www.thaiagj.org Thai Journal of Agricultural Science 2011, 44(3): 155-163 Longevity, Fecundity and Development Time of Eucalyptus Gall Wasp, Leptocybe invasa Fisher & La Salle (Hymenoptera: Eulophidae) in Kanchanaburi Province, Thailand B. Sangtongpraow 1, *, K. Charernsom 1 and S. Siripatanadilok 2 ¹Department of Entomology, Faculty of Agriculture, Kasetsart University, Bangkok 10900, Thailand ²Department of Forest Biology, Faculty of Forestry, Kasetsart University, Bangkok 10900, Thailand *Corresponding author. Email: [email protected] Abstract Biological information of L. invasa in Thailand is extremely limited. This study examined the longevity, fecundity and development time of this gall wasp. The galled leaves and gall wasps were sampled from Eucalyptus camaldulensis plantations in Kanchanaburi province and were conducted in the laboratory and ventilated greenhouse. The results showed that feeding L. invasa with honey solution could prolong the mean longevity of females (7.67±0.93 days) and that of males (5.67±0.47 days). The average potential fecundity of females was 158.70±4.62 eggs per a female. As size increased the potential fecundity of L. invasa tended to increased. The realized fecundity indicated that L. invasa is a pro-ovigenic species. The mean progeny per female was highest on the first day (30.47±7.41 progeny female -1 ) and lowest on the sixth day. The reproductive mode of L. invasa is deuterotoky. The development time of L. invasa from egg to adult was 45.96±0.52 days, ranging from 35 to 73 days. Keywords: eucalyptus gall wasp, Leptocybe invasa, longevity, fecundity, development time Introduction Eucalyptus camaldulensis trees are widely planted on state, private and farm land of Thailand. This fast growing tree species provides wood materials, other products and indirect benefits for domestic, commercial uses. Damages by Leptocybe invasa Fisher & La Salle (Hymenoptera: Eulophidae) in Eucalyptus plantations were first discovered in Israel in 2000, subsequently spreaded to Africa in 2002 and to India and Southeast Asia in 2004 (IFGTB, n.d.). L. invasa was first observed in Thailand during 2004-2005 and then spreaded to other parts of the country. At present, the damages are found in Lao PDR and Vietnam (Thu et al., 2009). L. invasa is the devastating pest of eucalypt and damages newly developed leaves and young twigs of E. camaldulensis. The wasp forms numerous galls on leaf petioles, midribs and young twigs and results in the deformity of leaf shape, abnormality of plant growth, reduction of wood yield and decreasing of owners’ incomes. Most E. camaldulensis plantations in Thailand use only some clones and monoculture system. Thai researchers, particularly in the private sectors, carry out E. camaldulensis improvement programs to obtain new clones which produce high wood yields and to tolerate L. invasa’s attack. However there are some difficulties to be solved. Some clones showed resistance in the first few years, they could tolerate L. invasa. Subsequently, they became susceptible because L. invasa adapted themselves and were able to produce new offsprings to damage those clones. Some countries in Mediterranean region use many measures

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.thaiagj.org Thai Journal of Agricultural Science 2011, 44(3): 155-163
Longevity, Fecundity and Development Time of Eucalyptus Gall Wasp, Leptocybe invasa Fisher & La Salle (Hymenoptera: Eulophidae) in
Kanchanaburi Province, Thailand
B. Sangtongpraow1,*, K. Charernsom1 and S. Siripatanadilok2
¹Department of Entomology, Faculty of Agriculture, Kasetsart University,
Bangkok 10900, Thailand ²Department of Forest Biology, Faculty of Forestry, Kasetsart University,
Bangkok 10900, Thailand
*Corresponding author. Email: [email protected]
Abstract Biological information of L. invasa in Thailand is extremely limited. This study examined the
