Research paper Methodological aspects of rat β-endorphin analysis—influence of diurnal variation Anja Finn a,c, ⁎ , Susanne Froelich Fabre b , Per M. Hellström c , Stefan Brené d a Peptide Laboratory, Section of Medical Rehabilitation, Department of Surgical Sciences, Sweden b AstraZeneca R&D, Department of Disease Biology, S-151 85 Södertälje, Sweden c Department of Medicine, Unit of Gastroenterology and Hepatology, Karolinska Universitetssjukhuset, Solna, S-171 76 Stockholm, Sweden d Department of Neurotec, Karolinska Institutet, Universitetssjukhuset, Huddinge, M98, 141 86 Huddinge, Sweden Received 15 September 2005; received in revised form 20 December 2005; accepted 2 March 2006 Available online 24 April 2006 Abstract β-Endorphin radioimmunoassays (RIAs) are widely performed following physical, emotional and environmental challenges in the rat. In the literature, a wide range of techniques have been described, but in the present study, we have focused on methodological aspects of β-endorphin RIAs, investigating various characteristics of human and rat specific antibodies. Initial studies verified that the RIA outcome was not appropriate when using non-species compatible components. Novel rat β-endorphin antibodies, r 4114 and r 4268, were raised in rabbits and characterised in terms of specificity, avidity and titer. Both of the new antisera showed 68.1% cross-reactivity with human β-endorphin. The ED 50 was 50 ± 8 pmol/l, and the mean ED 80 was 17 pmol/l for r 4268 but three-fold higher for r 4114. The intra-assay coefficient of variation (CV) was 7% at 100 pmol/l and the inter-assay CV was 10% at the same level for r 4268 and similar for r 4114. Using this novel rat β-endorphin RIA for analyses of diurnal influence and removal from the Animal House cage, no significant changes were observed in either the hypothalamus or peri-aqueductal grey regions. These results suggest that rat β-endorphin concentrations in these brain areas are not affected by order of removal or diurnal variation. © 2006 Elsevier B.V. All rights reserved. Keywords: Radioimmunoassay; β-Endorphin; Diurnal; Hypothalamus; PAG 1. Introduction β-Endorphin radioimmunoassays (RIAs) are com- monly used to study the regulation of β-endorphin levels in the hypothalamus and the peri-aqueductal grey (PAG) areas of the brain, which are regions involved in the adaptation to individual challenges (Berglund et al., 1989; Popp and Erickson, 1998; Miller et al., 2003; Laurent et al., 2004). However, crucial methodological aspects of the RIA technique used are normally not described. There are three types of β-endorphin RIAs in use: (1) commercially available KITs (Olive et al., 2001; Miller et al., 2003); (2) assays with separate compounds as antisera, labeled peptide and standard peptide. Some of these are in-house and some are commercial (Barrett et al., 2000; Medina et al., 1998; Wardlaw et al., 1996; Journal of Immunological Methods 312 (2006) 118 – 125 www.elsevier.com/locate/jim ⁎ Corresponding author. B215, R&D, AstraZeneca, S-151 85 Södertälje, Sweden. Tel.: +46 8 55323240; fax: +46 8 55328949. E-mail address: [email protected] (A. Finn). 0022-1759/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.jim.2006.03.006

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Immunological Methods 312 (2006) 118–125www.elsevier.com/locate/jim
Research paper
Methodological aspects of rat β-endorphin analysis—influence ofdiurnal variation
Anja Finn a,c,⁎, Susanne Froelich Fabre b, Per M. Hellström c, Stefan Brené d
a Peptide Laboratory, Section of Medical Rehabilitation, Department of Surgical Sciences, Swedenb AstraZeneca R&D, Department of Disease Biology, S-151 85 Södertälje, Sweden
c Department of Medicine, Unit of Gastroenterology and Hepatology, Karolinska Universitetssjukhuset, Solna, S-171 76 Stockholm, Swedend Department of Neurotec, Karolinska Institutet, Universitetssjukhuset, Huddinge, M98, 141 86 Huddinge, Sweden
Received 15 September 2005; received in revised form 20 December 2005; accepted 2 March 2006Available online 24 April 2006
Abstract
β-Endorphin radioimmunoassays (RIAs) are widely performed following physical, emotional and environmentalchallenges in the rat. In the literature, a wide range of techniques have been described, but in the present study, wehave focused on methodological aspects of β-endorphin RIAs, investigating various characteristics of human and rat specificantibodies. Initial studies verified that the RIA outcome was not appropriate when using non-species compatiblecomponents.
