Modulation of the effects of methylmercury on rat neurodevelopment by co-exposure with Labrador Tea (Rhododendron tomentosum ssp. subarcticum) Paleah Black a,b , Lijie Niu b , Manisha Sachdeva b , David Lean b , Raymond Poon a , Wayne J. Bowers a , Hing Man Chan c , John T. Arnason b , Guillaume Pelletier a,⇑ a Hazard Identification Division, Environmental Health Science and Research Bureau, Health Canada, Ottawa, ON, Canada b Department of Biology, University of Ottawa, Ottawa, ON, Canada c Community Health Sciences Program, University of Northern British Columbia, Prince George, BC, Canada article info Article history: Received 13 May 2011 Accepted 15 June 2011 Available online 21 June 2011 Keywords: Rat Neurodevelopment Behaviour Methylmercury Rhododendron tomentosum ssp. subarcticum Phytochemicals abstract Seafood is an important source of nutrients for many populations worldwide. Unfortunately the contam- ination of seafood with methylmercury (MeHg) has raised health concerns, particularly for developing infants. The modification of MeHg toxicity by nutrients, including antioxidants, has been reported in both epidemiological and animal exposure studies. We used a rat perinatal exposure model to investigate the modulation of developmental MeHg toxicity by an extract of Rhododendron tomentosum ssp. subarcticum, a plant rich in antioxidants traditionally consumed by Inuit. Sprague–Dawley rat dams were exposed daily to 2 mg MeHg/kg bw and 100 mg R. tomemtosum extract/kg bw administered either separately or jointly, from the first day of gestation until weaning. MeHg exposure was associated with impaired pup growth and survival, decreased grip strength, hyperactivity and altered exploratory behaviour, delayed acoustic startle response, elevated brain N-methyl-D-aspartate receptor (NMDA-R) levels, and increased serum lipid peroxidation. R. tomemtosum extract co-exposure modulated MeHg’s effects on oxi- dative stress and brain NMDA-R levels and may have affected MeHg pharmacokinetic. However, modu- lation of these molecular endpoints did not lead to significant alterations of MeHg’s effects on rat neurobehaviour. Crown Copyright Ó 2011 Published by Elsevier Ltd. All rights reserved. 1. Introduction There is a growing body of literature on the effects of dietary nutrients on MeHg toxicity. Most of these studies focussed on com- pounds commonly found in MeHg-contaminated food sources or which may potentially interfere with MeHg molecular mechanisms of toxicity (selenium, fatty acids, thiol-containing compounds and vitamins). Divergent findings were often reported, but globally these studies suggest that nutrients can modulate the adverse out- comes resulting from exposure to MeHg (Chapman and Chan, 2000; Rice, 2008). Among the best characterized mechanisms of MeHg neurotox- icity are excitotoxicity and oxidative stress (Yin et al., 2007; Aschner et al., 2007). Perturbation of glutamate transport in astro- cytes by MeHg can lead to over-stimulation and dysfunction of the N-methyl-D-aspartate receptors (NMDA-R) (Yin et al., 2007). The NMDA-R is part of the glutamatergic system which is crucial for neuronal plasticity, learning and memory (Scheetz and Constantine-Paton, 1994). During fetal development, NMDA-Rs are also involved in the establishment of neuronal circuitry (Haberny et al., 2002), which might constitute an additional target of MeHg toxicity. While NMDA-R mediated excitotoxicity can lead to the generation of reactive oxygen species (Gasso et al., 2001), MeHg can also generate oxidative stress through direct perturba- tion of mitochondrial functions (Aschner et al., 2007). The resultant oxidative stress can lead to cell membrane damage, calcium dereg- ulation, enzyme and cell signalling interference, microtubule disas- sembly, and ultimately, cell death (do Nascimento et al., 2008). MeHg co-exposure studies provide further evidence on the role of glutamate excitotoxicity and oxidative stress in MeHg-induced neurotoxicity. MeHg-induced neurotoxicity can be mitigated by co-exposure with NMDA-R antagonists (Zhang et al., 2003; Faro et al., 2002). Various purified phytochemicals or plant extracts have been shown to confer some protection against NMDA recep- tor agonist-induced excitotoxicity and oxidative stress in vivo and 0278-6915/$ - see front matter Crown Copyright Ó 2011 Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.fct.2011.06.035 Abbreviations: BROD, benzyloxyresorufin-O-dealkylase; BW, body weight; EROD, ethoxyresorufin-O-deethylase; GD, gestational day; GSH, reduced glutathi- one; MeHg, methylmercury; NMDA-R, N-methyl-D-aspartate receptor; PND, post- natal day; PPI, prepulse inhibition; PROD, pentoxyresorufin-O-dealkylase; TBARS, thiobarbituric acid reactive substances; THg, total mercury. ⇑ Corresponding author. Address: Environmental Health Centre, 50 Colombine Driveway, P.L. 0803B, Tunney’s Pasture, Ottawa, ON, Canada K1A 0L2. Tel.: +1 613 941 8063; fax: +1 613 948 9413. E-mail address: [email protected] (G. Pelletier). Food and Chemical Toxicology 49 (2011) 2336–2342 Contents lists available at ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Food and Chemical Toxicology 49 (2011) 2336–2342
Contents lists available at ScienceDirect
Food and Chemical Toxicology
journal homepage: www.elsevier .com/locate/ foodchemtox
Modulation of the effects of methylmercury on rat neurodevelopment byco-exposure with Labrador Tea (Rhododendron tomentosum ssp. subarcticum)
Paleah Black a,b, Lijie Niu b, Manisha Sachdeva b, David Lean b, Raymond Poon a, Wayne J. Bowers a,Hing Man Chan c, John T. Arnason b, Guillaume Pelletier a,⇑a Hazard Identification Division, Environmental Health Science and Research Bureau, Health Canada, Ottawa, ON, Canadab Department of Biology, University of Ottawa, Ottawa, ON, Canadac Community Health Sciences Program, University of Northern British Columbia, Prince George, BC, Canada
a r t i c l e i n f o
Article history:Received 13 May 2011Accepted 15 June 2011Available online 21 June 2011
Keywords:RatNeurodevelopmentBehaviourMethylmercuryRhododendron tomentosum ssp. subarcticumPhytochemicals
0278-6915/$ - see front matter Crown Copyright � 2doi:10.1016/j.fct.2011.06.035
Abbreviations: BROD, benzyloxyresorufin-O-deaEROD, ethoxyresorufin-O-deethylase; GD, gestationalone; MeHg, methylmercury; NMDA-R, N-methyl-D-asnatal day; PPI, prepulse inhibition; PROD, pentoxyrethiobarbituric acid reactive substances; THg, total me⇑ Corresponding author. Address: Environmental H
Driveway, P.L. 0803B, Tunney’s Pasture, Ottawa, ON,941 8063; fax: +1 613 948 9413.
E-mail address: [email protected] (G
a b s t r a c t
Seafood is an important source of nutrients for many populations worldwide. Unfortunately the contam-ination of seafood with methylmercury (MeHg) has raised health concerns, particularly for developinginfants. The modification of MeHg toxicity by nutrients, including antioxidants, has been reported in bothepidemiological and animal exposure studies. We used a rat perinatal exposure model to investigate themodulation of developmental MeHg toxicity by an extract of Rhododendron tomentosum ssp. subarcticum,a plant rich in antioxidants traditionally consumed by Inuit. Sprague–Dawley rat dams were exposeddaily to 2 mg MeHg/kg bw and 100 mg R. tomemtosum extract/kg bw administered either separately orjointly, from the first day of gestation until weaning. MeHg exposure was associated with impairedpup growth and survival, decreased grip strength, hyperactivity and altered exploratory behaviour,delayed acoustic startle response, elevated brain N-methyl-D-aspartate receptor (NMDA-R) levels, andincreased serum lipid peroxidation. R. tomemtosum extract co-exposure modulated MeHg’s effects on oxi-dative stress and brain NMDA-R levels and may have affected MeHg pharmacokinetic. However, modu-lation of these molecular endpoints did not lead to significant alterations of MeHg’s effects on ratneurobehaviour.
