MOLECULAR AND CELLULAR BIOLOGY, Apr. 1993, p. 2504-2514 0270-7306/93/042504-11$02.00/0 Copyright © 1993, American Society for Microbiology Ligand-Dependent Recruitment of the Arnt Coregulator Determines DNA Recognition by the Dioxin Receptor MURRAY WHITELAW, INGEMAR PONGRATZ, ANNA WILHELMSSON, JAN-AKE GUSTAFSSON, AND LORENZ POELLINGER* Department of Medical Nutrition, Karolinska Institutet, Huddinge University Hospital F-60, Novum, 5-141 86 Huddinge, Sweden Received 23 October 1992/Returned for modification 18 December 1992/Accepted 6 January 1993 The intracellular basic region/helix-loop-helix (bHLH) dioxin receptor mediates signal transduction by dioxin (2,3,7,8-tetrachlorodibenzo-p-dioxin) and functions as a ligand-activated DNA binding protein directly interacting with target genes by binding to dioxin response elements. Here we show that the partially purified, ligand-bound receptor alone could not bind target DNA. In contrast, DNA binding by the receptor could be induced by addition of a cytosolic auxiliary activity which functionally and biochemically corresponded to the bHLH factor Arnt. While Arnt exhibited no detectable affinity for the dioxin response element in the absence of the dioxin receptor, it strongly promoted the DNA binding function of the ligand-activated but not the ligand-free receptor forms. Arnt also functionally reconstituted in vitro the DNA binding activity of a mutant, nuclear translocation-deficient dioxin receptor phenotype in cytosolic extracts from a dioxin-resistant hepatoma cell line. Importantly, coimmunoprecipitation experiments showed that Arnt physically interacted in solution with the ligand-activated dioxin receptor but failed to heterodimerize with the ligand-free, hsp9O-associated receptor form. Mutational analysis suggested that the functional interaction between these two factors occurred via the bHLH motif of Arnt. These data suggest that dioxin receptor activity is governed by a complex pattern of combinatorial regulation involving repression by hsp9O and then by ligand-dependent recruitment of the positive coregulator Arnt. The dioxin receptor system also provides the first example of signal-controlled dimerization of bHLH factors. Signal transduction by dioxins (most notably 2,3,7,8- tetrachlorodibenzo-p-dioxin [TCDD]) is mediated by the intracellular dioxin (or aryl hydrocarbon) receptor. The receptor binds dioxin and its planar aromatic congeners in a saturable manner with high affinity (for reviews, see refer- ences 22 and 49). The potent toxicity of dioxins is well established in animal models but is a matter of debate in humans (for a recent review, see reference 19). In animals the toxic effects are typified by thymic wasting and immune suppression, severe epithelial disorders, and tumor promo- tion (for a review, see reference 51). At the molecular level, dioxins are very potent inducers of transcription of a battery of target genes encoding xenobiotic metabolizing enzymes such as cytochrome P-450IA1, glutathione S-transferase Ya, aldehyde dehydrogenase, and quinone oxidoreductase (for a review, see reference 34). In addition, dioxin appears to transcriptionally regulate the expression of the growth mod- ulatory genes for interleukin-1 and plasminogen activator inhibitor-2 (59). The dioxin induction response is mediated by single or multiple copies of dioxin-inducible transcrip- tional control elements (xenobiotic response elements [XREs]) in target promoters (16, 18, 47). In analogy to the current model of action of steroid hormone receptors (for a review, see reference 3), the ligand-activated dioxin receptor appears to transmit the gene induction signal from the cytoplasm to the nucleus, where it interacts with its cognate DNA response element to activate transcription (for re- views, see references 34 and 49). Finally, the endogenous ligand, if any, of the dioxin receptor has not been identified (for a review, see reference 49). The detailed mechanism of signal transduction through the * Corresponding author. dioxin receptor is currently unclear. The ligand-binding form of receptor is a ubiquitous -100-kDa protein which was recently shown to contain a putative basic region/helix-loop- helix (bHLH) motif (8, 15). In contrast, the zinc finger motif (27, 38 [and references therein]) is highly conserved among members of the steroid receptor superfamily. Thus, the dioxin receptor belongs to a distinct class of ligand-activated nuclear receptors. Moreover, the receptor is structurally related to the bHLH factor Arnt which functionally comple- ments C4 mutant hepatoma cells which are nonresponsive to dioxin and express a nuclear translocation-deficient (nt-) dioxin receptor phenotype (8, 15, 30). Arnt has therefore been postulated to govern nuclear translocation of the li- gand-activated dioxin receptor form (30). Interestingly, in analogy to a distinct subgroup of the zinc finger receptors including the glucocorticoid receptor (for a review, see reference 3), the latent (i.e., non-DNA binding) form of dioxin receptor is found complexed with hsp90 (11, 48, 61). Exposure to dioxin in vivo (13, 18, 26, 45, 61) or in vitro treatment with dioxin under high-ionic-strength condi- tions (10, 52) leads to release of hsp90 and a transformed dioxin receptor form with high affinity for XRE sequences. This DNA binding form of the dioxin receptor has a native molecular mass of about 200 kDa (26, 55), indicating that it either represents a homodimer or a heterodimeric complex with a factor of similar molecular mass. The latter possibility is favored by DNA cross-linking studies which have identi- fied two distinct proteins (-100 and -110 kDa) in contact with the XRE target sequence (14). A good candidate for a receptor partner is the -85-kDa structurally related bHLH factor Arnt. In line with this model, Arnt has recently been reported to be a component of the nuclear dioxin receptor complex after treatment of target cells with dioxin in vivo (57). 2504 Vol. 13, No. 4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MOLECULAR AND CELLULAR BIOLOGY, Apr. 1993, p. 2504-25140270-7306/93/042504-11$02.00/0Copyright © 1993, American Society for Microbiology
Ligand-Dependent Recruitment of the Arnt CoregulatorDetermines DNA Recognition by the Dioxin Receptor
MURRAY WHITELAW, INGEMAR PONGRATZ, ANNA WILHELMSSON, JAN-AKE GUSTAFSSON,AND LORENZ POELLINGER*
Department of Medical Nutrition, Karolinska Institutet, Huddinge University Hospital F-60,Novum, 5-141 86 Huddinge, Sweden
Received 23 October 1992/Returned for modification 18 December 1992/Accepted 6 January 1993
The intracellular basic region/helix-loop-helix (bHLH) dioxin receptor mediates signal transduction bydioxin (2,3,7,8-tetrachlorodibenzo-p-dioxin) and functions as a ligand-activated DNA binding protein directlyinteracting with target genes by binding to dioxin response elements. Here we show that the partially purified,ligand-bound receptor alone could not bind target DNA. In contrast, DNA binding by the receptor could beinduced by addition of a cytosolic auxiliary activity which functionally and biochemically corresponded to thebHLH factor Arnt. While Arnt exhibited no detectable affinity for the dioxin response element in the absenceof the dioxin receptor, it strongly promoted the DNA binding function of the ligand-activated but not theligand-free receptor forms. Arnt also functionally reconstituted in vitro the DNA binding activity of a mutant,nuclear translocation-deficient dioxin receptor phenotype in cytosolic extracts from a dioxin-resistant hepatomacell line. Importantly, coimmunoprecipitation experiments showed that Arnt physically interacted in solutionwith the ligand-activated dioxin receptor but failed to heterodimerize with the ligand-free, hsp9O-associatedreceptor form. Mutational analysis suggested that the functional interaction between these two factors occurredvia the bHLH motif of Arnt. These data suggest that dioxin receptor activity is governed by a complex patternof combinatorial regulation involving repression by hsp9O and then by ligand-dependent recruitment of thepositive coregulator Arnt. The dioxin receptor system also provides the first example of signal-controlleddimerization of bHLH factors.
Signal transduction by dioxins (most notably 2,3,7,8-tetrachlorodibenzo-p-dioxin [TCDD]) is mediated by theintracellular dioxin (or aryl hydrocarbon) receptor. Thereceptor binds dioxin and its planar aromatic congeners in a
saturable manner with high affinity (for reviews, see refer-ences 22 and 49). The potent toxicity of dioxins is wellestablished in animal models but is a matter of debate inhumans (for a recent review, see reference 19). In animalsthe toxic effects are typified by thymic wasting and immunesuppression, severe epithelial disorders, and tumor promo-tion (for a review, see reference 51). At the molecular level,dioxins are very potent inducers of transcription of a batteryof target genes encoding xenobiotic metabolizing enzymessuch as cytochrome P-450IA1, glutathione S-transferase Ya,aldehyde dehydrogenase, and quinone oxidoreductase (for a
review, see reference 34). In addition, dioxin appears totranscriptionally regulate the expression of the growth mod-ulatory genes for interleukin-1 and plasminogen activatorinhibitor-2 (59). The dioxin induction response is mediatedby single or multiple copies of dioxin-inducible transcrip-tional control elements (xenobiotic response elements[XREs]) in target promoters (16, 18, 47). In analogy to thecurrent model of action of steroid hormone receptors (for a
review, see reference 3), the ligand-activated dioxin receptorappears to transmit the gene induction signal from thecytoplasm to the nucleus, where it interacts with its cognateDNA response element to activate transcription (for re-
views, see references 34 and 49). Finally, the endogenousligand, if any, of the dioxin receptor has not been identified(for a review, see reference 49).The detailed mechanism of signal transduction through the
* Corresponding author.
dioxin receptor is currently unclear. The ligand-binding formof receptor is a ubiquitous -100-kDa protein which was
recently shown to contain a putative basic region/helix-loop-helix (bHLH) motif (8, 15). In contrast, the zinc finger motif(27, 38 [and references therein]) is highly conserved among
members of the steroid receptor superfamily. Thus, thedioxin receptor belongs to a distinct class of ligand-activatednuclear receptors. Moreover, the receptor is structurallyrelated to the bHLH factor Arnt which functionally comple-ments C4 mutant hepatoma cells which are nonresponsive todioxin and express a nuclear translocation-deficient (nt-)dioxin receptor phenotype (8, 15, 30). Arnt has thereforebeen postulated to govern nuclear translocation of the li-gand-activated dioxin receptor form (30).