Novel rat β-endorphin antibodies, r 4114 and r 4268, were raised in rabbits and characterised in terms of specificity, avidity andtiter. Both of the new antisera showed 68.1% cross-reactivity with human β-endorphin. The ED50 was 50±8 pmol/l, and the meanED80 was 17 pmol/l for r 4268 but three-fold higher for r 4114. The intra-assay coefficient of variation (CV) was 7% at 100 pmol/land the inter-assay CV was 10% at the same level for r 4268 and similar for r 4114.
Using this novel rat β-endorphin RIA for analyses of diurnal influence and removal from the Animal House cage, no significantchanges were observed in either the hypothalamus or peri-aqueductal grey regions. These results suggest that rat β-endorphinconcentrations in these brain areas are not affected by order of removal or diurnal variation.© 2006 Elsevier B.V. All rights reserved.
Keywords: Radioimmunoassay; β-Endorphin; Diurnal; Hypothalamus; PAG
1. Introduction
β-Endorphin radioimmunoassays (RIAs) are com-monly used to study the regulation of β-endorphinlevels in the hypothalamus and the peri-aqueductal grey(PAG) areas of the brain, which are regions involved in
⁎ Corresponding author. B215, R&D, AstraZeneca, S-151 85Södertälje, Sweden. Tel.: +46 8 55323240; fax: +46 8 55328949.
E-mail address: [email protected] (A. Finn).
0022-1759/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.jim.2006.03.006
the adaptation to individual challenges (Berglund et al.,1989; Popp and Erickson, 1998; Miller et al., 2003;Laurent et al., 2004). However, crucial methodologicalaspects of the RIA technique used are normally notdescribed. There are three types of β-endorphin RIAs inuse: (1) commercially available KITs (Olive et al., 2001;Miller et al., 2003); (2) assays with separate compoundsas antisera, labeled peptide and standard peptide. Someof these are in-house and some are commercial (Barrettet al., 2000; Medina et al., 1998; Wardlaw et al., 1996;

119A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
Eskeland et al., 1992); (3) RIAs developed in-house(Glämsta et al., 1993; Karuri et al., 1998).
β-Endorphin is derived from the precursor pro-opiomelanocortin (POMC) and represents the C-termi-nal 31 residues of POMC and β-lipotropin (β-LPH).Furthermore, the N-terminal region of β-endorphin (1–5) (human and rat) shows homology to met-enkephalin(Fig. 1). The primary structure differs slightly betweenspecies (Strand, 1999), and significant differencesrelevant from a methodological perspective are presentin the C-terminal domain (Owens and Smith, 1987)(Fig. 1). In the case of rat and human β-endorphin, itwas previously noted that there is potential risk inassuming equal cross-reactivity between these species(Ogawa et al., 1979). However, a number of later studiesfound in the literature present results using approacheswhich combine less compatible antisera or no definedstandards in their β-endorphin assays (Sacerdote et al.,1987; Berglund et al., 1989; Jessop et al., 1994;Wardlaw et al., 1996; Johansson et al., 1997; Poppand Erickson, 1998; Ceccarelli et al., 2004). Although itmakes comparisons between different studies difficult, itshould be noted that the divergence in the methods usedand their results may be attributed to a range of factorssuch as availability of standards and antibodies andmethodological experience at the present time. Thecamel antibody described by Ogawa et al. (1979), wasreported to show 100% cross-reactivity with ovine β-endorphin but only 30% with human β-endorphin,although it was not tested against rat β-endorphin. It wasassumed that the camel and rat sequence werehomologous, although the peptides differ at aminoacid residue 26 (Fig. 1). However, manufacturers havenow produced antibodies to camel β-endorphin thatshow 100% cross-reactivity with rat β-endorphin.
In the present study, methodological aspects of β-endorphin radioimmunoassays (RIAs) were assessed inorder to clarify some of the differences between theantibodies in use. Initially, we measured β-endorphin inthe hypothalamus in Lewis and Fisher rats with anantibody directed against human β-endorphin. Howev-
Met-EnkH-Tyr-Gly-Gly-Phe-Met-Thr-Ser-Glu-Lys-Ser-Gln-Ile-Ile-Lys-Asn-Val26-His27-Lys-Lys-Gly-Gln31 -O
-Ala26-Tyr27-Lys-Lys-Gly-Glu31
-Ala26-His27-Lys-Lys-Gly-Glu31
Fig. 1. Amino acid sequences of β-endorphin in rat, human and camel. Thenkephalin. Significant differences, relevant from a methodological perspect
er, methodological validation showed lack of parallel-ism, and as results may depend on the choice ofantiserum, novel antibodies against rat β-endorphinwere raised and characterised.
