1 Involvement of histone acetylation in ovarian steroid-induced decidualization of human endometrial stromal cells* Nozomi Sakai, Tetsuo Maruyama‡, Rei Sakurai, Hirotaka Masuda, Yurie Yamamoto, Aki Shimizu, Ikuko Kishi, Hironori Asada, Satoshi Yamagoe§, Yasunori Yoshimura From Department of Obstetrics and Gynecology, School of Medicine, Keio University, Shinanomach 35, Shinjuku-ku, Tokyo 160-8582 and the §Department of Bioactive Molecules, National Institute of Infectious Diseases, 1-23-1 Toyama, Shinjuku-ku, Tokyo 162-8640, Japan. ‡To whom correspondence should be addressed: Department of Obstetrics and Gynecology, School of Medicine, Keio University, 35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan. Tel: +81-3-3353-1211, ext 62916; Fax: +81-3-3226-1667; E-mail: [email protected] Running title: Histone acetylation in decidualization Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc. JBC Papers in Press. Published on February 27, 2003 as Manuscript M211715200 by guest on July 5, 2019 http://www.jbc.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1
Involvement of histone acetylation in ovarian steroid-induced
decidualization of human endometrial stromal cells*
Nozomi Sakai, Tetsuo Maruyama‡, Rei Sakurai, Hirotaka Masuda,
Yurie Yamamoto, Aki Shimizu, Ikuko Kishi, Hironori Asada,
Satoshi Yamagoe§, Yasunori Yoshimura
From Department of Obstetrics and Gynecology, School of Medicine, Keio
University, Shinanomach 35, Shinjuku-ku, Tokyo 160-8582 and the
§Department of Bioactive Molecules, National Institute of Infectious Diseases,
1-23-1 Toyama, Shinjuku-ku, Tokyo 162-8640, Japan.
‡To whom correspondence should be addressed:
Department of Obstetrics and Gynecology,
School of Medicine, Keio University,
35 Shinanomachi, Shinjuku-ku, Tokyo 160-8582, Japan.
Tel: +81-3-3353-1211, ext 62916; Fax: +81-3-3226-1667;
E-mail: [email protected]
Running title: Histone acetylation in decidualization
Copyright 2003 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on February 27, 2003 as Manuscript M211715200 by guest on July 5, 2019
http://ww
w.jbc.org/
Dow
nloaded from

2
Summary
Histone acetyltransferases and histone deacetylases (HDACs) determine the
acetylation status of histones, regulating gene transcription. Decidualization is the
progestin-induced differentiation of estrogen-primed endometrial stromal cells (ESC),
which is crucial for implantation and maintenance of pregnancy. We here show that
trichostatin A (TSA), a specific HDAC inhibitor, enhances the up-regulation of
decidualization markers such as insulin-like growth factor binding protein-1 (IGFBP-1)
and prolactin in a dose-dependent manner that is directed by 17β-estradiol (E2) plus
progesterone (P4) in cultured ESC, but not glandular cells, both isolated from human
endometrium. Morphological changes resembling decidual transformation were also
augmented by co-addition of TSA. Acid urea triton gel analysis and immunoblot using
acetylated histone type-specific antibodies demonstrated that treatment with E2+P4
significantly increased the levels of acetylated H3 and H4 whose increment was
augmented by co-treatment with TSA. Chromatin immunoprecipitation assay revealed that
treatment with E2+P4 increased the amount of proximal progesterone-responsive region of
IGFBP-1 promoter associated with acetylated H4, which was dramatically enhanced by
co-addition of TSA. Taken together, our results suggest that histone acetylation is deeply
involved in differentiation of human ESC and that TSA has a potential as an enhancer of
decidualization through promotion of progesterone action.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

3
Introduction
Decidualization is the progestin-induced differentiation of fibroblast-like stromal
cells of the proliferative estrogen-primed endometrium into decidual cells that are easily
distinguishable histologically as the larger and rounder cells appearing around the spiral
arteries and eventually spreading through the most part of the endometrium in the late
luteal phase of the menstrual cycle (1). Following embryo implantation, decidualization
persists and extends throughout the endometrium, leading to the formation of the
pregnancy decidua. This morphological change is accompanied by the biochemical
expression of a number of bioactive substances (2) that in turn act as local regulators of
both decidual and placental functions (3,4). Thus, decidualization is critical for embryo
implantation and maintenance of pregnancy.
In the presence of estrogen and progestin, endometrial stromal cells (ESC)1
isolated from human cycling endometrium can exhibit morphological and functional
changes in vitro that mimic in vivo decidual transformation (5,6). With development of
this in vitro model, many studies have addressed the molecular mechanisms underlying
decidualization. Increasing bodies of evidence indicates that a number of growth factors,
cytokines, proteinases for extracellular matrices, and peptide hormones are produced by
decidualized ESC (2-4). As the cause or consequence, decidua-specific signaling
pathways involving signaling molecules such as cAMP-protein kinase A (7-9), c-Src
tyrosine kinase (10-12), and STATs (13,14) become activated during decidualization.
Recent microarray technologies have explosively accelerated identification of dynamically
regulated genes in the process of decidualization (15,16). However, little is known about
the molecular events involved in the regulation of transcription and expression of those
decidualization-associated genes.
A key event in the regulation of eukaryotic gene expression is the
posttranslational modification of nucleosomal histones, which converts regions of
chromosomes into transcriptionally active or inactive chromatin. The protein building
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

4
blocks of the nucleosome are the core histones H2A, H2B, H3, and H4. The amino-
terminal tails of these histones house the sites for acetylation, methylation,
phosphorylation, and ADP-ribosylation. Among these well-known covalent modifications
of histones, the most well studied is the acetylation of ε-amino groups on conserved
lysine residues in the histones’ amino-terminal tail domains (17). There are two classes of
enzymes involved in determining the state of acetylation of histones: histone acetyl
transferases (HATs) and histone deacetylases (HDACs) that have been identified in the
past several years (17), coinciding with the discovery of specific HDAC inhibitors such
as trapoxin and trichostatin A (TSA) (18). These inhibitors increase acetylated histones in
many cell types, thereby inducing expression of specific pre-programmed genes, which,
in turn, lead to cell growth arrest, differentiation, and apoptotic cell death (19). Along
with availability of these materials, recent dramatic advances highlight the involvement of
HATs in transcriptional activation and HDACs in transcriptional repression (17).
However, there has been no report on histone acetylation in decidualization except a
single study demonstrating in 1979 that histones H2B and H4 become acetylated
immediately upon in vivo decidualization in rat (20).
In this study, we have investigated the effect of TSA on ESC in terms of ovarian
steroid-induced differentiation induction to address how histone acetylation is involved in
this process. We here show that TSA advances ovarian steroid-induced decidualization of
ESC isolated from human cycling endometrium. Histones H3 and H4 became acetylated
upon decidualization and the acetylated H4 was associated with the ovarian steroid-
induced promoter activation of IGFBP-1, a typical decidualization marker gene, both of
which were augmented by co-treatment with TSA. These results provide evidence
suggesting that histone acetylation is deeply involved in the process of decidualization of
human ESC.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

