-6.tex;18/11/1998;
13:21;p.7

84
& Yen, 1990; Jaffe & Gardinali, 1990; Jaffé & Gal-lardo, 1993). Series of methyl esters of both 17α(H),21β(H)- and 17β(H), 21α(H)-hopanoid acids wereidentified; for example, components with fragmentsions at m/z 191 and 369, at either m/z 235 or 249, andat m/z 441 or 455 ([M-15]∗) with a molecular ion atm/z 456 or 470 (Figure 3g, h; cf. Barakat & Yen, 1990)are assigned as methyl 17α(H), 21β(H)-hopanoateandmethyl 17α(H), 21β(H)-homohopanoate, respectively.In the first of these mass spectra, the presence of ionsat m/z 207, 275, 290 and 444 (Figure 3g) can beattributed to a co-eluting carboxysteroid (3β-carboxy-24-methylcholestaneas an Me ester; Dany et al., 1990;Trifilieff et al., 1992; Schaeffer et al., 1993; Barakat& Rullkötter, 1994b), whereas the origin of m/z 468is unclear (possibly a homohopenoic acid?). Othersteroidal acids with carboxylation in their side chain(Table 2) were recognized in m/z 217 mass chro-matograms (cf. Seifert et al., 1971,1972; Boon et al.,1978b). The carbon number of these steroidal acidswas assigned from their molecular ions, but no de-termination of their stereochemical configuration wasmade.
Results
Eijnur Salt Lake quaternary sediment
Bulk characteristicsThe organic carbon (TOC) content of this shallowblack mud was moderately high (0.87%; Table 1).Carboxylic acids were prevalent organic constituents(981 ppm; Table 1) and composed a significant pro-portion (>20%; Table 1) of the extractable organicmatter.
Straight-chain carboxylic acidsCarboxylic acids in ESL sediment (Figure 2) arepredominatelyn-alkanoic acids ranging to C34. Thepronounced even carbon number predominance in thehigher carbon number range (>C20) of then-alkanoicacids is characterized by two measures of their distri-bution, namely CPIf and R22f (Table 3). Both valuesare high (6.3 and 9.5, respectively) for ESL reflectinga strong even over odd predominance (EOP) and theprominence ofn-C22- C16 and C18 components aremajor components among the lower carbon numberrange of alkanoic acids (<C20) and are accompaniedby their n-alkenoic acid counterparts, with C16:1 dom-inant; C18:1 (peak 20, Figure 2) is present as the front
shoulder of the more prominent phytanoic acid (isopC20; peak 21, Figure 2).
Branched-chain carboxylic acidsThe major branched acids are comparatively short-chain, odd-numbered iso- and anteiso-componentsthat are more abundant that theirn-alkanoic acid coun-terparts, as illustrated by the high values of the ratiosfor (i + ai C15)/n-C15 and (i + ai C17)/n-C17 (2.01 and2.51, respectively; Table 3).
Acyclic isoprenoid carboxylic acidsThe predominant acid in the sample was phytanoicacid (3, 7, 11 ,15-tetramethylhexadecanoic acid, isopC20, PhA; peak 21, Figure 2). It coeluted with mi-nor amounts of C18:1. Markedly smaller amounts ofpristanoic acid (2, 6, 10, 14-tetramethylpentadecanoicacid, isopC19, PrA; peak 18, Figure 2) were detected,yielding a high PhA/PrA ratio (>25). Other acydicisoprenoid acids of higher molecular weight (isop C21,isop C23, isop C25, and isop C30) were minor con-stituents. Consequently, the ratio of phytanoic acid tothe C18 n-alkanoic acid is markedly higher than thatof its C25 counterpart to the C21 n-alkanoic acid (2.26and<0.1, respectively; Table 3).
Polycyclic carboxylic acidsCarboxylic acids with steroid and hopanoid affinitieswere tentatively identified as minor components of thesample. Their low concentrations and weak spectraprecluded confirmation of their carbon number andstereochemistry, but the GC retention time of the prin-cipal hopanoid acid suggested that it was C32 17β(H),21β(H)-bishomohopanoic acid.
Jianghan Eocene mudstones
Bulk characteristicsThe organic carbon (TOC) contents of these mud-stones lie in the range 0.25–0.71% (Table 1), valuesthat are comparable to an average shale (0.49%; Prattl& Davis, 1992). The exception is sample W2, ablack marlstone significantly richer in organic matter(4.01%; Table 1). The concentrations of carboxylicacids and their abundance in proportion to extractableorganic matter (Table 1) both show considerable vari-ation, and appear to bear no systematic relationship toeither TOC, or depth. The sample with the smallestamounts of carboxylic acids as a percentage of its ex-tractable organic matter is W2, the sample richest inorganic matter (Table 1).
hysh-6.tex; 18/11/1998; 13:21; p.8

85
Figure 4. Partial mass chromatograms from GC/MS analysis of the carboxylic acids (analyzed as methhyl esters) in sample W2 from theJianghan basin. a. reconstituted ion cgromatogram (RIC) for the elution region of C24–C34 n-alkanoic acids; b. mass fragmentogram for m/z191 depicting the distributions ofαβ- andβα-hopanoid acids.
Table 3. Ratios of carboxylic acids in the Recent (Ejinur salt lake) and Tertiary(Jianghan) hypersaline sediments.
Sample CPIf R22f i+aiC15/ i+aiC17/ isop C20/ isop C25/
code n-C15 n-C17 n-C18 n-C21
ESL 6.3 9.5 2.0 2.5 2.3 <0.1
W1 4.0 4.3 0.29 0.19 0.62 0.45
W2 2.4 2.0 0.19 0.44 0.16 0.40
W3 1.9 1.6 0.49 0.32 0.78 1.3
E8 1.7 1.8 0.25 0.22 0.37 0.64
E9 1.8 2.3 0.21 0.50 0.05 0.79
E6 3.1 2.0 0.20 0.30 0.31 1.6
E5 1.4 2.3 0.15 0.25 0.37 2.8
E15 1.4 1.9 0.14 0.43 0.18 1.2
FA = total free fatty acids;EOM = extractable organic matter
CPIf = 12
[(C20+C22+C24+C26+C28)(C21+C23+C25+C27+C29)
+ (C22+C24+C26+C28+C30)(C23+C25+C27+C29+C32)
]R22f = 2n-C22/(n-C21 + n-C23)isop C20/n-C18 = 3, 7, 11, 15-tetramethylhexadecanoic acid /n-C18isop C25/n-C21 = 3, 7, 11, 15, 19-pentamethyleicosanoic acid /n-C21i + ai C15/n-C15 = (iso+ anteisoC15) / n-C15i + ai C17/n-C17 = (iso+ anteisoC17) / n-C17
hysh-6.tex; 18/11/1998; 13:21; p.9

86
Straight-chain carboxylic acidsn-Alkanoic acids in the C13 to C34 range are prominentconstituents of all the samples. Their carbon num-ber distributions vary, but they all exhibit a strongeven over odd carbon number preference (EOP) withC16, C18 and C22 as the three most abundant ho-mologues (Figure 2). Two measures of EOP are theuniformly high values for CPIf and R22f which liein the range 1.4 to 4.0 and 1.6 to 4.3, respectively(Table 3). Neither measure shows any depth trendor correspondence to other bulk characteristics. Thedominantn-alkenoic acid in the samples is C16 (peak14; Figure 2); less amounts of C18, which partiallycoelutes with phytanoic acid (isop C20; peak 21, Fig-ure 2), are also present. No polyunsaturated alkenoicacids were detected.
Branched carboxylic acidsVarious methylalkanoic acids occur in the samples,including components withiso- and anteiso- substi-tution, and others with methyl branching at C-8 orC-10 (Figure 2; Table 2), although their distributionsdiffer markedly between samples. However, the ra-tios of iso- andanteiso-C15 and C17 components totheir n-alkanoic acid counterparts all fall within com-parably narrow ranges (0.14 to 0.49 and 0.19 to 0.50,respectively; Table 3), although there appears to be nodirect correspondence between the two values for eachsample.
Acyclic isoprenoid carboxylic acidsPhytanoic acid (isop C20; peak 21, Figure 2, 3d) isthe most prominent acylic isoprenoid acid in the sam-ples. Most samples also contain C19 (pristanoic), C21,C24 (Figure 3e), C25 (Figure 3f), C29, and C30 ho-mologues with regular, or head-to-tail structures invarying abundance. Values for the isop C20/n-C18 ratio(0.05 to 0.78) are uniformly less than unity (Table 3)indicating that the abundances of acyclic isoprenoidacids are subordinate ton-alkanoic acids in the lowercarbon range (<C20). However, the relative proportionof acyclic isoprenoid acids appears to be greater inthe higher carbon range (>C20), as shown by the isopC25/n-C21 ratios (0.40 to 2.82; Table 3).
Polycyclic carboxylic acidsC30–C33 hopanoid acids are present as minor con-stituents of samples from Eq3 (Figure 4). The predom-inant isomers are 17α(H), 21β(H) components whichare more abundant than their 17β(H), 21α(H) counter-parts. They are accompanied by less amounts of other
polycyclic components, including a C29 sterane withside chain carboxylation, a 3β-carboxy C29 steraneand, perhaps, a C31 homohopenoid acid.
Discussion
Carboxylic acids in ancient sediments: occurrencesand origins
Carboxylic acids (>C10) are recognized as promi-nent components of contemporary lacustrine sedi-ments (e.g. Cranwell, 1973, 1974; Brooks et al., 1976;Matsuda & Koyama, 1977a, b; Matsuda, 1978; Kawa-mura et al., 1980; Meyers et al., 1980; Cardoso et al.,1983; Kawamura & Ishiwatari, 1981, 1984) and havealso been reported in Neogene lake sediments thathave experienced little diagenetic alteration (Logan& Eglinton, 1994). They occur as free acids, bothsaturated and unsaturated, and as esters in triacylglyc-erols, phospholipids and glycolipids (e.g. Cranwell& Volkman, 1981; Cranwell, 1983; Fukushima &Ishiwatari, 1984; Zeng et al., 1992a, b), componentsall derived directly from biological sources. Someof these components have been recognized as con-stituents of ancient or mature sediments (Burlingameet al., 1969; Kvenvolden, 1970; Douglas et al., 1970;Haug et al., 1971; Van Dorsselaer et al., 1974; Barneset al., 1979; McEvoy, 1983), although there arecomparatively few reports of the presence of thesecarboxylic acids in petroleum source rocks (Suzuki& Taguchi, 1984; Barakat & Rullkötter, 1994a, b)by comparison with the plethora of publications con-cerning their hydrocarbon constituents. However, awide variety of carboxylic acids, including most of thecomponents recognized in the JB samples, are welldocumented constituents of petroleums (e.g. Seifert,1975; Schmitter et al., 1978). The majority of theseacids are considered to be products formed by ox-idation of hydrocarbons during biodegradation (e.g.Connan, 1984), rather than components inherited fromsource rocks. The argument that carboxylic acids inpetroleums originate as postaccumulation products issupported by various lines of evidence. First, many ofthe acids in petroleums, such as suites of polycyclictriterpenoids (Schmitter et al., 1978, 1981), do notoccur in sedimentary rocks. Second, the concentra-tions and variety of carboxylic acids in biodegradedpetroleums are markedly greater than in petroleumsthat appear unaffected by such microbial processes(Mackenzie et al., 1983; Behar & Albrecht, 1984).
hysh-6.tex; 18/11/1998; 13:21; p.10

87
For example, they are more prevalent in shallow, de-graded oils than in equivalent, deeper accumulationsthat were derived from the same source rock (Behar& Albrecht, 1984). Third, other constituents foundin biodegraded petroleums, notably alkenoic acids(Mackenzie et al., 1983), are considered too labile tosurvive the processes of diagenesis, maturation andpetroleum generation. They must therefore be formedin the reservoired oils, presumably by microbial ox-idation of hydrocarbons, although other components,including macromolecular species and organo-sulfurcompounds (OSC; Sinninghe Damsté et al., 1989a, c),may also act as precursors.
The occurrence of carboxylic acids in ancientsediments, such as the JB samples, cannot be ex-plained by formation during post-accumulation oxi-dation processes and therefore prompts evaluation ofalternative criteria to explain their origin. Three in-dependent explanations for their occurrence can beoffered, namely: (i) an origin as extraneous ratherthan indigenous components of the sample, either pro-duced by oxidation, or introduced as contaminants,(ii) formation by diagenetic transformation processesfrom hydrocarbon or other precursors, and (iii) preser-vation of biosynthetic compounds inherited directlyfrom source organisms that survive burial, and escapealteration during compaction and diagenesis.
(i) Extraneous origin. The occurrence of car-boxylic acids, especially the appearance of labilealkenoic acids in the JB samples, raises the concernthat they could be either contaminants or oxidationartefacts formed during sampling, transportation orstorage. It is important, therefore, to first resolvethis issue prior to any further discussion of the geo-chemical significance of carboxylic acids in ancientsediments.
First, the carboxylic acids that might be expectedto be introduced as contaminants during handling areC16 and C18 alkanoic and alkenoic acids because theyare prominent components of human skin oils andfingerprints. These carboxylic acids do occur in thesamples, but their distributions do not match thosein the contaminant sources typical of handling. Also,few of the other accompanying carboxylic acids repre-sent known contaminants and their overall distributionpattern is unlike that associated with contamination.Thus, it is possible that a minor proportion of spe-cific carboxylic acids in the sediments (i-e. C16 andC18 alkanoic and alkenoic acids) may arise as con-taminants, but the majority of these constituents andall other components are not extraneous. It therefore
seems consistent to interpret the carboxylic acids asindigenous components of the samples in the absenceof conclusive evidence that relates them to contamina-tion.
Second, the possibility that the carboxylic acidsare artefacts, produced by oxidation of alkanes aftersampling can also be excluded based on specific char-acteristics of their distributions that are inconsistentwith this mode of formation. In particular, oxidationprocesses cannot account for observed differences inthe distributions of alkanoic acids and alkanes. Someselectivity between different carbon skeletons (e.g.n-alkyl components, acyclic isoprenoids, hopanoids)could occur if oxidation was microbially mediated,rather than physicochemical, but not to the extendobserved in the JB samples. A biological alterationprocess could involve preferential uptake of specificcompounds thereby creating a discrimination in therate of oxidation of homologs or different carbonskeletons that, in turn, could lead to discrepanciesin the distributions of alkanes and alkanoic acids.However, any kends for homologous series wouldbe expected to be uniform. For example, preferentialoxidation of lower homologs ofn-alkanes would cre-ate a distribution ofn-alkanoic acids biased in lowercarbon numbers. Neither the ESL, nor JB distribu-tions exhibit such characteristics. In the ESL samplethe homologous series ofn-alkanoic acids extends toC34, and is dominated by C24, whereas cooccurringn-alkanes range from C15 to C30, with C28 predominant(Wang et al., unpublished results). Similarly, the dis-tributions ofn-alkanes andn-alkanoic acids in the JBsamples show major discrepancies, in terms of boththe relative abundance of the various homologs, andthe carbon number of the dominant component. Acomplete dissimilarity in such distributions seems im-probable because of the common biosynthetic originsof all n-alkyl lipids, coupled with the possibility thatn-alkanoic acids are, in part, the diagenetic precursors ofn-alkanes (e.g. Shimoyama & Johns, 1972). Overall,the distinct, irregular and largely independent varia-tions in the abundance of both individual homologsand different compound classes argue against the pos-sibility that then-alkanoic acids are artefacts formedby oxidative processes. This conclusion is supportedby the consistent, if few, reports of similar distribu-tions of carboxylic acids in other ancient sediments(e.g. Suzuki & Taguchi, 1984; Barakat & Rullkötter,1994a, b).
(ii) Diagenetic origin. The discrepancy betweenthe recognized prevalence of carboxylic acids in re-
hysh-6.tex; 18/11/1998; 13:21; p.11

