UNCORRECTED PROOF Please cite this article in press as: D. Paris et al., Inhibition of A production by NF-B inhibitors, Neurosci. Lett. (2006), doi:10.1016/j.neulet.2006.12.029 ARTICLE IN PRESS +Model NSL 23749 1–6 Neuroscience Letters xxx (2006) xxx–xxx Inhibition of A production by NF-B inhibitors 3 Daniel Paris ∗ , Nikunj Patel, Amita Quadros, Monica Linan, Pancham Bakshi, Ghania Ait-Ghezala, Michael Mullan 4 5 Roskamp Institute, 2040 Whitfield Avenue, Sarasota, FL 34243, USA 6 Received 26 April 2006; received in revised form 9 October 2006; accepted 21 December 2006 7 Abstract 8 The transcription factor nuclear factor B (NF-B) is widely expressed in the nervous system and increased NF-B immunoreactivity has been observed in Alzheimer’s disease (AD) brains in the nuclei of neurons within the vicinity of diffuse -amyloid plaques. -Amyloid (A) peptides are the main constituent of senile plaques and are known to stimulate NF-B activity. In the present study, we investigated the effect of various NF-B inhibitors on the production of A 1–40 ,A 1–42 , secreted APP (sAPP and sAPP) and APP C-terminal fragments (APP-CTF) using CHO cells overexpressing the -amyloid precursor protein (APP). Our data show that NF-B inhibitors decrease both A 1–40 and A 1–42 production. In addition, we show that some NF-B inhibitors decrease sAPP and APP-CTF suggesting that they reduce the -secretase cleavage of APP. Altogether our data suggest that NF-B inhibitors may be of therapeutic importance for the treatment of AD pathology not only by blocking inflammatory processes but also by directly inhibiting the production of A peptides. 9 10 11 12 13 14 15 16 © 2006 Published by Elsevier Ireland Ltd. 17 Keywords: Alzheimer; APP processing; NF-B; Amyloid 18 19 The brains of Alzheimer’s disease (AD) patients are character- 20 ized by the accumulation of a 38–43-amino acids peptide termed 21 amyloid -peptide or A [16]. This peptide is the major com- 22 ponent of senile plaques found in AD and is proposed as a key 23 player in the pathobiology of AD since all familial forms of the 24 disease are associated with an increased A accumulation. A 25 is a proteolytic fragment of the large amyloid precursor pro- 26 tein (APP) [6]. It is cleaved by -secretase generating secreted 27 APP (sAPP) and the carboxyl terminal intracellular fragment 28 (APP-CTF) which is further cleaved by -secretase to produce 29 A peptides [17], whereas the -secretase cleavage of APP takes 30 place within the A sequence precluding A production, leading 31 to the secretion of sAPP and the accumulation of membrane- 32 coupled APP CTF-. The -secretase cleavage is not precise and 33 leads to the production of a series of A peptides of 38–43 amino 34 acids. A 1–42 is more fibrillogenic than shorter A peptides but 35 a higher proportion of A is produced as A 1–40 compared to 36 A 1–42 . Elevated A 1–42 concentrations are thought to drive the 37 formation of insoluble fibrils resulting in the deposition of A 38 as amyloid plaques. For this reason A 1–42 is considered to have 39 ∗ Corresponding author. Tel.: +1 941 752 2949; fax: +1 941 752 2948. E-mail address: [email protected] (D. Paris). a causative role in the etiology of AD and therapies particularly 40 targeting A 1–42 are expected to have the greatest impact on 41 -amyloid pathology. 42 The transcription factor NF-B is widely expressed in the 43 nervous system and particularly in synaptic terminals. In non- 44 stimulated cells, NF-B is sequestered in the cytoplasm by 45 inhibitory units called IB proteins. Stimulation of cells by var- 46 ious inducers causes IB phosphorylation and its subsequent 47 degradation by the proteasome [21]. Liberated NF-B is trans- 48 ported in the nucleus, where it induces transcription of target 49 genes, including IB as an autoregulatory loop [21]. 50 Studies of postmortem brain tissue from patients with AD 51 have revealed increased NF-B immunoreactivity in neurons 52 and astrocytes in the immediate vicinity of -amyloid plaques 53 [7,19]. Many genes newly induced in AD are under immediate- 54 early transcriptional control of NF-B [11] also suggesting that 55 NF-B pathway is activated in AD brains. Other studies have 56 shown that A peptides can activate NF-B in primary neu- 57 rons and astrocytes [2] suggesting a molecular mechanism by 58 which A may act during AD pathogenesis. Recently, it has been 59 reported that indomethacin (a non steroidal anti-inflammatory 60 drug) can lead to a reduction in the level of A peptides and NF- 61 B in the brains of a transgenic mouse model of AD (Tg2576) 62 whereas another anti-inflammatory compound, nimesulide had 63 1 0304-3940/$ – see front matter © 2006 Published by Elsevier Ireland Ltd. 2 doi:10.1016/j.neulet.2006.12.029

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

N
3
4
5
6
7
A8
oaNcIAi
9
10
11
12
13
14
15
16
©17
K18
19
T20
i21
a22
p23
p24
d25
i26
t27
A28
(29
A30
p31
t32
c33
l34
a35
a36
A37
f38
a39
1 02 d
OO
F
ARTICLE IN PRESS+ModelSL 23749 1–6
Neuroscience Letters xxx (2006) xxx–xxx
Inhibition of A� production by NF-�B inhibitors
Daniel Paris ∗, Nikunj Patel, Amita Quadros, Monica Linan, Pancham Bakshi,Ghania Ait-Ghezala, Michael Mullan
Roskamp Institute, 2040 Whitfield Avenue, Sarasota, FL 34243, USA
Received 26 April 2006; received in revised form 9 October 2006; accepted 21 December 2006
bstract
The transcription factor nuclear factor �B (NF-�B) is widely expressed in the nervous system and increased NF-�B immunoreactivity has beenbserved in Alzheimer’s disease (AD) brains in the nuclei of neurons within the vicinity of diffuse �-amyloid plaques. �-Amyloid (A�) peptidesre the main constituent of senile plaques and are known to stimulate NF-�B activity. In the present study, we investigated the effect of variousF-�B inhibitors on the production of A�1–40, A�1–42, secreted APP (sAPP� and sAPP�) and APP C-terminal fragments (APP-CTF) using CHO
ells overexpressing the �-amyloid precursor protein (APP). Our data show that NF-�B inhibitors decrease both A�1–40 and A�1–42 production.
