Membrane Alterations during Cornification of Mammalian Squamous Epithelia: A Freeze-Fracture, Tracer, and T hin-section Study PETER M. ELIAS,' N. SCOTT MCNUlT * AND DANIEL S. FRIEND 'Dermatology Section and 'Anatomic Pathology Service, Veterans Administration Hospital, San Francisco, California 94121; and 'DepartmentsofDerrnatology and 3Pathology, University of California School of Medicine, San Francisco, California ABSTRACT Tight junctions (zonulae occludentes) create a pericellular bar- rier to the diffusion of large molecules in non-keratinizing mammalian epithelia. However, in cornifying epithelia such as the epidermis, the importance of tight- junctional elements versus secreted intercellular lipid for barrier function is uncertain. In an attempt to resolve this question, we compared membrane struc- ture in the stratum granulosum and stratum corneum of epidermis, esophagus, and vagina of newborn and adult humans and mice under both normal and various experimental conditions. We incubated pieces of epidermis in organ cul- ture and infused tissues with lanthanum or horseradish peroxidase in vivo and in vitro. All were processed for electron microscopy of freeze-fracture replicas or thin sections. Lanthanum seeped outward to the stratum granulosum in all tis- sues examined-further apical migration was halted by lamellar-body contents in skin. A similar pattern of intercellular lamellar lipid deposition and mem- brane structure occurred in all epithelia studied. Freeze-fracture replicas of these obstructive regions revealed occasional, incomplete junctional strands (particularly in moist epithelia) and abundant lamellar material, but complete zonulae occludentes were never encountered. A possible relationship between moisture and tight junction formation was further suggested by organ culture experiments during which brief incubations stimulated an increase in the num- ber of junctional strands and diminished numbers of lamellar bodies. We con- clude that, in the epithelia studied, the deposition of secreted lamellar body con- tents forms the barrier to water-soluble tracer loss: tight-junctional elements are either absent or too fragmentary to constitute an effective barrier. In keratinizing strata of squamous epithe- lia the outer layers separate an aqueous inter- nal milieu from a dry external environment. Although the barrier to water loss is generally attributed to the stratum corneum (Scheup- lein and Blank, '71), observations from several morphological studies indicate that the subja- cent stratum granulosum may participate in this function as well (Schreiner and Wolff, '69; Squier, '73; Elias and Friend, '75). At points of lateral membrane fusion, in non-keratinizing and in some keratinizing epithelia, a continuous network of anastomos- ing strands comprises a pericellular permea- bility barrier (Farquhar and Palade, '64; Friend and Gilula, '72). In addition to these ANAT. REC., 189: 577-594. sealing elements (zonulae occludentes or tight junctions) mature and developing tissues may contain interrupted, isolated strands seen in thin sections as focal membrane fusions (fasciae or maculae occludentes). While the function of the zonula occludens is firmly allied to barrier function, the role of focal strands and fusion points is not fully known. They have been considered as scaffolding for forming gap junctions (Decker and Friend, ,751, fragments of "diluted" tight junctions (Revel et al., '73), and as both deteriorating or forming ones (Elias and Friend, '76). While tight junctions function as barriers Received June 3, '77. Accepted July 19, '77. 577

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Membrane Alterations during Cornification of Mammalian Squamous Epithelia: A Freeze-Fracture, Tracer, and T hin-sect ion Study
PETER M. ELIAS,' N. SCOTT MCNUlT * AND DANIEL S. FRIEND 'Dermatology Section and 'Anatomic Pathology Service, Veterans Administration Hospital, San Francisco, California 94121; and 'Departments ofDerrnatology and 3Pathology, University of California School of Medicine, San Francisco, California
ABSTRACT Tight junctions (zonulae occludentes) create a pericellular bar- rier to the diffusion of large molecules in non-keratinizing mammalian epithelia. However, in cornifying epithelia such as the epidermis, the importance of tight- junctional elements versus secreted intercellular lipid for barrier function is uncertain. In an attempt to resolve this question, we compared membrane struc- ture in the stratum granulosum and stratum corneum of epidermis, esophagus, and vagina of newborn and adult humans and mice under both normal and various experimental conditions. We incubated pieces of epidermis in organ cul- ture and infused tissues with lanthanum or horseradish peroxidase in vivo and in vitro. All were processed for electron microscopy of freeze-fracture replicas or thin sections. Lanthanum seeped outward to the stratum granulosum in all tis- sues examined-further apical migration was halted by lamellar-body contents in skin. A similar pattern of intercellular lamellar lipid deposition and mem- brane structure occurred in all epithelia studied. Freeze-fracture replicas of these obstructive regions revealed occasional, incomplete junctional strands (particularly in moist epithelia) and abundant lamellar material, but complete zonulae occludentes were never encountered. A possible relationship between moisture and tight junction formation was further suggested by organ culture experiments during which brief incubations stimulated an increase in the num- ber of junctional strands and diminished numbers of lamellar bodies. We con- clude that, in the epithelia studied, the deposition of secreted lamellar body con- tents forms the barrier to water-soluble tracer loss: tight-junctional elements are either absent or too fragmentary to constitute an effective barrier.
In keratinizing strata of squamous epithe- lia the outer layers separate an aqueous inter- nal milieu from a dry external environment. Although the barrier to water loss is generally attributed to the stratum corneum (Scheup- lein and Blank, '71), observations from several morphological studies indicate that the subja- cent stratum granulosum may participate in this function as well (Schreiner and Wolff, '69; Squier, '73; Elias and Friend, '75).
At points of lateral membrane fusion, in non-keratinizing and in some keratinizing epithelia, a continuous network of anastomos- ing strands comprises a pericellular permea- bility barrier (Farquhar and Palade, '64; Friend and Gilula, '72). In addition to these
ANAT. REC., 189: 577-594.
sealing elements (zonulae occludentes or tight junctions) mature and developing tissues may contain interrupted, isolated strands seen in thin sections as focal membrane fusions (fasciae or maculae occludentes). While the function of the zonula occludens is firmly allied to barrier function, the role of focal strands and fusion points is not fully known. They have been considered as scaffolding for forming gap junctions (Decker and Friend, ,751, fragments of "diluted" tight junctions (Revel et al., '73), and as both deteriorating or forming ones (Elias and Friend, '76).
While tight junctions function as barriers
Received June 3, '77. Accepted July 19, '77.
577

578 PETER M. ELIAS. N. SCOTT MCNUTT AND DANIEL S. FRIEND
in most non-keratinizing epithelia (Decker and Friend, '72; McNutt and Weinstein, '73; Staehelin, '741, they were encountered only rarely in neonatal murine epidermis (Elias and Friend, '75). Instead, a t the level where they would be anticipated, accumulations of intercellular lamellar-body (membrane-coat- ing granule) contents effectively impeded tracer (horseradish peroxidase, lanthanum) egress. Contrary to our findings in this tissue, however, three recent freeze-fracture studies of other keratinizing epithelia - murine esophagus (McNutt, '731, rat oral mucosa (Shimono and Clementi, '76), and human epi- dermis (Caputo, '75) -have established the presence of tight junctions in the stratum granulosum and have suggested a role in regu- lating tissue permeability to water. In the cur- rent study, we present morphological observa- tions on the cell membranes of keratinizing epithelia (with particular attention to the stratum corneum and stratum granulosum) in adult and neonatal mouse and human esopha- gus and epidermis, as well as in the mouse va- gina. The purpose of our investigation was to attempt to resolve the question: Do tight junctions or intercellular lipid sheets derived from lamellar bodies, or both, subserve the pericellular barrier to the diffusion of small tracers?