Interestingly, in analogy to a distinct subgroup of the zincfinger receptors including the glucocorticoid receptor (for a
review, see reference 3), the latent (i.e., non-DNA binding)form of dioxin receptor is found complexed with hsp90 (11,48, 61). Exposure to dioxin in vivo (13, 18, 26, 45, 61) or invitro treatment with dioxin under high-ionic-strength condi-tions (10, 52) leads to release of hsp90 and a transformeddioxin receptor form with high affinity for XRE sequences.This DNA binding form of the dioxin receptor has a nativemolecular mass of about 200 kDa (26, 55), indicating that iteither represents a homodimer or a heterodimeric complexwith a factor of similar molecular mass. The latter possibilityis favored by DNA cross-linking studies which have identi-fied two distinct proteins (-100 and -110 kDa) in contactwith the XRE target sequence (14). A good candidate for a
receptor partner is the -85-kDa structurally related bHLHfactor Arnt. In line with this model, Arnt has recently beenreported to be a component of the nuclear dioxin receptorcomplex after treatment of target cells with dioxin in vivo(57).
2504
Vol. 13, No. 4

LIGAND-DEPENDENT RECRUITMENT OF THE Arnt COREGULATOR 2505
We have previously shown that the isolated, hsp9O-freeform of dioxin receptor shows constitutive DNA bindingactivity and does not form a stable complex with dioxin (52).Therefore, hsp90 may serve as a cellular chaperone mole-cule, which, in addition to repressing the DNA bindingactivity of the receptor, stabilizes and maintains a ligand-binding configuration of the protein. To further explore themechanism of regulation of dioxin receptor activity, we haveinvestigated the DNA binding properties of the partiallypurified, hsp9O-associated form of dioxin receptor. We showhere that it was not possible to in vitro activate DNA bindingof this receptor form, even though it can bind ligand (52). Incomplementation assays, we identified a distinct 4S-5S ac-tivity which strongly enhanced the binding of the receptor toits responsive element. This activity corresponded by func-tional and biochemical criteria to the Arnt coregulator.Coimmunoprecipitation experiments demonstrated that Arntformed a strong complex with the ligand-activated dioxinreceptor in solution. Moreover, neither Arnt nor the dioxinreceptor showed any detectable affinity for the XRE targetsequence unless the heteromeric complex was formed. ThebHLH region of Arnt was required for functional interactionwith the dioxin receptor. Importantly, however, this inter-action also required the presence of ligand and depended onthe dose and receptor affinity of the employed ligand. Inagreement with these observations, Arnt failed to physicallyassociate with the ligand-free, hsp9O-containing dioxin re-ceptor form. Thus, hsp9o appears to prevent Arnt frominteracting with the receptor. Taken together, activation ofthe dioxin receptor appears to be a multistep process involv-ing ligand-induced release of hsp9O and subsequent interac-tion with the Arnt coregulator prior to binding to DNA targetsequences.
MATERIALS AND METHODS
Recombinant plasmids. The Arnt expression vector pBM5-NEO-M1-1 has previously been described (30). The in vitrotranscription vector pArntAHLH was constructed by sub-cloning of a HindIII fragment containing the full-length Arntcoding region from pBM5-NEO-M1-1 into pGem7Z (Pro-mega), excision of a 390-bp BclI fragment from the Arntcoding region, and subsequent religation of the plasmid. TheArntAHLH coding region was then excised as a ClaI-XbaIfragment and inserted into the corresponding sites in pCMV4(2) to create the expression vector pCMVArntAHLH. TheXRE-driven reporter gene was created by substituting theglucocorticoid response element in pMMTV-hGH2 (1) with asingle, synthetic wild-type or point-mutated XRE element tocreate the reporter constructs pXRE-MMTV-hGH or pXM1-MMTV-hGH, respectively.
Cells, transient transfection, and extract preparation. Thewild-type, dioxin-responsive Hepa lclc7 cell line and themutant, dioxin-resistant nt- C4 cell line derived from it (25)were grown in minimum essential medium as describedpreviously (61). Cells were grown to near confluence in anatmosphere of 6% CO2. Cells were transiently transfectedwith 1 ,ug of the wild-type or mutant Arnt expression vectorand 1 ,ug of the wild-type or point-mutated XRE reportergene, respectively, with lipofectin (Bethesda Research Lab-oratories). Secreted hGH levels were assayed in the cellmedium by a radioimmunoassay (Pharmacia). Cytosolicextracts were prepared by homogenization of untreated cellsin 1 volume of TEG buffer (20 mM Tris-HCl [pH 7.4], 1 mMEDTA, 10% [wt/vol] glycerol, 1 mM dithiothreitol) andcentrifugation at 120,000 x g for 45 min. The resulting
supernatant was taken as the cytosolic fraction and eitherused immediately or frozen in small aliquots at -70°C.
Fractionation of the dioxin receptor and the auxiliary factoron sucrose density gradients. Wild-type or mutant cell cytosol(500 ,ul; about 4 mg of protein per ml of cytosol) was layeredon 10 to 40% (wtlvol) linear sucrose gradients prepared inTEG buffer containing 50 mM NaCl. The gradients werecentrifuged at 300,000 x g to a cumulative centrifugal effectof 1.7 x 1012 rad2/s in a Beckman L8-60 ultracentrifuge.Fractions were collected by gravity flow, starting from thebottom of the gradients. 14C-labeled immunoglobulin G (IgG)(6.6S) and bovine serum albumin (4.4S) were used as exter-nal sedimentation marker proteins.DNA binding assay. The DNA binding activity of the
dioxin receptor was monitored by a gel mobility shift assayperformed essentially as described previously (26, 44). DNAbinding reactions were assembled with the indicated proteinfractions in 10 mMN-2-hydroxyethylpiperazine-N'-2-ethane-sulfonic acid (HEPES; pH 7.9), 5% (vol/vol) glycerol, 0.5mM dithiothreitol, 2.5 mM MgCl2, 1 mM EDTA, 0.08%(wt/vol) Ficoll, and 4 mM spermidine at a final concentrationof 60 mM NaCl in a final volume ranging between 20 and 50pl. A 36-bp 32P-3'-end-labeled, double-stranded oligonucle-otide XRE (10) spanning the dioxin-responsive XRE1 ele-ment of the rat cytochrome P-450IA1 upstream-promoterregion (18) was added to the reactions as a specific probe inthe presence of 1 pug of poly(dI-dC) (Pharmacia) nonspecificcompetitor DNA and incubated for 30 min at 25°C. Boundand free DNAs were electrophoretically separated underconditions described previously (26). The double-strandedoligonucleotide XM1 (10) containing a single-point mutationof the XRE target sequence was used in DNA bindingcompetition experiments.
Expression of Arnt by in vitro translation. Wild-type ormutated Arnt mRNAs were generated from pBM5-NEO-Mi-1 or pArntAHLH using T7 polymerase and used for invitro synthesis of labeled or unlabeled Arnt proteins in rabbitreticulocyte lysates (Promega) in the presence of either[35S]methionine (New England Nuclear) or 20 to 30 ,uMunlabeled methionine under conditions suggested by themanufacturers. For sodium dodecyl sulfate polyacrylamidegel electrophoresis (SDS-PAGE) analysis, prestained and14C-labeled Mr marker proteins were purchased from Bio-Rad and Amersham, respectively.
Noncovalent and covalent labeling of the dioxin receptor.The crude or -9S dioxin receptor from wild-type or mutanthepatoma cells was noncovalently labeled by incubation for2 to 3 h at 25°C with the indicated concentration of nonra-dioactive dioxin or [3H]dioxin (Chemsyn, Lenexa, Kans.;specific activity, 40 Ci/mmol). The affinity ligand 2-azido-3-[ I]2-iodo-7,8-dibromo-dibenzo-p-dioxin ([125 ]dioxin, spe-
cific activity, -2,000 Ci/mmol) was synthesized and purifiedessentially as previously described (50, 52). The crudecytosolic dioxin receptor was covalently labeled by incuba-tion for 1 h at 0 to 4°C with [1251]dioxin and subsequent UVirradiation at 330 nm for 15 min as previously described (48,50). Prior to UV irradiation, the sample was treated withdextran-coated charcoal to remove nonbound ligand.
Dioxin receptor antiserum, immunoblot, and immunopre-cipitation experiments. Peptide corresponding to amino acids12 to 31 of the murine dioxin receptor (8, 15) was synthesizedand coupled to ovalbumin (62), and antisera were preparedfrom rabbits by standard techniques (28). In the immunoblotexperiments, proteins were separated by SDS-PAGE andelectrophoretically transferred to nitrocellulose membranes.The membranes were subsequently incubated with preim-
VOL. 13, 1993

2506 WHITELAW ET AL.
mune or anti-receptor antisera at 1:10 dilutions and stainedby peroxidase-conjugated swine anti-rabbit immunoglobu-lins (Dako). For the immunoprecipitation experiments, cy-tosolic extracts from nt- mutant C4 cells were treated with10 nM TCDD at 25°C for 3 h, in the presence of the proteaseinhibitors aprotonin (5 p,g/ml) and 1 mM phenylmethylsulfo-nyl fluoride. In vitro-translated [35S]methionine-labeled Arnt(5 ,u) was added to 35 VI of untreated or ligand-treated C4cytosol (6 mg of protein per ml) and incubated at 25°C for 20min. Immune or preimmune serum (20 VI) was then addedand gently shaken for 1 h. Immunoprecipitation was carriedout by the addition of 100 [lI of a 50% slurry of proteinA-Sepharose in PEG buffer (20 mM sodium phosphate [pH7.2], 1 mM EDTA, 10% [wt/vol] glycerol) containing 150 mMNaCl, 1% Triton X-100, and 1 mM dithiothreitol. After beingshaken for 30 min, the resin was washed four times with thesame buffer and the immunoprecipitated proteins were sep-arated through an SDS-7.5% polyacrylamide gel. For fluo-rography, gels were fixed in 20% methanol-10% acetic acid,immersed in Amplify (Amersham) for 30 min, dried, andexposed to film.
Safety precautions. In experiments involving the use ofdioxin, special handling procedures were employed (61 andreferences therein), and contaminated materials were dis-posed of by high-temperature incineration.