D P
Rn addition, we show that some NF-�B inhibitors decrease sAPP� and APP-CTF� suggesting that they reduce the �-secretase cleavage of APP.ltogether our data suggest that NF-�B inhibitors may be of therapeutic importance for the treatment of AD pathology not only by blocking
nflammatory processes but also by directly inhibiting the production of A� peptides.2006 Published by Elsevier Ireland Ltd.
a 40
t 41
� 42
43
n 44
s 45
i 46
i 47
d 48
p 49
g 50
51
h 52
a 53
[ 54
e 55
N 56
OR
RE
CTE
eywords: Alzheimer; APP processing; NF-�B; Amyloid
he brains of Alzheimer’s disease (AD) patients are character-zed by the accumulation of a 38–43-amino acids peptide termedmyloid �-peptide or A� [16]. This peptide is the major com-onent of senile plaques found in AD and is proposed as a keylayer in the pathobiology of AD since all familial forms of theisease are associated with an increased A� accumulation. A�s a proteolytic fragment of the large amyloid precursor pro-ein (APP) [6]. It is cleaved by �-secretase generating secretedPP� (sAPP�) and the carboxyl terminal intracellular fragment
APP-CTF�) which is further cleaved by �-secretase to produce� peptides [17], whereas the �-secretase cleavage of APP takeslace within the A� sequence precluding A� production, leadingo the secretion of sAPP� and the accumulation of membrane-oupled APP CTF-�. The �-secretase cleavage is not precise andeads to the production of a series of A� peptides of 38–43 aminocids. A�1–42 is more fibrillogenic than shorter A� peptides buthigher proportion of A� is produced as A�1–40 compared to
UN
C
Please cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
�1–42. Elevated A�1–42 concentrations are thought to drive theormation of insoluble fibrils resulting in the deposition of A�s amyloid plaques. For this reason A�1–42 is considered to have
∗ Corresponding author. Tel.: +1 941 752 2949; fax: +1 941 752 2948.E-mail address: [email protected] (D. Paris).
s 57
r 58
w 59
rd�w
304-3940/$ – see front matter © 2006 Published by Elsevier Ireland Ltd.oi:10.1016/j.neulet.2006.12.029
causative role in the etiology of AD and therapies particularlyargeting A�1–42 are expected to have the greatest impact on-amyloid pathology.
The transcription factor NF-�B is widely expressed in theervous system and particularly in synaptic terminals. In non-timulated cells, NF-�B is sequestered in the cytoplasm bynhibitory units called I�B proteins. Stimulation of cells by var-ous inducers causes I�B phosphorylation and its subsequentegradation by the proteasome [21]. Liberated NF-�B is trans-orted in the nucleus, where it induces transcription of targetenes, including I�B as an autoregulatory loop [21].
Studies of postmortem brain tissue from patients with ADave revealed increased NF-�B immunoreactivity in neuronsnd astrocytes in the immediate vicinity of �-amyloid plaques7,19]. Many genes newly induced in AD are under immediate-arly transcriptional control of NF-�B [11] also suggesting thatF-�B pathway is activated in AD brains. Other studies have
hown that A� peptides can activate NF-�B in primary neu-ons and astrocytes [2] suggesting a molecular mechanism byhich A� may act during AD pathogenesis. Recently, it has been
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
eported that indomethacin (a non steroidal anti-inflammatory 60
rug) can lead to a reduction in the level of A� peptides and NF- 61
B in the brains of a transgenic mouse model of AD (Tg2576) 62
hereas another anti-inflammatory compound, nimesulide had 63

ED
IN+ModelN
2 nce L
n64
N65
i66
t67
w68
t69
e70
t71
w72
e73
o74
w75
A76
t77
o78
79
i80
s81
a82
a83
o84
o85
N86
s87
t88
o89
K90
i91
n92
l93
i94
t95
e96
N97
s98
s99
k100
a101
[102
t103
104
(105
b106
c107
M108
U109
s110
A111
B112
i113
i114
w115
e116
k117
t118
m119
d120
s 121
t 122
p 123
r 124
N 125
f 126
w 127
A 128
� 129
p 130
m 131
z 132
A 133
c 134
A 135
136
k 137
u 138
i 139
c 140
a 141
t 142
a 143
144
s 145
B 146
( 147
w 148
( 149
o 150
d 151
E 152
t 153
a 154
i 155
m 156
C 157
l 158
A 159
s 160
161
m 162
p 163
N 164
A 165
N 166
e 167
c 168
W 169
m 170
r 171
t 172
i 173
NC
OR
RE
CT
ARTICLESL 23749 1–6
D. Paris et al. / Neuroscie
o effect on either A� peptides or NF-�B [18] suggesting thatF-�B activation blockade could reduce the amyloid pathology
n Tg2576 mice. Since A� itself is known to induce NF-�B,he reduced NF-�B activity observed in Tg2576 mice treatedith indomethacin could alternatively be secondary to the reduc-
ion of A� accumulation observed. We therefore investigated theffect of various NF-�B inhibitors on the production of A� pep-ides using Chinese Hamster Ovary cells stably transfected withildtype APP 751 (7W CHO), overproducing human A�. The
ffect of NF-�B inhibitors was also tested on the accumulationf APP C-terminal fragments (APP-CTF) in order to determinehether NF-�B inhibitors could impact �-secretase activity.dditionally, we investigated the effect of NF-�B inhibitors on
he secretion of sAPP� and sAPP� to determine a possible effectf NF-�B inhibitors on �-secretase or �-secretase activities.
