Molecular Basis for Pacemaker Cells in Epithelia* Received for publication, September 24, 2001, and in revised form, February 11, 2002 Published, JBC Papers in Press, February 15, 2002, DOI 10.1074/jbc.M109207200 M. Fatima Leite‡, Keiji Hirata§, Thomas Pusl§, Angela D. Burgstahler§, Keisuke Okazaki¶, J. Miguel Ortega, Alfredo M. Goes, Marco A. M. Prado**, David C. Spray‡‡, and Michael H. Nathanson§ §§ From the ‡Department of Physiology and Biophysics, Universidade Federal de Minas Gerais (UFMG), 31270-901 Belo Horizonte, Brazil, the §Departments of Medicine and Cell Biology, Yale University, New Haven, Connecticut 06520-8019, the ¶Department of Surgery, University of Occupational and Environmental Health, Kitakyushu, 8078555, Japan, the Department of Biochemistry and Immunology, UFMG, Belo Horizonte, Brazil, the **Department of Pharmacology, UFMG, Belo Horizonte, Brazil, and the ‡‡Department of Neuroscience, Albert Einstein College of Medicine, Bronx, New York 10461 Intercellular signaling is highly coordinated in excit- able tissues such as heart, but the organization of inter- cellular signaling in epithelia is less clear. We examined Ca 2 signaling in hepatoma cells expressing the hepato- cyte gap junction protein connexin32 (cx32) or the car- diac gap junction protein cx43, plus a fluorescently tagged V 1a vasopressin receptor (V 1a R). Release of ino- sitol 1,4,5-trisphosphate (InsP 3 ) in wild type cells in- creased Ca 2 in the injected cell but not in neighboring cells, while the Ca 2 signal spread to neighbors when gap junctions were expressed. Photorelease of caged Ca 2 rather than InsP 3 resulted in a small increase in Ca 2 that did not spread to neighbors with or without gap junctions. However, photorelease of Ca 2 in cells stimulated with low concentrations of vasopressin re- sulted in a much larger increase in Ca 2 , which spread to neighbors via gap junctions. Cells expressing tagged V 1a R similarly had increased sensitivity to vasopressin, and could signal to neighbors via gap junctions. Higher concentrations of vasopressin elicited Ca 2 signals in all cells. In cx32 or cx43 but not in wild type cells, this signaling was synchronized and began in cells express- ing the tagged V 1a R. Thus, intercellular Ca 2 signals in epithelia are organized by three factors: 1) InsP 3 must be generated in each cell to support a Ca 2 signal in that cell; 2) gap junctions are necessary to synchronize Ca 2 signals among cells; and 3) cells with relatively in- creased expression of hormone receptor will initiate Ca 2 signals and thus serve as pacemakers for their neighbors. Together, these factors may allow epithelia to act in an integrated, organ-level fashion rather than as a collection of isolated cells. Cells within excitable tissues such as the heart must act in a coordinated fashion to carry out organ-level functions such as muscle contraction to maintain blood flow. Excitation-contrac- tion coupling in the heart is coordinated by anatomically de- fined pacemaker cells, and is mediated by cytosolic Ca 2 sig- naling in individual myocytes. Intercellular Ca 2 waves can be observed in the intact heart (1), and these Ca 2 signaling patterns spread from cell to cell via gap junctions (2) and may relate to normal and abnormal cardiac function (1). In individ- ual myocytes, intracellular Ca 2 release occurs principally via the ryanodine receptor (RyR) 1 (3). Although RyR are spread throughout myocytes, focal clusters generate Ca 2 sparks that can initiate cell-wide Ca 2 signals (4 – 6). Intra- and intercel- lular Ca 2 signaling follows a different paradigm in non-excit- able cells (3). Ca 2 signaling in epithelial organs and in many other tissues instead occurs principally via inositol 1,4,5- trisphosphate (InsP 3 ) and the InsP 3 receptor (InsP 3 R) (3). As in the heart, cell-to-cell spread of Ca 2 waves has been observed in epithelial organs, including the liver (7–9) and salivary gland (10). Intercellular Ca 2 signaling follows a complex pat- tern in the liver, much as it does in the heart. For example, vasopressin induces Ca 2 waves that spread from pericentral to periportal hepatocytes, opposite to the direction of blood flow (7, 9), and this may help direct canalicular motility and bile flow (11). Altered intercellular Ca 2 signaling may contribute to the pathophysiology of certain disease states, since agents that block gap junction conductance also alter tissue function (12–14), and because certain cholestatic liver diseases are char- acterized by decreased expression of gap junctions and im- paired transmission of intercellular Ca 2 signals (15). Here we examined the conditions necessary to organize intercellular Ca 2 signals in a cell system in which Ca 2 signaling is medi- ated entirely via InsP 3 and the InsP 3 R. EXPERIMENTAL PROCEDURES Materials—[Arg 8 ]vasopressin, tetracycline, penicillin, and strepto- mycin were obtained from Sigma; fluo-3/AM, fluo-4/AM, fura-2/AM, rhodamine-conjugated phalloidin, and caged compounds (NPE-caged InsP 3 and DM-nitrophen-EDTA) were obtained from Molecular Probes (Eugene, OR). Dulbecco’s modified Eagle’s medium, Liebovitz 15 (L-15) medium, and other tissue culture reagents were from Invitrogen (Basel, Switzerland). All other chemicals were of the highest quality commer- cially available. Antibodies—InsP 3 receptors were labeled using an antibody from affinity-purified specific rabbit polyclonal antiserum directed against the 18 COOH-terminal residues of the rat type II InsP 3 receptor (16), which was kindly provided by Richard Wojcikiewicz (SUNY, Syracuse, NY). Vasopressin V 1a receptors were labeled using an affinity-purified polyclonal antibody directed against the rat hepatocyte V 1a receptor (7), which was kindly supplied by Carlos Gonzalez (Universidad Austral de * This work was supported by National Institutes of Health Grants DK45710, DK34989, DK57751, DK41918, RR04224, and TW01452 and an Established Investigator Grant from the American Heart Associa- tion. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. §§ To whom correspondence should be addressed: 333 Cedar St., Rm. 1080 LMP, Yale University School of Medicine, New Haven, CT 06520- 8019. Tel.: 203-737-6060; Fax: 203-785-4306; E-mail: michael.nathan- [email protected]. 1 The abbreviations used are: RyR, ryanodine receptor; InsP 3 , inositol 1,4,5-trisphosphate; InsP 3 R, inositol 1,4,5-trisphosphate receptor; cx32, connexin32; cx43, connexin43; CHO, Chinese hamster ovary; V 1a R, V 1a vasopressin receptor; RFP, red fluorescence protein. THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 18, Issue of May 3, pp. 16313–16323, 2002 © 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A. This paper is available on line at http://www.jbc.org 16313

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Basis for Pacemaker Cells in Epithelia*
Received for publication, September 24, 2001, and in revised form, February 11, 2002Published, JBC Papers in Press, February 15, 2002, DOI 10.1074/jbc.M109207200
M. Fatima Leite‡, Keiji Hirata§, Thomas Pusl§, Angela D. Burgstahler§, Keisuke Okazaki¶,J. Miguel Ortega�, Alfredo M. Goes�, Marco A. M. Prado**, David C. Spray‡‡,and Michael H. Nathanson§ §§
From the ‡Department of Physiology and Biophysics, Universidade Federal de Minas Gerais (UFMG), 31270-901 BeloHorizonte, Brazil, the §Departments of Medicine and Cell Biology, Yale University, New Haven, Connecticut 06520-8019,the ¶Department of Surgery, University of Occupational and Environmental Health, Kitakyushu, 8078555, Japan, the�Department of Biochemistry and Immunology, UFMG, Belo Horizonte, Brazil, the **Department of Pharmacology,UFMG, Belo Horizonte, Brazil, and the ‡‡Department of Neuroscience, Albert Einstein College of Medicine,Bronx, New York 10461
Intercellular signaling is highly coordinated in excit-able tissues such as heart, but the organization of inter-cellular signaling in epithelia is less clear. We examinedCa2� signaling in hepatoma cells expressing the hepato-cyte gap junction protein connexin32 (cx32) or the car-diac gap junction protein cx43, plus a fluorescentlytagged V1a vasopressin receptor (V1aR). Release of ino-sitol 1,4,5-trisphosphate (InsP3) in wild type cells in-creased Ca2� in the injected cell but not in neighboringcells, while the Ca2� signal spread to neighbors whengap junctions were expressed. Photorelease of cagedCa2� rather than InsP3 resulted in a small increase inCa2� that did not spread to neighbors with or withoutgap junctions. However, photorelease of Ca2� in cellsstimulated with low concentrations of vasopressin re-sulted in a much larger increase in Ca2�, which spreadto neighbors via gap junctions. Cells expressing taggedV1aR similarly had increased sensitivity to vasopressin,and could signal to neighbors via gap junctions. Higherconcentrations of vasopressin elicited Ca2� signals in allcells. In cx32 or cx43 but not in wild type cells, thissignaling was synchronized and began in cells express-ing the tagged V1aR. Thus, intercellular Ca2� signals inepithelia are organized by three factors: 1) InsP3 mustbe generated in each cell to support a Ca2� signal in thatcell; 2) gap junctions are necessary to synchronize Ca2�
signals among cells; and 3) cells with relatively in-creased expression of hormone receptor will initiateCa2� signals and thus serve as pacemakers for theirneighbors. Together, these factors may allow epitheliato act in an integrated, organ-level fashion rather thanas a collection of isolated cells.