88
cent sediments and their apparent paucity in ancientsediments can be attributed to diagenetic alterationprocesses, including degradation (Hedges & Prahl,1993), transformation into other components, for ex-ample by decarboxylation reactions (e.g. Didyk et al.,1978), or perhaps by incorporation into larger molec-ular species and kerogen (e.g. Ishiwatari et al., 1980;Tissot & Welte, 1984). Laboratory experiments in-tended to simulate diagenesis show an initial increasein the abundance of carboxylic acids on mild heat-ing and a subsequent decline in their abundance athigher temperatures (Kawamura & Ishiwatari, 1985).These results can be interpreted as an initial domi-nance of degradative processes leading to alteration ofcarboxylic acids, followed by thermal release of car-boxylic acids, presumably from precursors among thepolar lipids or bound in the kerogen. They demonstratethat there are additional sources of carboxylic acidswithin macromolecular structures, but do not excludethe possibility that some of the carboxylic acids inimmature sediments may survive sediment diagenesis.Thus, the sedimentary assemblage of carboxylic acidscan be to expected to evolve with increasing thermalalteration and will be greatly influenced by the charac-ter and importance of potential precursors within thepolar lipids and/or kerogen. This consideration raisesthe possibility that the distributions and abundance ofcarboxylic acids in salt lake sediments may differ fromother environments partly because of differences in thenature of their organic matter. In particular it seemsreasonable to suggest that the formation of OSC byincorporation of sulfur into unsaturated lipids duringearly diagenesis, a process prevalent in saline envi-ronments (e.g. Sinninghe Damsté et al., 1989c), mayplay a role in the preservation of carboxylic acids ortheir potential precursors. For example, the reactionof sulfur species with specific unsaturated lipid struc-tures can form either intramolecular bonds yieldingOSC such as alkylthiophenes and alkylthiolanes, orinter-molecular sulfur bridges that may contribute tokerogen formation (e.g. Tegelaar et al., 1989). Bothof these processes are recognized from the specia-tion of OSC found in recent and immature sediments.The absence of such OSC in more mature sedimentsdemonstrates that they do not persist but degradewith increasing thermal maturity, presumably becauseof the comparative weakness of carbon–sulfur bondsrelative to carbon–carbon bonds.
However, despite the prevalence of OSC in theJB (e.g. Sinninghe Damsté et al., 1989b), it is un-clear whether the circumstantial co-occurrence of car-
boxylic acids and OSC reflects any causal relationshipin the absence of specific evidence for the generationof carboxylic acids from monomeric or cross-linkedpolymeric or macromolecular OSC precursors.
(iii) Biosynthetic origin. One purpose of this pa-per is to compare the characteristics of ancient salinedeposits (i.e. JB) with a modern analog, namelythe ESL. It is reasonable to presume that all of thecomponents of ESL are either formed biosyntheti-cally, or are first-stage degradation products that canbe expected to retain the structural integrity of theirimmediate biosynthetic precursors. Where these com-pounds, their homologs or similar analogs, occur inthe JB samples they can be attributed to biosyntheticorigins and may represent refactory carboxylic acidsthat can survive diagenetic alteration processes. Incontrast the carboxylic acids in ancient saline depositsthat do not occur in modern sediments from analogousenvironments can be interpreted as products of diage-netic processes acting on biosynthetic precursors, or ascomponents unrepresented in the ESL sample, perhapsbecause their source organisms are not present there.
In summary, it appears that sedimentary carboxylicacids derive from a combination of biosynthetic anddiagenetic origins. Irrespective of their precise bio-logical origins and relationships to other constituents,such as potential precursors including hydrocarbonsand OSC, the presence of carboxylic acids in theJB samples indicates that under favorable circum-stance they can occur in sediments buried to significantdepths, i.e. 5 km.
Sources of carboxylic acids
Source organisms and salt lake environments
Examination of carboxylic acids in the ESL samplepermits comparison with previous studies of their dis-tributions in contemporary sediments, and leads toan evaluation of compositional differences and sim-ilarities between the carboxylic acids of saline ver-sus marine or freshwater environments, and of vari-ous saline settings. Most investigations of carboxylicacid compositions in contemporary environments havebeen concerned with distributions and abundance infreshwater, (e.g. Meyers & Ishiwatari, 1993, 1994),estuarine or marine settings (e.g. Brooks et al., 1976;Boon et al., 1975a, 1978a; Perry et al., 1979), whereasstudies of saline or hypersaline environments are com-paratively rare (e.g. Johnson & Calder, 1973; Cardosoet al., 1978; Philp et al., 1978; Oremland et al., 1988).
hysh-6.tex; 18/11/1998; 13:21; p.12

89
One focus here is the search for evidence amongthe carboxylic acids of contributions to sedimentaryorganic matter that can be attributed to organisms thatare restricted to, or proliferate in, saline environments,notably halophilic Archaea that include heterotrophichalobacteria and methanogens (Javor, 1989). Halobac-teria are recognized in many contemporary salt lakeenvironments somewhat similar to ESL, including theDead Sea (Kates, 1978; Kaplan & Baedecker, 1970;Nissenbaum et al., 1972; Anderson et al., 1977;Javor, 1989) and Zabuye Salt Lake in Tibet, west-ern China (Zheng et al., 1985). Most importantlytheir lipid constituents include series of components,especially various acyclic isoprenoids, that are re-stricted to Archaea (e.g. Volkman & Maxwell, 1986).In general, the specificity of biosynthetic pathwaysmanifest in the restriction of particular componentsto a limited range of classes, families or species oforganisms provides opportunities to interpret the ori-gins of sedimentary organic matter where structuralcharacteristics diagnostic of their precise biologicalsources are preserved (e.g. Eglinton, 1969; Brassell& Eglinton, 1983a, b, 1986; Brassell et al., 1987; tenHaven et al., 1987a). Similarly, detailed knowledgeof the compositions of living organisms, especiallyevidence that specific components are restricted intheir biological occurrence coupled with discrepan-cies in the compound distributions of different typesof organism, provides the necessary framework forassignment of selected components in sediments to adirect origin from biological sources, and, by infer-ence, recognition of other biologically unprecedentedcompounds as products formed by post-depositionalprocesses. However, diagnostic structural character-istics can persist in diagenetic products permittingtheir occurrence to be directly related to contribu-tions from the organisms recognized as the sourceof their precursors (e.g. Brassell, 1992). Such pre-cursor/product relationships depend on a conclusivestructural inheritance, for example between hydrocar-bon products (e.g. steranes) and their functionalizedprecursors (e.g. sterols; Mackenzie et al., 1982; Bras-sell, 1992) and on their biological specificity. Themajority of classes of biomarkers found in sediments,including carboxylic acids, tend to contain compo-nents with diagnostic structural characteristics (e.g.chain length, alkylation patterns; positions of unsat-uration; stereochemistry; ring structure; e.g. Brassell& Eglinton, 1980; Cranwell, 1982; Brassell, 1992)or features of their distribution patterns (e.g. carbonnumber predominance or preference, modality; e.g.
Brassell & Eglinton, 1980) that permit their assign-ment to an origin from either bacteria (prokaryotes),algae, or or terrestrial vascular plants. Alternatively,individual components or compound distributions canderive from a combination of these three broad bio-logical categories, making them non-specific as sourceindicators (Brassell & Eglinton, 1986) unless thereare environmental circumstances (e.g. transportationmechanisms, redox conditions, packaging) that affectthe comparative survival of components from differentbiological sources.
Non-specific carboxylic acidsC16(palmitic) and C18 (stearic)n-alkanoic acids arewidespread in organisms, including animals, vascu-lar plants, bacteria, algae, cyanobacteria and fungi(Harwood & Russell, 1984). For example, palmiticacid (C16) is the predominant carboxylic acid in manyspecies of marine algae (e.g. Smith et al., 1983a;Wood, 1988), yet it also dominates then-alkanoicacid distributions of many vascular plants (e.g. Ker-win, 1994). Similarly, the C16 n-alkanoic and C16:1alkenoic acids are prominent constitutents of the com-plex lipids of sulfate reducing bacteria (SRB), such asDesulfovibrio desulfuricans(Boon et al., 1977a), andmany other species of bacteria (O’Leary & Wilkinson,1988; Wilkinson, 1988; Lechevalier & Lechevalier,1988). The prominence of the same C16 componentin such a diverse range of organisms illustrates its lackof biological specificity and suggests that interpreta-tions of such lower molecular weightn-alkanoic acidsas indicators of contributions from algae and bacte-ria rather than from higher plants (e.g. Leenheer &Meyers, 1983) among free lipids may be erroneous(Brassell & Eglinton, 1986). These compounds tend tobe more prone to rapid degradation during early-stagediagenesis (e.g. Philp et al., 1978; Cranwell, 1978)and are less likely to survive extensive transportationthan the waxy, higher homologs (e.g.>C26) whichare more typically characteristic of then-alkanoic aciddistributions of vascular plants (Cranwell, 1978). Forexample, the refactory, waxy, highern-alkanoic acidhomologs of vascular plants were found to be the dom-inant carboxylic acids of sediment layers underlyingcyanobacterial mats despite their low abundance in theupper living layers (e.g. Philp et al., 1978). Similarly,in marine environments it is reasonable to interpret thepresence of labile organic constituents in sedimentsas autochthonous rather than allochthonous compo-nents (Brassell et al., 1980) although lower molecularweightn-alkanoic acids can be better preserved among
hysh-6.tex; 18/11/1998; 13:21; p.13

90
bound lipids (Cranwell, 1978; Meyers & Ishiwatari,1993).
These considerations bear on the occurrence oflower n-alkanoic andn-alkenoic acid homologs (i.e.C14– C20) in both the ESL and JB samples. Theirwidespread occurrence in biota could be presumedto indicate that they are non-specific markers, andtherefore not diagnostic of their biological source.However, their prominence relative to higher (>C20)n-alkanoic acids argues against the dominance of con-tributions of organic matter from allochthonous, ter-restrial sources and suggests that, in these instances, ahigher plant origin for these constituents can perhapsbe excluded. Yet, this interpretation cannot distin-guish between algal and bacterial origins for the loweracids homologs, nor can it eliminate the possibilityfor the JB samples that these components are break-down products formed from complex vascular plantlipids that are only liberated during diagenetic alter-ation. These issues cannot be resolved based on theoverall occurrences, distributions and individual abun-dances of carboxylic acids (or other components) inthe samples, although examination of carbon isotopiccompositions elsewhere has enabled recognition, dis-crimination, and deconvolution of disparate origins forn-alkanes (e.g. Collister et al., 1992), and could beequally useful in assessing the biological sources ofn-alkanoic acids.
Bacterial carboxylic acids.Both iso- andanteiso- alkanoic and alkenoic acids aregenerally regarded as constituents of bacterial origin(e.g. Cranwell, 1973; Brassell & Eglinton, 1986; Ker-win, 1994). For example, numerous types of bacteria(e.g. O’Leary & Wilkinson, 1988; Wilkinson, 1988),including SRB such asDesulfovibrio desulfuricans,containiso- andanteiso- alkenoic acids (Boon et al.,1977a, 1978a) and represent a likely biological sourcefor these components in anoxic sediments depositedin environments where sulfate reduction is prevalent,like Solar Lake, Gavish Sabkha and the Black Sea(e.g. Boon et al., 1983; de Leeuw et al., 1985; Gillan& Johns, 1986), and in other modern Chinese saltlakes, such as Dachaidan Lake and Zabuye Salt Lake,Tibet (Ma et al., 1987; Zheng et al., 1985). The in-tensity and depth zonation of sulfate reduction in ESLhas not been assessed, but the black coloration of thesample from 11 m depth with a TOC content of only0.87% (Table 1) probably arises from a pronouncedsulfide content, consistent with sulfate reduction. Theprevalence of branched carboxylic acids throughout
the Kingdom Eubacteria (e.g. Kerwin, 1994), notwith-standing their occurrence in algae, protozoa, and in-sects (e.g. Kerwin, 1994), however, precludes theirexplicit assignation to SRB rather than other potentialbacterial sources.
The widespread occurrence ofisoandanteisoalka-noic acids offers a ready explanation for their pres-ence in the ESL sample, and raises the possibilitythat they may survive diagenesis. However, it is alsonecessary to consider carbon skeletons among boundlipids that might explain their occurrence in the JBsamples following generation during diagenetic alter-ation of organic matter. For example,iso andanteisoalkanoic acids are found among the glycolipids andphospholipids of the purple photosynthetic bacteriumChlorobium and of layers of microbial mats (Zenget al., 1992a,b). In addition,iso and anteisocarbonskeletons, notably C15 components, are found in thelipid chains of monoglycerol ethers in SRB (Lang-worthy et al., 1983; Langworthy, 1985; Holzer, 1983)and occur in kerogen (Chappe et al., 1982; Chappe &Albrecht, 1982). Thus, there are a variety of bacte-ria that contain polar lipids that could act as potentialsources of these compounds following their diageneticbreakdown.
The three branched alkanoic acids (C15, C16 andC23) methylated at C-10 in the JB samples couldrepresent components of direct biological origin. 10-Methylalkanoic acids have been reported in SRB(Dowling et al., 1986), in Mycobacteria (Campbell &Noworal, 1969; Brennan, 1988), in thiobacilli (Kergeret al., 1986), and in the actinomycete Microbispora(Fowler & Douglas, 1987), although the constituentsrecognized in these organisms possess carbon num-ber distributions different from those in the sediments.The occurrence of a C23 homolog in the ancient sed-iments also prompts comparison with the series of10-methylalkanes, ranging from C21 to C35, identi-fied in pyrolysates of recent hypersaline sedimentsfrom Big Soda Lake, Nevada (Oremland et al., 1988),raising the question whether they derive from a com-mon biological source. For example, they could bothoriginate from some polar lipid or macromolecularprecursor that liberates the 10-methylalkanes on py-rolysis, but can also break down during diagenesis toform 10-methylalkanoic acids. However, the generaldifferences in the distribution patterns between the BigSoda Lake and JB samples, especially their carbonnumber ranges, and relative abundances (Figure 2) ar-gues against a common biosynthetic precursor. In ad-dition, the recognition of two 8-methylalkanoic acids
hysh-6.tex; 18/11/1998; 13:21; p.14