For this study, we tested the effect of different NF-�Bnhibitors (NF-�B SN50, parthenolide, hypoestoxide, cap-aicin, andrographolide, Caffeic Acid Phenethyl Ester (CAPE),rtemisinin, celastrol, 6-amino-4-(4-phenoxyphenyl-ethyl-mino)quinazoline (quinazoline), isohelenin, kamebakaurin)n the production of A�1–40 and A�1–42 peptides by CHO cellsverexpressing APP. NF-�B SN50 is a selective, cell permeableF-�B inhibitor peptide which contains the nuclear localization
equence (NLS) of the transcription factor NF-�B p50 linkedo the hydrophobic region (h-region) of the signal peptidef Kaposi fibroblast growth factor (K-FGF). The N-terminal-FGF h-region confers cell-permeability, while the NLS
nhibits translocation of the NF-�B active complex into theucleus [14]. Parthenolide and isohelenin are two sequiterponeactone known to block NF-�B activation [3]. Andrographolides a bicyclic diterpenoid lactone, which inhibits NF-�B bindingo DNA [8]. CAPE is an active component of propolis from hon-ybee hives that prevents the translocation of the p65 subunit ofF-�B to the nucleus [15]. Artemisinin (a potent anti-malarial
esquiterpene) and celastrol (sesquiterpene ester) have beenhown to block NF-�B activation [1,10]. Kamebakaurin is aaurane diterpene, which prevents the DNA binding activity ofctivated NF-�B but does not block its nuclear translocation13]. Quinazoline is a low molecular weight inhibitor of NF-�Branscriptional activation [20].
Briefly, 7W CHO cells were maintained in DMEMATCC, Manassas, VA, USA) supplemented with 10% fetalovine serum (Invitrogen, Carlsbad, CA, USA), 1× Peni-illin–Streptomycin Fungizone mixture (Cambrex, Rockland,E, USA) and 0.3% Geneticin (Invitrogen, Carlsbad, CA,SA). Cells were plated on 96 well-culture plates at a den-
ity of 5 × 104 cells per well in 200 �L of culture medium.ll the inhibitors used in this study were purchased from EMDiosciences Inc. (San Diego, CA, USA). The concentration of
nhibitors used to achieve NF-�B inhibition were chosen accord-ng to referenced publications. For each inhibitor, experimentsere performed in quadruplicate and A�1–40 and A�1–42 lev-
ls were evaluated following 18 h of incubation using ELISA
UPlease cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
its (Invitrogen Biosource Inc., Carlsbad, CA, USA) accordingo the recommendations of the manufacturer. For the measure-
ent of A�1–40, samples were diluted 20-fold with the sampleiluent provided in the kits whereas for A�1–42 determination,
cdp�
PR
OO
F
PRESSetters xxx (2006) xxx–xxx
amples were diluted only two-fold. Results were expressed ashe percentage of A�1–40 or A�1–42 measured in control sam-les. No direct toxicity (monitored by lactate-dehydrogenaseelease in the culture medium) was observed for the doses ofF-�B inhibitors tested (data not shown). Interestingly, the dif-
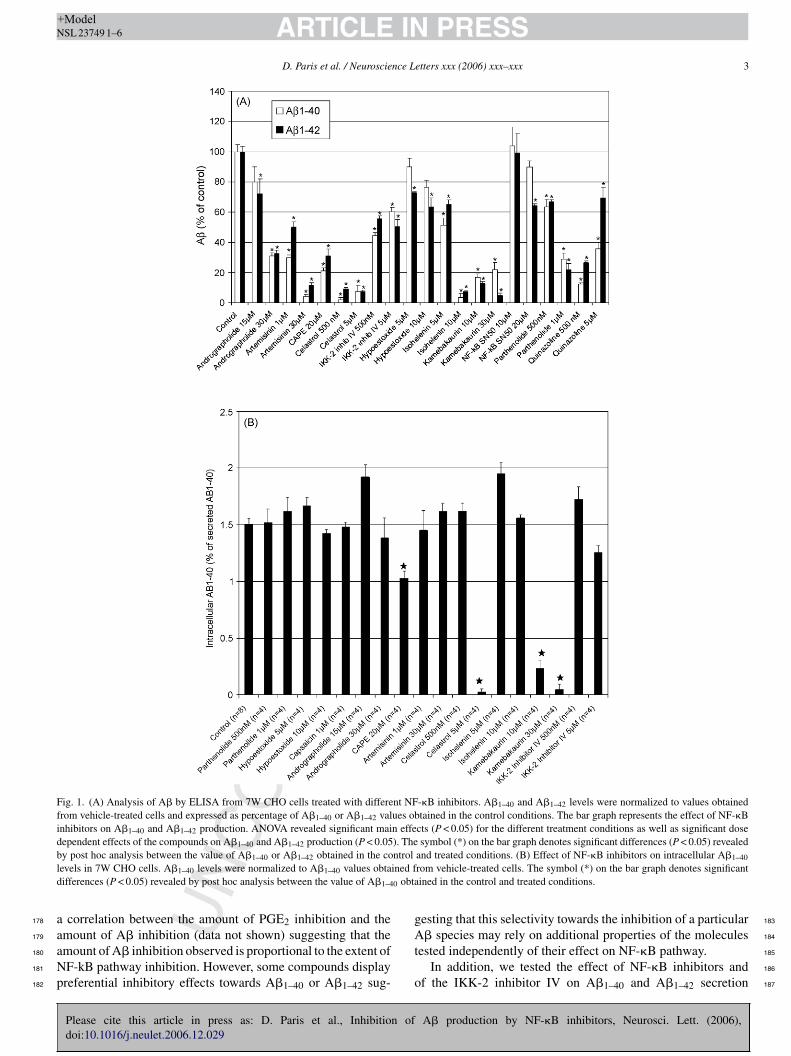
erent NF-�B inhibitors tested appear to diminish A� productionith different potency and show a differential effect towards�1–40 and A�1–42 production (Fig. 1A). For instance, NF-B SN50 at 20 �M inhibits A�1–42 (∼40% inhibition) moreotently than A�1–40 (∼9%). Similarly, hypoestoxide displaysore potency towards A�1–42 than A�1–40 inhibition. Quina-
