Editor’s Choice The Editor takes a closer look at some of this month’s articles Safety of inhaled steroids: watch the dose The inhaled route for lipophilic topically active steroids has transformed the management of airway disease providing a safe and effective alternative to systemic treatment with glucocorticoids. However their excellent safety record has led to a tendency to prescribe in doses that are higher than the risk-benefit ratio would warrant. The paper by Weatherall and colleagues (pp. 1451–1458) pointing out a dose response relationship between the risk of fractures and inhaled steroid dose is a timely reminder that these drugs are not without their dangers and the minimum dose to control airway inflammation should be used. Ion channels: a new target for asthma The complexity of ion channel biology is bewildering to non-experts yet at the core of all cellular functions is a flow of ions across the cell membrane. It is axiomatic therefore that both the abnormalities in immune regulation and the disordered function in structural cells that underlies the asthma process are controlled by ion channels. In a novel and exciting observation Sel et al (pp. 1548–1558) have found that the channel TRPC6 which is highly expressed in the lung is involved in both airway hyperresponsiveness and ovalbumin induced inflammation, offering a new target for therapeutic intervention. Histamine3 receptor: a new tune for an old mediator Histamine is the grandfather of allergy-associated mediators but the discovery that there are at least four histamine receptors has opened up new lines of investigation and treatment. However our un- derstanding of the role of the H3 and H4 receptors in allergic disease remains limited. This interesting study by Suzuki et al (pp. 1476–1482) makes the case for histamine receptors being present on mucus glands and being involved in mucus hyper- secretion, an aspect of rhinitis not well controlled with H1 blockers. We await a new class of anti- histamines for the allergy clinic. Gene expression in allergy: CISH goes the Th2 cell With hens egg allergy as a model using gene chip technology, Nakajima and colleagues (pp. 1499–1506) have investigated the genes differentially expressed in Th2 cells. They found that the gene most closely associated with allergy to hens egg was CIS(H) a member of the suppressors of cytokine signalling (SOCS) family of signal transduction molecules. This is one of the first times that this molecule has been implicated in Th2 function and opens up a new avenue to regulate the activation of these key allergy related cells. Cows milk allergy: soybean may not be the answer Cows milk allergy is a major cause of morbidity in infants and parents often turn to soybean as an alternative. In this study Curciarello et al (pp. 1559–1565) have investigated the cross-reactive components of conventional soybean and a cultivar (Raiden) which may be less allergenic. They found that while Raiden contains less cross- reactive components than conventional soybean both strains contain a subunit-b-conglycinin which cross- reacted with cows milk specific IgE antibodies. Back to the drawing board. EC This logo highlights the Editor’s Choice articles on the cover and the first page of each of the articles. Clinical and Experimental Allergy Volume 38, September 2008 Immunohistochemistry for histamine H3 receptor in human nasal mucosa with allergic rhinitis (see fig 2 in Suzuki et al, pp. 1476–1482).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Editor’s ChoiceThe Editor takes a closer look at some of this month’s articles
Safety of inhaled steroids: watch the dose
The inhaled route for lipophilic topically active steroids has transformed the management of airway diseaseproviding a safe and effective alternative to systemic treatment with glucocorticoids. However their excellent safetyrecord has led to a tendency to prescribe in doses that are higher than the risk-benefit ratio would warrant. Thepaper by Weatherall and colleagues (pp. 1451–1458) pointing out a dose response relationship between the risk offractures and inhaled steroid dose is a timely reminder that these drugs are not without their dangers and theminimum dose to control airway inflammation should be used.
Ion channels: a new target for asthma
The complexity of ion channel biology is bewildering to non-experts yet at the core of all cellular functions is a flowof ions across the cell membrane. It is axiomatic therefore that both the abnormalities in immune regulation and thedisordered function in structural cells that underlies the asthma process are controlled by ion channels. In a noveland exciting observation Sel et al (pp. 1548–1558) have found that the channel TRPC6 which is highly expressed inthe lung is involved in both airway hyperresponsiveness and ovalbumin induced inflammation, offering a newtarget for therapeutic intervention.
Histamine3 receptor: a new tune for an oldmediator
Histamine is the grandfather of allergy-associatedmediators but the discovery that there are at leastfour histamine receptors has opened up new linesof investigation and treatment. However our un-derstanding of the role of the H3 and H4 receptorsin allergic disease remains limited. This interestingstudy by Suzuki et al (pp. 1476–1482) makes thecase for histamine receptors being present onmucus glands and being involved in mucus hyper-secretion, an aspect of rhinitis not well controlledwith H1 blockers. We await a new class of anti-histamines for the allergy clinic.
Gene expression in allergy: CISH goes the Th2 cell
With hens egg allergy as a model using gene chip technology, Nakajima and colleagues (pp. 1499–1506) haveinvestigated the genes differentially expressed in Th2 cells. They found that the gene most closely associated withallergy to hens egg was CIS(H) a member of the suppressors of cytokine signalling (SOCS) family of signaltransduction molecules. This is one of the first times that this molecule has been implicated in Th2 function andopens up a new avenue to regulate the activation of these key allergy related cells.
Cows milk allergy: soybean may not be the answer
Cows milk allergy is a major cause of morbidity in infants and parents often turn to soybean as an alternative.In this study Curciarello et al (pp. 1559–1565) have investigated the cross-reactive components of conventionalsoybean and a cultivar (Raiden) which may be less allergenic. They found that while Raiden contains less cross-reactive components than conventional soybean both strains contain a� subunit-b-conglycinin which cross-reacted with cows milk specific IgE antibodies. Back to the drawing board.
�EC This logo highlights the Editor’s Choice articles on the cover and the first page of each of the articles.
Clinical and Experimental Allergy Volume 38, September 2008
Immunohistochemistry for histamine H3 receptor in human nasalmucosa with allergic rhinitis (see fig 2 in Suzuki et al, pp. 1476–1482).

ORIGINAL PAPER
Loss of classical transient receptor potential 6 channel reduces allergicairway responseS. Sel�1, B. R. Rostw1, A. O. Yildirimz, B. Sel�, H. Kalwaw, H. Fehrenbachz, H. Renz�, T. Gudermannw and A. Dietrichw�Department of Clinical Chemistry and Molecular Diagnostics, Philipps—University Marburg, Marburg, Germany, wInstitute for Pharmacology and Toxicology,
Philipps—University Marburg, Marburg, Germany and zClinical Research Group, Chronic Airway Diseases, Medical Faculty, Philipps-University Marburg,
Marburg, Germany
Clinical andExperimental
Allergy
Correspondence:Alexander Dietrich, Institute forPharmacology and Toxicology,Philipps—University Marburg, Karl-von-Frisch-Str.1, 35033 Marburg, Germany.E-mail: [email protected] this as: S. Sel, B. R. Rost, A. O.Yildirim. B. Sel, H. Kalwa, H. Fehrenbach,H. Renz, T. Gudermann and A. Dietrich,Clinical and Experimental Allergy, 2008(38) 1548–1558.
Summary
Background Non-selective cation influx through canonical transient receptor potentialchannels (TRPCs) is thought to be an important event leading to airway inflammation.TRPC6 is highly expressed in the lung, but its role in allergic processes is still poorlyunderstood.Objective The purpose of this study was to evaluate the role of TRPC6 in airwayhyperresponsiveness (AHR) and allergic inflammation of the lung.Methods Methacholine-induced AHR was assessed by head-out body plethysmography ofwild type (WT) and TRPC6� /� mice. Experimental airway inflammation was induced byintraperitoneal ovalbumin (OVA) sensitization, followed by OVA aerosol challenges. Allergicinflammation and mucus production were analysed 24 h after the last allergen challenge.Results Methacholine-induced AHR and agonist-induced contractility of tracheal rings wereincreased in TRPC6� /� mice compared with WT mice, most probably due to compensatoryup-regulation of TRPC3 in airway smooth muscle cells. Most interestingly, when comparedwith WT mice, TRPC6� /� mice exhibited reduced allergic responses after allergen challengeas evidenced by a decrease in airway eosinophilia and blood IgE levels, as well as decreasedlevels of T-helper type 2 (Th2) cytokines (IL-5, IL-13) in the bronchoalveolar lavage. However,lung mucus production after allergen challenge was not altered by TRPC6 deficiency.Conclusions TRPC6 deficiency inhibits specific allergic immune responses, pointing to animportant immunological function of this cation channel in Th2 cells, eosinophils, mast cellsand B cells.