91
(C23 and C24) illustrates that other mid-chain posi-tions of methylation occur in the JB samples, althoughthis range of methyl-branched alkanes is less exten-sive than that observed in Proterozoic sediments andmodern cyanobacteria mats (Fowler & Douglas, 1987;Summons, 1987; Summons et al., 1988; Sheia et al.,1990; Robinson & Eglinton, 1990). The specificityof the position of branching at C-10 or C-8 supportsthe possibility of a direct biosynthetic origin becausebiosynthetic pathways leading to selective methyla-tion at preferred sites are prevalent in bacteria. Forexample, cyanobacteria synthesize specific methyl-branched alkanes, most notably 7- and 8- methylhep-tadecane (Han, 1970; Sheia et al., 1990). However, thediscrepancy in the carbon number distributions of theC-10 and C-8 methyl branched carboxylic acids (C15,C16 and C23 versus C23 and C24) argues against theircommon origin.
An alternative possibility is that these componentsare formed by diagenetic methylation, comparable tothat responsible for C-3 alkylation of steranes (Sum-mons & Capon, 1988, 1991; Dahl et al., 1992).3-Methylsteranes appear to involve methyl additionat the positions of unsaturation in12-sterenes, com-ponents that are formed by dehydation of stanols(Dastillung & Albrecht, 1977; Brassell, 1992). Com-parable alkylation of17 and19 n-alkenoic acids (orcyclopropylalkanoic acids) could occur to yield 8- and10-methylalkanoic acids, respectively, but would tendto produce a carbon number distribution similar tothat of the precursorn-alkenoic acids which is notobserved in the JB samples.
In summary, the precise origin of the 10- and 8-methyl alkanoic acids in the JB samples is unclear,but their positions of branching are consistent with ei-ther a direct biosynthetic origin from bacterial lipidsor formation as products of bacterial activity.
Sedimentary hopanoids are generally held to de-rive principally from prokaryotes (e.g. Ourisson et al.,1979), although other biological sources may con-tribute particular components. Studies of theδ13Ccompositions of hopanoids in both sediments and pe-troleums reveal that individual components originatefrom different bacterial sources, including methan-otrophs (Freeman et al., 1990; Hayes et al., 1992;Schoell et al., 1992), demonstrating a diversity oforigins for hopanoids in a single sample. In con-trast, the carbon isotopic composition of diploptene(hop22(29)-ene) demonstrates that it derives from ter-restrial sources (Prahl et al., 1992). The hopanoidacids recognized in recent lacustrine and marine
sediments, especially the prominent C32 component17β(H),21β(H)-bishomohopanoid acid, are generallyattributed to bacterial sources (Brooks et al., 1977;Quirk et al., 1980; Brassell et al., 1983; Schaef-fer et al., 1993; Brassell, 1994). Unlike the moreprominent biomarkers discussed above, sedimentaryhopanoids typically originate from aerobic prokary-otes rather than anaerobic microbes, although specifichopane constituents also derive from methanotrophicbacteria, as confirmed by their carbon isotopic signa-tures (e.g. Freeman et al., 1990). Hopanoids, espe-cially C31 and higher homologues, are held to arise asdegradation products of bacterial polyalcohols, mostnotably bacteriohopanetetraol (e.g. Ourisson et al.,1979; Brassell & Eglinton, 1984). They include com-ponents formed from biological precursors during theearly stages of diagenesis; a comparable origin ishere presumed for the C32 hopanoid acid in the ESLsample. Other hopanoids are liberated from insolubleorganic matter with increasing thermal maturation askerogen breaks down (cf. Seifert & Moldowan, 1980;Tannenbaum et al., 1986; Mycke et al., 1987), ac-companied by changes in hopanoid stereochemistry.Specifically, the less thermally stable configuration(17β(H), 21β(H)) formed biosynthetically graduallydisappears while the more stable (17β(H), 21α(H))and most stable (17α(H), 21β(H)) hopanoid configu-rations increase accompanied by isomerization of thepreferred biological orientation at C-22 (22R) into anequilibrium mixture of 22R and 22S isomers (Macken-zie et al., 1980; Seifert & Moldowan, 1980). Similartransformations occur with hopanoid acids, except thatthe isomerization rates appear slower than those of thehydrocarbons (e.g. Schaeffer et al., 1993). Generalsimilarities in the carbon number distributions, relativeabundances and stereochemistries of hopanoid acidsin the JB samples (Figure 4) to co-occurring hopanessuggests that the acids derive from the same sourceas their hydrocarbon counterparts. They probably rep-resent the products of parallel, diagenetic pathwaysgenerated from functionalized precursors, althoughformation of the acids might also perhaps proceed viaintermediates that involve sulfur incorporation (e.g.Valisolalao et al., 1984; Sinninghe Damsté et al.,1989c). The apparent absence of 17β(H), 21β(H)-hopanoid acids in the JB samples, although consistentwith their thermal instability, contrasts with publishedresults for other ancient sediments. The occurrence ofa series of 17β(H), 21β(H)-hopanoid acids in shalesfrom the Misoa Formation, Venezuela (Jaffé & Gar-dinali, 1990) is attributed to their generation during
hysh-6.tex; 18/11/1998; 13:21; p.15

92
kerogen breakdown from insoluble precursors whichhave retained their original 17α(H),21α(H) stereo-chemistry. The presence of 17β(H), 21β(H)-hopanoidacids in shallow petroleums (cf. Jaffé et al., 1988)could reflect an inheritance from source rocks, but isattributed to entainment from immature sources Jafféet al., 1988; Jaffé & Gallardo, 1993). Alternatively,these hopanoid acids may be formed, directly or in-directly, by microbes presumed to be active in thebiodegradation process.
Carboxylic acids from ArchaeaThe occurrence of series of higher homologs (>C20)of acyclic isoprenoid hydrocarbons and alcohols insediments and petroleum is generally attributed tocontributions from Archaea, principally methanogensand/or halophiles (e.g. Brassell et al., 1981; De Rosaet al., 1982, 1986; ten Haven et al., 1985; Holzer& Tornabene, 1979; Albaigés, 1980; Wang et al.,1988c). In addition, the high abundance of phytaneitself has also been related to contributions from Ar-chaea (ten Haven et al., 1987b). The prominence ofboth phytane (isop C20) and higher acyclic isoprenoidhomologs (C21–C30) as constituents of both rocks andoil samples from the JB has been attributed to Archaea(e.g. Fu et al., 1986; Wang et al., 1988a, c; Brassellet al., 1988). For example, phytane itself can amountto as much as 15% of the total EOM (Wang et al.,1988a). The presence of a suite of isoprenoid acids inthe ESL sample demonstrates that they are formed dur-ing early-stage diagenesis and several features of thecarboxylic acid distributions of the JB samples closelyresemble those of the acyclic isoprenoid alkanes andmay therefore be related to comparable sources. Phy-tanoic acid is the predominant carboxylic acid in ESL,comprisingc. 3% of its EOM, and it is also prominentin JB sediments (Figure 2; Table 3). The parallelismto the abundance of phytane suggests that phytanoicacid may also derive from Archaea. However, theether-linked isoprenoid lipids in Archaea cannot yieldisoprenoid acids directly by cleavage reactions, norhave such acids been reported in these microbes. De-spite these constraints the structural similarity of theisoprenoid cores of the ether lipids to the sedimen-tary components implies a precursor/product relation-ship, especially as no other organisms are known tobiosynthesize the required precursors, such as C25isoprenoids. Phytanoic acid could derive from the2,3-di-O-phytanyl-sn-glycerol that is widespread inArchaea (e.g. De Rosa et al., 1982), or from asymmet-ric glycerol ether lipids containing both phytanyl and
2, 6, 10, 14, 18-pentamethyleicosanyl chains (i.e. theC20, C25 diether (2-O-sesterpanyl-3-O-phytanyl-sn-glycerol; De Rosa et al., 1982) that are characteristicof strains of the alkalophilic halophiles also occurin species of methanogens (e.g. Smith, 1988). Thesource of phytanoic acid in normal marine environ-ments is generally held to be chlorophyll (e.g. Boonet al., 1975b; Didyk et al., 1978), but its origin fromhalobacterial lipids was convincing demonstrated forDead Sea sediments (Anderson et al., 1977), and canbe extended to other hypersaline enviromnents. Onepossibly is that these ether core lipids are liberatedas alcohols (Risatti et al., 1984) that then undergodiagenetic transformation into carboxylic acids. Alter-natively, they may derive from comparable hydrocar-bon precursors among Archaea, notably methanogens(e.g. Holzer et al., 1979; Albaigés, 1980; Brassellet al., 1981; Oremland, 1988). Methanogens are morelikely sources of the C30 isoprenoids (e.g. Holzer et al.,1979; Albaigés, 1980; Brassell et al., 1981; Orem-land, 1988) which are not found in halophiles (e.g.Smith, 1988). Also, sulfur incorporation at sites ofunsaturation may play an important role in the preser-vation of the isoprenoid skeletons in the ESL and JBsamples. This phenomenon can occur in marine sed-iments (Brassell et al., 1986c), but is perhaps bestrecognized and documented in various ancient hyper-saline environments (e.g. Sinninghe Damsté et al.,1987a), including JB (Sinninghe Damsté et al., 1989b)although its rôle in the preservation of precursors ofisoprenoids acids is unknown.
The inference that Archaea are among the bi-ological sources of organic matter within the ESLand JB samples is consistent with the expectationthat halobacteria and methanogens would be likelycontributors of organic matter within these salt lakeenvironments (Javor, 1989). Halobacteria have beenobserved in some of the inland modern salt lakes inarid area of China, environmentally similar to ESL(Zheng et al., 1985). For example, bacilliform andglobular halobacteria have been reported in the ZabuyeSalt Lake, Tibet, western China (Zheng et al., 1985).Species of halophilic Archaea tend to populate all hy-persaline environments, including various salt ponds,the Gavish Sabkha and the Dead Sea (Javor, 1989)and can therefore be expected to contribute to theorganic matter in such ecosystems, a presumption con-firmed by biomarker analyses (e.g. Nissenbaum et al.,1972; de Leeuw et al., 1985). The minor concen-trations, or absence, of Archaea-derived isoprenoidacids in the ESL sample may reflect the diagenetic
hysh-6.tex; 18/11/1998; 13:21; p.16

