Downloaded from www.microbiologyresearch.org by IP: 54.144.197.59 On: Thu, 21 Apr 2016 03:50:37 Journal of General Virology (1997), 78, 2761–2770. Printed in Great Britain ........................................................................................................................................................................................................................................................................................... Molecular cloning of a defective hepatitis C virus genome from the ascitic fluid of a patient with hepatocellular carcinoma Chau-Ting Yeh, Su-Chuan Lu, Chia-Ming Chu and Yun-Fan Liaw Liver Research Unit, Chang-Gung Memorial Hospital and Medical College, 199 Tung Hwa North Road, Taipei, Taiwan A defective hepatitis C virus (HCV) genome in the ascitic fluid of a patient with hepatocellular car- cinoma was cloned and sequenced up to the 3« poly(U) stretch. When compared with the published Taiwanese HCV sequence, this defective genome contained deletions of single nucleotides at eight sites, double nucleotides at two sites, triple nucleo- tides at four sites, quadruple nucleotides at one site and replacement of a short stretch of sequence at one site. For comparison, the corresponding regions containing these mutations were also cloned from a serum sample from this patient. Except for deletions of two triple nucleotides in the hypervariable region, the reading frames of all serum-derived clones were intact. The defective HCV genome encoded a truncated core protein with Introduction Hepatitis C virus (HCV) is a major cause of non-A, non-B hepatitis worldwide. In a majority of patients, the virus causes chronic hepatitis which may lead to complications such as cirrhosis and hepatocellular carcinoma (Houghton, 1996). Since molecular cloning of HCV was achieved in 1989, several complete or nearly complete genomic sequences have been published (Chen et al., 1992 ; Choo et al., 1989, 1991 ; Hayashi et al., 1993 ; Inchauspe et al., 1991; Kato et al., 1990; Ogata et al., 1991 ; Okamoto et al., 1991, 1992 a ; Takamizawa et al., 1991). Comparison of the nucleotide and polyprotein amino acid sequences of these isolates revealed considerable het- erogeneity, leading to the classification of multiple genotypes Author for correspondence : Chau-Ting Yeh. Fax ›886 3 3282824. The GenBank accession number for the nucleotide sequence of the defective ascitic sequence reported in this paper is U89019 ; the numbers for the serum-derived partial sequences C-fs, HVR-fs, NS2-fs and NS3-fs are U89015, U89018, U89017 and U89016, respectively. 90 amino acid residues (the last 20 amino acid residues came from a different reading frame), whereas the serum-derived genome encoded a full- length core protein. When expressed in Huh-7 cells, these two proteins were localized to the nucleus and cytoplasm, respectively. Using specific primer-sets, ascites- and serum-derived genomes were each detected alone in ascitic fluid and serum samples, respectively, whereas both sequences were present in ascitic mononuclear cells. The defective sequence thus constituted the major virus population in the ascitic fluid whereas a putative helper genome coexisted with it inside the ascitic mononuclear cells. This sequence is possibly a defective and interfering genome. (Bukh et al., 1995 ; Okamoto et al., 1992 b ; Simmonds et al., 1993). The genomic RNA of HCV is about 9–4 kb in length. With the help of an internal ribosome entry site, a polyprotein of about 3000 amino acid residues is expressed, which is proteolytically processed to produce several structural and nonstructural proteins (Ali & Siddiqui, 1995 ; Shimotohno et al., 1995 ; Wang & Siddiqui, 1995). The long open reading frame is followed by a poly(U) homopolymeric tract, except for HCV-1, which contains a poly(A) tract (Han et al., 1991). Recently, a highly conserved sequence-element following the 3«-terminal poly(U) tract was identified (Kolykhalov et al., 1996 ; Tanaka et al., 1995). This sequence is predicted to form a stable stem–loop structure and is believed to be required for authentic HCV replication. When HCV sequences were cloned from serum, defective sequences were occasionally identified but in a very small proportion of the virus population (Martell et al., 1992). Interestingly, the nucleotide sequence of the core region from the tissue of a hepatocellular carcinoma revealed deletions and mutations resulting in truncated proteins (Ru $ ster et al., 1996). When the hypervariable regions of HCV sequences from 0001-4980 # 1997 SGM CHGB

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
Journal of General Virology (1997), 78, 2761–2770. Printed in Great Britain. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Molecular cloning of a defective hepatitis C virus genome fromthe ascitic fluid of a patient with hepatocellular carcinoma
Chau-Ting Yeh, Su-Chuan Lu, Chia-Ming Chu and Yun-Fan Liaw
Liver Research Unit, Chang-Gung Memorial Hospital and Medical College, 199 Tung Hwa North Road, Taipei, Taiwan
A defective hepatitis C virus (HCV) genome in theascitic fluid of a patient with hepatocellular car-cinoma was cloned and sequenced up to the 3«poly(U) stretch. When compared with the publishedTaiwanese HCV sequence, this defective genomecontained deletions of single nucleotides at eightsites, double nucleotides at two sites, triple nucleo-tides at four sites, quadruple nucleotides at one siteand replacement of a short stretch of sequence atone site. For comparison, the correspondingregions containing these mutations were alsocloned from a serum sample from this patient.Except for deletions of two triple nucleotides in thehypervariable region, the reading frames of allserum-derived clones were intact. The defectiveHCV genome encoded a truncated core protein with
IntroductionHepatitis C virus (HCV) is a major cause of non-A, non-B
hepatitis worldwide. In a majority of patients, the virus causeschronic hepatitis which may lead to complications such ascirrhosis and hepatocellular carcinoma (Houghton, 1996). Sincemolecular cloning of HCV was achieved in 1989, severalcomplete or nearly complete genomic sequences have beenpublished (Chen et al., 1992 ; Choo et al., 1989, 1991 ; Hayashiet al., 1993 ; Inchauspe et al., 1991 ; Kato et al., 1990 ; Ogata etal., 1991 ; Okamoto et al., 1991, 1992a ; Takamizawa et al.,1991). Comparison of the nucleotide and polyprotein aminoacid sequences of these isolates revealed considerable het-erogeneity, leading to the classification of multiple genotypes
Author for correspondence: Chau-Ting Yeh.
Fax 886 3 3282824.
The GenBank accession number for the nucleotide sequence of the
defective ascitic sequence reported in this paper is U89019; the
numbers for the serum-derived partial sequences C-fs, HVR-fs, NS2-fs
and NS3-fs are U89015, U89018, U89017 and U89016, respectively.