AAntibody: aDR
Protein: I C)
Kd: ¢ E
205 -
116-
80 -
B
kD:117 -80 -50- No
M 1
PIS--i
0-CC)R 0
3. E Kd:- 205
- R -116
- 80
2 3 4
6.6 S 4.5 S
- Rec
10SDG FRACTIONS
15 20 0X0
C
Protein: c) 9 S FractionI
Dioxin: - t -
- R
RESULTS
Failure to activate the isolated, hsp9O-associated dioxinreceptor form to a DNA binding species by ligand treatment invitro. In an effort to in vitro reconstitute the process ofactivation of the latent dioxin receptor to a DNA bindingform, we partially purified the -9S (63), ligand-free form ofreceptor by fractionation on sucrose density gradients of acytosolic extract from untreated Hepa lclc7 hepatoma cells.This form of receptor is tightly associated with hsp90 (11, 48,61) and shows dioxin-binding activity in vitro (52). The -9Sreceptor form was identified by immunoblot analysis of theindividual sucrose gradient fractions. To this end, we usedan anti-receptor antiserum, the specificity of which is shownin Fig. 1A. The -95-kDa receptor was specifically recog-nized by the antiserum but not by preimmune serum follow-ing fractionation of a crude cytosolic extract from Hepalclc7 cells by SDS-PAGE (Fig. 1A, compare lanes 1 and 3).A similar immunoblot analysis of Hepa lclc7 cell cytosolfollowing fractionation on a sucrose gradient demonstratedthat the ligand-free dioxin receptor was recovered as adistinct symmetric peak in the 9S region of the gradient (Fig.1B).
In cytosolic extracts from a number of cells, it is possibleto activate the DNA binding function of the dioxin receptorby in vitro treatment with ligand and salt at moderatetemperatures (10, 18, 44, 52). Thus, in gel mobility shiftassays the ligand-free dioxin receptor in crude cytosol fromuntreated Hepa lclc7 cells is inactive and shows no detect-able affinity for the XRE target sequence, whereas it isactivated to maximal levels of DNA binding activity byexposure to 5 to 10 nM dioxin in vitro (10) (Fig. 1C, comparelanes 1 and 2). Remarkably, however, it was not possible toconvert the isolated 9S dioxin receptor to a DNA bindingspecies by ligand treatment in vitro (Fig. 1C, compare lanes2 to 7) by the protocol which readily activated the receptor incrude cytosol. Even in the presence of very high concentra-tions (100 nM) of dioxin, no DNA binding activity wasdetected in the 9S fraction (Fig. 1C, lane 7). Although this
S.,,.. - F1 2 3 4 5 6 7
FIG. 1. Failure to activate the -9S dioxin receptor form byligand treatment in vitro. (A) Immunoblot analysis of dioxin recep-tor. Crude cytosolic extracts (-100 ,ug of protein) from wild-type(wt Hepa; lanes 1 and 3) or nt- mutant C4 (mut C4; lanes 2 and 4)hepatoma cells were fractionated on an SDS-7.5% polyacrylamidegel and analyzed on immunoblots. Note that the anti-receptorantiserum (otDR; lanes 1 and 2) recognized a single band (indicatedby R) in both wild-type and mutant cell extracts, whereas no stainingof the filters was obtained with preimmune serum (PIS; lanes 3 and4). (B) Immunoblot analysis of ligand-free dioxin receptor followingfractionation on a sucrose density gradient (SDG). A cytosolicextract (500 ,ul; -4 mg of protein per ml) from untreated wild-typehepatoma cells was fractionated on a 10 to 40% (wt/vol) sucrosegradient, and the individual fractions were analyzed by immunoblotanalysis as described above with the anti-receptor antiserum. Theposition of the receptor (Rec) is indicated. On top of the blots areindicated the position of sedimentation marker proteins (albumin,4.5S; immunoglobulin G, 6.6S) run on separate gradients. (C) DNAbinding analysis. Ligand-free cytosol from wild-type hepatoma cellswas fractionated on sucrose gradients as described above. Thereceptor sedimenting in the 9S fraction was incubated in the absence(lane 3) or presence of increasing concentrations of dioxin (0.1 to 100nM; lanes 4 to 7). As a control, the crude input cytosolic extract wastreated in the absence (lane 1) or presence (lane 2) of 5 nM dioxin.DNA binding activities were analyzed by gel mobility shift analysiswith a 32P-labeled XRE oligonucleotide probe. The relative mobili-ties of the receptor-dependent XRE complex (R) and the free probe(F) are indicated.
form of receptor avidly binds ligand (52) (data not shown),ligand binding alone is not sufficient for activation.We conclude from these experiments that the partially
purified dioxin receptor showed very low, if any, intrinsicaffinity for DNA. To bind efficiently to DNA, the receptor
MOL. CELL. BIOL.
1
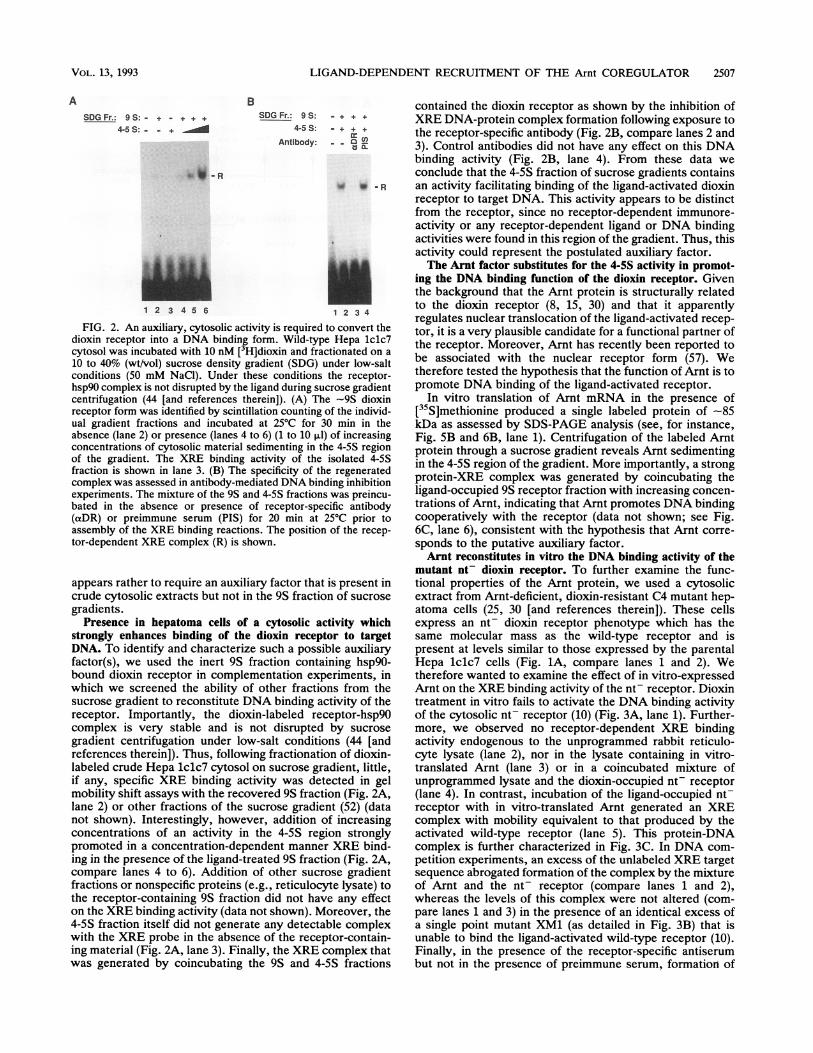
LIGAND-DEPENDENT RECRUITMENT OF THE Arnt COREGULATOR 2507
BSDGFr.: 9S:- + - + + +
4-5S: +
9 S:4-5 S:
Antibody:
I R
1 2 -1 4 5 fi
FIG. 2. An auxiliary, cytosolic activity is required tdioxin receptor into a DNA binding form. Wild-typecytosol was incubated with 10 nM [3H]dioxin and frac10 to 40% (wt/vol) sucrose density gradient (SDG) u
conditions (50 mM NaCI). Under these conditionshsp90 complex is not disrupted by the ligand during succentrifugation (44 [and references therein]). (A) Thereceptor form was identified by scintillation counting cual gradient fractions and incubated at 25°C for 3(absence (lane 2) or presence (lanes 4 to 6) (1 to 10 ,ul)concentrations of cytosolic material sedimenting in thof the gradient. The XRE binding activity of thefraction is shown in lane 3. (B) The specificity of thecomplex was assessed in antibody-mediated DNA bindexperiments. The mixture of the 9S and 4-5S fractionsbated in the absence or presence of receptor-spec(aDR) or preimmune serum (PIS) for 20 min at 2assembly of the XRE binding reactions. The positiontor-dependent XRE complex (R) is shown.
appears rather to require an auxiliary factor thatcrude cytosolic extracts but not in the 9S fractioigradients.