93
state of the sample, which is probably insufficient forsuch components to have been generated from anybound precursors. Alternatively, the low abundancesmay indicate that methanogenic activity in this modernsalt lake is only poorly established at the depth sam-pled. For example, the sample may originate from thezone of sulfate reduction, as is reflected by the muchhigher abundance ofiso and anteiso monomethylbranched acids, rather than from a deeper zone wheremethanogenesis becomes dominant. In contrast, thehigh concentration of C25 sesterterpane (2, 6, 10, 14,18-pentamethyleicosane)and its carboxylic acid coun-terpart may derive from alkaline halophilic Archaea,although a methanogenic source cannot be excluded(e.g. Risatti et al., 1984).
Algal and terrestrial carboxylic acidsThe biological origin of steroidal acids in sedimentsand petroleums was contentious until comparativelyrecently. Arguments for an animal origin for cholanicacid (Seifert, 1973) were refuted (Boon et al., 1978b),but the restriction of steroidal compounds to eukary-otes (e.g. Mackenzie et al., 1982) provides convincingevidence that these components do not derive frombacteria. Thus, the presence of a 3β-carboxysteroid(Dany et al., 1990; Schaeffer et al., 1993; Barakat& Rullkötter, 1994a) and other minor, tentativelyassigned steroidal acids in the JB samples are here pre-sumed to reflect contributions from eukaryotic organ-isms. In hypersaline environments eukaryotic sourcesmay include the green alga Dunaliella, diatoms, andprotozoa (Javor, 1989), in addition to allochthonoussources of plant matter that may be presumed to derivefrom surrounding drainage areas. The low abundanceof steroidal carboxylic acids, however, does not pro-vide a quantitative measure of the importance of con-tributions from eukaryotic sources to recent or ancienthypersaline sediments, primarily because these com-ponents appear to be generated from kerogen (Barakat& Rullkötter, 1994a). Thus, their occurrence andabundance is contingent on that of precursor ester-bound steroids, which appear among bound lipids butare less prevalent in kerogen than many other lipidclasses (e.g. Mycke & Michaelis, 1986). Furthermore,formation of 3β-carboxysteroids depends on incor-poration of a modified (i.e. C-3 substituted) steroidstructure (Barakat & Rullkötter, 1994a) and thereforerequires a three-step transformation process consistingof sterol modification, incorporation, and subsequentliberation. However, the best documented steroidalconstituents of kerogens appear to be compounds in-
corporated via ether, rather than ester-linked, at C-3(e.g. Mycke & Michaelis, 1986). All of these consid-erations may bear on the fact that the minor amounts ofsteroidal acids in the JB samples are in sharp contrastto the dominance of steranes among their aliphatic hy-drocarbons (e.g. Fu et al., 1986; Brassell et al., 1988;Wang et al., 1988b, c).
Molecular signals of diagenetic alteration anddepositional environment
Inter-comparison of JB and ESL samplesInevitably many of the differences between modernand ancient sediments are a function of diagenetic al-teration. Thus, it is critical to attempt to assess theeffects of diagenetic overprinting on the original com-position of the ancient sediments and to exclude suchinfluences, or compensate for them, in considerationsof molecular signatures that reflect the environmentof deposition. Allowance can be made for the ab-sence in recent sediments of components formed bydiagenetic transformation, but assessment of the ef-fect of diagenesis on quantitative measures are moreproblematic and perhaps irresolvable for many com-pounds. For example, the abundances of C15 andC17 branched monomethyl acids and of C20 and C25acyclic isoprenoid acids relative to their neighboringn-alkanoic acids (Table 3) reveal marked differencesbetween the ESL sediment and JB samples. The highervalues in ESL for three of these ratios may be at-tributed to the fact that this sediment is too imma-ture for n-alkanoic acids to have been released fromprecursor lipids such as triacylglycerols and phospho-lipids (cf. Zeng et al., 1992b). Alternatively, the C15and C17 branched monomethyl and C20 acyclic iso-prenoid acids may be less abundant because they, andother components derived from their inferred sources(i.e. sulfate-reducing bacteria and halophiles), havebeen diluted or modiffed by subsequent diageneticprocesses in the shallow subsurface.
Irrespective of the cause(s) for the differences inthese molecular abundances between the samples, itis impossible to determine whether the ratios for theESL sediment would decrease to values comparableto those of the JB samples with increasing diageneticalteration. Thus, the equivalence between the data setsand any inferences regarding similarities in their de-positional characteristics cannot be tested. The fourthmeasure (isop C25/C21) shows the opposite behaviourwith the value for the ESL sediment markedly lowerthan that of the JB samples. Apart from diagenetic
hysh-6.tex; 18/11/1998; 13:21; p.17

94
considerations, it is also possible that this differencerepresents sampling or environmental differences. As-suming that the C25 isoprenoid alkanoic acid (isopC25) originates from alkaline halobacteria, its lowabundance may reflect the influence of lake alkalin-ity and/or salinity on the populations of halobacteriarather than any post-depositional differences. For ex-ample, the alkalinity of the Eocene Jianghan salt lakemay have been higher than that of the modern Ejinursalt lake. Alternatively, if the C25 isoprenoid alkanoicacid (isop C25) derives from methanogens, it wouldbe expected to be a minor constituent if the ESL sed-iment collected at shallow depth was taken from anhorizon where methanogenesis was poorly developed.These considerations demonstrate the need for de-tailed investigation of the species-dependence of dif-ferent dominant lipids of Archaea to better constraintheir utility and significance in efforts to reconstructdepositional environments.
Despite some significant differences in their mole-cular distributions, the suites of carboxylic acids inthe ESL and JB environments bear a major resem-blance. Perhaps the comparable prominence of acylicisoprenoid acids represents the most pronounced com-mon characteristic. However, it is also important tonote that components which record and characterizehigher plant contributions from gymnosperms and an-giosperms, namely diterpenoid (e.g. Simoneit, 1977;Brassell et al., 1983) and triterpenoid (Corbet et al.,1980; Schmitter et al., 1981) acids, respectively, werenot identified in either environment, in contrast to theirabundance in samples from other depositional regimes(e.g. Corbet et al., 1980; Brassell et al., 1987).
Intra-comparison of JB samplesA key question in the assessment of discrepanciesin the distributions of carboxylic acids between theJB samples is whether the observed differences (Fig-ure 2; Table 3) reflect varying degrees of diageneticalteration or significant changes in depositional en-vironment. Alternatively, minor differences may besimply a function of the expected stratigraphic vari-ation (cf. Brassell et al., 1986b; Moldowan et al.,1986; Curiale & Odermatt, 1989; Miranda & Walters,1992). However, the sampling intervals between theJB samples and their collection from different wellsand separate formations (Table 1) restrict considera-tion of such factors to broad generalities. Assessmentof stratigraphic changes in geochemical characteris-tics can seek to identify systematic depth trends thatare consistent with diagenetic influences and distin-
guish them from fluctuations that appear independentof depth. For example, there is a general reductionin carboxylic acid concentrations with depth (Table 1)consistent with their gradual loss, although this trendis irregular, especially given the high value for sampleE9. The TOC contents (Table 1) of the samples varyerratically, and appear, in part, to reflect lithologicaldifferences. Similarly, the ratios reflecting the relativeproportions ofn-, monomethyl branched, and acyclicisoprenoid alkanoic acids are broadly similar and showno coherent depth trend (Table 3; Figure 5). In general,the comparability between the distributions of the var-ious families of carboxylic acids in the samples arguesagainst any radical change in depositional conditionsthroughout the sequence, although their capability toprovide diagnostic evidence of major diagenetic trendsremains untested.
SalinityAn even over odd predominance (EOP) amongn-alkanes is generally regarded (Sheng et al., 1980; tenHaven et al., 1988; Brassell et al., 1988; de Leeuw &Damsté, 1990; Li et al., 1992) as a feature of hyper-saline environments, including the JB (Brassell et al.,1988), especially when coupled with a dominance ofdocosane (C22) (ten Haven et al., 1985; Brassell et al.,1988; de Leeuw & Damsté, 1990). Values greater than1.5 for the index R22 = 2C22/(C21 + C23) have beenproposed as an indication of hypersaline depositionalconditions (ten Haven et al., 1985, 1988). However,similar characteristics inn-alkanoic acid distributions,namely the predominance of C22 (docosanoic acid)and other even homologs in the JB samples and ESLsediment, do not necessarily convey the same informa-tion about the depositional environment. In particular,biosynthetic pathways produce a dominance of even-numberedn-alkanoic acids, which, after decarboxy-lation, yield n-alkane distributions with a predom-inance of odd-numbered homologs. A pronouncedEOP forn-alkanes is therefore a geochemical, ratherthan biological, phenomenon typically attributed totheir formation fromn-alkanoic acid precursors (Welte& Ebhardt, 1968; Kvenvolden, 1970; Albaigés andTorradas, 1977; Sheng et al., 1980), probably viaOSC intermediates, notablyn,n-dialkylthiophenes andn,n- dialkylthiolanes (Sinninghe Damsté et al., 1987a,1989b, c). It is held to be dependent on depositionalconditions that enhance the preservation of precursorlipids with an EOP and favor their selective transfor-mation ton-alkanes. Thus, it is the result of a discreteprocess occurring in hypersaline environments that is
hysh-6.tex; 18/11/1998; 13:21; p.18
95
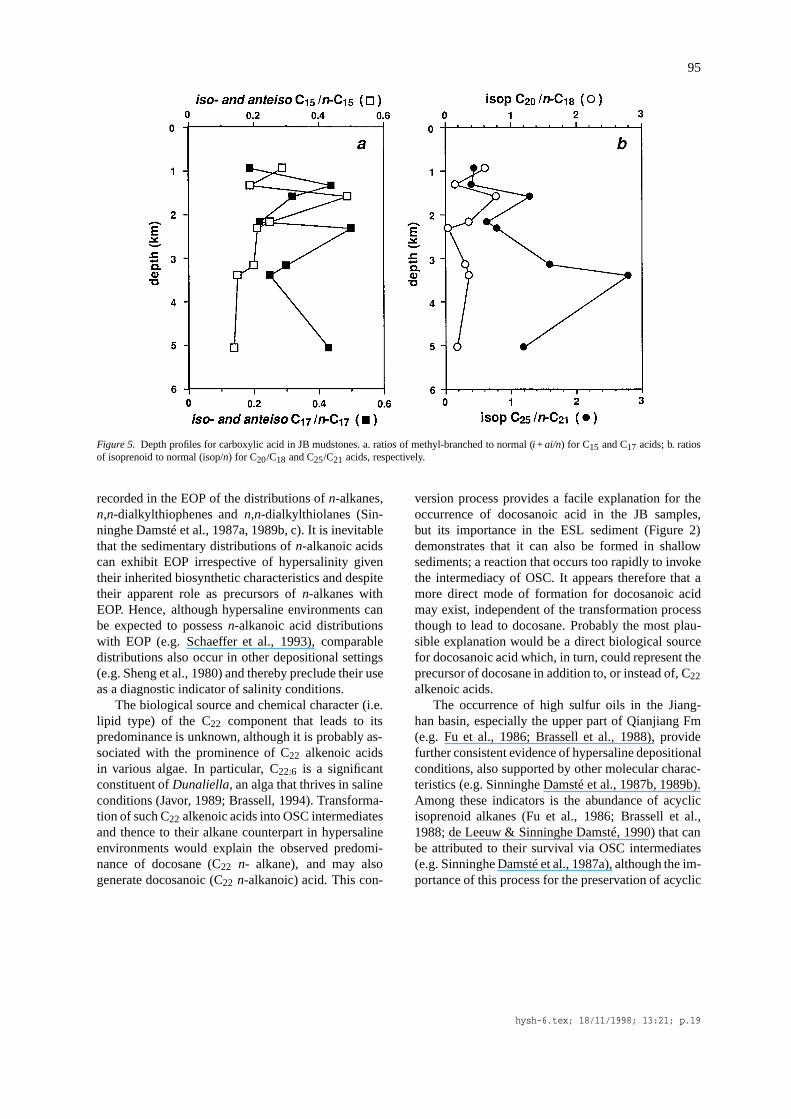
Figure 5. Depth profiles for carboxylic acid in JB mudstones. a. ratios of methyl-branched to normal (i + ai/n) for C15 and C17 acids; b. ratiosof isoprenoid to normal (isop/n) for C20/C18 and C25/C21 acids, respectively.
recorded in the EOP of the distributions ofn-alkanes,n,n-dialkylthiophenes andn,n-dialkylthiolanes (Sin-ninghe Damsté et al., 1987a, 1989b, c). It is inevitablethat the sedimentary distributions ofn-alkanoic acidscan exhibit EOP irrespective of hypersalinity giventheir inherited biosynthetic characteristics and despitetheir apparent role as precursors ofn-alkanes withEOP. Hence, although hypersaline environments canbe expected to possessn-alkanoic acid distributionswith EOP (e.g. Schaeffer et al., 1993), comparabledistributions also occur in other depositional settings(e.g. Sheng et al., 1980) and thereby preclude their useas a diagnostic indicator of salinity conditions.
The biological source and chemical character (i.e.lipid type) of the C22 component that leads to itspredominance is unknown, although it is probably as-sociated with the prominence of C22 alkenoic acidsin various algae. In particular, C22:6 is a significantconstituent ofDunaliella, an alga that thrives in salineconditions (Javor, 1989; Brassell, 1994). Transforma-tion of such C22 alkenoic acids into OSC intermediatesand thence to their alkane counterpart in hypersalineenvironments would explain the observed predomi-nance of docosane (C22 n- alkane), and may alsogenerate docosanoic (C22 n-alkanoic) acid. This con-
version process provides a facile explanation for theoccurrence of docosanoic acid in the JB samples,but its importance in the ESL sediment (Figure 2)demonstrates that it can also be formed in shallowsediments; a reaction that occurs too rapidly to invokethe intermediacy of OSC. It appears therefore that amore direct mode of formation for docosanoic acidmay exist, independent of the transformation processthough to lead to docosane. Probably the most plau-sible explanation would be a direct biological sourcefor docosanoic acid which, in turn, could represent theprecursor of docosane in addition to, or instead of, C22alkenoic acids.
The occurrence of high sulfur oils in the Jiang-han basin, especially the upper part of Qianjiang Fm(e.g. Fu et al., 1986; Brassell et al., 1988), providefurther consistent evidence of hypersaline depositionalconditions, also supported by other molecular charac-teristics (e.g. Sinninghe Damsté et al., 1987b, 1989b).Among these indicators is the abundance of acyclicisoprenoid alkanes (Fu et al., 1986; Brassell et al.,1988; de Leeuw & Sinninghe Damsté, 1990) that canbe attributed to their survival via OSC intermediates(e.g. Sinninghe Damsté et al., 1987a), although the im-portance of this process for the preservation of acyclic
hysh-6.tex; 18/11/1998; 13:21; p.19