5
Experimental procedures
Reagents—TSA was obtained from Wako Bio-chemicals (Osaka, Japan) and
dissolved in ethanol. An antibody against histone H3 acetylated at both lysine residue 9
(K9) and K14 and anti-acetylated histone H4 antibodies specifically reacting with
acetylated K5, K8, K12, or K16 were purchased from Upstate Biotechnology Inc (Lake
Placid, NY). Antibodies recognizing histone H2A acetylated at K5 or H2B acetylated at
K5, K12, K15 and K20 were obtained from Chemicon International Inc (Temecula, CA).
All the oligonucleotides were synthesized by Invitrogen (Tokyo, Japan).
Preparation and primary cultures of human endometrial stromal cells (ESC) and
endometrial glandular cells (EGC)—Endometrial specimens were obtained from
consenting patients undergoing endometrial biopsies or total abdominal hysterectomy for
benign gynecological disease. The use of these human specimens was approved by the
Keio University Ethics Committee. ESC were isolated from human cycling endometria as
previously described (21). In brief, tissue samples were washed with Dulbecco’s
Modified Eagles’ Medium (DMEM) and minced into small pieces of less than 1 mm3. The
tissues were then incubated for 2 hours at 37 °C in DMEM containing 0.2% (w/v)
collagenase (Wako), 0.05% DNase I (Life technologies, Gaithursberg, MD), 1%
antibiotic-antimycotic mixture (Life technologies), and 10% fetal bovine serum (FBS).
After enzymatic digestion, cell clumps were dispersed by pipetting. Most of ESC that
were present as single cells or small aggregates were strained through a 70 µm cell
strainer (Falcon 2350; Becton Dickinson. Flanklin Lakes, NJ), which allowed the ESC to
pass through while intact glands were retained. The filtrates were washed twice and
inoculated into 6-cm dishes or each well of six-well plates.
Glands were recovered from the cell strainer by backwashing with DMEM
without FBS. To disperse glands into a single cell suspension, the gland-containing
fraction was first resuspended in DMEM without FCS, spun at 800 × g for 10 min and
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

6
the pellet suspended in 3 ml of 0.25% trypsin/EDTA solution (Sigma, St.Louis, MO).
Glands were disrupted by thoroughly pippetting. The trypsin was inactivated with 3 ml of
DMEM/10%FBS. The EGC were pelleted again and resuspended in DMEM/10%FBS/1%
antibiotic-antimycotic mixture and seeded into a 6 cm dish coated with collagen type I.
The isolated ESC and EGC were pre-cultured for about two days to be grown to
subconfluence in DMEM supplemented with 10% FBS and 1% antibiotic-antimycotic
mixture. The cells were then cultured in the absence or the presence of 10 nM 17β-
estradiol (E2) plus 1 µM P4 or in combination with TSA for different periods according to
the experimental protocol.
RT-PCR—Total RNA was extracted from cell cultures using TRIzol® reagent
(Life Technologies) according to the manufacturer’s instruction. Semi-quantitative RT-
PCR was carried out with 0.3 µg of total cellular RNA using the OneStep RT-PCR kit
(Qiagen, Hilden, Germany) according to the manufacturer’s recommendations. Primers
used to amplify human IGFBP-1, prolactin (PRL), β-actin, and matrix metalloproteinase-
2 (MMP-2) were as follows: IGFBP-1, 5’-AACCTCTGCACGCCCTCACC-3’ (P1) and
5’-AGGGATCCTCTTCCCATTCCAAGGGTAGA-3’ (P2); PRL, 5’-
GCCCCCTTGCCCATCTGTCC-3’ (P4) and 5’-AGAAGCCGTTTGGTTTGCTCC-3’
(P5); β-actin, 5’-CCCAGGCACCAGGGCGTGATC-3’ and 5’-
TCAAACATGATCTGGGTCAT-3’; MMP-2, 5’-TGGGAGCATGGCGATGGATA-3’
and 5’-ACAGTGGACATGGCGGTCTCA-3’, respectively. Preliminary experiments
determined the optimum PCR cycle number within the linear range of amplification for
each gene being measured. After PCR amplification, 15 µl aliquots were electrophoresed
in 3% agarose gels, followed by photographic recording of the gels stained with ethidium
bromide. Gel photos were scanned and densitometric analyses of PCR products were
performed using the image analysis program NIH Image, version 1.62 (Research
Services Branch, National Institutes of Health).
Construction of Competitive Templates—To quantitate the mRNA level of
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

7
IGFBP-1 in ESC untreated or treated with various reagents, we adopted competitive PCR
methods (22) using competitive external standard that consisted of a shortened IGFBP-1
DNA fragment as illustrated in Figure 2A. To prepare this competitive template, a 39-bp
primer (5’-AACCTCTGCACGCCCTCACCCACGGAGATAACTGAGGAG-3’; P3, Fig.
2A) was designed to include 19 bp of the binding sequence (underlined, matching the
white region in Fig. 2A) addended to the 5' end of a sequence corresponding to bases 275
- 294 of IGFBP-1 cDNA (the black region). Thus, using P3 as the 5' primer and P2 as
the 3' primer, 343 bp of the IGFBP-1 sequence was amplified. With the appended 20 bp
of primer P3, this resulted in a 363-bp amplicon that included the P1 and P2 binding
sequences at its 5' and 3' ends, respectively (Fig. 2A). Similarly, to construct a
competitive template for PRL cDNA, 40-bp primer (5’-
GCCCCCTTGCCCATCTGTCCCGGTATACCCATGGCCGGGG-3’; P6) was
designed to contain 20 bp of the binding sequence addended to the 5' end of a sequence
corresponding to bases 79 - 98 of PRL cDNA. With this primer as the forward primer
and P5 as the reverse primer, 253 bp of the PRL sequence was amplified. With the
appended 20 bp of primer P6, this resulted in a 273-bp amplicon that included the P4 and
P5 binding sequences at its 5' and 3' ends, respectively.
Competitive RT-PCR and quantitation—Total RNA was extracted from cell
cultures using TRIzol® reagent according to the manufacturer’s instruction. RT-PCR was
performed using the OneStep RT-PCR kit (Qiagen) according to the manufacturer’s
recommendations. For each sample, serial dilutions of competitive template generated as
described above were added to 300 ng of total RNA on which reverse transcription was
performed. After the reaction for 30 min at 50 °C, the samples were heated for 15 min at
90 °C as the initial PCR activation step. PCR amplifications were then performed with a
set of primer pairs, P1 and P2, under the following conditions: 60 s at 94 °C, 60 s at
56 °C, and 90 s at 72 °C for 23 cycles, followed by 7 min at 72 °C as the last primer
extension step, generating a 446-bp product from the target cDNA and a 363-bp product
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