Keywords airway hyperresponsiveness, B cells, eosinophils, IgE, IL-5, IL-13, ovalbumin-induced experimental airway inflammation, Th2 cells, TRPC6Submitted 6 September 2007; revised 9 April 2008; accepted 10 April 2008
Introduction
The transient receptor potential (TRP) family of ionchannels is a growing group of structurally and evolutio-narily related cation channels comprised of several sub-families including the TRPC, TRPM and TRPV classes ofchannels [1, 2]. TRP-type ion channels are assembled ashomo- or heterotetramers of subunits, each spanning theplasma membrane six times. The founding members ofthis channel family are the insect TRP and TRPL channels,which are responsible for photoreceptor depolarization in
response to light. Mammalian TRPCs (C stands for cano-nical or classical) are the closest mammalian structuralrelatives of insect TRPs [2]. Among the TRPC channels,TRPC3, 6 and 7 are 75% identical and gated by signaltransduction pathways that activate C-type phospholi-pases (PLCs) [3, 4], as well as by direct exposure todiacylglycerols (DAG) [5]. TRPC3, 6 and 7 interact physi-cally and, upon co-expression, co-assemble to form func-tional channels [6].
In contrast to members of other TRP families, thefunctional importance of most members of the TRPCsubfamily is still poorly understood. A TRPC channel forwhich considerable evidence has accumulated for a
Experimental Models of Allergic Disease
�EC
1These authors contributed equally to the work.
Clinical and Experimental Allergy, 38, 1548–1558doi: 10.1111/j.1365-2222.2008.03043.x
�c 2008 The Authors
Journal compilation �c 2008 Blackwell Publishing Ltd

specific role is TRPC6, which has been proposed toregulate smooth muscle function [7]. The TRPC6 cDNAwas originally isolated from mouse brain, but TRPC6mRNA is also highly expressed in the lung [8]. AlthoughTRPC6 expression has been demonstrated in human air-way smooth muscle cells and differentiated bronchialsmooth muscle cells [9], as well as in pulmonary arterialsmooth muscle cells [10], data for a physiological role ofTRPC6 in lung are rare. Recently, we were able todemonstrate a unique physiological role of TRPC6 in theregulation of the acute hypoxic vasoconstriction byanalysing a TRPC6-deficient mouse model [11]. These datawere in clear contrast to the systemic vasculature, whereup-regulation of TRPC3 was partially able to compensatefor the TRPC6 deficiency resulting in higher contractilityof aortic smooth muscle cells, as well as smooth musclecells from cerebral arteries [12].
TRPC6 is also expressed in neutrophils [13], eosinophils[14] and T lymphocytes [15], which are involved in airwayinflammation leading to pathophysiological processes likeasthma and chronic obstructive pulmonary disease(COPD) in the lung. However, experimental data support-ing a unique functional role of TRPC6 expressed inleucocytes [13–15] are lacking.
For this reason, we analysed agonist-induced airwayhyperresponsiveness (AHR) and allergic airway inflam-mation [16] in a TRPC6-deficient mouse model. Whileairway contractility was enhanced in TRPC6� /� micecompared with wild-type (WT) mice, most probably be-cause of up-regulation of TRPC3 in the lung, antigen-specific IgE blood levels, as well as Th2 cytokine (IL-5 andIL-13) levels in the bronchoalveolar lavage (BAL) weredecreased in ovalbumin (OVA)-sensitized and -challengedTRPC6-deficient mice. Moreover, airway eosinophiliawas decreased in TRPC6-deficient mice, while lung mucusproduction after allergic inflammation was not altered.In conclusion, we provide first experimental evidencethat TRPC6 has a unique non-redundant role in allergicairway inflammation and may represent a new drugtarget for airway inflammatory diseases like asthma andCOPD.
Material and methods
Animals
TRPC6� /� mice and their WT litter mates were bred andgenotyped as described previously [12]. For allergic sensi-tization and airway allergen challenges, the targetedmutation was crossed 10 times into the BALB/c back-ground to establish an inbred colony carrying theTRPC6� /� allele. Age- and sex-matched mice were usedin all experiments. All animal experiments were approvedby the local animal care committee.
Head-out body plethysmography
Bronchial responsiveness (BR) to b-methacholine wasessentially tested using head-out body plethysmographyas described before [17]. Methacholine was aerosolizedwith a nebulizer in increasing concentrations from 0 to200 mg/mL in phosphate-buffered saline (PBS) and themethacholine concentration that caused a 50% reductionin baseline midexpiratory airflow (MCh50 mg/mL) isreported.
Assessment of airway smooth muscle responsiveness byanalysis of the contractility of tracheal rings
Airway smooth muscle responsiveness was assessed bystimulation with increasing concentrations of methacho-line as described [18].
Isolation of smooth muscle cells and epithelial cells frommurine tracheae
Airway smooth muscle cells from murine tracheae wereisolated as described for tracheae from guinea-pigs [19]and identified by their immunoreactivity with antibodiesagainst a-smooth muscle actin. Epithelial cells were isolatedand cultured on collagen-coated transwells as described[20] in MTEC medium [DMEM-Ham’s F12 (1 : 1 v/v)medium with penicillin/streptomycin] with a supplemen-tal mix for airway epithelial cell growth medium(C-21060; Promocell, Heidelberg, Germany).
Cultivation of cell lines and mast cells
Cell lines EL4 (murine T lymphocytes; ATCC no. TIB-39),RAW 264.7 (murine macrophages; ECACC no. 85011428)and WEHI-231 (lymphoblasts from a mouse B cell lympho-ma, ECACC no. 85022107) were cultured according to theinstructions of the suppliers [American Tissue CultureCollection (ATCC), Manassas, VA, USA, and EuropeanCollection of Cell Cultures (ECAC), provided by Sigma-Aldrich, Taufkirchen, Germany]. First-strand cDNA of pri-mary mast cells, which were cultured as described [21], wasa kind gift from Dr R. Mailhammer (GSF-National ResearchCenter for Environmet and Health, Munich, Germany).
Quantitative real-time polymerase chain reaction
Total RNA and first-strand synthesis and real-time PCRwas carried out as described [12]. The following primerspairs were used for the amplification of specific fragmentsfrom the first-strand synthesis: TRPC1: C1F (50-TGG GCCCAC TGC AGA TTT CAA) and C1R (50-AAG ATG GCC ACGTGC GCT AAG GAG); TRPC2: C2F (50-TTG CCT CCC TCATCT TCC TCA CCA) and C2R (50-CCG CAA GCC CTC GATCCA CAC CT), TRPC3: C3F (50-AGC CGA GCC CCT GGA
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
TRPC6� /� mice show reduced allergic airway responses 1549

AAG ACA C) and C3R (50-CCG ATG GCG AGG AAT GGAAGA C); TRPC4: C4F (50-GGG CGG CGT GCT GCT GAT)and C4R (50-CCG CGT TGG CTG ACT GTA TTG TAG);TRPC5: C5F (50-AGT CGC TCT TCT GGT CTG TCT TT) andC5R (50-TTT GGG GCT GGG AAT AAT G); TRPC6: C6F (50-GAC CGT TCA TGA AGT TTG TAG CAC) and C6R (50-AGTATT CTT TGG GGC CTT GAG TCC), TRPC7: C7F (50-GTGGGC GTG CTG GAC CTG) and C7R (50-AGA CTG TTG CCGTAA GCC TGA GAG) as well as glyceraldehydephosphatedehydrogenase (GAPDH): GAPDHF (50-GTG AAG GTCGGT GTG AAC G) and GAPDHR (50-GGT GAA GAC ACCAGT AGA CTC) and b-actin ACTF, 50-CCA ACC GTG AAAAGA TGA CC, ACTR, 50-GTG GTA CGA CCA GAG GCATAC as reference genes. All primers were tested usingdiluted first-strand synthesis from brain RNA (10–1000-fold) to confirm linearity of the reaction. Real-time PCRwas performed using the 2� master mix from theQuantitect SYBR Green PCR kit (Qiagen, Hilden, Germany)containing a HotStar Taq polymerase, buffer, nucleotides,5 mM MgCl2 (final 2.5 mM) and SYBR Green. Ten picomolesof each primer pair and 0.2 mL from the first-strandsynthesis were added to the reaction mixture and PCRwas carried out in a light-cycler apparatus (Roche, Man-nheim, Germany) using the following conditions: 15 minof initial activation and 45 cycles of 12 s at 94 1C, 30 s at50 1C, 30 s at 72 1C and 10 s at 80 1C each. Fluorescenceintensities were recorded after the extension step at 80 1Cafter each cycle to exclude the fluorescence of primerdimers melting at temperatures lower than 80 1C. Samplescontaining primer dimers were excluded by melting curveanalysis and identification of the products by agarose gelelectrophoresis. Crossing points were determined by asoftware program. The relative gene expression wasquantified using the formula: (2e(crossing point GAPDH� cross-
ing point X))� 100 = % of reference gene expression.