longevity, fecundity and development time of this gall wasp. The galled leaves and gall wasps were sampled from Eucalyptus camaldulensis plantations in Kanchanaburi province and were conducted in the laboratory and ventilated greenhouse. The results showed that feeding L. invasa with honey solution could prolong the mean longevity of females (7.67±0.93 days) and that of males (5.67±0.47 days). The average potential fecundity of females was 158.70±4.62 eggs per a female. As size increased the potential fecundity of L. invasa tended to increased. The realized fecundity indicated that L. invasa is a pro-ovigenic species. The mean progeny per female was highest on the first day (30.47±7.41 progeny female-1) and lowest on the sixth day. The reproductive mode of L. invasa is deuterotoky. The development time of L. invasa from egg to adult was 45.96±0.52 days, ranging from 35 to 73 days. Keywords: eucalyptus gall wasp, Leptocybe invasa, longevity, fecundity, development time
Introduction Eucalyptus camaldulensis trees are widely
planted on state, private and farm land of Thailand. This fast growing tree species provides wood materials, other products and indirect benefits for domestic, commercial uses. Damages by Leptocybe invasa Fisher & La Salle (Hymenoptera: Eulophidae) in Eucalyptus plantations were first discovered in Israel in 2000, subsequently spreaded to Africa in 2002 and to India and Southeast Asia in 2004 (IFGTB, n.d.). L. invasa was first observed in Thailand during 2004-2005 and then spreaded to other parts of the country. At present, the damages are found in Lao PDR and Vietnam (Thu et al., 2009).
L. invasa is the devastating pest of eucalypt and damages newly developed leaves and young twigs
of E. camaldulensis. The wasp forms numerous galls on leaf petioles, midribs and young twigs and results in the deformity of leaf shape, abnormality of plant growth, reduction of wood yield and decreasing of owners’ incomes. Most E. camaldulensis plantations in Thailand use only some clones and monoculture system.
Thai researchers, particularly in the private sectors, carry out E. camaldulensis improvement programs to obtain new clones which produce high wood yields and to tolerate L. invasa’s attack. However there are some difficulties to be solved. Some clones showed resistance in the first few years, they could tolerate L. invasa. Subsequently, they became susceptible because L. invasa adapted themselves and were able to produce new offsprings to damage those clones. Some countries in Mediterranean region use many measures

156 B. Sangtongpraow et al. Thai Journal of Agricultural Science
including biological control to manage this gall wasp (Kim et al., 2008; Protasov et al., 2008). Successful biological control of L. invasa requires biological knowledge of this gall wasp however this knowledge in Thailand is extremely limited.
This research aimed to determine the longevity, fecundity and development time of L. invasa. These biological parameters will be beneficial in appropriate parasitoid selection. The selected parasitoids should have better biological parameters than those of L. invasa. The successful biological control of L. invasa will be a supplement to E. camaldulensis improvement programs in order to provide integrated control of L. invasa.
Materials and Methods Longevity Assessment
Both female and male L. invasa were found in the study areas, as shown in Figure 1. The female differed from male in antenna, forewing, gaster shape and size. The female size (1.10-1.60 mm in length from head to abdomen tip) was larger than
male one (0.80-1.40 mm in length). In Israel, only female L. invasa was found and were 1.10-1.40 mm in length (Mendel et al., 2004) but discovered both female and male in Turkey (Doğanlar, 2005) and India (Gupta and Poorani, 2009). So this research on longevity of L. invasa covered both female and male.
Mean Longevity
The longevity of L. invasa refers to the period from adult, emerging from leaf gall of E. camaldulensis, to its expiration. The results in laboratory showed that the mean longevity of female and that of male L. invasa fed with honey solution, could prolong the largest mean longevity of female (7.67±0.93 days) and that of male (5.67±0.41 days).
When females and males were fed with no-diet, water, water+flowers, honey solution+flowers and flowers of E. camaldulensis, the means female longevity were 1.33, 1.56, 1.89, 6.11 and 2.00 days respectively and male longevity was 1.00, 1.11, 1.56, 5.22 and 1.22 days, respectively (Table 1).
A B
Figure 1 Leptocybe invasa found in Eucalyptus plantations in Kanchanaburi province : (A) female; and (B) male.
Table 1 Mean longevities (days) of female and male Leptocybe invasa fed with different diets.