oline, artemisinin (at 1 �M) and isohelenin (at 5 �M) inhibit�1–40 secretion more potently than A�1–42 whereas the other
ompounds tested display analogous effects on A�1–40 and�1–42 production.NF-�B activation can be blocked indirectly by inhibiting I
appa B kinase (IKK-2). We therefore tested a cell-permeablereidocarboxamido thiophene compound that acts as a potentnhibitor of IKK-2 ([5-(p-Fluorophenyl)-2-ureido]thiophene-3-arboxamide or IKK-2 inhibitor IV). IKK-2 inhibitor IV alsoppears to significantly inhibit both A�1–40 and A�1–42 produc-ion (Fig. 1A). These data support the concept that the secretionnd/or production of A�1–40 and A�1–42 is NF-�B dependent.
In order to verify that NF-�B inhibitors were impacting theecretion of A�, we measured the intracellular level of A�1–40.riefly, 7W CHO were treated with different NF-�B inhibitors
Fig. 1B). Following 18 h of incubation, cells were washedith PBS and lyzed with 100 �L of ice-cold M-PER® Reagent
Pierce, IL, USA) containing 1 mM phenylmethanesulfonyl flu-ride and 1 mM sodium orthovanadate. Cellular lysates wereiluted five-fold with the standard diluent provided in the A�1–40LISA kit (Biosource, CA) before intracellular A�1–40 quan-
ification. Data show that intracellular A�1–40 represent onlysmall fraction of the A�1–40 produced by the cells (approx-
mately 1.5% of the amount of A�1–40 secreted in the cultureedium). Interestingly, among the NF-�B inhibitors tested, onlyAPE, celastrol and kamebakaurin were able to significantly
ower intracellular A�1–40, but no intracellular accumulation of� was observed with any of the compounds suggesting that the
ecretion of A� is not impacted by NF-�B inhibition (Fig. 1B).In order to verify that NF-�B inhibition was achieved, we
easured the production of prostaglandin E2 (PGE2), a mainroduct of cyclooxygenase-2 (whose expression is regulated byF-�B) employing the same culture paradigm used to measure� production. This was necessarily an indirect way of assessingF-�B inhibition as the inhibitors are purported to achieve their
ffects by different mechanisms (some inhibit nuclear translo-ation of NF-�B, while others inhibit its binding to DNA).e analyzed conditioned cell culture media for PGE2 (a mainetabolite of cyclooxygenase-2, expression of which is NF-�B
egulated), using a commercially available ELISA according tohe strict recommendations of the manufacturer (Cayman Chem-cal, Ann Arbor, MI, USA), following treatment of 7W CHO
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
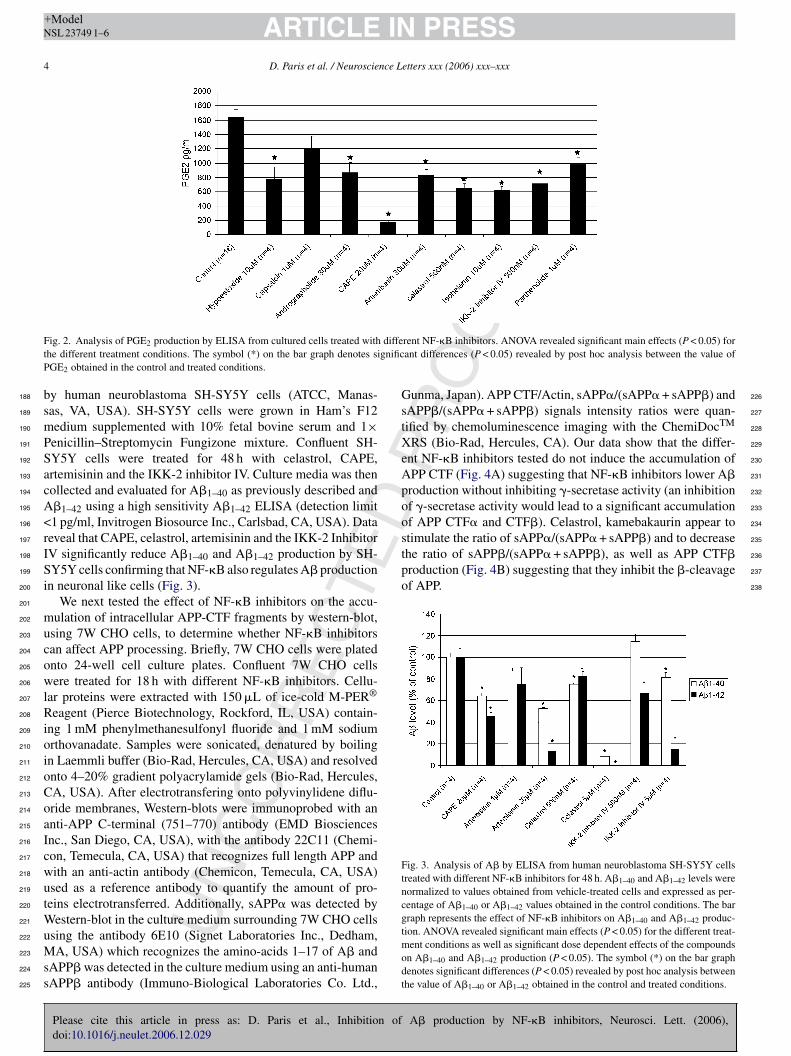
ells with various NF-�B inhibitors for 18 h. Our results (Fig. 2) 174
emonstrate a differential inhibition of 7W CHO cell PGE2 175
roduction showing that the NF-�B inhibitors tested inhibit NF- 176
B activity with different potencies [4]. Interestingly, we found 177
OR
RE
CTE
D P
RO
OF
ARTICLE IN PRESS+ModelNSL 23749 1–6