Cells within excitable tissues such as the heart must act in acoordinated fashion to carry out organ-level functions such asmuscle contraction to maintain blood flow. Excitation-contrac-tion coupling in the heart is coordinated by anatomically de-fined pacemaker cells, and is mediated by cytosolic Ca2� sig-
naling in individual myocytes. Intercellular Ca2� waves can beobserved in the intact heart (1), and these Ca2� signalingpatterns spread from cell to cell via gap junctions (2) and mayrelate to normal and abnormal cardiac function (1). In individ-ual myocytes, intracellular Ca2� release occurs principally viathe ryanodine receptor (RyR)1 (3). Although RyR are spreadthroughout myocytes, focal clusters generate Ca2� sparks thatcan initiate cell-wide Ca2� signals (4–6). Intra- and intercel-lular Ca2� signaling follows a different paradigm in non-excit-able cells (3). Ca2� signaling in epithelial organs and in manyother tissues instead occurs principally via inositol 1,4,5-trisphosphate (InsP3) and the InsP3 receptor (InsP3R) (3). As inthe heart, cell-to-cell spread of Ca2� waves has been observedin epithelial organs, including the liver (7–9) and salivarygland (10). Intercellular Ca2� signaling follows a complex pat-tern in the liver, much as it does in the heart. For example,vasopressin induces Ca2� waves that spread from pericentralto periportal hepatocytes, opposite to the direction of blood flow(7, 9), and this may help direct canalicular motility and bileflow (11). Altered intercellular Ca2� signaling may contributeto the pathophysiology of certain disease states, since agentsthat block gap junction conductance also alter tissue function(12–14), and because certain cholestatic liver diseases are char-acterized by decreased expression of gap junctions and im-paired transmission of intercellular Ca2� signals (15). Here weexamined the conditions necessary to organize intercellularCa2� signals in a cell system in which Ca2� signaling is medi-ated entirely via InsP3 and the InsP3R.
EXPERIMENTAL PROCEDURES
Materials—[Arg8]vasopressin, tetracycline, penicillin, and strepto-mycin were obtained from Sigma; fluo-3/AM, fluo-4/AM, fura-2/AM,rhodamine-conjugated phalloidin, and caged compounds (NPE-cagedInsP3 and DM-nitrophen-EDTA) were obtained from Molecular Probes(Eugene, OR). Dulbecco’s modified Eagle’s medium, Liebovitz 15 (L-15)medium, and other tissue culture reagents were from Invitrogen (Basel,Switzerland). All other chemicals were of the highest quality commer-cially available.
Antibodies—InsP3 receptors were labeled using an antibody fromaffinity-purified specific rabbit polyclonal antiserum directed againstthe 18 COOH-terminal residues of the rat type II InsP3 receptor (16),which was kindly provided by Richard Wojcikiewicz (SUNY, Syracuse,NY). Vasopressin V1a receptors were labeled using an affinity-purifiedpolyclonal antibody directed against the rat hepatocyte V1a receptor (7),which was kindly supplied by Carlos Gonzalez (Universidad Austral de
* This work was supported by National Institutes of Health GrantsDK45710, DK34989, DK57751, DK41918, RR04224, and TW01452 andan Established Investigator Grant from the American Heart Associa-tion. The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
§§ To whom correspondence should be addressed: 333 Cedar St., Rm.1080 LMP, Yale University School of Medicine, New Haven, CT 06520-8019. Tel.: 203-737-6060; Fax: 203-785-4306; E-mail: [email protected].
1 The abbreviations used are: RyR, ryanodine receptor; InsP3, inositol1,4,5-trisphosphate; InsP3R, inositol 1,4,5-trisphosphate receptor; cx32,connexin32; cx43, connexin43; CHO, Chinese hamster ovary; V1aR, V1avasopressin receptor; RFP, red fluorescence protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 277, No. 18, Issue of May 3, pp. 16313–16323, 2002© 2002 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org 16313

Chile) and Juan Saez (Catholic University, Santiago). Commerciallyavailable monoclonal antibodies (Chemicon, Temecula, CA) were usedto label connexin32 (cx32) and connexin43 (cx43).
Cell Culture—Wild type SkHep1 cells, SkHep1 cells stably trans-fected with cx32 (17, 18) or cx43 (19), and CHO cells were cultured inDulbecco’s modified Eagle’s medium supplemented with 10% fetal calfserum and penicillin/streptomycin (100 �g/ml). Cells were plated ontoglass coverslips, and experiments were performed 2–3 days after plat-ing. Coverslips containing the cells were transferred to L-15 mediumsupplemented with 10% fetal calf serum 1 h before each experiment,then loaded with a fluorescent Ca2� indicator and studied as describedbelow. Expression of cx32 was under the control of a tetracycline-sensitive promoter (17), so tetracycline (1 �g/ml) was either added to orremoved from the medium 24 h before experiments using these cells.Primary cultures of rat hepatocytes were isolated from liver by colla-genase perfusion, then plated onto glass coverslips and used within 2–4h as described previously (20).
Immunoblot Analysis—Wild type and SkHep1 cells transfected withcx32 or cx43 were harvested, dissolved in SDS, and the protein concen-tration of cell homogenate was determined as described previously (21).Proteins collected from the cells were separated by SDS-polyacrylamidegel electrophoresis using a 10% polyacrylamide gel and subsequentlytransferred to protein nitrocellulose membranes. The membranes wereblocked at 4 °C overnight and probed for 2 h with the primary antibody.The membranes then were washed, incubated for 1 h with peroxidase-conjugated secondary antibody, and revealed by enhanced chemilumi-nescence (Amersham Bioscience, Arlington Heights, IL).
Reverse Transcription and PCR Amplification—Total RNA fromSkHep1 cells was isolated using Trizol reagent according to the manufac-turer (Invitrogen, Grand Island, NY). First strand cDNA was synthesizedusing the primer oligo-d(T)16 and Moloney murine leukemia virus-reversetranscriptase. Degenerate primers were used to amplify a 530-bp productfrom a portion of the 3� region common to the three known RyRs (22–24).PCR amplification was performed in a PTC-100 automated thermocycler(MJ Research, Watertown, MA) using 2 �l of the first-strand cDNAreaction, 200 nM of each degenerate primer, 200 �M dNTPs, 2.5 mM
MgCl2, and 2.5 units of AmpliTaq DNA polymerase for a total volume of100 �l. The PCR samples were subjected to a hot start (2 min at 94 °C),followed by 30 cycles of 45 s at 94 °C, 1 min at 50 °C, and 1 min at 72 °C.The reaction was followed by a final extension at 72 °C for 10 min. ThePCR product was analyzed by agarose gel electrophoresis.
V1aR-RFP Construction and Transfection—The gene encoding the V1a
vasopressin receptor (V1aR) was fused in-frame to the amino terminus ofred fluorescence protein (RFP) in a pDsRed1-N1 vector (CLONTECH,Palo Alto, CA). The V1aR open reading frame initially was amplified byPCR from a plasmid containing the full-length cDNA using oligonucleo-tide primers to introduce HindIII/Kozak consensus and BamHI sites atthe 5� and 3� end of the cDNA, respectively. A typical Kozak consensussequence (underlined) was introduced into the receptor coding sequence toimprove translation. The forward primer was 5�-GCGCAAGCTTGCCG-CCACCATGAGTTTCCCGCGAGGCTCC-3� and the reverse primer was5�-CGGTGGATCCCGGAATAAGAAGTCTGTCTTTCGGCTCATGC-3�.To generate the V1aR-RFP chimera construct, the resulting PCR productwas digested with HindIII and BamHI and ligated into the HindIII andBamHI site of pDsRed-N1 followed by transformation in Escherichia coliDH5�. Positive clones were identified by miniplasmid preparation andrestriction enzyme analysis. Selected clones were used for maxiplasmidpreparation and subsequent transfection. Subconfluent monolayers ofSkHep1 cells were transfected with 1 �g of pV1aR-RFP DNA using effec-tene (Qiagen, Valencia, CA) according to the manufacturer’s instructions.Cells were examined by time lapse confocal microscopy 48 h aftertransfection. To obtain a pV1aR construct expressing the V1a receptorwithout fusion to RFP, the pV1aR-RFP plasmid was digested with NotI,followed by gel purification of the linearized band for further digestionwith BamHI. The final digestion product was purified and the extremitieswere turned blunt through Klenow DNA polymerase treatment. Afterligation and E. coli DH5� transformation, positive clones were identifiedby loss of the BamHI restriction site. The selected clone was transfectedinto 5 � 105 CHO cells using effectene, and suspensions of fura-2-loadedcells were used for cytosolic Ca2� measurements 48 h after transfection.
Confocal Immunofluorescence Microscopy—Immunofluorescence wasperformed on SkHep1 cells 2–3 days after plating, and on isolated rat
FIG. 1. SkHep1 cells as a model for InsP3-mediated Ca2� sig-naling. a, the InsP3R is expressed in SkHep1 cells. Western analysisusing type II-specific InsP3R receptor antibody CT2 identifies a singleband of the same size in lysates from SkHep1 cells and the positivecontrol, rat liver (60 �g each). b, confocal immunofluorescence imagedemonstrates that the InsP3R is distributed in a punctate fashionthroughout the cytosol of SkHep1 cells. InsP3R were labeled with anti-body CT2 and counterstained with Alexa 488-tagged secondary anti-body, shown in green. The specimens were co-labeled with rhodaminephalloidin, shown in red, to identify the actin cytoskeleton and plasmamembrane of SkHep1 cells. Nonspecific staining by the secondary an-tibody was not evident (not shown). c, SkHep1 cells do not express RyR.Reverse transcriptase-PCR was used to amplify RyR from SkHep1 RNAusing degenerate primers that recognize all three RyR isoforms. Apositive band for actin was observed (lane 1), indicating the integrity ofthe SkHep1 RNA. No band was seen using RyR primers (lane 2) or in
RNA or DNA negative controls (lanes 3 and 4, respectively). d, a PCRproduct of 530 bp in heart RNA indicates the presence of RyR (positivecontrol). No band was seen in RNA or DNA negative controls (lanes 2and 3).