Presence in hepatoma cells of a cytosolic ac
strongly enhances binding of the dioxin receptDNA. To identify and characterize such a possitfactor(s), we used the inert 9S fraction contaibound dioxin receptor in complementation exp(which we screened the ability of other fractioisucrose gradient to reconstitute DNA binding ac
receptor. Importantly, the dioxin-labeled rec
complex is very stable and is not disruptedgradient centrifugation under low-salt conditioreferences therein]). Thus, following fractionaticlabeled crude Hepa lclc7 cytosol on sucrose gra
if any, specific XRE binding activity was detmobility shift assays with the recovered 9S fractiilane 2) or other fractions of the sucrose gradiernot shown). Interestingly, however, addition o
concentrations of an activity in the 4-5S regipromoted in a concentration-dependent manner
ing in the presence of the ligand-treated 9S fracticompare lanes 4 to 6). Addition of other sucrc
fractions or nonspecific proteins (e.g., reticulocythe receptor-containing 9S fraction did not havon the XRE binding activity (data not shown). M44-5S fraction itself did not generate any detectalwith the XRE probe in the absence of the recep
ing material (Fig. 2A, lane 3). Finally, the XRE c
was generated by coincubating the 9S and 4-'
contained the dioxin receptor as shown by the inhibition of+ XRE DNA-protein complex formation following exposure to
- + the receptor-specific antibody (Fig. 2B, compare lanes 2 and- FL 3). Control antibodies did not have any effect on this DNA
binding activity (Fig. 2B, lane 4). From these data we
conclude that the 4-5S fraction of sucrose gradients contains--R an activity facilitating binding of the ligand-activated dioxin
receptor to target DNA. This activity appears to be distinctfrom the receptor, since no receptor-dependent immunore-activity or any receptor-dependent ligand or DNA bindingactivities were found in this region of the gradient. Thus, thisactivity could represent the postulated auxiliary factor.The Arnt factor substitutes for the 4-5S activity in promot-
ing the DNA binding function of the dioxin receptor. Giventhe background that the Arnt protein is structurally related
1 2 3 4 to the dioxin receptor (8, 15, 30) and that it apparentlyregulates nuclear translocation of the ligand-activated recep-
toconvert
te tor, it is a very plausible candidate for a functional partner ofHepa lcoc7 the receptor. Moreover, Arnt has recently been reported torl-al be associated with the nuclear receptor form (57). We
the low-salt therefore tested the hypothesis that the function of Arnt is to
the gradient promote DNA binding of the ligand-activated receptor.-9S dioxin In vitro translation of Arnt mRNA in the presence of
A the individ- [35S]methionine produced a single labeled protein of -85O min in the kDa as assessed by SDS-PAGE analysis (see, for instance,of increasing Fig. SB and 6B, lane 1). Centrifugation of the labeled Arnte 4-5S region protein through a sucrose gradient reveals Arnt sedimenting
elregenerated in the 4-5S region of the gradient. More importantly, a stronging inhibition protein-XRE complex was generated by coincubating the,nwas preincu- ligand-occupied 9S receptor fraction with increasing concen-
ific antibody trations of Arnt, indicating that Arnt promotes DNA bindingZ5YC prior to cooperatively with the receptor (data not shown; see Fig.of the recep- 6C, lane 6), consistent with the hypothesis that Arnt corre-
sponds to the putative auxiliary factor.Arnt reconstitutes in vitro the DNA binding activity of the
mutant nt- dioxin receptor. To further examine the func-is present in tional properties of the Arnt protein, we used a cytosolicn of sucrose extract from Arnt-deficient, dioxin-resistant C4 mutant hep-
atoma cells (25, 30 [and references therein]). These cellstivity which express an nt- dioxin receptor phenotype which has theor to target same molecular mass as the wild-type receptor and isble auxiliary present at levels similar to those expressed by the parentalining hsp9O- Hepa 1c1c7 cells (Fig. 1A, compare lanes 1 and 2). Weeriments, in therefore wanted to examine the effect of in vitro-expressedns from the Arnt on the XRE binding activity of the nt- receptor. Dioxintivity of the treatment in vitro fails to activate the DNA binding activityeptor-hsp90 of the cytosolic nt- receptor (10) (Fig. 3A, lane 1). Further-by sucrose more, we observed no receptor-dependent XRE bindingIns (44 [and activity endogenous to the unprogrammed rabbit reticulo-rn of dioxin- cyte lysate (lane 2), nor in the lysate containing in vitro-dient, little, translated Arnt (lane 3) or in a coincubated mixture ofected in gel unprogrammed lysate and the dioxin-occupied nt- receptoron (Fig. 2A, (lane 4). In contrast, incubation of the ligand-occupied nt-nt (52) (data receptor with in vitro-translated Arnt generated an XREf increasing complex with mobility equivalent to that produced by the[on strongly activated wild-type receptor (lane 5). This protein-DNAXRE bind- complex is further characterized in Fig. 3C. In DNA com-
on (Fig. 2A, petition experiments, an excess of the unlabeled XRE target:se gradient sequence abrogated formation of the complex by the mixturete lysate) to of Arnt and the nt- receptor (compare lanes 1 and 2),e any effect whereas the levels of this complex were not altered (com-oreover, the pare lanes 1 and 3) in the presence of an identical excess ofble complex a single point mutant XM1 (as detailed in Fig. 3B) that isitor-contain- unable to bind the ligand-activated wild-type receptor (10).omplex that Finally, in the presence of the receptor-specific antiserumSS fractions but not in the presence of preimmune serum, formation of
A
VOL. 13, 1993

2508 WHITELAW ET AL.
B XRE: 5'-CCAGGCTCTTCTCACGCAACI C-;GGGC-3
XM1:
C Protein: C4 + Arrit1JLLS
Competitor: x x
Antibody:
AXRE probe
+ - Ligand + - Arnt + n.s. DNA
StopDNA-bindingI I 2 _ r.. 20', I.t _ assy
2 h, r.t. 20O', r.t. 20O', r. t.I s a
,e too-v A.,* - R
bU - F
1 2 3 4 5
1 2 3 4 5 6
B Protein: wt C4 C4 + Arnt
Ligand: + - C-
-".1 . .ot s "''
. `..- F
FIG. 3. In vitro reconstitution of the DNA binding activity of thent- dioxin receptor phenotype by Arnt. (A) The nt- dioxin receptorin C4 mutant hepatoma cell cytosol (-4 mg of protein per ml) wastreated with 5 nM dioxin. Gel mobility shift assays were performedwith the 32P-labeled XRE probe in the presence of 5 p.l of theligand-treated nt- receptor alone (lane 1), 5 ,ul of unprogrammedreticulocyte lysate (R.L.; lane 2), 5 p.l of reticulocyte lysate con-
taining in vitro-translated Arnt (Arnt; lane 3), or reaction mixturescontaining the ligand-treated nt- receptor that had been coincubatedfor 20 min at 25°C with either 5 ,ul of unprogrammed reticulocytelysate (lane 4) or 5 p.l in vitro-translated Arnt (lane 5). (B) Sequencesof the wild-type XRE motif and a single-point mutant (XM1)deficient in binding the ligand-activated wild-type receptor. (C) Thespecificity of the reconstituted XRE complex was assessed by DNAcompetition and antibody experiments. The ligand-treated nt- re-ceptor was coincubated with in vitro-translated Arnt as describedabove in the absence (lanes 1 and 4) or in the presence of an excessof either the unlabeled wild-type XRE (lane 2) or XM1 mutant (lane3) oligonucleotides or receptor-specific (oaDR; lane 5) or preimmune(PIS; lane 6) antibodies. XRE binding activities were monitored bygel mobility shift analysis. The positions of the reconstituted,receptor-dependent complex (R) and the free probe (F) are indi-cated. As shown in the competition experiments, the protein-XREcomplexes of faster mobility than the receptor complex are theresult of nonspecific protein-DNA interactions.
this complex was disrupted (Fig. 3C, compare lanes 4 to 6),indicating that the nt- receptor was a component of the XREcomplex following addition of Arnt. Taken together, theseresults show that Arnt recruited the receptor into the XREbinding complex. Moreover, these experiments demonstratethat, individually, the receptor and Arnt proteins bind verypoorly, if at all, on their own to the DNA target sequence.The ligand regulates the functional interaction between the
dioxin receptor and Arnt. We next explored the cell-freereconstitution assay to investigate whether the receptorligand influenced the effect of Arnt on the DNA bindingactivity of the nt- dioxin receptor. The scheme for activa-tion of the receptor is detailed in Fig. 4A: incubation of theC4 cytosolic extract with ligand proceeded for 2 h at roomtemperature; Arnt was added and further incubated beforestandard gel mobility shift analysis. As expected, treatmentof the cytosolic nt- receptor with increasing concentrations(1 to 10 nM) of dioxin failed to induce an XRE complex in theabsence of Arnt (Fig. 4B, compare lanes 3 to 5), whileaddition of Arnt to the ligand-occupied nt- receptor pro-duced the receptor-dependent XRE complex which, in turn,increased in intensity at the higher concentration of dioxin(compare lanes 7 and 8). Moreover, the receptor-XRE
1 2 3 4 5 6 7 8 9
FIG. 4. Requirement of ligand for conversion of the dioxinreceptor into a DNA binding form by the Arnt coregulator. (A)Schematic diagram of the experimental protocol for ligand-depen-dent in vitro reconstitution of the DNA binding activity of the ntVmutant dioxin receptor. (B) Cytosol from mutant C4 cells (-6 mg ofprotein per ml) was treated with no dioxin (lanes 3 and 6) or with 1(lanes 4 and 7) or 10 (lanes 5 and 8) nM dioxin or with 500 nM,B-naphthoflavone (lane 9) and further incubated in the absence(lanes 3 to 5) or presence (lanes 6 to 9) of 5 p.l of reticulocyte lysatecontaining in vitro-translated Arnt. As a control, wild-type (wt)hepatoma cell cytosol (-6 mg of protein per ml) was treated with 10nM dioxin (lane 2). XRE binding activities were monitored by thegel mobility shift assay. The position of the receptor-dependentcomplex is indicated.
complex was also generated in the presence of a highconcentration (500 nM) of the weaker dioxin receptor ago-nist ,B-naphthoflavone. This complex was less intense thanthe corresponding dioxin-induced complex (compare lanes 8and 9), consistent with the about 50- to 100-fold-loweraffinity of 3-naphthoflavone for the dioxin receptor in vitro(20). For reference, the cytosolic, wild-type dioxin receptorwas in vitro activated by exposure to 10 nM dioxin. Inter-estingly, the resulting XRE complex did not only comigratewith the corresponding complex produced by Arnt and thent- receptor (compare lane 2 with lanes 7 to 9) but showedan intensity that was only slightly stronger than that gener-ated in the reconstituted system in the presence of anidentical concentration (10 nM) of dioxin (compare lanes 2and 8). Against the background that both the wild-type andC4 cytosolic extracts were matched with regard to proteinconcentration and that the receptor is expressed at similarlevels in both the wild-type and nt- mutant C4 cells (Fig.1A), these data clearly indicate that Arnt is a potent regula-tor which, in the presence of maximal ligand stimulation,appears to be both necessary and sufficient to restore theDNA binding activity of the nt- receptor to almost wild-typelevels.
It is important that the receptor-dependent XRE complexwas not detected if Arnt was incubated with the ligand-freent- receptor (Fig. 4B, compare lanes 6 to 8). Thus, func-tional interaction between Arnt and the dioxin receptor wasa strictly ligand-dependent event. In the case of the wild-type receptor, we have previously shown that the intensity
Ax
.0
+7 * c+ +
cc 0 0r
- R
MOL. CELL. BIOL.

