ORIGINAL ARTICLE Isolation of a CK2α Subunit and the Holoenzyme from the Mussel Mytilus galloprovincialis and Construction of the CK2α and CK2β cDNAs Regina-Maria Kolaiti & Andrea Baier & Ryszard Szyszka & Sophia Kouyanou-Koutsoukou Received: 30 March 2010 / Accepted: 7 September 2010 # Springer Science+Business Media, LLC 2010 Abstract Protein kinase CK2 is a ubiquitous, highly pleio- tropic, and constitutively active phosphotransferase that phosphorylates mainly serine and threonine residues. CK2 has been studied and characterized in many organisms, from yeast to mammals. The holoenzyme is generally composed of two catalytic (α and/or α′) and two regulatory (β) subunits, forming a differently assembled tetramer. The free and catalytically active α/α′ subunits can be present in cells under some circumstances. We present here the isolation of a putative catalytic CK2α subunit and holoenzyme from gills of the mussel Mytilus galloprovincialis capable of phosphorylating the purified recombinant ribosomal protein rMgP1. For further analysis of M. galloprovincialis protein kinase CK2, the cDNA molecules of CK2α and CK2β subunits were con- structed and cloned into expression vectors, and the recombi- nant proteins were purified after expression in Escherichia coli. The recombinant MgCK2β subunit and MgP1 were phosphorylated by the purified recombinant MgCK2α subunit. The mussel enzyme presented features typical for CK2: affinity for GTP, inhibition by both heparin and ATP competitive inhibitors (TBBt, TBBz), and sensitivity towards NaCl. Predicted amino acid sequence comparison showed that the M. galloprovincialis MgCK2α and MgCK2β subunits have similar features to their mammalian orthologs. Keywords Protein kinase CK2 . Catalytic CK2α subunit . Regulatory CK2β subunit . Ribosomal stalk phosphorylation . Mytilus galloprovincialis Introduction Protein kinase CK2 is a ubiquitous highly conserved Ser/ Thr protein kinase which is found in all eukaryotes so far investigated, from yeast to human (Allende and Allende 1995; Pinna 1997; Litchfield 2003). Mounting evidence indicates that the enzyme is a component of regulatory networks that are involved in a wide variety of cellular processes including cell cycle control, transcription, signal transduction, cell proliferation, and survival (Litchfield 2003; Filhol and Cochet 2009). A unique property of this kinase is that it utilizes ATP and GTP as nucleotide substrates. The number of different α and β subunits differs in various species. Human CK2 heterotetrameric structure consists of two catalytic subunits α and/or α′ (42 and 38 kDa, respectively) and two regulatory β subunits (28 kDa) existing as α 2 β 2 , αα′β 2 , or α′ 2 β 2 configurations (Allende and Allende 1995; Pinna 2003; Ahmad et al. 2008). Distinct isoenzymic forms of the CK2 catalytic subunit have been identified in many organisms (Lozeman et al. 1990; Maridor et al. 1991; Glover 1998). CK2 has been considered to be a tetrameric enzyme, with CK2α subunit exerting control over the catalytic activity of CK2 at a number of possible levels. However, there is convinc- ing evidence that the CK2 catalytic subunits exist as free subunits as well as in the tetrameric CK2 complex (Abramczyk et al. 2003). R.-M. Kolaiti and A. Baier gave equal contribution to this study. R.-M. Kolaiti : S. Kouyanou-Koutsoukou (*) Department of Genetics and Biotechnology, Faculty of Biology, University of Athens, Panepistimiopolis, Athens 15701, Greece e-mail: [email protected] A. Baier : R. Szyszka Department of Molecular Biology, Institute of Biotechnology, The John Paul II Catholic University of Lublin, Al. Krasnicka 102, 20-718 Lublin, Poland Mar Biotechnol DOI 10.1007/s10126-010-9321-z

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

ORIGINAL ARTICLE
Isolation of a CK2α Subunit and the Holoenzymefrom the Mussel Mytilus galloprovincialis and Constructionof the CK2α and CK2β cDNAs
Regina-Maria Kolaiti & Andrea Baier &
Ryszard Szyszka & Sophia Kouyanou-Koutsoukou
Received: 30 March 2010 /Accepted: 7 September 2010# Springer Science+Business Media, LLC 2010
Abstract Protein kinase CK2 is a ubiquitous, highly pleio-tropic, and constitutively active phosphotransferase thatphosphorylates mainly serine and threonine residues. CK2has been studied and characterized in many organisms, fromyeast to mammals. The holoenzyme is generally composed oftwo catalytic (α and/or α′) and two regulatory (β) subunits,forming a differently assembled tetramer. The free andcatalytically active α/α′ subunits can be present in cells undersome circumstances.We present here the isolation of a putativecatalytic CK2α subunit and holoenzyme from gills of themussel Mytilus galloprovincialis capable of phosphorylatingthe purified recombinant ribosomal protein rMgP1. Forfurther analysis of M. galloprovincialis protein kinase CK2,the cDNA molecules of CK2α and CK2β subunits were con-structed and cloned into expression vectors, and the recombi-nant proteins were purified after expression in Escherichiacoli. The recombinant MgCK2β subunit and MgP1 werephosphorylated by the purified recombinant MgCK2αsubunit. The mussel enzyme presented features typical forCK2: affinity for GTP, inhibition by both heparin and ATPcompetitive inhibitors (TBBt, TBBz), and sensitivity towardsNaCl. Predicted amino acid sequence comparison showed
that the M. galloprovincialis MgCK2α and MgCK2βsubunits have similar features to their mammalian orthologs.
Keywords Protein kinase CK2 . Catalytic CK2α subunit .
Regulatory CK2β subunit . Ribosomal stalkphosphorylation .Mytilus galloprovincialis
Introduction
Protein kinase CK2 is a ubiquitous highly conserved Ser/Thr protein kinase which is found in all eukaryotes so farinvestigated, from yeast to human (Allende and Allende1995; Pinna 1997; Litchfield 2003). Mounting evidenceindicates that the enzyme is a component of regulatorynetworks that are involved in a wide variety of cellularprocesses including cell cycle control, transcription, signaltransduction, cell proliferation, and survival (Litchfield2003; Filhol and Cochet 2009). A unique property of thiskinase is that it utilizes ATP and GTP as nucleotidesubstrates. The number of different α and β subunitsdiffers in various species. Human CK2 heterotetramericstructure consists of two catalytic subunits α and/or α′ (42and 38 kDa, respectively) and two regulatory β subunits(28 kDa) existing as α2β2, αα′β2, or α′2β2 configurations(Allende and Allende 1995; Pinna 2003; Ahmad et al.2008). Distinct isoenzymic forms of the CK2 catalyticsubunit have been identified in many organisms (Lozemanet al. 1990; Maridor et al. 1991; Glover 1998). CK2 hasbeen considered to be a tetrameric enzyme, with CK2αsubunit exerting control over the catalytic activity of CK2at a number of possible levels. However, there is convinc-ing evidence that the CK2 catalytic subunits exist as freesubunits as well as in the tetrameric CK2 complex(Abramczyk et al. 2003).
R.-M. Kolaiti and A. Baier gave equal contribution to this study.
R.-M. Kolaiti : S. Kouyanou-Koutsoukou (*)Department of Genetics and Biotechnology, Faculty of Biology,University of Athens,Panepistimiopolis,Athens 15701, Greecee-mail: [email protected]
A. Baier : R. SzyszkaDepartment of Molecular Biology, Institute of Biotechnology,The John Paul II Catholic University of Lublin,Al. Krasnicka 102,20-718 Lublin, Poland
Mar BiotechnolDOI 10.1007/s10126-010-9321-z

Although related, CK2α and CK2α′ are the products ofdifferent genes (Lozeman et al. 1990; Glover 1998). Threeisoforms of the catalytic α subunit, CK2α, CK2α′, andCK2α′′, have been identified in humans. Two of them,namely, CK2α and CK2α′, exhibit more than 90% aminoacid sequence identity at the N-terminal 330 amino acidsbut have unrelated C-terminal sequence (Lozeman et al.1990). The CK2α′′ subunit is almost identical to CK2α,and the only distinguishing feature between them lies in theunique sequence at the C-terminal 32 amino acids. CK2α′′is highly expressed in liver and correlates with normaltrafficking of hepatocellular membrane proteins (Shi et al.2001; Hilgard et al. 2004).