Epithelial Pacemakers16314

hepatocyte couplets 2–4 h after isolation. SkHep1 cells were fixed withacetone, blocked with phosphate-buffered saline containing 1% bovineserum albumin, then labeled with primary antibody for 1 h. SkHep1cells were labeled with anti-InsP3R antibodies to determine the subcel-lular distribution of InsP3Rs, or with anti-cx32 or anti-cx43 antibodiesto confirm that transfected SkHep1 cells expressed the intended gapjunction protein. Primary rat hepatocytes were fixed with Bouin’s fix-ative for labeling with anti-V1a receptor antibodies (7), but were other-wise processed similarly. After primary antibody incubation the cellswere rinsed with phosphate-buffered saline and incubated with Alexa488-conjugated secondary antibody (Molecular Probes). Specimenswere co-labeled with rhodamine-phalloidin to facilitate the identifica-tion of the plasma membrane. Negative controls were stained withsecondary antibodies alone, along with rhodamine-phalloidin. Speci-mens were examined using a Zeiss LSM 510 Laser Scanning ConfocalMicroscope equipped with a krypton/argon laser (Thornwood, NY). Toensure specificity of staining, images were obtained using confocalmachine settings at which no Alexa 488 fluorescence was detectable innegative control samples labeled with secondary antibodies alone. Spec-imens were serially excited at 488 nm and observed at 505–550 nm todetect Alexa 488, then excited at 568 nm and observed at �585 nm todetect rhodamine. This approach eliminated bleed-through of Alexa 488fluorescence into the longer wavelength (rhodamine) detection channel.
Cytosolic Ca2� Measurements—Cytosolic Ca2� was measured in wildtype SkHep1 cells and in SkHep1 stably transfected with cx32 or cx43using time lapse confocal microscopy (20, 25). Cells were incubated for 1 hat 37 °C with fluo-3/AM or fluo-4/AM (6 �M). Coverslips containing thecells were transferred to a custom-built perfusion chamber on the stage of
a Bio-Rad MRC-1024 confocal microscope (Bio-Rad, Hercules, CA) andobserved using a �63, 1.4 N.A. objective. The 488 nm line of a krypton/argon laser was used to excite the dye, and emission signals between 505and 550 nm were collected. In experiments in which some cells expressedV1aR-RFP, the 568-nm line of the laser was used to excite RFP, whileemission signals above 585 nm were collected. Using this approach, cellsexpressing the V1aR-RFP could be identified while Ca2� signaling in thoseand other cells could be monitored simultaneously. Neither autofluores-cence nor background signals were detectable at the machine settingsthat were used. Cells were stimulated either by perfusion with vasopres-sin, or else by flash photolysis (uncaging) of second messengers, and wereobserved at a rate of 2–10 frames/s. During two-photon flash photolysis,software constraints limited data collection to a rate of 1 frame/s.
Cytosolic Ca2� was measured in wild type CHO cells and in CHOcells transiently transfected with pV1aR or pV1aR-RFP using spectroflu-orometry (26). For these Ca2� measurements, CHO cells were loadedwith fura-2/AM (5 �M) for 1 h at 37 °C. Cell suspensions loaded withfura-2 were then transferred to a cuvette to monitor changes in thefura-2 fluorescence ratio in response to addition of vasopressin (100 nM),using an Hitachi F-2000 spectrofluorometer (Danbury, CT).
Flash Photolysis Studies of Caged InsP3 and Ca2�—The mechanicalstimulation associated with microinjection induces transient Ca2� sig-nals in epithelial cells (20), including in SkHep1 cells.2 Therefore, cellshere were loaded with a caged rather than the active form of InsP3 or
2 A. D. Burgstahler and Michael H. Nathanson, unpublishedobservation.
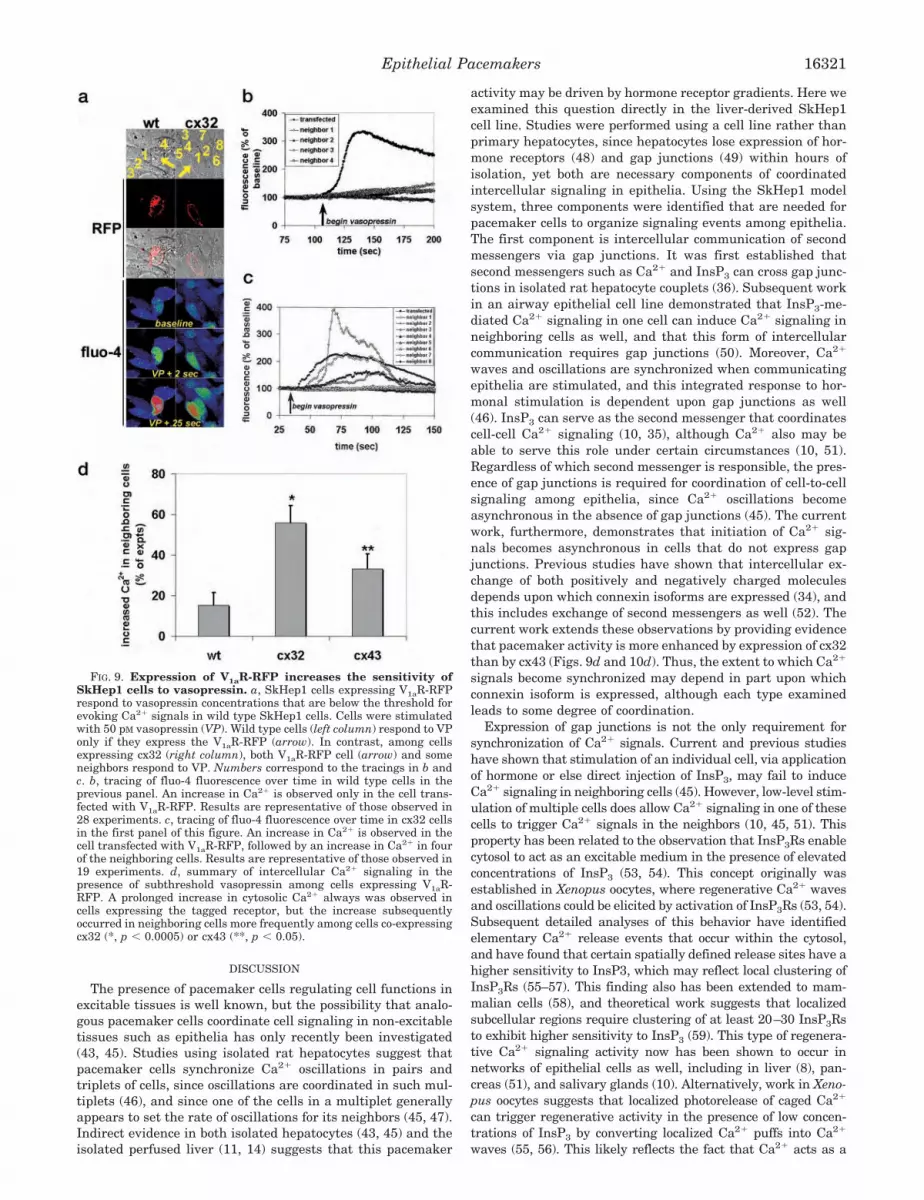
FIG. 2. Intercellular communication in wild type and transfected SkHep1 cells. a, inducible expression of cx32 in SkHep1 cells.Immunoblotting using cx32-specific antibodies identified a single band of the appropriate size. Expression occurs only in the absence of tetracycline, asdescribed previously (17). b, immunoblotting confirmed stable expression of cx43 in a separate SkHep1 cell line, as described previously (19). c, confocalimmunofluorescence confirmed localization of cx43 (shown in the micrograph) or cx32 (not shown) to cell-cell borders. Cx43 was labeled with amonoclonal antibody and counterstained with Alexa 488-tagged secondary antibody, shown in green. Cells were co-labeled with rhodamine phalloidin,shown in red, to identify cell-to-cell boundaries. d, expression of cx32 confers the ability to transfer Lucifer Yellow from cell to cell. Injection of LuciferYellow into a single cell (arrow) results in dye transfer to neighboring cells if cx32 is expressed (top row) but not if cx32 expression is suppressed (bottomrow). Lucifer Yellow fluorescence is detected by time lapse confocal microscopy. e, intercellular communication of Lucifer Yellow occurs only in SkHep1cells expressing cx32 or cx43. Lucifer Yellow (LY) fluorescence is detected only in the Lucifer Yellow-injected cell in both wild type (wt) and cx32 cellsincubated with tetracycline (cx32 � tet). Values are mean � S.E. (*, p � 10�7 relative to cx32 � tet or wt).