90 amino acid residues (the last 20 amino acidresidues came from a different reading frame),whereas the serum-derived genome encoded a full-length core protein. When expressed in Huh-7 cells,these two proteins were localized to the nucleus andcytoplasm, respectively. Using specific primer-sets,ascites- and serum-derived genomes were eachdetected alone in ascitic fluid and serum samples,respectively, whereas both sequences were presentin ascitic mononuclear cells. The defective sequencethus constituted the major virus population in theascitic fluid whereas a putative helper genomecoexisted with it inside the ascitic mononuclearcells. This sequence is possibly a defective andinterfering genome.
(Bukh et al., 1995 ; Okamoto et al., 1992b ; Simmonds et al.,1993). The genomic RNA of HCV is about 9±4 kb in length.With the help of an internal ribosome entry site, a polyproteinof about 3000 amino acid residues is expressed, which isproteolytically processed to produce several structural andnonstructural proteins (Ali & Siddiqui, 1995 ; Shimotohno et al.,1995 ; Wang & Siddiqui, 1995). The long open reading frameis followed by a poly(U) homopolymeric tract, except forHCV-1, which contains a poly(A) tract (Han et al., 1991).Recently, a highly conserved sequence-element following the3«-terminal poly(U) tract was identified (Kolykhalov et al.,1996 ; Tanaka et al., 1995). This sequence is predicted to forma stable stem–loop structure and is believed to be required forauthentic HCV replication.
When HCV sequences were cloned from serum, defectivesequences were occasionally identified but in a very smallproportion of the virus population (Martell et al., 1992).Interestingly, the nucleotide sequence of the core region fromthe tissue of a hepatocellular carcinoma revealed deletions andmutations resulting in truncated proteins (Ru$ ster et al., 1996).When the hypervariable regions of HCV sequences from
0001-4980 # 1997 SGM CHGB

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
C.-T. Yeh and othersC.-T. Yeh and others
ascites and serum samples were compared, we also discovereda defective sequence which constituted the major viruspopulation in the ascitic fluid of a patient with hepatocellularcarcinoma (Yeh et al., 1996). In this case, all 14 clones obtainedfrom the ascitic fluid contained a frameshift mutation, whereasall 10 clones from a serum sample of the same patient did not.This finding prompted us to clone the complete defectivesequence to understand its genetic structure. Selective regionsof HCV RNA from the serum sample were also cloned andsequenced in parallel for comparison.
Methods+ Patient. A male patient, 58 years of age, was diagnosed as havinghepatocellular carcinoma by liver biopsy and received surgical resectionin December 1993. Unfortunately, recurrence of liver tumours withmassive ascites was found in October 1994. Paracentesis was performedfor diagnostic purposes and also to relieve symptoms. A serum samplewas collected simultaneously. All samples were kept at ®70 °C until use.The serum was negative for hepatitis B virus surface antigen (HBsAg;Ausria-II, Abbott) but positive for anti-hepatitis C virus antibody (anti-HCV; UBI HCV EIA, United Biomedical).
+ RT nested PCR (RT–PCR), cloning and sequencing. Theascites and serum samples were first centrifuged to remove all cellularcomponents before analysis. RT–PCR was performed according to theprocedure described previously (Yeh et al., 1994b). Briefly, serum orascites samples (100 µl) were mixed with 600 µl extraction buffer (4 Mguanidinium thiocyanate, 2% Sarkosyl, 0±5 M mercaptoethanol, 5 µgtRNA, 5 mM sodium acetate, pH 6). After vortexing, the nucleic acid wasextracted with phenol–chloroform and then precipitated with an equalvolume of isopropyl alcohol. The pellet was dried under vacuum. For RT,10 U RNasin (HT Biotechnology) and 30 pmol RT primer were added tothe pellet together with 10±5 µl water. The mixture was warmed to 70 °Cfor 5 min. One µl 5 mM deoxynucleoside triphosphate (dNTP), 10 URNasin, 20 µg BSA and 1 µl Moloney murine leukaemia virus reversetranscriptase (20 U}µl) (Gibco BRL) were then added together with theprovided reaction buffer to a final volume of 20 µl. The mixture wasbrought to room temperature for 15 min, and then the reaction wascarried out at 37 °C for 90 min. After RT, PCR was done in two stepswith two sets of primers. The reaction was carried out with 10 µl cDNAmixture, 200 µM dNTP, 50 pmol of each primer, 2 U Super Tag (HTBiotechnology) and the provided PCR reaction buffer in a final volume of100 µl. A DNA thermal cycler (Perkin-Elmer Cetus) was used with a stepcycle programme of 94 °C for 30 s, 58 °C for 30 s and 72 °C for 60 s for25 cycles. For secondary PCR, 5 µl from the first PCR product was usedas the source of DNA template. The primers used for RT and nested PCRwere deduced from the published Taiwanese HCV sequence and areavailable from the authors on request. The RT primer was also used as thefirst step anti-sense PCR primer. After gel-purification by GeneClean(Bio101), the DNA fragments were cloned into pT7Blue T-Vector(Novagen). For each DNA fragment, 6–14 clones were obtained andsubmitted to dideoxy sequencing (Sequenase version 2.0 DNA sequenc-ing kit, US Biochemical) for both plus and minus strands. The cloning andsequencing procedures were described previously (Yeh et al., 1994a).
The cDNA for the extreme 5« end of the HCV genome was amplifiedaccording to the ‘one-sided ’ PCR method (Okamoto et al., 1992a). Thesingle-stranded cDNA corresponding to the 5« terminus was first tailedwith dATP homopolymer by a terminal deoxynucleotidyl transferase(Boehringer Mannheim). The tailed cDNA was then amplified by PCR
using primer A (5« AAGGATCCGTCGACATCGATTTTTTTTTTT-TTTTTTTTTT 3«) and primer ®238RT (5« GGGGTCCTGGAGG-CTGCACG 3«). Secondary PCR was performed with primer A« (5«AAGGATCCGTCGACATCGAT 3«) and primer ®259 (5« CACTC-ATACTAACGCCATGG 3«). To obtain clone CG14, a primer, poly(A)
"&
was used as RT and the first and second step anti-sense PCR primer. ThePCR sense primers for the first and second steps were 8958 (5«CCGACCCCGCTGGTTTATGT 3«) and 8978 (5« TGTGCCTACTCC-TACTTTCT 3«), respectively.