Knockout experiments in mice suggest that CK2α maycompensate the role of CK2α′ regarding viability. How-ever, some defects were detected in spermatogenesis (Xu etal. 1999). Similar effects were observed in Saccharomycescerevisiae when either gene for CK2α or CK2α′ wasblocked but the cells remained viable, while knockout ofboth genes was lethal. Similarly in mice, functional overlapbetween CK2α and CK2α′ is incomplete since yeasttemperature-sensitive alleles of CKA1 or CKA2 exhibitdistinct phenotypes (Glover 1998).
In humans, only one regulatory subunit CK2β has beenidentified, while multiple forms have been identified inother organisms such as S. cerevisiae (Bibby and Litchfield2005; Glover 1998). CK2β does not display an extensivehomology with the regulatory subunits of other proteinkinases but is highly conserved among various species andits amino acid sequence is completely identical betweenbirds and mammals (Maridor et al. 1991). Several lines ofevidence indicate that dimers of CK2β are at the core of thetetrameric CK2 complexes (Litchfield 2003; Glover 1998).CK2β contains at its N-terminus a phosphorylation sitecomprising the serine residues S2, S3, and possibly S4
(Boldyreff et al. 1993). Although the physiological rele-vance of this process remains unclear, it is evident thatCK2β is phosphorylated at this site in cells (Litchfieldet al. 1991). CK2β is also phosphorylated at S209, a sitethat is modulated in a cell cycle-dependent manner in vitroand in vivo by p34cdc2 (Litchfield et al. 1991; Meggio etal. 1995). It has been shown that CK2β enhances thestability of the catalytic subunits in the CK2 holoenzymeand modulates their substrate specificity. For instance,CK2β is capable to stimulate the CK2 holoenzyme activitytowards certain protein substrates, like topoisomerase II(Leroy et al. 1999), while it inhibits its activity towardscalmodulin (Bidwai et al. 1993; Marin et al. 1999).
CK2 seems to be also responsible for the phosphoryla-tion of the ribosomal stalk proteins P0, P1, and P2(Ballesta et al. 1999; Bou et al. 2000; Wojda et al. 2002).Phosphorylation of the ribosomal P-proteins has beendescribed for S. cerevisiae (Zieliński et al. 2002; Tchórzewski
et al. 2000; Nusspaumer et al. 2000; Ballesta et al. 2000), forZea mays (Szick-Miranda and Bailey-Serres 2001), and byus for the insects Ceratitis capitata and Bombyx mori(Gagou et al. 1999, 2000; Koumarianou et al. 2007). Arecent phosphoproteomic investigation of P-proteins fromPopulus dormant terminal buds identified, among others, thephosphorylation of the ribosomal P-proteins P0, P1, P2, andP3 at S and T sites (Liu et al. 2010). The identifiedphosphopeptides shared a common phosphorylation motif(S/T)XX(D/E), an indication that they may be phosphory-lated in vivo by protein kinase CK2.
For bivalves and especially the mussel Mytilus gallo-provincialis, no isolation of protein kinase CK2 has beenreferred, so far. However, Bardales et al. (2007) haverecently described the CK2-mediated phosphorylation of atype II regulatory subunit of cAMP-dependent proteinkinase using human CK2. In their study, they detected aCK2-like activity in the crude extract from mantle tissueusing GTP as phosphate donor. This activity was inhibitedby CK2 inhibitors, like emodin and apigenin.
We have recently investigated the ribosomal stalk proteinsMgP0, MgP1, andMgP2 ofM. galloprovincialis. We isolatedtheir cDNA molecules and purified the recombinant proteins(Kolaiti et al. 2009). Also, we showed that MgP1 and MgP2proteins are possibly phosphorylated and that MgP0 is over-expressed at stress conditions. It is known that cellular stresscaused by environmental contamination induces genotoxicdamage in marine bivalves like the mussel Mytilus edulis(Hagger et al. 2005) or spatial and seasonal variability inglobal protein synthesis as in the mussel M. galloprovincialis(Geret and Cosson 2002; Kalpaxis et al. 2004; Pytharopou-lou et al. 2006). To this direction, we have studied andpresent here the isolation and partial purification of aputative protein kinase CK2 from the gills of M. gallopro-vincialis as well as the construction of the cDNA moleculesof the catalytic CK2α and the regulatory CK2β subunits andthe purification of the recombinant proteins.
Materials and Methods
Materials
Restriction enzymes and T4 DNA-Ligase were obtained fromNew England Biolabs, pTA plasmid (pGEM) from Promega,expression vector pRSET from Invitrogen, Go Taq Flexi andPfu DNA Polymerase from Promega, and 5′/3′ RACE Kit—2nd Generation from ROCHE. Protein standards were fromFermentas GmbH (Germany) and [γ-32P]ATP (3,000 Ci/mmol) from Hartmann Analytics (Braunschweig, Germany).Chromatography equipment and accessories were purchasedfrom Pharmacia Biotech or Whatman. Further chemicals ifnot otherwise stated were from Sigma-Aldrich.
Mar Biotechnol

EMBL Nucleotide Sequence Database AccessionNumbers
M. galloprovincialis protein kinase CK2 catalytic subunitMgCK2α (mRNA): FN677519
M. galloprovincialis protein kinase CK2 regulatorysubunit MgCK2β (mRNA): FN677520
Animals—Animal Treatments
Adult mussels (70–80 mm in length) M. galloprovincialis(Lam.) were kindly provided by a marine farm (KastanisCo, Neraki, Attiki, Saronikos gulf, Greece) during theperiods of May and October 2008. The molluscs wereplaced in tanks containing seawater and transported to thelaboratory. Tissues were dissected out and immediatelyfrozen at −80°C until use.
Isolation and Partial Purification of Putative ProteinKinase CK2
Gills of M. galloprovincialis were homogenized 1:3 (m/v)in ice-cold buffer A containing 50 mM Tris–HCl buffer,pH 7.5, 6 mM 2-mercaptoethanol, 0.5 mM EDTA, and0.5 mM of phenyl-methylsulfonyl fluoride and disruptedby sonication. The homogenate was centrifuged at 3,000 gfor 10 min to remove cell debris. Afterwards, thesupernatant was again centrifuged at 100,000 g for 2 h toget the ribosome-free extract (S-100 fraction). Lipids wereremoved by filtration. In the first step of purification, thesupernatant was loaded onto a DEAE-cellulose column(DE-52) equilibrated with buffer A. Bound protein waseluted by a gradient from 0 to 500 mM NaCl in buffer A.The flow-through fraction (DE0) and the 350–450 mMNaCl fractions (DE400) were dialysed and further sepa-rately purified by P-cellulose chromatography (P11). Inboth cases, bound protein was eluted with a gradientfrom 0 to 1 M NaCl in buffer A. Active fractions fromeach P-cellulose column were further separately purifiedby heparin–agarose chromatography (Sigma). CK2enzymes were eluted from each column in 0.5-mL frac-tions with 250, 500, and 750 mM NaCl in buffer A.Fractions showing phosphorylation activity were dialysed,concentrated, and stored at −20°C in the presence ofglycerol. Fractions from this third step of the purificationprocess were then used for the characterization of nativeCK2.
Purification of Recombinant P1
Recombinant ribosomal stalk protein P1 from M. gallopro-vincialis (MgP1) was purified as previously described(Kolaiti et al. 2009).
Purification of Recombinant Human and Yeast CK2αSubunits
Recombinant CK2α subunits of human and yeast werepurified as previously described (Bosc et al. 2000; Sajnagaet al. 2008).
Protein Phosphorylation
CK2 activity was determined in a standard reaction mixture(50 μL of final volume) containing 20 mM Tris–HClbuffer, pH 7.5, 10 mM Mg2+, and 20 μM [γ-32P]ATP(specific radioactivity 500–1,000 cpm/pmol) in the presenceof rMgP1 as endogenous substrate. Incubation was per-formed at 37°C for 30 min. Afterwards, the reaction wasterminated by adding 10 μL of the SDS-PAGE loadingbuffer. Reaction mixtures were resolved in SDS-PAGEfollowed by coomassie blue staining and autoradiography.The phosphate incorporation level in the rMgP1 protein wasestimated by cutting off the corresponding band andmeasuring the radioactivity in a scintillation counter.
