This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Research Report
Miniature EPSCs in the lateral superior olive before hearingonset: Regional and cell-type-specific differences andheterogeneous neuromodulatory effects of ATP
Michael Kreinest⁎, Britta Müller, Jennifer Winkelhoff, Eckhard Friauf, Stefan LöhrkeAnimal Physiology Group, Department of Biology, University of Kaiserslautern, PO Box 3049, D-67653 Kaiserslautern, Germany
A R T I C L E I N F O A B S T R A C T
Article history:Accepted 24 July 2009Available online 6 August 2009
Spontaneous activity occurs in the mammalian auditory system prior to hearing onset andis relevant for neuronal differentiation. Growing evidence indicates that miniature events,i.e., action potential-independent synaptic activity, also have some developmentalrelevance. An intriguing question is whether these events are purely stochastic or ratherdisplay specific characteristics. We addressed this question and studied miniatureexcitatory postsynaptic currents (mEPSCs) in morphologically defined neurons of the ratlateral superior olive (LSO) during early neonatal life. To do so, whole-cell recordings fromneurons in acute slices were combined with Lucifer yellow fillings. mEPSCs were identifiedby their TTX insensitivity and their blockade by glutamate receptor antagonists. Altogether,60% of the LSO neurons displayed mEPSCs, and their presence correlated with the celllocation andmorphology. Their percentage was highest in themedial limb (86%) and lowestin the lateral limb (14%). Seventy-seven percent of the neurons with mEPSCs were bipolarcells, whereas 77% of thosewithoutmEPSCs weremultipolar cells. The neuromodulator ATPaffected the frequency of mEPSCs in 61% of the LSO neurons in a heterogeneous manner:both frequency increases and decreases occurred. These data provide further evidence forthe specificity of mEPSCs. Finally, we investigated whether missing cochlear input changesmEPSCs characteristics. Characterizing LSO neurons of CaV1.3-/- mice, which lack cochlea-driven nerve activity, we observed higher mEPSC frequencies and peak amplitudes,indicative of a compensatory response to deprivation. Together, our results demonstratespecific, rather than stochastic, characteristics of mEPSCs in the neonatal LSO, inaccordance with their potential developmental significance.
© 2009 Elsevier B.V. All rights reserved.
Keywords:Adenosine triphosphateAuditory brainstemP2 receptorCaV1.3-/-
LSO
B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
⁎ Corresponding author. Current address: Orthopaedic and Trauma Center, University Hospital Mannheim, Theodor-Kutzer-Ufer 1-3, 68167Mannheim, Germany.
E-mail address: [email protected] (M. Kreinest).Abbreviations: AP5,D-2-amino-5-phosphonovaleric acid; ATP, adenosine-5′-triphosphate; CaV1.3-/-, mice lacking the L-type calcium
channel isoform 1.3; CNQX, 6-cyano-7-nitroquinoxaline-2,3-dione; LSO, lateral superior olive; mEPSC, miniature excitatory postsynapticcurrent; MNTB, medial nucleus of the trapezoid body; NA, numerical aperture; P, postnatal day; PPADS, pyridoxal phosphate-6-azo(benzene-2,4-disulfonic acid); sPSC, spontaneous postsynaptic current; TTX, tetrodotoxin; WT, wild type
0006-8993/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.brainres.2009.07.083
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /b ra i n res
Author's personal copy
1. Introduction
Throughout the nervous system, epigenetic mechanisms,namely, activity-dependent processes, are involved in thedevelopment of neural circuits. Within the auditory system,the activity-dependent processes comprise acoustically evokedactivity as well as spontaneous activity (Friauf and Lohmann,1999; Kandler, 2004; Walmsley et al., 2006). Prior to hearingonset, when synaptic connections become established andrefined, spontaneous activity is the only source for generatingsynaptic responses. In the caudal auditory brainstem, actionpotential activity before hearing onset occurs, e.g., in chickens(Lippe, 1994) and gerbils (Kotak and Sanes, 1995; Woolf andRyan, 1985). A disturbed development of the medial nucleus ofthe trapezoid body (MNTB) was demonstrated in congenitallydeafmice, which lack spontaneous auditory nerve activity (Leaoet al., 2004, 2006). LSO neuron survival in organotypic slicecultures generated before hearing onset crucially depends onsynaptic activity (Ehrlich et al., 1998; Lohmann et al., 1998). Inthese studies, synaptic activity was mimicked by an increasedconcentration of extracellular potassium ions, leading toprolonged depolarizations (Löhrke et al., 1998).
Spontaneous synaptic responses can be action potential-dependent or action potential-independent. The actionpotential-independent synaptic responses represent minia-ture events (mEPSCs or mIPSCs). Growing experimentalevidence led to the suggestion that such miniature eventsmay be functionally relevant during the period of synapsematuration (Otsu and Murphy, 2003; Leao et al., 2005). In theanteroventral cochlear nucleus,mEPSCs occur during a criticalperiod prior to hearing onset and display age-related changes(Lu et al., 2007). In the developing LSO, it is unknown whetherminiature events occur and whether they show some charac-teristic features that can be related to nuclear region, cell type,and age. Such features would provide further evidence insupport of the hypothesis thatminiature events do notmerelyreflect a leaky evoked-release mechanism, i.e., stochasticcharacteristics, but may have a function of their own (Otsuand Murphy, 2003). In the present study, we concentrated onmEPSCs and characterized the nature of these events inmorphologically identified LSO neurons of neonatal rats. Todo so, we combined whole-cell patch-clamp recordings inacute brainstem slices with intracellular fluorescent dye fills.We also assessed whether mEPSC characteristics werechanged upon application of a neuromodulator. We appliedATP because its effects on neurons are well documented inseveral systems (Gu and MacDermott, 1997; Hugel andSchlichter, 2000; Brockhaus et al., 2004; Mendoza-Fernandezet al., 2000; Zhang et al., 2003). Effects on spontaneous PSCswere only recently reported in MNTB neurons (Watano et al.,2004), fostering our study. Finally, we compared mEPSCs fromLSO neurons of normal mice with those obtained from micelacking the L-type calcium channel isoform 1.3 (CaV1.3-/- mice),which leads to the loss of transmitter release from cochlearinner hair cells to auditory nerve fibers (Platzer et al., 2000;Brandt et al., 2003). Therefore, the auditory brainstem structuresof these mice are deprived of cochlea-driven action potentialactivity normally present before hearing onset (Beutner andMoser, 2001; Kros et al., 1998; Marcotti et al., 2003). In theMNTB,it was shown that the deprivation affects thematuration of fast
glutamatergic neurotransmission via the calyx of Held (Erazo-Fischer et al., 2007). We assessed whether the deprivation alsoaffects mEPSC frequency and peak amplitude in LSO neurons,possibly in a compensatory manner.
2. Results
2.1. Spontaneous synaptic activity in LSO neurons ofnormal rats
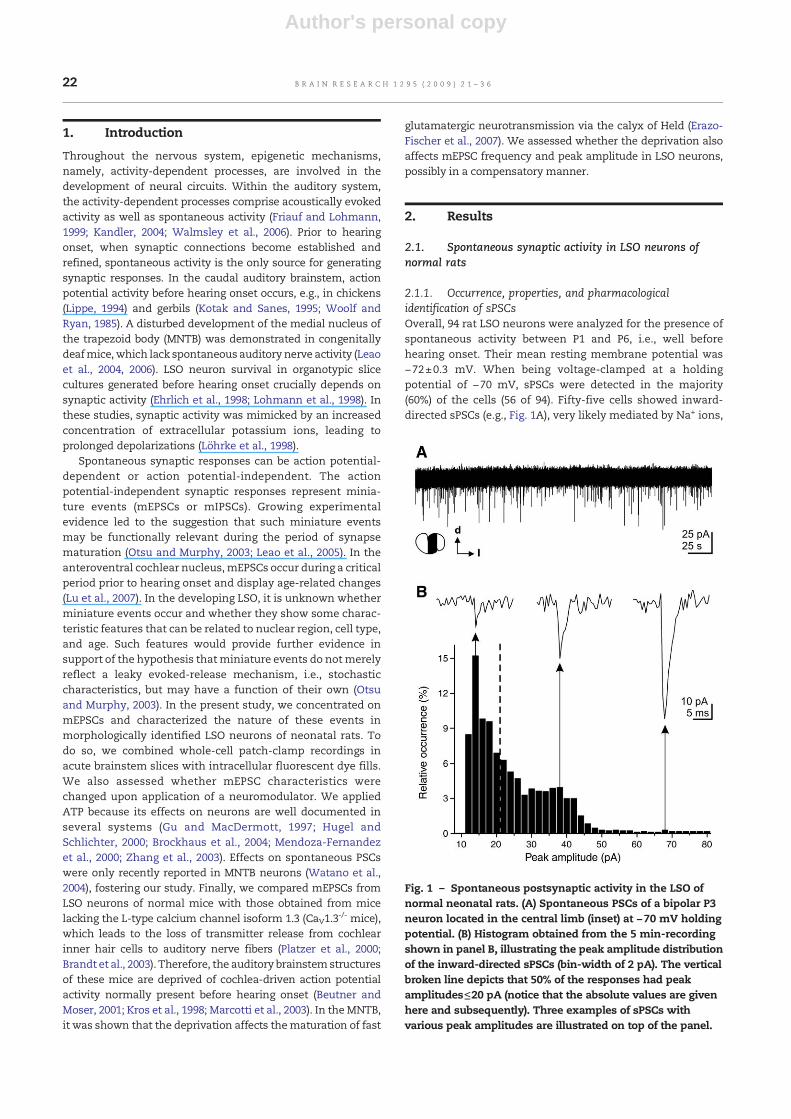
2.1.1. Occurrence, properties, and pharmacologicalidentification of sPSCsOverall, 94 rat LSO neurons were analyzed for the presence ofspontaneous activity between P1 and P6, i.e., well beforehearing onset. Their mean resting membrane potential was−72±0.3 mV. When being voltage-clamped at a holdingpotential of −70 mV, sPSCs were detected in the majority(60%) of the cells (56 of 94). Fifty-five cells showed inward-directed sPSCs (e.g., Fig. 1A), very likely mediated by Na+ ions,
Fig. 1 – Spontaneous postsynaptic activity in the LSO ofnormal neonatal rats. (A) Spontaneous PSCs of a bipolar P3neuron located in the central limb (inset) at −70 mV holdingpotential. (B) Histogram obtained from the 5 min-recordingshown in panel B, illustrating the peak amplitude distributionof the inward-directed sPSCs (bin-width of 2 pA). The verticalbroken line depicts that 50% of the responses had peakamplitudes≤20 pA (notice that the absolute values are givenhere and subsequently). Three examples of sPSCs withvarious peak amplitudes are illustrated on top of the panel.