D. Paris et al. / Neuroscience Letters xxx (2006) xxx–xxx 3
Fig. 1. (A) Analysis of A� by ELISA from 7W CHO cells treated with different NF-�B inhibitors. A�1–40 and A�1–42 levels were normalized to values obtainedfrom vehicle-treated cells and expressed as percentage of A�1–40 or A�1–42 values obtained in the control conditions. The bar graph represents the effect of NF-�Binhibitors on A�1–40 and A�1–42 production. ANOVA revealed significant main effects (P < 0.05) for the different treatment conditions as well as significant dosedependent effects of the compounds on A�1–40 and A�1–42 production (P < 0.05). The symbol (*) on the bar graph denotes significant differences (P < 0.05) revealedb ntroll ined fd 0 obta
a178
a179
a180
N181
p182
g 183
NCy post hoc analysis between the value of A�1–40 or A�1–42 obtained in the co
evels in 7W CHO cells. A�1–40 levels were normalized to A�1–40 values obtaifferences (P < 0.05) revealed by post hoc analysis between the value of A�1–4
correlation between the amount of PGE2 inhibition and the
UPlease cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
mount of A� inhibition (data not shown) suggesting that themount of A� inhibition observed is proportional to the extent ofF-kB pathway inhibition. However, some compounds displayreferential inhibitory effects towards A�1–40 or A�1–42 sug-
At
o
and treated conditions. (B) Effect of NF-�B inhibitors on intracellular A�1–40
rom vehicle-treated cells. The symbol (*) on the bar graph denotes significantined in the control and treated conditions.
esting that this selectivity towards the inhibition of a particular
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
� species may rely on additional properties of the molecules 184
ested independently of their effect on NF-�B pathway. 185
In addition, we tested the effect of NF-�B inhibitors and 186
f the IKK-2 inhibitor IV on A�1–40 and A�1–42 secretion 187
ED
F
ARTICLE IN PRESS+ModelNSL 23749 1–6
4 D. Paris et al. / Neuroscience Letters xxx (2006) xxx–xxx
F diffet gnificP
b188
s189
m190
P191
S192
a193
c194
A195
<196
r197
I198
S199
i200
201
m202
u203
c204
o205
w206
l207
R208
i209
o210
i211
o212
C213
o214
a215
I216
c217
w218
u219
t220
W221
u222
M223
s224
s225
G 226
s 227
t 228
X 229
e 230
A 231
p 232
o 233
o 234
stimulate the ratio of sAPP�/(sAPP� + sAPP�) and to decrease 235
the ratio of sAPP�/(sAPP� + sAPP�), as well as APP CTF� 236
production (Fig. 4B) suggesting that they inhibit the �-cleavage 237
of APP. 238
Fig. 3. Analysis of A� by ELISA from human neuroblastoma SH-SY5Y cellstreated with different NF-�B inhibitors for 48 h. A�1–40 and A�1–42 levels werenormalized to values obtained from vehicle-treated cells and expressed as per-centage of A�1–40 or A�1–42 values obtained in the control conditions. The bargraph represents the effect of NF-�B inhibitors on A�1–40 and A�1–42 produc-
NC
OR
RE
CT
ig. 2. Analysis of PGE2 production by ELISA from cultured cells treated withhe different treatment conditions. The symbol (*) on the bar graph denotes siGE2 obtained in the control and treated conditions.
