Le potentiel de compétition apparente entre l’orignal (Alces alces) et le caribou forestier (Rangifer tarandus caribou) varie le long d’un gradient latitudinal Mémoire Caroline Gagné Maîtrise en biologie Maître ès sciences (M.Sc.) Québec, Canada © Caroline Gagné, 2014

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Le potentiel de compétition apparente entre l’orignal (Alces alces) et le caribou forestier (Rangifer tarandus
caribou) varie le long d’un gradient latitudinal
Mémoire
Caroline Gagné
Maîtrise en biologie Maître ès sciences (M.Sc.)
Québec, Canada
© Caroline Gagné, 2014

iii
RÉSUMÉ
À la suite d’une coupe forestière, les parterres en régénération peuvent subir un
enfeuillement favorable à l’orignal. L’accroissement des populations de loups qui s’en suit
affecte alors les populations du caribou forestier. Ce projet vise à évaluer si la contribution
de la coupe forestière à cette compétition apparente varie selon un gradient latitudinal.
Nous avons modélisé la sélection d’habitat d’orignaux, de loups et de caribous. Après la
prise en compte de l’âge de la coupe, l’enfeuillement des parterres diminuait avec la
latitude. La sélection des parterres de coupe par les orignaux et les loups augmentait en
fonction de leur enfeuillement. Les caribous évitaient les coupes, particulièrement celles
riches en feuillus. La cooccurrence de l’orignal et du loup dans les coupes augmente donc
en fonction de l’enfeuillement. Ainsi, la contribution de la coupe forestière à la compétition
apparente dans le système orignal-loup-caribou devrait diminuer vers le nord de la forêt
boréale.


v
ABSTRACT
Moose benefit from deciduous vegetation that becomes increasingly available during the
regeneration of logged forests. This numerical response of moose populations can lead to
an increase in gray wolf abundance, which impacts forest-dwelling caribou populations.
This study evaluates if the contribution of logging to such apparent competition could vary
along the latitudinal axis. We modelled habitat selection behaviour of moose, wolves and
caribous monitored by telemetry. After controlling for time since harvest, deciduous
vegetation availability decreased with increasing latitude. Moose and wolves avoided
cutovers with low deciduous vegetation availability, while they selected harvested stands
with high deciduous vegetation availability. Caribou avoided cutovers, and their avoidance
increased with the level of deciduous regeneration. The co-occurrence between moose and
wolves in cutovers increased with deciduous vegetation availability. Therefore, the
contribution of logging to the apparent competition in the moose-wolf-caribou system
should be stronger in southern than in northern boreal forest.


vii
TABLE DES MATIÈRES
RÉSUMÉ .............................................................................................................................. iii ABSTRACT ........................................................................................................................... v TABLE DES MATIÈRES .................................................................................................... vii LISTE DES TABLEAUX ..................................................................................................... ix LISTE DES FIGURES .......................................................................................................... xi REMERCIEMENTS ........................................................................................................... xiii AVANT-PROPOS ................................................................................................................ xv INTRODUCTION .................................................................................................................. 1
Les gradients géographiques ........................................................................................... 1
Les perturbations anthropiques ....................................................................................... 2
Le caribou forestier comme sujet d’étude ....................................................................... 4
Objectif de l’étude ........................................................................................................... 7
Approche empirique ........................................................................................................ 8 CHAPITRE PRINCIPAL ....................................................................................................... 9
Résumé .......................................................................................................................... 10
Abstract ......................................................................................................................... 12
Introduction ................................................................................................................... 14
Methodology ................................................................................................................. 16
Results ........................................................................................................................... 21
Discussion ..................................................................................................................... 28
Acknowledgements ....................................................................................................... 31
References ..................................................................................................................... 31 CONCLUSION GÉNÉRALE .............................................................................................. 35
Perspectives ................................................................................................................... 40 BIBLIOGRAPHIE GÉNÉRALE .......................................................................................... 41


ix
LISTE DES TABLEAUX
Table 1. Parameter estimates of a generalized linear model characterizing the influence of
time since harvest and latitude on the weighted mean Enhanced Vegetation Index of
cutovers in the boreal forest of the Côte-Nord region, Québec, Canada. For each
covariate, model coefficients (β), standard errors (SE) and P-values are presented. ... 23
Table 2. Parameter estimates of the top-ranking Resource Selection Functions describing
the annual habitat selection by 15 moose, 11 wolves and 37 forest-dwelling caribou
that were monitored with GPS collars in Québec’s boreal forest, Canada. Each of the
model covariates is presented with their selection coefficients (β ± SE). .................... 25
Table 3. Pourcentage de chaque milieu composant les trois régions (Nord : 5763481 à
5858391 ; Milieu : 5499818 à 5763481 ; Sud : 5312850 à 5499818) dans l’aire d’étude
selon, ainsi que le pourcentage d’utilisation de chacun des milieux par l’orignal, le
loup gris et le caribou forestier. .................................................................................... 38


xi
LISTE DES FIGURES
Figure 1. Location of the study area within the Côte-Nord region of Québec’s boreal forest,
Canada (red line). White line delimits the area where radio-collared moose, wolves
and caribou were tracked. Three reference points of the latitudinal gradient (South:
5352900 UTM; Middle: 5556575 UTM; North: 5760253 UTM) are represented as
reference for some of the analyses and results reported. .............................................. 17
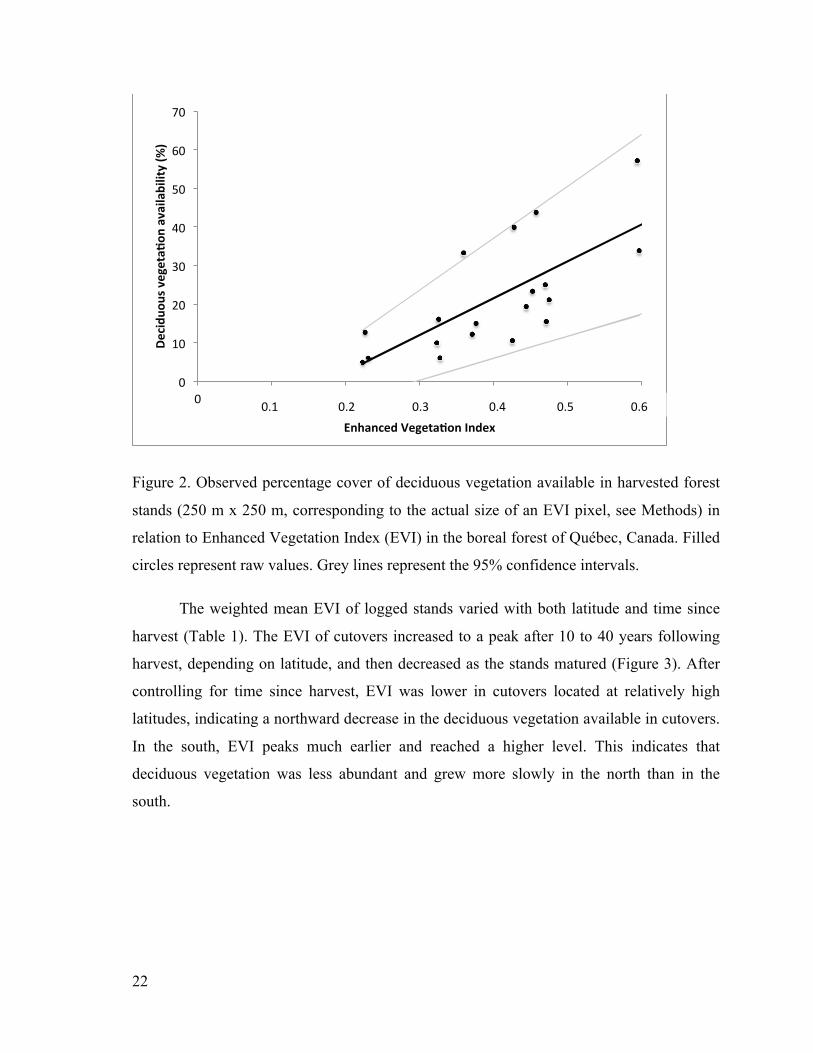
Figure 2. Observed percentage cover of deciduous vegetation available in harvested forest
stands (250 m x 250 m, corresponding to the actual size of an EVI pixel, see Methods)
in relation to Enhanced Vegetation Index (EVI) in the boreal forest of Québec,
Canada. Filled circles represent raw values. Grey lines represent the 95% confidence
intervals. ........................................................................................................................ 22
Figure 3. The influence of time since harvest and latitude on the weighted mean Enhanced
Vegetation Index (EVI) of cutovers in the boreal forest of the Côte-Nord region,
Québec, Canada. The South region corresponds to the first quartile of latitudes; the
Middle region to the second and third quartile while the North region corresponds to
the fourth quartile of the 139,725 cutovers studied. Filled coloured diamonds are mean
values of each 10-year class of every region. Lines represent the predicted
relationships from generalised linear models between EVI and time since harvest, for
stands from different latitudes (South: 5,419,517 UTM; Middle: 5,527,884 UTM;
North: 5,619,927 UTM, see Figure 1). ......................................................................... 24
Figure 4: Variation in the relative probability of occurrence in 10 land cover types, as a
function of the Enhanced Vegetation Index (EVI), a proxy for the availability of
deciduous vegetation, for A) moose, B) wolf and C) caribou in the boreal forest of the
Côte-Nord region, Québec, Canada. ............................................................................. 27
Figure 5: Variation in the relative probability of co-occurrence in 10 land cover types, as a
function of the Enhanced Vegetation Index (EVI), a proxy for the availability of
deciduous vegetation, for A) moose and wolves, and B) caribou and wolves in the

xii
boreal forest of the Côte-Nord region, Québec, Canada. .............................................. 28

xiii
REMERCIEMENTS
Je tiens d’abord à remercier mon directeur de recherche, Daniel Fortin, de m’avoir offert un
projet aussi intéressant. Ton soutien et ta patience m’ont permis de mener à terme ce projet
ayant des implications directes sur la conservation du caribou forestier. Ton dévouement
pour la recherche de qualité et ton désir de toujours repousser les barrières établies sont
contagieux.
J’aimerais aussi remercier Julien Mainguy, mon codirecteur. Tu as toujours été là
pour m’appuyer et m’apporter de nouvelles perspectives malgré le fait que tu attaquais un
nouvel emploi chargé au Ministère des Forêts, de la Faune et des Parcs. Ton aide précieuse
durant le court mais intense séjour de terrain a été très appréciée.
Un gros merci à Philippe Goulet qui m’a aidé à obtenir et démêler toutes les notions
de foresterie qui m’étaient complètement inconnues. Je ne crois pas que Julien et moi
aurions réussi à terminer cette semaine de terrain sans ta présence et tes conseils de
forestier. Merci aussi à Pierre Racine pour son aide avec PostGIS, un logiciel très utile mais
un peu complexe pour des débutants en informatique comme moi. Tu as toujours pris le
temps pour que je comprenne bien ce que je faisais au lieu de seulement m’envoyer des
codes à rouler.
Cette expérience aux études supérieures n’aurait certainement pas été la même sans
la présence de mes collègues de laboratoire : Chrystel Losier, Jerod Merkle, Marie Sigaud,
Hélène Le Borgne, Orphé Bichet, Olivia Tardy, Julie Martineau, Angélique Dupuch, Tom
Mason et Florian Barnier. Un merci tout particulier à Chrystel dont le projet a pavé la voie
au mien et qui a répondu avec patience à mes milliers de questions. Merci à Jerod pour ton
aide avec R et tes nombreux commentaires. Merci à toi et Marie de m’avoir aussi bien
accueilli dans votre bureau et de m’avoir intégré à vos conversations de bisons. Un gros
merci à Julie avec qui j’ai vécu cette expérience du début à la fin, pour tes nombreux
encouragements dans les moments les plus durs et ton enthousiasme sans fin lors des
réussites. Finalement, un grand merci à vous tous pour les nombreux moments partagés que

xiv
ce soit autour de notre lunch ou d’une bière bien méritée!
Merci à Jean-Pierre Tremblay et à David Pothier pour leurs conseils suite à la
lecture de mon proposé. Aussi, un grand merci à Steeve Côté et encore une fois à David
Pothier pour leurs commentaires suite à l’évaluation de mon mémoire.
Finalement, un très grand merci à ma famille et mes amis à Montréal qui ont
su faire la différence pendant mes absences. Merci à ma petite sœur, MJ, qui m’a toujours
poussé à continuer malgré les difficultés et qui en sait maintenant autant que moi sur le
caribou, le loup et l’orignal à force de lire et relire mes différentes épreuves. Merci à ma
grande sœur et mes parents pour leurs encouragements à toujours aller plus loin. Merci
papa d’avoir transmis à tes filles ton amour pour les animaux et la nature. Un très gros
merci à Nathalie et Richard qui ont pris soin de mes chevaux à Montréal pendant la
semaine alors que j’étais à Québec. Sans vous, je n’aurais pu dormir aussi tranquillement
alors que j’étais à 3 h de mes trois petits monstres. Merci à toi maman de t’être occupée de
Brette et d’avoir inclus Goliath dans ta routine (même si tu le trouvais très impressionnant).

xv
AVANT-PROPOS
Ce mémoire de maitrise vise à évaluer si la contribution de la coupe forestière à la
compétition apparente affectant le caribou forestier varie le long d’un axe latitudinal. Il
contient trois parties : une introduction générale, un chapitre principal présenté sous forme
d’article scientifique en anglais et une conclusion générale. Le chapitre principal a été
rédigé en collaboration avec les personnes suivantes : mon directeur de recherche, le Dr
Daniel Fortin, professeur au département de biologie de l’Université Laval et le Dr Julien
Mainguy, biologiste au Ministère des Forêts, de la Faune et des Parcs. Étant donné mon
engagement à chacune des étapes du projet, je suis la première auteure de l’article.