Ba21 influx experiments, design and electroporation ofsiRNA vectors expressing small interfering RNAs
Cells were loaded with fura-2-acetoxymethylester andanalysed as described previously [12, 22]. The design andsequences for the production of an siRNA homologous toTRPC3 are described elsewhere [12]. The unrelated siRNA(50-ACC GAC GAG CAG CTA GAA C) was selected by itslow homology to sequences of a genomewide screen.Plasmids containing the siRNA expression cassettes underthe control of an H1 promoter were electroporated using anucleofactor (Amaxa GmbH, Cologne, Germany) and theprotocol and solutions for human aortic smooth musclecells as described by the supplier.
Allergic sensitization and airway allergen challenges
TRPC6� /� BALB/c mice and WT mice were sensitized toovalbumin (OVA) by three intraperitoneal (i.p.) injections
of OVA adsorbed to aluminium hydroxide (Pierce Biotech-nology, Rockford, IL, USA) and diluted in PBS (50 mg OVAgrade VI per injection; Sigma, Tauf-Kirchen, Germany) ondays 0, 14 and 21. Mice were challenged with 1% OVAaerosols in PBS on days 27, 28, and 29 before analysis ondays 30 and 31. Sham sensitization was carried out withsterile aluminium hydroxide in PBS, and PBS aerosolswere used as sham challenges.
Measurements of ovalbumin-specific serumimmunoglobulin levels
Blood was drawn from the tail vein and OVA-specific Iglevels (IgE, IgG1 and IgG2a) were determined using ELISAtechniques as described previously [23].
Isolation of bronchoalveolar lavage fluid andquantification of eosinophilia and cytokine levels
Sensitized and airway allergen-challenged mice werekilled 24–48 h after the last challenge, and the tracheaswere cannulated. BAL was performed by flushing thelungs and airways three times with 800 mL of F12KNutrient Mixture (Invitrogen, Carlsbad, CA, USA) supple-mented with 15% foetal calf serum (FCS), 1% glutamine,100 U/mL penicillin and 100 mg/mL streptomycin (PAALaboratories, Colbe, Germany). Cells were spun down ontoglass slides with a cytospin (Thermo Electron Corp.,Waltham, MA, USA), followed by staining with Diff-Quick(Dade, Marburg, Germany), to identify immune cells. Thepercentage of leucocytes was determined microscopicallyusing standard cytological criteria. In cell-free lavagefluids, IL-5 (BD Biosciences, San Jose, CA, USA) andIL-13 (Invitrogen) were measured by ELISA according tothe manufacturer’s instructions.
In vitro ovalbumin restimulation of splenic mononuclearcells
Splenic mononuclear cells (MNCs) were isolated andrestimulated with OVA as described previously [23]. TotalRNA and first-strand synthesis and real-time PCR wascarried out as described [12]. Cytokine levels in cellculture supernatants were analysed as described above.
Flow cytometric analysis of single-cell suspensions fromthe bone marrow, thymus and spleen
The spleen and thymus from control and TRPC6� /� micewere removed and placed in 60-mm culture dishes con-taining resuspension buffer (PBS supplemented with 1%FCS). Single-cell suspensions of the spleen and thymuswere prepared by passing disrupted organs through a 70 mmmesh nylon filter. The bone marrow was flushed from the
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
1550 S. Sel et al
femur and tibia with a 23-gauge needle. The remainingcell aggregates were removed by passing cell suspensionsthrough a 70-mm nylon mesh filter. After washing, cellswere resuspended in PBS at 1�107 cells/mL.One� 106 cells were incubated in PBS containing 5% ofrat serum for 5 min at room temperature and subsequentlyincubated with the appropriate antibody coupled to fluor-escent dyes for 10 min at room temperature. After addingFACS-lysis buffer (BD Bioscience) and an additionalincubation for 10 min at room temperature, cells werewashed once and analysed using a FACSortTM flowcytometer and CELLQuestTM software (BD Bioscience).The following antibodies were used: fluorescein isothio-cyanate (FITC)-conjugated rat anti-mouse CD45, PE-con-jugated rat anti-mouse CD45, PE-conjugated rat anti-mouse CD8, PerCP-conjugated rat anti-mouse CD45R,PerCP-conjugated rat anti-mouse CD4 and FITC-conju-gated rat anti-mouse Ly-6G. All antibodies were pur-chased from BD Bioscience.
Analysis of mucus production
The lungs were formaldehyde fixed by instillation at aconstant pressure of a 20-cm fluid column. Subsequently,the trachea was ligated, and the lungs were immersed infixative. Organ slices, tissue block and fields of view werecollected according to the principle of ‘systematic uniformrandom sampling’ in order to assure that the sample wasrepresentative of the whole organ. Paraffin-embedded lungsections were stained with Periodic acid–Schiff (PAS). Thevolume of PAS-stained epithelial mucus (Vmucus) per sur-face area of airway epithelial basal membrane (Sep) wasdetermined by means of point and intersection countingusing a computer-assisted stereology tool box [24] (CAST-Grid 2.0, Olympus, Kobenhavn, Denmark) and subse-quent calculation according to the formula: Vmucus/Sep =LpSPmucus/2SIep with SPmucus = sum of all points hittingmucus, SIep = sum of all intersections of test lines withepithelial basal membrane, and Lp = test-line length atfinal magnification.
Statistics
Results are presented as standard error of the mean (SEM).Student’s unpaired t-test was used to determine thesignificance of differences between groups.
Results
Increased airway responsiveness and contractility oftracheal rings in TRPC6� /� mice
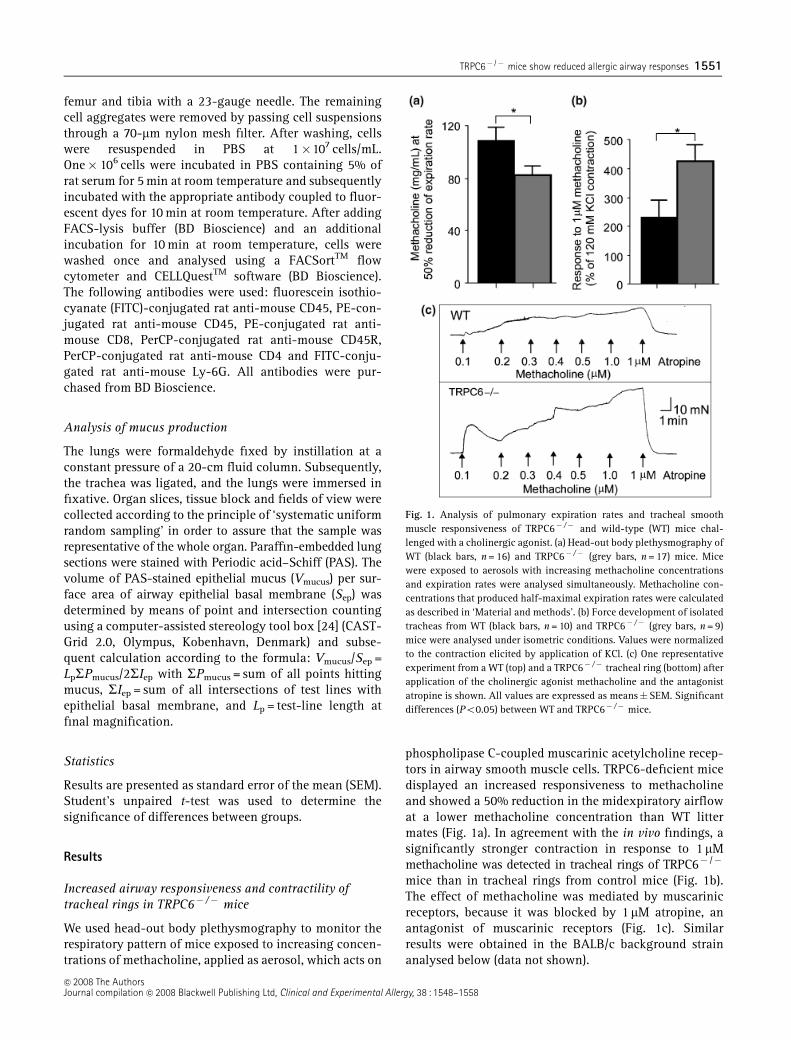
We used head-out body plethysmography to monitor therespiratory pattern of mice exposed to increasing concen-trations of methacholine, applied as aerosol, which acts on
phospholipase C-coupled muscarinic acetylcholine recep-tors in airway smooth muscle cells. TRPC6-deficient micedisplayed an increased responsiveness to methacholineand showed a 50% reduction in the midexpiratory airflowat a lower methacholine concentration than WT littermates (Fig. 1a). In agreement with the in vivo findings, asignificantly stronger contraction in response to 1 mMmethacholine was detected in tracheal rings of TRPC6� /�
mice than in tracheal rings from control mice (Fig. 1b).The effect of methacholine was mediated by muscarinicreceptors, because it was blocked by 1 mM atropine, anantagonist of muscarinic receptors (Fig. 1c). Similarresults were obtained in the BALB/c background strainanalysed below (data not shown).