Epithelial Pacemakers 16315

FIG. 3. InsP3 initiates a Ca2� wave that spreads to neighboring SkHep1 cells via gap junctions. a, intercellular Ca2� signaling inSkHep1 cells expressing cx32. Caged InsP3 (1 mM) was co-injected with Texas Red into one of the cells (top panel), identified by confocal microscopy.Sequential confocal images of the Ca2� dye fluo-4 (bottom panels) identify increases in cytosolic Ca2� in the injected cell and its neighbors followingUV flash photolysis of the InsP3. Fluo-4 fluorescence in this and subsequent confocal images is pseudocolored according to the color scale shownbelow. b, tracing of the increases in fluo-4 fluorescence in the cells indicated in the previous panel. Ca2� signals spread to a similar number of cellsin each experiment (see panel f), despite variability in the percent increase in fluo-4 fluorescence among experiments. c, expanded time scale of theprevious tracing, which represents events from t � 34 to t � 38 s. Observe that an increase in Ca2� begins in the injected cell, then spreads to itsneighbors. d, Ca2� signaling in SkHep1 cells in which cx32 expression is suppressed by tetracycline. Caged InsP3 is injected into the cell indicated
Epithelial Pacemakers16316

Ca2�. Caged InsP3 was microinjected into cells, then uncaged by UVflash photolysis, while a cell-permeant form of caged Ca2� was loadedinto cells, then uncaged by two-photon flash photolysis. For InsP3
studies, NPE-InsP3 (1 mM) was dissolved in an intracellular-like buffer(150 mM KCl plus 1 mM Hepes), along with Texas Red (0.4 mg/ml) as amarker of successful microinjection. A series 5171 Eppendorf microma-nipulator (Westbury, NY) was used for positioning, and an Eppendorfseries 5242 microinjector was used for pressure microinjections. In-jected cells were allowed to recover for at least 5 min before flashphotolysis. NPE-InsP3 was photolyzed using a mercury arc lamp (75 W)coupled to a 1-mm quartz fiberoptic cable through a high-speed shutterand filterwheel (26). The flash duration varied from 100 to 200 ms, topermit maximal uncaging. For caged Ca2� experiments, cells wereloaded for 1 h with the Ca2� cage DM-nitrophen (2 �M) plus fluo-4/AM(6 �M). Ca2� was uncaged by two-photon flash photolysis using a Bio-Rad MRC 1024 confocal microscope (Hercules, CA) adapted for two-photon excitation with a Spectra-Physics Tsunami titanium:sapphirelaser and a Millenia X pump laser (Mountain View, CA). The titanium:sapphire laser was mode-locked with a pulse width of 90 fs and tuned toa wavelength of 730 nm, since that wavelength is optimal for two-photon release of Ca2� from DM-nitrophen (27). A pump laser power of7.2 W was used, which resulted in a power level of 600 mW exiting thetitanium:sapphire laser. The beam was passed through a 32% neutraldensity filter, and a power level of 2–4 mW was detected at the focalplane. The calculated volume of uncaging at the focal point was 0.02–0.03 fl, so caged Ca2� was photoreleased at multiple adjacent points toelevate cytosolic Ca2� in predetermined subcellular regions (28). Ca2�
signals induced by uncaged InsP3 or Ca2� were monitored by time lapseconfocal microscopy as described previously (25, 26).
Statistics—Values listed are mean � S.E., except where otherwisenoted. Statistical comparisons were made using Student’s t test, orpaired t test where appropriate.
RESULTS
Characterization of Ca2� Release Channels in SkHep1Cells—Expression of the InsP3R was examined in SkHep1 cellsby immunoblot (Fig. 1a). SkHep1 cells were found to expressthe type II InsP3R, which is the most prevalent isoform in liver(16, 29). The subcellular distribution of the InsP3 receptor wasinvestigated by confocal immunofluorescence. SkHep1 cellswere labeled with the InsP3R antibody used for immunoblots,and co-labeled with rhodamine phalloidin to identify the actincytoskeleton (Fig. 1b). The InsP3R was diffusely distributedthroughout the cytosol. No labeling was seen in negative con-trol tissues stained with secondary but not primary antibodies.In contrast, reverse transcriptase-PCR failed to detect RyRexpression in SkHep1 cells (Fig. 1, c and d). These findingsdemonstrate that the InsP3R is the only intracellular Ca2�
release channel in this cell type. This in turn suggests thatSkHep1 cells are a model system in which intracellular Ca2�
release is mediated entirely by InsP3Rs, and that Ca2� signal-ing in these cells would not be influenced by subcellular gradi-ents in InsP3R expression, as occur in some epithelia (30–33).
Intercellular Communication in SkHep1 Cells—The SkHep1hepatoma cell line normally is communication deficient, butwas stably transfected with cx32 under the control of a tetra-cycline-sensitive promoter (17) or with cx43 (19), as described
FIG. 4. Ca2� does not act as an intercellular messenger inunstimulated SkHep1 cells. a, serial confocal images of SkHep1 cx32cells before, during, and after Ca2� is uncaged in a single cell. All of thecells are loaded with the cell-permeant form of the Ca2� cage DM-Nitrophen (1 �M), but two-photon excitation is used to restrict theregion in which Ca2� is photoreleased to the (rectangular) area indi-cated. b, tracing of fluo-4 fluorescence in the cell subjected to two-photon flash photolysis, plus two neighboring cells. The increase inCa2� is restricted to the cell in which Ca2� is uncaged. Result isrepresentative of that seen in 29 separate experiments using cellsexpressing either cx32 or cx43.
FIG. 5. Ca2� does not act as a global intracellular messenger inunstimulated SkHep1 cells. a, illustration of the limited range overwhich free Ca2� travels in a single SkHep1 cell. Serial confocal imagesshow a SkHep1 cell before, during, and 240 ms after photorelease ofCa2� in region 1. Each square is 6 � 6 �m. b, tracing of fluo-4 fluores-cence in the regions of the cell shown in the previous panel. A smalltransient increase in Ca2� is detected in the region subjected to flashphotolysis, and the increase becomes progressively attenuated in re-gions farther from the flash site. Result is representative of that seen in8 cells.
by the arrow, then released by flash photolysis. Serial confocal images demonstrate an increase in Ca2� in the microinjected cell, which did notspread to neighboring cells. e, tracing of fluo-4 fluorescence in the cells shown in the previous panel. An increase in cytosolic Ca2� was detectedin the microinjected cell (filled circle) but not in neighboring cells (open symbols). f, summary of experiments using caged InsP3. InsP3-induced Ca2�
signals spread to neighboring SkHep1 cells expressing cx32 or cx43 but not in cx32 cells with tetracycline or wild type cells (*, p � 0.0001).
Epithelial Pacemakers 16317
previously. We verified expression levels of cx32 and cx43 inthese cell lines by immunoblot (Fig. 2, a and b). Single bandswere detected at 32 kDa in cx32-transfected cells and at 43 kDain cx43-transfected cells, and cx32 expression occurred only inthe absence of tetracycline. The subcellular location of cx32 andcx43 in these cells was verified by confocal immunofluorescence(Fig. 2c). Both connexin isoforms were detected in a punctatedistribution almost entirely along cell-to-cell borders, which isthe appropriate location for gap junctions (15). Cell-to-celltransmission of microinjected Lucifer Yellow was examined asa functional measure of gap junction expression in each trans-fected cell line (Fig. 2, d and e). This small fluorescent dye isbiologically inactive and is transmitted across cx32 and cx43gap junctions with similar efficacy (34). Lucifer Yellow injectedinto cx32 cells spread to 3.04 � 0.37 neighboring cells within 3min (mean � S.E.; n � 26), whereas Lucifer Yellow injectedinto cx43 cells spread to 3.80 � 0.45 neighboring cells duringthe same time interval (n � 40; p � 0.2). Lucifer Yellow injectedinto non-transfected (wild type) SkHep1 cells spread to no(0.0 � 0.0) neighboring cells (n � 30; p � 10�8 relative to bothcx32 and cx43 cells), while Lucifer Yellow injected into cx32cells in tetracycline spread to only 0.29 � 0.07 cells (n � 49; p �10�7 relative to cx32 cells without tetracycline). These studiesconfirm functional expression of cx32 and cx43 in SkHep1 cells,and suggest that intercellular communication is established innearly all of these cells.
Intercellular Signaling by InsP3 and Ca2� in UnstimulatedCells—NPE-caged InsP3 was injected into wild type and cx43-and cx32-transfected SkHep1 cells, then the InsP3 was liber-ated by UV flash photolysis. The resulting increase in cytosolicCa2� in the injected and neighboring cells was detected by timelapse confocal microscopy (Fig. 3). The Ca2� increase spread to3.2 � 1.6 cells in the cx32 group (n � 17), 2.0 � 1.3 cells in thecx43 group (n � 9; p � 0.5 relative to the cx32 group), and to0.8 � 0.9 cells in the wild type SkHep1 cells (n � 11; p � 0.0001relative to both transfected cell lines). The Ca2� increase didnot spread to any other cells in cx32 cells treated with tetra-cycline (n � 8; p � 0.0001 relative to cx32 cells without tetra-cycline), similar to what was observed in the wild type. InsP3-induced Ca2� signals spread to neighboring cells within 0.60 �0.13 s (n � 26). These findings demonstrate that InsP3 canmediate the spread of Ca2� signals among SkHep1 cells via gapjunctions, similar to what has been shown previously in othercell lines (35). Furthermore, InsP3-mediated Ca2� signals canspread across either cx32 or cx43, similar to what has beenobserved in other cell types as well (36, 37).
