The International Journal of Biochemistry & Cell Biology 40 (2008) 1743–1752 Available online at www.sciencedirect.com Insights into the mechanism and regulation of pyruvate carboxylase by characterisation of a biotin-deficient mutant of the Bacillus thermodenitrificans enzyme Abdussalam Adina-Zada a , Sarawut Jitrapakdee b , Kathy H. Surinya c , Matthew J. McIldowie a , Matthew J. Piggott a , W. Wallace Cleland d , John C. Wallace c , Paul V. Attwood a,∗ a School of Biomedical, Biomolecular and Chemical Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia b Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailand c School of Molecular Biosciences, University of Adelaide, Adelaide, SA 5005, Australia d Institute for Enzyme Research and Department of Biochemistry, University of Wisconsin-Madison, Madison, WI 53726, USA Received 8 December 2007; accepted 2 January 2008 Available online 6 January 2008 Abstract Pyruvate carboxylase is a biotin-dependent enzyme in which the biotin is carboxylated by a putative carboxyphosphate interme- diate that is formed in a reaction between ATP and bicarbonate. The resultant carboxybiotin then transfers its carboxyl group to pyruvate to form oxaloacetate. In the Bacillus thermodenitrificans enzyme the biotin is covalently attached to K1112. A mutant form of the enzyme (K1112A) has been prepared which is not biotinylated. This mutant did not catalyse the complete reaction, but did catalyse ATP-cleavage and the carboxylation of free biotin. Oxaloacetate decarboxylation was not catalysed, even in the presence of free biotin, suggesting that only the biotin carboxylation domain of the enzyme is accessible to free biotin. This mutant allowed the study of ATP-cleavage both coupled and not coupled to biotin carboxylation. Kinetic analyses of these reactions indicate that the major effect of the enzyme activator, acetyl CoA, is to promote the carboxylation of biotin. Acetyl CoA reduces the K m s for both MgATP and biotin. In addition, pH profiles of the ATP-cleavage reaction in the presence and absence of free biotin revealed the involvement of several ionisable residues in both ATP-cleavage and biotin carboxylation. K1112A also catalyses the phospho- rylation of ADP from carbamoyl phosphate. Stopped-flow studies using the fluorescent ATP analogue, formycin A-5 -triphosphate, in which nucleotide binding to the holoenzyme was compared to K1112A indicated that the presence of biotin enhanced binding. Attempts to trap the putative carboxyphosphate intermediate in K1112A using diazomethane were unsuccessful. © 2008 Elsevier Ltd. All rights reserved. Keywords: Pyruvate carboxylase; ATP-cleavage; Biotin carboxylation; Acetyl CoA regulation; Stopped-flow ∗ Corresponding author at: Biochemistry and Molecular Biology (M310), School of Biomedical, Biomolecular and Chemical Sciences, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia. Tel.: +61 8 6488 3329; fax: +61 8 6488 1148. E-mail addresses: [email protected] (A. Adina-Zada), [email protected] (S. Jitrapakdee), [email protected] (K.H. Surinya), [email protected] (M.J. McIldowie), [email protected] (M.J. Piggott), [email protected] (W.W. Cleland), [email protected] (J.C. Wallace), [email protected] (P.V. Attwood). 1357-2725/$ – see front matter © 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.biocel.2008.01.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
dpocottbtriA©
K
1
The International Journal of Biochemistry & Cell Biology 40 (2008) 1743–1752
Available online at www.sciencedirect.com
Insights into the mechanism and regulation of pyruvate carboxylaseby characterisation of a biotin-deficient mutant of the Bacillus
thermodenitrificans enzyme
Abdussalam Adina-Zada a, Sarawut Jitrapakdee b, Kathy H. Surinya c,Matthew J. McIldowie a, Matthew J. Piggott a, W. Wallace Cleland d,
John C. Wallace c, Paul V. Attwood a,∗a School of Biomedical, Biomolecular and Chemical Sciences, The University of Western Australia,
35 Stirling Highway, Crawley, WA 6009, Australiab Department of Biochemistry, Faculty of Science, Mahidol University, Bangkok 10400, Thailand
c School of Molecular Biosciences, University of Adelaide, Adelaide, SA 5005, Australiad Institute for Enzyme Research and Department of Biochemistry, University of Wisconsin-Madison, Madison, WI 53726, USA
Received 8 December 2007; accepted 2 January 2008Available online 6 January 2008
bstract
Pyruvate carboxylase is a biotin-dependent enzyme in which the biotin is carboxylated by a putative carboxyphosphate interme-iate that is formed in a reaction between ATP and bicarbonate. The resultant carboxybiotin then transfers its carboxyl group toyruvate to form oxaloacetate. In the Bacillus thermodenitrificans enzyme the biotin is covalently attached to K1112. A mutant formf the enzyme (K1112A) has been prepared which is not biotinylated. This mutant did not catalyse the complete reaction, but didatalyse ATP-cleavage and the carboxylation of free biotin. Oxaloacetate decarboxylation was not catalysed, even in the presencef free biotin, suggesting that only the biotin carboxylation domain of the enzyme is accessible to free biotin. This mutant allowedhe study of ATP-cleavage both coupled and not coupled to biotin carboxylation. Kinetic analyses of these reactions indicate thathe major effect of the enzyme activator, acetyl CoA, is to promote the carboxylation of biotin. Acetyl CoA reduces the Kms for
oth MgATP and biotin. In addition, pH profiles of the ATP-cleavage reaction in the presence and absence of free biotin revealedhe involvement of several ionisable residues in both ATP-cleavage and biotin carboxylation. K1112A also catalyses the phospho-ies usi ′
ylation of ADP from carbamoyl phosphate. Stopped-flow stud n which nucleotide binding to the holoenzyme was compared to K1ttempts to trap the putative carboxyphosphate intermediate in K1112008 Elsevier Ltd. All rights reserved.eywords: Pyruvate carboxylase; ATP-cleavage; Biotin carboxylation; Acety
∗ Corresponding author at: Biochemistry and Molecular Biology (M310), SThe University of Western Australia, 35 Stirling Highway, Crawley, WA 600
E-mail addresses: [email protected] (A. Adina-Zada), [email protected]@adelaide.edu.au (K.H. Surinya), [email protected] ([email protected] (W.W. Cleland), [email protected] (J
357-2725/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.biocel.2008.01.001
ng the fluorescent ATP analogue, formycin A-5 -triphosphate,
112A indicated that the presence of biotin enhanced binding.2A using diazomethane were unsuccessful.l CoA regulation; Stopped-flow
chool of Biomedical, Biomolecular and Chemical Sciences,9, Australia. Tel.: +61 8 6488 3329; fax: +61 8 6488 1148.mucc.mahidol.ac.th (S. Jitrapakdee),.J. McIldowie), [email protected] (M.J. Piggott),.C. Wallace), [email protected] (P.V. Attwood).