MATERIAL AND METHODS
Experimental tissue The following mammalian keratinizing epi-
thelia were studied: (1) adult mouse (Swiss albino) epidermal ear skin, because of its rela- tive hairlessness; (2) adult mouse thoracic esophagus (ICR, JAX, CF-1 strains); (3) adult human esophagus and epidermis (fresh surgi- cal specimens) from glabrous sites; (4) new- born Swiss albino mouse epidermis; and (5 ) adult mouse vagina.
Methods for thin sections Several different fixation procedures were
used, all resulting in comparable preservation. Each type of tissue was fixed by a t least two (but not all) of the methods described below (see figure explanations).
Initial fixation in 1.5% glutaral- dehyde in 0.1 M cacodylate buffer with 4% su- crose for four to five hours, followed by post fixation in 2% osmium-potassium ferrocya- nide (Karnovsky, '68) for one and one-half hours in the dark. This method clearly dis- tinguishes membranes, glycogen, lamellar bodies, and horseradish peroxidase.
Method 1
Method 2 Thin slices of tissue were fixed in 1.5% glutaraldehyde in cacodylate buffer for one to one and one-half hours, then incubated in concanavalin A (Con A, Miles Labs., 0.1 mg/ ml in 0.1 M phosphate buffer), a t room temper- ature on a rotary shaker. After 15 minutes horseradish peroxidase (HRP) (Worthington) was added to make a final concentration of 0.05 mg/ml, and incubation was continued for an additional 15 minutes (Huet and Herxberg, '73). After three rapid rinses in phosphate buffer, the slices were incubated in hydrogen peroxide-diaminobenzidine solution (Graham and Karnovsky, '681, and subsequently post- fixed in osmium tetroxide-potassium ferro- cyanide as noted above.
Method 3 Some specimens from all tissues were fixed in 1.5% glutaraldehyde in 0.1 M cacodylate buffer containing 4% sucrose, then washed overnight in 0.1 M cacodylate buffer containing 7% sucrose. Subsequently they were stained en bloc with ethanolic uranyl acetate (4%), dehydrated in graded ethanols, and embedded in Epon 812.
Tissues were fixed in 2% formal- dehyde-2.5% glutaraldehyde-0.008% picric acid (It0 and Karnovsky, '68), in 0.1 M cacody- late buffer, pH 7.4, for four to six hours.
Method 5 Tannic acid (1%) was used with glutaraldehyde as previously described (Til- ney et al., '73).
Some epidermal specimens were fixed directly in osmium vapor for two hours a t room temperature without prior aldehyde fixation (Elias and Friend, '75). Sections were cut with diamond knives, stained with lead citrate and uranyl acetate, and examined in either a JEM-100B, Philips 201, or Siemens 1A electron microscope.
Tracer methods-lanthanum 1. Applied simultaneously with fixative
Only murine esophagi were prepared ac- cording to this method. Cross-sectioned slices fixed by immersion (Method 4) for 15 minutes were transferred to 2% formaldehyde-2.5% glutaraldehyde with 1% lanthanum nitrate (pH 7.41, prepared according to the method of Revel and Karnovsky ('67) in 0.1 M cacodylate buffer, and fixed for four hours. Tissues were then immersed overnight in 2% lanthanum ni- trate in 0.1 M sodium cacodylate (pH 7.4), postfixed in 1% osmium tetroxide-1% lantha- num nitrate in 0.1 M cacodylate buffer, rapidly dehydrated in a graded series of ethanol-water fixtures and embedded in epoxy plastic (Spurr, '69).
Method 4
Method 6

MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 579
Fig. 1 Freeze-fracture replica of neonatal mouse epidermis. Lamellar bodies fill the apical cytoplasm of cells in the mid-to-upper stratum granulosum W). The profiles are ellipsoidal, ranging in diameter from 0.1- 0.3 jan. In this figure the shadowing angle is reversed from the usual upward direction in order to maintain the basal-apical orientation presented in figure 2. X 25,000.
Fig. 2 Freeze-fracture replica of adult mouse esophagus. The structural features of lamellar bodies in the stratum granulosum of esophagus are similar to those in epidermis. Again, lamellar bodies are ellipsoidal, and almost always fracture through their limiting membrane, revealing P and E face images with very few associ- ated particles. SG, stratum granulosum. X 40,000.
2. Zqjection before f ia t ion 4% lanthanum nitrate in 0.05 M Tris-HC1 buffer with 4% sucrose, pH 7.6. Thirty minutes
These experiments were performed with after intracutaneous injection, the tissues adult and neonatal mouse epidermis. The were excised, fixed in 1.5% glutaraldehyde in injection fluid consisted of freshly prepared cacodylate buffer, and processed as above.

580 PETER M. ELIAS, N. SCOTT MCNUTT AND DANIEL S. FRIEND
Tracer methods- horseradish peroxidase Horseradish peroxidase (Sigma, 25 mg/ml
phosphate-buffered saline) was injected in- tracutaneously into neonatal mice in 0.05 ml aliquots. After 30 minutes, the tissues were excised, fixed in glutaraldehyde-cacodylate as above, and washed in 0.1 M cacodylate buffer with 7% sucrose overnight. Forty-micron sec- tions were cut with a Smith-Farquhar tissue chopper, incubated in diaminobenzidine, post- fixed in 1% osmium tetroxide, and processed as described above.
Freeze-fracture techniques After 15 minutes or 2 hours of fixation
in either glutaraldehyde-paraformaldehyde- picric acid or 1.5% glutaraldehyde-cacodylate, cubes of tissue were mounted on cardboard discs, and then fractured and replicated in a Balzers freeze-etch apparatus (Balzers High Vacuum Corp., Santa Ana, California) either a t - 100°C or - 115OC. Platinum-carbon rep- licas were cleaned in bleach or by sequential treatment with absolute methanol and bleach.
Organ culture experiments In order to determine whether environmen-
tal factors can influence the permeability bar- rier in keratinizing epithelia, we obtained 1- cm2 pieces of fresh neonatal mouse-skin and floated them dermis-side-down on metal rafts in plastic Petri dishes. Specimens were main- tained in 100% humidity at 37OC under 5% C02/95% air for 0 to 72 hours. Waymouth’s me- dium supplemented with 10% fetal calf serum, 1% amphotericin B, penicillin (100 U/ml), and streptomycin (50 pg/ml) was employed. Addi- tional samples were incubated in the same media, supplemented with vitamin A acid (Hoffman-LaRoche, 20 U/ml final concentra- tion) and prepared as previously described to ascertain whether vitamin A has the same ef- fect on mouse epidermis in vitro as i t does on chick skin (the stimulus of tight junction for- mation) (Elias and Friend, ’76). The meta- plastic potency of vitamin A was gauged in parallel cultures of 14-day embryonic chick shank skin. At intervals of 0,6, 16,48, and 72 hours, specimens were removed, fixed for 15 to 45 minutes in 1.5% cacodylate-buffered glu- taraldehyde, and processed for freeze-fracture and thin-section electron microscopy as de- tailed above.