1
INTRODUCTION
Les gradients géographiques
Les gradients géographiques décrivant la variation spatiale de la disponibilité de la
végétation sont des facteurs importants à considérer lors de l’étude de processus
biologiques (Connell et al., 1964). Par exemple, sur le territoire européen, il a été démontré
que l’abondance de chevreuils (Capreolus capreolus) fluctue selon un gradient longitudinal
en corrélation avec la productivité du couvert de végétation (Melis et al., 2009). La densité
des populations de chevreuils décline d’ouest en est passant de 5380 individus/100 km2 à 11
individus/100 km2. Les gradients géographiques de productivité d’un système donné
peuvent aussi influencer le comportement de certaines espèces. En effet, Singh et al. (2012)
ont démontré qu’en Fennoscandinavie, les déplacements des orignaux (Alces alces) étaient
influencés par un gradient latitudinal ; les orignaux retrouvés au nord ont tendance à migrer
annuellement étant donné le manque de ressources alimentaires alors que les orignaux
retrouvés plus au sud, où le milieu est plus productif et le climat plus doux, choisissent un
mode de vie plutôt sédentaire. Les gradients géographiques de disponibilité de la végétation
peuvent donc affecter l’abondance et les comportements de quête alimentaire de différentes
espèces, notamment chez les herbivores.
Les interactions trophiques, particulièrement celles liées à la prédation, peuvent
aussi être influencées par les gradients géographiques de disponibilité de la végétation. Un
exemple classique de cet effet est la variation de l’amplitude des cycles d’abondances de
campagnols des champs (Microtus agrestis), étudiée par Hanski et al. (2001) en
Fennoscandinavie. Au nord, les prédateurs spécialistes concentrent leurs efforts de
recherche de nourriture sur ces petits rongeurs, exacerbant l’amplitude de leurs cycles
d’abondance. Alors qu’au sud, où la végétation est plus productive, les prédateurs sont plus
généralistes – en majeure partie due à la présence de proies alternatives – ce qui diminue la
cyclicité des abondances de petits rongeurs (Turchin et al., 1997). La variation liée à la
disponibilité des végétaux le long de l’axe latitudinal peut donc avoir un impact sur les
relations trophiques.

2
Les perturbations anthropiques
Certaines perturbations anthropiques peuvent toutefois altérer ces comportements en
modifiant le patron de répartition des espèces. La modification des habitats par l’Homme
constituera d’ailleurs probablement le phénomène ayant le plus de répercussions sur la
biodiversité d’ici 2100 (Pimm et al., 2000; Sala et al., 2000). Sur la planète, plus de 40 %
de la superficie des prairies, des forêts et des tourbières auraient déjà été transformées en
zones agricoles ou urbaines au cours des derniers siècles en réaction aux besoins croissants
des humains (Chapin III et al., 2000). Par ailleurs, Stohlgren et al. (2005) ont démontré que
les variables anthropiques, telle que la densité de population humaine, étaient des facteurs
importants à considérer pour décrire la diversité et la répartition des espèces de plantes
vasculaires aux États-Unis. Alors qu’aucune corrélation entre la diversité des plantes
vasculaires et le gradient latitudinal n’a été trouvée, il existait une forte corrélation positive
entre la densité des espèces et celle de la population humaine. L’accroissement de la
population humaine et l’augmentation de la modification des habitats ont donc modifié le
gradient latitudinal originel de répartition de la diversité des espèces.
Ces modifications de l’habitat peuvent influencer la répartition des espèces de
plusieurs façons. Tout d’abord, les conditions environnementales peuvent devenir
inadéquates pour certaines espèces après la modification des caractéristiques du paysage.
Par exemple, le tigre de l’Asie (Panthera tigris) est en voie d’extinction et a disparu de
plusieurs territoires, dont l’île de Bali (IUCN, 2011). Son extinction sur l’île de Bali est
attribuable en bonne partie à la transformation de son habitat de prédilection, la forêt
tropicale, en zone agricole (Seidensticker, 1986). Toutefois, ces modifications sont parfois
profitables pour d’autres espèces qui occupent par la suite ces milieux modifiés. Ainsi, dans
un tout autre habitat, les populations de bernaches du Canada (Branta canadensis) sont
nombreuses au printemps dans le sud du Québec où elles profitent des zones agricoles pour
se nourrir de céréales et de graminées (Giroux et al., 1996), alors qu’auparavant celles-ci
n’avaient pas accès à cet apport de nourriture d’origine anthropique. Les modifications
apportées à l’habitat par l’homme peuvent donc générer des impacts positifs ou négatifs
selon les besoins de l’espèce considérée.

3
En plus de modifier l’utilisation de l’espace et la répartition des espèces, les
perturbations anthropiques peuvent aussi modifier les interactions trophiques. Tout d’abord,
les perturbations anthropiques peuvent avoir des effets sur les interactions trophiques
directes comme dans le cas du retrait d’un prédateur dans un écosystème. Par exemple,
Dulvy et al. (2004) ont démontré qu’en retirant les poissons prédateurs, les perturbations
anthropiques ont enclenché la prolifération de leurs proies principales, les étoiles de mer,
dans les îles océaniques de Fidji. Les perturbations peuvent également avoir un impact sur
les interactions trophiques indirectes, comme la compétition apparente. La compétition
apparente est caractérisée par un système où deux proies n’interagissent pas pour l’accès
aux ressources disponibles, mais partagent un prédateur commun (Holt, 1984). Lorsque la
population de l’une des deux espèces de proies subit un accroissement, cela génère souvent
une augmentation de la population de prédateurs, ce qui peut avoir un impact sur la
population de la seconde proie. À titre d’exemple, il existe plusieurs insectes phytophages
dont les populations ne sont pas régulées par la disponibilité des ressources nutritives, mais
plutôt par des prédateurs communs, les guêpes parasitoïdes (Hymenoptera sp.) (Holt et al.,
1993).
Plusieurs études ont démontré que les perturbations anthropiques pouvaient avoir
des conséquences indirectes sur les réseaux trophiques à travers la compétition apparente.
Par exemple, à la suite de l’introduction du sanglier (Sus scrofa), les îles californiennes
Channel ont connu un accroissement des populations d’aigles royaux (Aquila chrysaetos)
ce qui a causé le déclin du renard gris insulaire (Urocyon littoralis) (Roemer et al., 2001).
Bryant et al. (2005) ont également démontré que le phénomène de compétition apparente
affectant la marmotte de l’île de Vancouver (Marmota vancouverensis) avait été déclenché
par la coupe forestière en augmentant les populations de cerfs à queue noire (Odocoileus
hemionus columbianus) ainsi que les populations de leurs prédateurs communs, le loup gris
et le couguar (Puma concolar). Aussi, les populations de cerfs de Virginie (Odocoileus
virginianus) du sud de la Colombie-Britannique semblent avoir été favorisées par la
transformation des habitats forestiers en champs agricoles, ce qui a occasionné une
augmentation des populations du prédateur présent, le couguar, et une diminution de la
présence du cerf mulet (Odocoileus hemionus), une proie alternative (Robinson et al.,
2002).

4
Plusieurs études se sont penchées sur la présence d’un gradient géographique de la
disponibilité de la végétation ayant une influence sur la démographie ou l’utilisation de
l’habitat d’une espèce (Melis et al., 2009; Singh et al., 2012) ainsi que les effets des
perturbations anthropiques sur les interactions trophiques directes ou indirectes
(Hebblewhite et al., 2005; Dulvy et al., 2004), mais peu se sont penchées sur l’effet
multiplicatif de ces deux facteurs. Cette étude vise à évaluer si les conséquences des
perturbations anthropiques affectant une interaction trophique indirecte varient le long d’un
gradient géographique. Pour ce faire, nous avons utilisé un exemple classique de
compétition apparente exacerbée par une perturbation anthropique, soit le cas du caribou
forestier et de la coupe forestière ; ainsi que la variation de l’influence de celle-ci le long
d’un gradient latitudinal de disponibilité en végétation décidue.
Le caribou forestier comme sujet d’étude
Le caribou des bois, d’écotype forestier, est considéré comme une espèce menacée au
Canada (COSEWIC, 2011) et comme une espèce vulnérable au Québec (MRNF, 2005). Il
habite la forêt boréale canadienne où ses populations sont en déclin (Courtois et al., 2003;
Hervieux et al., 2013). Il est fortement associé à des forêts de conifères en fin de succession
(forêts matures) où le lichen est leur principale source de nourriture (Courtois et al., 2008;
Hébert et al., 2013; Rominger et al., 1996). Leur domaine vital est souvent constitué de lacs
et de tourbières qui servent de refuge à la prédation en hiver (Rettie et al., 2000; Fortin et
al., 2008). Il s’agit d’une espèce avec un faible taux de recrutement (Bergerud, 1974) et une
sensibilité marquée aux modifications environnementales, ce qui en fait une espèce modèle
pertinente pour étudier l’impact des coupes forestières sur ses relations avec son
environnement et les autres espèces qui s’y retrouvent.
La coupe forestière semble être étroitement associée au retrait du caribou forestier
vers le nord (Courtois et al., 2008; Schaefer, 2003). La sensibilité du caribou forestier à la
présence de coupes dans son habitat est un phénomène bien connu (Vors et al., 2007).
Celui-ci évite généralement les milieux en régénération et en intègre ainsi une très faible
proportion dans son domaine vital (Hins et al., 2009; Beauchesne et al., 2013; Courtois et
al., 2008; Dyer et al., 2001). La coupe forestière diminue drastiquement la disponibilité

5
d’une des ressources primordiales du caribou, le lichen arboricole, en plus d’avoir
possiblement des effets négatifs à long terme sur la présence de lichens terricoles (Courtois
et al., 2008). Les individus des populations comprises dans des milieux perturbés ont
tendance à augmenter la superficie de leur domaine vital et à diminuer leur fidélité à ce
domaine (Courtois et al., 2007; Faille et al., 2010). La présence de milieux en régénération
dans leurs domaines vitaux augmenterait d’ailleurs le risque de mortalité (Dussault et al.,
2012; Wittmer et al., 2007). L’une des principales causes de l’impact de la coupe forestière
sur la répartition du caribou serait liée à l’augmentation de la pression de prédation que la
coupe forestière génère en forêt boréale (Seip, 1992; Bergerud, 1974).
Cette pression de prédation amenée par la compétition apparente entre le caribou et
l’orignal via le loup serait engendrée par l’enfeuillement des parterres de coupe forestière.
L’enfeuillement est un phénomène résultant d’une baisse significative du nombre de
conifères présents dans un secteur donné à la suite d’une coupe forestière, ce qui entraîne
une augmentation des espèces feuillues intolérantes à l’ombre comme le bouleau à papier
(Betula papyrifera) et le peuplier faux-tremble (Populus tremuloides), soient des ressources
alimentaires de prédilection pour l’orignal (Boucher et al., 2009; Laquerre et al., 2009;
Potvin et al., 2004). Une plus grande richesse est aussi observée dans la strate arbustive,
représentée par plusieurs espèces comme l’érable à épis (Acer spicatum), l’aulne (Alnus
sp.), l’amélanchier (Amelanchier sp.), le saule (Salix sp.) et le sorbier américain (Sorbus
americana) (Hart et al., 2008; Dussart et al., 2002; Reich et al., 2001). De plus, les
communautés végétales diffèrent toujours de celles des forêts matures plus de trente ans
après la perturbation (McRae et al., 2001; Elson et al., 2007; Boan et al., 2011).
L’enfeuillement des parterres de coupe peut différer selon la latitude à laquelle ces
parcelles se retrouvent. Par exemple, le peuplier faux-tremble et le bouleau à papier se
retrouvent surtout sous forme arbustive lorsque la latitude augmente (Jobidon, 1994).
Parallèlement, Huang et al. (2010) ont démontré que la température pouvait influencer la
croissance radiale de ces deux mêmes espèces à différentes latitudes. La productivité du
peuplier faux-tremble diminue aussi avec l’augmentation de la latitude (Lapointe-Garant et
al., 2010). Toutefois, peu d’études se sont penchées sur l’effet des gradients géographiques
sur l’enfeuillement général des parterres de coupe. Ces différences de régénération des

6
peuplements forestiers dues à la latitude sont importantes à considérer. C’est cette
régénération qui sera le moteur de la compétition apparente entre le caribou et l’orignal en
forêt boréale puisque c’est l’enfeuillement des parterres de coupe qui attire l’orignal dans
ce milieu.
Les milieux ayant subi un enfeuillement à la suite d’une coupe forestière deviennent
particulièrement riches en nourriture pour l’orignal (Serrouya et al., 2011; Bjørneraas et al.,
2011; Crête, 1977; Bowman et al., 2010), ce qui peut amener une augmentation des
populations d’orignaux dans le milieu (Potvin et al., 2005). Dans les secteurs coupés, les
orignaux favorisent les milieux riches en régénération où le couvert latéral est élevé et le
brout feuillu abondant (Courtois et al., 1998; Potvin et al., 2004). De plus, les orignaux
semblent profiter des bordures entre deux habitats distincts qui sont disponibles à la suite
d’une coupe forestière (Courtois et al., 2002). Ainsi, ils utilisent généralement les parterres
de coupe enfeuillés comme parcelles de nourriture et le couvert forestier des forêts matures
environnantes comme couvert protecteur (Crête, 1977). Finalement, il a été démontré que
des facteurs abiotiques de l’habitat, comme la latitude et le climat, peuvent aussi influencer
la présence et la condition physique des orignaux par leurs effets sur la disponibilité en
nourriture (Mansson et al., 2012; Dou et al., 2013; Saether, 1985). Ainsi, les facteurs
influençant l’enfeuillement des parterres de coupe pourraient avoir des effets directs sur la
sélection d’habitat de l’orignal.
Les loups sont au sommet du réseau trophique de la forêt boréale et régulent les
populations de plusieurs herbivores (Messier, 1994). L’orignal étant généralement la proie
privilégiée des loups (Courbin, 2013; Bergerud et al., 1986), les populations de ces derniers
augmentent habituellement à la suite d’une hausse des populations d’orignaux dans un
secteur donné (Messier, 1985). Les loups sont reconnus pour utiliser les routes et chemins
forestiers, qui sont souvent nombreux près des parterres de coupe, afin de faciliter leurs
déplacements et ainsi améliorer leur efficacité de prédation (James et al., 2000). Ils
sélectionnent donc les milieux coupés, où la nourriture de leur proie privilégiée sera
abondante et leurs déplacements facilités (Neufeld, 2006; Bowman et al., 2010). Étant
donné qu’ils sélectionnent les milieux privilégiés où l’orignal devrait être présent, la
sélection d’habitat du loup devrait s’ajuster à celle de l’orignal pour maximiser ses chances