Diet Female Male
No-diet 1.33±0.17 a 1.00±0.00 a Water 1.56±0.18 a 1.11±0.11 a Water+flowers 1.89±0.26 a 1.56±0.18 a Honey solution 7.67±0.93 b 5.67±0.41 b Honey solution+flowers 6.11±0.79 b 5.22±0.47 b Flowers 2.00±0.40 a 1.22±0.15 a
Female: F=27.434; df =5; P-value=0.000, Male: F=64.463; df =5; P-value=0.000. Means followed by the same letter within each column were not significantly different (Tukey’s HSD, P=0.05).

Vol. 44, No. 3, 2011 Fisher & La salle in Kanchanaburi province, Thailand 157
The statistical analysis of the means of female longevity and the means of male longevity indicated that P–values=0.000 were lower than P=0.05, thus different diets had significant effect on the means female and male longevity of L. invasa.
The reasons to use six diets in this study were that honey/or nectar of flowers was a common diet of most insects. Moreover those diets were consistent with natural diets found in E. camaldulensis plantations.
Mendel et al. (2004) studied only the longevity of female L. invasa in Israel and reported that honey solution prolonged the mean female longevity of (6.50 days). The other five diets; no-diet, water, honey solution plus young leaves, flowers, and young leaves of E. camaldulensis could extend female longevity but to a lesser extent. The difference in the means of female longevity in Thailand and that in Israel was possibly explained in terms of body sizes because the body size had a positive correlation with longevity (Jervis, 2005). The size of female L. invasa in Thailand (1.10-1.60 mm in length) was larger than that in Israel (1.10-1.40 mm in length). The larger size of L. invasa might consume more diet and might be stronger and led to the longer longevity.
The findings from this study suggested that the longevity of L. invasa could be extended by feeding with honey solution. Thus L. invasa can be
used as host for mass rearing of the parasitoids in the laboratory. Finally, the parasitoids from mass production will be introduced to E. camaldulensis plantations to use as biological control agent of L. invasa.
Survival Patterns
This term denotes the percent of survivors at each day within the range of longevity when L. invasa is fed with a diet. It was found that female and male L. invasa could be divided into two groups of survivors. Survivors with the longest time were among the females that were fed with honey solution and honey solution plus flowers, ranging from 1 to 13 days and 1 to 11 days, respectively. The estimated 50% female survival periods were 5 and 4 days (Figure 2). Survivors with the shortest time were among the female that were supplied with no-diet, water, water plus flowers, and flowers, ranging from 1 to 4 days. The estimated 50% female survival period was ~2 days.
The longest survival times were among males that were fed with honey solution and honey solution plus flowers, ranging from 1 to 8 days. The estimated 50% male survival periods were 4 and 3 days, respectively. The shortest survival periods were among the males that were supplied with no-diet, water, water plus flowers, and flowers, ranging from 1 to 3 days only. The estimated 50% male survival period was ~1 day (Figure 3).
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10 11 12 13
Days
% S
urvi
vors
No dietWaterWater+flowersHoney solutionHoney+flowersFlowers
Figure 2 Survival patterns of female Leptocybe invasa fed with different diets.

158 B. Sangtongpraow et al. Thai Journal of Agricultural Science
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8
Days
% S
urvi
vors
No dietWaterWater+flowersHoney solutionHoney+flowersFlowers
Figure 3 Survival patterns of male Leptocybe invasa fed with different diets.
The findings from the survival patterns suggested that honey solution and honey solution plus flowers of E. camaldulensis were the best diets to prolong survival periods of female and male L. invasa. However, feeding with honey solution alone was more practical and more convenient than feeding with the later diet.
Mendel et al. (2004) reported from Israel that the longest survivals were among the females that were fed with honey solution, ranging from 1 to 8 days. The different findings between the survival patterns of the female L. invasa found in this research and that in Israel might be concerned with the difference in body sizes and/or environmental conditions.
Fecundity of L. invasa Potential Fecundity
The potential fecundity denotes the number of mature eggs in an ovary (or egg load) that can be potentially laid by an adult female L. invasa. It can be counted directly in the ovary by dissecting all of the eggs in ovaries of adult female L. invasa emerging from leaf galls.