TERMINOLOGY
The term freeze-fracture refers to the frac-
ture and rapid surface-replication of frozen tissues before the sublimation of ice from the fractured surface becomes detectable. Freeze- etching refers to the use of controlled sublima- tion (“etching”) of ice from the fractured sur- face before replication (Branton et al., ’66).
Frozen plasma membranes fracture into two leaflets or “faces”: one plasma membrane leaflet (P-face) remains attached to the cyto- plasm, while the other leaflet (E-face) re- mains adjacent to the extracellular space (Branton et al., ’76).
The identification of junctions in thin sec- tions is sometimes difficult, due to variations in tissue fixation, staining, dehydration, sec- tioning, and photography which may obscure the subtle differences between tight and gap junctions (Friend and Gilula, ’72). Classifica- tion of cellular junctions has been recent- ly reviewed (McNutt and Weinstein, ’73; Staehelin, ’74; Gilula, ’74). The gap junction (nexus, communicating junction) is composed of globular subunits, often hexagonally packed, with a 90-100 A center-to-center spac- ing, as first described by Revel and Karnovsky (‘67). In contrast, zonulae occludentes- i.e., “tight” junctions consist of a continuous net- work of intramembranous fibrils joining the membranes of adjacent cells and sealing the lateral intercellular spaces from luminal sur- faces (Staehelin, ’74). Tight junctions facili- tate the assemblage of individual cells into epithelial sheets, impede the admixture of in- tercellular and luminal fluid contents, and
Fig. 3 Section of neonatal mouse stratum granulosum, stained with concanavalin A plus horseradish peroxidase (Con A + HRP). The internal organization of lamellar body contents lying within the intercellular space is revealed by staining with Con A + HRP. Discs do not stain, but the enveloping matrix is darkened revealing the internal orga- nization of secreted packets in detail. The internal struc- ture of lamellar bodies remains unchanged after secretion. SG, stratum granulosum. X 63,000.
Fig. 4 Freeze-fracture replicas of neonatal mouse epi- dermis. Level corresponds to figure 3. Lamellar body limit- ing membrane fuses with plasma membrane (single arrow), and contents are expelled into the intercellular space (dou- ble arrows). Bundles of tonofilaments (tfl are in the cyto- plasm near to the E faces of desmosomes (D). Insert depicts internal structure of a fractured lamellar body revealing packing of discs into several discrete arrays. SG, stratum granulosum. X 62,500. Insert X 55,000.
Figs. 5.6 Freeze-fracture replicas of adult mouse va- ginal epithelium. The fine structure of this epithelium is similar to epidermis and esophagus, though far fewer la- mellar bodies and correspondingly much less intercellular lamellar material is present. Figure 5 depicts a lamellar body during the process of expulsion, and figure 6 a later stage with lamellar material aggregated in the intercellu- lar space (arrows). Figure 5 X 86,000; figure 6 X 115,000.

MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 581

582 PETER M. ELIAS, N. SCOTT MCNUTT AND DANIEL S. FRIEND
may participate in the generation of trans- epithelial electrical gradients (Martinez- Palomo et al., '71). Discontinuous or isolated strands (maculae occludentes) also exist, but are relatively ineffective in that they do not fulfill an occlusive function. Only continuous, unbroken strands encircle the cell forming a complete seal. The relative inability of small ions and water to penetrate the interstices of certain epithelia generally correlates with the complexity of the fibrillar network of tight junctions between cells (Claude and Good- enough, '73), albeit this correlation is by no means invariable (e.g., Mollgard et al., '76).
RESULTS
Membrane structure in the stratum granulosum 1. Thin section and freeze-Facture observations
The structure of the stratum granulosum in the various epithelia we surveyed differed only slightly. In all tissues, the cytoplasm con- tains prominent keratohyalin jg-anules en- meshed in dense bundles of tonofilaments, as well as myriads of lamellar bodies (LB). As previously described (Matoltsy, ,651, these bodies are 0.1 to 0.3-p ovoid structures (figs. 1, 2) containing lamellated materials which are secreted into the intercellular spaces in the mid-to-upper part of the stratum granulosum (figs. 3-6). The internal structure of these or- ganelles and their extruded contents are best appreciated in tissues treated with Con A + HRP, which disclose variable numbers of short discs. In tissues so-treated, the homoge- nous matrix is selectively blackened, often causing both intracellular and secreted lamel- lar material to stand out in relief (fig. 3). As lamellar bodies secrete their contents, it ini- tially congregates within large, cup-shaped dilations of the intercellular space which dis- tend regions between desmosomes (fig. 3).
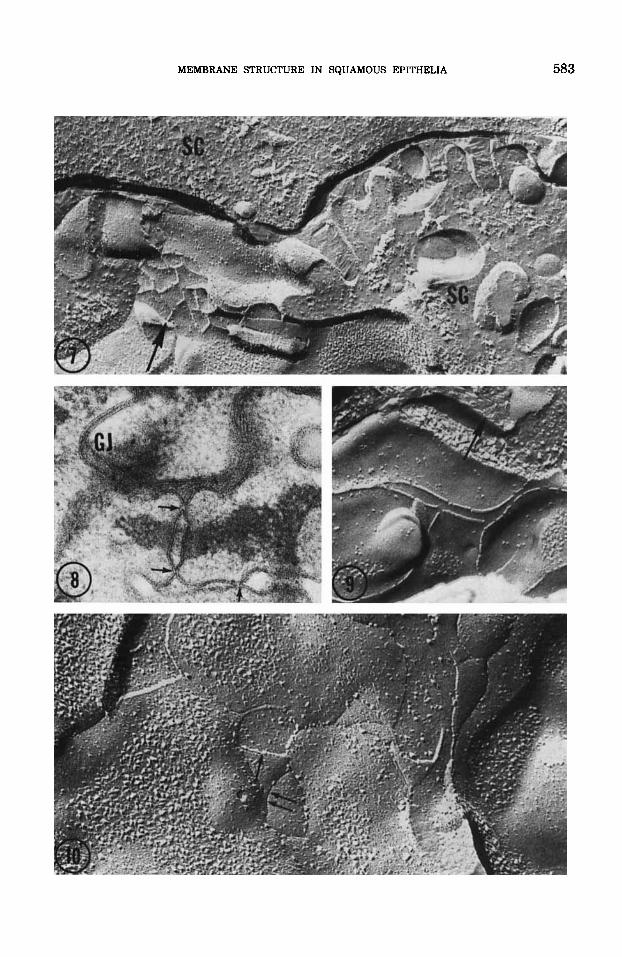
Freeze-fracture replicas of murine and human epidermis and esophagus as well as mouse vagina do not show complete occluding junctions. However, occasional replicas do exhibit incomplete and isolated networks con- sisting of one-to-four P-face strands or corre- sponding E-face grooves (figs. 7-10). Replicas of the esophagus and vagina (figs. 9-10] reveal these elements more frequently than do freeze-fractures of the epidermis (fig. 7). Such meshes, however, were always seen to be frag- mentary and often located immediately be- neath the secreted contents of lamellar bodies (figs. 7-91.