Crown Copyright � 2011 Published by Elsevier Ltd. All rights reserved.
1. Introduction
There is a growing body of literature on the effects of dietarynutrients on MeHg toxicity. Most of these studies focussed on com-pounds commonly found in MeHg-contaminated food sources orwhich may potentially interfere with MeHg molecular mechanismsof toxicity (selenium, fatty acids, thiol-containing compounds andvitamins). Divergent findings were often reported, but globallythese studies suggest that nutrients can modulate the adverse out-comes resulting from exposure to MeHg (Chapman and Chan,2000; Rice, 2008).
Among the best characterized mechanisms of MeHg neurotox-icity are excitotoxicity and oxidative stress (Yin et al., 2007;
011 Published by Elsevier Ltd. All
lkylase; BW, body weight;day; GSH, reduced glutathi-partate receptor; PND, post-sorufin-O-dealkylase; TBARS,rcury.ealth Centre, 50 Colombine
Canada K1A 0L2. Tel.: +1 613
. Pelletier).
Aschner et al., 2007). Perturbation of glutamate transport in astro-cytes by MeHg can lead to over-stimulation and dysfunction ofthe N-methyl-D-aspartate receptors (NMDA-R) (Yin et al., 2007).The NMDA-R is part of the glutamatergic system which is crucialfor neuronal plasticity, learning and memory (Scheetz andConstantine-Paton, 1994). During fetal development, NMDA-Rsare also involved in the establishment of neuronal circuitry(Haberny et al., 2002), which might constitute an additional targetof MeHg toxicity. While NMDA-R mediated excitotoxicity can leadto the generation of reactive oxygen species (Gasso et al., 2001),MeHg can also generate oxidative stress through direct perturba-tion of mitochondrial functions (Aschner et al., 2007). The resultantoxidative stress can lead to cell membrane damage, calcium dereg-ulation, enzyme and cell signalling interference, microtubule disas-sembly, and ultimately, cell death (do Nascimento et al., 2008).
MeHg co-exposure studies provide further evidence on the roleof glutamate excitotoxicity and oxidative stress in MeHg-inducedneurotoxicity. MeHg-induced neurotoxicity can be mitigated byco-exposure with NMDA-R antagonists (Zhang et al., 2003; Faroet al., 2002). Various purified phytochemicals or plant extractshave been shown to confer some protection against NMDA recep-tor agonist-induced excitotoxicity and oxidative stress in vivo and
rights reserved.

P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342 2337
in vitro (Silva et al., 2008; Campos-Esparza et al., 2009; Chen et al.,2008; Cho and Lee, 2004) and interestingly, similar findings havebeen reported following co-exposure with MeHg (Lucena et al.,2007; Yongjin et al., 2008; Farina et al., 2005). Possible mecha-nisms involved in the mitigation of MeHg effects by phytochemi-cals may include the reduction of reactive oxygen species,activation of enzymatic antioxidant systems, restoration of themitochondrial membrane potential, and modulation of cell signal-ling pathways (Campos-Esparza et al., 2009). Dietary phytochemi-cals may also affect additional endpoints such as MeHgbioavailability and pharmacokinetics (Passos et al., 2007; Lairdet al., 2009).
Canadian Inuit are exposed to a variety of persistent organicpollutants and methylmercury (MeHg) through the consumptionof their traditional diet (Van Oostdam et al., 2005). Due to geo-graphical, economical and cultural reasons, low consumption offruits and vegetables in Canadian Arctic communities can limittheir plant-based antioxidant intake. Rhododendron tomentosumssp. subarcticum, generally known as Northern Labrador Tea, isan abundant woody tundra plant commonly used by Canadian In-uit as a traditional medicinal tea for the prevention and treatmentof respiratory illnesses (Black et al., 2008). Extracts of R. tomento-sum have high free radical scavenging activity in vitro and containmany well characterized antioxidant phytochemicals such as quer-cetin and caffeic acid (Black et al., 2011). Consumption of thisplant, which is already part of the cultural background of manyNorthern Communities, may therefore represent a convenientand practical method to increase phytochemical-based antioxidantintake in Canadian Arctic populations exposed to MeHg.
The current study was designed to test if co-exposure with anti-oxidants present in R. tomentosum extract, hereafter referred to as‘‘Tea’’, can mitigate methylmercury-induced neurotoxicity in a ratperinatal exposure model. Dams were dosed daily throughout ges-tation and lactation with MeHg and Tea administered either indi-vidually or together. The effects of co-exposure on pup growth,development, tissue mercury concentrations and oxidative status,neuromuscular functions, neurobehaviour and brain NMDA recep-tor levels were assessed.
2. Materials and methods
2.1. Plant extract preparation
R. tomentosum ssp. subarcticum was collected in a single batch in July 2006 nearIqaluit, Nunavut, Canada and taxonomically verified. Plant vouchers were preparedand deposited at the University of Montreal Herbarium (call # IQAL04-1A andIQAL04-1B). Dried plant material was ground using a filter pore size of 1 mm andextracted in 80% ethanol twice for 24 h. The solvent was removed by freeze-dryingand the homogenized extract stored in amber vials at �20 �C. Phytochemical con-tent and antioxidant properties of the extract were characterized and quantified(Black et al., 2011).
2.2. Animal treatment and tissue collection
All experimental procedures adhered to the Canadian Council on Animal CareGuidelines and were approved in advance by Health Canada’s Animal Care Commit-tee. Adult (11 weeks) male and female Sprague–Dawley rats were obtained fromCharles River (Saint-Constant, Canada) and kept in 35 � 30 � 17 cm hanging poly-carbonate cages on shaved wood beddings with ad libitum access to tap waterand rat chow (Labdiet 5018, Purina Mills). The room was maintained at 22 ± 1 �Cand 50 ± 10% humidity on a reversed 12-h light cycle (dark phase from 7:00 to19:00).
After a 2-week acclimatization period, breeding was initiated by introducingtwo females into the male’s home cage. Females were checked daily for the pres-ence of a vaginal plug or sperm, which was defined as gestational day 0 (GD 0).At GD 0 females were housed individually and randomly assigned to one of the fourtreatment groups; MeHg (2.0 mg/kg bw, n = 12), Tea (100 mg/kg bw, n = 10),MeHg + Tea (2.0 mg MeHg/kg bw & 100 mg Tea extract/kg bw, n = 11), or Control(vehicle, n = 10). Dams were weighed and dosed daily from GD 1 to PND 21. R.tomentosum extract dissolved in high-grade 100% ethanol (500 mg/ml) was appliedto small graham cookies (Teddy Graham, Nabisco Ltd., Toronto, Canada) and the
ethanol was allowed to evaporate overnight in a fume hood. MeHg (Sigma–Aldrich,St. Louis, MO, USA) dissolved in corn oil (2 mg/ml) was then applied to the cookie.Control and Tea-treated dams were dosed with a cookie containing the sameweight-adjusted volume of corn oil. Offspring were weighed and counted at partu-rition, and litters were culled to four males and four females on PND 4. Dams andpups (grouped by gender) were weighed daily, and morbidity monitored by veter-inary technicians. At PND 14 and PND 21 two males and two females from each lit-ter were sacrificed under isoflurane anaesthesia. Brains were cut along thelongitudinal axis and half brains were kept for the analysis of Hg residues, oxidativestress and NMDA-R. At PND 21 the dams were also sacrificed using the same pro-cedure. Blood, brain and liver tissues were collected, flash frozen in liquid nitrogenand stored at �80 �C.