y human neuroblastoma SH-SY5Y cells (ATCC, Manas-as, VA, USA). SH-SY5Y cells were grown in Ham’s F12edium supplemented with 10% fetal bovine serum and 1×enicillin–Streptomycin Fungizone mixture. Confluent SH-Y5Y cells were treated for 48 h with celastrol, CAPE,rtemisinin and the IKK-2 inhibitor IV. Culture media was thenollected and evaluated for A�1–40 as previously described and�1–42 using a high sensitivity A�1–42 ELISA (detection limit1 pg/ml, Invitrogen Biosource Inc., Carlsbad, CA, USA). Dataeveal that CAPE, celastrol, artemisinin and the IKK-2 InhibitorV significantly reduce A�1–40 and A�1–42 production by SH-Y5Y cells confirming that NF-�B also regulates A� production
n neuronal like cells (Fig. 3).We next tested the effect of NF-�B inhibitors on the accu-
ulation of intracellular APP-CTF fragments by western-blot,sing 7W CHO cells, to determine whether NF-�B inhibitorsan affect APP processing. Briefly, 7W CHO cells were platednto 24-well cell culture plates. Confluent 7W CHO cellsere treated for 18 h with different NF-�B inhibitors. Cellu-
ar proteins were extracted with 150 �L of ice-cold M-PER®
eagent (Pierce Biotechnology, Rockford, IL, USA) contain-ng 1 mM phenylmethanesulfonyl fluoride and 1 mM sodiumrthovanadate. Samples were sonicated, denatured by boilingn Laemmli buffer (Bio-Rad, Hercules, CA, USA) and resolvednto 4–20% gradient polyacrylamide gels (Bio-Rad, Hercules,A, USA). After electrotransfering onto polyvinylidene diflu-ride membranes, Western-blots were immunoprobed with annti-APP C-terminal (751–770) antibody (EMD Biosciencesnc., San Diego, CA, USA), with the antibody 22C11 (Chemi-on, Temecula, CA, USA) that recognizes full length APP andith an anti-actin antibody (Chemicon, Temecula, CA, USA)sed as a reference antibody to quantify the amount of pro-eins electrotransferred. Additionally, sAPP� was detected by
estern-blot in the culture medium surrounding 7W CHO cells
UPlease cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
sing the antibody 6E10 (Signet Laboratories Inc., Dedham,A, USA) which recognizes the amino-acids 1–17 of A� and
APP� was detected in the culture medium using an anti-humanAPP� antibody (Immuno-Biological Laboratories Co. Ltd.,
tmodt
PR
OOrent NF-�B inhibitors. ANOVA revealed significant main effects (P < 0.05) for
ant differences (P < 0.05) revealed by post hoc analysis between the value of
unma, Japan). APP CTF/Actin, sAPP�/(sAPP� + sAPP�) andAPP�/(sAPP� + sAPP�) signals intensity ratios were quan-ified by chemoluminescence imaging with the ChemiDocTM
RS (Bio-Rad, Hercules, CA). Our data show that the differ-nt NF-�B inhibitors tested do not induce the accumulation ofPP CTF (Fig. 4A) suggesting that NF-�B inhibitors lower A�roduction without inhibiting �-secretase activity (an inhibitionf �-secretase activity would lead to a significant accumulationf APP CTF� and CTF�). Celastrol, kamebakaurin appear to
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
ion. ANOVA revealed significant main effects (P < 0.05) for the different treat-ent conditions as well as significant dose dependent effects of the compounds
n A�1–40 and A�1–42 production (P < 0.05). The symbol (*) on the bar graphenotes significant differences (P < 0.05) revealed by post hoc analysis betweenhe value of A�1–40 or A�1–42 obtained in the control and treated conditions.

EC
TED
ARTICLE IN+ModelNSL 23749 1–6
D. Paris et al. / Neuroscience L
Fig. 4. Analysis of APP processing after treatment of CHO cells with differentNF-�B inhibitors. (A) Lanes 1: control; 2: parthenolide 1 �M; 3: andro-grapholide 30 �M; 4: CAPE 20 �M; 5: artemisinin 30 �M; 6: celastrol 5 �M; 7:isohelenin 10 �M; 8: kamebakaurin 30 �M. (B) The chemoluminescent signalintensities of the ratios of sAPP�/(sAPP� + sAPP�), sAPP�/(sAPP� + sAPP�),full length APP (FL APP)/actin and APP-CTF� + APP-CTF� (APP CTF)/actinare represented on the histogram. The symbol (*) on the bar graph denotes signif-icant differences (P < 0.05) revealed by post hoc analysis between the value ofthe chemoluminescent signals obtained in the control and treated conditions.DdA
239
(240
s241
N242
a243
o244
w245
s246
r247
c248
N249
A250
c251
o252
t 253
p 254
a 255
a 256
f 257
b 258
t 259
H 260
g 261
d 262
I 263
p 264
w 265
n 266
b 267
A 268
269
e 270
S 271
C 272
a 273
m 274
R 275
276
277
278
279
280
281
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
NC
OR
Rata analysis revealed that CAPE, celastrol and kamebakaurin significantlyecreased sAPP� and APP-CTF but increased sAPP� production. Full lengthPP increment was observed following CAPE and celastrol treatment.
