Protein S-Nitrosylation in plants: Current progresses and challenges Jian Feng 1 , Lichao Chen 2,3 and Jianru Zuo 2,3 * 1. Sainsbury Laboratory, University of Cambridge, Cambridge CB2 1LR, UK 2. State Key Laboratory of Plant Genomics and National Center for Plant Gene Research, CAS Center for Excellence in Molecular Plant Sciences, Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China 3. The University of Chinese Academy of Sciences, Beijing 100049, China Abstract Nitric oxide (NO) is an important signaling molecule regulating diverse biological processes in all living organisms. A major physiological function of NO is executed via protein S-nitrosylation, a redox-based the past decade, significant progress has been made in functional characterization of S-nitrosylated proteins in plants. Emerging evidence indicates that protein S- nitrosylation is ubiquitously involved in the regulation of plant development and stress responses. Here we review current understanding on the regulatory mechanisms of protein S-nitrosylation in various biological processes in plants and highlight key challenges in this field. Edited by: Zhizhong Gong, China Agricultural University, China Received Dec. 25, 2018; Accepted Jan. 14, 2019; Online on Jan. 21, 2019 INTRODUCTION In 1774, Joseph Priestly published his book “Experiments and Observations on Different Kinds of Air” (Volume 1), in which he described the discovery of several airs, including ‘nitrous air’ (Priestly 1774), currently known as ‘nitric oxide’ (NO), which simply consists of one oxygen atom and one nitrogen atom. For a long time since its discovery, NO has been known as a colorless, relatively unstable gaseous molecule with a free radical that makes it relatively reactive and is thought as an air pollutant. In the 1980s, the importance of NO was recognized as a signaling molecule in mammals widely present in different tissues and organs with diverse physiological roles in the nervous system, immune system, cardiovascular system and endocrine system (Schmidt and Walter 1994; Stamler et al. 1997). Later studies identified NO as a highly conserved signaling molecule found in all living organisms. In higher plants, NO has been found to play important regulatory roles in almost all aspects of the life cycle as well as responses to biotic and abiotic stresses with diverse, yet not fully understood, mechanisms (Freschi 2013; Puyaubert and Baudouin 2014; Yu et al. 2014; Domingos et al. 2015; Trapet et al. 2015; Asgher et al. 2017; Fancy et al. 2017; Begara-Morales et al. 2018). Based on our current understanding, NO executes its physiological roles mainly through covalent post- translational modifications, including tyrosine nitration, metal nitrosylation, and S-nitrosylation. Tyrosine nitra- tion involves the addition of a nitro group at the ortho position of the phenolic hydroxyl group of tyrosine to produce 3-nitrotyrosine (Radi 2013). Peroxynitrite (OONO - ) and nitrogen dioxide (NO 2 ), two reactive © 2019 Institute of Botany, Chinese Academy of Sciences JIPB Journal of Integrative Plant Biology Invited Expert Review posttranslational modification by covalently adding a NO molecule to a reactive cysteine thiol of a target protein. S-nitrosylation is an evolutionarily conserved mechanism modulating multiple aspects of cellular signaling. During *Correspondence: J ianru Z uo ( [email protected]) doi: 10.1111/jipb.12780 December 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net FA: Free Access FA nitrogen species, generated by reactions of NO with superoxide radicals (O 2 .- ) and O 2 , respectively, are major NO donors to mediate tyrosine nitration (Liu et al. 1998 Radi 2013; Bartesaghi and Radi 2018). Tyrosine nitration High-Impact Article

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Protein S-Nitrosylation in plants: Currentprogresses and challengesJian Feng1, Lichao Chen2,3 and Jianru Zuo2,3*
1. Sainsbury Laboratory, University of Cambridge, Cambridge CB2 1LR, UK2. State Key Laboratory of Plant Genomics and National Center for Plant Gene Research, CAS Center for Excellence in Molecular Plant Sciences,Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China3. The University of Chinese Academy of Sciences, Beijing 100049, Chinadoi: 10.1111/jipb.12780
Abstract Nitric oxide (NO) is an important signalingmolecule regulating diverse biological processes in allliving organisms. A major physiological function of NOis executed via protein S-nitrosylation, a redox-based
the past decade, significant progress has been madein functional characterization of S-nitrosylated proteins
in plants. Emerging evidence indicates that protein S-nitrosylation is ubiquitously involved in the regulation ofplant development and stress responses. Here we reviewcurrent understanding on the regulatory mechanisms ofprotein S-nitrosylation in various biological processes inplants and highlight key challenges in this field.
Edited by: Zhizhong Gong, China Agricultural University, ChinaReceived Dec. 25, 2018; Accepted Jan. 14, 2019; Online on Jan. 21,2019
INTRODUCTION
In 1774, Joseph Priestly published his book “Experimentsand Observations on Different Kinds of Air” (Volume 1), inwhich he described the discovery of several airs,including ‘nitrous air’ (Priestly 1774), currently knownas ‘nitric oxide’ (NO), which simply consists of oneoxygen atom and one nitrogen atom. For a long timesince its discovery, NO has been known as a colorless,relatively unstable gaseous molecule with a free radicalthat makes it relatively reactive and is thought as an airpollutant. In the 1980s, the importance of NO wasrecognized as a signaling molecule in mammals widelypresent in different tissues and organs with diversephysiological roles in the nervous system, immunesystem, cardiovascular system and endocrine system(Schmidt and Walter 1994; Stamler et al. 1997). Later
studies identified NO as a highly conserved signalingmolecule found in all living organisms. In higher plants,
NO has been found to play important regulatory roles inalmost all aspects of the life cycle as well as responsesto biotic and abiotic stresses with diverse, yet not fullyunderstood, mechanisms (Freschi 2013; Puyaubert andBaudouin 2014; Yu et al. 2014; Domingos et al. 2015;Trapet et al. 2015; Asgher et al. 2017; Fancy et al. 2017;Begara-Morales et al. 2018).
Based on our current understanding, NO executesits physiological roles mainly through covalent post-translational modifications, including tyrosine nitration,metal nitrosylation, and S-nitrosylation. Tyrosine nitra-tion involves the addition of a nitro group at the orthoposition of the phenolic hydroxyl group of tyrosine toproduce 3-nitrotyrosine (Radi 2013). Peroxynitrite(OONO
-
) and nitrogen dioxide (NO2), two reactive
© 2019 Institute of Botany, Chinese Academy of Sciences
JIPB Journal of IntegrativePlant Biology
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX
Invited Ex
pert
Rev
iew
December 2019 | Volume 61 | Issue 12 | 1199-1216
*Correspondence:
posttranslational modification by covalently adding a NOmolecule to a reactive cysteine thiol of a target protein.S-nitrosylation is an evolutionarily conserved mechanismmodulating multiple aspects of cellular signaling. During
*Correspondence: J ianru Z uo ( [email protected])doi: 10.1111/jipb.12780
December 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
FA: Free Access
FA
nitrogen species, generated by reactions of NO withsuperoxide radicals (O2
.-) and O2, respectively, aremajorNO donors to mediate tyrosine nitration (Liu et al. 1998Radi 2013; Bartesaghi and Radi 2018). Tyrosine nitration
High-Im
pact
Article