In-gel Kinase Assay
Detection of protein kinase after SDS-PAGE was performedusing the modified methods of Wang and Erikson (1992)and Kameshita and Fujisawa (1989). Protein fractionscontaining putative MgCK2 were resolved on 12.5%SDS-PAGE containing 0.2 mg/mL rMgP1. After electro-phoresis, the gel was washed twice with 50 mM Tris–HCl,pH 8.0 (buffer B), containing 20% isopropanol and twicewith 100 mL buffer B containing 5 mM 2-mercaptoethanolfor 30 min to remove SDS. Afterwards, the gel was washedtwice with 75 mL 6 M guanidine in buffer B for 30 min todenature proteins, which were later renaturated by soakingthe gel in 100 mL buffer B containing 0.04% Tween for30 min at room temperature and overnight at 4°C. The gelwas washed thereafter in 100 mL of the same fresh solutionfor 30 min and then it was equilibrated in 20 mLphosphorylation buffer (20 mM Tris–HCl, pH 7.5, 10 mMMgCl2) for 30 min. The reaction was carried out in 20 mLof phosphorylation buffer containing 20 μM [γ-32P]ATP(0.1 mCi). After a 1-h reaction at 37°C, the gel was washedten times with 100 mL 10% TCA. Finally, the gel was driedand exposed to Kodak X-ray film to detect labeled bands.
Immunodetection of CK2 Subunits
After SDS-PAGE, the proteins were transferred for 1.5 honto Immobilon P transfer membrane (0.45 μm, Millipore)in transfer buffer (24 mM Tris, 19.5 mM glycine, 0.037%SDS, 20% methanol). The membrane was incubated for 1 hin TBS buffer (10 mM Tris–HCl, pH 7.5, 0.9% NaCl) with
Mar Biotechnol

1%milk and for 1 h with the monoclonal anti-CK2α and anti-CK2β or anti-His antibodies (Sigma) (1:10,000 dilution inTBS with 1% milk). Afterwards, the membrane was washedwith TBS with 1% milk and 0.1% Tween 20. The boundantibodies were detected with anti-mouse antibodies conju-gated with alkaline phosphatase (Sigma) (1:10,000 dilution).
Construction of M. galloprovincialis CK2α and CK2βcDNAs
MgCK2α Subunit In order to determine the 5′- and 3′-endof MgCK2α, a conservative part of the cDNA was firstconstructed by PCR and cloned in pGEM vector using thedegenerated primers MgCK2AF1 (5′-CTAGGCCGGGGWAAATACAGTGAAGT-3 ′) and MgCK2AR1 (5 ′-TCWGCCAAACCCCAGTCTATTAGACG-3′), designedaccording to the conserved regions of CK2a cDNAs fromother eukaryotic species and total cDNA pool from M.galloprovincialis gills as template. The 404-bp PCRproduct was subjected to sequence analysis and found tocorrespond to CK2α. To determine the sequences at the 5′-and 3′-ends of the MgCK2α mRNA, rapid amplification ofcDNA ends (RACE) assays was carried out using the 5′/3′RACE Kit and specific homologue primers designedaccording to the constructed MgCK2α sequence. Amplifi-cation of the 3′-end was carried out using an oligo(dT)containing adapter primer for first-strand cDNA synthesisof poly(A) RNA. The first round of cDNA amplificationwas performed using 250 pmol each of an abridgeduniversal amplification primer (anchor) and 500 pmol ofthe gene-specific primer MgCK2αRACE1′F (5′-GGAAAATCTGCGTGGTGG-3′) and the second amplificationwith 250 pmol of the same anchor primer and 500 pmol ofa gene-speci f ic pr imer MgCK2αRACE1F (5 ′ -GCTAATGGCGATTGTTAAAGACC-3′). The prominentband on an agarose gel was subcloned in pGEM vectorand sequenced. To amplify the MgCK2α 5′ end, 250 pmolof the gene-specific oligonucleotide MgCK2αRACEa (5′-CCATACTGTGACAATAATCTAATGC-3′) was used tosynthesize the first strand of cDNA that was then purifiedand used for amplification of the 5′-end. For the first roundof amplification, PCR was performed with 250 pmol ofoligo-dT primer (5′/3′ RACE Kit) and 500 pmol ofMgCK2αRACE1 (5 ′-GGTCTTTAACAATCGCCATTAGC-3′). For the second amplification, a nested PCR wasperformed with 250 pmol of the anchor primer and 500 pmolof a gene-specific primer MgCK2αRACE1′ (5′-CCACCACGCAGATTTTCC-3′). The prominent band on an agarosegel was subcloned in pGEM vector and sequenced.
MgCK2β Subunit To amplify the M. galloprovincialisCK2β cDNA, the primers MgCK2βF (5′-ATGAGTAGCTCAGAAGTATC-3′) and MgCK2βR (5′-TTATCGTTTAT
TATTACTATGGC-3′) were designed identical to the firstand last nucleotides of the published EST of M. gallopro-vincialis, FL594940 (Venier, et al. 2009), that was found tocorrespond to the regulatory subunit of protein kinase CK2after BLAST search. The initial cDNA pool was used astemplate for standard PCR with Pfu DNA polymerase usingthe above primers. The PCR product was subcloned in pTAplasmid (pGEM), sequenced, and found to correspond toCK2β by comparing data from the Swiss-Prot databaseusing the BLAST and the CLUSTAL W programs.
Cloning of the MgCK2α and MgCK2β cDNAs in pRSETExpression Vectors
In order to over-express the MgCK2α andMgCK2β proteinsin Escherichia coli, their cDNAs were first subcloned inpTA plasmids adding XhoI and EcoRI restriction sites withappropriate primers. MgCK2α: MgCK2αXhoI (5′-CCGCTCGAGCGGATGCCGGCAACCAGTAAAGC-3′)and MgCK2αEcoRI (5′-CCGGAATTCCGGTTATCGGGGTATACC-3′); MgCK2β: MgCK2βXhoI (5′-CCGCTCGAGCGGATGAGTAGCTCAG-3′) and MgCK2βEcoRI (5′-CCGGAATTCCGGTTATCGTTTATTATTACTATGGC-3′).The inserted XhoI and EcoRI restriction sites are italicized.The initial cDNA pool was used as template for standardPCR with Pfu DNA Polymerase using the above primers. A-ends were then added to the PCR products. The resultingplasmids (pGEM) were digested with XhoI and EcoRI andthe appropriate fragments were subcloned into pRSETexpression vector.
Purification of the Recombinant MgCK2α and MgCK2βProteins After Expression in E. coli
E. coli BL21(DE3) strains carrying the pRSET expressionvectors with histidine-tagged MgCK2α or MgCK2βproteins were grown until OD600=0.6–0.8. IPTG was thenadded to a final concentration of 0.1–0.3 mM and cellswere further cultivated at 37°C for 4 h. Cells wereharvested by centrifugation at 5,000 g. The cell pelletswere suspended in buffer A, homogenized, and disruptedby sonication. After centrifugation at 10,000 g for 15 min,the supernatant was incubated with Ni-NTA beads (Sigma)equilibrated with buffer A overnight at 4°C. The mixture wasthen applied onto an empty column and allowed to drain bygravity. The matrix was washed with buffer A and boundprotein was eluted with a gradient from 0 to 500 mMimidazole. The purity of the eluted fractions was analyzed bySDS-PAGE. For further purification, the eluted fractionswere combined, concentrated, and separated by P-cellulose(MgCK2α) and DEAE-cellulose (MgCK2β), respectively.Finally, the eluted fractions were analyzed by SDS-PAGE,immunobloting, and kinase assay.