As several other non steroidal anti-inflammatory drugsNSAIDs) can also inhibit NF-�B activation (such as sulindaculphide or R-flurbiprofen), it remains possible that selectiveSAIDs presenting an inhibitory activity towards NF-�B will be
ble to lower A� levels independently of their inhibitory actionn cyclooxygenase activities. However, NSAIDs lower A�1–42ith low efficiency (IC50 generally greater than 50 �M) [22] and
ome cyclooxygenase-2 selective NSAIDs have been shown toaise A�1–42 levels by 200% [12]. Unfortunately also to date,linical trials designed to inhibit cyclooxygenase activity with
UPlease cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
SAIDs have failed to demonstrate efficacy in the treatment ofD patients [9]. Among the NF-�B inhibitors that we tested,
elastrol appears the most potent (more than 90% inhibitionf A�1–40 and A�1–42 with 500 nM in 7W CHO cells). Acute
[
[
PR
OO
F
PRESSetters xxx (2006) xxx–xxx 5
reatment with celastrol in rodent has revealed that this com-ound can decrease brain NF-�B immunostaining, astrogliosisnd neurodegeneration in mouse models of Parkinson’s diseasend Huntington’s disease [5]. Neuroinflammation is a prominenteature of AD pathology and NF-�B blockade is also expected toe beneficial by decreasing astrogliosis and microgliosis in addi-ion to reducing A� accumulation as outlined in this manuscript.owever, NF-�B influences the expression of a complex array ofenes in the nervous system and the consequences of a chronicown regulation of the NF-�B pathway are not well established.t is therefore premature to consider NF-�B as a viable thera-eutic target for AD. Further work will be required to determinehether NF-�B inhibitors can lower A� levels and improve cog-ition in transgenic animal models of AD and whether they cane safely used in humans.
cknowledgments
We thank Dr. Michael Wolfe (Center for Neurologic Dis-ases, Brigham and Women’s Hospital, Harvard Medicalchool, Boston, Massachusetts, USA) for providing the 7WHO cells used in this study. The authors are grateful to Mr.nd Mrs. Roskamp for their generous support which helped toake this work possible.
eferences
[1] E. Aldieri, D. Atragene, L. Bergandi, C. Riganti, C. Costamagna, A. Bosia,D. Ghigo, Artemisinin inhibits inducible nitric oxide synthase and nuclearfactor NF-kB activation, FEBS Lett. 552 (2003) 141–144.
[2] K.R. Bales, Y. Du, R.C. Dodel, G.M. Yan, E. Hamilton-Byrd, S.M. Paul,The NF-kappaB/Rel family of proteins mediates Abeta-induced neurotox-icity and glial activation, Brain Res. Mol. Brain Res. 57 (1998) 63–72.
[3] P.M. Bork, M.L. Schmitz, M. Kuhnt, C. Escher, M. Heinrich, Sesquiterpenelactone containing Mexican Indian medicinal plants and pure sesquiterpenelactones as potent inhibitors of transcription factor NF-kappaB, FEBS Lett.402 (1997) 85–90.
[4] S.H. Choi, R. Langenbach, F. Bosetti, Cyclooxygenase-1 and -2 enzymesdifferentially regulate the brain upstream NF-kappa B pathway and down-stream enzymes involved in prostaglandin biosynthesis, J. Neurochem. 98(2006) 801–811.
[5] C. Cleren, N.Y. Calingasan, J. Chen, M.F. Beal, Celastrol protects againstMPTP and 3-nitropropionic acid-induced neurotoxicity, J. Neurochem. 94(2005) 995–1004.
[6] G. Evin, A. Zhu, R.M. Holsinger, C.L. Masters, Q.X. Li, Proteolytic pro-cessing of the Alzheimer’s disease amyloid precursor protein in brain andplatelets, J. Neurosci. Res. 74 (2003) 386–392.
[7] I. Ferrer, E. Marti, E. Lopez, A. Tortosa, NF-kB immunoreactivity isobserved in association with beta A4 diffuse plaques in patients withAlzheimer’s disease, Neuropathol. Appl. Neurobiol. 24 (1998) 271–277.
[8] M.A. Hidalgo, A. Romero, J. Figueroa, P. Cortes, I.I. Concha, J.L. Hancke,R.A. Burgos, Andrographolide interferes with binding of nuclear factor-kappaB to DNA in HL-60-derived neutrophilic cells, Br. J. Pharmacol. 144(2005) 680–686.
[9] J.J. Hoozemans, M.K. O’Banion, The role of COX-1 and COX-2 inAlzheimer’s disease pathology and the therapeutic potentials of non-steroidal anti-inflammatory drugs, Curr. Drug Targets CNS Neurol. Disord.4 (2005) 307–315.