Fig. 1. Analysis of pulmonary expiration rates and tracheal smoothmuscle responsiveness of TRPC6� /� and wild-type (WT) mice chal-lenged with a cholinergic agonist. (a) Head-out body plethysmography ofWT (black bars, n = 16) and TRPC6� /� (grey bars, n = 17) mice. Micewere exposed to aerosols with increasing methacholine concentrationsand expiration rates were analysed simultaneously. Methacholine con-centrations that produced half-maximal expiration rates were calculatedas described in ‘Material and methods’. (b) Force development of isolatedtracheas from WT (black bars, n = 10) and TRPC6� /� (grey bars, n = 9)mice were analysed under isometric conditions. Values were normalizedto the contraction elicited by application of KCl. (c) One representativeexperiment from a WT (top) and a TRPC6� /� tracheal ring (bottom) afterapplication of the cholinergic agonist methacholine and the antagonistatropine is shown. All values are expressed as means�SEM. Significantdifferences (Po0.05) between WT and TRPC6� /� mice.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
TRPC6� /� mice show reduced allergic airway responses 1551
Up-regulated TRPC3-levels in tracheal smooth musclecells of transient receptor potential channel 6-deficientmice
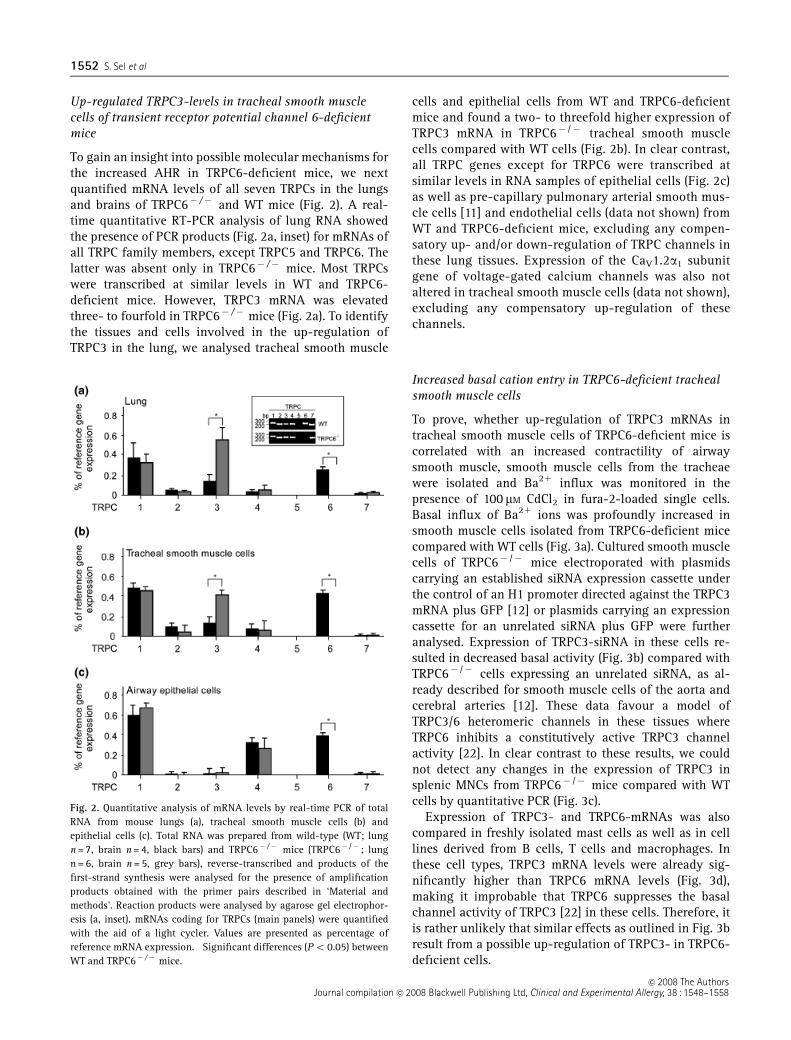
To gain an insight into possible molecular mechanisms forthe increased AHR in TRPC6-deficient mice, we nextquantified mRNA levels of all seven TRPCs in the lungsand brains of TRPC6� /� and WT mice (Fig. 2). A real-time quantitative RT-PCR analysis of lung RNA showedthe presence of PCR products (Fig. 2a, inset) for mRNAs ofall TRPC family members, except TRPC5 and TRPC6. Thelatter was absent only in TRPC6� /� mice. Most TRPCswere transcribed at similar levels in WT and TRPC6-deficient mice. However, TRPC3 mRNA was elevatedthree- to fourfold in TRPC6� /� mice (Fig. 2a). To identifythe tissues and cells involved in the up-regulation ofTRPC3 in the lung, we analysed tracheal smooth muscle
cells and epithelial cells from WT and TRPC6-deficientmice and found a two- to threefold higher expression ofTRPC3 mRNA in TRPC6� /� tracheal smooth musclecells compared with WT cells (Fig. 2b). In clear contrast,all TRPC genes except for TRPC6 were transcribed atsimilar levels in RNA samples of epithelial cells (Fig. 2c)as well as pre-capillary pulmonary arterial smooth mus-cle cells [11] and endothelial cells (data not shown) fromWT and TRPC6-deficient mice, excluding any compen-satory up- and/or down-regulation of TRPC channels inthese lung tissues. Expression of the CaV1.2a1 subunitgene of voltage-gated calcium channels was also notaltered in tracheal smooth muscle cells (data not shown),excluding any compensatory up-regulation of thesechannels.
Increased basal cation entry in TRPC6-deficient trachealsmooth muscle cells
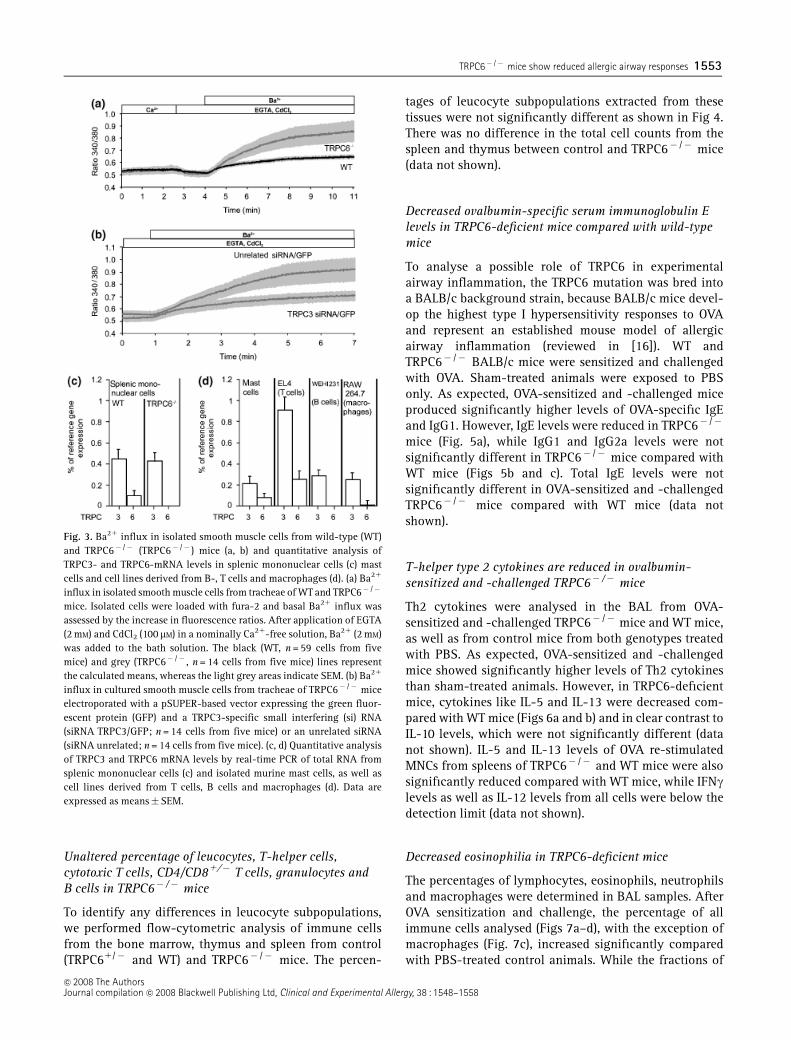
To prove, whether up-regulation of TRPC3 mRNAs intracheal smooth muscle cells of TRPC6-deficient mice iscorrelated with an increased contractility of airwaysmooth muscle, smooth muscle cells from the tracheaewere isolated and Ba21 influx was monitored in thepresence of 100 mM CdCl2 in fura-2-loaded single cells.Basal influx of Ba21 ions was profoundly increased insmooth muscle cells isolated from TRPC6-deficient micecompared with WT cells (Fig. 3a). Cultured smooth musclecells of TRPC6� /� mice electroporated with plasmidscarrying an established siRNA expression cassette underthe control of an H1 promoter directed against the TRPC3mRNA plus GFP [12] or plasmids carrying an expressioncassette for an unrelated siRNA plus GFP were furtheranalysed. Expression of TRPC3-siRNA in these cells re-sulted in decreased basal activity (Fig. 3b) compared withTRPC6� /� cells expressing an unrelated siRNA, as al-ready described for smooth muscle cells of the aorta andcerebral arteries [12]. These data favour a model ofTRPC3/6 heteromeric channels in these tissues whereTRPC6 inhibits a constitutively active TRPC3 channelactivity [22]. In clear contrast to these results, we couldnot detect any changes in the expression of TRPC3 insplenic MNCs from TRPC6� /� mice compared with WTcells by quantitative PCR (Fig. 3c).