The outcome of a β-endorphin RIA depends onseveral aspects of the experimental conditions. A diurnalpattern and variation in β-endorphin release has beenreported in rat plasma (Barrett et al., 2000) and brainareas (Kerdelhue et al., 1983). Therefore, we set out toinvestigate the concentrations of β-endorphin-likeimmunoreactivity in the hypothalamus and PAG ofrats using these newly produced antibodies. Theimportance of diurnal rhythm as well as the order ofremoval from their Animal House cage was assessed.The latter is a model of environmental stress (Borsini etal., 1989; Brodin et al., 1994).
2. Methods
2.1. Animals and study design
Male Fisher and Lewis rats were obtained fromTaconic M&B, Ry, Denmark, and male Sprague–Dawley rats were from B&K Universal AB, Sollentuna,Sweden. The light–dark cycle in the animal housingfacilities was a 12-h-light–12-h-dark schedule, withlights on at 0700 h daily. All rats were provided withfood and water ad libitum. The study protocols wereapproved by the Ethics Committee for animal researchin Stockholm, Sweden. Fisher and Lewis rats wereindividually housed and Sprague–Dawley rats (230–250 g) were housed socially with 4–5 animals in eachcage (Table 1). The rats were not exposed to anytreatment except the daily laboratory routines. After 4weeks of individual housing, the Fisher and Lewis ratswere sacrificed. The brain was rapidly removed, thehypothalamus dissected by hand and directly put on dryice. After 1 week of adaptation to local routines, the SDrats were sacrificed in consecutive order by decapitationdaytime or at night, beginning at 0700 or 1900 h,respectively. The brains were rapidly removed and
Thr-Pro-Leu-Val-Thr-Leu-Phe-Lys-Asn-Ala-H (rat 1-31)
-OH (human 26-31)
-OH (camel 26-31)
e N-terminal region of β-endorphin (1–5) shows homology to met-ive, are present in the C-terminal domain (aa 26–31).

Table 1Summary of study design and β-endorphin concentrations in hypothalamic (HT) and the peri-aqueductal grey (PAG) areas in the male rat brain
β-endorphin pmol/mg protein
Study number Group Strain Housing Time of death Extraction Antibody Standard HT mean±S.D. Dilution PAG mean±S.D.
1 a Lewis Singly 1200–1330 h Acid h A40 Rat 3.04±0.35 1/50 ×1 b Lewis Singly 1200–1330 h Acid h A40 Human 3.30±0.34 1/50 ×1 a Lewis Singly 1200–1330 h Acid h A40 Rat 2.01±0.29 1/10 ×1 b Lewis Singly 1200–1330 h Acid h A40 Human 1.20±0.11 1/10 ×2 a Lewis Singly 1200–1330 h Acid h A40 Rat 3.72±0.95 1/50 ×2 b Fisher Singly 1200–1330 h Acid h A40 Rat 2.97±0.25 1/50 ×3 a SD Group 0700–0830 h Neutral r 4114 Rat 14.42±7.74 1/10 1.50±0.793 b SD Group 1900–2030 h Neutral r 4114 Rat 10.33±0.89 1/10 1.09±0.65
SD=Sprague–Dawley, h=human, r=rat.
120 A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
placed in a cold mould (Brain Blocker, AgnThos,Lidingö, Sweden) where they could be sliced in 2.5-mmcoronal sections by a thin stainless steel razor blade. Thehypothalamus and PAG regions were dissected out andimmediately frozen on dry ice. All the tissues werestored at −70 °C until extraction and RIA.
2.2. Extraction of tissue samples
Tissues from studies 1 and 2 (Table 1) were cut intosmall pieces and placed into a pre-warmed tube with2 ml 1 M acetic acid. The samples were placed intoboiling water for 10 min. Homogenization wasperformed using a steel rod in the tube and a Vortexmixer, followed by centrifugation (4 °C, 2800×g) for10 min. Supernatants were decanted into a new tube,and the pellet was reconstituted in 2 ml of 0.05 Mphosphate buffer (pH 7.4) including 0.015% EDTA andreboiled for another 10 min. After repeat centrifugation,the second supernatant was pooled with the corres-ponding one from the first boiling process, lyophilizedand kept at −20 °C. The samples were dissolved in0.05 M phosphate buffer (pH 7.4) before RIA. Tissuesfrom study 3 (Table 1) were extracted in reverse order,e.g., first with phosphate buffer and then reboiled inacetic acid, to be able to obtain peptides that require aneutral extraction. Changing the order of the extractiontechnique for β-endorphin, we did not note anydifferences since the peptide data are correlated to theamount of total protein.