has been used as a marker of oxidant burden in human
diseases (Ischiropoulos 2009). In plants, a number of
proteins modified by tyrosine nitration has been
identified in proteomic studies (Romero-Puertas et al.
2007; Chaki et al. 2011; Lozano-Juste et al. 2011; Tanou
et al. 2012; Begara-Morales et al. 2013) and several
tyrosine nitration-modified proteins have been func-
tionally characterized (Melo et al. 2011; Begara-Morales
et al. 2015; Castillo et al. 2015; Holzmeister et al. 2015;
Takahashi et al. 2017a, 2017b).
More attention has been given to S-nitrosylation, a
redox-based protein posttranslational modification by
covalently forming an S-linked NO group to the reactive
thiols of a cysteine residue. A cyclic GMP (cGMP)-
dependent NO signaling mechanism has been pro-
posed, of which NO enhances the activity of soluble
guanylate cyclase (sGC) to boost the intracellular level
of cGMP, eventually activating downstream Ca2þ-modulated signaling (Wendehenne et al. 2001; Ahern
et al. 2002; Besson-Bard et al. 2008). A key step of this
signaling pathway is NO-mediatedmetal nitrosylation of
sGC (Ahern et al. 2002; Besson-Bard et al. 2008), a
regulatory mechanism similar to that of protein
S-nitrosylation.
Similar to other posttranslational modifications,
S-nitrosylation regulates protein activities by various
mechanisms, including stability, biochemical activity,
conformation change, subcellular localization, and
protein–protein interaction (Hess et al. 2005; Astier
et al. 2011; Astier et al. 2012b; Lamotte et al. 2014). In
plants, a number of S-nitrosylated proteins involved in
various physiological processes have been functionally
characterized (Table 1). In this review, we will outline
the current progress in NO metabolism and functional
characterization of representative S-nitrosylated pro-
teins in plants.
METABOLISM OF NITRIC OXIDE
In mammalian cells, biosynthesis of NO is mainlydependent on nitric oxide synthase (NOS), whichcatalyzes the generation of NO by converting L-arginineto L-citrulline (Nathan and Xie 1994; Mayer and
Hemmens 1997; Wendehenne et al. 2001). Althoughthe production of NO in plants appears to be sensitive
to animal NOS inhibitors, structural and functional
analogues of NOS-like proteins have not been found inany land plants (Besson-Bard et al. 2008; Gas et al. 2009;
Frohlich and Durner 2011; Jeandroz et al. 2016; Astier
et al. 2018). Instead, currently available evidence
suggests that NO in land plants is produced from
nitrite via nonenzymatic or enzymatic pathways, of
which the latter is catalyzed by nitrate reductase (NR),
nitrite-NO reductase (Ni-NOR) and xanthine oxidore-
ductase (XOR) (Besson-Bard et al. 2008; Gupta et al.2011; Mur et al. 2013; Fancy et al. 2017).
In higher plants, NR is considered as a key enzyme of
nitrate assimilation (Wendehenne et al. 2004; Besson-
Bard et al. 2008). Cytosolic NR reduces nitrate (NO3-)
to nitrite (NO2-) in an NAD(P)H-dependent manner.
NR contains an N-terminal molybdenum-containing
domain, a b-type cytochrome domain and a C-terminal
flavin adenine dinucleotide (FAD) domain, each con-
nected by a hinge region (Figure 1A). All of these
domains function as redox centers to transfer electrons
from NAD(P)H to nitrate (Dean and Harper 1988;Campbell and Kinghorn 1990; Campbell 1996; Yamasakiand Sakihama 2000; Rockel et al. 2002). Evidenceobtained from both genetic and pharmacological
studies suggests that NR plays an important role inNO production (Desikan et al. 2002; Bright et al. 2006;Yamamoto-Katou et al. 2006; Kolbert et al. 2008;Seligman et al. 2008; Zhao et al. 2009;Wang et al. 2010).NR is capable of reducing NO2
- to NO and ONOO- (Deanand Harper 1988; Rockel et al. 2002). In Chlamydomonas,a different mechanism has been proposed that theamidoxime reducing component (renamed as nitricoxide-forming nitrite reductase, NOFNiR) catalyzes NOproduction from nitrite using NAD(P)H-NR as electrondonors. This dual NR-NOFNiR system catalyzes theproduction of NO from nitrite in the presence of arelatively high concentration of nitrate, which inhibitsthe activity of NR (Chamizo-Ampudia et al. 2016). Intobacco (Nicotiana tabacum L. cv. Samsun) roots, theplasma membrane-bound NR is associated with anitrite:NO oxidoreductase (Ni:NOR), which may reduceapoplastic nitrite and contribute to NO production(Stohr et al. 2001). Moreover, a peroxisomal enzymeXOR is also involved in the production of NO (Corpaset al. 2001; Harrison 2002; Corpas et al. 2008). Similar toNR, XOR is an oxidoreductase using molybdenum as a
2 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1207
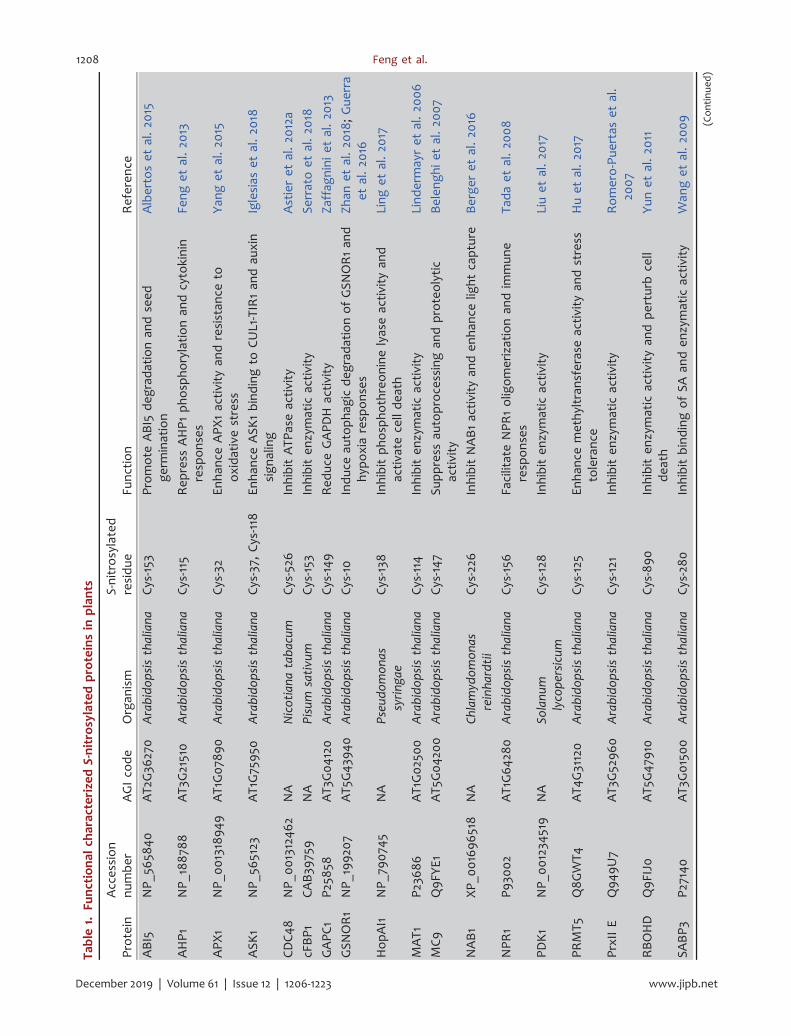
Table1.
Function
alcharacterizedS-nitrosylated
proteins
inplan
ts
Protein
Accession
numbe
rAGIco
deOrgan
ism
S-nitrosylated
residu
eFu
nction
Referen
ce
ABI5
NP_
5658
40AT2G36
270
Arabido
psisthaliana
Cys-153
Prom
oteABI5
degrad
ationan
dseed
germ
ination
Albertoset
al.20
15
AHP1
NP_
1887
88AT3G21510
Arabido
psisthaliana
Cys-115
Rep
ress
AHP1
phosph
orylationan
dcytokinin
respon
ses
Feng
etal.20
13
APX
1NP_
00131894
9AT1G07
890
Arabido
psisthaliana
Cys-32
Enha
nceAPX
1activity
andresistan
ceto
oxidativestress
Yang
etal.20
15
ASK
1NP_
565123
AT1G7595
0Arabido
psisthaliana
Cys-37,Cy
s-118
Enha
nceASK
1bind
ingto
CUL1-TIR1an
dau
xin
sign
aling
Iglesias
etal.20
18
CDC4
8NP_
00131246
2NA
Nicotiana
taba
cum
Cys-526
InhibitATP
aseactivity
Astieret
al.20
12a
cFBP1
CAB39
759
NA
Pisum
sativum
Cys-153
Inhibiten
zymatic
activity
Serratoet
al.20
18
GAPC
1P2
5858
AT3G04
120
Arabido
psisthaliana
Cys-149
Red
uceGAPD
Hactivity
Zaffag
nini
etal.20
13
GSN
OR1
NP_
1992
07AT5
G43
940
Arabido
psisthaliana
Cys-10
Indu
ceau
toph
agicde
grad
ationof
GSN
OR1an
dhy
poxiarespon
ses
Zhan
etal.20
18;Gue
rra
etal.20
16
Hop
AI1
NP_
7907
45NA
Pseudo
mon
assyring
aeCy
s-138
Inhibitph
osph
othreo
nine
lyaseactivity
and
activate
cellde
ath
Ling
etal.20
17
MAT1
P236
86AT1G02
500
Arabido
psisthaliana
Cys-114
Inhibiten
zymatic
activity
Lind
ermayret
al.20
06
MC9
Q9F
YE1
AT5
G04
200
Arabido
psisthaliana
Cys-147
Supp
ress
autoproc
essing
andproteo
lytic
activity
Belen
ghie
tal.20
07
NAB1
XP_0
0169
6518
NA
Chlamydom
onas
reinha
rdtii
Cys-226
InhibitNAB1activity
anden
hancelight
capture
Berge
ret
al.20
16
NPR
1P9
3002
AT1G64
280
Arabido
psisthaliana
Cys-156
FacilitateNPR
1oligom
erizationan
dim
mun
erespon
ses
Tada
etal.20
08
PDK1
NP_
001234
519
NA
Solanu
mlycope
rsicum
Cys-128
Inhibiten
zymatic
activity
Liuet
al.20
17
PRMT5
Q8G
WT4
AT4
G31120
Arabido
psisthaliana
Cys-125
Enha
ncemethy
ltransferase
activity
andstress
tolerance
Huet
al.20
17
PrxIIE
Q94
9U7
AT3G5296
0Arabido
psisthaliana
Cys-121
Inhibiten
zymatic
activity
Rom
ero-Pu
ertaset
al.
2007
RBOHD
Q9F
IJ0
AT5
G47
910
Arabido
psisthaliana
Cys-89
0Inhibiten
zymatic
activity
andpe
rturbcell
death
Yunet
al.20
11
SABP3
P27140
AT3G0150
0Arabido
psisthaliana
Cys-28
0Inhibitbind
ingof
SAan
den
zymatic
activity
Wan
get
al.20
09 (Con
tinu
ed)
Protein S-nitrosylation in plants 3
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1208
co-factor. Under anaerobic conditions, XOR catalyzesthe conversion of nitrite to NO, using NADH or xanthineas reducing agents (Godber et al. 2000a; Godber et al.2000b) (Figure 1B).
Under certain conditions, NO can be producedthrough non-enzymatic pathways (Figure 1B). In thealeurone layers of barley (Hordeum vulgare), non-enzymatic synthesis of NO is facilitated by reductionof apoplastic nitrite under acidic conditions (Bethkeet al. 2004a). In tobacco leaves, the mitochondrialelectron transport is proposed to contribute to thenitrite-dependent NO production, whereas other non-enzymatic sources of NO include reduction of nitrite toNOby carotenoids, ascorbic acid or polyamines (Cooneyet al. 1994; Crawford 2006; Tun et al. 2006). Currently,the mechanisms and the physiological significance ofnon-enzymatic production of NO remain largely unclear.
The homeostasis of NO is tightly controlled by itsbiosynthesis and turnover. Although the biosynthesisof NO is likely using different mechanisms in animaland plant cells, the turnover of bioactive NO speciesis controlled by a conserved mechanism. In allliving organisms, S-nitrosoglutathione (GSNO), derivedfrom the reaction of NO and glutathione (GSH), is amajor reservoir of bioactive NO species (Hess et al.2005; Gupta 2011), which is irreversibly degraded byS-nitrosoglutathione reductase (GSNOR). GSNORs, aclass of highly conserved NADH-dependent reductasespresented in all living organisms, catalyze the reduc-tion of GSNO to glutathione disulfide (GSSG) andammonia (NH3) in the presence of GSH (Liu et al. 2001;Sakamoto et al. 2002) (Figure 1C). Mutations in GSNORgenes cause severe defects in development and stressresponses in both animals and plants (Liu et al. 2004;Feechan et al. 2005; Que et al. 2005; Lee et al. 2008;Chen et al. 2009; Kwon et al. 2012), illustrating thecritical role of NO homeostasis.
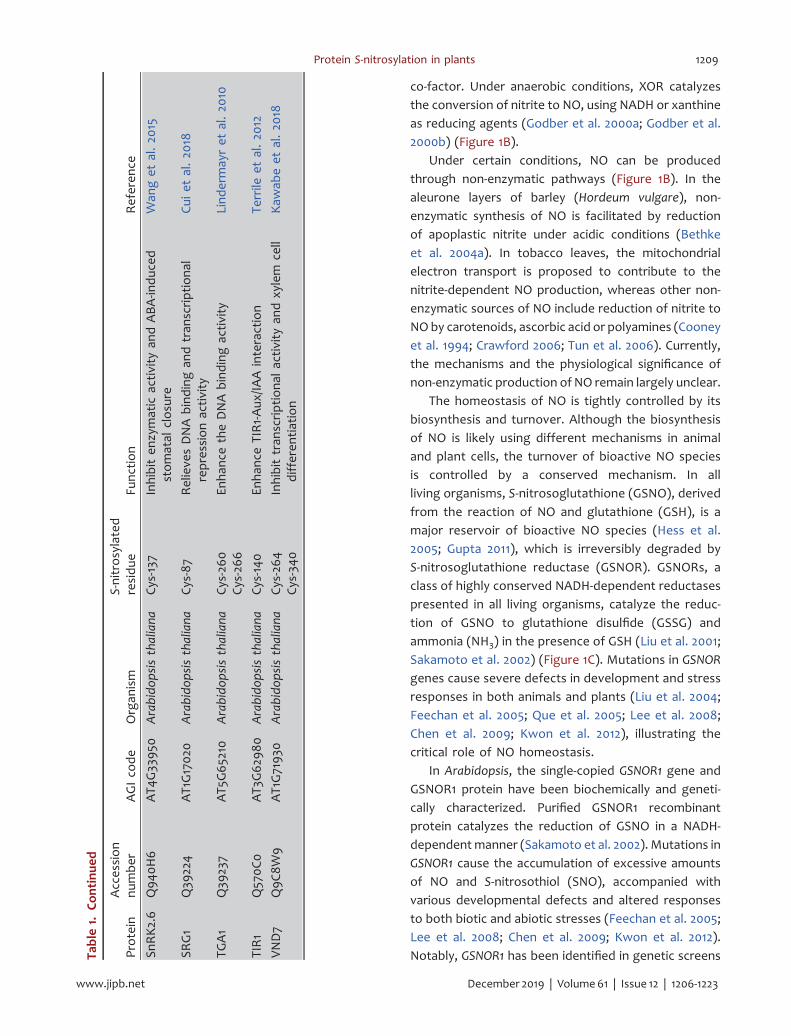
In Arabidopsis, the single-copied GSNOR1 gene andGSNOR1 protein have been biochemically and geneti-cally characterized. Purified GSNOR1 recombinantprotein catalyzes the reduction of GSNO in a NADH-dependentmanner (Sakamoto et al. 2002). Mutations inGSNOR1 cause the accumulation of excessive amountsof NO and S-nitrosothiol (SNO), accompanied withvarious developmental defects and altered responsesto both biotic and abiotic stresses (Feechan et al. 2005;Lee et al. 2008; Chen et al. 2009; Kwon et al. 2012).Notably, GSNOR1 has been identified in genetic screensTa
ble1.
Continue
d
Protein
Accession
numbe
rAGIco
deOrgan
ism
S-nitrosylated
residu
eFu
nction
Referen
ce
SnRK2.6
Q94
0H6
AT4
G3395
0Arabido
psisthaliana
Cys-137
Inhibiten
zymatic
activity
andABA-in
duced
stom
atal
closure
Wan
get
al.20
15
SRG1
Q39
224
AT1G1702
0Arabido
psisthaliana
Cys-87
RelievesDNAbind
ingan
dtran
scriptiona
lrepression
activity
Cuie
tal.20
18
TGA1
Q39
237
AT5
G65
210
Arabido
psisthaliana
Cys-26
0Cy
s-26
6En
hancetheDNAbind
ingactivity
Lind
ermayret
al.20
10
TIR1
Q570C
0AT3G62
980
Arabido
psisthaliana
Cys-140
Enha
nceTIR1-Aux
/IAAinteraction
Terrile
etal.20
12
VND7
Q9C
8W9
AT1G7193
0Arabido
psisthaliana
Cys-26
4Cy
s-34
0Inhibittran
scriptiona
lactivity
andxylem
cell
differen
tiation
Kaw
abeet
al.20
18
4 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1209
for thermotolerance-defective and oxidative-stress-resistant mutants, designated as SENSITIVE TO HOTTEMPERATURE5 (HOT5) and PARAQUAT RESISTANT2(PAR2), respectively (Lee et al. 2008; Chen et al.2009), implying a tight link of NO with abiotic stressresponses (see also below).
As mentioned above, the nitrogen assimilationenzyme NR reduces NO2
- to NO, hence playing a criticalrole in NO production in higher plants. SeveralArabidopsis mutants have been characterized to showan increased NO or SNO level, including gsnor1/hot5/par2, nox1 (for NO overproducer1) (He et al. 2004), andnia1 nia2, of which NIA1 and NIA2 are two closely relatedgenes encoding NRs (Wilkinson and Crawford 1991).Analysis of these mutants revealed that nitrogenassimilation is regulated by SNO via suppressing nitratetransport and reduction (Frungillo et al. 2014). More-over, GSNOR1 itself is S-nitrosylated (Frungillo et al.2014; Guerra et al. 2016; Zhan et al. 2018), correlatedto the (S)NO-induced inhibition of the enzymaticactivity, suggesting that this negative regulatorymechanism plays an important role for the homeostasisof S-nitrosothiols via nitrogen assimilation (Frungilloet al. 2014). While multiple cysteine residues of GSNOR1are found to be S-nitrosylated, none of the singlesubstitutional mutations in these cysteine residuesshows an apparent effect on the enzymatic activity(Guerra et al. 2016; Zhan et al. 2018). A triple mutation inthree highly conserved cysteine residues abolishes theresponsiveness to NO in the inhibition of the GSNOR1enzymatic activity in an in vitro assay (Guerra et al.2016). Moreover, NO induces autophagic degradationof GSNOR1 (Zhan et al. 2018). Therefore, both theenzymatic activity and stability of GSNOR1 are nega-tively regulated by NO, indicating that a positivefeedback regulatory loop modulates the accumulationNO (Figure 1C), which is likely a key during earlysignaling for the burst of NO in response to variousstimuli.
In addition to the GSNOR-mediated NO turnover,other NO scavenging mechanisms have also beendescribed. In a genetic screen for Arabidopsis NO-insensitive mutants, continuous no-unstressed1 wasfound to be allelic to altered meristem program1 (Liuet al. 2013), a mutant showing a pleiotropic phenotypewith an increased cytokinin level (Helliwell et al. 2001).Subsequent analyses found that ONOO-, a reactive NOintermediate NO, directly interacts with the bioactive
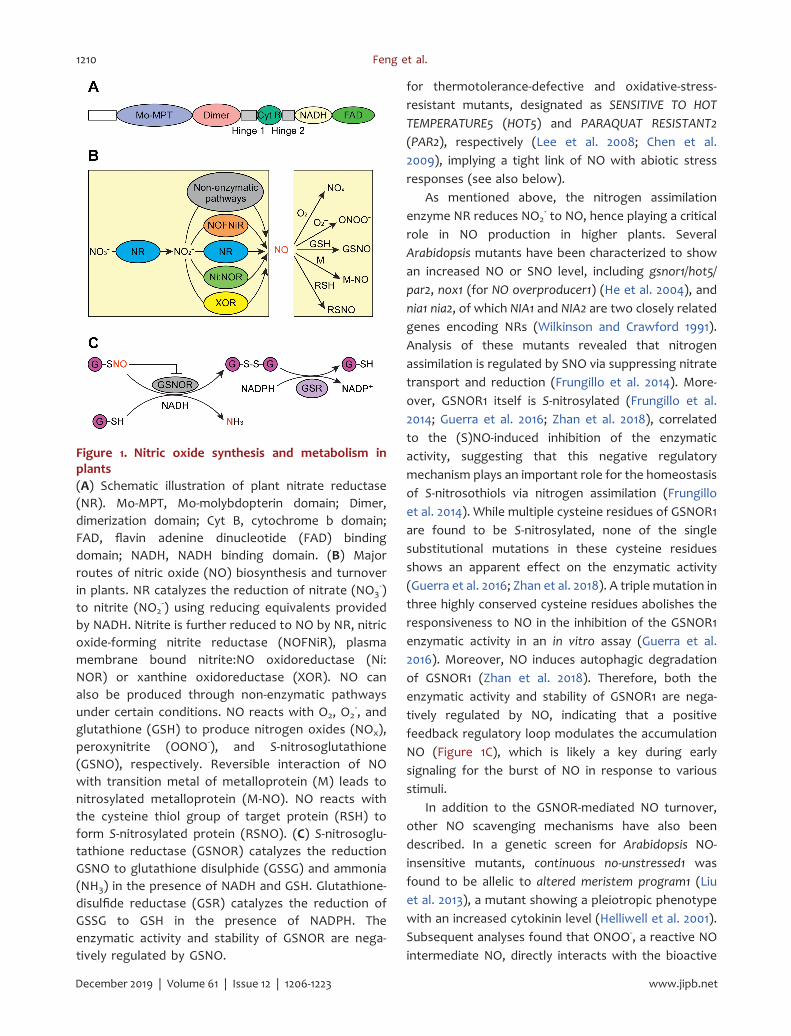
Figure 1. Nitric oxide synthesis and metabolism inplants(A) Schematic illustration of plant nitrate reductase(NR). Mo-MPT, Mo-molybdopterin domain; Dimer,dimerization domain; Cyt B, cytochrome b domain;FAD, flavin adenine dinucleotide (FAD) bindingdomain; NADH, NADH binding domain. (B) Majorroutes of nitric oxide (NO) biosynthesis and turnoverin plants. NR catalyzes the reduction of nitrate (NO3
-)to nitrite (NO2
-) using reducing equivalents providedby NADH. Nitrite is further reduced to NO by NR, nitricoxide-forming nitrite reductase (NOFNiR), plasmamembrane bound nitrite:NO oxidoreductase (Ni:NOR) or xanthine oxidoreductase (XOR). NO canalso be produced through non-enzymatic pathwaysunder certain conditions. NO reacts with O2, O2
-, andglutathione (GSH) to produce nitrogen oxides (NOx),peroxynitrite (OONO-), and S-nitrosoglutathione(GSNO), respectively. Reversible interaction of NOwith transition metal of metalloprotein (M) leads tonitrosylated metalloprotein (M-NO). NO reacts withthe cysteine thiol group of target protein (RSH) toform S-nitrosylated protein (RSNO). (C) S-nitrosoglu-tathione reductase (GSNOR) catalyzes the reductionGSNO to glutathione disulphide (GSSG) and ammonia(NH3) in the presence of NADH and GSH. Glutathione-disulfide reductase (GSR) catalyzes the reduction ofGSSG to GSH in the presence of NADPH. Theenzymatic activity and stability of GSNOR are nega-tively regulated by GSNO.
Protein S-nitrosylation in plants 5
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1210