Expression of TRPC3- and TRPC6-mRNAs was alsocompared in freshly isolated mast cells as well as in celllines derived from B cells, T cells and macrophages. Inthese cell types, TRPC3 mRNA levels were already sig-nificantly higher than TRPC6 mRNA levels (Fig. 3d),making it improbable that TRPC6 suppresses the basalchannel activity of TRPC3 [22] in these cells. Therefore, itis rather unlikely that similar effects as outlined in Fig. 3bresult from a possible up-regulation of TRPC3- in TRPC6-deficient cells.
Fig. 2. Quantitative analysis of mRNA levels by real-time PCR of totalRNA from mouse lungs (a), tracheal smooth muscle cells (b) andepithelial cells (c). Total RNA was prepared from wild-type (WT; lungn = 7, brain n = 4, black bars) and TRPC6� /� mice (TRPC6� /� ; lungn = 6, brain n = 5, grey bars), reverse-transcribed and products of thefirst-strand synthesis were analysed for the presence of amplificationproducts obtained with the primer pairs described in ‘Material andmethods’. Reaction products were analysed by agarose gel electrophor-esis (a, inset). mRNAs coding for TRPCs (main panels) were quantifiedwith the aid of a light cycler. Values are presented as percentage ofreference mRNA expression. �Significant differences (Po 0.05) betweenWT and TRPC6� /� mice.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
1552 S. Sel et al
Unaltered percentage of leucocytes, T-helper cells,cytotoxic T cells, CD4/CD81/� T cells, granulocytes andB cells in TRPC6� /� mice
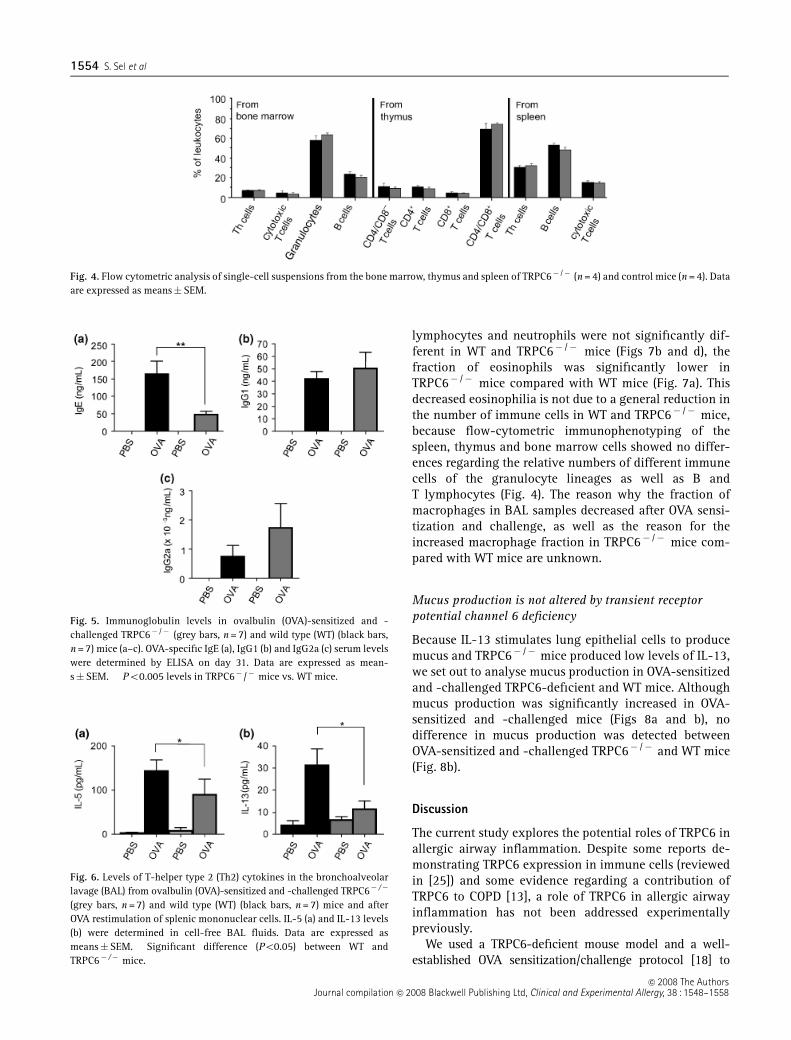
To identify any differences in leucocyte subpopulations,we performed flow-cytometric analysis of immune cellsfrom the bone marrow, thymus and spleen from control(TRPC61/� and WT) and TRPC6� /� mice. The percen-
tages of leucocyte subpopulations extracted from thesetissues were not significantly different as shown in Fig 4.There was no difference in the total cell counts from thespleen and thymus between control and TRPC6� /� mice(data not shown).
Decreased ovalbumin-specific serum immunoglobulin Elevels in TRPC6-deficient mice compared with wild-typemice
To analyse a possible role of TRPC6 in experimentalairway inflammation, the TRPC6 mutation was bred intoa BALB/c background strain, because BALB/c mice devel-op the highest type I hypersensitivity responses to OVAand represent an established mouse model of allergicairway inflammation (reviewed in [16]). WT andTRPC6� /� BALB/c mice were sensitized and challengedwith OVA. Sham-treated animals were exposed to PBSonly. As expected, OVA-sensitized and -challenged miceproduced significantly higher levels of OVA-specific IgEand IgG1. However, IgE levels were reduced in TRPC6� /�
mice (Fig. 5a), while IgG1 and IgG2a levels were notsignificantly different in TRPC6� /� mice compared withWT mice (Figs 5b and c). Total IgE levels were notsignificantly different in OVA-sensitized and -challengedTRPC6� /� mice compared with WT mice (data notshown).
T-helper type 2 cytokines are reduced in ovalbumin-sensitized and -challenged TRPC6� /� mice
Th2 cytokines were analysed in the BAL from OVA-sensitized and -challenged TRPC6� /� mice and WT mice,as well as from control mice from both genotypes treatedwith PBS. As expected, OVA-sensitized and -challengedmice showed significantly higher levels of Th2 cytokinesthan sham-treated animals. However, in TRPC6-deficientmice, cytokines like IL-5 and IL-13 were decreased com-pared with WT mice (Figs 6a and b) and in clear contrast toIL-10 levels, which were not significantly different (datanot shown). IL-5 and IL-13 levels of OVA re-stimulatedMNCs from spleens of TRPC6� /� and WT mice were alsosignificantly reduced compared with WT mice, while IFNglevels as well as IL-12 levels from all cells were below thedetection limit (data not shown).