2.3. Antisera–rat β-endorphin
Antisera were raised in-house (Glämsta et al.,1993) in female New Zealand rabbits in parallel withthose previously described in detail (Finn et al.,2004). Briefly, synthetic rat β-endorphin 1–31 (SC106, Neosystem, Strasbourg, France), was coupled to
porcine thyroglobulin using the glutaraldehyde meth-od (Adrian, 1997). First, rat β-endorphin peptide wasmixed with phosphate buffer. Thereafter, thyroglobu-lin was added and vigorously mixed with the additionof ice-cold glutaraldehyde. The suspension wasstirred avoiding light and the reaction product wasdialyzed, diluted and emulsified drop-wise withFreund's complete adjuvant. The conjugate wasadministered subcutaneously, in four rabbits,0.25 ml in each of four sites in the neck region toavoid unwanted tissue reaction. Booster doses of thesame conjugate administered in Freund's incompleteadjuvant were given three times at 8-week intervals.Ten days after the second booster dose, the rabbitswere bled from an ear vein and serum was assessedfor antibodies.
2.4. Radioimmunoassay
2.4.1. Human β-endorphin-LIHuman β-endorphin antibody (A 40, Eurodiagnos-
tica, Malmö, Sweden) raised in rabbits with a synthetichuman β-endorphin peptide (Peninsula Labs, SanCarlos, CA, USA) as an immunogen was used in thetwo first studies. The immunization protocol wascarried out as described by the manufacturer. Thecross-reactivity with rat β-endorphin was not given butstated to be 100% for human β-endorphin and <1% forβ-lipotropin (human). The antibody was used withhuman as well as rat β-endorphin peptide to createstandard curves in study 1 (Fig. 2) and for the assay ofstudy 2.
2.4.2. Rat β-endorphin-LIA standard curve for the in-house rat β-endorphin
(range 3.9–1000 pmol/l) was prepared by serial dilutionswith phosphate buffer (pH 7.4). 100 μL of standardsolution or extracted sample (HT diluted 1/10) were

0
0.2
0.4
0.6
0.8
1
0
0.2
0.4
0.6
0.8
1
0 3.9 7.8 15.6 31.2 62.5 125 250 500 1000 0 3.9 7.8 15.6 31.2 62.5 125 250 500 1000
rat ab 4114/rat STDrat ab 4268/rat STD
human ab/rat STDhuman ab/human STD
A. rat β-endorphin B. β-endorphin, human versus rat STD
Concentration (pmol/l)
Re s pon s e
Fig. 2. Validation. (A) Standard curves using newly produced β-endorphin specific rat antibodies and rat standard. (B) Standard curves of rat andhuman β-endorphin analysed with human specific β-endorphin antibody A 40. (Study 1).
121A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
incubated with 200 μl of antiserum solution (r 4114 wasused for study 3) at 4 °C for 48 h. The tracer labeling wascarried out using the chloramine-T method (Eurodiag-nostica, Malmö, Sweden on request and Phoenix Pharm,Karlsruhe, Germany) and 200 μl of the tracer were addedand incubated at 4 °C for 24 h. Separation was performedusing 500 μl of a second antibody, Suspension-3(Pharmacia Biotech, Uppsala, Sweden) or 50 μl Sac-cel (HPLC, study 3) (IDS Ltd, Boldon, Tyne and Wear,UK) for 30 min at room temperature. After the additionof 1 ml H2O, the tubes were centrifuged and decanted.The bound fraction was assessed for 3 min in a gamma-counter (PerkinElmer-Wallac OY, Turku, Finland). Allsamples were assayed in duplicate.
2.5. Protein determination
The protein concentration of tissue homogenates wasanalysed according to a slightly modified method (Finnet al., 2004) of Lowry. Peptide concentrations werecalculated and expressed as pmol/mg protein.
2.6. High-performance liquid chromatography
Reverse-phase HPLC for rat β-endorphin wasperformed using Genesis C 18 300 Å, 4, 150 mm(Sorbent, Västra Frölunda, Sweden) eluted with a25-min linear gradient of acetonitrile (10–60%) in watercontaining 0.1% TFA. Thereafter, a washing period wasincluded for 35 min with a linear gradient of acetonitrile(60–70%) in water containing 0.1% TFA. Samples werepassed through Millipore GS filters, (0.45 μm) beforechromatography and 100 μl of the synthetic peptide(standard solution with TFA) were injected. The flowwas maintained by two Gilson pumps controlled by a
gradient programmer (Unipoint System Software, Gil-son, Middletown, WI, USA). Fractions of 1 ml werecollected at an elution rate of 1 ml/min and, thesefractions were lyophilized and re-dissolved in 100 μlassay buffer before analysis.