96
isoprenoid alkanoic acids in hypersaline environmentsis unclear. For the ESL and JB samples the relativeproportions of acydic isoprenoid alkanoic acids andn-alkanoic acids vary markedly; such differences mayarise from differences in salinity, but other factors,especially source inputs and diagenetic alteration reac-tions most probably control their ratios. For example,phytanoic acid (isop C20) dominates octadecanoic (n-C18) acid in the ESL sediment (isopC20/C18 = 2.3;Table 3), but is subordinate to it in the JB samples (isopC20/C18 ranges from 0.05 to 0.78; Table 3; Figure 5b).In contrast, the proportion of the C25 isoprenoid acid(isop C25) relative to heneicosanoic (n-C21) acid ismarkedly higher in the JB samples (isop C25/C21ranges from 0.40 to 2.8; Table III; Figure 5b) than inthe ESL sediment (isop C25/C21<0.1; Table 3). Thesediscrepant ratios demonstrate that any assessment ofsalinity from such parameters is fraught with difficul-ties associated with inadequacies in our understandingof the precise sources of specific carboxylic acids andtheir diagenetic transformations. Thus, it appears thatthe distributions of carboxylic acids can augment therange of measures that denote hypersaline depositionalconditions (cf. ten Haven et al., 1985, 1988; de Leeuw& Sinninghe Damsté, 1990), but do not provide a moreprecise environmental characterization.
Conclusions
The range and distributions of carboxylic acids in aRecent mud from Eijnur salt lake and in Eocene mud-stones from the Jianghan basin record evidence ofbacterial contributions to the organic matter in thesehypersaline environments supplemented by algal orhigher plant sources. The structural specificity of someof these components helps constrain their microbialorigins. For example, several of the components areconsistent with an origin from sulfate-reducing bac-teria. Others, especially the higher homologs (>C21)of acyclic isoprenoid acids, attest to contributionsfrom halophilic and/or methanogenic Archaea. Ar-chaea, however, do not synthesize carboxylic acidswhich implies that these diagnostic tail-to-tail iso-prenoid structures must represent diagenetic productsprobably formed from polar lipid precursors. Theiroccurrence in the Jianghan basin samples may per-haps be attributed to their early incorporation intoOSC (or kerogen?) and subsequent release duringdiagenesis. The preferential preservation associatedwith this process involving OSC may further help ex-
plain why such acyclic isoprenoid acids tend to beprevalent in other ancient sediments from hypersalinesettings. Thus, specific chacteristics of the carboxylicacid distributions, primarily the abundance of acyclicisoprenoids, appear to be representative of hyper-saline conditions, although they cannot yet be used forquantitative measurement of salinity.
Acknowledgments
We thank Dr Shuichang Zhang (Institute of PetroleumExploration and Development, Beijing, PRC) andMr Jigang Jiang (Jianghan Institute of Petrolum Ex-ploration & Development, PRC) for providing someof the core samples from the Jianghan basin and Prof.Dapeng Sun (Qinghai Salt Lake Institute, CAS) forassistance with sampling Ejinur Salt Lake sediments.We also acknowledge research support from the Davidand Lucile Packard Foundation through a Fellowshipin Science and Engineering (SCB).
References
Albaigés, J., 1980. Identification and geochemical significance oflong chain acyclic isoprenoid hydrocarbons in crude oils. InA. G. Douglas & J. R. Maxwell Pergamon (eds), Advances inOrganic Geochemistry 1979, Oxford: 19–28.
Albaigés, J. & J. Torradas, 1977. Geochemical characterization ofSpanish crude oils. In R. Campos & J. Goñi (eds), Advances inOrganic Geochemistry 1975 (eds). Enadimsa, Madrid: 99–115.
Anderson, R., M. Kates, M. J. Baedecker, I. R. Kaplan & R. G. Ack-man, 1977. The stereoisometric composition of phytany chainsin lipids of Dead Sea sediments. Geochim. Cosmochim. Acta 41:1381–1390.
Apon, J. M. B. & N. Nicolaides, 1975. The determination ofpositional isomers of methyl esters by capillary GC/MS. J.Chromatogr. Sci. 13: 467–473.
Barakat, A. O. & T. F. Yen, 1990. Distribution of pentacyclic triter-penoids in Green River oil shale kerogen. Org. geochem. 15:299–311.
Barakat, A. O. & J. Rullkötter, 1994a. Free aliphatic acids insulfur-rich lacustrine sediments: their origin and relation tohydrocarbons. Energy & Fuels 8: 474–480.
Barakat, A. O. & J. Rullkötter, 1994b. Occurrence of bound 3β-carboxysteroids in geological samples. Energy & Fuels 8: 481–486.
Behar, F. H. & P. Albrecht, 1984. Correlations between car-boxylic acids and hydrocarbons in several crude oils; alter-ation by biodegradation. In P. A. Schenck, J. W. de Leeuw &G. W. M. Lijmbach (eds), Advances in Organic Geochemistry1983. Org. Geochem. 6: 597–604.
Barnes, P. J., S. C. Brassell, P. Comet, G. Eglinton, J. McEvoy,J. R. Maxwell, A. M. K. Wardroper & J. K. Volkman (1979). Pre-liminary lipid analysis of core sections 18, 24 and 30 from Hole402A. In L. Montadert, D. G. Roberts et al. (eds), Initial Reportsof the Deep Sea Drilling Project, Vol. XLVIII, U.S. GovernmentPrinting Office, Washington D.C.: 965–976.
hysh-6.tex; 18/11/1998; 13:21; p.20

97
Bobbie, R. & D. C. White, 1980. Characterization of benthic mi-crobial community structure by high-resolution gas chromatog-raphy of fatty acid methyl esters. Appl. Envir. Microbiol. 39:1212–1222.
Boon, J. J., J. W. de Leeuw & P. A. Schenck, 1975a. Organic geo-chemistry of Walvis Bay diatomaceous ooze – I. Occurrence andsignificance of the fatty acids. Geochim. Cosmochim. Acta 39:1559–1565.
Boon, J. J., W. I. C. Rijpstra, J. W. de Leeuw & P. A. Schenck,1975b. Phytenic acid in sediments. Nature 258: 414–416.
Boon, J. J., J. W. de Leeuw, G. J. v.d. Hoek & J. H. Vosjan, 1977a.Significance and taxonomic value of iso and anteiso monoenoicfatty acids and branchedβ-hydroxy acids inDesulfovibrio desul-furicans. J. Bacteriol. 129: 1183–1191.
Boon, J. J., B. van de Graaf, P. J. W. Schuyl, F. de Lange &J. W. de Leeuw, 1977b. The mass spectrometry of iso and anteisomonoenoic fatty acids. Lipids 12: 717–721.
Boon, J. J., W. Liefkens, W. I. C. Rijpstra, M. Baas & J. W. deLeeuw, 1978a. Fatty acids ofDesulfovibrio desulfuricansasmarker molecules in sedimentary environments. In W. E. Krum-bein (ed.), Environmental Biogeochemistry and Geomicrobiol-ogy. Vol. 1: The Aquatic Environment, Ann Arbor Science, AnnArbor: 355–372.
Boon, J. J., J. W. de Leeuw & A. L. Burlingame, 1978b. Organicgeochemistry of Walvis Bay diatomaceous ooze – III. Structuralanalysis of the monoenoic and polycyclic fatty acids. Geochim.Cosmochim. Acta 42: 631–644.
Boon, J. J., H. Hines, A. L. Burlingame, J. Klok, W. I. C. Ri-jpstra, J. W. de Leeuw, K. E. Edmunds & G. Eglinton, 1983.Organic geochemical studies of Solar Lake laminated cyanobac-terial mats. In M. Bjorøy et al. (eds), Advances in OrganicGeochemistry 1981, Wiley, Chichester: 207–227.
Brassell, S. C., 1992. Biomarkers in recent and ancient sediments:the importance of the diagenetic continuum. In J. Whelan &J. W. Farrington (eds), Organic Matter: Productivity, Accu-mulation, and Preservation in Recent and Ancient Sediments,Columbia University Press, New York: 339–367.
Brassell, S. C., 1994. Isopentenoids and Geochemistry. In W. D. Nes(ed.), Isopentenoids and Other Natural Products: Evolution andFunction. American Chemical Society Symposium Series 562:2–30.
Brassell, S. C. & G. Eglinton, 1980. Environmental chemistry– an interdisciplinary subject. Natural and pollutant organiccompounds in contemporary aquatic environments. In J. Al-baigés (ed.), Analytical Techniques in Environmental Chemistry,Pergamon, Oxford: 1–22.
Brassell, S. C. & G. Eglinton, 1983a. The potential of organic geo-chemical compounds as sedimentary indicators of upwelling. InE. Suess & J. Thiede (eds), Coastal Upwelling, its SedimentRecord, Part A, Plenum, New York: 545–571.
Brassell, S. C. & G. Eglinton, 1983b. Steroids and triterpenoids indeep sea sediments as environmental and diagenetic indicators.In M. Bjoróy et al. (eds), Advances in Organic Geochemistry1981, Wiley, Chichester: 684–697.
Brassell, S. C. & G. Eglinton, 1984. Lipid indicators of mi-crobial activity in marine sediments. In J. E. Hobbie &P. J. leB. Williams, Plenum, New York: 481–503.
Brassell, S. C. & G. Eglinton, 1986. Molecular geochemical in-dicators in sediments. In M. L. Sohn (ed.), Organic MarineGeochemistry, American Chemical Society Symposium Series305: 1–32.
Brassell, S. C., P. A. Comet, G. Eglinton, P. J. Isaacson, J. McEvoy,J. R. Maxwell, I. D. Thomson, P. J. C. Tibbetts & J. K. Volk-man, 1980. The origin and fate of lipids in the Japan Trench.
In A. G. Douglas & J. R. Maxwell (eds), Advances in OrganicGeochemistry 1979, Pergamon, Oxford: 375–391.
Brassell, S. C., A. M. K. Wardroper, I. D. Thomson, J. R. Maxwell& G. Eglinton, 1981. Specific acyclic isoprenoids as biologicalmarkers of methanogenic bacteria in marine sediments. Nature290: 693–696.
Brassell, S. C., G. Eglinton & J. R. Maxwell, 1983. The geo-chemistry of terpenoids and steroids. Biochem. Soc. Trans. 11:575–586.
Brassell, S. C., G. Eglinton, I. T. Marlowe, U. Pflaumann &M. Sarnthein, 1986a. Molecular stratigraphy: a new tool forclimatic assessment. Nature 320: 129–133.
Brassell, S. C., G. Eglinton & Fu Jiamo, 1986b. Biological markercompounds as indicators of the depositional history of theMaoming oil shale. In D. Leythaeuser & J. Rullkötter (eds),Advances in Organic Geochemistry 1985, Org. Geochem. 10:927–941.
Brassell, S. C., C. A. Lewis, J. W. de Leeuw, F. de Lange & J. S. Sin-ninghe Damsté, 1986c. Isoprenoid thiophenes: novel products ofsediment diagenesis? Nature 320: 160–162.
Brassell, S. C., G. Eglinton & V. J. Howell, 1987. Paleoenvironmen-tal assessment for marine organic-rich sediments using molecularorganic geochemistry. In J. Brooks & A. J. Fleet (eds), MarinePetroleum Source Rocks, Geological Society Special PublicationNo. 24, Blackwell, Oxford: 79–98.
Brassell, S. C., Sheng Guoying, Fu Jiamo & G. Eglinton, 1988.Biological markers in lacustrine Chinese oil shales. In A. J. Fleet,K. Kelts & M. R. Talbot (eds), Lacustrine Petroleum SourceRocks, Geological Society Special Publication No. 40, Black-well, Oxford: 299–308.
Brennan, P. J., 1988. Mycobacterium and other actinomycetes. InC. Ratledge & S. G. Wilkinson (eds), Microbial Lipids Vol. 1,Academic, London: 203–298.
Brooks, P. W., G. Eglinton, S. J. Gaskell, D. J. McHugh,J. R. Maxwell & R. P. Philp, 1976. Lipids of recent sedi-ments, Part I: straight-chain hydrocarbons and carboxylic acidsof some temperate lacustrine and subtropical lagoonal/tidal flatsediments. Chem. Geol. 18: 21–38.
Brooks, P. W., G. Eglinton, S. J. Gaskell, D. J. McHugh,J. R. Maxwell & R. P. Philp, 1977. Lipids of recent sedi-ments, Part II: branched and cyclic alkanes and alkanoic acidsof some temperate lacustrine and subtropical lagoonal/tidal flatsediments. Chem. Geol. 20: 189–204.
Burlingame, A. L., P. A. Haug, H. K. Schnoes & B. R. T. Simoneit,1969. Fatty acids derived from the Green River Formation oilshale by extractions and oxidations – a review. In P. A. Schenck& I. Havenaar (eds), Advances in Organic Geochemistry 1968,Pergamon, Oxford: 85–128.
Campbell, I. M. & J. Noworal, 1969. Composition of the saturatedand mono-unsaturated fatty acids ofMycobacterium phlei. J.Lipid Res. 10: 593–598.
Cardoso, J. N., C. D. Watts, J. R. Maxwell, R. Goodfellow, G. Eglin-ton & S. Golubic, 1978. A biogeochemical study of the AbuDhabi algal mats: a simplified ecosystem. Chem. Geol. 23:273–291.
Cardoso, J. N., S. J. Gaskell, M. M. Quirk & G. Eglinton, 1983. Hy-drocarbon and fatty acid distributions in Rostherne lake sediment(England). Chem. Geol. 38: 107–128.
Chappe, B. & P. Albrecht, 1982. Polar lipids of Archaebacteria insediments and petroleums. Science 217: 65–66.
Chappe, B., W. Michaelis & P. Albrecht, 1980. Molecular fossilsof Archaebacteria as selective degradation products of kerogen.In A. G. Douglas & J. R. Maxwell (eds), Advances in OrganicGeochemistry 1979, Pergamon, Oxford: 265–274.
hysh-6.tex; 18/11/1998; 13:21; p.21

