Late glacial, Holocene and modern dino£agellate cyst assemblages in the Aegean^Marmara^Black Sea corridor : statistical analysis and re-interpretation of the early Holocene Noah’s Flood hypothesis Peta J. Mudie a , Andre ¤ Rochon a; , Ali E. Aksu b , Helen Gillespie b a Geological Survey of Canada^Atlantic, Bedford Institute of Oceanography, 1 Challenger Drive, Dartmouth, NS, Canada B2Y 4A2 b Department of Earth Sciences, Memorial University of Newfoundland, St. John’s, NF, Canada A1B 3X5 Abstract Understanding of the history of water exchange between the Mediterranean and Black seas has been hampered by the lack of continuous microfossil records for Holocene cores from the Marmara Sea Gateway (Aegean^ Marmara^Black Sea corridor), and by the lack of core-top data linking modern microfossil assemblages with sea surface conditions. Based on molluscs, an abrupt transition from freshwater to marine conditions at ca. 7.5 ka has been postulated, with interpretation of this event as the basis for the story of Noah’s Flood. We have re-examined this hypothesis using organic-walled dinoflagellate cysts that show excellent preservation and moderate to high diversity in Pleistocene^Holocene sediments of the Marmara Sea Gateway. Principle component analysis of 16 core-top samples shows that an assemblage dominated by cysts of autotrophic gonyaulacoids distinguishes the hypersaline Aegean^ Mediterranean water (salinity 31^38), whereas heterotrophic protoperidinioids characterise the low-salinity Marmara^ Black Sea water (salinity V14^25). About 150 samples from eight cores, with multiple radiocarbon ages spanning the past V33 000 years, show correlatable major changes in cyst assemblages along the Marmara^Black Sea corridor. Variation in length and shape of gonyaulacoid species processes decreases upcore from basal units where there are no- analogue assemblages of cysts with highly variable spine development. It is shown that these variable and no-analogue assemblages correspond to brackish conditions (salinity V4^12) when calibrated against N 18 O data and salinity estimates derived from planktonic foraminifera. Salinity reconstructions indicate that the Aegean and Marmara seas were connected by V11 ka when the Marmara Sea was a brackish or low-salinity sea (V12^17) and that the Black Sea was flowing into the Marmara Sea by at least 10.2^9.5 ka. There is no evidence to support either the idea that the Black Sea was a large freshwater lake suitable for farming in the early Holocene or that a sudden ( 6 500-yr) flooding of the Black Sea by a 100-m-high waterfall of Mediterranean water occurred at 7.5 ka. It is shown that there is a need for precise use of salinity terms when reconstructing the history of saline lakes in the context of Neolithic human occupation so that the likelihood of agricultural settlement can be evaluated realistically. ȣ 2003 Elsevier B.V. All rights reserved. Keywords: palynology; dino£agellate cysts; eastern Mediterranean; Aegean Sea; Black Sea; Marmara Sea; Noah’s Flood; palaeosalinity 0034-6667 / 03 / $ ^ see front matter ȣ 2003 Elsevier B.V. All rights reserved. doi :10.1016/S0034-6667(03)00117-9 * Corresponding author. Present address: ISMER-UQAR, 310 Alle ¤e des Ursulines, Rimouski, PQ, Canada G5L 3A1. Fax: +1-418-724-1842. E-mail address: [email protected] (A. Rochon). PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart Review of Palaeobotany and Palynology 128 (2004) 143^167 R Available online at www.sciencedirect.com www.elsevier.com/locate/revpalbo

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
Late glacial, Holocene and modern dino£agellate cystassemblages in the Aegean^Marmara^Black Sea corridor:
statistical analysis and re-interpretation of the earlyHolocene Noah’s Flood hypothesis
Peta J. Mudie a, Andre¤ Rochon a;�, Ali E. Aksu b, Helen Gillespie b
a Geological Survey of Canada^Atlantic, Bedford Institute of Oceanography, 1 Challenger Drive, Dartmouth, NS, Canada B2Y 4A2b Department of Earth Sciences, Memorial University of Newfoundland, St. John’s, NF, Canada A1B 3X5
Abstract
Understanding of the history of water exchange between the Mediterranean and Black seas has been hamperedby the lack of continuous microfossil records for Holocene cores from the Marmara Sea Gateway (Aegean^Marmara^Black Sea corridor), and by the lack of core-top data linking modern microfossil assemblages with seasurface conditions. Based on molluscs, an abrupt transition from freshwater to marine conditions at ca. 7.5 ka hasbeen postulated, with interpretation of this event as the basis for the story of Noah’s Flood. We have re-examined thishypothesis using organic-walled dinoflagellate cysts that show excellent preservation and moderate to high diversity inPleistocene^Holocene sediments of the Marmara Sea Gateway. Principle component analysis of 16 core-top samplesshows that an assemblage dominated by cysts of autotrophic gonyaulacoids distinguishes the hypersaline Aegean^Mediterranean water (salinity 31^38), whereas heterotrophic protoperidinioids characterise the low-salinity Marmara^Black Sea water (salinity V14^25). About 150 samples from eight cores, with multiple radiocarbon ages spanning thepast V33 000 years, show correlatable major changes in cyst assemblages along the Marmara^Black Sea corridor.Variation in length and shape of gonyaulacoid species processes decreases upcore from basal units where there are no-analogue assemblages of cysts with highly variable spine development. It is shown that these variable and no-analogueassemblages correspond to brackish conditions (salinity V4^12) when calibrated against N
18O data and salinityestimates derived from planktonic foraminifera. Salinity reconstructions indicate that the Aegean and Marmara seaswere connected by V11 ka when the Marmara Sea was a brackish or low-salinity sea (V12^17) and that the BlackSea was flowing into the Marmara Sea by at least 10.2^9.5 ka. There is no evidence to support either the idea that theBlack Sea was a large freshwater lake suitable for farming in the early Holocene or that a sudden (6 500-yr) floodingof the Black Sea by a 100-m-high waterfall of Mediterranean water occurred at 7.5 ka. It is shown that there is a needfor precise use of salinity terms when reconstructing the history of saline lakes in the context of Neolithic humanoccupation so that the likelihood of agricultural settlement can be evaluated realistically.@ 2003 Elsevier B.V. All rights reserved.
Keywords: palynology; dino£agellate cysts; eastern Mediterranean; Aegean Sea; Black Sea; Marmara Sea; Noah’s Flood;palaeosalinity
0034-6667 / 03 / $ ^ see front matter @ 2003 Elsevier B.V. All rights reserved.doi:10.1016/S0034-6667(03)00117-9
* Corresponding author. Present address: ISMER-UQAR, 310 Alle¤e des Ursulines, Rimouski, PQ, Canada G5L 3A1.Fax: +1-418-724-1842. E-mail address: [email protected] (A. Rochon).
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
Review of Palaeobotany and Palynology 128 (2004) 143^167
R
Available online at www.sciencedirect.com
www.elsevier.com/locate/revpalbo

1. Introduction
The Aegean Sea, the Marmara Sea and theBlack Sea are deep (s 1^2 km), semi-enclosed,marginal marine basins located in the easternMediterranean region between ca. 34.5 and46‡N latitude (Figs. 1 and 2). The low-salinity(V17^22) inland Black Sea is about twice thesize of the hypersaline (salinity s 36) AegeanSea that forms the northern branch of the easternMediterranean Sea. The tiny, almost land-lockedMarmara Sea links these two large seas. Narrowstraits with shallow sills (45^75 m depth) presentlyconnect the Marmara Sea with the Aegean andBlack seas. This ‘Marmara Sea Gateway’ (thestraits of Dardanelles and Bosphorus, next tothe deep intervening Marmara Sea, see Fig. 3)provides a set of natural £ow valves that regulatesthe history of water exchange and salinity of theBlack Sea (Aksu et al., 2002a). The timing andcauses of reconnection of the Black and Mediter-ranean seas during Holocene sea-level rise hasparticularly important implications for Europeanand Middle East archaeology in addition to pa-laeoclimatic reconstruction and sedimentology(e.g. genesis of sapropelic mud). For example, ithas been postulated that the Black Sea was aland-locked freshwater lake that attracted earlyNeolithic agricultural settlement before ca. 7500yr BP (years Before Present, i.e. AD 1950), atwhich time it would suddenly (in less than 2 years)have been reconnected to the Mediterranean Seaby a deluge of seawater pouring over a 100-m-high sill at the east end of the Bosphorus Strait(Ryan and Pitman, 1999; Wilson, 2001). It is fur-ther speculated that this catastrophic event wasthe historical basis for the biblical story of Noah’sFlood and that it triggered the spread of agricul-ture north, south and east from a Black Sea epi-centre, contrary to most archaeological evidence(e.g. Bogucki, 1996).
In fact, the primary evidence for Ryan and Pit-man’s postulation comes from mollusc shells inthe Holocene sapropel of 5 cores from the outernorthern Black Sea shelf, all of which have agesof V7150 P 100 yr BP (Ryan et al., 1997). Thesemolluscs include the so-called freshwater Caspianspecies Dreissena polymorpha, D. rostriformis and
the marine species Mytilus lineatus and M. gallo-provincialis. Changes in these mollusc assemblageswere thus interpreted as indicating a successionfrom freshwater to the present low-salinity marineenvironment. In fact, modern populations of so-called freshwater Dreissena species (Mills et al.,1996) live in estuarine areas of the Black andAzov seas with salinities ranging from about1 (freshwater) to 7 (brackish water) and they sur-vive in the so-called freshwater Caspian Sea (e.g.Ballard et al., 2000) which now has a salinity ofabout 13 and is clearly a brackish water inlandlake. Other shell ages were obtained from bulkdredging o¡ Sinop in the southern Black Sea (Bal-lard et al., 2000). Here the so-called freshwaterDreissena species and lacustrine Turricaspia caspiaspecies had ages of 15.5^10.5 ka, while the marineMytilus, Papillicardium, Parvicardium, Modiolu-lus, Bittium and Abra species had ages of ca.2.1^6.8 ka, but there were no shells or ages forthe interval from 9.3 to 7.48 ka immediately be-fore the purported marine £ooding. Ryan and Pit-man’s interpretation also contradicts seismostrati-graphic evidence for a complex and progressivereconnection of the Aegean Sea, the MarmaraSea and the Black Sea during the past 12 ka(Aksu et al., 1999a, 2002a; Hiscott and Aksu,2002). In addition, Dreissena and other Caspianmolluscs with ages of V8.1 ka were found at 18 mdepth in cores from near the east of the Bospho-rus Strait, showing that the water level in theBlack Sea was not far below present sea leveljust before 7.1 ka (Go«ru«r et al., 2001), contraryto the 3100 m depth postulated by Ryan andPitman.
Other studies have attempted to use siliceous orcalcareous microfossils in Black Sea cores to in-terpret the abrupt change from silty banded mudto organic-rich marine sapropel at V7 ka (e.g.Ross and Degens, 1974 and references therein).In the Black Sea, however, the diversity of earlyHolocene calcareous microfossils is restricted toseven coccolith species (Bukry, 1974) that occuronly in sediments younger than V7 ka, and tofour benthic foraminifer species (Georghian,1974). More than 60 benthic foraminifer speciesoccur in shelf sediments of the Marmara Sea butnone were found in the early to mid-Holocene
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167144