2.7. Statistics
Due to the skewness of the data, the Mann–WhitneyU-test was used for statistical evaluation of straindifferences. Diurnal variation was calculated using theWilcoxon matched pairs test. P>0.05 was consideredstatistically different.
3. Results
An overview of mean values and standard deviationsof peptide concentrations established in the differentstudy groups is presented in Table 1.
3.1. β-Endorphin (rat) antisera
Antiserum r 4268 displayed a titer of 1:37,000(final dilution in the assay tube). The avidityconstant was 0.16×1012 M according to Scatchard.The ED50 was 50±8 pmol/l and the mean ED80 was17 pmol/l. The intra-assay coefficient of variation (CV)was 28% at 10 pmol/l (limit of quantification) and 7% at100 pmol/l and the inter-assay CV were 13% and 10%,respectively.
Antiserum r 4114 had a titer of 1:18,000, and anavidity constant of 0.035×1012 M. The ED50 was 174±5 pmol/l and ED80 was 40 pmol/l. The intra-assay CVwas 6% at levels between 15 and 250 pmol/l and inter-assay CV was 13% at ED50 levels. The cross-reactivity
A.
0
500
1000
1500
2000
25001 3 5 7 9
11 13 15 17 19 21 23 25
fractions
fractions
Co
nce
ntr
atio
n (
pm
ol/l
)
rat STD
humanSTD
B.
0
50
100
150
200
250
300
350
400
9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24
Co
nce
ntr
atio
n (
pm
ol/l
)
HT
PAG
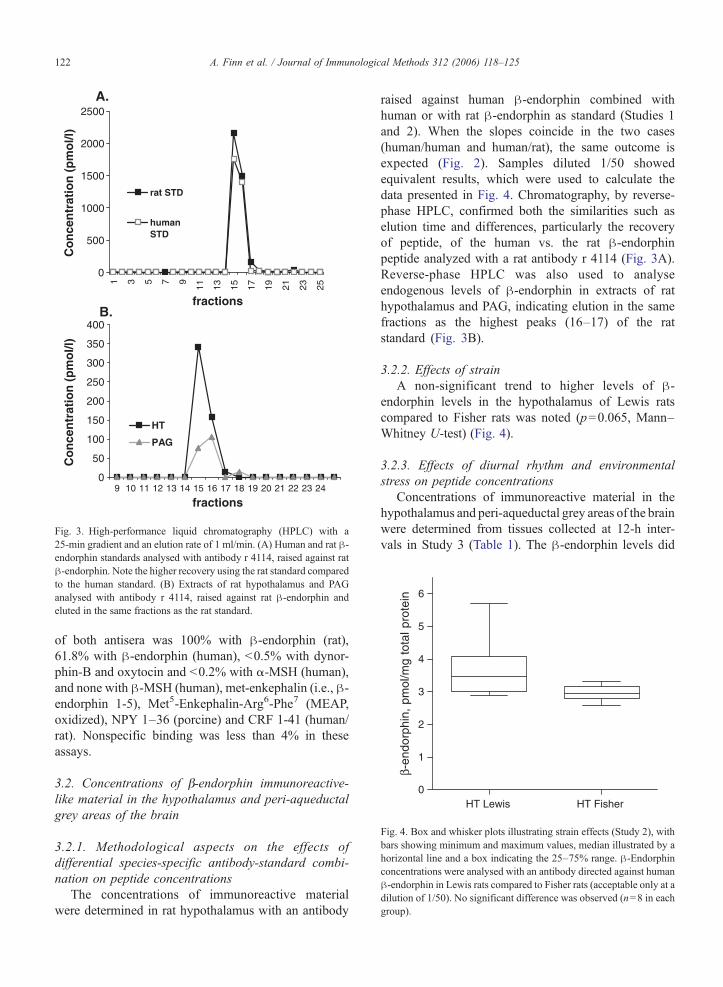
Fig. 3. High-performance liquid chromatography (HPLC) with a25-min gradient and an elution rate of 1 ml/min. (A) Human and rat β-endorphin standards analysed with antibody r 4114, raised against ratβ-endorphin. Note the higher recovery using the rat standard comparedto the human standard. (B) Extracts of rat hypothalamus and PAGanalysed with antibody r 4114, raised against rat β-endorphin andeluted in the same fractions as the rat standard.