The next group of studies investigated whether Ca2� can actdirectly as an intercellular messenger. DM-nitrophen-EDTA(caged Ca2�) was loaded into wild type and cx43- and cx32-transfected SkHep1 cells, then the Ca2� was liberated by two-photon flash photolysis as Ca2� was observed by time lapseconfocal microscopy (Fig. 4, a and b). Use of two-photon exci-tation allowed uncaging of Ca2� to be restricted to individualcells, even though all cells were loaded with caged Ca2� (28). Incontrast to what was observed after photolysis of NPE-InsP3,uncaged Ca2� never spread to adjacent cells in either the cx32group (n � 13) or the cx43 group (n � 16). This is consistentwith the idea that the range of action of InsP3 is greater thanthat of Ca2� (38). To examine the effective range of action ofCa2� in this system in more detail, Ca2� was uncaged indiscrete 40 (6.3 � 6.3) �m2 regions in SkHep1 cells, and thespread of Ca2� to elsewhere within the cell was monitored (Fig.5, a and b). Increases in cytosolic Ca2� dissipated rapidly, andextended no more than 8.7 � 1.1 (n � 8) �m away from the siteof uncaging. Together, these findings suggest that InsP3 rather
than Ca2� acts as an intercellular messenger in unstimulatedepithelial cells.
Local Ca2� Release Can Trigger Intercellular Ca2� Signals—Although the range of action of Ca2� is limited in unstimulatedcells, Ca2� acts as a co-agonist for the InsP3 receptor (39–41).As a result, the effects of Ca2� are potentiated in the presenceof InsP3. Therefore, we examined whether local release of Ca2�
can trigger intracellular and intercellular Ca2� waves duringhormonal stimulation. Ca2� was photoreleased in individualcells both before and during stimulation with a “subthreshold”concentration of vasopressin (50 pM to 1 nM) that was too low toincrease Ca2� by itself (Fig. 6, a and b). Photorelease of cagedCa2� increased fluo-3 fluorescence by 25 � 5% in the absence ofvasopressin, and by 236 � 26% during subthreshold stimula-tion with vasopressin (n � 31; p � 0.0001 by paired t test).Similarly, the increase in fluo-3 fluorescence lasted 9.1 � 0.9 sin the absence of vasopressin, and over 2 min during vasopres-sin stimulation (p � 0.0001 by paired t test). Thus, althoughrelease of caged Ca2� induced a small, transient increase inCa2� in unstimulated cells, it instead triggered a much largerand prolonged increase in Ca2� in cells stimulated with sub-threshold amounts of vasopressin. This demonstrates that lo-cal, subcellular release of Ca2� increases the sensitivity of
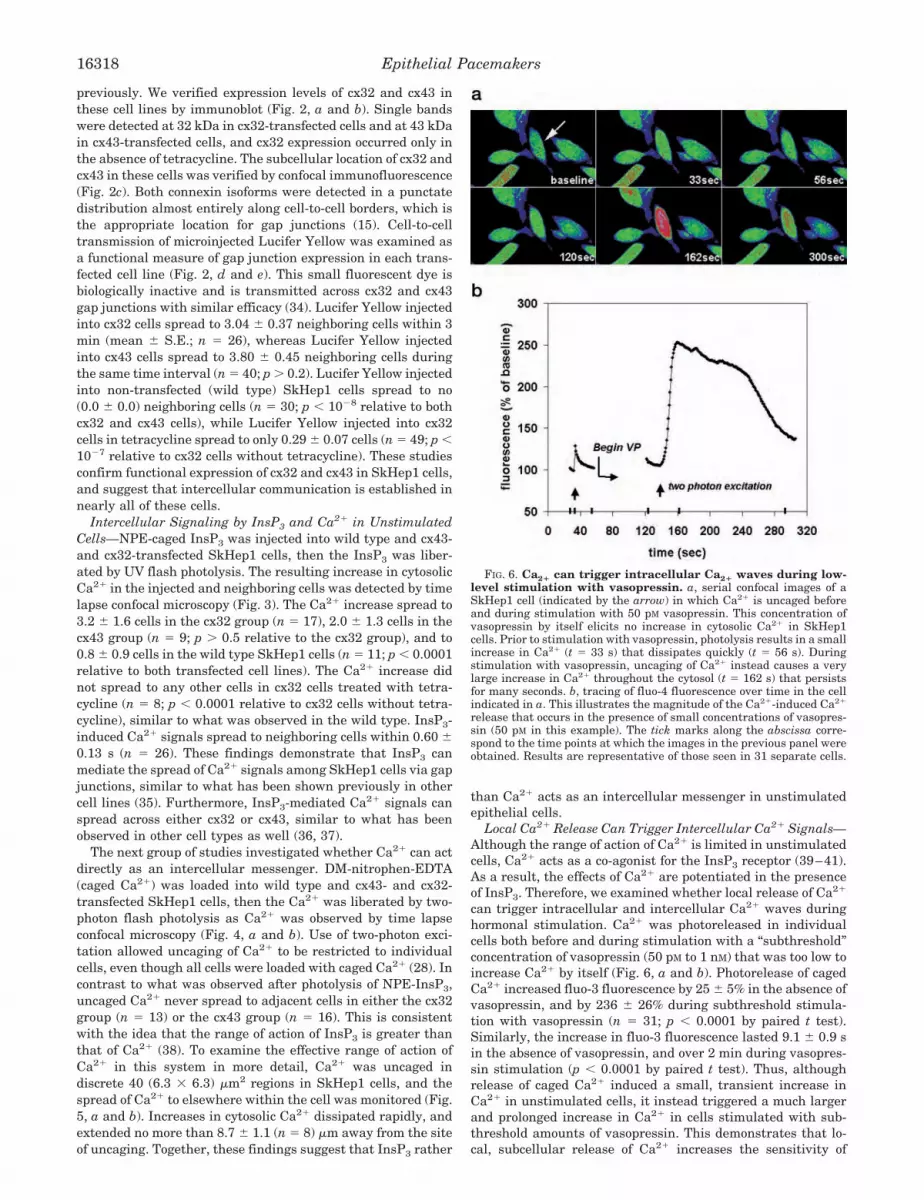
FIG. 6. Ca2� can trigger intracellular Ca2� waves during low-level stimulation with vasopressin. a, serial confocal images of aSkHep1 cell (indicated by the arrow) in which Ca2� is uncaged beforeand during stimulation with 50 pM vasopressin. This concentration ofvasopressin by itself elicits no increase in cytosolic Ca2� in SkHep1cells. Prior to stimulation with vasopressin, photolysis results in a smallincrease in Ca2� (t � 33 s) that dissipates quickly (t � 56 s). Duringstimulation with vasopressin, uncaging of Ca2� instead causes a verylarge increase in Ca2� throughout the cytosol (t � 162 s) that persistsfor many seconds. b, tracing of fluo-4 fluorescence over time in the cellindicated in a. This illustrates the magnitude of the Ca2�-induced Ca2�
release that occurs in the presence of small concentrations of vasopres-sin (50 pM in this example). The tick marks along the abscissa corre-spond to the time points at which the images in the previous panel wereobtained. Results are representative of those seen in 31 separate cells.
Epithelial Pacemakers16318

individual SkHep1 cells to vasopressin. Next we investigatedwhether these vasopressin-plus-Ca2�-induced Ca2� signalscan spread to other cells as well. For these studies, Ca2� wasphotoreleased in individual cells during stimulation with sub-threshold vasopressin, then Ca2� signaling was monitored inthose cells as well as in neighboring cells (Fig. 7, a–c). The Ca2�
increase spread to 1.7 � 0.3 cells in the cx32 group (n � 10),and to 2.1 � 0.3 cells in the cx43 group (n � 19; p � 0.4 relativeto the cx32 group), but did not spread to any other wild typeSkHep1 cells (n � 6; p � 0.0005 relative to both transfected celllines). Similarly, the Ca2� increase spread to no other cells incx32 cells treated with tetracycline (n � 4; p � 0.0005 relativeto cx32 cells without tetracycline). The time delay betweenphotorelease of Ca2� in an individual cell and Ca2� signaling inneighboring cells was 2.8 � 0.3 s in cells expressing cx32 and2.3 � 0.3 s in cells expressing cx43 (p � 0.3), which is similarto the time interval required for InsP3-induced Cai2� signals tospread in these cells (Fig. 3, a–c). Thus, unlike what wasobserved in unstimulated cells, Ca2� triggered global intracel-lular as well as intercellular Ca2� waves in cells stimulated
with low concentrations of vasopressin. This ability of localCa2� signals to enable individual cells to act as pacemakers fortheir neighbors depended upon expression of gap junctions aswell.
