Retrospective eses and Dissertations Iowa State University Capstones, eses and Dissertations 1-1-2000 Lethal and sublethal effects of ivermectin in a freshwater oligochaete, Lumbriculus variegatus Jing Jing Iowa State University Follow this and additional works at: hps://lib.dr.iastate.edu/rtd is esis is brought to you for free and open access by the Iowa State University Capstones, eses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective eses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Recommended Citation Jing, Jing, "Lethal and sublethal effects of ivermectin in a freshwater oligochaete, Lumbriculus variegatus" (2000). Retrospective eses and Dissertations. 17903. hps://lib.dr.iastate.edu/rtd/17903

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Retrospective Theses and Dissertations Iowa State University Capstones, Theses andDissertations
1-1-2000
Lethal and sublethal effects of ivermectin in afreshwater oligochaete, Lumbriculus variegatusJing JingIowa State University
Follow this and additional works at: https://lib.dr.iastate.edu/rtd
This Thesis is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University DigitalRepository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University DigitalRepository. For more information, please contact [email protected].
Recommended CitationJing, Jing, "Lethal and sublethal effects of ivermectin in a freshwater oligochaete, Lumbriculus variegatus" (2000). Retrospective Thesesand Dissertations. 17903.https://lib.dr.iastate.edu/rtd/17903

Lethal and sub lethal effects of ivermectin in a
freshwater oligochaete, Lumbriculus variegatus
by
Jing Ding
A thesis submitted to the graduate faculty
in partial fulfillment of the requirements for the degree of
MASTER OF SCIENCE
Major: Toxicology
Major Professor: Walter H. Hsu
Iowa State University
Ames, Iowa
2000
Copyright© Jing Ding, 2000. All right reserved

11
Graduate College Iowa State University
This is to certify that the Master' s thesis of
Jing Ding
has met the thesis requirements of Iowa State University
Signatures have been redacted for privacy
Signatures have been redacted for privacy

111
TABLE OF CONTENTS
REVIEW OF LITERATURE 1
Ivermectin 1
Structure, physical and chemical characteristics of ivermectin 1
Application of ivermectin 3
Antiparasitic effect of ivermectin and mechanism of action 4
Other actions of ivermectin 5
Safety of ivermectin in mammals 5
Environmental fate and concern of ivermectin 6
Freshwater oligochaete, Lumbriculus variegatus 7
Locomotor behaviors in L. variegatus 7
Giant-intemeuron pathways and escape reflex 8
Non-giant intemeuron pathways and other locomotor behaviors 9
LETHAL AND SUBLETHAL EFFECTS OF IVERMECTIN IN A FRESHWATER OLIGOCHAETE, LUMBRICULUS VARIEGATUS 10
Introduction 10
Methods and Materials 12
Materials 12
Animal maintenance and selection 12
Treatment 12
Lethal effect 13
Behavioral testing 13
Electrophysiological testing 15
Data analysis 16
Results 17
Lethal effect 1 7
Sublethal effects 18
Effect of ivermectin on locomotor behaviors controlled by non-giant-intemeuron pathways 18
Effect of ivermectin on behavior controlled by giant-intemeuron pathways 23
Discussion
CONCLUSIONS
28
33

LITERATURE CITED
ACKNOWLEDGEMENTS
IV
34
41

1
REVIEW OF LITERATURE
Ivermectin
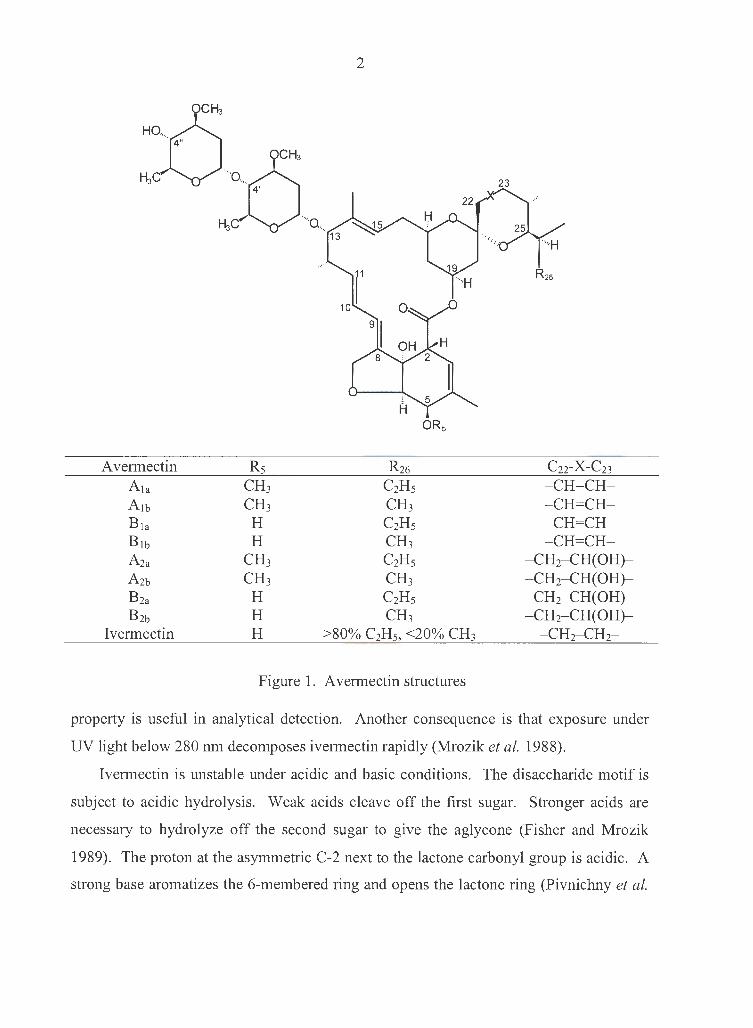
Ivermectin (22,23-dihydroavermectin B 1) is a member of the avermectin family. It is
the drug of choice for the treatment and prevention of many nematode and arthropod
parasite infections (Chabala et al. 1980; Campbell et al. 1983, 1984, 1991, 1993; Ottesen
et al. 1994). The anthelmintic activity of the avermectins was first described in 1979
(Burg et al. 1979; Egerton et al. 1979; Miller et al. 1979). Ivermectin was introduced
commercially in 1981.
Structure, physical and chemical characteristics of ivermectin
Fermentation of the actinomycete Streptomyces avermitilis produces four
homologous pairs of closely related, complex, 16-membered, macrocyclic lactones:
avermectin A1, A2, B 1 and B2. Based on the sidechain composition at carbon position 25
(Fig. 1), the four pairs are further divided into the major components "a" (a secondary
butyl substituent at C-25, more than 80%) and minor components "b" (an isopropyl
group, less than 20%). Avermectin B 1a has the highest potency against many of endo
and ectoparasites of animals, as well as many agricultural mite and insect pests. It is also
the starting material for synthesizing ivermectin.
Because it is very difficult and impractical to separate the major components "a"
from the minor components "b", and because the biological activities of the two
components are almost indistinguishable, the avermectins are often used as a mixture of
the two components (more than 80% of component "a", less than 20% of "b"). In such
cases they are just named as A1, A2, B 1 and B2. Ivermectin (22,23-dihydroavermectin B 1)
is used as such a mixture.
The avermectins are highly lipophilic, and dissolve in most organic solvents. Their
water solubility is correspondingly low - only 6 to 9 ppb (Fisher and Mrozik 1989).
lvermectin contains two double bonds conjugated as an 8,9,10,11-diene function,
which results in a strong UV absorption at 245 nm (Fisher and Mrozik 1989). This
HO,. 4"
Avermectin A1a
A1b
Bia
B1b
A2a
A2b
B2a
B2b
Ivermectin
OCH:i
OCH3
4'
H3C ·•,,a,,
Rs CH3 CH3
H H
CH3 CH3 H H H
2
13
C2Hs CH3 C2Hs CH3 C2Hs CH3 C2Hs CH3
ORs
>80% C2Hs, <20% CH3
Figure 1. A vermectin structures
C22-X-C23 - CH=CH-- CH=CH-- CH=CH--CH=CH-
- CH2- CH(OH)-- CH2- CH(OH)-- CH2- CH(OH)-- CH2- CH(OH)-
- CH2-CH2-
property is useful in analytical detection. Another consequence is that exposure under
UV light below 280 nm decomposes ivermectin rapidly (Mrozik et al. 1988).
Ivermectin is unstable under acidic and basic conditions. The disaccharide motif is
subject to acidic hydrolysis. Weak acids cleave off the first sugar. Stronger acids are
necessary to hydrolyze off the second sugar to give the aglycone (Fisher and Mrozik
1989). The proton at the asymmetric C-2 next to the lactone carbonyl group is acidic. A
strong base aromatizes the 6-membered ring and opens the lactone ring (Pivnichny et al.

3
1983, 1988). Consequently, strong bases can be used to destroy ivermectin residues for
safe disposal.
Application of ivermectin
Synthetic compounds dominated the anthelmintic market until the late 1970s. The
discovery of the avermectins changed that dramatically. Ivermectin has been used most
widely in domestic animals in the control of parasitic diseases that cause losses of
hundreds of millions of dollars annually. Its use in human medicine has also received
considerable attention because of its control of onchocerciasis (river blindness), one of
the most insidious and intractable of tropical diseases.
Ivermectin is used against a wide range of nematode and arthropod parasites in
domestic animals (Chabala et al. 1980; Campbell et al. 1983, 1984, 1993), as well as in
laboratory and exotic mammals and in birds, fish, and reptiles (Soll 1989). Essentially all
important nematode species are susceptible, both immature and mature stages. In
addition to its activity against nematode parasites, ivermectin is also active against
several important ectoparasites, such as grubs, screwworms, lice, mites and ticks.
Because of its activity against both endoparasitic and ectoparasitic organisms, it has
become known as the first "endectocide". Ivermectin is active when given orally,
intramuscularly, subcutaneously, or even topically.
Ivermectin was first reported in human trials against onchocerciasis in 1982 (Aziz et
al. 1982a, 1982b). Its clinical trials and usage have been well reviewed (Campbell 1991,
1993). Its potent microfilaricidal activity against the other major filarial parasites of
humans has also been recognized (Campbell 1991; Ottesen et al. 1994). Preliminary
studies have shown that ivermectin is effective against some human intestinal nematodes,
Ascaris lumbricoides, Strongyloides stercoralis, Trichuris trichiura and Enterobius
vermicularis, but weak against other species, Ancylostoma duodenale and Necator
americanus (Campbell 1993). The efficacy of ivermectin against ectoparasitic
infestations (mites, lice) of humans has also been reported (Ottesen et al. 1994).