LIGAND-DEPENDENT RECRUITMENT OF THE Arnt COREGULATOR 2509
BLabeled Arnt:
Ligand: +TCDD -TCDD
Antibody: - o aFLkD: 200- a
98- @
69 - t
46- a
M 1 2 3 4 5
FIG. 5. The ligand-activated dioxin receptor and the Arnt coreg-ulator are tightly associated in solution. (A) Immunoprecipitation ofthe dioxin receptor. The nt- dioxin receptor in cytosol (-2 mg ofprotein per ml) from C4 mutant hepatoma cells was covalentlylabeled with [1 I]dioxin in the absence or presence of the unlabeledcompetitor ligand prior to immunoprecipitation with receptor-spe-cific (aDR) antibodies or preimmune serum (PIS), as indicated.Input material (lane 1) or the immunoprecipitated complexes wereseparated on a 7.5% SDS-polyacrylamide gel and analyzed byautoradiography. The position of the receptor (R) is indicated. (B)Coimmunoprecipitation experiments. Arnt was labeled by in vitrotranslation in the presence of [35S]methionine and incubated with 10
of a cytosolic extract (-6 mg of protein per ml) from C4 mutanthepatoma cells that had been pretreated in the absence (lanes 4 and5) or presence (lanes 2 and 3) of 20 nM dioxin. These reactionmixtures were immunoprecipitated with the receptor-specific anti-bodies (aDR) or preimmune serum (PIS) and analyzed by SDS-PAGE and subsequent fluorography. As a reference, the labeled,input Arnt material was also analyzed on the gel (lane 1). Themolecular mass marker lanes (M) are indicated.
of the receptor-XRE complex is directly proportional to thedose and receptor affinity of the ligand used (10). Employingin vitro-translated Arnt in artificial conversion of the nt-receptor to a DNA binding species therefore seems tosuccessfully and faithfully mimic the dioxin activation pro-cess. Finally, it is noteworthy that Arnt fails to promote theXRE binding activity of the ligand-free nt- dioxin receptor.Since the ligand-free dioxin receptor is characterized by astable heteromeric complex with hsp90 (52 [and referencestherein]), it is interesting to consider that hsp90 may preventArnt from functionally interacting with the receptor by stericinterference or by simply masking a possible dimerizationinterface.Arnt forms a stable physical complex with the ligand-
activated dioxin receptor in solution. The results of thereconstitution experiments suggest that the positive effect ofArnt on the XRE binding activity of ligand-occupied recep-tor may be due to direct interactions between these twoproteins. To investigate this possibility, we employed thereceptor-specific polyclonal antibodies in immunoprecipita-tion assays. In control experiments the nt- dioxin receptorpresent in crude cytosol from C4 mutant hepatoma cells wascovalently labeled with the photoaffinity ligand [125I]dioxin.By this procedure the receptor was visualized as an -95-kDaprotein by SDS-PAGE and subsequent autoradiography(Fig. SA, lane 1), in perfect agreement with the molecularmass determination of the nt- receptor by immunoblotanalysis (Fig. 1A). Thus, these two independent assaysconfirmed that the nt- receptor shows wild-type propertieswith regard to ligand-binding activity and molecular mass.Immunoprecipitation of the covalently labeled material withthe receptor-specific antibodies and protein A-Sepharoseproduced the labeled -95-kDa ligand-receptor complex (Fig.SA, lane 2). This complex was not recovered by the recep-
tor-specific antibodies, however, if the labeling reaction ofthe receptor was performed in the presence of an excess ofthe nonradioactive high-affinity ligand 2,3,7,8-tetrachloro-dibenzofuran (lane 3), nor was it precipitated by the preim-mune antibodies (lane 4). Subjecting either the crude inputC4 cytosol or the immunoprecipitate produced by the recep-tor-specific antibodies to SDS-PAGE, followed by transferto a filter and subsequent immunodetection, provided asingle -95-kDa band. No receptor band was seen by thistechnique when preimmune serum was used to immunopre-cipitate crude C4 cytosol (data not shown).To examine whether Arnt can physically interact with the
ligand-activated dioxin receptor in the absence of DNA,35S-radiolabeled Arnt was synthesized by in vitro translationand incubated with the ligand-activated, nonradioactive nt-dioxin receptor. Immunoprecipitation of the receptor by thereceptor antibodies revealed that the labeled -85-kDa Arntprotein coprecipitated with the receptor (Fig. SB, lane 2).Moreover, the labeled Arnt protein was not detected follow-ing immunoprecipitation with control antibodies (lane 3),indicating that Arnt formed a strong complex with theligand-activated receptor independent of the XRE sequence.To investigate the role of the receptor ligand in this process,we performed similar coimmunoprecipitation experimentsemploying the ligand-free nt- receptor and the radiolabeledArnt protein. Little or no coimmunoprecipitation of Arntwas observed under these conditions (Fig. SB, lanes 4 and5), strongly arguing that physical association between Arntand the receptor is a ligand-dependent event and that thedimerization interface may be masked in the latent receptorform. A strict requirement of ligand for Arnt-receptor inter-action was also observed in a precipitation assay employinga biotinylated XRE probe and streptavidin-agarose (data notshown). Upon addition of the ligand-occupied nt- receptor,it was also possible to specifically shift the sedimentationposition of the labeled Arnt protein to the bottom fractions ofa sucrose gradient with the receptor-specific receptor anti-bodies (data not shown). In conclusion, the results obtainedby these different experimental approaches clearly indicatethat the Arnt and dioxin receptor proteins physically asso-ciate with each other in solution, resulting in conversion ofthe receptor to its DNA binding form. The data furtherstrongly suggest that this interaction is ligand dependent.The bHLH region of Arnt is required for functional inter-
action with the dioxin receptor. To study the role of thebHLH motif in Arnt-dioxin receptor interaction we con-structed the Arnt deletion mutant ArntAHLH. As shownschematically in Fig. 6A, the deletion encompasses theentire 56-amino-acid bHLH consensus motif (43) of Arnt.Upon in vitro translation of the deletion mutant mRNA, a[3 S]methionine-labeled protein of -75 kDa versus the -85-kDa wild-type protein was detected (Fig. 6B, compare lanes1 and 2), in accordance with the calculated molecular massof the deletion mutant. DNA binding reconstitution experi-ments using either the ligand-occupied nt- receptor or thepartially purified, ligand-occupied -9S form of wild-typereceptor demonstrated that deletion of the bHLH domain-containing region of Arnt led to a complete loss of XREbinding (Fig. 6C), indicating that this region of Arnt isrequired for binding of the receptor to an isolated XREelement. Thus, these results support the notion that thebHLH motif constitutes the dimerization interface betweenthese two regulatory factors.
In transient transfection experiments with an XRE-con-taining pXRE-MMTV-hGH reporter gene, we observedstrong activation in wild-type Hepa 1c1c7 cells in the pres-
ALabeled Dioxin Receptor:
0 TAntibody: - n X.Excess TCDF:. - - + -
kD: 200-
- R98- -M
69 -
46 -
30 -
M 1 2 3 4
VOL. 13, 1993
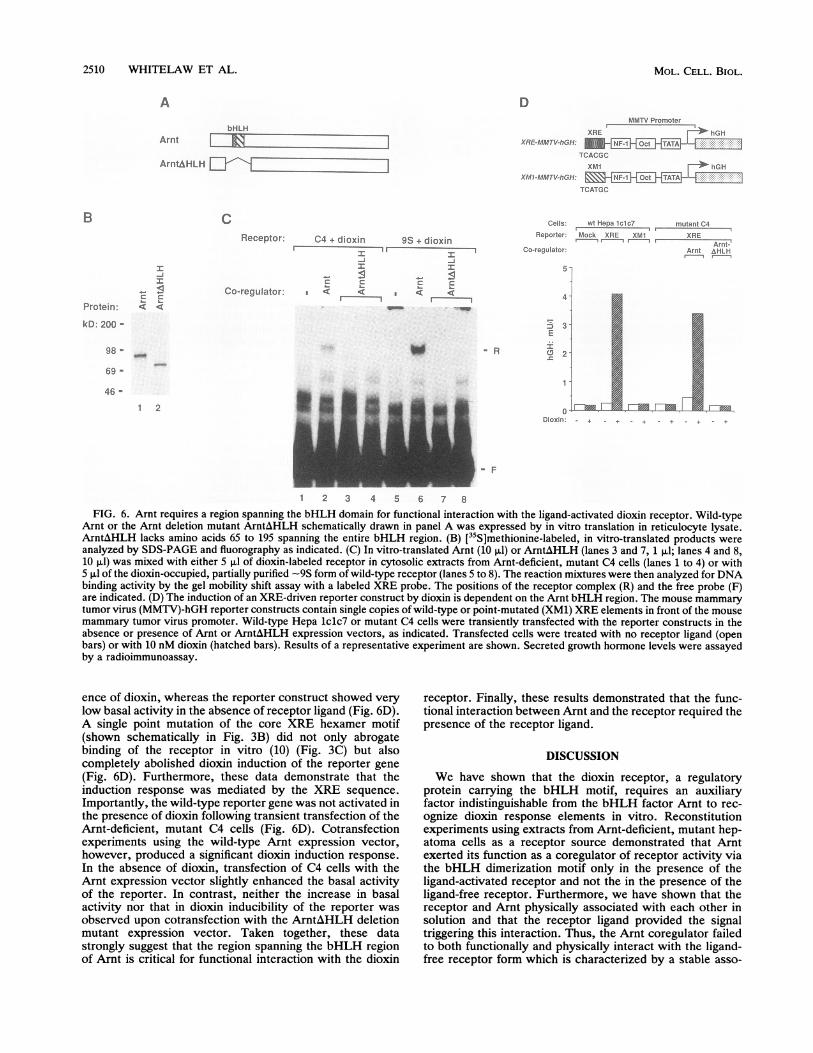
2510 WHITELAW ET AL.
A
bH LH1Arnt L E
ArntAHLHEI<'{
B
I-J
- SC C
Protein: c: <
kD: 200 -
98 -
69 -
46 -
1 2
DMMTV Prornoter
XRE I -3> hGHfXRE-MMTV-hGHI-::
TCACGCXM1 l*R hGH
XM1-MMTV-hGH: c T
TCATGC
CReceptor; C4 + dioxin 9S + dioxin
r~~~r-- 1
I =g9 - SC C
Co-regulator: <: <r, : ci
V
Cells: wt Hepa 1c1c7 mutant C4Reporter: Mock XRE XM1 XRE
Aro1 t-'Co-regulator: Arrnt AHLH
r-
5 -
- R
U_
- F
1 2 3 4 5 6 7 8FIG. 6. Arnt requires a region spanning the bHLH domain for functional interaction with the ligand-activated dioxin receptor. Wild-type
Arnt or the Arnt deletion mutant ArntAHLH schematically drawn in panel A was expressed by in vitro translation in reticulocyte lysate.ArntAHLH lacks amino acids 65 to 195 spanning the entire bHLH region. (B) [35S]methionine-labeled, in vitro-translated products wereanalyzed by SDS-PAGE and fluorography as indicated. (C) In vitro-translated Arnt (10 ,ul) or ArntAHLH (lanes 3 and 7, 1 ,ul; lanes 4 and 8,10 pul) was mixed with either 5 p,l of dioxin-labeled receptor in cytosolic extracts from Arnt-deficient, mutant C4 cells (lanes 1 to 4) or with5 p,l of the dioxin-occupied, partially purified -9S form of wild-type receptor (lanes 5 to 8). The reaction mixtures were then analyzed for DNAbinding activity by the gel mobility shift assay with a labeled XRE probe. The positions of the receptor complex (R) and the free probe (F)are indicated. (D) The induction of an XRE-driven reporter construct by dioxin is dependent on the Arnt bHLH region. The mouse mammarytumor virus (MMTV)-hGH reporter constructs contain single copies of wild-type or point-mutated (XM1) XRE elements in front of the mousemammary tumor virus promoter. Wild-type Hepa lclc7 or mutant C4 cells were transiently transfected with the reporter constructs in theabsence or presence of Arnt or ArntAHLH expression vectors, as indicated. Transfected cells were treated with no receptor ligand (openbars) or with 10 nM dioxin (hatched bars). Results of a representative experiment are shown. Secreted growth hormone levels were assayedby a radioimmunoassay.