l of Bioc
1744 A. Adina-Zada et al. / The International Journa1. Introduction
Pyruvate carboxylase (PC) is a biotin-dependentenzyme that catalyses the carboxylation of pyruvate inthe series of reaction steps shown in Fig. 1. Biotin iscarboxylated in a reaction that involves the formationof a putative carboxyphosphate intermediate from ATPand bicarbonate. Incorporation of 18O from HC18O3
−into Pi provides strong evidence for the existenceof carboxyphosphate in biotin carboxylation reactions(Kaziro, Hass, Boyer, & Ochoa, 1962) and there is alsoother, less direct, evidence (for reviews see Attwood,1995 and Attwood & Wallace, 2002), however the inter-mediate has never been trapped in a biotin-dependentcarboxylase. The carboxybiotin then moves to the site ofthe pyruvate carboxylation reaction where transfer of thecarboxyl group results in the formation of oxaloacetate.
Recently, the molecular structure of pyruvate car-boxylase from R. etli has been determined (St Mauriceet al., 2007). The authors of this article presented evi-dence that the carboxybiotin from one subunit binds atthe site of pyruvate carboxylation in the active site ofits dimer partner subunit. In addition, only one of thedimer pairs of subunits in the tetrameric enzyme boundthe activator, acetyl CoA and was configured to allowthis cross-subunit binding of carboxybiotin (St Mauriceet al., 2007).
Although there have been detailed studies of themechanism of action of pyruvate carboxylase and theregulation of its activity by acetyl CoA (Attwood, 1993;Attwood & Cleland, 1986; Branson & Attwood, 2000;Branson, Nezic, Jitrapakdee, Wallace, & Attwood, 2004;Legge, Branson, & Attwood, 1996), these have all faceddifficulties in differentiating between attributes of theATP-cleavage reaction and those of biotin carboxylationitself. Studies have been carried out on the biotin car-boxylase subunit of acetyl CoA carboxylase, howeverit is apparent that the biotin carboxylation reaction ismarkedly different to that occurring when it is associ-ated with the other subunits of the enzyme (Blanchard,Chapman-Smith, Wallace, & Waldrop, 1999).
In the present work, we have mutated lysine (K1112)to alanine (K1112A) in pyruvate carboxylase from Bacil-lus thermodenitrificans. The biotin is normally attachedby post-translational modification, via formation of an
Fig. 1. Reaction scheme for the carboxylation of pyruvate, showing the initial cintermediate, followed by transfer of the carboxy group to pyruvate to form o
hemistry & Cell Biology 40 (2008) 1743–1752
amide bond to the N6 of K1112 in the wild-type enzyme(WT); the K1112A mutant cannot be biotinylated andthus K1112A is an apoenzyme, with an intact polypep-tide chain, that only lacks the biotin prosthetic group. Inthe present work, we describe the characterisation of theK1112A mutant, including its ability to catalyse ATP-cleavage, carboxylate free biotin and the role played byacetyl CoA in the biotin carboxylation reaction.
2. Materials and methods
2.1. Chemicals and reagents
The formycin A was purchased from Berry & Asso-ciates, Inc (MI, USA) and formycin A-5′-triphosphatewas synthesised from formycin A as described by Ward,Cerami, Reich, Acs, and Altwerger (1969). The sodiumsalt of acetyl CoA was purchased from Sigma and allother reagents were of analytical quality.
2.2. Construction of WT and K1112A mutant
The wild-type B. thermodenitrificans PC constructwith N-terminal 9His tag and two extra glycineswas generated by PCR-amplification of PC genefrom genomic DNA. The gene was cloned fromtwo overlapping PCR fragments using two pairsof primers designed from the published sequence(Kondo, Kazuta, Saito, & Fuji, 1997). BacF1 (5′-gccggtaccatatgcatcaccatcaccatcaccatcaccatggtggtaag-acaagacgaattcgcaaagtgctcgta-3′; underline indicatesNdeI restriction site, bold indicates an ATG start codon)and BacR1 (5′-ctacggcgttggcagaacggagcaaca-3′) weresynthesized to PCR-amplify a 1.8 kb 5′-end of theBacPC gene whilst BacF2 (5′-ttggcgacacgagtccgcacgg-tcgat-3′) and BacR2 (5′-ccccgggtaccttatttggacaactcc-atgagcaaatcg-3′ (the underlined indicates KpnI restric-tion site) were synthesized to PCR-amplify a 1.7 kb3′-end of the BacPC gene. These two fragmentswere sequenced and assembled by digestion withNdeI/BamHI and with BamHI/KpnI, respectively, and
arboxylation of biotin via formation of the putative carboxyphosphatexaloacetate.
ligated to NdeI/KpnI digested pET17b (Novagen),yielding a wild-type full length construct.
K1112A was constructed by site-directed muta-genesis using a 1.7 kb 3′-end of the BacPC con-

l of Bioc
sfccgpnTBsv
BB1c
2
vicwttIwp
ftNt(p
wb0llutpbw0i
tb
A. Adina-Zada et al. / The International Journa
truct as the template. The oligonucleotides usedor creating this mutant were K1112A-For (5′-acggaagcaatggccatggaaacgacc-3′, bold indicates theodon changed for alanine) and K1112A-Rev (5′-gtcgtttccatggccattgcttccgtg-3′). The mutagenesis waserformed using a Quick-Change site-directed mutage-esis, as previously described (Jitrapakdee et al., 2007).he BamHI-KpnI mutagenic cassette then replaced theamHI-KpnI wild-type fragment in the full length con-truct. The nucleotide sequence of the mutant waserified by DNA sequencing.
The wild-type 9His BacPC and 9His K1112AacPC constructs were transformed to E. coli RosettaL21 (Stratagene) and selected on LB agar containing00 �g ml−1 ampicillin and 30 �g ml−1 chlorampheni-ol.
.3. Expression and purification of WT and K1112A
WT Bacillus thermodenitrificans and K1112A pyru-ate carboxylase were expressed by growth of the E. colin terrific broth media, containing 100 �g ml−1 ampi-illin and 30 �g ml−1 chloramphenicol. The culturesere grown at 37 ◦C to an OD600 = 0.5. In the case of
he WT expression, biotin was added to a final concen-ration of 1 mg l−1 and the cultures were induced withPTG to a final concentration of 0.1 mM. The culturesere transferred to a 20 ◦C shaking incubator overnightrior to cell harvesting.
The harvested cells were incubated with lysozymeollowed by cell breakage using a Bead-BeaterTM inhe presence of a protease inhibitor cocktail (Sigma).ucleic acids were removed from the lysate by pro-
amine sulphate precipitation as described previouslyBranson et al., 2004). The proteins in the lysate wererecipitated using ammonium sulphate (57% saturated).