98
Chicarelli, M. I., L. P. Damasceno & J. N. Cardoso, 1984. Diage-netic chemistry of the Paraiba Valley oil shale. In P. A. Schenck,J. W. de Leeuw & G. W. M. Lijmbach (eds), Advances in OrganicGeochemistry 1983, Org. Geochem. 6: 153–155.
Collister, J. W., R. E. Summons, E. Lichtfouse & J. M. Hayes, 1992.An isotopic biogeochemical study of the Green River oil shale.In C. B. Eckardt, J. R. Maxwell, S. R. Larter & D. A. C. Manning(eds), Advances in Organic Geochemistry 1991, Org. Geochem.19: 265–276.
Connan, J., 1984. Biodegradation of crude oils in reservoirs. InJ. Brooks & D. Welte (eds), Advances in Petroleum Geochem-istry, Vol. 1, Academic Press, London: 299–336.
Cooper, W. J. & M. Blumer, 1968. Linear, iso and anteiso fattyacids in recent sediments of the North Atlantic. Deep Sea Res.15: 535–540.
Corbet, B., P. Albrecht & G. Ourisson, 1980. Photochemical andphotomimetic fossil triterpenoids in sediments and petroleum. J.Am. Chem. Soc. 102: 1171–1173.
Cranwell, P. A., 1973. Branched-chain and cyclopropanoid acids ina recent sediment. Chem. Geol. 11: 307–313.
Cranwell, P. A., 1974. Monocarboxylic acids in lake sediments:indicators, derived from terrestrial and aquatic biota, of paleoen-vironmental trophic levels. Chem. Geol. 14: 1–14.
Cranwell, P. A., 1978. Extractable and bound lipid componentsin freshwater sediment. Geochim. Cosmochim. Acta 42: 1523–1532.
Cranwell, P. A., 1981. Lipid geochemistry of sediments from UptonBroad, a small productive lake. Org. Geochem. 3: 79–89.
Cranwell, P. A., 1982. Lipids of aquatic sediments and sedimentingparticulates. Prog. Lipid Res. 21: 271–308.
Cranwell, P. A., 1983. Alkyl esters in Recent sediments of two pro-ductive lakes. In M. Bjorøy et al. (eds), Advances in OrganicGeochemistry 1981, Wiley, Chichester: 299–308.
Cranwell, P. A., 1984. Alkyl esters, mid-chain ketones and fattyacids in late gacial and postglacial lacustrine sediments. InP. A. Schenck, J. W. de Leeuw & G. W. M. Lijmbach (eds),Advances in Organic Geochemistry 1983, Org. Geochem. 6:115–124.
Cranwell, P. A. & J. K. Volkman, 1981. Alkyl and steryl esters inrecent lacustrine sediment. Chem. Geol. 32: 29–43.
Cox, R. E., J. R. Maxwell, G. Eglinton, C. T. Pillinger, R. G. Ack-man & S. N. Hooper, 1970. The geological fate of chlorophyll:the absolute sterochemistries of a series of acyclic isoprenoidacids in a 50 million year old lacustrine sediment. J. Chem. Soc.Chem. Commun.: 1639–1641.
Curiale, J. A. & J. R. Odermatt, 1989. Short-term biomarker vari-ability in the Monterey Formation, Santa Maria Basin. Org.Geochem. 14: 1–13.
Cyr, T. D. & O. P. Strausz, 1984. Bound carboxylic acids in theAlberta oil sands. Org. Geochem. 7: 127–140.
Dahl, B., J. M. Moldowan, M. A. McCaffrey & P. A. Lipton, 1992.3-Alkyl steranes in petroleum: evidence for a new class of naturalproducts. Nature 355: 154–157.
Dany, F., J. Riolo, J. M. Trendel & P. Albrecht, 1990. 3β-carboxysteranes, a novel family of fossil steroids. J. Chem. Soc.,Chem. Commun.: 1228–1230.
Dastillung, M. & P. Albrecht, 1977.12-sterenes as diageneticintermediates in sediments. Nature 269: 678–679.
DeBaar, H. J. W., J. W. Farrington & S. G. Wakeham, 1983. Verticalflux of fatty acids in the North Atlantic Ocean. J. Mar. Res. 41:19–41.
De Rosa, M., A. Gambacorta, B. Nicolaus, H. N. M. Ross,W. D. Grant & J. D. Bu’lock, 1982. An asymmetric archaebacte-
rial diether from alkaliphilic halophiles. J. Gen. Microbiol. 128:343–348.
De Rosa, M., A. Gambacorta & A. Gliozzi, 1986. Structure, Biosyn-thesis, and physicochemical properties of archaebacterial lipids.Microbiol. Rev. 50: 70–80.
Didyk, B. M., B. R. T. Simoneit, S. C. Brassell & G. Eglinton,1978. Organic geochemical indicators of palaeoenvironmentalconditions of sedimentation. Nature 272: 216–222.
Douglas, A. G., K. Douraghi-Zadeh, G. Eglinton, J. R. Maxwell& J. N. Ramsay, 1970. Fatty acids in sediments including theGreen River shale (Eocene) and Scottish torbanite (Carbonifer-ous). In G. D. Hobson & G. C. Speers (eds), Advances in OrganicGeochemistry 1966, Pergamon Press, Oxford: 315–334.
Douglas, A. G., M. Blumer, G. Eglinton & K. Douraghi-Zadeh,1971. gas chromatographic-mass spectrometric characterizationof naturally-occurring acyclci isoprenoid carboxylic acids. Tetra-hedron 27: 1071–1092.
Dowling, N. J. E., F. Widdel & D. C. White, 1986. Phospholipidester-linked fatty acid biomarkers of acetate-oxidizing sulphate-reducers and other sulphide-forming bacteria. J. Gen. Microbiol.132: 1815–1825.
Eglinton, G., 1969. Hydrocarbons and fatty acids in living organ-isms and recent and ancient sediments. In P. A. Schenck &I. Havenaar (eds), Advances in Organic Geochemistry 1968,Pergamon Press, Oxford: 1–24.
Erwin, J. A., 1973. Comparative biochemistry of fatty acids ineukaryotic microorganisms. In J. A. Erwin (ed.), Lipids andBiomembranes of Eukaryotic Microorganisms, Pt. 2, AcademicPress, New York, London: 45–145.
Fowler, M. G. & A. G. Douglas, 1987. Saturated hydrocarbon bio-markers in oils of late Precambrian age from Eastern Siberia.Org. Geochem. 11: 201–213.
Freeman, K. H., J. M. Hayes, J. M. Trendel & P Albrecht, 1990. Ev-idence from GC-MS carbon isotopic measurements for multipleorigins of sedimentary hydrocarbons. Nature 353: 254–256.
Fu Jiamo, Sheng Guoying, Peng Pingan, S. C. Brassell, G. Eglinton& Jiang Jigang, 1986. Pecularities of salt lake sediments as po-tential source rocks in China. In D. Leythaeuser & J. Rullkötter(eds), Advances in Organic Geochemistry 1985, Org. Geochem.10: 119–126.
Fu Jiamo, Sheng Guoying, Xu Jiayou, Jia Rongfen, Fan Shanfa,Peng Ping’an, G. Eglinton & A. P. Gowar, 1992. Biomarker com-pounds as indicators of paleoenvironments. Chinese J. Geochem.11: 1–12.
Fukushima, K. & R. Ishiwatari, 1984. Acid and alcohol composi-tions of wax esters in sediments from different environments.Chem. Geol. 47: 41–56.
Gaskell, S. J., R. J. Morris, G. Eglinton, S. E. Calvert, 1985. Thegeochemistry of a recent sediment off northwest Africa. An as-sessment of the source of input and early diagenesis. Deep SeaRes. 22: 777–789.
Gaskell, S. J., M. M. Rhead, P. W. Brooks & G. Eglinton, 1986.Diagenesis of oleic acid in an estuarine sediment. Chem. Geol.17: 319–324.
Gillan, F. T. & R. B. Johns, 1986. Chemical markers for marinebacteria: fatty acids and pigments. In R. B. Johns (ed.), Biolog-ical Markers in the Sedimentary Record, Elsevier, Amsterdam:291–310.
Goossens, H., R. R. Düren, J. W. de Leeuw & P. A. Schenck, 1989.Lipids and their mode of occurrence in bacteria and sediments– II. Lipids in the sediment of a stratified, freshwater lake. Org.Geochem. 14: 27–41.
hysh-6.tex; 18/11/1998; 13:21; p.22

99
Han, J. C.-Y., 1970. Chemical studies of terrestrial and extrater-restrial life. Ph.D. Thesis, University of California, Berkeley,California, U.S.A.
Harwood, J. L. & N. J. Russell, 1984. Lipids in Plants and Microbes.London, Allen & Unwin.
Haug, P., H. K. Schnoes & A. L. Burlingame, 1971. Studies of theacidic components of the Colorado Green River formation oilshale: mass spectrometric identification of the methyl esters ofthe extractable acids. Chem. Geol. 7: 213–236.
ten Haven, H. L., J. W. de Leeuw & P. A. Schenck, 1985. Organicgeochemical studies of a Messinian evaporitic basin, northernApennines (Italy) I: Hydrocarbon biomarkers for a hypersalineenvironment. Geochimica Cosmochimica Acta 49: 2188–2191.
ten Haven, H. L., M. Baas, M. Kroot, J. W. de Leeuw, P. A. Schenck& J. Ebbing, 1987a. Late Quaternary Mediterranean sapropels.III: assessment of source of input and palaeotemperature as de-rived from biological markers. Geochim. Cosmochim. Acta 51:803–810.
ten Haven, H. L., J. W. de Leeuw, J. Rullkötter, J. S. SinningheDamsté, 1987b. Restricted utility of the pristane/phytane ratio asa palaeoenvironmental indicator. Nature 330: 641–643.
ten Haven, H. L., J. W. de Leeuw, J. S. Sinninghe Damsté,P. A. Schenck, S. E. Palmer & J. E. Zumberge, 1988. Applica-tion of biological markers in the recognition of paleohypersalineenvironments. In A. J. Fleet, K. Kelts & M. R. Talbot (eds),Lacustrine Petroleum Source Rocks, Geological Society SpecialPublication No. 40, Blackwell, Oxford: 123–130.
Hayes, J. M., K. H. Freeman, B. N. Popp & C. H. Hoham, 1990.Compound-specific isotopic analyses: a novel tool for recon-struction of ancient biogeochemical processes. In B. Durand &F. Behar (eds), Advances in Organic Geochemistry 1989, Org.Geochem. 16: 1115–1128.
Hedges, J. I. & F. G. Prahl, 1993. Early diagenesis: consequencesfor applications of molecular biomarkers. In M. H. Engel &S. A. Macko (eds), Organic Geochemistry, Principles and Ap-plications, Plenum Press, New York: 237–253.
Holzer, G., 1983. Lipids from methane-producing and sulfate-reducing bacteria and their geochemical significance. In P. Mac-Carthy (ed.), Research in Chemistry and Geochemistry at Col-orado School of Mines, a Tribute to L. J. Beckham, Pt. 2,Colorado School of Mines Quarterly 78: 9–13.
Holzer, G., J. Oró & T. G. Tornabene, 1979. Gas chromatographic-mass spectrometric analysis of neutral lipids from methanogenicand thermoacidophilic bacteria. J. Chromatogr. 186: 795–809.
Ishiwatari, I., S. Yamamoto & K. Fukushima, 1980. Possible inter-actions of insoluble organic geopolymers (kerogen) with organiccompounds in the aquatic environment – I. Interaction with fattyacid methyl esters. In A. G. Douglas & J. R. Maxwell (eds),Advances in Organic Geochemistry 1979, Advances in OrganicGeochemistry 1979, Pergamon, Oxford: 495–503.
Jaffé, P. R., P. Albrecht & J. L. Oudin, 1988. Carboxylic acids as in-dicators of migration. II: Case of the Mahakam Delta, Indonesia.Geochim. Cosmochim. Acta 52: 2599–2607.
Jaffé, R. & M. T. Gallardo, 1993. Application of carboxylic acidbiomarkers as indicators of biodegradation and migration ofcrude oils from the Maracaibo Basin, western Venezula. Org.Geochem. 20: 973–984.
Jaffé, R. & M. T. Gardinali, 1990. Generation and maturation ofcarboxylic acids in ancient sediments from the Maracaibo Basin,Venezuela. In B. Durand & F. Behar (eds), Advances in OrganicGeochemistry 1989, Org. Geochem. 16: 211–218.
Javor, B., 1989. Hypersaline Environments: Microbiology andBiogeochemistry. Springer, Berlin.
Jiang Jigang, 1985. About the source of oil in the QianjiangFormation. Acta Petrolei Sinica 6: 23–30 (in Chinese).
Jiang Jigang & Zhang Qian, 1982. The generation and evolution ofpetroleum of the saline Qianjiang Formation in Jianghan Basin.Oil & Gas Geol. Sinica 4: 1–15.
Johnson, R. W. & J. A. Calder, 1973. Early diagenesis of fattyacids and hydrocarbons in a salt marsh environment. Geochim.Cosmochim. Acta 37: 1943–1955.
Kaplan, I. R. & M. J. Baedecker, 1970. Biological productivity inthe Dead Sea, Part II. Evidence for phosphatidyl glycerophos-phate lipid in sediment. Israel J. Chem. 8: 529–533.
Kates, M., 1978. The phytanyl ether-linked polar lipids and iso-prenoid neutral lipids of extremely halophilic bacteria. Prog.Chem. Fats Other Lipids 15: 301–342.
Kawamura, K. & R. Ishiwatari, 1981. Polyunsaturated fatty acidsin a lacustrine sediment as a possible indicator of paleoclimate.Geochim. Cosmochim. Acta 45: 149–155.
Kawamura, K. & R. Ishiwatari, 1984. Fatty acid geochemistry ofa 200 m sediment core from Lake Biwa, Japan. Early diagene-sis and paleoenvironmental information. Geochim. Cosmochim.Acta 48: 252–266.
Kawamura, K. & R. Ishiwatari, 1985. Distribution of lipid classcompounds in bottom sediments of freshwater lakes with differ-ent tropic status in Japan. Chem. Geol. 51: 123–133.
Kawamura, K., R. Ishiwatari & M. Yamazaki, 1980. Identificationof polyunsaturated fatty acids insurface lacustrine sediments.Chem. Geol. 28: 31–39.
Kerger, B. D., P. D. Nichol, C. P. Anthworth, E. Bock, J. C. Cox,T. A. Langworthy & D. C. White, 1986. Signature fatty acids inthe polar lipids of acid-producingThiobacillusspp.: methyoxy,cyclopropyl, alpha-hydroxycyclopropyl and branched and nor-mal monoenoic fatty acids. FEMS Microbiol. Ecol. 38: 67–77.
Kerwin, J. L., 1994. Evolution of structure and function of fattyacids and their metabolites. In W. D. Nes (ed.), Isopentenoidsand Other Natural Products: Evolution and Function, AmericanChemical Society Symposium Series 562: 163–201.
Kvenvolden, K. A., 1970. Evidence for transformations of nor-mal fatty acids in sediments. In G. D. Hobson & G. C. Speers(eds), Advances in Organic Geochemistry 1966, PergamonPress, Oxford: 335–366.
Langworthy, T. A., 1985. Lipids of Archaebacteria. In C. R. Woese& R. S. Wolfe (eds), Archaebacteria, Academic Press, New York:459–497.
Langworthy, T. A., G. Holzer, J. G. Zeikus & T. G. Tornabene,1983. Iso- and anteiso-branched glycerol diethers of the ther-mophilic aerobe Thermodisulfoto-bacterium commune system.Appl. Microbiol. 101: 1–17.
de Leeuw, J. W. & J. S. Sinninghe Damsté, 1990. Organic sulfurcompounds and other biomarkers as indicators of palaeosalinity:a critical evaluation. In W. L. Orr & C. M. White (eds), Geo-chemistry of Sulfur in Fossil Fuels, American Chemical SocietySymposium Series 429: 417–443.
de Leeuw, J. W., J. S. Sinninghe Damsté, J. Klok, P. A. Schenck& J. J. Boon, 1985. Biogeochemistry of Gavish Sabkha sedi-ments I. Studies on neutral reducing sugars and lipid moietiesby gas chromatography-mass spectrometry. In G. M. Friedman& W. E. Krumbein (eds), Hypersaline Ecosystems. The GavishSabhka, Springer, Berlin: 350–367.
Lechevalier, H. & M. P. Lechevalier, 1988. Chemotaxonomic useof lipids – an overview. In C. Ratledge & S. G. Wilkinson (eds),Microbial Lipids, Vol. 1, Academic, London: 869–902.
Leenheer, M. J. & P. A. Meyers, 1983. Comparison of lipid com-positions in marine and lacustrine sediments. In M. Bjorøy
hysh-6.tex; 18/11/1998; 13:21; p.23

