This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/authorsrights

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Molecular characterization and expression analysis of a GTP-bindingprotein (MiRab5) in Mangifera indica
Zhao-liang Liu a,1, Cong Luo a,1, Long Dong a, Can Van Toan a, Peng-xiao Wei a, Xin-hua He a,b,⁎a College of Agriculture, Guangxi University, Nanning 530004, PR Chinab Guangxi Crop Genetic Improvement and Biotechnology Laboratory, Nanning 530007, PR China
a b s t r a c ta r t i c l e i n f o
Article history:Received 4 November 2013Received in revised form 14 February 2014Accepted 17 February 2014Available online 18 February 2014
Keywords:Mangifera indicaMiRab5 geneFruit ripeningStressProkaryotic expression
The Rab family, the largest branch of Ras small GTPases, plays a crucial role in the vesicular transport in plants.The members of Rab family act as molecular switches that regulate the fusion of vesicles with target membranesthrough conformational changes. However, little is known about the Rab5 gene involved in fruit ripening andstress response. In this study, the MiRab5 gene was isolated from stress-induced Mangifera indica. The full-length cDNA sequence was 984 bp and contained an open reading frame of 600 bp, which encoded a 200amino acid protein with a molecular weight of 21.83 kDa and a theoretical isoelectric point of 6.99. The deducedamino acid sequence exhibited high homology with tomato (91% similarity) and contains all five characteristicRabmotifs. Real-timequantitative RT-PCR analysis demonstrated thatMiRab5wasubiquitously expressed in var-ious mango tree tissues at different levels. The expression ofMiRab5was up-regulated during later stages of fruitripening.Moreover,MiRab5was generally up-regulated in response to various abiotic stresses (cold, salinity, andPEG treatments). Recombinant MiRab5 protein was successfully expressed and purified. SDS-PAGE and westernblot analysis indicated that the expressed protein was recognized by the anti-6-His antibody. These results pro-vide insights into the role of the MiRab5 gene family in fruit ripening and stress responses in the mango plant.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The small GTP-binding proteins act as molecular switches that per-form regulatory functions in various biological processes including cellgrowth and differentiation, and organization of the cytoskeleton. TheRab protein is the largest branch of Ras small GTPases, which plays acrucial role in regulating vesicle transport and signal transductions(Dale et al., 2004). At least 60 distinct Rab proteins have been identifiedin eukaryotes (Segev, 2001). The genome of Arabidopsis thaliana con-tains 57 Rab members that can be grouped into eight clades (RabA–RabH), each with counterparts in yeast and animals (Rutherford andMoore, 2002; Vernoud et al., 2003). Among them, RabF (Rab5) is thelargest group with multifunctional proteins (Stenmark et al., 1994). In
A. thaliana, three identified homolog genes AtRabF1 (Ara6), AtRabF2a(Rha1), andAtRabF2b (Ara7), are similar to Rab5 andRab22 ofmammals(Rutherford and Moore, 2002; Vernoud et al., 2003). Ara7 and Rha1 areorthologs of Rab5 with conserved structure and high similarity. Ara6with neither a cysteine residue at the C-terminus nor a complete hyper-variable region is known as a plant-unique type contributing to theunique structural features (Hoepflinger et al., 2013; Kotzer et al.,2004; Sohn et al., 2003; Ueda et al., 2001).
Rab5 group is responsible for endosomal trafficking in yeast, animal,and plant cells (Schwartz et al., 2007). Ara6 and Ara7 are localized in theendosomes (Ueda et al., 2001). Trafficking pathway from endosomes tothe plasma membrane in A. thalianamediated by Ara6 is similar to thatin animals (Ebine et al., 2012). The rice OsRab5 participates in the bio-synthesis of carbon and nitrogen storage reserves in developing endo-sperm (Doroshenk et al., 2012; Morris et al., 2010). The tissue-specificexpression patterns of Rha1 in wild-type plants indicated that Rha1 isinvolved in auxin signal transduction (Qi et al., 2005).
In some cases, the different isoforms of same Rab protein were in-volved in dissimilar processes, while for three isoforms of Rab5 havebeen detected functionally equivalent (Bucci et al., 1995; Marshall,1995). Improved tolerance to stress has been detected in AtRab7 trans-genic plants and in prokaryotes (Mazel et al., 2004; Peng et al., 2011).But only a few is known about Rab5 in plant response to abiotic stress,for instance, the expression of Mcrab5b was induced by salt stress inMesembryanthemum crystallinum (Bolte et al., 2000).
Gene 540 (2014) 86–91
Abbreviations: SDS-PAGE, sodium dodecyl sulfate-polyacrylamide gel electrophoresis;PEG, polyethylene glycol; cDNA–SCoT, complementary deoxyribonucleic acid succinylCoA:3-oxoacid CoA transferase; RACE, rapid-amplification of cDNA ends; dCTP,deoxycytidine triphosphate; qRT-PCR, real-time quantitative-polymerase chain reaction;ORF, open reading frame; NJ, neighbor-joining; Kan, kanamycin; IPTG, isopropyl β-D-1-thiogalactopyranoside; CA, constitutive-active; DN, dominant-negative; DAB, chromo-meric substrate-3, 3′-diaminobenzidine.⁎ Corresponding author at: College of Agriculture, Guangxi University, Nanning 530004,
PR China.E-mail address: [email protected] (X. He).