cytokinin compound zeatin, resulting in the reduced NOlevel, thereby modulating NO homeostasis (Liu et al.2013). In Chlamydomonas reinhardtii, two truncatedhemoglobins, THB1 and THB2, function with NR toconvert NO into nitrate (Sanz-Luque et al. 2015).Moreover, the NO-derived compounds react withunsaturated fatty acids to produce nitro-fatty acids(NO2-FAs), which act as NO donors in both animals andplants (Villacorta et al. 2007; Kelley et al. 2008; Bakeret al. 2009; Sanchez-Calvo et al. 2013; Mata-Perez et al.2016a). The presence of NO2-FAs [in particular, nitro-oleic acid (NO2-OA), nitro-linoleic acids (NO2-LA) andnitro-linolenic acid (NO2-Ln)] has been confirmed inArabidopsis, pea and rice by mass spectrometry (Mata-Perez et al. 2016b). NO2-Ln is involved in regulatingresponses to abiotic stresses and its level is induced bysalt, heavy metal, low temperature and mechanicalwounding, suggesting an important role of NO2-FAs inthe adaption of abiotic stresses in plants (Mata-Perezet al. 2016b).
OVERVIEW OF S-NITROSYLATION,TRANSNITROSYLATION ANDDENITROSYLATION
A major physiological activity of NO is mediated byS-nitrosylation, a redox-based posttranslational modifi-cation that covalently links a NO group to the reactiveCys thiol (-SH) to form S-nitrosothiol (-SNO) (Figure 2A).This highly conserved posttranslational mechanismplays crucial roles in diverse physiological and patho-logical processes by modulating protein activities.
S-nitrosylation is a dynamic and reversible process,involving nitrosylation, denitrosylation and transnitro-sylation (Hess et al. 2005; Benhar et al. 2009; Stomberskiet al. 2019).
S-nitrosylation has been thought mainly as a nonen-
zymatic process. The formation of an S-nitrosothiol can
be directly mediated by NO, or indirectly mediated by
various higher nitrogen oxides (NOx), metal-NO inter-
mediates, SNOs or ONOO-(Wang et al. 2006). The
addition of NO donors (usually GSNO) to purified
recombinant proteins causes the S-nitrosylation of the
target in the absence of any co-factors or enzyme-like
proteins, a process similar to that of auto-phosphor-
ylation. More critically, yet no highly specific enzymes
catalyzing S-nitrosylation have been identified (but, see
below, for transnitrosylation). This possible nonenzy-
matic-reaction feature is unique to S-nitrosylation, which
is possibly attributed to the fact that S-nitrosylation (and
otherNO-mediatedmodifications) is largelydrivenby the
redox potentials between a NO donor molecule and a
modified target (Stamler et al. 2001; Hess et al. 2005;
Benhar et al. 2009), whereas most, if not all, other
posttranslational modifications, are energy-consuming
processes. Possiblybecauseof itsnonenzymatic-reaction
nature, at least in part, the redox-based S-nitrosylation
has been recently proposed to act predominantly as
transient intermediates prior to the formation of
disulfide bonds (Wolhuter et al. 2018).A key question in this newly emerged research field
is the specificity or selectivity of the nonenzymatic-reaction modification, i.e., how to select and determinea modified target protein. Currently available evidence
Figure 2. Biochemistry of protein S-nitrosylation, denitrosylation and transnitrosylation(A) Schematic representation of S-nitrosylation and denitrosylation. S-nitrosylation is the covalent incorporation ofa NO group to the thiol group of a cysteine residue to form S-nitrosothiol (SNO). Thioredoxin (Trx) denitrosylatesS-nitrosylated proteins through its dithiol moiety, forming a reduced protein thiol (-SH) and oxidized Trx; oxidizedTrx is reduced by NADPH-dependent Trx reductase (NTR). (B) Schematic representation of transnitrosylation. Atransnitrosylase carrying an SNO group transfers the NO moiety to a target protein.
6 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1211

suggests that the specificity of protein S-nitrosylation ismainly determined by the structure of a target and thelocal NO concentration. Several motifs have beenproposed by analysis of known S-nitrosylated proteinsand peptides, largely identified by the proteomicapproach in various organisms (Lindermayr et al.2005; Hao et al. 2006; Romero-Puertas et al. 2008;Palmieri et al. 2010; Fares et al. 2011; Puyaubert et al.2014; Chen et al. 2015; Hu et al. 2015; Ib�a~nez-Vea et al.2018). One of the commonly known motifs is (H/K/R)CX(D/E) (X as any amino acid residue) in mammals andcharacterized as a modified cysteine residue flanking byacidic and/or basic amino acid residues in a hydrophobicmicroenvironment (Hess et al. 2005; Hao et al. 2006;Seth and Stamler 2011). However, several motifs thatcontain acidic, but not basic, amino acid residuesflanking the S-nitrosylated cysteine residues have beenproposed, based on a large-scale proteomic study inArabidopsis (Hu et al. 2015). It remains unclear whetherthese variations are caused by differences betweenanimal and plant models or other unknown reasons.
The second factor for the determination ofS-nitrosylation is the local NO concentration near themodified target, and a higher local NO concentration isconsidered to induce S-nitrosylation of a target (Hesset al. 2005; Iwakiri et al. 2006; Benhar et al. 2009). Inanimal cells, the S-nitrosylation of a target protein istemporally and spatially linked to the NOS activity.Under certain circumstances, a target protein physicallyinteracts with NOS, which directly transfers the de novosynthesized NO group to the modified cysteine residue(Kim et al. 2005; Benhar et al. 2009; Martinez-Ruiz et al.2013; Jia et al. 2014). Similar selective mechanisms havenot been discovered in plants.
Although S-nitrosylation has long considered as anonenzymatic reaction, recent studies suggest thepresenceof transnitrosylation, inwhich a transitionmetal(e.g., Fe2þ, Cu2þ) complex or an S-nitrosylated proteinmediates the transfer of its bearingNOmoiety to anotherprotein and nitrosylates the latter (Figure 2B). During thisprocess, a nitroso group is transferred from a transitionmetal nitrosyl to a Cys residue or between two Cysresidues, whereas the protein that carries and transfersthe NO group to its target is termed as a transnitrosylase(Anand and Stamler 2012; Stomberski et al. 2019). Severaltransnitrosylases have been identified and functionallycharacterized in bacterial and mammalian cells, includinghemoglobins, cytoglobin, neuroglobin, ceruloplasmin,
cytochrome c, cyclin-dependent kinase 5 (CDK5), caspase3, thioredoxin, and glycerol-3-phosphatase-dehydrase(GAPDH) (Anand and Stamler 2012). Two transnitrosylasecomplexes have been recently characterized. In humancells, a heterotrimeric S-nitrosylase complex consists ofthree subunits, inducible NO synthase (iNOS), S100A8,and transnitrosylase S100A9. During the transnitrosyla-tion, S100A9 is first S-nitrosylated by NO released fromiNOS and the S-nitrosylated S100A9 subsequently trans-nitrosylates the substrates containing a conservedmotif that is coordinately recognized by S100A8 andS100A9 (Jia et al. 2014). More recently, a multiplexenzymatic S-nitrosylation machinery was identified inE. coli, which includes NO synthase, SNO-proteins (SNOsynthase), and transnitrosylases to regulate a subset ofphysiological activities under anaerobic conditions (Sethet al. 2018). In both cases, the transnitrosylase complexescatalyze both de novo S-nitrosylation and transnitrosyla-tion. In general, all transnitrosylases characterized thusfar show distinctive structural features and no commonor conserved domains/motifs specific to this class ofproteins have been identified, a case different fromthat for the enzymes that catalyze other types ofposttranslational modifications, such as phosphorylation.It remains largely elusive for the mechanisms of selectiveS-nitrosylation and transnitrosylation. Nevertheless,transnitrosylation is an important enzymatic mechanismto determine the specificity of S-nitrosylation. To date,no transnitrosylase has been found in plants.
Similar to other posttranslational modifications, thedecoration of cysteine residues with NO is a reversibleprocess, of which the removal of NO from the modifiedcysteine residue of a protein is termed as denitrosyla-tion. Denitrosylation is mediated by the thioredoxin(Trx) system consisting of Trx and Trx reductase (TrxR).Trx denitrosylates SNO proteins through its dithiolmoiety, forming a reduced protein thiol and oxidizedTrx. The oxidized Trx in turn is reduced by TrxRand NADPH (Benhar et al. 2009) (Figure 2A). InArabidopsis, TRXh3 and TRXh5 selectively denitrosylatethe S-nitrosylated proteins to regulate plant immunity(Kneeshaw et al. 2014) (see also below).
During the past decade, a number of S-nitrosylatedproteins have been functionally characterized in plants,mostly in Arabidopsis (Table 1). Progresses on thestudies of the representative members of theseproteins involved in various physiological or signalingprocesses are summarized below.
Protein S-nitrosylation in plants 7
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1212