2.3. Pup early development measures
Eye and ear opening were assessed on a daily basis in all pups, beginning onPND 11. Grip strength was assessed in the same subset of pups at both PND 12and 15 using the previously described forepaw suspension test (Bowers et al.,2004). Briefly, each pup was suspended by placing its forepaws on a horizontal3 mm radius metal wire, 30 cm above soft bedding. The time the pup held ontothe wire was recorded for a maximum of 60 s. Testing was completed during thefirst 4 h of the pup’s active dark cycle.
2.4. Pup behaviour assessment
Behavioural tests were performed on a subset of one male and one female pupfrom each litter during the first 4 h of their active dark cycle under 20-watt red bulblighting. Tests were conducted in a sound attenuated room with background whitenoise generated by 2 speakers (80 dB at source) located in opposite corners of theroom.
The open field test was conducted on PND 16 pups to determine spontaneousmotor activity and exploratory behaviour (Russell, 1973). Animals were tested indi-vidually by placing the pup in the start corner of a clean 17 � 17 � 1200 polypropyl-ene chamber. Chambers were equipped with infrared beams and detectors dividingthe floor area into 289 one inch squares (model ENV-515, Med Associates Inc., USA).The location of the pup was monitored for 25 min by recording photocell beambreaks, using Activity Monitor Software 4.33 (Med Associates Inc., USA) on a com-puter located in an adjacent room. Ambulatory time was defined as the time thepup spent crossing the squares and vertical counts as the number of vertical rear-ings detected by a separate set of infrared beams located 1.500 above the chamber’sfloor. Thigmotaxis (preference for the periphery) was estimated by the amount oftime spent within 2 inches of the walls.
The acoustic startle test was conducted on PND 20 pups to evaluate sensorimo-tor gating and acoustic reactivity (Geyer and Swerdlow, 2001). Automated startlechambers (model 6500-0091-E, San Diego Instruments Inc., USA) with a ventilatedclear holding tube mounted on a motion detector and a wideband audio speakerwere used. The pup was allowed to acclimatize for 5 min with 65 dB of backgroundwhite noise. Startle responses of each pup were measured over 120 trials consistingof a prepulse stimulus (0, 70, 85, and 100 dB) followed 50 ms later with a 50 msstartle stimulus (105, 112, and 118 dB). Individual trials were spaced by randomintervals ranging from 5 to 10 s. The first and last 12 trials consisted of startle onlytrials (prepulse value = 0 dB) with 3 trials at each startle intensity (105, 112, and118 dB). The remaining trials consisted of prepulse trials with 8 trials at each star-tle-prepulse combination (4 prepulse and 3 startle intensities). In addition, 10 nulltrials with no acoustic stimuli were randomly presented throughout the testingperiod. Data was recorded on a computer in an adjacent room (SR-LAB Startle ReflexSystem 5.0, San Diego Instruments Inc., USA). Startle response time and amplitudewere averaged over all trials for each startle-prepulse combination. Amplitude wasnormalized to pup weight by dividing the amplitude by the weight of the pup theday of the trial. The prepulse inhibition (PPI), or the suppression of the startle re-sponse by a prepulse, was calculated as % PPI = [1�(startle amplitude forpulse + prepulse trial/startle amplitude for pulse only trial)] � 100.
2.5. Mercury quantification
Total Hg (THg) was quantified by thermal decomposition followed by dual stepgold amalgamation and detection by cold vapor atomic absorption spectrophotom-etry using a SP-3D mercury analyzer (Nippon Instruments, Japan) as described pre-viously (Basu et al., 2005; Al-Reasi et al., 2007). Approximately 10 mg ofunprocessed sample was placed in a ceramic boat covered with layers of additiveM (sodium carbonate and calcium hydroxide) and additive B (aluminum oxide),and placed into the thermal decomposition chamber for analysis.
MeHg concentration was assessed as described previously (Cai et al., 1996;Al-Reasi et al., 2007). Briefly, the samples were subject to alkaline digestionfollowed by acid leaching. Organomercuric species were extracted in dichlorometh-ane, filtered and placed in amber vials for analysis by capillary gas chromatography(HP6890 Series, Agilent, USA) coupled with atomic fluorescence spectrometry (PSAMerlin Millennium Detector, PS Analytical, Orpington, UK)
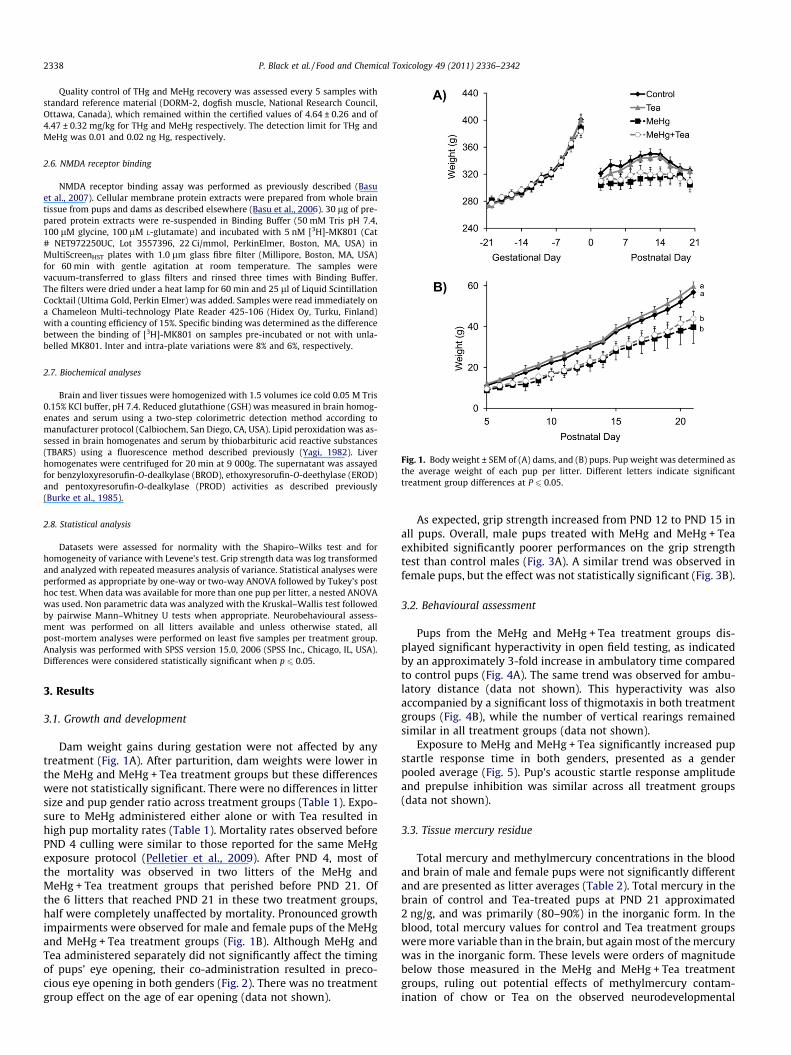
Fig. 1. Body weight ± SEM of (A) dams, and (B) pups. Pup weight was determined asthe average weight of each pup per litter. Different letters indicate significanttreatment group differences at P 6 0.05.
2338 P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342
Quality control of THg and MeHg recovery was assessed every 5 samples withstandard reference material (DORM-2, dogfish muscle, National Research Council,Ottawa, Canada), which remained within the certified values of 4.64 ± 0.26 and of4.47 ± 0.32 mg/kg for THg and MeHg respectively. The detection limit for THg andMeHg was 0.01 and 0.02 ng Hg, respectively.