ence of dioxin, whereas the reporter construct showed verylow basal activity in the absence of receptor ligand (Fig. 6D).A single point mutation of the core XRE hexamer motif(shown schematically in Fig. 3B) did not only abrogatebinding of the receptor in vitro (10) (Fig. 3C) but alsocompletely abolished dioxin induction of the reporter gene(Fig. 6D). Furthermore, these data demonstrate that theinduction response was mediated by the XRE sequence.Importantly, the wild-type reporter gene was not activated inthe presence of dioxin following transient transfection of theArnt-deficient, mutant C4 cells (Fig. 6D). Cotransfectionexperiments using the wild-type Arnt expression vector,however, produced a significant dioxin induction response.In the absence of dioxin, transfection of C4 cells with theArnt expression vector slightly enhanced the basal activityof the reporter. In contrast, neither the increase in basalactivity nor that in dioxin inducibility of the reporter wasobserved upon cotransfection with the ArntAHLH deletionmutant expression vector. Taken together, these datastrongly suggest that the region spanning the bHLH regionof Arnt is critical for functional interaction with the dioxin
receptor. Finally, these results demonstrated that the func-tional interaction between Arnt and the receptor required thepresence of the receptor ligand.
DISCUSSION
We have shown that the dioxin receptor, a regulatoryprotein carrying the bHLH motif, requires an auxiliaryfactor indistinguishable from the bHLH factor Arnt to rec-ognize dioxin response elements in vitro. Reconstitutionexperiments using extracts from Arnt-deficient, mutant hep-atoma cells as a receptor source demonstrated that Arntexerted its function as a coregulator of receptor activity viathe bHLH dimerization motif only in the presence of theligand-activated receptor and not the in the presence of theligand-free receptor. Furthermore, we have shown that thereceptor and Arnt physically associated with each other insolution and that the receptor ligand provided the signaltriggering this interaction. Thus, the Arnt coregulator failedto both functionally and physically interact with the ligand-free receptor form which is characterized by a stable asso-
MOL. CELL. BIOL.

LIGAND-DEPENDENT RECRUITMENT OF THE Arnt COREGULATOR 2511
ciation with hsp9O. Therefore, this model provides the firstexample of signal-regulated dimerization of bHLH factors.
Regulation of dioxin receptor function by Arnt. AlthoughArnt has been implicated in the regulation of nuclear trans-location of the dioxin receptor (30), the mechanism by whichit modulates dioxin receptor function has hitherto beenunclear. The present experiments demonstrate that both theligand-activated dioxin receptor and Arnt individually exhib-ited poor, if any, intrinsic XRE binding activity. The forma-tion of a heteromeric protein complex in vitro between thesetwo factors, however, dramatically increased the bindingaffinity of the complex for the XRE target sequence. On thebasis of these criteria, Arnt is not only a physical partner ofthe receptor but also functions as a critical coregulatorprotein serving to target the receptor to its DNA responseelement.Given the background that the nuclear dioxin receptor has
a native molecular mass of approximately 200 kDa (26, 55),the Arnt-receptor complex most probably represents a het-erodimer. It is formally possible, however, that this complexmay harbor additional, low-molecular-weight components.Once the XRE binding heteromeric complex has beenformed, it appears that both the receptor and Arnt are able tointeract directly with the XRE target sequence, since UVDNA cross-linking experiments have indicated the specificbinding of both an -110 and -100-kDa protein, respectively,to an XRE probe following dioxin activation (14). Interest-ingly, however, the -100-kDa component (presumablyArnt) is only detected at late times and at very high energiesofUV irradiation (14, 61, 62). These data suggest that, withinthe ligand-activated, heteromeric complex, Arnt may exhibita significantly lower affinity than the receptor protein for theXRE target sequence.
Deletion experiments indicated that the region containingthe bHLH region of Arnt was critical for XRE binding and/orfunctional interaction with the dioxin receptor. Among pro-teins in the growing family of factors bearing the bHLHmotif, bHLH-mediated heterodimerization processes appearto constitute a critical mechanism of regulation of the DNAbinding activity of these factors. For instance, c-Myc (whichcontains a bHLH region contiguous with a leucine zippermotif) has to heterodimerize with the structurally relatedMax factor to efficiently bind E box (CACGTG)-relatedDNA sequences (5, 6, 31, 54). In similar fashion, the bHLHprotein E12 appears to be a functional and physical partnerprotein of the bHLH myogenic factor MyoD in the activa-tion of muscle-specific gene expression. While the E12 factoritself shows low affinity for E box sequences, the bindingactivity of the heterodimeric complex is greatly enhanced(35, 42, 56). Finally, the bHLH factor TFEB binds to arelated sequence motif as a heterodimeric complex with thebHLH factor TFE3 (17). Thus, a strikingly distinct andrestricted pattern of heterodimerization has been observedamong these factors, even though they are capable ofbinding to very similar, if not identical, target DNA se-quences in vitro. It may be noteworthy that the core of theconserved motif (GCGTGA [see reference 49 for a review])in several dioxin response elements bears some similarity tothe E box motif (see the underlined sequence). However, theArnt-receptor complex does not bind the E box, nor does thein vitro-expressed bHLH factor USF (which strongly bindsto the E box motif and lacks a known dimerization partnerprotein [23]) affect the XRE binding activity of the receptor(data not shown).Mechanism of activation of the dioxin receptor. Dioxin
receptor activity appears to be highly controlled by a com-
Latent Receptor. Ligand-Activated300 kD RecepLigand Binding Activity 200 kDNo DNA Binding Activity DNA Binding Activity
-I ~~~~~~~Dioxin
TCACGC TCACGC TCACGC,Target DNA Target DNA Target DNA
FIG. 7. Hypothetical model for ligand-dependent activation ofthe dioxin receptor into a DNA binding form. The ligand binding,but non-DNA binding -300-kDa form of dioxin receptor (DR) isstably associated with hsp90 (represented as a putative dimer) and isdefined as the latent (inactive) receptor form. Ligand-induced acti-vation of the receptor into the -200-kDa DNA binding, nuclear formappears to involve the following critical steps: (i) binding of ligand(dioxin), (ii) release of hsp90, and (iii) subsequent interaction withthe Arnt coregulator. The bHLH motifs of the receptor and Arnt areindicated. In our model, the receptor is derepressed simply bydissociation of hsp9O, resulting in interaction with Arnt. It is alsoconceivable that the ligand induces a specific receptor conformationthat is critical for dimerization with Arnt. See text for details.
plex pattern of protein-protein interactions resulting in eithernegative or positive regulation. Although the mechanism ofactivation of the dioxin receptor remains obscure, it is clearthat the decision to convert from a latent into a functionalreceptor form is strictly governed by the presence of recep-tor ligand (10). A striking feature of the latent dioxin receptoris that it is stably associated with hsp9O in a large, -300-kDacomplex (11, 48, 61). In analogy to the glucocorticoidreceptor (12 [and references therein]), ligand-induced acti-vation of the DNA binding activity of the dioxin receptorinvolves the release of hsp90 (52, 61). These observationscan be interpreted to suggest that hsp90 passively inhibitsArnt from interacting with the receptor simply by sterichindrance, resulting in inhibition of the XRE binding func-tion. As detailed in the model in Fig. 7, this mode ofrepression is readily reversed upon binding of ligand. Alter-natively, hsp90 maintains the ligand-free receptor polypep-tide in a specific conformation, in which the dimerizationinterface with Arnt may be masked. In support of the latterscheme, we have recently demonstrated that hsp90 main-tains and stabilizes the dioxin receptor in a ligand bindingconformation, whereas the hsp90-free form shows no detect-able dioxin binding activity in vitro (52). Conversely, artifi-cially induced release of hsp9o results in derepression of thereceptor, manifested in constitutive XRE binding activity(52). In support of this model, it has recently been shownthat purified hsp90 chaperones protein folding in vitro (60).Given the background that Arnt-deficient hepatoma cells arecharacterized by a nuclear translocation-deficient dioxinreceptor phenotype in vivo (30 [and references therein]),these data raise the interesting possibility, that, in theabsence of ligand, hsp9O maintains the receptor in a specific,repressed conformation that precludes DNA binding, nu-clear localization, or both receptor activities, by preventingArnt from interacting with the receptor. We are currentlyattempting to examine this hypothesis in closer detail.
VOL. 13, 1993

2512 WHITELAW ET AL.
Common functional properties of coregulators of ligand-activated nuclear receptor proteins. Although both the dioxinreceptor and Arnt are bHLH factors, there are strikingfunctional similarities to the mechanism of gene regulationby members of the steroid receptor (here referred to as zincfinger receptors) superfamily. Most importantly, the retinoidX receptor (RXR) appears to function as a critical coregula-tor of a broad numiber of zinc finger receptors including theretinoic acid, thyroid hormone, vitamin D, and peroxisomeproliferator-activated receptors. In a fashion similar to thatof Arnt, RXR seems to enhance or modulate the DNAbinding activity in vitro of partner zinc finger receptors bythe formation of a heterodimeric complex (7, 21, 24, 32, 33,36, 39, 64, 65). Since many of the partner receptors of RXR(such as the thyroid hormone receptor) are constitutivenuclear proteins (see reference 3 for a review), it is presentlyunclear whether RXR plays any role in the nuclear translo-cation process of its partner receptors. In the case of thedioxin receptor system it will be of interest to examinewhether Arnt associates with the receptor in the cytoplasmicor nuclear compartment of target cells. Clearly, there existeda 4-5S cytosolic activity that functionally corresponded toArnt with regard to promoting the DNA binding activity ofthe dioxin receptor in vitro. While there is only indirectevidence that the Arnt coregulator may be important fornuclear import of the ligand-activated dioxin receptor (30[and references therein]), it has been established that distinctcytosolic factors are required for the two critical steps duringnuclear translocation processes: targeting of the nuclearenvelope and import through the nuclear pore (41 [andreferences therein]).