Mar Biotechnol
Results and Discussion
Isolation and Partial Purification of Putative Protein KinaseCK2 from the Gills of M. galloprovincialis
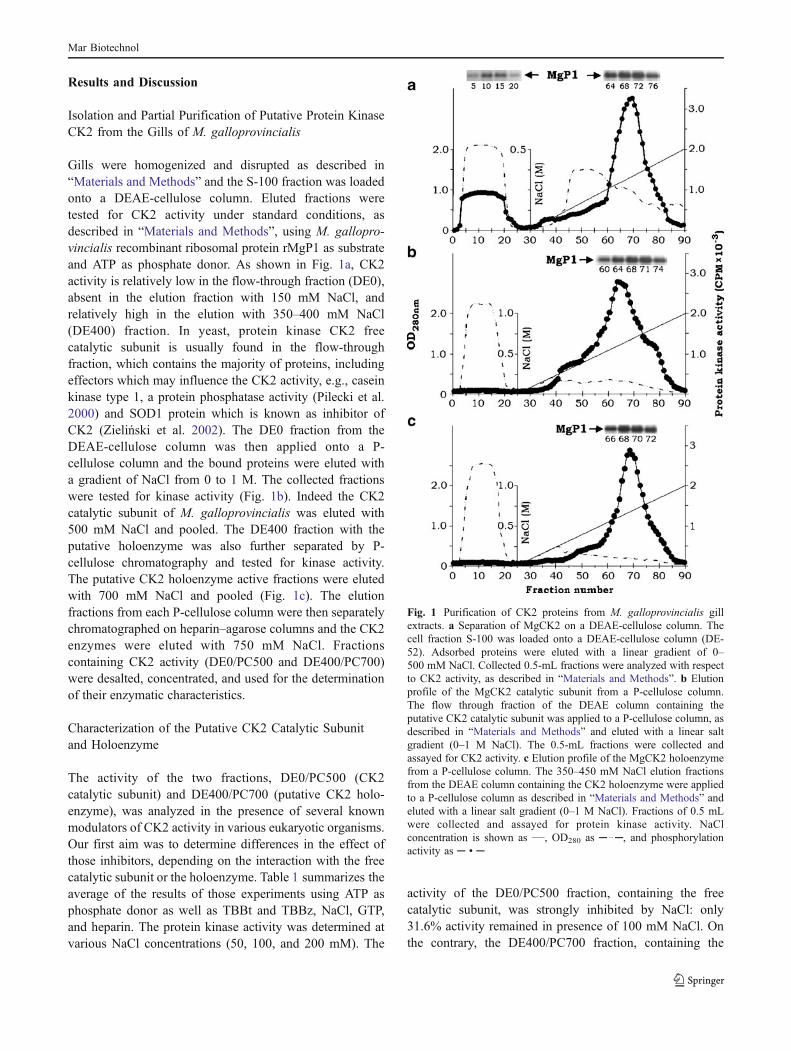
Gills were homogenized and disrupted as described in“Materials and Methods” and the S-100 fraction was loadedonto a DEAE-cellulose column. Eluted fractions weretested for CK2 activity under standard conditions, asdescribed in “Materials and Methods”, using M. gallopro-vincialis recombinant ribosomal protein rMgP1 as substrateand ATP as phosphate donor. As shown in Fig. 1a, CK2activity is relatively low in the flow-through fraction (DE0),absent in the elution fraction with 150 mM NaCl, andrelatively high in the elution with 350–400 mM NaCl(DE400) fraction. In yeast, protein kinase CK2 freecatalytic subunit is usually found in the flow-throughfraction, which contains the majority of proteins, includingeffectors which may influence the CK2 activity, e.g., caseinkinase type 1, a protein phosphatase activity (Pilecki et al.2000) and SOD1 protein which is known as inhibitor ofCK2 (Zieliński et al. 2002). The DE0 fraction from theDEAE-cellulose column was then applied onto a P-cellulose column and the bound proteins were eluted witha gradient of NaCl from 0 to 1 M. The collected fractionswere tested for kinase activity (Fig. 1b). Indeed the CK2catalytic subunit of M. galloprovincialis was eluted with500 mM NaCl and pooled. The DE400 fraction with theputative holoenzyme was also further separated by P-cellulose chromatography and tested for kinase activity.The putative CK2 holoenzyme active fractions were elutedwith 700 mM NaCl and pooled (Fig. 1c). The elutionfractions from each P-cellulose column were then separatelychromatographed on heparin–agarose columns and the CK2enzymes were eluted with 750 mM NaCl. Fractionscontaining CK2 activity (DE0/PC500 and DE400/PC700)were desalted, concentrated, and used for the determinationof their enzymatic characteristics.
Characterization of the Putative CK2 Catalytic Subunitand Holoenzyme
The activity of the two fractions, DE0/PC500 (CK2catalytic subunit) and DE400/PC700 (putative CK2 holo-enzyme), was analyzed in the presence of several knownmodulators of CK2 activity in various eukaryotic organisms.Our first aim was to determine differences in the effect ofthose inhibitors, depending on the interaction with the freecatalytic subunit or the holoenzyme. Table 1 summarizes theaverage of the results of those experiments using ATP asphosphate donor as well as TBBt and TBBz, NaCl, GTP,and heparin. The protein kinase activity was determined atvarious NaCl concentrations (50, 100, and 200 mM). The
activity of the DE0/PC500 fraction, containing the freecatalytic subunit, was strongly inhibited by NaCl: only31.6% activity remained in presence of 100 mM NaCl. Onthe contrary, the DE400/PC700 fraction, containing the
Fig. 1 Purification of CK2 proteins from M. galloprovincialis gillextracts. a Separation of MgCK2 on a DEAE-cellulose column. Thecell fraction S-100 was loaded onto a DEAE-cellulose column (DE-52). Adsorbed proteins were eluted with a linear gradient of 0–500 mM NaCl. Collected 0.5-mL fractions were analyzed with respectto CK2 activity, as described in “Materials and Methods”. b Elutionprofile of the MgCK2 catalytic subunit from a P-cellulose column.The flow through fraction of the DEAE column containing theputative CK2 catalytic subunit was applied to a P-cellulose column, asdescribed in “Materials and Methods” and eluted with a linear saltgradient (0–1 M NaCl). The 0.5-mL fractions were collected andassayed for CK2 activity. c Elution profile of the MgCK2 holoenzymefrom a P-cellulose column. The 350–450 mM NaCl elution fractionsfrom the DEAE column containing the CK2 holoenzyme were appliedto a P-cellulose column as described in “Materials and Methods” andeluted with a linear salt gradient (0–1 M NaCl). Fractions of 0.5 mLwere collected and assayed for protein kinase activity. NaClconcentration is shown as ___, OD280 as ─ ..─, and phosphorylationactivity as ─ • ─
Mar Biotechnol
holoenzyme, possessed an increasing activity at the sameconcentration, compared to the control without NaCl. At aconcentration of 200 mM NaCl, inhibition up to 80% wasobserved. It is known that the regulatory subunit isresponsible for stabilization and influences the activity ofthe tetrameric complex (Bibby and Litchfield 2005).Similar results have been obtained in the case of recon-stituted human and maize holoenzymes α2β2 (Riera et al.2001). The difference in the behavior between bothfractions was also measurable using the selective CK2inhibitors TBBz and TBBt (Zień et al. 2003). TBBzinhibited the free catalytic subunit and holoenzyme withIC50 values of 0.8 and 1 μM, respectively. In higher con-centrations, the putative holoenzyme was more effectivelyinhibited. The putative free catalytic subunit was inhibitedmore than 50% by 0.5 μM TBBt. In the case of theholoenzyme, the IC50 was below 0.3 μM. Our resultscorroborate with former studies including several CK2isoforms from yeast to human where the selective inhibitors
Table 1 Comparison of native free catalytic subunit CK2α and CK2holoenzyme activities of the mussel M. galloprovincialis
Effector MgCK2α (DE0/PC500) MgCK2 holoenzyme(DE400/PC700)
% of activity
25 μM ATP (control) 100.00 100.00
ATP/GTP (1:1) 85.4±2.2 62.1±3.1
ATP/GTP (1:2) 83.5±1.9 54.9±2.5
ATP/GTP (1:4) 79.5±1.7 53.5±2.7
1 μM TBBz 45.7±0.8 50.1±0.9
5 μM TBBz 39.6±0.7 20.3±0.5
20 μM TBBz 34.7±0.5 14.0±0.5
0.5 μM TBBt 45.5±1.0 17.2±0.5
2.5 μM TBBt 44.5±0.8 9.3±0.3
10.0 μM TBBt 32.0±0.6 4.8±0.2
0.2 ng/μL heparin 71.3±2.0 57.3±0.9