4
Antiparasitic effect of ivermectin and mechanism of action
Ivermectin reduces motor activity of the parasites so that they are excreted from the
host (Martin 1993). Early electrophysiological studies using muscle or nerve
preparations from lobsters, insects, large nematodes or mammalian brains (Fritz et al.
1979; Kass et al. 1980, 1984; Duce and Scott 1985; Mellin et al. 1983; Abalis et al. 1986;
Albert et al. 1986; Krusek and Zemkova 1994; Huang and Casida 1997) suggested that
ivermectin interferes with synaptic transmission, and that this is the result of increased
cell membrane permeability to chloride ions, which is explained by the opening of
GABA-gated er channels.
It has recently been shown, however, that ivermectin also activates Cl- channels that
are not regulated by GABA. Specific ivermectin binding sites have been identified in the
free-living nematode Caenorhabditis elegans and in mammalian brain tissue. The
affinity of ivermectin for the nematode site (apparent dissociation constant, ~ = 0.26
nM), is about 100 times greater than that for the mammalian brain site (Schaeffer and
Haines 1989). The failure of GABA to compete with ivermectin for binding to the
nematode (Cully and Paress 1991) suggests that the GABA and ivermectin receptors are
significantly different. The mode of action of ivermectin has also been studied in
specially prepared oocytes of the frog Xenopus laevis. After mRNA of C. elegans is
injected into an oocyte, a Cl- current can be activated in the membrane of the oocyte by
ivermectin and L-glutamate, but not GABA (Arena et al. 1991, 1992). These studies
together suggest that GABA plays little, if any, part in the antiparasitic action of
ivermectin. They also lead to the discovery of a novel type of er channels, namely a
glutamate-gated ivermectin-sensitive Cl- channel.
Glutamate-gated ivermectin-sensitive Cl- channels have been described in many
invertebrates, such as nematodes, insects, crustaceans, and mollusks (Cleland 1996).
Although their physiological role has not been fully determined, they are reported to be
involved in generating rhythmic firing of the neurons within the crustacean
stomatogastric ganglion (Cleland and Selverston, 1995, 1998). Two subunits (GluCla
and GluCl~) have been cloned from C. elegans (Cully et al. 1994). Gene structure
comparisons and phylogenetic analyses reveal that these channels are related to

5
vertebrate glycine channels/receptors. Since these channels only exist in invertebrates,
they may mediate the powerful anthelmintic and insecticidal action of ivermectin, while
accounting for the low toxicity to mammals.
Other actions of ivermectin
It was reported that ivermectin specifically binds to retinal-binding protein isolated
from parasitic worms of the family Filarioidae, but not the host organism (Sani and Vaid
1988). lvermectin was also reported to be a specific inhibitor of protein kinase C isolated
from rat brain (Ellis et al. 1987). The high concentration of ivermectin required for this
action (IC50 : 1 µM) suggests that this may not be of physiological significance. In
sarcoplasmic reticulum (SR) of rat and rabbit skeletal muscle, ivermectin directly
activates ryanodine receptor Ca2+ channels, and reduces Ca2+ uptake by the SR Ca2+ -
Mg2+-ATPase (Ahem et al. 1999). Once again, the high dose of ivermectin (10 µM) may
not be pharmacologically significant.
Safety of ivermectin in mammals
Clinical signs of the acute toxicity of ivermectin in mammals may be related to its
effect on GABA-gated c1- channels in the mammalian brain and spinal cord. Common
signs include ataxia, tremors, and in severe cases, coma and death (Lankas and Gordon
1989; Pulliam and Preston 1989). Rodents are more sensitive to ivermectin toxicity than
are other species. Severe reactions and death occur in mice at dosages as low as 200
µg/kg- the dosage that is routinely used in treating parasitic nematodes in animals and
onchocerciasis in human (LD50 = 25 mg/kg) (Campbell 1993). However, doses of 2.0
mg/kg or higher are needed for acute toxicity in other species, such as rats, rabbits, rhesus
monkeys, horses, cattle, sheep, goats, swine and dogs (Lankas and Gordon 1989; Pulliam
and Preston 1989).
Results of some subchronic, developmental and reproductive toxicity studies are
summarized in Table 1 (Lankas and Gordon 1989). It is concluded that ivermectin is
well tolerated in most mammals.

6
Table 1. No-observed-effect level (mg/kg/day) of toxicity studies with ivermectin
Species Subchronic studies Developmental and reproductive studies
0.1 Mouse -
(maternotoxicity)
Rat 0.4 5.0 (maternotoxicity
(splenic enlargement) and developmental toxicity)
Rabbit 1.5
-(developmental toxicity)
Dog 0.5
(mydriasis) -
Rhesus monkey >1.2
(treatment-related effects) -
Environmental fate and concern of ivermectin
When ivermectin is administered (orally, topically, or subcutaneously) to food
producing animals, the drug enters the environment as a result of excretion. Most of it (at
least 90%) is excreted as intact drug in the feces of the treated animal. The remainder is
excreted in urine (Campbell 1993).
The distribution and movement of a chemical in the environment depends on the
compound's physical and chemical properties. Ivermectin's low water solubility predicts
tight binding to soils, as evidenced by the high partition coefficient (Ko = concentration
in soil / concentration in water = 227 to 333) and high organic-carbon binding constant
(Koc = 12,600 to 15,700) (Halley et al. 1989). The strong soil binding results in the
minimal leaching into ground water. But run-off in surface water with eroding sediments
should be expected.
Ivermectin is degraded by sunlight and soil microbes. Depending on the
environmental conditions, the degradation half-life of ivermectin varies from 7 - 14 d in
summer to 91 - 217 d in winter (Halley et al. 1989).
Because of its potent antiparasitic effect mediated through glutamate-gated c1-
channels, which are present in many invertebrates (Cleland 1996), ivermectin raises
concerns about its impact on non-target organisms, especially invertebrates, and
associated environmental consequences. It has been shown that dung from ivermectin
treated animals failed to degrade in the normal way and this failure was associated with
-

7
the absence of dung-degrading invertebrate community, which includes many species of
insects and oligochaetes (Wall and Strong 1987; Strong 1993). This may exert a threat to
pastureland ecology.
Although lethal levels of ivermectin have been determined in some non-target
organisms - daphnia, fish, earthworms, birds and insects (Halley et al. 1993), the effects
of sub lethal doses have hardly been recognized at the present time. In the present study,
we examined the lethal and sublethal behavioral and electrophysiological effects of
ivermectin in a freshwater oligochaete, Lumbriculus variegatus.
Freshwater oligochaete, Lumbriculus variegatus
Blackworms, Lumbriculus variegatus (Phylum Annelida, Class Oligochaeta), are
common in ponds, lakes, and marshes of North America and Europe. They freely crawl
within and feed on submerged and decaying vegetation, such as rotting leaves and logs
(Drewes and Fourtner 1989).
Locomotor behaviors in L. variegatus
Generally, L. variegatus moves toward food sources and away from strong light and
open spaces. When suddenly touched or threatened, it uses a variety of responses to
move to safety. The first response, seen when worms are covered or in contact with
substrate, is an escape reflex involving rapid body shortening and withdrawal of the
worm's head or tail end. However, other reflex responses may occur depending on the
worm's environmental context. On wet surfaces or in confined spaces under water,
worms crawl forward or backward when lightly touched at tail or head end, respectively.
However, when worms are submerged in "open water" condition, tail stimulation evokes
rhythmic waves of helical swimming, while head stimulation evokes a body reversal
response (Drewes, 1999a). Each of these reflex behaviors is apparently controlled by a
different neural pathway (Fig. 2).

Stimulus
Touch
Cl)
~ 0
~ (1) z c 0 Cl)
~ (1)
VJ
8
,-----------------~
I Non-giant i ;----;
I ,Cll, Cl)
···II::~ Intemeuron i • • ••; § 1' • •• ~ : I~
Pathways I : ;:j a I : I (1) µ.. ------------------ I: z ,-----------------~
I
: Giant ; 6 : Intemeuron -+-1 o ,' I p h :;E: at ways : :
I ____ 1
CNS = Ventral Nerve Cord
Behaviors
Swimming, Reversal, Crawling
Escape withdrawal
Figure 2. Diagram of neural pathways in L. variegatus and locomotor behaviors controlled by different neural pathways
Giant-interneuron pathways and escape reflex
Rapid escape responses in oligochaetes are mediated by two independent giant
intemeuron pathways, the medial giant fiber (MGF) system and the lateral giant fiber
(LGF) system (Drewes 1984; Zoran and Drewes 1987; Drewes and Fourtner 1989;
Drewes and Brinkhurst 1990). Both MGF and LGF pathways consist of tandemly
aligned large caliber axons derived from intemeurons in each segment. The tandem
axonal units are connected at intersegmental boundaries by electrical synapses (gap
junctions). These connections cause the fibers to act as a functional syncytium, allowing
uninterrupted conduction of action potentials along the whole length of the ventral nerve
cord (Drewes 1999b ). The MGF is excited by tactile stimulation to the anterior 30% of
the worm's body, while the sensory field for the LGF extends over the posterior 70% of
the body (Drewes and Fourtner 1990). The LGF is also excited by photosensory inputs
(Drewes and Fourtner 1989). In each segment the giant fibers activate motor neurons,
which innervate body wall longitudinal muscles and bring about body shortening
(Drewes 1999b ).

9
Non-giant interneuron pathways and other locomotor behaviors
Tactile stimulation anywhere within the posterior one-half of the worm consistently
evokes swimming responses when the worm is in "open water" conditions. Swimming
movements consist of a series of alternating left-handed and right-handed, rapid and
rhythmic waves of helical body bending. Each wave rapidly progresses from head to tail,
thus propelling the worm forward (Drewes 1999a).
Touching to anterior segments while in "open water" evokes body reversal, which
consists of a stereotyped sequence of bending movements that effectively reverses head
and tail positions. Although there is little net movement of the body center, reversal may
reposition the head away from the predatory threat and prepare the worm for swimming
away from the threatening stimulus (Drewes 1999a).
Crawling movements are seen when the worm is in substantial contact with substrate
or sediment, thus restricting other movements, such as swimming. Crawling consists of a
series of rhythmic peristaltic waves of circular and longitudinal muscle contraction. Each
wave begins at the anterior or posterior end and rapidly progresses posteriorly or
anteriorly along the body (forward or backward crawling, respectively).
The specific network of neurons in an animal's nervous system that controls
rhythmic movements, such as the worm's swimming and crawling, is called a central
pattern generator (Young 1989; Drewes 1999b). Central pattern generators for
locomotion in invertebrates, such as annelids and arthropods, are usually in the ventral
nerve cord but do not involve giant nerve fibers.

10
LETHAL AND SUBLETHAL EFFECTS OF
IVERMECTIN IN A FRESHWATER OLIGOCHAETE,
LUMBRICULUS VARIEGATUS
Introduction
Ivermectin, (22, 23-dihydroavermectin B 1a), a semisynthetic avermectin analog, is a
potent anthelmintic and insecticide against nematode and arthropod parasites. It is
widely used to improve the health care of livestock and companion animals (Chabala et
al. 1980; Campbell et al. 1983, 1984, 1993). Its use also extends to human medicine
(Campbell et al. 1991; Ottesen et al. 1994). Although ivermectin's mode of action is not
fully understood, it is generally believed that ivermectin reduces excitability of muscle or
nerve through the opening of CC channels (Arena et al. 1995; Brownlee et al. 1997; Duce
& Scott 1985; Kass et al. 1980; Martin et al. 1997).
The environmental effects and fates of ivermectin have also been studied (Halley et
al. 1989, 1993; Roncalli 1989; Strong 1993; Wall & Strong 1987). In the vast majority
of these studies, mortality has been the endpoint. Daphnia magna was particularly
sensitive to ivermectin with 48 h LC50 of 0.025 ppb (~ 0.03 nM, Halley et al. 1993). Fish
were less sensitive [48 h LC50 for bluegill (Lepomis macrochirus) and rainbow trout
(Oncorhynchus mykiss) were 4.8 ppb and 3.0 ppb, respectively], and earthworms (Eisenia
foetida, 28 d LC50 315 ppm in soil) were relatively insensitive to ivermectin (Halley et al.
1993). Without analysis of sublethal end-points (e.g., effects on growth, development,
fecundity, morphology, behavior, or physiology), we can not fully understand the
complex biological actions or predict ecological impacts of environmental toxicants.
This is especially true for a chemical like ivermectin, which exerts antiparasitic effects
not by immediately killing the target organisms but by reducing their motor activities so
that the parasites are excluded from the host (Martin 1993).
Among a few studies concerning sublethal effects of ivermectin on non-target
organisms, most have examined development and reproduction, especially in dung
dwelling insect populations, that may be threatened by drug residues in the manure from

11
ivermectin-treated animals (Halley et al. 1993). Ivermectin residues inhibited larval
development of bushfly, M vetustissima and M. domestica (Wardhaugh et al. 1993,
1996; Wardhaugh and Mahon 1998). Adult dung beetles (C. hispanus, Bubas bubalus,
or Onthophagus binodis) survived exposure to dung containing ivermectin residue, but
the rate of oviposition was reduced (Ridsdill-Smith 1988, 1993; Wardhaugh and
Rodriguez-Menendez 1988).
Locomotor capabilities are logical focal points for studies of sublethal effects of
ivermectin, because the most predominant effects shown in target organisms are reduced
motor activities (Martin 1993). However, effects of ivermectin on motor activity on non
target organisms are poorly understaood. In this study, we examined sublethal effects of
ivermectin on locomotor behaviors of a non-target invertebrate, Lumbriculus variegatus
(Family Lumbriculidae, common name: mud worm, blackworm).
Features that make L. variegatus especially suitable for this study include: (i) their
ubiquitous inhabitation in North America and Europe, and introduction into Africa,
Australia, and New Zealand (Brinkhurst and Jamieson 1971); (ii) their freshwater benthic
habitat, in which they are vulnerable to the possible runoff of ivermectin within eroding
sediments; (iii) their ease in laboratory rearing, maintenance and handling; (iv) their
defined patterns of locomotor behaviors, namely helical swimming, body reversal
(Drewes 1999a), and crawling (Drewes and Cain 1999); (v) the presence of giant nerve
fibers (intemeurons) that mediate rapid escape responses (Drewes 1984; Drewes and
Fourtner 1989; Drewes and Brinkhurst 1990); (vi) and the capability of non-invasive
electrophysiological testing of escape reflex function (Drewes 1984; Zoran and Drewes
1987; Rogge and Drewes 1993).
Our specific objectives were to: (i) determine lethal concentrations of ivermectin in
L. variegatus; (ii) examine sublethal effects of ivermectin on locomotor behaviors:
swimming, reversal and crawling; (iii) verify the involvement of Cl- channels using
picrotoxin, a Cl- channel blocker, and (iv) examine electrophysiologically the effects of
ivermectin on giant nerve fiber pathways.