The potential fecundity in a pair of ovaries of a female L. invasa was shown in Figure 4. In the laboratory, our research found that the average potential fecundity of the females, of all sizes and ages, was 158.70±4.62 eggs per female, ranging from 39 to 298 eggs per female.
To use hind tibia length to indicate female size (Jervis, 2005), the average of all female sizes of L. invasa was 0.28 mm, ranging from 0.20-0.35 mm. We were able to divide the female sizes into 3 classes as follows: small size (0.20-0.24 mm), medium size (0.25-0.29 mm), and large size (0.30 mm or more). The percentage of small, medium, large females was 17.55:48.58:33.86 (Table 2). The results suggested that female sizes in the research area were mostly medium and large sizes. Increasing of female sizes tended to increase potential fecundity of the female L. invasa (Table 2).
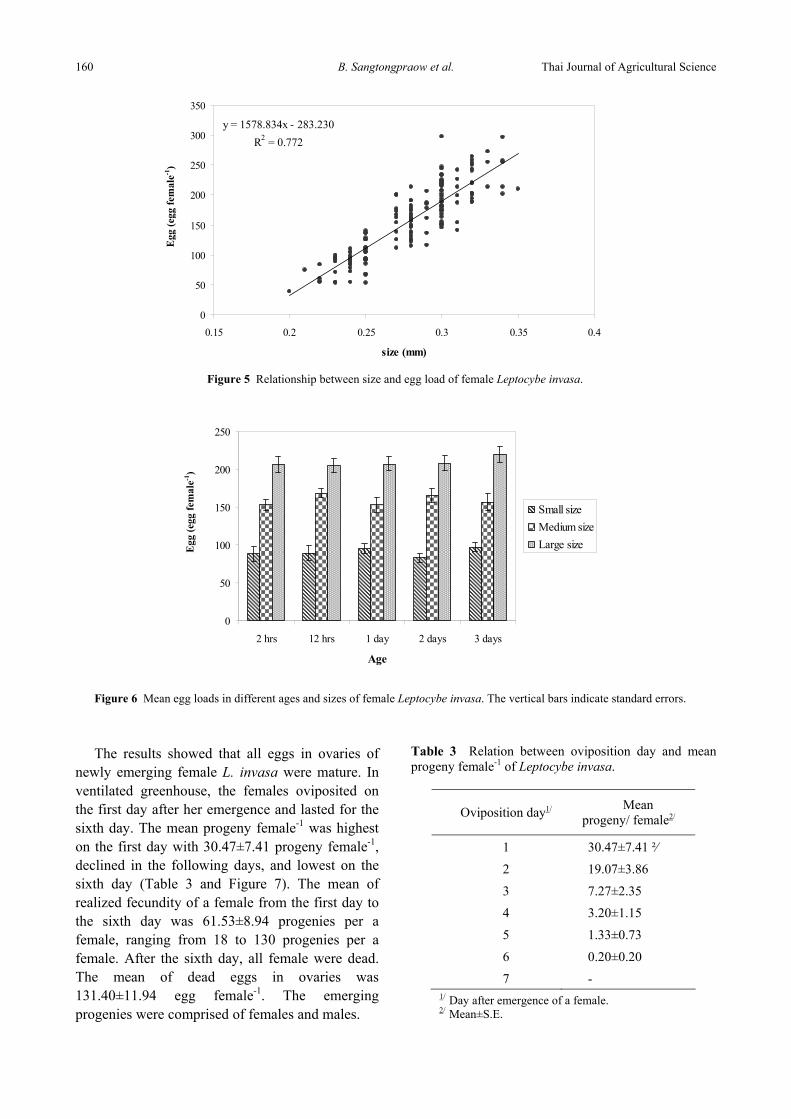
The effects of female sizes of L. invasa (1 day-old age) on potential fecundity were determined by ANOVA regression analysis. The results showed that sizes had significant effects on potential fecundity at P=0.05 (F=532.257; P-value=0.000). There was significant positive relationship between sizes and potential fecundities of female (y =1578.834x – 283.230; R²=0.772; n=159). The larger females tended to produce more eggs than the smaller ones (Figure 5). The R²=0.772 indicated that female sizes influenced on egg loads at about 77.20%.
