Cell Tissue Res (1991) 265:219 230 Cell and Tissue Research Springer-Verlag 1991 Localization of neuropeptide Y-immunoreactive cells and fibres in the brain of the Japanese quail* N. Aste 1, C. Viglietti-Panzica 1, A. Fasoio 2, C. Andreone 2, H. Vaudry 3, G. Pelletier 4, and G.C. Panzica 1 1 Department of Human Anatomy and Physiology, University of Torino, Corso Massimo D'Azeglio 52, I-10126 Torino, Italy 2 Department of Animal Biology, University of Torino, Torino, Italy 3 Laboratory of Molecular Endocrinology, URA CNRS 650, University of Rouen, Mont-Saint-Aignan, France 4 Laboratory of Molecular Endocrinology, Hospital Center Laval University, Sainte-Foy, Quebec, Canada Accepted April 2, 1991 Summary. In the present study, we have demonstrated, by means of the biotin-avidin method, the widespread distribution of neuropeptide Y (NPY)-immunoreactive structures throughout the whole brain of the Japanese * A preliminary report of this study was presented at the 15th Conference of European Comparative Endocrinologists, Leuven, Belgium, September 1990 Offprint requests to." N. Aste Abbreviations: AA archistriatum anterius; A C nucleus accumbens; AM nucleus anterior medialis; APP avian pancreatic polypeptide; CNS central nervous system; CO chiasma opticum; CP com- missura posterior; CPi cortex piriformis; DIC differential interfer- ential contrast; DLA1 nucleus dorsolateralis anterior thalami, pars lateralis; DLAm nucleus dorsolateralis anterior thalami, pars me- dialis; E ectostriatum; EW nucleus of Edinger-Westphal; FLM fasciculus longitudinalis medialis; GCt substantia grisea centralis; GLv nucleus geniculatus lateralis, pars ventralis ; HA hyperstriatum accessorium; Hp hippocampus; HPLC high performance liquid chromatography; HV hyperstriatum ventrale; IF nucleus infundi- bularis; IO nucleus isthmo-opticus; IP nucleus interpeduncularis; IR immunoreactive; LA nucleus lateralis anterior thalami; LC nu- cleus linearis caudalis; LFS lamina frontalis superior; LH lamina hyperstriatica; LHRH luteinizing hormone-releasing hormone; LoC locus coeruleus; LPO lobus paraolfactorius; ME eminentia mediana; N neostriatum; NC neostriatum caudale; NPY neu- ropeptide Y; NIII nervus oculomotorius; NV nervus trigeminus; NVI nervus facialis; NVIIIc nervus octavus, pars cochlearis; nIV nucleus nervi oculomotorii; niX nucleus nervi glossopharyngei; nBOR nucleus opticus basalis (ectomamilaris) ; nCPa nucleus com- missurae pallii; nST nucleus striae terminalis; OM tractus occipito- mesencephalicus; OS nucleus olivaris superior; PA palaeostriatum augmentatum; PBS phosphate-buffered saline; POA nucleus praeopticus anterior; POM nucleus praeopticus medialis; POP nu- cleus praeopticus periventricularis; PP pancreatic polypeptide; PYY polypeptide YY; PVN nucleus paraventricularis magnocellu- laris; PVO organum paraventriculare; R nucleus raphes; ROT nu- cleus rotundus; RP nucleus reticularis pontis caudalis; Rpc nucleus reticularis parvocellularis ; RPgc nucleus reticularis pontis caudalis, pars gigantocellularis; RPO nucleus reticularis pontis oralis; SCd nucleus subcoeruleus dorsalis; SCv nucleus subcoeruleus ventralis; SCNm nucleus suprachiasmaticus, pars medialis; SCNl nucleus su- prachiasmaticus, pars lateralis; SL nucleus septalis lateralis; SM nucleus septalis medialis; Ta nucleus tangentialis; TeO tectum opti- cum; Tn nucleus taeniae; TPc nucleus tegmenti pedunculo-pontin- us, pars compacta; TSM tractus septo-mesencephalicus; TV nucle- us tegmenti ventralis; VeL nucleus vestibularis lateralis; VLTnucle- us ventrolateralis thalami; VMN nucleus ventromedialis hypotha- lami quail (Coturnix coturnix japonica). The prosencephalic region contained the highest concentration of both NPY-containing fibres and perikarya. Immunoreactive fibres were observed throughout, particularly within the paraolfactory lobe, the lateral septum, the nucleus taen- iae, the preoptic area, the periventricular hypothalamic regions, the tuberal complex, and the ventrolateral thala- mus. NPY-immunoreactive cells were represented by: a) small scattered perikarya in the telencephalic portion (i.e. archistriatal, neostriatal and hyperstriatal regions, hippocampus, piriform cortex); b) medium-sized cell bodies located around the nucleus rotundus, ventrolater- al, and lateral anterior thalamic nuclei; c) small clustered cells within the periventricular and medial preoptic nu- clei. The brainstem showed a less diffuse innervation, although a dense network of immunopositive fibres was observed within the optic tectum, the periaqueductal re- gion, and the Edinger-Westphal, linearis caudalis and raphes nuclei. Two populations of large NPY-containing perikarya were detected: one located in the isthmic re- gion, the other at the boundaries of the pons with the medulla. The wide distribution of NPY-immunoreactive structures within regions that have been demonstrated to play a role in the control of vegetative, endocrine and sensory activities suggests that, in birds, this neu- ropeptide is involved in the regulation of several aspects of cerebral functions. Key words: Neuropeptide Y (NPY) - Immunocytochem- istry Avian brain - Japanese quail, Coturnix coturnix japonica Neuropeptide Y (NPY) is a 36 amino acid peptide that was originally isolated from porcine brain extracts (Tate- moto et al. 1982; Tatemoto 1982). It shares considerable structural and biological homologies with several pan- creatic polypeptides (PP), such as avian PP (APP), which was the first to be isolated (Kimmel et al. 1975), and polypeptide YY (PYY) (Tatemoto et al. 1982; Tatemoto 1982; Allen et al. 1987). NPY is one of the most abundant peptides character- ized in the central and peripheral nervous system of ver-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Cell Tissue Res (1991) 265:219 230 Cell and Tissue Research �9 Springer-Verlag 1991
Localization of neuropeptide Y-immunoreactive cells and fibres in the brain of the Japanese quail* N. Aste 1, C. Viglietti-Panzica 1, A. Fasoio 2, C. Andreone 2, H. Vaudry 3, G. Pelletier 4, and G.C. Panzica 1
1 Department of Human Anatomy and Physiology, University of Torino, Corso Massimo D'Azeglio 52, I-10126 Torino, Italy 2 Department of Animal Biology, University of Torino, Torino, Italy 3 Laboratory of Molecular Endocrinology, URA CNRS 650, University of Rouen, Mont-Saint-Aignan, France 4 Laboratory of Molecular Endocrinology, Hospital Center Laval University, Sainte-Foy, Quebec, Canada
Accepted April 2, 1991
Summary. In the present study, we have demonstrated, by means of the biotin-avidin method, the widespread distribution of neuropeptide Y (NPY)-immunoreactive structures throughout the whole brain of the Japanese
* A preliminary report of this study was presented at the 15th Conference of European Comparative Endocrinologists, Leuven, Belgium, September 1990
Offprint requests to." N. Aste
Abbreviations: AA archistriatum anterius; A C nucleus accumbens; A M nucleus anterior medialis; A P P avian pancreatic polypeptide; CNS central nervous system; CO chiasma opticum; CP com- missura posterior; CPi cortex piriformis; DIC differential interfer- ential contrast; DLA1 nucleus dorsolateralis anterior thalami, pars lateralis; DLAm nucleus dorsolateralis anterior thalami, pars me- dialis; E ectostriatum; E W nucleus of Edinger-Westphal; F L M fasciculus longitudinalis medialis; GCt substantia grisea centralis; GLv nucleus geniculatus lateralis, pars ventralis ; HA hyperstriatum accessorium; Hp hippocampus; H P L C high performance liquid chromatography; H V hyperstriatum ventrale; IF nucleus infundi- bularis; IO nucleus isthmo-opticus; IP nucleus interpeduncularis; IR immunoreactive; LA nucleus lateralis anterior thalami; L C nu- cleus linearis caudalis; LF S lamina frontalis superior; L H lamina hyperstriatica; L H R H luteinizing hormone-releasing hormone; LoC locus coeruleus; LPO lobus paraolfactorius; M E eminentia mediana; N neostriatum; N C neostriatum caudale; N P Y neu- ropeptide Y; NII I nervus oculomotorius; N V nervus trigeminus; N V I nervus facialis; NVIIIc nervus octavus, pars cochlearis; n IV nucleus nervi oculomotorii; n i X nucleus nervi glossopharyngei; nBOR nucleus opticus basalis (ectomamilaris) ; nCPa nucleus com- missurae pallii; n S T nucleus striae terminalis; O M tractus occipito- mesencephalicus; OS nucleus olivaris superior; PA palaeostriatum augmentatum; P B S phosphate-buffered saline; POA nucleus praeopticus anterior; P O M nucleus praeopticus medialis; POP nu- cleus praeopticus periventricularis; PP pancreatic polypeptide; P Y Y polypeptide YY; P V N nucleus paraventricularis magnocellu- laris; PVO organum paraventriculare; R nucleus raphes; R O T nu- cleus rotundus; R P nucleus reticularis pontis caudalis; Rpc nucleus reticularis parvocellularis ; RPgc nucleus reticularis pontis caudalis, pars gigantocellularis; RPO nucleus reticularis pontis oralis; SCd nucleus subcoeruleus dorsalis; SCv nucleus subcoeruleus ventralis; SCNm nucleus suprachiasmaticus, pars medialis; SCNl nucleus su- prachiasmaticus, pars lateralis; SL nucleus septalis lateralis; S M nucleus septalis medialis; Ta nucleus tangentialis; TeO tectum opti- cum; Tn nucleus taeniae; TPc nucleus tegmenti pedunculo-pontin- us, pars compacta; T S M tractus septo-mesencephalicus; T V nucle- us tegmenti ventralis; VeL nucleus vestibularis lateralis; VLTnucle- us ventrolateralis thalami; V M N nucleus ventromedialis hypotha- lami
quail ( C o t u r n i x c o t u r n i x j a p o n i c a ) . The prosencephalic region contained the highest concentration of both NPY-containing fibres and perikarya. Immunoreactive fibres were observed throughout, particularly within the paraolfactory lobe, the lateral septum, the nucleus taen- iae, the preoptic area, the periventricular hypothalamic regions, the tuberal complex, and the ventrolateral thala- mus. NPY-immunoreactive cells were represented by: a) small scattered perikarya in the telencephalic portion (i.e. archistriatal, neostriatal and hyperstriatal regions, hippocampus, piriform cortex); b) medium-sized cell bodies located around the nucleus rotundus, ventrolater- al, and lateral anterior thalamic nuclei; c) small clustered cells within the periventricular and medial preoptic nu- clei. The brainstem showed a less diffuse innervation, although a dense network of immunopositive fibres was observed within the optic tectum, the periaqueductal re- gion, and the Edinger-Westphal, linearis caudalis and raphes nuclei. Two populations of large NPY-containing perikarya were detected: one located in the isthmic re- gion, the other at the boundaries of the pons with the medulla. The wide distribution of NPY-immunoreactive structures within regions that have been demonstrated to play a role in the control of vegetative, endocrine and sensory activities suggests that, in birds, this neu- ropeptide is involved in the regulation of several aspects of cerebral functions.