2. Tracer perfusion studies Both colloidal lanthanum and horseradish
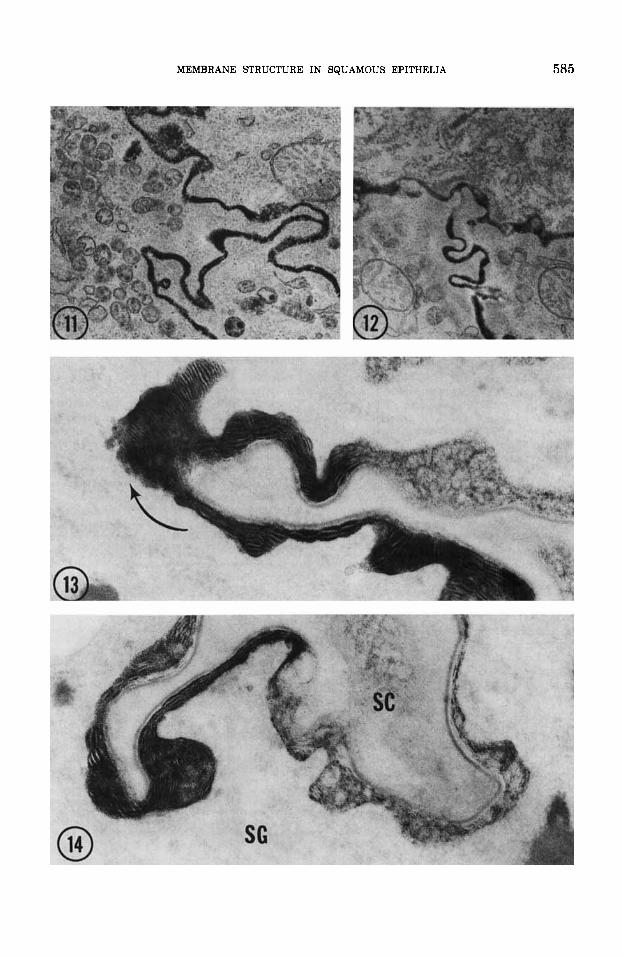
peroxidase filter outward to identical levels in the mid-to-upper stratum granulosum (figs. 11-13). In all the epithelia examined, tracer movement is blocked a t this level by the ex- truded contents of lamellar bodies (figs. 12, 13). Tight junctions are absent here. Colloidal lanthanum, the smallest of the tracers em- ployed, permeates gap junctions and the in- ter-discal matrix regions of secreted lamellar body contents for variable distances (fig. 13). In esophageal tissue, tracer sometimes pene- trates the most distal recesses of stratum granulosum interstices, even occasionally reaching the stratum corneum (fig. 143. No such intrusion occurs in epidermis, where tracer fails to reach the stratum corneum in all circumstances thus far tested.
Membrane alterations during transition from stratum granulosum to stratum corneum
(figs. 15-19). 1. Alterations in intercellular lamellar bodies
After secretion into the intercellular spac- es, lamellar bodies initially reside in packets corresponding in dimension and appearance to intracellular lamellar bodies (figs. 3, 6, 7). In freeze-fracture replicas, the newly-secreted lamellar body contents produce minimal deviations of the fracture plane into the in- tercellular space (figs. 4, 6, 7, 9, 15). How- ever during subsequent apical migration and
~ ~~ ~~~
Fig. 7 Freeze-fracture replicas of neonatal mouse epi- dermis. The interface of the stratum granulosum (SG) and the stratum corneum (SC) is depicted. Cross-fractured ar- rays of expelled intercellular lamellar body contents dis- tend the intercellular space. Immediately beneath this re- gion a small network of strands (arrow) is found. This type of image is extremely rare in epidermis, and when encoun- tered, such strands are rudimentary, never forming com- plete tight junctions, and are always situated immediately beneath expelled lamellar body contents. (cf. figs. 8 and 9). X 52,500.
Fig. 8 Thin sections of neonatal mouse epidermis. Ex- pelled lamellar body contents in the intercellular space is stained with Con A + HRP above three apparent mem- brane fusion points (arrows) in the mid-stratum gran- ulosum. Gap junctions (GJ) are plentiful at levels in all these keratinizing epithelia. X 60,000.
Fig. 9 Freeze-fracture replica of adult mouse esoph- agus. This image roughly corresponds to figures 7 and 8, de- picting focal strands immediately beneath expelled inter- cellular lamellar body material (arrow). X 82,500. Fig. 10 Freeze-fracture replica of adult mouse vagina.
Focal networks of strands (single arrow) and grooves (dou- ble arrow) are found more frequently in vagina than in epi- dermis. Again, these structures predominate in the mid-to- upper stratum granulosum immediately beneath regions of lamellar body expulsion. X 95,000.
MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 583

584 PETER M. ELIAS, N. SCOTT MCNUTT AND DANIEL S. FRIEND
transformation of granular cells into cornified cells, definite alterations are seen in both thin sections and replicas. At the granular-cor- nified cell layer interface and between the first and second layer of cornified cells, the in- tercellular discs coalesce into broad lamellae (figs. 15, 161, deviating the fracture plane from the cell membrane into the intercellular lipid sheets (Elias and Friend, '75). Transition images (short discs extending to broad lamel- lae) are infrequently encountered (fig. 16).
2. Unique features of membranes in transitional
The foregoing descriptions apply equally to esophagus, vagina, and epidermis. However, in human and murine epidermis, an additional feature is found a t the junction of the stratum granulosum and stratum corneum (figs. 17- 19). A network of strands, bearing superficial resemblance to tight junctions, emanates from plaques which correspond to E faces of desmosomes or often decorates E faces of cor- nified cell plasma membranes (figs. 17, 19). These strands are broad, flat and more serpen- tine than true tight junctional strands, and corresponding indentations or grooves in adja- cent leaflets are not observed.
Membrane structure in the stratum corneum Analysis of membrane structure in the
stratum corneum in routinely processed mate- rial is difficult. Routine processing solutions, which admirably preserve other tissues, often fail to penetrate or may selectively destroy the stratum corneum, conveying the false impression that this layer consists of loosely adherent squames. Only well-fixed specimens reveal the stratum corneum to be a cohesive sheet of geometrically packed squames (Chris- tophers and Kligman, '64; MacKenzie, '69; Menton and Eisen, '70). Freeze-fracture re- veals a similar image (fig. 20).