1 The first two authors contributed equally to this paper.
http://dx.doi.org/10.1016/j.gene.2014.02.0220378-1119/© 2014 Elsevier B.V. All rights reserved.
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene

Author's personal copy
Mango (Mangifera indica L.) crop is widely cultivated in tropicalareasworldwide. However, its fruit quality and productivity are affectedby environmental stress such as cold, salinity, and drought. Cloning andexpression analysis of critical abiotic stress genes are a basis for geneticimprovement of mango crop. In previous research, we obtained a frag-ment of MiRab5 gene from cDNA–SCoT differential display of abioticstress. In this study, a modified 5′ RACE (rapid-amplification of cDNAends) was performed and the full-length cDNA sequence of MiRab5genewas isolated fromM. indica. We also discussed the expression pat-terns in different organs and under various abiotic stress treatments.
2. Materials and methods
2.1. Plant materials
Samples were collected from the orchard in Guangxi University,Nanning, Guangxi, in southeastern China. For transcriptional level of dif-ferent tissues and organ, samples were collected from young leaves(YL), old leaves (OL), young stems (YS), old stems (OS), flowers (FL),and fruits (F1, F3 and F6) 10, 30, 60 d after flowering from 11-year-oldM. indica L. cv. ‘SiJi’ grafted plants (on rootstock ‘Tu’ mango plants)trees. For transcriptional level of fruits at different stages, fruits (F1–F7) were sampled from 10, 20, 30, 40, 50, 60 and 70 d after flowering,respectively. Abiotic stress treatments were conducted on one-year-oldM. indica L. cv. ‘SiJi’ grafted plants grown in soil-filled pots outdoors.For low temperature stress, the plants were transferred to growthchambers adjusted to 4 °C (16 h light and 8 h dark photoperiod at alight intensity of 400 μmolm−1 s−1). Salinity treatment was conductedby watering each pot with 300 mM NaCl. Drought resistance wasevaluated by watering each pot with 300 g L−1 polyethylene glycol(PEG-6000). Plants that were not subjected to stress treatments servedas control under the same growth conditions. Different treated groupsamples were collected at 0, 24, 48, and 72 h after treatments, respec-tively. Each sample was immediately frozen in liquid nitrogen andstored at −80 °C in a refrigerator until analysis by real-time quantita-tive RT-PCR and other methods.
2.2. RNA extraction and cDNA synthesis
Total RNA from themango sampleswas isolated according to amod-ified hot borate method (Wan andWilkins, 1994). Integrity of the RNAwas checked by electrophoresis in a 1% agarose gel and ultraviolet spec-trophotometer (Eppendorf, Germany) analysis. High-quality total RNA(1 μg) from the samples was used to synthesize first-strand cDNAusing M-MLV virus reverse transcriptase (Takara, China) following themanufacturer's instructions with primer AUP1.
2.3. Cloning and sequence analysis
MiRab5 cDNA fragmentwas amplified by cDNA–SCoT analysis in ourprevious study and only lacked the 5′ end sequence. Amodified 5′ RACE(rapid-amplification of cDNA ends) was performed in this study to am-plify the 5′ end fragment (Luo et al., 2011). First-strand cDNAwas puri-fied, and homopolymer dC-tails were added to the 3′ ends of the cDNAby terminal deoxynucleotidyl transferase with dCTP (Takara, China).Two nested gene specific primers (MiRabd1 and MiRabd2), for MiRab5were designed based on the known sequence (Table 1). Two adaptorprimers, AP1 and AP2, were designed to conduct nested PCR with thegene-specific primers (Table 1). The first 5′ RACE was performedusing the tailed cDNA as a template with the primers MiRabd2 andAP1. Products of the first 5′ RACE were diluted 40 times and used astemplate for the second 5′ RACE amplification with the primersMiRabd1 and AP2. After two rounds of nested 5′ RACE amplification,the products of the two reactions were verified by agarose gel electro-phoresis, and the target fragments were selected by size based on thenested primers. Subsequently, the product was cloned and sequenced.
After obtaining the 5′ end fragments, the full-length MiRab5 sequencewas amplified by RT-PCR. The PCR was run in a T-Professionalthermocycler (Biometra, Germany) under the following conditions: ini-tial denaturation of 4min at 95 °C; then 36 cycles of 40 s at 95 °C, 40 s at56 °C, and 1 min at 72 °C; and a final extension of 10 min at 72 °C. Theresulting colonies were screened by PCR with the sequencing primersM13F and M13R to identify clones that contained inserts of the appro-priate size. The PCR reaction mixture (25 μL) contained 1× DreamTaqbuffer (Fermentas), 2.5 μM dNTPs, 10 μM of each primer, 1 μL of bacte-rial colony as template, and 0.625 U of DreamTaq. The PCR cycling con-ditions are as follows: After an initial denaturation of 95 °C for 4 min,there were 30 cycles of 94 °C for 30 s, 57 °C for 30 s, and 72 °C for2 min, followed by 72 °C for 8 min.