Key words: Neuropeptide Y (NPY) - Immunocytochem- istry Avian brain - Japanese quail, C o t u r n i x c o t u r n i x
j a p o n i c a
Neuropeptide Y (NPY) is a 36 amino acid peptide that was originally isolated from porcine brain extracts (Tate- moto et al. 1982; Tatemoto 1982). It shares considerable structural and biological homologies with several pan- creatic polypeptides (PP), such as avian PP (APP), which was the first to be isolated (Kimmel et al. 1975), and polypeptide YY (PYY) (Tatemoto et al. 1982; Tatemoto 1982; Allen et al. 1987).
NPY is one of the most abundant peptides character- ized in the central and peripheral nervous system of ver-

220
tebrates (for a more extensive literature, see Danger et al. 1990). Moreover, the NPY-gene shows strong evolution- ary conservation as demonstrated by the comparison of its sequence in mammals , chicken, horned shark, and goldfish (Larhammar et al. 1990). The widespread cen- tral distribution of both NPY-immunoreact ive (IR) peri- karya and fibres has been demonstrated in many mam- malian species, particularly in rat (Allen et al. 1983; Pel- letier et al. 1984b; Chronwall et al. 1985; De Quidt and Emerson 1986a, b), monkey (Smith et al. 1985; Bons et al. 1990), cat (Whale and Albus 1984), and man (Ad- rian et al. 1983; Dawbarn et al. 19'84; Chan-Palay and Yasargil 1986). In particular, the largest numbers of N P Y - I R structures have been observed in the cortex, septum, h ippocampus and nucleus arcuatus, in the peri- ventricular regions of the thalamus and hypothalamus, in the bed nucleus of stria terminalis, in the pineal gland and along the olfactory and visual pathways. These mor- phological observations together with physiological stu- dies (for reviews, see Gray and Morley 1986; Danger et al. 1990) suggest that N P Y plays a role as a neu- rotransmitter or a neuromodula tor involved in several central nervous system (CNS) functions, such as the con- trol of the secretion of several pituitary hormones (ad- renocorticotropic hormone: Wahlestedt e tal . 1987; growth hormone, tyreostimulating hormone, prolactin: Fuxe et al. 1982; luteinizing hormone-releasing hormone [LHRH] : Kalra 1985; McDonald et al. 1985), and in sexual behaviour (Clark et al. 1985; Kalra et al. 1988; Dornan and Malsbury 1989; Poggioli et al. 1990). More- over, N P Y partly mimics the action of noradrenaline in the control of food and water intake (for a review see Leibovitz 1989), of L H R H secretion (Kalra and Crowley 1984; McDona ld et al. 1985), and of vasocon- striction and blood pressure (Ekblad et al. 1984); it also colocalizes with catecholamines (Everitt e tal . 1984; Sawchenko et al. 1985).
Although the distribution of N P Y has been elucidat- ed in lower vertebrates, such as reptiles (Reiner and Oli- vier 1987), amphibians (Danger et al. 1985; Caillez et al. 1987; Perroteau et al. 1988) and fishes (Vallarino et al. 1988; Noe et al. 1989; Pontet et al. 1989), few studies are available regarding birds. Some deal with the whole- brain distribution (chicken: Kuenzel and McMurt ry 1988; pigeon: Lorenz and Skoftisch 1990), others only involve circumscribed brain regions (house sparrow, su- prachiasmatic nucleus: Cassone and Moore 1988; pi- geon, forebrain: Anderson and Reiner 1990; chicken, optic pathway: B1/ihser and Tobies 1990). Moreover, two physiological investigations have demonstrated that, in birds, intracerebroventricularly injected N P Y is involved in the control of feeding (Kuenzel et al. 1987) and plasma insulin levels (Kuenzel and McMurt ry 1988).
The aim of the present study was to provide a more complete picture of the distribution of N P Y within the avian CNS. In view of the effect of N P Y in controlling several aspects of the mammal ian reproduction and of its possible role as one of the modula tors of sexual behaviour in birds, we paid special attention to struc- tures involved in the control of reproduction.
Materials and methods
Two male and 3 female sexually active adult Japanese quails (Cotur- nix coturnix japonica) were used. Birds were injected with 0.1 ml heparin solution (30 mg/ml) (Parke-Davis) into the wing vein and deeply anaesthetized with an overdose of Hypnodil| (Janssen Phar- maceutica, Belgium). The animals were then perfused transcardial- ly with a saline solution (NaC1 150 mM in water), followed by Heidenhain's (SUSA) or Bouin's fixatives (Romeis 1968). The dis- sected brains were stored in the same fixative at 4~ for 24 h and embedded in Histosec (Merck). Specimens were serially sec- tioned in a frontal plane, at a thickness of 10 gm; one section per 50 gm was processed for immunocytochemistry. The adjacent section was Nissl-stained to improve the identification of nervous structures.
Immunocytochemistry
After inhibition of endogenous peroxidase activity with 7.5% hy- drogen peroxide and 2.28% periodic acid, deparaffinized sections were incubated with a normal goat serum (Vectastain Labtek) dilut- ed 1:100 in 0.01 M phosphate-buffered saline (PBS), pH 7.2, in a moist chamber, for 20 rain at room temperature (RT).
The primary antibody was raised in rabbit against synthetic porcine NPY; its production and the results of the assays per- formed to control its specificity and cross-reactivity with other neuropeptides have been previously described (Pelletier et al. 1984b). The anti-NPY serum was employed at a dilution of 1:800 in PBS, and the incubation was carried out overnight at RT in a moist chamber. After several rinses in PBS, the sections were incubated for 30 min with a secondary biotinylated antibody (Vec- tastain Labtek, 1:250), followed by a avidin-biotinylated peroxi- dase complex (Vectastain Labtek, 1:250). The peroxidase activity was revealed with a solution containing 0A 5 mg/ml 3',3'-diamino- benzidine and 0.025% hydrogen peroxide in 0.05 M TRIS-HC1 pH 7.6.
Control reactions were performed by replacing the anti-NPY serum with normal serum, and by omitting the secondary serum.
