INFORMAnON TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus. sorne thesis and dissertatiOn copies are in typewriter face. while athers may be trom any type of computer printer. The quallty of thf. reproduction 1. dependent upon the quallty of the capy submltted. Broken or indistind print, colored or poor quality illustrations and photographs. print bleedthrough. substandard margins, and improper alignment can adversely affect reproduction. ln the unlikely event that the author did not send UMI a complete manuscript and there are missing pages. these will be noted. Aise, if unauthorized copyright material had ta be removed. a note will indicate the deletion. Oversize materials (e.g., maps, drawings, chans) are reproduced by sectioning the original, beginning al the upper left-hand corner and continuing from left ta right in equal sections with small overtaps. Photographs induded in tfIe original manuscript have been reproduced xerographically in this capy. Higher quality 6- x g- black and white photographie prints are available for any photographs or illustrations appearing in this copy for an additional charge. Contad UMI directJy to arder. Bell & Howell Information and Leaming 300 North Z8eb Raad. Ann Arbor, MI 48106-1346 USA UMI e 800-521-0600

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

INFORMAnON TO USERS
This manuscript has been reproduced from the microfilm master. UMI films the
text directly from the original or copy submitted. Thus. sorne thesis and
dissertatiOn copies are in typewriter face. while athers may be trom any type of
computer printer.
The quallty of thf. reproduction 1. dependent upon the quallty of the capy
submltted. Broken or indistind print, colored or poor quality illustrations and
photographs. print bleedthrough. substandard margins, and improper alignment
can adversely affect reproduction.
ln the unlikely event that the author did not send UMI a complete manuscript and
there are missing pages. these will be noted. Aise, if unauthorized copyright
material had ta be removed. a note will indicate the deletion.
Oversize materials (e.g., maps, drawings, chans) are reproduced by sectioning
the original, beginning al the upper left-hand corner and continuing from left ta
right in equal sections with small overtaps.
Photographs induded in tfIe original manuscript have been reproduced
xerographically in this capy. Higher quality 6- x g- black and white photographie
prints are available for any photographs or illustrations appearing in this copy for
an additional charge. Contad UMI directJy to arder.
Bell & Howell Information and Leaming300 North Z8eb Raad. Ann Arbor, MI 48106-1346 USA
UMIe
800-521-0600


".
•
.'
Postnatal Developmental LocalizatioD or LRP-2in the Efferent Oueu and Epididymis of the Male Rat
and the Oviduct and Uterus of the Female Rat
Marc LustigDepartment ofAnatomy and CclI Biology
McGill University~ Montreal1997
A thesis submitted to the Fac:ulty of Graduate Studies andResearch in partial fuJfillment of the requirements
furtheMwn~ofSàenœ
@ Marc Lustig 1997

1+1 National Ubraryof Canada
Acquisitions andBibliographie services
395 Wellington StreetOUawaON K1A0N4Canada
Bibliothèque nationaledu Canada
Acquisitions etservices bibliographiques
395. rue WellingtonOttawa ON K1A ON4Canada
The author bas granted a oonexclusive licence allowing theNational Library of Canada toreproduce, loan, distnbute or sellcopies of this thesis in microform,paper or electronic formats.
The author retains ownership of thecopyright in this thesis. Neither thethesis nor substantial extracts from itmay be printed or otherwisereproduced without the author' spemusslon.
L'auteur a accordé une licence nonexclusive permettant à laBibliothèque nationale du Canada dereproduire, prêter, distribuer ouvendre des copies de cette thèse sousla fonne de microfiche/film, dereproduction sur papier ou sur fonnatélectronique.
L'auteur conserve la propriété dudroit d'auteur qui protège cette thèse.Ni la thèse ni des extraits substantielsde ceDe-ci ne doivent être imprimésou autrement reproduits sans sonautorisation.
0-612-44213-6
Canad!t

•
•
•
Shol1 dlle: Developmelllailocali:atioll ofLRP-2 in the rat reprodl4clive system
Il

•
•
•
ABSlRACT
Law density lipoprotein receptor-related protein-2, LRP-2 (gp330/megalin) is a
eell surface receptor involved in the intemalization of apolipoprotein J (apoJ/clusterin).
Apo J is found in the luminal fluids of the testis and epididymis, as weil as on the plasma
m~:"llbrane of the sperm. It has already been suggested that an apo J/LRP- . interaction
may be involved in sperm maturation whereby apo J redistributes cholesterol on the
sperm membrane which is necessary tor the capacitation and acrosomal reaction in the
oviduct. This research examined the postnatal developmentallocalization ofLRP-2 in the
efferent duets and in the epididymis as well as the developmentallocalization of LRP-2 in
the oviduct, and the uterus. The pattern of immunolocalization was then analyzed in
terrns age dependent tàctors. Immunocytochemistry was performed using polyclonal
antibody to LRP-2 on the efferent ducts and epididymides of male rats (aged 7-90 days)
and the oviduct and uterus of female rats (aged 7-56 days). Localization of LRP-2 in the
efferent duets and epididymis revealed a regionally specifie expression from day 28 to the
nl. Jre 90 day old. There was a complete absence of LRP..2 in the re:..: l,ms of the
proximal initial segment and distal caput which resembled the lack of immunoreactivity
of apo 1 in the same regions. The greatest intensity of immunoperoxidase reaction
oceurred in the distal initial segment, intermediate zone and proximal caput. where it is
known chat apo J is most abundantly seereted. LRP-2 localization was tirst c1early
evident in the female on day 29. Furthermore, after the initiation of the estrus cycle,
marked by the first ovulation, usually between 36·39 days, significant differences in
loealization of LRP·2. in different regions at different stages of the cycle were
demonstrated. These results provide new information with respect to thp postnatal
development localization of LRP·2 in the male and female reproductive tract~ and support
tH. concept that LRP-2 mediated endocytosis of apo J may play a rCPl': in sperm
maturation.
UI