1.0 ng/μL heparin 62.0±1.6 42.5±0.7
50 mM NaCl 38.2±0.6 143.9±8.1
100 mM NaCl 31.6±0.5 136.0±5.4
200 mM NaCl 16.6±0.4 16.5±0.5
All experiments were performed three times. Summarized are theaverage of the results and the standard deviation between the threeexperiments
Fig. 2 Detection of the CK2 activities in protein fractions isolatedfrom M. galloprovincialis gill extracts with an in-situ phosphorylationmethod. On the 12.5% SDS-PAGE containing 0.2 mg/mL rMgP1were separated: ∼3 μg of the MgCK2α fraction (DE0/PC500) after P-cellulose and heparin–agarose chromatography (lane 1), 0.5 μg ofrecombinant S. cerevisiae CK2α (lane 2), 0.4 μg of recombinanthuman CK2α (lane 3), and ∼22 μg of the S-100 fraction (lane 4) fromgill extracts. Separated proteins were treated with guanidine chlorideand allowed to renature overnight. The renatured proteins wereincubated with 2.5 mL of reaction buffer containing [γ-32P]ATP asdescribed in “Materials and Methods”. The dried gel was autoradio-graphed. Arrows indicate the positions of protein markers
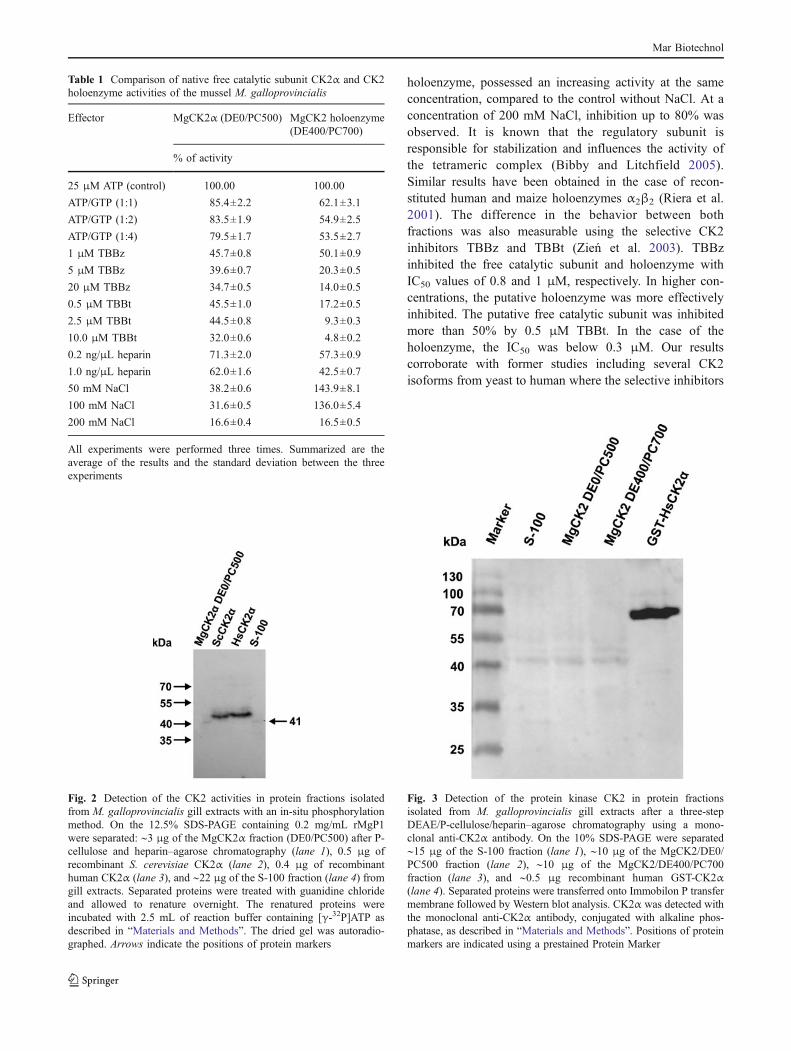
Fig. 3 Detection of the protein kinase CK2 in protein fractionsisolated from M. galloprovincialis gill extracts after a three-stepDEAE/P-cellulose/heparin–agarose chromatography using a mono-clonal anti-CK2α antibody. On the 10% SDS-PAGE were separated∼15 μg of the S-100 fraction (lane 1), ∼10 μg of the MgCK2/DE0/PC500 fraction (lane 2), ∼10 μg of the MgCK2/DE400/PC700fraction (lane 3), and ∼0.5 μg recombinant human GST-CK2α(lane 4). Separated proteins were transferred onto Immobilon P transfermembrane followed by Western blot analysis. CK2α was detected withthe monoclonal anti-CK2α antibody, conjugated with alkaline phos-phatase, as described in “Materials and Methods”. Positions of proteinmarkers are indicated using a prestained Protein Marker
Mar Biotechnol

TBBt and TBBz showed strong inhibitory effects with IC50
values at 1 μM or even below (Olsen et al. 2008; Guerra etal. 1999). Using heparin, a well-known modulator of CK2activity (Wojda et al. 1999), the presence of 1 ng/μL
inhibited almost 40% of the DE0/PC500 catalytic subunitactivity and 60% of the putative holoenzyme activity. Thesevalues are much lower than those known for the human ormaize holoenzyme (Riera et al. 2001). Protein kinase CK2is one of the few kinases which are able to utilize both ATPand GTP as co-substrate (Gschwendt et al. 1995; owninvestigations). A comparison of the Km values for ATPobtained for both CK2α and the holoenzyme showedtypical values. Whereas the Km of CK2α was about14 μM, that of the holoenzyme lied at about 8 μM. Theinfluence of several GTP concentrations was examinedusing non-labeled GTP as counterpart for ATP, as describedin the standard procedure. GTP was added in the same, indouble, and in quadruple concentrations with ATP. In thecase of the free catalytic subunit, the competitive effect wasrather low, with only 20% inhibition at the highestconcentration, suggesting that ATP is preferred as phos-
Table 2 Comparison of M. galloprovincialis MgCK2α and MgCK2βpredicted amino acid sequences with homologue sequences from B.mori, H. sapiens, C. intestinalis, and Danio rerio
MgCK2α MgCK2β
Identity(%)
Similarity(%)
Identity(%)
Similarity(%)
Bombyx mori 85 96 83 97
Homo sapiens 85 96 92 97
Ciona intestinalis 82 96 90 99
Danio rerio 84 95 92 98
Fig. 4 Alignment of thepredicted amino acid sequenceof M. galloprovincialis (Mg)CK2α catalytic subunit withhomologue sequences of B. mori(Bm—AB206394), H. sapiens(Hs—BT019792), D. rerio(Dr—BC165881), andC. intestinalis (Ci—AY092081).The gaps required for alignmentare indicated as dashes. TheATP binding motif (GXGXXS)is shaded, amino acids forputative nuclear localizationsignal (PVKKKKI) are boxed,while the most highly conservedamino acid motif D173WG175,specific to members of the CK2family, and G 197PE199, acommon feature of the family,are marked with an arrow.Identical amino acids areindicated by an asterisk,strongly conserved by a colon,and weakly conserved by a dot
Mar Biotechnol

phate donor. On the contrary, the holoenzyme was up to40% blocked at equal ATP and GTP concentrations.
Further evidence for the existence of CK2 activity inboth enzyme preparations was gained by an in-situphosphorylation assay. In this experiment, the ribosomalprotein MgP1 was co-polymerized with polyacrylamide, asdescribed in “Materials and Methods”. After electrophoresis,renaturation, and phosphorylation reactions, the proteinband corresponding to the CK2 catalytic subunit could bedetected by autoradiography. Human and yeast CK2α wereincluded as positive controls. As seen in Fig. 2, both theS-100 and the DE0/PC500 fractions contained a kinasewith MW of about 41 kDa. This method indicates that theputative CK2 catalytic subunit has a MW over 40 kDa,similar to the human and yeast enzymes. In a secondexperiment, anti-CK2α antibodies were used to detect theCK2α subunit in the S-100, DE0/PC500, and DE400/PC700 fractions. Human CK2α was used again as positivecontrol. As seen in Fig. 3, a band of around 41 kDa wasdetected by the anti-CK2α antibody in all three fractions.The results described above reinforce the hypothesis thatthe protein kinase CK2 of M. galloprovincialis has beendetected and partially purified. S-100, DE0/PC500, andDE400/PC700 fractions were also investigated for thepresence of CK2β using anti-CK2β antibodies. The CK2βsubunit was detected only in the S-100 and the DE400/PC700 fractions as a band of approximately 26 kDa and
was absent in the DE0/PC500 fraction as expected (datanot shown). This emphasizes our former results concerningthe free catalytic subunit and the holoenzyme.