12
Methods and Materials
Materials
The following chemicals were used: ivermectin stock solution (10 mg/ml in 40%
glycerol formal and 60% propylene glycol, Merck AgVet, Rahway, NJ) and picrotoxin
(Sigma Chemical Co., St. Louis, MO).
Animal maintenance and selection
L. variegatus were reared in the laboratory from asexually reproducing colonies.
The worms were kept in aerated aquaria containing pieces of brown paper towel (21 -
23°C) and fed three to five times per week with sinking fish food.
Medium-sized worms (::::; 4 - 5 cm long) were removed from rearing tanks 12 - 24 h
prior to testing and placed in Petri dishes containing distilled water to allow clearance of
gut contents. Worms were visually screened for uniformity in segmentation pattern.
Worms showing recent segment regeneration or any obvious morphological defects were
not used.
Treatment
Ivermectin solutions were prepared by diluting ivermectin stock solution in distilled
water. Additional propylene glycol was added to all the solutions of lower ivermectin
concentrations so that the volume of vehicle in all solutions was the same. Control
solutions also had the same volume of vehicle as the ivermectin solutions. In ivermectin
and picrotoxin antagonism experiments, picrotoxin was dissolved in distilled water.
After worms were exposed in the picrotoxin solutions for 60 min, ivermectin and/or
propylene glycol were added to the solutions. Concentration levels of ivermectin and/or
picrotoxin were determined according to preliminary range-finding experiments (Table
2). All concentrations reported are nominal; no analytical procedures were performed to
verify the actual concentrations of the chemicals. However, all aqueous solutions were
freshly prepared immediately before the experiments.

13
Table 2. Treatments used in the present study
Experiment Treatments
LCso Ivermectin: 180, 320, 560, 1000, 1800 nM
Swimming and reversal Ivermectin: vehicle, 0.3, 3, 30, 300 nM
Swimming frequency Ivermectin: vehicle, 0.03, 0.3, 3, 30 nM
Antagonism by picrotoxin of All combinations of two levels of ivermectin (vehicle, 30 nM) and six levels of picrotoxin
ivermectin on swimming frequency (0, 1, 10, 100, 1000, 10000 nM)
Crawling frequency and speed Ivermectin: 0, 10, 30, 100, 300 nM
Antagonism by picrotoxin of All combinations of two levels of ivermectin ivermectin on crawling frequency and (vehicle, 300 nM) and six levels of speed picrotoxin (0, 1, 3, 10, 30, 100 µM)
Treatment was carried out in covered glass Petri dishes (9 cm in diameter, 2 cm in
depth) with one worm per container of 100 ml (for swimming and reversal tests) or 50 ml
(for other tests) solution. Individual worms were randomly assigned to the treatments.
Each treatment was replicated ten to 21 times. In swimming and reversal tests, the
worms were examined directly in the treatment dishes. In other tests (swimming
frequency, crawling and electrophysiology), the worms were quickly rinsed twice in
distilled water and temporarily removed from the treatment dishes for behavioral or
electrophysiological testing.
Lethal effect
Fifty worms were randomly assigned to five treatments (Table 2), ten
worms/treatment. Observations were made after 24 and 72 h of exposure. Mortality was
determined as decomposition of the worms.
Behavioral testing
Swimming and Reversal
Helical swimming and body reversal behaviors in L. variegatus were studied as
previously described (Drewes 1999a). The worms' ability to initiate swimming and/or
reversal episodes was tested before and at various times after treatment (0, 1, 3 and 8 h).
-

14
In each test, a worm was touched ten times with a thin rubber probe alternately at its
anterior or posterior end to evoke reversal and swimming, respectively. The interval
between successive touches was three to five seconds. A response to a touch stimulus
was scored as successful only when the worm showed stereotypical patterns of swimming
or reversal movements.
Swimming frequency and pattern
To quantify possible effects of ivermectin on swimming frequency, a worm was
placed in the middle of a plastic Petri dish (14 cm in diameter, 2.5 cm in depth)
containing 200 ml of distilled water. Swimming responses were evoked twice by tactile
stimulation to the posterior end of the worm using a rubber probe (Drewes 1999a). The
worm was allowed to rest about 2 min after it was moved into the dish and between the
two trials. The process was recorded on VHS videotape using a video cassette recorder
(MITSUBISHI, model HS-U650) connected to a camcorder (LXI, model 934.53796290),
and replayed frame-by-frame on a video monitor (NEC, model XM-2950) after testing to
examine the swimming pattern and frequency (number of helical body waves produced
per second). Each worm's responses were measured before and at a selected time after
treatment (0 and 3 h in ivermectin-alone experiment, 0 and 1.5 h in ivermectin-picrotoxin
antagonism experiment). The ratio of the mean frequency after treatment to the mean
frequency before treatment was defined as relative swimming frequency for each worm.
When exposed to higher concentrations of ivermectin, some worms failed to swim in one
trial or both trials. In such cases, only successful trials were used to calculate relative
swimming frequency. If the worm failed twice, the failures were recorded as a separate
category. These failures were not used for calculation of mean swimming frequency.
Crawling
In the crawling test, a worm was placed next to a smooth strip of Plexiglass (180 x
40 x 6 mm) which rested on a piece of thoroughly wetted filter paper (Whatman #1).
Any excess water was removed, thus confining the worm within the surface tension of a
narrow band of water between the Plexiglass and paper. A straight rubber band (5 mm
long, 0.5 mm in diameter, attached to a wooden applicator stick) was used to brush the
worm's tail, so the worm would crawl forward in a straight line along the Plexiglass. The

15
frequency of brushing was 3.6 ± 0.1 strokes/sec (n = 20) as determined from videotape
replay. This stimulation lasted 10 - 15 sec, or until the worm had crawled 4 - 6 cm. The
same procedure was repeated once, and the worm was allowed to rest for about 1 minute
between trials. Crawling behavior was recorded on videotape, and later replayed, frame
by-frame, after testing to measure the crawling speed (distance moved per second) and
frequency (number of peristaltic waves of contraction produced per second). Each trial
consisted of one to three episodes of continuous crawling movements. Episodes that had
relatively constant crawling frequency were used to calculate speed and frequency. Only
the episode with the highest crawling speed was used for analysis. Each parameter was
measured twice, once before (0 min) and once after the treatment (15, 30, 60, 120 or 180
min in ivermectin-alone experiment, 90 min in ivermectin-picrotoxin antagonism
experiment). Each of these values represents the highest value obtained from one to six
episodes performed by each worm. Relative crawling frequency and relative crawling
speed were defined as previously described for relative swimming frequency.
Electrophysiological testing
Techniques for noninvasive electrophysiological recording were used as previously
described (Drewes, 1984; Zoran and Drewes, 1987; Rogge and Drewes, 1993). Briefly, a
worm was placed next to a smooth strip of Plexiglass ( 4 x 1 cm) on a printed circuit
board recording grid. Excess water was removed, thus trapping the worm in surface
tension along a narrow band of water between the Plexiglass and electrode grid. The
worm's giant fiber system was activated indirectly through sensory stimulation or
directly through electric stimulation. Evoked spikes, as well as muscle potentials, were
detected by two pairs of recording electrodes. Signals were amplified, filtered and
displayed as two channels on a digital oscilloscope (TENMA, model 72-915 20MHz).
Conduction velocity of giant fibers
The worms' medial and lateral giant fiber (MGF and LGF) systems were activated
by tactile stimulation to the anterior and posterior ends of the worms, respectively. Giant
fiber conduction velocity was measured at a mid-body location over a 10 mm conduction
distance. To obtain velocity, conduction distance was divided by conduction time, as

16
indicated on the oscilloscope screen by the peak-to-peak interval between spikes in the
two recording channels. Each worm was measured before and after the treatment (0 and
3 h). Mean velocity (five measurements per worm) was then converted to relative
conduction velocity, which was defined as the ratio of the mean velocity at any time after
treatment compared to the mean velocity in the same worm before treatment. Therefore,
by definition, the relative velocity before treatment in each worm was 1.0.
Muscle potentials
Using one pair of metallic electrodes on the grid surface, twin pulses ( duration 100
µs, inter-pulse interval 10 ms) from an electronic stimulator (Model SD9, Grass Medical
Instruments, Quincy, MA) were applied to the anterior end of a worm. The voltage of the
pulse was adjusted so that only MGF spikes, but not LGF spikes, were evoked. Peak-to
peak amplitude of the muscle potential associated with the second of the two evoked
MGF spikes was measured on the oscilloscope screen. To account for variation in signal
attenuation, muscle potential amplitudes were referenced to the average amplitudes of the
all-or-none MGF spikes. Usually, the ratio between the amplitude of the muscle potential
and the amplitude of the MGF spike is 0.72 ± 0.08 (n = 10). Duplicate measurements of
this ratio from each worm were obtained before and after treatment (0 and 3 h), and then
expressed as a relative value (mean ratio after treatment / mean ratio before treatment =
relative muscle potential).
Data analysis
Mean lethal concentration (LC50) and 95% confidence interval were calculated using
the method described by Well (Well 1952). Mean inhibitory concentrations (IC50) were
calculated using a computer program (phrmcalc.bas).
In all cases, including figures, parametric data were expressed as means ± SEM and
analyzed by ANOV A. The conservative F value was used to establish significance for
the treatment effect. Then the least significant difference test was used to determine
significance of each concentration level. In picrotoxin and ivermectin antagonism
experiments, data were analyzed using the SAS Proc General Linear Means (GLM)
procedure. Significance of interaction between ivermectin and picrotoxin was

17
established using two-way factorial analysis. Simple effect comparisons were then used
to evaluate significance of each concentration level of picrotoxin. x2 -test was used to
evaluate significance of the non-parametric swimming pattern change. The significance
level was set at P < 0.05.
Results
Lethal effect
lvermectin concentrations of 560 nM or higher were lethal to worms (Fig. 3). LCso
at 72 h post-ivermectin treatment was 560 nM (95% confidence interval: 440 - 720 nM).
A characteristic of ivermectin's lethal effect in L. variegatus was that the concentration
range for 0 - 90% death was narrow, only representing a three-fold difference. It was
also evident that onset of ivermectin-induced mortality in L. variegatus was rather
delayed. At a concentration of 1000 nM and 1800 nM, only one and three out of ten
treated worms, respectively, died within 24 h of exposure. Most worms (eight and six,
respectively) died between 24 and 72 h after treatment. In another experiment, ten
10 Cl)
..c +-' 8 cu Q)
0 6 -0 I.... Q) 4 ..c E :::::, 2 z
0
0.0 500.0
72 hrs, LC50 = 560 nM
• <'.>.
1000.0
lvermectin (nM)
1500.0
• 6.
24 hrs
2000.0
Figure 3. Lethal effect of ivermectin in Lumbriculus variegatus. Ten worms were treated in each concentration group.