GROWTH AND DEVELOPMENT
The important role of NO in plant growth anddevelopment has been well documented. NO is widelyinvolved in regulating multiple aspects of development,including seed germination, root development, flower-ing time, pollen tube guidance, and leaf senescence(Besson-Bard et al. 2008; Astier et al. 2011; Mur et al.2013; Yu et al. 2014). In Arabidopsis, loss-of-functionmutations in the GSNOR1 gene cause severe develop-mental defects, including dramatically altered root andshoot architecture, reduced hypocotyl elongation, andreduced fertility (Lee et al. 2008; Chen et al. 2009; Kwonet al. 2012). These defects are closely associated withthe impaired auxin signaling and polar auxin transport ingsnor1, correlating with reduced levels of PIN proteins(Shi et al. 2015; Ni et al. 2017).
VASCULAR-RELATED NAC-DOMAIN (VND), a smallfamily of NAM/ATAF/CUC (NAC) transcription factors,are important regulators of xylem vessel differentia-tion. Overexpression of VND6 and VND7 induces trans-differentiation into xylem vessels, and this phenotype issuppressed by a gsnor1 loss-of-function mutation, whichin turn affects xylem vessel development. Correlated tothese observations, Cys-264 and Cys-320 of VND7 areS-nitrosylated analyzed in an in vitro biotin-switchassay and these two residues are directly related tothe transactivation activity of VND7, implying thatS-nitrosylation plays an importance role in modulatingxylem vessel cell differentiation (Kawabe et al. 2018). Anumber of VND6/7-regulated genes have been identi-fied, including METACASPASE9 (MC9), which is specifi-cally expressed in developing xylem and plays animportant role in xylem cell death (Bollhoner et al.2013). S-nitrosylation of MC9 at Cys-147 suppresses theMC9 autoprocessing and proteolytic activity, therebydirectly regulating the activity of MC9 (Belenghi et al.2007). Together, these findings suggest a tight linkbetween NO signaling and xylem development.
PHYTOHORMONE SIGNALING
S-nitrosylation has been found as an important mecha-nism to regulate signaling of phytohormones, includingauxin and cytokinin. Auxin signaling is activated byproteasomal degradation of the transcriptional repress-ors Aux/IAAs via the auxin-dependent interaction withreceptors TIR1/AFBs, leading to the alterations of
downstream auxin-responsive genes. While auxin
induces the accumulation of NO, the combined
treatment with sodium nitroprusside (SNP), a NO
donor, and auxin enhances the degradation of AUX/
IAA proteins and the expression of auxin-responsive
genes (Terrile et al. 2012). Further studies indicate that
the S-nitrosylation of the auxin receptor TIR1 at Cys-140
and Arabidopsis SKP1-like (ASK1) protein at Cys-37 and
Cys-118 enhances the formation of the SCFTIR1/AFB-Aux/
IAAs complex, thereby facilitating proteasomal degra-
dation of Aux/IAA repressors (Iglesias et al. 2018).
Cytokinin signaling is mediated by a two-component
system-based multiple-step phosphorelay involved in
sequential transferring of phosphoryl groups from the
receptors to the downstream components histidine
phosphotransfer proteins (AHPs) and response regula-
tors (ARRs). Of those components, AHP1 is S-nitrosylated
at Cys-115, which represses its phosphorylation and
subsequent transferring of the phosphoryl group to
ARR1. The NO-inhibited phosphorylation of AHP1 and
ARR1 causes a compromised cytokinin response, indicat-
ing that redox signaling and cytokinin signaling coordi-
nate plant growth and development (Feng et al. 2013).
Notably, ONOO- and zeatin directly interact, which in
principle causes the reduction of both bioactive NO and
cytokinin species (Liu et al. 2013), further supporting a
negative regulatory role of NOon the cytokinin pathway.
The observation that NO-mediated S-nitrosylation posi-
tively and negatively regulates auxin signaling and
cytokinin signaling, respectively, is consistent with the
notion that these two phytohormones are considered to
play agonistic roles in regulating plant growth and
development.
IMMUNE RESPONSES
Plant immune responses are often associated withredox status changes and pathogen infection triggers arapid burst of NO and reactive oxygen species (ROS).The regulatory roles of NO on plant immunity have beencomprehensively reviewed and, in particular, theinteractions between ROS and NO signaling havebeen long thought of as an important means toregulate stress responses (Delledonne et al. 1998;Delledonne et al. 2001; Wendehenne et al. 2004;Zaninotto et al. 2006; Besson-Bard et al. 2008; Leitneret al. 2009; Wang et al. 2013; Wendehenne et al. 2014;
8 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1213

Yu et al. 2014; Qi et al. 2018). The Arabidopsis gsnor1mutants and the tomato GSNOR-silenced plantsshow altered immune responses, indicating the directinvolvement of NO in response to pathogens (Feechanet al. 2005; Liu et al. 2017). However, a GSNOR1transgene rescues the SNO-dependent phenotype ofpar2 (an allele of gsnor1), but not the nox1 mutantphenotype, whereas a gsnor1-3 nox1-1 double mutantshows an additive phenotype, indicating that GSNO andNO may have distinctive molecular targets duringimmune responses and development (Yun et al.2016). It remains unknown how plants distinguish thesetwo types of signals.
The burst of ROS is a key event during the onsetof immune responses. However, a higher intracellularlevel of ROS is also toxic to cells. To minimize its toxiceffects, the intracellular ROS level is tightly controlledby various mechanisms, of which ROS-scavengingenzymes, including ascorbate peroxidase (APX), cata-lase, glutathione peroxidase, peroxidase and superox-ide dismutase, play a dominant role in regulating thehomeostasis of ROS (Mittler et al. 2004; Mittler et al.2011). The chloroplast-localized peroxiredoxin II E (PrxIIE), a member of the peroxiredoxin family, is aperoxidase that efficiently removes H2O2. PrxII E isS-nitrosylated at Cys-121, leading to the inhibition ofboth the peroxidase and ONOO- reductase activities.During pathogen infection, S-nitrosylation also inhibitsthe ONOO- detoxification activity of PrxII E, resulting inan increase of ONOO--mediated tyrosine nitration andlipid peroxidation (Romero-Puertas et al. 2007). Nota-bly, cytosolic PrxII B and mitochondria localized PrxII Fare also found to be S-nitrosylated in Arabidopsis cellculture extracts treated with GSNO in a proteomicstudy, suggesting that this family of proteins issubjected to the regulation by NO (Lindermayr et al.2005).
When challenged by pathogens, gsnor1-3 and nox1,two mutants accumulating excessive amount ofintracellular NO, show increased cell death but reducedROS accumulation. This phenotype is directly associatedwith NO-regulated activity of the NADPH oxidaseRBOHD. The S-nitrosylation of RBOHD at Cys-890inhibits its enzyme activity, resulting in reduced ROSbiosynthesis. Conversely, the non-nitrosylatable mutantRBOHDC890A shows the increased ROS production andcell death (Yun et al. 2011). In the ROS-scavengingsystem, the S-nitrosylation of APX1 at Cys-32 positively
regulates its enzymatic activity to reduce H2O2 into H2O,resulting in enhanced resistance to oxidative stress andcompromised immune signaling (Yang et al. 2015).These observations indicate that the biosynthesisand turnover of ROS are coordinated by NO viaS-nitrosylation.
The mitogen-activated protein kinase (MAPK) cas-cades play central roles in plant immune signalingagainst pathogens. Multiple effectors have beenidentified to target MAPK cascades, including HopAI1that dephosphorylates MPK3 and MPK6 by thephosphothreonine lyase activity to suppress immunesignaling (Zhang et al. 2007). HopAI1 is S-nitrosylated atCys-138 during the onset of the hypersensitive responseto inhibit its phosphothreonine lyase activity, whichrelieves the suppressing effect on MAPK signaling toactivate immune responses (Ling et al. 2017). TheS-nitrosylation of tobacco CDC48 at Cys-526 and tomatoPDK1 at Cys-128 has also been shown to be involved inregulating defense responses and cell death (Astieret al. 2012a; Liu et al. 2017; Rosnoblet et al. 2017).
Salicylic acid (SA) is an important signaling moleculeregulating plant immune responses and the accumula-tion of SA is rapidly induced by pathogen infection. Ingsnor1-3, SA-induced gene expression is compromised,correlated to the reduced SA level, suggestive of acritical role of S-nitrosylation in SA biosynthesis andsignaling (Feechan et al. 2005). In the SA signalingpathway, a number of proteins have been shown to beregulated by S-nitrosylation. The S-nitrosylation of SA-binding protein3 at Cys-280 inhibits its binding affinity toSA and its carbonic anhydrase activity, of which thelatter is required for the pathogen resistance (Wanget al. 2009). NON-EXPRESSOR OF PATHOGENESIS-RELATED 1 (NPR1) is a master regulator of SA-induceddefense responses, which, together with its paralogsNPR3 and NPR4, function as the SA receptors. It hasbeen reported that NPR1 is localized in the cytoplasm asan oligomer connected by intermolecular disulphidebonds in the absence of pathogen challenge. Uponpathogen attack, the NPR1 oligomer is reduced tomonomer in response to the increased SA level and thealtered cellular redox status and subsequently trans-located into the nucleus to activate the expression ofdownstream genes (Mou et al. 2003). During theprocess, S-nitrosylation at Cys-156 is reported tofacilitate NPR1 oligomer assembly and the substitutionof Cys-156 with Ser results in constitutive nuclear
Protein S-nitrosylation in plants 9
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1214

localization of NPR1 monomer and enhanced resis-
tance (Tada et al. 2008). The S-nitrosylated NPR1 and
other S-nitrosylated proteins are selectively denitrosy-
lated by the oxidoreductase thioredoxin TRX-h5,
thereby resetting SA signaling. As a consequence,
overexpression of TRX-h5 selectively rescues the
immune deficiencies in nox1, but not in gsnor1, through
selective protein denitrosylation (Kneeshaw et al.
2014).
Downstream of the SA receptors, two important
components have been found being regulated by
S-nitrosylation. Upon translocating into the nucleus,
NPR1 and NPR3/4 interact with and positively or
negatively regulate a group of transcription factors
TGAs. The S-nitrosylation and S-glutathionylation of
TGA1 at Cys-260 and Cys-266, respectively, inhibit
intramolecular disulfide bond formation between these
two cysteine residues, leading to the enhanced DNA-
binding activity of TGA1 (Lindermayr et al. 2010). A
recent study shows that SRG1, a zinc finger transcription
factor, interacts with the general transcriptional
corepressor TOPLESS to repress the expression of
target genes. The S-nitrosylation of SRG1 at Cys-87
decreases its DNA-binding affinity and transcriptional
repression activity, thereby allowing a higher expres-
sion of downstream target genes (Cui et al. 2018).
In both the TGA1 and SRG1 cases, S-nitrosylation acts
to positively and negatively regulate the activity of
a transcriptional activator and a transcriptional repres-
sor, respectively, which collectively boosts immune
responses, illustrating the importance of NO-mediated
S-nitrosylation in SA signaling.
ABIOTIC STRESS RESPONSES
Similar to that of immune responses, when chal-lenged by various abiotic stresses, plants rapidlyrespond by the burst of ROS and NO, which playcritical roles in modulating growth and developmentunder adverse environmental conditions. To date, NO-mediated S-nitrosylation has been found to be largelyinvolved in regulation of biotic and abiotic stressresponses. As discussed above, the homeostasis ofROS is regulated by the S-nitrosylation of RBOHD,PrxII E and APX1, which modulates the biosynthesis orturnover of ROS (Romero-Puertas et al. 2007; Yunet al. 2011; Yang et al. 2015).
The homeostasis of NO is regulated by GSNOR,a master regulator of NO signaling, whose enzymaticactivity and stability are in turn regulated by S-nitrosylation (Guerra et al. 2016; Zhan et al. 2018).Among the several identified S-nitrosylated cysteineresidues in ArabidopsisGSNOR1 (Guerra et al. 2016; Zhanet al. 2018), the regulatory role of Cys-10 has beenstudied in detail (Zhan et al. 2018). Analysis of the crystalstructure of GSNOR1 shows that Cys-10 is located in ab-sheet covering an ATG8-interacting motif (AIM)(Guerra et al. 2016; Zhan et al. 2018). The S-nitrosylationof Cys-10 causes local conformational changes, render-ing the AIM accessible by ATG8 and subsequently beingrecruited into autophagosome for degradation in anATG8-dependent manner. Whereas NO induces auto-phagic degradation of GSNOR1, the substitution ofCys-10 with Ser abolishes the responsiveness to NO andtherefore stabilizes the GSNOR1C10S mutant protein. AGSNOR1C10S transgene rescues all abnormalities of thegsnor1-3 mutant, but with an impaired response to NOunder low oxygen conditions during seed germination,suggesting that S-nitrosylation-mediated autophagicdegradation of GSNOR1 is involved in hypoxia responses(Zhan et al. 2018).
S-methionine adenosyltransferase (MAT) catalyzesthe biosynthesis of S-adenosylmethionine, a methyldonor for thebiosynthesis of ethylene.TheS-nitrosylationof MAT1 at Cys-114 results in reversible inhibition of itsenzymatic activity. Cys-114 is absent in MAT2 and MAT3,which do not show an inhibitory effect of NO on theirenzymatic activities (Lindermayr et al. 2006). Because ofpossible functional redundancy, it remains unclearwhether or not a mutation in Cys-114 will affect theintracellular concentration of S-adenosylmethionine.Nevertheless, as NO regulates ethylene production, thisstudy highlights a linkbetweenethylene andNOsignaling(Lindermayr et al. 2006).
Abscisic acid (ABA) plays important roles in regulat-ing plant development and stress responses. Uponbinding by ABA, the ABA receptors interact with andsuppress the activity of protein phosphatase 2C,resulting in the activation of the SnRK2-type proteinkinases, which subsequently phosphorylate and acti-vate downstream components. NO inhibits the kinaseactivity of SnRK2.6/OST1 by the S-nitrosylation at Cys-137that is adjacent to the kinase catalytic site, therebynegatively regulating ABA signaling (Wang et al. 2015).Interestingly, ABA enhances the S-nitrosylation of
10 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1215