In this study we used hind tibial length as a substitute for female size of L. invasa. Usually width or length of some body parts such as head, thorax, or hind tibia are used to represent the body size (Jervis, 2005). Unfortunately, the insects in Family Eulophidae change their width and length

Vol. 44, No. 3, 2011 Fisher & La salle in Kanchanaburi province, Thailand 159
Table 2 Female size average, percentage and potential fecundity in each size class of Leptocybe invasa.
Size class (mm)
Female size average (mm) Percentage Potential fecundity1/
(egg female-1) Small (0.20–0.24) 0.239±0.002 17.55 90.64±3.32
(39–140) Medium (0.25–0.29) 0.279±0.001 48.58 160.58±3.82
(112–213) Large (0.30 or more) 0.311±0.002 33.86 209.03±4.41
(141–298) 1/ Mean±S.E., with range in the bracket.
Figure 4 Mature eggs in a pair of ovaries of Leptocybe invasa.
of heads from normal to crumping and distorted heads after death. This made it difficult to take head measurement. The length of hind tibia was unchanged and it related to body size. Thus, hind tibia length was reasonable to be used as representative of body size.
The effects of female ages and sizes on egg loads were shown in Figure 6. In each age group, mean egg loads increased with increasing sizes; but in each size group, mean egg loads in different ages were slightly different. By two-way analysis of variance, ages had no effect on egg loads at P=0.05 (F=0.281, df =4, P-value=0.890). Sizes had significant effects on egg loads at P=0.05 (F=211.973, df =2, P-value=0.000). Age x size had no effect on egg loads at P=0.05 (F=0.416, df =8, P-value=0.910). This finding was in consistent with Bernado et al. (2005) who studied on the potential fecundity of Thripobius semiluteus Bouček (Hymenoptera: Eulophidae) and reported that ages had no significant effect on mean egg loads.
Realized Fecundity Generally it refers to the total number of eggs
actually laid over the life-time of an adult female. Practically, the realized fecundity is determined by counting the number of eggs laid in host per day until a female dies and then calculate the total number of eggs laid in host over the life time of a female.
Takagi (1985) and Hardy et al. (1992) determined the realized fecundity of parasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae) without counting the number of eggs. They counted the number of adult offsprings produced and combined with dead eggs, so deriving the estimated number of eggs originally deposited. The female L. invasa laid her eggs in eucalyptus leaf tissues, so the eggs laid in each day and the dead eggs in the tissues were difficult to detect. Thus this research determined the realized fecundity of L. invasa by counting the total number of emerging progenies over the life-time of a female and expressed in terms of progenies per a female.
160 B. Sangtongpraow et al. Thai Journal of Agricultural Science
y = 1578.834x - 283.230R2 = 0.772
0
50
100
150
200
250
300
350
0.15 0.2 0.25 0.3 0.35 0.4
size (mm)
Figure 5 Relationship between size and egg load of female Leptocybe invasa.
0
50
100
150
200
250
2 hrs 12 hrs 1 day 2 days 3 days
Age
Small sizeMedium sizeLarge size
Figure 6 Mean egg loads in different ages and sizes of female Leptocybe invasa. The vertical bars indicate standard errors. The results showed that all eggs in ovaries of
newly emerging female L. invasa were mature. In ventilated greenhouse, the females oviposited on the first day after her emergence and lasted for the sixth day. The mean progeny female-1 was highest on the first day with 30.47±7.41 progeny female-1, declined in the following days, and lowest on the sixth day (Table 3 and Figure 7). The mean of realized fecundity of a female from the first day to the sixth day was 61.53±8.94 progenies per a female, ranging from 18 to 130 progenies per a female. After the sixth day, all female were dead. The mean of dead eggs in ovaries was 131.40±11.94 egg female-1. The emerging progenies were comprised of females and males.
Table 3 Relation between oviposition day and mean progeny female-1 of Leptocybe invasa.