7
de rencontre avec sa proie principale et ainsi être corrélée à l’ampleur de l’enfeuillement
des parterres de coupe.
Plusieurs études se sont concentrées sur les conséquences de la compétition
apparente, déclenchée par la coupe forestière, sur la survie du caribou forestier en forêt
boréale (Latham et al., 2013; James et al., 2000; Neufeld, 2006). En effet, la présence de
coupe forestière rend la séparation spatiale entre le caribou et l’orignal de plus en plus
difficile à obtenir, en augmentant la proportion de jeunes forêts au sein de l’habitat du
caribou forestier (Peters et al., 2013; James et al., 2004). La sélection des parterres de
coupe forestière par l’orignal et le loup (Neufeld, 2006; Serrouya et al., 2011) ainsi que
l’évitement du caribou forestier des milieux fréquentés par l’orignal (James et al., 2004)
sont des phénomènes bien connus. Toutefois, il existe peu d’informations sur les
changements possibles de ces relations le long d’un axe latitudinal. Notre étude est basée
sur l’hypothèse que la contribution de la coupe forestière à l’interaction trophique indirecte
qu’est la compétition apparente varie selon un gradient latitudinal, et ce, à travers des
différences au niveau de l’enfeuillement des parterres de coupe. Ainsi, nous nous attendons
à ce qu’un degré d’enfeuillement moindre au nord soit moins propice à l’utilisation des
parterres de coupe par l’orignal, ce qui devrait diminuer le potentiel de compétition
apparente à des latitudes plus élevées.
Objectif de l’étude
Cette étude vise à évaluer si la contribution de la coupe forestière au phénomène de
compétition apparente entre l’orignal et le caribou forestier varie selon un gradient
latitudinal. Je vais d’abord évaluer si l’enfeuillement des parterres de coupes est influencé
par un gradient latitudinal. Puis, je vérifierai si ces différences potentielles d’enfeuillement
des parterres de coupe entraînent une différence comportementale de la sélection de ces
milieux pour l’orignal, le loup et le caribou forestier. La sélection de l’orignal pour des
milieux à haute régénération en feuillus à l’intérieur des coupes (Potvin et al., 2004) laisse
penser que ceux-ci sélectionneront plus fortement les coupes les plus enfeuillées.
Parallèlement, en forêt boréale, les loups ont tendance à avoir un plus grand chevauchement
spatial avec l’orignal (Basille et al., 2012), ce qui nous amène à penser que ceux-ci

8
ajusteront leur sélection de l’habitat en fonction des milieux privilégiés par les orignaux.
Finalement, selon la théorie de la ségrégation spatiale (James et al., 2004; Peters et al.,
2013), les caribous forestiers devraient éviter plus fortement les milieux sélectionnés par les
orignaux, dont les parcelles de coupe forestière avec une grande régénération en feuillus.
Approche empirique
L’Indice de Végétation Amélioré (Enhanced Vegetation Index, EVI) a été utilisé pour
estimer l’enfeuillement général des parcelles de coupes. Une évaluation de la relation entre
le degré d’enfeuillement d’un parterre de coupe et la valeur d’EVI qui lui est associé a
également été réalisée par des observations directes sur le terrain dans un souci de
validation. Puis, nous avons évalué l’impact de la latitude et du temps écoulé depuis la
coupe sur la moyenne pondérée de l’EVI de 139 725 parterres de coupe. Nous avons
également utilisé une base de données provenant du suivi télémétrique d’un total de 15
orignaux, 11 loups et 37 caribous équipés de colliers GPS afin d’étudier leur sélection de
l’habitat, notamment en lien avec l’enfeuillement des parterres de coupe. Pour ce faire, la
sélection de l’habitat des trois espèces a été modélisée à l’aide de Fonctions de Sélection
des Ressources (Resource Selection Function, RSF). En comparant les caractéristiques des
milieux utilisés avec celles des milieux disponibles, les RSF nous permettent de quantifier
la sélection d’habitat des individus pour chacune des espèces suivies (Manly et al., 2002).
Les milieux disponibles ont été décrits en projetant des points aléatoires à l’intérieur du
polygone minimal convexe 100 % (MCP) de chaque individu pour les caribous et les
orignaux, et à l’intérieur du MCP 100 % de la meute pour les loups. L’influence de
l’enfeuillement des parterres de coupe sur leur sélection par les trois espèces a été
déterminée grâce à l’ajout d’une interaction entre la sélection des parterres de coupe et
l’EVI pour chaque point GPS considéré. Finalement, la cooccurrence entre espèces a été
évaluée en fonction de l’enfeuillement des parterres de coupe.

9
CHAPITRE PRINCIPAL
The potential for human-induced apparent competition between forest-
dwelling caribou and moose in the boreal forest varies along a latitudinal
gradient
Caroline Gagné1, Julien Mainguy2 and Daniel Fortin1
1Chaire de Recherche Industrielle CRSNG, Université Laval en Sylviculture et Faune,
Département de biologie, Université Laval, Québec, QC, G1V 0A6, Canada.
2Ministère des Forêts, de la Faune et des Parcs, Direction générale de l’expertise sur la
faune terrestre et ses habitats, Direction de la faune terrestre et de l’avifaune, 880 chemin
Sainte-Foy, Québec, QC, G1S 4X4, Canada.

10
Résumé
Une proie peut avoir un impact sur une autre espèce de proie à travers un prédateur
commun. Cette compétition apparente peut être engendrée par des activités humaines, et
tout dépendamment de leur force, peut générer la disparition locale d’une espèce de proie.
Alors que des gradients géographiques affectant la force des interactions directes sont bien
documentés, il existe peu d’exemples où la menace de la compétition apparente varie le
long d’un axe latitudinal. Pourtant, cette information est nécessaire pour mieux comprendre
les systèmes complexes et mieux orienter les efforts de conservation d’espèces vulnérables.
Nous avons utilisé une combinaison d’analyses spatiales de la disponibilité de la végétation
décidue et d’analyses de sélection d’habitat par des animaux munis de colliers
télémétriques pour démontrer que le potentiel de la coupe forestière d’engendrer la
compétition apparente entre le caribou forestier (Rangifer tarandus caribou) et l’orignal
(Alces alces) à travers le loup gris (Canis lupus) devrait diminuer avec une augmentation de
la latitude. Durant la succession écologique suivant la coupe forestière, il y a souvent une
augmentation de la disponibilité de la végétation décidue consommée par l’orignal tandis
que le lichen consommé par les caribous diminue en abondance. Nous avons utilisé les
Fonctions de Sélection de Ressources pour déterminer les conséquences de la variation
spatiale de l’enfeuillement des parterres de coupe sur le comportement de sélection
d’habitat de chacune des espèces. Après avoir tenu compte du temps écoulé depuis la
coupe, nous avons démontré que l’enfeuillement moyen des parterres de coupe diminuait
avec une augmentation de la latitude. Les orignaux et les loups suivis par télémétrie
augmentaient leurs sélections des parterres de coupe qui contenaient un plus fort
enfeuillement. Au contraire, la probabilité d’occurrence des caribous restait faible dans les
parterres de coupe, et ce, quelle que soit l’intensité de l’enfeuillement. Les réactions
comportementales sont telles que la probabilité relative de cooccurrence entre les orignaux
et les loups étaient seulement faiblement associée aux parterres de coupe forestière à des
latitudes élevées, contrairement aux basses latitudes. De plus, la cooccurrence entre les
caribous et les loups demeuraient généralement faible dans les coupes forestières. Notre
étude démontre que l’influence de la coupe forestière sur la compétition apparente entre le
caribou forestier et l’orignal à travers la prédation du loup gris devrait diminuer avec une

11
augmentation de la latitude. Nos résultats démontrent l’importance de considérer les
gradients géographiques de disponibilité de la végétation pour aider à mieux comprendre
l’effet des perturbations anthropiques sur les interactions trophiques et à mettre au point des
stratégies de conservation des espèces qui seront adéquates sur de vastes territoires.

12
Abstract
Prey species can affect one another through a shared predator. This apparent competition
may be triggered by human activities, and, depending on its strength, can result in the local
extinction of one of the prey populations. While geographical gradients in the strength of
direct interactions are well documented for many taxa, there are few demonstrations that
the threat of apparent competition can vary along a latitudinal axis. Yet this information is
needed to better understand complex systems and guide conservation efforts towards
vulnerable species. We used spatial analyses of deciduous vegetation availability and
habitat selection by radio-collared animals to demonstrate that the potential for logging
activities to trigger the apparent competition between forest-dwelling caribou (Rangifer
tarandus caribou) and moose (Alces alces) through gray wolf (Canis lupus) predation
should decrease with an increase in latitude. During the ecological succession that follows
forest harvesting, there is often an increase in availability of the deciduous vegetation
consumed by moose, and a decrease in the abundance of lichen consumed by caribou. We
used a Resource Selection Function to determine the consequences of deciduous vegetation
availability following harvest on the habitat selection behaviour of each species. After
controlling for time since harvest, we found that the mean availability of deciduous
vegetation in logged stands decreased with increasing latitude, and all three species reacted
to this change. Radio-collared moose and wolves increased their selection for cutovers as
they contained more deciduous vegetation. In contrast, the probability of occurrence of
radio-collared caribou remained low in cutovers regardless of deciduous vegetation
availability. The behavioural responses were such that the relative probability of co-
occurrence between moose and wolves became only weakly associated with cutovers at
high latitudes, unlike at low latitudes. Furthermore, co-occurrence between caribou and
wolves remained generally low in cutovers. Overall, our study demonstrates that the
potential influence of logging activities on the apparent competition between forest-
dwelling caribou and moose through wolf predation should decrease with increasing
latitude. Our findings show that considering geographical gradients in vegetation
availability can help to better understand the effect of human activities on trophic

13
interactions, and to develop strategies for species conservation that remain effective over
vast areas.

14
Introduction
Analyses of biological processes across geographical gradients provide a powerful
approach for understanding a broad range of topics from evolutionary ecology to ecosystem
function. One of the most important geographical gradients is the spatial variation in
vegetation availability (Connell et al., 1964). Indeed, such a gradient can have a profound
influence, for instance, on how predators impact herbivore prey species (Melis et al., 2009).
For example, the influence of predation on the temporal dynamics of field vole (Microtus
agrestis) populations in Fenno-scandinavia varies along a latitudinal vegetation gradient
(Hanski et al., 2001). In northern parts of their range, specialist predators target this small
species, resulting in large fluctuations in their abundance. In contrast, in southern parts of
their range where plant productivity is higher, predators tend to be generalists – due to the
presence of a range of alternative prey species – and have a stabilizing effect on field vole
population dynamics (Turchin et al., 1997). Some latitudinal gradients are also altered by
anthropogenic activities. In fact, the interactions between humans and natural processes can
be such that the level of human activities is often a better predictor of terrestrial plant
diversity than latitudinal or climate gradients (Stohlgren et al., 2005). Given the increasing
human footprint on wildlife habitat (Chapin III et al., 2000), there is high conservation
value in identifying the mechanisms by which anthropogenic activities can affect ecological
processes along geographical gradients. This information can provide a strong basis for the
development of strategies that could reduce the effects of humans on natural ecosystems.
By modifying vegetation availability through land use and biotic exchange (Sala et
al., 2000), anthropogenic disturbances can alter trophic interactions. For instance, in Banff
National Park (Alberta, Canada) human activities have excluded wolves (Canis lupus) from
certain areas, resulting in local increases in elk (Cervus canadensis) density. Such an
increase in herbivore density has led to increased browsing and a marked decline in aspen
(Populus spp.) recruitment and riparian songbird diversity (Hebblewhite et al., 2005).
Likewise, human activities can trigger apparent competition, whereby two species do not
necessarily compete directly for resources but are preyed upon by the same predator (Holt,
1984). For example, the human introduction of feral pigs (Sus scrofa) to the Channel
Islands in California caused an increase in the abundance of golden eagles (Aquila

15
chrysaetos) that resulted in increased predation on, and ultimately the decline of, the island
fox (Urocyon litoralis) (Roemer et al., 2001).
A classic example of apparent competition driven by human disturbance involves
forest-dwelling caribou (Rangifer tarandus caribou), moose (Alces alces) and gray wolves
(Seip, 1992). When old growth forests are converted to early-seral forests by logging,
cutovers get colonized by deciduous shrubs and trees, primarily trembling aspen (Populus
tremuloides) and paper birch (Betula papyrifera), which are high-quality food for moose
(Potvin et al., 2004; Bjørneraas et al., 2011). As a result, local moose abundance increases,
triggering a numeric response in wolves (Messier, 1985; Bowman et al., 2010). In addition
to increasing wolf abundance, forest harvesting allow wolves to hunt more efficiently by
using the extensive road network created by logging activities (James et al., 2000; Apps et
al., 2013). Overall, by fragmenting and modifying forest ecosystems, logging reduces the
spatial segregation between caribou and both moose and wolf (i.e. spatial segregation
hypothesis) (James et al., 2004; Peters et al., 2013), increasing the predation pressure on
caribou, a specie considered to be an alternative prey for wolves (Seip, 1992). The effect of
apparent competition on a vulnerable prey specie is moderated by the asymmetry of the
interaction (DeCesare et al., 2010). Regarding the moose-caribou system, logging activities
favor moose by providing their prime forage, i.e. deciduous vegetation (Bjørneraas et al.,
2011). Whereas for caribou, their resources decrease following harvest with an abrupt
decline of arboreal lichen and a possible negative long-term effect on terrestrial lichen
(Courtois et al., 2008).
The trophic interactions between caribou, moose and wolves have received
considerable attention in recent years due to the importance of boreal forest management
and the endangered status of forest-dwelling caribou (e.g. Apps et al., 2013; Latham et al.,
2013). Forest-dwelling caribou are known to have declined over most of Canada’s boreal
forest (Peters et al., 2013; Wittmer et al., 2013; Vors et al., 2009), and are considered
threatened (COSEWIC, 2011). Caribou conservation thus leads to significant constraints on
the logging industry. A question of socioeconomic importance is therefore whether or not
timber harvest will continue to drive the apparent competition documented between the
caribou and moose, as logging progresses towards the north.