18
worms were treated with 2,400 nM (extrapolated 72 h LC99) ivermectin for 8 h, and then
transferred to distilled water. All of them recovered.
Sublethal effects
Our subsequent experiments focused on ivermectin's sublethal effects (0.03 nM -
300 nM). During the first hour after exposure to 300 nM ivermectin, worms became
flaccidly paralyzed. The worms ' bodies became thinner and longer. Spontaneous
movement was totally inhibited, as well as swimming, reversal and crawling. In worms
exposed to lower concentrations of ivermectin, the effects were similar, but less extreme
and more delayed in onset. In contrast to the ivermectin effects on slow locomotor
movements, rapid escape reflexes appeared less affected by these exposures (see last
section ofresults).
Effect of ivermectin on locomotor behaviors controlled by non-giant
interneuron pathways
Swimming and reversal
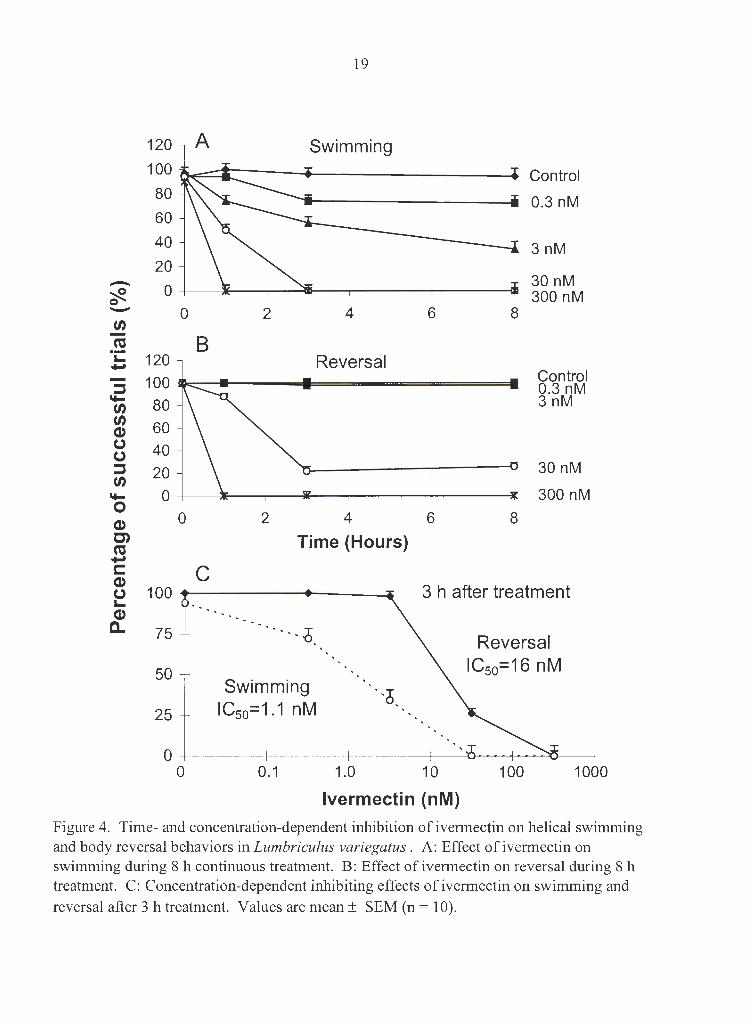
Ivermectin inhibited helical swimming and body reversal behaviors in a time- and
concentration-dependent manner (Fig. 4). There were no observable behavioral changes
within 10 min of exposure to 300 nM ivermectin. But within 60 min of ivermectin
administration, swimming and reversal were abolished. Ivermectin at 30 nM also
abolished swimming (within 3 h) but not reversal. The IC50 at 3 h for swimming and
reversal were 1.1 nM and 16 nM, respectively.
Swimming frequency and swimming pattern
Normal swimming movements consist of a series of alternating left-handed and
right-handed, rapid, and rhythmic waves of helical body bending. Each wave rapidly
progresses from head to tail, thus propelling the worm forward (Drewes 1999a). The
swimming frequency in untreated worms was remarkably consistent, 10.7 ± 0.1 Hz (n =
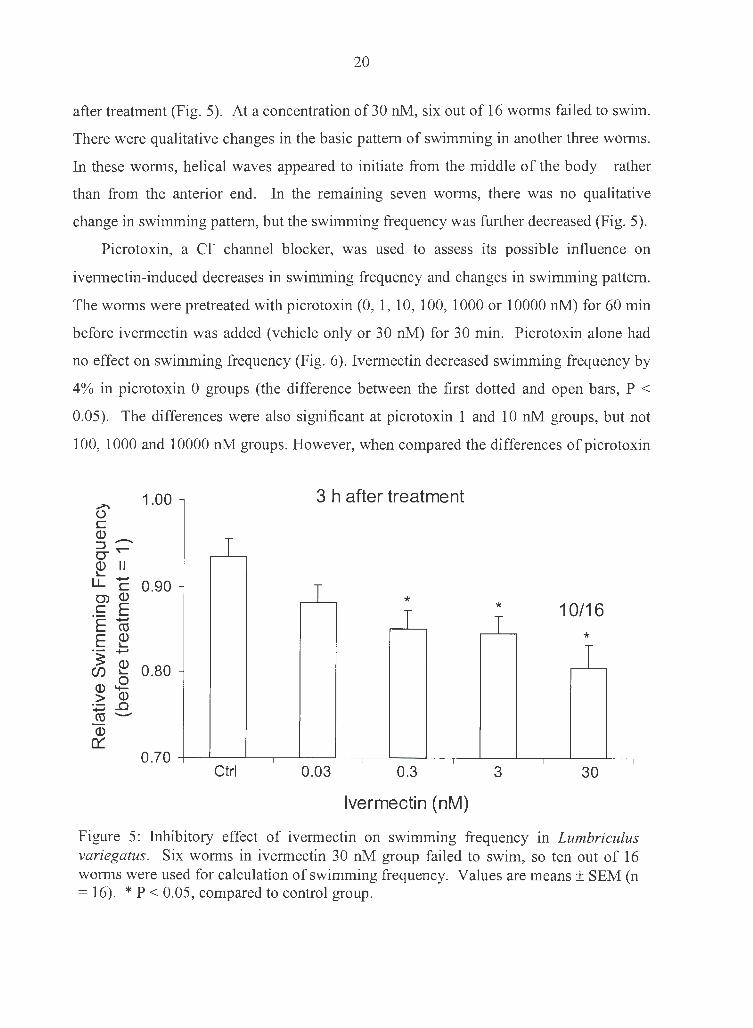
80). Ivermectin at 0.3 nM or higher significantly decreased the swimming frequency 3 h
19
120 A Swimming 100 Control 80 0.3 nM 60
40 3nM 20 - 0
30 nM ~ 300 nM 0 - 0 2 4 6 8 (/) -ca B ·;:: 120 Reversal ...,
100 Control :l 0.3 nM
II+- 80 3nM (/) (/)
60 Q) 0 40 0 :l 20 30 nM (/)
II+- 0 300 nM 0
2 4 6 8 Q) 0 C') Time (Hours) ca ..., C: C Q) 0 100 3 h after treatment :.... Q)
a.. 75 ... i Reversal
50 IC50=16 nM Swimming ·.6
25 IC50=1.1 nM
0 0 0.1 1.0 10 100 1000
lvermectin (nM)
Figure 4. Time- and concentration-dependent inhibition of ivermectin on helical swimming and body reversal behaviors in Lumbriculus variegatus . A: Effect of ivermectin on swimming during 8 h continuous treatment. B: Effect of ivermectin on reversal during 8 h treatment. C: Concentration-dependent inhibiting effects of ivermectin on swimming and reversal after 3 h treatment. Values are mean± SEM (n = 10).
20
after treatment (Fig. 5). At a concentration of 30 nM, six out of 16 worms failed to swim.
There were qualitative changes in the basic pattern of swimming in another three worms.
In these worms, helical waves appeared to initiate from the middle of the body rather
than from the anterior end. In the remaining seven worms, there was no qualitative
change in swimming pattern, but the swimming frequency was further decreased (Fig. 5).
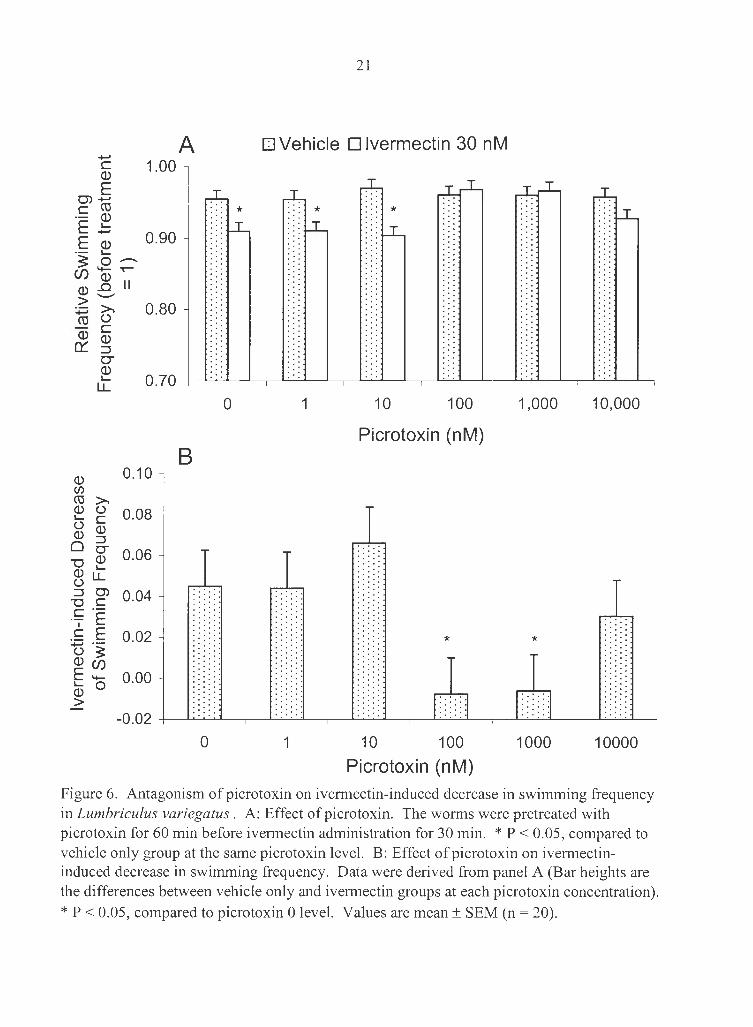
Picrotoxin, a er channel blocker, was used to assess its possible influence on
ivermectin-induced decreases in swimming frequency and changes in swimming pattern.
The worms were pretreated with picrotoxin (0, 1, 10, 100, 1000 or 10000 nM) for 60 min
before ivermectin was added (vehicle only or 30 nM) for 30 min. Picrotoxin alone had
no effect on swimming frequency (Fig. 6) . Ivermectin decreased swimming frequency by
4% in picrotoxin 0 groups (the difference between the first dotted and open bars, P <
0.05). The differences were also significant at picrotoxin 1 and 10 nM groups, but not
100, 1000 and 10000 nM groups. However, when compared the differences of picrotoxin
1.00 3 h after treatment ...,.., (.) C
(1) -:J r-CJ" (1) II I...
LL C 0.90 en © * -~ E * 10/16 E ro E (1) * I... ·- +-'
31: (1) 0.80 (/) I...
0 (1) '+-> (1)
+-' ..Q cu ---(1)
0:: 0.70
Ctrl 0.03 0.3 3 30
lvermectin (nM)
Figure 5: Inhibitory effect of ivermectin on swimming frequency in Lumbriculus variegatus. Six worms in ivermectin 30 nM group failed to swim, so ten out of 16 worms were used for calculation of swimming frequency. Values are means± SEM (n = 16). * P < 0.05, compared to control group.
A ...... 1.00 -C
Q)
E 0) ...... C C'O
E Q) !.... ......
0.90 -E Q)
-~ !....
0 -Cf) '+- 't'""" Q)
Q) ..c II
> ---...... >, 0.80 -C'O (.)
Q) C Q)
O'.'. ::J O" Q)
0.70 !....
LL
B Q) 0.10 Cl)
C'O >, Q) (.) 0.08 !.... C (.) Q) Q)
::J 0 O" 0.06 l:J Q)
!.... Q) LL (.) ::J 0) 0.04 l:J C C E I C E 0.02 t5 -~ Q) (./) E ._ 0.00 !.... 0 Q) >
-0.02
0
21
• Vehicle • lvermectin 30 nM
* * * LI... LI...
I ' ' '
0 1 10 100
Picrotoxin (nM)
*
1 10 100
Picrotoxin (nM)
T J_ r-:-:-:-
1,000
*
1000
' '
10,000
10000
Figure 6. Antagonism of picrotoxin on ivermectin-induced decrease in swimming frequency in Lumbriculus variegatus . A: Effect of picrotoxin. The worms were pretreated with picrotoxin for 60 min before ivermectin administration for 30 min. * P < 0.05, compared to vehicle only group at the same picrotoxin level. B: Effect of picrotoxin on ivermectininduced decrease in swimming frequency. Data were derived from panel A (Bar heights are the differences between vehicle only and ivermectin groups at each picrotoxin concentration).
* P < 0.05, compared to picrotoxin 0 level. Values are mean± SEM (n = 20).
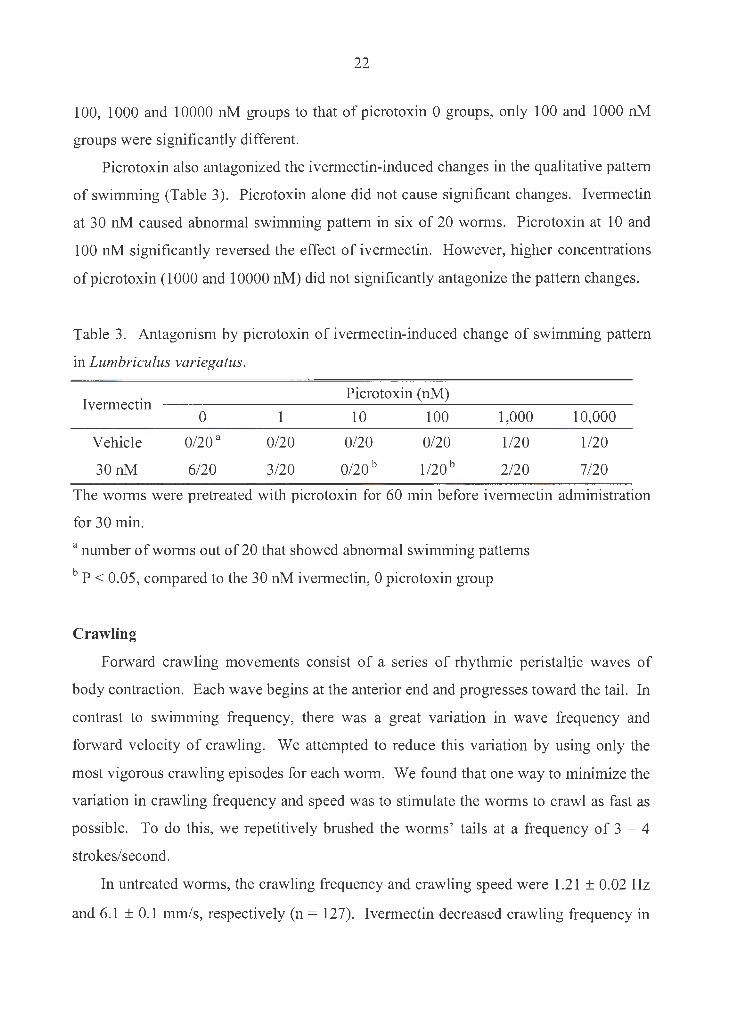
22
100, 1000 and 10000 nM groups to that of picrotoxin 0 groups, only 100 and 1000 nM
groups were significantly different.
Picrotoxin also antagonized the ivermectin-induced changes in the qualitative pattern
of swimming (Table 3). Picrotoxin alone did not cause significant changes. Ivermectin
at 30 nM caused abnormal swimming pattern in six of 20 worms. Picrotoxin at 10 and
100 nM significantly reversed the effect of ivermectin. However, higher concentrations
ofpicrotoxin (1000 and 10000 nM) did not significantly antagonize the pattern changes.
Table 3. Antagonism by picrotoxin of ivermectin-induced change of swimming pattern
in Lumbriculus variegatus.
Ivermectin Picrotoxin (nM)
0 1 10 100 1,000 10,000
Vehicle 0/20a 0/20 0/20 0/20 1/20 1/20
30nM 6/20 3/20 0/20 b 1/20 b 2/20 7/20
The worms were pretreated with picrotoxin for 60 min before ivermectin administration
for 30 min.
a number of worms out of 20 that showed abnormal swimming patterns
b P < 0.05, compared to the 30 nM ivermectin, 0 picrotoxin group
Crawling
Forward crawling movements consist of a series of rhythmic peristaltic waves of
body contraction. Each wave begins at the anterior end and progresses toward the tail. In
contrast to swimming frequency, there was a great variation in wave frequency and
forward velocity of crawling. We attempted to reduce this variation by using only the
most vigorous crawling episodes for each worm. We found that one way to minimize the
variation in crawling frequency and speed was to stimulate the worms to crawl as fast as
possible. To do this, we repetitively brushed the worms' tails at a frequency of 3 - 4
strokes/ second.
In untreated worms, the crawling frequency and crawling speed were 1.21 ± 0.02 Hz
and 6.1 ± 0.1 mrn/s, respectively (n = 127). Ivermectin decreased crawling frequency in