•
•
•
RÉsUMÉ
Le récepteur des lipoprotéines de tàit le densité. le LRP-2 (gp330 ou mégaline) est
localisé à la surface de la cellule et contribue à interioriser l'apolipoprotéine l (apo J ou
dusterin). L'apo J est présente dans les liquides intratubulaires du testicule et de
l-épididyme ainsi que dans la membrane cytoplasmique des spermatozoïdes. li a déjà été
suggéré que l'interaction de l'apo J et du LRP-2 doit jouer Wl rôle dans la maturation des
spermatozoïdes en favorisant la redistribution du cholestérol de la membrane
cytoplasmique. réarrangement des molécules qui serait nécessaire à la capacitation et à la
réaction de l'acrosome des spermatozoïdes dans l'oviducte. Au cours de la présente
recherche nous avons examiné la distribution du LRP-2 pas immunocytochimie dans les
canaux efférents et répididyme de rats mâles en croissance (i.e. de 7 à 90 jours) ainsi que
dans l'oviducte et 1· utérus de rates (de 7 à 56 jours) au cours du cycle ovarien. Chez le
mâle de 18 à 90 jours on note l'absence de LRP-2 dans les régions épididymaires
suivantes. le segment initial proximal et le segment caput distal ce qui correspond à
l'absence dïmmWloréactivité de l'apo J dans ces memes régions. Pas ailleurs
1ïmmunoréactivitê pour le LRP-2 est intense dans le segment initial distal. dans la zone
intermédiaire et le segment caput proximal où l'apo J est secréteé abondamment. Chez la
femelle la présence du LRP-~ apparaît à 29 jours dans roviducte et l'utérus. Par la suite
aprés la premiére ovulation, vers 36-39 jours, on remarque une variation marquée de
1ïmmunoréactivité du LRP-1 dans les diverses régions du tractus génital au cours du
cycle ovarien. Ces observations supportent bien l'hypothèse que l'endocytose de l'apo J
tàcilitée par le LRP-1 peut jouer un rôle dans ta maturation des spermatozoïdes lors de
leur transit dans les systèmes reproducteurs mâles et femelles .
iv

•
•
•
10 myfami{v and Gillian
v

•
•
•
;\C~~OWLEQG[M[NTS
There are several people who [ would like to thank.. whose help was indispensable
during the course of this Master's Thesis.
Firstly. [ would like ta thank Dr. Clermont for his suggestions, his teachings and
mast importantly his advice.. '''to always be critical of your work.·"
The assistance of my fellow graduate students.. Xing Quaa. Jana Fuska.. Chi Chi
Wosu.. Jason Boman, Andrea Mueller is greatly appreciated.
[ would like ta thank Dr. Hermo for his suggestions and the time he took to lend
his knowledge of reproductive biology.
Grateful acknowledgement is given to Dr. El AIfy for ail of his help during
dissection and for his demonstrations ofmale and female reproductive tract histology.
The clerical assistance and desktop support of Gillian Douglas is greatly
appreciated.
Finally, [ would like to give my sincere thanks ta my supervisor and fiiend Dr.
Carlos MoraJes. who at any time.. wouId give me his undivided attention.. ta answer aJl of
my questions and ta share his vast insight.
VI

• T;\BL[ OF CONTENTS
Page
Short Titl~ .. ... .. .. li
..\bstract .... .. .. . .. .. ... . . .. .. .. III
Resume: vDedic3tion _ __ , vi.~ckno\\·ledgements viiTable of Contents .. viiiList of Abbreviations '" ,. ... x
•
•
INTRODUCTION .REVIEW OF LITER.:\TURE .
LRP-2 .As A.\tlember afthe LDL SlIperjàmi(v ..Stnlcrure .. .
Figure 1 , - .LIgands and Binding Acnvtty ..
Figure 2 '" .Possible Function .
Apolipoprotein JEndoc..vrosls b..vLRP-2 .lnteracnon afLRP-2 wlth Apo./ ln Reproductive Eplthelia
Objectives .
~(ale Rat Reproductive Tract. .Figure 3 ..
Efferent Duets ..lVon clliated ,,'ells .Cilialed ceils .
Epididymis .Structure ..Pnnclpai ce//s ..~Varrow ce/ls ..Apical ceils .Clear cells .Halo cells ..Basal cel/s .Epididymai Functions ..Epididymal Regulation androgens ..Deve/opmenta/ Aspects oj.'4ale Rep,oducnve Tract .
Female Rat Reproductive Tract .Figure.J .
Oviducts .Stnlcture ..
Uterus .Strocrure .Developmento/ Aspects ofthe Female Reproductive Tract.
Estrus Cycle .
vu
133344567
889
Il11121212131313141415l5l5151611
1919202020202122

DISCUSSION .. 35Posma(a/ Deve/opmenta/ Loca/izanon olLRP-2 ln the E.lJerent Ductsand Epuiidymls ofthe l\lla/e Rat 35Posmatal Deve/opmenra/ Loca/ization ofLRP-2 zn the Ovrductsand UtenlS ofthe Female Rat 38
•
•
D,estnls .Proestnls .. , '" '" '" '" ,Estnls .~\tletf!."J·tnlS .
Figure 5 '" .
~l-\TERIALS :\~D ~IETHODS ...4nlma/s .vagInal Smear _ .Pt!!':tùslOn and DIssection ofAllale and Female Rat .
L~l lmmunoc)tochemistry " ..Deh.vdration ..Embeddlng .Immunocytochemlstry .
RESULTS .Posmatal Developmental Localizanon ofLRP-2 ln the Efferent Duetsand Ep,didymis ofthe ~'dale Rat .
Table 1 .Figure 6 , , , .
Posmatal Developmenta/ Localizarion olLRP-2 in the Ovidllctsand Utenls afthe Femafe RaI .
Table 2 '" ., .
222221,~_J
23
27
213031
3234
•
CONCLUSION ...1
PLATES AND LEGENDS ~3
ORIGINAL CONTRIBUTIONS _ 64
IliFERENCES 65
VIII

•
•
•
1.(51 OF A88REYL\TION5
a.1~IRAP- alpha 2-~[acroglabulin Receptor Associated Pratein
a2MR- alpha 2-~facraglabulin Receptor
Apa E· Apolipoprotein E
Apo J- Apolipoprotein J
DAB· Diaminobenzidine
EGF- Epithelial Growth Factor
fSH· Follicle Stimulating Hormone
Gp 330- Glycoprotein 330
LDL- Low Density Lipoprotein
LDlR- Law Density Lipoprotein Receptor
LH- Leutinizing Hormone
LRP-I- Low Density Lipoprotein Receptor Related Protein-l
LRP-2- Law Density Lipoprotein Rec.eptor Related Protein-2
PAI-I- Plasminogen Aetivator in complex with Type 1 Inhibitor
RAP· Receptor Associated Protein
SGP·1- Sulfated Glycoprotein-l
SGP-2- Sulfated Glycoprotein-2
TBS- Tris-base Saline
VLDL- Very Law Density Lipopratein
VLDLR· Very Low Density Lipaprotein Receptor
IX