HT Lewis HT Fisher 0
1
2
3
4
5
6
β-en
dorp
hin,
pm
ol/m
g to
tal p
rote
in
Fig. 4. Box and whisker plots illustrating strain effects (Study 2), withbars showing minimum and maximum values, median illustrated by ahorizontal line and a box indicating the 25–75% range. β-Endorphinconcentrations were analysed with an antibody directed against humanβ-endorphin in Lewis rats compared to Fisher rats (acceptable only at adilution of 1/50). No significant difference was observed (n=8 in eachgroup).
122 A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
of both antisera was 100% with β-endorphin (rat),61.8% with β-endorphin (human), <0.5% with dynor-phin-B and oxytocin and <0.2% with α-MSH (human),and none with β-MSH (human), met-enkephalin (i.e., β-endorphin 1-5), Met5-Enkephalin-Arg6-Phe7 (MEAP,oxidized), NPY 1–36 (porcine) and CRF 1-41 (human/rat). Nonspecific binding was less than 4% in theseassays.
3.2. Concentrations of β-endorphin immunoreactive-like material in the hypothalamus and peri-aqueductalgrey areas of the brain
3.2.1. Methodological aspects on the effects ofdifferential species-specific antibody-standard combi-nation on peptide concentrations
The concentrations of immunoreactive materialwere determined in rat hypothalamus with an antibody
raised against human β-endorphin combined withhuman or with rat β-endorphin as standard (Studies 1and 2). When the slopes coincide in the two cases(human/human and human/rat), the same outcome isexpected (Fig. 2). Samples diluted 1/50 showedequivalent results, which were used to calculate thedata presented in Fig. 4. Chromatography, by reverse-phase HPLC, confirmed both the similarities such aselution time and differences, particularly the recoveryof peptide, of the human vs. the rat β-endorphinpeptide analyzed with a rat antibody r 4114 (Fig. 3A).Reverse-phase HPLC was also used to analyseendogenous levels of β-endorphin in extracts of rathypothalamus and PAG, indicating elution in the samefractions as the highest peaks (16–17) of the ratstandard (Fig. 3B).
3.2.2. Effects of strainA non-significant trend to higher levels of β-
endorphin levels in the hypothalamus of Lewis ratscompared to Fisher rats was noted (p=0.065, Mann–Whitney U-test) (Fig. 4).
3.2.3. Effects of diurnal rhythm and environmentalstress on peptide concentrations
Concentrations of immunoreactive material in thehypothalamus and peri-aqueductal grey areas of the brainwere determined from tissues collected at 12-h inter-vals in Study 3 (Table 1). The β-endorphin levels did

HT morning HT evening0
5
10
15
20
25
30
35
40
β-e
ndor
phin
, pm
ol/m
g to
tal p
rote
in
PAG morning PAG evening0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
β-en
dorp
hin,
pm
ol/m
g to
tal
prot
ein
A.
B.
Fig. 5. Box and whisker plots illustrating circadian effects (Study 3),with bars showing minimum and maximum values, median illustratedby a horizontal line and a box indicating the 25–75% range. (A) β-Endorphin concentrations in the hypothalamus were analysed withnewly produced rat antibody r 4114. No significant differences wereobserved (n=8 in each group). (B) β-Endorphin concentrations in theperi-aqueductal grey (PAG) area of the brain, were analysed withnewly produced rat antibody r 4114. No significant differences wereobserved (n=8 morning group, n=7 evening group).
123A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
not show any significant differences between the lightand dark phases. A non-significant trend towardshigher levels of β-endorphin levels were noted in themorning in HT, but not in PAG (p=0.093 HT,p=0.735, PAG, Wilcoxon matched pairs test) (Fig.5). Regression analysis of β-endorphin levels inrelation to “order of removal” showed no significantdeviation from zero (data not shown).
4. Discussion
This report concerns species-dependent aspects ofRIA, illustrated by the example of β-endorphin
assays, including the development of two new ratantibodies. The choice of antiserum for RIA method-ology is critical. Appropriate β-endorphin specificantisera should not cross-react or interact with the N-terminal domain of the peptide. Thus, most of theantisera are typically directed against the C-terminal partof the peptide. Furthermore, due to sequence diver-gence, the specificity of an antiserum is dependent onwhich species is the source of the β-endorphin that isused as immunogen.
Results from our present studies clearly show thata RIA needs to be validated for the species to whichit is applied (Figs. 2 and 3A). Comparing the slopesof the human vs. rat standard curves in studies 1and 2, an initial similarity was noted between thecurves. However, an increased deviation was seenwith increased peptide concentrations, indicating aweakening compatibility and reduced cross-reactivityin the human antibody/rat standard case (Fig. 2B).This will have important consequences when analys-ing levels in that range. Due to this initialexperience of antisera-standard incompatibility, anovel antibody against rat β-endorphin was raisedand characterized.