sediments of the deep basins (Kaminski et al.,2002). Preservation of marine diatoms is alsopoor in the Black and Marmara seas (Maynard,1974; Pilskaln and Pike, 2001; Aksu, unpub-lished) and there is a strong bias towards repre-sentation of the thicker-walled freshwater diatomspecies e.g. Stephanodiscus and Astera that mayobscure the evidence for a brackish or mixed(fresh and salt water) palaeoenvironment. Palae-oceanographic studies have also been hindered bythe lack of core-top data linking modern plank-tonic microfossil assemblages with sea surfaceconditions in the Marmara Gateway region (Ka-minski et al., 2002).
Previous palynological studies of the Aegean^Marmara^Black Sea corridor (Wall and Dale,1974; Traverse, 1988; Mudie et al., 2001,2002a,b) showed that organic-walled dino£agel-late cysts (dinocysts) are abundant and well pre-served in both surface and late Pleistocene^Holo-cene sediments throughout the region, anddescribed informal assemblage zones. Dinocystspecies diversity (N, the number of species) ranges
from 9^12 in modern samples from all these seasbut some species appear to be restricted to eitherthe Mediterranean Sea or Black Sea, whereas theMarmara Sea contains more mixed assemblages.These past studies also show that in organic-richmarginal seas such as the Marmara Sea and theBlack Sea, preservation of thin-walled protoperi-dinioid species is uniformly good (Aksu et al.,1995, 1999b; Mudie et al., 2002a).
The purpose of this paper is to provide a de-tailed account of the present-day oceanography ofthe eastern Mediterranean and Marmara Gate-way regions and a statistical analysis of the dino-cyst assemblage distributions in relation to surfacesalinity. We also describe the morphological var-iation of key indicator species in the context ofpublished laboratory experiments on cyst^thecarelationships of the Spiniferites ramosus^Spinife-rites membranaceus complex (Lewis et al., 2001).These new data are then applied to interpretationof major changes in cyst assemblages during thepast V33 ka in sediment cores with multiple ra-diocarbon ages (see Tables 1A and 1B) from the
Fig. 1. Map of the Mediterranean region showing the bathymetry in metres and the location of the study area (large squarebox).
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 145
Marmara Gateway in order to document the his-tory of surface water exchange between the Med-iterranean and Black seas. We also show that thesalinity terminology employed by geologists formarginal seas (e.g. freshwater or brackish lake
for ponded coastal waters with a salinity of 27)is incompatible with that used by biologists, farm-ers and water quality agencies who restrict theterm of freshwater to waters with a salinity ofless than 2 and refer to more saline waters as
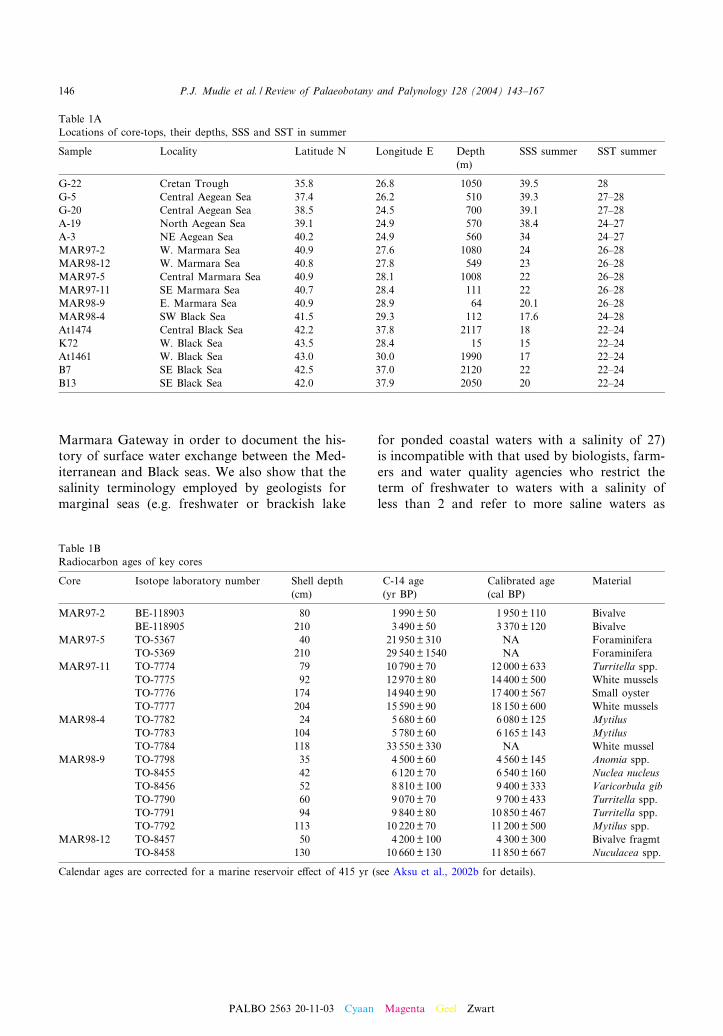
Table 1ALocations of core-tops, their depths, SSS and SST in summer
Sample Locality Latitude N Longitude E Depth SSS summer SST summer(m)
G-22 Cretan Trough 35.8 26.8 1050 39.5 28G-5 Central Aegean Sea 37.4 26.2 510 39.3 27^28G-20 Central Aegean Sea 38.5 24.5 700 39.1 27^28A-19 North Aegean Sea 39.1 24.9 570 38.4 24^27A-3 NE Aegean Sea 40.2 24.9 560 34 24^27MAR97-2 W. Marmara Sea 40.9 27.6 1080 24 26^28MAR98-12 W. Marmara Sea 40.8 27.8 549 23 26^28MAR97-5 Central Marmara Sea 40.9 28.1 1008 22 26^28MAR97-11 SE Marmara Sea 40.7 28.4 111 22 26^28MAR98-9 E. Marmara Sea 40.9 28.9 64 20.1 26^28MAR98-4 SW Black Sea 41.5 29.3 112 17.6 24^28At1474 Central Black Sea 42.2 37.8 2117 18 22^24K72 W. Black Sea 43.5 28.4 15 15 22^24At1461 W. Black Sea 43.0 30.0 1990 17 22^24B7 SE Black Sea 42.5 37.0 2120 22 22^24B13 SE Black Sea 42.0 37.9 2050 20 22^24
Table 1BRadiocarbon ages of key cores
Core Isotope laboratory number Shell depth C-14 age Calibrated age Material(cm) (yr BP) (cal BP)
MAR97-2 BE-118903 80 1 990 P 50 1 950 P 110 BivalveBE-118905 210 3 490 P 50 3 370 P 120 Bivalve
MAR97-5 TO-5367 40 21 950 P 310 NA ForaminiferaTO-5369 210 29 540 P 1540 NA Foraminifera
MAR97-11 TO-7774 79 10 790 P 70 12 000 P 633 Turritella spp.TO-7775 92 12 970 P 80 14 400 P 500 White musselsTO-7776 174 14 940 P 90 17 400 P 567 Small oysterTO-7777 204 15 590 P 90 18 150 P 600 White mussels
MAR98-4 TO-7782 24 5 680 P 60 6 080 P 125 MytilusTO-7783 104 5 780 P 60 6 165 P 143 MytilusTO-7784 118 33 550 P 330 NA White mussel
MAR98-9 TO-7798 35 4 500 P 60 4 560 P 145 Anomia spp.TO-8455 42 6 120 P 70 6 540 P 160 Nuclea nucleusTO-8456 52 8 810 P 100 9 400 P 333 Varicorbula gibTO-7790 60 9 070 P 70 9 700 P 433 Turritella spp.TO-7791 94 9 840 P 80 10 850 P 467 Turritella spp.TO-7792 113 10 220 P 70 11 200 P 500 Mytilus spp.
MAR98-12 TO-8457 50 4 200 P 100 4 300 P 300 Bivalve fragmtTO-8458 130 10 660 P 130 11 850 P 667 Nuculacea spp.
Calendar ages are corrected for a marine reservoir e¡ect of 415 yr (see Aksu et al., 2002b for details).
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167146

brackish (2^7) or saline (s 7). When correct sa-linity terms are applied, it is evident that the earlyHolocene Black Sea contained water unsuitablefor agriculture and human consumption.
2. Present oceanography
2.1. Water circulation
Water circulation in the semi-enclosed seas ofthe Mediterranean region is primarily driven bysalinity gradients (Fairbridge, 1966; Robinsonand Golnaraghi, 1994). Winter cooling generatesplumes that cause mixing at the boundaries ofhypersaline and less saline water masses, however,and it can cause deep convection mixing in latewinter (Schott et al., 1994). These mixing process-es ultimately govern the rate of salt storage orremoval from the deep Mediterranean basins.
Most of the water in the Aegean^Marmara^Black Sea corridor originates from the Mediterra-nean Sea (Fig. 1). The Mediterranean Sea has alagoon-type circulation (see Garrels et al., 1975)that is driven by entry of North Atlantic SurfaceWater (NASW, salinity 35) at Gibraltar. The sa-linity of the NASW increases as it £ows eastwardthrough the Alboran Sea and along the NorthAfrican margin where evaporation, E, greatly ex-ceeds precipitation, P (Garrels et al., 1975; Fair-bridge, 1966). This dense hypersaline water (s 37)sinks to ¢ll the deep (1.5^5 km) basins within theMediterranean Sea with a homogeneous warm sa-line bottom water layer (V13.5‡C; 38.4^39). Abranch of the NASW water £ows northward tothe west of Italy where evaporation and coolingproduce an Intermediate Western Basin watermass beneath the Tyrrhenian Sea. Anotherbranch is the source of the intermediate watermass in the Ionian Sea east of Italy. The remain-ing NASW £ows southeast to the Levantine Ba-sin, becoming more saline (38.7^39) and movingnorthwards into the Aegean where winter coolingproduces a third body of distinctive bottomwater. The net salt balance of the system is keptin a steady state by periodic boundary-layer mix-ing, by downwelling of cold, low-salinity runo¡and corresponding upwelling of saline water,
and by the net out£ow of saline (36) bottom waterin the Strait of Gibraltar.
The Black Sea is a large (432 000 km2) basinwith a deep plain (2^2.2 km) £anked by a wide(V100 km), shallow shelf (6 200 m) in the NW,and with steep slopes elsewhere (Fig. 2). TheBlack Sea has an estuarine-type circulation inwhich river runo¡ (V350 km3/yr) and excess P(300 km3/yr) greatly exceed water loss by E(V350 km3/yr) and sustain an out£ow ofV300^600 km3/yr of hyposaline (17.9) surfacewater. The Black Sea is linked to the MaramaraSea (Fig. 3A) by the narrow (2 km) BosphorusStrait, with maximum depth of 45 m, throughwhich relatively low-salinity (22.5) mixed Aegeanand Marmara seawater £ows as V300 km3/yr ofbottom water (Stanev and Peneva, 2002; Oº zsoy etal., 1995). This mixed Aegean and Marmara Seawater is much less saline than the Aegean Seawater (32^37) but it is much denser than thevery low-salinity surface Black Sea water (14^20), The denser, more saline water of Aegean ori-gin therefore pours over the SW Black Sea shelfand is diluted by mixing with a cold-water layer atabout 100 m depth. The incoming water thensinks towards the abyssal plain, generating intru-sions and secondary circulations down to 500 m(Oº zsoy et al., 1995; Eremeev, 1995). These insta-bilities drive large-scale vertical circulation andmixing across the halocline (i.e. the boundary be-tween watermasses of di¡erent salinity). A bottomconvection layer V300 m thick is maintained bygeothermal heat £uxes in the southern basin (Topet al., 1990). This heat £ow circulation homoge-nises the temperature and salinity of the bottomwater (Oº zsoy et al., 1995), but upward mixing ofthis saline (22.5) layer is very slow, with a renewaltime of about 2000 years.
The Sea of Marmara is a small sea (V11 500km2) with a wide shelf (V50 km) to the southand very narrow shelf (5^10 km) in the north.Three deep (s 1000 m) basins are separated byshallower sills (600 m). Water exchange takesplace as a two-layered £ow (Fig. 3A). Warm, hy-persaline (38^39) Mediterranean water from thenortheast Aegean Sea enters the eastern Darda-nelles Strait and plunges below a surface layer(10^30 m thick) of out£owing low-salinity Black
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 147
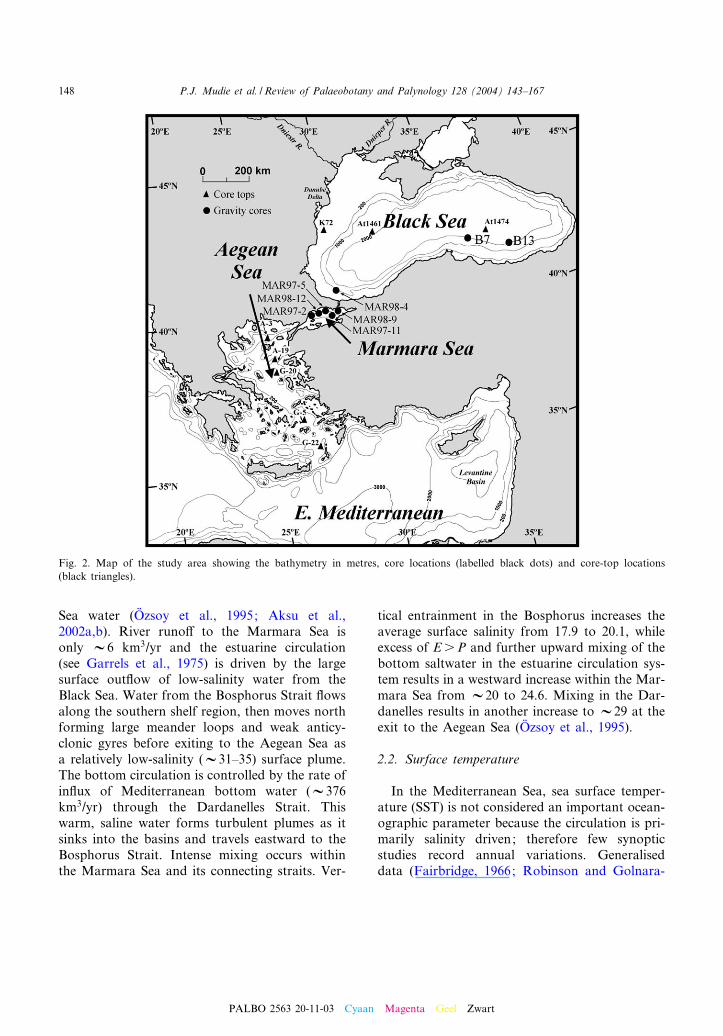
Sea water (Oº zsoy et al., 1995; Aksu et al.,2002a,b). River runo¡ to the Marmara Sea isonly V6 km3/yr and the estuarine circulation(see Garrels et al., 1975) is driven by the largesurface out£ow of low-salinity water from theBlack Sea. Water from the Bosphorus Strait £owsalong the southern shelf region, then moves northforming large meander loops and weak anticy-clonic gyres before exiting to the Aegean Sea asa relatively low-salinity (V31^35) surface plume.The bottom circulation is controlled by the rate ofin£ux of Mediterranean bottom water (V376km3/yr) through the Dardanelles Strait. Thiswarm, saline water forms turbulent plumes as itsinks into the basins and travels eastward to theBosphorus Strait. Intense mixing occurs withinthe Marmara Sea and its connecting straits. Ver-
tical entrainment in the Bosphorus increases theaverage surface salinity from 17.9 to 20.1, whileexcess of EsP and further upward mixing of thebottom saltwater in the estuarine circulation sys-tem results in a westward increase within the Mar-mara Sea from V20 to 24.6. Mixing in the Dar-danelles results in another increase to V29 at theexit to the Aegean Sea (Oº zsoy et al., 1995).
2.2. Surface temperature
In the Mediterranean Sea, sea surface temper-ature (SST) is not considered an important ocean-ographic parameter because the circulation is pri-marily salinity driven; therefore few synopticstudies record annual variations. Generaliseddata (Fairbridge, 1966; Robinson and Golnara-
Fig. 2. Map of the study area showing the bathymetry in metres, core locations (labelled black dots) and core-top locations(black triangles).
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167148