•
•
•
INTRODUCTION
The location of LRP-2 has been studied most extensively in rats by
immunocyrochemistry using monoclonal and polyclonal anti-LRP-2 antibodies. LRP-2 bas been
localized on a restricted group of epithelia incJuding renal proximal tubules, type rI
pneumonocytes, yolk sac, ciliary epithelium of the eye, endometrial lining ceUs, efferent ducts
and epididymis, and endometriallining ceUs (Zheng et aL, 1994, Morales et aL, 1991). LRP-2 is
coocentrated 00 the apical portions of tbese epithelia often in the region of the coaœd pits, and
less prominently in subapical vesicles (Zheng et al 1994).
Kounnas et al. (1994) demonstrated that LRP-2 is a large 600 kDa celI surface protein,
strueturally homologous to homologous ta a.2MRILRP-l belonging to the low density lipoprotein
receptorfamily. LRP-2 and alMRILRP-l bath bind and internalize an overlapping set ofproteins
including the 40 kDa receptor-associated protein, lasminogen activator, lactoferrin and
apolipoprotein E by receptor-mediated endocytosis (Christensen et al. 1992, Kounnas et al.
1992).
Kounnas et al. (1995) identified another ligand, apolipoprotein 1, (also known as
Clusterin, and SGP-2) to be endocytosed by the LRP-2 receptor. Apo 1 is a 10kOa heterodimeric
protein synthesized and secreted by Sertoli celIs (Sylvester et al. 1984, Griswold et al. 1988,
Sylvester et al. 1991).
It is believed that apo l is involved in lipid transport which plays a mie in sperm
maturation. (de Silva et al. 1990, Fritz and Murphy 1993, Henno et al. 1991, Henno et al. 1994,
Kounnas 1995, and Sylvester et al. (991). The physiological mie of apo J in the reproductive
system remaios to be elucidated. However a possible funetion of apo 1 is to change the content of
cholesterol in the male and female reproductive tract prior to fertilizatioD. In~ spermalozoa
require a pcriod of residence in the epididymis and in the female reproductive tract to mature and
acquire the ability ta recognize and fertilize the egg.
1

• Using light microscope immunocytochemistry it bas been demonstrated in adult rats that
expression of apo 1 varied between regions of the epididymis. This pattern of apo J expression
suggested the presence of age~ependent luminal factors (Le. spennatozo~ various epididymal
proteins, and androgens) as cause for the differential expression (Henno et al. 1994). Sïnce LRP-2
bas been shown to endocytose apo J it is valuable to study the localization of LRP-2 both
regionallyand developmentally.
This research e:<amines the postnatal developmentallocalization of LRP-2 in the efferent
ducts and in the epididymis in order ta evaluate the effect of androgen levels and epididymal
factors, namely apo 1, which May regulate it. The same objective is used to investigate the female
postnatal developmentallocaIization of LRP-2 in the proximal 3Ild distal ovidu~ uterus, and the
uterine glands. The pattern of immunoloca1ization was then analyzed in terms ofthe ditferent ages
of the rats and the differeot stages of the female esttus cycle. Immunocytochemistty was
perfonned using a polyclonal antibody ta LRP-2 00 the efferent ducts and epididymides of male
• rats (aged 7-90 days) and the oviduct and uterus of female rats (aged 7-56 days) to determine the
pattern of localization of LRP-2 and ta demonstrate the changes in staining of LRP-2 with respect
to regional and developmental variations.
As the focus of titis thesis, a review of LRP-2 in tenns of its homology to other LDL-like
receptors, structure, ligands and binding aetivity, function and ifs interaction with apo J are given.
Subsequently, the review of the literature will describe the anatomy, histology and physiological
processes of the efferent ducts and epididymis of the male rat reproductive~ as weU as the
oviduets. uterus and hormonal cycle of the female rat reproductive tract as these are the areas in
which localization of LRP-2 will he studied. Following the review of the literature, sections on
the materials and methods employed, results, discussion and summary of this thesis are included.
• 2

•
•
•
REVIEW OF LITERATURE
Low Density Receptor-Related Protein-2/Mecalin/Gp 330
LRP·2: A member ofIlle LDL ,eceptorfamily
The superfamily of Low Density Lipoprotein endocytic receptors is composed of severa!
known members (including LDL~ a2MRILRP-I,VLDL~ vitellogenin receptor, a
Caenorhabditis e/egans protein and LRP-2) the structures ofwhich have been deduced by cDNA
cloning and sequencing. In 1988 Herz et al. c10ned a large protein structure closely related ta
LDtlt and cal1ed it LDLR-related protein (aIso known as a2-Macroglobulin Receptor).
Evidence demonstrating that LRP cao function as a receptor for chylomicron remnantsIB
migrating very low density lipoprotein, (B-VLDL) rich in apolipoprotein E (Apo-E) was provided
by Kowal et al 1989. LRP-2 (Op 330~ megalin) was subsequently discovered whena2MRJLRP
was affinity purified and revealed the presence of a 40-kDa protein designatech2MRAP (Jensen
et al. 1989). cOua cloning of this protein revealed that this was in fact a sequence of rat epithelial
glycoprotein 330 (Gp330), which bad previously been shown ta be the autoantigen in an induced
buman glomeruJar nephritis, and in the rat model Heymann nephritis (Pietromonaco 1990). This
finding verified an earlier report (Kerjaschki and Farquahar 1982) that immunoglobulin, extracted
from glomeruli of Heymann Nephritis rats, recognized a high molecular mass proximal tubule
glycoprotein with an estimated sUe of 330 kDa, hence the original name Gp 330. The estimates
of330 kDa were confirmed by eleettophoretic mobility studies to be much larger, al 550-600 kDa
(Orlando et al., 1992). Raychowdbury et al. (1989) panially cloned the rat gp330 cDNA. They
produced tryptic digests of immuno-affinity-purified rat gp330 and designed synthetic
oligonucleotides for screening a cDNA library. Sequencing of severa! positive clones revealed
two non-overlapping 1.4 and 2.9 kb segments with strang bomology ta LDLR and alMR (Herz et
al., 1988, Strickland et al., (991). However~ the encoding amino 3Cid sequence was completely
diffèrent from that reported by Pietromonaco et al. (1990) wbo at the same tinte cloned theic own
ratgp330 cDNA sequence isolated from an expression cDNA library using an anti-LRP-2 [gO. It
3

• was later demonstrated chat the sequence reported by Raychowdhury (1989) \Vas homologous ta
human RAP and thus not a part ofgp330. Christensen et ai. (1992) then demonstrated in the rena!
tubules that binding~ endocytosis and lysosomal degnadation of RAP was mediated by LRP-2.
LRP-2: Strllcture
Although the precise biological functions of LRP-2 are still unclear. the complete cloning
and sequencing of this receptor (Saito et al., 1994) has confinned the structural homology to
a2~IRILRP .. l (Figure 1).
•< a~ ~"',.1 :&oq.no~s
~K.atCl' 109~na
•
FigUre 2: Simplified scitem:ltic dr:1\\1ng of the pOSSIble proc::sstng :ma subsequent FJtcof R.-\P md other ccmmon lipnds in the ~ndcc:ytic companmentS of tRP·: md3.!~IRJlRP.l. This model suggests dtlt R.-\P binds ta the receptor LRP·2 :md/or :1Z.~IRJLRP.l JI1d mav ~SlSt the tolding prccess. subsc:qucndy dissoc:UUing mà bcingdegr:uic:d. AIso R.\P rmLY inhlDlt the binciing of other proœins during reeeplorproccssang. whereby the rcc:c:ptcrs \,jould ref=se R.-\P for dcgr:ld:aàon. thcr=anc:rreeyc:ling mc:msc[vcs. me! xhleving Jffinity for other ligands. This figure W3S _teciftom ~loc:sttUp ~t JI.• (994).
The deduced 4660-aa sequence. consists ofa probable N-terminal signal peptide sequence
(25 aa)r an extracelluJar region {4400 33):0 a single transmembr:me dommn (22 aa)f and a C
tenninal ~1Oplasmic tail (213 aa). The extracelluJar region contains three types of cysteine-rich
repeats char:u:teristic of the low density lipoprotein receptor (LDLR) gene family: 36 LDLR
lig3l1d-binding repeats fonning fourclusters ofputative lig6U1d-binding domains f 16 growth factor
4