SnRK2.6/OST1, correlated to the observation that ABAinduces the burst of NO, suggesting that ABA and NOform a negative regulatory loop to modulate ABAsignaling. Consistently, an increased S-nitrosylation ofSnRK2.6/OST1 has also been found in gsnor1 thatshows the impaired ABA-induced stomatal closure.The S-nitrosylated cysteine residue is highly conservedin land plants, mammals and several yeast species,suggesting that S-nitrosylation-mediated inhibition mayrepresent an evolutionarily conserved mechanism forprotein kinase regulation (Wang et al. 2015).
Seed germination and subsequent seedling establish-ment are critical processes regulated by ABA and NO. Ingeneral, whereas ABA inhibits, NO promotes, seedgermination and seedling establishment. The promotiverole of NO on seed germination have been reported in anumber of species (Beligni and Lamattina 2000; Bethkeet al. 2004b; Lozano-Juste and Leon2010). InArabidopsis,the ABI5 transcription factor is a key regulator ofseed germination and seedling establishment, and theexpression of ABI5 is directly regulated by the ERF VIItranscription factors (Gibbs et al. 2014). The increasedNOlevel is sensed by ERF VII, which are then targeted forproteasomal degradation via the N-end rule pathway,leading to the reduced expression of ABI5, therebypromoting seed germination (Gibbs et al. 2014). Thestability of ABI5 is in turn regulated by NO throughS-nitrosylation. S-nitrosylation of ABI5 at Cys-153 pro-motes its proteasomal degradation, resulting in theenhancement of seed germination and seedling estab-lishment (Albertos et al. 2015). Therefore, boththe expression of ABI5 and the stability of ABI5 proteinare negatively regulated by NO, thus revealing themolecular basis of NO-promoted seed germination.In both cases as highlighted above, NO-mediatedS-nitrosylationof SnRK2.6/OST1 andABI5plays a negativeregulatory role in ABA signaling, thus enhancing thestress resistance (Albertos et al. 2015; Wang et al. 2015).
Crosstalks between different types of proteinposttranslational modifications play an importantrole in regulating cellular signaling. A recent studyrevealed that S-nitrosylationmodulates protein argininemethylation during stress responses (Hu et al. 2017).Protein arginine methylation is catalyzed by a group ofhighly conserved protein arginine methyltransferases(PRMTs), of which PRMT5 has been well characterizedin higher eukaryotes. Mutations in the ArabidopsisPRMT5 gene causes a pleiotropic phenotype, including
hypersensitivity to salt stress, associated with reduced
arginine methylation of small nuclear ribonucleoprotein
Sm-like4 and altered pre-mRNA splicing of a subset
of stress-related genes (Zhang et al. 2011). PRMT5 is
S-nitrosylated at Cys-125, leading to the increased
methyltransferase activity. A PRMT5C125S transgene,
encoding a non-nitrosylatable mutant protein, fully
rescues the developmental defects of prmt5 under
normal growth conditions. However, the PRMT5C125S
transgenic plants are hypersensitive to NaCl and ABA,
but insensitive to NO that enhances resistance to stress,
indicating that S-nitrosylation at Cys-125 is essential for
the PRMT5-regulated stress responses. Moreover, salt
stress-induced arginine symmetric demethylation is also
abolished in PRMT5C125S transgenic plants, correlated to
aberrant pre-mRNA splicing of stress-related genes (Hu
et al. 2017). This study illustrates the importance of the
crosstalk between S-nitrosylation and arginine methyl-
ation in modulating stress responses.
CHLOROPHYLL METABOLISM ANDPHOTOSYNTHESIS
Chlorophyll metabolism and photosynthesis are largely
oxidation-reduction reaction. Emerging evidence sug-
gests that these two processes are extensively
regulated by the redox-based S-nitrosylation. In a site-
specific nitrosoproteomic studies, a large number of
proteins related to chlorophyll metabolism and photo-
synthesis have been identified as S-nitrosylated proteins
in Arabidopsis (Hu et al. 2015). Nearly one third of
proteins annotated as related to chlorophyll metabo-
lism were identified as S-nitrosylated proteins in the
nitrosoproteomic analysis and the S-nitrosylation of
several of these proteins has been further confirmed by
the biotin-switch assay (Hu et al. 2015). Whereas several
proteins are S-nitrosylated at a higher level in gsnor1
than in wild-type plants, the allelic gsnor1/hot5/par2
mutants show a reduced chlorophyll level and yellowing
leaves, suggesting that NO plays an important role in
chlorophyll metabolism (Lee et al. 2008; Hu et al. 2015).Again, nitrosoproteomic studies have revealed the
S-nitrosylated proteins significantly enriched in thephotosynthesis process, correlated to the alteredphotosynthetic properties in gsnor1 (Lindermayr et al.2005; Fares et al. 2011; Hu et al. 2015). In particular, alarge number of key components of both light and dark
Protein S-nitrosylation in plants 11
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1216

reactions of photosynthesis is S-nitrosylated (Hu et al.
2015). In the dark reaction (also known as the Calvin
cycle), the identified S-nitrosylated proteins cover in all
three phases of the Calvin cycle (carbon fixation,
reduction, and the regeneration of ribulose), including
the Rubisco complex [ribulose bisphosphate carboxyl-
ase, phosphoglycerate kinase, and glyceraldehyde-3-
phosphate dehydrogenase (GAPDH)]. These results
suggest that key components of photosynthesis are
extensively regulated by S-nitrosylation (Hu et al. 2015).Several S-nitrosylated proteins in chlorophyll metab-
olism and photosynthesis have been further character-ized by the biotin-switch assay (Abat et al. 2008; Abatand Deswal 2009; Hu et al. 2015). In the green algaeChlamydomonas reinhardtii, the translation of LHCBM, amajor light-harvesting protein at PSII, is controlledby light conditions. NAB1, a cytosolic RNA-bindingprotein, represses the translation of LHCBM in responseto high light exposure. Under low light conditions,S-nitrosylation of NAB1 at Cys-226 causes the decreasedrepressor activity, resulting in the accumulation of ahigher level of LHCBM and efficient light capture.Conversely, high light triggers denitrosylation of NAB1mediated by TRX-h1. This light-modulated nitrosylation-denitrosylation identifies NAB1 as a key elementfor redox-regulated photosynthetic light harvesting(Berger et al. 2016).
FUTURE CHALLENGES
In recent years, significant progress has been made inunderstanding the biochemistry and biological func-tionality of protein S-nitrosylation in plants. However, akey technical bottleneck is the lack of methods that arehighly sensitive, low noise/signal ratio, and easy-handling for the analysis of protein S-nitrosylation.Currently, the most often-used method for the analysisof S-nitrosylation is the biotin-switch assay (Jaffrey andSnyder 2001). This method involves several key steps,including the block of free thiol groups of cysteineresidues by methylation, followed by reducing themodified thiols with a strong reductant, usually ascorbicacid, and then decorating the reduced thiol with a biotinmolecule. The biotin-labelled protein or peptides arethen detected by immunoblotting using anti-biotinantibodies or by mass spectrometry. While the biotin-switch assay has been improved in some degrees
(Huang and Chen 2010), this original method is time-consuming and tedious, especially unsuitable formonitoring dynamic changes of this important post-translational modification in vivo. The development ofhigh sensitivity and user-friendly new methods istherefore technically demanding and emergent.
In addition to the methodological requirements,there are several challenging questions that need to befocused on in plant NO biology. First, do land plantshave a NOS-like system for NO biosynthesis? This hasbeen a long-time puzzle in NO biology. Until very recent,genomics studies indicate that the NOS-like genes arepresented in algae but disappeared since plantsconquered land (Rensing et al. 2008; Jeandroz et al.2016; Bowman et al. 2017; Nishiyama et al. 2018). Inresponse to this enigma, it has been proposed that landplants may have evolved and utilized a nitrateassimilation and reduction mechanism for NO biosyn-thesis, rather than using the conserved NOS enzymesfound in animals (Jeandroz et al. 2016). Perhaps, thehunt for NOS-like proteins or the unambiguousdemonstration of the absence of NOS-like genes inland plants will still be hard tasks.
Second, whereas S-nitrosylation has been consid-ered mainly as a non-enzymatic reaction, the recentlydiscovered transnitrosylation in animals has revealed animportant mechanism to determine the specificity ofS-nitrosylation. However, no transnitrosylase has beenfound in plants. Identification and characterization ofpotential transnitrosylases in plants will be criticalfor understanding the biochemical basis of selectiveS-nitrosylation.
Third, GSNOR is a master regulator of NO signaling.Both the enzymatic activity and the stability ofArabidopsis GSNOR1 are negatively regulated by NO(Guerra et al. 2016; Zhan et al. 2018). While this positiveregulatory feedback mechanism is important foramplifying the NO signal during early signaling, itremains completely unknown about the mechanismsthat turn down NO signaling. Addressing any of thesethree questions will be challenging and undoubtedlymajor breakthroughs in plant NO biology.
In response to these challenges, we expect thatbreakthrough progress will be achieved within the next5–10 years in addressing some key questions in plant NObiology. It may be optimistic to reveal, at least partially,the molecular mechanisms of selective S-nitrosylationandnegative regulationofNOsignaling in thenear future.
12 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1217