Identification of nucleotide sequences was established using theNCBI Blast program [http://www.ncbi.nlm.nih.gov/BLAST] (Altschulet al., 1990). The conserved domain was analyzed using InterProScan[http://www.ebi.ac.uk/Tools/pfa/iprscan] (Zdobnov and Apweiler,2001). The theoretical isoelectric point (pI) and molecular weight(Mw) for mature peptides were calculated using the PeptideMass pro-gram [http://us.expasy.org/tools/peptide-mass.html] (Wilkins et al.,1997). Multiple sequence alignments were carried out on Clustalx 2.0and GenDoc 2.7, and a phylogenetic tree was constructed using theneighbor-joining (NJ) method in the MEGA 5.2 (Tamura et al., 2007).
2.4. Real-time quantitative RT-PCR
The cDNA templates were synthesized using the total RNA isolatedfrom leaves and stems at 0, 24, 48, and 72 h after treatment with300 mM NaCl and 30% PEG-6000 at 4 °C. The synthesized cDNA wasdiluted 1:40 with water, and 2 μL of the diluted cDNA was used as atemplate for quantitative real-time PCR analysis. PCR reactions wereperformed in a total volume of 20 μL containing 10 μL SYBR PremixEx Taq, 0.4 μL of each primer (5 μM), 2 μL of cDNA, and 7.2 μL ofnuclease-free water. Real-time quantitative RT-PCR experiments wereperformed with SYBR Green PCR Master Mix (TaKaRa, China) on anABI 7500 real-time PCR machine (Applied Biosystems, America). Therelative gene expression was evaluated with the 2−ΔΔCT method(Livak and Schmittgen, 2001). Three biological replicates were usedfor the analysis. All data were expressed as mean ± standard error,and the transcript levels were normalized against the mango MiACT1gene, which was used as an internal control (Luo et al., 2013). Expres-sion level was calculated as the average ratio between the relative tran-script abundance of the treated and control samples. The qRT-PCRconditions were as follows: initial denaturation at 95 °C for 3 min and40 cycles of 10 s at 95 °C, 30 s at 57 °C, and 30 s at 72 °C.
Table 1PCR primers in this study.
Primer ID Sequence (5′–3′) Application
AUP1 GGCCACGCGTCGACTAGTACTTTTTTTTTTTTTTTTTT
Reverse transcriptase
AP1 AAGCAGTGGTATCAA CAGAGTACGCGGGGGGGGGG
Adaptor primers
AP2 AAGCAGTGGTATCAACGC AGAGT Adaptor primersMiRabd1 AGTTTGAGATTTGGGATACTGC 5′ RACEMiRabd2 TTTGAGCGGGCTAAGAAATGGG 5′ RACEqRab5F TTTGAGCGGGCTAAGAAATG Real time PCRqRab5R CTTGCATCCAGCAAATCAGA Real time PCRMiACT1F CCCAAGGCTAACAGAGAGAAGATG Real time PCRMiACT1R ATCACCAGAATCCAGCACA ATACC Real time PCRM13F CGCCAGGGTTTTCCCAGTCACGAC PMD-18 T universal primerM13R AGCGGATAACAATTTCACACAGGA PMD-18 T universal primerRab–BamH CGGGATCCATGGCTACCACCGGAAAC RecombinantRab–Sal GCGTCGACAGAGCAACAGGATGCAC Recombinant
87Z. Liu et al. / Gene 540 (2014) 86–91

Author's personal copy
2.5. Construction, expression, and purification of recombinant protein
Constructed recombinant MiRab5 protein as follows: the codingregion of MiRab5 was PCR-amplified, restriction-digested, and then li-gated with identical restriction-digested pET-30a vector to generatepET-Rab5 protein. The PCR primers used for the construction of sixhistidine-tagged recombinant proteins were Rab5–BamH and Rab5–Sal(Table 1). The in-frame fusion between the histidine tag and MiRab5was also verified by nucleotide sequencing. A single positive clone waspicked with a sterilized toothpick and transferred into 10 mL of LB/Kan(50 mg L−1) culture liquid. The culture was placed in a shaker at 37 °Cwith a constant speed of 200 rmin−1.When the OD600 value of the cul-ture reached 0.6, IPTGwas added at afinal concentration of 0.5mM.Afterthe addition of IPTG, the culture was divided into several aliquots andplaced at 28 °C in a shaker at 200 r min−1 continually for 4 h. Solublebody proteins were extracted from 1 mL of culture liquid, and the ex-pression levels were analyzed by SDS-PAGE. The recombinant proteinswere affinity-purified by Ni-NTA Agarose (Qiagen).