8
from the competitor cDNA (Figure 2A). With a set of primer pairs, P4 and P5, PCR
amplifications were performed under the following conditions: 60 s at 94 °C, 60 s at
56 °C, and 90 s at 72 °C for 26 cycles, followed by 10 min at 72 °C as the last primer
extension step, generating a 385-bp product from the target cDNA and a 273-bp product
from the competitor cDNA. PCR products separated on 3% agarose gel were visualized
by ethidium bromide staining and photographed under UV illumination. The intensity of
the bands on the image from FAS-III MINI (TOYOBO, Tokyo, Japan) was measured
with the public domain NIH Image program, version 1.62. To confirm the detection
range of competitive RT-PCR, the relationship between the amount of cDNA generated
and the initial concentrations of total RNA used were determined.
For analyzing the results, the log of the ratio of amplified target to competitor
products was graphed as a function of the known amount of competitor added to the PCR
reaction (Fig. 2C). CA-Cricket Graph III (Computer Associates) was used for the
regression analysis and calculation of x intercepts. When the log ratio equals zero, the
concentrations of the target (originally from the RT reaction) and the competitor are equal
(Fig. 2C). The amount of cDNA synthesized from 300 ng of the initial RNA sample was
calculated from the graph (Fig. 2C).
Northern blotting—Total RNA was extracted using TRIzol® reagent according
to the manufacturer’s instruction. Ten micrograms of total RNA were electrophoresed,
transferred to Maximum Strength Nytran® nylon (Schleicher & Schuell, Keene, NH) by
TurboBlotter® system (Schleicher & Schuell) as described previously (23). The filter was
hybridized with the human IGFBP-1 probe labeled by a Gene Images random prime-
labeling module and detected by a Gene Images CDP-star detection module (Amersham
Bioscineces) according to the manufacturer' s instructions. The membranes were exposed
to x-ray film for 30-60 min.
Extraction of cellular histones—Histones of Hela cells and cultured ESC were
extracted according to the procedure of Yoshida et al. (24). ESC (2 × 106) in 60-mm
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

9
culture dish were treated without or with E2+P4 in combination with TSA for 8 hours,
harvested using a rubber policeman, collected by centrifugation at 700 ∞ g for 10 min,
and washed once with phosphate-buffered saline. The washed cells were suspended in
200 µl of ice-cold lysis buffer (10 mM Tris-HCl, 50 mM sodium bisulfite, 1% Triton X-
100, 10 mM MgCl2, 8.6% sucrose, pH 6.5). After pestle homogenization, the nuclei
were collected by centrifugation at 1,000 × g for 10 min, washed three times with the cold
lysis buffer, and once with 10 mM Tris-HCl, 13 mM EDTA pH 7.4, successively. The
pellet was suspended in 100 µl of ice-cold H2O using a Vortex mixer, and concentrated
H2SO4 was added to the suspension to give a concentration of 0.4 N. After incubation at
4 °C for at least 1 hour, the suspension was centrifuged for 5 min at 15,000 rpm using a
microcentrifuge, and the supernatant was taken and mixed with 1 ml of acetone. After
overnight incubation at -20 °C, the coagulated material was collected by
microcentrifugation and air-dried. This acid-soluble histone fraction was dissolved in 30
µl of H2O. Protein was quantitated using a protein assay kit (Bio-Rad).
Acid Urea Triton (AUT) Gel Electrophoresis—Acetylation of cellular histones
from Hela cells and ESC treated without or with E2+P4 in combination with TSA was
analyzed by slab gel electrophoresis using AUT gel as described elsewhere (24). The
extracted cellular histones were incubated with the same volume of loading buffer (7.4 M
urea, 1.4 M NH3, 10 mM dithiothreitol) for 5 min and were then added with a l/8 volume
of 1% pyronine G in glacial acetic acid. The mixture was applied onto of the upper gel (1
M acetic acid, 6.3 M urea, 4.4% acrylamide) and electrophoresed in 0.2 M glycine, 1 M
acetic acid. Gels were stained with Coomassie Brilliant Blue R-250 (Bio-Rad), dried, and
photographed.
Immunoblotting—Acid-extracted histones from Hela cells treated with TSA or
ESC treated without or with E2+P4 in combination with TSA were resolved by 15% SDS-
PAGE, transferred to Immobilon-P PVDF membrane (Millipore, Bedford, MA) and
probed with acetylated histone type-specific antibodies as indicated. Proteins were
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

10
visualized using a goat anti-rabbit secondary antibody conjugated to HRP and an
enhanced chemiluminescence detection system.
Chromatin Immunoprecipitation (ChIP) assay—ChIP assays were performed
according to the protocol for the acetyl-histone H4 ChIP Assay Kit (Upstate
Biotechnology). ESC were treated without or with 500 ng/ml TSA or in combination with
E2+P4 for 60 min in 10 cm dishes. Following treatment, cells were cross-linked by
addition of formaldehyde into the medium at a final concentration of 1% and incubated for
15 min at 37 °C. Cells were washed with ice-cold phosphate-buffered saline containing
protease inhibitors and resuspended in 200 µl of ChIP Lysis buffer (1% SDS, 10 mM
EDTA, 50 mM Tris-HCl, pH 8.0, with protease inhibitors). The lysates were divided into
two tubes and sonicated utilizing an ultrasonic processor at amplitude setting 30 with five
10-s bursts followed by five 6-s bursts at amplitude setting 40. The sonicated lysates
were then diluted to 800 µl with ChIP dilution buffer (0.01% SDS, 1.1% Triton X-100,
1.2mM EDTA, 16.7mM Tris-HCl, pH 8.0, 167 mM NaCl). One ml of each sample was
precleared by incubating with 40 µl salmon sperm DNA/protein A-agarose beads for 30
min at 4 °C with rotation. Three µl of anti-acetyl histone H4 antibody (Upstate
Biotechnology, Lake Placid, NY) was added, and immunoprecipitation was done
overnight at 4 °C with rotation. Immune complexes were collected with 30 µl of salmon
sperm DNA/protein A-agarose and washed once for 5 min on a rotating platform with 1
ml each of the following buffers in sequence: Low Salt Wash Buffer (0.1% SDS, 1%
Triton X-100, 2 mM EDTA, 20 mM Tris-HCl, pH 8.0, 150 mM NaCl), High Salt Wash
Buffer (0.1% SDS, 1% Triton X-100, 2 mM EDTA, 20 mM Tris-HCl, pH 8.0, 1500
mM NaCl), LiCl Wash Buffer (250 mM LiCl, 1% Nonidet P-40, 1% sodium
deoxycholate, 1 mM EDTA, 10 mM Tris-HCl, pH 8.0) and twice with TE (10 mM Tris-
HCl, pH 8.0, 1 mM EDTA). Immune complexes were eluted, and cross-links were
reversed by heating at 65 °C for 4 hours and subjected to proteinase K treatment. DNA
was recovered by phenol/chloroform extraction followed by ethanol precipitation and was
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