16
Here we used a combination of spatial analyses of deciduous vegetation availability
and habitat selection by radio-collared animals to determine if the potential influence of
logging activities on the apparent competition between forest-dwelling caribou (Rangifer
tarandus caribou) and moose (Alces alces) through wolf (Canis lupus) predation can be
expected to decrease with increasing latitude.
Methodology
STUDY AREA
The study region (31,575 km2, Figure 1) is located in the boreal forest of Québec, Canada
(51°N, 69°W). It is a typical Canadian Precambrien Shield landscape with rolling hills and
elevations varying from 300 to 1,000 m. The mean temperatures in February and July are -
20°C and 15°C respectively (Environnement Canada, 2014). The mean annual precipitation
is 715 mm, of which a third falls as snow (Courbin et al., 2009). The forest is dominated by
black spruce (Picea mariana) and balsam fir (Abies balsamea), with jack pine (Pinus
banksiana), trembling aspen (Populus tremuloides), paper birch (Betula papyrifera) and
eastern larch (Larix laricina) also present (Fortin et al., 2008). Moose density was
evaluated at 4.3 individuals/100 km2 in 1989 in a part of the study region (Gingras et al.,
1989). Caribou density was later estimated at 4.7 individuals/100 km2 based on an aerial
survey conducted in March 2014 over another part of the study region in which some of the
collared animals were monitored. Four packs of wolves were found in the same region with
four to six individuals per pack. Black bears (Ursus americanus) are also present in the area
but they are not expected to prey on adult moose or caribou. Logging has been the main
source of anthropogenic disturbances in the region since the 1990s, particularly south of
51°N latitude.

17
Figure 1. Location of the study area within the Côte-Nord region of Québec’s boreal forest,
Canada (red line). White line delimits the area where radio-collared moose, wolves and
caribou were tracked. Three reference points of the latitudinal gradient (South: 5352900
UTM; Middle: 5556575 UTM; North: 5760253 UTM) are represented as reference for
some of the analyses and results reported.
66°0'0"W
67°0'0"W
67°0'0"W
68°0'0"W
68°0'0"W
69°0'0"W
69°0'0"W
70°0'0"W
70°0'0"W71°0'0"W
52°0'0"N 52°0'0"N
51°0'0"N 51°0'0"N
50°0'0"N 50°0'0"N
49°0'0"N 49°0'0"N
®
TextTextTextText
South
Middle
North
0 20 40 60 8010Kilometers

18
ASSESSING POST-CUT VEGETATION GROWTH
To evaluate the availability of deciduous vegetation during post-harvest forest succession,
we considered 139,725 cutovers with information about the year of logging, ranging from
1925 to 2011. We used the centroid of each cutovers to define the latitude. To quantify the
deciduous vegetation availability in the regenerating harvested stands, we used the
Enhanced Vegetation Index (EVI) of the Moderate Resolution Imaging Spectro-radiometer
(MODIS) data set, which has a 250 m resolution. EVI was developed from the Normalized
Difference Vegetation Index (NDVI) to optimize the vegetation signal, improve sensitivity
in high biomass region and to take into account canopy background and atmospheric
influences (Huete et al., 2002). Twenty EVI pixels were visited to determine the
relationship between EVI and the percentage of cover of deciduous vegetation (shrubs and
trees). Within the area covered by each pixel, we visually estimated the percentage of
deciduous vegetation in nine circular (10-m radius) plots, and used average values in
subsequent analysis.
We used generalized linear models with Gaussian error structure on the 139,725
cutovers to assess how weighted mean EVI of cutovers covaried with time since harvest
and latitude. We used EVI from July 28th to ensure that foliage was fully developed. If a
cutover was distributed across more than one EVI pixel, weighted mean EVI (based on the
proportion of each cutover in each EVI grid cell) was calculated. All geographical
information system (GIS) work was conducted using POSTGIS 2.0.1 (Obe et al., 2011). The
model also included a quadratic term for ‘time since harvest’ because we expected
deciduous vegetation availability to peak in early to mid-seral forests, before decreasing as
the forest matures. We added an interaction term between ‘time since harvest’ and ‘latitude’
to examine whether the temporal changes in EVI during forest succession differed along
the latitudinal axis.
ANIMAL MONITORING
We fitted 15 adult female moose with Global Positionning System (GPS) collars (model
3300L, Lotek Engineering Inc., Newmarket, ON, Canada) that registered a location every
hour. Each moose was followed for an average of 27.8 months (range: 8 to 45 months)

19
from January 2005 to February 2008. We also followed a total of 37 female caribou from
March 2005 to March 2012 with GPS collars (model 3300, Lotek Engineering Inc.,
Newmarket, ON, Canada; model TGW 4680, Telonics Inc., Mesa, AZ, USA; or Global
Star model Track M, Lotek Engineering Inc., Newmarket, ON, Canada). The collars were
programmed to transmit a location every 1, 2, 3, 4 or 8 hours depending on collar type and
year. Each caribou was monitored using telemetry for an average of 23.1 months (range: <
1 to 56 months). Finally, 11 adult wolves (6 females, 5 males), from 4 packs, were followed
for an average of 19.6 months (range: < 1 to 49 months) from March 2005 to November
2011. Nine individuals were fitted with GPS collars transmitting a location every 6 hours
every day of the week (model GPS3300, Lotek Engineering Inc.) or 4 hours five days a
week, and every hour for the rest of the week (model GPS3300SW, Lotek Engineering
Inc.). The two remaining wolves were fitted with Argos collars (model TGW-3580,
Telonics Inc.), transmitting every 10 hours. All individuals from the three species were
captured in winter with a net-gun fired from a helicopter.
HABITAT CHARACTERISTICS
Habitat characteristics at GPS locations were identified using a Landsat Thematic Mapper
image taken in 2000 with a 25 m resolution (Ressources naturelles Canada, 2000). The
satellite image was composed of 52 land cover classes, which we reclassified into 8 broad
land cover types: 1) open areas, 2) water bodies (including wetlands), 3) shrublands, 4)
lichen heath communities, 5) closed canopy mature conifer forests, 6) open mature forests
with lichen, 7) open mature forests without lichen and 8) mixed forests. The satellite image
was updated every year with information provided by the forestry companies of the region,
adding two land cover types: cuts (0-50 years) and burned areas (0-50 years). We used a
50-year threshold for disturbance because the GPS-collared animals we monitored were
found in an area where cutovers dated from 1965 to 2011. As well as being assigned a land
cover type, each location was also attributed an EVI value to estimate the deciduous
vegetation availability.
HABITAT SELECTION MODEL

20
We used Resource Selection Functions (RSFs) to determine the habitat selection of caribou,
moose and wolves. RSFs compare characteristics of observed GPS points (scored 1) and
random (scored 0) locations (Manly et al., 2002). Random locations for a given moose and
caribou were drawn within the 100% Minimum Convex Polygon (MCP) annual range of an
individual, whereas for wolves, random locations were drawn within the 100% MCP of all
pack members. Although the MCP should exceed the actual annual home range of each
individual (Burgman et al., 2003), our intent was not to determine the actual home range,
but rather to delineate an area that was available to the individual. We thereby investigated
habitat selection between Johnson’s (1980) second- and third-order of selection. We
incorporated an interaction between the “cuts” land cover type and EVI, to determine
whether or not the use of cutovers was influenced by the availability of deciduous
vegetation. We did not detect any multicollinearity issues, as variance inflation factor were
systematically <10 (Cohen et al., 2013).
RSFs were implemented using mixed-effect logistic regression. For the moose and
the caribou RSFs, we included a random intercept for the interaction between individual
and year to take into account the correlations between an individual’s locations within a
given year (Gillies et al., 2006) and to correct for the different collar schedules for a given
species. The RSFs for moose and caribou took the following functional form:
w(x) = exp(β0 + β1x1ij + … + βn(xnij × EVIij) + γ0jk),
where w(x) is the relative probability of occurrence, β0 is the mean, βn is the coefficient
estimated for the nth variable, xnij is the value of the nth variable for the ith location for the jth
individual, EVIij is the deciduous vegetation availability for the ith location for the jth
individual and γ0jk is the random intercept for the jth individual of the kth year. For wolves,
we used the same general model structure, with the exception that “individuals within
packs” and “pack within a year” were both modeled as random effects because a given
pack could include more than one collared wolf (Gillies et al., 2006; Hebblewhite et al.,
2008). Analyses were performed using the ‘glmer’ function in the « lme4 » package (Bates
et al., 2012) in R program version 3.1.0 (R Development Core Team, 2010).

21
To assess whether the availability of post-harvest deciduous vegetation had an
impact on habitat selection in the three species, we investigated the relative support of two
Resource Selection Functions. The first was a general landscape model, which included the
10 land cover types as covariates, with open conifer forests without lichen as the reference
category. In the second model, we added an interaction between cutovers and EVI. We
identified the model with the most empirical support using the Bayesian Information
Criterion (BIC). With large sample sizes, BIC is a more appropriate metric of model
parsimony than the Akaike Information Criterion (Schwarz, 1978; Boyce et al., 2002). As
suggested by Boyce et al. (2002), we tested model robustness using k-fold cross-validation,
fitting RSFs using 80% of the data before testing their predictive on the remaining 20% of
data. The process was done 100 times and average rs (r̅s) are reported. For each species, we
used the top-ranking RSF to estimate the relative probability of occurrence w(x), with
predicted values being scaled between 0 and 1 following:
! = ! x !!!"#!!"#!!!"#
,
where w(x) was the relative probability of use of a given pixel, and wmin and wmax were,
respectively, the smallest and largest values of w(x) for that species over the study area. We
also assessed the relative probability of co-occurrence between moose and wolves, and
caribou and wolves by multiplying the relative probability of occurrence of the two species.
Results
POST-CUT VEGETATION GROWTH
Harvested stands with higher weighted mean EVI were covered by a larger proportion of
deciduous vegetation (r2=0.56, n=20, Figure 2). EVI in cutovers thus provides a valid
estimate of deciduous vegetation availability.
22
Figure 2. Observed percentage cover of deciduous vegetation available in harvested forest
stands (250 m x 250 m, corresponding to the actual size of an EVI pixel, see Methods) in
relation to Enhanced Vegetation Index (EVI) in the boreal forest of Québec, Canada. Filled
circles represent raw values. Grey lines represent the 95% confidence intervals.
The weighted mean EVI of logged stands varied with both latitude and time since
harvest (Table 1). The EVI of cutovers increased to a peak after 10 to 40 years following
harvest, depending on latitude, and then decreased as the stands matured (Figure 3). After
controlling for time since harvest, EVI was lower in cutovers located at relatively high
latitudes, indicating a northward decrease in the deciduous vegetation available in cutovers.
In the south, EVI peaks much earlier and reached a higher level. This indicates that
deciduous vegetation was less abundant and grew more slowly in the north than in the
south.

23
Table 1. Parameter estimates of a generalized linear model characterizing the influence of
time since harvest and latitude on the weighted mean Enhanced Vegetation Index of
cutovers in the boreal forest of the Côte-Nord region, Québec, Canada. For each covariate,
model coefficients (β), standard errors (SE) and P-values are presented.
Variable β SE P Intercept 3.65 3.18e-02 <0.001
Time since harvest (years) -3.66e-02 1.38e-03 <0.001
Time since harvest2 -4.02e-05 6.49e-07 <0.001
Latitude (UTM) -5.88e-07 5.69e-09 <0.001
Time since harvest × Latitude 6.88e-09 2.46e-10 <0.001

24
Figure 3. The influence of time since harvest and latitude on the weighted mean Enhanced
Vegetation Index (EVI) of cutovers in the boreal forest of the Côte-Nord region, Québec,
Canada. The South region corresponds to the first quartile of latitudes; the Middle region to
the second and third quartile while the North region corresponds to the fourth quartile of
the 139,725 cutovers studied. Filled coloured diamonds are mean values of each 10-year
class of every region. Lines represent the predicted relationships from generalised linear
models between EVI and time since harvest, for stands from different
latitudes (South: 5,419,517 UTM; Middle: 5,527,884 UTM; North: 5,619,927 UTM, see
Figure 1).
HABITAT SELECTION
The RSF best describing the relative probability of occurrence of the three species
considered the landscape covariates together with EVI in cutovers (Table 2). These models
were robust to cross-validation, with r̅s > 0.90 for all species (Table 2). By contrast, the
models considering only landscape received essentially no empirical support for any
species (ΔBIC: moose= 2,177; wolves= 204; caribou= 3,807).

25
The top-ranking RSF revealed that, relative to open conifer forests without lichen
(the model’s reference category), moose selected mixed/deciduous forests, whereas they
avoided closed-canopy mature conifer forests, open mature conifer forests with lichen,
lichen heath communities and water bodies (Table 2). Similarly, wolves selected
mixed/deciduous forests, but also burned areas and open areas. Caribou selected open
mature conifer forests with lichen, lichen heath communities and shrublands, whereas they
avoided closed-canopy mature conifer forests, burned areas, mixed/deciduous forests, open
areas and water bodies.
Table 2. Parameter estimates of the top-ranking Resource Selection Functions describing
the annual habitat selection by 15 moose, 11 wolves and 37 forest-dwelling caribou that
were monitored with GPS collars in Québec’s boreal forest, Canada. Each of the model
covariates is presented with their selection coefficients (β ± SE).
Variablea Resource selection function
Moose Wolf Caribou
Open areas -0.004 ±0.046 0.175 ±0.083* -0.249 ±0.031*
Water bodies -1.972 ±0.028* -0.041 ±0.031 -1.806 ±0.011*
Shrublands 0.026 ± 0.026 -0.276 ±0.049* 0.261 ±0.016*
Lichen heath -1.629 ± 0.102* 0.138 ±0.106 0.544 ±0.037*
Closed mature conifer -0.361±0.011* -0.034 ±0.026 -0.270 ±0.006*
Open conifer with lichen -1.377 ±0.021* -0.014 ± 0.025 0.458 ±0.008*
Mixed 1.023 ± 0.012* 0.134 ±0.028* -0.407 ±0.010*
Burned areas (0-50 years) -0.190 ± 0.017* 0.189 ±0.019* -0.755 ±0.012*
Cuts (0-50 years) -1.505 ±0.038* -0.250 ±0.073* -1.575 ±0.092*
Cuts × EVIb 3.654 ±0.097* 0.988 ±0.178* -3.543 ±0.777*
k-fold: r̅s 0.901 0.910 0.944 aReference category is open conifer forests without lichen bEnhanced Vegetation Index averaged over the cutblock * 95% confidence intervals exclude 0

26
The response of all species to cutovers varied with local EVI estimate, revealing
that their response to cutblocks depends on deciduous tree availability. Moose avoided
cutovers with low EVI values, ranking similarly to lichen heath communities, water or open
mature conifer forests with lichen (Table 2; Figure 4A). As EVI increased, however, moose
increased their selection for cutovers. In our study region, weighted mean EVI of cutovers
is expected to peak between 0.322 and 0.506 depending on latitudes (Figure 3).
Accordingly, moose either did not select cutovers or exhibited little selection at high
latitude, even when the available deciduous vegetation had reached its maximum. Wolves
showed a similar response: cutovers with a high EVI were the most selected cover types;
whereas those with a low EVI tended to be strongly avoided (Table 2; Figure 4B). Caribou
avoided cutovers with low EVI estimates, and avoided even more strongly cutovers as they
became invaded by deciduous vegetation that yielded relatively high EVI values (Table 2;
Figure 4C). Accordingly, the co-occurrence of moose and wolves in cutovers increased
with increasing EVI values (Figure 5A), with only mixed/deciduous forests having a higher
probability of co-occurrence at high latitude (i.e. lower EVI values). The co-occurrence of
caribou and wolves slightly decreased with high EVI values (Figure 5B), remaining the
land cover type where it is less likely to see them coexist.