The outcome of an RIA may also depend ondifferential affinity/avidity between the antisera. Adifference between our two novel rat β-endorphinantisera was found which could be related to thisfactor. We consider the avidity constant to beappropriate at levels above 1011 M in order toachieve sensitivity high enough to permit alterationsof β-endorphin levels in a biological setting.Polyclonal antisera always have several affinities. Astandard curve with a late or a poor slope hasnumerous affinities at that level, compared to serawith an early slope such as antibody r 4268. This wasillustrated in the three-fold or larger difference inED50 concentrations (Fig 2A) obtained using thedifferent rat antibodies. The ‘in-house’ rat β-endor-phin antisera also showed higher assay CVs at lowconcentrations. However, endogenous levels of β-endorphin were 4–10 times higher. The rat antibodiesexhibited a larger detection range together withspecificity, thereby permitting studies of variationsbetween individuals, as shown in the final results instudy 3.
In order to verify if the assayed concentrations areof biological significance, the effects of differentinterventions were addressed by analysing β-endor-phin concentrations in rat tissue. In study 3, SpragueDawley males were used to investigate the effects ofdiurnal rhythm and environmental stress (Borsini et

124 A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
al., 1989; Brodin et al., 1994). Trends towardsbiphasic changes in peptidergic concentrations werefound in both the hypothalamus and PAG during the24-h period. This is consistent with reports of diurnalfluctuations of β-endorphin release in plasma andlevels in rat brain (Kerdelhue et al., 1983; Barrett etal., 2000). Also, the increase of PAG β-endorphin isconsistent with an increased sympathetic tone (Cov-entry et al., 2001).
In summary, this study has focused on methodo-logical aspects of a β-endorphin RIA. Novel rat β-Endorphin antibodies, r 4114 and r 4268, were raisedand characterized in terms of specificity, avidity andtiter. Both of the new antisera showed 68.1% cross-reactivity with human β-endorphin. Studies withantisera directed against human β-endorphin todetermine levels of the peptide in the rat hypothal-amus clearly demonstrated that the RIA outcome wasonly reliable in a narrowly defined range when usingnon-species compatible components. Increased sensi-tivity and an improved detection range were obtainedwhen using species-specific antibody and standard,illustrating the importance of a proper analysisprocedure. When analysing diurnal influence on β-endorphin concentration in the brain, no significantchanges were seen in β-endorphin concentrations inrat hypothalamus or PAG.
Both of the rat β-endorphin specific antisera (r 4114and r 4268) described here are available for validation ofRIAs on request.
Acknowledgement
This work was initiated and supported by Dr. T.Lundeberg and mainly financed by the Foundation forAcupuncture and CBT, FoUU, Rehab. Medicine,Karolinska Hospital and Swedish Research Council(S. Brené).
References
Adrian, T.E., 1997. Production of antisera using peptide conjugates.Methods Mol. Biol. Neuropept. Protoc. 73, 239.
Barrett, T., Stephen, K., Voudouris, N., 2000. Does melatoninmodulate beta-endorphin, corticosterone and pain treshold? LifeSci. 66, 467.
Berglund, L.A., Millington, W.R., Simpkins, J.W., 1989. Gonadalsteroid and chronic morphine treatment do not change theposttranslational processing of β-endorphin in the rat brain. LifeSci. 44, 601.
Borsini, F., Lecci, A., Volterra, G., Meli, A., 1989. A model tomeasure anticipatory anxiety in mice? Psychopharmacology 98,207.
Brodin, E., Rosen, A., Schott, E., Brodin, K., 1994. Effects ofsequential removal of rats from a group cage, and of individualhousing of rats, on substance P, cholecystokinin and somatostatinlevels in the periaqueductal grey and limbic regions. Neuropep-tides 4, 253.
Ceccarelli, I., Lariviere, W.R., Fiorenzani, P., Sacerdote, P., Aloisi,A.M., 2004. Effects of long-term exposure of lemon essential oilodor on behavioral, hormonal and neuronal parameters in maleand female rats. Brain Res. 1001, 78.
Coventry, T.L., Jessop, D.S., Finn, D.P., Crabb, M.D., Kinoshita,H., Harbuz, M.S., 2001. Endomorphins and activation of thehypothalamus–pituitary–adrenal axis. J. Endocrinol. 169, 85.
