Markedly Elevated Lipid Transfer Inhibitor Protein in Hypercholesterolemic Subjects Is Mitigated by Plasma Triglyceride Levels Richard E. Morton, Valéria Nunes, Lahoucine Izem, Eder Quintão Abstract—Lipid transfer inhibitor protein (LTIP, apolipoprotein F) regulates the interaction of cholesteryl ester transfer protein (CETP) with lipoproteins and is postulated to enhance the ability of CETP to stimulate reverse cholesterol transport. The factors that regulate LTIP levels and control its biosynthesis are unknown. Here, we demonstrate that plasma LTIP is dramatically increased (3-fold) in hypercholesterolemic subjects with normal to mildly elevated plasma triglyceride (TG) levels compared with control subjects. LTIP in these subjects is not correlated with the extent of hypercholesterolemia or with low density lipoprotein (LDL), high density lipoprotein, or CETP levels. However, unlike CETP, LTIP levels correlate negatively with plasma TG levels. This association does not appear to reflect decreased LTIP synthesis, inasmuch as conditions that stimulate TG synthesis and secretion (200 mmol/L oleate) do not reduce LTIP secretion by SW872 or Caco-2 cells. In contrast, native or acetyl LDL stimulates LTIP secretion 2-fold. Importantly, although plasma LTIP typically resides on LDL, up to 25% of LTIP is bound to very low density lipoprotein when this lipoprotein is enriched in cholesteryl esters, as occurs in hypercholesterolemia. In summary, LTIP levels are markedly elevated by hypercholesterolemia; however, plasma TG levels attenuate this response. We hypothesize that this arises from an increased association of LTIP with very low density lipoprotein, leading to a more rapid clearance of the inhibitor from circulation. (Arterioscler Thromb Vasc Biol. 2001;21:1642-1649.) Key Words: hypercholesterolemia n cholesteryl ester transfer protein n lipid transfer inhibitor protein n triglycerides n lipoprotein metabolism C holesteryl ester transfer protein (CETP) mediates the flux of cholesteryl esters (CEs) and triglycerides (TGs) between lipoproteins. 1,2 CETP facilitates a heteroexchange reaction, resulting in the net transfer of CEs toward CE-poor particles (VLDLs) with the reciprocal movement of TGs to CE-rich particles (LDLs or HDLs). 3,4 Strong in vitro evidence demonstrates an important role for CETP in facilitating VLDL catabolism to LDLs and in promoting HDL metabo- lism and reverse cholesterol transport. 3–5 However, the over- all impact of CETP activity on atherogenesis has remained controversial, inasmuch as CETP can potentially facilitate processes that would appear to be proatherogenic and antiatherogenic. CETP activity is regulated by another plasma component, lipid transfer inhibitor protein (LTIP). We recently purified and cloned LTIP and demonstrated its identity with apoF. 6 Although LTIP was first identified simply by its capacity to suppress CETP activity in binary lipid transfer assays, 7 it now appears that LTIP plays a more complex role in regulating CETP. CETP has little preference for interacting with differ- ent lipoprotein classes under steady-state conditions, 8 and within a mixture of lipoproteins, CETP mediates transfer events between lipoprotein classes at rates that are largely determined by their relative concentrations. 9 This finding contrasts with that seen in plasma, in which HDL appears to be a preferred CETP substrate. 10 –12 We have recently dem- onstrated that LTIP activity accounts for this discrepancy. 9 This is hypothesized to occur because LTIP preferentially suppresses the interaction of CETP with LDL. 13 Because VLDL concentrations are rate limiting in normal plasma to the CE-TG exchange process, 14 the suppression of transfers with LDL results in a stimulation of lipid exchange between VLDL and HDL. 9,15 Therefore, LTIP is a regulator of CETP function in that it controls the rate of individual lipid transfer reactions. We have proposed that LTIP augments the anti- atherogenic capacities of CETP by stimulating reverse cho- lesterol transport. 9,15 CETP synthesis is strongly upregulated by cholesterol, 16,17 and elevated CETP levels are commonly observed in hyper- cholesterolemic subjects. 18,19 This appears to be an adaptive response to enhance mechanisms responsible for sterol ho- meostasis. Because the beneficial actions of CETP are likely to be enhanced by LTIP, it seems reasonable that LTIP levels may be increased by similar stimuli. At present, nothing is Received May 14, 2001; revision accepted July 16, 2001. From the Department of Cell Biology (R.E.M., L.I.), Lerner Institute, Cleveland Clinic Foundation, Cleveland, Ohio, and the Lipids Laboratory (V.N., E.Q.), University of São Paulo Medical School, São Paulo, Brazil. Correspondence to Richard E. Morton, PhD, Cell Biology, NC10, Cleveland Clinic Foundation, 9500 Euclid Ave, Cleveland, OH 44195. E-mail [email protected] © 2001 American Heart Association, Inc. Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org 1642 by guest on August 20, 2015 http://atvb.ahajournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Markedly Elevated Lipid Transfer Inhibitor Protein inHypercholesterolemic Subjects Is Mitigated by
Plasma Triglyceride LevelsRichard E. Morton, Valéria Nunes, Lahoucine Izem, Eder Quintão
Abstract—Lipid transfer inhibitor protein (LTIP, apolipoprotein F) regulates the interaction of cholesteryl ester transferprotein (CETP) with lipoproteins and is postulated to enhance the ability of CETP to stimulate reverse cholesteroltransport. The factors that regulate LTIP levels and control its biosynthesis are unknown. Here, we demonstrate thatplasma LTIP is dramatically increased (3-fold) in hypercholesterolemic subjects with normal to mildly elevated plasmatriglyceride (TG) levels compared with control subjects. LTIP in these subjects is not correlated with the extent ofhypercholesterolemia or with low density lipoprotein (LDL), high density lipoprotein, or CETP levels. However, unlikeCETP, LTIP levels correlate negatively with plasma TG levels. This association does not appear to reflect decreasedLTIP synthesis, inasmuch as conditions that stimulate TG synthesis and secretion (200mmol/L oleate) do not reduceLTIP secretion by SW872 or Caco-2 cells. In contrast, native or acetyl LDL stimulates LTIP secretion 2-fold.Importantly, although plasma LTIP typically resides on LDL, up to 25% of LTIP is bound to very low densitylipoprotein when this lipoprotein is enriched in cholesteryl esters, as occurs in hypercholesterolemia. In summary, LTIPlevels are markedly elevated by hypercholesterolemia; however, plasma TG levels attenuate this response. Wehypothesize that this arises from an increased association of LTIP with very low density lipoprotein, leading to a morerapid clearance of the inhibitor from circulation.(Arterioscler Thromb Vasc Biol. 2001;21:1642-1649.)
Key Words: hypercholesterolemian cholesteryl ester transfer proteinn lipid transfer inhibitor proteinn triglyceridesn lipoprotein metabolism
Cholesteryl ester transfer protein (CETP) mediates theflux of cholesteryl esters (CEs) and triglycerides (TGs)
between lipoproteins.1,2 CETP facilitates a heteroexchangereaction, resulting in the net transfer of CEs toward CE-poorparticles (VLDLs) with the reciprocal movement of TGs toCE-rich particles (LDLs or HDLs).3,4 Strong in vitro evidencedemonstrates an important role for CETP in facilitatingVLDL catabolism to LDLs and in promoting HDL metabo-lism and reverse cholesterol transport.3–5 However, the over-all impact of CETP activity on atherogenesis has remainedcontroversial, inasmuch as CETP can potentially facilitateprocesses that would appear to be proatherogenic andantiatherogenic.