ghi, 1994; Malanotte-Rizzole et al., 1996) suggestthat average winter SST is cooler in the west (12^13‡C) than in the east (15^18‡C). Summer SSTaverages V25‡C (range 22^28‡C, with highestvalues in the Levantine region). In the AegeanSea, the winter SST is about 14.5^19‡C (e.g.Aksu et al., 1995; Lascaratos et al., 1999) andAugust SST ranges from 24 to 28‡C. In the Mar-mara Sea, average winter SST is V5‡C and thesummer temperature maximum (September) isusually V26‡C, with rare increases to 29‡C (Oº z-soy et al., 1995). In the Black Sea, February andAugust SSTs normally are V3^8‡C and 22^24‡C,respectively, but ice may occur along the coasts ofthe Black Sea, Bosphorus Strait and eastern Mar-mara Sea (Black Sea Pilot, 1990).
2.3. Surface salinity
Mudie et al. (2002a) show a map of the sea
surface salinity (SSS) in summer for the studyarea. In most of the eastern Mediterranean andAegean seas, there is little seasonal variation insalinity (38.5^39), and the halocline is below 200m (Lascaratos et al., 1999; Aksu et al., 2002a).Near the western Dardanelles Strait, however,there is a relatively low-salinity surface plume(0^V20 m depth) in which the salinity variesfrom V31 in summer to 35 in winter, dependingon the volume of river runo¡ to the Black Sea.
The catchment area of the Black Sea includesabout half of Europe, as well as the large Dniestr,Dnieper and Don rivers from Russia (see Aksu etal., 2002a; Stanev and Peneva, 2002). During thelast century, P3E varied from 0 to 600 km3/yr,with a mean of V300 km3/yr. Summer surfacesalinity during a normal year (e.g. September1991; Uº nlu«ata et al., 1993) ranged from V15o¡ the Danube delta to 17 in a plume from theDnieper delta to Bosphorus Strait, but SSS overmost areas was V17.5 to s 18, with maxima of20^22 in the cyclonic gyres of the southern BlackSea (Arthur and Dean, 1998; Black Sea Pilot,1990). During periods of £ooding in spring, how-ever, the shelf salinity may be reduced to 14. Thehalocline in the southern Black Sea is gradualfrom V70^130 m depth, with the depth increas-ing eastwards from the Bosphorus shelf (Aksu etal., 2002a).
In contrast, the halocline in the Marmara Gate-way is very sharp (Fig. 3). Over the Marmara Sea,the surface low-salinity layer (22^24) is only about10 m thick (Oº zsoy et al., 1995) and the salinityincreases from 26 to 36 in the halocline betweendepths of 10 to 20 m. Below this, the salinity is37.4^38.6. In the Bosphorus Strait, SSS can rangefrom 15 in February to 21 in October. In theDardanelles Strait, the average SSS ranges from24.6 to 29.
2.4. Nutrients and productivity
Satellite images for 1990 and 1997 show thatsummer phytoplankton production in the Black,Marmara and northern Aegean seas is much high-er (1^s 10 mg/m3 Chlorophyll-a) than in otherdeepwater areas of the Mediterranean. Mudie etal. (2002a) have shown that thecate organic-
Fig. 3. Diagram of the £ow directions (straight arrows), sa-linity (numbers in parentheses) and water exchange (spiralarrows) in the Marmara Sea Gateway, after Oº zsoy et al.,1995. (A) Present circulation. (B) Probable circulation atV9.5 ka.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 149

walled dino£agellate diversity is also correspond-ingly higher in the study area than in the northernMediterranean (48 and 35 species, respectively). Itis di⁄cult to evaluate the link between nutrientsand the core-top dinocyst assemblages, however,because there have been large anthropogenicchanges during the past 50 years (Daskalov,2002). Nutrient discharge to the MediterraneanSea also increased from about 1940 to 1970(Haas and Zuckman, 1990). Representative long-term nutrient values, however, can be obtainedfrom historical records. In 1964, phosphate(PO4^P) values in spring were 0.033^0.065 WM(micromoles) according to Fairbridge (1966) inthe western and eastern basins of the Mediterra-nean Sea, while values of 0.15^0.41 WM were re-corded for the surface 100 m of the Black Sea in1940^1949 (Fonselius, 1974). From 1986 to 1991,the mean concentrations of PO4^P and NO3 en-tering the Bosphorus from the Black Sea werereported as 0.17 P 0.17 WM and 1.55 P 1.36 WM,respectively (Oº zsoy et al., 1995).
3. Methods
Six gravity cores (Fig. 2; Tables 1A and 1B)were collected in the Marmara Sea and the BlackSea from the RV Koca Piri Reis of the Institute ofMarine Sciences and Technology, Dokuz Eylu« lUniversity, using a 4-m-long gravity corer. Thesecores were stored upright and were shipped to theMemorial University of Newfoundland (MUN),where they were split, described and photo-graphed. The sedimentology and calcareous mi-crofossils in these cores are described by Aksu etal. (2002b). Two box cores were studied from thesoutheastern Black Sea. The sedimentology ofthese cores is described by Duman (1994), andinitial dinocyst studies were reported by Mudieet al. (2001). Core-top data for interpretation ofthe dinocyst ecostratigraphies and associatedsporomorph assemblages were obtained fromthese eight cores and from ¢ve Aegean Sea sites(see Aksu et al., 1995) and three other Black Seasites (triangles in Fig. 2), including published dataof Wall and Dale (1974) and Traverse (1974).
The cores were sampled at 10-cm intervals, so
that 10 samples were studied for each metre of acore. Palynomorphs were extracted from samplesof 2^5 cm3 volume, using standard methods forQuaternary marine sediments (Rochon et al.,1999): initial sieving at 20- and 125-Wm meshsizes, then digestion in HCl, and adding exoticspores to obtain estimates of palynomorph con-centration per cm3, and ¢nally, digestion in 52%HF. Palynomorph preservation is good in mostsections of the cores, but sandy intervals oftencontain oxidised and abraded grains. Some levelsalso contain reworked pre-Quaternary (mainlyPliocene) pollen or spores and, more rarely, dino-£agellate cysts. These reworked pre-Quaternarypalynomorphs (see Traverse, 1974) were primarilyrecognised by their distinctive morphology, but£attened and/or yellowish grains of extant pollentypes were also scored as reworked.
One or two slides of each processed samplewere routinely counted (by P.J.M. and A.R.) atU160^250 magni¢cation, until a minimum of 300exotic spores was reached. This yielded counts of100^200 for total dinocysts in the middle to upperHolocene samples. In some brownish sandy sedi-ments of the Marmara Sea late glacial to earlyHolocene deltaic intervals (e.g. MAR98-09; seeHiscott et al., 2002), the Quaternary palyno-morphs appeared strongly oxidised (thin-walledand very pale, even when stained) and the countswere as low as 25^50 for two slides and s 1000exotic spores. Dinocyst nomenclature follows thatused by Williams et al. (1998) and Rochon et al.(1999) except where noted. Other palynomorphscounted included pollen, spores of ferns andmosses, microforaminiferal linings, acritarchs,fungal remains and freshwater algae, includingcoenobia of Pediastrum and Botryococcus. Thespatial and temporal distributions of these terres-trial and freshwater taxa are reported in Mudie etal. (2002b).
For the principal component analysis of thecore-top data, the software from Guiot (1990)was used on log-transformed dinocyst percentagedata. The analysis provides a cross-correlationmatrix between taxa, which allows identi¢cationof those that are in phase or in opposition. Atotal of 26 taxa were used in the analysis. Thelog-transformation of percentage data was used
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167150