In the preparation of WT the precipitated proteinsere resuspended in avidin affinity column loadinguffer (25 mM KH2PO4, 1 mM EDTA, 0.5 mM DTE,.1 mM PMSF, pH 7.2). The protein solution was dia-yzed against the loading buffer further overnight andoaded onto a 25-ml Avidin-Sepharose column. The col-mn was washed by loading buffer until OD280 ≈ 0 andhe enzyme was eluted by addition of 1 mM biotin. Theurified enzyme fractions were pooled and precipitatedy 57% saturated ammonium sulphate. All operationsere performed at 4 ◦C. The enzyme was dissolved in.1 M Tris–Cl, 1 mM DTE, 30% glycerol, pH 7.8, frozen
n liquid nitrogen and stored at −80 ◦C.In the preparation of K1112A, the precipitated pro-eins were resuspended in Ni-NTA column loadinguffer (0.3 M NaCl, 50 mM NaH2PO4, 1 mM DTE,
hemistry & Cell Biology 40 (2008) 1743–1752 1745
and 0.1 mM PMSF, pH 8.0). The solution was loadedonto a 20 ml column of Ni-loaded Profinity Imac resin(Bio-Rad Laboratories). The column was washed withwashing buffer (0.3 M NaCl, 50 mM NaH2PO4, 5 mMimidazole, 1 mM DTE, 0.1 mM PMSF, pH 8.0) untilOD280 ≈ 0 and the enzyme was eluted using a gradi-ent 0–250 mM imidazole. The purified enzyme fractionswere pooled and precipitated by ammonium sulphate.All operations were performed at 4 ◦C. The enzyme pre-cipitate was dissolved in 0.1 M Tris–Cl, 1 mM DTE, 30%glycerol, pH 7.8, frozen in liquid nitrogen and stored at−80 ◦C.
Prior to assay, the enzymes were transferred to 0.1 MTris–Cl, pH 7.8 by centrifuging through Sephadex G-25as described by Helmerhorst and Stokes (1980).
2.4. Western blotting
Western analysis of whole cell lysates of WT andK1112A mutant was performed as previously described(Jitrapakdee et al., 2007). The biotinylated PC wasdetected using conjugated steptavidin-HRP while theHis tag was detected using NiNTA conjugated with alka-line phosphatase (Qiagen).
2.5. Assay of ATP-cleavage
ADP formation was measured in coupled reactionswith pyruvate kinase and lactate dehydrogenase essen-tially as described by Attwood and Graneri (1992). Oneunit of enzyme activity is defined as the amount ofenzyme required to catalyse the formation of 1 �molof ADP min−1 at 30 ◦C. All assays were performedat 30 ◦C and in 0.1 M Tris–Cl, pH 7.8, with 0–50 mMbiotin; 20 mM NaHCO3 in the presence or absence of0.25 mM acetyl CoA. Note that in the experiments where[ATP] was varied, the free Mg2+ concentration was main-tained at 5 mM. Assays were initiated by the additionof K1112A to a final concentration of 1 mg ml−1 andspecific activities were calculated in milliunits mg−1
(mU mg−1).
2.6. Assay of ADP phosphorylation by carbamoylphosphate
These assays were performed at 30 ◦C as describedby Attwood and Graneri (1991). One unit of enzymeactivity is defined as the amount of enzyme requiredto catalyse the formation of 1 �mol of ATP min−1 at
30 ◦C. All assays were performed in 0.1 M Tris–Cl, pH7.8 at 30 ◦C with 5 mM MgADP; 10 mM biotin; 10 mMMgCl2 in the presence or absence of 0.25 mM acetylCoA. Assays were initiated by the addition of K1112A to
l of Bioc
1746 A. Adina-Zada et al. / The International Journaa final concentration of 2 mg ml−1 and specific activitieswere calculated in mU mg−1.
2.7. Assay of carboxylation of free biotin
The conditions for this radioactive assay for the deter-mination of the rate of carboxylation of free biotin wereessentially as described by Stoll, Ryder, Edwards, andLane (1968). All assays were performed at 30 ◦C andin 0.1 M Tris–Cl, pH 7.8 with the following concen-trations of components unless stated otherwise: 50 mMbiotin; 5 mM MgATP; 10 mM MgCl2; 20 mM NaHCO3;10 �Ci ml−1 NaH14CO3; 0.25 mM acetyl CoA. Assayswere initiated by the addition of K1112A to a final con-centration of 1 mg ml−1. After incubation of the reactionmixture at 30 ◦C for different periods of time, the reac-tion was terminated by a rapid transfer of 0.1 ml aliquotsto 0.9 ml stop solution – a mixture of water and n-octanol (8:1), then kept on ice for 30 min. To removethe excess of H14CO3
−/14CO2, CO2 gas was bubbledthrough the solutions for 40 min on ice. Aliquots (0.5-ml) of gassed solutions were then added to scintillationfluid and the radioactivity due to 14C-carboxybiotin wascounted. Controls were performed in which aliquots ofreaction mixture were transferred to stop solution beforethe initiation of reaction by addition of K1112A. Theenzyme was then added to these quenched aliquots andthe radioactivity measured in these controls was sub-tracted from the samples. The specific radioactivity ofNaH14CO3 was determined by measuring the radioac-tivity of aliquots of reaction mixture with known numberof moles of total NaHCO3. One unit of enzyme activitywas defined as the amount of enzyme required to catal-yse the formation of 1 �mol of carboxybiotin min−1 at30 ◦C. The enzyme specific activities were calculated inmU mg−1.
2.8. Assay of oxaloacetate decarboxylation
The conditions for this spectrophotometric assay ofoxamate-induced oxaloacetate decarboxylation, wherepyruvate formation is measured in a coupled reac-tion with lactate dehydrogenase, were essentially asdescribed by Attwood and Cleland (1986). One unit ofenzyme activity is defined as the amount of enzymerequired to catalyse the formation of 1 �mol of pyru-vate min−1 at 30 ◦C. All assays were performed at 30 ◦Cand in 0.1 M Tris–Cl, pH 7.8 with: 0.2 mM oxaloacetate;
10 mM biotin; 2 mM oxamate; 0.25 mM acetyl CoA.Assays were initiated by the addition of K1112A to afinal concentration of 1 mg ml−1, but prior to its addi-tion a background rate of oxaloacetate decarboxylationhemistry & Cell Biology 40 (2008) 1743–1752
was established, and this rate was subtracted from therate in the presence of K1112A.
2.9. Radioactive assay of pyruvate carboxylatingactivity
The reaction was performed as described by Leggeet al. (1996). Oxaloacetate formation was measuredin a coupled reaction with malate dehydrogenase. Allassays were performed at 30 ◦C and in 0.1 mM Tris–Cl,pH 7.8 with the following concentrations of compo-nents unless stated otherwise: 5 mM MgATP; 10 mMbiotin; 10 mM MgCl2; 20 mM NaHCO3; 10 �Ci ml−1
NaH14CO3; 0.25 mM acetyl CoA; 10 mM pyruvate;0.2 mM NADH; 1 unit of malate dehydrogenase. Assayswere initiated by the addition of K1112A to final con-centration of 1 mg ml−1. After incubation of reactionmixture at 30 ◦C for different periods of time, the reac-tion was quenched by a rapid addition of 50-�l aliquots to50 �l quench solution −1.5 M semicarbazide/1 M HCl.The tubes containing quenched solutions were then leftuncapped in the fume hood for 30 min to allow the 14CO2released on acidification to escape. After this time 50 �lwas spotted on to a 2.5 cm square of Whatman 3 MMfilter paper and dried under a drying lamp for 5 min.The filters were then placed in scintillation fluid, andthe radioactivity in 14C-malate/14C-oxaloacetate on thefilters was counted. Controls were performed in whichaliquots of reaction mixture were transferred to stopsolution before the initiation of reaction by K1112A.The enzyme was then added to these quenched aliquotsand the radioactivity measured in these controls wassubtracted from the samples. Specific radioactivity ofNaH14CO3 was determined by measuring the radioac-tivity of aliquots of reaction mixture with known molesof total NaHCO3.