•
•
•
repeats separated by 8 YWTD spacer regions, and L C-tenninal epidennal growth factor repeat.
The c~1oplasmic tail contains two copies of the (FX) NPXY motif, which represents a signal for
coated pit-mediated intemalization and an 3dditional simHar motif. The overall structure of LRP
2 is similar ta that of the LDLR-related protein (LRP)/a2-macroglobulin receptor and shows even
greater similarity to the Caenorhabditis elegans protein, reported as a homologue of LRP-l
(Figure 1) (Saïte et al. 1994). However, LRP-2 differs from these proteins in (i) the cysteine-rich
repeat arrangements found in the extreme extracelluJar N- and C- tenninal regions, (H) the
distribution pattern of cysteine residues in the YWTD spacer regioDS, (iii) the location of the
RX(KIR)R consensus recognition sequence of furin, a precursor processing endoprotease, and (iv)
the length and structure of the cytoplasmic tail (Saito et al. 1994). These researchers who
uncovered the complete cloning and sequencing suggested gp330 be called megalin (from the
Greek mega), since it was the largest plasma membrane protein identified 50 fur in vertebrates.
LRP-2: Ligands and Binding Aetivity
In light of the structural homology it was hypothesized that LRP-2 binds severa! of the
ligands which interact with other LDL fumily receptors, namely LRP- L. This was quickly
verifie~ and the overlapping speetrum of ligands include RAP (Christensen et al. 1992, Kounnas
et al 1992, Willnow et al. 1992, Orlando et al. 1992, Morales et al. 1996), plasminogen aetivator
in complex with type-l inhibitor (PAI-l) (Willnow et al. 1992) lipoprotein lipase (Beiseigel et al.
1989), laetoferrin (Willnow et al. 1992) and apolipoprotein E-enriched lipoprotein particles
(Beiseigei et al. 1989). These very different types of ligands ail expose the regions of LRP-l and
LRP-2, wlùch contain positively charged residues. Receptor associated protein (RAP) inhibits the
binding of the ligands to LRP-l and LRP-2 (Moestrup et al. 1991., Willnow et al. 1992., Moestrup
et al 1993, Morales et al. 1996). It is this quality which has made RAP a very useful tooi for
studying ligand binding ta LRP-l and/or LRP-2.
The localization of LRP-2 in coated pits, the structural homology and the overlapping
ligand specificity with the recycling endoc~1osis-mediating receptors LRP-l and LDL~ strongly
suggest similar mechanisms of endocytosis and funetion. After studying the analogous binding
s

•
•
and ~ndoC)1osis of multiple proteins (RAPt uPA:PAI-l. and aprotinïn) by both a2MRlLRP-L
and LRP-2. [n 1994 f ~(oestnlp et al. hypothesized that RAP may have a priming roie for
functional ~ndoc~1osis. It was suggested by this group that RAP. found in the E~ binds to newly
synthesized LRP-2 and cx2...~lRILRP-l and May assist the folding process~ subsequenrly
dissociating as a cbaperone Molecule. The function of RAP May also be to protect the LDLR
fumily receptors against the binding of other proteins during receptor processing, whereby the
receptors wouJd telease Rap for degradation~ and after recycJing themselves achieve affinity for
other ligands (~(oestrup et aL, 1994). (Figure 2)
cr aCIl'4~&nd&n' ~•• fd.IU A cnoaOa ~~~ (dAU Sot ~all1 ~wtft.i-=r :ewIC !CûU 5.: iZlocul
- ~ splCltt rtp'" C-c:'!1'u.)
• 00WèS"I"~"1 nNmKlDnl\e~
c: .:'Y'QD•..MIIC lII1LeNPn.oNPX't-uu.
, l·;(·j(/R.i(~
~• Sè' 53"
~
•
Figure 1: Schematic represencoon of protein structures of the major kno,\n membersor the LDLR gene fumily Amino acid identity percenages are shown bet\\'ecn thesuuctUt:llly conserved domains of l.1t LRP-2 and the C.,ûegans protein (row 1).bct\\"een C.~!egans protein and human LRP-l (row 1)~ between LRP-L and LRP-2 (row3). 3S weU 3S bet\Veen human LDLR md VLDtR (row ~). Four purative ligandbinding domains of LRP...2 are indieated ([-CV). EGF. epidermal groVith filctor. (Thisfigure ",-as Jdapted tram Saito ~ aL~ L994).
6

•
•
•
LRP-2: Possible Functions
Despite the overlap in ligand binding, differences in the expression pattern of LRP-l and LRP-2
suggest important functional differences in vivo. LRP-l is expressed to a variable extent in
various tissues including fibroblasts and macrophages, but is most abundant in bepatocytes and
neurons. In contrast, LRP-2 is primarily found on the apical surfaces of epithelia such as the
glomerulus and proximal tubule in the kidney, the type II pneumonocytes, Clara cells in the lung,
ependymal region in the brain, yolk sac, and specifie epitbelia witbin the maie and female
reproductive tracts (Zheng et al.~ 1994, Morales et aL, 1997). Altbough the biologica1 functions of
these related receptors is still unclear~ it seems that part of the function of LRP-2 May be related ta
ifs location on apical surfaces ofepithelia which are exposed to fluid-filled spaces.
To explore possible LRP-2-specific physiological processes, LRP-2 knockout micc were
used by Willnow et al. (1996). The LRP-2 knockouts developed abnonnalities in epithelial
tissues including lung and kidney that normally express the protein and they die peri-natally ftam
respiratory insufficiency. In the b~ impaired proliferation of neuroepithelium resulted in a
disorder, characterized by lack ofolfàctory bulbs, brain fusion, and a common ventricular system.
Similar syndromes in humans are caused by insufficient supply of cholesterol during
development. The findings of Willnow et al. (1996) suggest that LRP-2, which is ideally situated
to Mediate the uptake of cholesterol-rich lipoproteins, is required for normal cholesterol transport
into the developing embryo.
The finding by Stefansson et aI.( 1995)t that mainly LRP-2 (no other members of the
LDlR family) mediates the endocytosis of low density lipoproteins via the interaction with
apolipoprotein B-IOO t also supports an obvious function of LRP-2 in the metabolism of
lipoproteins.
A possible unique funetio~ different than the other members of the LDLR family is also
supported by the identification of LRP-2 as an endocytic receptor for the intemalization and
degradation of apolipoprotein I/clusterin (Kounnas et al. 1995t Morales et al.t (996).
Apolipoprotein 1 can bind several proteins, including a subclass of high density lipoproteins (de
7