11
subjected as a template for PCR amplification to detect the presence of promoter regions
immunoprecipitated with anti-acetyl histone H4 antibody. The following primers were
used for PCR analysis of immunoprecipitated promoter regions I and II as illustrated in
Figure 5A: region I Fw-(886), 5' -GGAAGGCCATGGAGGGAGTG-3' , and Rv-(619),
5' -TCTCCTCTCTGGAGGGGCAG-3' ; region II Fw-(263), 5' -
CGTCATCCCCCTCCCAGCTGAG-3' , and Rv-(33), 5' -
GCACAGGCCGCGCCACTTGCACC-3' . PCR amplifications were carried out under
the following conditions: 60 s at 94 °C, 60 s at 50 °C, and 90 s at 72 °C for 30 cycles,
preceded by 3 min at 95 °C and followed by 10 min at 72 °C. PCR products separated on
3% agarose gel were visualized by ethidium bromide staining and photographed under
UV illumination. The intensity of the bands on the image from FAS-III MINI was
measured with the public domain NIH Image program, version 1.62.
Statistical Analysis—Data were analyzed by Wilcoxon rank-sum test or Kruskal-
Wallis test followed by post hoc Dunn' s test. A p value of less than 0.05 was considered
significant. by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

12
Results
Promotion of ovarian steroid-induced decidualization of cultured ESC by
TSA—To explore the role of histone acetylation in decidualization, we examined the
effect of TSA, a selective and potent histone deacetylase inhibitor, on mRNA expression
of IGFBP-1, a typical biochemical marker of decidualization, in ESC cultured in the
absence or presence of E2+P4.
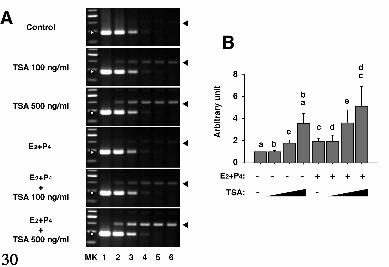
Semi-quantitative RT-PCR analysis revealed that IGFBP-1 mRNA was up-
regulated by E2+P4 in 48 hours (Fig. 1A). Co-addition with TSA provoked a further
enhanced induction whose magnitude was comparable to that obtained from decidualized
ESC treated with E2+P4 for 14 days (Fig. 1A). TSA alone was capable to induce IGFBP-
1 mRNA at a relatively high concentration (500 ng/ml) (Fig. 1A). In contrast, the levels
of β-actin mRNA expression were almost constant throughout the treatment (Fig. 1A).
Consistent with the RT-PCR data, northern blot analysis showed that TSA augmented
E2+P4 -induced IGFBP-1 mRNA expression (Fig. 1B).
PRL, another typical decidualization marker, was up-regulated by E2+P4 in 48
hours, which was further augmented by co-addition of TSA (Fig. 1C). In contrast,
neither TSA alone nor in combination with E2+P4 affected the expression of matrix
metalloproteinase II mRNA (Fig. 1D) that has been reported to be regulated by cytokines,
but not ovarian steroid hormones per se (25). In addition to up-regulation of
decidualization markers, TSA alone or in combination with E2+P4 induces morphological
changes that resemble the decidual transformation of ESC (Fig. 1E).
Synergistic enhancement of the ovarian steroid-induced IGFBP-1 and PRL
mRNA expression by TSA —We then investigated more quantitatively using competitive
RT-PCR whether TSA enhances E2+P4-induced IGFBP-1 mRNA expression in a dose-
dependent manner and examined whether the effect of TSA is additive or synergistic.
As illustrated in Fig. 2A, competitor DNA was generated by PCR using a
chimeric primer P3 and the downstream primer P2. Both the competitor and the target
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

13
sequence of IGFBP-1 cDNA derived from ESC were competitively co-amplified by PCR
using primers P1 and P2. The amplified product of competitor is 83 bp shorter than the
target sequence. Fig. 2B demonstrates the representative RT-PCR data where serial
dilutions of the competitor are added to the fixed amount of sample cDNA from ESC
treated without or with either E2+P4, TSA, or both, co-amplified and then examined by
electrophoresis. In the competitive RT-PCR analysis of mRNA for IGFBP-1, the
equivalent points, i.e. , the log ratio equals zero, in which similar intensities of the bands
of cDNA and competitor are detected (Figs. 2, B and C), were observed between 8 and
32 picograms of competitor when cells were treated with TSA in combination with E2+P4
(Fig. 2C). The competitive RT-PCR analysis using four different samples revealed that
TSA synergistically enhanced E2+P4-induced IGFBP-1 mRNA expression in a dose-
dependent manner (Fig. 2D).
Fig. 3A shows the representative RT-PCR data on PRL where serial dilutions of
the competitor, which is 112 bp shorter than the target sequence, are added to the fixed
amount of sample cDNA, co-amplified and then examined by electrophoresis. The
competitive RT-PCR analysis using three different samples revealed that TSA
synergistically enhanced E2+P4-induced PRL mRNA expression in a dose-dependent
manner (Fig. 3B).
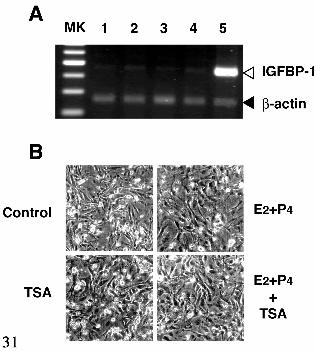
No effects of TSA on EGC—To investigate whether these molecular events
provoked by TSA are specific for ESC, we examined the effect of TSA on morphological
and biochemical changes in EGC isolated from endometrial specimens simultaneously
used for isolation of ESC. RT-PCR analysis revealed that TSA did not affect mRNA
expression of both IGFBP-1 (Fig. 4A) and PRL (data not shown) in EGC. Likewise,
TSA alone, ovarian steroid hormones, or co-treatment did not provoke any dramatically
morphological changes (Fig. 4B). These results suggest that the effect of TSA is selective
for ESC.
Acetylation of core histones of ESC upon treatment with ovarian steroids in
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