100
et al. (eds), Advances in Organic Geochemistry 1981, Wiley,Chichester: 309–316.
Leenheer, M. J., K. D. Flessland & P. A. Meyers, 1984. Compari-son of lipid character of sediments from the Great Lakes and theNorthwestern Atlantic. Org. Geochem. 7: 141–150.
Li Ren Wei, Wu Hanguo, Lin Daxing, Wang Zhizhen, Chen Rijin,Tian Xiangyou & Zhang Rufan, 1992. Biomarker features of theChinese Mesozoic, Cenozoic saline lake deposits. In Advancesin Geoscience, Inst. of Geology, Chinese Academy of Sciences:275–295.
Logan, G. A. & G. Eglinton, 1994. Biogeochemistry of theMiocene lacustrine deposit, at Clarkia northern Idaho, U.S.A.Org. Geochem. 21: 857–870.
Ma Xiuzhen, Yin Mingshan & Wang Dazhen, 1987. Sulfate-reducing bacteria and their geobiochemical role of salt lakesediments of China. Acta Sedimentologica Sinica 5: 57–61.
Mackenzie, A. S., S. C. Brassell, G. Eglinton & J. R. Maxwell,1982. Chemical fossils: the geological fate of steroids. Science217: 491–504.
Mackenzie, A. S., R. L. Patience, J. R. Maxwell, M. Vandenbroucke& B. Durand, 1980. Molecular parameters of maturation in theToarcian shales, Paris Basin, France – I. Changes in the config-urations of acyclic isoprenoid alkanes, steranes and triterpanes.Geochim. Cosmochim. Acta 44: 1709–1721.
Mackenzie, A. S., G. A. Wolff & J. R. Maxwell, 1983. Fatty acids insome biodegraded petroleums. Possible origins and significance.In M. Bjorøy et al. (eds), Advances in Organic Geochemistry1981, Wiley, Chichester: 637–649.
Matsuda, H., 1978. Early diagenesis of fatty acids in lacustrinesediments: III. Changes in fatty acid composition in the sedi-ments from a brackish water lake. Geochim. Cosmochim. Acta42: 1027–1034.
Matsuda, H. & T. Koyama, 1977a. Early diagenesis of fatty acidsin lacustrine sediments – I. Identification and distribution offatty acids in recent sediment from a freshwater lake. Geochim.Cosmochim. Acta 41: 777–783.
Matsuda, H. & T. Koyama, 1977b. Early diagenesis of fatty acidsin lacustrine sediments – II. A statistical approach to changesin fatty acid composition from recent sediments and sourcematerials. Geochim. Cosmochim. Acta 41: 1825–1834.
McEvoy, J., 1983. The origin and diagenesis of organic lipids insediments from the San Miguel Gap. Ph.D. Thesis. University ofBristol, U.K.
Meyers, P. A. & N. Takeuchi, 1979. Fatty acids and hydrocarbons insurficial sediments of Lake Huron. Org. Geochem. 1: 127–138.
Meyers, P. A. & R. Ishiwatari, 1993. The early diagenesis oforganic matter in lacustrine sediments. In M. H. Engel &S. A. Macko (eds), Organic Geochemistry: Principles and Ap-plications, Plenum, New York: 185–209.
Meyers, P. A. & R. Ishiwatari, 1994. Lacustrine organic geochem-istry – an overview of indicators of organic matter sources anddiagenesis in lake sediments. Org. Geochem. 20: 867–900.
Meyers, P. A., H. B. Maring & R. A. Bourbonniere, 1980. Alkaneand alkanoic acid variations with depth in modern sedimentsof Pyramid Lake. In A. G. Douglas & J. R. Maxwell (eds),Advances in Organic Geochemistry 1979, Pergamon, Oxford:365–374.
Miranda, R. M. & C. C. Walters, 1992. Geochemical variations insedimentary organic matter within a ‘homogeneous’ shale core(Tuscaloosa Formation, Upper Cretaceous, Mississippi, U.S.A.Org. Geochem. 18: 899–911.
Moldowan, J. M., P. Sundararaman & M. Schoell, 1986. Sen-sitivity of biomarker properties to depositional environmentand/or source input in the Lower Toarcian of SW-Germany.
In D. Leythaeuser & J. Rullkötter (eds), Advances in OrganicGeochemistry 1985. Org. Geochem. 10: 915–926.
Mycke, B. & W. Michaelis, 1986. Molecular fossils fromchemical degradation of macromolecular organic matter. InD. Leythaeuser & J. Rullkötter (eds), Advances in OrganicGeochemistry 1985. Org. Geochem. 10: 847–858.
Mycke, B., F. Narjes & W. Michaelis, 1987. Bacteriohopanetetrolfrom chemical degradation of an oil shale kerogen. Nature 326:179–181.
Nissenbaum, A., M. J. Baedecker & I. R. Kaplan, 1972. Organicgeochemistry of Dead Sea sediments. Geochim. Cosmochim.Acta 36: 709–727.
O’Leary, W. M. & S. G. Wilkinson, 1988. Gram-positive bacteria.In C. Ratledge & S. G. Wilkinson (eds), Microbial Lipids, Vol.1, Academic, London: 117–201.
Oremland, R. S., 1988. Biogeochemistry of methanogenic bacteria.In A. J. B. Zehnder (ed.), Biology of Anaerobic Microorganisms,Wiley, New York: 641–705.
Oremland, R. S., J. E. Cloern, Z. Sofer, R. L. Smith, C. W. Culbert-son, J. Zehr, L. Miller, B. Cole, R. Harvey, N. Iversen, M. Klug,D. J. DesMarais & G. Rau, 1988. Microbial and biogeochemicalprocessed in Big Soda Lake, Nevada. In A. J. Fleet, K. Kelts &M. R. Talbot (eds), Lacustrine Petroleum Source Rocks, Geo-logical Society Special Publication No. 40, Blackwell, Oxford:59–75.
Ourisson, G., P. Albrecht & M. Rohmer, 1979. The hopanoids:palaeochemistry and biochemistry of a group of natural products.Pure Appl. Chem. 51: 709–729.
Parkes, R. J. & J. Taylor, 1983. The relationship between fattyacid distributions and bacterial respiratory types in contemporarymarine sediments. Estuarine, Coastal Shelf Sci. 3: 311–319.
Perry, G. J., J. K. Volkman, R. B. Johns & H. J. Bavor, 1979.Fatty acids of bacterial origin in contemporary marine sediments.Geochim. Cosmochim. Acta 43: 1715–1725.
Philp, R. P., S. Brown, M. Calvin, S. C. Brassell & G. Eglin-ton, 1978. Hydrocarbon and fatty acid distributions in recentlydeposited algal mats at Laguna Guerrero, Baja California. InW. E. Krumbein (ed.), Environmental Biogeochemistry and Ge-omicrobiology: Vol. 1. The Aquatic Environment, Ann ArborScience, Ann Arbor: 255–270.
Philp, R. P. & Z. Fan, 1987. Geochemical investigation of oils andsource rocks from Qianjiang Depression of Jianghan Basin, aterrigenous saline basin, China. Org. Geochem. 11: 549–562.
Prahl, F. G., J. M. Hayes & T. M. Xie, 1992. An indicator of terrige-nous organic carbon in Washington coastal sediments. Limnol.Oceanogr. 37: 1290–1300.
Prattl, L. M. & C. L. Davis, 1992. Intertwined fates of metals, sulfur,and organic carbon in black shales. In L. M. Pratt, J. B. Comer& S. C. Brassell (eds), Geochemistry of Organic Matter inSediments and Sedimentary Rocks, SEPM Short Course 27:1–27.
Quirk, M. M., A. M. K. Wardroper, P. W. Brooks, R. E. Wheatley& J. R. Maxwell, 1980. Transformations of acyclic and cyclicisoprenoids in recent sedimentary environments. In R. Daumas(ed.), Biogéochimie de la matière organique à l’interface eau-sédiment marin, Éditions du CNRS, Paris: 225–232.
Ratledge, C. & S. G. Wilkinson, 1988. An overview of microbiallipids. In C. Ratledge & S. G. Wilkinson (eds), Microbial Lipids,Vol. 1, Academic, London: 3–22.
Risatti, J. B., S. J. Rowland, D. A. Yon & J. R. Maxwell, 1984.Stereochemical studies of acyclic isoprenoids – XII. Lipids ofmethanogenic bacteria and possible contributions to sediments.In P. A. Schenck, J. W. de Leeuw & G. W. M. Lijmbach. Org.Geochem. 6: 93–104.
hysh-6.tex; 18/11/1998; 13:21; p.24