22 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
Fig. 2 – sPSCs are resistant to TTX. (A) sPSCs of a bipolar P2 neuron located in the medial limb were not influenced by 0.5 μMTTX (5-min bath application indicated by the horizontal bar); thus, they were identified as miniature PSCs (mPSCs). (B) mPSCsfrom the recording in panel A at higher time resolution, illustrating events (triangles) before (i), during (ii), and after (iii) TTXapplication. (C and D) Frequency histogram (C, bin-width of 10 s) and averagemPSCs (D) obtained from the recording in panel A,showing no TTX effect on mPSC frequency and peak amplitude. In panel D, numbers in brackets represent numbers of mPSCsincluded for averaging. (E and F) Normalized data based on 6 neurons, demonstrating that neither the mPSC frequency (E) northe mPSC amplitude (F) was significantly altered by TTX.
23B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
yet not Cl- ions (Na+ reversal potential=+92 mV; Cl- reversalpotential=−87 mV). In one cell, only outward-directed sPSCsoccurred, whereas three neurons showed inward-directedsPSCs as well as outward-directed sPSCs. These outward-directed sPSCs had small amplitudes and were very likelymediated by Cl- influx. We therefore performed our subse-quent analysis exclusively on the inward-directed sPSCs seenin the 55 cells.
A representative peak amplitude distribution of sPSCs froma neuron in the central LSO limb is illustrated in Fig. 1B. Fiftypercent of the sPSCs had peak amplitudes≤20 pA (Fig. 1B;vertical broken line). Overall, the sPSCs displayedmedian peakamplitudes ranging from 11 to 32 pA (n=55 cells). Like in thisexample, sPSC peak amplitudes in all cells were distributed ina non-bell-shaped manner. Because of these low values, wehypothesized that they were miniature PSCs (mPSCs). Some
indirect a priori support for our hypothesis was provided bydata obtained in P7 bushy and stellate cells from theanteroventral cochlear nucleus, where mEPSCs had meanpeak amplitudes of 61 pA and 58 pA, respectively (Lu et al.,2007), i.e., of the same magnitude as our data. To experimen-tally identify the nature of the sPSCs in the LSO, we blockedaction potential activity by applying the sodium channelblocker TTX (0.5 μM). In the neuron illustrated in Figs. 2A–D,the mean sPSC frequency was not significantly altered by a 5-min bath application of TTX (control: 0.3±0.04 Hz; with TTX:0.4±0.04 Hz). More importantly, the variability of sPSC peakamplitudes (Fig. 2B) and themean sPSC peak amplitude (Fig. 2D)were also not changed by TTX. Altogether, the mean sPSCfrequency (Fig. 2E) and themean sPSC peak amplitude (Fig. 2F) of6 cells tested (4 bipolar cells, 1multipolar cell, 1 non-defined cell)were not significantly influenced by TTX (100%±4% of control,
Fig. 3 – mPSCs are mediated by glutamate receptors. (A) mPSCs of a bipolar P2 neuron located in the medial limb werereversibly blocked by 20 μM CNQX and 30 μM AP5 (5-min bath coapplication), demonstrating that they were mediated byionotropic glutamate receptors and represented excitatory mPSCs (mEPSCs). (B) mEPSCs (triangles) before (i), during (ii), andafter (iii) the coapplication of CNQX and AP5 (from the recording shown in panel A at higher time resolution). (C) Frequencyhistogram from the recording shown in panel A, illustrating an almost complete blockade of mEPSCs in the presence of thetwo glutamate receptor antagonists (p=3.3 × 10-12; bin-width of 10 s). (D) Summary based on 5 cells, showing a significant(p=4.2 × 10-4) blockade of mEPSCs by CNQX and AP5 (normalized data).
24 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
Author's personal copy
and 103%±9% of control, respectively), demonstrating that theobserved sPSCs were not mediated by action potential-triggeredtransmitter release, yet rather represented mPSCs.
In order to clarify whether glutamate receptors mediatedthese inward-directed mPSPs, we coapplied the ionotropicglutamate receptor antagonists CNQX (20 μM) and AP5 (30 μM)to the bath in some recordings. In the cell shown in Figs. 3A–C,the coapplication resulted in a reduction of its mean mPSCfrequency from 2.6±0.1 to 0.2±0.1 Hz (Fig. 3C; p=3.3 × 10-12,Mann and Whitney's U-test). In a total of five neurons, the
mean mPSC frequency was significantly reduced by CNQXand AP5 (7±2% of control; p=4.2 × 10-4, Student's paired t-test;Fig. 3D). Together, these results showed that the spontaneousevents from LSO neurons were mEPSCs, mainly mediatedthrough ionotropic glutamate receptors.
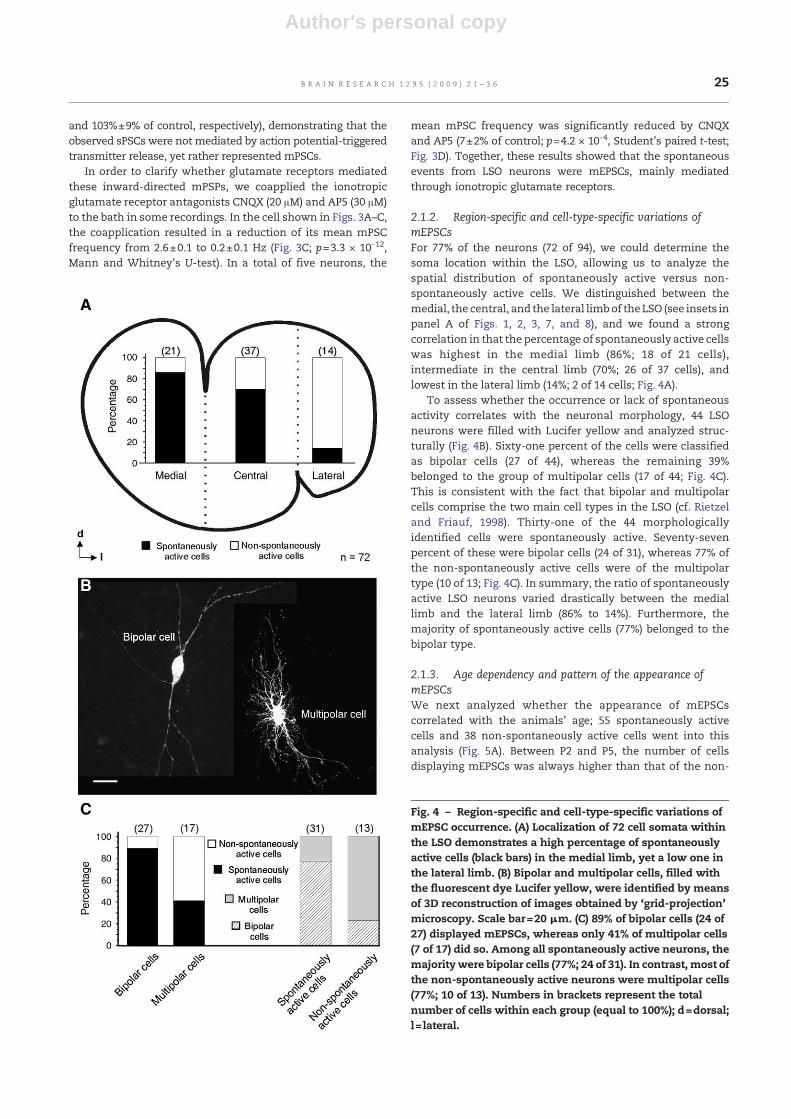
2.1.2. Region-specific and cell-type-specific variations ofmEPSCsFor 77% of the neurons (72 of 94), we could determine thesoma location within the LSO, allowing us to analyze thespatial distribution of spontaneously active versus non-spontaneously active cells. We distinguished between themedial, the central, and the lateral limbof the LSO (see insets inpanel A of Figs. 1, 2, 3, 7, and 8), and we found a strongcorrelation in that the percentage of spontaneously active cellswas highest in the medial limb (86%; 18 of 21 cells),intermediate in the central limb (70%; 26 of 37 cells), andlowest in the lateral limb (14%; 2 of 14 cells; Fig. 4A).