2.10. Stopped-flow measurements of Mg.formycinA-5′-triphosphate (MgFTP) binding
The fluorescence measurement of the kinetics offormycin A-5′-triphosphate binding were as describedby Geeves, Branson, and Attwood (1995), except exper-iments were performed at 30 ◦C. A KinTek SF-2004instrument (KinTek Corporation, USA) was used inthese measurements. Excitation was at 310 nm and a350 nm cut-on filter was used to monitor emission.
2.11. Carboxyphosphate trapping
The attempts to trap the putative carboxyphosphateintermediate were based on the diazomethane methyla-

l of Biochemistry & Cell Biology 40 (2008) 1743–1752 1747
ttstAa(mat3stCo5odtmosecic(ficp&a
2
blv
v
fM
cas
Table 1Kinetic parameters for the ATP-cleavage reaction catalysed byK1112A in the absence and presence of acetyl CoA
Kinetic parameter − Acetyl CoA + Acetyl CoA
V1 (mU mg−1) 4.8 ± 0.4 3.8 ± 0.4V2 (mU mg−1) 12 ± 2 8.0 ± 0.5
KMgATPi (mM) 0.4 ± 0.1 0.5 ± 0.1
KMgATP (mM) 1.8 ± 0.6 0.6 ± 0.1Kbiotin (mM) 18 ± 13 3.3 ± 1.3
Parameters derived from the fits of ATP-cleavage data, in the pres-
lysine biotinylation site in K1112A completely abol-
A. Adina-Zada et al. / The International Journa
ion method employed by Powers and Meister (1976)o trap the intermediate in the carbamoyl phosphateynthetase reaction. The components of reaction mix-ure were 50 mM triethanolamine-Cl, pH 7.8; 5 mMTP; 20 mM NaHCO3; 10 mM MgCl2. A thiophosphatenalogue of ATP, adenosine 5′-O-(3-thiophosphate)ATP�S) was also used instead of ATP in one experi-ent. The K1112A stock solution was desalted throughG25 Sephadex column prior to use. The reaction solu-
ions and K1112A stock solution were equilibrated at0 ◦C for 10 min prior combining them. The reaction wastarted by addition of 0.585 ml of enzyme (36 mg ml−1)o 1.415 ml of reaction mixture to a final volume of 2 ml.ontrol reactions were also performed with no additionf the enzyme. After allowing the reaction to proceed formin the reaction mixture was quickly mixed with 1 mlf tetrahydrofuran and 1 ml of diazomethane solution iniethyl ether. The resulting suspension was then mixedhoroughly and allowed to react for 5 min. The reactionixture was extracted with 2 ml of ethyl acetate and the
rganic layer was decanted and dried with anhydrousodium sulphate. The extract was analysed directly bylectrospray ionization mass spectrometry, employingollision-induced dissociation (ESI-MS/MS) by directnfusion into an ABI QSTAR Pulsari instrument and gashromatography–electron ionisation mass spectrometryGC–EI-MS) on a Shimadzu GCMS-QP2010 instrumenttted with a 30 metre Restek Rtx®-5MS 0.25 mm IDolumn. Authentic trimethyl carboxyphosphate was pre-ared by modification of the published procedure (Ogita
Knowles, 1988), using sodium dimethyl phosphatend neat methyl chloroformate.
.12. Kinetic data analysis
The data obtained for ATP-cleavage with ATP andiotin as varied substrates were analysed by non-lineareast-squares fits of the data to Eq. (1) and the resultantalues for the kinetic parameters are given in Table 1.
= (V1[MgATP] + V2[MgATP][biotin])
(KMgATPi Kbiotin + Kbiotin[MgATP]
+ KMgATP[biotin] + [MgATP][biotin])
(1)
The kinetic parameters given in Table 2 were obtainedrom non-linear regression fits of the data to the
ichaelis–Menten equation.
The stopped-flow traces of the increase in fluores-ence on binding of MgFTP to K1112A and WT werenalysed by non-linear least-squares fits of the data to aingle first-order exponential.
ence and absence of acetyl CoA to Eq. (1). Values of parameters areshown ± the standard error of the estimate derived from the regressionanalyses.
The pH profiles were analysed by non-linear least-squares regression fits of the data to Eqs. (3)–(6):
v = C
1 + K1/[H+](2)
v = C
1 + [H+]/K1 + K2/[H+](3)
v = C
1 + [H+]/K1 + [H+]2/(K1K2) + K3/[H+]
(4)
v = C
1 + [H+]/K1 + [H+]3/(K1K
22) + K3/[H+]
(5)
3. Results and discussion
3.1. Sequence variation of B. thermodenitrificansPC and abrogation of biotinylation caused byK1112A mutation
On sequencing the wild-type construct several aminoacid differences were noted compared to the sequencepublished by Kondo et al. (1997): F90L, F108L, A151R,V425G, F426I, S565T, D603E, A604R, H643D, G729Aand Y739H (where the first amino acid is that in the pre-viously published sequence and the second is that wefound). However, these amino acid changes observed inour clone are highly conserved in PCs from other Bacil-lus species deposited in the NCBI database (data notshown).
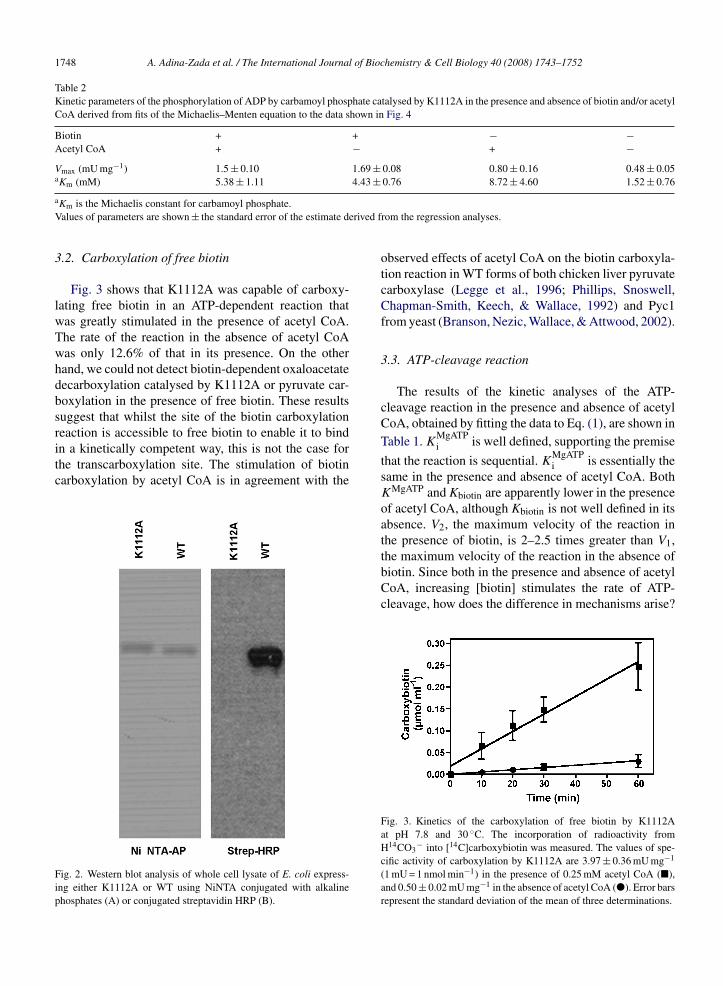
Fig. 2 shows the Western blot analysis of wholecell lysate of E. coli expressing K1112A or WT usingconjugated streptavidin-HRP to detect the biotin moi-ety on these enzymes. As expected, mutation of this
ished biotinylation in vivo whilst the biotinylation of WTis evident. An identical blot was also stained with NiNTAconjugated with alkaline phosphatase to detect equallyexpressed WT and mutant isoforms of PC.