23
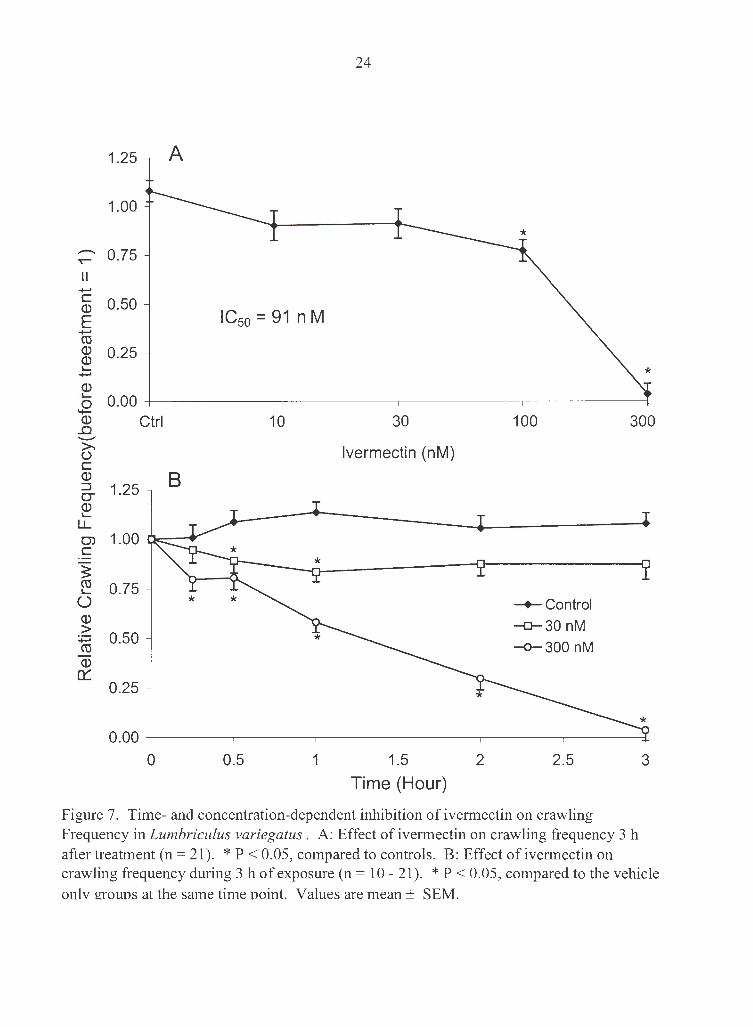
a time- and concentration-dependent manner (Fig. 7). Crawling was totally inhibited
after 3 h exposure to 300 nM ivermectin. IC50 at 3 h for crawling frequency was 91 nM.
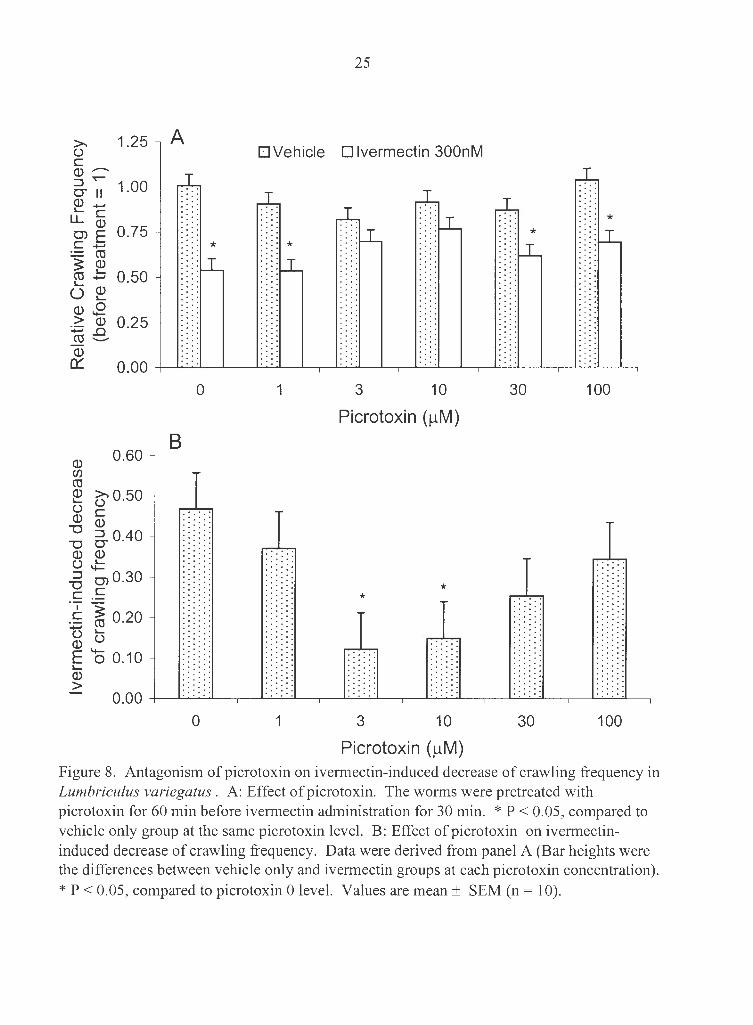
Picrotoxin antagonized the ivermectin-induced decrease in crawling frequency. The
worms were pretreated with picrotoxin (0, 1, 3, 10, 30 or 100 µM) for 60 min before
ivermectin co-exposure (vehicle only or 300 nM) for 30 min. Picrotoxin alone at all
concentrations studied did not change crawling frequency (Fig. 8). Ivermectin at 300 nM
significantly decreased crawling frequency by 47% in picrotoxin 0 group. Picrotoxin (3
and 10 µM) significantly antagonized the effect of ivermectin. However, picrotoxin at 1,
30, and 100 µM failed to do so (Fig. 8).
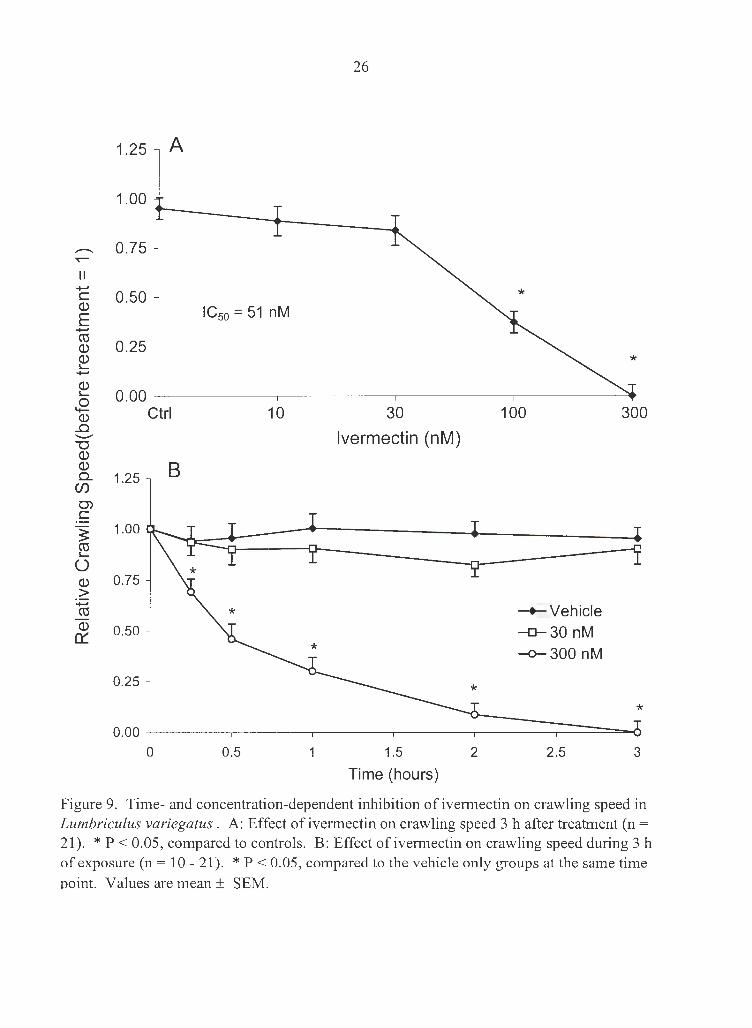
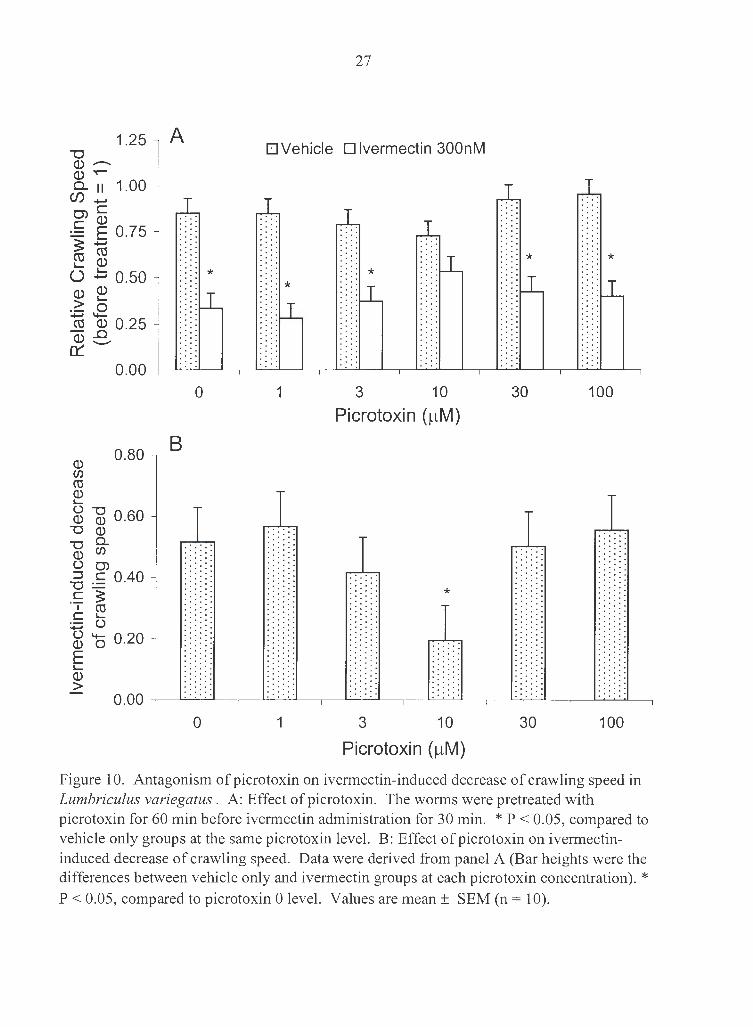
Ivermectin' s inhibitory effect on crawling speed was similar to its effect on crawling
frequency (Fig. 9). IC50 for crawling speed at 3 h was 51 nM. Ivermectin-induced
decrease in crawling speed was significantly reduced from 52% to 19% by 10 µM
picrotoxin (Fig. 10).
Effect of ivermectin on behavior controlled by giant-interneuron
pathways
The worms retained escape reflex function throughout the 3 h of treatment with 300
nM ivermectin. That is, while crawling on substrate, worms were capable of rapidly
withdrawing head or tail in response to tactile stimulation. However, responses were
different from those in normal worms in two ways. First, the escape response was not
followed by any slower locomotor movements (swimming, reversal or crawling), which
usually occurred immediately after escape responses in normal worms. Second, while
normal worms had little difficulty in rapid withdrawing in response to repeated tactile
stimulation, the escape withdrawal in treated worms was hardly noticeable after four or
five repeated stimuli. When the worms were allowed to rest for 1 or 2 min, their escape
reflex reinstated.
MGF and LGF spiking, recorded noninvasively, was used as an indicator of the
function of giant-intemeuron pathways. In untreated worms, conduction velocity of
MGF and LGF was 8.8 ± 0.1 m/s and 6.4 ± 0.1 mis (n = 60), respectively. Ivermectin up
24
1.25 A
1.00
- 0.75 T"""
II +-' C 0.50 Q)
E IC50 = 91 n M +-' Cl:l Q) 0.25 Q) I....
+-'
Q) I....
0.00 .8 Q) Ctrl 10 30 100 300
..0 --->- lvermectin (nM) (.) C Q) B ::::l 1.25 O" Q) I....
LL O') 1.00 C
~ Cl:l 0.75 I.... u ~control Q)
-o-30 nM > :.;::::; 0.50 -o--300 nM Cl:l
Q)
0:: 0.25
0.00
0 0.5 1 1.5 2 2.5 3
Time (Hour)
Figure 7. Time- and concentration-dependent inhibition of ivermectin on crawling Frequency in Lumbriculus variegatus . A: Effect of ivermectin on crawling frequency 3 h after treatment (n = 21). * P < 0.05, compared to controls. B: Effect of ivermectin on crawling frequency during 3 h of exposure (n = 10 - 21). * P < 0.05, compared to the vehicle onlv grouos at the same time ooint. Values are mean± SEM.
» 1.25 A () C Q) -:::J 'I""""
O" II 1.00 Q) +-' i,.._ C
LL Q) o, E 0.75 C +-< * ~ ~ ~.::: 0.50 u ~ Q) .E -~ © 0.25 +-' ..0 cu -----Q)
25
• Vehicle • lvermectin 300nM
* *
*
0:: 0. 00 -+-...,__.._~~~~-~~~ ~ ---~~~ ~~-~-.--~~~--.~~~~._.,
Q) (/) cu
0.60
~ ~0.50 U C Q) Q)
""O -o 6- 0.40 Q) Q) (.) .!:: :::J o, 0.30
""O C C
c ~ 0.20 t5 IQ) (.)
E o 0.10 I-Q) >
0 1
B
0 1
3 10 30 100
Picrotoxin (µM)
* *
3 10 30 100
Picrotoxin (µM) Figure 8. Antagonism of picrotoxin on ivermectin-induced decrease of crawling frequency in Lumbriculus variegatus . A: Effect of picrotoxin. The worms were pretreated with picrotoxin for 60 min before ivermectin administration for 30 min. * P < 0.05, compared to vehicle only group at the same picrotoxin level. B: Effect of picrotoxin on ivermectininduced decrease of crawling frequency. Data were derived from panel A (Bar heights were the differences between vehicle only and ivermectin groups at each picrotoxin concentration).
* P < 0.05, compared to picrotoxin 0 level. Values are mean± SEM (n = 10).
II +-' C Q.)
E +-' ro Q.) Q.) I...
+-'
~ s Q.)
..0 ---"'O Q.) Q.) 0..
Cl)
0) C
~ ro I...
0 Q.) >
+-' ro Q.)
0::
26
1.25 A
1.00
0.75
0.50 IC50 = 51 nM
0.25
0.00 ---+------~------~------~-----~
Ctrl
1.25 B
1.00
0.75
0.50
0.25
0.00
0
10
0.5
30
lvermectin (nM)
1.5
Time (hours)
100 300
-+-Vehicle -o-30 nM -o-300 nM
*
2 2.5 3
Figure 9. Time- and concentration-dependent inhibition of ivermectin on crawling speed in Lumbriculus variegatus. A: Effect of ivermectin on crawling speed 3 h after treatment (n = 21). * P < 0.05, compared to controls. B: Effect of ivermectin on crawling speed during 3 h of exposure (n = 10 - 21). * P < 0.05, compared to the vehicle only groups at the same time ooint. Values are mean ± SEM.
1.25 ""C Q) --Q) ~
0.. 11 1.00 (f) ..... 0) C C Q)
=-= E 0.75 :s: ..... cu cu I- Q)
U .b 0.50
Q) Cl)
cu Q) I-
0.25
0.80
~ -g 0.60 ""C Q)
""C Cl. Q.) Cl)
() CJ)
°"5 C 0.40 C :S:
I (0 C "-...., ()
() -Q.) 0 E I-Q)
>
0.20
A
*
0
B
0
27
• Vehicle • lvermectin 300nM
* * * : *
1 3 10 30 100
Picrotoxin (µM)
*
1 3 10 30 100
Picrotoxin (µM)
Figure 10. Antagonism of picrotoxin on ivermectin-induced decrease of crawling speed in Lumbriculus variegatus . A: Effect of picrotoxin. The worms were pretreated with picrotoxin for 60 min before ivermectin administration for 30 min. * P < 0.05, compared to vehicle only groups at the same picrotoxin level. B: Effect of picrotoxin on ivermectininduced decrease of crawling speed. Data were derived from panel A (Bar heights were the differences between vehicle only and ivermectin groups at each picrotoxin concentration). * P < 0.05, compared to picrotoxin 0 level. Values are mean± SEM (n = 10).