To assess whether the occurrence or lack of spontaneousactivity correlates with the neuronal morphology, 44 LSOneurons were filled with Lucifer yellow and analyzed struc-turally (Fig. 4B). Sixty-one percent of the cells were classifiedas bipolar cells (27 of 44), whereas the remaining 39%belonged to the group of multipolar cells (17 of 44; Fig. 4C).This is consistent with the fact that bipolar and multipolarcells comprise the two main cell types in the LSO (cf. Rietzeland Friauf, 1998). Thirty-one of the 44 morphologicallyidentified cells were spontaneously active. Seventy-sevenpercent of these were bipolar cells (24 of 31), whereas 77% ofthe non-spontaneously active cells were of the multipolartype (10 of 13; Fig. 4C). In summary, the ratio of spontaneouslyactive LSO neurons varied drastically between the mediallimb and the lateral limb (86% to 14%). Furthermore, themajority of spontaneously active cells (77%) belonged to thebipolar type.
2.1.3. Age dependency and pattern of the appearance ofmEPSCsWe next analyzed whether the appearance of mEPSCscorrelated with the animals' age; 55 spontaneously activecells and 38 non-spontaneously active cells went into thisanalysis (Fig. 5A). Between P2 and P5, the number of cellsdisplaying mEPSCs was always higher than that of the non-
Fig. 4 – Region-specific and cell-type-specific variations ofmEPSC occurrence. (A) Localization of 72 cell somata withinthe LSO demonstrates a high percentage of spontaneouslyactive cells (black bars) in the medial limb, yet a low one inthe lateral limb. (B) Bipolar and multipolar cells, filled withthe fluorescent dye Lucifer yellow, were identified by meansof 3D reconstruction of images obtained by ‘grid-projection’microscopy. Scale bar=20 μm. (C) 89% of bipolar cells (24 of27) displayed mEPSCs, whereas only 41% of multipolar cells(7 of 17) did so. Among all spontaneously active neurons, themajoritywere bipolar cells (77%; 24 of 31). In contrast, most ofthe non-spontaneously active neurons were multipolar cells(77%; 10 of 13). Numbers in brackets represent the totalnumber of cells within each group (equal to 100%); d=dorsal;l= lateral.
25B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
spontaneously active cells, and the average ratio of sponta-neously versus non-spontaneously active cells amounted to1.7±0.2 (range: 1.2 to 2.0; Fig. 5A).
Regarding the mean mEPSC frequency, a weak agedependency was observed (Fig. 5B, r=0.2, n=55), i.e., slightlyhigher mean frequencies occurred at older ages compared toyounger ages (e.g., P5, 1.6±0.6 Hz, n=6; P1, 0.5±0.2 Hz, n=3).
No significant difference in mEPSC frequency was observedbetween the two morphological cell types (bipolar cells: 0.9±0.2 Hz, n=24; multipolar cells: 0.7±0.4 Hz, n=7; cf. Fig. 5B).
To assess whether mEPSCs of bipolar and multipolar cellsoccur in a randomor in a temporally patterned fashion (e.g., inbursts), we plotted the sequence of their inter-event intervals.Representative examples of a bipolar cell and a multipolar cell
Fig. 5 – Age dependency and inter-event intervals of mEPSCs. (A) Between P2 and P5, spontaneously active cells werepredominant. In each age column, numbers in brackets represent the ratio between spontaneously active and non-spontaneously active cells; n.p.=not possible to determine. (B) The mean mEPSC frequency of all spontaneously active cellsincreased slightly with age. The correlation coefficient r was obtained by the depicted exponential regression line andamounted to 0.2. (C) Sequence of 345 inter-event intervals within a 5-min period, obtained from a P3 bipolar cell. (D) Histogramof the inter-event intervals depicted in panel C; 50% of the values are ≤0.6 s (vertical broken line; bin-width of 100 ms). (E)Sequence of 117 inter-event intervals within a 5-min period, obtained from a P3multipolar cell. (F) Histogram of the inter-eventintervals depicted in panel E; 50% of the values are ≤1.5 s (vertical broken line; bin-width of 300 ms).
26 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
Author's personal copy
are shown in Figs. 5C and E, and do not reveal any rhythmicity.The bipolar and multipolar cell featured mEPSCs with 50% ofinter-event intervals≤0.6 s and≤1.5 s, respectively (Figs. 5Dand F, vertical broken lines). On average, mEPSCs occurredwith 50% of inter-event intervals≤1.4±0.2 s in bipolar cells(n=24) and≤1.6±0.4 s in multipolar cells (n=7) which was notsignificantly different. These short intervals were irregularlyinterrupted by longer inter-event intervals (Figs. 5C and E),suggesting no temporally patterned, but rather a possiblyrandom appearance of mEPSCs in both neuron types. Togeth-er, the results about mEPSCs during the first postnatal weekcan be summarized as follows: First, cells exhibiting mEPSCsare prevailing between P2 and P5. Second, there is only a slightincrease in the frequency of mEPSCs with age. Third, bipolarandmultipolar cells show a temporally random appearance ofmEPSCs.
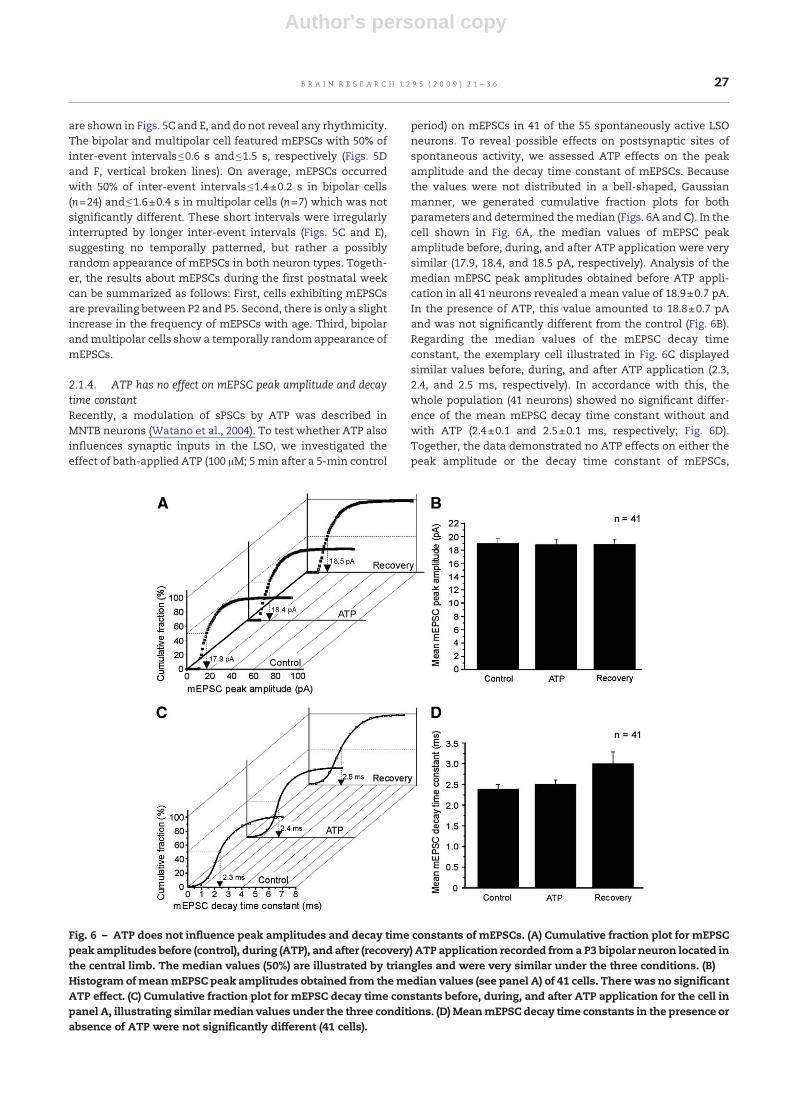
2.1.4. ATP has no effect on mEPSC peak amplitude and decaytime constantRecently, a modulation of sPSCs by ATP was described inMNTB neurons (Watano et al., 2004). To test whether ATP alsoinfluences synaptic inputs in the LSO, we investigated theeffect of bath-applied ATP (100 μM; 5 min after a 5-min control
period) on mEPSCs in 41 of the 55 spontaneously active LSOneurons. To reveal possible effects on postsynaptic sites ofspontaneous activity, we assessed ATP effects on the peakamplitude and the decay time constant of mEPSCs. Becausethe values were not distributed in a bell-shaped, Gaussianmanner, we generated cumulative fraction plots for bothparameters and determined themedian (Figs. 6A and C). In thecell shown in Fig. 6A, the median values of mEPSC peakamplitude before, during, and after ATP application were verysimilar (17.9, 18.4, and 18.5 pA, respectively). Analysis of themedian mEPSC peak amplitudes obtained before ATP appli-cation in all 41 neurons revealed a mean value of 18.9±0.7 pA.In the presence of ATP, this value amounted to 18.8±0.7 pAand was not significantly different from the control (Fig. 6B).Regarding the median values of the mEPSC decay timeconstant, the exemplary cell illustrated in Fig. 6C displayedsimilar values before, during, and after ATP application (2.3,2.4, and 2.5 ms, respectively). In accordance with this, thewhole population (41 neurons) showed no significant differ-ence of the mean mEPSC decay time constant without andwith ATP (2.4±0.1 and 2.5±0.1 ms, respectively; Fig. 6D).Together, the data demonstrated no ATP effects on either thepeak amplitude or the decay time constant of mEPSCs,
Fig. 6 – ATP does not influence peak amplitudes and decay time constants of mEPSCs. (A) Cumulative fraction plot for mEPSCpeak amplitudes before (control), during (ATP), and after (recovery) ATP application recorded froma P3 bipolar neuron located inthe central limb. The median values (50%) are illustrated by triangles and were very similar under the three conditions. (B)Histogram ofmeanmEPSC peak amplitudes obtained from themedian values (see panel A) of 41 cells. There was no significantATP effect. (C) Cumulative fraction plot for mEPSC decay time constants before, during, and after ATP application for the cell inpanel A, illustrating similarmedian values under the three conditions. (D) MeanmEPSC decay time constants in the presence orabsence of ATP were not significantly different (41 cells).