14
combination with TSA—To ascertain whether TSA treatment leads to acetylation of core
histones in ESC, acid-extractable proteins were analyzed on AUT gel (Fig. 5A). As noted
before (26), histone H4 exhibited the best resolution among other histones. Before TSA
treatment, H4 derived from Hela cells were primarily in the unacetylated or mono-
acetylated form (Fig. 5A lane 1). Eight hours of TSA treatment at 100 ng/ml led to
dramatic increase in di-, tri- and tetra-acetylated forms (lane 2). Likewise, H4 derived
from ESC treated with control vehicles for 8 hours consists largely of unacetylated or
mono-acetylated forms (lane 3), while di-, tri-, and tetra-acetylated forms appeared upon
8-hour stimulation with 500 ng/ml of TSA (lane 4). Also, treatment with E2+P4 showed
some increase in di- and tetra-acetylated forms (lane 5), both whose increment and
changes in the acetylation profiles were further enhanced by co-addition of TSA (lane 6).
With immunoblot analysis using acetylated histone type-specific antibodies, we
further determined which core histones and lysine residues underwent acetylation during
E2+P4-induced decidualization. As shown in Figure 5B, H2B, H3, and H4 (K8 and K12)
were preferentially acetylated upon eight-hour treatment with E2+P4. Co-addition of TSA
further enhanced the increase in acetylation levels of those core histones. Although TSA
alone induced an increase in acetylation levels of histones, a significant enhancement of
the signals by co-addition of E2+P4 was observed in H3 and H4. The Coomassie-stained
gel, at the bottom, indicates that the amount of protein loaded in each lane was similar
(Fig. 5B)
Densitometric analysis of the individual band intensities obtained from three or
more independent experiments revealed that treatment with E2+P4 or TSA alone
significantly increased the levels of acetylated H3 (K9 and K14) and H4 (K8), which
were further augmented by treatment in combination (Fig. 5C). H2B and K5 and K12 of
H4 also became acetylated, although it did not reach statistical significance by
nonparametric testing.
Augmentation of ovarian steroid-induced H4 acetylation by TSA in the
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

15
progesterone responsive region of the IGFBP-1 promoter—To determine whether the
levels of histone acetylation are increased in response to either E2+P4, TSA, or both at the
IGFBP-1 promoter, we used ChIP assays with an antibody specific for the acetylated
form of histone H4. Fig. 6A shows a schematic representation of the 5’-flanking of the
human IGFBP-1 gene promoter containing two functional progesterone responsive
elements as described elsewhere (27,28). Taking advantage of this information, the
progesterone-responsive fragment (region II) in the IGFBP-1 promoter was targeted for
PCR amplification, while non-progesterone-responsive fragment (region I) was also
targeted as controls to verify the progesterone dependence. As shown in Fig. 6B, there
are no differences in the amount of region I and II sequences in inputs across non-treated
and treated cells (lanes 1–4, 9–12). Importantly, treatment with E2+P4 increased the
amount of region II promoter associated with acetylated histone H4 (lane 14) when
compared with control vehicles (lanes 13) or TSA alone (lanes 14). The association was
dramatically enhanced by co-addition of TSA (lane 16). In contrast, neither E2+P4 nor
in combination with TSA had any appreciable effect on the amount of region I promoter
associated with acetylated histone H4 (lanes 5–8).
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

16
Discussion
In this study, we demonstrated using immunoblot analysis with acetylated
histone type-specific antibodies that H3 and H4 became significantly acetylated at K9/K14
and K8, respectively, in response to ovarian steroid hormones. The mild but significant
acetylation was further enhanced by co-addition of TSA. Individual HATs are known to
possess acetylation preferences for specific sites and substrates (29,30). For instance,
SRC-1 preferentially acetylates K9 and K14 of H3, while CBP/p300 has a broader
preference for K5 of H2A, K12 and K15 of H2B, K14 and K18 of H3, and K5 and K8
of H4 (29,30). Conversely, no HAT but CBP/p300 has been so far reported to acetylate
H4 on K8 (29,30). Given the acetylation sites upon treatment with ovarian steroids as
presented here, SRC-1 and CBP/p300 may be likely candidates for HATs responsible for
histone acetylation in the process of decidualization.
As for SRC-1, uterine decidual response is impaired in SRC-1 null mice (31). In
addition, SRC-1e, one of the SRC-1 isoforms, significantly enhanced decidual PRL
promoter activity in response to 8-bromo cAMP (32). These collectively suggest that
SRC-1 is at least in part required for decidualization, supporting our current idea. CBP
(CREB-binding protein) was originally discovered as a factor interacting with the
phosphorylated form of the transcription factor CREB (cAMP response element-binding
protein)(33). It is well known that CBP plays a pivotal role not only in cAMP-regulated
gene expression by interacting with the phosphorylated form of CREB but also in
expression of numerous genes by associating with many transcription factors including
nuclear steroid receptors (34). p300 was isolated independently as factors interacting with
the adenovirus E1A protein (35). The high degree of relatedness between CBP and p300
was soon recognized, and subsequent studies indicated that they are largely
interchangeable in function (36). Since cAMP is known to be one of the potent inducers
of decidualization (7-9), we postulate that CBP/p300 may also participate in the process
of decidualization. As the first step to prove our hypothesis, we have confirmed that
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