27
Figure 4: Variation in the relative probability of occurrence in 10 land cover types, as a
function of the Enhanced Vegetation Index (EVI), a proxy for the availability of deciduous
vegetation, for A) moose, B) wolf and C) caribou in the boreal forest of the Côte-Nord
region, Québec, Canada.

28
Figure 5: Variation in the relative probability of co-occurrence in 10 land cover types, as a
function of the Enhanced Vegetation Index (EVI), a proxy for the availability of deciduous
vegetation, for A) moose and wolves, and B) caribou and wolves in the boreal forest of the
Côte-Nord region, Québec, Canada.
Discussion
Although there are numerous examples of latitudinal gradients in the intensity of direct
biotic interactions (e.g., decreases in predation risk with increasing latitude) involving
many taxa (Schemske et al., 2009), geographical gradients in indirect interactions remain
poorly documented. Yet the fate of vulnerable prey populations may depend on the

29
intensity of indirect trophic interactions (DeCesare et al., 2010). We provided an example
in which the intensity of an indirect interaction varies along a geographical gradient. More
specifically, we demonstrated how the potential role of human activities in triggering
apparent competition involving the threatened forest-dwelling caribou should decline
progressively as the industry moves northwards (Schaefer, 2003).
Despite the impacts of apparent competition on caribou being the subject of many
studies in recent years (e.g. Apps et al., 2013; Peters et al., 2013; Latham et al., 2013;
James et al., 2004), to our knowledge, this is the first report of a latitudinal gradient in the
potential for apparent competition. We based this conclusion on an investigation of some
key elements underlying the process of human-induced apparent competition between
moose and caribou: 1) a latitudinal decrease in primary productivity, and 2) the response of
wolves, moose and caribou to this decrease.
First, we found that deciduous forage availability (based on EVI, which covaried
with the percentage of deciduous vegetation) in cutovers was lower at northern than
southern latitudes, a trend that we detected even along a rather short 400-km latitudinal
gradient located in a single ecoregion. Net primary productivity generally decreases with
increasing latitude (Rosenzweig, 1968). Accordingly, the growth of two prime deciduous
forage species for moose, trembling aspen and paper birch, varies with latitude, taking
more ‘shrubby’ forms (Jobidon, 1994) and decreasing in productivity at northerly latitudes
(Huang et al., 2010; Lapointe-Garant et al., 2010). The availability of deciduous vegetation
is predicted to be higher in the southern part of our study area throughout the first 75 years
after harvest than at any point in time in the northern part of this area.
Second, wolves, caribou, and moose all responded to the lower productivity of
deciduous vegetation in cutovers at high latitudes. Indeed, the selection of moose for
cutovers decreased with the availability of deciduous vegetation, which is lower at high
compared to low latitudes. This change in habitat selection behaviour is consistent with
previous reports that moose most strongly select cutovers with high lateral cover and high
deciduous regeneration (Potvin et al., 2004; Courtois et al., 1998). We demonstrate that this
response should reduce the relative probability of encounter between moose and wolves in

30
cutovers at high latitudes, thereby reducing the relative contribution of cutovers on the rate
of moose consumption by wolves (see Joly et al., 2003) in higher latitudes. The
consumption of moose by wolves is central to its apparent competition with the threatened
caribou, which should vary along the latitudinal axis. Furthermore, the lower availability of
deciduous vegetation in cutovers at high latitudes should result in a smaller (if any)
numerical response of moose towards the north (Potvin et al., 2004). As the availability of
deciduous forage decreases, the species would then rely on balsam fir (Crête et al., 1997),
which is a relatively poor food source (Peek, 1974). This consumption of poor food source
by moose would also reduce the potential for human-induced apparent competition.
Although we show that the potential for apparent competition triggered by deciduous
vegetation availability following harvest decreases with increasing latitude, we also
recognize that forestry activities produce extended road networks which can improve
predation efficiency of wolves and still impact caribou (James et al., 2000).
Understanding the complex interactions involved in food webs is fundamental to the
development of effective measures aimed at restoring or protecting ecosystem properties
despite human activities. Our study demonstrates that the impact of human activities on
indirect trophic interactions can vary along the latitudinal axis. According to Gosz (1992),
most of our current knowledge about ecosystem structure and function has been generated
from studies conducted over small areas. By revealing that predator-prey interactions
should vary along geographical gradients (Fortin et al., 2008), our study indicates why
particular management or conservation guidelines developed based on a particular
population may be less effective when applied to other populations. Even when developed
from studies conducted over large spatial extents, those guidelines may not be effective if
they only reflect the “average” response of multiple populations (e.g. Environnement
Canada, 2011). Indeed, when a geographical gradient in biological processes (e.g., direct
and indirect trophic interactions) affects a particular species, the use of a unique guideline
reflecting an average response should be most effective to manage the populations
established at the median location of all studied populations. Moving away from such a
location may eventually result in guidelines that become too permissive for adequate
species conservation. Understanding the dynamics, structure, and function of ecosystems
along geographical gradient should therefore increase our ability to manage these systems.

31
Acknowledgements
We are grateful to Philippe Goulet for assistance in obtaining and compiling forestry data.
We also thank Pierre Racine for his help with PostGIS, Bruno Baillargeon, Dominic
Grenier, Daniel Dorais, François-Alexis Behm and Richard Audy for their help in the field.
The NSERC–Université Laval Industrial Research Chair in Silviculture and Wildlife
(Chaire de recherche industrielle CRSNG-Université Laval en sylviculture et en faune) and
its partners founded this research.
References
APPS, C., MCLELLAN, B., KINLEY, A., SERROUYA, R., SEIP, D. & WITTMER, H. 2013. Spatial factors related to mortality and population decline of endangered mountain caribou. Journal of Wildlife Management, 77, 7, 1409-1419.
BATES, D., MAECHLER, M. & BOLKER, B. 2012. lme4: Linear mixed-effects models using S4 classes [En ligne]. Disponible: http://CRAN.R-project.org/package=lme4%5D.
BJØRNERAAS, K., SOLBERG, E. J., HERFINDAL, I., MOORTER, B., ROLANDSEN, C., TREMBLAY, J.-P., SKARPE, C., SÆTHER, B., ERIKSEN, R. & ASTRUP, R. 2011. Moose (Alces alces) habitat use at multiple temporal scales in a human-altered landscape. Wildlife Biology, 17, 1, 44-54.
BOWMAN, J., RAY, J. C., MAGOUN, A. J., JOHNSON, D. S. & DAWSON, F. N. 2010. Roads, logging, and the large-mammal community of an eastern Canadian boreal forest. Canadian Journal of Zoology, 88, 5, 454-467.
BOYCE, M., VERNIER, P., NIELSEN, S. E. & SCHMIEGELOW, F. 2002. Evaluating resource selection functions. Ecological Modelling, 157, 2, 281-300.
BURGMAN, M. & FOX, J. 2003. Bias in species range estimates from minimum convex polygons: implications for conservation and options for improved planning. Animal Conservation, 6, 1, 19-28.
CHAPIN III, F. S., ZAVALETA, E. S., EVINER, V. T., NAYLOR, R. L., VITOUSEK, P. M., REYNOLDS, H. L., HOOPER, D. U., LAVOREL, S., SALA, O. E. & HOBBIE, S. E. 2000. Consequences of changing biodiversity. Nature, 405, 6783, 234-242.
COHEN, J., COHEN, P., WEST, S. & AIKEN, L. 2013. Applied multiple regression/correlation analysis for the behavioral sciences, Routledge.
CONNELL, J. & ORIAS, E. 1964. The ecological regulation of species diversity. American Naturalist, 98, 903, 399-414.
COSEWIC. 2011. Canadian species at risk [En ligne]. Disponible: http://www.cosewic.gc.ca/fra/sct1/SearchResult_f.cfm?commonName=&scienceName=rangifer+tarandus&Submit=Soumettre [Consulté le 22/11/2012].

32
COURBIN, N., FORTIN, D., DUSSAULT, C. & COURTOIS, R. 2009. Landscape management for woodland caribou: the protection of forest blocks influences wolf-caribou co-occurrence. Landscape Ecology, 24, 10, 1375-1388.
COURTOIS, R., GINGRAS, A., FORTIN, D., SEBBANE, A., ROCHETTE, B. & BRETON, L. 2008. Demographic and behavioural response of woodland caribou to forest harvesting. Canadian Journal of Forest Research, 38, 11, 2837-2849.
COURTOIS, R., OUELLET, J. P. & GAGNÉ, B. 1998. Characteristics of cutovers used by moose (Alces Alces) in early winter. Alces, 34, 1, 201-211.
CRÊTE, M. & COURTOIS, R. 1997. Limiting factors might obscure population regulation of moose (Cervidae: Alces alces) in unproductive boreal forests. Journal of Zoology, 242, 4, 765-781.
DECESARE, N., HEBBLEWHITE, M., ROBINSON, H. & MUSIANI, M. 2010. Endangered, apparently: the role of apparent competition in endangered species conservation. Animal Conservation, 13, 4, 353-362.
ENVIRONNEMENT CANADA 2011. Évaluation scientifique aux fins de la désignation de l'habitat essentiel de la population boréale du caribou des bois (Rangifer tarandus caribou) au Canada; Mise à jour 2011.
ENVIRONNEMENT CANADA. 2014. La météo au Canada [En ligne]. Disponible: http://meteo.gc.ca/canada_f.html [Consulté le 2014-01-26].
FORTIN, D., COURTOIS, R., ETCHEVERRY, P., DUSSAULT, C. & GINGRAS, A. 2008. Winter selection of landscapes by woodland caribou: behavioural response to geographical gradients in habitat attributes. Journal of Applied Ecology, 45, 5, 1392-1400.
GILLIES, C., HEBBLEWHITE, M., NIELSEN, S., KRAWCHUK, M., ALDRIDGE, C., FRAIR, J., SAHER, J., STEVENS, C. & JERDE, C. 2006. Application of random effects to the study of resource selection by animals. Journal of Animal Ecology, 75, 4, 887-898.
GINGRAS, A., AUDY, A. & COURTOIS, R. 1989. Inventaire aérien de l'orignal dans la zone de chasse 19 à l'hiver 1987-1988. Ministère du Loisir de la Chasse et de la Pêche.
GOSZ, J. 1992. Gradient analysis of ecological change in time and space: implications for forest management. Ecological Applications, 2, 3, 248-261.
HANSKI, I., HENTTONEN, H., KORPIMÄKI, E., OKSANEN, L. & TURCHIN, P. 2001. Small-rodent dynamics and predation. Ecology, 82, 6, 1505-1520.
HEBBLEWHITE, M. & MERRILL, E. 2008. Modelling wildlife–human relationships for social species with mixed‐effects resource selection models. Journal of Applied Ecology, 45, 3, 834-844.
HEBBLEWHITE, M., WHITE, C., NIETVELT, C., MCKENZIE, J., HURD, T., FRYXELL, J., BAYLEY, S. & PAQUET, P. 2005. Human activity mediates a trophic cascade caused by wolves. Ecology, 86, 8, 2135-2144.
HOLT, R. D. 1984. Spatial heterogeneity, indirect interactions, and the coexistence of prey species. American Naturalist, 124, 3, 377-406.
HUANG, J., TARDIF, J., BERGERON, Y., DENNELER, B., BERNINGER, F. & GIRARDIN, M. 2010. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Global Change Biology, 16, 2, 711-731.

33
HUETE, A., DIDAN, K., MIURA, T., RODRIGUEZ, E. P., GAO, X. & FERREIRA, L. G. 2002. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 83, 1, 195-213.
JAMES, A. R., BOUTIN, S., HEBERT, D. M., RIPPIN, A. B. & WHITE, J. 2004. Spatial separation of caribou from moose and its relation to predation by wolves. Journal of Wildlife Management, 68, 4, 799-809.
JAMES, A. R. C. & STUART-SMITH, A. K. 2000. Distribution of caribou and wolves in relation to linear corridors. Journal of Wildlife Management, 64, 1, 154-159.
JOBIDON, R. 1994. Autécologie de quelques espèces de compétition d'importance pour la régénération forestière au Québec: revue de littérature.
JOHNSON, D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61, 1, 65-71.
JOLY, D. O. & PATTERSON, B. R. 2003. Use of selection indices to model the functional response of predators. Ecology, 84, 6, 1635-1639.
LAPOINTE-GARANT, M. P., HUANG, J. G., GEA‐IZQUIERDO, G., RAULIER, F., BERNIER, P. & BERNINGER, F. 2010. Use of tree rings to study the effect of climate change on trembling aspen in Quebec. Global Change Biology, 16, 7, 2039-2051.
LATHAM, A. D. M., LATHAM, M. C., KNOPFF, K. H., HEBBLEWHITE, M. & BOUTIN, S. 2013. Wolves, white‐tailed deer, and beaver: implications of seasonal prey switching for woodland caribou declines. Ecography, 36, 12, 1276-1290.
MANLY, B., MCDONALD, L., THOMAS, D., MCDONALD, T. & ERICKSON, W. 2002. Resource selection by animals: statistical analysis and design for field studies. Kluwer: Boston, MA.
MELIS, C., JĘDRZEJEWSKA, B., APOLLONIO, M., BARTOŃ, K. A., JĘDRZEJEWSKI, W., LINNELL, J. D., KOJOLA, I., KUSAK, J., ADAMIC, M. & CIUTI, S. 2009. Predation has a greater impact in less productive environments: variation in roe deer, (Capreolus capreolus) population density across Europe. Global Ecology and Biogeography, 18, 6, 724-734.
MESSIER, F. 1985. Social organization, spatial distribution, and population density of wolves in relation to moose density. Canadian Journal of Zoology, 63, 5, 1068-1077.
OBE, R. & HSU, L. S. 2011. PostGIS in action. Manning Publications. Stanford, CT, USA. PEEK, J. 1974. A review of moose food habits studies in North America. Naturaliste
Canadien, 101, 1-2, 195-215. PETERS, W., HEBBLEWHITE, M., DECESARE, N., CAGNACCI, F. & MUSIANI, M.
2013. Resource separation analysis with moose indicates threats to caribou in human altered landscapes. Ecography, 36, 4, 487-498.
POTVIN, F. & COURTOIS, R. 2004. Winter presence of moose in clear-cut black spruce landscapes: related to spatial pattern or to vegetation. Alces, 40, 61-70.
R DEVELOPMENT CORE TEAM 2010. R: a language and environment for Statistical Computing. Vienna.
RESSOURCES NATURELLES CANADA 2000. Service canadien des forêts - Centre de foresterie des Laurentides (RNCAN), 2006.Cartographie intermédiaire en 52 classes de la couverture terrestre du Québec (CCTQ) réalisée dans le cadre du projet Observation de la Terre pour le développement durable des forêts (OTTD) à partir