27B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
implying that there is no influence on the spontaneousactivity mediated at postsynaptic sites.
2.1.5. Heterogeneous ATP effects on mEPSC frequencyIn a next series of experiments, we analyzed ATP effects onmEPSC frequency to test whether ATP may modulate thespontaneous activity at presynaptic sites. In 39% of the LSOneurons (16 of 41; for cell-type-specific effects see Table 1),ATP had no significant effect on mEPSC frequency (data notshown). In the remaining 25 neurons (61%), the mEPSCfrequency was significantly affected by ATP, yet the effectswere heterogeneous. In 9 neurons (22%), the frequency wasdecreased (Table 1). In the exemplary cell shown in Fig. 7Aand B, the mean mEPSC frequency decreased significantly
from 1.7±0.1 to 1.4±0.1 Hz in the presence of ATP (p=0.01,Mann and Whitney's U-test). The mean mEPSC frequency ofall 9 neurons decreased significantly to 62±6% of the control(Fig. 7C; p=1.0 × 10-4, Student's paired t-test). This ATP effectwas almost fully reversible (99±12% of control).
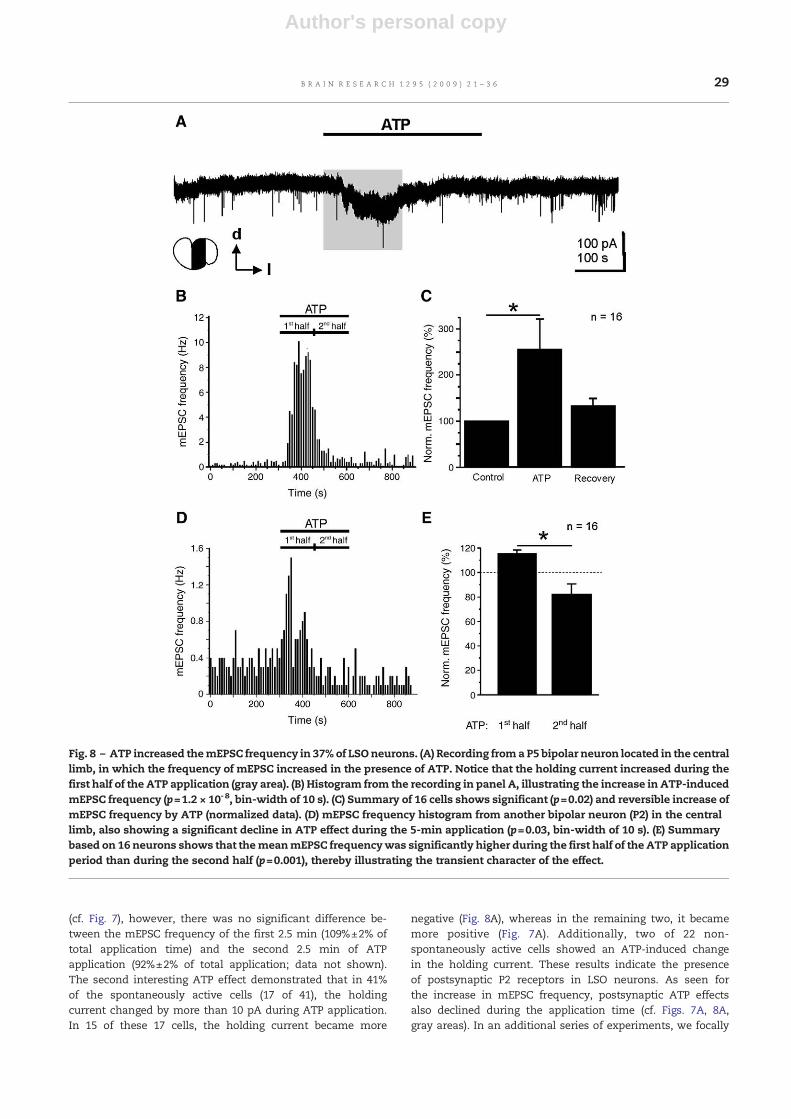
In another group comprising 16 neurons (39%), the mEPSCfrequency was significantly increased by ATP (Fig. 8 andTable 1). In the cell illustrated in Figs. 8A and B, the meanmEPSC frequency increased considerably from 0.3±0.03 to3.1±0.6 Hz during ATP application (p=1.2 × 10-8, Mann andWhitney's U-test). On average, the mEPSC frequency in-creased to 256%±62% of control (Fig. 8C; p=0.02, Student'spaired t-test). This ATP-induced increase was almost fullyreversible (129±16% of control).
Aside from the ATP-induced augmentation and depressionof mEPSC frequency, we observed two further interesting ATPeffects. First, the ATP-induced increase in mEPSC frequencydeclined during the 5-min ATP application (Figs. 8B and D).Within a total of 16 neurons, the mean mEPSC frequency wassignificantly higher during the first half of ATP applicationthan during the second half (116%±2% vs. 82%±4% of totalapplication time; p=0.001, Student's paired t-test; notice that100% represents the average frequency during the totalapplication time), demonstrating the transient nature of theATP effect on the mEPSC frequency (Fig. 8E). Regarding the9 cells with an ATP-induced decrease in mEPSC frequency
Table 1 – Cell-type-specific effects of ATP on mEPSCfrequency. Values represent the number of cells testedwith ATP.
Cell type mEPSC frequency Total
Increase Decrease None
Bipolar cells 6 6 8 20Multipolar cells 3 2 2 7Nondefined cells 7 1 6 14Total 16 9 16 41
Fig. 7 – ATP decreased the mEPSC frequency in 21% of LSO neurons. (A) Recording from a P4 bipolar neuron located in themedial limb, in which themEPSC frequency decreased in the presence of ATP. Notice that the holding current decreased duringthe first half of the ATP application (gray area). (B) Histogram from the recording shown in panel A, illustrating the ATP-induceddecrease in mEPSC frequency (p=0.01; bin-width of 10 s). (C) Summary based on 9 cells shows significant (p=1.0 × 10-4) andreversible decrease of mEPSC frequency by ATP (normalized data).
28 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
Author's personal copy
(cf. Fig. 7), however, there was no significant difference be-tween the mEPSC frequency of the first 2.5 min (109%±2% oftotal application time) and the second 2.5 min of ATPapplication (92%±2% of total application; data not shown).The second interesting ATP effect demonstrated that in 41%of the spontaneously active cells (17 of 41), the holdingcurrent changed by more than 10 pA during ATP application.In 15 of these 17 cells, the holding current became more
negative (Fig. 8A), whereas in the remaining two, it becamemore positive (Fig. 7A). Additionally, two of 22 non-spontaneously active cells showed an ATP-induced changein the holding current. These results indicate the presenceof postsynaptic P2 receptors in LSO neurons. As seen forthe increase in mEPSC frequency, postsynaptic ATP effectsalso declined during the application time (cf. Figs. 7A, 8A,gray areas). In an additional series of experiments, we focally
Fig. 8 – ATP increased themEPSC frequency in 37% of LSOneurons. (A) Recording froma P5 bipolar neuron located in the centrallimb, in which the frequency of mEPSC increased in the presence of ATP. Notice that the holding current increased during thefirst half of the ATP application (gray area). (B) Histogram from the recording in panel A, illustrating the increase in ATP-inducedmEPSC frequency (p=1.2 × 10-8, bin-width of 10 s). (C) Summary of 16 cells shows significant (p=0.02) and reversible increase ofmEPSC frequency by ATP (normalized data). (D) mEPSC frequency histogram from another bipolar neuron (P2) in the centrallimb, also showing a significant decline in ATP effect during the 5-min application (p=0.03, bin-width of 10 s). (E) Summarybased on 16 neurons shows that themeanmEPSC frequencywas significantly higher during the first half of the ATP applicationperiod than during the second half (p=0.001), thereby illustrating the transient character of the effect.