Morphometry
The immunohistochemically stained sections were subjected to a morphometric analysis by means of an image analyser based upon an EPSON AX (80286) computer, connected to an Olympus BHS microscope through a CCD TV camera OVC), a digitizing board (PC Vision plus, Imaging Technology Inc., Woburn, MA, USA), and a commercially available morphometry program (Image Mea- sure, Microscience, Inc., Washington, USA). Measurements of im- munopositive neurons were made employing the x 60 objective, but only on those cells in which the nucleus was clearly detectable. The program provides values of the area (A), perimeter (P), diame- ter of the equivalent circle, and shape factor (4 ~ A/P 2) of the detected objects, their means and standard errors (s.c.). Neurons were measured in 2 females fixed with the SUSA fluid. Differences between the mean values for each animal were about 5%; for this reason the morphometric data were pooled in a single group containing cells measured in the two birds.
The sections were observed and photographed by means of a Zeiss Axioplan microscope equipped with differential interferen- tial contrast (DIC). A Kodak Wratten 44 gelatine filter was em- ployed to increase the contrast of non-DIC pictures. The exact location of positive cell groups and nerve fibres was determined both on adjacent NissI-stained sections and by employing phase- contrast objectives.

221
Results
NPY-IR structures (fibres and cell bodies) were distrib- uted within the whole quail brain, with some regional differences. NPY fibres were also detected in the walls of the cerebral blood vessels. However, the cerebellum exhibited no immunopositivity. The intensity of the im- munostaining of cell bodies was not affected by the em- ployed fixatives, nor by individual variations, but was strikingly dependent on the topographical location of the elements. No distinct sexual difference was observed in the present limited number of birds.
The distribution of the NPY-IR elements is reported in Fig. I (a-n) showing different levels of the quail brain. The nomenclature adopted in the present study is that developed by Kuenzel and Masson (1988) for the chicken brain, with some minor modifications for the hypotha- lamic region (Panzica et al. 1991).
Telencephalon
The telencephalon contained the most extensive popula- tion of NPY-IR cell bodies in the brain, and a supply of IR fibres showing strong differences in their distribu- tion and content within its subdivisions, hnmunoreactive cell bodies were observed within the hyperstriatum ac- cessorium (HA), the hyperstriatum ventrale (HV) and the neostriatum (N). Several scattered cells were visible in the most rostral portion of the telencephalon, where these three regions did not show clear subdivisions. These perikarya were generally small (n = 50, mean di- ameters +_ s.e. = 10.6+_0.14 ~tm) and round or polygonal, some having long, thin processes bearing thinner branches (Fig. 2 a), and some supplied with short pro- cesses. This region showed the lowest density of NPY-IR fibres within the telencephalon. These fibres were char- acterized by a dotted pattern and were dispersed among the immunonegative elements. Sometimes, beaded IR fibres almost completely surrounded immunonegative cell bodies.
At more caudal levels (Fig. ~ a, b), the HA, HV and N were well differentiated, A large number of small, round or elongated IR perikarya bearing short processes were observed within these areas. In particular, they were scattered throughout the lateral and dorsal portion of the N, in the medial and ventral aspects of the HA, and between the lamina frontalis superior (LFS) and the lamina hyperstriatica (LH) bordering the HV. In this last region, we observed several fusiform perikarya with their major axis paralleling one of the two laminae. A moderate density of NPY-IR fibres was detected within these regions, but no close contacts with the surrounding elements were evident.
The hippocampus (Hp) presented many scattered small (n=50, mean diameter+_s.e.=ll.59+0.13 gm), round or irregular, intensely stained, IR cell bodies. These cells generally had long and thin processes. In the same region, the IR fibres showed a medium density
and their distribution pattern was similar to that pre- viously described for HA, HV, and N.
Some IR perikarya were located within the cortex piriformis (Fig. 2c) close to the pial surface or in a deeper portion. They were round or elongated, bipolar or multipolar with thin processes giving a stellated ap- pearance.
The archistriatal and paleostriatal regions contained various clusters of weakly IR neurons, and a large amount of IR fibres. Several oval, bipolar or multipolar cells with short thick processes were located both within the nucleus taeniae (Fig. 2b) and the archistriatum aug- mentatum. A very dense network of thick IR fibres was found in the lobus paraolfactorius (LPO) representing the most heavily innervated region of the whole telence- phalon. Several negative cell bodies were located among a particularly tight network of IR axons and nerve termi- nals (Fig. 3).
The septal region showed a dense NPY innervation in its lateral portion (Figs. 1 d, e, 2d) where immuno- positive terminals were visualized in close proximity to negative cells (Fig. 2e). Because of the large number of terminals near the cell bodies and the major processes, it was difficult to decide whether some of them were true IR neurons. A few small round weakly IR perikarya were scattered within the nucleus accumbens and the nucleus of stria terminalis. In this last nucleus, a medium density of NPY-IR fibres was also present. A large sup- ply of fibres was detected within the nucleus commissur- ae pallii (nCPa).
Diencephalon
Thalamus. The periventricular and the lateral portions of the thalamus contained a large supply of NPY-IR structures (both fibres and perikarya), whereas the mid- dle regions showed only a few scattered fibres. The nuclei of the lateral thalamus were surrounded by several, in- tensely stained cell bodies, showing some common fea- tures, such as the cell shape and size (n = 50, mean diame- ter+_s.e. = 17.03_+0.4 gm), and the dendritic arboriza- tion. The IR neurons were located partly bordering the nucleus lateralis anterior thalami (LA), distributed close to the ventral (sometimes within the optic tract), lateral, and dorsal boundaries of it. Some were scattered within the lateral portion of the nucleus ventrolateralis thalami (VLT) and between the other thalamic nuclei identified in Nissl-stained adjacent sections (Fig. 1 d, e). The main population of neurons was composed of small clusters in proximity to the nucleus rotundus (ROT) and by a single layer of IR cells surrounding it (Figs. 1 f, 4a). These elements showed a complex rostro-caudal ar- rangement around this large thalamic structure. IR neu- rons were clustered ventrally to the ROT at the level of its most anterior boundaries; then, moving caudally, a single layer of neurons almost completely surrounded the ROT. Lastly, two clusters were located at its ventral and dorsolateral caudal aspects. The cell bodies border- ing the ROT were elongated or rounded, and bipolar
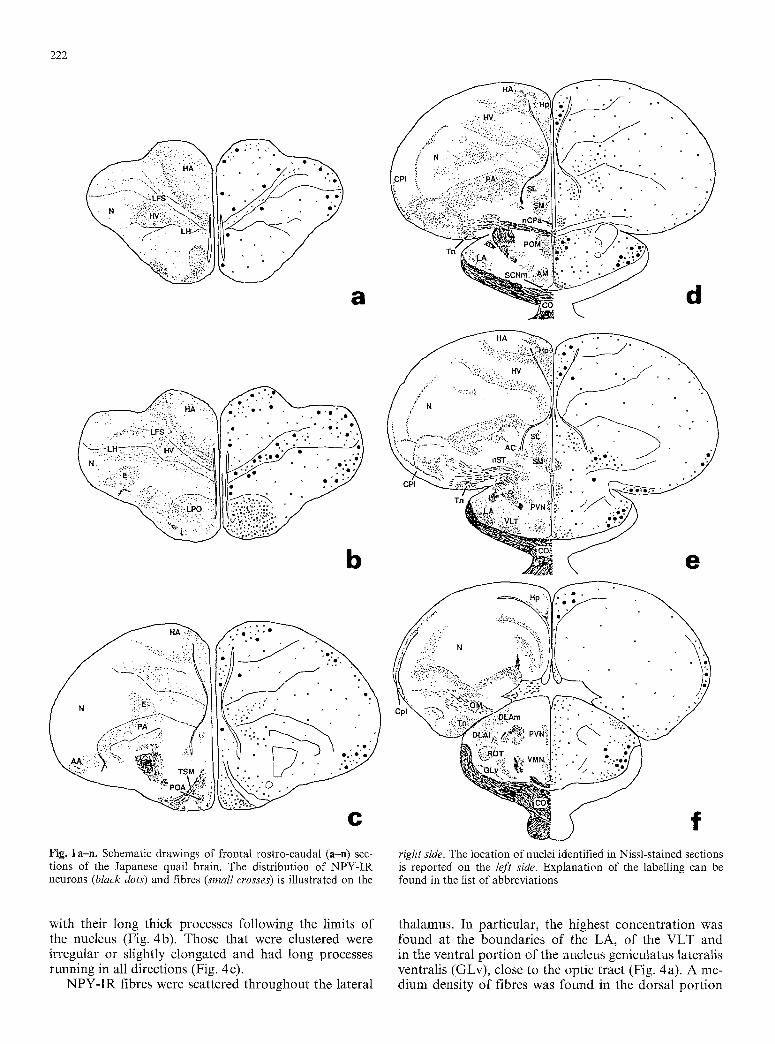
222
b e
Q N J~.:;'g'.'::; ....... :.
e f Fig. 1 an. Schematic drawings of frontal rostro-caudal (a-n) sec- right side. The location of nuclei identified in Nissl-stained sections tions of the Japanese quail brain. The distribution of NPY-IR is reported on the left side. Explanation of the labelling can be neurons (black dots) and fibres (small crosses) is illustrated on the found in the list of abbreviations
with their long thick processes following the limits of the nucleus (Fig. 4b). Those that were clustered were irregular or slightly elongated and had long processes running in all directions (Fig. 4 c).