+ Construction of DNA plasmids encoding truncated and full-length core proteins. To connect the two clones CG1 and C-fs (Fig.1), 20 ng gel-purified DNA fragments from the two correspondingRT–PCR reactions were mixed together in 200 µM dNTP, 2 U Super Tagand the provided PCR reaction buffer in a final volume of 100 µl. Afterthe reaction had been performed in a DNA thermal cycler with a stepcycle programme of 94 °C for 30 s, 60 °C for 60 s and 72 °C for 60 s for10 cycles, 50 pmol of primer 642 (5« TGATCATGTCCGCTGCCTCA3«) and primer B (5« CACCATGAGCACAAATCCTAAACC 3«) wereadded and the reaction was performed for a further 25 cycles. Theresulting product was cloned into pT7Blue T-Vector. After verificationby dideoxy sequencing, the fragment containing the core reading framewas isolated by restriction enzyme digestion (HindIII–SmaI, from thepolylinker sites of pT7Blue T-Vector) and inserted into HindIII–XbaI(blunted) polylinker sites of the vector pRc}CMV (Invitrogen). Clonesderived from both the serum and ascites samples were connected andinserted into pRc}CMV to generate pCMV-SC and pCMV-AC,respectively. A plasmid, pCMV-RCEβ (a gift from J. Ou, USC, LosAngeles, CA), containing HCV-RH isolate from nucleotide 1 to 1292 (aa1–430) (Lo et al., 1995), was also included in this study for comparison.
+ DNA transfection and immunofluorescence analysis. Huh-7cells were maintained in a 1 :1 mixture of Dulbecco’s modified essentialmedium and F12 medium supplemented with 10% FBS. DNA plasmidswere transfected by using the standard CaPO
%precipitation method
(Graham & van der Ed, 1973). Cells were glycerol-shocked for 2 min at4–6 h after transfection. For immunofluorescence analysis, cells weregrown on cover slips (Yeh et al., 1990). At 48 h post-transfection, the cellswere fixed in acetone at ®20 °C for 2 min. Mouse monoclonal antibodyto HCV core fragment (aa 1–120 ; Biogenesis) (1 :500 dilution) and FITC-conjugated goat anti-mouse antibody (Jackson ImmunoResearch Labora-tories) (1 :20 dilution) were used as the primary and secondary antibodies,respectively.
+ Specific detection of serum- or ascites-derived clones. Tospecifically detect either the serum- or ascites-derived clones in samples,two sets of primers were designed according to the hypervariableregions : specific primers for serum-derived clones, 5« AGTGATGGC-CGTGCGCCTACGCTTG 3« [sense ; nt 1171 to 1198 (positionscorresponding to the published Taiwanese strain) ; see also Fig. 3] and 5«AAGACAGCAGGCACACCATGA 3« (anti-sense ; nt 1438 to 1418) ;specific primers for ascites-derived clones, 5« GCAGCAGGCATAC-CACCCGCGGCTTCA 3« (sense ; nt 1171 to 1198) and 5« GGGCCTC-CATGCTCAGCATAG 3« (anti-sense ; nt 1438 to 1418). These two setsof primers had the same sequences as those of the serum and ascites-derived clones but had 20 and 10 nucleotide mismatches between eachother for the sense and anti-sense primers, respectively. These two setsof primers were used in the second step PCR for specific detection, whilethe primers for RT and first step PCR were the same as those for cloningHVR-fs (available from the authors on request). The step cycleprogramme of the first step PCR was modified to 94 °C for 30 s, 58 °Cfor 30 s and 72 °C for 60 s for 22 cycles, and the second step PCR to94 °C for 30 s, 62 °C for 30 s and 72 °C for 30 s for 20 cycles.
CHGC

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
Defective HCV genome in ascitesDefective HCV genome in ascites
Fig. 1. Strategy for PCR amplification of an ascitic HCV genome. Primers used for RT and nested PCR are available from theauthors on request. In total, 19 clones were obtained: the 5« end, C-fs, HVR-fs, NS2-fs, NS3-fs and CG1–CG14. For the serumsample, only nine clones were obtained: C-fs, HVR-fs, NS2-fs, NS3-fs, CG1 and CG10–CG13.
To determine the sensitivity of this PCR experiment, HVR-fs clonesfrom both ascitic fluid and serum samples were used as templates for in
vitro transcription using MEGAscript T7 Kit (Ambion) to producefragments of RNA. These RNA fragments were diluted serially to obtainconcentrations of 10"!–10" copies}ml.
To confirm its specificity, the PCR product was further analysed bydirect sequencing. The gel-purified DNA fragment was analysed usingSequiTherm Cycle Sequencing kits (Epicentre Technologies). Theprocedure was performed according to the manual provided by themanufacturer and [α-$&S]dATP was used for radiolabelling. The primerused for direct sequencing was from either one of the second step PCRprimers.
+ Minus-strand RT–PCR. The experiment was performed accordingto a previously described procedure with minor modifications (Yeh et al.,1996). Briefly, ascitic fluid was centrifuged at 3000 g for 10 min. Thepellet (containing mononuclear cells) was washed once with TBS (10 mMTris–HCl pH 7±2, 150 mM NaCl) and dispersed in 250 µl water beforebeing submitted to RT–PCR. The outer primers used in minus-strandRT–PCR were 5« CCCTGTGAGGAACTACTGTC 3« (sense ; nt ®299to ®282) and 5« CGGCAACAGGTAAACTCCAC 3« (anti-sense ; nt114 to 95). The inner primers were 5« TTCACGCAGAAAGCGTCTAG3« (sense ; nt ®279 to ®260) and 5« ACGATCTGACCACCGCCCGG3« (anti-sense ; nt 92 to 73). The sense, outer primer was used for RT, afterwhich 5 U RNase A was added. The mixture was incubated at 37 °C for30 min before further PCR reaction.