CETP activity is regulated by another plasma component,lipid transfer inhibitor protein (LTIP). We recently purifiedand cloned LTIP and demonstrated its identity with apoF.6
Although LTIP was first identified simply by its capacity tosuppress CETP activity in binary lipid transfer assays,7 it nowappears that LTIP plays a more complex role in regulatingCETP. CETP has little preference for interacting with differ-ent lipoprotein classes under steady-state conditions,8 andwithin a mixture of lipoproteins, CETP mediates transfer
events between lipoprotein classes at rates that are largelydetermined by their relative concentrations.9 This findingcontrasts with that seen in plasma, in which HDL appears tobe a preferred CETP substrate.10–12 We have recently dem-onstrated that LTIP activity accounts for this discrepancy.9
This is hypothesized to occur because LTIP preferentiallysuppresses the interaction of CETP with LDL.13 BecauseVLDL concentrations are rate limiting in normal plasma tothe CE-TG exchange process,14 the suppression of transferswith LDL results in a stimulation of lipid exchange betweenVLDL and HDL.9,15 Therefore, LTIP is a regulator of CETPfunction in that it controls the rate of individual lipid transferreactions. We have proposed that LTIP augments the anti-atherogenic capacities of CETP by stimulating reverse cho-lesterol transport.9,15
CETP synthesis is strongly upregulated by cholesterol,16,17
and elevated CETP levels are commonly observed in hyper-cholesterolemic subjects.18,19 This appears to be an adaptiveresponse to enhance mechanisms responsible for sterol ho-meostasis. Because the beneficial actions of CETP are likelyto be enhanced by LTIP, it seems reasonable that LTIP levelsmay be increased by similar stimuli. At present, nothing is
Received May 14, 2001; revision accepted July 16, 2001.From the Department of Cell Biology (R.E.M., L.I.), Lerner Institute, Cleveland Clinic Foundation, Cleveland, Ohio, and the Lipids Laboratory (V.N.,
E.Q.), University of São Paulo Medical School, São Paulo, Brazil.Correspondence to Richard E. Morton, PhD, Cell Biology, NC10, Cleveland Clinic Foundation, 9500 Euclid Ave, Cleveland, OH 44195. E-mail
[email protected]© 2001 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol.is available at http://www.atvbaha.org
1642 by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from
known about the factors that influence LTIP biosynthesis.The purpose of the present study is to examine the responseof LTIP to hyperlipidemia. The data show that LTIP levelsare markedly increased in hypercholesterolemic subjects andalso demonstrate an intriguing relationship between LTIP andplasma TG levels.
MethodsLipoprotein Isolation and DerivationLipoproteins were isolated from fresh human plasma by sequentialultracentrifugation.20 In some instances, lipoproteins were labeledwith cholesteryl[1a,2a(n)-3H]oleate (50 Ci/mmol, Amersham Corp)before isolation.7 Labeled and unlabeled lipoproteins were exten-sively dialyzed against 0.9% NaCl, 0.01% EDTA, and 0.02% NaN3,pH 7.4, and stored at 4°C. LDL was biotinylated with NHS-biotin, aspreviously described,21 or acetylated by incubation with aceticanhydride.22
Human Subjects
Brazilian SubjectsTwenty-three patients with primary hypercholesterolemia were stud-ied. None of the patients had diabetes mellitus or renal, hepatic, orother secondary causes of hyperlipidemia. Some (14 of 23) of thesesubjects are a subset of a previously reported group.23 Additionally,46 normolipidemic control subjects in good clinical health andwithout any known cardiovascular risk factors were studied. Forreasons explained below, these controls are reported as 2 separategroups (control 1 and control 2, Table 1). None of these subjects hadtaken drugs known to interfere with lipid metabolism for at least 2months. Venous blood samples were drawn after an overnight fast,and plasma was obtained by low-speed centrifugation. Antibioticsand protease inhibitors were added.23 Plasma cholesterol and TGlevels were measured with commercially available kits, and LDLcholesterol, apoB, and apoA-I levels were assayed as previouslydescribed.23 For most subjects, plasma was stored at270°C untilfurther analysis. Lipoproteins were isolated from a subset of freshplasma samples as described above.
Subjects From the United StatesBlood was collected from 7 healthy normolipidemic subjects. Aftercentrifugation to isolate plasma, LDL and HDL cholesterol levelswere measured by the LDL-Direct method (Isolabs, Inc). A pool ofplasma was made by combining equal volumes of each donorplasma. After the addition of phenylmethylsulfonyl fluoride andparaoxon (1 mmol/L final for each), aliquots were frozen at280°C.
All subjects signed a formal written consent approved by theethics committees from the respective institutions. See Table 1 forcharacterization of these subject groups.
Preparation of Lipoprotein-Deficient PlasmaLipoprotein-deficient plasma fractions were prepared by a modifieddivalent cation precipitation method.24 Briefly, 100 mL of freshlyprepared 6.5% dextran sulfate (Mr '53105) was added to 1 mLplasma, mixed briefly, and incubated for 30 minutes on ice. Subse-quently, 61mL of 2 mol/L MnCl2 was added, mixed briefly, and
incubated for 60 minutes on ice. After centrifugation (51 500g) for25 minutes at 4°C, the supernatant was transferred to a second tube,and 122mL of 15% BaCl2 was added, mixed briefly, and incubatedon ice for 30 minutes. After centrifugation as described above, thesupernatant was dialyzed against freshly prepared 50 mmol/L Tris-HCl, 150 mmol/L NaCl, 0.02% EDTA, and 0.02% azide, pH 7.4buffer (Tris/saline buffer). The dialysis consisted of two 2-hour stepsversus 1 L of buffer each and then finally (overnight) versus 2 L ofbuffer. During each step, performed at 4°C, 1 g Chelex-100 (Bio-RadLaboratories) was added to the dialysis buffer. The following day,dialysates were filtered through 0.45-mm syringe-type filters (What-man Inc) to remove particulate material. Samples were stored on iceand assayed the same day for LTIP activity. This method quantita-tively removed lipoproteins from normal and hyperlipidemic sam-ples, as determined by lipid assay.
Assay of LTIP Activity in Lipoprotein-Deficient PlasmaLTIP activity in lipoprotein-deficient plasma was determined bymeasuring its capacity to suppress exogenous CETP.10,25This assaywas modified slightly as follows. Samples (30 to 40mL) werecombined with 500mL of “assay mix,” without or with partiallypurified CETP25 (sufficient to facilitate'20% CE transfer) andTris/saline buffer (pH 7.4 at 37°C) to yield a final assay volume of700 mL. The assay mix consisted of the following: 10mg [3H]CELDL, 30 mg unlabeled LDL, 40mg HDL, 14 mL of 100 mmol/LEDTA (pH 7.4), 200mL of 3.5% BSA, 100mL H2O, and sufficientTris/saline buffer to make 500mL. All components were kept on iceuntil use. After gentle mixing, samples were incubated at 37°C for 2hours. Subsequently, tubes were placed in ice water for 15 minutes,and then the donor and acceptor lipoproteins were separated byselective precipitation.26 LTIP assays were performed over a 3-weekperiod and used the same assay reagents. All samples were incubatedin quadruplicate; duplicate samples received exogenous CETP,whereas the second duplicate was incubated without the additionof CETP.
LTIP activity was calculated by comparing the activity of exog-enously added CETP in tubes containing lipoprotein-deficientplasma (after subtracting the CETP activity measured in sampleswithout added CETP) to tubes containing exogenous CETP alone.To facilitate the comparison of values between assays and to correctfor the nonlinear response of LTIP in this assay,7,10,21these percentinhibition values were then read against a dose-response curvegenerated in each assay by using lipoprotein-deficient plasma pre-pared from pooled normolipidemic plasma (distinct from the plasmapool described in Table 1). An aliquot of this “standard” plasma wasprocessed alongside each group of plasma samples. Because LTIPactivity declines on long-term storage at270°C, LTIP activities arereported as normalized values (ie, relative to the mean value of acontrol group of the same storage age) to facilitate comparison ofsamples stored for different times. The hypercholesterolemic sam-ples are storage-age–matched to those in control 1 (21 subjects);hypercholesterolemic LTIP values are compared with only thiscontrol group to simplify data analysis.