2.6. NMDA receptor binding
NMDA receptor binding assay was performed as previously described (Basuet al., 2007). Cellular membrane protein extracts were prepared from whole braintissue from pups and dams as described elsewhere (Basu et al., 2006). 30 lg of pre-pared protein extracts were re-suspended in Binding Buffer (50 mM Tris pH 7.4,100 lM glycine, 100 lM L-glutamate) and incubated with 5 nM [3H]-MK801 (Cat# NET972250UC, Lot 3557396, 22 Ci/mmol, PerkinElmer, Boston, MA, USA) inMultiScreenHST plates with 1.0 lm glass fibre filter (Millipore, Boston, MA, USA)for 60 min with gentle agitation at room temperature. The samples werevacuum-transferred to glass filters and rinsed three times with Binding Buffer.The filters were dried under a heat lamp for 60 min and 25 ll of Liquid ScintillationCocktail (Ultima Gold, Perkin Elmer) was added. Samples were read immediately ona Chameleon Multi-technology Plate Reader 425-106 (Hidex Oy, Turku, Finland)with a counting efficiency of 15%. Specific binding was determined as the differencebetween the binding of [3H]-MK801 on samples pre-incubated or not with unla-belled MK801. Inter and intra-plate variations were 8% and 6%, respectively.
2.7. Biochemical analyses
Brain and liver tissues were homogenized with 1.5 volumes ice cold 0.05 M Tris0.15% KCl buffer, pH 7.4. Reduced glutathione (GSH) was measured in brain homog-enates and serum using a two-step colorimetric detection method according tomanufacturer protocol (Calbiochem, San Diego, CA, USA). Lipid peroxidation was as-sessed in brain homogenates and serum by thiobarbituric acid reactive substances(TBARS) using a fluorescence method described previously (Yagi, 1982). Liverhomogenates were centrifuged for 20 min at 9 000g. The supernatant was assayedfor benzyloxyresorufin-O-dealkylase (BROD), ethoxyresorufin-O-deethylase (EROD)and pentoxyresorufin-O-dealkylase (PROD) activities as described previously(Burke et al., 1985).
2.8. Statistical analysis
Datasets were assessed for normality with the Shapiro–Wilks test and forhomogeneity of variance with Levene’s test. Grip strength data was log transformedand analyzed with repeated measures analysis of variance. Statistical analyses wereperformed as appropriate by one-way or two-way ANOVA followed by Tukey’s posthoc test. When data was available for more than one pup per litter, a nested ANOVAwas used. Non parametric data was analyzed with the Kruskal–Wallis test followedby pairwise Mann–Whitney U tests when appropriate. Neurobehavioural assess-ment was performed on all litters available and unless otherwise stated, allpost-mortem analyses were performed on least five samples per treatment group.Analysis was performed with SPSS version 15.0, 2006 (SPSS Inc., Chicago, IL, USA).Differences were considered statistically significant when p 6 0.05.
3. Results
3.1. Growth and development
Dam weight gains during gestation were not affected by anytreatment (Fig. 1A). After parturition, dam weights were lower inthe MeHg and MeHg + Tea treatment groups but these differenceswere not statistically significant. There were no differences in littersize and pup gender ratio across treatment groups (Table 1). Expo-sure to MeHg administered either alone or with Tea resulted inhigh pup mortality rates (Table 1). Mortality rates observed beforePND 4 culling were similar to those reported for the same MeHgexposure protocol (Pelletier et al., 2009). After PND 4, most ofthe mortality was observed in two litters of the MeHg andMeHg + Tea treatment groups that perished before PND 21. Ofthe 6 litters that reached PND 21 in these two treatment groups,half were completely unaffected by mortality. Pronounced growthimpairments were observed for male and female pups of the MeHgand MeHg + Tea treatment groups (Fig. 1B). Although MeHg andTea administered separately did not significantly affect the timingof pups’ eye opening, their co-administration resulted in preco-cious eye opening in both genders (Fig. 2). There was no treatmentgroup effect on the age of ear opening (data not shown).
As expected, grip strength increased from PND 12 to PND 15 inall pups. Overall, male pups treated with MeHg and MeHg + Teaexhibited significantly poorer performances on the grip strengthtest than control males (Fig. 3A). A similar trend was observed infemale pups, but the effect was not statistically significant (Fig. 3B).
3.2. Behavioural assessment
Pups from the MeHg and MeHg + Tea treatment groups dis-played significant hyperactivity in open field testing, as indicatedby an approximately 3-fold increase in ambulatory time comparedto control pups (Fig. 4A). The same trend was observed for ambu-latory distance (data not shown). This hyperactivity was alsoaccompanied by a significant loss of thigmotaxis in both treatmentgroups (Fig. 4B), while the number of vertical rearings remainedsimilar in all treatment groups (data not shown).
Exposure to MeHg and MeHg + Tea significantly increased pupstartle response time in both genders, presented as a genderpooled average (Fig. 5). Pup’s acoustic startle response amplitudeand prepulse inhibition was similar across all treatment groups(data not shown).
3.3. Tissue mercury residue
Total mercury and methylmercury concentrations in the bloodand brain of male and female pups were not significantly differentand are presented as litter averages (Table 2). Total mercury in thebrain of control and Tea-treated pups at PND 21 approximated2 ng/g, and was primarily (80–90%) in the inorganic form. In theblood, total mercury values for control and Tea treatment groupswere more variable than in the brain, but again most of the mercurywas in the inorganic form. These levels were orders of magnitudebelow those measured in the MeHg and MeHg + Tea treatmentgroups, ruling out potential effects of methylmercury contam-ination of chow or Tea on the observed neurodevelopmental
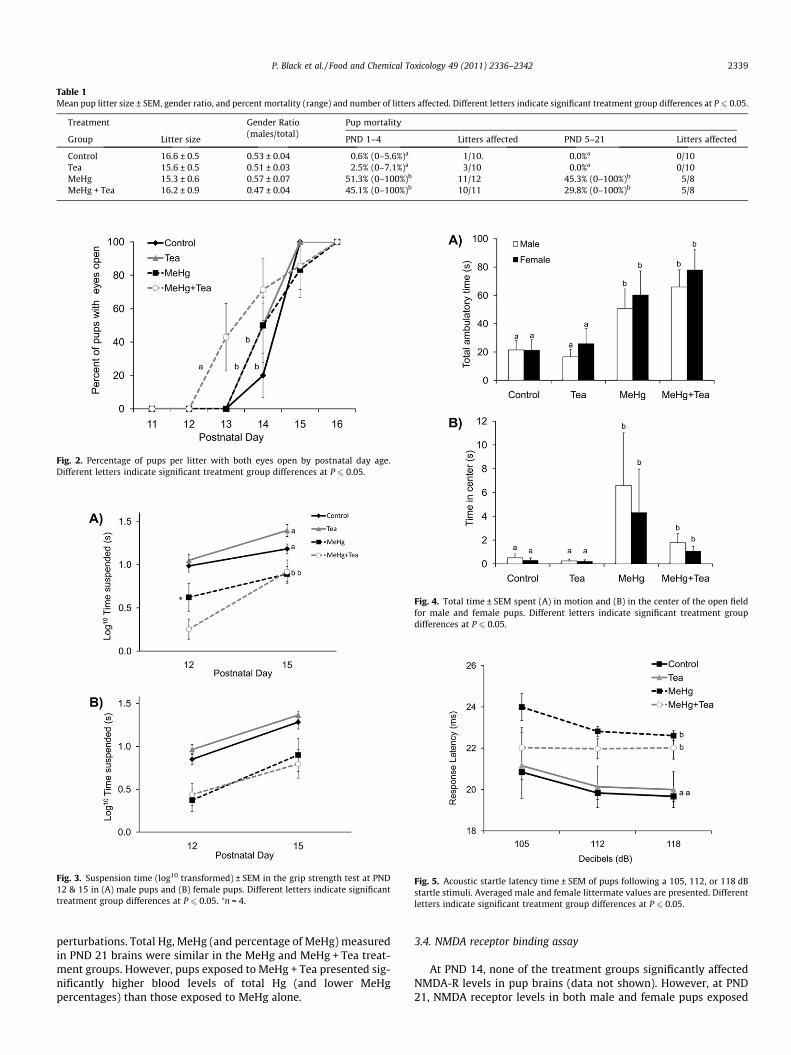
Table 1Mean pup litter size ± SEM, gender ratio, and percent mortality (range) and number of litters affected. Different letters indicate significant treatment group differences at P 6 0.05.