NPY-IR fibres were scattered throughout the lateral
thalamus. In particular, the highest concentration was found at the boundaries of the LA, of the VLT and in the ventral portion of the nucleus geniculatus lateralis ventralis (GLv), close to the optic tract (Fig. 4a). A me- dium density of fibres was found in the dorsal portion
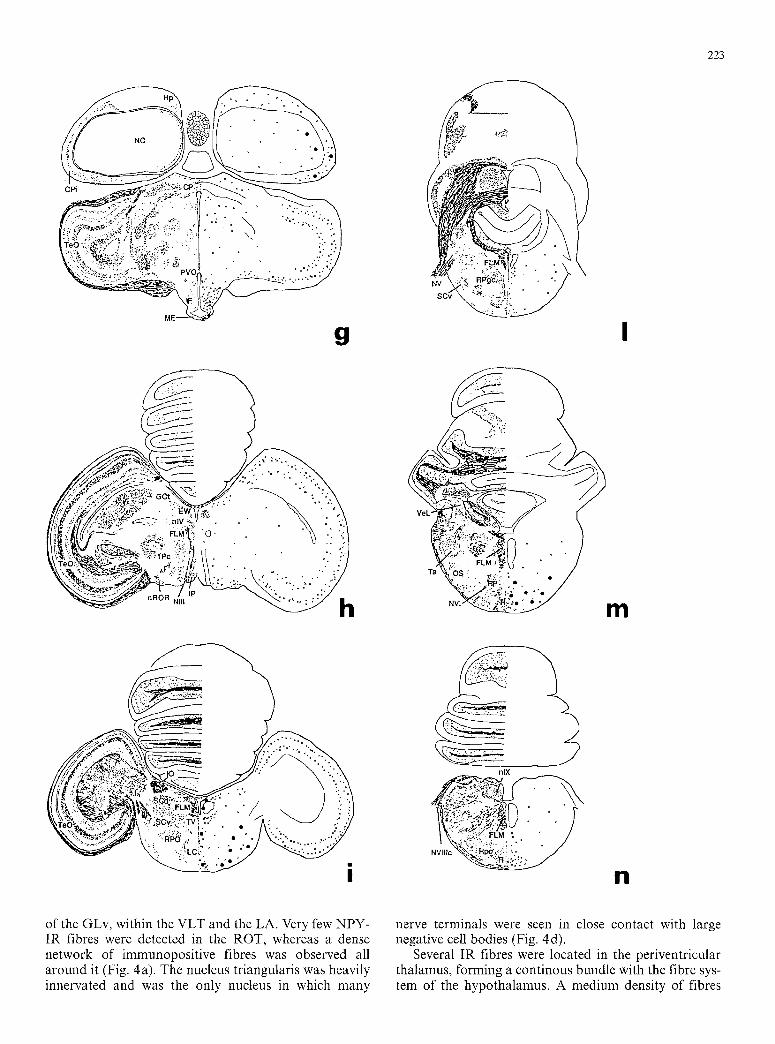
223
�9 g I
~ ~ ._~: , m ~ ~-/..,. '.....x I ~ ~ ~ //i... ~ :-::..X ~ - ~
2
I n
of the GLv, within the VLT and the LA. Very few NPY- IR fibres were detected in the ROT, whereas a dense network of immunopositive fibres was observed all around it (Fig. 4a). The nucleus triangularis was heavily innervated and was the only nucleus in which many
nerve terminals were seen in close contact with large negative cell bodies (Fig. 4d).
Several IR fibres were located in the periventricular thalamus, forming a continous bundle with the fibre sys- tem of the hypothalamus. A medium density of fibres
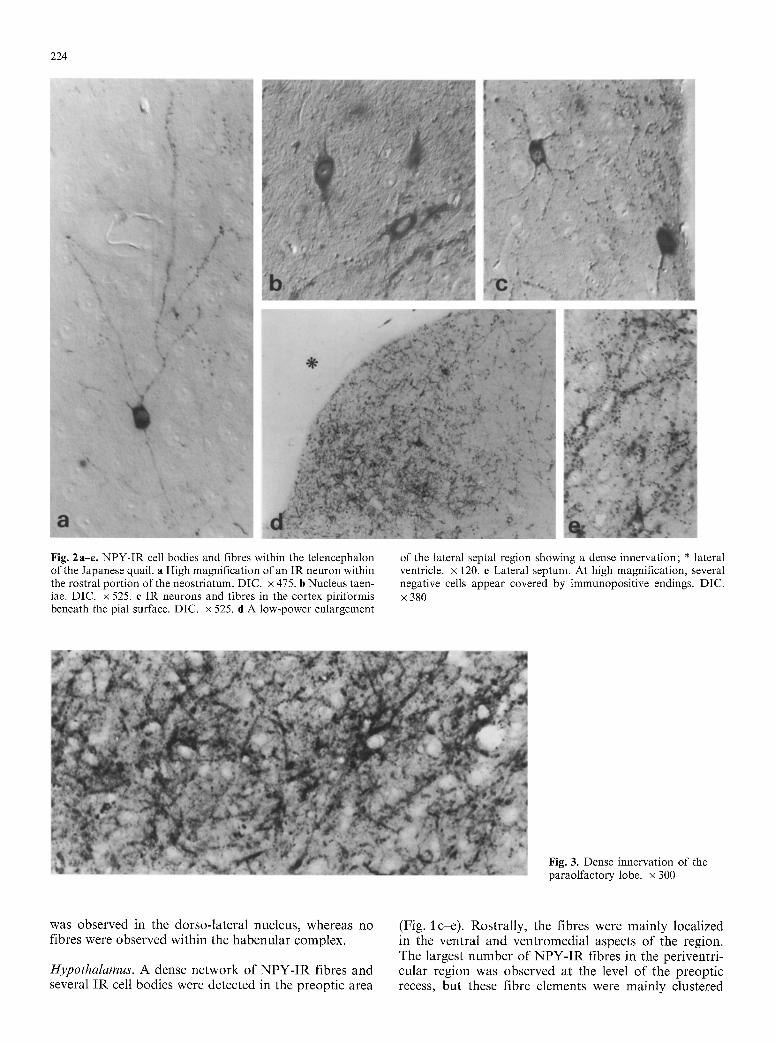
224
Fig. 2a-e. NPY-IR cell bodies and fibres within the telencephalon of the Japanese quail, a High magnification of an IR neuron within the rostral portion of the neostriatum. DIC. • 475. b Nucleus taen- iae. DIC. x 525. e IR neurons and fibres in the cortex piriformis beneath the pial surface. DIC. x 525. d A low-power enlargement
of the lateral septal region showing a dense innervation; * lateral ventricle, x 120. e Lateral septum. At high magnification, several negative cells appear covered by immunopositive endings. DIC. x 380
Fig. 3. Dense innervation of the paraolfactory lobe. x 300
was obse rved in the do r so - l a t e r a l nucleus, whereas no fibres were obse rved wi th in the h a b e n u l a r complex .