Electrophoresis and Immunoblot AnalysesSamples were separated on 7.5% SDS-PAGE gels, electrotrans-ferred, and reacted with a 1:500 dilution of anti-LTIP antibody 2
TABLE 1. Characteristics of Study Groups
GroupTotal Subjects
n (Male1Female) Age, y BMI, kg/m2
Levels, mg/dL
TC TG
Hypercholesterolemic 23 (1419) 55.4611.7 (34–82) 26.062.7 (20–38) 252.8626.9 (203–314) 159.1628.7 (80–238)
Control 1 21 (3118) 32.765.0 (20–48) 23.862.9 (18–28) 158.0625.9 (116–199) 90.9644.5 (19–223)
Control 2 25 (0125) 26.564.8 (19–35) 22.362.0 (19–25) 157.9623.6 (108–199) 74.6637.3 (35–157)
Control pool 7 (314) 35.966.0 (27–42) ND 172.7625.5 (125–199) 82.0635.4 (38–129)
BMI indicates body mass index; HDLc, HDL cholesterol; LDLc, LDL cholesterol; VLDLc, VLDL cholesterol; and ND, not determined.Values are mean6SD (range).*LDLc1VLDLc.
Morton et al LTIP Elevation in Hypercholesterolemia 1643
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from

antisera as previously described.6 To measure LTIP on intactlipoproteins, 2 methods were used. Lipoproteins were resolved byagarose gel electrophoresis, transferred to nitrocellulose, and reactedwith an equal volume mix of anti-LTIP antibody 1 and antibody 2IgG.6 Alternatively, aliquots (5mL) of isolated lipoproteins contain-ing #1.6 mg protein were applied to nitrocellulose with a slot-blotapparatus. After 30 minutes, wells were washed with 50mL PBS,and then the membrane was blocked and reacted with anti-LTIPantisera as described before.6 Reaction products were visualized withenhanced chemiluminescent reagent (NEN Life Science Products,Inc). X-ray film images were captured by a ScanMaker III (MicrotekLaboratory Inc) scanner and quantified with NIH Image software.
Cell CultureSW872 adipocytes (American Type Culture Collection No. HTB 92)were grown in 100-mm dishes containing DMEM/F-12 medium with5% calf serum and antibiotics. At confluence, cells were washedtwice with serum-free medium and then incubated with the indicatedagent in OptiMEM (GIBCO-BRL). After 24 hours, cells werewashed twice in medium and then incubated in serum-free mediumfor 48 hours. This conditioned medium was centrifuged to removecell debris and then assayed for LTIP activity and mass. LTIPactivity was determined in a transfer assay containing [3H]LDL asdonor and biotinylated LDL (10mg cholesterol each) as acceptor.21
LTIP protein in conditioned medium (concentrated 10 times) wasdetected by SDS-PAGE/Western blot as described above, except thatblots were reacted with 1:700 dilutions of anti-LTIP antibody 1 andantibody 2 antisera. CETP activity in conditioned medium wasmeasured by its capacity to promote CE transfer from LDL to HDL1
during long-term incubation (18 hours). CE transfer activity waslinear with the dose of medium and due fully to CETP, as assessedby monoclonal antibody against the transfer protein. The percentageof CE transfer was calculated as previously described.1
Cholesterol synthesis was measured by determining the extent of[14C]acetate incorporation into cholesterol. Cells, preincubated for 24hours in medium containing 5% lipoprotein-deficient serum, re-ceived 0.5mCi [14C]acetate (NEN Life Science Products, Inc) with orwithout 100 mg LDL in DMEM/F-12 medium for 24 hours.Afterward, cells were washed, and lipids were extracted27 andfractionated by thin-layer chromatography.3 Measurements of TGsynthesis followed the same approach but monitored the incorpora-tion of [9,10-3H(n)]oleic acid (NEN).
Other MethodsProtein, total and free cholesterol, CE, and TGs were quantified bypublished methods or with commercial kits, as previously de-scribed.9 CETP mass was determined by radioimmunoassay28 in thelaboratory of Dr Ruth McPherson (University of Ottawa HeartInstitute, Ottawa, Canada).
A nonparametric Mann-Whitney unpaired test was used to eval-uate the difference between groups, unless indicated otherwise.Reported probability values are based on 2-tailed calculations.Results were considered significant atP,0.05. Linear correlationcoefficients were determined by the Pearson (parametric) method.
ResultsLTIP Activity in Hypercholesterolemic SubjectsIncreased CETP mass and activity in response to hypercho-lesterolemia are well documented.4,19 We have proposed that
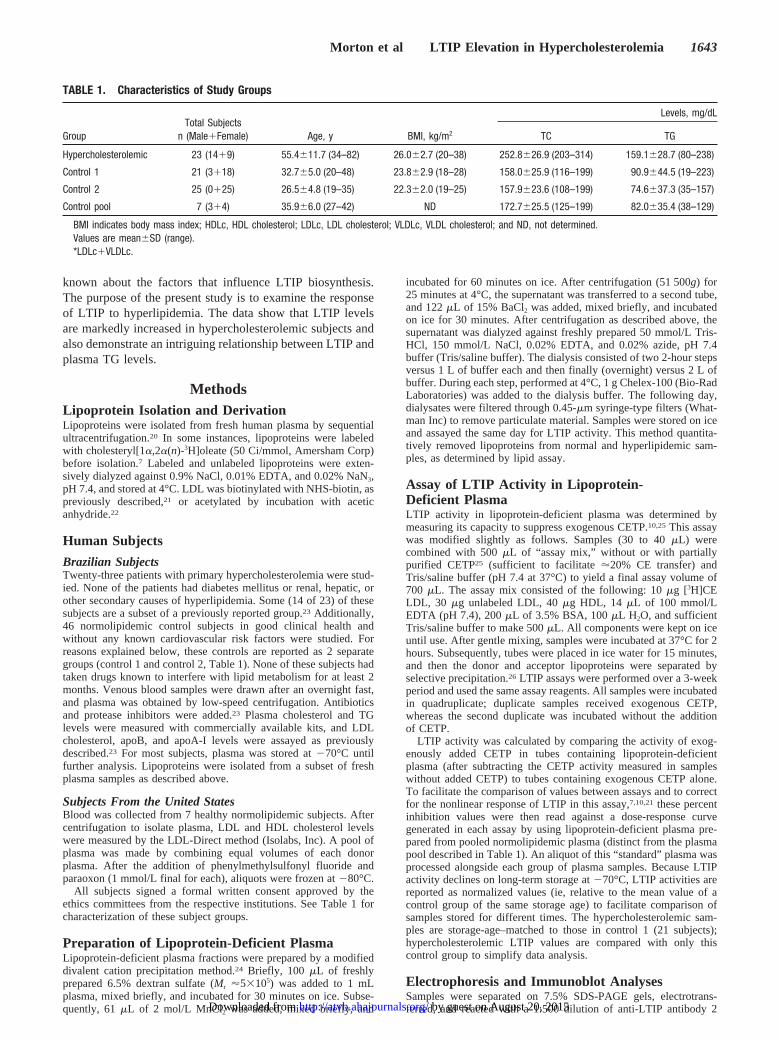
LTIP modulates the activity and function of CETP such thatthe pathways associated with reverse cholesterol transport areenhanced.9,15This suggests that LTIP levels may be increasedunder conditions of cholesterol excess, such as hypercholes-terolemia. To investigate this possibility, LTIP activity wasmeasured in lipoprotein-deficient plasma fractions from con-trol and hyperlipidemic subjects. Plasma cholesterol and TGlevels in hypercholesterolemic subjects average 160% and192% of control values, respectively, although the mean TGlevel of this group was within accepted normal ranges (Table1). LTIP levels in control subjects recruited in Brazil variednearly 4-fold (Figure 1), similar to that previously reportedfor normolipidemic subjects.25 A pool of plasma from 7normolipidemic individuals recruited at the Cleveland Clinichad very similar LTIP levels (Figure 1). The 5 data pointspresented for this plasma pool were determined on separatelyprepared lipoprotein-deficient plasma fractions. Variability inthese assay values (118.5615.2% [mean6SD], n55) reflectsthe combined reproducibility of the lipoprotein-deficientplasma preparation and the LTIP assay. Notably, comparedwith LTIP activity in control subjects, LTIP activity inhypercholesterolemic subjects was increased 3-fold(300.9690.2% [mean6SD],P,0.0001; n514; Figure 1).LTIP activity did not correlate with subject age or weight(body mass index) in either the hypercholesterolemic orcontrol group. Furthermore, LTIP activity in male and femalecontrol subjects was similar (100612.8% [n57] versus91.168.7% [n54], respectively). Thus, it is not apparent,within the limitations of sample size, that the differences inage, body mass index, and sex between these 2 study groupscontribute significantly to the large differences in LTIPactivity observed.