to increase the weight of less represented taxa,while decreasing that of the ubiquitous ones.
4. Results
4.1. Core top assemblages
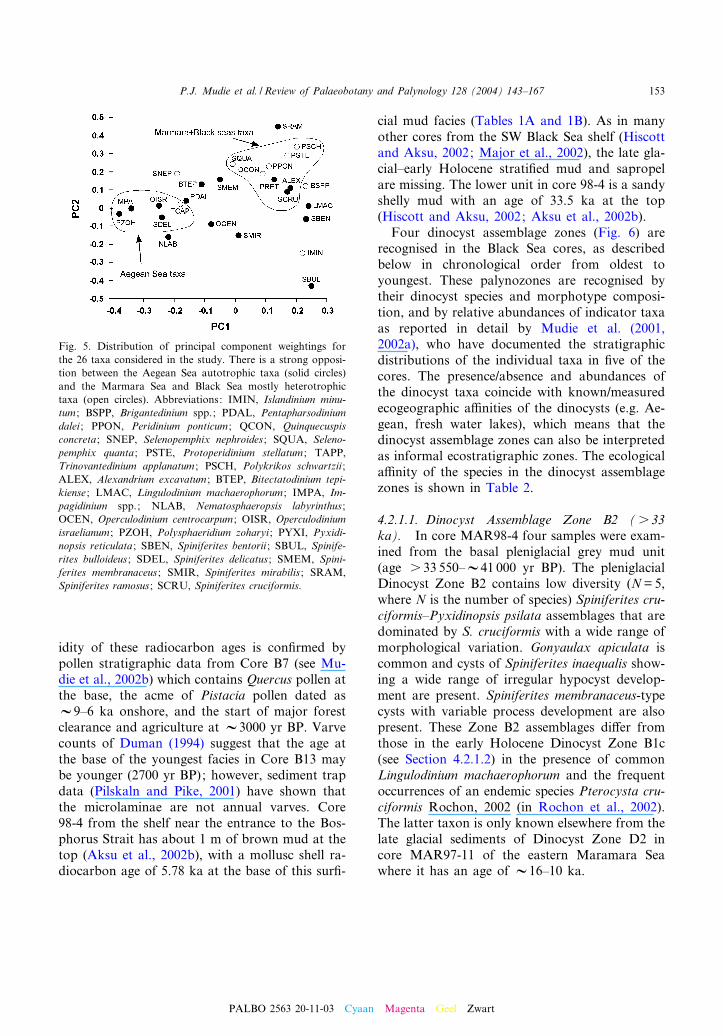
The core-top samples represent the depositionof dinocysts over time intervals ranging from aminimum of V26 years (MAR97-2) to 150 years(B7), with an average of about 130 years (seeAksu et al., 1995; Mudie et al., 2002b, for sedi-mentation rates at most sites). The relative abun-dances of the dinocyst taxa (Fig. 4) suggest thepresence of two geographic groups of species. AnAegean group is characterised by the gonyaula-coid taxa Operculodinium israelianum, Poly-sphaeridium zoharyi, Impagidinium spp. (I. aculea-tum, I. patulum, I. sphaericum, I. strialatum),Spiniferites delicatus and the protoperidinioid Tri-novantedinium applanatum. A Marmara/Black Seagroup of taxa is characterised by the protoperidi-nioids Protoperidinium (Stelladinium) stellatum,Selenopemphix quanta, Pyxidinopsis reticulata,Quinquecuspis concreta, Islandinium minutum andPeridinium ponticum, the colonial polykrikoidPolykrikos schwartzii, and three gonyaulacoids(Alexandrium excavatum-type cyst, Spiniferitesbulloideus and Spiniferites cruciformis). The prin-cipal component analysis (Fig. 5) con¢rms thepresence of two distinct dinocyst assemblages.Principal components 1 (PC1) and 2 (PC2) repre-sent 55.3% and 11.6% of the total variance, re-spectively. The taxa weightings for PC1 andPC2 (Fig. 5) show the strong disparity betweenthe Aegean Sea (negative weighting) and the Mar-mara/Black sea taxa (positive weighting) alongPC1, but there is no clear trend with respect toPC2. The main taxa in the high-salinity AegeanSea PC1 group are the autotrophic gonyaulacoidsO. israelianum, S. delicatus, Impagidinium spp.and P. zoharyi, and the protoperidinioid T. appla-natum. The principal component analysis also re-veals that the autrophic gonyaulacoid Nemato-sphaeropsis labyrinthus is closely correlated withthe Aegean group. The low-salinity Marmara^Black Sea’s PC1 group is dominated by the
known or presumed heterotrophic species P. stel-latum, Q. concreta, Peridinium ponticum, S. quan-ta, and P. schwartzii, and by the autotrophic taxaSpiniferites bentorii, Pyxidinopsis reticulata, andAlexandrium excavatum-type cyst. The principlecomponent analysis further shows that heterotro-phic protoperidinioid Brigantedinium spp. areclosely correlated with this group. Most of theother taxa are ubiquitous neritic gonyaulacoidspecies that are present in many of the core-topsamples and are occasionally common in both thehigh and low-salinity environments, includingOperculodinium centrocarpum, Spiniferites mirabi-lis, Lingulodinium machaerophorum, Spiniferitesmembranaceus, and the cyst of Pentapharsodiniumdalei.
4.2. Stratigraphic distribution
4.2.1. Black Sea coresThree cores from the Black Sea were studied:
cores B7, B13 and MAR98-4 (Fig. 6). Previousstudies (e.g. Ross et al., 1970; Wall and Dale,1974; Duman, 1994; Mudie et al., 2001; Mudieet al., 2002a) show that the Cores B7 and B13from the southeastern basin contain three litho-facies, which correspond to facies 1^3 at Wall andDale’s sites 1474K, 1445P and 1451G. The oldestfacies is an olive-grey to grey mud unit with dis-continuous silt laminae. Palynostratigraphic cor-relations with reference stratigraphies onshore(Mudie et al., 2002b) now show that the bottomof core B7 is 6 9800 yr BP based on the presenceof Quercus pollen. The lower half of this facies incore B13 contains major turbidites and reworkedpre-Quaternary palynomorphs, as also found inthe cores studied by Wall and Dale (1974) whogive an age of V13 ka for the base of this litho-facies based on extrapolation and cross-correla-tion with other radiocarbon dated cores fromthe Black Sea. The mid-Holocene and the young-est sediments in cores B7 and B13 are a sapropeland microlaminated carbonate mud, respectively.The lower boundaries of these facies in Core B7are assigned ages of 7140 P 180 and 3300 P 130 yrBP based on chronostratigraphic correlation withcores 1474 and 1462 that are dated by bulk or-ganic carbon (Ross et al., 1970). The general val-
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 151
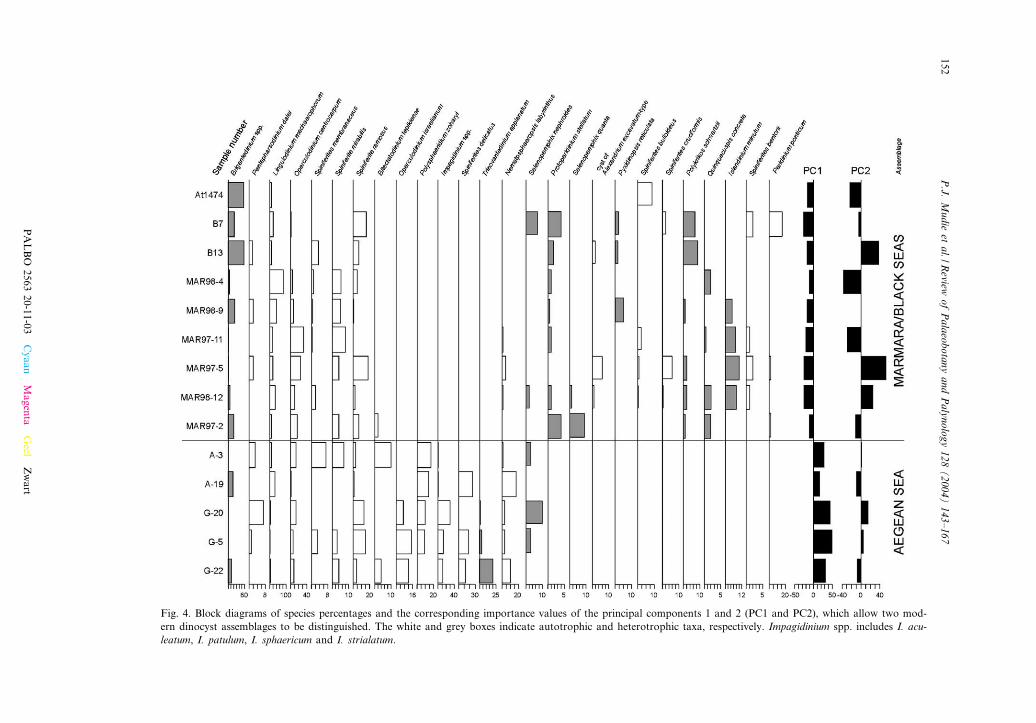
Fig. 4. Block diagrams of species percentages and the corresponding importance values of the principal components 1 and 2 (PC1 and PC2), which allow two mod-ern dinocyst assemblages to be distinguished. The white and grey boxes indicate autotrophic and heterotrophic taxa, respectively. Impagidinium spp. includes I. acu-leatum, I. patulum, I. sphaericum and I. strialatum.
PA
LB
O2563
20-11-03C
yaanM
agentaG
eelZ
wart
P.J.
Mudie
etal./R
eviewof
Palaeobotany
andPalynology
128(2004)
143^167152
idity of these radiocarbon ages is con¢rmed bypollen stratigraphic data from Core B7 (see Mu-die et al., 2002b) which contains Quercus pollen atthe base, the acme of Pistacia pollen dated asV9^6 ka onshore, and the start of major forestclearance and agriculture at V3000 yr BP. Varvecounts of Duman (1994) suggest that the age atthe base of the youngest facies in Core B13 maybe younger (2700 yr BP); however, sediment trapdata (Pilskaln and Pike, 2001) have shown thatthe microlaminae are not annual varves. Core98-4 from the shelf near the entrance to the Bos-phorus Strait has about 1 m of brown mud at thetop (Aksu et al., 2002b), with a mollusc shell ra-diocarbon age of 5.78 ka at the base of this sur¢-
cial mud facies (Tables 1A and 1B). As in manyother cores from the SW Black Sea shelf (Hiscottand Aksu, 2002; Major et al., 2002), the late gla-cial^early Holocene strati¢ed mud and sapropelare missing. The lower unit in core 98-4 is a sandyshelly mud with an age of 33.5 ka at the top(Hiscott and Aksu, 2002; Aksu et al., 2002b).
Four dinocyst assemblage zones (Fig. 6) arerecognised in the Black Sea cores, as describedbelow in chronological order from oldest toyoungest. These palynozones are recognised bytheir dinocyst species and morphotype composi-tion, and by relative abundances of indicator taxaas reported in detail by Mudie et al. (2001,2002a), who have documented the stratigraphicdistributions of the individual taxa in ¢ve of thecores. The presence/absence and abundances ofthe dinocyst taxa coincide with known/measuredecogeographic a⁄nities of the dinocysts (e.g. Ae-gean, fresh water lakes), which means that thedinocyst assemblage zones can also be interpretedas informal ecostratigraphic zones. The ecologicala⁄nity of the species in the dinocyst assemblagezones is shown in Table 2.
4.2.1.1. Dinocyst Assemblage Zone B2 (s 33ka). In core MAR98-4 four samples were exam-ined from the basal pleniglacial grey mud unit(age s 33 550^V41 000 yr BP). The pleniglacialDinocyst Zone B2 contains low diversity (N= 5,where N is the number of species) Spiniferites cru-ciformis^Pyxidinopsis psilata assemblages that aredominated by S. cruciformis with a wide range ofmorphological variation. Gonyaulax apiculata iscommon and cysts of Spiniferites inaequalis show-ing a wide range of irregular hypocyst develop-ment are present. Spiniferites membranaceus-typecysts with variable process development are alsopresent. These Zone B2 assemblages di¡er fromthose in the early Holocene Dinocyst Zone B1c(see Section 4.2.1.2) in the presence of commonLingulodinium machaerophorum and the frequentoccurrences of an endemic species Pterocysta cru-ciformis Rochon, 2002 (in Rochon et al., 2002).The latter taxon is only known elsewhere from thelate glacial sediments of Dinocyst Zone D2 incore MAR97-11 of the eastern Maramara Seawhere it has an age of V16^10 ka.
Fig. 5. Distribution of principal component weightings forthe 26 taxa considered in the study. There is a strong opposi-tion between the Aegean Sea autotrophic taxa (solid circles)and the Marmara Sea and Black Sea mostly heterotrophictaxa (open circles). Abbreviations: IMIN, Islandinium minu-tum ; BSPP, Brigantedinium spp.; PDAL, Pentapharsodiniumdalei ; PPON, Peridinium ponticum ; QCON, Quinquecuspisconcreta ; SNEP, Selenopemphix nephroides ; SQUA, Seleno-pemphix quanta ; PSTE, Protoperidinium stellatum ; TAPP,Trinovantedinium applanatum ; PSCH, Polykrikos schwartzii ;ALEX, Alexandrium excavatum ; BTEP, Bitectatodinium tepi-kiense ; LMAC, Lingulodinium machaerophorum ; IMPA, Im-pagidinium spp.; NLAB, Nematosphaeropsis labyrinthus ;OCEN, Operculodinium centrocarpum ; OISR, Operculodiniumisraelianum ; PZOH, Polysphaeridium zoharyi ; PYXI, Pyxidi-nopsis reticulata ; SBEN, Spiniferites bentorii ; SBUL, Spinife-rites bulloideus ; SDEL, Spiniferites delicatus ; SMEM, Spini-ferites membranaceus ; SMIR, Spiniferites mirabilis ; SRAM,Spiniferites ramosus ; SCRU, Spiniferites cruciformis.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 153

4.2.1.2. Dinocyst Assemblage Zone B1c (6 9.8^7.1 ka). The ¢ve (core B13) to seven (core B7)samples from the early Holocene Black Sea dino-cyst Assemblage Zone B1c are marked by lowdiversity dinocyst assemblages (N= 2^5) that aredominated (V80%) by Spiniferites cruciformis(V80%) and spherical cysts of Pyxidinopsis psila-ta (see Mudie et al., 2001), with rare Peridiniumponticum, Brigantedinium spp. and Spiniferitesmirabilis except at the top where other gonyaula-coids (Spiniferites bentorii, S. inaequalis, Spinife-rites spp. and Operculodinium centrocarpum) in-
crease to V40%. The freshwater dinocystspecies Gonyaulax apiculata (Plate I, 8,9) is occa-sionally present in B1c, along with abundant thin-walled yellowish spherical palynomorphs calledbaggy saccocysts (Traverse, 1988; Mudie et al.,2002a) that include peridinioid dinocysts of prob-able brackish or freshwater origin. The cysts ofS. cruciformis (Plate I, 1^7) show variable devel-opment of the periphragm and processes (Wall etal., 1973; Mudie et al., 2001). Detached endocystsand morphotypes with very short spines (S. cru-ciformis forms 4 and 5 of Mudie et al., 2001; Plate
Fig. 6. Chronostratigraphic distribution of dinocyst assemblage zones in the Marmara Sea Gateway cores, showing their correla-tion and the temporal shifts from three di¡erent brackish water assemblages B2 (cross-hatch), D2 and B1c (¢ne hachure) to low-salinity marine assemblages B1b (light stipple) and D1b (light hachure) and the modern assemblages B1a (salinity 17^22) andD1a (salinity 22^24) with dark and medium stipples, respectively. The cores were sampled at 10-cm intervals, so that 10 sampleswere studied for each metre of a core (for details, see Mudie et al., 2002a). Note that some of the oldest assemblage zones areonly shown in part because of space restrictions.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167154
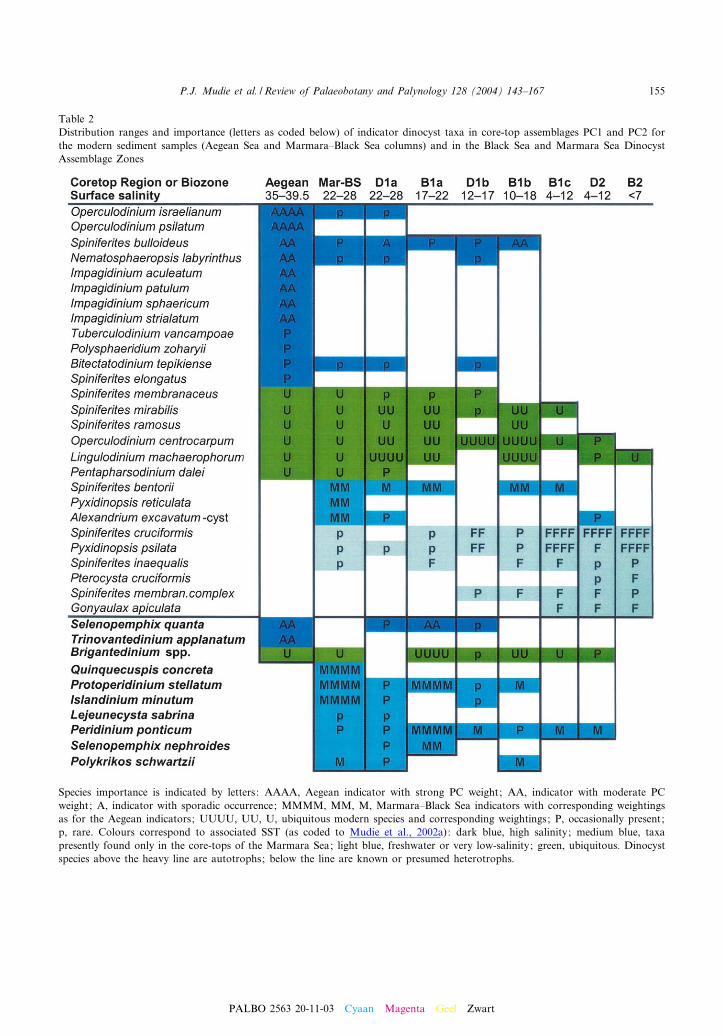
Table 2Distribution ranges and importance (letters as coded below) of indicator dinocyst taxa in core-top assemblages PC1 and PC2 forthe modern sediment samples (Aegean Sea and Marmara^Black Sea columns) and in the Black Sea and Marmara Sea DinocystAssemblage Zones
Species importance is indicated by letters: AAAA, Aegean indicator with strong PC weight; AA, indicator with moderate PCweight; A, indicator with sporadic occurrence; MMMM, MM, M, Marmara^Black Sea indicators with corresponding weightingsas for the Aegean indicators; UUUU, UU, U, ubiquitous modern species and corresponding weightings; P, occasionally present;p, rare. Colours correspond to associated SST (as coded to Mudie et al., 2002a): dark blue, high salinity; medium blue, taxapresently found only in the core-tops of the Marmara Sea; light blue, freshwater or very low-salinity; green, ubiquitous. Dinocystspecies above the heavy line are autotrophs; below the line are known or presumed heterotrophs.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 155