•
•
•
Silva et al.~ 1990, James et al.~ 1991; Stuart et aL~1992). One hypothesis from this report
conceives that LRP-2 Mediates clearance of apo J in complex with other Molecules, likely
lipoproteins and cholesterol. The ability to endocytose apo J introduces other very plausible
funetions of LRP-2~ which may expIain the colocalization of LRP-2 and apo l in tissues of the
male and femaJe reproductive systems, which is the focus ofthis thesis.
Apolipoprotein J/ClusterinlSGP-2
ApoJ: Endocytosis by LRP-2
As noted, apoüpoprotein J (cluste~ SGP-2) is a heterodimeric 70 kDa protein which bas
been shown to be specifically endocytosed by LRP-2~ not LRP or any other LDL receptor
superfamily members (Kounnas 1995). Solid phase binding assays and LRP-2 bound to ApoJ
transferred to nitrocellulose after SDS-polyacrylamide gel electrophoresis confinned that LRP-2
bound to Apo J with high affinity (Kd=14.2) (KoUDDas et al. 1995). It was revealed in the same
report that LRP showed no binding in either type of assay. Funhennore, Kounnas et al. (1995)
demonstrated that ooly cultured cell lines that express LRP-2 endocytosed and degraded
radiolabelled 125I-Apo J. Also, when F9 cells were trealed with retinoic 3Cid and dibutyryl cyclic
AMP, to inCtease expression levels of LRP-2, the receptor displayed an increased ability to
intemalize and degrade Apo l, RAP and LRP-2 antibodies. The precise funetion of Apo 1 is
currently unknown, however severa! hypotheses bave arisen. Icone and Tschopp (1992) suggest
that expression of Apo J is up-regulated at sites undergoing tissue remodeling occurring in
conjunction with apoptosis or folloWÎDg injury. French et al. (1992) proposed that Apo J may he
involved in the removal ofdebris resulting from apoptosis. 80th ofthese theories suggest a raie of
Apo J in complex with complement componcnts or lipid debrislapoptotic bodies~ to be
endocytosed by LRP-2 receptor.
LRP-2 Interaction with ApolipoproteinJ in Urogmital Epithelia: A possible role in spermmaturation
The colocalization of apo J and LRP-2 in specialized epithelia of the male and female
reproductive tracts (Zheng et al. 1994, Kounnas et al. 1994) bas a1so evoked speculation tbat the
8

•
•
•
apo IILRP-2 binding has an important role in the reproductive process, namely, the maturation of
sperm. Apolipoprotein I is synthesized and secreted by Sertoli cells (Sylvester et al. 1984,
Griswold et al. 1988, Sylvester et al. 1991). However, SOS PAGE analysis of proteins in the
epidiymal cells by Sylvester et al. (1984) revealed that a lower moiecular weight from of apo I,
differing in glycosylation, was present in the epididymis (Sylvester et al. 1984). The same authors
later reported the presence ofapo l mRNA in epididymal cells by in situ hybridization.
It is believed tbat both testicuJar and epididymal forms of apo J, are involved in lipid
transport which in some way plays an important role in sperm maturation. (de Silva et al. 1990,
Fritz and Murphy 1993, Hermo et al. 1991, Henno et al. 1994, Kounnas 1995, and Sylvester et al.
1991) Although uncertain, this mie May be essential for proteeting the spermatozoa, improving
and providing mobility, which are acquired by spennatozoa during its transit from the testis to the
epididymis. Furthennore, apo l May function ta redistribute spenn membrane cholesterol and
phospholipids during its residence in the uterus and oviduct prior to fertilization.
From research conducted in our laboratory it is has been suggested that apo I secreted by
Senoli cells binds to the plasma membrane of spermatozoa. Upon reaching the efferent duets,
apo I dissociates from spenn altering the lipid content of its plasma membrane. A less
glycosylated fonn of apo I is then secreted by the distal initial segment, intennediate zone and
capot epididymis which is again bound by spenn. LRP-2 is expressed in different regions of the
epidiymis where, once again, it endocytoses apo J. A simHar series of events may accur in the
female reproductive tract. Apo 1, secreœd in the glands of the uterus, may altemate binding sites
from the SPermatozoa membrane, to LRP-2 receptors in the uterus~ distal and proximal oviduct in
order to transport spenn plasma membrane lipids and proteins, possibly giving the spenn the
ability ta recognize and fertilize the egg or activating acrosome reaction.
Objectives
The objective of this research is to examine the postnatal developmental loc:alization of
LRP-2 in the efferent duets and in the epididymis in order to evaluate the effeet of age (possibly
related ta androgen levels) and epididymal factors, namely apo l, which May regulate it.
9

•
•
•
Concomitandy, another objective is ta investigate the femaIe postnatal developmentallocalization
of LRP-2 in the proximal and distal oviduct, uterusy and the uterine glands. The pattern of
immunolocalization was then analyzed in terms of the different ages of the rats and the different
stages of the female estrus cycle. Immunocytochemistly was perfonned using polyclonal antibody
to LRP-2 on the etÏerent ducts and epididymides of male rats (aged 7-90 days) and the oviduet
and uterus offemale rats (aged 7-56 days) to determine the pattern oflocalization of LRP-2 and
ta demonstrate the changes in immunoreactivity of LRP-2 with respect ta rcgional and
developmental variations.
10

• ~laJe Reproductive System or the Rat
In order to compare the localization of LRP-2 in the etTerent duets and epididymis of rats from 7
90 days (pre-postpubertal) to 3I1drogen levels and epididymal factors an inspection of the male
reproductive system follows.
Testls
Efferent Quers
Couda
Corpus
Initiai SeQment"proximal
distal areaarea 1
Intermediote "Zone
{
prOximal.Caput
distal
•
Figure J: SchentUic representatioQ of the testis7 efferent duets7 and epidiymisof the adult rat. The epididymis is divided into various regiODS7 which are theinitial segment. intermediaœ zone7 caput. corpus 3I1d cauda into a proximal (p>and distal (d) 31'e3.
• Il