1748 A. Adina-Zada et al. / The International Journal of Biochemistry & Cell Biology 40 (2008) 1743–1752
Table 2Kinetic parameters of the phosphorylation of ADP by carbamoyl phosphate catalysed by K1112A in the presence and absence of biotin and/or acetylCoA derived from fits of the Michaelis–Menten equation to the data shown in Fig. 4
Biotin + + − −Acetyl CoA + − + −Vmax (mU mg−1) 1.5 ± 0.10 1.69 ± 0.08 0.80 ± 0.16 0.48 ± 0.05aKm (mM) 5.38 ± 1.11 4.43 ± 0.76 8.72 ± 4.60 1.52 ± 0.76
erived f
aKm is the Michaelis constant for carbamoyl phosphate.Values of parameters are shown ± the standard error of the estimate d
3.2. Carboxylation of free biotin
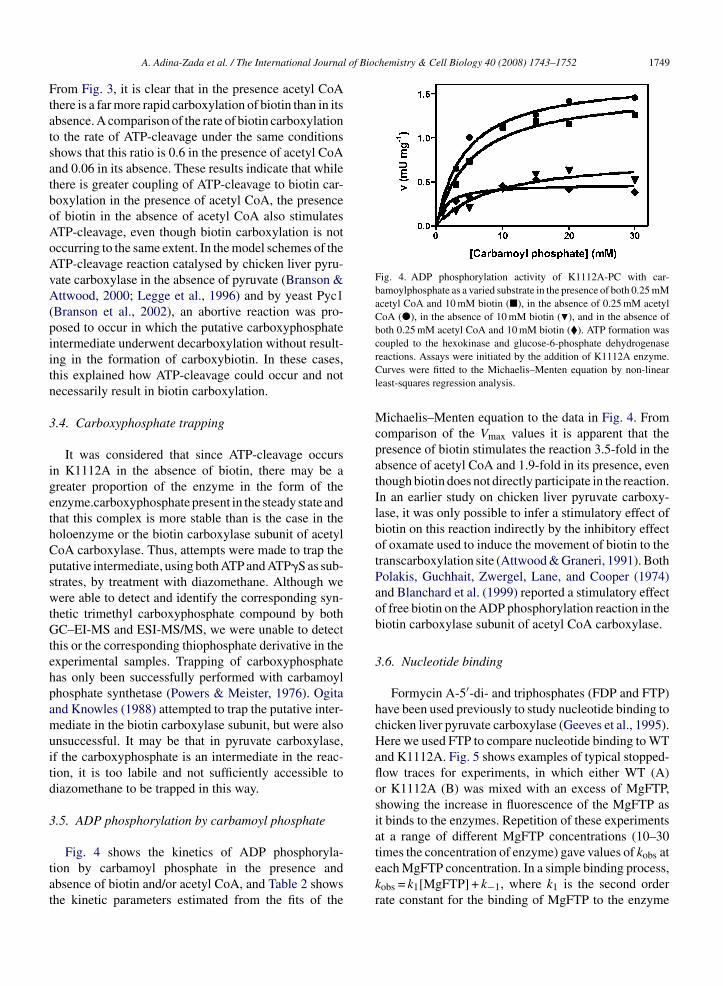
Fig. 3 shows that K1112A was capable of carboxy-lating free biotin in an ATP-dependent reaction thatwas greatly stimulated in the presence of acetyl CoA.The rate of the reaction in the absence of acetyl CoAwas only 12.6% of that in its presence. On the otherhand, we could not detect biotin-dependent oxaloacetatedecarboxylation catalysed by K1112A or pyruvate car-boxylation in the presence of free biotin. These resultssuggest that whilst the site of the biotin carboxylationreaction is accessible to free biotin to enable it to bind
in a kinetically competent way, this is not the case forthe transcarboxylation site. The stimulation of biotincarboxylation by acetyl CoA is in agreement with theFig. 2. Western blot analysis of whole cell lysate of E. coli express-ing either K1112A or WT using NiNTA conjugated with alkalinephosphates (A) or conjugated streptavidin HRP (B).
rom the regression analyses.
observed effects of acetyl CoA on the biotin carboxyla-tion reaction in WT forms of both chicken liver pyruvatecarboxylase (Legge et al., 1996; Phillips, Snoswell,Chapman-Smith, Keech, & Wallace, 1992) and Pyc1from yeast (Branson, Nezic, Wallace, & Attwood, 2002).
3.3. ATP-cleavage reaction
The results of the kinetic analyses of the ATP-cleavage reaction in the presence and absence of acetylCoA, obtained by fitting the data to Eq. (1), are shown inTable 1. KMgATP
i is well defined, supporting the premise
that the reaction is sequential. KMgATPi is essentially the
same in the presence and absence of acetyl CoA. BothKMgATP and Kbiotin are apparently lower in the presenceof acetyl CoA, although Kbiotin is not well defined in itsabsence. V2, the maximum velocity of the reaction inthe presence of biotin, is 2–2.5 times greater than V1,
the maximum velocity of the reaction in the absence ofbiotin. Since both in the presence and absence of acetylCoA, increasing [biotin] stimulates the rate of ATP-cleavage, how does the difference in mechanisms arise?Fig. 3. Kinetics of the carboxylation of free biotin by K1112Aat pH 7.8 and 30 ◦C. The incorporation of radioactivity fromH14CO3
− into [14C]carboxybiotin was measured. The values of spe-cific activity of carboxylation by K1112A are 3.97 ± 0.36 mU mg−1
(1 mU = 1 nmol min−1) in the presence of 0.25 mM acetyl CoA (�),and 0.50 ± 0.02 mU mg−1 in the absence of acetyl CoA (�). Error barsrepresent the standard deviation of the mean of three determinations.
l of Biochemistry & Cell Biology 40 (2008) 1743–1752 1749
FtatsatboAoAvA(piitn
3
igethCpswtGtehpamuitd
3
tat
Fig. 4. ADP phosphorylation activity of K1112A-PC with car-bamoylphosphate as a varied substrate in the presence of both 0.25 mMacetyl CoA and 10 mM biotin (�), in the absence of 0.25 mM acetylCoA (�), in the absence of 10 mM biotin (�), and in the absence ofboth 0.25 mM acetyl CoA and 10 mM biotin (�). ATP formation wascoupled to the hexokinase and glucose-6-phosphate dehydrogenase
A. Adina-Zada et al. / The International Journa
rom Fig. 3, it is clear that in the presence acetyl CoAhere is a far more rapid carboxylation of biotin than in itsbsence. A comparison of the rate of biotin carboxylationo the rate of ATP-cleavage under the same conditionshows that this ratio is 0.6 in the presence of acetyl CoAnd 0.06 in its absence. These results indicate that whilehere is greater coupling of ATP-cleavage to biotin car-oxylation in the presence of acetyl CoA, the presencef biotin in the absence of acetyl CoA also stimulatesTP-cleavage, even though biotin carboxylation is notccurring to the same extent. In the model schemes of theTP-cleavage reaction catalysed by chicken liver pyru-ate carboxylase in the absence of pyruvate (Branson &ttwood, 2000; Legge et al., 1996) and by yeast Pyc1
Branson et al., 2002), an abortive reaction was pro-osed to occur in which the putative carboxyphosphatentermediate underwent decarboxylation without result-ng in the formation of carboxybiotin. In these cases,his explained how ATP-cleavage could occur and notecessarily result in biotin carboxylation.