28
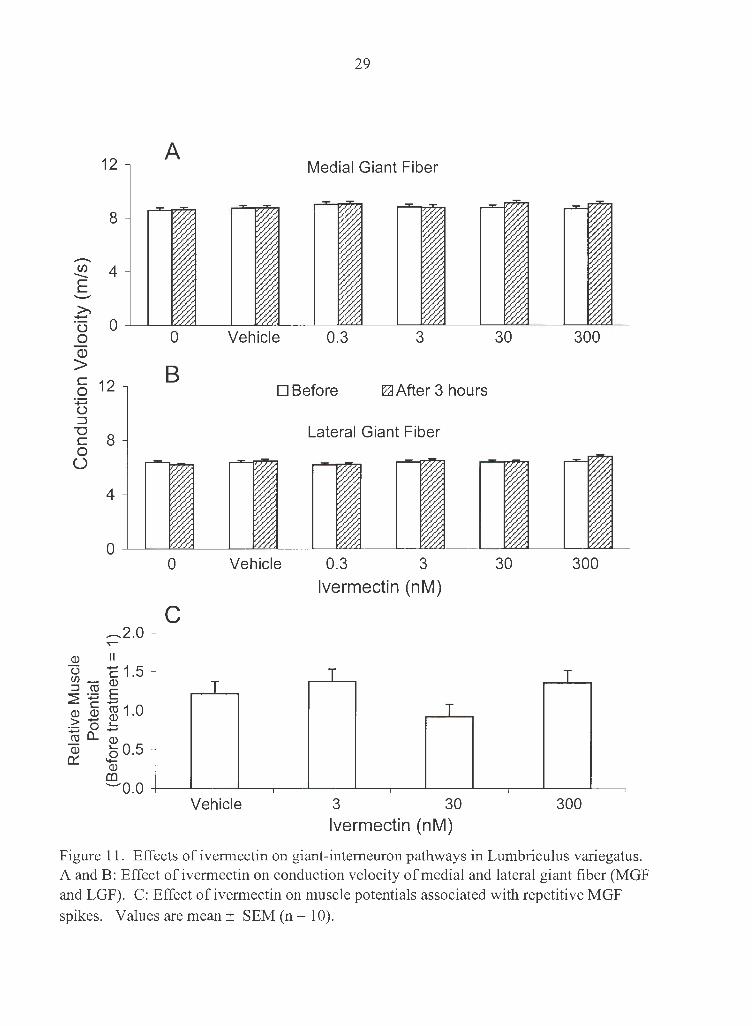
300 nM did not change either MGF or LGF conduction velocity (Fig. 1 lA and 11B).
There were also no noticeable changes related to the function of sensory inputs and motor
output associated with the giant fiber pathways, i.e. , no changes were observed in the
sensitivity to tactile stimulation, or muscle potentials associated with multiple spiking in
the MGF (Fig. 11 C).
Discussion
Numerous environmental fate and effect studies on non-target organisms have been
carried out in the development of ivermectin as an antiparasitic agent (Campbell 1989;
Halley et al. 1993). Lethal level has been determined in some non-target organisms.
Among them, Daphnia magna was most sensitive with 48 h LC50 of 0.025 ppb (= 0.03
nM, Halley et al. 1993). Fish were less sensitive (48 h LC50 for bluegill and rainbow
trout were 4.8 ppb and 3.0 ppb, respectively), and earthworms (28 d LD50 315 ppm in
soil) were relatively insensitive to ivermectin (Halley et al. 1993). Our results showed
that L. variegatus was much less sensitive to ivermectin than Daphnia magna and fish
(72 h LC50 of 560 nM ~ 490 ppb ), but much more sensitive than earthworms. However,
there were differences between the exposure method (aqueous solution vs. soil) and time
scale (3 d vs. 28 d), which makes the comparison to earthworms less meaningful.
Although the most predominant effects shown in target organisms are reduced motor
activities, there are few studies in the literature examining sub lethal effects of ivermectin
on non-target organisms. Among a few studies concerning sublethal effects of ivermectin
on non-target organisms, most have examined development and reproduction in dung
dwelling insect populations (Halley et al. 1993). The present study was the first attempt
to examine the effects of ivermectin on locomotor behaviors of a non-target invertebrate,
L. variegatus.
We developed a set of methods to measure the locomotor behaviors in L. variegatus.
These worms are benthic inhabitants in ponds, lakes, and marshes of North America and
29
12 A
Medial Giant Fiber
8
-.!:!:_ 4 E >,
+-' ·c::; 0 _j__--'--__.lL.'-LL..L_------'----"'-"-"-'u..._- --'--_.1.L.'-L..LL_ -----'-____j"---'--'--''-'------- --'------"'-"c.<..<..l._- -----'--'-"--"-"-u..__
0 Q)
> C 12 0
t5 ::::l
-g 8 0 u
4
-2.0 Cl.) 11
~ c 1.5 :::J cu Cl.)
~:.::::E C...., Cl.) Cl.) m 1.0
.2: 0 !:; cu Cl. Cl.)
Cl.) 0 0.5 0::: ,_
Cl.)
co --o.o
-
-
-
-
0 Vehicle 0.3 3 30 300
B • Before • After 3 hours
Lateral Giant Fiber
0 Vehicle 0.3 3 30 300
lvermectin (nM)
C
T T T
T
I I I I
Vehicle 3 30 300
lvermectin (nM)
Figure 11. Effects of ivermectin on giant-intemeuron pathways in Lumbriculus variegatus. A and B: Effect of ivermectin on conduction velocity of medial and lateral giant fiber (MGF and LGF). C: Effect of ivermectin on muscle potentials associated with repetitive MGF spikes. Values are mean ± SEM (n = 10).