• Efferent Duets
The etTerent ducts conneeting the rete testis ta the epididymis are composed of 4 ta 20
tubules (Hemeida et al., 1978; Ilio and Hess, 1994; StotTel and Friess, 1994). There is a sharp
demareation between the rete testis and the efferent ducts. The former is lined by low cuboidal
epithelium. which abruptly changes to tall columnar non-ciliated and ciliated cells of the etTerent
ducts.
Nondliated cells
The efferent duct is lined predominandy by nonciliated cells. These cells possess a loose
network of rough endoplasmic reticulum cistemae as weil as a large Golgi apparatus with a weil
developed trans-Golgi network (Rambourg et al., (987). Furtbennore, these cells contain a highly
endocytic apparatus including tubular coated pits, numerous apical tubules, endosomes,
multivesicular bodies and secondary lysosomes (Henno and Morales, 1984; Hermo et al., 1988a).
• Thus. nonciliated cells are actively involved in the endocytosis of proteins from the tubuJar fluid
such as SGP-l (lgdoura et al., 1993), SGP..2 (Hermo et al., 1991) and androgen binding protein
(Pelliniemi et al., 1981). However, no evidenee of transcytosis has been observed in these cells
(Henno and Morales, 1984; Henno et aI.~ 1988a).
Ci/iated cells
The ciliated celis of the efferent duct, like the nonciliated cells, are tall eolumnar bowever,
fewer in number. They are easily reeognizable by their dark staining appearancc, apical
micmvilli~ abundant cHia interspersed with long mierovilli (Robaire and Henno, 1988). Although
their endocytic apparatus is less elaborate than that of the nonciliated cells, ciliated eelIs are aIso
involved in tluid-pbase and absorptive endocytosis (Henno et al.~ 1985). Therefore, in addition to
their mie in moving luminal fluid and spennatozoa through the duet via their ciliary actio~ these
cells cao modify the composition ofthe luminal tluid due ta endocyWsis.
•12

•
•
Epididymis
Structure
The epididymis is a highly coiled du~ which provides a healthy environment for
spennatozoa maturation and storage. It was shown, in the mid and lale 1960's, that the acquisition
of spermatozoa motility 3I1d ability to fertilize eggs was not simply due to passage of time but
ratber exposure to the luminal contents of the epididymis (Bedfo~ 1967; Orgebin-Crist, 1967;
Orgebin-Cri~ 1969). Since then, many histological, biochemical and endocrinological studies
have been done on mammalian and non-mammalian species. The rat has been most frequently
used since its differences with other mammals appear to be only quantitative in nature (Robaire
and Hermo, 1988).
The epididymis is divided ioto five major regions including the initial segment,
intennediate zone. cap~ corpus and cauda (Figure 3). The epithelium is composed of severa!
different types of ceUs that vary in morphological appearance. relative disaibution and functions.
The cell types identified under light microscopy (Reid and Cleland, 1957) are principal, narrow,
apical.. clear, halo and basal cells. A brief description of the major distribution. histology and
function is presented below.
Principal cells
Principal cells as indicated by the name makc up the majority of the epithelial cells in the
epididymis. Depending on the region, principal cells show distinctive morphologica1 and
functional features (Hermo et al., 1994). In the initial segmen~ tbese cells account for 80% of the
total composition ofcelIs, yel, this number declines to 69% in the corpus and cauda (Robaire and
Henno. 1988).
Principal celIs of the initial segment are taU columnar witb a few microvilli fanning a
brush border, in addition the lumen is small. Their nuclei appear at the basal region of the cell
(Reid and Cleland, 1957; Hamilto~ 1975; Sun and Flickinger, 1980). Furthermore, the nucleus
is round and pale-stained with a pronounced nucleolus. Apical junetional complexes between
principal cells form the blood-epididYmal barrier (HotTer and Hinton, 1984; Cyr et al., 1995;
Pelletier, 1995). The function of the principal ceUs is secretion and endocytosis as will he
• discussed below (Robaire and Hermo, (988). In the initial segment, these eeUs are characterized
13

•
•
•
by an extensive secretory apparatus and a less distinguished endocytic system which is composed
ofcoated pits and vesicles~ endosomes~ multivesicular bodies and lysosomes (Hoffer et al.~ 1973;
Moore and Bedford~ 1979; Hermo et al., 1994).
In the intennediate zone~ principal cells show a striking difference from those in adjacent
regions. In addition te the structural difference ofthe Golgi apparatus and endoplasmic reticulum,
the presence of large endosomes (2mm and greater in diameter) in the apical region of the cell is
typical ofthe intennediate zone (Hermo~ 1995).
In the caput corpus~ cauda, the principal cells have a moderately-stained and irreguiar
shaped nucleus and contain numerous large dense supranuclear granules, identified as lysosomes
(Frien~ 1969; Henno et al., 1994). The height of the principal cells gradually decreases and thus
these celis become cuboidal in the cauda region. Funbennore, the lumen inereases in size from
the initial segment to the cauda regioD.
It has been shown that principal cells differ strueturally along the length of the epididymis
and display region-specifie expression of different lysosomal enzymes within their lysosomes and
these differ not ooly from each other but from other eell types (Hermo et al., 1992c: Igdoura et aI.~
1995; Henno and Adamali, 1997).
Nan'owcells
Narrow celis are found exclusively in the initial segment and the intermediate zone and
constitute about 3% ofthe total epithelial population in the proximal initial segment and 6% in the
intennediate zone. They are identified by their slender narrow appearance~ deep-staining
cytoplasm and dense elongated nucleus loeated in the upper half of the cell. Narrow cells May be
involved in the protection of the epididymal epithelium or modify the pH of the lumen resulting
in the quiescence of spenn motility in the proximal end of the epididymis (Cohen et al.~ 1976;
Adamali and Henno, 1996).
Apical cells
Apical eells are found in the initial segment and the intennediate zone. These goblet
shaped eells constitute 10% of the total epithelial population in the initial segment and 1% in the
14

•
•
•
intermediate zone (Adamali and Henno7 1996). Apical cells nuclei are pale round or oblong and
located in the upper balf of the cytoplasm. However7 the specifie funetion of these cells is still
unknown.
Clearcells
Cleac cells are found only in the capu~ corpus., cauda epididymides. ln the capu~ they
account for 5% of the cells, 6.5% in the corpus and approximately 10% in the cauda region
(Robaire and Hermo, 1988). The primary function of these cells is to endocytose a varlety of
substances, sucb as immobilin (Hermo et al'7 1992b). As with principal cells7 clear cells,
depending on the region of the epididymis, serve different funetions as they show regional
specificity in their endocytic uptake of various substances (Le~ 1978; Flinckinger et a1'7 1988;
Hermo et al. 7 1992b; Vierula et al., 1995).
Halo cells
Halo cells are distributed throughout the epididymal epithelium and constitute less than
10% of the total cell number in any region (Robaire and Hermo. 1988). Halo cells are thought ta
be intraepithelial monocytes due ta their identical appearance ta circulating monocytes in the
human blood., but their origin and funetion œmain obscure (Robaire and Hermo, 1988).
Basal cells
Basal cells are found throughout the epididymis and account for 12% of the total cells in
the initial segment and 21% in the corpus and cauda epididymis. (Robaire and Herm07 1988).
lbese cells are small round or elongated ceIls with a large nucleus and a small amount of
cytoplasm. They line the basement membrane and do not reach the lumen (Robaire and Hermo,
1988). They possess an endocytic apparatus7 however,. a functional lOle in secretion or
endocytosis bas yet to he detennined.
Epididymtlllllnctions
There are numerous functions ascribed to the principal cells of the epididymis. These
include secretion~ endocytosis, and the conversion and metabolism of steroids. AU these
functions provide the appropriate enviromnent for spermatozoa maturation and storage.
IS