2.6. Western blot analysis
We evaluated the expression of the recombinant MiRab5 protein bySDS-PAGE and revealed that it was pET-Rab5 protein using westernblot. Protein sample (20 μg) was resolved by SDS-PAGE in a 15% poly-acrylamide gel and then blotted on polyvinylidene difluoride filtersusing a semi-dry blotting device (Hoefer TE77). The membrane wasincubated with anti-6× His antibody (origene) (1:1000; CwBiotech,China) and then with goat anti-mouse HRP-conjugated IgG secondaryantibody (1:1000; CwBiotech, China). The method used was accordingto one-step Western Kit HRP (Mouse) of Cwbiotech. The color was de-tected by DAB (chromomeric substrate-3, 3′-diaminobenzidine).
3. Results
3.1. MiRab5 gene sequence and structural analysis
In our previous study, MiRab5 cDNA fragment was obtained bycDNA–SCoT analysis. Its 5′ end sequencewas amplifiedwith a modified5′ RACE as described above. The full-length cDNA sequence was 984 bpand contained an open reading frame (ORF) of 600 bp,which encoded a200 amino acid proteinwith amolecularweight of 21.83 kDa and a the-oretical pI of 6.99.
The deduced amino acid sequence exhibited high homologywith to-mato Rab5 (Sl-RabF2a). The deduced polypeptide contains the struc-tures of typical GTP-binding sites: G1 domain (GDXGXGKX) and G3domain (DTAG) protein motifs are involved in the binding of phospho-ryl residues; the G4 domain (NKXD) and G5 domain (ETSA) are in-volved in the binding of guanine; and the G2 domain is known to bindother proteinous activators or inhibitors. Among the five characteristicRab motifs, G2, G3, and G5 are highly conserved (Fig. 1).
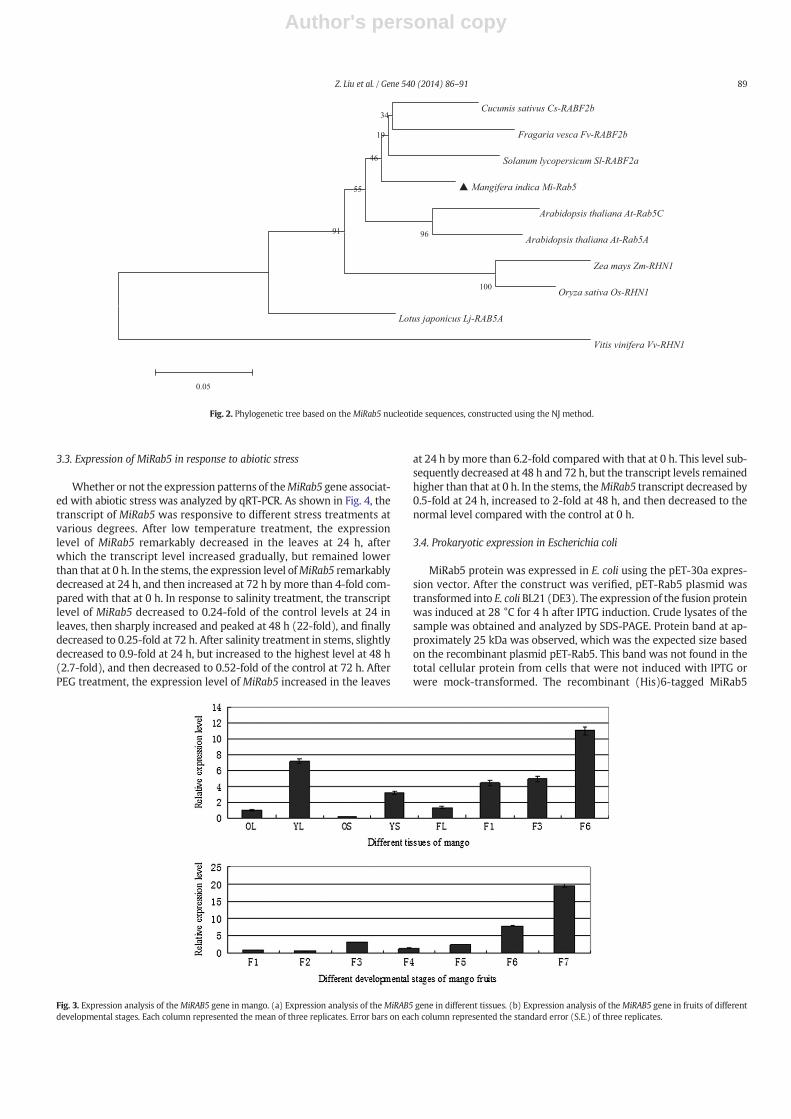
Phylogenetic tree based on MiRab5 protein nucleotide sequenceswas constructed using the NJ method (Fig. 2). We conclude that theisolated clone is a homolog of mango Rab5. The protein sequence ofMiRab5 was compared with selected Rab5 protein sequences availablefrom current sequence databases.