ACKNOWLEDGEMENTS
We apologize to colleagues whose work could not be
cited owing to space limitations. This work was
supported by grants from the National Natural Science
Foundation of China (31830017 and 31521001), Chinese
Academy of Sciences (XDB27030207), and State Key
Laboratory of Plant Genomics (SKLPG2016-22).
REFERENCES
Abat JK, Deswal R (2009) Differential modulation of S-nitrosoproteome of Brassica juncea by low temperature:Change in S-nitrosylation of Rubisco is responsible for theinactivation of its carboxylase activity. Proteomics 9:4368–4380
Abat JK, Mattoo AK, Deswal R (2008) S-nitrosylated proteinsof a medicinal CAM plant Kalanchoe pinnata– ribulose-1,5-bisphosphate carboxylase/oxygenase activity targeted forinhibition. FEBS J 275: 2862–2872
Ahern GP, Klyachko VA, Jackson MB (2002) cGMP andS-nitrosylation: Two routes for modulation of neuronalexcitability by NO. Trends Neurosci 25: 510–517
Albertos P, Romero-Puertas MC, Tatematsu K, Mateos I,Sanchez-Vicente I, Nambara E, Lorenzo O (2015) S-nitro-sylation triggers ABI5 degradation to promote seedgermination and seedling growth. Nat Commun 6: 8669
Anand P, Stamler JS (2012) Enzymatic mechanisms regulatingprotein S-nitrosylation: Implications in health and disease.J Mol Med (Berl) 90: 233–244
Asgher M, Per TS, Masood A, Fatma M, Freschi L, Corpas FJ,Khan NA (2017) Nitric oxide signaling and its crosstalk withother plant growth regulators in plant responses to abioticstress. Environ Sci Pollut Res Int 24: 2273–2285
Astier J, Besson-Bard A, Lamotte O, Bertoldo J, Bourque S,Terenzi H, Wendehenne D (2012a) Nitric oxide inhibits theATPase activity of the chaperone-like AAAþ ATPaseCDC48, a target for S-nitrosylation in cryptogein signallingin tobacco cells. Biochem J 447: 249–260
Astier J, Gross I, Durner J (2018) Nitric oxide production inplants: An update. J Exp Bot 69: 3401–3411
Astier J, Kulik A, Koen E, Besson-Bard A, Bourque S, JeandrozS, Lamotte O, Wendehenne D (2012b) Protein S-nitro-sylation: What’s going on in plants? Free Radic Biol Med53: 1101–1110
Astier J, Rasul S, Koen E, Manzoor H, Besson-Bard A, LamotteO, Jeandroz S, Durner J, Lindermayr C, Wendehenne D(2011) S-nitrosylation: An emerging post-translationalprotein modification in plants. Plant Sci 181: 527–533
Baker PR, Schopfer FJ, O’Donnell VB, Freeman BA (2009)Convergence of nitric oxide and lipid signaling: Anti-inflammatory nitro-fatty acids. Free Radic Biol Med 46:989–1003
Bartesaghi S, Radi R (2018) Fundamentals on the biochemistryof peroxynitrite and protein tyrosine nitration. Redox Biol14: 618–625
Begara-Morales JC, Chaki M, Sanchez-Calvo B, Mata-Perez C,Leterrier M, Palma JM, Barroso JB, Corpas FJ (2013)Protein tyrosine nitration in pea roots during developmentand senescence. J Exp Bot 64: 1121–1134
Begara-Morales JC, Chaki M, Valderrama R, Sanchez-Calvo B,Mata-Perez C, Padilla MN, Corpas FJ, Barroso JB (2018)Nitric oxide buffering and conditional nitric oxide releasein stress response. J Exp Bot 69: 3425–3438
Begara-Morales JC, Sanchez-Calvo B, Chaki M, Mata-Perez C,Valderrama R, Padilla MN, Lopez-Jaramillo J, Luque F,Corpas FJ, Barroso JB (2015) Differential molecularresponse of monodehydroascorbate reductase and gluta-thione reductase by nitration and S-nitrosylation. J ExpBot 66: 5983–5996
Belenghi B, Romero-Puertas MC, Vercammen D, Brackenier A,Inze D, Delledonne M, Van Breusegem F (2007) Meta-caspase activity of Arabidopsis thaliana is regulated by S-nitrosylation of a critical cysteine residue. J Biol Chem 282:1352–1358
Beligni MV, Lamattina L (2000) Nitric oxide stimulates seedgermination and de-etiolation, and inhibits hypocotylelongation, three light-inducible responses in plants.Planta 210: 215–221
Benhar M, Forrester MT, Stamler JS (2009) Protein denitro-sylation: Enzymatic mechanisms and cellular functions.Nat Rev Mol Cell Biol 10: 721–732
Berger H, De Mia M, Morisse S, Marchand CH, Lemaire SD,Wobbe L, Kruse O (2016) A light switch based on protein S-nitrosylation fine-tunes photosynthetic light harvesting inChlamydomonas. Plant Physiol 171: 821–832
Besson-Bard A, Pugin A, Wendehenne D (2008) New insightsinto nitric oxide signaling in plants.Annu Rev Plant Biol 59:21–39
Bethke PC, Badger MR, Jones RL (2004a) Apoplastic synthesisof nitric oxide by plant tissues. Plant Cell 16: 332–341
Bethke PC, Gubler F, Jacobsen JV, Jones RL (2004b) Dormancyof Arabidopsis seeds and barley grains can be broken bynitric oxide. Planta 219: 847–855
Bollhoner B, Zhang B, Stael S, Denance N, Overmyer K, GoffnerD, Van Breusegem F, Tuominen H (2013) Post mortemfunction of AtMC9 in xylem vessel elements. New Phytol200: 498–510
Bowman JL, Kohchi T, Yamato KT, Jenkins J, Shu S, Ishizaki K,Yamaoka S, Nishihama R, Nakamura Y, Berger F, Adam C,Aki SS, Althoff F, Araki T, Arteaga-Vazquez MA, Balasubr-manian S, Barry K, Bauer D, Boehm CR, Briginshaw L,Caballero-Perez J, Catarino B, Chen F, Chiyoda S, ChovatiaM, Davies KM, DelmansM, Demura T, Dierschke T, Dolan L,Dorantes-Acosta AE, Eklund DM, Florent SN, Flores-Sandoval E, Fujiyama A, Fukuzawa H, Galik B, GrimanelliD, Grimwood J, Grossniklaus U, Hamada T, Haseloff J,Hetherington AJ, Higo A, Hirakawa Y, Hundley HN, Ikeda Y,Inoue K, Inoue SI, Ishida S, Jia Q, Kakita M, Kanazawa T,Kawai Y, Kawashima T, Kennedy M, Kinose K, Kinoshita T,
Protein S-nitrosylation in plants 13
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1218

Kohara Y, Koide E, Komatsu K, Kopischke S, Kubo M,Kyozuka J, Lagercrantz U, Lin S-S, Lindquist E, Lipzen AM,Lu C-W, De Luna E, Martienssen RA, Minamino N, MizutaniM, Mizutani M, Mochizuki N, Monte I, Mosher R, NagasakiH, Nakagami H, Naramoto S, Nishitani K, Ohtani M,Okamoto T, Okumura M, Phillips J, Pollak B, Reinders A,R€ovekamp M, Sano R, Sawa S, Schmid MW, Shirakawa M,Solano R, Spunde A, Suetsugu N, Sugano S, Sugiyama A,Sun R, Suzuki Y, Takenaka M, Takezawa D, Tomogane H,Tsuzuki M, Ueda T, Umeda M, Ward JM, Watanabe Y,Yazaki K, Yokoyama R, Yoshitake Y, Yotsui I, Zachgo S,Schmutz J (2017) Insights into land plant evolutiongarnered from the Marchantia polymorpha genome. Cell171: 287–304e215
Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ (2006) ABA-induced NO generation and stomatal closure in Arabi-dopsis are dependent on H2O2 synthesis. Plant J 45: 113–122
Campbell WH (1996) Nitrate reductase bochemistry comes ofage. Plant Physiol 111: 355–361
Campbell WH, Kinghorn KR (1990) Functional domains ofassimilatory nitrate reductases and nitrite reductases.Trends Biochem Sci 15: 315–319
CastilloMC, Lozano-Juste J, Gonzalez-GuzmanM, Rodriguez L,Rodriguez PL, Leon J (2015) Inactivation of PYR/PYL/RCARABA receptors by tyrosine nitration may enable rapidinhibition of ABA signaling by nitric oxide in plants. SciSignal 8: ra89
Chaki M, Valderrama R, Fernandez-Ocana AM, Carreras A,Gomez-Rodriguez MV, Lopez-Jaramillo J, Begara-MoralesJC, Sanchez-Calvo B, Luque F, Leterrier M, Corpas FJ,Barroso JB (2011) High temperature triggers the metabo-lism of S-nitrosothiols in sunflower mediating a process ofnitrosative stress which provokes the inhibition offerredoxin-NADP reductase by tyrosine nitration. PlantCell Environ 34: 1803–1818
Chamizo-Ampudia A, Sanz-Luque E, Llamas A, Ocana-CalahorroF, Mariscal V, Carreras A, Barroso JB, Galvan A,Fernandez E (2016) A dual system formed by the ARCand NR molybdoenzymes mediates nitrite-dependentNO production in Chlamydomonas. Plant Cell Environ 39:2097–2107
Chen R, Sun S,Wang C, Li Y, Liang Y, An F, Li C, Dong H, Yang X,Zhang J, Zuo J (2009) The Arabidopsis PARAQUATRESISTANT2 gene encodes an S-nitrosoglutathione reduc-tase that is a key regulator of cell death. Cell Res 19:1377–1387
Chen YJ, Lu CT, Su MG, Huang KY, Ching WC, Yang HH,Liao YC, Chen YJ, Lee TY (2015) dbSNO 2.0: A resourcefor exploring structural environment, functional anddisease association and regulatory network of proteinS-nitrosylation. Nucleic Acids Res 43: D503–D511
Cooney RV, Harwood PJ, Custer LJ, Franke AA (1994) Light-mediated conversion of nitrogen dioxide to nitric oxide bycarotenoids. Environ Health Perspect 102: 460–462
Corpas FJ, Barroso JB, del Rio LA (2001) Peroxisomes as asource of reactive oxygen species and nitric oxide signalmolecules in plant cells. Trends Plant Sci 6: 145–150
Corpas FJ, Palma JM, Sandalio LM, Valderrama R, Barroso JB,Del Rio LA (2008) Peroxisomal xanthine oxidoreductase:characterization of the enzyme from pea (Pisum sativumL.) leaves. J Plant Physiol 165: 1319–1330
Crawford NM (2006) Mechanisms for nitric oxide synthesis inplants. J Exp Bot 57: 471–478
Cui B, Pan Q, Clarke D, Villarreal MO, Umbreen S, Yuan B,ShanW, Jiang J, Loake GJ (2018) S-nitrosylation of the zincfinger protein SRG1 regulates plant immunity. NatCommun 9: 4226
Dean JV, Harper JE (1988) The conversion of nitrite to nitrogenoxide(s) by the constitutive NAD(P)H-nitrate reductaseenzyme from soybean. Plant Physiol 88: 389–395
Delledonne M, Xia Y, Dixon RA, Lamb C (1998) Nitric oxidefunctions as a signal in plant disease resistance. Nature394: 585–588
Delledonne M, Zeier J, Marocco A, Lamb C (2001) Signalinteractions between nitric oxide and reactive oxygenintermediates in the plant hypersensitive disease resis-tance response. Proc Natl Acad Sci USA 98: 13454–13459
Desikan R, Griffiths R, Hancock J, Neill S (2002) A new role foran old enzyme: Nitrate reductase-mediated nitric oxidegeneration is required for abscisic acid-induced stomatalclosure in Arabidopsis thaliana. Proc Natl Acad Sci USA 99:16314–16318
Domingos P, Prado AM, Wong A, Gehring C, Feijo JA (2015)Nitric oxide: A multitasked signaling gas in plants. MolPlant 8: 506–520
Fancy NN, Bahlmann AK, Loake GJ (2017) Nitric oxide functionin plant abiotic stress. Plant Cell Environ 40: 462–472
Fares A, Rossignol M, Peltier JB (2011) Proteomics investiga-tion of endogenous S-nitrosylation in Arabidopsis. Bio-chem Biophys Res Commun 416: 331–336
Feechan A, Kwon E, Yun BW, Wang Y, Pallas JA, Loake GJ(2005) A central role for S-nitrosothiols in plant diseaseresistance. Proc Natl Acad Sci USA 102: 8054–8059
Feng J, Wang C, Chen Q, Chen H, Ren B, Li X, Zuo J (2013)S-nitrosylation of phosphotransfer proteins repressescytokinin signaling. Nat Commun 4: 1529
Freschi L (2013) Nitric oxide and phytohormone interactions:Current status and perspectives. Front Plant Sci 4: 398
Frohlich A, Durner J (2011) The hunt for plant nitric oxidesynthase (NOS): Is one really needed? Plant Sci 181:401–404
Frungillo L, Skelly MJ, Loake GJ, Spoel SH, Salgado I (2014) S-nitrosothiols regulate nitric oxide production and storagein plants through the nitrogen assimilation pathway. NatCommun 5: 5401
Gas E, Flores-Perez U, Sauret-Gueto S, Rodriguez-ConcepcionM (2009) Hunting for plant nitric oxide synthase providesnew evidence of a central role for plastids in nitric oxidemetabolism. Plant Cell 21: 18–23
Gibbs DJ, Md Isa N, Movahedi M, Lozano-Juste J, MendiondoGM, Berckhan S, Marin-de la Rosa N, Vicente Conde J,Sousa Correia C, Pearce SP, Bassel GW, Hamali B, Talloji P,Tome DF, Coego A, Beynon J, Alabadi D, Bachmair A,
14 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1219