29B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
Author's personal copy
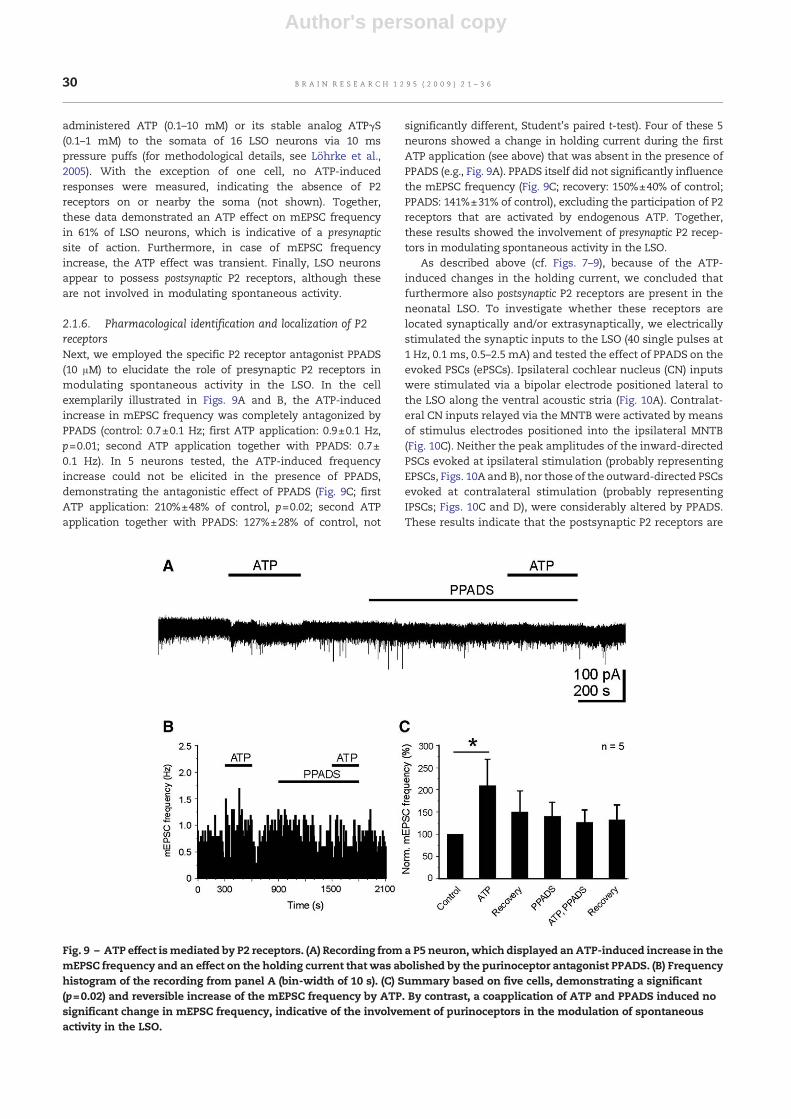
administered ATP (0.1–10 mM) or its stable analog ATPγS(0.1–1 mM) to the somata of 16 LSO neurons via 10 mspressure puffs (for methodological details, see Löhrke et al.,2005). With the exception of one cell, no ATP-inducedresponses were measured, indicating the absence of P2receptors on or nearby the soma (not shown). Together,these data demonstrated an ATP effect on mEPSC frequencyin 61% of LSO neurons, which is indicative of a presynapticsite of action. Furthermore, in case of mEPSC frequencyincrease, the ATP effect was transient. Finally, LSO neuronsappear to possess postsynaptic P2 receptors, although theseare not involved in modulating spontaneous activity.
2.1.6. Pharmacological identification and localization of P2receptorsNext, we employed the specific P2 receptor antagonist PPADS(10 μM) to elucidate the role of presynaptic P2 receptors inmodulating spontaneous activity in the LSO. In the cellexemplarily illustrated in Figs. 9A and B, the ATP-inducedincrease in mEPSC frequency was completely antagonized byPPADS (control: 0.7±0.1 Hz; first ATP application: 0.9±0.1 Hz,p=0.01; second ATP application together with PPADS: 0.7±0.1 Hz). In 5 neurons tested, the ATP-induced frequencyincrease could not be elicited in the presence of PPADS,demonstrating the antagonistic effect of PPADS (Fig. 9C; firstATP application: 210%±48% of control, p=0.02; second ATPapplication together with PPADS: 127%±28% of control, not
significantly different, Student's paired t-test). Four of these 5neurons showed a change in holding current during the firstATP application (see above) that was absent in the presence ofPPADS (e.g., Fig. 9A). PPADS itself did not significantly influencethe mEPSC frequency (Fig. 9C; recovery: 150%±40% of control;PPADS: 141%±31% of control), excluding the participation of P2receptors that are activated by endogenous ATP. Together,these results showed the involvement of presynaptic P2 recep-tors in modulating spontaneous activity in the LSO.
As described above (cf. Figs. 7–9), because of the ATP-induced changes in the holding current, we concluded thatfurthermore also postsynaptic P2 receptors are present in theneonatal LSO. To investigate whether these receptors arelocated synaptically and/or extrasynaptically, we electricallystimulated the synaptic inputs to the LSO (40 single pulses at1 Hz, 0.1 ms, 0.5–2.5 mA) and tested the effect of PPADS on theevoked PSCs (ePSCs). Ipsilateral cochlear nucleus (CN) inputswere stimulated via a bipolar electrode positioned lateral tothe LSO along the ventral acoustic stria (Fig. 10A). Contralat-eral CN inputs relayed via the MNTB were activated by meansof stimulus electrodes positioned into the ipsilateral MNTB(Fig. 10C). Neither the peak amplitudes of the inward-directedPSCs evoked at ipsilateral stimulation (probably representingEPSCs, Figs. 10A and B), nor those of the outward-directed PSCsevoked at contralateral stimulation (probably representingIPSCs; Figs. 10C and D), were considerably altered by PPADS.These results indicate that the postsynaptic P2 receptors are
Fig. 9 – ATP effect ismediated by P2 receptors. (A) Recording froma P5 neuron, which displayed an ATP-induced increase in themEPSC frequency and an effect on the holding current that was abolished by the purinoceptor antagonist PPADS. (B) Frequencyhistogram of the recording from panel A (bin-width of 10 s). (C) Summary based on five cells, demonstrating a significant(p=0.02) and reversible increase of the mEPSC frequency by ATP. By contrast, a coapplication of ATP and PPADS induced nosignificant change in mEPSC frequency, indicative of the involvement of purinoceptors in the modulation of spontaneousactivity in the LSO.
30 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
unlikely to be located synaptically in the CN-LSO projections,but rather extrasynaptically and/or at other input synapses tothe LSO.
2.2. Spontaneous Synaptic Activity in LSO Neurons ofCaV1.3-/- Mice
The results presented so far demonstrated that mEPSCs showregion-specific and cell-type-specific variations in the neona-tal LSO, indicating that these events should not be consideredasmerely random activity with little, if any, relevance. Rather,they could be involved in the development of synapticconnectivity and could thereby act in conjunction with theaction potential-dependent inputs. We reasoned that areduction of the latter inputs could affect the properties ofminiature events, possibly in a compensatory manner (cf.Hartman et al., 2006; Leao et al., 2004, 2005). To address thisissue, we comparedmEPSCs from LSO neurons of normalmicewith those from CaV1.3-/- mice, which display a lack oftransmitter release from cochlear inner hair cells to auditorynerve fibers. The auditory brainstem structures in CaV1.3-/-
mice are thus deprived of cochlea-driven action potentialactivity and auditory brainstem responses are absent (Platzeret al., 2000).
Fig. 10 – Peak amplitudes of evoked EPSCs and IPSCs are not altered by PPADS. (A) Series of 40 EPSCs (gray traces) evoked byrepetitive electrical stimulation (1 Hz) of the ipsilateral LSO inputs (asterisk in inset marks position of the stimulus electrode)before (control) and in the presence of PPADS. The peak amplitudes of the mean evoked inward-directed currents (solid traces)differed onlymarginally (control: −62 pA; PPADS: −74 pA). (B) Summary of the PPADS effect on eEPSC peak amplitudes obtainedfrom 5 cells (recordings from cell 3 are shown in panel A). (C) IPSCs in response to repetitive stimulation (40 times at 1 Hz) of thecontralateral CN inputs to the LSO (asterisk in inset marks position of the stimulus electrode). Under control conditions, themean peak amplitude of the evoked outward-directed currents (solid traces) amounted to 82 pA and was only marginallyreduced by PPADS (80 pA). (D) Summary of the PPADS effect on eIPSC peak amplitude obtained from five neurons (recordingsfrom cell 3 are shown in panel C). Stimulus artifacts are truncated for clarity (parallel bars in panels A and C).
Fig. 11 – Frequency and peak amplitude of mEPSCs fromLSO neurons are significantly increased in P3-4 CaV1.3-/-
mice. (A) In CaV1.3-/- mice, the mean mEPSC frequency wassignificantly higher than in age-matched WT counterparts(p=0.04). (B) The mean mEPSC peak amplitude was alsosignificantly larger in the Cav1.3-/- than in the WT animals(p=0.03). Numbers in brackets correspond to number ofcells.
31B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
We analyzed animals at P3–4, an age at which inner haircells are able to generate spontaneous action potentials(Marcotti et al., 2003). CaV1.3-/- animals displayed a significantlyhigher mean mEPSC frequency than WT animals (4.0±1.4 Hz,n=7, compared to 0.2±0.04 Hz, n=6; p=0.04, Student's unpairedt-test; Fig. 11A). In addition, their mean mEPSC peak amplitudewas significantly higher than in the WT animals (20.7±1.9 pA,compared to 14.8±1.2 pA; p=0.03, Student's unpaired t-test;Fig. 11B). These results demonstrate that mEPSCs are signifi-cantly altered in neonatal LSO neurons of CaV1.3-/- mice,showing a compensatory reaction of action potential-indepen-dent activity in response to peripheral deprivation.
3. Discussion
In this study, we have analyzed spontaneous mEPSC activity inthe LSO of rats and mice during early postnatal life, whensynapse maturation takes place without hearing experience.Rather than occurring in a stochasticmanner, merely reflectinga leaky release mechanism, these miniature events are region-specific and cell-type-specific. This is in accord with the ideathat the miniature events may be of some particular functionalrelevance. Furthermore, the neuromodulator ATP modulatesthe miniature events in several heterogeneous ways, involvingpresynaptic P2 purinoceptors. Finally, data obtained fromCaV1.3-/- mice show compensatory effects in the LSO todeprivation of peripheral activation, corroborating the ideathat the mEPSCs represent functionally relevant activity.