Although not suitable for immunoassays, the anti-LTIPantibodies available6 do permit an estimation of LTIP massby Western blotting. Among 9 individuals, selected fromvarious ongoing studies to represent a wide range in LTIPactivities, immunodetectable LTIP protein in lipoprotein-
Figure 1. LTIP activity in control and hypercholesterolemic sub-jects. LTIP activity was measured in lipoprotein-deficient plasmafractions prepared from control group 1 (n513) and hypercho-lesterolemic (n514) subjects as described in Methods. A pool ofcontrol subjects (control pool, see Table 1) was prepared from 7normolipidemic subjects. LTIP activity values are presented rela-tive to the mean value of the control samples (percentage ofcontrol). Control lipoprotein-deficient plasma (30 mL) suppressedexogenous CETP by 22.9%. Each data point is the mean ofduplicate determinations. Bars reflect the mean value for eachgroup. LTIP activity in hypercholesterolemic subjects was 3-foldhigher than that in control subjects (P,0.0001).
TABLE 1. (Continued)
HDLc LDLc VLDLc
44.869.0 (34–62) 177.0628.8 (128–241) 31.268.8 (16–43)
47.6610.5 (30–72) 93.4621.7 (65–134) 18.869.3 (10–47)
49.0613.1 (32–77) 93.6620.9 (59–128) 15.268.3 (7–41)
44.5614.6 (31–74) 119.8623.3* (87–149) z z z
1644 Arterioscler Thromb Vasc Biol. October 2001
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from
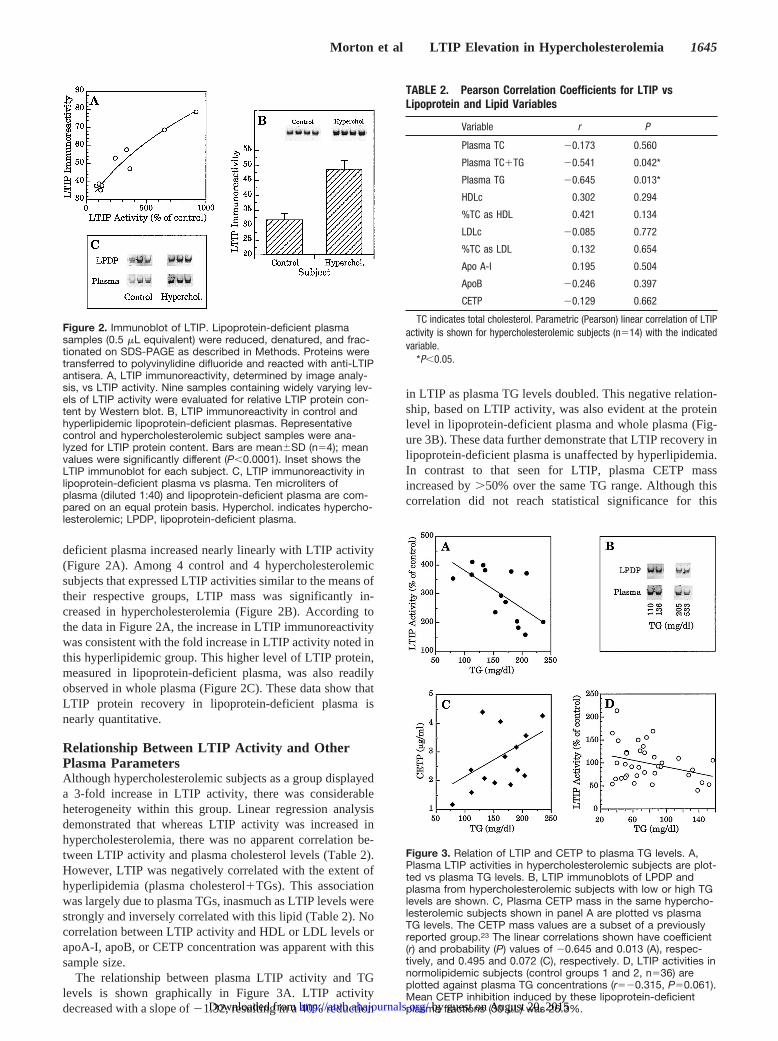
deficient plasma increased nearly linearly with LTIP activity(Figure 2A). Among 4 control and 4 hypercholesterolemicsubjects that expressed LTIP activities similar to the means oftheir respective groups, LTIP mass was significantly in-creased in hypercholesterolemia (Figure 2B). According tothe data in Figure 2A, the increase in LTIP immunoreactivitywas consistent with the fold increase in LTIP activity noted inthis hyperlipidemic group. This higher level of LTIP protein,measured in lipoprotein-deficient plasma, was also readilyobserved in whole plasma (Figure 2C). These data show thatLTIP protein recovery in lipoprotein-deficient plasma isnearly quantitative.
Relationship Between LTIP Activity and OtherPlasma ParametersAlthough hypercholesterolemic subjects as a group displayeda 3-fold increase in LTIP activity, there was considerableheterogeneity within this group. Linear regression analysisdemonstrated that whereas LTIP activity was increased inhypercholesterolemia, there was no apparent correlation be-tween LTIP activity and plasma cholesterol levels (Table 2).However, LTIP was negatively correlated with the extent ofhyperlipidemia (plasma cholesterol1TGs). This associationwas largely due to plasma TGs, inasmuch as LTIP levels werestrongly and inversely correlated with this lipid (Table 2). Nocorrelation between LTIP activity and HDL or LDL levels orapoA-I, apoB, or CETP concentration was apparent with thissample size.
The relationship between plasma LTIP activity and TGlevels is shown graphically in Figure 3A. LTIP activitydecreased with a slope of21.32, resulting in a 40% reduction
in LTIP as plasma TG levels doubled. This negative relation-ship, based on LTIP activity, was also evident at the proteinlevel in lipoprotein-deficient plasma and whole plasma (Fig-ure 3B). These data further demonstrate that LTIP recovery inlipoprotein-deficient plasma is unaffected by hyperlipidemia.In contrast to that seen for LTIP, plasma CETP massincreased by.50% over the same TG range. Although thiscorrelation did not reach statistical significance for this
Figure 3. Relation of LTIP and CETP to plasma TG levels. A,Plasma LTIP activities in hypercholesterolemic subjects are plot-ted vs plasma TG levels. B, LTIP immunoblots of LPDP andplasma from hypercholesterolemic subjects with low or high TGlevels are shown. C, Plasma CETP mass in the same hypercho-lesterolemic subjects shown in panel A are plotted vs plasmaTG levels. The CETP mass values are a subset of a previouslyreported group.23 The linear correlations shown have coefficient(r) and probability (P) values of 20.645 and 0.013 (A), respec-tively, and 0.495 and 0.072 (C), respectively. D, LTIP activities innormolipidemic subjects (control groups 1 and 2, n536) areplotted against plasma TG concentrations (r520.315, P50.061).Mean CETP inhibition induced by these lipoprotein-deficientplasma fractions (30 mL) was 26.3%.