Construction and Analysis of the M. galloprovincialisCK2α and CK2β cDNAs
The cDNA molecules of the catalytic CK2α and theregulatory CK2β subunits of M. galloprovincialis wereconstructed by the 5′ and 3′ RACE and RT-PCR methods,respectively, as described in “Materials and Methods”. ΤheCK2α cDNA is 1,071-bp long encoding for a predictedpolypeptide of 356 amino acids, while the CK2β cDNA is666-bp long encoding for a predicted polypeptide of 221amino acids. Comparison of the CK2β cDNA sequence toone published EST of M. galloprovincialis, FL594940(Venier et al. 2009), revealed 98% homology, with ninenucleotide changes (157 from A to G, 294 from T to C, 312from A to G, 315 from C to T, 438 from A to C, 441 fromG to A, 552 from C to T, 546 from G to A, and 567 from Ato G). These changes did not alter the encoded amino acidsequence, except for the residue 53 that switched from I toV. This change does not possibly affect the regulatorysubunit as it is not included in a regulatory motif. Thesedifferences may be due to DNA polymorphism rather thanto artificial mutations during PCR amplification sinceproofreading polymerase was used.
Fig. 5 Alignment of the predicted amino acid sequence of M.galloprovincialis (Mg) regulatory subunit CK2β with homologuesequences of B. mori (Bm—AB206395), H. sapiens (Hs—NM_001320), D. rerio (Dr—S76877), and C. intestinalis (Ci—AF360544). The gaps required for alignment are indicated as dashes.The cysteine residues thought to hold the zinc finger in place areindicated in the text with arrowheads. The potential autophosphor-
ylation site of MgCK2β (SSSEE) is in bold. Residues E20-K33
responsible for the export of CK2 as an ectokinase are boxed. Theacidic stretch D55-D64, responsible for down-regulating the CK2activity, is also boxed. Serine residues near the C terminus that couldbe phosphorylated are shaded. Identical amino acids are indicated byan asterisk, strongly conserved by colon, and weakly conserved by adot
Mar Biotechnol

Alignments of the predicted amino acid sequence of theCK2α and CK2β subunits of M. galloprovincialis with thecorresponding sequences of Homo sapiens (Hs), Drosophilamelanogaster (Dm), B. mori (Bm), and Ciona intestinalis(Ci) showed considerable similarity, as presented in Table 2.The primary structure of the predicted MgCK2α proteinpresents the characteristic subdomains and the conservedfeatures of a typical protein kinase CK2 (Fig. 4): (1) theATP binding motif G44XGXXS49, (2) a lysine-rich segment70PVKKKKIR77, responsible for the recognition of substratedeterminant, nuclear localization, and interaction with hsp90and heparin, (3) a T125LTDYD130 autophosphorylationsegment, (4) residues R153, D154, and H158, importantelements of the catalytic loop, (5) conserved sequencesD173WG175 and G197PE199, responsible for the formation ofCK2α activation loop, and (6) conservative residues R189,R193, and K196 of the C-terminal loop, responsible for therecognition of acidic residue at position n+1 in the phos-phoacceptor sequence (Hanks and Quinn 1991; Allende andAllende 1995; Martel et al. 2001).
The primary structure of MgCK2β subunit contains alsoall typical conserved features characteristic for CK2βsubunits (Fig. 5): (1) S2S3SEE main phosphorylation sites,(2) aa 5–20, Nopp140 interaction site, (3) the highlyconserved E20-K33 amino acids used for export of CK2 asan ectokinase, (4) aa 46–54 destruction box, (5) aa 55–64acidic loop responsible for down regulation of CK2activity, (6) amino acids 106–116, 124, 134, 141, 145–149, and 152 involved in the interaction with cell cycleregulators p21 and p53, (7) aa 143–148, dimer interfaceresidues, (8) aa 187–192, region containing CK2α bindingresidues, (9) aa 187–205, responsible for binding A-Rafand Mos, (10) four cysteine residues, C109, C114, C137, andC140, responsible for zinc finger formation, and (11)
proximal to the C terminus serine residues, phosphorylatedby p34cdc2 in vitro (Soderling 1990; Litchfield et al. 1991;Boldyreff et al. 1993; Meggio et al. 1995; Kobe et al. 1997;Sarno et al. 2000; Bolanos-Garcia et al. 2006).
Purification of the Recombinant M. galloprovincialis CK2αand CK2β Proteins After Expression in E. coli
The purification of the recombinant proteins MgCK2α andMgCK2β was done as described in “Materials andMethods”. As shown in Fig. 6a, it is obvious that therecombinant proteins MgCK2α and CK2β are homogenousafter the two purification steps (affinity and ion exchangechromatography).
Fig. 7 Phosphorylation of rMgP1 and rMgCK2β proteins byrMgCK2α. About 3 μg of pure recombinant MgP1 (lanes 1 and 2)were phosphorylated by 0.2 μg of recombinant MgCK2α in theabsence (lane 1) and the presence (lane 2) of 0.5 μg pure recombinantMgCK2β subunit. Lane 3 shows the phosphorylation of MgCK2βalone by rMgCK2α. Phosphorylated proteins were resolved in 12.5%SDS-PAGE followed by coomassie blue staining and autoradiography.Arrows show the positions of protein markers
Fig. 6 Detection of purified rCK2α and rCK2β proteins of M.galloprovincialis. On the 10% SDS-PAGE were separated ∼2 μg ofrMgCK2β (lane 2) and ∼2 μg of rMgCK2α (lane 3). Separatedproteins were stained with coomassie (a) or transferred ontoImmobilon P transfer membrane followed by Western blot analysis.
Positions of the CK2 subunits were detected with anti-His (b) or anti-CK2α and anti-CK2β (c) antibodies conjugated with alkalinephosphatase as described in “Materials and Methods”. Molecularweights of protein markers are indicated using a prestained proteinmarker
Mar Biotechnol

Properties of the Recombinant CK2α and CK2β Proteinsof M. galloprovincialis
The identity of the recombinant MgCK2α and MgCK2βproteins was confirmed by immunoblotting using themonoclonal anti-His antibody (Fig. 6b) as well as themonoclonal anti-CK2α and anti-CK2β antibodies (Fig. 6c).Both proteins were detected at expected molecular weights,MgCK2α at about 42 kDa and MgCK2β at 29 kDa. Thecalculated MWs from the predicted amino acid sequenceswith the additional 6xHis residues were 42.8 and 26.4 kDa,respectively. Taking into account several further aspectswhich may influence the migration of proteins on SDS-PAGE, as well as the insufficient reliability of the proteinstandards, the statistical values are confirmed.
M. galloprovincialis recombinant CK2α and CK2βproteins were also examined for their functionality. In astandard phosphorylation assay, as described in “Materialsand Methods”, rMgCK2α was incubated in the presence ofrMgP1 and/or rMgCK2β. Figure 7 shows the phosphory-lation of rMgP1 alone or in the presence of rMgCK2βsubunit. rMgP1 phosphorylation was increased by theaddition of the regulatory rMgCK2β subunit. Due to thepresence of the consensus sequence in the regulatoryrMgCK2β subunit, its autophosphorylation could also bedetected. The observed CK2β stimulating effect is notobvious for all protein substrates. Calmodulin has beenused to discriminate between CK2 free catalytic subunitand holoenzyme, as it is phosphorylated by α-subunits, butno phosphorylation is detected in the presence of β-subunit(Bidwai et al. 1993; Marin et al. 1999).
Conclusions
The results obtained by our investigations showed for thefirst time in marine invertebrates the isolation and partialpurification of the protein kinase CK2 from the gills ofthe mussel M. galloprovincialis. Active fractions contain-ing the CK2α catalytic subunit and the holoenzyme wereisolated by DEAE-cellulose, P-cellulose, and heparin–agarose chromatography. The properties of both isoformswere examined in phosphorylation assays, using as asubstrate M. galloprovincialis purified recombinant ribo-somal protein rMgP1 (Kolaiti et al. 2009). Both enzymaticfractions were tested in the presence of several knownmodulators of the CK2 activity, like GTP, heparin, andATP-competitive inhibitors (TBBz and TBBt) (Wojda et al.1999; Zień et al. 2003; Riera et al. 2001). The experimentalevidence led to the conclusion that the M. galloprovincialisCK2α catalytic subunit was obtained in the DE0/PC500fraction and the holoenzyme in the DE400/PC700 frac-tion. Additionally, the isolated CK2 fraction showedsimilar MW in both, an in-situ phosphorylation assay
and an immuno-detection assay with a specific anti-CK2α antibody.