Blastp search of the GenBank database showed that the deducedprotein sequence of this cDNA was highly homologous to those ofknown Rab5 proteins with overall sequence identities of 91%, 90%, and90% to Solanum lycopersicum (Sl-RabF2a), Cucumis sativus (Cs-RabF2b),and Fragaria vesca (Fv-RabF2b) proteins, respectively. Therefore, wenamed this protein M. indica Rab5 (MiRab5, GenBank ID: KF768563).
3.2. Expression analysis of the MiRab5 gene in different mango tissues
Tissue-specific expression analysis of the MiRab5 gene was per-formed by quantitative real-time reverse transcription PCR (qRT-PCR).The results indicated that the expression pattern ofMiRab5was ubiqui-tous in all test samples with significant differences (Fig. 3). The tran-script level in young leaves (7.2-fold), young stems (3.2-fold), flower(1.4-fold), fruits F1 (4.5-fold), fruits F3 (5-fold) and fruits F6 (11-fold)was much higher than that in old leaves. Moreover, the transcriptlevel in fruits F1, F3 and F6 was distinctly increased.
To investigate whether MiRab5 was associated with fruit develop-ment or not, the transcript levels in fruits at different developmentalstages were examined. The results demonstrated that the transcriptlevel was decreased to 0.7-fold in fruits F2, then increased to 3.3-foldin fruits F3, but after dropping to 1.4-fold in fruits F4, then gradually in-creased to the highest level in fruits F7 (20-fold) much higher than thatin fruits F1. Taken together,MiRab5 was ubiquitously expressed in var-ious tissues with different expression levels. The level of MiRab5 tran-scription was much higher in young leaves than old leaves, so did inyoung stems and old stems. However, the expression level was up-regulated in the later stages during the fruit ripening.
Fig. 1. Amino acid sequence comparison of selected Rab5 subfamily proteins. Amino acid residues identical in all Rab5 proteins are shown on a black background, whereas residues con-served inmost members are shown on a gray background. The GDP/GTP binding domains (G1–G5) are indicated by dashed lines and are labeled. GenBank accession numbers: At-Rab5C(NP_193699.1), Cs-RabF2b (XP_004145451.1), Fv-RabF2b (XP_004293732.1), Sl-RabF2b (XP_004233205.1), Zm-RHN1 (ACF78199.1), At-Rab5A (NP_199326.1), Os-RHN1 (NP_001050846.1),Vv-RHN1 (XP_002284702.2), Mi-Rab5 (KF768563), Lj-Rab5A (CAA98166.1).
88 Z. Liu et al. / Gene 540 (2014) 86–91
Author's personal copy
3.3. Expression of MiRab5 in response to abiotic stress
Whether or not the expression patterns of theMiRab5 gene associat-ed with abiotic stress was analyzed by qRT-PCR. As shown in Fig. 4, thetranscript of MiRab5 was responsive to different stress treatments atvarious degrees. After low temperature treatment, the expressionlevel of MiRab5 remarkably decreased in the leaves at 24 h, afterwhich the transcript level increased gradually, but remained lowerthan that at 0 h. In the stems, the expression level ofMiRab5 remarkablydecreased at 24 h, and then increased at 72 h by more than 4-fold com-pared with that at 0 h. In response to salinity treatment, the transcriptlevel of MiRab5 decreased to 0.24-fold of the control levels at 24 inleaves, then sharply increased and peaked at 48 h (22-fold), and finallydecreased to 0.25-fold at 72 h. After salinity treatment in stems, slightlydecreased to 0.9-fold at 24 h, but increased to the highest level at 48 h(2.7-fold), and then decreased to 0.52-fold of the control at 72 h. AfterPEG treatment, the expression level of MiRab5 increased in the leaves
at 24 h bymore than 6.2-fold compared with that at 0 h. This level sub-sequently decreased at 48 h and 72 h, but the transcript levels remainedhigher than that at 0 h. In the stems, theMiRab5 transcript decreased by0.5-fold at 24 h, increased to 2-fold at 48 h, and then decreased to thenormal level compared with the control at 0 h.
3.4. Prokaryotic expression in Escherichia coli
MiRab5 protein was expressed in E. coli using the pET-30a expres-sion vector. After the construct was verified, pET-Rab5 plasmid wastransformed into E. coli BL21 (DE3). The expression of the fusion proteinwas induced at 28 °C for 4 h after IPTG induction. Crude lysates of thesample was obtained and analyzed by SDS-PAGE. Protein band at ap-proximately 25 kDa was observed, which was the expected size basedon the recombinant plasmid pET-Rab5. This band was not found in thetotal cellular protein from cells that were not induced with IPTG orwere mock-transformed. The recombinant (His)6-tagged MiRab5
Fig. 3. Expression analysis of theMiRAB5 gene in mango. (a) Expression analysis of theMiRAB5 gene in different tissues. (b) Expression analysis of theMiRAB5 gene in fruits of differentdevelopmental stages. Each column represented the mean of three replicates. Error bars on each column represented the standard error (S.E.) of three replicates.