3.1. Morphological and regional differences ofspontaneously active LSO cells
Seven morphological cell types have been identified in the ratLSO (Rietzel and Friauf, 1998). Bipolar cells represent the mainpopulation, followed by multipolar cells. Only these two celltypes were identified and analyzed in the present study.Interestingly, 89% of the bipolar cells showed spontaneousactivity, yet only 41% of the multipolar cells. Regarding thetemporal pattern of spontaneous activity, no difference wasobserved between the two types. Both showed continuallymEPSCs with mainly short intervals; burst-like activity wasnot present. The higher percentage of bipolar cells withmEPSCs compared to multipolar cells may reflect differencesin the maturation of the two cell types. Indeed, the number ofprimary dendrites declines in bipolar cells, but not inmultipolar cells between P4 and P36 (Rietzel and Friauf,1998). This anatomical study further showed that much ofthe dendritic reorganization takes place at P4–15, thus beingmainly independent of acoustic input (hearing onset in ratsoccurs at around P12; Blatchley et al., 1987). The regional andcell-type-specific occurrence of miniature events during thefirst postnatal week is consistent with the idea that theycontribute to synapse maturation in a particular manner attimes when there is no sound-evoked activity. In accordancewith this, a higher percentage of bipolar cells, which displaydendritic pruning, receive mEPSCs compared to multipolarcells which do not show pruning. In cultured hippocampalneurons, dendritic protein synthesis is inhibited by mEPSCs(Sutton et al., 2004). If the same holds for LSO neurons, one can
assume that the reduction of dendritic arborization isaccompanied by a reduction of dendritic protein synthesis.
We observed regional differences in the occurrence ofmEPSCs within the LSO: the percentage of spontaneouslyactive cells declined from the medial limb to the lateral limb.The regional differences in spontaneous activity can beexplained by the facts that mostly bipolar cells are spontane-ously active (present study) and that this cell type does notexist in the lateral limb (Rietzel and Friauf, 1998). Thus,distribution differences between specific cell types, ratherthan differences in maturation (the dorsomedial, high-frequency region matures earlier than the ventrolateral,low-frequency region), are likely to produce the regionaldifferences in mEPSC occurrence.
3.2. Heterogeneous effects of ATP in the LSO
In the adult rat superior olivary complex, P2 receptor subunitshave been demonstrated by serial analysis of gene expres-sion, in situ hybridization, and immunohistochemistry (Koehlet al., 2004; Yao et al., 2000). P2 receptors can be separatedinto two major types, namely, ionotropic P2X receptors andmetabotropic P2Y receptors (reviews: Burnstock, 2006; Khakh,2001; Lechner and Boehm, 2004; North, 2002). They areactivated by extracellular ATP, which can act as a neuro-transmitter as well as a neuromodulator. NeuromodulatingATP effects were reported in spinal cord neurons (Gu andMacDermott, 1997; Hugel and Schlichter, 2000), cerebellarPurkinje cells (Brockhaus et al., 2004), and hippocampalpyramidal neurons (Mendoza-Fernandez et al., 2000; Zhanget al., 2003). Recently, Watano and colleagues (2004) showedan ATP-induced increase of excitatory and inhibitory inputsin MNTB principal cells that was mediated through P2Xreceptors.
We observed two different ATP effects in the LSO: (1) 41% ofspontaneously active cells showed a change in holdingcurrent; (2) in 61% of spontaneously active cells, the frequencyof mEPSCs was altered, but neither their amplitude nor theirdecay time constant was changed. The first effect implies thepresence of postsynaptic purinoceptors, similar to the situa-tion in spinal cord dorsal horn neurons (Hugel and Schlichter,2000) and hypocretin/orexin neurons of the hypothalamus(Wollmann et al., 2005). In the vast majority of LSO neurons,the holding current became more negative, suggestive of theactivation of inward-directed, depolarizing cation conduc-tances via functional P2X receptors. Because the ATP effectson the holding currents were observed mostly with bathapplication, and only once if ATP or ATPγS was focally appliedonto the cell somata, we suggest that P2X receptors are locatedin distal processes of the LSO neurons. However, our experi-ments further show that these receptors are neither present inthe ipsilateral CN-LSO projection nor in the contralateral CN-LSO projection. Thus, postsynaptic purinoceptors in LSOneurons appear to be located extrasynaptically or to be partof other afferents (e.g., originating from the lateral nucleus ofthe trapezoid body or the ventral nucleus of the trapezoidbody; see Thompson and Schofield, 2000).
The second ATP effect (on mEPSC frequency) is suggestiveof the presence of presynaptic purinoceptors in the LSO. Abouttwo-thirds of the LSO cells showed an increase in mEPSC
32 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
frequency by ATP, whereas in one-third, ATP decreased themEPSC frequency. Moreover, in the bipolar cells, 50% showedno ATP effect on the mEPSC frequency, whereas 25% showedan augmentation and another 25% a depression. These resultsdemonstrate the heterogeneity of ATP effects in the LSO, evenwithin one cell type. Heterogeneous ATP effects were alsofound in the rat MNTB (Watano et al., 2004). In this nucleus, noATP-induced depression of spontaneous activity was found.This is in contrast to the situation in the LSO and indicatesnucleus-specific neuromodulatory roles of ATP in the auditorybrainstem. It is possible that different P2 receptor types, and/or P2 receptors containing various subunits, are present in theMNTB and the LSO. In the MNTB, P2X1–6 receptor subunitswere revealed by immunohistochemistry (Yao et al., 2000).Moreover, pharmacologically identified P2X1 and P2X3 recep-tor subunits in MNTB neurons were responsible for theaugmentation of inhibitory and excitatory postsynaptic cur-rents, respectively (Watano et al., 2004).
The composition of the P2 receptors modulating thespontaneous activity in the LSO is unknown. Concerning theaugmentation of mEPSCs frequency, ionotropic P2X receptorsmay be involved, like in the MNTB (see above), the spinalcord (Gu and MacDermott, 1997; Nakatsuka and Gu, 2001;Nakatsuka et al., 2003), or the hippocampus (Rodrigues et al.,2005). They may be composed of P2X4 and P2X6 subunits,which have been described in the LSO (Koehl et al., 2004). Onthe other hand, the reduction of mEPSC frequency may bemediated throughmetabotropic P2Y receptors, known to havepresynaptic effects on glutamate release in hippocampalneurons (Mendoza-Fernandez et al., 2000; Zhang et al., 2003;Rodrigues et al., 2005). Regarding the observed transient effectof ATP on the increase in mEPSC frequency, and the decline ofATP-induced changes in holding current, there are twopossibilities (cf. Wollmann et al., 2005). The transient ATPeffects may be due to the desensitization of P2X receptors(Vial et al., 2004) or to the reduction in extracellular ATP byectonucleotidases (Dunwiddie et al., 1997).
3.3. Functional considerations
The results of our recordings indicate a continuous, asyn-chronous release of glutamate in the developing auditorybrainstem before the onset of hearing. Although this release isnot elicited by spike-driven activity and may thus reflect lowlevels of excitatory synaptic input, possibly triggered bypresynaptic store-operated calcium channels (Emptage et al.,2001; Llano et al., 2000), it may be functionally related tonormal development. Evidence for this conclusion is providedby the fact that the mEPSCs demonstrate specific character-istics, rather than being stochastic. Miniature synaptic eventsare involved in the clustering of glutamate receptors atneuromuscular junctions (Saitoe et al., 2001) and sufficientto maintain dendritic spines in hippocampal pyramidal cells(McKinney et al., 1999). The functional significance of minia-ture synaptic activity in central auditory neurons was recentlyinvestigated by Lu and coworkers (2007), who found develop-mental changes of mEPSCs in bushy cells of the mouseanteroventral cochlear nucleus across a critical period priorto hearing onset. Furthermore, after cochlea ablation, theauthors observed the emergence of mEPSCs impinging on
dendrites of surviving bushy cells. Altogether, these studiesdemonstrate the attendance of miniature synaptic activity indevelopmental processes. The present study provides furtherhints. In CaV1.3-/- mice lacking cochlea-driven nerve activity,we found a significantly increased mEPSC frequency at P3–4compared toWTmice. Onemay argue that the effect is causedlocally by the lack of CaV1.3 channels in the LSO and not due tomissing input originating from the cochlea. In fact, CaV1.3channels are expressed in the rat auditory brainstem. But inthe developing LSO, their contribution to voltage-gated Ca2+
influx amounts to less than 20% of the whole amplitude (BrittaMüller, personal communication), making it unlikely that theeffect is caused locally. Results of a preliminary anatomicalstudy show a drastic shrinkage of the LSO in CaV1.3-/- mice(Müller et al., 2007). The shrinkage may be explained by areduced protein synthesis. As miniature synaptic events caninhibit dendritic protein synthesis (Sutton et al., 2004), theincreased mEPSC frequency in CaV1.3-/- mice may be causallyrelated to the shrinkage of the LSO.
In conclusion, to our knowledge, we provide the firstanalysis of spontaneous mEPSC activity in LSO neurons inprehearing animals at times when synapses mature. Wealso show the heterogeneous modulation of the mEPSCs byATP via P2 purinoceptors. Because of their regional andcell-type-specific nature, the mEPSCs are likely of somefunctional significance during the period when the neuro-nal auditory brainstem circuits develop. It will be veryinteresting to see whether mIPSCs also display specific,rather than stochastic, features in the developing auditorybrainstem.