For a better understanding of the properties andregulatory mechanisms of CK2α and the holoenzyme, weconstructed the cDNAs of MgCK2α and MgCK2βsubunits. Both proteins were over-expressed in E. coli andpurified to homogeneity. Analysis of their predicted aminoacid sequences showed that the MgCK2α catalytic subunitpresents the characteristic subdomains and the conservedfeatures of a typical protein kinase CK2 (Hanks and Quinn1991; Sarno et al. 1996), similar to the regulatory MgCK2βsubunit that also presents the conserved features of CK2βsubunits (Litchfield et al. 1991; Boldyreff et al. 1993;Meggio et al. 1995; Kobe et al. 1997).
These promising results need further investigation, andour future plans are to analyze in depth the properties of theenzyme subunits at normal and under stress conditions.Protein kinase CK2 is known to be upregulated in severaldiseases or by oxidative stress (Gerber et al. 2000). Bivalvemolluscs, like the sea mussel M. galloprovincialis, are usedas biomarkers in the assessment of environmental quality asthey are able to accumulate in their tissues a wide range ofpollutants (Gourgou et al. 2010; Torres et al. 2002).Therefore, the sea mussel M. galloprovincialis is anexcellent candidate for the investigation of environmentalstress impact on the expression of protein kinase CK2.
Acknowledgements The authors are grateful to Dr. Y. Drosos,Miss A. Sielouvari, and Dr. K. Kubiński for scientific help anddiscussion. 4,5,6,7-Tetrabromo-1H-benzotriazole (TBBt) and 4,5,6,7-tetrabromo-1H-benzimidazole (TBBz) were generous gifts of ZygmuntKazimierczuk (Department of Experimental Pharmacology, PolishAcademy of Sciences, Medical Research Center, Warsaw, Poland). Thiswork was supported by the University of Athens, the Special Account forResearch Grants of Athens University (SARG, 70/4/7803 to SK), and theGreek National Fellowship Foundation (IKY) (fellowship to RMK).
References
Abramczyk O, Zień P, Zieliński R, Pilecki M, Hellman U, Szyszka R(2003) The protein kinase 60S is a free catalytic CK2α′ subunitand form an inactive complex with superoxide dismutase SOD1.Biochem Biophys Res Commun 307:31–40
Ahmad KA, Wang G, Unger G, Slaton J, Ahmed K (2008) Proteinkinase CK2—a key suppressor of apoptosis. Adv Enzyme Regul48:179–187
Allende JE, Allende CC (1995) Protein kinase CK2: an enzyme withmultiple substrates and a puzzling regulation. FASEB J 9:313–323
Ballesta JP, Rodriguez-Gabriel MA, Bou G, Briones E, Zambrano R,Remacha M (1999) Phosphorylation of the yeast ribosomal stalk.Functional effects and enzymes involved in the process. FEMSMicrobiol Rev 23:537–550
Ballesta JPG, Guarinos E, Zurdo J, Parada P, Nusspaumer G, LaliotiVS, Perez-Fernandez J, Remacha M (2000) Structure of the yeastribosomal stalk. In: Garrett R, Douthwaite S, Matheson A, LiljasA, Moore PB, Noller HF (eds) The ribosome structure, function,
Mar Biotechnol

antibiotics and cellular interactions. American Society forMicrobiology Press, Washington, pp 115–125
Bardales JR, Hellman U, Villamarin JA (2007) CK2-mediatedphosphorylation of a type II regulatory subunit of cAMP-dependentprotein kinase from the mollusk Mytilus galloprovincialis. ArchBiochem Biophys 461:130–137
Bibby AC, Litchfield DW (2005) The multiple personalities of theregulatory subunit of protein kinase CK2: CK2 dependent andCK2 independent roles reveal a secret identity for CK2β. Int JBiol Sci 1:67–79
Bidwai AP, Reed JC, Glover CVC (1993) The phosphorylation ofcalmodulin by the catalytic subunit of casein kinase II is inhibitedby the regulatory subunit. Arch Biochem Biophys 300:265–270
Bolanos-Garcia VM, Fernandez-Recio J, Allende JE, Blundell TL(2006) Identifying interaction motifs in CK2β—a ubiquitouskinase regulatory subunit. Trends Biochem Sci 31:654–661
Boldyreff B, James P, Staudenmann W, Issinger O-G (1993) Ser2 is theautophosphorylation site in the beta subunit from bicistronicallyexpressed human casein kinase-2 and from native rat liver caseinkinase-2 beta. Eur J Biochem 218:515–521
Bosc DG, Graham KC, Saulnier RB, Zhang C, Prober D, Gietz RD,Litchfield DW (2000) Identification and characterization of CKIP-1,a novel pleckstrin homology domain-containing protein that interactswith protein kinase CK2. J Biol Chem 275:14295–14306
Bou G, Remacha M, Ballesta JP (2000) Ribosomal stalk proteinphosphorylating activities in Saccharomyces cerevisiae. ArchBiochem Biophys 375:83–89
Filhol O, Cochet C (2009) Cellular functions of protein kinase CK2: adynamic affair. Cell Mol Life Sci 66:1830–1839
Gagou ME, Rodriguez-Gabriel MA, Ballesta JP, Kouyanou S (1999)Isolation and expression of the genes encoding the acidicribosomal phosphoproteins P1 and P2 of the medfly Ceratitiscapitata. Gene 226:365–373
Gagou M, Ballesta JP, Kouyanou S (2000) Cloning and characteriza-tion of the ribosomal protein CcP0 of the medfly Ceratitiscapitata. Insect Mol Biol 9:47–55
Gerber DA, Souquere-Besse S, Puvion F, Dubois MF, Bensaude O,Cochet C (2000) Heat-induced relocalization of protein kinaseCK2. Implication of CK2 in the context of cellular stress. J BiolChem 275:23919–23926
Geret F, Cosson RP (2002) Induction of specific isoforms ofmetallothionein in mussel tissues after exposure to cadmium ormercury. Arch Environ Contam Toxicol 42:36–42
Glover CVC (1998) On the physiological role of casein kinase II inSaccharomyces cerevisiae. Prog Nucleic Acid Res Mol Biol59:95–133
Gourgou E, Aggeli IK, Beis I, Gaitanaki C (2010) Hyperthermia-induced Hsp70 and MT20 transcriptional upregulation aremediated by p38-MAPK and JNKs in Mytilus galloprovincialis(Lamarck); a pro-survival response. J Exp Biol 213:347–357
Gschwendt M, Kittstein W, Kielbassa K, Marks F (1995) Proteinkinase C delta accepts GTP for autophosphorylation. BiochemBiophys Res Commun 206:614–620
Guerra B, Boldryff B, Sarno S, Cesaro L, Issinger OG, Pinna LA(1999) CK2: a protein kinase in need of control. Pharmacol Ther82:303–313
Hagger JA, Atienzar FA, Jha AN (2005) Genotoxic, cytotoxic,developmental and survival effects of tritiated water in the earlylife stages of the marine mollusc, Mytilus edulis. Aquat Toxicol74:205–217
Hanks SK, Quinn AM (1991) Protein kinase catalytic domainsequence database: identification of conserved features ofprimary structure and classification of family members. MethodsEnzymol 200:38–62
Hilgard P, Czaja MJ, Gerken G, Stockert RJ (2004) Proapoptoticfunction of protein kinase CK2alpha′′ is mediated by a JNK
signaling cascade. Am J Physiol Gastrointest Liver Physiol287:192–201
Kalpaxis DL, Theos C, Xaplanteri MA, Dinos GP, Catsiki AV,Leotsinidis M (2004) Biomonitoring of Gulf of Patras, N.Peloponnesus, Greece. Application of a biomarker suite includingevaluation of translation efficiency in Mytilus galloprovincialiscells. Environ Res 94:211–220