Leon J, Gray JE, Theodoulou FL, Holdsworth MJ (2014)Nitric oxide sensing in plants is mediated by proteolyticcontrol of group VII ERF transcription factors.Mol Cell 53:369–379
Godber BL, Doel JJ, Durgan J, Eisenthal R, Harrison R (2000a)A new route to peroxynitrite: A role for xanthineoxidoreductase. FEBS Lett 475: 93–96
Godber BL, Doel JJ, Sapkota GP, Blake DR, Stevens CR,Eisenthal R, Harrison R (2000b) Reduction of nitrite tonitric oxide catalyzed by xanthine oxidoreductase. J BiolChem 275: 7757–7763
Guerra D, Ballard K, Truebridge I, Vierling E (2016) S-nitrosationof conserved cysteines modulates activity and stability ofS-nitrosoglutathione reductase (GSNOR). Biochemistry55: 2452–2464
Gupta KJ (2011) Protein S-nitrosylation in plants: Photo-respiratory metabolism and NO signaling. Sci Signal 4: doi:10.1126/scisignal.2001404
Gupta KJ, Fernie AR, Kaiser WM, van Dongen JT (2011) On theorigins of nitric oxide. Trends Plant Sci 16: 160–168
Hao G, Derakhshan B, Shi L, Campagne F, Gross SS (2006)SNOSID, a proteomic method for identification of cysteineS-nitrosylation sites in complex proteinmixtures. Proc NatlAcad Sci USA 103: 1012–1017
Harrison R (2002) Structure and function of xanthineoxidoreductase: Where are we now? Free Radic BiolMed 33: 774–797
He Y, Tang RH, Hao Y, Stevens RD, Cook CW, Ahn SM, Jing L,Yang Z, Chen L, Guo F, Fiorani F, Jackson RB, Crawford NM,Pei ZM (2004) Nitric oxide represses the Arabidopsis floraltransition. Science 305: 1968–1971
Helliwell CA, Chin-Atkins AN,Wilson LW, Chapple R, Dennis ES,Chaudhury A (2001) The Arabidopsis AMP1 gene encodesa putative glutamate carboxypeptidase. Plant Cell 13:2115–2125
Hess DT, Matsumoto A, Kim SO, Marshall HE, Stamler JS(2005) Protein S-nitrosylation: Purview and parameters.Nat Rev Mol Cell Biol 6: 150–166
Holzmeister C, Gaupels F, Geerlof A, Sarioglu H, Sattler M,Durner J, Lindermayr C (2015) Differential inhibition ofArabidopsis superoxide dismutases by peroxynitrite-medi-ated tyrosine nitration. J Exp Bot 66: 989–999
Hu J, Huang X, Chen L, Sun X, Lu C, Zhang L, Wang Y, Zuo J(2015) Site-specific nitrosoproteomic identification ofendogenously S-nitrosylated proteins in Arabidopsis. PlantPhysiol 167: 1731–1746
Hu J, Yang H, Mu J, Lu T, Peng J, Deng X, Kong Z, Bao S, Cao X,Zuo J (2017) Nitric oxide regulates protein methylationduring stress responses in plants.Mol Cell 67: 702–710 e704
Huang B, Chen C (2010) Detection of protein S-nitrosationusing irreversible biotinylation procedures (IBP). FreeRadic Biol Med 49: 447–456
Ib�a~nez-VeaM, Huang H,Mart�ınez deMorentin X, P�erez E, GatoM, Zuazo M, Arasanz H, Fern�andez-Irigoyen J, Santamar�ıaE, Fernandez-Hinojal G, Larsen MR, Escors D, Kochan G(2018) Characterization of macrophage endogenous
S-nitrosoproteome using a cysteine-specific phosphonateadaptable tag in combination with TiO2 chromatography.J Proteome Res 17: 1172–1182
Iglesias MJ, Terrile MC, Correa-Aragunde N, Colman SL,Izquierdo-Alvarez A, Fiol DF, Paris R, Sanchez-Lopez N,Marina A, Calderon Villalobos LIA, Estelle M, Lamattina L,Martinez-Ruiz A, Casalongue CA (2018) Regulation ofSCF(TIR1/AFBs) E3 ligase assembly by S-nitrosylation ofArabidopsis SKP1-like1 impacts on auxin signaling. RedoxBiol 18: 200–210
Ischiropoulos H (2009) Protein tyrosine nitration: An update.Arch Biochem Biophys 484: 117–121
Iwakiri Y, Satoh A, Chatterjee S, Toomre DK, Chalouni CM,Fulton D, Groszmann RJ, Shah VH, Sessa WC (2006) Nitricoxide synthase generates nitric oxide locally to regulatecompartmentalized protein S-nitrosylation and proteintrafficking. Proc Natl Acad Sci USA 103: 19777–19782
Jaffrey SR, Snyder SH (2001) The biotin switch method forthe detection of S-nitrosylated proteins. Sci STKE 2001:doi: 10.1126/stke.2001.86.pl1
Jeandroz S, Wipf D, Stuehr DJ, Lamattina L, Melkonian M,Tian Z, Zhu Y, Carpenter EJ, Wong GK-S, Wendehenne D(2016) Occurrence, structure, and evolution of nitric oxidesynthase-like proteins in the plant kingdom. Sci Signal 9:re2–re2
Jia J, Arif A, Terenzi F, Willard B, Plow EF, Hazen SL, Fox PL(2014) Target-selective protein S-nitrosylation by se-quence motif recognition. Cell 159: 623–634
Kawabe H, Ohtani M, Kurata T, Sakamoto T, Demura T (2018)Protein S-nitrosylation regulates xylem vessel cell differ-entiation in Arabidopsis. Plant Cell Physiol 59: 17–29
Kelley EE, Batthyany CI, Hundley NJ, Woodcock SR, Bonacci G,Del Rio JM, Schopfer FJ, Lancaster JR, Jr., Freeman BA,TarpeyMM (2008) Nitro-oleic acid, a novel and irreversibleinhibitor of xanthine oxidoreductase. J Biol Chem 283:36176–36184
Kim SF, Huri DA, Snyder SH (2005) Inducible nitric oxidesynthase binds, S-nitrosylates, and activates cyclooxygen-ase-2. Science 310: 1966–1970
Kneeshaw S, Gelineau S, Tada Y, Loake GJ, Spoel SH (2014)Selective protein denitrosylation activity of thioredoxin-h5modulates plant immunity. Mol Cell 56: 153–162
Kolbert Z, Bartha B, Erdei L (2008) Exogenous auxin-inducedNO synthesis is nitrate reductase-associated in Arabidopsisthaliana root primordia. J Plant Physiol 165: 967–975
Kwon E, Feechan A, Yun BW, Hwang BH, Pallas JA, Kang JG,Loake GJ (2012) AtGSNOR1 function is required formultiple developmental programs in Arabidopsis. Planta236: 887–900
Lamotte O, Bertoldo JB, Besson-Bard A, Rosnoblet C, Aime S,Hichami S, Terenzi H, Wendehenne D (2014) Protein S-nitrosylation: Specificity and identification strategies inplants. Front Chem 2: 114
Lee U, Wie C, Fernandez BO, Feelisch M, Vierling E (2008)Modulation of nitrosative stress by S-nitrosoglutathionereductase is critical for thermotolerance and plant growthin Arabidopsis. Plant Cell 20: 786–802
Protein S-nitrosylation in plants 15
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1220

Leitner M, Vandelle E, Gaupels F, Bellin D, Delledonne M(2009) NO signals in the haze: Nitric oxide signalling inplant defence. Curr Opin Plant Biol 12: 451–458
Lindermayr C, Saalbach G, Bahnweg G, Durner J (2006)Differential inhibition of Arabidopsismethionine adenosyl-transferases by protein S-nitrosylation. J Biol Chem 281:4285–4291
Lindermayr C, Saalbach G, Durner J (2005) Proteomicidentification of S-nitrosylated proteins in Arabidopsis.Plant Physiol 137: 921–930
Lindermayr C, Sell S, Muller B, Leister D, Durner J (2010) Redoxregulation of the NPR1-TGA1 system of Arabidopsisthaliana by nitric oxide. Plant Cell 22: 2894–2907
Ling T, Bellin D, Vandelle E, Imanifard Z, Delledonne M (2017)Host-mediated S-nitrosylation disarms the bacterial effec-torHopAI1 to reestablish immunity.Plant Cell 29: 2871–2881
Liu JZ, Duan J, NiM, Liu Z, QiuWL,Whitham SA, QianWJ (2017)S-nitrosylation inhibits the kinase activity of tomatophosphoinositide-dependent kinase 1 (PDK1). J BiolChem 292: 19743–19751
Liu L, Hausladen A, Zeng M, Que L, Heitman J, Stamler JS(2001) A metabolic enzyme for S-nitrosothiol conservedfrom bacteria to humans. Nature 410: 490–494
Liu L, Yan Y, Zeng M, Zhang J, Hanes MA, Ahearn G,McMahon TJ, Dickfeld T, Marshall HE, Que LG, Stamler JS(2004) Essential roles of S-nitrosothiols in vascularhomeostasis and endotoxic shock. Cell 116: 617–628
Liu WZ, Kong DD, Gu XX, Gao HB, Wang JZ, Xia M, Gao Q, TianLL, Xu ZH, Bao F, Hu Y, Ye NS, Pei ZM, He YK (2013)Cytokinins can act as suppressors of nitric oxide inArabidopsis. Proc Natl Acad Sci USA 110: 1548–1553
Liu X, Miller MJ, Joshi MS, Thomas DD, Lancaster JR, Jr. (1998)Accelerated reaction of nitric oxide with O2 within thehydrophobic interior of biological membranes. Proc NatlAcad Sci USA 95: 2175–2179
Lozano-Juste J, Colom-Moreno R, Leon J (2011) In vivo proteintyrosine nitration in Arabidopsis thaliana. J Exp Bot 62:3501–3517
Lozano-Juste J, Leon J (2010) Enhanced abscisic acid-mediatedresponses in nia1nia2noa1-2 triple mutant impaired in NIA/NR- and AtNOA1-dependent nitric oxide biosynthesis inArabidopsis. Plant Physiol 152: 891–903
Martinez-Ruiz A, Araujo IM, Izquierdo-Alvarez A, Hernansanz-Agustin P, Lamas S, Serrador JM (2013) Specificity inS-nitrosylation: A short-range mechanism for NO signal-ing? Antioxid Redox Signal 19: 1220–1235
Mata-Perez C, Sanchez-Calvo B, Begara-Morales JC,Carreras A, Padilla MN, Melguizo M, Valderrama R,Corpas FJ, Barroso JB (2016a) Nitro-linolenic acid is anitric oxide donor. Nitric Oxide 57: 57–63
Mata-Perez C, Sanchez-Calvo B, Padilla MN, Begara-MoralesJC, Luque F, Melguizo M, Jimenez-Ruiz J, Fierro-Risco J,Penas-Sanjuan A, Valderrama R, Corpas FJ, Barroso JB(2016b) Nitro-fatty acids in plant signaling: nitro-linolenicacid induces the molecular chaperone network inArabidopsis. Plant Physiol 170: 686–701
Mayer B, Hemmens B (1997) Biosynthesis and action of nitricoxide in mammalian cells. Trends Biochem Sci 22: 477–481
Melo PM, Silva LS, Ribeiro I, Seabra AR, Carvalho HG (2011)Glutamine synthetase is a molecular target of nitric oxidein root nodules ofMedicago truncatula and is regulated bytyrosine nitration. Plant Physiol 157: 1505–1517
Mittler R, Vanderauwera S, GolleryM, Van Breusegem F (2004)Reactive oxygen gene network of plants. Trends Plant Sci9: 490–498
Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB,Vandepoele K, Gollery M, Shulaev V, Van Breusegem F(2011) ROS signaling: The new wave? Trends Plant Sci 16:300–309
Mou Z, Fan W, Dong X (2003) Inducers of plant systemicacquired resistance regulate NPR1 function through redoxchanges. Cell 113: 935–944
Mur LA, Mandon J, Persijn S, Cristescu SM, Moshkov IE,Novikova GV, Hall MA, Harren FJ, Hebelstrup KH, Gupta KJ(2013) Nitric oxide in plants: An assessment of the currentstate of knowledge. AoB Plants 5: pls052
Nathan C, Xie QW (1994) Nitric oxide synthases: Roles, tolls,and controls. Cell 78: 915–918
Ni M, Zhang L, Shi YF, Wang C, Lu Y, Pan J, Liu JZ (2017)Excessive cellular S-nitrosothiol impairs endocytosis ofauxin efflux transporter PIN2. Front Plant Sci 8: 1988
Nishiyama T, Sakayama H, de Vries J, Buschmann H, Saint-Marcoux D, Ullrich KK, Haas FB, Vanderstraeten L, BeckerD, Lang D, Vosolsob�e S, Rombauts S, Wilhelmsson PKI,Janitza P, Kern R, Heyl A, R€umpler F, Villalobos LIAC, ClayJM, Skokan R, Toyoda A, Suzuki Y, Kagoshima H, Schijlen E,Tajeshwar N, Catarino B, Hetherington AJ, Saltykova A,Bonnot C, Breuninger H, Symeonidi A, Radhakrishnan GV,Van Nieuwerburgh F, Deforce D, Chang C, Karol KG,Hedrich R, Ulvskov P, Gl€ockner G, Delwiche CF, Petr�a�sek J,Van de Peer Y, Friml J, Beilby M, Dolan L, Kohara Y, SuganoS, Fujiyama A, Delaux P-M, Quint M, Theißen G, HagemannM, Harholt J, Dunand C, Zachgo S, Langdale J, Maumus F,Van Der Straeten D, Gould SB, Rensing SA (2018) The Charagenome: Secondary complexity and implications for plantterrestrialization. Cell 174: 448–464e424
Palmieri MC, Lindermayr C, Bauwe H, Steinhauser C, Durner J(2010) Regulation of plant glycine decarboxylase byS-nitrosylation and glutathionylation. Plant Physiol 152:1514–1528
Priestly J (1774) Experiments and Observations on DifferentKinds of Air. W. Bowyer and J. Nichols, London
Puyaubert J, Baudouin E (2014) New clues for a cold case:Nitric oxide response to low temperature. Plant CellEnviron 37: 2623–2630
Puyaubert J, Fares A, R�ez�e N, Peltier J-B, Baudouin E (2014)Identification of endogenously S-nitrosylated proteins inArabidopsis plantlets: Effect of cold stress on cysteinenitrosylation level. Plant Sci 215–216: 150–156
Qi J, Song CP, Wang B, Zhou J, Kangasj€arvi J, Zhu JK, Gong Z(2018) Reactive oxygen species signaling and stomatalmovement in plant responses to drought stress andpathogen attack. J Integr Plant Biol 60: 805–826
16 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1221