4. Experimental procedures
All protocols were approved by the animal care and usecommittees responsible for our institution (Landesuntersu-chungsamt Rheinland-Pfalz, Germany). Treatment of theanimals was in accordance with the German law for conduct-ing animal experiments and also followed the NIH guide forthe care and use of laboratory animals. Both the number ofanimals and their suffering were reduced to the possibleminimum.
4.1. Tissue preparation
Brainstem slices were prepared from Sprague-Dawley rats (normalrats), C57BL/6N wild-type mice (WT mice), and CaV1.3 knockoutmicewith C57BL/6N background (CaV1.3-/-mice; Platzer et al., 2000).Pups of both genderswere used between postnatal day (P) 1 and P6(the day of birth was taken as P0). The animals were bred andhoused in our facility. They were deeply anesthetized with anoverdose of ketamine (1 mg g–1 body weight intraperitoneal),decapitated, and their brainwasdissected.Coronal brainstemslices(300 μm thick) containing the LSO were cut in ice-cold preparationsolution (for composition, see below) with a vibrating microtome(VT-1000, Leica, Bensheim, Germany), and transferred into achamber filled with carboxygenated (95% O2, 5% CO2) extracellularsolution (for composition, seebelow).After 60minat approximately34 °C, the slices were stored at room temperature (20–23 °C) for atleast 30min, after which recordings begun.
33B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
4.2. Electrophysiology
Brainstem slices were transferred to the recording chamber(volume: 2 mL) and continuously superfused with extracellu-lar solution (rate: 3–4 mL min-1). The chamber was mountedon an upright microscope (Eclipse E600FN, Nikon, Tokyo,Japan) equipped with differential interference contrast optics(Nikon objectives: 4× CFI Achromat, 0.1 NA; 60× CFI Fluor W,1.0 NA) and an infrared video camera system (CCD cameraC5405-01, Hamamatsu, Herrsching, Germany; or CCD cameraVX44, PCO computer optics, Germany; PC frame grabber card,pciGrabber-4plus, PHYTEK, Mainz, Germany). Patch pipetteswere pulled from borosilicate glass capillaries (GB150-8P,Science Products, Hofheim, Germany) with a vertical puller(PP-83, Narishige, Tokyo, Japan). They had resistances of 5–8 MΩ when filled with intracellular solution (for composition,see below) and were connected via an Ag/AgCl wire to anAxopatch-1D patch-clamp amplifier (Molecular Devices,Union City, CA, USA) or an EPC 9 patch-clamp amplifier(HEKA Elektronik, Lambrecht, Germany). Because of the largedifferences in chloride concentration between intracellularand extracellular solution (for composition, see below), theliquid junction potential amounted to 16.7 mV (calculatedafter Barry, 1994) and was corrected either online (record-ings with EPC 9) or offline (recordings with Axopatch-1D).Signals were digitized, low-pass filtered, and sampled at2.0–2.9 kHz.
The presence of spontaneous activity was monitored withwhole-cell voltage clamp recordings at a holding potential of−70 mV lasting between 5 and 60 min. Events with peakamplitudes≥8–20 pA (depending on background noise) and afrequency≥0.1 Hz within a 5-min period were defined assPSCs. Cells showing such events were designated as sponta-neously active; those without such events were designated asnon-spontaneously active. Peak amplitude, decay time con-stant, and frequency of sPSCswere analyzedwith the softwareMiniAnalysis (Synaptosoft, Leonia, NJ, USA). Decay timeconstant was determined as the period between the peakamplitude and 37% of the peak amplitude. Drugs were bath-applied by switching between different inflow channels. Atthe single-cell level, drug effects on sPSC parameters weredetermined by comparing values from 5-min periods before,during, and after drug application. For frequency analysis,sPSC counts were obtained from successive 10-s periods, andmean frequencies per 5-min period (average of thirty 10-speriods) were calculated. For peak amplitude and decay timeconstant, mean values were determined from 5-min periods.Data were statistically analyzed with the software WinSTATfür Excel 1999.3 (Fitch Software, Zierenberg, Germany) and aregiven as mean±standard error of the mean. In the figures,statistical differences between data groups are indicated byasterisks (⁎p<0.05; ⁎⁎p<0.01; and ⁎⁎⁎p<0.001).
4.3. Cell fillings and morphological analysis
LSO neurons were filled with the fluorescent dye Lucifer yellow(0.01% w/v in intracellular solution) via diffusion during theelectrophysiological measurements. After recordings, sliceswere incubated for 2 h in extracellular solution at 4 °C indarkness. Thereafter, they were fixated with 4% (w/v) parafor-
maldehyde in 0.1 M phosphate buffer (pH 7.4) for 48 h at 4 °C indarkness. Fixated slices were mounted on slides, embedded inVectashield HardSet (Vector Laboratories, Burlingame, CA, USA),and protected with coverslips.
Dye-filled cells were visualized with an inverted micro-scope (Axiovert 200M, Zeiss, Göttingen, Germany), equippedwith epifluorescence (mercury arc lamp FluoArc HBO 100;filter settings, excitation 450–490 nm, beam splitter 510 nm,emission 520 nm, Zeiss) and ‘grid projection’ technology(ApoTome, Zeiss). Between 5 and 48 optical sections (0.5 μmdistance) were used for 3D reconstructions (AxioVision 4.3,Zeiss) of the LSO neurons. They were identified as bipolar ormultipolar cells based on criteria described previously(Rietzel and Friauf, 1998).
4.4. Solutions and drugs
The preparation solution consisted of (in [mM]) NaHCO3, 25;NaH2PO4, 1.25; KCl, 2.5; MgCl2, 1; CaCl2, 2; D-glucose, 260; Na-pyruvate, 2; myo-inositol, 3; and kynurenic acid, 1; pH 7.4 whenbubbled with 95% O2–5% CO2. The extracellular solution had thefollowing composition (in [mM]): NaCl, 125; NaHCO3, 25; NaH2PO4,1.25; KCl, 2.5; MgCl2, 1; CaCl2, 2; D-glucose, 10; Na-pyruvate, 2;myo-inositol, 3; ascorbic acid, 0.4; pH 7.4. The intracellular solutionscontained (in [mM]) potassium gluconate, 135; EGTA, 1; MgCl2, 2;HEPES, 10; CaCl2, 0.1; Na2ATP, 2; pH 7.3 with KOH. The followingdrugs were dissolved in extracellular solution (final concentration;supplier): DL-2-amino-5-phosphonopentanic acid (AP5; 30 μM;Sigma-Aldrich, Steinheim, Germany), adenosine 5′-triphosphate(ATP; 100 μM; Tocris, Bristol, UK), 6-cyano-7-nitro-quinoxaline-2,3-dione (CNQX; 20 μM; Sigma-Aldrich), pyridoxal phosphate-6-azo-phenyl-2′-4′-disolfonic acid (PPADS; 10 μM; Sigma-Aldrich), andtetrodotoxin (TTX; 0.5 μM; Tocris).
Acknowledgments
This work was supported in part by a grant from the EuropeanCommunity (Cavnet, MRTN-CT-206 -035367 to E.F). We greatlyacknowledge the gift of CaV1.3-/- mice by Jörg Striessnig(Innsbruck, Austria). We further thank Sascha Ehrhardt forexcellent technical support and Drs. Ekkehard Neuhaus andTorsten Möhlmann for providing the ApoTome and introduc-ing us to it.
R E F E R E N C E S
Barry, P.H., 1994. JPCalc, a software for calculating liquid junctionpotential corrections in patch-clamp, intracellular, epithelialand bilayer measurements and for correcting junctionpotential measurements. J. Neurosci. Meth. 51, 107–116.
Beutner, D., Moser, T., 2001. The presynaptic function of mousecochlear inner hair cells during development of hearing.J. Neurosci. 21, 4593–4599.
Blatchley, B.J., Cooper, W.A., Coleman, J.R., 1987. Development ofauditory brainstem response to tone pip stimuli in the rat. Dev.Brain Res. 32, 75–84.
Brandt, A., Striessnig, J., Moser, T., 2003. CaV1.3 channels areessential for development and presynaptic activity of cochlearinner hair cells. J. Neurosci. 23, 10832–10840.
34 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
Brockhaus, J., Dressel, D., Herold, S., Deitmer, J.W., 2004. Purinergicmodulation of synaptic input to Purkinje neurons in ratcerebellar brain slices. Eur. J. Neurosci. 19, 2221–2230.
Burnstock, G., 2006. Purinergic signalling. Brit. J. Pharmacol. 147,S172–S181.
Dunwiddie, T.V., Diao, L., Proctor, W.R., 1997. Adenine nucleotidesundergo rapid, quantitative conversion to adenosine in theextracellular space in rat hippocampus. J. Neurosci. 17,7673–7682.
Ehrlich, I., Ilic, V., Lohmann, C., Friauf, E., 1998. Development ofglycinergic transmission in organotypic cultures from auditorybrain stem. Neuroreport 9, 2785–2790.
Emptage, N.J., Reid, C.A., Fine, A., 2001. Calcium stores inhippocampal synaptic boutons mediate short-term plasticity,store-operated Ca2+ entry, and spontaneous transmitterrelease. Neuron 29, 197–208.