34
des images Landsat ca. 2000. La couverture terrestre au nord de la limite terrestre des forêts fut complétée dans le cadre d'un partenariat avec le MDDEP.
ROEMER, G. W., COONAN, T. J., GARCELON, D. K., BASCOMPTE, J. & LAUGHRIN, L. 2001. Feral pigs facilitate hyperpredation by golden eagles and indirectly cause the decline of the island fox. Animal Conservation, 4, 4, 307-318.
ROSENZWEIG, M. L. 1968. Net primary productivity of terrestrial communities: prediction from climatological data. American Naturalist, 102, 923, 67-74.
SALA, O. E., CHAPIN, F. S., ARMESTO, J. J., BERLOW, E., BLOOMFIELD, J., DIRZO, R., HUBER-SANWALD, E., HUENNEKE, L. F., JACKSON, R. B. & KINZIG, A. 2000. Global biodiversity scenarios for the year 2100. Science, 287, 5459, 1770-1774.
SCHAEFER, J. A. 2003. Long‐term range recession and the persistence of caribou in the Taiga. Conservation Biology, 17, 5, 1435-1439.
SCHEMSKE, D. W., MITTELBACH, G. G., CORNELL, H. V., SOBEL, J. M. & ROY, K. 2009. Is there a latitudinal gradient in the importance of biotic interactions? Annual Review of Ecology Evolution, and Systematics, 40, 245-269.
SCHWARZ, G. 1978. Estimating the dimension of a model. The Annals of Statistics, 6, 2, 461-464.
SEIP, D. R. 1992. Factors limiting woodland caribou populations and their interrelationships with wolves and moose in southeastern British Columbia. Canadian Journal of Zoology, 70, 8, 1494-1503.
STOHLGREN, T. J., BARNETT, D., FLATHER, C., KARTESZ, J. & PETERJOHN, B. 2005. Plant species invasions along the latitudinal gradient in the United States. Ecology, 86, 9, 2298-2309.
TURCHIN, P. & HANSKI, I. 1997. An empirically based model for latitudinal gradient in vole population dynamics. American Naturalist, 149, 5, 842-874.
VORS, L. S. & BOYCE, M. S. 2009. Global declines of caribou and reindeer. Global Change Biology, 15, 11, 2626-2633.
WITTMER, H. U., SERROUYA, R., ELBROCH, L. M. & MARSHALL, A. J. 2013. Conservation strategies for species affected by apparent competition. Conservation Biology, 27, 2, 254-260.

35
CONCLUSION GÉNÉRALE
Il existe plusieurs exemples de systèmes où l’intensité des interactions trophiques directes
est influencée par un gradient latitudinal, notamment par une variation de la pression de
prédation. Par exemple, la prédation effectuée sur des appâts constitués de larves par des
fourmis a été étudiée sur un gradient latitudinal de plus de 40 degrés (Jeanne, 1979). Le
taux de prédation par les fourmis était significativement plus élevé dans les zones tropicales
que dans les zones tempérées. Ceci s’explique par le fait qu’un appât de nourriture était
exploité par les fourmis en significativement moins de temps dans les régions situées près
de l’Équateur. De façon similaire, lorsque l’on compare les taux de prédation observés sur
des nids de plusieurs espèces d’oiseaux provenant d’une part des régions tropicales
(Kulesza, 1990; Robinson et al., 2000) et de l’autre des zones tempérées (Conway et al.,
2000), nous remarquons que les espèces tropicales subissent des taux de prédation des nids
significativement plus élevés (Schemske et al., 2009). Toutefois, il existe peu d’études qui
se sont concentrées sur l’influence de gradients géographiques sur les interactions
trophiques indirectes alors que l’intensité de ces relations peut fortement influencer la
persistance d’espèces vulnérables (DeCesare et al., 2010). Notre étude démontre qu’une
interaction trophique indirecte telle la compétition apparente peut varier le long de
gradients géographiques. En effet, nos résultats révèlent que le rôle potentiel des
perturbations anthropiques dans la mise en place d’une compétition apparente entre le
caribou forestier et l’orignal devrait diminuer alors que l’industrie forestière progresse vers
le nord. La région la plus au sud de la forêt boréale canadienne a subi une exploitation
intensive au cours des différentes années et cette exploitation continue de progresser vers le
nord (Schaefer, 2003). Nos résultats sont donc d’une importance accrue puisqu’ils
démontrent que les conséquences de la coupe forestière sur les interactions trophiques
affectant le caribou pourraient varier selon un axe latitudinal.
Au cours des dernières années, un grand nombre d’études se sont penchées sur les
conséquences des perturbations anthropiques sur la conservation du caribou et plus
particulièrement, sur les différents aspects de la compétition apparente. Plusieurs
informations tirées de ces études ont amélioré les stratégies de conservation par exemple au

36
niveau des différents risques de mortalités associés à certaines saisons (Latham et al.,
2013), des chevauchements spatiaux entre l’orignal et le caribou qui sont plus fréquents
dans les milieux perturbés (Peters et al., 2013) ou alors de l’utilisation de zones en altitude
ainsi que des marais et des tourbières comme refuge spatial par le caribou pour diminuer le
risque de mortalité (Apps et al., 2013; James et al., 2004). Toutefois, à notre connaissance,
notre étude est la première à démontrer la présence d’un gradient latitudinal influençant le
potentiel de compétition apparente entre le caribou forestier et l’orignal. Nous avons
démontré que les conséquences de la coupe forestière sur le potentiel de compétition
apparente devraient diminuer alors que les activités d’exploitation progressent vers le nord
étant donné la diminution décelable de l’enfeuillement des parterres de coupe. Pour ce
faire, nous avons d’abord déterminé qu’un gradient latitudinal influençait la moyenne
pondérée de l’Indice de Végétation Amélioré (Enhanced Vegetation Index, EVI), un indice
qui covariait étroitement avec la proportion des parterres de coupe forestière couverte par
de la végétation décidue dans notre région d’étude. L’enfeuillement des parterres de coupe
augmentait avec une diminution de la latitude ; les parterres de coupes dans les forêts
boréales du sud étant colonisés par plus de végétation feuillue. Finalement, nous avons
utilisé la modélisation de la sélection d’habitat pour évaluer la réaction comportementale
des trois espèces de grands mammifères impliquées dans la compétition apparente soit
l’orignal, le loup ainsi que le caribou forestier, face à ces variations dans l’intensité de
l’enfeuillement. Comme prédit, un enfeuillement plus important des parterres de coupe a
résulté en une plus grande sélection de ces parcelles par l’orignal et le loup alors qu’il a
engendré un plus grand évitement chez le caribou forestier.
Nos résultats démontrent que l’enfeuillement des parterres de coupes diminue avec
une augmentation de la latitude. Certaines études ont déjà établi la présence d’un gradient
latitudinal influençant la croissance des feuillus. Par exemple, lorsque la latitude augmente,
la présence de formes arborescentes d’espèces feuillues comme le peuplier faux-tremble ou
le bouleau à papier, diminue de plus en plus pour laisser place à des formes plutôt
arbustives, plus courantes dans les forêts du nord (Jobidon, 1994). La croissance radiale de
ces deux espèces diffère également selon la latitude et la température à laquelle l’arbre se
retrouve (Huang et al., 2010) et la productivité du peuplier faux-tremble diminue aussi avec
une augmentation de la latitude (Lapointe-Garant et al., 2010). Nous avons toutefois su

37
détecter ce patron au long d’un gradient latitudinal plutôt court de 400 km. La disponibilité
en végétation décidue reste plus élevée dans la partie sud que dans la partie nord de notre
aire d’étude au cours des 75 premières années suivant la coupe forestière.
xxxUne baisse de l’enfeuillement des parterres de coupe forestière à haute latitude
devrait engendrer une diminution de l’accroissement des populations d’orignaux lorsque
l’on se dirige vers le nord (Potvin et al., 2004). En effet, une diminution de l’enfeuillement
devrait inciter les orignaux à consommer davantage une nourriture de moins bonne qualité,
comme le sapin baumier (Crête et al., 1997; Peek, 1974). Ceci pourrait ainsi grandement
diminuer le potentiel de compétition apparente engendré par les perturbations anthropiques.
En effet, la sélection des parcelles de coupe forestière par l’orignal augmentait en fonction
de l’enfeuillement des parterres. Cette variation dans la réaction comportementale vient
corroborer les différentes études faites au niveau de la sélection de l’orignal à l’intérieur
même de la parcelle de coupe, qui notaient une attraction plus particulière pour les milieux
riches en feuillus (Potvin et al., 2004; Courtois et al., 1998). De plus, nos résultats
démontrent que la probabilité de cooccurrence entre l’orignal et le loup au niveau des
parterres de coupe forestière diminue avec une augmentation de la latitude. Ceci devrait
réduire la contribution relative de la coupe forestière sur le taux de consommation
d’orignaux par les loups (voir Joly et al., 2003) dans les régions plus au nord de la forêt
boréale. La consommation d’orignaux par le loup est un point central influençant la
compétition apparente avec le caribou forestier ; ce phénomène devrait donc varier selon un
axe latitudinal. Nos résultats sont d’autant plus importants étant donné la grande utilisation
des coupes forestières par les orignaux et les loups. En effet, malgré une faible
représentation de ce milieu dans notre aire d’étude (moins de 2%), le temps d’utilisation de
celui-ci par les orignaux et les loups sont élevé (8,1 et 15,6% respectivement). Il est donc
important de bien comprendre les variables qui influencent la régénération des parterres de
coupe. Bien que le potentiel de compétition apparente engendré par l’enfeuillement des
parterres de coupe forestière diminue avec une augmentation de la latitude, nous
reconnaissons que les réseaux routiers façonnés par l’industrie forestière pourraient
améliorer l’efficacité de prédation des loups et ainsi avoir un impact négatif sur le caribou
(James et al., 2000).
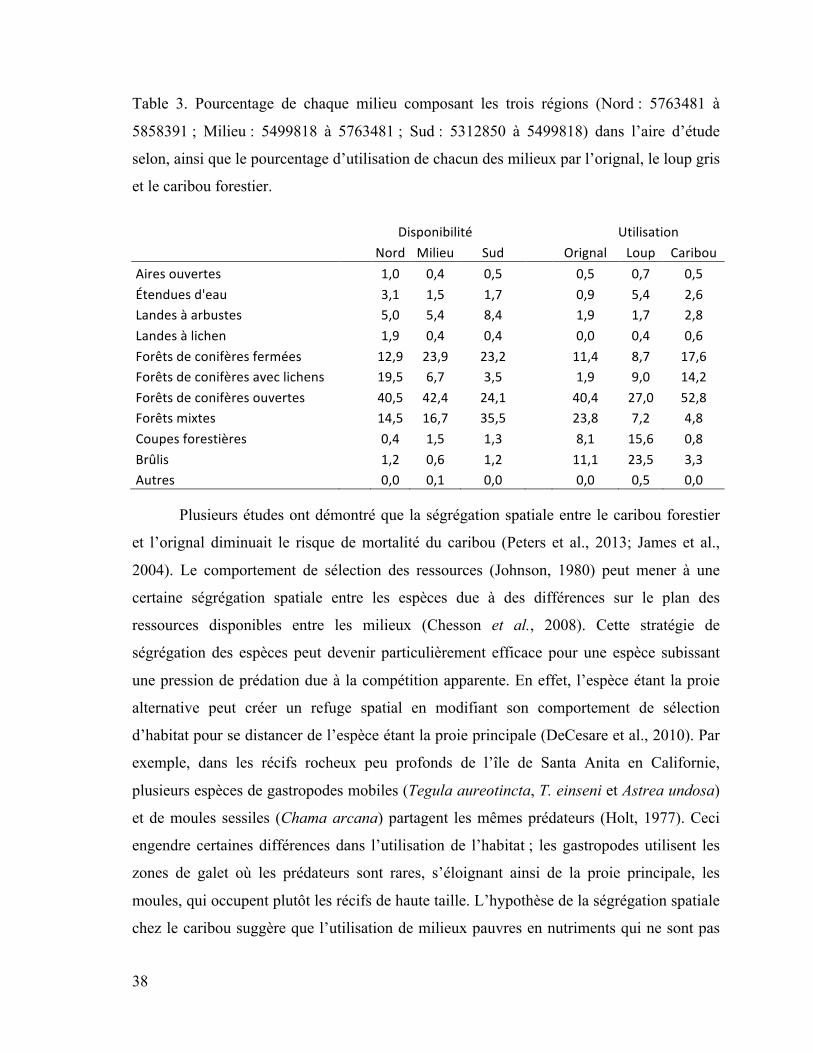
38
Table 3. Pourcentage de chaque milieu composant les trois régions (Nord : 5763481 à
5858391 ; Milieu : 5499818 à 5763481 ; Sud : 5312850 à 5499818) dans l’aire d’étude
selon, ainsi que le pourcentage d’utilisation de chacun des milieux par l’orignal, le loup gris
et le caribou forestier.
Disponibilité Utilisation Nord Milieu Sud Orignal Loup Caribou Aires ouvertes
1,0 0,4 0,5
0,5 0,7 0,5
Étendues d'eau
3,1 1,5 1,7
0,9 5,4 2,6 Landes à arbustes
5,0 5,4 8,4
1,9 1,7 2,8
Landes à lichen
1,9 0,4 0,4
0,0 0,4 0,6 Forêts de conifères fermées
12,9 23,9 23,2
11,4 8,7 17,6
Forêts de conifères avec lichens
19,5 6,7 3,5
1,9 9,0 14,2 Forêts de conifères ouvertes
40,5 42,4 24,1
40,4 27,0 52,8
Forêts mixtes
14,5 16,7 35,5
23,8 7,2 4,8 Coupes forestières
0,4 1,5 1,3
8,1 15,6 0,8
Brûlis
1,2 0,6 1,2
11,1 23,5 3,3 Autres 0,0 0,1 0,0 0,0 0,5 0,0
Plusieurs études ont démontré que la ségrégation spatiale entre le caribou forestier
et l’orignal diminuait le risque de mortalité du caribou (Peters et al., 2013; James et al.,
2004). Le comportement de sélection des ressources (Johnson, 1980) peut mener à une
certaine ségrégation spatiale entre les espèces due à des différences sur le plan des
ressources disponibles entre les milieux (Chesson et al., 2008). Cette stratégie de
ségrégation des espèces peut devenir particulièrement efficace pour une espèce subissant
une pression de prédation due à la compétition apparente. En effet, l’espèce étant la proie
alternative peut créer un refuge spatial en modifiant son comportement de sélection
d’habitat pour se distancer de l’espèce étant la proie principale (DeCesare et al., 2010). Par
exemple, dans les récifs rocheux peu profonds de l’île de Santa Anita en Californie,
plusieurs espèces de gastropodes mobiles (Tegula aureotincta, T. einseni et Astrea undosa)
et de moules sessiles (Chama arcana) partagent les mêmes prédateurs (Holt, 1977). Ceci
engendre certaines différences dans l’utilisation de l’habitat ; les gastropodes utilisent les
zones de galet où les prédateurs sont rares, s’éloignant ainsi de la proie principale, les
moules, qui occupent plutôt les récifs de haute taille. L’hypothèse de la ségrégation spatiale
chez le caribou suggère que l’utilisation de milieux pauvres en nutriments qui ne sont pas