Hypothalamus. A dense n e t w o r k o f N P Y - I R f ibres and several I R cell bodies were de tec ted in the p reop t i c a rea
(Fig. 1 ~ e ) . Ros t ra l ly , the f ibres were ma in ly loca l ized in the vent ra l and v e n t r o m e d i a l aspects o f the region. The larges t n u m b e r o f N P Y - I R fibres in the per ivent r i - cu la r r eg ion was obse rved a t the level o f the p r e o p t i c recess, bu t these f ibre e lements were m a i n l y c lus tered
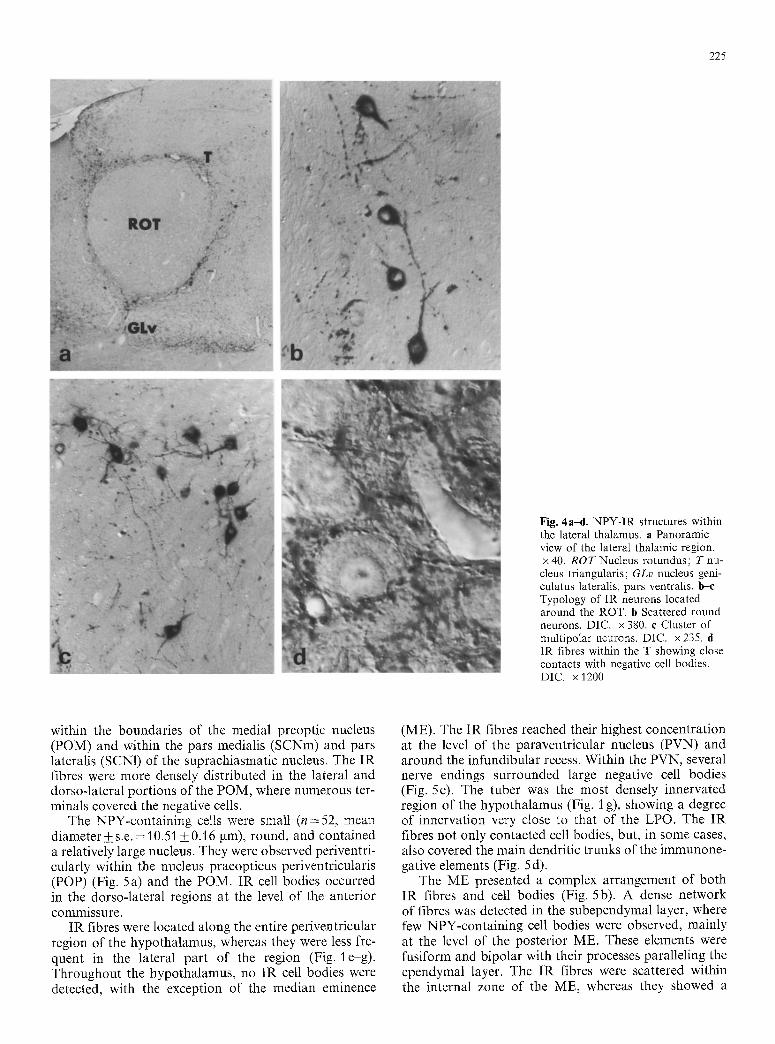
225
Fig. 4a-d. NPY-IR structures within the lateral thalamus, a Panoramic view of the lateral thalamic region. x 40. R O T Nucleus rotundus; T nu- cleus triangularis; GLv nucleus geni- culatus lateralis, pars ventralis, b--e Typology of IR neurons located around the ROT. b Scattered round neurons. DIC. • 380. e Cluster of multipolar neurons. DIC. x 235. d IR fibres within the T showing close contacts with negative cell bodies. DIC. x 1200
within the boundaries of the medial preoptic nucleus (POM) and within the pars medialis (SCNm) and pars lateralis (SCN1) of the suprachiasmatic nucleus. The IR fibres were more densely distributed in the lateral and dorsoqateral portions of the POM, where numerous ter- minals covered the negative cells.
The NPY-containing cells were small (n--52, mean diameter + s.e. = 10.51 _+ 0.16 ~tm), round, and contained a relatively large nucleus. They were observed periventri- cularly within the nucleus praeopticus periventricularis (POP) (Fig. 5 a) and the POM. IR cell bodies occurred in the dorso-lateral regions at the level of the anterior commissure.
IR fibres were located along the entire periventricular region of the hypothalamus, whereas they were less fre- quent in the lateral part of the region (Fig. l e g ) . Throughout the hypothalamus, no IR cell bodies were detected, with the exception of the median eminence
(ME). The IR fibres reached their highest concentration at the level of the paraventricular nucleus (PVN) and around the infundibular recess. Within the PVN, several nerve endings surrounded large negative cell bodies (Fig. 5c). The tuber was the most densely innervated region of the hypothalamus (Fig. 1 g), showing a degree of innervation very close to that of the LPO. The IR fibres not only contacted cell bodies, but, in some cases, also covered the main dendritic trunks of the immunone- gative elements (Fig. 5 d),
The ME presented a complex arrangement of both IR fibres and cell bodies (Fig. 5b). A dense network of fibres was detected in the subependymal layer, where few NPY-containing cell bodies were observed, mainly at the level of the posterior ME. These elements were fusiform and bipolar with their processes paralleling the ependymal layer. The IR fibres were scattered within the internal zone of the ME, whereas they showed a
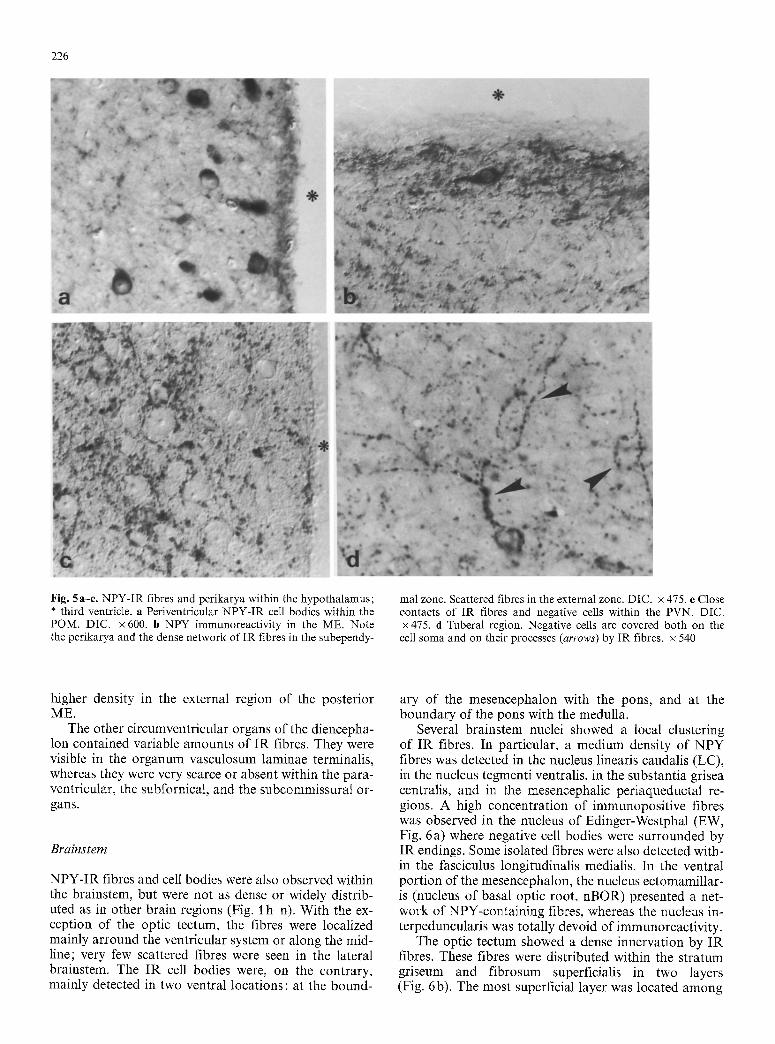
226
Fig. 5a-e. NPY-IR fibres and perikarya within the hypothalamus; * third ventricle, a Periventricular NPY-IR cell bodies within the POM. DIC. x 600. b NPY immunoreactivity in the ME. Note the perikarya and the dense network of IR fibres in the subependy-
mal zone. Scattered fibres in the external zone. DIC. x 475. e Close contacts of IR fibres and negative cells within the PVN. DIC. • 475. d Tuberal region. Negative cells are covered both on the ceil soma and on their processes (arrows) by IR fibres, x 540
higher density in the external region of the posterior ME.
The other circumventricular organs of the diencepha- lon contained variable amounts of IR fibres. They were visible in the organum vasculosum laminae terminalis, whereas they were very scarce or absent within the para- ventricular, the subfornical, and the subcommissural or- gans.
Brainstem
N P Y - I R fibres and cell bodies were also observed within the brainstem, but were not as dense or widely distrib- uted as in other brain regions (Fig. I h-n). With the ex- ception of the optic tectum, the fibres were localized mainly arround the ventricular system or along the mid- line; very few scattered fibres were seen in the lateral brainstem. The I R cell bodies were, on the contrary, mainly detected in two ventral locations: at the bound-
ary of the mesencephalon with the pons, and at the boundary of the pons with the medulla.
Several brainstem nuclei showed a local clustering of I R fibres. In particular, a medium density of N P Y fibres was detected in the nucleus linearis caudalis (LC), in the nucleus tegmenti ventralis, in the substantia grisea centralis, and in the mesencephalic periaqueductal re- gions. A high concentrat ion of immunoposi t ive fibres was observed in the nucleus of Edinger-Westphal (EW, Fig. 6a) where negative cell bodies were surrounded by I R endings. Some isolated fibres were also detected with- in the fasciculus longitudinalis medialis. In the ventral port ion of the mesencephalon, the nucleus ectomamillar- is (nucleus of basal optic root, nBOR) presented a net- work of NPY-containing fibres, whereas the nucleus in- terpeduncularis was totally devoid of immunoreactivity.