Vasopressin Receptor Expression and Establishment of Pace-maker Cells—Since increased hormone receptor expression canlead to increased InsP3 production (42, 43), we expressed V1a
vasopressin receptors in SkHep1 cells as an alternative way toincrease their sensitivity to vasopressin. The rat liver V1a re-ceptor (44) was tagged with RFP to identify cells expressing thereceptor. The V1aR-RFP construct first was transiently ex-pressed in CHO cells, which do not normally express vasopres-sin receptors. Transfected cells were readily identified by RFPfluorescence, which was concentrated around and beneath theplasma membrane (Fig. 8a). This fluorescence distribution pat-tern was similar to the subcellular distribution of the nativeV1a receptor, as determined by confocal immunofluorescence offreshly isolated rat hepatocytes (Fig. 8b). To determinewhether function was preserved in the tagged receptor, CHOcells were loaded with fluo-3 to monitor cytosolic Ca2�, then
FIG. 7. Ca2� can trigger intercellular Ca2� waves during low-level stimulation with vasopressin. a, serial confocal images of SkHep1cells expressing cx43 in which Ca2� is uncaged in one of the cells (indicated by the arrow) before and during stimulation with vasopressin (1 nM).Prior to stimulation, photolysis results in a small increase in Ca2� that is detected at the time of the flash, then dissipates. During stimulation withvasopressin, uncaging of Ca2� in the same cell now causes a very large increase in Ca2� throughout the cytosol that persists for many seconds. Asustained increase in Ca2� now spreads to neighboring cells 1 and 2 as well. This concentration of vasopressin by itself also elicits no increase incytosolic Ca2� in SkHep1 cells. b, tracing of fluo-4 fluorescence over time in the cells indicated in the previous panel. The tick marks along theabscissa correspond to the time points at which the images in the previous panel were obtained. This illustrates that Ca2�-induced Ca2� releasecan spread to neighboring cells when the cells are primed with small, subthreshold concentrations of vasopressin (50 pM-1 nM). Results arerepresentative of those seen in 29 separate cells. c, summary of intercellular Ca2� signaling in cells subjected to photolysis of caged Ca2� in thepresence of subthreshold vasopressin. A prolonged increase in cytosolic Ca2� always was observed in the cell in which Ca2� was uncaged, but theincrease spread only among cells expressing cx32 or cx43 (*, p � 0.0005).
Epithelial Pacemakers 16319
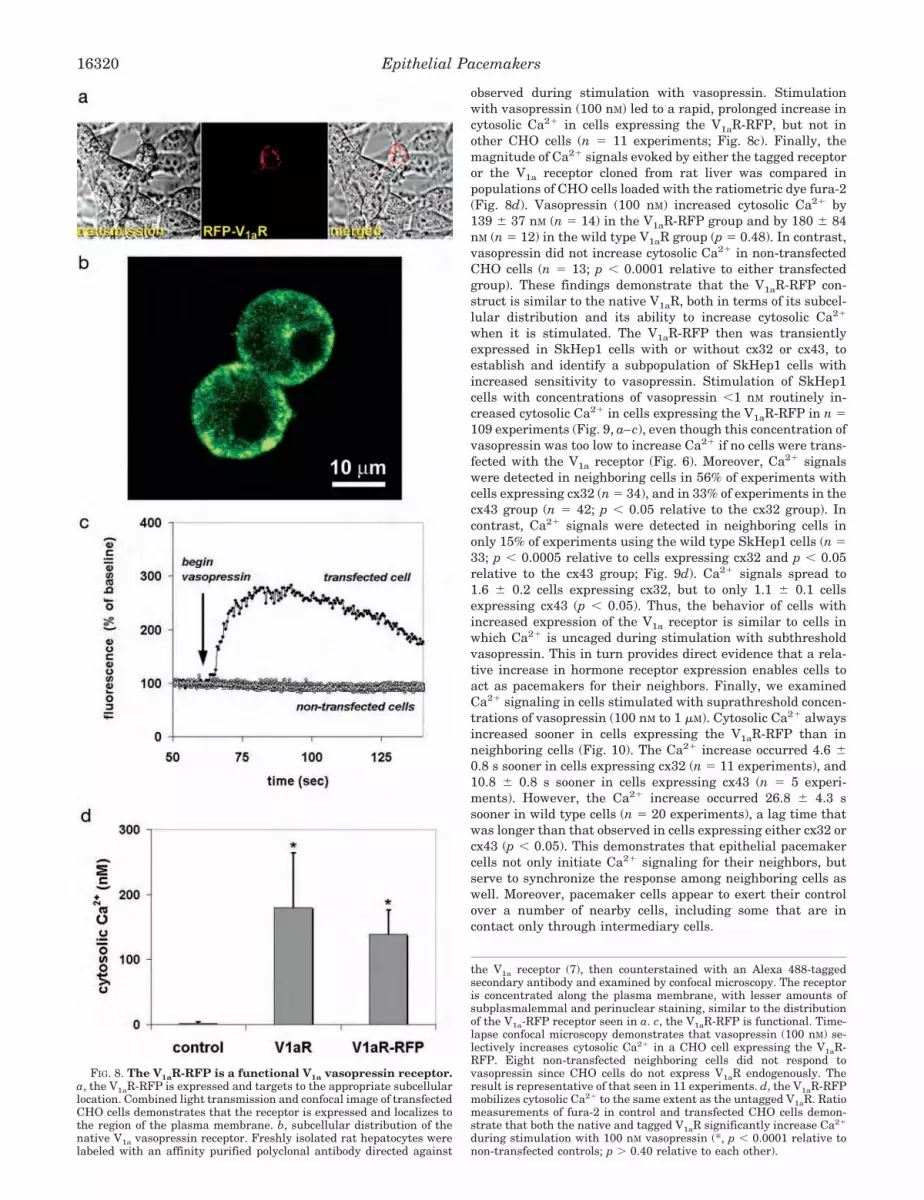
observed during stimulation with vasopressin. Stimulationwith vasopressin (100 nM) led to a rapid, prolonged increase incytosolic Ca2� in cells expressing the V1aR-RFP, but not inother CHO cells (n � 11 experiments; Fig. 8c). Finally, themagnitude of Ca2� signals evoked by either the tagged receptoror the V1a receptor cloned from rat liver was compared inpopulations of CHO cells loaded with the ratiometric dye fura-2(Fig. 8d). Vasopressin (100 nM) increased cytosolic Ca2� by139 � 37 nM (n � 14) in the V1aR-RFP group and by 180 � 84nM (n � 12) in the wild type V1aR group (p � 0.48). In contrast,vasopressin did not increase cytosolic Ca2� in non-transfectedCHO cells (n � 13; p � 0.0001 relative to either transfectedgroup). These findings demonstrate that the V1aR-RFP con-struct is similar to the native V1aR, both in terms of its subcel-lular distribution and its ability to increase cytosolic Ca2�
when it is stimulated. The V1aR-RFP then was transientlyexpressed in SkHep1 cells with or without cx32 or cx43, toestablish and identify a subpopulation of SkHep1 cells withincreased sensitivity to vasopressin. Stimulation of SkHep1cells with concentrations of vasopressin �1 nM routinely in-creased cytosolic Ca2� in cells expressing the V1aR-RFP in n �109 experiments (Fig. 9, a–c), even though this concentration ofvasopressin was too low to increase Ca2� if no cells were trans-fected with the V1a receptor (Fig. 6). Moreover, Ca2� signalswere detected in neighboring cells in 56% of experiments withcells expressing cx32 (n � 34), and in 33% of experiments in thecx43 group (n � 42; p � 0.05 relative to the cx32 group). Incontrast, Ca2� signals were detected in neighboring cells inonly 15% of experiments using the wild type SkHep1 cells (n �33; p � 0.0005 relative to cells expressing cx32 and p � 0.05relative to the cx43 group; Fig. 9d). Ca2� signals spread to1.6 � 0.2 cells expressing cx32, but to only 1.1 � 0.1 cellsexpressing cx43 (p � 0.05). Thus, the behavior of cells withincreased expression of the V1a receptor is similar to cells inwhich Ca2� is uncaged during stimulation with subthresholdvasopressin. This in turn provides direct evidence that a rela-tive increase in hormone receptor expression enables cells toact as pacemakers for their neighbors. Finally, we examinedCa2� signaling in cells stimulated with suprathreshold concen-trations of vasopressin (100 nM to 1 �M). Cytosolic Ca2� alwaysincreased sooner in cells expressing the V1aR-RFP than inneighboring cells (Fig. 10). The Ca2� increase occurred 4.6 �0.8 s sooner in cells expressing cx32 (n � 11 experiments), and10.8 � 0.8 s sooner in cells expressing cx43 (n � 5 experi-ments). However, the Ca2� increase occurred 26.8 � 4.3 ssooner in wild type cells (n � 20 experiments), a lag time thatwas longer than that observed in cells expressing either cx32 orcx43 (p � 0.05). This demonstrates that epithelial pacemakercells not only initiate Ca2� signaling for their neighbors, butserve to synchronize the response among neighboring cells aswell. Moreover, pacemaker cells appear to exert their controlover a number of nearby cells, including some that are incontact only through intermediary cells.
the V1a receptor (7), then counterstained with an Alexa 488-taggedsecondary antibody and examined by confocal microscopy. The receptoris concentrated along the plasma membrane, with lesser amounts ofsubplasmalemmal and perinuclear staining, similar to the distributionof the V1a-RFP receptor seen in a. c, the V1aR-RFP is functional. Time-lapse confocal microscopy demonstrates that vasopressin (100 nM) se-lectively increases cytosolic Ca2� in a CHO cell expressing the V1aR-RFP. Eight non-transfected neighboring cells did not respond tovasopressin since CHO cells do not express V1aR endogenously. Theresult is representative of that seen in 11 experiments. d, the V1aR-RFPmobilizes cytosolic Ca2� to the same extent as the untagged V1aR. Ratiomeasurements of fura-2 in control and transfected CHO cells demon-strate that both the native and tagged V1aR significantly increase Ca2�
during stimulation with 100 nM vasopressin (*, p � 0.0001 relative tonon-transfected controls; p � 0.40 relative to each other).