ResultsCloning of a defective ascitic HCV genome up to thepoly(U) stretch by RT–PCR
We cloned the HCV genome from the ascitic fluid of apatient with hepatocellular carcinoma (patient-HCC) by theprocedure of RT followed by nested PCR (Fig. 1). RT–PCRusing the primers deduced from the published Taiwanese clonewas quite efficient. Fragments as long as 1±2 kb (CG10) wereamplified but attempts to produce longer fragments failed. Tento sixteen clones were sequenced for each fragment. For eachset of clones derived from fragments CG4, NS2-fs, CG7,CG10, CG11 and CG12, one to two clones were different fromthe others in the same set and contained substitutions of up tofour nucleotides. For HVR-fs, the region with the most diversesequences, sequence analysis revealed five different kinds ofclones with eight, three, one, one and one clones (of the 16analysed) of each kind. However, there were only zero to sixnucleotide differences among them. For the remaining frag-
ments, all clones derived from the same fragment were identicaland no sequence variation was observed.
The HCV genome contained 9400 nucleotides plus apoly(U) stretch (Fig. 2). When compared with the previouslypublished Taiwanese strain (Chen et al., 1992), severalnucleotide deletions were identified. The 5« noncoding regionwas very conserved with only two nucleotide substitutions (nt®222 and ®1) and one single nucleotide deletion (between nt®171 and ®170). Strikingly, in the coding region, there werefive sites of deletions which were capable of disrupting thereading frame (marked in Fig. 2 ; see also Fig. 3) : a singlenucleotide deletion in C-fs, HVR-fs and NS3-fs, deletion of anucleotide duplet in NS2-fs and deletion of a nucleotidequadruplet in NS3-fs. These frameshift mutations were ob-served in all of the 10 C-fs, 16 HVR-fs, 12 NS2-fs and 11 NS3-fs sequenced clones. Unusual mutations were also found inclone NS3-fs, in which five nucleotides (nt 3401 to 3405 in theTaiwanese strain) were replaced by a short stretch of sequencecontaining 13 nucleotides (Fig. 3).
If these frameshift mutations were ignored, there were stillsix sites at which mutations led to amino acid deletions (Table1). At four of them (NS3-fs, CG1, CG11 and CG13), nucleotidetriplets were deleted, whereas at the other two (CG10 andCG12), separate but nearby nucleotide deletions resulted inboth amino acid deletion and missense mutations.
Cloning of HCV RNA fragments from a serum sampleof the same patient
Using the same cloning and sequencing strategy, weselectively cloned the corresponding fragments containingimportant deletions from the serum sample of patient-HCC. Intotal, seven, seven, eleven, six, eight, seven, seven, eight andsix clones were analysed for C-fs, CG1, HVR-fs, NS2-fs, NS3-fs, CG10, CG11, CG12 and CG13, respectively. Except forHVR-fs, only one to five nucleotide substitutions were foundamong clones derived from the same fragment. Surprisingly,none of the frameshift mutations detected in the ascitic cloneswere found in the serum-derived clones (Fig. 3). In the 5« non-coding region, the sequence was identical to that of the asciticclone except for the absence of the deletion between nt ®171and ®170 of the ascitic clone. In C-fs, all deletions, includingthe frameshift mutation, were absent in serum-derived clones.In HVR-fs, which contained the hypervariable region, 10
CHGD

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
C.-T. Yeh and othersC.-T. Yeh and others
Fig. 2. For legend see facing page.
CHGE

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
Defective HCV genome in ascitesDefective HCV genome in ascites
Fig. 2. Nucleotide and deduced amino acid sequences of the defective ascitic HCV genome. Nucleotides are numbered fromthe initiation codon of the truncated core reading frame. The start codons of the putative core to NS5 regions are marked by ‘ r ’followed by an arrow. Nucleotide mutations which were not present in the reported Taiwanese strain (Chen et al., 1992) aremarked as follows: ^, single nucleotide deletion ; #, deletion of nucleotide duplet ;j, deletion of nucleotide triplet ;U, deletionof nucleotide quadruplet ; … , replacement of oligonucleotides.
different sequences were observed in the serum-derived cloneswith up to 16 nucleotide substitutions among clones. However,only two kinds of predicted amino acid sequences were presentand most of the nucleotide substitutions did not result inheterogeneity at the amino acid level. Two single amino acid
deletions (nt 1183 to 1185 and 1220 to 1222) were observedfor all clones but no frameshift mutation was found. Only themost prevalent clone for HVR-fs is shown in Fig. 3. For NS2-fs and NS3-fs, both deletions leading to frameshift mutationsand the unusual mutations at nt 3401 to 3405 (replacement of
CHGF

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
C.-T. Yeh and othersC.-T. Yeh and others
Fig. 3. Comparison of the nucleotide sequences of Taiwanese, ascities- and serum-derived clones. Only regions containingframeshift mutations in the ascitic and serum clones are shown. Nucleotides are numbered according to the Taiwanese strain(without parentheses) and the defective ascitic genome as in Fig. 2 (in parentheses). –, Same nucleotide as in Taiwanesestrain ; /, nucleotide deletion ; ¯¯, regions used for designing specific primer-sets to distinguish ascitic from serum-derivedclones. The region where replacement of oligonucleotides occurred (in NS3-fs) are marked by dots and the noveloligonucleotide is underlined.
oligonucleotides) were absent. No mutation leading to aminoacid deletions was found. For NS5 regions (CG10, CG11,CG12 and CG13), none of the four amino acid deletions (Table1) was present in the serum-derived clones. Instead, thepredicted amino acid residues in these four sites were the sameas those in the Taiwanese strain.
The defective HCV genome encodes a truncatednuclear core protein
The frameshift mutation in C-fs resulted in early terminationof translation of the genome such that the only protein productwould be a truncated core protein of 90 amino acid residues(Fig. 2). The last 20 amino acid residues came from a different
reading frame and were not related to the original HCV coreprotein. This protein contained all three putative nuclearlocalization signals (Ravaggi et al., 1994). When we expressedit in Huh-7 cells, the protein was indeed localized to thenucleus (Fig. 4). In contrast, the serum-derived intact coreprotein was localized to the cytoplasm.
Specific detection of ascites- or serum-derived clonesin ascitic fluid, serum and ascitic mononuclear cells
To understand whether HCV replicated in the asciticmononuclear cells of this patient, minus-strand-specific RT–PCR was performed. Minus strand HCV RNA was onlydetected when ascitic mononuclear cells were collected from
CHGG

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
Defective HCV genome in ascitesDefective HCV genome in ascites
Table 1. Summary of amino acid ‘deletions’ in the ascitic HCV clone
Since there were frameshift mutations, the reading frame was in fact interrupted before these amino acid residues could be translated. Therefore,these ‘deletions ’ exist only when the frameshift mutations are ignored.