The optic tectum showed a dense innervation by I R fibres. These fibres were distributed within the s t ratum griseum and f ibrosum superficialis in two layers (Fig. 6b). The most superficial layer was located among
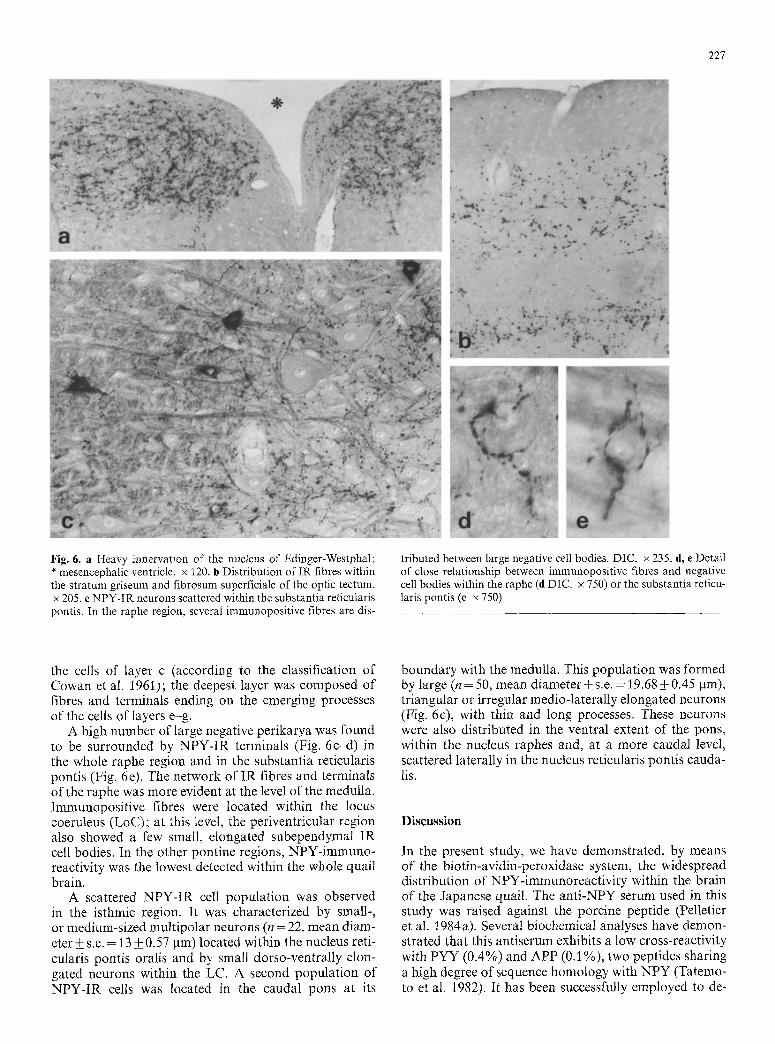
227
Fig. 6. a Heavy innervation of the nucleus of Edinger-Westphal; * mesencephalic ventricle, x 120. b Distribution of IR fibres within the stratum griseum and fibrosum superficiale of the optic tectum. x 205. c NPY-IR neurons scattered within the substantia reticularis pontis. In the raphe region, several immunopositive fibres are dis-
tributed between large negative cell bodies. DIC. x 235. d, e Detail of close relationship between immunopositive fibres and negative cell bodies within the raphe (d DIC. x 750) or the substantia reticu- laris pontis (e x 750)
the cells of layer c (according to the classification of Cowan et al. 1961); the deepest layer was composed of fibres and terminals ending on the emerging processes of the cells of layers e-g.
A high number of large negative perikarya was found to be surrounded by NPY-IR terminals (Fig. 6c-d) in the whole raphe region and in the substantia reticularis pontis (Fig. 6e). The network of IR fibres and terminals of the raphe was more evident at the level of the medulla. Immunopositive fibres were located within the locus coeruleus (LoC); at this level, the periventricular region also showed a few small, elongated subependymal IR cell bodies. In the other pontine regions, NPY-immuno- reactivity was the lowest detected within the whole quail brain.
A scattered NPY-IR cell population was observed in the isthmic region. It was characterized by small-, or medium-sized multipolar neurons (n = 22, mean diam- eter +_ s.e. = 13 + 0.57 ~tm) located within the nucleus reti- cularis pontis oralis and by small dorso-ventrally elon- gated neurons within the LC. A second population of NPY-IR cells was located in the caudal pons at its
boundary with the medulla. This population was formed by large (n = 50, mean diameter_+ s.e. = 19.68 • 0.45 ~tm), triangular or irregular medio-laterally elongated neurons (Fig. 6c), with thin and long processes. These neurons were also distributed in the ventral extent of the pons, within the nucleus raphes and, at a more caudal level, scattered laterally in the nucleus reticularis pontis cauda- lis.
Discussion
In the present study, we have demonstrated, by means of the biotin-avidin-peroxidase system, the widespread distribution of NPY-immunoreactivi ty within the brain of the Japanese quail. The anti-NPY serum used in this study was raised against the porcine peptide (Pelletier et al. 1984a). Several biochemical analyses have demon- strated that this antiserum exhibits a low cross-reactivity with PYY (0.4%) and APP (0.1%), two peptides sharing a high degree of sequence homology with NPY (Tatemo- to et al. 1982), It has been successfully employed to de-

228
tect NPY-like immunoreactivity in a variety of verte- brates: fishes (Vallarino et al. ]988; Pontet et al. 1989), amphibians (Danger et al. 1985; Caillez et al. 1987; Per- roteau et al. 1988), and mammals (Pelletier et al. 1984a, b; L~ger et al. 1987; Bons et al. 1990). It thus appears that the antigenic determinants of NPY have been highly conserved in vertebrates. Accordingly, biochemical data show that NPY-like peptides of fishes and amphibians (Danger et al. 1985; Perroteau et al. 1988; Vallarino et al. 1988; Noe et al. 1989; Pontet et al. 1989) have a retention time in HPLC similar to that of porcine or human NPY (differing from each other only by a single amino acid, Corder et al. 1984). In addition, a recent study has demonstrated the strong conservation of the NPY-gene sequence in fishes, birds and mammals (Lar- hammar et al. 1990).
NPY-immunoreactivity was observed throughout the whole brain of the Japanese quail, with the exception of the cerebellum. IR fibres were found to be spread, with local differences in their density, from the telence- phalon to the medulla, whereas IR cell bodies were de- tected only in certain specific regions. In the present study, birds were not subjected to treatments to increase the peptide content (e.g. intraventricular injection of col- chicine), in order to prevent any interference with the physiological situation. Indeed, recent studies have dem- onstrated that the administration of colchicine in rats can increase not only the immunoreactivity for some peptides of the PVN, but also the synthesis of the mRNA for those peptides (Ceccatelti et al. 1991), suggesting an influence of the drug on the expression of the genetic message.
To date, very few studies have been undertaken con- cerning either the distribution of NPY in birds or its physiological role (Kuenzel etal. 1987; Kuenzel and McMurtry 1988). It is hence difficult to compare the present results in quail with those in other species (B1/ihser and Tobies 1990; Lorenz and Skofitsch 1990). The only complete studies published deal with the pigeon telencephalon (Anderson and Reiner 1990) and with some regions of the chicken brain (Kuenzel and McMurtry 1988). They describe a distribution of IR cell bodies substantially similar to that observed in the quail.
The prosencephalic regions of the quail brain pre- sented the highest concentration of both IR fibres and cell bodies. In the telencephalon, NPY-IR structures were distributed both within the cortex-equivalent por- tions (Reiner et al. 1984) and within the basal ganglia (paleostriatal region). The quail telencephalon showed smaller numbers of IR cells than that of the pigeon (An- derson and Reiner 1990), probably because of the use of colchicine in the pigeon study. In contrast to the pi- geon, the quail LPO showed no IR cell bodies; however, it cannot be excluded that the dense innervation of the region masks the existence of a few immunopositive peri- karya. The septal region was also endowed with IR fibres (mainly in the lateral septum, the nucleus accum- bens, and the nucleus striae terminalis) and presented scattered faintly IR cell bodies.
The presence of such a population of IR neurons in cortex- and striatal-equivalent parts of the avian brain
is to some extent comparable with the distribution ob- served in rodents (De Quidt and Emson 1986b). NPY is known to have inhibitory effects on mammalian corti- cal target structures (Cotmers et al. 1987), and the pres- ent data, together with those in the pigeon, suggest that it might play a similar role in the intrinsic circuitry of the avian telencephalon.
In the diencephalon, IR cell bodies cluster in two more clearly identifiable areas: the preoptic periventricu- lar region, where they are mainly associated with the POM and POP, and the lateral thalamic region (involved in the thalamofugal optic pathway; Kuenzel 1989), sur- rounding the ROT and the LA. IR fibres reach the high- est density in the periveutricular hypothalamus and in the lateral anterior thalamus (e.g. the GLv, the LA, and the DLA1). Thus, the optic pathway (Karten et al. 1973) appears to be one of the major targets of the system. Indeed both primary centres, such as the GLv, nBOR and optic tectum, and secondary centres, such as the EW and the anterior lateral thalamus, are strongly inner- vated. The presence of high levels of immunoreactivity within the PVN and periventricular hypothalamic re- gions confirms, as demonstrated in chicken (Kuenzel et al. 1987), a direct involvement of the NPY in the con- trol of feeding in birds. In avian species, other functions regulated by diencephalic centres demonstrating a large supply of NPY-immunoreactivity are water intake (Den- bow and Myers 1982), thermoregulation (Sato and Si- mon 1988), and the control of hypophysial secretion (Korf et al. 1988). Recent studies in chicken (Merckaert and Vandesande 1990) have demonstrated that cultured adenohypophysial cells contain NPY-binding sites. To- gether with the identification of NPY-IR fibres in the external zone of the ME, these data suggest that the peptide has a possible direct role as a hypophysiotrophic factor.