.4. Carboxyphosphate trapping
It was considered that since ATP-cleavage occursn K1112A in the absence of biotin, there may be areater proportion of the enzyme in the form of thenzyme.carboxyphosphate present in the steady state andhat this complex is more stable than is the case in theoloenzyme or the biotin carboxylase subunit of acetyloA carboxylase. Thus, attempts were made to trap theutative intermediate, using both ATP and ATP�S as sub-trates, by treatment with diazomethane. Although weere able to detect and identify the corresponding syn-
hetic trimethyl carboxyphosphate compound by bothC–EI-MS and ESI-MS/MS, we were unable to detect
his or the corresponding thiophosphate derivative in thexperimental samples. Trapping of carboxyphosphateas only been successfully performed with carbamoylhosphate synthetase (Powers & Meister, 1976). Ogitand Knowles (1988) attempted to trap the putative inter-ediate in the biotin carboxylase subunit, but were also
nsuccessful. It may be that in pyruvate carboxylase,f the carboxyphosphate is an intermediate in the reac-ion, it is too labile and not sufficiently accessible toiazomethane to be trapped in this way.
.5. ADP phosphorylation by carbamoyl phosphate
Fig. 4 shows the kinetics of ADP phosphoryla-ion by carbamoyl phosphate in the presence andbsence of biotin and/or acetyl CoA, and Table 2 showshe kinetic parameters estimated from the fits of the
reactions. Assays were initiated by the addition of K1112A enzyme.Curves were fitted to the Michaelis–Menten equation by non-linearleast-squares regression analysis.
Michaelis–Menten equation to the data in Fig. 4. Fromcomparison of the Vmax values it is apparent that thepresence of biotin stimulates the reaction 3.5-fold in theabsence of acetyl CoA and 1.9-fold in its presence, eventhough biotin does not directly participate in the reaction.In an earlier study on chicken liver pyruvate carboxy-lase, it was only possible to infer a stimulatory effect ofbiotin on this reaction indirectly by the inhibitory effectof oxamate used to induce the movement of biotin to thetranscarboxylation site (Attwood & Graneri, 1991). BothPolakis, Guchhait, Zwergel, Lane, and Cooper (1974)and Blanchard et al. (1999) reported a stimulatory effectof free biotin on the ADP phosphorylation reaction in thebiotin carboxylase subunit of acetyl CoA carboxylase.
3.6. Nucleotide binding
Formycin A-5′-di- and triphosphates (FDP and FTP)have been used previously to study nucleotide binding tochicken liver pyruvate carboxylase (Geeves et al., 1995).Here we used FTP to compare nucleotide binding to WTand K1112A. Fig. 5 shows examples of typical stopped-flow traces for experiments, in which either WT (A)or K1112A (B) was mixed with an excess of MgFTP,showing the increase in fluorescence of the MgFTP asit binds to the enzymes. Repetition of these experimentsat a range of different MgFTP concentrations (10–30
times the concentration of enzyme) gave values of kobs ateach MgFTP concentration. In a simple binding process,kobs = k1[MgFTP] + k−1, where k1 is the second orderrate constant for the binding of MgFTP to the enzyme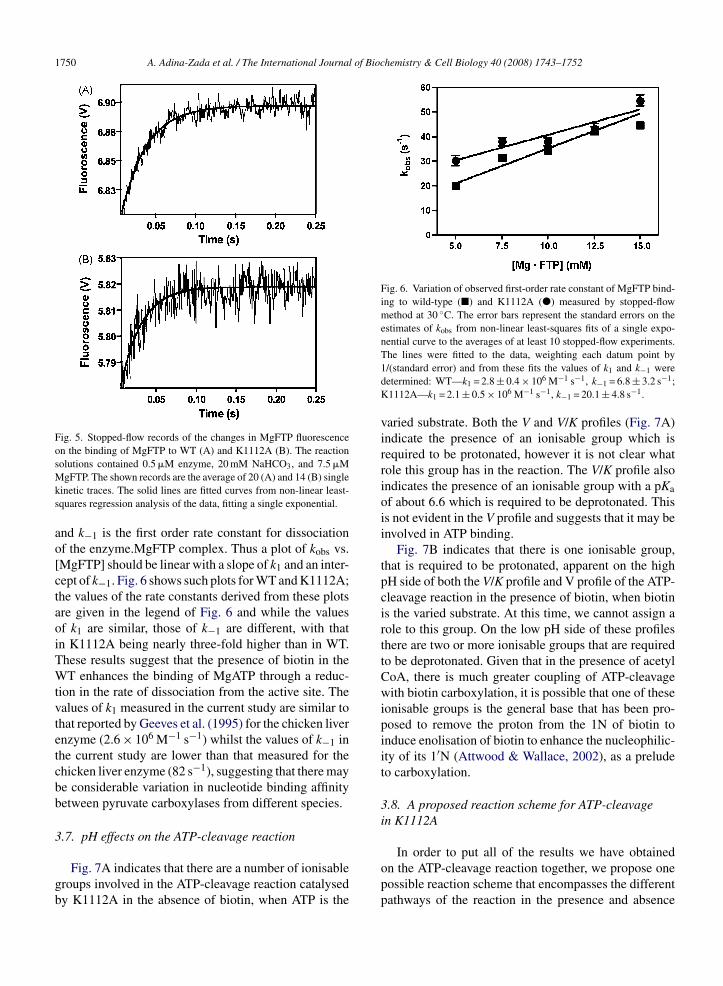
1750 A. Adina-Zada et al. / The International Journal of Biochemistry & Cell Biology 40 (2008) 1743–1752
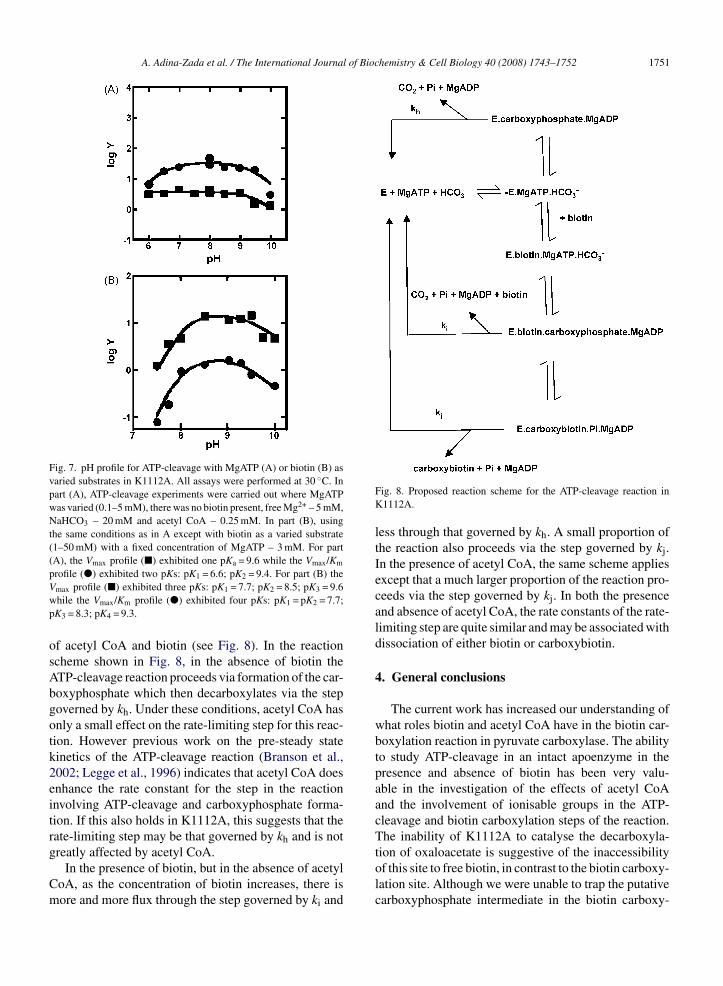
Fig. 5. Stopped-flow records of the changes in MgFTP fluorescenceon the binding of MgFTP to WT (A) and K1112A (B). The reaction
Fig. 6. Variation of observed first-order rate constant of MgFTP bind-ing to wild-type (�) and K1112A (�) measured by stopped-flowmethod at 30 ◦C. The error bars represent the standard errors on theestimates of kobs from non-linear least-squares fits of a single expo-nential curve to the averages of at least 10 stopped-flow experiments.