Nucleotide Amino acid Sequence changeRegion (clone) no.* no.* TW strainUAS clone†
Core (CG1) 433–435 145 GGG (G)! del.NS3 (NS3-fs) 3721–3723 1241 CCG (P)! del.NS5 (CG10) 6802–6825 2268–2275 GTATCCGTTCCGGCGGAGATCCTG
(6795–6815) (VSVPAEIL)!GTGTCGTCCGGCGGAGATCTG
(VSSGGDL)NS5 (CG11) 7903–7908 2635–2636 GGTTTC (GF)!GTC (V)
(7893–7895)NS5 (CG12) 8170–8184 2724–2728 GCTGCGAAGCTCCAG (AAKLQ)!
(8157–8168) GCTGGAAGCCAG (AGSQ)NS5 (CG13) 8821–8823 2942 GGC (G)! del.
* Numbered according to the Taiwanese HCV strain. The nucleotide numbers in the ascitic clone (as in Fig. 2) are also given in parentheses forclones CG10, CG11 and CG12.† Predicted amino acid residues are shown in parentheses after the nucleotide sequence and are underlined. TW strain, Taiwanese HCV strain ; ASclone, defective ascitic genome ; del., deleted.
(a) (b)
pCMV-RCEb
pCMV-AC
pCMV-SC
Core
E1
E2/NS1
Non-HCV sequence
pCMV-RCEb
pCMV-AC
pCMV-SC
Fig. 4. Immunofluorescence analysis of serum-derived HCV core protein and ascites-derived truncated HCV core protein. (a)Proteins expressed by pCMV-RCEβ, pCMV-AC and pCMV-SC. Although E1 and/or E2/NS1 were encoded by pCMV-RCEβ andpCMV-SC, they were expected to be cleaved away by the host signal peptidase after expression. (b) Immunofluorescenceanalysis of HCV core proteins expressed by pCMV-RCEβ, pCMV-AC and pCMV-SC.
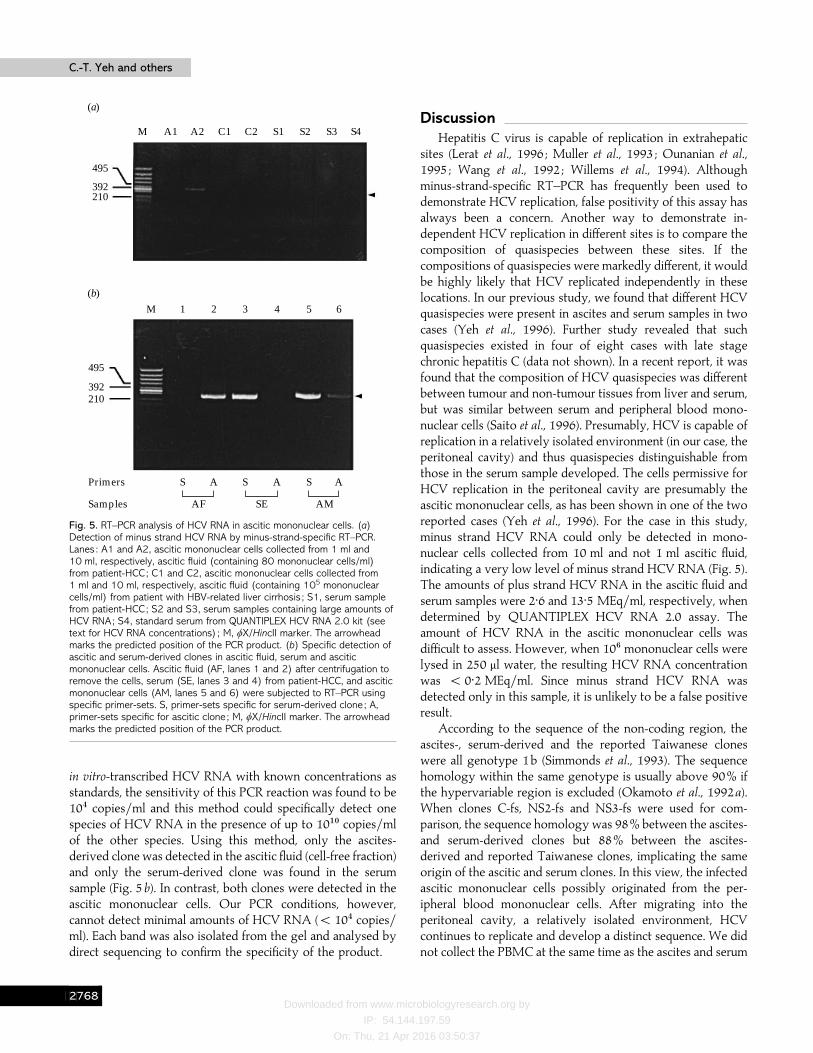
10 ml ascitic fluid but was not detectable when only 1 mlascitic fluid was used (Fig. 5a, lanes A1 and A2). Ascites frompatients with hepatitis B-related liver cirrhosis with similaramounts of mononuclear cells, and serum samples with highHCV RNA levels were included as controls (Fig. 5a, lanes C1,C2 and S1–4). The HCV RNA concentrations for S1, S2, S3and S4, determined by QUANTIPLEX HCV RNA 2.0 assay,were 13±5, 120, 200 and 30 MEq}ml, respectively. The HCV
RNA concentration in cell-free ascitic fluid was 2±6 MEq}ml,whereas for 10' mononuclear cells (collected from 10 ml asciticfluid) dissolved in 250 µl water it was ! 0±2 MEq}ml. Thiswas the mixture submitted to minus-strand-specific RT–PCRfor sample A2.