Oviposition day1/ Mean progeny/ female2/
1 30.47±7.41 ²⁄
2 19.07±3.86
3 7.27±2.35
4 3.20±1.15
5 1.33±0.73
6 0.20±0.20
7 - 1/ Day after emergence of a female. 2/ Mean±S.E.
Egg
(egg
fem
ale-1
)
Egg
(egg
fem
ale-1
)

Vol. 44, No. 3, 2011 Fisher & La salle in Kanchanaburi province, Thailand 161
0
5
10
15
20
25
30
35
40
1 2 3 4 5 6 7
Oviposition day Figure 7 Mean progeny female-1 as related to oviposition day of Leptocybe invasa. The vertical bars indicate standard errors. The realized fecundity of L. invasa indicated
that this gall wasp species is a pro-ovigenic species. The reasons are that a) all eggs in ovaries of newly emerging female were mature and b) the females oviposite on the first day after her emergence and the mean progeny female-1 is highest on the first day or on early adult life.
The mean number of female progenies:male progenies was 40.80:20.73. Thus the sex ratio of female: male progeny was 1.97:1. The number of female progenies was higher than the male one. The non mating female had offspring of both sexes. From the observation on mating behavior, the male offsprings did not mate with female offsprings because the male offspring were non-functional. So the reproductive mode of L. invasa is deuterotoky. Luck et al. (1993) and Jervis (2005) explained that deuterotoky means the female produces more female offsprings than male offsprings, both sexes of offsprings developed from unfertilized eggs and the male offsprings were non-functional.
Development of L. invasa
Development of female and male L. invasa in this study covered from egg to young larva, mature larva to prepupa, pupa, and to adult stage (adult emerging from E. camaldulensis tissues).
Oviposition behavior The research found that the adult female L.
invasa oviposites in newly developed leaves and in young twigs of E. camaldulensis since the first day of her emergence (Figure 8). The females insert eggs in vascular bundles of leaf petioles and midribs, and young twigs. The development of eggs inside vascular bundles stimulates the development of galls, forming typical bump-shape on the petioles, midribs, and young twigs. The larvae of L. invasa which develop inside the galls take away mineral salts and nutrients from plant organs. The development of eggs to adult L. invasa occurs in plant galls. Egg overloading and heavy galling gave rise to the severe reduction of leaf and shoot growth (Figure 9) and prevented the development and growth of infested trees.
Developmental time from egg to adult
The results showed that the mean development time of L. invasa from egg to adult in ventilated greenhouse was 45.96±0.52 days, ranging from 35 to 73 days. The egg stage, young larval stage, mature larval to prepupal stage, and pupal stage took 10.94±0.25, 18.07±1.16, 30.13±1.05 and 37.70±1.07 days, respectively. The development time was minimum in egg stage and maximum in larval stage and in pupal stage (Figure 10).
Egg
(egg
fem
ale-1
)

162 B. Sangtongpraow et al. Thai Journal of Agricultural Science
Figure 8 Oviposition behavior of a female Leptocybe invasa on the petiole of a newly developed leaf of Eucalyptus camaldulensis.
Figure 9 Damage to leaves of Eucalyptus camaldulensis by Leptocybe invasa.
Figure 10 Development time at each stage of Leptocybe invasa in leaf gall of Eucalyptus camaldulensis.
Mendel et al. (2004) reported that the mean development time of L. invasa, at room temperature in Israel, was 132.60 days. The differences of the mean development time of L. invasa in this study and in Israel probably resulted from the differences in temperature and the environmental conditions of the researches.
The study on longevity, fecundity and development time of L. invasa was undertaken from March 2009 to January 2011.
Conclusions
Feeding L. invasa with honey solution could
prolong the largest mean longevity of females (7.67±0.93 days) and that of males (5.67±0.47 days). The estimated 50% female and male survival periods were 5 days and 4 days, respectively. The average potential fecundity of females, of all sizes and ages, was 158.70±4.62 eggs per a female, ranging from 39 to 298 egg female-1. Increasing of sizes tended to increase potential fecundities of female L. invasa. The realized fecundity indicated that L. invasa is a pro-ovigenic species. The mean progeny female-1 was highest on the first day (30.47±7.41 progeny female-1) and lowest on the sixth day. The reproductive mode of L. invasa is deuterotoky. The developmental time of L. invasa from egg to adult was 45.96±0.52 days, ranging from 35 to 73 days. The developmental time was the longest in larval stage and in pupal stage.