Figure 2. Immunoblot of LTIP. Lipoprotein-deficient plasmasamples (0.5 mL equivalent) were reduced, denatured, and frac-tionated on SDS-PAGE as described in Methods. Proteins weretransferred to polyvinylidine difluoride and reacted with anti-LTIPantisera. A, LTIP immunoreactivity, determined by image analy-sis, vs LTIP activity. Nine samples containing widely varying lev-els of LTIP activity were evaluated for relative LTIP protein con-tent by Western blot. B, LTIP immunoreactivity in control andhyperlipidemic lipoprotein-deficient plasmas. Representativecontrol and hypercholesterolemic subject samples were ana-lyzed for LTIP protein content. Bars are mean6SD (n54); meanvalues were significantly different (P,0.0001). Inset shows theLTIP immunoblot for each subject. C, LTIP immunoreactivity inlipoprotein-deficient plasma vs plasma. Ten microliters ofplasma (diluted 1:40) and lipoprotein-deficient plasma are com-pared on an equal protein basis. Hyperchol. indicates hypercho-lesterolemic; LPDP, lipoprotein-deficient plasma.
TABLE 2. Pearson Correlation Coefficients for LTIP vsLipoprotein and Lipid Variables
Variable r P
Plasma TC 20.173 0.560
Plasma TC1TG 20.541 0.042*
Plasma TG 20.645 0.013*
HDLc 0.302 0.294
%TC as HDL 0.421 0.134
LDLc 20.085 0.772
%TC as LDL 0.132 0.654
Apo A-I 0.195 0.504
ApoB 20.246 0.397
CETP 20.129 0.662
TC indicates total cholesterol. Parametric (Pearson) linear correlation of LTIPactivity is shown for hypercholesterolemic subjects (n514) with the indicatedvariable.
*P,0.05.
Morton et al LTIP Elevation in Hypercholesterolemia 1645
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from
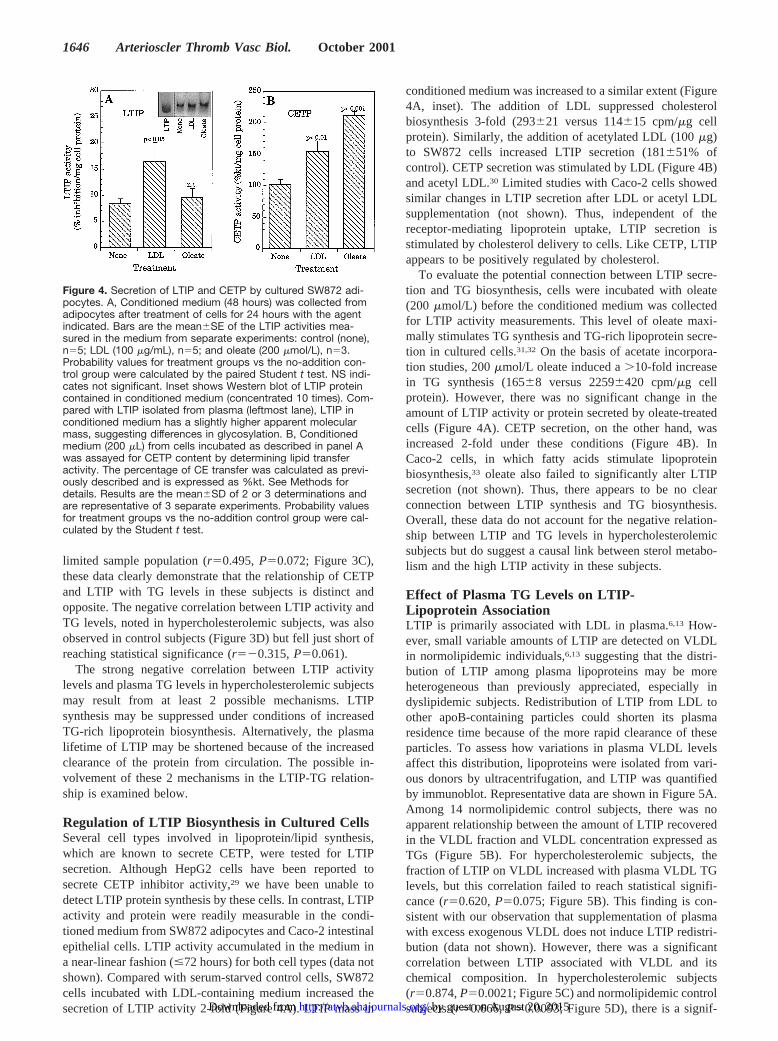
limited sample population (r50.495,P50.072; Figure 3C),these data clearly demonstrate that the relationship of CETPand LTIP with TG levels in these subjects is distinct andopposite. The negative correlation between LTIP activity andTG levels, noted in hypercholesterolemic subjects, was alsoobserved in control subjects (Figure 3D) but fell just short ofreaching statistical significance (r520.315,P50.061).
The strong negative correlation between LTIP activitylevels and plasma TG levels in hypercholesterolemic subjectsmay result from at least 2 possible mechanisms. LTIPsynthesis may be suppressed under conditions of increasedTG-rich lipoprotein biosynthesis. Alternatively, the plasmalifetime of LTIP may be shortened because of the increasedclearance of the protein from circulation. The possible in-volvement of these 2 mechanisms in the LTIP-TG relation-ship is examined below.
Regulation of LTIP Biosynthesis in Cultured CellsSeveral cell types involved in lipoprotein/lipid synthesis,which are known to secrete CETP, were tested for LTIPsecretion. Although HepG2 cells have been reported tosecrete CETP inhibitor activity,29 we have been unable todetect LTIP protein synthesis by these cells. In contrast, LTIPactivity and protein were readily measurable in the condi-tioned medium from SW872 adipocytes and Caco-2 intestinalepithelial cells. LTIP activity accumulated in the medium ina near-linear fashion (#72 hours) for both cell types (data notshown). Compared with serum-starved control cells, SW872cells incubated with LDL-containing medium increased thesecretion of LTIP activity 2-fold (Figure 4A). LTIP mass in
conditioned medium was increased to a similar extent (Figure4A, inset). The addition of LDL suppressed cholesterolbiosynthesis 3-fold (293621 versus 114615 cpm/mg cellprotein). Similarly, the addition of acetylated LDL (100mg)to SW872 cells increased LTIP secretion (181651% ofcontrol). CETP secretion was stimulated by LDL (Figure 4B)and acetyl LDL.30 Limited studies with Caco-2 cells showedsimilar changes in LTIP secretion after LDL or acetyl LDLsupplementation (not shown). Thus, independent of thereceptor-mediating lipoprotein uptake, LTIP secretion isstimulated by cholesterol delivery to cells. Like CETP, LTIPappears to be positively regulated by cholesterol.
To evaluate the potential connection between LTIP secre-tion and TG biosynthesis, cells were incubated with oleate(200 mmol/L) before the conditioned medium was collectedfor LTIP activity measurements. This level of oleate maxi-mally stimulates TG synthesis and TG-rich lipoprotein secre-tion in cultured cells.31,32 On the basis of acetate incorpora-tion studies, 200mmol/L oleate induced a.10-fold increasein TG synthesis (16568 versus 22596420 cpm/mg cellprotein). However, there was no significant change in theamount of LTIP activity or protein secreted by oleate-treatedcells (Figure 4A). CETP secretion, on the other hand, wasincreased 2-fold under these conditions (Figure 4B). InCaco-2 cells, in which fatty acids stimulate lipoproteinbiosynthesis,33 oleate also failed to significantly alter LTIPsecretion (not shown). Thus, there appears to be no clearconnection between LTIP synthesis and TG biosynthesis.Overall, these data do not account for the negative relation-ship between LTIP and TG levels in hypercholesterolemicsubjects but do suggest a causal link between sterol metabo-lism and the high LTIP activity in these subjects.