Specific primer-sets were designed for second step PCR tospecifically detect ascites- or serum-derived clones in thesamples using more stringent conditions (see Methods). Using
CHGH
Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
C.-T. Yeh and othersC.-T. Yeh and others
M A1 A2 C1 C2 S1 S2 S3 S4
495
392210
495
392210
M 1 2 3 4 5 6
S A S A S A
AF SE AM
Primers
Samples
(a)
(b)
Fig. 5. RT–PCR analysis of HCV RNA in ascitic mononuclear cells. (a)Detection of minus strand HCV RNA by minus-strand-specific RT–PCR.Lanes : A1 and A2, ascitic mononuclear cells collected from 1 ml and10 ml, respectively, ascitic fluid (containing 80 mononuclear cells/ml)from patient-HCC; C1 and C2, ascitic mononuclear cells collected from1 ml and 10 ml, respectively, ascitic fluid (containing 105 mononuclearcells/ml) from patient with HBV-related liver cirrhosis ; S1, serum samplefrom patient-HCC; S2 and S3, serum samples containing large amounts ofHCV RNA; S4, standard serum from QUANTIPLEX HCV RNA 2.0 kit (seetext for HCV RNA concentrations) ; M, φX/HincII marker. The arrowheadmarks the predicted position of the PCR product. (b) Specific detection ofascitic and serum-derived clones in ascitic fluid, serum and asciticmononuclear cells. Ascitic fluid (AF, lanes 1 and 2) after centrifugation toremove the cells, serum (SE, lanes 3 and 4) from patient-HCC, and asciticmononuclear cells (AM, lanes 5 and 6) were subjected to RT–PCR usingspecific primer-sets. S, primer-sets specific for serum-derived clone ; A,primer-sets specific for ascitic clone ; M, φX/HincII marker. The arrowheadmarks the predicted position of the PCR product.
in vitro-transcribed HCV RNA with known concentrations asstandards, the sensitivity of this PCR reaction was found to be10% copies}ml and this method could specifically detect onespecies of HCV RNA in the presence of up to 10"! copies}mlof the other species. Using this method, only the ascites-derived clone was detected in the ascitic fluid (cell-free fraction)and only the serum-derived clone was found in the serumsample (Fig. 5b). In contrast, both clones were detected in theascitic mononuclear cells. Our PCR conditions, however,cannot detect minimal amounts of HCV RNA (! 10% copies}ml). Each band was also isolated from the gel and analysed bydirect sequencing to confirm the specificity of the product.
DiscussionHepatitis C virus is capable of replication in extrahepatic
sites (Lerat et al., 1996 ; Muller et al., 1993 ; Ounanian et al.,1995 ; Wang et al., 1992 ; Willems et al., 1994). Althoughminus-strand-specific RT–PCR has frequently been used todemonstrate HCV replication, false positivity of this assay hasalways been a concern. Another way to demonstrate in-dependent HCV replication in different sites is to compare thecomposition of quasispecies between these sites. If thecompositions of quasispecies were markedly different, it wouldbe highly likely that HCV replicated independently in theselocations. In our previous study, we found that different HCVquasispecies were present in ascites and serum samples in twocases (Yeh et al., 1996). Further study revealed that suchquasispecies existed in four of eight cases with late stagechronic hepatitis C (data not shown). In a recent report, it wasfound that the composition of HCV quasispecies was differentbetween tumour and non-tumour tissues from liver and serum,but was similar between serum and peripheral blood mono-nuclear cells (Saito et al., 1996). Presumably, HCV is capable ofreplication in a relatively isolated environment (in our case, theperitoneal cavity) and thus quasispecies distinguishable fromthose in the serum sample developed. The cells permissive forHCV replication in the peritoneal cavity are presumably theascitic mononuclear cells, as has been shown in one of the tworeported cases (Yeh et al., 1996). For the case in this study,minus strand HCV RNA could only be detected in mono-nuclear cells collected from 10 ml and not 1 ml ascitic fluid,indicating a very low level of minus strand HCV RNA (Fig. 5).The amounts of plus strand HCV RNA in the ascitic fluid andserum samples were 2±6 and 13±5 MEq}ml, respectively, whendetermined by QUANTIPLEX HCV RNA 2.0 assay. Theamount of HCV RNA in the ascitic mononuclear cells wasdifficult to assess. However, when 10' mononuclear cells werelysed in 250 µl water, the resulting HCV RNA concentrationwas ! 0±2 MEq}ml. Since minus strand HCV RNA wasdetected only in this sample, it is unlikely to be a false positiveresult.
According to the sequence of the non-coding region, theascites-, serum-derived and the reported Taiwanese cloneswere all genotype 1b (Simmonds et al., 1993). The sequencehomology within the same genotype is usually above 90% ifthe hypervariable region is excluded (Okamoto et al., 1992a).When clones C-fs, NS2-fs and NS3-fs were used for com-parison, the sequence homology was 98% between the ascites-and serum-derived clones but 88% between the ascites-derived and reported Taiwanese clones, implicating the sameorigin of the ascitic and serum clones. In this view, the infectedascitic mononuclear cells possibly originated from the per-ipheral blood mononuclear cells. After migrating into theperitoneal cavity, a relatively isolated environment, HCVcontinues to replicate and develop a distinct sequence. We didnot collect the PBMC at the same time as the ascites and serum
CHGI

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
Defective HCV genome in ascitesDefective HCV genome in ascites
samples were obtained. However, PBMC collected 3 monthslater were available and only serum-derived clones weredetected by the specific primer-sets method. This result wasconsistent with the previous report (Saito et al., 1996).However, when ascitic mononuclear cells were examined, bothserum- and ascites-derived clones were present (" 10%
copies}ml). Judging from the relative densities of PCRproducts, the ascites-derived clone was the minor species andthe serum-derived clone was the predominant one (Fig. 5b).The ascitic clone possibly developed after migration ofmononuclear cells to the peritoneal cavity.
A striking finding was that there are deletions leading toframeshifts in the ascitic clone and the open reading frame isterminated shortly after the first frameshift mutation. As aresult, a truncated core protein is produced (90 aa). Theseframeshift mutations appeared in every clone from the samePCR fragments, indicating that they were the major species ofthe virus population. A major concern was whether thesemutations resulted from PCR, cloning or sequencing artifacts.Several factors argued against this. Firstly, all sequence datawere obtained from sequencing of both strands. Secondly,many deletions were in triplets and represented single aminoacid deletions on the same putative polyprotein reading frame(except for the one in CG11). Finally, using the same cloningand sequencing strategy, these deletions were absent in theserum-derived clones. Therefore, this ascitic HCV genome notonly constituted the major virus population in the ascitic fluidbut was also defective. Since both ascites- and serum-derivedclones were present in the ascitic mononuclear cells but onlythe defective ascitic clone was present in the ascitic fluid (afterremoval of the cellular components), the serum-derived clonein the ascitic mononuclear cells probably served as a helpersequence which enabled the encapsidation and secretion of thedefective genome. Conversely, the maturation of helper virusitself was interfered with and the helper sequence failed to exitthe mononuclear cells. At present, the molecular mechanismfor selective secretion of a defective genome is not clear. It ispossible that the area responsible for encapsidation in thedefective genome (encapsidation signal) is not occupied by thetranslation machinery and is more readily accessed by HCVcore protein. Alternatively, a clonal origin of cells maypreferentially allow replication of the defective genomebecause of the presence of certain cellular factors.