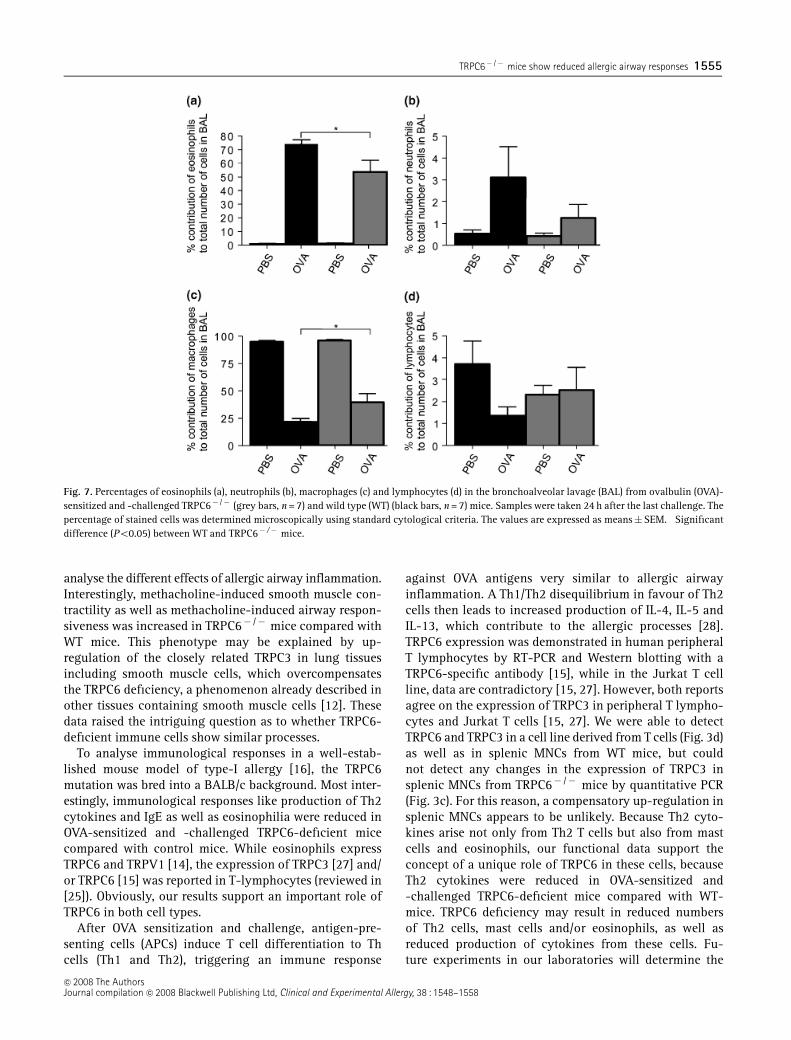
Decreased eosinophilia in TRPC6-deficient mice
The percentages of lymphocytes, eosinophils, neutrophilsand macrophages were determined in BAL samples. AfterOVA sensitization and challenge, the percentage of allimmune cells analysed (Figs 7a–d), with the exception ofmacrophages (Fig. 7c), increased significantly comparedwith PBS-treated control animals. While the fractions of
Fig. 3. Ba21 influx in isolated smooth muscle cells from wild-type (WT)and TRPC6� /� (TRPC6� /� ) mice (a, b) and quantitative analysis ofTRPC3- and TRPC6-mRNA levels in splenic mononuclear cells (c) mastcells and cell lines derived from B-, T cells and macrophages (d). (a) Ba21
influx in isolated smooth muscle cells from tracheae of WTand TRPC6� /�
mice. Isolated cells were loaded with fura-2 and basal Ba21 influx wasassessed by the increase in fluorescence ratios. After application of EGTA(2 mM) and CdCl2 (100 mM) in a nominally Ca21-free solution, Ba21 (2 mM)was added to the bath solution. The black (WT, n = 59 cells from fivemice) and grey (TRPC6� /� , n = 14 cells from five mice) lines representthe calculated means, whereas the light grey areas indicate SEM. (b) Ba21
influx in cultured smooth muscle cells from tracheae of TRPC6� /� miceelectroporated with a pSUPER-based vector expressing the green fluor-escent protein (GFP) and a TRPC3-specific small interfering (si) RNA(siRNA TRPC3/GFP; n = 14 cells from five mice) or an unrelated siRNA(siRNA unrelated; n = 14 cells from five mice). (c, d) Quantitative analysisof TRPC3 and TRPC6 mRNA levels by real-time PCR of total RNA fromsplenic mononuclear cells (c) and isolated murine mast cells, as well ascell lines derived from T cells, B cells and macrophages (d). Data areexpressed as means�SEM.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
TRPC6� /� mice show reduced allergic airway responses 1553
lymphocytes and neutrophils were not significantly dif-ferent in WT and TRPC6� /� mice (Figs 7b and d), thefraction of eosinophils was significantly lower inTRPC6� /� mice compared with WT mice (Fig. 7a). Thisdecreased eosinophilia is not due to a general reduction inthe number of immune cells in WT and TRPC6� /� mice,because flow-cytometric immunophenotyping of thespleen, thymus and bone marrow cells showed no differ-ences regarding the relative numbers of different immunecells of the granulocyte lineages as well as B andT lymphocytes (Fig. 4). The reason why the fraction ofmacrophages in BAL samples decreased after OVA sensi-tization and challenge, as well as the reason for theincreased macrophage fraction in TRPC6� /� mice com-pared with WT mice are unknown.
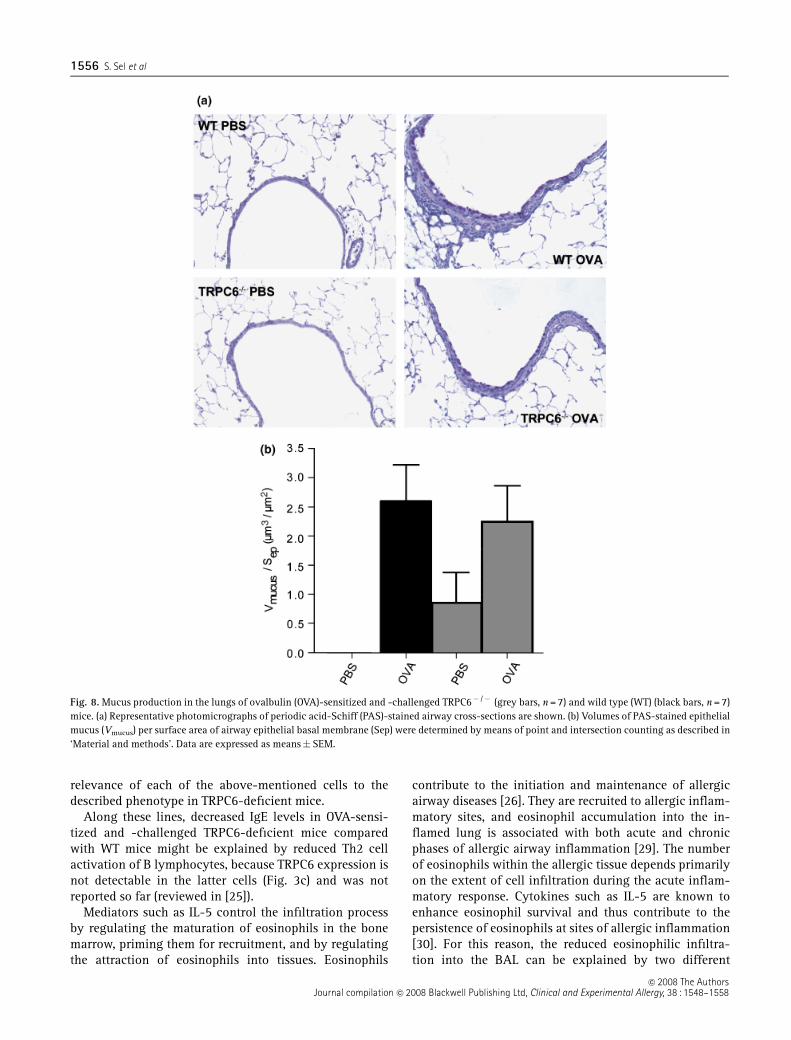
Mucus production is not altered by transient receptorpotential channel 6 deficiency
Because IL-13 stimulates lung epithelial cells to producemucus and TRPC6� /� mice produced low levels of IL-13,we set out to analyse mucus production in OVA-sensitizedand -challenged TRPC6-deficient and WT mice. Althoughmucus production was significantly increased in OVA-sensitized and -challenged mice (Figs 8a and b), nodifference in mucus production was detected betweenOVA-sensitized and -challenged TRPC6� /� and WT mice(Fig. 8b).