Acknowledgments
The authors wish to express the gratitude to
SCG Paper Public Company Limited in Ban Pong district, Ratchaburi province, and Siam Forestry Company in Kanchanaburi province, for generous supports throughout the research. We are most grateful to Miss Titiporn Saimanee, Mr. Wichan Charoenkitsupat, Dr. Thananan Akhadejdamrong, Mr. Yuttana Singchada, and their colleagues for their kind helps in field works, moral support, and beautiful friendship throughout this research work.
References
Bernardo, U., G. Viggiani and R. Sasso. 2005.
Biological parameters of Thripobius semiluteus Bouček (Hym., Eulophidae), a larval endoparasitoid

Vol. 44, No. 3, 2011 Fisher & La salle in Kanchanaburi province, Thailand 163
of Heliothrips haemorrhoidalis (Bouché) (Thysan., Thripidae). J. Appl. Entomol. 129: 250-257.
Doğanlar, O. 2005. Occurrence of Leptocybe invasa Fisher and La Salle, 2004 (Hymenoptera: Chalcidoidea) on Eucalyptus camaldulensis in Turkey, with a description of the male sex. Zool. Middle East 35: 112-113.
Grabenweger, G., H. Hopp, S. Schmolling, T. Koch, H. Balder and B. Jäckel. 2009. Laboratory rearing and biological parameters of the eulophid Pnigalio agraules, a parasitoid of Cameraria ohridella. J. Appl. Entomol. 133: 1-9.
Hardy, I.C.W., N.T. Griffiths and H.C.J. Godfray. 1992. Clutch size in a parasitoid wasp: a manipulation experiment. J. Anim. Ecol. 61: 121-129.
Institute of Forest Genetics and Tree Breeding (IFGTB). n.d. Press Note on Gall Insect Problem in Eucalyptus – Present Status and Future Course of Action. Available Source: http://ifgtb.icfre.gov.in.
Jervis, M.A. 2005. Insects as Natural Enemies: A Practical Perspective. Springer, Netherlands.
Kim, I.K., Z. Mendel, A. Protasov, D. Blumberg and J. La Salle. 2008. Taxonomy, biology and efficacy of two Australian parasitoids of the Eucalyptus gall wasp, Leptocybe invasa Fisher & La Salle (Hymenoptera: Eulophidae: Tetrastichinae). Zootaxa 1910: 1-20.
Luck, R.F., R. Stouthamer and L.P. Nunney. 1993. Sex determination and sex ratio patterns in parasitic Hymenoptera, pp. 442-476. In D.L. Wrensch and M.A. Ebbert, eds.), Evolution and Diversity of Sex Ratio in Insects ad Mites. Chapman and Hall, New York.
Mendel, Z., A. Protasov, N. Fisher and J. La Salle. 2004. Taxonomy and biology of Leptocybe invasa gen. & sp.n. (Hymenoptera: Eulophidae), an invasive gall inducer on Eucalyptus. Aust. J. Entomol. 43: 101-113.
Protasov, A., M. Doğanlar, J. La Salle and Z. Mendel. 2008. Occurrence of two local Megastigmus species parasitic on the eucalyptus gall wasp, Leptocybe invasa in Israel and Turkey. Phytoparasitica 36: 449-459.
Takagi, M. 1985. The reproductive strategy on the gregarious parasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae): optimum number of eggs in a single host. Oecologia 68: 1-6.
Thu, P.Q., B. Dell and T.I. Burgess. 2009. Susceptibility of 18 eucalypt species to the gall wasp Leptocybe invasa in the nursery and young plantations in Vietnam. Sci. Asia 35: 113-117.
Manuscript received 11 May 2011, accepted 20 October 2011
Now online at http://www.thaiagj.org
Related Documents