Treatment Gender Ratio(males/total)
Pup mortality
Group Litter size PND 1–4 Litters affected PND 5–21 Litters affected
Control 16.6 ± 0.5 0.53 ± 0.04 0.6% (0–5.6%)a 1/10. 0.0%a 0/10Tea 15.6 ± 0.5 0.51 ± 0.03 2.5% (0–7.1%)a 3/10 0.0%a 0/10MeHg 15.3 ± 0.6 0.57 ± 0.07 51.3% (0–100%)b 11/12 45.3% (0–100%)b 5/8MeHg + Tea 16.2 ± 0.9 0.47 ± 0.04 45.1% (0–100%)b 10/11 29.8% (0–100%)b 5/8
Fig. 2. Percentage of pups per litter with both eyes open by postnatal day age.Different letters indicate significant treatment group differences at P 6 0.05.
Fig. 3. Suspension time (log10 transformed) ± SEM in the grip strength test at PND12 & 15 in (A) male pups and (B) female pups. Different letters indicate significanttreatment group differences at P 6 0.05. ⁄n = 4.
Fig. 4. Total time ± SEM spent (A) in motion and (B) in the center of the open fieldfor male and female pups. Different letters indicate significant treatment groupdifferences at P 6 0.05.
Fig. 5. Acoustic startle latency time ± SEM of pups following a 105, 112, or 118 dBstartle stimuli. Averaged male and female littermate values are presented. Differentletters indicate significant treatment group differences at P 6 0.05.
P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342 2339
perturbations. Total Hg, MeHg (and percentage of MeHg) measuredin PND 21 brains were similar in the MeHg and MeHg + Tea treat-ment groups. However, pups exposed to MeHg + Tea presented sig-nificantly higher blood levels of total Hg (and lower MeHgpercentages) than those exposed to MeHg alone.
3.4. NMDA receptor binding assay
At PND 14, none of the treatment groups significantly affectedNMDA-R levels in pup brains (data not shown). However, at PND21, NMDA receptor levels in both male and female pups exposed
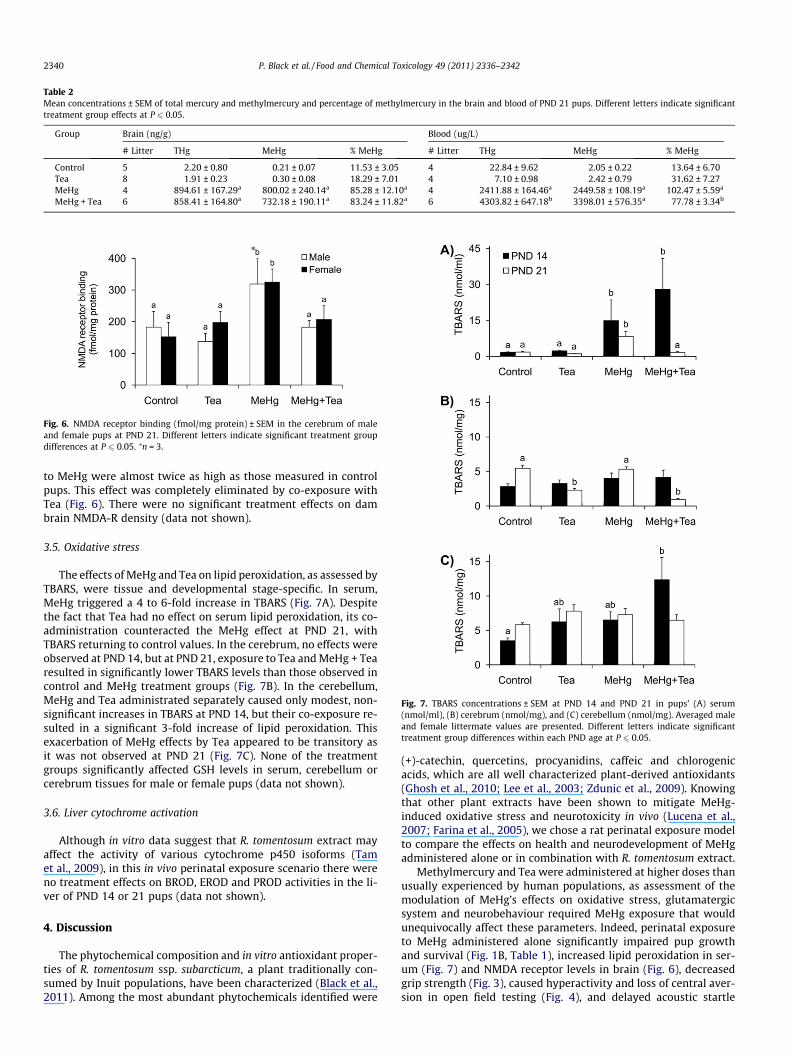
Table 2Mean concentrations ± SEM of total mercury and methylmercury and percentage of methylmercury in the brain and blood of PND 21 pups. Different letters indicate significanttreatment group effects at P 6 0.05.
Group Brain (ng/g) Blood (ug/L)
# Litter THg MeHg % MeHg # Litter THg MeHg % MeHg
Control 5 2.20 ± 0.80 0.21 ± 0.07 11.53 ± 3.05 4 22.84 ± 9.62 2.05 ± 0.22 13.64 ± 6.70Tea 8 1.91 ± 0.23 0.30 ± 0.08 18.29 ± 7.01 4 7.10 ± 0.98 2.42 ± 0.79 31.62 ± 7.27MeHg 4 894.61 ± 167.29a 800.02 ± 240.14a 85.28 ± 12.10a 4 2411.88 ± 164.46a 2449.58 ± 108.19a 102.47 ± 5.59a
MeHg + Tea 6 858.41 ± 164.80a 732.18 ± 190.11a 83.24 ± 11.82a 6 4303.82 ± 647.18b 3398.01 ± 576.35a 77.78 ± 3.34b
Fig. 6. NMDA receptor binding (fmol/mg protein) ± SEM in the cerebrum of maleand female pups at PND 21. Different letters indicate significant treatment groupdifferences at P 6 0.05. ⁄n = 3.
Fig. 7. TBARS concentrations ± SEM at PND 14 and PND 21 in pups’ (A) serum(nmol/ml), (B) cerebrum (nmol/mg), and (C) cerebellum (nmol/mg). Averaged maleand female littermate values are presented. Different letters indicate significanttreatment group differences within each PND age at P 6 0.05.
2340 P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342
to MeHg were almost twice as high as those measured in controlpups. This effect was completely eliminated by co-exposure withTea (Fig. 6). There were no significant treatment effects on dambrain NMDA-R density (data not shown).