17
transcripts of SRC-1 and CBP/p300 were present in human ESC (unpublished
observation2), consistent with recent data as reported elsewhere (37,38).
Since TSA is thought to inhibit histone deacetylase activity specifically, it has
been widely used as a tool to study the consequences of histone acetylation in vivo (18).
Intriguingly, the expression of only about 2% of expressed genes is changed (increased
or decreased) two-fold or more in cells cultured with TSA when compared with untreated
control cells (39). In agreement, we here show that treatment of ESC with E2+P4 up-
regulates IGFBP-1 and PRL, but neither MMP-2 nor β-actin, suggesting the gene-
selective effect of TSA. However, one can criticize that the effect of TSA may not be
specific for ESC in that TSA may provoke global histone acetylation and therefore induce
number of genes including IGFBP-1 and PRL in any types of cells. To address this
question, we examined the effect of TSA on EGC simultaneously isolated from the same
specimens. Unlike ESC, EGC did not express IGFBP-1 and PRL genes in response to
neither TSA nor E2+P4. Thus, induction of IGFBP-1 and PRL by TSA is ESC-specific.
Molecular determinants to induce pre-programmed genes of ESC, but not EGC, upon
decidualization remain to be elucidated.
Nuclear steroid hormone receptors recruit several HATs as well as HDACs upon
ligand binding and thereby cause an increase in the levels of histone acetylation around the
nuclear hormone responsive region (34). Subsequently, DNA that is tightly wrapped
around a deacetylated histone core relaxes and the accumulation of acetylated histones in
nucleosomes containing this region leads to expression of target genes of steroid
hormones (34). Consistent with this model, in the presence of E2+P4, H4 becomes
acetylated in the proximal region containing putative progesterone responsive elements,
but not the distal region of IGFBP-1 gene, as determined by ChIP assay. This acetylation
in the proximal region is further enhanced by co-addition of TSA, suggesting that
transcriptional synergy by TSA and E2+P4 may require the progesterone responsive
element.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

18
Even in the absence of hormones, a relatively high dose of TSA provokes both
induction of IGFBP-1 mRNA and acetylation of several core histones including H4. This
seems to be inconsistent with our ChIP data in that treatment with TSA alone does not
induce the acetylation of H4 in the proximal as well as distal region of IGFBP-1 promoter.
At present, we do not have direct evidence to account for this discrepancy. One possibility
is that, in the absence of ovarian steroid hormones, a high dose of TSA may affect the
core histones present in the regulatory regions of IGFBP-1 different from those tested in
this study. Indeed, it has been reported that a putative hypoxia-inducible factor responsive
element present in the first intron of the IGFBP-1 gene is functionally involved in the
IGFBP-1 gene regulation (40). Alternatively, Sp1 sites have been demonstrated to play a
crucial role in TSA-induced transcription of the telomerase catalytic subunit in several
types of normal human cells (41). Intriguingly, IGFBP-1 promoter possesses SP1 sites
between –2830 to 2630 bp of its distal region (42). Given that the Sp1 sites are extremely
active in decidualized stromal cells, which accounts for >95% of the total induction
(42,43), it is possible that, in the absence of ovarian steroid hormones, TSA might induce
transcription of IGFBP-1 via the distal promoter SP1 sites that were not tested by ChIP
assay in this study.
In conclusion, TSA advances P4-induced decidualization of E2-primed ESC
through potentiation of progesterone action, suggesting that histone acetylation is deeply
involved in ovarian steroid-induced decidualization. The present study may provide a clue
for possible differentiation therapies to target histone acetylation and deacetylation for
endometrium-derived diseases such as endometriosis, endometrial cancer, and infertility
due to endometrial defects of implantation.
Acknowledgements
We thank Ms. Shino Kuwabara for help with preparation of the manuscript.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

19
References
1. Noyes, R. W., Hertig, A. T., and Rock, J. (1950) Fertil Steril 1 , 3-25
2. Gurpide, E., Tabanelli, S., and Tang, B. (1992) Human endometrial stromal
cells. Hormones in Gynecological Endocrinology (Genazzani, A. R., and Petraglia, F.,
Eds.), Partenon Press, Casterton Hall. Carnforth, Lancs, UK
3. Tabibzadeh, S. (1991) Endocr Rev 1 2 , 272-90
4. Giudice, L. C. (1994) Fertil Steril 6 1 , 1-17
5. Huang, J. R., Tseng, L., Bischof, P., and Janne, O. A. (1987) Endocrinology
1 2 1 , 2011-7.
6. Irwin, J. C., Kirk, D., King, R. J. , Quigley, M. M., and Gwatkin, R. B.
(1989) Fertil Steril 5 2 , 761-8
7. Tang, B., Guller, S., and Gurpide, E. (1993) Endocrinology 1 3 3 , 2197-203
8. Telgmann, R., Maronde, E., Tasken, K., and Gellersen, B. (1997)
Endocrinology 1 3 8 , 929-37
9. Brar, A. K., Frank, G. R., Kessler, C. A., Cedars, M. I. , and Handwerger, S.
(1997) Endocrine 6 , 301-7
10. Maruyama, T., Yoshimura, Y., Yodoi, J. , and Sabe, H. (1999) Endocrinology
1 4 0 , 2632-2636
11. Maruyama, T., Yoshimura, Y., and Sabe, H. (1999) Endocrinology 140,
5982-90
12. Yamamoto, Y., Maruyama, T., Sakai, N., Sakurai, R., Shimizu, A., Hamatani,
T., Masuda, H., Uchida, H., Sabe, H., and Yoshimura, Y. (2002) Mol Hum Reprod 8,
1117-24.
13. Jabbour, H. N., Critchley, H. O., and Boddy, S. C. (1998) J Clin Endocrinol
Metab 8 3 , 2545-53.
14. Mak, I. Y., Brosens, J. J. , Christian, M., Hills, F. A., Chamley, L., Regan, L.,
and White, J. O. (2002) J Clin Endocrinol Metab 8 7 , 2581-8.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