Cucumis sativus Cs-RABF2b
Fragaria vesca Fv-RABF2b
Solanum lycopersicum Sl-RABF2a
Mangifera indica Mi-Rab5
Arabidopsis thaliana At-Rab5C
Arabidopsis thaliana At-Rab5A
Zea mays Zm-RHN1
Oryza sativa Os-RHN1
Lotus japonicus Lj-RAB5A
Vitis vinifera Vv-RHN1
100
9691
55
46
34
19
0.05
Fig. 2. Phylogenetic tree based on theMiRab5 nucleotide sequences, constructed using the NJ method.
89Z. Liu et al. / Gene 540 (2014) 86–91

Author's personal copy
protein was purified by a Ni-NTA column and eluted using imidazolebuffer. The supernatant protein of pET-Rab5 expressed in E. coli BL 21was compared with precipitate protein to examine the distribution ofthe expressed recombinant protein in soluble. The soluble supernatantwas collected and analyzed by SDS-PAGE. And the soluble protein waspurified by chromatography on nickel agarose column (Fig. 5a). West-ern blot analysis demonstrated that the recombinant pET-Rab5 proteinwas recognized by the antibody against the His6 tag and was indeed25 kDa (Fig. 5b). Therefore, recombinant MiRab5 was successfullyexpressed and purified.
4. Discussion
To understand the Rab5 gene of mango, the characterization andexpression ofMiRab5were analyzed. The deduced polypeptide containsall five characteristic Rabmotifs including the structures of typical GTP-binding sites: G1 domain (GDXGXGKX), G2 domain (TIGAAFF), G3domain (DTAG), G4 (NKXD), and G5 domain (ETSA). The C-terminal(XCCX) and the hypervariable regions are highly conserved. TheC-terminal region always ends in conserved motifs containing cys-teine residues, which is essential for membrane binding and function.
The hypervariable region determines the subcellular localization ofeach protein (Chavrier et al., 1991; Musha et al., 1992; Soldati et al.,1993; Ueda et al., 2000). The deduced amino acid sequence shares thehighest degree of similarity (91%)with S. lycopersicum and the samede-grees of similarity (90%) with both C. sativus and F. vesca.
The transcripts of MiRab5 displayed higher levels in young leavesthan old leaves, young stems than old stems, but ripened fruits muchhigher than unripe fruit. It may suggest thatMiRab5 is involved in mor-phogenetic processes such as organ formation and fruit ripening. Stud-ies demonstrated that a number of Rab proteins played an importantrole in different organ development processes of plants, such asmango, tomato, tobacco, grape and peach (Abbal et al., 2008; Falchiet al., 2010; Graaf et al., 2005; Lu et al., 2001; Zainal et al., 1996). But lit-tle was known about Rab5 involved in fruit ripening. The Rab5 geneswere localized in the plasma membrane and early endosomes, andubiquitously expressed in mammalian (Bucci et al., 1999). A Rab5 wasisolated from Cichorium leaf tissues by differential display to identifycDNA corresponding to genes that expressed during early somaticembryogenesis (Randoux et al., 2002).
The present data displayed that theMiRab5 gene was obviously up-regulated in mango induced by low temperature, salinity, and PEG.Therefore, MiRab5 might be involved in responding to cold, salinity,and dehydration in mango. The expression level of McRab5b inM. crystallinum increased under salt stress treatment (Bolte et al.,2000). ARA6 is required for the salt stress response, which mediateda trafficking pathway from the endosomes to the plasma membrane(Ebine et al., 2011). However, the mechanism of plant responding tovarious abiotic stresses is very complex because the stress-induceddamage repair processes involved in various proteins.
In prokaryotic expression,MiRab5 gene was cloned into prokaryoticexpression pET-30a vector and transformed into the host bacteria E. coliBL21 (DE3). Recombinant protein MiRab5 was successfully expressedand purified. SDS-PAGE and western blot analysis indicated that thespecific protein was expressed and recognized by the antibody againstthe His6 tag as predicted.
In summary, a Rab5-like novel geneMiRab5was isolated and charac-terized from mango. The MiRab5 protein had high identity to otherRab5 homologs and contained five conserved motifs of Rab. Moreover,the ubiquitously expressed MiRab5 was influenced by organ formation
Fig. 4. Expression analysis of the MiRAB5 gene under low temperature, NaCl, and PEGtreatments. Each column represented themean of three replicates. Error bars on each col-umn represented the standard error (S.E.) of three replicates.
Fig. 5. Purified proteins and western blot. (a). Lane M, protein molecular weight marker;Lane 1, pET-Rab5 (+). (b). Lane M, protein molecular weight marker; Lane 1, BL21;Lane 2, pet30a; Lane 3, pET-Rab5 (+). “(+)” meant that protein bands appeared byIPTG induction.