Plate I. Environmental Scanning Electron Microscope micrographs of the freshwater and brackish water species Spiniferites cruci-formis, S. inaequalis, and Gonyaulax apiculata. Scale bar = 10 Wm.
1. Spiniferites cruciformis form 5 of Mudie et al. (2001), with only traces of paratabulation.2. Spiniferites cruciformis form 4 of Mudie et al. (2001).3. Spiniferites cruciformis with reduced processes.4. Spiniferites cruciformis form 3 of Mudie et al. (2001).5. Spiniferites cruciformis form 2 of Mudie et al. (2001).6. Spiniferites inaequalis. Dorsal view showing the archeopyle.7. Spiniferites inaequalis. Ventral view showing the sulcus.8. Gonyaulax apiculata. Right lateral view showing the paratabulation and the characteristic reticulation pattern on the cyst
wall.9. Gonyaulax apiculata. Left lateral view showing part of the archeopyle and the antapical plate.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167156
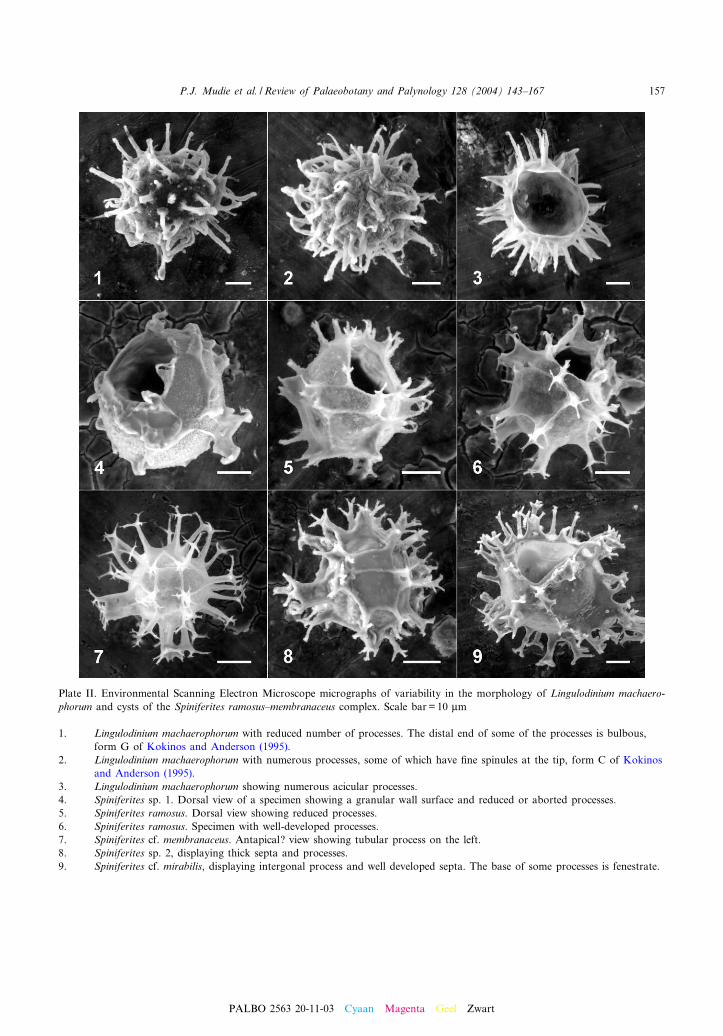
Plate II. Environmental Scanning Electron Microscope micrographs of variability in the morphology of Lingulodinium machaero-phorum and cysts of the Spiniferites ramosus^membranaceus complex. Scale bar = 10 Wm
1. Lingulodinium machaerophorum with reduced number of processes. The distal end of some of the processes is bulbous,form G of Kokinos and Anderson (1995).
2. Lingulodinium machaerophorum with numerous processes, some of which have ¢ne spinules at the tip, form C of Kokinosand Anderson (1995).
3. Lingulodinium machaerophorum showing numerous acicular processes.4. Spiniferites sp. 1. Dorsal view of a specimen showing a granular wall surface and reduced or aborted processes.5. Spiniferites ramosus. Dorsal view showing reduced processes.6. Spiniferites ramosus. Specimen with well-developed processes.7. Spiniferites cf. membranaceus. Antapical? view showing tubular process on the left.8. Spiniferites sp. 2, displaying thick septa and processes.9. Spiniferites cf. mirabilis, displaying intergonal process and well developed septa. The base of some processes is fenestrate.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 157

I, 1,2) comprise 10^50% of the S. cruciformis cystsin Dinocyst Assemblage Zone B1c. These S. cru-ciformis morphotypes with extremely reduced pro-cesses and the presence of marine (S. mirabilis)and low-salinity taxa (P. ponticum and Spiniferitesspp. with reduced processes as presently found inthe Baltic Sea) distinguishes the Black Sea B1cassemblage from the freshwater Spiniferites cruci-formis^Gonyaulax apiculata assemblage found inlate glacial sediments of northern Greece (Kouliet al., 2001; Mudie et al., 2002a). In particular, ithas been noted that S. cruciformis forms 4 and 5(Plate I, 1,2) have not been reported for fresh-water lakes and are believed to be indicators ofthe presence of some seawater (see Mudie et al.,2001, 2002a). Assemblage B1c corresponds to sur-face salinities of V12^17 according to salinityestimates obtained from planktonic foraminiferapreserved together with the dinocysts in perigla-cial mud at the base of the Marmara Sea coreMAR97-5 (Mudie et al., 2001) and coresMAR98-12 and 97-11 (this paper). In some BlackSea cores, oxygen isotopic measurements on cal-citic carbonate (Deuser, 1972) suggested that thesalinity of the Black Sea during the deposition ofthe early Holocene sediment was V5^7, while iondi¡usion studies (cited by Deuser, 1972) indicateda salinity of V3.5 from 20 000 to 8000 yr BP.
4.2.1.3. Dinocyst Assemblage Zone B1b (7.1^V3ka). The middle Holocene lithofacies in coresB7 and B13 is a 35^50-cm-thick laminated sapro-pel with a TOC content of 3^8 g l31 and abruptcontacts at top and base. Three to ¢ve sampleswere examined from this lithofacies in cores B7and B13, respectively. The average marine dino-cyst concentrations are high (6000/cm3) but £uc-tuate from 1000 to 10 000. The sapropel containsDinocyst Assemblage Zone B1b that is dominatedby Lingulodinium machaerophorum (10^20%),Operculodinium centrocarpum sensu Wall andDale 1966 (10^20%), and by Spiniferites taxathat are common in the Aegean (Spiniferites ben-torii, S. bulloideus, S. ramosus, S. hyperacanthusand S. mirabilis, 8^20%). Spiniferites cruciformis israre (6 10%) and represented mostly by forms 3and 4 (Plate I, 2,4) with medium or short process-es, whereas the Spiniferites spp. with variable pro-
cesses are common (Plate II, 4). Brigantediniumspp. increase and other heterotrophic protoperidi-nioids and polykrikoids make their ¢rst appear-ances up-section, e.g. Protoperidinium stellatumand Polykrikos kofoidi. The stenohaline speciesPyxidinopsis psilata is still occasionally present(6 10%). The processes of L. machaerophorumare variable in length and shape (Plate II, 1^3)as reported for low-salinity environments (seeMudie et al., 2001 for details) although none arecompletely lacking the development of spineshafts. Process length is also variable in O. cen-trocarpum, with most cysts having only longspines, some cysts having spines from 1 to 6 Wmlong, and rare cysts having truncate spines 1^2Wm long. Salinity estimates for this zone rangefrom 15^18 (Mudie et al., 2001) to 10^12 (Deuser,1972).
4.2.1.4. Dinocyst Assemblage Zone B1a (V3^0ka). The youngest sediment unit and the surfacesediment in cores B7 and B13 are organic-richmicrolaminated coccolith muds. Dinocyst abun-dances of V1000 cysts/cm3 are lower than inthe mid-Holocene sapropel although TOC re-mains s 2% and there is only a slight reductionin Mn levels (Arthur and Dean, 1998). Three sam-ples from this lithofacies show that heterotrophicprotoperidinioids dominate, forming AssemblageZone B1a with a preponderance of Brigantediniumspp., Protoperidinium stellatum, Selenopemphixquanta, S. nephroides, together with Peridiniumponticum (5^10%) and with morphotypes of Lin-gulodinium machaerophorum and Operculodiniumcentrocarpum that have processes of variablelength. The low-salinity indicators Spiniferites cru-ciformis and Pyxidinopsis psilata become rare orabsent above the base of this palynozone. Isotopicdata (Deuser, 1972) indicate salinities of 17^19and values of 18^22 were given by Wall et al.(1973) for Facies A in core 1745. These salinityestimates are in close agreement with present-daysurface salinities (Tables 1A and 1B).
In core 98^4, the youngest sediment unit con-sists of about 30 cm of olive mud with 1.2% TOC(Abrajano et al., 2002) and at the base it has anage of about 6 ka based on mollusc data. Thelowest sample contains an assemblage identical
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167158

to that in Dinocyst Zone B1a described above.Above 5680 yr BP, however, Lingulodinium ma-chaerophorum, Peridinium ponticum, Brigantedi-nium spp. and other protoperidinioids are stillthe dominant taxa as in cores B7 and B13, butSpiniferites ramosus and S. mirabilis are also com-mon and the youngest assemblages at this siteclose to the entrance of the Bosphorus Strait aremore like those of Dinocyst Assemblage Sub-zoneD1a in the Marmara Sea (see Section 4.2.2.3 be-low).
4.2.2. Marmara Sea coresIn the Marmara Sea, two cores were studied
from the western basin (MAR97-2) and saddle(MAR98-12). In the eastern Marmara Sea, threecores (MAR97-11, 98-7 and 98-9) were studiedfrom shelf areas and one core (MAR97-5) wasfrom the deep eastern basin (Fig. 2). Multipleradiocarbon ages for mollusc shells show thatthe sediments cover an age range from 15 590 to1990 yr BP (Tables 1A and 1B; Fig. 6).
4.2.2.1. Dinocyst Assemblage Zone D2 (V29^10.2 ka). Sediments of the last glacial maximumand late glacial intervals (V29 000^11 000 yr BP)in cores MAR98-12 (5 samples), MAR97-5 (13samples) and MAR97-11 (17 samples) containlow concentrations of both freshwater and marinedinocysts. This Dinocyst Assemblage Zone D2 isdominated (40^80%) by Spiniferites cruciformis(mostly forms 1 and 2 of Mudie et al., 2001,with long processes and expanded periphragms;Plate I, 5) and there are minor amounts of Peri-dinium ponticum, Pyxidinopsis psilata and Gonyau-lax apiculata. Spiniferites mirabilis and other go-nyaulacoid salt tolerant Aegean species are absentbut there are low percentages of the Spiniferitesmembranaceus-type species with variable spinemorphology, and cysts of Lingulodinium machae-rophorum and Operculodinium centrocarpum arepresent. Pterocysta cruciformis (Rochon et al.,2002) is also occasionally present, and in coreMAR98-12 there are a few Spiniferites inaequalis.
4.2.2.2. Dinocyst Assemblage Zone D1: Sub-zoneD1b (V10.2^5.7 ka). Dinocyst assemblagesthroughout the Holocene sediments are dominat-
ed by the wide-ranging species Lingulodinium ma-chaerophorum, Operculodinium centrocarpum andSpiniferites ramosus, with some Brigantediniumand other protoperidinioid cysts. Two Holocenedinocyst sub-zones are recognised. The early Ho-locene Sub-zone D1b has shell ages of 10 200 to6120 yr BP but exact dating of the upper assem-blage zone boundary is di⁄cult because the agesmust be extrapolated from an interval in whichsedimentation rates are rather variable in theMarmara Sea cores (see Fig. 6). In particular,the extrapolated age of 7285 yr BP for coreMAR98-9 is considered unreliable, and we haveprovisionally set the age of the assemblage sub-zone top at 5674 yr BP, that is the average valuefor the three other cores containing this sub-zone.Dinocyst Assemblage Sub-zone D1b is dominatedby O. centrocarpum, with variable amounts of thelow-salinity indicators Peridinium ponticum, Pyxi-dinopsis psilata and Spiniferites cruciformis. Spi-niferites mirabilis and protoperidinioid species arerare and Polykrikos is absent. Spiniferites spp.with variable spine morphology are common.There are also coinciding peaks of the freshwaterindicators Pediastrum, Sigmopollis psilata andTetraploa-type fungal spores at about 10 000^9000 yr BP. These peaks also correspond withthe maxima of Sigmopollis and Tetraploa-typespores found in the early Holocene sediments ofBlack Sea core B-7 (Mudie et al., 2002a). Thefreshwater species Gonyaulax apiculata is alsopresent at the base of core 98-09. Salinity is esti-mated as 12^17 based on correlation between thepercentages of Spiniferites cruciformis morpho-types and foraminiferal proxy-salinity data.
4.2.2.3. Dinocyst Assemblage Zone D1: Sub-zone1a (V5.7^0 ka). In the youngest sub-zone,D1a, Lingulodinium machaerophorum is dominantexcept near the Aegean Sea, where Operculodi-nium centrocarpum is the most abundant species.Relatively high percentages (5^20%) of Spinife-rites mirabilis and S. hyperacanthus accompanythese taxa. There are also rare occurrences ofOperculodinium israelianum and/or Bitectatodiniumtepikiense (MAR98-12, MAR97-11 and MAR98-9), and a diversity of heterotrophic taxa, includingIslandinium minutum, Polykrikos kofoidi, Seleno-
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 159