Erazo-Fischer, E., Striessnig, J., Taschenberger, H., 2007. The role ofphysiological afferent nerve activity during in vivo maturationof the calyx of Held synapse. J. Neurosci. 14, 1725–1737.
Friauf, E., Lohmann, C., 1999. Development of auditory brainstemcircuitry. Cell Tiss. Res. 297, 187–195.
Gu, J.G., MacDermott, A.B., 1997. Activation of ATP P2X receptorselicits glutamate release from sensory neuron synapses.Nature 389, 749–753.
Hartman, K.N., Pal, S.K., Burrone, J., Murthy, V.N., 2006.Activity-dependent regulation of inhibitory synaptictransmission in hippocampal neurons. Nat. Neurosci. 9,642–649.
Hugel, S., Schlichter, R., 2000. Presynaptic P2X receptors facilitateinhibitory GABAergic transmission between cultured rat spinalcord dorsal horn neurons. J. Neurosci. 15, 2121–2130.
Kandler, K., 2004. Activity-dependent organization of inhibitorycircuits: lessons from the auditory system. Curr. Opin.Neurobiol. 14, 96–104.
Khakh, B.S., 2001. Molecular physiology of P2X receptors and ATPsignalling at synapses. Nat. Rev. Neurosci. 2, 165–174.
Koehl, A., Schmidt, N., Rieger, A., Pilgram, S.A., Letunic, I., Bork, P.,Soto, F., Friauf, E., Nothwang, H.G., 2004. Gene expressionprofiling of the rat superior olivary complex using serialanalysis of gene expression. Eur. J. Neurosci. 12, 3244–3258.
Kotak, V.C., Sanes, D.H., 1995. Synaptically evoked prolongeddepolarizations in the developing auditory systemJ. Neurophysiol. 74, 1611–1620.
Kros, C.J., Ruppersberg, J.P., Rüsch, A., 1998. Expression of apotassium current in inner hair cells during development ofhearing in mice. Nature 394, 281–284.
Leao, R.N., Oleskevich, S., Sun, H., Bautista, M., Fyffe, R.E.W.,Walmsley, B., 2004. Differences in glycinergic mIPSCs in theauditory brain stem of normal and congenitally deaf neonatalmice. J. Neurophysiol. 91, 1006–1012.
Leao, R.N., Leao, F.N., Walmsley, B., 2005. Non-random nature ofspontaneous mIPSCs in mouse auditory brainstem neuronsrevealed by recurrence quantification analysis. Proc. R Soc. B272, 2551–2559.
Leao, R.N., Sun, H., Svahn, K., Berntson, A., Youssoufian, M.,Paolini, A.G., Fyffe, R.E.W., Walmsley, B., 2006. Topographicorganization in the auditory brainstem of juvenile mice isdisrupted in congenital deafness. J. Physiol. 571, 563–578.
Lechner, S.G., Boehm, S., 2004. Regulation of neuronal ion channelvia P2Y receptors. Purin. Signal 1, 31–41.
Lippe, W.R., 1994. Rhythmic spontaneous activity in the developingavian auditory system. J. Neurosci. 14, 1486–1495.
Llano, I., González, J., Caputo, C., Lai, F.A., Blayney, L.M., Tan,Y.P., Marty, A., 2000. Presynaptic calcium stores underlielarge-amplitude miniature IPSCs and spontaneous calciumtransients. Nature Neurosci. 3, 1256–1265.
Lohmann, C., Ilic, V., Friauf, E., 1998. Development of atopographically organized auditory network in slice cultureis calcium dependent. J. Neurobiol. 34, 97–112.
Löhrke, S., Kungel, M., Friauf, E., 1998. Electrical membraneproperties of trapezoid body neurons in the rat auditory brainstem are preserved in organotypic slice cultures. J. Neurobiol.36, 395–409.
Löhrke, S., Srinivasan, G., Oberhofer, M., Doncheva, E., Friauf, E.,2005. Shift from depolarizing to hyperpolarizing glycine actionoccurs at different perinatal ages in superior olivary complexnuclei. Eur. J. Neurosci. 22, 2708–2722.
Lu, Y., Harris, J.A., Rubel, E.W., 2007. Development of spontaneousminiature EPSCs in mouse AVCN neurons during a criticalperiod of afferent-dependent neuron survival. J. Neurophysiol.97, 635–646.
Marcotti, W., Johnson, S.L., Rüsch, A., Kros, C.J., 2003. Sodium andcalcium currents shape action potentials in immature mouseinner hair cells. J. Physiol. 552, 743–761.
McKinney, R.A., Capogna, M., Dürr, R., Gähwiler, B.H., Thompson,S.M., 1999. Miniature synaptic events maintain dendriticspines via AMPA receptor activation. Nature Neurosci. 2,44–49.
Mendoza-Fernandez, V., Andrew, R.D., Barajas-López, C., 2000.ATP inhibits glutamate synaptic release by acting at P2Yreceptors in pyramidal neurons of hippocampal slices. J.Pharmacol. Exp. Ther. 293, 172–179.
Müller, B., Friauf, E., Löhrke, S., 2007. CaV1.3-/- mice show adisturbed development in the lateral superior olive of theauditory brainstem. In: Kriegelstein, K., Hoffmann, K.-P. (Eds.),Göttingen Neurobiology Report 2007. Proceedings of the 7thMeeting of the German Neuroscience Society 2007, 31stGöttingen Neurobiology Conference. Georg Thieme Verlag,Stuttgart, p T19-9B.
Nakatsuka, T., Gu, J.G., 2001. ATP P2X receptor-mediatedenhancement of glutamate release and evoked EPSCs in dorsalhorn neurons of the rat spinal cord. J. Neurosci. 21, 6522–6531.
Nakatsuka, T., Tsuzuki, K., Ling, J.X., Sonobe, H., Gu, J.G., 2003.Distinct roles of P2X receptors in modulating glutamaterelease at different primary sensory synapses in rat spinalcord. J. Neurophysiol. 89, 3243–3252.
North, R.A., 2002. Molecular physiology of P2X receptors. Physiol.Rev. 82, 1013–1067.
Otsu, Y., Murphy, T.H., 2003. Miniature transmitter release:accident of nature or careful design? Sci. STKE pe54.
Platzer, J., Engel, J., Schrott-Fischer, A., Stephan, K., Bova, S., Chen,H., Zheng, H., Striessnig, J., 2000. Congenital deafness andsinoatrial node dysfunction in mice lacking class D L-type Ca2+
channels. Cell 102, 89–97.Rietzel, H.-J., Friauf, E., 1998. Neuron types in the rat lateral
superior olive and developmental changes in the complexity oftheir dendritic arbors. J. Comp. Neurol. 390, 20–40.
Rodrigues, R.J., Almeida, T., Richardson, P.J., Oliveira, C.R., Cunha,R.A., 2005. Dual presynaptic control by ATP of glutamaterelease via facilitatory P2X1, P2X2/3, and P2X3 and inhibitoryP2Y1, P2Y2, and/or P2Y4 receptors in the rat hippocampus.J. Neurosci. 25, 6286–6295.
Saitoe, M., Schwarz, T.L., Umbach, J.A., Gundersen, C.B., Kidokoro,Y., 2001. Absence of junctional glutamate receptor clusters inDrosophila mutants lacking spontaneous transmitter release.Science 293, 514–517.
Sutton, M.A., Wall, N.R., Aakula, G.N., Schuman, E.M., 2004.Regulation of dendritic protein synthesis by miniature synapticevents. Science 304, 1979–1983.
Thompson, A.M., Schofield, B.R., 2000. Afferent projections of thesuperior olivary complex. Microsc. Res. Tech. 51, 330–354.
Vial, C., Roberts, J.A., Evans, R.J., 2004. Molecular properties ofATP-gated P2X receptor ion channels. Trends Pharmacol. Sci.25, 487–493.
Walmsley, B., Berntson, A., Leao, R.N., Fyffe, R.E.W., 2006.Activity-dependent regulation of synaptic strength andneuronal excitability in central auditory pathways. J. Physiol.572, 313–321.
35B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6

Author's personal copy
Watano, T., Calvert, J.A., Vial, C., Forsythe, I.D., Evans, R.J., 2004.P2X receptor subtype-specific modulation of excitatory andinhibitory synaptic inputs in the rat brainstem. J. Physiol. 558,745–757.
Wollmann, G., Acuna-Goycolea, C., van den Pol, A.N., 2005. Directexcitation of hypocretin/orexin cells by extracellular ATP atP2X receptors. J. Neurophysiol. 94, 2195–2206.
Woolf, N.K., Ryan, A.F., 1985. Ontogeny of neural dischargepatterns in the ventral cochlear nucleus of the mongoliangerbil. Dev. Brain Res. 17, 131–147.
Yao, S.T., Barden, J.A., Finkelstein, D.I., Bennett, M.R., Lawrence,A.J., 2000. Comparative study on the distribution patternsof P2X1-P2X6 receptors immunoreactivity in the brainstemof the rat and the common marmoset (Callithrix jacchus):association with catecholamine cell groups. J. Comp. Neurol.427, 485–507.
Zhang, J., Wang, H., Ye, C., Ge, W., Chen, Y., Jiang, Z., Wu, C.,Poo, M., Duan, S., 2003. ATP released by astrocytes mediatesglutamatergic activity-dependent heterosynaptic suppression.Neuron 40, 971–982.
36 B R A I N R E S E A R C H 1 2 9 5 ( 2 0 0 9 ) 2 1 – 3 6
Related Documents