30
Europe. They freely crawl within submerged and decaying vegetation, such as rotting
leaves and logs (Drewes and Fourtner 1989). When touched or threatened, these worms
use a variety of locomotor responses to move to safety. Their responses are context
specific. When the worm's tail is extended above the sediments, it responds to the tactile
stimulation or shadow by a rapid withdrawal ( escape response). On wet surfaces or in
confined spaces under water, the worm crawls forward or backward when touched in tail
or head regions, respectively. In open spaces under water, however, tail stimulation
evokes helical swimming, while head stimulation evokes body reversal (Drewes 1999a).
These locomotor behaviors are highly stereotyped, thus making them ideal for sublethal
toxicological tests (see Materials and Methods). The presence of giant nerve fibers that
mediate escape response (Drewes and Fourtner 1989; Drewes and Brinkhurst 1990) and
the capability of noninvasive electrophysiological testing (Drewes 1984; Zoran and
Drewes 1987; Rogge and Drewes 1993) provide us with additional advantages in
accessing sub lethal effects of neurotoxicants, such as ivermectin.
Our results showed that ivermectin impaired the worm's locomotor behaviors. The
behavioral endpoints were much more sensitive than LC50. LC50 at 72 h was 560 nM,
while 3 h IC50s for swimming, reversal, crawling speed and crawling frequency were 1. 1,
16, 51 and 91 nM, respectively. At 0.3 nM (lower than LC50 by more than three orders of
magnitude), ivermectin decreased swimming frequency significantly. The differences in
sensitivity to ivermectin suggested that motor behaviors in L. variegatus are probably
independent. Therefore, it is important to have multiple measurements to obtain an
overall picture of the effect of ivermectin. Our results demonstrated that L. variegatus is
a sensitive model for assessing sublethal locomotor behavioral effects of environmental
toxicants on aquatic invertebrates.
It was noted that the onset of paralytic effect of ivermectin in L. variegatus was
rather delayed. Within 10 min of exposure to 300 nM ivermectin, there was no
observable behavioral change. It is in sharp contrast to some other neuroactive chemicals,
e.g. 4-aminopyridine, cadmium chloride, carbofuran, chloroform and diazinon, which
caused behavioral changes in L. variegatus almost immediately upon exposure ( < 1 min)
(Rogge and Drewes 1993). It was evident that the onset of ivermectin-induced mortality

31
in L. variegatus was also delayed. A possible explanation for the slow actions of
ivermectin is the availability of the chemical to the worms. Although ivermectin is
lipophilic, it is a relatively large molecule (MW :::::: 870) compared to 4-aminopyridine,
cadmium chloride, carbofuran, chloroform and diazinon (MW: 94, 183, 221, 119 and
304, respectively). Therefore, it might be difficult for ivermectin to penetrate the worm's
cuticle. Nevertheless, it is also likely that some of the biological processes involved in
ivermectin's paralytic and lethal effects are delayed or slowly developing. These
processes are still elusive.
Although the mode of action of ivermectin is not fully understood, many studies
have demonstrated that Cl- channels are involved in its antiparasitic effects (Arena et al.
1995; Brownlee et al. 1997; Duce & Scott 1985; Kass et al. 1980; Martin et al. 1997). In
the present study we used picrotoxin, a Cl- channel blocker, to antagonize ivermectin's
effects on locomotor behaviors. Our results were consistent with the involvement of c1-
channels. However, the antagonism of picrotoxin to ivermectin was only effective in a
limited range of concentration (Fig. 6, 8, 10 and Table 2) and time scales. In our
preliminary experiments, picrotoxin failed to reverse ivermectin-induced effects on
swimming and crawling if the exposure to ivermectin lasted 3 h. A possible explanation
is that picrotoxin at high concentrations has some additional effects mediated by action
sites other than Cl- channels. Ivermectin (:-s; 300 nM) had no effects on MGF-evoked
muscle potentials (Fig. SC), but picrotoxin at 100 µM alone decreased the muscle
potentials (unpublished data). Picrotoxin (5 mM) potentiates contraction while inhibiting
voltage-dependent tubular Ca2+ current in frog skeletal muscle fibers (Jacquemond et al.
1996); it (10 µM) decreases the intensity of methylation of phospholipids
(phosphatidylethanolamine) in rat olfactory cortex (Gerasimova et al. 1993). Another
possibility is that ivermectin has action sites other than Cl- channels (Ellis et al. 1987;
Sani and Vaid 1988; Ahern et al. 1999), thus a Cl- blocker can only reduce but not
abolish the effect of ivermectin.
lvermectin-sensitive c1- channels are present in nerve and/or muscle cells in many
invertebrates, such as nematodes, insects, crustaceans, and mollusks (Cleland 1996).
Although their physiological role has not been fully determined, they are reported to be

32
involved in generating rhythmic firing of the neurons within the crustacean
stomatogastric ganglion (Cleland and Selverston, 1995, 1998). Our results showed that
ivermectin decreased swimming frequency and crawling frequency, suggesting that
ivermectin-sensitive Cl- channels are involved in the neuropathways that control
swimming and crawling.
It was interesting to observe that the escape reflex behavior controlled by giant
interneuron pathways was still intact even after treatment with 300 nM ivermectin.
Electrophysiological studies confirmed that ivermectin had no effects on the conduction
velocity of MGF or LGF, or the muscle potentials evoked during multiple firing of MGF
action potentials, suggesting that ivermectin-sensitive c1- channels are not crucially
involved in the escape reflex functions of giant interneurons. This is not surprising if one
takes a closer look at the giant interneuron system. The MGF and LGF pathways are
derived from the electrically connected large axons of interneurons in each segment.
They function as a syncytium, rapidly conducting nerve action potentials, without
interruption, along their length (Drewes 1999b ). In such a straightforward system, the
main emphasis seems to be speed and reliability; negative feedback via inhibition may be
unnecessary or inconsequential. On the other hand, some locomotor behaviors controlled
by non-giant interneuron pathways (swimming and crawling) are slower, rhythmic, and
probably subject to modulatory influence. Specific networks of neurons in an animal's
central nervous system, which control coordinated (and often rhythmic) pattern of
movements, are termed central pattern generators (Young 1989). Negative feedback and
other modulatory controls are usually utilized in such networks.
It has been reported that ivermectin has inhibitory effects on the pharyngeal muscle
through opening Cl- channels in the parasitic nematode Ascaris suum (Adelsberger et al.
1997; Brownlee et al. 1997). In Lubriculus, muscle potentials are normally associated
with repetitive MGF spikes, but our results showed no significant changes in the
appearance of these potentials following ivermectin treatment. However, our
observations were based on muscle activities recorded from the body surface. This
method is indirect, and may not sufficiently sensitive to detect subtle changes in
membrane potential of muscle fibers.

33
CONCLUSIONS
In conclusion, our results demonstrated that (1) sublethal behavioral effects were
much more sensitive endpoints than was mortality in assessing ivermectin's potential
neurobiological and ecological impacts; (2) locomotor behaviors controlled by non-giant
intemeuron pathways were sensitive to ivermectin whereas those controlled by giant
intemeurons did not appear to be affected at the concentrations studied; and (3) er channels appeared to be involved in ivermectin's inhibitory effects.

34
LITERATURE CITED
Abalis IM, Eldefrawi AT, Eldefrawi ME (1986) Actions of avermectin Bia on the
gamma-aminobutyric acidA receptor and chloride channels in rat brain. J Biachem
Taxical 1(1):69-82
Adelsberger H, Scheuer T, Dudel J (1997) A patch clamp study of a glutamatergic
chloride channel on pharyngeal muscle of the nematode Ascaris suum. Neurasci
Lett 230(3) : 183-6
Ahem GP, Junank:ar PR, Pace SM, Curtis S, Mould JA, Dulhunty AF (1999) Effects of
ivermectin and midecamycin on ryanodine receptors and the Ca2+-ATPase in
sarcoplasmic reticulum of rabbit and rat skeletal muscle. J Physial (Land) 514 (Pt
2): 313-26
Albert J, Lingle CJ, Marder E, O'Neil MB (1986) A GABA-activated chloride
conductance not blocked by picrotoxin on spiny lobster neuromuscular
preparations. Br J Pharmacal 87( 4): 771-9
Arena JP, Liu KK, Paress PS, Cully DF (1991) Avermectin-sensitive chloride currents
induced by Caenorhabditis elegans RNA in Xenopus oocytes. Mal Pharmacal
40(3): 368-74
Arena JP, Liu KK, Paress PS, Schaeffer JM, Cully DF (1992) Expression of a glutamate
activated chloride current in Xenopus oocytes injected with Caenorhabditis elegans
RNA: evidence for modulation by avermectin. Brain Res Mal Brain Res 15(3-4):
339-48
Arena JP, Liu KK, Paress PS, Frazier EG, Cully DF, Mrazik H, Schaeffer JM (1995) The
mechanism of action of avermectins in Caenorhabditis elegans: correlation between
activation of glutamate-sensitive chloride current, membrane binding, and
biological activity. J Parasital 81(2): 286-94
Aziz MA, Diallo S, Diop IM, Lariviere M, Porta M (1982) Efficacy and tolerance of
ivermectin in human onchocerciasis. Lancet 2(8291): 171-3
Aziz MA, Diallo S, Lariviere M, Diop IM, Porta IM, Gaxotte P (1982) Ivermectin in
onchocerciasis. Lancet 2(8313): 1456-7