3.5. Oxidative stress
The effects of MeHg and Tea on lipid peroxidation, as assessed byTBARS, were tissue and developmental stage-specific. In serum,MeHg triggered a 4 to 6-fold increase in TBARS (Fig. 7A). Despitethe fact that Tea had no effect on serum lipid peroxidation, its co-administration counteracted the MeHg effect at PND 21, withTBARS returning to control values. In the cerebrum, no effects wereobserved at PND 14, but at PND 21, exposure to Tea and MeHg + Tearesulted in significantly lower TBARS levels than those observed incontrol and MeHg treatment groups (Fig. 7B). In the cerebellum,MeHg and Tea administrated separately caused only modest, non-significant increases in TBARS at PND 14, but their co-exposure re-sulted in a significant 3-fold increase of lipid peroxidation. Thisexacerbation of MeHg effects by Tea appeared to be transitory asit was not observed at PND 21 (Fig. 7C). None of the treatmentgroups significantly affected GSH levels in serum, cerebellum orcerebrum tissues for male or female pups (data not shown).
3.6. Liver cytochrome activation
Although in vitro data suggest that R. tomentosum extract mayaffect the activity of various cytochrome p450 isoforms (Tamet al., 2009), in this in vivo perinatal exposure scenario there wereno treatment effects on BROD, EROD and PROD activities in the li-ver of PND 14 or 21 pups (data not shown).
4. Discussion
The phytochemical composition and in vitro antioxidant proper-ties of R. tomentosum ssp. subarcticum, a plant traditionally con-sumed by Inuit populations, have been characterized (Black et al.,2011). Among the most abundant phytochemicals identified were
(+)-catechin, quercetins, procyanidins, caffeic and chlorogenicacids, which are all well characterized plant-derived antioxidants(Ghosh et al., 2010; Lee et al., 2003; Zdunic et al., 2009). Knowingthat other plant extracts have been shown to mitigate MeHg-induced oxidative stress and neurotoxicity in vivo (Lucena et al.,2007; Farina et al., 2005), we chose a rat perinatal exposure modelto compare the effects on health and neurodevelopment of MeHgadministered alone or in combination with R. tomentosum extract.
Methylmercury and Tea were administered at higher doses thanusually experienced by human populations, as assessment of themodulation of MeHg’s effects on oxidative stress, glutamatergicsystem and neurobehaviour required MeHg exposure that wouldunequivocally affect these parameters. Indeed, perinatal exposureto MeHg administered alone significantly impaired pup growthand survival (Fig. 1B, Table 1), increased lipid peroxidation in ser-um (Fig. 7) and NMDA receptor levels in brain (Fig. 6), decreasedgrip strength (Fig. 3), caused hyperactivity and loss of central aver-sion in open field testing (Fig. 4), and delayed acoustic startle

P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342 2341
response time (Fig. 5) in absence of effects on startle amplitude andprepulse inhibition. These findings were generally consistent withthe published literature on the effects of MeHg (Johansson et al.,2007; Pelletier et al., 2009; Beyrouty and Chan, 2006; Farinaet al., 2004).
Except for one tissue and time-specific decrease in lipid perox-idation (Fig. 7), exposure to Tea alone had no significant effect onany of the endpoint measured in the pups. Tea co-administrationmay have altered MeHg pharmacokinetics: Total mercury concen-tration was higher and the relative proportion of MeHg lower inthe blood of the MeHg + Tea treatment group (Table 2). Surpris-ingly, increased mercury levels in the MeHg + Tea animals didnot add to the adverse effects of MeHg on pup development andneurobehaviour, as Tea co-administration did not affect MeHg’simpact on pups growth and mortality (Fig. 1B, Table 1), gripstrength (Fig. 3) ambulatory behaviour in open field testing(Fig. 4) and acoustic startle response latency (Fig. 5). However,Tea co-exposure abolished MeHg effect on brain NMDA receptorbinding (Fig. 6) and depending on the tissue and time point as-sessed, either exacerbated or antagonized MeHg effects on lipidperoxidation (Fig. 7). Finally, although exposure to MeHg or Teaadministered alone had no effect on the timing of eye opening,co-exposure resulted in precocious eye opening (Fig. 2). This obser-vation has also been reported following MeHg co-exposure withselenium and vitamin E (Beyrouty and Chan, 2006) and polychlori-nated biphenyl 126 (Vitalone et al., 2008). However, the etiology,significance and consequences of this phenomenon remainobscure.
Most reports describing animal exposure to purified phyto-chemicals or phytochemical extracts generally attributed beneficialeffects at least in part to their antioxidant properties (Ghosh et al.,2010; Khalatbary et al., 2010). Although R. tomentosum extractexhibited high in vitro antioxidant potency, (Black et al., 2011)our observations in developing rat pups at PND 14 and 21 revealeda more complex situation. In the cerebrum, exposure to Tea admin-istered alone or in combination with MeHg lowered lipid peroxida-tion below levels observed in the control group at PND 21, while ithad no effects at PND 14. Results were more surprising in PND 14cerebellum, where both Tea and MeHg produced a non-significantincrease in lipid peroxidation when administered separately, anda significant 3-fold increase when administered together. Exposureto MeHg significantly increased lipid peroxidation in serum at bothPND 14 and 21, and Tea co-exposure abolished MeHg-induced in-crease in lipid peroxidation only at PND 21. Overall, Tea co-expo-sure potentiated the effect of MeHg on lipid peroxidation at PND14 and antagonized it at PND 21. Several possible mechanisms, suchas different concentrations of mercury and phytochemicals at PND14 and PND 21, may explain these observations. Interestingly, thetherapeutic dose-range of phytochemicals can be narrow (Ossolaet al., 2009) and phytochemical potentiation of MeHg effects on li-pid peroxidation has also been reported in adult mouse cerebellum(Martins et al., 2009).
Perinatal exposure to MeHg resulted in hyperactivity and loss ofthigmotaxis (preference for the periphery) in open field testing.These observations are commonly reported following rat exposureto NMDA-R antagonists (Koros et al., 2007; Adamec et al., 1999;Plaznik et al., 1994) and interestingly, MeHg exposure also affectedNMDA-R binding in vitro. However, in our study Tea counteractedMeHg’s effect on NMDA-R binding but not on open field hyperac-tivity or thigmotaxis, arguing against a direct causal relationshipbetween MeHg-induced alterations of NMDA-R binding and spon-taneous behaviour.
Total mercury and methylmercury concentrations measured inthe blood of MeHg + Tea exposed pups were higher than thosemeasured in MeHg-treated pups, but this trend was statisticallysignificant only for total mercury. The proportion of total mercury
present as methylmercury was also significantly lower in theMeHg + Tea treatment group, suggesting that MeHg demethylationmay have been affected. Interestingly, MeHg demethylation ratecan be modulated by oxidative stress and reactive oxygen speciesin vitro (Shapiro and Chan, 2008; Yasutake and Hirayama, 2001),and in vivo alteration of MeHg bioavailability and pharmacokinet-ics by diet and phytochemicals has also been observed, including inhuman volunteers (Rowland et al., 1984; Canuel et al., 2006).Although MeHg demethylation and pharmacokinetic was not theprimary focus of this investigation, our preliminary observationsfollowing exposure to high levels of MeHg and Tea warrant furtherinvestigations at lower, more environmentally relevant doses ofMeHg and Tea.