Kameshita I, Fujisawa H (1989) A sensitive method for detection ofcalmodulin-dependent protein kinase II activity in sodiumdodecyl sulfate-polyacrylamide gel. Anal Biochem 183:139–143
Kobe B, Heierhorst J, Kemp BE (1997) Intrasteric regulation ofprotein kinases. Adv Second Messenger Phosphoprotein Res31:29–40
Kolaiti RM, Lucas JM, Kouyanou-Koutsoukou S (2009) Molecularcloning of the ribosomal P-proteins MgP1, MgP2, MgP0, andsuperoxide dismutase (SOD) in the mussel Mytilus galloprovin-cialis and analysis of MgP0 at stress conditions. Gene 430:77–85
Koumarianou P, Marcos AG, Ballesta JP, Kouyanou-Koutsoukou S(2007) In vivo analysis of the acidic ribosomal proteins BmP1and BmP2 of the silkworm Bombyx mori in the yeastSaccharomyces cerevisiae. Gene 388:27–33
Leroy D, Alghisi GC, Roberts E, Filhol-Cochet O, Gasser SM (1999)Mutations in the C-terminal domain of topoisomerase II affectmeiotic function and interaction with the casein kinase 2 betasubunit. Mol Cell Biochem 191:85–95
Litchfield DW (2003) Protein kinase CK2: structure, regulation androle in cellular decisions of life and death. Biochem J 369:1–15
Litchfield DW, Lozeman FJ, Cicirelli MF, Harrylock M, Ericsson LH,Piening CJ, Krebs EG (1991) Phosphorylation of the beta subunitof casein kinase II in human A431 cells: identification of theautophosphorylation site and a site phosphorylated by p34cdc2. JBiol Chem 266:20380–20389
Liu CC, Lu TC, Li HH, Wang HX, Liu GF, Ma L, Yang CP, Wang BC(2010) Phosphoproteomic identification and phylogenetic analy-sis of ribosomal P-proteins in Populus dormant terminal buds.Planta 231:571–581
Lozeman FJ, Litchfield DW, Piening C, Takio K, Walsh KA, KrebsEG (1990) Isolation and characterization of human cDNA clonesencoding the α and α´ subunits of casein kinase II. Biochemistry29:8436–8447
Maridor G, Park W, Krek W, Nigg EA (1991) Casein kinase II. cDNAsequences, developmental expression and tissue distribution ofmRNAs for α, α´ and β subunits of the chicken enzyme. J BiolChem 266:2362–2368
Marin O, Meggio F, Pinna LA (1999) Structural features underlyingthe unusual mode of calmodulin phosphorylation by proteinkinase CK2: a study with synthetic calmodulin fragments.Biochem Biophys Res Commun 256:442–446
Martel V, Filhol O, Nueda A, Gerber D, Benitez MJ, Cochet C (2001)Visualization and molecular analysis of nuclear import of proteinkinase CK2 subunits in living cells. Mol Cell Biochem 227:81–90
Meggio F, Boldyreff B, Marin O, Issinger O-G, Pinna LA (1995)Phosphorylation and activation of protein kinase CK2 byp34cdc2 are independent events. Eur J Biochem 230:1025–1031
Nusspaumer G, Remacha M, Ballesta JP (2000) Phosphorylation andN-terminal region of yeast ribosomal protein P1 mediate itsdegradation, which is prevented by protein P2. EMBO J 19:6075–6084
Olsen BB, Rasmussen T, Niefind K, Issinger O-G (2008) Biochemicalcharacterization of CK2α and α′ paralogues and their derivedholoenzymes: evidence for the existence of a heteroteramericCK2α′-holoenzyme forming trimeric complexes. Mol Cell Bio-chem 316:37–47
Pilecki M, Grzyb A, Zień P, Sekuła A, Szyszka R (2000) Yeast proteinphosphatase active with acidic ribosomal proteins. J BasicMicrobiol 40:1–10
Mar Biotechnol

Pinna LA (1997) Protein kinase CK2. Int J Biochem Cell Biol29:551–554
Pinna LA (2003) The raison d’être of constitutively active proteinkinases: the lesson of CK2. Acc Chem Res 36:378–384
Pytharopoulou S, Kouvela EC, Sazakli E, Leotsinidis M, Kalpaxis DL(2006) Evaluation of the global protein synthesis in Mytilusgalloprovincialis in marine pollution monitoring: seasonal vari-ability and correlations with other biomarkers. Aquat Toxicol80:33–41
Riera M, Peracchia G, de Nadal E, Ariño J, Pages M (2001) Maizeprotein kinase CK2: regulation and functionality of three betaregulatory subunits. Plant J 25:365–374
Sajnaga E, Kubiński K, Szyszka R (2008) Catalytic activity of mutantsof yeast protein kinase CK2α. Acta Biochim Pol 55:767–776
Sarno S, Vaglio P, Meggio F, Issinger OG, Pinna LA (1996) Proteinkinase CK2 mutants defective in substrate recognition. Purifica-tion and kinetic analysis. J Biol Chem 271:10595–10601
Sarno S,Marin O, Boschetti M, PaganoMA,Meggio F, Pinna LA (2000)Cooperative modulation of protein kinase CK2 by separate domainsof its regulatory beta-subunit. Biochemistry 39:12324–12329
Shi X, Potvin B, Huang T, Hilgard P, Spray DC, Suadicani SO,Wolkoff AW, Stanley P, Stockert RJ (2001) A novel casein kinase2 alpha-subunit regulates membrane protein traffic in the humanhepatoma cell line HuH-7. J Biol Chem 276:2075–2082
Soderling TR (1990) Protein kinases. Regulation by autoinhibitorydomains. J Biol Chem 265:1823–1826
Szick-Miranda K, Bailey-Serres J (2001) Regulated heterogeneity in12-kDa P-protein phosphorylation and composition of ribosomesin maize (Zea mays L.). J Biol Chem 276:10921–10928
Tchórzewski M, Boguszewska A, Dukowski P, Grankowski N (2000)Oligomerization properties of the acidic ribosomal P-proteinsfrom Saccharomyces cerevisiae: effect of P1A protein phosphor-
ylation on the formation of the P1A–P2B hetero-complex.Biochim Biophys Acta 1499:63–73
Torres MA, Testa CP, Gáspari C, Masutti MB, Panitz CM, Curi-Pedrosa R, de Almeida EA, Di Mascio P, Filho DW (2002)Oxidative stress in the mussel Mytella guyanensis from pollutedmangroves on Santa Catarina Island, Brazil. Mar Pollut Bull44:923–932
Venier P, De Pittà C, Bernante F, Varotto L, De Nardi B, Bovo G, Roch P,Novoa B, Figueras A, Pallavicini A, Lanfranchi G (2009)MytiBase:a knowledgebase of mussel (M. galloprovincialis) transcribedsequences. BMC Genomics 10:72. doi:10.1186/1471-2164-10-72
Wang HC, Erikson RL (1992) Activation of protein serine/threoninekinases p42, p63, and p87 in Rous sarcoma virus-transformedcells: signal transduction/transformation-dependent MBP kinases.Mol Biol Cell 3:1329–1337
Wojda I, Cytryńska M, Frajnt M, Jakubowicz T (1999) Phosphory-lation of yeast ribosomal proteins by CKI and CKII in thepresence of heparin. Acta Biochim Pol 46:211–215
Wojda I, Cytryńska M, Frajnt M, Jakubowicz T (2002) Protein kinasesCKI and CKII are implicated in modification of ribosomal proteinsof the yeast Trichosporon cutaneum. Acta Biochim Pol 49:947–957
Xu X, Toselli PA, Russell LD, Seldin DC (1999) Globozoospermia inmice lacking the casein kinase II alpha’ catalytic subunit. NatGenet 23:118–121
Zieliński R, Pilecki M, Kubiński K, Zień P, Hellman U, Szyszka R(2002) Inhibition of yeast ribosomal stalk phosphorylation byCu–Zn superoxide dismutase. Biochem Biophys Res Commun296:1310–1316
Zień P, Abramczyk O, Domańska K, Bretner M, Szyszka R (2003)TBBz but not TBBt discriminates between two molecular formsof CK2 in vivo and its implications. Biochem Biophys ResCommun 312:623–628
Mar Biotechnol
Related Documents