pemphix quanta, S. nephroides and Protoperidi-nium stellatum. Salinities are the same as thepresent range of 22-28, or possibly, periodicallyhigher during the intervals with Aegean speciesthat are presently not found in the MarmaraSea (e.g. Operculodinium israelianum, Tectatodi-nium pellitum, P. stellatum, Nematosphaeropsislabyrinthus).
5. Discussion
5.1. Core top distributions
Principle component analysis of 16 core-topsamples shows that core-top assemblages domi-nated by cysts of autotrophic gonyaulacoids dis-tinguish the modern sediments overlain by high-salinity Aegean^Mediterranean water, whereasheterotrophic protoperidinioids characterise thelow-salinity Marmara^Black Sea region. The re-cent changes in ¢shing practices and nutrient dis-charge make it di⁄cult to specify the main envi-ronmental factor governing this distribution.However, the relative uniformity in dinocyst as-semblages from the high-resolution core 97-02(Mudie et al., 2002a), shows that there has beenno major change in production of resting cystsduring the past 2000 years at this site near theentrance to the Aegean Sea and downstream ofthe highly polluted Black Sea. The absence ofmajor shifts in cyst assemblages during the past30 years suggests that changes in nutrient levelsand concomitant changes in food web dynamicsare not primary factors governing cyst assemblagecomposition. Therefore, we can reasonably com-pare the modern cyst distributions with the otherpresent-day environmental parameters describedearlier in this paper: temperature, strati¢cationand salinity.
First, if we assume that most cysts are pro-duced in the surface water layer during summer,it is clear that SST does not di¡er greatly betweenthe Marmara^Black Sea areas with high loadingsfor PC-1 (mostly heterotrophic protoperidinioidsand ubiquitous or opportunistic gonyaulacoids)and the Aegean PC-2 assemblages (mostly go-nyaulacoids restricted to subtropical/temperate re-
gions and salinity s 32). Major di¡erences inwater strati¢cation are also only evident withinthe Marmara Sea where the surface layer is about10 m thick compared to 100 m or more elsewhere.Beneath this shallow halocline there is no signi¢-cant change in cyst distribution among the 11Marmara^Black Sea core-tops presently analysed;however, a stronger link may appear when thenumber of core tops is increased to 100 in thefuture, using new samples from the MarmaraGateway collected in the summer of 2002 (Aksu,pers. commun., January 2003).
At present, it appears that surface salinity ac-counts for the largest di¡erence in environmentalparameters between the distributions of PC-1 and-2 in the Aegean^Black Sea corridor. This ¢ndingis consistent with other studies of dinocyst assem-blages in warm temperate and tropical environ-ments. For example, Marret (1994) found thatsalinity in the range of 29^35 was more importantthan SST (23^28‡C) in the eastern EquatorialAtlantic, where cyst assemblages are generallydominated by Brigantedinium spp., Spiniferitesdelicatus, Polykrikos kofoidi, and Lingulodiniummachaerophorum. Here the distributions of Oper-culodinium israelianum, Spiniferites membranaceusand Selenopemphix nephroides are linked with sal-inities of 30^34, whereas Spiniferites delicatus,P. kofoidi, Spiniferites mirabilis, S. ramosus, andLejeunecysta sabrina are found in areas with sa-linity s 34. These tropical distributions onlypartly match those in our study area where thecore-top distribution of O. israelianum is re-stricted to areas with salinities of s 34, and whereS. mirabilis, S. ramosus and S. membranaceus areessentially ubiquitous, occurring in salinities of17^38. Tuberculodinium vancampoae is ubiquitousand euryhaline in the Equatorial region, but inour study area, it is found only in the hypersalinewaters of the southern Aegean Sea.
Nematosphaeropsis labyrinthus, Impagidiniumaculeatum and I. patulum are restricted to oceanicenvironments in both the Equatorial Atlantic andin our study area where only the deep basins ofthe central and southern Aegean Sea (A-19 toG-22 in Tables 1A and 1B) have relatively stableoceanic-type conditions. Dodge (1994) also foundthat Impagidinium spp. and N. labyrinthus are as-
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167160

sociated with warm temperate waters (16^21‡C)and upwelling conditions in the eastern NorthAtlantic Ocean, and I. aculeatum, I. strialatum,I. patulum and N. labyrinthus are common inlate Holocene sediments of oceanic sectors ofthe western and central Mediterranean seas (Tu-ron and Londeix, 1988; Zonneveld, 1996). In con-trast, no Impagidinium species were found in anyof the Marmara Gateway core-tops or Late Pleis-tocene core samples from the Marmara Sea andBlack Sea, although three species are present inthe late Holocene sediments of the Aegean Sea(Aksu et al., 1995), and Marret et al. (2003) hasrecently described some endemic Impagidiniumspecies from Holocene sediments in the CaspianSea. N. labyrinthus is also rare east of the AegeanSea, occurring in only a few samples from theMarmara Sea. Another widespread North Atlan-tic species, Spiniferites elongatus, is also notablyabsent from our entire study area, although inother regions it is known from waters with a sa-linity as low as 20. Two other widespread NorthAtlantic species, Bitectatodinium tepikiense andTrinovantedinium applanatum, are very rare inthe lower salinity waters east of the Aegean Sea.These two species range from tropical/subtropicalto polar waters but do not occur in waters with asalinity of less than 28.
In a survey of modern dino£agellates in theeastern North Atlantic, Dodge (1994) notedthat: (1) only the gonyaulacoids show a clear in-£uence of temperature, and (2) protoperidinioidsare rare in the warm temperate region. This dom-inance of gonyaulacoids over protoperidinioids isevident in surface samples from the Aegean sectorof our study area, but it does not persist down-core where the gonyaulacoid:protoperidinioid ra-tio is only slightly less than 1 throughout Assem-blage Zones D1a and B1a (see Table 2). Thisapparent link between temperature and speciesrichness of protoperidinioids in middle^low lati-tudes requires more study before it can be appliedwidely.
Data from previous studies also suggest thatthere are regional variations associated with salin-ity within the Mediterranean Sea. The nutrientand SST values given in Section 2.4, and otherdata sources (e.g. Civili, 1992), show that all Med-
iterranean Sea water beyond a few polluted coast-al areas is oligotrophic compared to the Aegean^Black Sea corridor, and the annual temperaturerange is V14^28‡C throughout. In the coastalarea (6 200 m) of the Tyrrhenian Sea, where sa-linity is normally 37.8^38.2 (rarely 35^36 afterrain), the main organic-walled cysts are Operculo-dinium centrocarpum, Brigantedinium spp. andother protoperidinioid taxa, and Spiniferites is rel-atively rare (Montresor et al., 1998). Further o¡-shore, at depths of s 600 m in the Adriatic Sea,eastern Ionian and northern Levantine basins,surface salinity is only slightly reduced during£ooding of the Po River (Zonneveld, 1996).Here, O. centrocarpum and Impagidinium spp.dominate together with common Spiniferitesspp., and although Brigantedinium spp. are alsocommon, other protoperidinioids are rare in thelate Holocene assemblages, showing that in theo¡shore areas there has been no major changein the dominance of gonyaulacoids as a result ofrecent increases in nutrient-rich e¥uent dischargeconditions. Similar dinocyst assemblages arefound in calcareous mud at the top of a core at1590 m depth on the Rhone Delta (Morzadec-Kerfourn, 1992) and in cores from the AlboranSea (855^1375 m depth) near the Strait of Gibral-tar where summer SST and SSS are slightly lower(21^22‡C and 36.3, respectively). Here, however,Spiniferites mirabilis, S. hyperacanthus, S. delica-tus and S. bentorii are co-dominant, and Lingulo-dinium machaerophorum is also common (Turonand Londeix, 1988). In contrast, O. centrocarpumwas not found in recent sediments from nearshoreareas o¡ the desert regions of Tunisia and Israel,where L. machaerophorum, Polysphaeridium zo-haryi, Tuberculodinium vancampoae and Spinife-rites spp. are the most common taxa (Morzadec-Kerfourn, 1979, 1992; Rossignol-Strick, 1964).There is a clear need for more studies of thesesub-regional variations in salinity in relation tothe major di¡erences in dinocyst assemblageswithin the Aegean^Marmara^Black Sea corridor.
5.2. Morphological variations
Several morphotypes, principally of Spiniferites,have been identi¢ed in the core-top dinocyst as-
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 161

semblages. Cysts resembling S. bulloideus andS. mirabilis but with short, acuminate, underde-veloped processes, and over-developed septa(Plate II, 4,7^9) cannot be assigned to a knownspecies or sub-speci¢c taxon. Several early Holo-cene intervals in the Marmara Sea cores also con-tain a high percentage of these unusual Spinife-rites morphotypes (e.g. Plate II, 4,5) that are alsocommon in Assemblage Zones B1b and B1a ofthe Black Sea cores. Previous studies (Wall etal., 1973; Mudie et al., 2001) have also reporteda wide range in variation of spine length for Spi-niferites cruciformis, Operculodinium centrocarpumand Lingulodinium machaerophorum in the BlackSea and the Marmara Sea. It has been suggestedthat these variations are a response to reducedsalinity (Dale, 1996), changes in strati¢cationand/or stress in the physical environment (Mudieet al., 2001). Observations on natural and labora-tory populations also show that dino£agellatescan produce a wide range of cyst morphologies,depending upon the temperature and/or salinityconditions (Lewis et al., 1999) but without a uni-directional pattern. Under laboratory conditions,several morphotypes may co-exist in one culture,but a particular form may dominate over theothers. The high-salinity gradient observed be-tween the Aegean and Black Sea may explainthe morphological di¡erences that we observe inour samples. However, we know relatively littleabout either the ecology of those morphotypes,or the salinity and/or temperature conditions re-sponsible for such variations. Morphological var-iations in the process length and shape of O. cen-trocarpum, S. bulloideus and Pyxidinopsis psilatain recent sediments of the Baltic Sea and Norwe-gian ¢ords strongly suggest that these changesoccur where the average salinity is less than about6^9 (Matthiessen and Brenner, 1996; Dale, 1996;Nehring, 1997). Likewise, the late glacial low-salinity S. cruciformis cysts from the Black Seaand Marmara Sea tend to show a wide range ofwithin-population variation in process develop-ment, including apparent abnormalities, but pres-ently it is not possible to relate this variation tosalinity with quantitative signi¢cance (see Mudieet al., 2001). Therefore, the interpretation offossil assemblages and palaeoenvironments re-
main limited until more quantitative data areavailable.
Cyst assemblages dominated by Spiniferites cru-ciformis, S. inaequalis and Pyxidiella psilata arenot present during the middle^late Holocene.Although we know that these taxa are rare todayin the low-salinity Black Sea and the MarmaraSea, and P. psilata occurs in the Baltic region ata salinity of 1^5 (Dale, 1996 and unpublisheddata), we have no modern analogue to illustratethe environments in which those species had theiroptimum development. In Mudie et al. (2001) wetried to compensate for this limitation by calibrat-ing percent abundance of variant morphotypesagainst salinity estimates from measured changesin the N O18 content of planktonic foraminifera.Although our results may give too wide a range ofsalinity estimate (11^17) compared to the results(18^22) from new transfer functions for salinity(Sperling et al., 2003), our quantitative dinocystdata are more re¢ned than the salinity estimatesbased on the presence of the coccolith Emilianiahuxleyi (salinity range 11^38) or so-called fresh-water mollusks (1^13). New studies of so-called‘freshwater’ Caspian mollusks, e.g. Dreisseniapolymorpha and Dreissena rostriformis, also showthat some populations have a plastic response tochanging salinity and adjust to changes from 1 to12 over about three breeding generations (Mills etal., 1996). For example, D. rostriformis grimmiihas adapted to the increased salinity of the Cas-pian Sea and D. rostriformis grimmii presently in-habits waters with a salinity of 12.7. The greaterwater-column depth of planktonic foraminifera(25^250 m) further limits the interpretation ofcorrelation between near-surface salinity changeand morphological variation in autotrophic dino-£agellates, and reduces the accuracy for use intransfer functions for quantitative reconstructionof palaeoenvironments.
5.3. Cyst preservation in the study area
Previous studies (Abrajano et al., 2002; Mudieet al., 2002a) have shown that preservation ofthin-walled protoperidinioid species is uniformlygood in the organic-rich, marginal seas of ourstudy area. It is also evident that elevated Mn
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167162