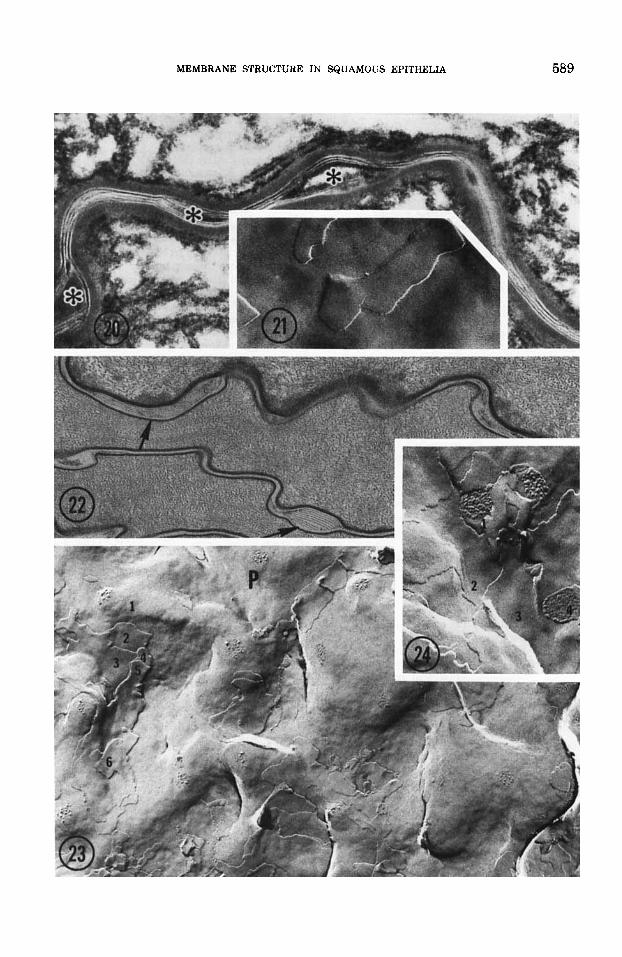
Both in well-fixed tissues and replicas the intercellular spaces of the stratum corneum appear to be filled with broad lamellar sheets (figs. 20-24). The sheets are more extensive in the epidermis than in the esophagus. Conse- quently, the plasma membranes of esophageal cornified cells are readily visualized in rep- licas (figs. 23, 241, but in epidermis, the plasma membrane is only rarely seen because the fracture plane usually remains in the in- tercellular space (fig. 21) (Elias and Friend, '75).
regions of epidermis
1. Plasma membrane structure of cornified cells A trilaminar image of the plasma mem-
brane is often difficult to see in thin sections of the stratum corneum (figs. 20, 22). Often the cytoplasmic dense zone hides the mem- brane's inner leaflet. Moreover, dense inter- cellular lamellae may equally obscure the outer membrane leaflet (fig. 20). However, all leaflets of the plasma membrane can be seen in replicas and thin sections after brief treat- ment of tissues with solvents (Elias et al., '77). During progression from the inner to the outer cornified cell layer in the esophagus and va- gina, the number of desmosomal aggregates and random intramembranous particles grad- ually diminishes. In the upper stratum cor- neum, plasma membrane fracture faces are devoid of membrane-associated particles and junctional structures (figs. 23, 24). No gap or tight junctions have been found in the stra- tum corneum.
2. Intercellular regions of the stratum corneum Thin sections and freeze-fracture replicas of
epidermis reveal two to six, 40-60 A smooth la- mellar sheets between cornified cells. Com- paratively, lamellae in esophagus and vagina are not as extensive (cf. figs. 20 and 21 with 22-24). In the former, groups of five or six la- mellae are segregated into ballooned inter- cellular regions interrupted by narrower re- gions where the intercellular spaces contain only one to three lamellae (fig. 22). In epider- mis, a minimum of four lamellae span the in- tercellular space (fig. 20). In all the epithelia, lamellar sheets are interrupted by dilatations filled with non-lamellar electron dense, homo- geneous or flocculent material (fig. 20).
In vitro modulations Despite the overall similarity of esophageal,
vaginal, and epidermal membrane structure, Figs. 11,12 Thin section of neonatal mouse epidermis.
Animal was injected 20 minutes earlier with horseradish peroxidase which percolates upward through the stratum granulosum until further egress ceases where lamellar bcdy contents engorge the intercellular space Wig. 12). Fix- ation: osmium tetroxide-potassium ferrocyanide. Figures 11 and 12 X 45,000.
Figs. 13,14 Thin sections of adult mouse esophagus. Lanthanum applied with fixative (glutaraldehyde plus picric acid) percolates upward to the mid-stratum gran- ulosum and permeates discs lying within the intercellular spaces for variable distances until egress ceases (fig. 13). Rarely, lanthanum appears to reach the interface of the stratum granulosum (SG) and stratum corneum (SO. Figure 13 X 60,000; figure 14 X 90,000.
MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 585

586 PETER M. ELIAS, N. SCOTT MCNUTT AND DANIEL S. FRIEND
the number of tight junction elements in esophagus and vagina exceeds that in epider- mis. Since the luminal surface of mucosal ke- ratinizing epithelia faces a more moist envi- ronment than the epidermis, we wondered if a more humid milieu might promote the expres- sion of true tight junctions. Therefore, we in- cubated pieces of epidermis for variable pe- riods of time in organ culture in a humid enclosure. In several parallel experiments ret- inoic acid was added to some organ culture media (Elias and Friend, '76).
In all specimens, either with or without added vitamin A, an increase in tight-junc- tional elements is apparent within six hours (fig. 25). Tight junction proliferation occurred only in the mid-to-upper stratum granulosum, i.e., in sites where the few pre-existing strands are located, and by 48 to 72 hours, belts or net- works resembling true zonulae occludentes are common (fig. 25). The increase in tight junction elements is accompanied by a dimin- ution in lamellar bodies and their secreted contents. In some replicas of the granular-cor- nified layer interface, forming focal junction- al strands appear to transect sheets of inter- cellular lamellar material (fig. 26). Except for the increased number of tight-junctional ele- ments, no other changes in junctional struc- tures are noted during these incubations. Addition of retinoic acid to organ cultures fails to enhance or hasten tight or gap junc- tion generation, as it does in embryonic chicken skin (Elias and Friend, '76).
Occasional replicas of 48 and 72 hour reti- noic acid-treated cultures displayed a unique feature in the stratum granulosum: three to six parallel arrays of crystalline rodlets tra- verse the nuclear envelope stopping abruptly without register to adjacent intramembra- nous particles and without impinging on nu- clear pores (fig. 27). The arrays apparently traverse the perinuclear space, since comple- mentary patterns appear on both of the envelopes (fig. 28). Similar structures were not encountered a t other levels in the epider- mis, in control cultures, or in cultures incu- bated for less than 48 hours.
DISCUSSION
Unique aspects of membrane structure in keratinizing layers of
squamous epithelia The present study describes striking dif-
ferences between the membranes and special- ized junctions of the stratum granulosum and
stratum corneum, in comparison with the membranes and junctions of the stratum basale and stratum spinosum. Although the plasma membranes of basal and spinous cells possess the same features (Breathnach et al., '73) as membranes in other epithelia (McNutt and Weinstein, '731, they change drastically during cornification: First, gap junctions do not persist into the stratum corneum; second, tight-junctional strands (occasionally en- countered in these epithelia a t the level of the mid-stratum granulosum) disappear during transformation of granular cells into cornified cells; and, third, an additional dramatic dif- ference is the progressive depletion of mem- brane-associated particles and desmosomes within the plasma membranes during apical migration into the stratum corneum.