Effect of Plasma TG Levels on LTIP-Lipoprotein AssociationLTIP is primarily associated with LDL in plasma.6,13 How-ever, small variable amounts of LTIP are detected on VLDLin normolipidemic individuals,6,13 suggesting that the distri-bution of LTIP among plasma lipoproteins may be moreheterogeneous than previously appreciated, especially indyslipidemic subjects. Redistribution of LTIP from LDL toother apoB-containing particles could shorten its plasmaresidence time because of the more rapid clearance of theseparticles. To assess how variations in plasma VLDL levelsaffect this distribution, lipoproteins were isolated from vari-ous donors by ultracentrifugation, and LTIP was quantifiedby immunoblot. Representative data are shown in Figure 5A.Among 14 normolipidemic control subjects, there was noapparent relationship between the amount of LTIP recoveredin the VLDL fraction and VLDL concentration expressed asTGs (Figure 5B). For hypercholesterolemic subjects, thefraction of LTIP on VLDL increased with plasma VLDL TGlevels, but this correlation failed to reach statistical signifi-cance (r50.620,P50.075; Figure 5B). This finding is con-sistent with our observation that supplementation of plasmawith excess exogenous VLDL does not induce LTIP redistri-bution (data not shown). However, there was a significantcorrelation between LTIP associated with VLDL and itschemical composition. In hypercholesterolemic subjects(r50.874,P50.0021; Figure 5C) and normolipidemic controlsubjects (r50.666,P50.0093; Figure 5D), there is a signif-
Figure 4. Secretion of LTIP and CETP by cultured SW872 adi-pocytes. A, Conditioned medium (48 hours) was collected fromadipocytes after treatment of cells for 24 hours with the agentindicated. Bars are the mean6SE of the LTIP activities mea-sured in the medium from separate experiments: control (none),n55; LDL (100 mg/mL), n55; and oleate (200 mmol/L), n53.Probability values for treatment groups vs the no-addition con-trol group were calculated by the paired Student t test. NS indi-cates not significant. Inset shows Western blot of LTIP proteincontained in conditioned medium (concentrated 10 times). Com-pared with LTIP isolated from plasma (leftmost lane), LTIP inconditioned medium has a slightly higher apparent molecularmass, suggesting differences in glycosylation. B, Conditionedmedium (200 mL) from cells incubated as described in panel Awas assayed for CETP content by determining lipid transferactivity. The percentage of CE transfer was calculated as previ-ously described and is expressed as %kt. See Methods fordetails. Results are the mean6SD of 2 or 3 determinations andare representative of 3 separate experiments. Probability valuesfor treatment groups vs the no-addition control group were cal-culated by the Student t test.
1646 Arterioscler Thromb Vasc Biol. October 2001
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from
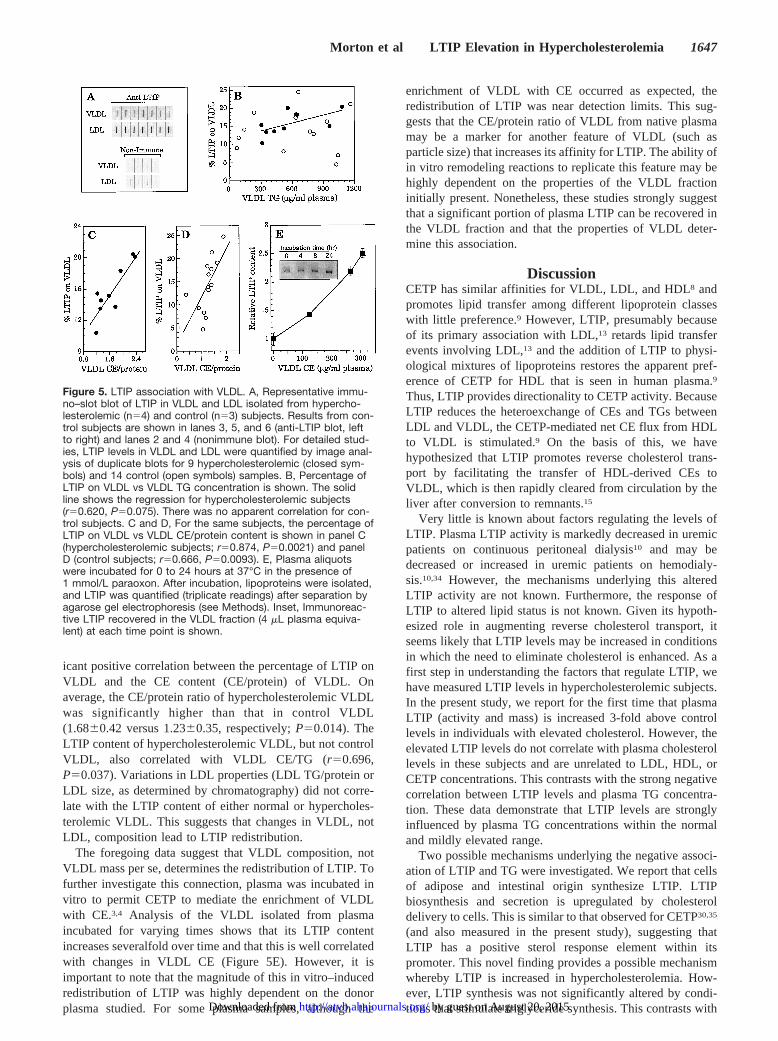
icant positive correlation between the percentage of LTIP onVLDL and the CE content (CE/protein) of VLDL. Onaverage, the CE/protein ratio of hypercholesterolemic VLDLwas significantly higher than that in control VLDL(1.6860.42 versus 1.2360.35, respectively;P50.014). TheLTIP content of hypercholesterolemic VLDL, but not controlVLDL, also correlated with VLDL CE/TG (r50.696,P50.037). Variations in LDL properties (LDL TG/protein orLDL size, as determined by chromatography) did not corre-late with the LTIP content of either normal or hypercholes-terolemic VLDL. This suggests that changes in VLDL, notLDL, composition lead to LTIP redistribution.
The foregoing data suggest that VLDL composition, notVLDL mass per se, determines the redistribution of LTIP. Tofurther investigate this connection, plasma was incubated invitro to permit CETP to mediate the enrichment of VLDLwith CE.3,4 Analysis of the VLDL isolated from plasmaincubated for varying times shows that its LTIP contentincreases severalfold over time and that this is well correlatedwith changes in VLDL CE (Figure 5E). However, it isimportant to note that the magnitude of this in vitro–inducedredistribution of LTIP was highly dependent on the donorplasma studied. For some plasma samples, although the
enrichment of VLDL with CE occurred as expected, theredistribution of LTIP was near detection limits. This sug-gests that the CE/protein ratio of VLDL from native plasmamay be a marker for another feature of VLDL (such asparticle size) that increases its affinity for LTIP. The ability ofin vitro remodeling reactions to replicate this feature may behighly dependent on the properties of the VLDL fractioninitially present. Nonetheless, these studies strongly suggestthat a significant portion of plasma LTIP can be recovered inthe VLDL fraction and that the properties of VLDL deter-mine this association.
DiscussionCETP has similar affinities for VLDL, LDL, and HDL8 andpromotes lipid transfer among different lipoprotein classeswith little preference.9 However, LTIP, presumably becauseof its primary association with LDL,13 retards lipid transferevents involving LDL,13 and the addition of LTIP to physi-ological mixtures of lipoproteins restores the apparent pref-erence of CETP for HDL that is seen in human plasma.9
Thus, LTIP provides directionality to CETP activity. BecauseLTIP reduces the heteroexchange of CEs and TGs betweenLDL and VLDL, the CETP-mediated net CE flux from HDLto VLDL is stimulated.9 On the basis of this, we havehypothesized that LTIP promotes reverse cholesterol trans-port by facilitating the transfer of HDL-derived CEs toVLDL, which is then rapidly cleared from circulation by theliver after conversion to remnants.15
Very little is known about factors regulating the levels ofLTIP. Plasma LTIP activity is markedly decreased in uremicpatients on continuous peritoneal dialysis10 and may bedecreased or increased in uremic patients on hemodialy-sis.10,34 However, the mechanisms underlying this alteredLTIP activity are not known. Furthermore, the response ofLTIP to altered lipid status is not known. Given its hypoth-esized role in augmenting reverse cholesterol transport, itseems likely that LTIP levels may be increased in conditionsin which the need to eliminate cholesterol is enhanced. As afirst step in understanding the factors that regulate LTIP, wehave measured LTIP levels in hypercholesterolemic subjects.In the present study, we report for the first time that plasmaLTIP (activity and mass) is increased 3-fold above controllevels in individuals with elevated cholesterol. However, theelevated LTIP levels do not correlate with plasma cholesterollevels in these subjects and are unrelated to LDL, HDL, orCETP concentrations. This contrasts with the strong negativecorrelation between LTIP levels and plasma TG concentra-tion. These data demonstrate that LTIP levels are stronglyinfluenced by plasma TG concentrations within the normaland mildly elevated range.