FIG. 8. The V1aR-RFP is a functional V1a vasopressin receptor.a, the V1aR-RFP is expressed and targets to the appropriate subcellularlocation. Combined light transmission and confocal image of transfectedCHO cells demonstrates that the receptor is expressed and localizes tothe region of the plasma membrane. b, subcellular distribution of thenative V1a vasopressin receptor. Freshly isolated rat hepatocytes werelabeled with an affinity purified polyclonal antibody directed against
Epithelial Pacemakers16320
DISCUSSION
The presence of pacemaker cells regulating cell functions inexcitable tissues is well known, but the possibility that analo-gous pacemaker cells coordinate cell signaling in non-excitabletissues such as epithelia has only recently been investigated(43, 45). Studies using isolated rat hepatocytes suggest thatpacemaker cells synchronize Ca2� oscillations in pairs andtriplets of cells, since oscillations are coordinated in such mul-tiplets (46), and since one of the cells in a multiplet generallyappears to set the rate of oscillations for its neighbors (45, 47).Indirect evidence in both isolated hepatocytes (43, 45) and theisolated perfused liver (11, 14) suggests that this pacemaker
activity may be driven by hormone receptor gradients. Here weexamined this question directly in the liver-derived SkHep1cell line. Studies were performed using a cell line rather thanprimary hepatocytes, since hepatocytes lose expression of hor-mone receptors (48) and gap junctions (49) within hours ofisolation, yet both are necessary components of coordinatedintercellular signaling in epithelia. Using the SkHep1 modelsystem, three components were identified that are needed forpacemaker cells to organize signaling events among epithelia.The first component is intercellular communication of secondmessengers via gap junctions. It was first established thatsecond messengers such as Ca2� and InsP3 can cross gap junc-tions in isolated rat hepatocyte couplets (36). Subsequent workin an airway epithelial cell line demonstrated that InsP3-me-diated Ca2� signaling in one cell can induce Ca2� signaling inneighboring cells as well, and that this form of intercellularcommunication requires gap junctions (50). Moreover, Ca2�
waves and oscillations are synchronized when communicatingepithelia are stimulated, and this integrated response to hor-monal stimulation is dependent upon gap junctions as well(46). InsP3 can serve as the second messenger that coordinatescell-cell Ca2� signaling (10, 35), although Ca2� also may beable to serve this role under certain circumstances (10, 51).Regardless of which second messenger is responsible, the pres-ence of gap junctions is required for coordination of cell-to-cellsignaling among epithelia, since Ca2� oscillations becomeasynchronous in the absence of gap junctions (45). The currentwork, furthermore, demonstrates that initiation of Ca2� sig-nals becomes asynchronous in cells that do not express gapjunctions. Previous studies have shown that intercellular ex-change of both positively and negatively charged moleculesdepends upon which connexin isoforms are expressed (34), andthis includes exchange of second messengers as well (52). Thecurrent work extends these observations by providing evidencethat pacemaker activity is more enhanced by expression of cx32than by cx43 (Figs. 9d and 10d). Thus, the extent to which Ca2�
signals become synchronized may depend in part upon whichconnexin isoform is expressed, although each type examinedleads to some degree of coordination.
Expression of gap junctions is not the only requirement forsynchronization of Ca2� signals. Current and previous studieshave shown that stimulation of an individual cell, via applicationof hormone or else direct injection of InsP3, may fail to induceCa2� signaling in neighboring cells (45). However, low-level stim-ulation of multiple cells does allow Ca2� signaling in one of thesecells to trigger Ca2� signals in the neighbors (10, 45, 51). Thisproperty has been related to the observation that InsP3Rs enablecytosol to act as an excitable medium in the presence of elevatedconcentrations of InsP3 (53, 54). This concept originally wasestablished in Xenopus oocytes, where regenerative Ca2� wavesand oscillations could be elicited by activation of InsP3Rs (53, 54).Subsequent detailed analyses of this behavior have identifiedelementary Ca2� release events that occur within the cytosol,and have found that certain spatially defined release sites have ahigher sensitivity to InsP3, which may reflect local clustering ofInsP3Rs (55–57). This finding also has been extended to mam-malian cells (58), and theoretical work suggests that localizedsubcellular regions require clustering of at least 20–30 InsP3Rsto exhibit higher sensitivity to InsP3 (59). This type of regenera-tive Ca2� signaling activity now has been shown to occur innetworks of epithelial cells as well, including in liver (8), pan-creas (51), and salivary glands (10). Alternatively, work in Xeno-pus oocytes suggests that localized photorelease of caged Ca2�
can trigger regenerative activity in the presence of low concen-trations of InsP3 by converting localized Ca2� puffs into Ca2�
waves (55, 56). This likely reflects the fact that Ca2� acts as a
FIG. 9. Expression of V1aR-RFP increases the sensitivity ofSkHep1 cells to vasopressin. a, SkHep1 cells expressing V1aR-RFPrespond to vasopressin concentrations that are below the threshold forevoking Ca2� signals in wild type SkHep1 cells. Cells were stimulatedwith 50 pM vasopressin (VP). Wild type cells (left column) respond to VPonly if they express the V1aR-RFP (arrow). In contrast, among cellsexpressing cx32 (right column), both V1aR-RFP cell (arrow) and someneighbors respond to VP. Numbers correspond to the tracings in b andc. b, tracing of fluo-4 fluorescence over time in wild type cells in theprevious panel. An increase in Ca2� is observed only in the cell trans-fected with V1aR-RFP. Results are representative of those observed in28 experiments. c, tracing of fluo-4 fluorescence over time in cx32 cellsin the first panel of this figure. An increase in Ca2� is observed in thecell transfected with V1aR-RFP, followed by an increase in Ca2� in fourof the neighboring cells. Results are representative of those observed in19 experiments. d, summary of intercellular Ca2� signaling in thepresence of subthreshold vasopressin among cells expressing V1aR-RFP. A prolonged increase in cytosolic Ca2� always was observed incells expressing the tagged receptor, but the increase subsequentlyoccurred in neighboring cells more frequently among cells co-expressingcx32 (*, p � 0.0005) or cx43 (**, p � 0.05).
Epithelial Pacemakers 16321

co-agonist for the InsP3R (40, 60). The current work uses two-photon flash photolysis to demonstrate that highly localized in-creases in Ca2� can trigger InsP3-induced Ca2� signals in mam-malian cells as well. We, furthermore, found that localized,subcellular release of Ca2� is sufficient to trigger not only globalCa2� signaling within an individual cell, but also cell-cell signal-ing, as long as the cell network communicates via gap junctionsand is primed by InsP3. The current work, furthermore, estab-lishes SkHep1 cells as a model system for investigating cell-cellsignaling via InsP3 by demonstrating that InsP3Rs are distrib-uted uniformly throughout each cell, and that RyR are notexpressed.
Signaling patterns that occur in organs and tissues in vivoare more complex than can be accounted for simply by gapjunctional communication among excitable cells. For example,intercellular Ca2� waves in liver are oriented in specific direc-
tions (7, 9). In addition, the direction of such Ca2� wavesdepends upon which hormone is used to elicit the waves (9).Similarly, cell-cell spread of Ca2� waves among isolated clus-ters of hepatocytes occurs in a reproducible pattern, but thepattern varies depending on the hormonal stimulus (61). Cellswithin the group that initiates Ca2� signals have increasedhormonal sensitivity, and several indirect lines of evidencesuggest this is due to increased hormone receptor expression (7,42, 43). The current findings directly demonstrate that in-creased expression of hormone receptor enables a cell to behaveas a pacemaker for its neighbors. Such oriented intercellularsignaling is crucial for proper tissue function, since defects inintercellular signaling can alter critical functions such as glu-cose production or bile secretion in liver (14, 62, 63), or amylaserelease in pancreas (64). Thus, intercellular integration of Ca2�
signaling allows cells to exhibit organ-level behavior, rather
FIG. 10. Establishment of pacemaker cells by overexpression of V1a vasopressin receptors. a, SkHep1 cells expressing V1aR-RFP actas pacemakers for their neighbors. Cells here express cx32 and are stimulated with 100 nM vasopressin (VP). Serial confocal images demonstratethat an increase in Ca2� occurs first in the cell expressing the V1aR-RFP (arrow), but all other cells respond with a prompt, sustained increase inCa2� soon afterward. b, tracing of the increases in fluo-4 fluorescence in the cells is indicated in a. c, expanded time scale of the previous tracingreveals that the increase in Ca2� in the cell expressing V1aR-RFP precedes the Ca2� signal in its neighbors by several seconds. The result isrepresentative of that seen in 16 experiments using cx32 or cx43 cells. The lag time between Ca2� signaling in V1aR-RFP cells and their neighborsis significantly longer in wild type SkHep1 cells (n � 20 experiments). d, summary of intercellular Ca2� signaling in the presence of suprathresholdvasopressin (100 nM) among cells expressing V1aR-RFP. An increase in cytosolic Ca2� always occurred first in cells expressing the tagged receptor,but the delay between the Ca2� signal in the V1aR-RFP cell and its neighbors was significantly shorter if cells co-expressed cx32 or cx43(*, p � 0.05).
Epithelial Pacemakers16322

than to behave merely as a collection of single cells. Moreover,the distribution of pacemaker cells within an organ permitshormones to evoke distinct responses from that particular or-gan, even though each hormone may activate the same secondmessenger pathways at the single cell level.
Acknowledgments—We thank Warren Zipfel for advice regardingtwo-photon flash photolysis, Richard Wojcikiewicz for kindly supply-ing InsP3R antibody CT2, and Carlos Gonzalez and Juan Saez forkindly supplying an antibody directed against the V1a vasopressinreceptor.