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
10] H.Z. Jin, B.Y. Hwang, H.S. Kim, J.H. Lee, Y.H. Kim, J.J. Lee, Anti 307
inflammatory constituents of Celastrus orbiculatus inhibit the NF-kappaB 308
activation and NO production, J. Nat. Prod. 65 (2002) 89–91. 309
11] B. Kaltschmidt, M. Uherek, B. Volk, P.A. Baeuerle, C. Kaltschmidt, Tran- 310
scription factor NF-kappaB is activated in primary neurons by amyloid 311

IN+ModelN
6 nce L
312
313
314
[315
316
317
318
319
[320
321
322
323
[324
325
326
327
[328
329
330
331
[332
[ 333
334
[ 335
336
337
338
339
[ 340
341
342
[ 343
344
345
346
[ 347
348
349
ARTICLESL 23749 1–6
D. Paris et al. / Neuroscie
beta peptides and in neurons surrounding early plaques from patientswith Alzheimer disease, Proc. Natl. Acad. Sci. U.S.A. 94 (1997) 2642–2647.
12] T. Kukar, M.P. Murphy, J.L. Eriksen, S.A. Sagi, S. Weggen, T.E. Smith,T. Ladd, M.A. Khan, R. Kache, J. Breard, M. Dodson, S. Merit, V.V.Ozols, P.Z. Anastasiadis, P. Das, A. Fauq, E.H. Koo, T.E. Golde, Diversecompounds mimic Alzheimer disease-causing mutations by augmentingAbeta42 production, Nat. Med. 11 (2005) 545–550.
13] J.H. Lee, T.H. Koo, B.Y. Hwang, J.J. Lee, Kaurane diterpene, kamebakau-rin, inhibits NF-kappa B by directly targeting the DNA-binding activity ofp50 and blocks the expression of antiapoptotic NF-kappa B target genes,J. Biol. Chem. 277 (2002) 18411–18420.
14] Y.Z. Lin, S.Y. Yao, R.A. Veach, T.R. Torgerson, J. Hawiger, Inhibition ofnuclear translocation of transcription factor NF-kappa B by a synthetic pep-tide containing a cell membrane-permeable motif and nuclear localizationsequence, J. Biol. Chem. 270 (1995) 14255–14258.
15] K. Natarajan, S. Singh, T.R. Burke, D. Grunberger, B.B. Aggarwal, Caffeic
UN
CO
RR
EC
TED
Please cite this article in press as: D. Paris et al., Inhibition ofdoi:10.1016/j.neulet.2006.12.029
acid phenethyl ester is a potent and specific inhibitor of activation of nucleartranscription factor NF-kappa B, Proc. Natl. Acad. Sci. U.S.A. 93 (1996)9090–9095.
16] D.J. Selkoe, Amyloid beta-peptide is produced by cultured cells duringnormal metabolism: a reprise, J. Alzheimer Dis. 9 (2006) 163–168.
[
F
PRESSetters xxx (2006) xxx–xxx
17] S. Sinha, I. Lieberburg, Cellular mechanisms of beta-amyloid productionand secretion, Proc. Natl. Acad. Sci. U.S.A. 96 (1999) 11049–11053.
18] S. Sung, H. Yang, K. Uryu, E.B. Lee, L. Zhao, D. Shineman, J.Q. Tro-janowski, V.M. Lee, D. Pratico, Modulation of nuclear factor-kappa Bactivity by indomethacin influences Abeta levels but not Abeta precur-sor protein metabolism in a model of Alzheimer’s disease, Am. J. Pathol.165 (2004) 2197–2206.
19] K. Terai, A. Matsuo, P.L. McGeer, Enhancement of immunoreactivityfor NF-kappa B in the hippocampal formation and cerebral cortex ofAlzheimer’s disease, Brain Res. 735 (1996) 159–168.
20] M. Tobe, Y. Isobe, H. Tomizawa, T. Nagasaki, H. Takahashi, T. Fukazawa,H. Hayashi, Discovery of quinazolines as a novel structural class ofpotent inhibitors of NF-kappa B activation, Bioorg. Med. Chem. 11 (2003)383–391.
21] I.M. Verma, J.K. Stevenson, E.M. Schwarz, D. Van Antwerp, S. Miyamoto,Rel/NF-kappa B/I kappa B family: intimate tales of association and disso-ciation, Genes Dev. 9 (1995) 2723–2735.
PR
OO
A� production by NF-�B inhibitors, Neurosci. Lett. (2006),
22] S. Weggen, J.L. Eriksen, P. Das, S.A. Sagi, R. Wang, C.U. Pietrzik, K.A. 350
Findlay, T.E. Smith, M.P. Murphy, T. Bulter, D.E. Kang, N. Marquez- 351
Sterling, T.E. Golde, E.H. Koo, A subset of NSAIDs lower amyloidogenic 352
Abeta42 independently of cyclooygenase activity, Nature 414 (2001) 353
212–216. 354
Related Documents











![Fingolimod Affects Transcription of Genes Encoding Enzymes ...Aβ [27]. Aβ peptides modulate the enzymes of sphingolipid metabolism and S1P receptors in cellular models; thus, Aβ’s](https://static.cupdf.com/doc/110x72/60c8a68d47f86855c059212d/fingolimod-affects-transcription-of-genes-encoding-enzymes-a-27-a-peptides.jpg)