Two possible mechanisms underlying the negative associ-ation of LTIP and TG were investigated. We report that cellsof adipose and intestinal origin synthesize LTIP. LTIPbiosynthesis and secretion is upregulated by cholesteroldelivery to cells. This is similar to that observed for CETP30,35
(and also measured in the present study), suggesting thatLTIP has a positive sterol response element within itspromoter. This novel finding provides a possible mechanismwhereby LTIP is increased in hypercholesterolemia. How-ever, LTIP synthesis was not significantly altered by condi-tions that stimulate triglyceride synthesis. This contrasts with
Figure 5. LTIP association with VLDL. A, Representative immu-no–slot blot of LTIP in VLDL and LDL isolated from hypercho-lesterolemic (n54) and control (n53) subjects. Results from con-trol subjects are shown in lanes 3, 5, and 6 (anti-LTIP blot, leftto right) and lanes 2 and 4 (nonimmune blot). For detailed stud-ies, LTIP levels in VLDL and LDL were quantified by image anal-ysis of duplicate blots for 9 hypercholesterolemic (closed sym-bols) and 14 control (open symbols) samples. B, Percentage ofLTIP on VLDL vs VLDL TG concentration is shown. The solidline shows the regression for hypercholesterolemic subjects(r50.620, P50.075). There was no apparent correlation for con-trol subjects. C and D, For the same subjects, the percentage ofLTIP on VLDL vs VLDL CE/protein content is shown in panel C(hypercholesterolemic subjects; r50.874, P50.0021) and panelD (control subjects; r50.666, P50.0093). E, Plasma aliquotswere incubated for 0 to 24 hours at 37°C in the presence of1 mmol/L paraoxon. After incubation, lipoproteins were isolated,and LTIP was quantified (triplicate readings) after separation byagarose gel electrophoresis (see Methods). Inset, Immunoreac-tive LTIP recovered in the VLDL fraction (4 mL plasma equiva-lent) at each time point is shown.
Morton et al LTIP Elevation in Hypercholesterolemia 1647
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from

CETP secretion, which is increased in this situation (Izem andMorton30, Faust and Albers,36 and present study). Theseresults show that LTIP biosynthesis is not decreased underconditions in which TG-rich lipoprotein secretion is en-hanced31,33 and does not support this process as a plausiblemechanism for the negative association of plasma LTIP andTG levels.
An alternative mechanism for the negative correlation ofLTIP and plasma TG levels is that a portion of LTIPredistributes to VLDL as its concentration increases. In-creased association with VLDL could decrease LTIP levels,because this lipoprotein is more rapidly cleared from circu-lation. We observed that LTIP is variably associated with theVLDL fraction when evaluated in multiple individuals butthat this is poorly correlated with the concentration of VLDL(expressed as TGs) in plasma. However, we report theinteresting finding that LTIP association with VLDL is wellcorrelated with the CE content of the VLDL fraction. Thiscorrelation was highly significant for hypercholesterolemicand control subjects. This suggests that LTIP association withVLDL is determined by specific properties of VLDL. Ele-vated CE content of VLDL can result either from an increasein CE within the nascent VLDL particle or from elevatedremodeling by CETP-mediated lipid transfer processes. Bothof these processes are likely in hypercholesterolemia.14,37
However, the success of in vitro attempts to induce theredistribution of LTIP from LDL to VLDL by enrichingVLDL in CE have not been consistently successful and arehighly dependent on the properties of the starting plasma.This strongly suggests that the CE content of plasma VLDLmay generally reflect other properties of VLDL (such asparticle size) that are not easily manipulated in vitro withtransfer reactions. Nonetheless, these studies clearly showthat the distribution of LTIP among VLDL and LDL isvariable and dependent on the physicochemical propertiesof VLDL.
We suggest that the redistribution of LTIP to VLDLparticles may explain the negative correlation of LTIP andplasma TG levels in hypercholesterolemic subjects. Wedemonstrate that LTIP progressively associates with VLDLas it becomes enriched in CE. Although a similar percentageof LTIP is associated with VLDL in hypercholesterolemicand control subjects for a given VLDL CE content (Figure 5Cand 5D), the 3-fold higher LTIP levels in hypercholesterol-emic subjects results in a significant increase in total LTIPassociated with VLDL. Additionally, the CE content issignificantly higher in hypercholesterolemic VLDL than incontrol VLDL, as has been reported in other hyperlipidemicsituations.38,39 This may occur for 2 reasons: First, plasmaCETP concentrations are increased.19,23 Second, because therates of net lipid transfer in plasma are strongly influenced bythe concentration of VLDL TG,14 CETP-mediated remodel-ing of VLDL composition is likely to be increased in thesesubjects and progressively enhanced as TG concentrationsincrease. Because VLDL and its remnants are more rapidlycleared from circulation than LDL, we hypothesize that thenegative correlation of LTIP with plasma TG levels can beexplained by the increased turnover of LTIP from the plasmacompartment that is due to the redistribution of LTIP toparticles with shorter plasma lifetimes. If such a mechanismis verified, this may provide an explanation for the pro-
foundly decreased plasma LTIP levels in patients undergoingcontinuous ambulatory peritoneal dialysis. These patients,because of the infusion of glucose, have significantly in-creased VLDL synthesis and flux40 as well as VLDL levelsthat are elevated in their CE content.10
In summary, we report the novel finding that plasma LTIPlevels are markedly increased in hypercholesterolemic sub-jects and that the extent of this increase is partially mitigatedby plasma elevated TG concentrations. In cultured cells,LTIP synthesis and secretion are increased by cholesteroldelivery, suggesting that the high levels of LTIP in hyper-cholesterolemia reflect the presence of a positive sterolresponse element in the LTIP gene. The negative correlationof LTIP levels with plasma TG levels may be related to apartial redistribution of LTIP from its normal localization onLDL to VLDL, resulting in a more rapid turnover of LTIPfrom the plasma compartment. This plausible testable hypoth-esis is under investigation. Increased turnover of LTIP couldcompromise the antiatherogenic potential of this protein,15
providing another link between elevated plasma TG levelsand coronary heart disease.
AcknowledgmentsThis research was supported by grant HL-60934 from the NationalHeart, Lung, and Blood Institute, National Institutes of Health, anda grant from S. Paulo State Research Foundation (FAPESP, No.99/10735-9). The authors are grateful to Drs Alexandre Carrilho,Eliana C. de Faria, and Águeda Zaratin for the selection and care ofthe patients.
References1. Morton RE, Zilversmit DB. Purification and characterization of lipid
transfer protein(s) from human lipoprotein-deficient plasma.J Lipid Res.1982;23:1058–1067.
2. Hesler CB, Swenson TL, Tall AR. Purification and characterization of ahuman plasma cholesteryl ester transfer protein.J Biol Chem. 1987;262:2275–2282.
3. Morton RE. Interaction of lipid transfer protein with plasma lipoproteinsand cell membranes.Experientia. 1990;46:552–560.
4. Tall A. Plasma lipid transfer proteins.Annu Rev Biochem. 1995;64:235–257.
5. Tall AR. An overview of reverse cholesterol transport.Eur Heart J.1998;19(suppl A):A31–A35.
6. Wang X, Driscoll DM, Morton RE. Molecular cloning and expression oflipid transfer inhibitor protein reveals its identity with apolipoprotein F.J Biol Chem. 1999;274:1814–1820.