•
•
•
The synthesis and secretion of ions (Wang et al., 1978)~ small organic molecuJes
(Marquis and Fri~ 1965; Hinton and Setchell, 1980)y and a variety of diffèrent glycoproteins
(Flickinger, 1983; Henno et al., (994), such as immobilin (Henno et al., 1992b) and apo J (Hermo
et al., (991) have been weIl documented. The list of genes synthesized and/or secreted proteins,
by the epididymis is ever growing. The synthesis and secretion of proteins in the epididymis bas
been proven using morphologjcal and biochemical methods (Flickinger, 1985; Turner, 1991;
Hermo et al., 1994; Syntin et al., (996), Northen blot analysis and in situ hybridization and
immunocytochemistry (Lea et al., 1978; Henno et al., 1991, 1992b; Rankin et al., 1992; Vierula
et al.y (995). In various cases, these secreted proteins become associated with the spennatozoa in
specifie regions and this might promote spenn maturation and/or spenn-egg interactions.
In addition ta secretion, principal cells endocytose a number of proteins, sorne of wmch
are derived from the lestis (Robaire and Henno, 1988; Henno et al., 1994). Furthermore, sorne
proteins secreted in the proximal epididymal region are reabsorbed in the distal regions (Hermo et
al., 1992b). From the vast 3IOount of studies in the literature~ it is clear that endocytosis plays an
important cole in the epididymis. It bas also become evident that endocytosis is often a region
specifie event judging from the disaibution of lysosomal enzymes in principal and other epithelial
cells. It could thus be concluded that it is the coordinated activity of secretion and endocytosis of
various substances by epithelial cells along the epididymis that influences the luminal
environment where the final maturation of the spenn occurs as weil as their protection and
storage.
Storage and protection are other important functions ofthe epididymis. The spennatozoa
storage site is the cauda of the epididymis. It takes 3 to 19 days for spenn to travel through the
epididymis and spennatozoa can be stored for as long as 30 days before they are reabsorbed
(Orgebin..cri~ 1975). Spennatozoa are also protected within the epididymal environment
established by a blood-epididymal barrier (Hoffer and Hinto~ 1984; Cyr et al., 1995) which
prevents sperm damage.
EpididYllltll Regullltioll (tlnJrogells)
16

•
•
•
It has been established that the epididymis is dependent on androgens for creating a
luminal environment inducive for sperm maturation. Androgens regulate growth and
differentiation~ normal morphology, ttansport of ions and synthesis and secretion of various
proteins including those involved in adhesion and sperm functions (Cyr et al.~ 1993; Cornwall
and Hann~ 1995; Hinton and Palladino, 1995: lobaire and Viger~ 1995; Orgebin-Cri~ 1996).
Knowledge of the funetions of each protein and their regulation is of importance in
comprehending the key events leading co spenn maturatio~ protection and storage ofspenn under
normal conditions. Although rat epididymis is the model system in this study, increasing
evidence is pointing to the faet that similar regions and cell types are constituents of the buman
epididymis. The various epithelial cells even appear in the appropriate regions~ sbare similar
structural features and funetions including region specifie differences in expression for some
secretery proteins as weil as acquiring their full fertilizing capability in the same regions (Palacios
et al.~ 1991, 1993: Yeung et al., 1991, 1993, 1994: Krull et al .. 1993: Q'Bryan et al., 1994;
Turner, (995).
DevelopmentalAspects ofthe Male Reproductive T,act
The postnatal developmenta1 aspects of the male reproductive~ (Knorr et aJ.~ 1970;
Papp et al., 1994: Sun and Flickinger~ 1979: Henno et al.~ 1992a) have been described in rats
ranging from birth to adulthood (day 90). As early as day 39, a full complement ofgenn cells and
Sertoli cells can be found fonning the seminiferous epithelium but few spermatozoa are released
into the lumen of the semioiferous tubules (Papp et a1.~ 1994). It is by day 49 that an adult male
pattern is acquired in terms of size of the tubule~ and number of elongated spennatids per tubule
(Papp et al., 1994). Ciliated cells of the efferent duet can be seen at day 21 (Hermo et al., 1992a).
Nonciliared cells become adult in appearance by day 49 with Many endosomes and dense
lysosomes in the supranuclear region. Interestingly, their Golgi apparatus is weil developed by
day 21 (Hermo et a1.~ 1992a).
In the epididymis, columnar cells differentiate iota principal cells throughout the
epididymis by day 39 when they acquire a distinct brosh border, prominent Golgi apparatus and
17

• endosomes and lysosomes (Sun and Filickinger~ 1979; Hermo et a1.~ 1992a). In the caput, corpus
and proximal cauda, elear cells are fully differentiated only by day 49 (Hermo et al., 1992a).
Clear cells of the distal cauda are fully mature by day 39 even though no spennatozoa are found in
the lumen. This can be due ta the large number of degenerating cells 611ing the lumen which
often were recognized as being spennatids and spennatocytes (Henno et a1.~ 1992a, 1994b). ln the
epididymis~ spermatozoa arrive in the initial segment and caput by day 49 and reach the cauda on
day 56 (Robaire and Henno~ 1988).
Taken together these observations suggest that it is a eombination ofdifferent factors such
as the level of androgens which increases fram day 21 te 56 (Scheer and Robaire~ 1980: Jean
Faucher et al., 1985)~ the presence of spennatozaa and Sertoli cell produets that is needed for the
structural differentiation of the various epididymal epithelial eell types.
•
• 18

• Female Reproductive System of the Rat
The localizarion of LRP·2 was aIso explored in the fem31e rat system. Therefore a review of the
anatomy~ histology~ physiology and development of the femaJe reproductive system of the rat is
also provided.
•
OV1'Y -----,
Uterine Ham-----\
} Ovidue:t
•
------UtatUI
Vagin.' 1
Figure .a: Schematic representation of the ovary t oviduet. uterine hom, uterusand vagÏJ1i1 of the 3duJt r:LL During titis study the oviduct wu divided iDto rworegions: pro:<imal (p) and distal (d), and the uterine harns and uterus weregrouped into uterus collectively.
19