REFERENCES
1. Kaneko, T., Tanaka, H., Oyamada, M., Kawata, S., and Takamatsu, T. (2000)Circ. Res. 86, 1093–1099
2. Lamont, C., Luther, P. W., Balke, C. W., and Wier, W. G. (1998) J. Physiol.(Lond.) 512, 669–676
3. Berridge, M. J. (1993) Nature 361, 315–3254. Cheng, H., Lederer, W. J., and Cannell, M. B. (1993) Science 262, 740–7445. Blatter, L. A., Huser, J., and Rıos, E. (1997) Proc. Natl. Acad. Sci. U. S. A. 94,
4176–41816. Mackenzie, L., Bootman, M. D., Berridge, M. J., and Lipp, P. (2001) J. Physiol.
(Lond.) 530, 417–4297. Nathanson, M. H., Burgstahler, A. D., Mennone, A., Fallon, M. B., Gonzalez,
C. B., and Saez, J. C. (1995) Am. J. Physiol. Gastrointest. Liver Physiol. 269,G167–G171
8. Robb-Gaspers, L. D., and Thomas, A. P. (1995) J. Biol. Chem. 270, 8102–81079. Motoyama, K., Karl, I. E., Flye, M. W., Osborne, D. F., and Hotchkiss, R. S.
(1999) Am. J. Physiol. Regul. Integr. Comp. Physiol. 276, R575–R58510. Zimmermann, B., and Walz, B. (1999) EMBO J. 18, 3222–323111. Serriere, V., Berthon, B., Boucherie, S., Jacquemin, E., Guillon, G., Claret, M.,
and Tordjmann, T. (2001) FASEB J. 15, 1484–148612. Meda, P., Bosco, D., Chanson, M., Giordano, E., Vallar, L., Wollheim, C., and
Orci, L. (1990) J. Clin. Invest. 86, 759–76813. Meda, P., Bruzzone, R., Chanson, M., Bosco, D., and Orci, L. (1987) Proc. Natl.
Acad. Sci. U. S. A. 84, 4901–490414. Nathanson, M. H., Rios-Velez, L., Burgstahler, A. D., and Mennone, A. (1999)
Gastroenterology 116, 1176–118315. Fallon, M. B., Nathanson, M. H., Mennone, A., Saez, J. C., Burgstahler, A. D.,
and Anderson, J. M. (1995) Am. J. Physiol. Cell Physiol. 268, C1186–C119416. Wojcikiewicz, R. J. H. (1995) J. Biol. Chem. 270, 11678–1168317. Fishman, G. I., Gao, Y., Hertzberg, E. L., and Spray, D. C. (1995) Cell Adhes.
Commun. 3, 353–36518. Eghbali, B., Kessler, J. A., and Spray, D. C. (1990) Proc. Natl. Acad. Sci.
U. S. A. 87, 1328–133119. Fishman, G. I., Spray, D. C., and Leinwand, L. A. (1990) J. Cell Biol. 111,
589–59820. Schlosser, S. F., Burgstahler, A. D., and Nathanson, M. H. (1996) Proc. Natl.
Acad. Sci. U. S. A. 93, 9948–995321. Laemmli, U. K. (1970) Nature 227, 680–68522. Leite, M. F., Dranoff, J. A., Gao, L., and Nathanson, M. H. (1999) Biochem. J.
337, 305–30923. Giannini, G., Conti, A., Mammarella, S., Scrobogna, M., and Sorrentino, V.
(1995) J. Cell Biol. 128, 893–90424. Bennett, D. L., Cheek, T. R., Berridge, M. J., De Smedt, H., Parys, J. B.,
Missiaen, L., and Bootman, M. D. (1996) J. Biol. Chem. 271, 6356–636225. Hirata, K., Nathanson, M. H., and Sears, M. L. (1998) Proc. Natl. Acad. Sci.
U. S. A. 95, 8381–838626. Hagar, R. E., Burgstahler, A. D., Nathanson, M. H., and Ehrlich, B. E. (1998)
Nature 396, 81–8427. Brown, E. B., Shear, J. B., Adams, S. R., Tsien, R. Y., and Webb, W. W. (1999)
Biophys. J. 76, 489–49928. Leite, M. F., Burgstahler, A. D., and Nathanson, M. H. (2002) Gastroenterology
122, 415–42729. Dufour, J.-F., Luthi, M., Forestier, M., and Magnino, F. (1999) Hepatology 30,
1018–102630. Hirata, K., Nathanson, M. H., Burgstahler, A. D., Okazaki, K., Mattei, E., and
Sears, M. L. (1999) Invest. Ophthalmol. Vis. Sci. 40, 2046–205331. Nathanson, M. H., Fallon, M. B., Padfield, P. J., and Maranto, A. R. (1994)
J. Biol. Chem. 269, 4693–469632. Yule, D. I., Ernst, S. A., Ohnishi, H., and Wojcikiewicz, R. J. H. (1997) J. Biol.
Chem. 272, 9093–909833. Lee, M. G., Xu, X., Zeng, W. Z., Diaz, J., Wojcikiewicz, R. J. H., Kuo, T. H.,
Wuytack, F., Racymaekers, L., and Muallem, S. (1997) J. Biol. Chem. 272,15765–15770
34. Elfgang, C., Eckert, R., Lichtenberg-Frate, H., Butterweck, A., Traub, O.,Klein, R. A., Hulser, D. F., and Willecke, K. (1995) J. Cell Biol. 129,805–817
35. Boitano, S., Dirksen, E. R., and Sanderson, M. J. (1992) Science 258, 292–29536. Saez, J. C., Connor, J. A., Spray, D. C., and Bennett, M. V. L. (1989) Proc. Natl.
Acad. Sci. U. S. A. 86, 2708–271237. Charles, A. C., Naus, C. C., Zhu, D., Kidder, G. M., Dirksen, E. R., and
Sanderson, M. J. (1992) J. Cell Biol. 118, 195–20138. Allbritton, N. L., Meyer, T., and Stryer, L. (1992) Science 258, 1812–181539. Finch, E. A., Turner, T. J., and Goldin, S. M. (1991) Science 252, 443–44640. Bezprozvanny, I., Watras, J., and Ehrlich, B. E. (1991) Nature 351, 751–75441. Iino, M. (1990) J. Gen. Physiol. 95, 1103–112242. Tordjmann, T., Berthon, B., Combettes, L., and Claret, M. (1996) Gastroenter-
ology 111, 1343–135243. Tordjmann, T., Berthon, B., Jacquemin, E., Clair, C., Stelly, N., Guillon, G.,
Claret, M., and Combettes, L. (1998) EMBO J. 17, 4695–470344. Morel, A., O’Carroll, A.-M., Brownstein, M. J., and Lolait, S. J. (1992) Nature
356, 523–52645. Tordjmann, T., Berthon, B., Claret, M., and Combettes, L. (1997) EMBO J. 16,
5398–540746. Nathanson, M. H., and Burgstahler, A. D. (1992) Mol. Biol. Cell 3, 113–12147. Clair, C., Chalumeau, C., Tordjmann, T., Poggioli, J., Erneux, C., Dupont, G.,
and Combettes, L. (2001) J. Cell Sci. 114, 1999–200748. Ishac, E. J. N., Lazar-Wesley, E., and Kunos, G. (1992) J. Cell. Physiol. 152,
79–8649. Saez, J. C., Gregory, W. A., Watanabe, T., Dermietzel, R., Hertzberg, E. L.,
Reid, L., Bennett, M. V. L., and Spray, D. C. (1989) Am. J. Physiol. CellPhysiol. 257, C1–C11
50. Sanderson, M. J., Charles, A. C., and Dirksen, E. R. (1990) Cell Regul. 1,585–596
51. Yule, D. I., Stuenkel, E., and Williams, J. A. (1996) Am. J. Physiol. CellPhysiol. 271, C1285–C1294
52. Paemeleire, K., Martin, P. E. M., Coleman, S. L., Fogarty, K. E., Carrington,W. A., Leybaert, L., Tuft, R. A., Evans, W. H., and Sanderson, M. J. (2000)Mol. Biol. Cell 11, 1815–1827
53. Lechleiter, J., Girard, S., Peralta, E., and Clapham, D. (1991) Science 252,123–126
54. Lechleiter, J. D., and Clapham, D. E. (1992) Cell 69, 283–29455. Marchant, J., Callamaras, N., and Parker, I. (1999) EMBO J. 18, 5285–529956. Marchant, J. S., and Parker, I. (2001) EMBO J. 20, 65–7657. Yao, Y., Choi, J., and Parker, I. (1995) J. Physiol. (Lond.) 482, 533–55358. Thomas, D., Lipp, P., Tovey, S. C., Berridge, M. J., Li, W., Tsien, R. Y., and
Bootman, M. D. (2000) Curr. Biol. 10, 8–1559. Swillens, S., Dupont, G., Combettes, L., and Champeil, P. (1999) Proc. Natl.
Acad. Sci. U. S. A. 96, 13750–1375560. Finch, E. A., and Goldin, S. M. (1994) Science 265, 813–81561. Combettes, L., Tran, D., Tordjmann, T., Laurent, M., Berthon, B., and Claret,
M. (1994) Biochem. J. 304, 585–59462. Eugenin, E. A., Gonzalez, H., Saez, C. G., and Saez, J. C. (1998) Am. J. Physiol.
Gastrointest. Liver Physiol. 37, G1109–G111663. Stumpel, F., Ott, T., Willecke, K., and Jungermann, K. (1998) Hepatology 28,
1616–162064. Chanson, M., Fanjul. M., Bosco, D., Nelles, E., Suter, S., Willecke, K., and
Meda, P. (1998) J. Cell Biol. 141, 1267–1275
Epithelial Pacemakers 16323
Related Documents