Role of ligand in the interaction of nuclear receptors withpartner factors. One important feature of the dioxin receptorsystem appears to be that the receptor strictly requiredligand for functional interaction with the Arnt coregulator.Moreover, the level of receptor-dependent XRE bindingactivity that was reconstituted by addition of Arnt to thereceptor closely mirrored the dose and receptor affinity ofthe employed ligand. This mechanism of regulation of recep-tor activity has so far not been observed with regard to theeffect of the RXR coregulator on zinc finger receptor func-tion. However, the recent observation that the 9-cis retinoicacid (a ligand of RXR [29, 37]) induces homodimer formationof RXR (66) opens the interesting possibility that the ligandplays a more central role than previously anticipated inregulating the formation of homo- or heteromeric zinc fingerreceptor complexes.
Finally, by analogy to the regulation of RXR function by9-cis retinoic acid, one may consider the possibility that theArnt protein could bind a natural ligand and thus represent anovel class of bHLH orphan receptors. This therefore leadsto the intriguing question of whether the interaction of thereceptor and Arnt in fact reflects the physical convergence oftwo distinct signal transduction pathways. Clearly, however,Arnt does not bind dioxin (30), nor did dioxin treatmentinduce any Arnt-dependent XRE binding activity (data notshown).
Implications for the dioxin signal transduction process.Combinatorial regulation of dioxin receptor activity by pro-tein-protein interactions with hsp90 and the Arnt coregulatormay create the basis for a very complex and cell-type-specific pattern of modulation of receptor function. Forinstance, given the complex regulatory networks of otherbHLH factors, it is an interesting possibility that the recep-tor, Arnt, or both proteins interact with additional, as-yet-unidentified dimerization partners in target cells. Such puta-
tive partners may or may not be structurally related to Arntand the receptor. For instance, the Drosophila neural regu-lator sim is structurally related to the dioxin receptor andArnt (8, 15, 30, 43), implying that putative partner proteinsmay be found in developmental regulatory pathways. Underconditions in which Arnt or the receptor is limiting, suchpatterns of protein-protein interactions may result in differ-ential regulation of the receptor and possibly novel phenom-ena of cross-coupling of distinct regulatory pathways withthe dioxin signalling process. In the case of the zinc fingerreceptor family, a number of its members have been re-ported to be negatively regulated by interaction with thec-Fos/c-Jun complex belonging to the leucine zipper familyof DNA binding proteins (reviewed in references 40 and 58).Moreover, the bHLH factor MyoD is similarly repressed bydirect interaction with c-Jun but not c-Fos (4). Finally, theDNA-binding activity of the dioxin receptor appears to bemediated by protein kinase C-dependent phosphorylationprocesses (4a, 9, 46, 53).The efficiency of the putative Arnt-receptor heterodimer
formation may constitute a crucial step in regulation ofreceptor activity. Interestingly, the amount of added Arntprotein was clearly critical for efficient in vitro promotion ofthe DNA binding activity of the receptor (data not shown),indicating that a threshold level had to be surpassed for thefunctional effect to occur. These preliminary observationsraise the interesting possibility that the level of expression ofArnt (or the pool of Arnt available to dimerize with thereceptor) may be limiting for the receptor following ligandactivation and may thus provide an important mechanism fordetermining cellular sensitivity and responsiveness to diox-ins.
ACKNOWLEDGMENTS
M.W. and I.P. are equal first authors. We thank Oliver Hankinson(UCLA) for providing the full-length Arnt cDNA clone and wild-type and C4 mutant Hepa 1clc7 cells and Asa Andersson andKristina Sjoblom for excellent technical assistance. We also appre-ciate the comments from Anna Berghard, Anders Berkenstam, andMartin Gottlicher on the manuscript.M.W. was supported by a fellowship from the Swedish Cancer
Society. This work was supported by grants from the SwedishCancer Society.
REFERENCES1. Alksnis, M., T. Barkhem, P.-E. Stromstedt, H. Ahola, E. Kutoh,
J.-A. Gustafsson, L. Poellinger, and S. Nilsson. 1991. High levelexpression of functional full length and truncated glucocorticoidreceptor in Chinese hamster ovary cells: demonstration ofligand-induced down-regulation of expressed receptor mRNAand protein. J. Biol. Chem. 266:10078-10085.
2. Andersson, S., D. L. Davis, H. Dahiback, H. Jornvall, and D. W.Russel. 1989. Cloning, structure and expression of the mitochon-drial cytochrome P-450 sterol 26-hydroxylase, a bile acid bio-synthetic enzyme. J. Biol. Chem. 264:8222-8229.
3. Beato, M. 1989. Gene regulation by steroid hormones. Cell56:335-344.
4. Bengal, E., L. Ransone, R. Schafmann, V. J. Dwarki, S. J.Tapscott, H. Weintraub, and I. M. Verma. 1992. Functionalantagonism between c-Jun and MyoD proteins: a direct physicalinteraction. Cell 68:507-519.
4a.Berghard, A., K. Gradin, I. Pongratz, M. Whitelaw, and L.Poellinger. 1993. Cross-coupling of signal transduction path-ways: the dioxin receptor mediates induction of cytochromeP-450IA1 expression via a protein kinase C-dependent mecha-nism. Mol. Cell. Biol. 13:677-689.
5. Blackwood, E. M., and R. N. Eisenman. 1991. Max: a helix-loop-helix zipper protein that forms a sequence-specific DNA bindingcomplex with Myc. Science 251:1211-1217.
MOL. CELL. BIOL.

LIGAND-DEPENDENT RECRUITMENT OF THE Arnt COREGULATOR 2513
6. Blackwood, E. M., B. Luscher, and R. N. Eisenman. 1992. Mycand Max associate in vivo. Genes Dev. 6:71-80.
7. Bugge, T. H., J. Pohl, 0. Lonnoy, and H. G. Stunnenberg. 1992.RXRax, a promiscuous partner of retinoic acid and thyroidhormone receptors. EMBO J. 11:1409-1418.
8. Burbach, K. M., A. Poland, and C. A. Bradfield. 1992. Cloningof the Ah receptor cDNA reveals a distinctive ligand-activatedtranscription factor. Proc. Natl. Acad. Sci. USA 89:8185-8189.
9. Carrier, F., R. A. Owens, D. W. Nebert, and A. Puga. 1992.Dioxin-dependent activation of murine Cypla-1 gene transcrip-tion requires protein kinase C-dependent phosphorylation. Mol.Cell. Biol. 12:1856-1863.
10. Cuthill, S., A. Wilhelmsson, and L. Poellinger. 1991. Role of theligand in intracellular receptor function: receptor affinity deter-mines activation in vitro of the latent dioxin receptor to aDNA-binding form. Mol. Cell. Biol. 11:401-411.
11. Denis, M., S. Cuthill, A.-C. Wikstrom, L. Poellinger, and J.-A.Gustafsson. 1988. Association of the dioxin receptor with the mr90,000 heat shock protein: a structural kinship with the glu-cocorticoid receptor. Biochem. Biophys. Res. Commun. 155:801-807.
12. Denis, M., L. Poellinger, A. C. Wikstrom, and J.-A. Gustafsson.1988. Requirement of hormone for thermal conversion of theglucocorticoid receptor to a DNA-binding state. Nature (Lon-don) 333:686-688.
13. Denison, M. S., J. M. Fisher, and J. J. Whitlock. 1988. Induc-ible, receptor-dependent protein-DNA interactions at a dioxin-responsive transcriptional enhancer. Proc. Natl. Acad. Sci.USA 85:2528-2532.
14. Elferink, C. J., T. A. Gasiewicz, and J. P. Whitlock, Jr. 1990.Protein-DNA interactions at a dioxin-responsive enhancer: ev-idence that the transformed Ah receptor is heteromeric. J. Biol.Chem. 265:20708-20712.
15. Ema, M., K. Sogawa, Y. Watanabe, Y. Chujoh, N. Matsushita,0. Gotoh, Y. Funae, and Y. Fujii-Kuriyama. 1992. cDNAcloning and structure of mouse putative Ah receptor. Biochem.Biophys. Res. Commun. 184:246-253.
16. Favreau, L. V., and C. B. Pickett. 1991. Transcriptional regula-tion of the rat NAD(P)H:quinone reductase gene: identificationof regulatory elements controlling basal level expression andinducible expression by planar aromatic compounds and phe-nolic antioxidants. J. Biol. Chem. 266:4556-4561.
17. Fischer, D. E., C. S. Carr, L. A. Parent, and P. A. Sharp. 1991.TFEB has DNA binding and oligomerization properties of aunique helix-loop-helix/leucine zipper family. Genes Dev.5:2342-2352.
18. Fujisawa-Sehara, A., M. Yamane, and Y. Fujii-Kuriyama. 1988.A DNA-binding factor specific for xenobiotic responsive ele-ments of P-450c gene exists as a cryptic form in cytoplasm: itspossible translocation to nucleus. Proc. Natl. Acad. Sci. USA85:5859-5863.
19. Gallo, M. A., R. J. Scheuplein, and K. A. van der Heiden. 1991.Biological basis for risk assessment of dioxins and relatedcompounds. Banbury report 35. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
20. Gasiewicz, T. A., and R. A. Neal. 1982. The examination andquantification of tissue cytosolic receptors for 2,3,7,8-tetrachlo-rodibenzo-p-dioxin using hydroxylapatite. Anal. Biochem. 124:1-11.
21. Gearing, K. L., M. Gittlicher, M. Teboul, E. Widmark, andJ.-A. Gustafsson. Proc. Natl. Acad. Sci. USA, in press.
22. Green, S. 1992. Nuclear receptors and chemical carcinogenesis.Trends Pharmacol. Sci. 13:251-255.
23. Gregor, P. D., M. Sawadogo, and R. G. Roeder. 1990. Theadenovirus major late transcription factor USF is a member ofthe helix-loop-helix group of regulatory proteins and binds toDNA as a dimer. Genes Dev. 4:1730-1740.
24. Hallenbeck, P. L., M. M. Marks, R. E. Lippoldt, K. Ozato, andV. M. Nikodem. 1992. Heterodimerization of thyroid hormone(TH) receptor with H-2RIIBP (RXR3) enhances DNA bindingand TH-dependent transcriptional activation. Proc. Natl. Acad.Sci. USA 89:5572-5576.
25. Hankinson, 0. 1983. Evidence that benzo(a)pyrene-resistant,
aryl hydrocarbon hydroxylase-deficient variants of mouse hep-atoma line, Hepa-1, are mutational in origin. Somatic CellGenet. 9:497-514.