levels in our cores correspond with the boundariesof the sapropelic units in contrast to the Levan-tine Basin, where a ‘£oating’ Mn boundary marksthe oxidised sapropel top (Zonneveld et al., 2001).In the Black Sea cores B13 and B7, dinocyst con-centrations are lower in the carbonate mud sedi-ments of Assemblage Zone B1a than in the sap-ropel containing Assemblage Zone B1b, althoughthe TOC in Zone B1a is s 2% and the Mn levelsare only slightly lower than in the sapropel (Ar-thur and Dean, 1998). Despite this reduction incyst concentration in Zone B1a, the relative abun-dances of oxidation-sensitive protoperidinioidsare much higher than in the sapropel. Conversely,Brigantedinium is also common in the lower inter-val (Last Glacial Maximum s 15 000 yr BP) ofthe deltaic sequence in core MAR97-11 (see Mu-die et al., 2002a), although the TOC content ofthis unit is only about 0.25% compared to theoverlying sapropelic and surface muds (1^1.5%).It is therefore clear that, in the organic-rich, ¢ne-grained sediments of the Marmara Gatewaychanges in cyst concentrations and assemblagecompositions are not simply the result of varia-tions in bottom water redox conditions that allowbetter preservation of thin-walled cysts, but re£ectmajor shifts in palaeoclimatic conditions such asSST and SSS. Consequently, in the MarmaraGateway, di¡erential diagenesis of thin-walledtaxa does not bias the interpretation of assem-blages in the sapropelic units as reported for theeastern Mediterranean Sea by Zonneveld et al.(2001).
5.4. Marmara Gateway salinity models
Global sea level and the depths of channelsconnecting the Aegean Sea, the Marmara Seaand the Black Sea are the primary factors thatcontrol the salinity history of the Marmara Gate-way. The climate also controls the amount of gla-cial meltwater and river runo¡ to the Black Sea,and the balance between precipitation and evap-oration (P3E). Water exchange via the MarmaraGateway controls the amount of salt input to theBlack Sea, and the rate of upward mixing of sa-line bottom water in the Black Sea further regu-lates the volume of water exchange and salt re-
moval (see Boudreau and Leblond, 1989; Mudieet al., 2002a). Major et al. (2002) review the var-ious palaeoceanographic models that describe thesalinity history of the Marmara Gateway since thelast glaciation. These authors show the need for asensitive geochemical indicator of marine water todistinguish between the Ryan and Pitman Noah’sFlood ‘freshwater lake’ model and the semi-per-sistent out£ow model (e.g. Aksu et al., 2002a). Itshould be noted that although the Noah’s Floodmodel (Ryan et al., 1997; Ryan and Pitman,1999) embodies the Biblical idea of a very rapid(40 days and 40 nights) £ooding which cannot betested with the resolution of our data (maximumresolution is V15 years at site MAR97-2), otherelements of the Flood model can be tested by ourpalynological data. These elements include thelowering of the Black Sea water level to V100m below the Bosphorus Strait entrance becauseof dry conditions in the watershed immediatelyprior to 7.5 ka, the time of the purported £ood,and the presence of a freshwater lake supportinglarge Neolithic agricultural settlements that disap-peared when drowned by saline MediterraneanSea water.
Mudie et al. (2002b) have shown that the pollenassemblages in the Black Sea and the MarmaraSea cores indicate the presence of temperate mesicforest in this region by 9.5 ka which contradictsthe hypothesis of cold and dry conditions that arerequired for 3100 m water level lowering in theRyan and Pitman model. The abundance of Che-nopodiaceae pollen in the early Holocene assem-blages also suggests that saltmarshes were wide-spread on the shorelines, in agreement with thedinocyst evidence for brackish water conditions(Mudie et al., 2001, 2002a and this paper). Wehave also demonstrated (Mudie et al., 2002a)that the total removal of salt from the BlackSea during the interval of glacial sea-level lower-ing would require a much longer time of fresh-water export than is provided by the model ofRyan and Pitman. Several authors (e.g. Aksu etal., 2002a,b and references therein) have also usedseismostratigraphic data to infer that water was£owing out of the Black Sea into the Aegean Seaby about 10 ka.
In our earlier paper (Mudie et al., 2002a), we
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 163

mentioned the importance of using appropriateterminology to describe the salinity of water inmodels that concern the relationship betweenwater salinity, archaeological settlements andagriculture. In this paper, before discussing theecostratigraphic importance of our dinocyst as-semblage data, we further emphasise the need toapply appropriate salinity terms when describinglow-salinity water bodies. Although oceanogra-phers sometimes refer to low-salinity inlandwaters as freshwater lakes (e.g. the Bras d’OrLake, Nova Scotia, SSS = 27), it must be remem-bered that in water management and agriculturalterminology, freshwater has a salinity of 6 2,brackish water is V2^7, and water with a higherdissolved salt content is saline. It is therefore lessconfusing to use water management terminologyfor the salinity in the Marmara Gateway becauseNoah’s Flood was supposed to have destroyedNeolithic farming communities on the shores ofthe Black Sea and changed the direction of agri-cultural settlement in Europe and Asia (Ryan etal., 1997; Wilson, 2001). In fact, there is no un-equivocal evidence for Black Sea water with asalinity of 6 2 at the time of the purported earlyHolocene Neolithic settlement. Modern agricul-tural disasters, such as the failed irrigation proj-ects of the Indus Valley and Colorado Desert,show that even with complicated tools and powersupplies, irrigation agriculture is not normallysustainable in warm climate regions in the absenceof abundant freshwater (salinity 6 1) for £ushingthe salts that accumulate in clay-containing soils(see Mudie, 1974 and references therein). Further-more, it is unlikely that irrigation technology ex-isted in the Middle East before about V4.6 ka,and even in areas of well-developed Bronze Ageirrigation, toxic levels of salt build-up occurredafter about 400 years of irrigation (Jorgensenand Yasin al-Tikiriti, 2002). Around the BlackSea at 7500 yr BP, the availability of freshwaterwould have been con¢ned to a narrow striparound the deltas of the Danube and large north-ern rivers. The coastal plain south of the DanubeDelta may have supported small Neolithic farm-ing communities around 9 ka (Bozilova et al.,1996), but these were ephemeral (gone byV8 ka), and they would also have been far inland
from the lowered Black Sea shoreline commun-ities purportedly drowned at 7.5 ka (Ryan andPitman, 1999). The pollen records from cores inthe Black Sea and the Marmara Sea (Mudie et al.,2002b) and the deltas of the southwesternUkraine (Kremenetski, 1995) show no evidenceof sustained forest clearance or agriculture beforeV4 ka when the ¢rst signs of agriculture (cereals)and horticulture (olives and walnuts) appear.
The data in Table 2 show that only the plen-iglacial glacial Dinocyst Assemblage Zone B2contains virtually no dinocyst species that are to-day common in brackish or saline waters (onlyLingulodinium machaerophorum and Spiniferitesmembranaceus-type cysts are present). In ZoneB2 the occurrence of the freshwater species Go-nyaulax apiculata together with Spiniferites cruci-formis forms 1^3, that are common in late glacialGreek lakes (Kouli et al., 2001) and Pyxidiellapsilata that occurs today in freshwater (salinity6 2) ponds of inner Norwegian ¢ords, is strongevidence for a very low-salinity environment.However, the concurrent presence of the ubiqui-tous marine species L. machaerophorum and thelow-salinity species Spiniferites inaequalis that ispresently found in the Black Sea and MarmaraSea suggest that there was some mixing of saltwater into the surface layer. Similarly, the lateglacial^early postglacial Zones D2 and B1c alsocontain G. apiculata and are dominated by S. cru-ciformis forms 1^2 and P. psilata. Also, thesezones have few ubiquitous marine species andno hypersaline Aegean taxa. However, the pres-ence of the ubiquitous marine species Operculodi-nium centrocarpum and L. machaerophorum, nor-mally found in salinities of s 7 (e.g. Dale, 1996),clearly indicates that the inland basins were notentirely freshwater from as early as V16^10 ka inthe Marmara Sea and V9.8^7 ka in the BlackSea. The question remains, however, whether thebrackish water environments were isolated lakesor interlinked basins receiving periodic inputs ofAegean Sea water. Mudie et al. (2002a) haveshown, however, that it would require s 10 000years to reduce the surface salinity of the BlackSea to a salinity of 1 even when it was continu-ously linked to the Marmara and Aegean seas andbeing continuously £ushed by freshwater. If the
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167164

basins were isolated lakes, the removal of the saltwould not be possible except by precipitation asmineral deposits.
The postglacial Marmara Sea assemblage Sub-zone D1b and the mid-Holocene Black Sea ZoneB1b clearly re£ect the in£ux of Aegean Sea waterand considerable mixing of brackish water andubiquitous marine taxa by at least 9.5 ka. Thistrend has increased during the past to 4000 (Mar-mara Sea) and to 3000 years (Black Sea), and hasalso been accompanied by increases in protope-ridinioid species that may re£ect greater primaryproductivity. In conclusion, it is clear that even ata semi-quantitative level, organic-walled dinocystsare very useful markers of surface water salinityin the Marmara Gateway, and our data clearlysupport the semi-persistent over£ow model thatindicates water exchange between the MarmaraSea and the Black Sea by at least 9.5 ka (Fig.3B). Future work will include analysis of 100core-top samples that have been collected fromtransects from V50^200 m over the shelves ofthe Black Sea, the Marmara Sea and the AegeanSea to obtain quantitative transfer functions (seeRochon et al., 1999) for backtracking the historyof salinity and SST. Furthermore, we stronglyadvocate the employment of a precise salinity ter-minology when relating the history of salinitychanges to human settlement and land use.
Acknowledgements
We thank the o⁄cers and crew of the RV KocaPiri Reis for their assistance in data collection oncruises MAR97 and MAR98. We acknowledgeresearch and ship time funds from the NaturalSciences and Engineering Research Council ofCanada, travel funds from the Dean of Science,Memorial University of Newfoundland, a grantfrom the Piri Reis Foundation for Maritime andMarine Resources Development and Education,and funding support of the Geological Survey ofCanada. This paper was improved by the helpfulreviews of Drs. Rob Fensome and Graham Wil-liams (GSCA) and Dr. Suzanne Leroy (BrunelUniversity). This is Geological Survey of CanadaContribution No. 2003057.
References
Abrajano, T., Aksu, A.E., Hiscott, R.N., Mudie, P.J., 2002.Aspects of carbon isotope biogeochemistry of late Quater-nary sediments from the Marmara Sea and Black Sea. Mar.Geol. 190, 151^164.
Aksu, A.E., Yas›ar, D., Mudie, P.J., Gillespie, H., 1995. Lateglacial^Holocene paleoclimatic and paleoceanographic evo-lution of the Aegean Sea: Micropaleontological and stableisotopic evidence. Mar. Micropalaeontol. 25, 1^28.
Aksu, A.E., Hiscott, R.N., Yas›ar, D., 1999a. Oscillating Qua-ternary water levels of the Marmara Sea and vigorous out-£ow into the Aegean Sea from the Marmara Sea^Black Seadrainage corridor. Mar. Geol. 153, 275^302.
Aksu, A.E., Abrajano, J., Mudie, P.J., Yas›ar, D., 1999b. Or-ganic geochemical and palynological evidence for terrige-nous origin of the organic matter in Aegean Sea sapropelS1. Mar. Geol. 153, 303^318.
Aksu, A.E., Hiscott, R.N., Mudie, P.J., Rochon, A., Kamin-ski, M.A., Abrajano, T., Yas›ar, D., 2002a. Persistant Holo-cene out£ow from the Black Sea to the Eastern Mediterra-nean contradicts Noah’s Flood hypothesis. GSA Today 12,4^9.
Aksu, A.E., Hiscott, R.N., Mudie, P.J., Gillespie, H., Yas›ar,D., 2002b. Last glacial^Holocene sea surface temperatureand salinity variations in the Black and Marmara Sea: Sta-ble isotopic, planktonic foraminiferal and coccolith evi-dence. Mar. Geol. 190, 119^149.
Arthur, M.A., Dean, W.E., 1998. Organic-matter productionand preservation and evolution of anoxia in the HoloceneBlack Sea. Paleoceanography 13, 395^411.
Ballard, R.D., Coleman, D.F., Rosenberg, G.D., 2000. Fur-ther evidence of abrupt Holocene drowning of the Black Seashelf. Mar. Geol. 170, 253^261.
Black Sea Pilot, 1990. Marmara Denizi, Black Sea and Azov-skoye More with adjacent coasts of Turkey, Bulgaria, Ro-mania and USSR, 12th ed. Great Britain Hydrographic De-partment, Taunton, 231 pp.
Bogucki, P., 1996. The spread of early farming in Europe. Am.Sci. 84, 242^253.
Boudreau, B.P., Leblond, P.H., 1989. A simple evolutionarymodel for water and salt in the Black Sea. Paleoceanography4, 157^166.
Bozilova, E., Filipova, M., Filipovich, L., Tonkov, S., 1996.Bulgaria. In: Berglund, B.E., Birks, H.J.B., Ralska-Jasiewic-zowa, M., Wright, H.E. (Eds.), Palaeoecological events dur-ing the past 15000 years: Regional syntheses of palaeoco-logical studies of lakes and mires in Europe. Wiley,Chichester, pp. 701^728.
Bukry, D., 1974. Coccoliths as paleosalinity indicators ^ evi-dence from the Black Sea. In: Degens, E.T., Ross, D.A.(Eds.), The Black Sea: Geology, Chemistry, and Biology.Am. Assoc. Pet. Geol. Mem. 20, Tulsa, OK, pp. 364^380.
Civili, F.S., 1992. Mediterranean water quality: The Mediter-ranean Action Plan and its environmental assessment com-ponent. In: Fabbri, P., Fiero, G. (Eds.), Semi-EnclosedSeas: Exchange of Environmental Experiences between
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 165