35
Brinkhurst RO, Jamieson BGM (1971) Aquatic oligochaeta of the world. University of
Toronto Press, toronto, Canada. pp 174-187
Brownlee DJ, Holden-Dye L, Walker RJ (1997) Actions of the anthelmintic ivermectin
on the pharyngeal muscle of the parasitic nematode, Ascaris suum. Parasitology
115 ( Pt 5): 553-61
Burg RW, Miller BM, Baker EE, Birnbaum J, Currie SA, Hartman R, Kong YL,
Monaghan RL, Olson G, Putter I, Tunac JB, Wallick H, Stapley EO, Oiwa R,
Omura S (1979) Avermectins, new family of potent anthelmintic agents: producing
organism and fermentation. Antimicrob Agents Chemother 15(3): 361-7
Campbell WC, Fisher MH, Stapley EO, Albers-Schonberg G, Jacob TA (1983)
Ivermectin: a potent new antiparasitic agent. Science 221(4613): 823-8
Campbell WC, Benz GW (1984) Ivermectin: a review of efficacy and safety. J Vet
Pharmacol Ther 7(1): 1-16
Campbell WC (1989) Ivermectin and abamectin. Spriner-Verlag New York Inc., New
York, USA
Campbell WC (1991) Ivermectin as an antiparasitic agent for use in humans. Annu Rev
Microbiol 45 : 445-74
Campbell WC (1993) Ivermectin, an antiparasitic agent. Med Res Rev 13(1): 61-79
Chabala JC, Mrozik H, Tolman RL, Eskola P, Lusi A, Peterson LH, Woods MF, Fisher
MH, Campbell WC, Egerton JR, Ostlind DA (1980) Ivermectin, a new broad
spectrum antiparasitic agent. J Med Chem 23(10): 1134-6
Cleland TA, Selverston AI (1995) Glutamate-gated inhibitory currents of central pattern
generator neurons in the lobster stomatogastric ganglion. J Neurosci 15(10): 6631-
6639
Cleland TA (1996) Inhibitory glutamate receptor channels. Mol Neurobiol 13: 97-135
Cleland TA, Selverston AI (1998) Inhibitory glutamate receptor channels in cultured
lobster stomatogastric neurons. J Neurophysiol 79(6): 3189-3196
Cully DF, Paress PS (1991) Solubilization and characterization of a high affinity
ivermectin binding site from Caenorhabditis elegans. Mo! Pharmacol 40(2): 326-
32

36
Cully DF, Vassilatis DK, Liu KK, Paress PS, Van der Ploeg LH, Schaeffer JM, Arena JP
(1994) Cloning of an avermectin-sensitive glutamate-gated chloride channel from
Caenorhabditis elegans. Nature 371(6499): 707-11
Drewes CD (1984) Escape reflexes in earthworms and other annelids. In Eaton RC (ed)
Neural mechanisms of startle behavior. Plenum Press, New York, USA. pp 43-91
Drewes CD, Fourtner CR (1989) Hindsight and rapid escape in a freshwater oligochaete.
Biol Bull 177: 363-371
Drewes CD, Brinkhurst RO (1990) Giant nerve fibers and rapid escape reflexes in newly
hatched aquatic oligochaetes, Lumbriculus variegatus (Family Lumbriculidae).
Invert Reprod Devel 17: 91-95
Drewes CD, Fourtner CR (1990) Morphallaxis in an aquatic oligochaete, Lumbriculus
variegatus: Reorganization of escape reflexes in regenerating body fragments.
Devel Biol 128: 94-103
Drewes CD (1999a) Helical sw1mmmg and body reversal behaviors in Lumbriculus
variegatus (Annelida: Clitellata: Lumbriculidae). Hydrobiologia 406(?): 263-268
Drewes CD (1999b) Non-invasive recording of giant nerve fiber action potentials from
freely moving oligochaetes. In Karcher SJ(ed) Tested Studies for Laboratory
Teaching Association for Biology Laboratory Education (ABLE), Volume 20, pp.
45-62
Drewes C, Cain K (1999) As the worm turns. Am Biol Teacher 61(6): 438-442
Duce IR, Scott RH (1985) Actions of dihydroavermectin B1a on insect muscle. Br J
Pharmacol 85(2): 395-401
Egerton JR, Ostlind DA, Blair LS, Eary CH, Suhayda D, Cifelli S, Riek RF, Campbell
WC (1979) Avermectins, new family of potent anthelmintic agents: efficacy of the
B1a component. Antimicrob Agents Chemother 15(3): 372-8
Ellis C, Nathwani B, Morrice N, Parker P, Evans FJ, Aitken A (1987) Ivermectin: an
inhibitor of protein kinase C: a potential target enzyme for onchocerciases
chemotherapy. Brit J Pharmacol 91: 22
Fisher MH, Mrozik H (1989) Chemistry. In Campbell WC (ed) Jvermectin and
abamectin Springer-Verlag New York Inc. New York, USA pp. 1-23

37
Fritz LC, Wang CC, Gorio A (1979) Avermectin B1a irreversibly blocks presynaptic
potentials at the lobster neuromuscular junction by reducing muscle membrane
resistance. Proc Natl Acad Sci USA 76: 2062-2066
Gerasimova IA, Emel'yanov MA, Mokrushin AA, Pavlinova LI (1993) The influence of
picrotoxin on the methylation of phospholipids of the olfactory cortex of the brain
ofrats. Neurosci Behav Physiol 23(1): 63-5
Halley BA, jacob TA, Lu A YH (1989) The environment impact of the use of ivermectin:
environmental effects and fate . Chemosphere 19(7-8): 1543-1563
Halley BA, VandenHeuvel WJ, Wislocki PG (1993) Environmental effects of the usage
of avermectins in livestock. Vet Parasitol 48(1-4): 109-25
Huang J, Casida JE (1997) Avermectin B1a binds to high- and low-affinity sites with dual
effects on the gamma-aminobutyric acid-gated chloride channel of cultured
cerebellar granule neurons. J Pharmacol Exp Ther 281(1): 261-6
Jacquemond V, Oetliker H, Rougier 0, Takeda K (1996) Picrotoxin potentiates
contraction while inhibiting Ca current but increasing birefringence signal in frog
skeletal muscle fibers. Jpn J Physiol 46(1): 99-104
Kass IS, Stretton AO, Wang CC (1980) The effects of avermectin and drugs related to
acetylcholine and 4-aminobutyric acid on neurotransmission in Ascaris suum. Mo!
Biochem Parasitol 13(2): 213-25
Kass IS, Wang CC, Walrond JP, Stretton AO (1984) Avermectin B 1a, a paralyzing
anthelmintic that affects intemeurons and inhibitory motoneurons in Ascaris. Proc
Natl Acad Sci USA 77(10):6211-5
Krusek J, Zemkova H (1994) Effect of ivermectin on gamma-aminobutyric acid-induced
chloride currents in mouse hippocampal embryonic neurones. Eur J Pharmacol
259(2):121-8
Lankas GP, Gordon LR (1989) Toxicology. In Campbell WC (ed) Ivermectin and
abamectin Springer-Verlag New York Inc. New York, USA pp. 89-112
Martin RJ (1993) Neuromuscular transmission in nematode parasites and antinematodal
drug action. Pharmacol Ther 58(1): 13-50

38
Martin RJ, Robertson AP, Bjorn H (1997) Target sites of anthelmintics. Parasitology
114 Suppl: S 111-24
Mellin TN, Busch RD, Wang CC (1983) Postsynaptic inhibition of invertebrate
neuromuscular transmission by avermectin Bia- Neuropharmacology 22(1): 89-96
Miller TW, Chaiet L, Cole DJ, Cole LJ, Flor JE, Goegelman RT, Gullo VP, Joshua H,
Kempf AJ, Krellwitz WR, Monaghan RL, Ormond RE, Wilson KE, Albers
Schonberg G, Putter I (1979) Avermectins, new family of potent anthelmintic
agents: isolation and chromatographic properties. Antimicrob Agents Chemother
15(3): 368-71
Mrozik H, Eskola P, Reynolds GF, Arison BH, Smith GM, Fisher MH (1988)
Photoisomers of avermectins. J Org Chem 53: 1820-1923
Ottesen EA, Campbell WC (1994) Ivermectin in human medicine. J Antimicrob
Chemother 34(2): 195-203
Pivnichny JV, Shim JS, Zimmerman LA (1983) Direct determination of avermectins in
plasma at nanogram levels by high-performance liquid chromatography. J Pharm
Sci 72(12) : 1447-50
Pivnichny JV, Arison BH, Preiser FA, Shim JSK, Mrozik H (1988) Base-catalyzed
isomerisation of avermectins. J Agric Food Chem 36: 826-828
Pulliam JD, Preston JM (1989) Safety of ivermectin in Target animals. In Campbell WC
(ed) Jvermectin and abamectin Springer-Verlag New York Inc. New York, USA
pp. 149-161
Ridsdill-Smith TJ (1988) Survival and reproduction of Musca vetustissima Walker
(Diptera: Muscidae) and a scarabaeine dung beetle in dung of cattle treated with
avermectin B 1. J Aust Entomol Soc 27: 175-178
Ridsdill-Smith TJ (1993) Effects of avermectin residues in cattle dung on dung beetle
(Coleoptera: Scarabaeidae) reproduction and survival. Vet Parasitol 48(1-4) : 127-
37
Rogge RW, Drewes CD (1993) Assessing sublethal neurotoxicity effects m the
freshwater oligochaete, lumbriculus variegatus. Aquat Toxicol 26: 73-90

39
Roncalli RA (1989) Environmental aspects of ivermectin usage in livestock: effects on
cattle dung fauna. In Campbell WC ( ed), Ivermectin and Abamectin, Spriner
Verlag New York Inc., New York, USA, pp 173-181
Sani BP, Vaid A (1988) Specific interaction of ivermectin with retinal-binding protein
from filarial parasites. Biochem J249(3): 929-32
Schaeffer JM, Haines HW (1989) A vermectin binding in Caenorhabditis elegans. A two
state model for the avermectin binding site. Biochem Pharmacol 38(14): 2329-38
Soll MD (1989) Use of ivermectin in laboratory and exotic mammals and in birds, fish,
and reptiles. In Campbell WC (ed) Ivermectin and abamectin Springer-Verlag
New York Inc. New York, USA pp. 260-286
Strong L, Wall R (1988) Ivermectin in cattle treatment: nonspecific effects on
pastureland ecology. Aspects Appl Biol 17: 231-238
Strong L (1993) Overview: the impact of avermectins on pastureland ecology. Vet
Parasitol 48(1-4): 3-17
Wall R, Strong L (1987) Environmental consequences of treating cattle with the
antiparasitic drug ivermectin. Nature 327(6121): 418-21
Wardhaugh KG, Rodriguez-Menendez H (1988) The effects of the antiparasitic drug,
ivermectin, on the development and survival of the dung-breeding fly, Onthelia
cornicina (F.) and the scarabaeine dung beetles, Copris hispanus L., Bubas bubalus
(Oliver) and Onitis belial F. J Appl entomol 106: 381-389
Wardhaugh KG, Holter P, Whitby WA, Shelley K (1996) Effects of drug residues in the
faeces of cattle treated with injectable formulations of ivermectin and moxidectin
on larvae of the bush fly, Musca vetustissima and the house fly, Musca domestica.
Aust Vet J74(5): 370-4
Wardhaugh KG, Mahon RJ (1998) Comparative effects of abamectin and two
formulations of ivermectin on the survival of larvae of a dung-breeding fly. Aust
Vet J76(4): 270-2
Weil CS (1952) Tables for convenient calculation of median-effective dose (LD50 or
ED50) and instructions in their use. Biometrics 8: 249-263

40
Young D (1989) Nerve cells and animal behavior. Cambridge University Press, New
York, USA
Zoran MJ, Drewes CD (1987) Rapid escape reflexes in aquatic oligochaetes: variation in
design and function of evolutionarily conserved giant fiber systems. J Comp
Physiol A 161: 729-738

41
ACKNOWLEDGEMENTS
I would like to express my sincere gratitude and deepest appreciation to my major
professor, Dr. Walter H. Hsu, and my POS committee member, Dr. Charles D. Drewes,
for their guidance, support, and continuous help in solving the problems I faced during
my master program at Iowa State University. I will be forever grateful for their efforts. I
would also like to thank, Dr. Gary J. Atchison and Dr. Joel R Coats for their serving on
my POS committee and for their valuable time reading and correcting this thesis. I would
also like to thank Tammy J. Benson for her assisting in statistical analysis. My thanks
are extended to my fellow graduate students Dr. Sirintom Yibchokanun (Lek), Henrique
Cheng, and Ehab A. H. Abu Basha. Also I wish to thank Mr. Laverene Escher for his
technical assistance and friendship.
Related Documents