Discussion
The current study explores the potential roles of TRPC6 inallergic airway inflammation. Despite some reports de-monstrating TRPC6 expression in immune cells (reviewedin [25]) and some evidence regarding a contribution ofTRPC6 to COPD [13], a role of TRPC6 in allergic airwayinflammation has not been addressed experimentallypreviously.
We used a TRPC6-deficient mouse model and a well-established OVA sensitization/challenge protocol [18] to
Fig. 4. Flow cytometric analysis of single-cell suspensions from the bone marrow, thymus and spleen of TRPC6� /� (n = 4) and control mice (n = 4). Dataare expressed as means� SEM.
Fig. 5. Immunoglobulin levels in ovalbulin (OVA)-sensitized and -challenged TRPC6� /� (grey bars, n = 7) and wild type (WT) (black bars,n = 7) mice (a–c). OVA-specific IgE (a), IgG1 (b) and IgG2a (c) serum levelswere determined by ELISA on day 31. Data are expressed as mean-s�SEM. ��Po0.005 levels in TRPC6� /� mice vs. WT mice.
Fig. 6. Levels of T-helper type 2 (Th2) cytokines in the bronchoalveolarlavage (BAL) from ovalbulin (OVA)-sensitized and -challenged TRPC6� /�
(grey bars, n = 7) and wild type (WT) (black bars, n = 7) mice and afterOVA restimulation of splenic mononuclear cells. IL-5 (a) and IL-13 levels(b) were determined in cell-free BAL fluids. Data are expressed asmeans�SEM. �Significant difference (Po0.05) between WT andTRPC6� /� mice.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
1554 S. Sel et al
analyse the different effects of allergic airway inflammation.Interestingly, methacholine-induced smooth muscle con-tractility as well as methacholine-induced airway respon-siveness was increased in TRPC6� /� mice compared withWT mice. This phenotype may be explained by up-regulation of the closely related TRPC3 in lung tissuesincluding smooth muscle cells, which overcompensatesthe TRPC6 deficiency, a phenomenon already described inother tissues containing smooth muscle cells [12]. Thesedata raised the intriguing question as to whether TRPC6-deficient immune cells show similar processes.
To analyse immunological responses in a well-estab-lished mouse model of type-I allergy [16], the TRPC6mutation was bred into a BALB/c background. Most inter-estingly, immunological responses like production of Th2cytokines and IgE as well as eosinophilia were reduced inOVA-sensitized and -challenged TRPC6-deficient micecompared with control mice. While eosinophils expressTRPC6 and TRPV1 [14], the expression of TRPC3 [27] and/or TRPC6 [15] was reported in T-lymphocytes (reviewed in[25]). Obviously, our results support an important role ofTRPC6 in both cell types.
After OVA sensitization and challenge, antigen-pre-senting cells (APCs) induce T cell differentiation to Thcells (Th1 and Th2), triggering an immune response
against OVA antigens very similar to allergic airwayinflammation. A Th1/Th2 disequilibrium in favour of Th2cells then leads to increased production of IL-4, IL-5 andIL-13, which contribute to the allergic processes [28].TRPC6 expression was demonstrated in human peripheralT lymphocytes by RT-PCR and Western blotting with aTRPC6-specific antibody [15], while in the Jurkat T cellline, data are contradictory [15, 27]. However, both reportsagree on the expression of TRPC3 in peripheral T lympho-cytes and Jurkat T cells [15, 27]. We were able to detectTRPC6 and TRPC3 in a cell line derived from T cells (Fig. 3d)as well as in splenic MNCs from WT mice, but couldnot detect any changes in the expression of TRPC3 insplenic MNCs from TRPC6� /� mice by quantitative PCR(Fig. 3c). For this reason, a compensatory up-regulation insplenic MNCs appears to be unlikely. Because Th2 cyto-kines arise not only from Th2 T cells but also from mastcells and eosinophils, our functional data support theconcept of a unique role of TRPC6 in these cells, becauseTh2 cytokines were reduced in OVA-sensitized and-challenged TRPC6-deficient mice compared with WT-mice. TRPC6 deficiency may result in reduced numbersof Th2 cells, mast cells and/or eosinophils, as well asreduced production of cytokines from these cells. Fu-ture experiments in our laboratories will determine the
Fig. 7. Percentages of eosinophils (a), neutrophils (b), macrophages (c) and lymphocytes (d) in the bronchoalveolar lavage (BAL) from ovalbulin (OVA)-sensitized and -challenged TRPC6� /� (grey bars, n = 7) and wild type (WT) (black bars, n = 7) mice. Samples were taken 24 h after the last challenge. Thepercentage of stained cells was determined microscopically using standard cytological criteria. The values are expressed as means�SEM. �Significantdifference (Po0.05) between WT and TRPC6� /� mice.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
TRPC6� /� mice show reduced allergic airway responses 1555
relevance of each of the above-mentioned cells to thedescribed phenotype in TRPC6-deficient mice.
Along these lines, decreased IgE levels in OVA-sensi-tized and -challenged TRPC6-deficient mice comparedwith WT mice might be explained by reduced Th2 cellactivation of B lymphocytes, because TRPC6 expression isnot detectable in the latter cells (Fig. 3c) and was notreported so far (reviewed in [25]).
Mediators such as IL-5 control the infiltration processby regulating the maturation of eosinophils in the bonemarrow, priming them for recruitment, and by regulatingthe attraction of eosinophils into tissues. Eosinophils
contribute to the initiation and maintenance of allergicairway diseases [26]. They are recruited to allergic inflam-matory sites, and eosinophil accumulation into the in-flamed lung is associated with both acute and chronicphases of allergic airway inflammation [29]. The numberof eosinophils within the allergic tissue depends primarilyon the extent of cell infiltration during the acute inflam-matory response. Cytokines such as IL-5 are known toenhance eosinophil survival and thus contribute to thepersistence of eosinophils at sites of allergic inflammation[30]. For this reason, the reduced eosinophilic infiltra-tion into the BAL can be explained by two different
Fig. 8. Mucus production in the lungs of ovalbulin (OVA)-sensitized and -challenged TRPC6� /� (grey bars, n = 7) and wild type (WT) (black bars, n = 7)mice. (a) Representative photomicrographs of periodic acid-Schiff (PAS)-stained airway cross-sections are shown. (b) Volumes of PAS-stained epithelialmucus (Vmucus) per surface area of airway epithelial basal membrane (Sep) were determined by means of point and intersection counting as described in‘Material and methods’. Data are expressed as means� SEM.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
1556 S. Sel et al

mechanisms that are not mutually exclusive. First, lowerIL-5 levels from Th2 cells may lead to decreased eosino-phil maturation in the bone marrow and consequently intheir release into the blood (reviewed in [31, 32]) and,second, TRPC6 deficiency may decrease eosinophil adhe-sion and subsequent infiltration into the lung tissue. Veryrecently, evidence was provided that lysophosphatidyl-choline (LPC) regulates b2-integrin-mediated adhesion ofeosinophils by a non-store-operated Ca21 channel [14].Moreover, only TRPC6 and TRPV1 of the TRPC, M and Vfamilies were found to be expressed in eosinophils.Because capsaicin, a TRPV1 agonist, was not able toincrease the intracellular calcium concentration ([Ca21]i)in eosinophils, the authors concluded that LPC causesa non-store-operated Ca21 influx via TRPC6, which sub-sequently activates CD11b/CD18 of the b2-integrin sub-family to cause eosinophil adhesion [14]. Only specificdown-regulation of TRPC6 in eosinophils may answer thequestion, as to which of the mechanism(s) is/are requiredfor the TRPC6-dependent eosinophilia in inflammatorydiseases like allergic airway inflammation.
The pathogenesis of allergic airway inflammation iscomposed of two phases: (i) AHR and (ii) subsequentairway remodelling. To investigate the latter, we alsoanalysed goblet hyperplasia and resulting mucus hyper-secretion in airway epithelial cells. As IL-13 levels areresponsible for mucus production [33] and TRPC6 isexpressed in airway epithelial cells [9], we expected adecreased mucus production in TRPC6� /� mice. Much toour surprise, we were not able to detect altered mucusproduction and goblet cell hyperplasia in OVA-sensitizedand -challenged TRPC6-deficient mice as compared withWT mice. The TRPC6-dependent IL-13 level may not playa role in mucus production in airway epithelial cellsduring acute allergic airway inflammation, because tem-porary dissociation of IL-13-mediated mucus hypersecre-tion from AHR was described recently [34]. However, asmucus production and goblet hyperplasia are more pro-nounced in chronic airway inflammation, further studiesneed to be performed to investigate whether there isTRPC6-dependent mucus production in a mouse modelof chronic airway inflammation [35].