101
Robinson, N. & G. Eglinton, 1990. Lipid chemistry of Iceland hotspring microbial mats. Org. Geochem. 15: 291–298.
Schaeffer, P., Fache-Dany, J. M. Trende & P. Albrecht, 1993. Polarconstituents of organic matter rich marls from evaporitic seriesof the Mulhouse basin. Org. Geochem. 20: 1227–1236.
Schmitter, J. M., P. Arpino & G. Guiochon, 1978. Investigationof high molecular weight carboxylic acids in petroleum by dif-ferent combinations of chromatography (gas and liquid) andmass spectrometry (electron impact and chemical ionization). J.Chromatogr. 167: 149–158.
Schmitter, J. M., P. J. Arpino & G. Guiochon, 1981. Isolation ofdegraded pentacyclic triterpenoid acids in a Nigerian crude oiland their identification as tetracyclic carboxylic acids resultingfrom ring A cleavage. Geochim. Cosmochim. Acta 45: 1951–1955.
Schoell, M., M. A. McCaffrey, F. J. Fago & J. M. Moldowan,1992. Carbon isotopic compositions of 28,30-bisnorhopanes andother biological markers in a Monterey crude oil. Geochim.Cosmochim. Acta 56: 1391–1399.
Seifert, W. K., 1973. Steroid acids in petroleum, animal contributionto the origin of petroleum. Pure Appl. Chem. 34: 633–640.
Seifert, W. K., 1975. Carboxylic acids in petroleum and sediments.Prog. Chem. Org. Nat. Prod. 32: 1–49.
Seifert, W. K. & J. M. moldowan, 1980. The effect of thermal stresson souce rock quality as measured by hopane sterochemistry.In A. G. Douglas & J. R. Maxwell (eds), Advances in OrganicGeochemistry 1979, Pergamon, Oxford: 229–237.
Seifert, W. K. & R. M. Teeter, 1970a. Identification of polycyclicnaphthenic, mono-, and diaromatic crude oil carboxylic acids.Anal. Chem. 42: 180.
Seifert, W. K. & R. M. Teeter, 1970b. Identification of poly-cyclic aromatic and heterocyclic crude oil carboxylic acids. Anal.Chem. 42: 750–758.
Seifert, W. K., E. J. Gallegos & R. M. Teeter, 1971. First identifi-cation of a steroid carboxylic acid in petroleum. Angew. Chem.,Int. Ed. Engl. 10: 747–748.
Seifert, W. K., E. J. Gallegos & R. M. Teeter, 1972. Proof ofstructure of steroid carboxylic acids in a California petroleum bydeuterium labelling, synthesis and mass spectrometry. J. Chem.Soc. 94: 5880–5887.
Shaw, G. J., R. A. Franich, G. Eglinton, J. Allan & A. G. Douglas,1980. Diterpenoid acids in Yallourn lignite. In A. G. Douglas &J. R. Maxwell (eds), Advances in Organic Geochemistry 1979,Pergamon, Oxford: 281–286.
Sheia, J., S. C. Brassell & D. M. Ward, 1990. Mid-chain branchedmono- and dimethyl alkanes in hot spring cyanobacterial mats:a direct biogenic source for branched alkanes in ancient sedi-ments? Org. Geochem. 15: 223–231.
Sheng Guoying, Fan Shangfa, Liu Dehan, Su Nengxian & ZhouHongming, 1980. The geochemistry ofn-alkanes with an even-odd predominance in the Tertiary Shahejie Formation of northernChina. In A. G. Douglas & J. R. Maxwell (eds), Advances inOrganic Geochemistry 1979, Pergamon, Oxford: 115–121.
Shimoyama, A. & W. D. Johns, 1972. Formation of alkanes fromfatty acids in the presence of CaCO3. Geochim. Cosmochim.Acta 36: 87–91.
Simoneit, B. R. T., 1977. Diterpenoid compounds and otherlipids in deep-sea sediments and their geochemical significance.Geochim. Cosmochim. Acta 41: 463–476.
Simoneit, B. R. T., 1978. In J. P. Riley & R. Chester (eds), ChemicalOceanography, Vol. 7, Academic, London: 233–311.
Sinninghe Damsté, J. S., J. W. de Leeuw, A. C. Kock Van Dalen,M. A. de Zeeuw, F. de Lange, W. I. C. Rijpstra & P. A. Schenck,1987a. The occurrence and identification of series of organic
sulphur compounds in oils and sediment extracts. I. A studyof Rozel Point oil (USA). Geochim. Cosmochim. Acta 51:2361–2391.
Sinninghe Damsté, J. S., A. C. Kock Van Dalen, J. W. deLeeuw, P. A. Schenck, Sheng Guoying & S. C. Brassell, 1987b.The identification of mono-, di-, and trimethyl 2-methyl-2-(4,8,12-trimethyltridecyl)chromans and their occurrence in thegeosphere. Geochim. Cosmochim. Acta 51: 2393–2400.
Sinninghe Damsté, J. S., T. I. Eglinton, J. W. de Leeuw &P. A. Schenck, 1989a. Organic sulphur in macromolecular sed-imentary organic matter: I. Structure and origin of sulphur-containing moieties in kerogen, asphaltenes and coals as revealedby flash pyrolysis. Geochim. Cosmochim. Acta 53: 873–889.
Sinninghe Damsté, J. S., W. I. C. Rijpstra, J. W. de Leeuw& P. A. Schenck, 1989b. The occurrence and identificationof series of organic sulphur compounds in oils and sedimentextracts. II. Their presence in samples from hypersaline andnon-hypersaline depositional environments and their possible ap-plication as source, palaeoenvironmental and maturity indicators.Geochim. Cosmochim. Acta 53: 1423–1441.
Sinninghe Damsté, J. S., W. I. C. Rijpstra, A. C. Kock Van Dalen,J. W. de Leeuw & P. A. Schenck, 1989c. Quenching of labilefunctionalised lipids by inorganic sulphur species: evidence forthe formation of sedimentary organic sulphur compounds at theearly stage of diagenesis. Geochim. Cosmochim. Acta 53: 1443–1455.
Smith, P. F., 1988. Archaebacteria and other specialized bacteria. InC. Ratledge & S. G. Wilkinson (eds), Microbial Lipids, Vol. 1,Academic, London: 489–553.
Smith, D. J., G. Eglinton & R. J. Morris, 1983a. Interfacial sedimentand assessment of organic input from a highly productive watercolumn. Nature 304: 259–262.
Smith, D. J., G. Eglinton & R. J. Morris, 1983b. The lipid chemistryof an interfacial sediment from the Peru Continental Shelf: fattyacids, alcohol, aliphatic ketones and hydrocarbons. Geochim.Cosmochim. Acta 47: 2225–2232.
Summons, R. E., 1987. Branched alkanes from ancient and mod-ern sediments: isomer discrimination by GC/MS with multiplereaction monitoring. Org. Geochem. 11: 281–289.
Summons, R. E. & R. J. Capon, 1988. Fossil steranes with unprece-dented methylation in ring-A. Geochim. Cosmochim. Acta 52:2733–2736.
Summons, R. E., S. C. Brasell, G. Eglinton, E. Evans,R. J. Horodyski, N. Robinson & D. M. Ward, 1988. Distinc-tive hydrocarbon biomarkers from fossiliferous sediment of theLate Proterozoic Walcott member, Chuar Group, Grand Canyon,Arizona. Geochim. Cosmochim. Acta 52: 2625–2637.
Suzuki, N. & K. Taguchi, 1984. Diagenesis of extractable and boundfatty acids in possible source rocks in Japan. In P. A. Schenck,J. W. de Leeuw & G. W. M. Lijmbach (eds), Advances in OrganicGeochemistry 1983, Org. Geochem. 6: 125–133.
Tannenbaum, E., E. Ruth & I. R. Kaplan, 1986. Steranes andtriterpanes generated from kerogen pyrolysis in the absence andpresence of minerals. Geochim. Cosmochim. Acta 50: 805–812.
Tegelaar, E. W., J. W. de Leeuw, S. Derenne & C. Largeau, 1989.A reappraisal of kerogen formation. Geochim. Cosmochim. Acta53: 3075–3106.
Tissot, B. P. & D. H. Welte, 1984. Petroleum Formation andOccurrence. Springer, Berlin.
Tornabene, T. G., T. A. Langworthy, G. Holzer & J. Oró, 1979.Squalenes, phytanes and other isoprenoids as major neutral lipidsof methanogenic and thermoacidophilic ‘archaebacteria’. J. Mol.Evol. 13: 73–83.
hysh-6.tex; 18/11/1998; 13:21; p.25

102
Trifilieff, S., O. Sieskind & P. Albrecht, 1992. Biological markersin petroleum asphaltenes: possible mode of incorporation. InJ. M. Moldowan, P. Albrecht & R. P. Philp (eds), BiologicalMarkers in Sediments and Petroleum: a Tribute to Wolfgang K.Seifert, Prentice-Hall, Englewood Cliffs: 350–369.
Valisolalao, J., N. Perakis, B. Chappe & P. Albrecht, 1984. A novelsulfur containing C35 hopanoid in sediments. Tetrahedron Lett.25: 1183–1186.
Van Dorsselaer, A., 1975. Triterpènes de sédiments. Thèse deDoctorat ès-Sciences, l’Université Louis Pasteur, Strasbourg,France.
Van Dorsselaer, A., A. Ensminger, C. Spyckerelle, M. Dastillung,O. Sieskind, P. Arpino, P. Albrecht, G. Ourisson, P. W. Brooks,S. J. Gaskell, B. J. Kimble, R. P. Philp, J. R. Maxwell & G. Eglin-ton, 1974. Degraded and extended hopane derivatives (C27 toC35) as ubiquitous geochemical markers. Tetrahedron Lett. 14:1349–1352.
Van Vleet, E. S. & J. G. Quinn, 1976. Characterisation of monoun-saturated fatty acids from an estuarine sediment. Nature 262:127–128.
Van Vleet, E. S. & J. G. Quinn, 1979. Early diagenesis of fattyacids and isoprenoid alcohols in estuarine and coastal sediments.Geochim. Cosmochim. Acta 43: 289–303.
Volkman, J. K. & R. B. Johns, 1977. The geochemical significanceof positional isomers of unsaturated acids from an intertidal zonesediment. Nature 267: 693–694.
Volkman, J. K. & J. R. Maxwell, 1986. Acyclic isoprenoids as bio-logical markers. In R. B. Johns (ed.), Biological Markers in theSedimentary Record, Elsevier, Amsterdam: 1–42.
Volkman, J. K., R. B. Johns, F. T. Gillan, G. J. Perry & H. J. Bavor,1980. Microbial lipids of an intertidal sediment – I. Fatty acidsand hydrocarbons. Geochim. Cosmochim. Acta 44: 1133–1143.
Wakeham, S. G. & C. Lee, 1989. Organic geochemistry of par-ticulate matter in the ocean: The role of particles in oceanicsedimentary cycles. Org. Geochem. 14: 83–96.
Wang Ruiliang, Fu Jiamo & Sheng Guoying, 1988a. Quantitativecomposition and distribution of biological markers in sedimentsof saline basins. In Annual Research Reports of the Organic Geo-chemistry Laboratory 1987 Institute of Geochemistry, ChineseAcademy of Sciences: 109–120.
Wang Zhizhen, Li Renwei & Lin Daxing, 1988b. Isoprenoid hy-drocarbons and the relation between them and sedimentaryenvironment. Oil Gas Geol. sinica 9: 140–145.
Wang Tieguan, Fan Pu & F. M. Swain, 1988c. Geochemical char-acteristics of crude oils and source beds in different continentalfacies of four oil-bearing basins, China. In A. J. Fleet, K. Kelts& M. R. Talbot (eds), Lacustrine Petroleum Source Rocks, Ge-ological Society Special Publication No. 40, Blackwell, Oxford:309–325.
Welte, D. & G. Ebhardt, 1968. Distribution of long chain n-paraffinsand n-fatty acids in sediments from the Persian Gulf. Geochim.Cosmochim. Acta 32: 465–466.
Wood, B. J. B., 1988. Lipids of algae and protozoa. In C. Ratledge& S. G. Wilkinson (eds), Microbial Lipids, Vol. 1, Academic,London: 807–867.
Wilkinson, S. G., 1988. Gram-negative bacteria. In C. Ratledge& S. G. Wilkinson (eds), Microbial Lipids, Vol. 1, Academic,London: 299–488.
Zeng, Y. B., D. M. Ward, S. C. Brassell & G. Eglinton, 1992. Bio-geochemistry of hot spring environments. 2. Lipid compositionsof Yellowstone cyanobacterial andChloroflexusmats. Chem.Geol. 95: 327–345.
Zeng, Y. B., D. M. Ward, S. C. Brassell & G. Eglinton, 1992. Bio-geochemistry of hot spring environments. 3. Apolar and polarlipids in the biologically active layers of a cyanobacterial mat.Chem. Geol. 95: 347–360.
Zheng Mianping, Liu Wengao & Xiang Jun, 1985. The discovery ofthe halophilic algae and halobacteria at Zabuye Salt Lake Tibetand preliminary study on the geoecology. Acta Geologica Sinica59: 162–170. (in Chinese)
hysh-6.tex; 18/11/1998; 13:21; p.26
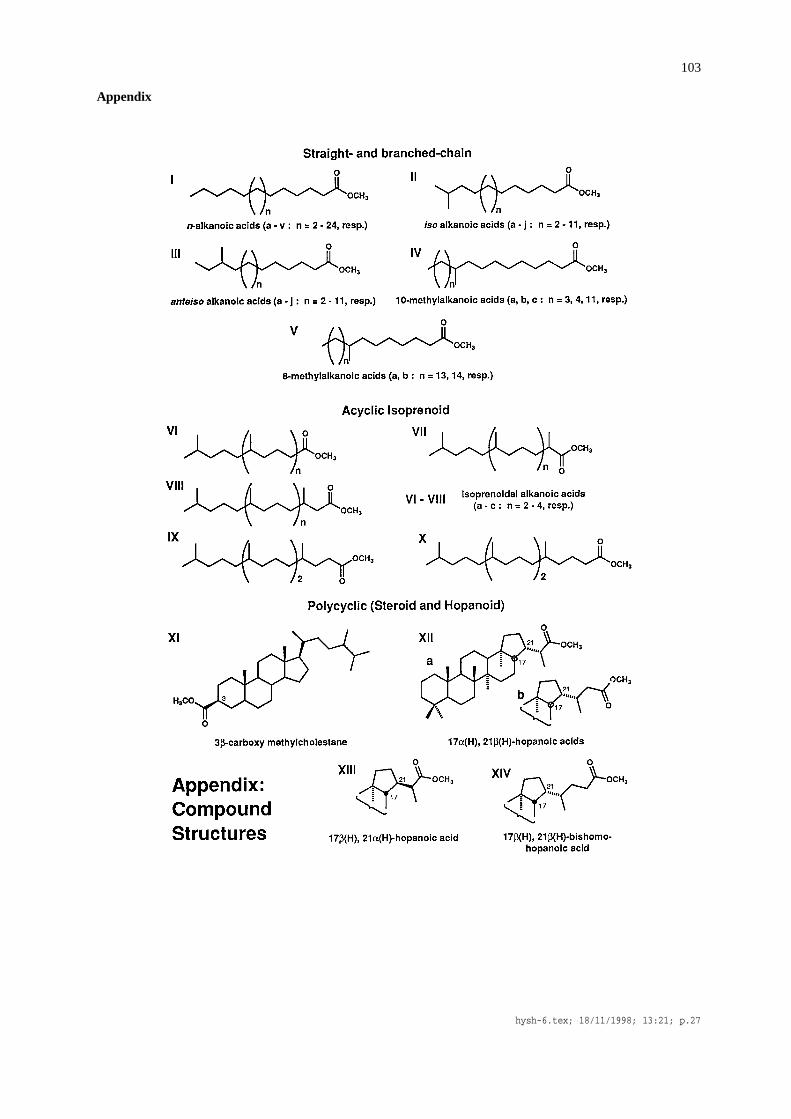
103
Appendix
hysh-6.tex; 18/11/1998; 13:21; p.27

hysh-6.tex; 18/11/1998; 13:21; p.28
Related Documents