In summary, perinatal exposure to R. tomentosum extractadministered alone had barely any effect on pups health and devel-opment, but it was nevertheless able to modulate methylmercury’seffects on a few molecular endpoints. However, evaluation of co-exposure effects was dependant on the developmental stage andendpoints assessed, as Tea co-administration had no significant ef-fects on most MeHg-induced perturbations, counteracted or poten-tiated MeHg’s effects on other endpoints and affected new,previously undisturbed parameters. The molecular mechanismsof MeHg toxicity are still incompletely characterized, and ourknowledge of the possible interactions with nutrients is even morerudimentary. Our results illustrate the complex nature of contam-inant–nutrients interaction and advocate for careful considerationof potentially beneficial and harmful effects of diet on contaminanttoxicity. A better understanding of the molecular mechanismsunderlying these interactions will ultimately lead to better humanhealth risk assessment and dietary advices for the mitigation ofmethylmercury toxicity in vulnerable populations.
Conflict of Interest
The authors have no conflict of interest to declare.
Acknowledgements
This study was funded in part by the Northern ContaminantsProgram from Indian and Northern Affairs Canada, and NSERC Dis-covery Grant. The authors want to thank Dr. Alain Cuerrier at theMontreal Botanical Gardens for taxonomic verification, EmmanuelYumvihoze at the University of Ottawa for technical assistance onmercury quantification, Karen Leingartner for liver enzymatic as-says and Drs. Phil Shwed and Daniel Desaulniers for critical reviewof this manuscript.
References
Adamec, R.E., Burton, P., Shallow, T., Budgell, J., 1999. NMDA receptors mediatelasting increases in anxiety-like behavior produced by the stress of predatorexposure – implications for anxiety associated with posttraumatic stressdisorder. Physiol. Behav. 65, 723–737.
Al-Reasi, H.A., Ababneh, F.A., Lean, D.R., 2007. Evaluating mercury biomagnificationin fish from a tropical marine environment using stable isotopes (delta13C anddelta15N). Environ. Toxicol. Chem. 26, 1572–1581.
Aschner, M., Syversen, T., Souza, D.O., Rocha, J.B., Farina, M., 2007. Involvement ofglutamate and reactive oxygen species in methyl mercury neurotoxicity. Braz. J.Med. Biol. Res. 40, 285–291.
Basu, N., Scheuhammer, A., Grochowina, N., Klenavic, K., Evans, D., O’Brien, M., Chan,H.M., 2005. Effects of mercury on neurochemical receptors in wild river otters(Lontra canadensis). Environ. Sci. Technol. 39, 3585–3591.
Basu, N., Scheuhammer, A.M., Rouvinen-Watt, K., Grochowina, N., Evans, R.D.,O’Brien, M., Chan, H.M., 2007. Decreased N-methyl-D-aspartic acid (NMDA)receptor levels are associated with mercury exposure in wild and captive mink.Neurotoxicology 28, 587–593.
Basu, N., Scheuhammer, A.M., Rouvinen-Watt, K., Grochowina, N., Klenavic, K.,Evans, R.D., Chan, H.M., 2006. Methylmercury impairs components of thecholinergic system in captive mink (Mustela vison). Toxicol. Sci. 91, 202–209.

2342 P. Black et al. / Food and Chemical Toxicology 49 (2011) 2336–2342
Beyrouty, P., Chan, H.M., 2006. Co-consumption of selenium and vitamin E alteredthe reproductive and developmental toxicity of methylmercury in rats.Neurotoxicol. Teratol. 28, 49–58.
Black, P.L., Arnason, J.T., Cuerrier, A., 2008. Medicinal plants used by the Inuit ofQikiqtaaluk (Baffin Island, Nunavut). Bot. 86, 157–163.
Black, P., Saleem, A., Dunford, A., Guerrero-Analco, J., Walshe-Roussel, B., Haddad, P.,Cuerrier, A., Arnason, J.T., 2011. Seasonal Variation of Phenolic Constituents andMedicinal Activities of Northern Labrador Tea, Rhododendron tomentosum ssp.subarcticum, an Inuit and Cree First Nations Traditional Medicine. Planta Med.doi:10.1055/s-0030-1270968.
Bowers, W.J., Nakai, J.S., Chu, I., Wade, M.G., Moir, D., Yagminas, A., Gill, S., Pulido, O.,Meuller, R., 2004. Early developmental neurotoxicity of a PCB/organochlorinemixture in rodents after gestational and lactational exposure. Toxicol. Sci. 77,51–62.
Burke, M.D., Thompson, S., Elcombe, C.R., Halpert, J., Haaparanta, T., Mayer, R.T.,1985. Ethoxy-, pentoxy- and benzyloxyphenoxazones and homologues: a seriesof substrates to distinguish between different induced cytochromes P-450.Biochem. Pharmacol. 34, 3337–3345.
Cai, Y., Jaff, R., Alli, A., Jones, R., 1996. Determination of organo mercurycompounds in aqueous samples by capillary gas chromatography—atomicfluorescence spectrometry following solid-phase extraction. Anal. Chim. Acta.334, 151–159.
Campos-Esparza, M.R., Sanchez-Gomez, M.V., Matute, C., 2009. Molecularmechanisms of neuroprotection by two natural antioxidant polyphenols. CellCalcium 45, 358–368.
Canuel, R., de Grosbois, S.B., Lucotte, M., Atikesse, L., Larose, C., Rheault, I., 2006.New evidence on the effects of tea on mercury metabolism in humans. Arch.Environ. Occup. Health 61, 232–238.
Chapman, L., Chan, H.M., 2000. The influence of nutrition on methyl mercuryintoxication. Environ. Health Perspect. 108 (Suppl 1), 29–56.
Chen, C.M., Lin, J.K., Liu, S.H., Lin-Shiau, S.Y., 2008. Novel regimen throughcombination of memantine and tea polyphenol for neuroprotection againstbrain excitotoxicity. J. Neurosci. Res. 86, 2696–2704.
Cho, J., Lee, H.K., 2004. Wogonin inhibits excitotoxic and oxidative neuronal damagein primary cultured rat cortical cells. Eur. J. Pharmacol. 485, 105–110.
do Nascimento, J.L., Oliveira, K.R., Crespo-Lopez, M.E., Macchi, B.M., Maues, L.A.,Pinheiro Mda, C., Silveira, L.C., Herculano, A.M., 2008. Methylmercuryneurotoxicity & antioxidant defenses. Indian J. Med. Res. 128, 373–382.
Farina, M., Franco, J.L., Ribas, C.M., Meotti, F.C., Missau, F.C., Pizzolatti, M.G., Dafre,A.L., Santos, A.R., 2005. Protective effects of Polygala paniculata extract againstmethylmercury-induced neurotoxicity in mice. J. Pharm. Pharmacol. 57, 1503–1508.
Farina, M., Soares, F.A., Zeni, G., Souza, D.O., Rocha, J.B., 2004. Additive pro-oxidativeeffects of methylmercury and ebselen in liver from suckling rat pups. Toxicol.Lett. 146, 227–235.
Faro, L.R., do Nascimento, J.L., Alfonso, M., Duran, R., 2002. Protection ofmethylmercury effects on the in vivo dopamine release by NMDA receptorantagonists and nitric oxide synthase inhibitors. Neuropharmacology 42, 612–618.
Gasso, S., Cristofol, R.M., Selema, G., Rosa, R., Rodriguez-Farre, E., Sanfeliu, C., 2001.Antioxidant compounds and Ca(2+) pathway blockers differentially protectagainst methylmercury and mercuric chloride neurotoxicity. J. Neurosci. Res.66, 135–145.
Geyer, M.A., Swerdlow, N.R., 2001. Measurement of startle response, prepulseinhibition, and habituation. In: Gerfen, C. (Ed.), Current Protocols inNeuroscience. John Wiley & Sons Inc., 8.7.1–8.7.15.