In conclusion, our data point to a unique and non-redundant role of TRPC6 in Th2 cells, mast cells and/oreosinophils that results in reduced Th2 cytokine produc-tion and decreased airway eosinophilia in OVA-sensitizedand -challenged TRPC6-deficient mice.
Acknowledgements
We wish to thank Winfried Lorenz, Tanja Rausch, AnjaSpieß-Naumann and Susanne Ziegler for excellent tech-nical assistance. The expertise of Drs Udo Herz and KhaiVi in head-out body-plethysmography and the analysis ofthe contractility of tracheal rings is greatly acknowledged.
We would like to thank Dr Reinhard Mailhammer (GSFNational Research Center for Environment and Health,Munich, Germany) for his gift of mast cell cDNA.
References
1 Nilius B, Owsianik G, Voets T, Peters JA. Transient receptorpotential cation channels in disease. Physiol Rev 2007;87:165–217.
2 Venkatachalam K, Montell C. TRP Channels. Annu Rev Biochem2007; 76:387–417.
3 Dietrich A, Kalwa H, Rost BR, Gudermann T. The diacylgylcerol-sensitive TRPC3/6/7 subfamily of cation channels: functionalcharacterization and physiological relevance. Pflugers Arch2005; 451:72–80.
4 Dietrich A, Mederos y Schnitzler M, Kalwa H, Storch U, Guder-mann T. Functional characterization and physiological relevanceof the TRPC3/6/7 subfamily of cation channels. Naunyn Schmie-debergs Arch Pharmacol 2005; 371:257–65.
5 Hofmann T, Obukhov AG, Schaefer M, Harteneck C, GudermannT, Schultz G. Direct activation of human TRPC6 and TRPC3channels by diacylglycerol. Nature 1999; 397:259–63.
6 Hofmann T, Schaefer M, Schultz G, Gudermann T. Subunitcomposition of mammalian transient receptor potential channelsin living cells. Proc Natl Acad Sci USA 2002; 99:7461–6.
7 Dietrich A, Chubanov V, Kalwa H, Rost BR, Gudermann T. Cationchannels of the transient receptor potential superfamily: theirrole in physiological and pathophysiological processes ofsmooth muscle cells. Pharmacol Ther 2006; 112:744–60.
8 Boulay G, Zhu X, Peyton M et al. Cloning and expression of anovel mammalian homolog of Drosophila transient receptorpotential (Trp) involved in calcium entry secondary to activationof receptors coupled by the Gq class of G protein. J Biol Chem1997; 272:29672–80.
9 Corteling RL, Li S, Giddings J, Westwick J, Poll C, Hall IP.Expression of transient receptor potential C6 and related tran-sient receptor potential family members in human airwaysmooth muscle and lung tissue. Am J Respir Cell Mol Biol 2004;30:145–54.
10 Wang J, Shimoda LA, Sylvester JT. Capacitative calcium entryand TRPC channel proteins are expressed in rat distal pulmonaryarterial smooth muscle. Am J Physiol Lung Cell Mol Physiol2004; 286:L848–58.
11 Weissmann N, Dietrich A, Fuchs B et al. Classical transientreceptor potential channel 6 (TRPC6) is essential for hypoxicpulmonary vasoconstriction and alveolar gas exchange. ProcNatl Acad Sci USA 2006; 103:19093–8.
12 Dietrich A, Mederos YSM, Gollasch M et al. Increased vascularsmooth muscle contractility in TRPC6� /� mice. Mol Cell Biol2005; 25:6980–9.
13 Li S, Gosling M, Poll C. Determining the functional role of TRPCchannels in primary cells. Pflugers Arch 2005; 451:43–52.
14 Zhu X, Learoyd J, Butt S et al. Regulation of eosinophil adhesionby lysophosphatidylcholine via a non-store-operated Ca21
channel. Am J Respir Cell Mol Biol 2007; 36:585–93.15 Gamberucci A, Giurisato E, Pizzo P et al. Diacylglycerol activates
the influx of extracellular cations in T-lymphocytes indepen-dently of intracellular calcium-store depletion and possibly
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
TRPC6� /� mice show reduced allergic airway responses 1557

involving endogenous TRP6 gene products. Biochem J 2002;364:245–54.
16 Herz U, Renz H, Wiedermann U. Animal models of type I allergyusing recombinant allergens. Methods 2004; 32:271–80.
17 Neuhaus-Steinmetz U, Glaab T, Daser A et al. Sequential devel-opment of airway hyperresponsiveness and acute airway ob-struction in a mouse model of allergic inflammation. Int ArchAllergy Immunol 2000; 121:57–67.
18 Herz U, Braun A, Ruckert R, Renz H. Various immunologicalphenotypes are associated with increased airway responsiveness.Clin Exp Allergy 1998; 28:625–34.
19 Devore-Carter D, Morway PF, Weiss EB. Isolation and character-ization of guinea-pig tracheal smooth muscle cells that retaindifferentiated function in long-term subculture. Cell Tissue Res1988; 251:325–31.
20 You Y, Richer EJ, Huang T, Brody SL. Growth and differentiationof mouse tracheal epithelial cells: selection of a proliferativepopulation. Am J Physiol Lung Cell Mol Physiol 2002;283:L1315–21.
21 Hultner L, Szots H, Van Snick J, Moeller J, Welle M, Dormer P.Interleukin 3 mediates interleukin 6 production in murine inter-leukin 3-dependent hemopoietic cells. Growth Factors 1989;2:43–51.
22 Dietrich A, Mederos y Schnitzler M, Emmel J, Kalwa H, HofmannT, Gudermann T. N-linked protein glycosylation is a majordeterminant for basal TRPC3 and TRPC6 channel activity. J BiolChem 2003; 278:47842–52.
23 Blumer N, Sel S, Virna S et al. Perinatal maternal applicationof Lactobacillus rhamnosus GG suppresses allergic airway in-flammation in mouse offspring. Clin Exp Allergy 2007; 37:348–57.
24 Weibel ER, Hsia CC, Ochs M. How much is there really? Whystereology is essential in lung morphometry. J Appl Physiol2007; 102:459–67.
25 Schwarz EC, Wolfs MJ, Tonner S et al. TRP channels in lympho-cytes. Handb Exp Pharmacol 2007; 179:445–56.
26 Rothenberg ME, Hogan SP. The eosinophil. Annu Rev Immunol2006; 24:147–74.
27 Philipp S, Strauss B, Hirnet D et al. TRPC3 mediates T-cellreceptor-dependent calcium entry in human T-lymphocytes. JBiol Chem 2003; 278:26629–38.
28 Ngoc PL, Gold DR, Tzianabos AO, Weiss ST, Celedon JC. Cyto-kines, allergy, and asthma. Curr Opin Allergy Clin Immunol2005; 5:161–6.
29 Kay AB. Asthma and inflammation. J Allergy Clin Immunol1991; 87:893–910.
30 Yamaguchi Y, Suda T, Ohta S, Tominaga K, Miura Y, Kasahara T.Analysis of the survival of mature human eosinophils: inter-leukin-5 prevents apoptosis in mature human eosinophils. Blood1991; 78:2542–7.
31 Rosenberg HF, Phipps S, Foster PS. Eosinophil trafficking inallergy and asthma. J Allergy Clin Immunol 2007; 119:1303–10.
32 Trivedi SG, Lloyd CM. Eosinophils in the pathogenesis of allergicairways disease. Cell Mol Life Sci 2007; 64:1269–89.
33 Wills-Karp M. Interleukin-13 in asthma pathogenesis. ImmunolRev 2004; 202:175–90.
34 Yang M, Rangasamy D, Matthaei KI et al. Inhibition of arginase Iactivity by RNA interference attenuates IL-13-induced airwayshyperresponsiveness. J Immunol 2006; 177:5595–603.
35 Wegmann M, Fehrenbach H, Fehrenbach A et al. Involvement ofdistal airways in a chronic model of experimental asthma. ClinExp Allergy 2005; 35:1263–71.
�c 2008 The AuthorsJournal compilation �c 2008 Blackwell Publishing Ltd, Clinical and Experimental Allergy, 38 : 1548–1558
1558 S. Sel et al
Related Documents










![Research Paper Deguelin Attenuates Allergic Airway ... · Research Paper Deguelin Attenuates Allergic Airway Inflammation via ... pathophysiology of asthma [4]. ... schematic diagram](https://static.cupdf.com/doc/110x72/5b396a827f8b9ab9068e82d4/research-paper-deguelin-attenuates-allergic-airway-research-paper-deguelin.jpg)