solutions contained 0.5 �M enzyme, 20 mM NaHCO3, and 7.5 �MMgFTP. The shown records are the average of 20 (A) and 14 (B) singlekinetic traces. The solid lines are fitted curves from non-linear least-squares regression analysis of the data, fitting a single exponential.
and k−1 is the first order rate constant for dissociationof the enzyme.MgFTP complex. Thus a plot of kobs vs.[MgFTP] should be linear with a slope of k1 and an inter-cept of k−1. Fig. 6 shows such plots for WT and K1112A;the values of the rate constants derived from these plotsare given in the legend of Fig. 6 and while the valuesof k1 are similar, those of k−1 are different, with thatin K1112A being nearly three-fold higher than in WT.These results suggest that the presence of biotin in theWT enhances the binding of MgATP through a reduc-tion in the rate of dissociation from the active site. Thevalues of k1 measured in the current study are similar tothat reported by Geeves et al. (1995) for the chicken liverenzyme (2.6 × 106 M−1 s−1) whilst the values of k−1 inthe current study are lower than that measured for thechicken liver enzyme (82 s−1), suggesting that there maybe considerable variation in nucleotide binding affinitybetween pyruvate carboxylases from different species.
3.7. pH effects on the ATP-cleavage reaction
Fig. 7A indicates that there are a number of ionisablegroups involved in the ATP-cleavage reaction catalysedby K1112A in the absence of biotin, when ATP is the
The lines were fitted to the data, weighting each datum point by1/(standard error) and from these fits the values of k1 and k−1 weredetermined: WT—k1 = 2.8 ± 0.4 × 106 M−1 s−1, k−1 = 6.8 ± 3.2 s−1;K1112A—k1 = 2.1 ± 0.5 × 106 M−1 s−1, k−1 = 20.1 ± 4.8 s−1.
varied substrate. Both the V and V/K profiles (Fig. 7A)indicate the presence of an ionisable group which isrequired to be protonated, however it is not clear whatrole this group has in the reaction. The V/K profile alsoindicates the presence of an ionisable group with a pKaof about 6.6 which is required to be deprotonated. Thisis not evident in the V profile and suggests that it may beinvolved in ATP binding.
Fig. 7B indicates that there is one ionisable group,that is required to be protonated, apparent on the highpH side of both the V/K profile and V profile of the ATP-cleavage reaction in the presence of biotin, when biotinis the varied substrate. At this time, we cannot assign arole to this group. On the low pH side of these profilesthere are two or more ionisable groups that are requiredto be deprotonated. Given that in the presence of acetylCoA, there is much greater coupling of ATP-cleavagewith biotin carboxylation, it is possible that one of theseionisable groups is the general base that has been pro-posed to remove the proton from the 1N of biotin toinduce enolisation of biotin to enhance the nucleophilic-ity of its 1′N (Attwood & Wallace, 2002), as a preludeto carboxylation.
3.8. A proposed reaction scheme for ATP-cleavagein K1112A
In order to put all of the results we have obtainedon the ATP-cleavage reaction together, we propose onepossible reaction scheme that encompasses the differentpathways of the reaction in the presence and absence
A. Adina-Zada et al. / The International Journal of Biochemistry & Cell Biology 40 (2008) 1743–1752 1751
Fig. 7. pH profile for ATP-cleavage with MgATP (A) or biotin (B) asvaried substrates in K1112A. All assays were performed at 30 ◦C. Inpart (A), ATP-cleavage experiments were carried out where MgATPwas varied (0.1–5 mM), there was no biotin present, free Mg2+ – 5 mM,NaHCO3 – 20 mM and acetyl CoA – 0.25 mM. In part (B), usingthe same conditions as in A except with biotin as a varied substrate(1–50 mM) with a fixed concentration of MgATP – 3 mM. For part(A), the Vmax profile (�) exhibited one pKa = 9.6 while the Vmax/Km
profile (�) exhibited two pKs: pK = 6.6; pK = 9.4. For part (B) theVwp
osAbgotk2eitrg
Cm
1 2
max profile (�) exhibited three pKs: pK1 = 7.7; pK2 = 8.5; pK3 = 9.6hile the Vmax/Km profile (�) exhibited four pKs: pK1 = pK2 = 7.7;K3 = 8.3; pK4 = 9.3.
f acetyl CoA and biotin (see Fig. 8). In the reactioncheme shown in Fig. 8, in the absence of biotin theTP-cleavage reaction proceeds via formation of the car-oxyphosphate which then decarboxylates via the stepoverned by kh. Under these conditions, acetyl CoA hasnly a small effect on the rate-limiting step for this reac-ion. However previous work on the pre-steady stateinetics of the ATP-cleavage reaction (Branson et al.,002; Legge et al., 1996) indicates that acetyl CoA doesnhance the rate constant for the step in the reactionnvolving ATP-cleavage and carboxyphosphate forma-ion. If this also holds in K1112A, this suggests that theate-limiting step may be that governed by kh and is not
reatly affected by acetyl CoA.In the presence of biotin, but in the absence of acetyloA, as the concentration of biotin increases, there isore and more flux through the step governed by ki and
Fig. 8. Proposed reaction scheme for the ATP-cleavage reaction inK1112A.
less through that governed by kh. A small proportion ofthe reaction also proceeds via the step governed by kj.In the presence of acetyl CoA, the same scheme appliesexcept that a much larger proportion of the reaction pro-ceeds via the step governed by kj. In both the presenceand absence of acetyl CoA, the rate constants of the rate-limiting step are quite similar and may be associated withdissociation of either biotin or carboxybiotin.