7. Morton RE, Zilversmit DB. A plasma inhibitor of triglyceride and cho-lesteryl ester transfer activities.J Biol Chem. 1981;256:11992–11995.
8. Morton RE. Binding of plasma-derived lipid transfer protein tolipoprotein substrates: the role of binding in the lipid transfer process.J Biol Chem. 1985;260:12593–12599.
9. Serdyuk AP, Morton RE. Lipid transfer inhibitor protein defines theparticipation of lipoproteins in lipid transfer reactions: CETP has nopreference for cholesteryl esters in HDL versus LDL.ArteriosclerThromb Vasc Biol. 1999;19:718–726.
10. Serdyuk AP, Morton RE. Lipid transfer inhibitor protein activity defi-ciency in normolipidemic uremic patients on continuous ambulatoryperitoneal dialysis.Arterioscler Thromb Vasc Biol. 1997;17:1716–1724.
11. Yen FT, Deckelbaum RJ, Mann CJ, Marcel YL, Milne RW, Tall AR.Inhibition of cholesteryl ester transfer protein activity by monoclonalantibody: effects on cholesteryl ester formation and neutral lipid masstransfer in human plasma.J Clin Invest. 1989;83:2018–2024.
12. Nichols AV, Smith L. Effect of very low-density lipoproteins on lipidtransfer in incubated serum.J Lipid Res. 1965;6:206–210.
13. Morton RE, Greene DJ. Regulation of lipid transfer between lipoproteinsby an endogenous plasma protein: selective inhibition among lipoproteinclasses.J Lipid Res. 1994;35:836–847.
14. Mann CJ, Yen FT, Grant AM, Bihain BE. Mechanism of plasma cho-lesteryl ester transfer in hypertriglyceridemia.J Clin Invest. 1991;88:2059–2066.
1648 Arterioscler Thromb Vasc Biol. October 2001
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from

15. Morton RE. Cholesteryl ester transfer protein and its plasma regulator:lipid transfer inhibitor protein.Curr Opin Lipidol. 99;10:321–327.
16. Quinet EM, Agellon LB, Kroon PA, Marcel YL, Lee YC, Whitlock ME,Tall AR. Atherogenic diet increases cholesteryl ester transfer proteinmessenger RNA levels in rabbit liver.J Clin Invest. 1990;85:357–363.
17. Jiang XC, Agellon LB, Walsh A, Breslow JL, Tall A. Dietary cholesterolincreases transcription of the human cholesteryl ester transfer proteingene in transgenic mice: dependence on natural flanking sequences.J Clin Invest. 1992;90:1290–1295.
18. Tall A, Granot E, Brocia R, Tabas I, Hesler C, Williams K, Denke M.Accelerated transfer of cholesteryl esters in dyslipidemic plasma.J ClinInvest. 1987;79:1217–1225.
19. McPherson R, Mann CJ, Tall AR, Hogue M, Martin L, Milne RW,Marcel YL. Plasma concentrations of cholesteryl ester transfer protein inhyperlipoproteinemia: relation to cholesteryl ester transfer protein activityand other lipoprotein variables.Arterioscler Thromb. 1991;11:797–804.
20. Havel RJ, Eder HA, Bragdon JH. The distribution and chemical compo-sition of ultracentrifugally separated lipoproteins in human serum.J ClinInvest. 1955;34:1345–1353.
21. Morton RE, Greene DJ. Enhanced detection of lipid transfer inhibitorprotein activity by an assay involving only low density lipoprotein.JLipid Res. 1994;35:2094–2099.
22. Fraenkal-Conrat H. Methods for investigating the essential groups forenzyme activity.Methods Enzymol. 1957;4:247–269.
23. Carrilho AJF, Medina WL, Nakandakare ER, Quintão ECR. Plasmacholesteryl ester transfer protein is lowered by treatment of hypercholes-terolemia with cholesteryramine.Clin Pharmacol Ther. 97;62:82–88.
24. Burstein M, Scholnick HR, Morfin R. Rapid method for the isolation oflipoproteins from human serum by precipitation with polyanions.J LipidRes. 1970;11:583–595.
25. Morton RE, Steinbrunner JV. Determination of lipid transfer inhibitorprotein activity in human lipoprotein-deficient plasma.ArteriosclerThromb. 1993;13:1843–1851.
26. Morton RE, Zilversmit DB. The separation of apolipoprotein D fromcholesteryl ester transfer protein.Biochim Biophys Acta. 1981;663:350–355.
27. Bligh EG, Dyer WJ. A rapid method of total lipid extraction and purifi-cation.Can J Biochem Phys. 59;37:911–917.
28. Marcel YL, McPherson R, Hogue M, Czarnecka H, Zawadzki Z, WeechPK, Whitlock ME, Tall AR, Milne RW. Distribution and concentration ofcholesteryl ester transfer protein in plasma of normolipemic subjects.J Clin Invest. 1990;85:10–17.
29. Faust RA, Cheung MC, Albers JJ. Secretion of cholesteryl ester transferprotein-lipoprotein complexes by human HepG2 hepatocytes.Atheroscle-rosis. 1989;77:77–82.
30. Izem, L, Morton RE. Regulation of CETP biosynthesis in SW872 adi-pocytes and Caco2 enterocytes.Circulation. 99;100(suppl I):I-256.Abstract.
31. Field FJ, Albright E, Mathur SN. Regulation of triglyceride-richlipoprotein secretion by fatty acids in CaCo-2 cells.J Lipid Res. 1988;29:1427–1437.
32. Bostrom K, Boren J, Wettesten M, Sjoberg A, Bondjers G, Wiklund O,Carlsson P, Olofsson SO. Studies on the assembly of apo B-100-containing lipoproteins in HepG2 cells.J Biol Chem. 88;263:4434–4442.
33. Levy E, Mehran M, Siedman E. Caco-2 cells as a model for intestinallipoprotein synthesis and secretion.FASEB J. 1995;9:626–635.
34. Mendez AJ, Perez GO, Hsia SL. Defect in cholesteryl ester transport inserum of patients with uremia receiving maintenance hemodialysis:increased inhibitor activity for cholesteryl ester transfer.J Lab Clin Med.1988;111:677–683.
35. Richardson MA, Berg DT, Johnston PA, McClure D, Grinnell BW.Human liposarcoma cell line, SW872, secretes cholesteryl ester transferprotein in response to cholesterol.J Lipid Res. 1996;37:1162–1166.
36. Faust RA, Albers JJ. Regulated vectorial secretion of cholesteryl estertransfer protein (LTP-1) by the CaCo-2 model of human enterocyteepithelium.J Biol Chem. 1988;263:8786–8789.
37. Parks JS, Rudel LL. Effect of fish oil on atherosclerosis and lipoproteinmetabolism.Atherosclerosis. 1990;84:83–94.
38. Bagdade JD, Ritter MC, Subbaiah PV. Accelerated cholesteryl estertransfer in plasma of patients with hypercholesterolemia.J Clin Invest.1991;87:1259–1265.
39. Bagdade JD, Ritter MC, Davidson M, Subbaiah PV. Effect of marinelipids on cholesteryl ester transfer and lipoprotein composition in patientswith hypercholesterolemia.Arterioscler Thromb. 1992;12:1146–1152.
40. Thomas ME, Moorhead JF. Lipids in CAPD. a review. In: Coles GA,Davies M, Williams JD, eds.CAPD: Host Defense, Nutrition and Ultra-filtration. Basel, Switzerland: Karger; 1990:92–99.
Morton et al LTIP Elevation in Hypercholesterolemia 1649
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from

Richard E. Morton, Valéria Nunes, Lahoucine Izem and Eder QuintãoMitigated by Plasma Triglyceride Levels
Markedly Elevated Lipid Transfer Inhibitor Protein in Hypercholesterolemic Subjects Is
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2001 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/hq1001.0967222001;21:1642-1649Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/21/10/1642World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on August 20, 2015http://atvb.ahajournals.org/Downloaded from
Related Documents