20
15. Popovici, R. M., Kao, L. C., and Giudice, L. C. (2000) Endocrinology 141,
3510-3.
16. Brar, A. K., Handwerger, S., Kessler, C. A., and Aronow, B. J. (2001)
Physiol Genomics 7 , 135-48.
17. Roth, S. Y., Denu, J. M., and Allis, C. D. (2001) Annu Rev Biochem 70, 81-
120.
18. Yoshida, M., Horinouchi, S., and Beppu, T. (1995) Bioessays 1 7 , 423-30.
19. Johnstone, R. W. (2002) Nat Rev Drug Discov 1 , 287-99.
20. Serra, M. J., Ledford, B. E., and Baggett, B. (1979) Biol Reprod 2 0 , 214-20.
21. Maruyama, T., Sachi, Y., Furuke, K., Kitaoka, Y., Kanzaki, H., Yoshimura,
Y., and Yodoi, J. (1999) Endocrinology 1 4 0 , 365-72
22. Jin, C. F., Mata, M., and Fink, D. J. (1994) PCR Methods Appl 3 , 252-5.
23. Maruyama, T., Kitaoka, Y., Sachi, Y., Nakanoin, K., Hirota, K., Shiozawa,
T., Yoshimura, Y., Fujii, S., and Yodoi, J. (1997) Mol Hum Reprod 3 , 989-93.
24. Yoshida, M., Kijima, M., Akita, M., and Beppu, T. (1990) J Biol Chem 265,
17174-9
25. Lockwood, C. J. , Krikun, G., Hausknecht, V. A., Papp, C., and Schatz, F.
(1998) Endocrinology 1 3 9 , 4607-13.
26. West, M. H., and Bonner, W. M. (1980) Biochemistry 1 9 , 3238-45.
27. Gao, J. , Mazella, J. , Suwanichkul, A., Powell, D. R., and Tseng, L. (1999)
Mol Cell Endocrinol 1 5 3 , 11-7.
28. Gao, J. , Mazella, J. , Tang, M., and Tseng, L. (2000) Mol Endocrinol 14,
1954-61.
29. Kuo, M. H., and Allis, C. D. (1998) Bioessays 2 0 , 615-26.
30. Sterner, D. E., and Berger, S. L. (2000) Microbiol Mol Biol Rev 6 4 , 435-59.
31. Xu, J. , Qiu, Y., DeMayo, F. J. , Tsai, S. Y., Tsai, M. J., and O' Malley, B. W.
(1998) Science 2 7 9 , 1922-5.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

21
32. Brosens, J. J. , Hayashi, N., and White, J. O. (1999) Endocrinology 140,
4809-20.
33. Chrivia, J. C., Kwok, R. P., Lamb, N., Hagiwara, M., Montminy, M. R., and
Goodman, R. H. (1993) Nature 3 6 5 , 855-9.
34. Robyr, D., Wolffe, A. P., and Wahli, W. (2000) Mol Endocrinol 1 4 , 329-47.
35. Eckner, R., Ewen, M. E., Newsome, D., Gerdes, M., DeCaprio, J. A.,
Lawrence, J. B., and Livingston, D. M. (1994) Genes Dev 8 , 869-84.
36. Vo, N., and Goodman, R. H. (2001) J Biol Chem 2 7 6 , 13505-8.
37. Wieser, F., Schneeberger, C., Hudelist, G., Singer, C., Kurz, C., Nagele, F.,
Gruber, C., Huber, J. C., and Tschugguel, W. (2002) Mol Hum Reprod 8 , 644-50.
38. Gregory, C. W., Wilson, E. M., Apparao, K. B., Lininger, R. A., Meyer, W.
R., Kowalik, A., Fritz, M. A., and Lessey, B. A. (2002) J Clin Endocrinol Metab 87,
2960-6.
39. Van Lint, C., Emiliani, S., and Verdin, E. (1996) Gene Expr 5 , 245-53.
40. Tazuke, S. I. , Mazure, N. M., Sugawara, J. , Carland, G., Faessen, G. H.,
Suen, L. F., Irwin, J. C., Powell, D. R., Giaccia, A. J. , and Giudice, L. C. (1998) Proc
Natl Acad Sci U S A 9 5 , 10188-93.
41. Takakura, M., Kyo, S., Sowa, Y., Wang, Z., Yatabe, N., Maida, Y., Tanaka,
M., and Inoue, M. (2001) Nucleic Acids Res 2 9 , 3006-11.
42. Gao, J., and Tseng, L. (1996) Mol Endocrinol 1 0 , 613-21.
43. Gao, J. G., Mazella, J. , Powell, D. R., and Tseng, L. (1994) DNA Cell Biol
1 3 , 829-37.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

22
Footnotes
This work was supported in part by grants from the Ministry of Education, Science, and
Culture of Japan (C1367143 to T.M. and B12470348 to Y.Y.), Keio Gijuku Academic
Development Funds (to T.M.), and grants from the Keio Health Counseling Center (to
T.M.). The costs of publication of this article were defrayed in part by the payment of
page charges. This article must therefore be hereby marked “advertisement” in accordance
with 18 U.S.C. Section 1734 solely to indicate this fact.
1The abbreviation used are: ESC, endometrial stromal cells; HDAC, histone deacetylase;
HAT, histone acetyltransferase; TSA, trichostatin A; IGFBP-1, insulin-like growth factor
binding protein-1; E2, 17-β estradiol; P4, progesterone; DMEM, Dulbecco’s Modified
Eagles’ Medium; FBS, fetal bovine serum; PCR, polymerase chain reaction; RT, reverse
transcriptase; PRL, prolactin; MMP-2, matrix metalloproteinase-2; PAGE,
polyacrylamide gel electrophoresis; PVDF, polyvinylidene difluoride; AUT, acid urea
triton; ChIP, Chromatin immunoprecipitation.
2Maruyama, T and Sakai, N.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

23
Figure legends
FIG. 1. TSA advances ovarian s teroid-induced decidualization of cultured
ESC.
A, RT-PCR analysis of IGFBP-1 mRNA derived from ESC treated without or
with E2+P4 in combination with TSA. ESC were treated for 48 hours with
control vehicles (lanes 1-4), E2+P4 (lanes 5-8), or in combination with TSA
(lanes 2 and 6, 10 ng/ml; 3 and 7, 100 ng/ml; 4 and 8, 500 ng/ml). As a positive
control, ESC were cultured for 14 days in the presence of E2+P4 (lane 9). Total
RNA was extracted and subjected to duplex RT-PCR for IGFBP-1 and β-actin
mRNA.
B, Northern blot analysis of IGFBP-1 mRNA derived from ESC treated without
or with E2+P4 in combination with TSA. ESC were treated for 48 hours with
control vehicles (lanes 1 and 2), E2+P4 (lanes 3 and 4), or in combination with
TSA (lanes 2 and 4, 100 ng/ml). Total RNA was extracted, electrophoresed,
transferred to the Nytran membrane, and then hybridized with IGFBP-1 cDNA
probe.
C, RT-PCR analysis of PRL mRNA derived from ESC treated without or with
E2+P4 in combination with TSA. ESC were treated for 48 hours with control
vehicles (lanes 1 and 2), E2+P4 (lanes 3 and 4), or in combination with TSA
(lanes 2 and 4, 500 ng/ml). Total RNA was extracted and subjected to duplex
RT-PCR for PRL and β-actin mRNA.
D, RT-PCR analysis of MMP-2 mRNA derived from ESC treated without or
with E2+P4 in combination with TSA. ESC were treated for 48 hours with
control vehicles (lanes 1-3), E2+P4 (lanes 4-6), or in combination with TSA
(lanes 2 and 5, 100 ng/ml; 4 and 6, 500 ng/ml). Total RNA was extracted and
subjected to RT-PCR for MMP-2 mRNA.
E, Phase contrast micrographs of ESC treated for 48 hours with control
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