90 Z. Liu et al. / Gene 540 (2014) 86–91

Author's personal copy
and on the ripening ofmango fruit. In addition, the expression pattern ofMiRab5 was obviously up-regulated by chilling, salinity, and PEG stresstreatments. In addition, recombinant protein MiRab5 was successfullyexpressed in and purified from E. coli. SDS-PAGE and western blot anal-ysis indicated that the molecular weight of the expressed specific pro-teins was as predicted. Further experiments involving proteininteraction and transgenic A. thalianamust be conducted to further elu-cidate the role of MiRab5, especially in regulating fruit ripening andstress response.
Conflict of interest
The author(s) declare(s) that there is no conflict of interests regard-ing the publication of this article.
Acknowledgments
This researchwas supported byGuangxi Natural Science Foundation(2011GXNSFA018115) and (2013GXNSFDA019011), the ScientificResearch Foundation of Guangxi University (XBZ120766) and Innova-tion Project of Guangxi graduate Education (GXU11T32561).
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.gene.2014.02.022.
References
Abbal, P., et al., 2008. Molecular characterization and expression analysis of the RabGTPase family in Vitis vinifera reveal the specific expression of a VvRabA protein. Jour-nal of Experimental Botany 59, 2403–2416.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic local alignmentsearch tool. Journal of Molecular Biology 215, 403–410.
Bolte, S., Schiene, K., Dietz, K.J., 2000. Characterization of a small GTP-binding protein ofthe rab 5 family in Mesembryanthemum crystallinum with increased level of expres-sion during early salt stress. Plant Molecular Biology 42, 923–936.
Bucci, C., et al., 1995. Co-operative regulation of endocytosis by three rab5 isoforms. FEBSLetters 366, 65–71.
Bucci, C., Chiariello, M., Lattero, D., Maiorano, M., Bruni, C.B., 1999. Interaction cloning andcharacterization of the cDNA encoding the human prenylated rab acceptor (PRA1).Biochemical and Biophysical Research Communications 258, 657–662.
Chavrier, P., Grovel, J.P., Stelzer, E., Simons, K., Gruenberg, J., Zerial, M., 1991. Hypervari-able C-terminal domain of Rab proteins acts as a targeting signal. Nature 353,769–772.
Dale, L.B., Seachrist, J.L., Babwah, A.V., Ferguson, S.S., 2004. Regulation of angiotensin IItype 1A receptor intracellular retention, degradation, and recycling by Rab5, Rab7,and Rab11 GTPases. Journal of Biological Chemistry 279, 13110–13118.
Doroshenk, K.A., et al., 2012. Endosomal trafficking pathway regulated by ARA6, a RAB5GTPase unique to plants. Small GTPases 3, 23–27.
Ebine, K., et al., 2011. A membrane trafficking pathway regulated by the plant-specificRAB GTPase ARA6. Nature Cell Biology 13, 853–859.
Ebine, K., Miyakawa, N., Fujimoto, M., Uemura, T., Nakano, A., Ueda, T., 2012. Endosomaltrafficking pathway regulated by ARA6, a RAB5 GTPase unique to plants. SmallGTPases 3, 23–27.
Falchi, R., Cipriani, G., Marrazzo, T., Nonis, A., Vizzotto, G., Ruperti, B., 2010. Identifi-cation and differential expression dynamics of peach small GTPases encodinggenes during fruit development and ripening. Journal of Experimental Botany61, 2829–2842.
Graaf, B.H., Cheung, A.Y., Andreyeva, T., Levasseur, K., Kieliszewski, M., Wu, H., 2005. Rab11 GTPase-regulated membrane trafficking is crucial for tip-focused pollen tubegrowth in tobacco. Plant Cell 17, 2564–2579.
Hoepflinger, M.C., et al., 2013. Molecular and biochemical analysis of the first ARA6 homo-logue, a RAB5 GTPase, from green algae. Journal of Experimental Botany 64,5553–5568.
Kotzer, A.M., Brandizzi, F., Neumann, U., Paris, N., Moore, I., Hawes, C., 2004. AtRabF2b(Ara7) acts on the vacuolar trafficking pathway in tobacco leaf epidermal cells.Journal of Cell Science 117, 6377–6389.
Livak, K.J., Schmittgen, D.T., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 25, 402–408.
Lu, C., Zainal, Z., Tucker, G.A., Lycett, G.W., 2001. Developmental abnormalities andreduced fruit softening in tomato plants expressing an antisense Rab11 GTPasegene. Plant Cell 13, 1819–1833.
Luo, C., He, X.H., Chen, H., Wei, Y.L., Li, M.G., 2011. A high-efficient method of RACEtechnique for obtaining the gene 5′ end. Journal of Plant Physiology 47, 409–414.
Luo, C., He, X.H., Chen, H., Hu, Y., Ou, S.J., 2013. Molecular cloning and expression analysisof four actin genes (MiACT) from mango. Biologia Plantarum 57, 238–244.