Although we have not analysed the virus sequences in thehepatocellular carcinoma tissue in this study, another grouphas performed such experiments (Ru$ ster et al., 1996) anddiscovered a mixed population of truncated core proteins inthe tumour tissue. Although nuclear localization of full-lengthcore protein is still unresolved, a truncated or processed coreprotein which contained the putative nuclear localizationsignals was localized to the nuclei (Ravaggi et al., 1994 ; Lo etal., 1995). It is likely that the carboxyl portion of HCV coreprotein, which contains a hydrophobic domain, is conforma-tionally interfering with the function of nuclear localization
signals. On the other hand, a recent study indicated that HCVcore protein cooperated with ras to transform primary ratembryo fibroblasts, but the molecular mechanism was notunderstood (Ray et al., 1996). We speculate that nucleartransport of HCV core protein plays a role in hepatocar-cinogenesis and a truncated core protein accumulates moreefficiently in the nuclei and facilitates its action.
In summary, we have identified a defective HCV genomewhich constituted the major virus population in the ascitic fluidfrom a patient with hepatocellular carcinoma. In contrast, anon-defective HCV genome existed in the patient’s serum andboth clones were present in the ascitic mononuclear cells. Thisfinding raised the possibility that the ascitic clone was adefective interfering HCV genome. With the aid of a helpersequence in the ascitic mononuclear cells, it could replicate andwas secreted into the ascitic fluid.
This work was supported by grants from the National Institute ofHealth, Taiwan (DOH85-HR-522) and from Chang Gung MedicalResearch Council (CMRP-483).
ReferencesAli, N. & Siddiqui, A. (1995). Interaction of polypyrimidine tract-bindingprotein with the 5« noncoding region of the hepatitis C virus RNAgenome and its functional requirement in internal initiation of translation.Journal of Virology 69, 6367–6375.
Bukh, J., Miller, R. H. & Purcell, R. H. (1995). Genetic heterogeneity ofhepatitis C virus : quasispecies and genotypes. Seminars in Liver Disease15, 41–63.
Chen, P.-J., Lin, M.-H., Tai, K.-F., Liu, P.-C., Lin, C.-J. & Chen, D.-S.(1992). The Taiwanese hepatitis C virus genome : sequence deter-mination and mapping the 5« termini of viral genomic and antigenomicRNA. Virology 188, 102–113.
Choo, Q.-L., Kuo, G., Weiner, A. J., Overby, L. R., Bradley, D. W. &Houghton, M. (1989). Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science 144, 359–362.
Choo, Q.-L., Richman, K. H., Han, J. H., Berger, K., Lee, C., Dong, C.,Gallegos, D., Coit, D., Medina-Selby, A., Barr, P. J., Weiner, A. J.,Bradley, D. W., Kuo, G. & Houghton, M. (1991). Genetic organizationand diversity of the hepatitis C virus. Proceedings of the National Academyof Sciences, USA 88, 2451–2455.
Graham, F. L. & van der Ed, A. J. (1973). A new technique for the assayof infectivity of human adenovirus 5 DNA. Virology 52, 456–467.
Han, J. H., Shyamada, V., Richman, K. H., Brauer, M. J., Irvine, B.,Urdea, M. S., Tekamp-Olson, P., Kuo, G., Choo, Q.-L. & Houghton, M.(1991). Characterization of the terminal regions of hepatitis C viralRNA: identification of conserved sequences in the 5« untranslated regionand poly(A) tails at the 3« end. Proceedings of the National Academy ofSciences, USA 88, 1711–1715.
Hayashi, N., Higashi, H., Kaminaka, K., Sugimoo, H., Esumi, M.,Komatsu, K., Hayashi, K., Sugitani, M., Suzuki, K., Tadao, O., Nozaki,C., Mizuno, K. & Shikata, T. (1993). Molecular cloning and het-erogeneity of the human hepatitis C virus (HCV) genome. Journal ofHepatology 17(Suppl. 3), S94–S107.
Houghton, M. (1996). Hepatitis C viruses. In Fields Virology, 3rd edn, pp.1035–1058. Edited by B. N. Fields, D. M. Knipe & P. M. Howley.Philadelphia : Lippincott–Raven.
CHGJ

Downloaded from www.microbiologyresearch.org by
IP: 54.144.197.59
On: Thu, 21 Apr 2016 03:50:37
C.-T. Yeh and othersC.-T. Yeh and others
Inchauspe, G., Zebedee, S., Lee, D.-H., Sugitani, M., Nasoff, M. &Prince, A. M. (1991). Genomic structure of the human prototype strainH of hepatitis C virus : comparison with American and Japanese isolates.Proceedings of the National Academy of Sciences, USA 88, 10292–10296.
Kato, N., Hijikata, M., Ootsuyama, Y., Nakagawa, M., Ohkoshi, S.,Sugimura, T. & Shimotohno, K. (1990). Molecular cloning of the humanhepatitis C virus genome from Japanese patients with non-A, non-Bhepatitis. Proceedings of the National Academy of Sciences, USA 87,9524–9528.
Kolykhalov, A. A., Feinstone, S. M. & Rice, C. M. (1996). Identificationof a highly conserved sequence element at the 3« terminus of hepatitis Cvirus genome RNA. Journal of Virology 70, 3363–3371.
Lerat, H., Berby, F., Trabaud, M. A., Vidalin, O., Major, M., Trepo, C. &Inchauspe, G. (1996). Specific detection of hepatitis C virus minusstrand RNA in hematopoietic cells. Journal of Clinical Investigation 97,845–851.
Lo, S.-Y., Masiarz, F., Hwang, S. B., Lai, M. M. C. & Ou, J. H. (1995).Differential subcellular localization of hepatitis C virus core geneproducts. Virology 213, 455–461.
Martell, M., Esteban, J. I., Quer, J., Genesca, J., Weiner, A., Esteban, R.,Guardia, J. & Gomez, J. (1992). Hepatitis C virus (HCV) circulates as apopulation of different but closely related genomes : quasispecies natureof HCV genome distribution. Journal of Virology 66, 3225–3229.