Ghosh, D., Ghosh, S., Sarkar, S., Ghosh, A., Das, N., Saha, K.D., Mandal, A.K., 2010.Quercetin in vesicular delivery systems: evaluation in combating arsenic-induced acute liver toxicity associated gene expression in rat model. Chem. Biol.Interact. 186, 61–71.
Haberny, K.A., Paule, M.G., Scallet, A.C., Sistare, F.D., Lester, D.S., Hanig, J.P., Slikker Jr,W., 2002. Ontogeny of the N-methyl-D-aspartate (NMDA) receptor system andsusceptibility to neurotoxicity. Toxicol. Sci. 68, 9–17.
Khalatbary, A.R., Tiraihi, T., Boroujeni, M.B., Ahmadvand, H., Tavafi, M., Tamjidipoor,A., 2010. Effects of epigallocatechin gallate on tissue protection and functionalrecovery after contusive spinal cord injury in rats. Brain Res. 1306, 168–175.
Koros, E., Rosenbrock, H., Birk, G., Weiss, C., Sams-Dodd, F., 2007. The selectivemGlu5 receptor antagonist MTEP, similar to NMDA receptor antagonists,induces social isolation in rats. Neuropsychopharmacology 32, 562–576.
Johansson, C., Castoldi, A.F., Onishchenko, N., Manzo, L., Vahter, M., Ceccatelli, S.,2007. Neurobehavioural and molecular changes induced by methylmercuryexposure during development. Neurotox Res. 11, 241–260.
Laird, B.D., Shade, C., Gantner, N., Chan, H.M., Siciliano, S.D., 2009. Bioaccessibility ofmercury from traditional northern country foods measured using an in vitro
gastrointestinal model is independent of mercury concentration. Sci. TotalEnviron. 407, 6003–6008.
Lee, K.W., Kim, Y.J., Kim, D.O., Lee, H.J., Lee, C.Y., 2003. Major phenolics in apple andtheir contribution to the total antioxidant capacity. J. Agric. Food Chem. 51,6516–6520.
Lucena, G.M., Franco, J.L., Ribas, C.M., Azevedo, M.S., Meotti, F.C., Gadotti, V.M.,Dafre, A.L., Santos, A.R., Farina, M., 2007. Cipura paludosa extract preventsmethyl mercury-induced neurotoxicity in mice. Basic Clin. Pharmacol. Toxicol.101, 127–131.
Martins, R.P., Braga Hde, C., da Silva, A.P., Dalmarco, J.B., de Bem, A.F., dos Santos,A.R., Dafre, A.L., Pizzolatti, M.G., Latini, A., Aschner, M., Farina, M., 2009.Synergistic neurotoxicity induced by methylmercury and quercetin in mice.Food Chem. Toxicol. 47, 645–649.
Ossola, B., Kaariainen, T.M., Mannisto, P.T., 2009. The multiple faces of quercetin inneuroprotection. Expert Opin. Drug Saf. 8, 397–409.
Passos, C.J., Mergler, D., Fillion, M., Lemire, M., Mertens, F., Guimaraes, J.R., Philibert,A., 2007. Epidemiologic confirmation that fruit consumption influencesmercury exposure in riparian communities in the Brazilian Amazon. Environ.Res. 105, 183–193.
Pelletier, G., Masson, S., Wade, M., Nakai, J., Alwiss, R., Mohottalage, S.,Kumarathasan, P., Black, P., Bowers, W.J., Chu, I., Vincent, R., 2009. Contrib-ution of methylmercury, polychlorinated biphenyls and organochlorinepesticides to the toxicity of a contaminant mixture based on Canadian Arcticpopulation blood profiles. Toxicol. Lett. 184, 176–185.
Plaznik, A., Palejko, W., Nazar, M., Jessa, M., 1994. Effects of antagonists at theNMDA receptor complex in two models of anxiety. Eur. Neuropsychophar-macol. 4, 503–512.
Rice, D.C., 2008. Overview of modifiers of methylmercury neurotoxicity: chemicals,nutrients, and the social environment. Neurotoxicology 29, 761–766.
Rowland, I.R., Robinson, R.D., Doherty, R.A., 1984. Effects of diet on mercurymetabolism and excretion in mice given methylmercury: role of gut flora. Arch.Environ. Health 39, 401–408.
Russell, P.A., 1973. Relationships between exploratory behaviour and fear: a review.Br. J. Psychol. 64, 417–433.
Scheetz, A.J., Constantine-Paton, M., 1994. Modulation of NMDA receptor function:implications for vertebrate neural development. FASEB J. 8, 745–752.
Shapiro, A.M., Chan, H.M., 2008. Characterization of demethylation ofmethylmercury in cultured astrocytes. Chemosphere 74, 112–118.
Silva, B., Oliveira, P.J., Dias, A., Malva, J.O., 2008. Quercetin, kaempferol andbiapigenin from Hypericum perforatum are neuroprotective against excitotoxicinsults. Neurotox. Res. 13, 265–279.
Tam, T.W., Liu, R., Arnason, J.T., Krantis, A., Staines, W.A., Haddad, P.S., Foster, B.C.,2009. Actions of ethnobotanically selected Cree anti-diabetic plants on humancytochrome P450 isoforms and flavin-containing monooxygenase 3. J.Ethnopharmacol. 126, 119–126.
Van Oostdam, J., Donaldson, S.G., Feeley, M., Arnold, D., Ayotte, P., Bondy, G., Chan,L., Dewaily, E., Furgal, C.M., Kuhnlein, H., Loring, E., Muckle, G., Myles, E.,Receveur, O., Tracy, B., Gill, U., Kalhok, S., 2005. Human health implications ofenvironmental contaminants in Arctic Canada: a review. Sci. Total Environ.351–352, 165–246.
Vitalone, A., Catalani, A., Chiodi, V., Cinque, C., Fattori, V., Goldoni, M., Matteucci, P.,Poli, D., Zuena, A., Costa, L.G., 2008. Neurobehavioral assessment of rats exposedto low doses of PCB126 and methyl mercury during development. Environ.Toxicol. Pharmacol. 25, 103–113.
Yagi, K., 1982. Assay for Serum Lipid Peroxide Level and its Clinical Significance. In:Yagi, K. (Ed.), Lipid Peroxide in Biology and Medicine. Academic Press, NewYork, USA, pp. 232–242.
Yasutake, A., Hirayama, K., 2001. Evaluation of methylmercury biotransformationusing rat liver slices. Arch. Toxicol. 75, 400–406.
Yin, Z., Milatovic, D., Aschner, J.L., Syversen, T., Rocha, J.B., Souza, D.O., Sidoryk, M.,Albrecht, J., Aschner, M., 2007. Methylmercury induces oxidative injury,alterations in permeability and glutamine transport in cultured astrocytes.Brain Res. 1131, 1–10.
Yongjin, L., Wei, S., Yindong, L., Yong, Z., Xuefeng, H., Chunmei, S., Hongbo, M.,Changwen, W., Yong, L., 2008. Neuroprotective effects of chlorogenic acidagainst apoptosis of PC12 cells induced by methylmercury. Env. Tox. Pharm. 26,13–21.
Zdunic, G., Godevac, D., Savikin, K., Novakovic, M., Milosavljevic, S., Petrovic, S.,2009. Isolation and identification of phenolic compounds from Hypericumricheri Vill. and their antioxidant capacity. Nat. Prod. Res. doi:10.1080/14786410802401390.
Zhang, J., Miyamoto, K., Hashioka, S., Hao, H.P., Murao, K., Saido, T.C., Nakanishi, H.,2003. Activation of mu-calpain in developing cortical neurons followingmethylmercury treatment. Brain Res. Dev. Brain Res. 142, 105–110.
Related Documents