26. Hapgood, J., S. Cuthill, M. Denis, L. Poellinger, and J.-A.Gustafsson. 1989. Specific protein-DNA interactions at a xeno-biotic-responsive element: copurification of dioxin receptor andDNA-binding activity. Proc. Natl. Acad. Sci. USA 86:60-64.
27. Haird, T., E. Kellenbach, R. Boelens, B. Maler, K. Dahlman,L. P. Freedman, K. R. Yamamoto, J.-A. Gustafsson, and R.Kaptein. 1990. Solution structure of the glucocorticoid receptorDNA binding domain. Science 249:157-160.
28. Harlow, E., and D. Lane. 1988. Antibodies: a laboratory man-ual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,N.Y.
29. Heyman, R. A., D. J. Mangelsdorf, J. A. Dyck, R. B. Stein, G.Eichele, R. M. Evans, and C. Thaller. 1992. 9-cis retinoic acid isa high affinity ligand for the retinoid X receptor. Cell 68:397-406.
30. Hoffman, E. C., H. Reyes, F. F. Chu, F. Sander, L. H. Conley,B. A. Brooks, and 0. Hankinson. 1991. Cloning of a factorrequired for activity of the Ah (dioxin) receptor. Science 252:954-958.
31. Kato, G. J., W. M. F. Lee, L. Chen, and C. V. Dang. 1992. Max:functional domains and interaction with c-Myc. Genes Dev.6:81-92.
32. Kliewer, S. A., K. Umesono, D. J. Mangelsdorf, and R. M.Evans. 1992. Retinoid X receptor interacts with nuclear recep-tors in retinoic acid, thyroid hormone and vitamin D3 signalling.Nature (London) 355:446-449.
33. Kliewer, S. A., K. Umesono, D. J. Noonan, R. A. Heyman, andR. M. Evans. 1992. Convergence of 9-cis retinoic acid andperoxisome proliferator signalling pathways through het-erodimer formation of their receptors. Nature (London) 358:771-774.
34. Landers, J. P., and N. J. Bunce. 1991. The Ah receptor and themechanism of dioxin toxicity. Biochem. J. 276:273-287.
35. Lassar, A. B., R. L. Davis, W. E. Wright, T. Kadesh, C. Murre,A. Voronova, D. Baltimore, and H. Weintraub. 1991. Functionalactivity of myogenic HLH proteins requires hetero-oligomeriza-tion with E12/E47-like proteins in vivo. Cell 66:305-315.
36. Laid, M., P. Kastner, R. Lyons, H. Nakshatri, M. Saunders, T.Zacharewski, J.-Y. Chen, A. Staub, J.-M. Garnier, S. Mader,and P. Chambon. 1992. Purification, cloning and RXR identityof the HeLa cell factor with which RAR or TR heterodimerizesto bind target sequences specifically. Cell 68:377-395.
37. Levin, A. A., L. J. Sturzenbecker, S. Kazmer, T. Bosakowski, C.Huselton, G. Allenby, J. Speck, C. Kratzeisen, M. Rosenberger,A. Lovey, and J. F. Grippo. 1992. 9-cis retinoic acid stereoiso-mer binds and activates the nuclear receptor RXRa Nature(London) 355:359-361.
38. Luisi, B. F., W. X. Xu, Z. Otwinowski, L. P. Freedman, K. R.Yamamoto, and P. B. Sigler. 1991. Crystallographic analysis ofthe interaction of the glucocorticoid receptor with DNA. Nature(London) 352:497-505.
39. Marks, M. S., P. L. Hallenbeck, T. Nagata, J. H. Segars, E.Appella, V. M. Nikodem, and K. Ozato. 1992. H-2RIIBP (RXR,)heterodimerization provides a mechanism for combinatorialdiversity in the regulation of retinoic acid and thyroid hormoneresponsive genes. EMBO J. 11:1419-1435.
40. Miner, J. N., and K. R. Yamamoto. 1991. Regulatory crosstalkat composite response elements. Trends Biochem. Sci. 16:423-426.
41. Moore, M. S., and G. Blobel. 1992. The two steps of nuclearimport, targeting to the nuclear envelope and translocationthrough the nuclear pore, require different cytosolic factors.Cell 69:939-950.
42. Murre, C. S., P. S. McCaw, H. Vassin, M. Caudy, L. Y. Jan,Y. N. Jan, C. V. Cabrera, J. N. Buskin, S. D. Hauschka, A. B.Lassar, H. Weintraub, and D. Baltimore. 1989. Interactionsbetween heterologous helix-loop-helix proteins generate com-plexes that bind specifically to a common DNA sequence. Cell58:537-544.
43. Nambu, J. R., J. 0. Lewis, K. A. Wharton, Jr., and S. T. Crews.
VOL. 13, 1993

2514 WHITELAW ET AL.
1991. The drosophila single-minded gene encodes a helix-loop-helix protein that acts as a master regulator of CNS midlinedevelopment. Cell 67:1157-1167.
44. Nemoto, T., G. G. F. Mason, A. Wilhelmsson, S. Cuthill, J.Hapgood, J.-A. Gustafsson, and L. Poellinger. 1990. Activationof the dioxin and glucocorticoid receptors to a DNA bindingstate under cell-free conditions. J. Biol. Chem. 265:2269-2277.
45. Neuhold, L. A., Y. Shirayoshi, K. Ozato, J. E. Jones, and D. W.Nebert. 1989. Regulation of mouse CYPlAl gene expression bydioxin: requirement of two cis-acting elements during induction.Mol. Cell. Biol. 9:2378-2386.
46. Okino, S. T., U. R. Pendurthi, and R. H. Tukey. 1992. Phorbolesters inhibit the dioxin receptor-mediated transcriptional acti-vation of the mouse Cypla-1 and Cypla-2 genes by 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Chem. 267:6991-6998.
47. Paulson, K. E., J. Darnell, Jr., T. Rushmore, and C. B. Pickett.1990. Analysis of the upstream elements of the xenobioticcompound-inducible and positionally regulated glutathioneS-transferase Ya gene. Mol. Cell. Biol. 10:1841-1852.
48. Perdew, G. H. 1988. Association of the Ah receptor with the 90kD heat shock protein. J. Biol. Chem. 263:13802-13805.
49. Poellinger, L., M. Gottlicher, and J.-A. Gustafsson. 1992. Thedioxin and peroxisome proliferator-activated receptors: nuclearreceptors in search of endogenous ligands. Trends Pharmacol.Sci. 13:241-245.
50. Poland, A., E. Glover, F. H. Ebetino, and A. S. Kende. 1986.Photoaffinity labelling of the Ah receptor. J. Biol. Chem.261:6352-6365.
51. Poland, A., and J. C. Knutson. 1982. 2,3,7,8-Tetrachlorodi-benzo-p-dioxin and related halogenated aromatic hydrocarbons:examination of the mechanism of toxicity. Annu. Rev. Pharma-col. Toxicol. 22:517-554.
52. Pongratz, l., G. G. F. Mason, and L. Poellinger. 1992. Dual rolesof the 90 kD heat shock protein in modulating functionalactivities of the dioxin receptor. J. Biol. Chem. 267:13728-13734.
53. Pongratz, I., P. E. Stromstedt, G. G. Mason, and L. Poellinger.1991. Inhibition of the specific DNA binding activity of thedioxin receptor by phosphatase treatment. J. Biol. Chem.266:16813-16817.
54. Prendergast, G. C., D. Lawe, and E. B. Ziff. 1991. Association ofMyn, the murine homolog of Max, with c-Myc stimulatesmethylation-sensitive DNA binding and Ras cotransformation.Cell 65:395-407.
55. Prokipcak, R. D., and A. B. Okey. 1988. Physicochemicalcharacterization of the nuclear form of Ah receptor from mousehepatoma cells exposed in culture to 2,3,7,8-tetrachlorodi-benzo-p-dioxin. Arch. Biochem. Biophys. 261:6352-6365.
56. Rashbass, J., M. V. Taylor, and J. B. Gurdon. 1992. The DNAbinding protein E12 co-operates with XMyoD in the activationof muscle-specific gene expression in Xenopus embryos. EMBOJ. 11:2981-2990.
57. Reyes, H., S. Reiz-Porszasz, and 0. Hankinson. 1992. Identifi-cation of the Ah receptor nuclear translocator protein (Arnt) asa component of the DNA binding form of the Ah receptor.Science 256:1193-1195.
58. Schule, R., and R. M. Evans. 1991. Cross-coupling of signaltransduction pathways: zinc finger meets leucine zipper. TrendsGenet. 7:377-381.
59. Sutter, T. R., K. Guzman, K. M. Dold, and W. F. Greenlee.1991. Targets for dioxin: genes for plasminogen activator inhib-itor-2 and interleukin-1,. Science 254:415-418.
60. Wiech, H., J. Buchner, R. Zimmermann, and U. Jakob. 1992.Hsp9O chaperones protein folding in vitro. Nature (London)358:169-170.
61. Wilhelmsson, A., S. Cuthill, M. Denis, A. C. Wikstrom, J.-A.Gustafsson, and L. Poellinger. 1990. The specific DNA bindingactivity of the dioxin receptor is modulated by the 90 kd heatshock protein. EMBO J. 9:69-76.
62. Wilhelmsson, A., and L. Poellinger. Unpublished data.63. Wilhelmsson, A., A.-C. Wikstrom, and L. Poellinger. 1986.
Polyanionic binding properties of the receptor for 2,3,7,8-tetrachlorodibenzo-p-dioxin: a comparison with the glucocorti-coid receptor. J. Biol. Chem. 261:13456-13463.
64. Yu, V. C., C. Delsert, B. Andersen, J. M. Holloway, 0. V.Devary, A. M. Nfiar, S. Y. Kim, J.-M. Boutin, C. K. Glass, andM. G. Rosenfeld. 1991. RXR,: a coregulator that enhancesbinding of retinoic acid, thyroid hormone, and vitamin Dreceptors to their cognate response elements. Cell 67:1251-1266.
65. Zhang, X.-K., B. Hoffman, P. B.-V. Tran, G. Graupner, and M.Pfahl. 1992. Retinoid X receptor is an auxiliary protein forthyroid hormone and retinoic acid receptors. Nature (London)355:441-446.
66. Zhang, X.-K., J. Lehmann, B. Hoffman, M. I. Dawson, J.Cameron, G. Graupner, T. Hermann, P. Tran, and M. Pfahl.1992. Homodimer formation of retinoid X receptor induced by9-cis retinoic acid. Nature (London) 358:587-591.
MOL. CELL. BIOL.
Related Documents