4. General conclusions
The current work has increased our understanding ofwhat roles biotin and acetyl CoA have in the biotin car-boxylation reaction in pyruvate carboxylase. The abilityto study ATP-cleavage in an intact apoenzyme in thepresence and absence of biotin has been very valu-able in the investigation of the effects of acetyl CoAand the involvement of ionisable groups in the ATP-cleavage and biotin carboxylation steps of the reaction.The inability of K1112A to catalyse the decarboxyla-
tion of oxaloacetate is suggestive of the inaccessibilityof this site to free biotin, in contrast to the biotin carboxy-lation site. Although we were unable to trap the putativecarboxyphosphate intermediate in the biotin carboxy-
l of Bioc
Academy of Sciences of the United States of America, 60,986–991.
1752 A. Adina-Zada et al. / The International Journa
lation reaction, the previously published evidence stillsubstantiates its existence as an intermediate.
Acknowledgements
This work was supported in part by the AustralianResearch Council grant DP0346807 to J.C.W., theNational Institutes of Health grant 1 R01 GM070455to W.W.C., J.C.W. and P.V.A.
References
Attwood, P. V. (1993). Locus of action of acetyl CoA in the biotin-carboxylation reaction of pyruvate carboxylase. Biochemistry, 32,12736–12742.
Attwood, P. V. (1995). The structure and the mechanism of action ofpyruvate carboxylase. International Journal of Biochemistry andCell Biology, 27, 231–249.
Attwood, P. V., & Cleland, W. W. (1986). Decarboxylation of oxalac-etate by pyruvate carboxylase. Biochemistry, 25, 8191–8196.
Attwood, P. V., & Graneri, B. D. L. A. (1991). Pyruvate carboxylasecatalysis of phosphate transfer between carbamoyl phosphate andADP. Biochemical Journal, 273, 443–448.
Attwood, P. V., & Graneri, B. D. L. A. (1992). Bicarbonate-dependentATP-cleavage catalysed by pyruvate carboxylase in the absence ofpyruvate. Biochemical Journal, 287, 1011–1017.
Attwood, P. V., & Wallace, J. C. (2002). Chemical and catalytic mecha-nisms of carboxyl transfer reactions in biotin-dependent enzymes.Accounts of Chemical Research, 35, 113–120.
Blanchard, C. Z., Chapman-Smith, A., Wallace, J. C., & Waldrop,G. L. (1999). The biotin domain peptide from the biotin carboxylcarrier protein of Escherichia coli acetyl-CoA carboxylase causes amarked increase in the catalytic efficiency of biotin carboxylase andcarboxyltransferase relative to free biotin. Journal of BiologicalChemistry, 274, 31767–31769.
Branson, J. P., & Attwood, P. V. (2000). Effects of Mg2+ on thepre-steady-state kinetics of the biotin carboxylation reaction ofpyruvate carboxylase. Biochemistry, 39, 7480–7491.
Branson, J. P., Nezic, M., Jitrapakdee, S., Wallace, J. C., & Attwood,P. V. (2004). Kinetic characterization of yeast pyruvate carboxy-lase isozyme Pyc1 and the Pyc1 mutant, C249A. Biochemistry, 43,1075–1081.
Branson, J. P., Nezic, M., Wallace, J. C., & Attwood, P. V. (2002).
Kinetic characterization of yeast pyruvate carboxylase isozymepyc1. Biochemistry, 41, 4459–4466.Geeves, M. A., Branson, J. P., & Attwood, P. V. (1995). Kineticsof nucleotide binding to pyruvate carboxylase. Biochemistry, 37,11846–11854.
hemistry & Cell Biology 40 (2008) 1743–1752
Helmerhorst, E., & Stokes, G. B. (1980). Microcentrifuge desalting: arapid, quantitative method for desalting small amounts of protein.Analytical Biochemistry, 104, 130–135.
Jitrapakdee, S., Adina-Zada, A., Besant, P. G., Surinya, K. H., Cle-land, W. W., Wallace, J. C., et al. (2007). Differential regulationof the yeast isozymes of pyruvate carboxylase and the locus ofaction of acetyl CoA. International Journal of Biochemistry andCell Biology, 39, 1211–1223.
Kaziro, Y., Hass, L. F., Boyer, P. D., & Ochoa, S. (1962). Mecha-nism of the propionyl carboxylase reaction. Journal of BiologicalChemistry, 237, 1460–1468.
Kondo, H., Kazuta, Y., Saito, A., & Fuji, K. (1997). Cloning andnucleotide sequence of bacillus stearothermophilus pyruvate car-boxylase. Gene, 191, 47–50.
Legge, G. B., Branson, J. P., & Attwood, P. V. (1996). Effects ofacetyl CoA on the pre-steady-state kinetics of the biotin car-boxylation reaction of pyruvate carboxylase. Biochemistry, 35,3849–3856.
Ogita, T., & Knowles, J. R. (1988). On the intermediacy of car-boxyphosphate in biotin-dependent carboxylations. Biochemistry,21, 8028–8033.
Phillips, N. F. B., Snoswell, M. A., Chapman-Smith, A., Keech, D. B.,& Wallace, J. C. (1992). Isolation of a carboxyphosphate interme-diate and the locus of acetyl CoA action in the pyruvate carboxylasereaction. Biochemistry, 31, 9445–9450.
Polakis, S. E., Guchhait, R. B., Zwergel, E. E., Lane, M. D., &Cooper, T. G. (1974). Acetyl coenzyme A carboxylase systemof Escherichia coli. Studies on the mechanisms of the biotincarboxylase- and carboxyltransferase-catalyzed reactions. Journalof Biological Chemistry, 249, 6657–6667.
Powers, S. G., & Meister, A. (1976). Identification of enzyme-bound activated CO2 as carbonic-phosphoric anhydride: isolationof the corresponding trimethyl derivative from the active site ofglutamine-dependent carbamyl phosphate synthetase. Proceedingsof the National Academy of Sciences of the United States of Amer-ica, 73, 3020–3024.
St Maurice, M., Reinhardt, L., Surinya, K. H., Attwood, P. V., Wallace,J. C., Cleland, W. W., et al. (2007). Domain architecture of pyruvatecarboxylase, a biotin-dependent multifunctional enzyme. Science,317, 1076–1079.
Stoll, E., Ryder, E., Edwards, J. B., & Lane, M. D. (1968). Liveracetyl coenzyme A carboxylase: activation of model partialreactions by tricarboxylic acids. Proceedings of the National
Ward, D. C., Cerami, A., Reich, E., Acs, G., & Altwerger, L. (1969).Biochemical studies of the nucleoside analogue, formycin. Journalof Biological Chemistry, 244, 3243–3250.
Related Documents