In the brainstem, the NPY system is represented by few scattered cell bodies. They are mainly located in the ventral isthmic region and at the ventral boundaries of the pons with the medulla. The fibre innervation pat- tern also appears to be less marked in comparison with upper levels. Outside the optic centres, the fibres are distributed within the median region, both periventricu- larly, and along the raphe region. This distribution of NPY immunoreactivity compared with that of catechol- amines and serotonin within the quail brainstem (Panzi- ca et al. 1990b; Cozzi et al. 1991) suggests a possible interaction of biogenic amines and NPY.
NPY-IR structures can be observed in several brain regions that are targets of sexual hormones or that are involved in the regulation of different reproductive activ- ities. In particular, they are observed in the telencephalic nucleus taeniae and in the nucleus striae terminalis, both of which are provided with sex steroid receptors (Watson and Adkins-Regan 1989 b; Balthazart et al. 1989).
In mammals, several data indicate a role for NPY in the modulation of luteinizing hormone secretion. This action is similar to that of catecholamines (Kalra and Crowley 1984; McDonald et al. 1985) and depends on both the presence and levels of circulating gonadal ster- oids. Moreover, double staining studies have shown

229
NPY fibres contacting LHRH cell bodies in the rat pre- optic area (Danger et al. 1990; Tsuruo et al. 1990). The identification of NPY pericellular terminals in regions where the main populations of LHRH perikarya have previously been observed in quail (lateral septum, LPO, nCPa, anterior preoptic region; Foster et al. 1988) sug- gests a role of this peptide in the control of the luteiniz- ing hormone release.
The presence of NPY fibres and cell bodies within the quail POM, a sexually dimorphic nucleus (Viglietti- Panzica et al. 1986; Panzica et al. 1990a, 1991), mainly involved in the control of male copulatory behaviour (Balthazart and Surlemont 1990; Watson and Adkins- Regan 1989a), is consistent with the idea that NPY may be involved in reproductive processes. The demonstra- tion that, in rodents, centrally injected NPY affects sex- ual behaviour (Clark et al. 1985; Kalra et al. 1988; Pog- gioli et al. 1990) provides more support for this assump- tion.
In conclusion, these morphological data demonstrate that NPY-IR structures are widely distributed within regions involved in the control of a variety of functions, such as visual input, food and water intake, thermoregu- lation, hypophysial secretion, and reproductive behav- iour. These findings suggest that, in birds as in mammals (Danger et al. 1990), NPY plays a role in the regulation of the vegetative, endocrine, and sensory systems.
Acknowledgements. This study was supported by grants from MURST (60% and 40%), CNR (90.02456.CT04), and EEC (SC1- 0230-C).
References
Adrian TE, Allen JM, Bloom SR, Ghatei MA, Rossor MN, Ro- berts GW, Crow TJ, Tatemoto K, Polak JM (1983) Neuropep- tide Y distribution in human brain. Nature 306:584-586
Allen YS, Adrian TE, Allen JM, Tatemoto K, Crow T J, Bloom SR, Polak JM (1983) Neuropeptide Y distribution in the rat brain. Science 221 : 877-879
Allen JM, Novotny J, Martin J, Heinrich G (1987) Molecular struc- ture of mammalian neuropeptide Y: analysis by molecular clon- ing and computer-aided comparison with crystal structure of avian homologue. Proc Natl Acad Sci USA 84:2532-2536
Anderson KD, Reiner A (1990) Distribution and relative abun- dance of neurons in the pigeon forebrain containing somatosta- tin, neuropeptide Y, or both. J Comp Neurol 299:261 282
Balthazart J, Surlemont C (1990) Copulatory behavior is controlled by the sexually dimorphic nucleus of the quail preoptic area. Brain Res Bull 25:7-14
Balthazart J, Gahr M, Surlemont G (1989) Distribution of estrogen receptors in the brain of the Japanese quail: an immunocyto- chemical study. Brain Res 501:205 214
Bl/ihser S, Tobies S (1990) Structural relationship between peptide- producing neurons, and vegetative and sensory systems in the brain of the domestic fowl. In: Elsner N, Roth G (eds) Brain - perception - cognition. Thieme, Stuttgart New York, pp 328
Bons N, Mestre N, Petter A, Danger JM, Pelletier G, Vaudry H (1990) Localization and characterization of neuropeptide Y in the brain of Microeebus murinus (primate, lemurian). J Comp Neurol 298 : 343-361
Caillez D, Danger JM, Andersen AC, Polak JM, Pelletier G, Kawa- mura K, Kikuyama S, Vaudry H (1987) Neuropeptide Y (NPY)-like immunoreactive neurons in the brain and pituitary of the amphibian Rana catesbeiana. Zool Sci 4:123 134
Cassone VM, Moore RY (1988) Retinohypothalamic projection and suprachiasmatic nucleus of the house sparrow, Passer do- mesticus. J Comp Neurol 266:171 182
Ceccatelli S, Cort6s R, H6kfelt T (1991) Effect of reserpine and colchicine on neuropeptide messenger RNA levels in the rat hypothalamic paraventricular nucleus. Mol Brain Res 9 : 57-69
Chan-Palay V, Yasargil G (1986) Immunocytochemistry of human brain tissue with a polyclonal antiserum against neuropeptide Y. Anat Embryol 174:27-33
Chronwall BM, DiMaggio DA, Massari V J, Pickel VM, Rugiero DA, O'Donohue TL (1985) The anatomy of neuropeptide Y in the rat brain. Neuroscience 15 : 115%1181
Clark JT, Kalra PS, Kalra SP (1985) Neuropeptide Y (NPY) stimu- lates feeding but inhibits sexual behavior in rats. Endocrinology 117 : 2435-2442
Colmers WF, Lukowiak K, Pittman QJ (1987) Presynaptic action of neuropeptide Y in area CA1 of the rat hippocampal slice. J Physiol (Lond) 383:285-299
Corder R, Emson PC, Lowry PJ (1984) Purification and character- ization of human neuropeptide Y from adrenal-medullary phaeochromocytoma tissue. Biochem J 219:69%706
Cowan WM, Adamson L, Powell TPS (1961) An experimental study of the avian visual system. J Anat 95:545 563
Cozzi B, Viglietti-Panzica C, Aste N, Panzica GC (1991) The sero- toninergic system in the brain of the Japanese quail. An immu- nohistochemical study. Cell Tissue Res 263:271-284
Danger JM, Guy J, Benyamina M, J~gou S, Leboulenger F, Cot+ J, Tonon MC, Pelletier G, Vaudry H (1985) Localization and identification of neuropeptide Y (NPY)-like immunoreactivity in the frog brain. Peptides 6:1225-1233
Danger JM, Tonon MC, Basille C, Jenks BG, Saint-Pierre S, Mar- tel JC, Fasolo A, Quirion R, Pelletier G, Vaudry H (1990) Neu- ropepfide Y: localization in the central nervous system and neuroendocrine functions. Fundam Clin Pharmacol 4: 307-340
Dawbarn D, Hung SP, Emson PC (1984) Neuropeptide Y: regional distribution, chromatographic characterization and immunocy- tochemical demonstration in post-mortem human brain. Brain Res 296:168 173
De Quidt ME, Emson PC (1986a) Distribution of neuropeptide Y-like immunoreaetivity in the central nervous system. I. Ra- dioimmunoassay and chromatographic characterization. Neu- roscience 18 : 52~543
De Quidt ME, Emson PC (1986b) Distribution of neuropeptide Y-like immunoreactivity in the rat central nervous system. II. Immunocytochemical analysis. Neuroscience 18 : 545-618
Denbow DM, Myers RD (1982) Eating, drinking and temperature responses to intracerebroventricular cholecystokinin in the chick. Peptides 3 : 739 743
Dornan WA, Malsbury CW (1989) Neuropeptides and male sexual behavior. Neurosci Biobehav Rev 13 : 1-15
Ekblad E, Edvinsson L, Wahlestedt C, Uddman R, Hgtkanson R, Sundler F (1984) Neuropeptide Y co-exists and co-operates with noradrenaline in perivascular nerve fibers. Regul Pept 8 : 225-235
Everitt BJ, H6kfelt T, Terenius L, Tatemoto K, Mutt V, Goldstein M (1984) Differential coexistence of neuropeptide Y (NPY)-like immunoreactivity with catecholamines in the central nervous system of the rat. Neuroscience 11 : 443-462
Foster RG, Panzica GC, Parry DM, Viglietti-Panzica C (1988) Immunocytochemical studies on the LHRH system of the Japa- nese quail : influence by photoperiod and aspects of sexual dif- ferentiation. Cell Tissue Res 253:327-335