24
vehicles (Control), TSA alone (500 ng/ml), E2+P4, or in combination with TSA
(500 ng/ml).
FIG. 2. TSA synergis tically enhances the ovarian s teroid-induced IGFBP-
1 mRNA expression in a dose-dependent manner.
A, Schematic representation of the preparation of competitive templates used as
external standards in each amplification reaction and the method of competitive
PCR. A chimeric primer P3, which matches the white-colored sequence located
103 bp downstream from the black-colored sequence and contains the black-
colored sequence at the 5’-end, and primer P2 were used for PCR amplification
to generate competitor using wild type cDNA as a template. The serially diluted
competitors generated were added to the samples as competitors. Both the target
sequence and competitor are competitively amplified by PCR using primers P1
and P2.
B, The representative data on competitive RT-PCR using total RNA derived
from ESC treated without or with E2+P4 in combination with TSA as indicated
for 48 hours. Serial dilutions of the competitor (lane 1, 512 pg; lane 2, 128 pg;
lane 3, 32 pg; lane 4, 8 pg; lane 5, 2 pg; lane 6, 0.5 pg; lane 7, 0 pg) were added
to the fixed amount of sample cDNA and then co-amplified and examined by
electrophoresis. The amplified product of competitor (white arrowheads) is 83
bp shorter than the target sequence (black arrowheads). MK, 100 bp ladder
marker.
C, The representative data plotted as a function of the log (target band
intensity/competitor band intensity) against the amount of competitor added to
the RT-PCR reactions. When the log ratio equals zero, the concentrations of the
target (originally from the RT reaction) and the competitor are equal. Thus, the
amount of cDNA synthesized from 300 ng of the initial RNA sample was
calculated from the graph as indicated by the arrow.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

25
D, Graphic representation of the mean relative expression levels of IGFBP-1
mRNA in ESC treated without or with E2+P4 in combination with various
concentrations of TSA (10, 100, or 500 ng/ml) for 48 hours. Mean (± SE,
n=4) ratio of IGFBP-1 mRNA signals, as determined by image analysis with
the ratios for treatment with E2+P4, set at 1. a, b, and c, p<0.01.
FIG. 3. TSA synergis tically enhances the ovarian s teroid-induced PRL
mRNA expression in a dose-dependent manner.
A, The representative data on competitive RT-PCR using total RNA derived
from ESC treated without or with E2+P4 in combination with TSA as indicated
for 48 hours. Serial dilutions of the competitor (lane 1, 32 pg; lane 2, 2 pg; lane
3, 0.13 pg; lane 4, 7.8 femtogram; lane 5, 0.49 femtogram; lane 6, 0 pg) were
added to the fixed amount of sample cDNA and then co-amplified and examined
by electrophoresis. The amplified product of competitor (white arrowheads) is
112 bp shorter than the target sequence (black arrowheads). MK, 100 bp ladder
marker.
B, Graphic representation of the mean relative expression levels of PRL mRNA
in ESC treated without or with E2+P4 in combination with various
concentrations of TSA (10, 100, or 500 ng/ml) for 48 hours. Mean (± SE,
n=5) ratio of PRL mRNA signals, as determined by image analysis with the
ratios for treatment with control vehicles, set at 1. a, b, c, d, and e, p<0.05.
FIG. 4. TSA dose not affect EGC.
A, RT-PCR analysis of IGFBP-1 mRNA derived from EGC treated without or
with E2+P4 in combination with TSA. EGC were treated for 48 hours with
control vehicles (lanes 1 and 2), E2+P4 (lanes 3 and 4), or in combination with
500 ng/ml TSA (lanes 2 and 4). As a positive control, ESC were cultured for 14
days in the presence of E2+P4 (lane 5). Total RNA was extracted and subjected
to duplex RT-PCR for IGFBP-1 and β-actin mRNA. MK, 100 bp ladder marker.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

26
B, Phase contrast micrographs of EGC treated without or with E2+P4 in
combination with 500 ng/ml TSA.
FIG. 5. Core his tones of ESC become acetylated upon treatment with
ovarian steroids in combination with TSA.
A, AUT gel analysis on acid-extractable histones from Hela cells treated without
(lane 1) or with 500 ng/ml TSA (lane 2) and ESC treated without (lane 3) or with
500 ng/ml TSA (lane 4), E2+P4 (lane 5), or both (lane 6).
B, Representative blots of acid-extracted histones from ESC treated without
(lane 1) or with 500 ng/ml TSA (lane 2), E2+P4 (lane 3), or both (lane 4) and
Hela cells treated with 500 ng/ml TSA (lane 5) using various antibodies against
acetylated histones as indicated. The Coomassie-stained gel, at the bottom,
indicates the amount of protein loaded in each lane was similar.
C, Graphic representation of the mean relative expression levels of IGFBP-1
mRNA in ESC treated without or with E2+P4 in combination with 500 ng/ml
TSA for 8 hours. The individual densitometric intensities (means ± SD) for at
least three independent experiments are shown as a –fold increase over those
observed for the control treatment. a, p<0.05 vs. control treatment. b, <0.01 vs.
control treatment. c, p<0.01, vs. treatment with TSA alone.
FIG. 6. TSA augments ovarian s teroid-induced H4 acetylation in the
progesterone responsive region of the IGFBP-1 promoter.
A, Schematic representation of the 5’-flanking of the human IGFBP-1 gene
promoter and two target promoter regions (region I, distal; region II, proximal)
for the ChIP assay. PRE, functional progesterone responsive element as
described elsewhere (27,28).
B, Immunoprecipitation of chromatin with antibodies directed against acetylated
histone H4. ESC were left untreated (lanes 1, 5, 9 and 13), or were treated for 1
hour with 500 ng/ml TSA (lanes 2, 6, 10 and 14), E2+P4 (lanes 3, 7, 11 and 15),
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from

27
or both (lanes 4, 8, 12 and 16). MK, 100 bp ladder marker.
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from





Shimizu, Ikuko Kishi, Hironori Asada, Satoshi Yamagoe and Yasunori YoshimuraNozomi Sakai, Tetsuo Maruyama, Rei Sakurai, Hirotaka Masuda, Yurie Yamamoto, Aki
human endometrial stromal cellsInvolvement of histone acetylation in ovarian steroid-induced decidualization of
published online February 27, 2003J. Biol. Chem.
10.1074/jbc.M211715200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on July 5, 2019http://w
ww
.jbc.org/D
ownloaded from
Related Documents