Muller, H. M., Pfaff, E., Goeser, T., Kallinwski, B., Solbach, C. &Thielmann, L. (1993). Peripheral blood leukocytes serve as a possibleextrahepatic site for hepatitis C virus replication. Journal of GeneralVirology 74, 669–676.
Ogata, N., Alter, H. J., Miller, R. H. & Purcell, R. H. (1991). Nucleotidesequence and mutation rate of the H strain of hepatitis C virus. Proceedingsof the National Academy of Sciences, USA 88, 3392–3396.
Okamoto, H., Okada, S., Sugiyama, Y., Kurai, K., Iizuka, H., Machida,A., Miyakawa, Y. & Mayumi, M. (1991). Nucleotide sequence of thegenomic RNA of hepatitis C virus isolated from a human carrier :comparison with reported isolates for conserved and divergent regions.Journal of General Virology 72, 2697–2704.
Okamoto, H., Kurai, K., Okada, S.-I., Yamamoto, K., Lizuka, H., Tanaka,T., Fukuda, S., Tsuda, F. & Mishiro, S. (1992a). Full-length sequence ofa hepatitis C virus genome having poor homology to reported isolates :comparative study of four distinct genotypes. Virology 188, 331–341.
Okamoto, H., Sugiyama, Y., Okada, S., Kurai, K., Akahane, Y., Sugai,Y., Tanaka, T., Sato, K., Tsuda, F., Miyakawa, Y. & Mayumi, M.(1992b). Typing hepatitis C virus by polymerase chain reaction withtype-specific primers : application to clinical surveys and tracinginfectious sources. Journal of General Virology 73, 673–679.
Ounanian, A., Gueddah, N., Rolachon, A., Thelu, M. A., Zarski, J. P. &Seigneurin, J. L. (1995). Hepatitis C virus RNA in plasma and bloodmononuclear cells in patients with chronic hepatitis C treated with alpha-interferon. Journal of Medical Virology 45, 141–145.
Ravaggi, A., Natoli, G., Primi, D., Albertini, A., Levrero, M. & Cariani,E. (1994). Intracellular localization of full-length and truncated hepatitisC virus core protein expressed in mammalian cells. Journal of Hepatology20, 833–836.
Ray, R. B., Lagging, L. M., Meyer, K. & Ray, R. (1996). Hepatitis C viruscore protein cooperates with ras and transforms primary rat embryofibroblasts to tumorigenic phenotype. Journal of Virology 70, 4438–4443.
Ru$ ster, B., Zeuzem, S. & Roth, W. K. (1996). Hepatitis C virussequences encoding truncated core proteins detected in a hepatocellularcarcinoma. Biochemical and Biophysical Research Communications 219,911–915.
Saito, S., Kato, N., Hijikata, M., Guji, T., Itabashi, M., Kondo, M.,Tanaka, K. & Shimothno, K. (1996). Comparison of hypervariableregions (HVR1 and HVR2) in positive- and negative-stranded hepatitis Cvirus RNA in cancerous and non-cancerous liver tissue, peripheral bloodmononuclear cells and serum from a patient with hepatocellularcarcinoma. International Journal of Cancer 67, 199–203.
Shimotohno, K., Tanji, Y., Hirowatari, Y., Komoda, Y., Kato, N. &Hijikata, M. (1995). Processing of the hepatitis C virus precursor protein.Journal of Hepatology 22, 87–92.
Simmonds, P., McOmish, F., Yap, P. L., Chan, S.-W., Lin, C. K.,Dusheiko, G., Saeed, A. A. & Holmes, E. C. (1993). Sequence variablilityin the 5« non-coding region of hepatitis C virus : identification of a newvirus type and restrictions on sequence diversity. Journal of GeneralVirology 74, 661–668.
Takamizawa, A., Mori, C., Fuke, I., Manabe, S., Murakami, S., Fujita, J.,Onishi, E., Andoh, T., Yoshida, I. & Okayama, H. (1991). Structure andorganization of the hepatitis C virus genome isolated from humancarriers. Journal of Virology 65, 1105–1113.
Tanaka, T., Kato, N., Cho, M.-J. & Shimotohno, K. (1995). A novelsequence found at the 3« terminus of the hepatitis C virus genome.Biochemical and Biophysical Research Communications 215, 744–749.
Wang, C. & Siddiqui, A. (1995). Structure and function of the hepatitisC virus internal ribosome entry site. Current Topics in Microbiology andImmunology 203, 99–115.
Wang, J. T., Sheu, J. C., Lin, J. T., Wang, T. H. & Chen, D. S. (1992).Detection of replicative form of hepatitis C virus RNA in peripheralblood mononuclear cells. Journal of Infectious Diseases 166, 1167–1169.
Willems, M., Peerlinck, K., Moshage, H., Deleu, I., Van den Eynde, C.,Vermylen, J. & Yap, S. H. (1994). Hepatitis C virus RNAs in plasma andin peripheral blood mononuclear cells of hemophiliacs with chronichepatitis C : evidence for viral replication in peripheral blood mononuclearcells. Journal of Medical Virology 42, 272–278.
Yeh, C.-T., Liaw, Y.-F. & Ou, J.-H. (1990). The arginine-rich domain ofhepatitis B virus precore and core proteins contains a signal for nucleartransport. Journal of Virology 64, 6141–6147.
Yeh, C.-T., Chiu, C.-T., Tsai, S.-L., Hong, S.-T., Chu, C.-M. & Liaw, Y.-F.(1994a). Absence of precore stop mutant in chronic dual (B and C) andtriple (B, C, and D) hepatitis virus infection. Journal of Infectious Diseases170, 1582–1585.
Yeh, C.-T., Han, C.-M., Lo, S.-Y., Ou, J.-H., Fan, K.-D., Sheen, I.-S., Chu,C.-M. & Liaw, Y.-F. (1994b). Early detection of anti-HCc antibody inacute hepatitis C virus (HCV) infection by western blot (immunoblot)using a recombinant HCV core protein fragment. Journal of ClinicalMicrobiology 32, 2235–2241.
Yeh, C.-T., Chu, C.-M. & Liaw, Y.-F. (1996). Distinct composition ofviral quasispecies between ascites and serum samples from patients withlate stage chronic hepatitis C. Biochemical and Biophysical ResearchCommunications 227, 524–529.
Received 2 June 1997; Accepted 17 July 1997
CHHA
Related Documents