Mediterranean and Caribbean Countries. Elsevier, Barking,pp. 6^51.
Dale, B., 1996. Dino£agellate cyst ecology: Modeling and geo-logical applications. In: Jansonius, J., McGregor, D.C.(Eds.), Palynology: Principles and Applications. Am. Assoc.Stratigr. Palynol. Found., Dallas, TX, 3, pp. 124^1275.
Daskalov, G.M., 2002. Over¢shing drives a trophic cascade inthe Black Sea. Mar. Ecol. Progr. Ser. 225, 53^63.
Deuser, W.G., 1972. Late Pleistocene and Holocene history ofthe Black Sea as indicated by stable isotopic Studies. J. Geo-phys. Res. 77, 1071^1077.
Dodge, J.D., 1994. Biogeography of marine armoured dino£a-gellates and dinocysts in the NE Atlantic and North Sea.Rev. Palaeobot. Palynol. 84, 169^180.
Duman, M., 1994. Late Quaternary chronology of the south-ern Black Sea basin. Geol. Mar. Lett. 14, 272^278.
Eremeev, V.N., 1995. Hydrology and circulation of waters inthe Black Sea. Bull. Inst. oce¤anogr. Monaco 15, 27^41.
Fairbridge, R.W., 1966. Encyclopedia of Oceanography. Rein-hold, New York, pp. 490^495.
Fonselius, S.H., 1974. Phosphorus in Black Sea. In: Degens,Ross (Eds.), The Black Sea ^ Geology, Chemistry and Biol-ogy. Am. Assoc. Pet. Geol. Mem. 20, Tulsa, OK, pp. 144^150.
Garrels, R.M., Mackenzie, F.T., Hunt, C., 1975. ChemicalCycles and the Global Environment. William Kaufmann,Los Altos, CA.
Georghian, M., 1974. Distribution pattern of benthonic fora-minifera on continental shelf of Black Sea o¡ Rumanianshore. In: Degens, E.T., Ross, D.A. (Eds.), The BlackSea: Geology, Chemistry and Biology. Am. Assoc. Pet.Geol. Mem. 20, Tulsa, OK, pp. 411^418.
Go«ru«r, N., Cagflatay, M.N., Emre, Oº ., Alpar, B., Sak|nc, M.,Iºslamos›lu, Y., Algan, O., Erkal, T., Kecer, M., Akko«k, R.,Karl|k, G., 2001. Is the abrupt drowning of the Black Seamshelf at 7150 yr BP a myth? Mar. Geol. 176, 65^73.
Guiot, J., 1990. Methods and programmes of statistics for pa-leoclimatology and paleoecology. In: Guiot, J., Labeyrie, L.(Eds.), Quanti¢cation des changements climatiques: me¤-thodes et programmes. Institut National des Sciences del’Univers (INSU^France) Monographie 1, Paris, 253 pp.
Haas, P.M., Zuckman, J., 1990. The Med is cleaner. Oceanus33, 40^42.
Hiscott, R.N., Aksu, A.E., 2002. Late Quaternary history ofthe Marmara Sea and Black Sea from high resolution seis-mic and gravity core studies. Mar. Geol. 190, 261^282.
Hiscott, R.N., Aksu, A.E., Yas›ar, D., Kaminski, M.A., Mu-die, P.J., Kostylev, V., MacDonald, J., Is›ler, F.I., Lord,A.R., 2002. Deltas south of the Bosphorus Strait recordpersistent Black Sea out£ow to the Marmara Sea sinceV10 ka. Mar. Geol. 190, 95^118.
Jorgensen, D.G., Yasin al-Tikiriti, W., 2002. A hydrologicaland archeologic study of climate change in Al Ain, UnitedArab Emirates. Glob. Planet. Change 35, 37^49.
Kaminski, M.A., Aksu, A., Box, M., Hiscott, R.N., Filipescu,S., Al-salameen, M., 2002. Late glacial to Holocene benthicforaminifera in the Marmara Sea: Implications for Black
Sea ^ Mediterranean Sea connections following the last de-glaciation. Mar. Geol. 190, 165^202.
Kokinos, J.P., Anderson, D.M., 1995. Morphological develop-ment of resting cysts in cultures of the marine dino£agellateLingulodinium polyedrum ( =L. machaerophorum). Palynol-ogy 19, 143^166.
Kouli, K., Brinkhuis, H., Dale, B., 2001. Spiniferites crucifor-mis : A fresh water dino£agellate cyst? Rev. Palaeobot. Paly-nol. 113, 273^286.
Kremenetski, C.V., 1995. Holocene vegetation and climate his-tory of southwestern Ukraine. Rev. Palaeobot. Palynol. 85,285^301.
Lascaratos, A., Roether, W., Nittis, K., Klein, B., 1999. Re-cent changes in deep water formation and spreading in theeastern Mediterranean Sea. Progr. Oceanogr. 44, 5^36.
Lewis, J., Rochon, A., Harding, I., 1999. Preliminary observa-tions of cyst^theca relationships in Spiniferites ramosus andSpiniferites membranaceus (Dinophyceae). Grana 38, 113^124.
Lewis, J., Rochon, A., Ellegaard, M., Mudie, P.J., Harding,I.C., 2001. Cyst^theca relationships in Bitectatodinium tepi-kiense (Dinophyceae). Eur. J. Phycol. 36, 137^146.
Major, C., Ryan, W., Lericolais, G., Hajdas, I., 2002. Con-straints on Black Sea out£ow to the Sea of Marmara duringthe last glacial^interglacial transition. Mar. Geol. 190, 19^34.
Malanotte-Rizzole, P., Robinson, A.R., Roether, W., Manca,B., Bergamasco, A., Brenner, S., Civitarese, G., Georgopou-lis, D., Haley, P.J., Kioroglou, S., Kontoyannis, H., Kress,N., Latif, M.A., Leslie, W.G., Oº zsoy, E., Ribera d’Alcala,M., Salihoglu, I., Sansone, E., Theocharis, A., 1996. Experi-ment in Eastern Mediterranean probes origin of deep watermasses. EOS Trans. Am. Geophys. Union 77, 305^311.
Marret, F., 1994. Distribution of dino£agellate cysts in recentmarine sediments from the east Equatorial Atlantic (Gulf ofGuinea). Rev. Palaeobot. Palynol. 84, 1^22.
Marret, F., Leroy, S., Chalie, F., Gasse, F., 2003. New organ-ic-walled dino£agellate cysts from recent sediments of Cen-tral Asian seas. Rev. Palaeobot. Palynol. (submitted).
Matthiessen, J., Brenner, W., 1996. Chlorococcalalgen und di-no£agellaten-zysten in rezenten Sedimenten des greifwalderBoddens (sudliche Ostesee). Senckenbergiana Mar. 27, 33^48.
Maynard, N.G., 1974. Diatoms in Pleistocene deep Black Seasediments. In: Degens, E.T., Ross, D.A. (Eds.), The BlackSea: Geology, Chemistry and Biology. Am. Assoc. Pet.Geol. Mem. 20, Tulsa, OK, pp. 389^395.
Mills, E.L., Rosenberg, G., Spidle, A.P., Ludyanskiy, P., Pli-gin, Y., May, B., 1996. A review of the biology and ecologyof the Quagga mussel (Dreissena bugensis), a second speciesof freshwater dreissenid introduced to North America. Am.Zool. 36, 271^286.
Montresor, M., Zinione, A., Sarno, D., 1998. Dino£agellatecyst production at a coastal Mediterranean site. J. PlanktonRes. 20, 2291^2312.
Morzadec-Kerfourn, M.T., 1979. Les kystes de Dino£agelle¤s.In: Burollet, P.F., Clairefond, P., Winnock, E. (Eds.), Lamer Pe¤lagienne. Ge¤ologie Me¤diterrane¤enne 6 (1), 221^248.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167166

Morzadec-Kerfourn, M.T., 1992. Upper Pleistocene and Holo-cene dino£agellate cyst assemblages in marine environmentsof the Mediterranean Sea and the Northwest Atlantic coast ofFrance. In: Head, M.J., Wrenn, J.H. (Eds.), Neogene andQuaternary Dino£agellate cysts and acritarchs. Am. Assoc.Stratigr. Palynol. Found., Dallas, TX, pp. 121^132.
Mudie, P.J., 1974. The potential economic uses of halophytes.In: Reimold, R.J., Queen, W.H. (Eds.), Ecology of Halo-phytes. Academic Press, New York, pp. 565^598.
Mudie, P.J., Aksu, A.E., Yas›ar, D., 2001. Late Quaternarydino£agellate cysts from the Black, Marmara and Aegeanseas: Variations in assemblages, morphology and paleosalin-ity. Mar. Micropalaeontol. 43, 155^178.
Mudie, P.J., Rochon, A., Aksu, A.E., Gillespie, H., 2002a.Dino£agellate cysts, freshwater algae and fungal spores assalinity indicators in Late Quaternary cores from Marmaraand Black Seas. Mar. Geol. 190, 203^231.
Mudie, P.J., Rochon, A., Aksu, A.E., 2002b. Pollen stratigra-phy of Late Quaternary cores from Marmara Sea: Land^seacorrelation and paleoclimatic history. Mar. Geol. 190, 233^260.
Nehring, S., 1997. Dino£agellate resting cysts from recent Ger-man coastal sediments. Bot. Mar. 40, 307^324.
Oº zsoy, E., Latif, M.A., Tugrul, S., Uº nluata, U., 1995. Ex-changes with the Mediterranean, £uxes, and boundary mix-ing processes in the Black Sea. Bull. Inst. Oce¤anogr. Mo-naco 15, 1^25.
Pilskaln, C.H., Pike, J., 2001. Formation of Holocene sedimen-tary laminae in the Black Sea and the role of the benthic£occulent layer. Paleoceanography 16, 1^19.
Robinson, A.R., Golnaraghi, M., 1994. The physical and dy-namical oceanography of the Mediterranean Sea. In: Mala-notte-Rizzole, P., Robinson, A.R. (Eds.), Ocean Processes inClimate Dynamics: Global and Mediterannean Examples.Kluwer, Dordrecht, pp. 255^306.
Rochon, A., de Vernal, A., Turon, J.L., Matthiessen. J., Head,M.J., 1999. Distribution of recent dino£agellate cysts in sur-face sediments from the North Atlantic Ocean and adjacentseas in relation to sea-surface parameters. Am. Assoc. Strat-igr. Palynol. Found., Contrib. Ser. 35, 152 pp.
Rochon, A., Mudie, P.J., Aksu, A.E., Gillespie, H., 2002.Pterocysta gen. nov. a new dino£agellate cyst from Pleisto-cene glacial-stage sediments of the Black and Marmara seas.Palynology 26, 95^105.
Ross, D.A., Degens, E.T., MacIlvane, J., 1970. Black Sea:Recent sedimentary history. Science 170, 163^165.
Ross, D.A., Degens, E.T., 1974. Recent sediments of the BlackSea. In: Degens, E.T., Ross, D.A. (Eds.), The Black Sea:Geology, Chemistry, and Biology. Am. Assoc. Pet. Geol.Mem. 20, Tulsa, OK, pp. 183^199.
Rossignol-Strick, M., 1964. Hystrichosphe'res du Quaternaireen Me¤diterrane¤e orientale, dans les se¤diments Ple¤istoce'nes etles boues marines actuelles. Rev. Micropale¤ontol. 7, 83^89.
Ryan, W., Pitman, W., 1999. Noah’s Flood: The New Scien-ti¢c Discoveries about the Event That Changed History.Simon and Schuster, New York, 318 pp.
Ryan, W.B.F., Pitmann, W.C., Major, C.O., Shimkus, K.,
Moskalenko, V., Jones, G.A., Dimitrov, P., Go«ru«r, N.,Sakinc, M., Yuce, H., 1997. An abrupt drowning of theBlack Sea shelf. Mar. Geol. 138, 119^126.
Schott, F., Visbeck, M., Send, U., 1994. Open ocean deepconvection, Mediterranean and Greenland seas. In: Mala-notte-Rizzole, P., Robinson, A.R. (Eds.), Ocean Processes inClimate Dynamics: Global and Mediterranean Examples.Kluwer, Dordrecht, pp. 203^225.
Sperling, M., Schmiedl, G., Hemleben, Ch., Meeis, K.C., Er-lenkeuser, H., Grootes, P.M., 2003. Black Sea impact on theformation of eastern Mediterranean sapropel S1? Evidencefrom the Marmara Sea. Palaeogeogr. Palaeoclimatol. Pa-laeoecol. 190, 9^21.
Stanev, E.V., Peneva, E.L., 2002. Regional sea level responseto global climatic change: Black Sea examples. Glob. Planet.Changes 32, 33^47.
Top, Z., Izdar, E., Ergun, M., Konuk, T., 1990. Evidence oftectonism from 3He and residence time of helium in theBlack Sea. EOS Trans. Am. Geophys. Union 71, 1020.
Turon, J.L., Londeix, L., 1988. Les assemblages de kystes dedino£agelle¤s en Me¤diterrane¤e occidentale (mer d’Alboran)mise en e¤vidence de l’e¤volution des pale¤oenvironnementsdepuis le dernier maximum glaciaire. Bull. Centr. Rech. Ex-plor.-Prod. Elf^Aquitaine 12, 313^344.
Traverse, A., 1974. Palynological investigation of two BlackSea cores. In: Degens, E.T., Ross, D.A. (Eds.), The BlackSea: Geology, Chemistry, and Biology. Am. Assoc. Pet.Geol. Mem. 20, Tulsa, OK, pp. 381^388.
Traverse, A., 1988. Paleopalynology. Unwin Hyman, Boston,600 pp.
Uº nlu«ata, U., Aubrey, D.G., Belberov, Z., Bologa, A., Ere-meev, V., Vinogradov, M., 1993. International program in-vestigates the Black Sea. EOS Trans. Am. Geophys. Union74, 401^412.
Wall, D., Dale, B., 1974. Dino£agellates in Late Quaternarydeep-water sediments of Black Sea. In: Degens, E.T., Ross,D.A. (Eds.), The Black Sea: Geology, Chemistry, and Biol-ogy. Am. Assoc. Pet. Geol. Mem. 20, Tulsa, OK, pp. 364^380.
Wall, D., Dale, B., Harada, K., 1973. Descriptions of newfossil dino£agellates from the Late Quaternary of the BlackSea. Micropaleontology 19, 18^31.
Williams, G.L., Lentin, J., Fensome, R., 1998. The Lentin andWilliams index of fossil dino£agellates, 1998 edition. Am.Assoc. Stratigr. Palynol. Found. Contrib. Ser. 34, 817 pp.
Wilson, I., 2001. Before the Flood. Orion Books, London, 336pp.
Zonneveld, K., 1996. Palaeoclimatic and palaeo-ecologicchanges in the Eastern Mediterranean and Arabian Sea re-gions during the last deglaciation: A palynological approachto land^sea correlation. University of Utrecht, LPP Contrib.ser. 3, 199 pp.
Zonneveld, K.A.F., Versteegh, G.J.M., de Lange, G.J., 2001.Palaeoproductivity and post-depositional aerobic organicmatter decay re£ected by dino£agellate cyst assemblages ofthe Eastern Mediterranean S1 sapropel. Mar. Geol. 172,181^195.
PALBO 2563 20-11-03 Cyaan Magenta Geel Zwart
P.J. Mudie et al. / Review of Palaeobotany and Palynology 128 (2004) 143^167 167
Related Documents