1. Role of focal strands Contrary to interpretations by other work-
ers (Hashimoto, '71; Caputo, '75; Shimono and Clementi, '761, the incomplete and focal na- ture of tight junction elements in stratified squamous epithelia undermines a role for them as a significant barrier to water loss. However, one might speculate that these strands have functions different from those related purely to barrier function. For exam- ple, incomplete collections of strands are fre- quently found in association with gap junc- tional elements during development (Decker and Friend, '75; Elias and Friend, '76). In- complete occluding junctions may segregate
Fig. 15 Freeze-fracture replica of neonatal mouse epi- dermis. During transition from stratum granulosum (SG) to stratum corneum (SC) the fracture pattern of intercellu- lar material suddenly shifts. In the SG the fracture plane remains in the plasma membrane, cross-fracturing the in- tercellular lamellar contents. However, in the SC the frac- ture plane detours to the intercellular space (Eli- and Friend, '75) revealing broad, smooth sheets. X 40,000.
Fig. 16 Freeze-fracture replica of neonatal mouse epi- dermis. The shift in fracture pattern of intercellular la- mellar material from cross-fractures to broad sheets usual- ly occurs suddently, but in this exceptional instance, lamel- lar material is partially cross-fractured, while spreading into broad sheets a t the interface of the two layers. X 50,000.
Figs. 17-19 Freeze-fracture replicas of adult mouse (fig. 17), adult human (fig. 18), and neonatal mouse (fig. 19) epidermis. In epidermis, but not in the other keratinizing epithelia, a t interface of the stratum granulosum and stratum corneum, serpiginous strands are occasionally encountered. These rivulets emanate from E face images of desmosomes and extend serpiginously for variable dis. tances, sometimes anastomosing with other strands to form a network (fig. 17). Corresponding grooves have not been identified. Figure 17 X 52,500; figure 18 X 50,000; figure 19 X 52,500.

MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 587

588 PETER M. ELIAS, N. SCOTT MCNUTT AND DANIEL S. FRIEND
the enzymatic and particle contents of por- tions of the membranes of cells by presumably limiting membrane fluidity within distinct regions as recently suggested (Pisam and Ripoche, '76). Moreover, focal strands may simply represent deteriorating, "diluted," or forming tight junctions (Revel et al., '73; Elias and Friend, '76). Finally, focal strands are elaborated without any obvious relation to permeability phenomena following treatment with proteolytic enzymes (Orci et al., '76; Shimono and Clementi, '77).
What might the function of focal strands be within keratinizing stratified squamous epi- thelia? First, these elements might form a focal physical barrier which limits backwards movement of (some) secreted lamellar body contents, thus forcing persistence of these or- ganelles into the stratum corneum. Their loca- tion, immediately subjacent to sites of lamel- lar body deposition in the intercellular space, supports this possibility. On the other hand, since the contents of granular cells are largely recycled prior to cornification (Cutright and Bauer, '671, too complete a barrier might pre- vent back-diffusion and reutilization of essen- tial metabolites. Considered in this light, a focal, physical barrier, such as these strands might form, but not a permeability barrier, would be a desirable feature. Secondly, these tight-junctional elements might simply be vestigial expressions of epidermal differentia- tion in lower vertebrates. For example, in frog skin tight junctions are essential for the maintenance of an electrochemical gradient (Martinez-Palomo et al., '71). A final possi- bility is that the occasional focal junctions encountered represent a response to hydration of the overlying stratum corneum. Two obser- vations in this and two other recent studies support this possibility. First, more tight junc- tion strands are found in the relatively hy- drated squamous epithelia, e.g., esophagus and oral mucosa (McNutt, '73; Shimono and Clementi, '76) than in epidermis (Elias and Friend, '75). Secondly, in this study we have described substantial increases in focal strands after even short exposures of epider- mis to a higher humidity in an organ culture environment. However, in vitro influences other than humidity also could account for these changes. Whether these alterations are reversible, or whether hydration in vivo will also induce similar modulations, remains to be determined.
2. Intercellular lamellae of the stratum corneum
A characteristic feature of all the keratiniz- ing epithelia studied is the system of parallel lamellar sheets which fills the intercellular spaces of the stratum corneum. In epidermis the sheets spread in broad bilayers through substantial intercellular domains completely obscuring the plasma membrane in most rep- licas. In contrast, such sheets are far less ex- tensive in esophagus and vagina, revealing typical membrane particles and desmosomes between discontinuities in the lamellae.
While the specific composition of these sheets is still not known, they appear to con- tain neutral lipid (Elias et al., '77) in suffi- cient quantities to induce deviation of the fracture plane to the intercellular space (Elias and Friend, '75). Biochemical analysis of iso- lated stratum corneum lipids also reveals a preponderance of neutral lipids (Kooyman, '33; Long, '73; Gray and Yardley, '75). Fur- thermore, a variety of histochemical tech- niques suggest that during cornification, lip- ids become segregated in membrane regions, and, that the material in these regions stains as neutral lipid (Elias et al., '77). This con- stellation of findings explains both the unique intercellular fracture plane, as well as the in- creased susceptibility of these regions to ex- traction during routine tissue processing.
Relation of lamellar body secretion to formation of broad sheets in stratum corneum
Although this study and previous studies
Figs. 20.21 Corresponding thin section and freeze- fracture replica of neonatal mouse epidermis. The inter- cellular spaces of the stratum corneum are filled with broad lamellar sheets of alternating electron-density and electron-lucency, The electron-dense lamina periodically bulges into lenticular clefts (*). Because the fracture plane in epidermis remains within the intercellular space only smooth, particle-free lamellae are seen, and the true plasma membrane is obscured. Tissue in figure 20 fixed by osmium vapor alone. Figure 20 X 47,000; figure 21 X 59,500.
Figs. 22-24 Corresponding thin section (fig. 22) and freeze-fracture replicas of adult mouse esophagus. In con- trast to epidermis, esophageal interstices contain shorter arrays of parallel laminae (fig. 22, arrows). Because lamel- lar material is more sparse, the fracture plane alternates between the plasma membrane and the intercellular apace. In figure 23 as many 88 five intercellular sheets overlie the P face, while in figure 24, at least two intercellular sheets separate P and E faces. Note corresponding P and E face images of two desmosomes. Fixation in figure 22: glutaral- dehyde-picric acid. Figure 22 x 52,000; figure 23 X 42,500; figure 24 X 67,500.
MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 589

590 PETER M. ELIAS. N. SCOTT MCNUTT AND DANIEL S. FRIEND
(Elias and Friend, '75; Lavker, '76), link la- mellar body secretion with the formation of broad sheets in the stratum corneum, it is clear that lamellar body contents undergo drastic structural and chemical changes prior to their eventual reorganization within the in- terstices of the stratum corneum. While still residing within the stratum granulosum, la- mellar bodies produce minimal deviations of the fracture plane in contrast to the extensive sheets of intercellular material fractured in stratum corneum. Furthermore, initially se- creted lamellar bodies contain polar lipids, but little neutral lipid, in contrast to the broad, presumably neutral lipid-rich sheets in cor- nified layers (Elias, '77). This shift in composi- tion in turn probably explains the alteration in freeze-fracture behavior. Further support for this contention is provided both by studies of freeze-fractured model lipids (e.g., Tocanne et al., '74) and images of fractured tissue neu- tral lipids (e.g., Ruska and Ruska, '71); i.e., polar lipids tend to cross-fracture, while neu- tral lipids fracture in broad lamellae.
Nature of strands at interface of stratum granulosum and stratum corneum
An unusual type of structure, bearing some resemblance to tight junctions, occurs a t the interface of the stratum granulosum and stratum corneum in both murine and human epidermis. These structures form either ser- piginous strands, or resemble "crows" feet indentations within the E fracture face (fig. 26). Although these strands have mistakenly been termed tight junctions in a t least one study (Caputo, '75), they can be distinguished from true tight-junctional elements by the following criteria: (1) the elevations are on the E rather than the P face; (2) the networks emanate from E face images of desmosomes (fig. 19); (3) the strands are serpiginous, rather than displayed in rigid, branching, and anastomosing networks; and (4) grooves on adjacent fracture faces, which would corre- spond to the strands, are missing. While pure- ly conjectural we feel that these alterations are either due to plastic deformation of the E face leaflet (Bullivant, '741, or representative of alterations in membrane lipid composition which may accompany changes in intercellu- lar lipids. The barrier within keratinizing epithelia: level
and subcellular locations I t is well established that the stratum cor-
neum forms an effective barrier to the per- cutaneous movement of water and other polar substances (Scheuplein and Blank, '71). How- ever, tracer studies (Schreiner and Wolff, '69; Squier, '73; Elias and Friend, '7.9, including this one, indicate that a t least in epidermis, the barrier to larger water-soluble tracer egress is already functional a t the level of the stratum granulosum. Whether these studies will ultimately reflect the route of egress of waterper se, remains to be determined. Inter- estingly, tracer did appear to reach the stra- tum corneum more readily in esophagus than in epidermis. If technical differences are not responsible for this disparity (tracer was ap- plied with fixative in esophagus and prior to fixative in epidermis), this observation may reflect the known porosity of extracutaneous, mucosal keratinizing epithelia in comparison to epidermis (Galey et al., '76). The structural basis for this difference may well reside in the observation reported here of less extensive in- tercellular lamellae in esophagus as opposed to epidermis. Furthermore, tracers also reach the stratum corneum in essential fatty acid deficient mice, who demonstrate markedly in- creased transepidermal water loss in associa- tion with less extensive intercellular lamellar material (Elias et al., '77). Thus, the quantita- tive relationship of intercellular lamellae with the level of tracer permeation further underscores the critical role of lamellar bodies in the formation of the permeability barrier in keratinizing epithelia.
With regard to inward penetration across the stratum corneum, the role of the cornified cell versus the intercellular compartment in transport phenomena is also in dispute. De-
Fig. 25 Freeze-fracture replica of neonatal mouse epi- dermis incubated for 48 hours. During short-term organ cultures of epidermis, typical tight junctions consisting of multiple anastoming strands begin to form as early as six hours in vitro, while lamellar body material diminishes in quantity (not illustrated). Figure 25 X 80,000.
Fig. 26 Freeze-fracture replica of neonatal mouse epi- dermis incubated for 72 hours. In this specimen forming tight junction elements (arrow) appear to transect a layer of intercellular lamellar material (ICS) which separates the P and E faces. X 60,000.
Figs. 27,28 Freeze-fracture replica of neonatal mouse epidermis incubated with retinoic acid for 72 hours. Peculiar arrays of parallel, crystalline rods or sheets tra- verse the nuclear envelope for variable distances without impinging on nuclear pores (fig. 27). Note that these struc- tures span the space separating the P and E faces of the nu- clear envelopes (fig. 28). These structures were found only in granular layer nuclei, and only in vitamin A-treated cul- tures. Figure 27 X 45,000; figure 28a X 80,000; figure 28b x 80,000.

MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 59 1

592 PETER M. ELIAS, N. SCOTT MCNU’IT AND DANIEL S. FRIEND
spite mathematical models supporting trans- cellular movement (Scheuplein and Blank, ’71), available evidence from tracer experi- ments supports an intercellular route: In in vitro perfusion experiments large, water-solu- ble substances were effectively excluded from normal stratum corneum, penetrating neither cells nor interstices (Elias and Friend, ’74). However, after either solvent- or detergent treatment of epidermis, which are known to destroy barrier function (Scheuplein and Blank, ’71), the same tracers percolate through regions of cellular detachment; the interiors of even the most severely injured cells are not penetrated (Elias and Friend, ’74). These studies suggest that the intercellu- lar route, though normally a formidable im- pediment to ingress of polar substances, is, nevertheless, the path of least resistance in damaged epidermis. Finally, in light of its pre- sumably lipid-rich contents, the intercellular route would appear to be the most likely path- way for the known, selective movement of hydrophobic substances.
ACKNOWLEDGMENTS
The technical assistance of Barbara Brown, Diana Cohen, and Irene Rudolph is greatly ac- knowledged. This study has been supported in part by NIH AM-19098, NIH HD-10445, NCI CA-16482, and VA institutional research funds.
LITERATURE CITED
Branton, D. 1966 Fracture faces of frozen membranes. Proc. Nat. Acad. Sci. (U.S.A.), 55: 1048-1056.
Branton, D., S. Bullivant and N. B. Gilula 1976 Freeze-etch nomenclature. Science, 109: 54-56.
Breathnach, A. S., T. Goodman, C. Stolinski and M. Gross 1973 Freeze-fracture replication of cells of stratum cor- neum of human epidermis. J. Anat., 114: 65-81.
Breathnach, A. S., C. Stolinski and M. Gross 1973 Ultra- structure of foetal and post-natal human skin as re- vealed by the freeze-fracture technique. Micron, 3: 287-304.
Bullivant, 5. 1974 Freeze-etching techniques applied to biological membranes. Phil. Trans. Roy. Soc. London (B), 268: 5-14.
Caputo, R., D. Peluchetti and M. Monti 1975 The junctions of normal human epidermis: a freeze-fracture study. J. Invest. Derm., 64: 282 (Abstract).
Christophers, E., and A. M. Kligman 1964 Visualization of the cell layers of the stratum corneum. J. Invest. Derm., 42: 407-409.
Claude, P., and D. A. Goodenough 1973 Fracture faces of zonulae occludentes from “tight” and “leaky” epithelia. J. Cell Biol., 58: 390-400.
Cutright, D. E., and H. Bauer 1967 Cell renewal in the oral mucosa and skin of the rat. 11. DNA conservation and reutilization during the renewal cycle in vivo and in tis- sue explants. Oral Surg., 23: 260.271.