Que LG, Liu L, Yan Y, Whitehead GS, Gavett SH, Schwartz DA,Stamler JS (2005) Protection from experimental asthmaby an endogenous bronchodilator. Science 308:1618–1621
Radi R (2013) Protein tyrosine nitration: biochemical mecha-nisms and structural basis of functional effects. Acc ChemRes 46: 550–559
Rensing SA, Lang D, Zimmer AD, Terry A, Salamov A, ShapiroH, Nishiyama T, Perroud P-F, Lindquist EA, Kamisugi Y,Tanahashi T, Sakakibara K, Fujita T, Oishi K, Shin-I T, KurokiY, Toyoda A, Suzuki Y, Hashimoto S-i, Yamaguchi K,Sugano S, Kohara Y, Fujiyama A, Anterola A, Aoki S, AshtonN, Barbazuk WB, Barker E, Bennetzen JL, Blankenship R,Cho SH, Dutcher SK, Estelle M, Fawcett JA, Gundlach H,Hanada K, Heyl A, Hicks KA, Hughes J, Lohr M, Mayer K,Melkozernov A, Murata T, Nelson DR, Pils B, Prigge M,Reiss B, Renner T, Rombauts S, Rushton PJ, Sanderfoot A,Schween G, Shiu S-H, Stueber K, Theodoulou FL, Tu H, Vande Peer Y, Verrier PJ, Waters E, Wood A, Yang L, Cove D,Cuming AC, Hasebe M, Lucas S, Mishler BD, Reski R,Grigoriev IV, Quatrano RS, Boore JL (2008) The Physcomi-trella genome reveals evolutionary insights into theconquest of land by plants. Science 319: 64–69
Rockel P, Strube F, Rockel A, Wildt J, Kaiser WM (2002)Regulation of nitric oxide (NO) production by plant nitratereductase in vivo and in vitro. J Exp Bot 53: 103–110
Romero-Puertas MC, Campostrini N, Matt�e A, Righetti PG,Perazzolli M, Zolla L, Roepstorff P, Delledonne M (2008)Proteomic analysis of S-nitrosylated proteins in Arabidop-sis thaliana undergoing hypersensitive response. Proteo-mics 8: 1459–1469
Romero-Puertas MC, Laxa M, Matte A, Zaninotto F, Finke-meier I, Jones AM, Perazzolli M, Vandelle E, Dietz KJ,Delledonne M (2007) S-nitrosylation of peroxiredoxin II Epromotes peroxynitrite-mediated tyrosine nitration. PlantCell 19: 4120–4130
Rosnoblet C, BegueH, Blanchard C, Pichereaux C, Besson-BardA, Aime S, Wendehenne D (2017) Functional characteriza-tion of the chaperon-like protein Cdc48 in cryptogein-induced immune response in tobacco. Plant Cell Environ40: 491–508
Sakamoto A, Ueda M, Morikawa H (2002) Arabidopsisglutathione-dependent formaldehyde dehydrogenase isan S-nitrosoglutathione reductase. FEBS Lett 515: 20–24
Sanchez-Calvo B, Barroso JB, Corpas FJ (2013) Hypothesis:Nitro-fatty acids play a role in plant metabolism. Plant Sci199–200: 1–6
Sanz-Luque E, Ocana-Calahorro F, de Montaigu A, Chamizo-Ampudia A, Llamas A, Galvan A, Fernandez E (2015) THB1, atruncated hemoglobin, modulates nitric oxide levels andnitrate reductase activity. Plant J 81: 467–479
Schmidt HH, Walter U (1994) NO at work. Cell 78: 919–925
Seligman K, Saviani EE, Oliveira HC, Pinto-Maglio CAF,Salgado I (2008) Floral transition and nitric oxide emissionduring flower development in Arabidopsis thaliana isaffected in nitrate reductase-deficient plants. Plant CellPhysiol 49: 1112–1121
Serrato AJ, Romero-Puertas MC, Lazaro-Payo A, Sahrawy M(2018) Regulation by S-nitrosylation of the Calvin-Bensoncycle fructose-1,6-bisphosphatase in Pisum sativum. RedoxBiol 14: 409–416
Seth D, Hess DT, Hausladen A, Wang L, Wang YJ, Stamler JS(2018) A multiplex enzymatic machinery for cellularprotein S-nitrosylation. Mol Cell 69: 451–464 e456
Seth D, Stamler JS (2011) The SNO-proteome: Causation andclassifications. Curr Opin Chem Biol 15: 129–136
Shi YF, Wang DL, Wang C, Culler AH, Kreiser MA, Suresh J,Cohen JD, Pan J, Baker B, Liu JZ (2015) Loss of GSNOR1function leads to compromised auxin signaling and polarauxin transport. Mol Plant 8: 1350–1365
Stamler JS, Lamas S, Fang FC (2001) Nitrosylation: theprototypic redox-based signaling mechanism. Cell 106:675–683
Stamler JS, Toone EJ, Lipton SA, Sucher NJ (1997) (S)NOsignals: Translocation, regulation, and a consensus motif.Neuron 18: 691–696
Stohr C, Strube F,Marx G, UllrichWR, Rockel P (2001) A plasmamembrane-bound enzyme of tobacco roots catalyses theformation of nitric oxide from nitrite. Planta 212: 835–841
Stomberski CT, Hess DT, Stamler JS (2019) Protein S-nitrosylation: Determinants of specificity and enzymaticregulation of S-nitrosothiol-based signaling. AntioxidRedox Signal 30: 1331–1351
Tada Y, Spoel SH, Pajerowska-Mukhtar K, Mou Z, Song J,Wang C, Zuo J, Dong X (2008) Plant immunity requiresconformational changes of NPR1 via S-nitrosylation andthioredoxins. Science 321: 952–956
Takahashi M, Shigeto J, Sakamoto A, Morikawa H (2017a)Selective nitration of PsbO1 inhibits oxygen evolution fromisolated Arabidopsis thylakoid membranes. Plant SignalBehav 12: e1304342
Takahashi M, Shigeto J, Sakamoto A, Morikawa H (2017b)Selective nitration of PsbO1, PsbO2, and PsbP1 decreasesPSII oxygen evolution and photochemical efficiency inintact leaves ofArabidopsis. Plant Signal Behav 12: e1376157
Tanou G, Filippou P, Belghazi M, Job D, Diamantidis G,Fotopoulos V, Molassiotis A (2012) Oxidative and nitro-sative-based signaling and associated post-translationalmodifications orchestrate the acclimation of citrus plantsto salinity stress. Plant J 72: 585–599
Terrile MC, Paris R, Calderon-Villalobos LI, Iglesias MJ,Lamattina L, Estelle M, Casalongue CA (2012) Nitric oxideinfluences auxin signaling through S-nitrosylation of theArabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxinreceptor. Plant J 70: 492–500
Trapet P, Kulik A, Lamotte O, Jeandroz S, Bourque S, Nicolas-Frances V, Rosnoblet C, Besson-Bard A, Wendehenne D(2015) NO signaling in plant immunity: A tale ofmessengers. Phytochemistry 112: 72–79
Tun NN, Santa-Catarina C, Begum T, Silveira V, Handro W, FlohEI, Scherer GF (2006) Polyamines induce rapid biosynthe-sis of nitric oxide (NO) in Arabidopsis thaliana seedlings.Plant Cell Physiol 47: 346–354
Protein S-nitrosylation in plants 17
www.jipb.net XXX 2019 | Volume XXXX | Issue XXXX | XXX-XXDecember 2019 | Volume 61 | Issue 12 | 1206-1223 www.jipb.net
1222

Villacorta L, Zhang J, Garcia-Barrio MT, Chen XL, Freeman BA,Chen YE, Cui T (2007) Nitro-linoleic acid inhibits vascularsmooth muscle cell proliferation via the Keap1/Nrf2signaling pathway. Am J Physiol Heart Circ Physiol 293:H770–H776
Wang P, Du Y, Hou YJ, Zhao Y, Hsu CC, Yuan F, Zhu X, Tao WA,Song CP, Zhu JK (2015) Nitric oxide negatively regulatesabscisic acid signaling in guard cells by S-nitrosylation ofOST1. Proc Natl Acad Sci USA 112: 613–618
Wang P, Du Y, Li Y, Ren D, Song C (2010) Hydrogen peroxide-mediated activation of MAP Kinase 6 modulates nitricoxide biosynthesis and signal transduction in Arabidopsis.Plant Cell 22: 2981–2998
Wang Y, Lin A, Loake GJ, Chu C (2013) H2O2-induced leaf celldeath and the crosstalk of reactive nitric/oxygen species.J Integr Plant Biol 55: 202–208
Wang Y, Yun BW, Kwon E, Hong JK, Yoon J, Loake GJ (2006) S-nitrosylation: An emerging redox-based post-translationalmodification in plants. J Exp Bot 57: 1777–1784
Wang YQ, Feechan A, Yun BW, Shafiei R, Hofmann A, Taylor P,Xue P, Yang FQ, Xie ZS, Pallas JA, Chu CC, Loake GJ (2009)S-nitrosylation of AtSABP3 antagonizes the expression ofplant immunity. J Biol Chem 284: 2131–2137
Wendehenne D, Durner J, Klessig DF (2004) Nitric oxide: Anew player in plant signalling and defence responses. CurrOpin Plant Biol 7: 449–455
Wendehenne D, Gao Q-M, Kachroo A, Kachroo P (2014) Freeradical-mediated systemic immunity in plants. Curr OpinPlant Biol 20: 127–134
Wendehenne D, Pugin A, Klessig DF, Durner J (2001) Nitricoxide: Comparative synthesis and signaling in animal andplant cells. Trends Plant Sci 6: 177–183
Wilkinson JQ, Crawford NM (1991) Identification of theArabidopsis CHL3 gene as the nitrate reductase structuralgene NIA2. Plant Cell 3: 461–471
Wolhuter K, Whitwell HJ, Switzer CH, Burgoyne JR, Timms JF,Eaton P (2018) Evidence against stable protein S-nitro-sylation as a widespread mechanism of post-translationalregulation. Mol Cell 69: 438–450 e435
Yamamoto-Katou A, Katou S, Yoshioka H, Doke N, Kawakita K(2006) Nitrate reductase is responsible for elicitin-inducednitric oxide production in Nicotiana benthamiana. PlantCell Physiol 47: 726–735
Yamasaki H, Sakihama Y (2000) Simultaneous production ofnitric oxide and peroxynitrite by plant nitrate reductase: In
vitro evidence for the NR-dependent formation of activenitrogen species. FEBS Lett 468: 89–92
Yang H, Mu J, Chen L, Feng J, Hu J, Li L, Zhou JM, Zuo J (2015)S-nitrosylation positively regulates ascorbate peroxidaseactivity during plant stress responses. Plant Physiol 167:1604–1615
Yu M, Lamattina L, Spoel SH, Loake GJ (2014) Nitric oxidefunction in plant biology: A redox cue in deconvolution.New Phytol 202: 1142–1156
Yun BW, Feechan A, Yin M, Saidi NB, Le Bihan T, Yu M, MooreJW, Kang JG, Kwon E, Spoel SH, Pallas JA, Loake GJ (2011)S-nitrosylation of NADPH oxidase regulates cell death inplant immunity. Nature 478: 264–268
Yun BW, Skelly MJ, Yin M, Yu M, Mun BG, Lee SU, Hussain A,Spoel SH, Loake GJ (2016) Nitric oxide and S-nitro-soglutathione function additively during plant immunity.New Phytol 211: 516–526
Zaffagnini M, Morisse S, Bedhomme M, Marchand CH,Festa M, Rouhier N, Lemaire SD, Trost P (2013) Mecha-nisms of nitrosylation and denitrosylation of cytoplasmicglyceraldehyde-3-phosphate dehydrogenase from Arabi-dopsis thaliana. J Biol Chem 288: 22777–22789
Zaninotto F, Camera SL, Polverari A, Delledonne M (2006)Cross talk between reactive nitrogen and oxygen speciesduring the hypersensitive disease resistance response.Plant Physiol 141: 379–383
Zhan N, Wang C, Chen L, Yang H, Feng J, Gong X, Ren B, Wu R,Mu J, Li Y, Liu Z, Zhou Y, Peng J, Wang K, Huang X, Xiao S,Zuo J (2018) S-nitrosylation targets GSNO reductase forselective autophagy during hypoxia responses in plants.Mol Cell 71: 142–154 e146
Zhang J, Shao F, Li Y, Cui H, Chen L, Li H, Zou Y, Long C, Lan L,Chai J, Chen S, Tang X, Zhou JM (2007) A Pseudomonassyringae effector inactivates MAPKs to suppress PAMP-induced immunity in plants. Cell Host Microbe 1: 175–185
Zhang Z, Zhang S, Zhang Y, Wang X, Li D, Li Q, Yue M, Li Q,Zhang YE, Xu Y, Xue Y, Chong K, Bao S (2011) Arabidopsisfloral initiator SKB1 confers high salt tolerance byregulating transcription and pre-mRNA splicing throughaltering histone H4R3 and small nuclear ribonucleoproteinLSM4 methylation. Plant Cell 23: 396–411
Zhao MG, Chen L, Zhang LL, Zhang WH (2009) Nitricreductase-dependent nitric oxide production is involvedin cold acclimation and freezing tolerance in Arabidopsis.Plant Physiol 151: 755–767
18 Feng et al.
XXX 2019 | Volume XXXX | Issue XXXX | XXX-XX www.jipb.netDecember 2019 | Volume 61 | Issue 12 | 1206-1223www.jipb.net
1223
Related Documents