Fuxe K, Anderssen K, Agnati LF, Eneroth P, Locatelli V, Cavic- cioli L, Masgani F, Tatemoto K, Mutt V (1982) The influence of cholecystokinin peptides and PYY on the amine turnover in discrete hypothalamic dopamine and noradrenaline nerve terminal system and possible relationship to neuroendocrine function. INSERM 110:65-69
Gray TS, Morley JE (1986) Neuropeptide Y: anatomical distribu- tion and possible function in mammalian nervous system. Life Sci 38 : 389-401

230
Kalra SP (1985) Neural circuits involved in the control of LHRH secretion: a model for estrous cycle regulation. J Steroid Bio- chem 23 : 733-742
Kalra SP, Crowley WR (1984) Norepinephrine-like effects of neu- ropeptide Y on LH release in the rat. Life Sci 35:1173-1176
Kalra SP, Clark JT, Sahu A, Dube MG, Kalra PS (1988) Control of feeding and sexual behaviors by neuropeptide Y : physiologi- cal implications. Synapse 2:254-257
Karten HJ, Hodos W, Nauta WJH, Revzen AM (1973) Neural connections of the "visual wulst" of the avian telencephalon. Experimental studies in the pigeon (Columba livia) and owl (Speotyto cunicularia). J Comp Neurol 150 : 253-278
Kimmel JR, Hayden LJ, Pollock HG (1975) Isolation and charac- terization of a new pancreatic polypeptide hormone. J Biol Chem 250 : 9369-9376
Korf H-W, Panzica GC, Viglietti-Panzica C, Oksche A (1988) Pat- tern of peptidergic neurons in the avian brain: clusters - local circuitries - projections. Basic Appl Histochem 32 : 55 75
Kuenzel WJ (1989) Neuroanatomical substrates involved in the control of food intake. Poultry Sci 68 : 926-937
Kuenzel WJ, Masson M (1988) A stereotaxic atlas of the brain of the chick (Gallus domesticus). Johns Hopkins University Press, Baltimore London
Kuenzel WJ, McMurtry J (1988) Neuropeptide Y: brain localiza- tion and central effects on plasma insulin levels in chicks. Phys- iol Behav 44 : 669-678
Kuenzel WJ, Douglass LW, Davison BA (1987) Robust feeding following central administration of neuropeptide Y or peptide YY in chicks, Gallus domesticus. Peptides 8 : 823-828
Larhammar D, Blomquist A, Lundell I (1990) Strong evolutionary conservation of neuropeptide Y between mammals, chicken, goldfish and horned shark. Neuroendocrinology 52 [Suppl 1]:53
L6ger L, Charnay Y, Danger JM, Vaudry H, Pelletier G, Dubois PM, Jouvet M (1987) Mapping of neuropeptide Y-like immuno- reactivity in the feline hypothalamus and hypophysis. J Comp Neurol 255:255-292
Leibowitz SF (1989) Hypothalamic neuropeptide Y and galanin: functional studies of coexistence with monoamines. In: Mutt V (ed) Neuropeptide Y. Raven, New York, pp 262281
Lorenz E, Skofitsch G (1990) Immunohistochemical localization of neuropeptide Y in the central nervous system of the dove (Columba livia). In: Elsner N, Roth G (eds) Brain perception
cognition. Thieme, Stuttgart New York, pp 326 McDonald JK, Lumpkin MD, Samson WK, McCann SM (1985)
Neuropeptide Y affects secretion of luteinizing hormone and growth hormone in ovariectomized rats. Proc Natl Acad Sci USA 82:561 564
Merckaert J, Vandesande F (1990) Study of a direct effect of neu- ropeptide Y (NPY) on the pituitary of the chicken (GalIus do- mesticus). 15th Conference of European Comparative Endo- crinologists, Leuven, p 148 (Abstract)
Noe BD, Milgram SL, Balasubramaniam A, Andrew PC, Calka J, McDonald JK (1989) Localization and characterization of neuropeptide Y-like peptides in the brain and islet organ of the anglerfish (Lophius americanus). Cell Tissue Res 257:303- 311
Panzica GC, Balthazart J, Viglietti-Panzica C (1990a) Anatomical and biochemical studies on the sexually dimorphic preoptic me- dial nucleus of the quail. In : Balthazart J (ed) Hormones, brain and behaviour in vertebrates. 1. Sexual differentiation, neu- roanatomical aspects, neurotransmitters and neuropeptides. Karger, Basel, pp 104-120
Panzica GC, Guglielmone R, Cozzi B, Aste N, Viglietti-Panzica C (1990b) Distribution of catecholamines and serotonin in the brain of the Japanese quail. A histochemical and immunohisto- chemical study. 15th Conference of European Comparative En- docrinologists, Leuven, p 100 (Abstract)
Panzica GC, Viglietti-Panzica C, Sanchez F, Sante P, Balthazart J (1991) Effects of testosterone on a selected neuronal popula-
tion within the preoptic sexually dimorphic nucleus of the Japa- nese quail. J Comp Neurol 303:443-456
Pelletier G, D6sy L, Kerk6rian L, Cot~ J (1984a) Immunocyto- chemical localization of neuropeptide Y (NPY) in the human hypothalamus. Cell Tissue Res 238:203 205
Pelletier G, Guy J, Allen YS, Polak JM (1984b) Electron micro- scopic immunocytochemical localization of neuropeptide Y (NPY) in the rat brain. Neuropeptides 4:319-324
Perroteau I, Danger JM, Biffo S, Pelletier G, Vaudry H, Fasolo A (1988) Distribution and characterization of neuropeptide Y immunoreactivity in the brain of the crested newt. J Comp Neurol 275 : 309-325
Poggioli R, Vergogni AV, Marrama D, Giuliani D, Bertolini A (1990) NPY-induced inhibition of male copulatory activity is a direct behavioural effect. Neuropeptides 16:169 172
Pontet A, Danger JM, Dubourg P, Pelletier G, Vaudry H, Kah O (1989) Distribution and characterization of neuropeptide Y- like immunoreactivity in the brain and pituitary of the goldfish. Cell Tissue Res 255:164-168
Reiner A, Oliver JR (1987) Somatostatin and neuropeptide Y are almost exclusively found in the same neurons in the teleneepha- lon of the turtles. Brain Res 426 : 149 156
Reiner A, Brauth SE, Karten HJ (1984) Evolution of the amniote basal ganglia. Trends Neurosci 7:320-325
Romeis B (1968) Mikroskopische Technik. Oldenbourg, M/.inchen Wien
Sato H, Simon E (1988) Thermal characterization and transmitter analysis of single units in the preoptic and anterior hypothala- mus of conscious ducks. Pflfigers Arch 411 : 34-41
Sawchenko PE, Swanson LW, Grzanna R, Howe PRC, Bloom SR, Polak JM (1985) Colocalization of neuropeptide Y immu- noreactivity in brainstem catecholaminergic neurons that pro- ject to the paraventricular nucleus of the hypothalamus, J Comp Neurol 241:131-153
Smith Y, Parent A, Kerk~rian L, Pelletier G (1985) Distribution of neuropeptide Y immunoreactivity in the basal forebrain and upper brainstem of the squirrel monkey (Saimiri sciureus). J Comp Neurol 236 : 71-79
Tatemoto K (1982) Neuropeptide Y: complete amino acid sequence of the brain peptide. Proc Natl Acad Sci USA 79 : 5485-5489
Tatemoto K, Calquist M, Mutt V (1982) Neuropeptide Y a novel brain peptide with structural similarities to peptide YY and pancreatic polypeptide. Nature 296 : 659-660
Tsuruo Y, Kawano H, Kagotani Y, Hisano S, Daikoku S, Chihara K, Zhang T, Yanaihara N (1990) Morphological evidence for neuronal regulation of luteinizing hormone-releasing hormone- containing neurons by neuropeptide Y in the rat septo-preoptic area. Neurosci Lett 110:261-266
Vallarino M, Danger JM, Fasolo A, Pelletier G, Saint-Pierre S, Vaudry H (1988) Distribution and characterization of neu- ropeptide Y in the fish brain. Brain Res 448 : 6%76
Viglietti-Panzica C, Panzica GC, Fiori MG, Calcagni M, Anselmet- ti GC, Balthazart J (1986) A sexually dimorphic nucleus in the quail preoptic area. Neurosci Lett 64:129-134
Wahlestedt C, Skagerberg G, Ekman R, Heilig M, Sundler F, H5k- anson R (1987) Neuropeptide Y in the area of the hypothalamic paraventricular nucleus activates the pituitary-adrenocortical axis in the rat. Brain Res 417:33-38
Watson JT, Adkins-Regan E (1989a) Activation of sexual behavior by implantation of testosterone propionate and estradiol ben- zoate into the preoptic area of the male Japanese quail (Cotur- nix japonica). Horm Behav 23 : 251-268
Watson JT, Adkins-Regan E (1989b) Neuroanatomical localization of sex steroid-concentrating cells in the Japanese quail (Coturnix japonica): autoradiography with [3H]-testosterone, [3H]-estra- diol, and [3H]-dihydrotestosterone. Neuroendocrinology 49:51 64
Whale P, Albus K (1984) Localization of a NPY-like immunoreac- tivity in the cat's central nervous system. Soc Neurosci Abstr 10:433
Related Documents