39
privilégiés par l’orignal ni par son prédateur le loup, permet une séparation spatiale entre le
caribou et l’orignal, diminuant ainsi la pression de prédation exercée sur le caribou (Seip,
1992; Bergerud et al., 1987). Nos résultats supportent cette hypothèse en démontrant que
les caribous évitent plus intensément les parcelles qui sont particulièrement privilégiées par
les orignaux. Ceci pourrait indiquer que la sélection naturelle favorise les caribous
forestiers qui se distancent des milieux où ils courent un risque plus élevé de croiser des
orignaux, diminuant ainsi les risques de prédation par le loup.
Comprendre les interactions complexes au niveau des réseaux trophiques permet de
développer des mesures de conservation efficaces visant à restaurer et protéger des
écosystèmes malgré les activités humaines. Nous avons démontré qu’il est important de
tenir compte des gradients géographiques lors de l’élaboration de plans d’aménagement sur
de grands territoires. En effet, la contribution de la coupe forestière à la compétition
apparente affectant le caribou forestier est influencée par un gradient latitudinal. Beaucoup
des connaissances scientifiques modernes sur les structures et le fonctionnement des
écosystèmes dérivent d’études effectuées sur de petits territoires et qui supposent que le
climat est constant et qu’il n’y a pas ou très peu de variabilité spatiale (Gosz, 1992). Les
plans d’aménagement établis en considérant ce savoir scientifique sont donc spécifiques à
de petites régions et à certaines populations. Ces plans d’aménagement peuvent se révéler
obsolètes s’ils ne tiennent pas compte de la plasticité possible dans la réaction
comportementale des espèces à travers des gradients géographiques, comme démontré dans
notre étude. Des plans d’aménagement basés sur des études conduites sur de grands
territoires peuvent aussi être moins efficaces si ces plans sont développés grâce à la réaction
moyenne de plusieurs populations. Par exemple, le seuil de perturbation (incluant coupes
forestières, chemins et autres structures anthropiques) de 35 % permettant la subsistance du
caribou forestier (c.-à-d., 60% des chances de la population ne décline pas) a été suggéré
par une étude faite sur plusieurs populations réparties à travers la forêt boréale canadienne
(e.g. Environnement Canada, 2011). Nos résultats démontrent que ce seuil de perturbation
pourrait être trop élevé dans les régions plus au sud de la forêt boréale où la contribution de
la coupe forestière à la compétition apparente affectant le caribou forestier est plus élevée.
De ce fait, il existe un risque qu’un plan de rétablissement unique appliqué sur de grands
secteurs soit trop permissif pour protéger adéquatement les populations au sud de la

40
location médiane des populations étudiées. Les gradients géographiques pourraient y avoir
de forts impacts sur les menaces affectant les espèces vulnérables, comme démontré dans
notre étude. Comprendre la dynamique, la structure et la fonction des écosystèmes le long
de gradients géographiques devrait augmenter notre capacité à gérer ces écosystèmes.
Perspectives
Cette étude permet de mieux comprendre les conséquences de la coupe forestière en forêt
boréale non seulement au niveau des peuplements, mais aussi sur le comportement des
grands mammifères. Nous avons démontré qu’un gradient latitudinal affectait
l’enfeuillement des parterres de coupe et que ces différences rencontrées au sein des
peuplements faisaient varier la contribution de la coupe forestière au potentiel de
compétition apparente affectant le caribou forestier en modifiant la sélection des parterres
de coupe par l’orignal et le loup. Il serait intéressant d’évaluer si le potentiel de compétition
apparente plus élevé au sud qu’au nord selon les résultats de notre étude se traduit
effectivement en un risque de mortalité plus élevé pour les caribous au sud. Étant donné
que des perturbations naturelles comme le feu peuvent aussi rajeunir les forêts et attirer les
orignaux ainsi que les loups, il serait intéressant d’évaluer s’il y a présence d’un gradient
latitudinal dans l’enfeuillement des parterres brûlés. Finalement, plusieurs autres variables
environnementales pourraient influencer l’enfeuillement des parterres, par exemple les
variations climatiques (p. ex., précipitations) attribuables aux oscillations nord-atlantiques
(NAO) ou même les conditions édaphiques, et il serait intéressant d’étudier leurs éventuels
impacts additionnels sur l’enfeuillement pour pouvoir mieux prédire les conséquences de la
coupe forestière sur les peuplements forestiers.

41
BIBLIOGRAPHIE GÉNÉRALE
APPS, C., MCLELLAN, B., KINLEY, A., SERROUYA, R., SEIP, D. & WITTMER, H. 2013. Spatial factors related to mortality and population decline of endangered mountain caribou. Journal of Wildlife Management, 77, 7, 1409-1419.
BASILLE, M., FORTIN, D., DUSSAULT, C., OUELLET, J. P. & COURTOIS, R. 2012. Ecologically based definition of seasons clarifies predator-prey interactions. Ecography, 36, 2, 001-010.
BATES, D., MAECHLER, M. & BOLKER, B. 2012. lme4: Linear mixed-effects models using S4 classes [En ligne]. Disponible: http://CRAN.R-project.org/package=lme4%5D.
BEAUCHESNE, D., JAEGER, J. & ST-LAURENT, M.-H. 2013. Disentangling woodland caribou movements in response to clearcuts and roads across temporal scales. PloS one, 8, 11.
BERGERUD, A. & PAGE, R. 1987. Displacement and dispersion of parturient caribou at calving as antipredator tactics. Canadian Journal of Zoology, 65, 7, 1597-1606.
BERGERUD, A. T. 1974. Decline of caribou in North America following settlement. Journal of Wildlife Management, 38, 4, 757-770.
BERGERUD, A. T. & ELLIOT, J. P. 1986. Dynamics of caribou and wolves in northern British Columbia. Canadian Journal of Zoology, 64, 7, 1515-1529.
BJØRNERAAS, K., SOLBERG, E. J., HERFINDAL, I., MOORTER, B., ROLANDSEN, C., TREMBLAY, J.-P., SKARPE, C., SÆTHER, B., ERIKSEN, R. & ASTRUP, R. 2011. Moose (Alces alces) habitat use at multiple temporal scales in a human-altered landscape. Wildlife Biology, 17, 1, 44-54.
BOAN, J. J., MCLAREN, B. E. & MALCOLM, J. R. 2011. Influence of post-harvest silviculture on understory vegetation: Implications for forage in a multi-ungulate system. Forest Ecology and Management, 262, 9, 1704-1712.
BOUCHER, Y., ARSENEAULT, D., SIROIS, L. & BLAIS, L. 2009. Logging pattern and landscape changes over the last century at the boreal and deciduous forest transition in Eastern Canada. Landscape Ecology, 24, 2, 171-184.
BOWMAN, J., RAY, J. C., MAGOUN, A. J., JOHNSON, D. S. & DAWSON, F. N. 2010. Roads, logging, and the large-mammal community of an eastern Canadian boreal forest. Canadian Journal of Zoology, 88, 5, 454-467.
BOYCE, M., VERNIER, P., NIELSEN, S. E. & SCHMIEGELOW, F. 2002. Evaluating resource selection functions. Ecological Modelling, 157, 2, 281-300.
BRYANT, A. & PAGE, R. 2005. Timing and causes of mortality in the endangered Vancouver Island marmot (Marmota vancouverensis). Canadian Journal of Zoology, 83, 5, 674-682.
BURGMAN, M. & FOX, J. 2003. Bias in species range estimates from minimum convex polygons: implications for conservation and options for improved planning. Animal Conservation, 6, 1, 19-28.
CHAPIN III, F. S., ZAVALETA, E. S., EVINER, V. T., NAYLOR, R. L., VITOUSEK, P. M., REYNOLDS, H. L., HOOPER, D. U., LAVOREL, S., SALA, O. E. & HOBBIE, S. E. 2000. Consequences of changing biodiversity. Nature, 405, 6783, 234-242.

42
CHESSON, P. & KUANG, J. 2008. The interaction between predation and competition. Nature, 456, 7219, 235-238.
COHEN, J., COHEN, P., WEST, S. & AIKEN, L. 2013. Applied multiple regression/correlation analysis for the behavioral sciences, Routledge.
CONNELL, J. & ORIAS, E. 1964. The ecological regulation of species diversity. American Naturalist, 98, 903, 399-414.
CONWAY, C. & MARTIN, T. 2000. Evolution of passerine incubation behavior: influence of food, temperature, and nest predation. Evolution, 54, 2, 670-685.
COSEWIC. 2011. Canadian species at risk [En ligne]. Disponible: http://www.cosewic.gc.ca/fra/sct1/SearchResult_f.cfm?commonName=&scienceName=rangifer+tarandus&Submit=Soumettre [Consulté le 22/11/2012].
COURBIN, N. 2013. Interactions entre le loup gris, le caribou forestier et l'orignal en forêt boréale aménagée. Université Laval.
COURBIN, N., FORTIN, D., DUSSAULT, C. & COURTOIS, R. 2009. Landscape management for woodland caribou: the protection of forest blocks influences wolf-caribou co-occurrence. Landscape Ecology, 24, 10, 1375-1388.
COURTOIS, R., DUSSAULT, C., POTVIN, F. & DAIGLE, G. 2002. Habitat selection by moose (Alces alces) in clear-cut landscapes. Alces, 38, 177-192.
COURTOIS, R., GINGRAS, A., FORTIN, D., SEBBANE, A., ROCHETTE, B. & BRETON, L. 2008. Demographic and behavioural response of woodland caribou to forest harvesting. Canadian Journal of Forest Research, 38, 11, 2837-2849.
COURTOIS, R., OUELLET, J. P., BRETON, L., GINGRAS, A. & DUSSAULT, C. 2007. Effects of forest disturbance on density, space use, and mortality of woodland caribou. Ecoscience, 14, 4, 491-498.
COURTOIS, R., OUELLET, J. P. & GAGNÉ, B. 1998. Characteristics of cutovers used by moose (Alces Alces) in early winter. Alces, 34, 1, 201-211.
COURTOIS, R., OUELLET, J. P., GINGRAS, A., DUSSAULT, C., BRETON, L. & MALTAIS, J. 2003. Historical changes and current distribution of caribou (Rangifer Tarandus) in Quebec. The Canadian Field-Naturalist, 117, 3, 400-414.
CRÊTE, M. 1977. Importance de la coupe forestière sur l'habitat hivernal de l'orignal dans le sud-ouest du Québec. Canadian Journal of Forest Research, 7, 2, 241-257.
CRÊTE, M. & COURTOIS, R. 1997. Limiting factors might obscure population regulation of moose (Cervidae: Alces alces) in unproductive boreal forests. Journal of Zoology, 242, 4, 765-781.
DECESARE, N., HEBBLEWHITE, M., ROBINSON, H. & MUSIANI, M. 2010. Endangered, apparently: the role of apparent competition in endangered species conservation. Animal Conservation, 13, 4, 353-362.
DOU, H., JIANG, G., STOTT, P. & PIAO, R. 2013. Climate change impacts population dynamics and distribution shift of moose (Alces alces) in Heilongjiang Province of China. Ecological Research, 28, 4, 1-8.
DULVY, N., FRECKLETON, R. & POLUNIN, N. 2004. Coral reef cascades and the indirect effects of predator removal by exploitation. Ecology letters, 7, 5, 410-416.
DUSSART, E. & PAYETTE, S. 2002. Ecological impact of clear-cutting on black spruce-moss forests in southern Québec. Ecoscience, 9, 4, 533-543.
DUSSAULT, C., PINARD, V., OUELLET, J. P., COURTOIS, R. & FORTIN, D. 2012. Avoidance of roads and selection for recent cutovers by threatened caribou: fitness-