Decker, R. S., and D. S. Friend 1974 Assembly of gap junc- tions during amphibian neurulation. J. Cell Biol., 62: 32-47.
Elias, P. M., R. Cunico, H. I. Maibach and B. E. Brown 1977 Increaaed transepidermal water loss correlates with de- creased intercellular lipid in essential fatty acid deficient mice. Clin. Res., 25: 281a.
Elias, P. M., and D. S. Friend 1974 Permeability barriers and pathways in mammalian epidermis. J. Cell Biol., 63: 93a.
1975 The permeability barrier in mammalian epidermis. J. Cell Biol., 65: 185-191.
1976 Vitamin A-induced mucous metaplasia: An in vitro system for modulating tight and gap junction differentiation. J. Cell Biol., 68: 173-188.
Elias, P. M., J. Goerke and D. S. Friend 1977 Permeability barrier lipids: Composition and influence on epidermal structure and function. J. Invest. Derm., in press.
Farquhar, M. G., and G. E. Palade 1964 Functional orga- nization of amphibian skin. Proc. Natl. Acad. Sci., 51: 569-577.
Friend, D. S., and B. Gilula 1972 Variation in tight and gap junctions in mammalian tissues. J. Cell Biol., 53: 758-776.
Galey, W. R., H. K. Lonsdale and S. Nocht 1976 The in vitro permeability of skin and basal mucosa to selected drugs and tritiated water. J. Invest. Derm., 67: 713.717.
Gilula, N. B. 1974 Junctions between cells. In: Cell Com- munication. R. P. Cox, ed. John Wiley & Sons, New York, pp. 1-29.
Graham, R. C., Jr., and M. J. Karnovsky 1966 The early stages of absorption of injected horseradish peroxidase in the proximal tubules of mouse kidney: Ultrastructural cytochemistry by a new technique. J. Histochem. Cyto- chem., 14: 291-302.
Gray, G. M., and H. J. Yardley 1975 Different populations of pig epidermal cells: isolation and lipid composition. J. Lipid Res., 16: 441-450.
Hashimoto, K. 1971 Intercellular spaces of the human epidermis as demonstrated with lanthanum. J. Invest. Derm., 57: 17-31.
Huet, C., and M. Herzberg 1973 Effects of enzymes and EDTA or ruthenium red and concanavalin A labeling of the cell surface. J. Ultrastr. Res., 42: 186-199.
Ito, S., and M. J. Karnovsky 1968 Formaldehyde-glutaral- dehyde fixatives containing trinitro compounds. J. Cell Biol., 39: 168a.
Karnovsky, M. J. 1971 Use of ferrocyanide-reduced osmium tetroxide in electron microscopy. Roc. 11th Meeting Am. Soc. Cell Biol., p. 146a.
1932 Lipids of the skin: some changes in the lipids of the epidermis during keratinization. Arch. Derm. Symph., 25: 444-450.
Lavker, R. S. 1976 Membrane coating granules: the fate of the discharged lamellae. J. Ultrastr. Res., 55: 79-86.
Long, V. J. W. 1970 Variations in lipid composition at different depths in the cow snout epidermis. J. Invest. Derm., 55: 269-273.
Martinez-Palomo, A., D. Erlij and H. Bracho 1971 Localiza- tion of permeability barriers in frog skin epidermis. J. Cell Biol., 50: 277-287.
Matoltsy, A. G., and P. E. Parakkal 1965 Membrane-coat- ing granules of keratinizing epithelia. J. Cell Biol., 24: 297-307.
McNutt, N. S. 1973 Freeze-etch study of plasma mem- brane differentiation and junction in a keratinized stratified squamous epithelium. J. Cell Biol., 59: 21Oa.
McNutt, N. S., and R. S. Weinstein 1973 Membrane ultra- structure a t mammalian intercellular junctions. Progr. Biophys. Mol. Biol., 26: 45-101.
Kooyman, D. J.

MEMBRANE STRUCTURE IN SQUAMOUS EPITHELIA 593
Menton, D., and A. Z. Eisen 1971 Structure and organiza- tion of mammalian stratum corneum. J. Ultrastr. Res., 35: 247-264.
Mollgard, K., D. N. Malinowski and N. R. Saunders 1976 Lack of correlation between tight junction morphology and permeability properties in developing choroid plexus. Nature, 264: 293-294.
Orci, L. 1976 The microanatomy of the islets of Langer- hans. Metabolism, 25: 1303-1313.
Pisam, M., and P. Ripoche 1976 Redistribution of surface micromolecules in dissociated epithelial cells. J. Cell Biol., 21: 907-920.
Revel, J.-P., and M. J. Karnovsky 1967 Hexagonal arrays of subunits in intercellular junctions of mouse heart and liver. J. Cell Biol., 35: 302-317.
Revel, J.-P., P. Yip and L. L. Chang 1973 Cell junction in the early chick embryo - a freeze etch study. Dev. Biol., 35: 302-317.
Ricardo-Martinez, I., Jr., and A. Peters 1971 Membrane coating granules and membrane modifications in keratin- izing epithelia. Am. J. Anat., 130: 93-120.
Ruska, V. C., and H. Ruska 1971 Beobachtungen hber Strukturem in zellularen Neutralfetten bei der Dar- stellung durch die GeFrieratztechnik. In: Ergebnisse der Hochvakuventech n ik und der Physik dummer Schichten. S. Auwarter, ed. Wissenschaftliche Verlags- gessel., Stuttgart, pp. 59-80.
Scheuplein, R. J., and I. H. Blank 1971 Permeability of the skin. Physiol. Rev., 51: 702-747.
Schreiner, E., and K. Wolff 1969 Die Permeabilitat des epi- dermalen intercellularraumes fur kleinmolekulares pro- teinen. Arch. Klin. Exp. Derm., 235: 78-88.
Shimono, M., and F. Clementi 1976 Intercellular junctions of oral epithelium. I. Studies with freeze-fracture and tracing methods of normal rat keratinized oral epithe- lium. J. Ultrastruc. Res., 56: 121-136.
Intercellular junctions of oral epithelium. 11. Ultrastructural changes in rat buccal epithelium induced by trypsin digestion. J. Ultrastruc. Res.,59: 101- 112.
Spurr, C. A. 1973 A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruc. Res., 26:
Squier, C. A. 1973 The permeability of keratinized and non-keratinized oral epithelium to horseradish peroxi- dase. J. Ultrastruc. Res., 43: 160-177.
Staehelin, L. A. 1974 The structure and function of in- tercellular junctions. Int. Rev. Cytol., 39: 191-283.
Tilney, L. G., J. Bryan and 0. J. Bush 1973 Microtubules: evidence for 13 protofilaments. J. Cell Biol., 59:
Tocanne, J. F., P. H. J. Th. Ververgaert, A. J. Verkleij and L. L. M. van Deenan 1974 Chem. Phys. Lipids, 12:
1977
31-43.
267-275.
201-219.
Related Documents