Marshall, C., 1995. The Ras/Raf/ERK pathway. In: Zerial, M., Huber, L.A. (Eds.), Guidebookto the Small GTPases. Oxford University Press, New York, pp. 65–73.
Mazel, A., Leshem, Y., Tiwari, B.S., Levine, A., 2004. Induction of salt and osmotic stresstolerance by overexpression of an intracellular vesicle trafficking protein AtRab7(AtRabG3e). Journal of Plant Physiology 134, 118–128.
Morris, R.T., Wyrick, J.J., Fukuda, M., Kumamaru, T., Satoh, H., 2010. Characterization of therice glup4 mutant suggests a role for the small GTPase RAB5 in the biosynthesis ofcarbon and nitrogen storage reserves in developing endosperm. Breeding Science60, 556–567.
Musha, T., Kawata, M., Takai, Y., 1992. The geranylgeranyl moiety but not the methylmoiety of the smg25A/rab3A protein is essential for the interactions with membraneand its inhibitory GDP/GTP exchange protein. Journal of Biological Chemistry 267,9821–9825.
Peng, X.J., Zeng, X., Ding, X., Li, S.B., Yu, C., Zhu, Y.L., 2011. Ectopic expression of a vesicletrafficking gene, OsRab7, from Oryza sativa, confers tolerance to several abiotic stress-es in Escherichia coli. African Journal of Biotechnology 10, 6941–6946.
Qi, X.P., Zhou, J., Jia, Q.J., Shou, H.X., Chen, H.M., Wu, P., 2005. The expression of Rha1 andthe subcellular localization could be affected by auxin suggested that Rha1 involvedin auxin signal transduction. Plant Science 69, 1136–1145.
Randoux, B., Quillet, M.C., Rambaud, C., Vasseur, L., Hilbert, J.L., 2002. Characterisation ofcDNAs homologous to RAB5-GTP binding protein expressed during early somaticembryogenesis in chicory. Plant Science 162, 413–422.
Rutherford, S., Moore, I., 2002. The Arabidopsis Rab GTPase family-another enigma varia-tion. Curr. Opin. Plant Biol. 5, 518–528.
Schwartz, S.L., Cao, C., Pylypenko, O., Rak, A., Wandinger-Ness, A., 2007. Rab GTPases at aglance. Journal of Cell Science 120, 3905–3910.
Segev, N., 2001. Ypt and Rab GTPases: insight into functions through novel interactions.Current Opinion in Cell Biology 13, 500–511.
Sohn, E.J., et al., 2003. Rha1, an Arabidopsis RAB5 homolog, plays a critical role in thevacuolar trafficking of soluble cargo proteins. Plant Cell 15, 1057–1070.
Soldati, T., Riederer, M.A., Pfeffer, S.R., 1993. Rab GDI — a solubilizing and recycling factorfor rab9 protein. Molecular Biology of the Cell 4, 425–434.
Stenmark, H., Parton, R.G., Steele-Mortimer, O., Lutcke, A., Gruenberg, J., Zerial, M., 1994.Inhibition of RAB5 GTPase activity stimulates membrane fusion in endocytosis.EMBO Journal 13, 1287–1296.
Tamura, K., Dudley, J., Nei, M., Kumar, S., 2007. MEGA4: Molecular Evolutionary GeneticsAnalysis (MEGA) software version 4.0. Molecular Biology and Evolution 24,1596–1599.
Ueda, T., Matsuda, N., Uchimiya, H., Nakano, A., 2000. Modes of interaction between theArabidopsis Rab protein, Ara4 and its putative regulator molecules revealed by ayeast expression system. Plant Journal 21, 341–349.
Ueda, T., Yamaguchi, M., Uchimiya, H., Nakano, A., 2001. Ara6, a plant-unique novel typeRab GTPase, functions in the endocytic pathway of Arabidopsis thaliana. EMBO Journal20, 4730–4741.
Vernoud, V, Horton, A.C, Yang, Z.B, Nielsen, E, 2003. Analysis of the Small GTPase GeneSuperfamily of Arabidopsis. Plant Physiology. 131, 1191–1208.
Wan, C.Y., Wilkins, T.A., 1994. A modified hot borate method significantly enhances theyield of high−quality RNA from cotton (Gossypium hirsutum L.). Analytical bio-chemistry 223, 7–12.
Wilkins, M.R., et al., 1997. Detailed peptide characterization using PEPTIDEMASS — aWorld-Wide-Web-accessible tool. Electrophoresis 18, 403–408.
Zainal, Z., Tucker, G.A., Lycett, G.W., 1996. A rab11-like gene is developmentally regulatedin ripening mango (Mangifera indica L.) fruit. Biochimica et Biophysica Acta, Molecu-lar Cell Research 1314, 187–190.
Zdobnov, E.M., Apweiler, R., 2001. InterProScan — an integration platform for thesignature-recognition methods in InterPro. Bioinformatics 17, 847–848.
91Z. Liu et al. / Gene 540 (2014) 86–91
Related Documents