43
rewarding or maladaptive behaviour? Proceedings of Biological Sciences, 279, 1746, 4481-4488.
DYER, S., O'NEILL, J., WASEL, S. & BOUTIN, S. 2001. Avoidance of industrial development by woodland caribou. Journal of Wildlife Management, 65, 3, 531-542.
ELSON, L., SIMON, N. & KNEESHAW, D. 2007. Regeneration differences between fire and clearcut logging in southeastern Labrador: a multiple spatial scale analysis. Canadian Journal of Forest Research, 37, 2, 473-480.
ENVIRONNEMENT CANADA 2011. Évaluation scientifique aux fins de la désignation de l'habitat essentiel de la population boréale du caribou des bois (Rangifer tarandus caribou) au Canada; Mise à jour 2011.
ENVIRONNEMENT CANADA. 2014. La météo au Canada [En ligne]. Disponible: http://meteo.gc.ca/canada_f.html [Consulté le 2014-01-26].
FAILLE, G., DUSSAULT, C., OUELLET, J. P., FORTIN, D., COURTOIS, R., ST-LAURENT, M. H. & DUSSAULT, C. 2010. Range fidelity: The missing link between caribou decline and habitat alteration? Biological Conservation, 143, 11, 2840-2850.
FORTIN, D., COURTOIS, R., ETCHEVERRY, P., DUSSAULT, C. & GINGRAS, A. 2008. Winter selection of landscapes by woodland caribou: behavioural response to geographical gradients in habitat attributes. Journal of Applied Ecology, 45, 5, 1392-1400.
GILLIES, C., HEBBLEWHITE, M., NIELSEN, S., KRAWCHUK, M., ALDRIDGE, C., FRAIR, J., SAHER, J., STEVENS, C. & JERDE, C. 2006. Application of random effects to the study of resource selection by animals. Journal of Animal Ecology, 75, 4, 887-898.
GINGRAS, A., AUDY, A. & COURTOIS, R. 1989. Inventaire aérien de l'orignal dans la zone de chasse 19 à l'hiver 1987-1988. Ministère du Loisir de la Chasse et de la Pêche.
GIROUX, J. F. & BERGERON, R. 1996. Spring diets of sympatric greater snow geese and Canada geese in southern Quebec. Canadian Journal of Zoology, 74, 5, 950-953.
GOSZ, J. 1992. Gradient analysis of ecological change in time and space: implications for forest management. Ecological Applications, 2, 3, 248-261.
HANSKI, I., HENTTONEN, H., KORPIMÄKI, E., OKSANEN, L. & TURCHIN, P. 2001. Small-rodent dynamics and predation. Ecology, 82, 6, 1505-1520.
HART, S. A. & CHEN, H. Y. H. 2008. Fire, logging, and overstory affect understory abundance, diversity, and composition in boreal forest. Ecological Monographs, 78, 1, 123-140.
HEBBLEWHITE, M. & MERRILL, E. 2008. Modelling wildlife–human relationships for social species with mixed‐effects resource selection models. Journal of Applied Ecology, 45, 3, 834-844.
HEBBLEWHITE, M., WHITE, C., NIETVELT, C., MCKENZIE, J., HURD, T., FRYXELL, J., BAYLEY, S. & PAQUET, P. 2005. Human activity mediates a trophic cascade caused by wolves. Ecology, 86, 8, 2135-2144.
HÉBERT, I. & WELADJI, R. 2013. The use of coniferous forests and cutovers by Newfoundland woodland caribou. Forest Ecology and Management, 291, 318-325.

44
HERVIEUX, D., HEBBLEWHITE, M., DECESARE, N., RUSSELL, M., SMITH, K., ROBERTSON, S. & BOUTIN, S. 2013. Widespread declines in woodland caribou (Rangifer tarandus caribou) continue in Alberta. Canadian Journal of Zoology, 91, 12, 872-882.
HINS, C., OUELLET, J. P., DUSSAULT, C. & ST-LAURENT, M.-H. 2009. Habitat selection by forest-dwelling caribou in managed boreal forest of eastern Canada: Evidence of a landscape configuration effect. Forest Ecology and Management, 257, 2, 636-643.
HOLT, R. D. 1977. Predation, apparent competition, and the structure of prey communities. Theoretical population biology, 12, 2, 197-229.
HOLT, R. D. 1984. Spatial heterogeneity, indirect interactions, and the coexistence of prey species. American Naturalist, 124, 3, 377-406.
HOLT, R. D. & LAWTON, J. H. 1993. Apparent competition and enemy-free space in insect host-parasitoid communities. American Naturalist, 142, 4, 623-645.
HUANG, J., TARDIF, J., BERGERON, Y., DENNELER, B., BERNINGER, F. & GIRARDIN, M. 2010. Radial growth response of four dominant boreal tree species to climate along a latitudinal gradient in the eastern Canadian boreal forest. Global Change Biology, 16, 2, 711-731.
HUETE, A., DIDAN, K., MIURA, T., RODRIGUEZ, E. P., GAO, X. & FERREIRA, L. G. 2002. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 83, 1, 195-213.
IUCN. 2011. The IUCN Red list of Threatened Species [En ligne]. Disponible: http://www.iucnredlist.org/details/15955/0 [Consulté le 10/11/2012].
JAMES, A. R., BOUTIN, S., HEBERT, D. M., RIPPIN, A. B. & WHITE, J. 2004. Spatial separation of caribou from moose and its relation to predation by wolves. Journal of Wildlife Management, 68, 4, 799-809.
JAMES, A. R. C. & STUART-SMITH, A. K. 2000. Distribution of caribou and wolves in relation to linear corridors. Journal of Wildlife Management, 64, 1, 154-159.
JEANNE, R. L. 1979. A latitudinal gradient in rates of ant predation. Ecology, 60, 6, 1211-1224.
JOBIDON, R. 1994. Autécologie de quelques espèces de compétition d'importance pour la régénération forestière au Québec: revue de littérature.
JOHNSON, D. H. 1980. The comparison of usage and availability measurements for evaluating resource preference. Ecology, 61, 1, 65-71.
JOLY, D. O. & PATTERSON, B. R. 2003. Use of selection indices to model the functional response of predators. Ecology, 84, 6, 1635-1639.
KULESZA, G. 1990. An analysis of clutch‐size in New World passerine birds. Ibis, 132, 3, 407-422.
LAPOINTE-GARANT, M. P., HUANG, J. G., GEA‐IZQUIERDO, G., RAULIER, F., BERNIER, P. & BERNINGER, F. 2010. Use of tree rings to study the effect of climate change on trembling aspen in Quebec. Global Change Biology, 16, 7, 2039-2051.
LAQUERRE, S., LEDUC, A. & HARVEY, B. D. 2009. Augmentation du couvert en peuplier faux-tremble dans les pessières noires du nord-ouest du Québec après coupe totale. Ecoscience, 16, 4, 483-491.

45
LATHAM, A. D. M., LATHAM, M. C., KNOPFF, K. H., HEBBLEWHITE, M. & BOUTIN, S. 2013. Wolves, white‐tailed deer, and beaver: implications of seasonal prey switching for woodland caribou declines. Ecography, 36, 12, 1276-1290.
MANLY, B., MCDONALD, L., THOMAS, D., MCDONALD, T. & ERICKSON, W. 2002. Resource selection by animals: statistical analysis and design for field studies. Kluwer: Boston, MA.
MANSSON, J., BUNNEFELD, N., ANDREN, H. & ERICSSON, G. 2012. Spatial and temporal predictions of moose winter distribution. Oecologia, 170, 2, 411-419.
MCRAE, D., DUCHESNE, L., FREEDMAN, B., LYNHAM, T. & WOODLEY, S. 2001. Comparisons between wildfire and forest harvesting and their implications in forest management. Environmental Reviews, 9, 4, 223-260.
MELIS, C., JĘDRZEJEWSKA, B., APOLLONIO, M., BARTOŃ, K. A., JĘDRZEJEWSKI, W., LINNELL, J. D., KOJOLA, I., KUSAK, J., ADAMIC, M. & CIUTI, S. 2009. Predation has a greater impact in less productive environments: variation in roe deer, (Capreolus capreolus) population density across Europe. Global Ecology and Biogeography, 18, 6, 724-734.
MESSIER, F. 1985. Social organization, spatial distribution, and population density of wolves in relation to moose density. Canadian Journal of Zoology, 63, 5, 1068-1077.
MESSIER, F. 1994. Ungulate population models with predation: a case study with the North American moose. Ecology, 75, 2, 478-488.
MRNF. 2005. Liste des espèces désignées menacées ou vulnérables au Québec [En ligne]. Disponible: http://www3.mrnf.gouv.qc.ca/faune/especes/menacees/liste.asp [Consulté le 18/11/2012].
NEUFELD, L. M. 2006. Spatial dynamics of wolves and woodland caribou in an industrial forest landscape in west-central Alberta. University of Alberta.
OBE, R. & HSU, L. S. 2011. PostGIS in action. Manning Publications. Stanford, CT, USA. PEEK, J. 1974. A review of moose food habits studies in North America. Naturaliste
Canadien, 101, 1-2, 195-215. PETERS, W., HEBBLEWHITE, M., DECESARE, N., CAGNACCI, F. & MUSIANI, M.
2013. Resource separation analysis with moose indicates threats to caribou in human altered landscapes. Ecography, 36, 4, 487-498.
PIMM, S. L. & RAVEN, P. 2000. Biodiversity: extinction by numbers. Nature, 403, 6772, 843-845.
POTVIN, F., BRETON, L. & COURTOIS, R. 2005. Response of beaver, moose, and snowshoe hare to clear-cutting in a Quebec boreal forest: a reassessment 10 years after cut. Canadian Journal of Forest Research, 35, 1, 151-160.
POTVIN, F. & COURTOIS, R. 2004. Winter presence of moose in clear-cut black spruce landscapes: related to spatial pattern or to vegetation. Alces, 40, 61-70.
R DEVELOPMENT CORE TEAM 2010. R: a language and environment for Statistical Computing. Vienna.
REICH, P. B., BAKKEN, P., CARLSON, D., FRELICH, L. E., FRIEDMAN, S. K. & GRIGAL, D. F. 2001. Influence of logging, fire, and forest type on biodiversity and productivity in southern boreal forests. Ecology, 82, 10, 2731-2748.
RESSOURCES NATURELLES CANADA 2000. Service canadien des forêts - Centre de foresterie des Laurentides (RNCAN), 2006.Cartographie intermédiaire en 52 classes

46
de la couverture terrestre du Québec (CCTQ) réalisée dans le cadre du projet Observation de la Terre pour le développement durable des forêts (OTTD) à partir des images Landsat ca. 2000. La couverture terrestre au nord de la limite terrestre des forêts fut complétée dans le cadre d'un partenariat avec le MDDEP.
RETTIE, W. J. & MESSIER, F. 2000. Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography, 23, 4, 466-478.
ROBINSON, H. S., WIELGUS, R. B. & GWILLIAM, J. C. 2002. Cougar predation and population growth of sympatric mule deer and white-tailed deer. Canadian Journal of Zoology, 80, 3, 556-568.
ROBINSON, W. D., ROBINSON, T. R., ROBINSON, S. K. & BRAWN, J. D. 2000. Nesting success of understory forest birds in central Panama. Journal of Avian Biology, 31, 2, 151-164.
ROEMER, G. W., COONAN, T. J., GARCELON, D. K., BASCOMPTE, J. & LAUGHRIN, L. 2001. Feral pigs facilitate hyperpredation by golden eagles and indirectly cause the decline of the island fox. Animal Conservation, 4, 4, 307-318.
ROMINGER, E. M., ROBBINS, C. T. & EVANS, M. A. 1996. Winter foraging ecology of woodland caribou in northeastern Washington. Journal of Wildlife Management, 60, 4, 719-728.
ROSENZWEIG, M. L. 1968. Net primary productivity of terrestrial communities: prediction from climatological data. American Naturalist, 102, 923, 67-74.
SAETHER, B.-E. 1985. Annual variation in carcass weight of Norwegian moose in relation to climate along a latitudinal gradient. Journal of wildlife management, 49, 4, 977-983.
SALA, O. E., CHAPIN, F. S., ARMESTO, J. J., BERLOW, E., BLOOMFIELD, J., DIRZO, R., HUBER-SANWALD, E., HUENNEKE, L. F., JACKSON, R. B. & KINZIG, A. 2000. Global biodiversity scenarios for the year 2100. Science, 287, 5459, 1770-1774.
SCHAEFER, J. A. 2003. Long‐term range recession and the persistence of caribou in the Taiga. Conservation Biology, 17, 5, 1435-1439.
SCHEMSKE, D. W., MITTELBACH, G. G., CORNELL, H. V., SOBEL, J. M. & ROY, K. 2009. Is there a latitudinal gradient in the importance of biotic interactions? Annual Review of Ecology Evolution, and Systematics, 40, 245-269.
SCHWARZ, G. 1978. Estimating the dimension of a model. The Annals of Statistics, 6, 2, 461-464.
SEIDENSTICKER, J. 1986. Large carnivores and the consequences of habitat insularization: ecology and conservation of tigers in Indonesia and Bangladesh. Symposium of the Zoological Society of London, 65, 105-125.
SEIP, D. R. 1992. Factors limiting woodland caribou populations and their interrelationships with wolves and moose in southeastern British Columbia. Canadian Journal of Zoology, 70, 8, 1494-1503.
SERROUYA, R., MCLELLAN, B. N., BOUTIN, S., SEIP, D. R. & NIELSEN, S. E. 2011. Developing a population target for an overabundant ungulate for ecosystem restoration. Journal of Applied Ecology, 48, 4, 935-942.
SINGH, N. J., BÖRGER, L., DETTKI, H., BUNNEFELD, N. & ERICSSON, G. 2012. From migration to nomadism: movement variability in a northern ungulate across its latitudinal range. Ecological Applications, 22, 7, 2007-2020.

47
STOHLGREN, T. J., BARNETT, D., FLATHER, C., KARTESZ, J. & PETERJOHN, B. 2005. Plant species invasions along the latitudinal gradient in the United States. Ecology, 86, 9, 2298-2309.
TURCHIN, P. & HANSKI, I. 1997. An empirically based model for latitudinal gradient in vole population dynamics. American Naturalist, 149, 5, 842-874.
VORS, L. S. & BOYCE, M. S. 2009. Global declines of caribou and reindeer. Global Change Biology, 15, 11, 2626-2633.
VORS, L. S., SCHAEFER, J. A., POND, B. A., RODGERS, A. R. & PATTERSON, B. R. 2007. Woodland caribou extirpation and anthropogenic landscape disturbance in Ontario. Journal of wildlife management, 71, 4, 1249-1256.
WITTMER, H. U., MCLELLAN, B. N., SERROUYA, R. & APPS, C. D. 2007. Changes in landscape composition influence the decline of a threatened woodland caribou population. Journal of Animal Ecology, 76, 3, 568-79.
WITTMER, H. U., SERROUYA, R., ELBROCH, L. M. & MARSHALL, A. J. 2013. Conservation strategies for species affected by apparent competition. Conservation Biology, 27, 2, 254-260.
Related Documents