Eskeland, N.L., Molineaux, C.J., Schachter, B.S., 1992. Regulation ofβ-endorphin secretion by corticotropin releasing factor in the intactrat testis. Endocrinology 130, 1173.
Finn, A., Ågren, G., Bjellerup, P., Vedin, I., Lundeberg, T., 2004.Production and characterization of antibodies for the specificdetermination of the opioid peptide Met5-Enkephalin–Arg6–Phe7.Scand. J. Clin. Lab. Invest. 64, 49.
Glämsta, E.L., Morkrid, L., Lantz, I., Nyberg, F., 1993. Concominantincrease of blood plasma levels of immunoreactive hemorphin-7and beta-endorphin following long distance running. Regul. Pept.19, 49.
Jessop, D.S., Lightman, S.L., Chowdrey, H.S., 1994. Effects of achronic inflammatory stress on levels of pro-opiomelanocortin-derived peptides in the rat spleen and thymus. J. Neuroimmunol.49, 197.
Johansson, P., Ray, A., Zhou, Q., Huang, W., Karlsson, K., Nyberg, F.,1997. Anabolic androgenic steroids increase β-endorphin levels inthe ventral tegmental area in the male rat brain. Neurosci. Res. 27,185.
Karuri, A.R., Agarwai, R.K., Engelking, L.R., Kumar, M.S.A., 1998.Effects of halothane and metoxyflurane on regional brain andspinal cord substance-P and beta-endorphine-like immunoreactiv-ities in the rat. Brain Res. Bull. 45, 501.
Kerdelhue, B., Karteszi, M., Pasqualini, C., Reinberg, A., Mezey, E.,Palkovits, M., 1983. Circadian variations in beta-endorphinconcentrations in pituitary and in some brain nuclei of the adultmale rat. Brain Res. 261, 243.
Laurent, V., Jaubert-Miazza, L., Desjardins, R., Day, R., Lindberg, I.,2004. Biosynthesis of proopiomelanocortin-derived peptides inprohormone convertase 2 and 7B2 null mice. Endocrinology 145,519.
Medina, F., Siddiqui, A., Scimonelli, T., Fenske, C., Wilson, C.A.,Celis, M.E., 1998. The inter-relationship between gonadalsteroids and POMC peptides, β-Endorphin and α-MSH, inthe control of sexual behavior in the female rat. Peptides 19,309.
Miller, R., Aaron, W., Toneff, T., Vishnuvardhan, D., Beinfeld, M.C.,Hook, V.Y.H., 2003. Obliteration of α-melanocyte-stimulatinghormone derived from POMC in pituitary brains of PC2-deficientmice. J. Neurochem. 86, 556.
Ogawa, N., Panerai, A.E., Lee, S., Forsbach, G., Havlicek, V., Friesen,H.G., 1979. Beta-Endorphin concentration in the brain of intactand hypophysectomized rats. Life Sci. 23–25, 317.
Olive, M.F., Koenig, H.N., Nannini, M.A., Hodge, C.W., 2001.Stimulation of endorphin neurotransmission in the nucleusaccumbens by ethanol, cocaine and amphetamine. J. Neurosci.21, 1.
Owens, P.C., Smith, R., 1987. Opioid peptides in blood andcerebrospinal fluid during acute stress. Baillière's Clin. Endocri-nol. Metab. 2, 415.

125A. Finn et al. / Journal of Immunological Methods 312 (2006) 118–125
Popp, L., Erickson, C.K., 1998. The effect of an acute ethanolexposure on the rat brain POMC opiopeptide system. Alcohol 2,139.
Sacerdote, P., Brini, A., Mantegazza, P., Panerai, A.E., 1987. A rolefor serotonin and beta-endorphin in the analgesia induced bysome tricyclic antidepressant drugs. Pharmacol. Biochem. Behav.1, 153.
Strand, F.L., 1999. The POMC-derived neuropeptides: ACTH, MSH,β-LPH, β-EP. Neuropeptides. Regulators of Physiological Pro-cesses. MIT Press, Cambridge, MA, p. 267.
Wardlaw, S.L., Kim, J., Sobieszczyk, S., 1996. Effect of morphine onproopiomelanocortin gene expression and peptide levels in thehypothalamus. Mol. Brain Res. 31, 140.
Related Documents


![Diurnal and Nocturnal Animals. Diurnal Animals Diurnal is a tricky word! Let’s all say that word together. Diurnal [dahy-ur-nl] A diurnal animal is an.](https://static.cupdf.com/doc/110x72/56649dda5503460f94ad083f/diurnal-and-nocturnal-animals-diurnal-animals-diurnal-is-a-tricky-word-lets.jpg)