• Oviduct
Structure
Whereas the male rat reproductive system is very similar to the human\> the female rat is
very different. The human oviduet consists of three segments. which are anatomically weIl
demarcated : infindibuJum. ampulla and isthmus. The rat oviduct is much smaller and more
convoluted. Only two histologically distinct regions are found. the proximal (closest 10 the ovary)
and the distal (closest to the uterus).
Proximally the mucous membrane fonns up to eighteen longitudinal folds. which often
branch. Height and number of folds decrease distally and almost disappear at the uterine end of
the oviduet. The epitheliwn coosists largely of a single layer of columnar cells~ which are often
ciliaœd. Ciliated ceUs are MOst numerous in the proximal oviduet. The wave of cilia is directed
toward the uterus. to aid transport of the ova distally. The nonciliated (peg) eells usuaIly contain
apical secretory vacuoles. Both types of epithelial cells lining the oviduet undergo cyclical
• variations in response ta changes in hormone levels. particuIarly estrogens.
The lamina propria of the oviduet contains numerous blood vessels and isolated muscle
bundles" wbich continue into mucosal falds. The muscuJar coat increases in thickness toward the
uterus. The mainly circular layers are complimented in the distal part of the oviduet by a
longitudinal layer ofmuscle.
Uterus
Structu,e
The uterus of the rat also greatly differs ftom that of the human uterus. In the rat it is
classified as a uterus duplex (Figure 4). The lumina of the uterine homs are completely separated
and open as a paired orifice. In this study the uterine horns and uterus are grouped collectively as
the uterus. The uterine endothelium is lined with a simple columnar epithelium with a miûW'e of
secretory and ciliaœd ceUs. This endometrium varies in tbickness during phases of the menstnaal
cycle. The surface epithelium invaginates into simple tubular glands~ wbich during the course of
• 20

•
•
•
this thesis will be referred te as uterine glands. The rat endometrial stroma is highly cellular and
cantains abundant intercellular ground substance.
DevelopmentlllAspects oftlte Femille Reproductive T,act
Honnones are the MOst important fadors involved in the development of the female rat
reproductive tract. The honnonal events associated with the natura! onset of puberty in the female
rat have been well documented. The initial ovulatio~ marks puberty in female rats. This occurs
during the interval of 36 to 40 days of age, as a result of the surge in gonadotropins fSHILH.
Preceding the preovulatory surge, increased uterine weight, growth in height of endomerial
luminal epithelium and the presence of increased mitotic figures in the surfàce epithelium and
endometrium have been reported to begin around clay 32 which strongly correlates with increases
in estrogen (Advis et al. 1979). The effect of esttogen and androgens was further demonstrated
when the onset ofpuberty (initial ovulation) was advanced to as early as 20 days by administering
estrogen (Ramirez and Sawyer, 1965) and aromatizable androgens (Knudscn and Mahesh, 1975)
to neonatai females.
At all ages prior ta Day 29 the epithelium lining the uterine lumen is cuboidal and devoid
of mitotic figures. At Day 30 hypertrophie aetivity of ail uterine tissue begins. The lumen
becomes more dilated and the epithelium changes to a tall~ colwnnar~ non-ciliated, with nuclei at
different levels ofthe cells (Knudsen et al. 1974).
Rapid cclI division was demonstrated by a rise of mitoses starting on day 30, then peaking al day
32 (Knudsen et al. 1974). From day 32 until the initial rise in esuogen and progesterone al the
time of the first ovulation (between da,,'s 36-39) no remarkable ditferences have been noted
throughout the oviduet and uterus.
The period between 36 and 39 postnatal days is consistendy the time when the femaie rat
exhibits hormonal changes leading to puberty. Circulating estrogen levels remain low while
serum progesterone rises between the first diestrus and early proestrus. While serum progesterone
levels decrease between early proesttus and late proestrus, estrogen concentration drastically
increases te ma.ximai values, the same time that uterine weight significantly rises. This rise of
21

• estrogen during late proestrus is followed immediately by the simultaneous surge of
gonadotropins FSH and LfL resulting in ovulation during estnls. Once ovulation has occurre~
hormone and gonadotropin levels retum te basal values and adulthood has been ac:hieved, the
estnls cycle in the female rat will continue for the life of the animal.
Estrus Cycle
The rat belongs te a group ofanimais with short (4 to 5 days) estrous cycles~ during which
a dozen or more eggs are ovuJated. These characteristics retlect the short life span and thus the
need for a high reproductive potential. During any phase of the cycle medium·sized Collicles (300
ta 500 um) are presen~ part of which undergoes atresia and the other part ovuJates. The ovarian
changes are accompanied by charaeteristic changes in the oviduet and uterus.These changes are
influenced by the different honnoDailevels ofeach stage of the estrus cycle(Figure 5).
Oiestrus (57 hours)
Hormonally. diestlUs is marked by a rise in estrogen~ while progesterone~ and the
• gODadOtropinS LH. and FSH remain low until Proestrus. During diestrus. corpora lutae and
follicles graw eontinuously. Regeneration of the DOW high columnar uterine epithelium is
completed. The leukocytic infiJtration of eodo and myometrium is reduced ta a minimum.
VaginaI epithelium is low. a smear contains small epithelial cells and leukocytes (Long and
Evans, 1922; AJlen.,1931:Gorbman and Bem, 1962). Part ofvaginal smear eells are contributed by
the epithelium of the cervi""< uteri (Hamilton~ 1947).
Proestrus ( 12 hours)
Proestrus is the most active stage in the 4-day estrus cycle of the rat. Estrogen reaches peak levels
in the early part of proestrus. SubsequentlYt the sharp rise in progesterone is followed by the LH
surget and the initial increase in FSa which is demonstrated in the late part ofproestrus. During
proestrus the uterine lumen is distended with fluid., the shape of the uterine epithelial cells
changes from eolumnac to cuboidal during this distension. Stroma and myometrium show an
extensive infiltration with leukocytest mostly eosinophilic granulocytest that persists tbroughout
• 22

•
•
•
esuus. Vaginal epithelium is thick. a comified layer develops beneath the superticial cells. The
vaginal smear cantains nucleated and relatively small epithelial cells.
Estrus (12 hours)
At the time of Estrus.. levels of sex hormones estrogen 3Ild progesterone have retumed ta basal
levels. Only FSH remains al peak levels. to fucilitate complete foIlicle maturation. During titis
period follicles grow ta their ma.ximal size. The uterine lumen is distended ma.~ally. The
vaginal epitheliwn is now ~overed by layers of comified eells; the vaginal smear therefore
contains moderare numbers of large comified squamous ecUs. Only al the end of proestrus and
during estnls will females accept the male animal.
Metestrus (21 hours)
During the early metesttus (15 hours) ovulation takes place. The fluid content of the oviduet
increases. The liquid content within the uterus diminishes~ the low cuboidal epithelium shows
vacuolar degeneratio~ leukocyte infiltration of stroma and the myometrium decreases. The
eomitied ceU layers of the vagina become detaehed~ the smear contlins a gre;tt number of large
squamous cells.
_f\1 ....t 1 CI l Pro 1 &
Oay Qf " • Oay C'tée
Figure~: Tc:mpor:ù changes in àrcul:umg concemr:mons of goaadoU'Cpinsme! steroid hormones. and in sc:wa1 ~cptivity. dlrcupou( the iDfcniIe:ovuWory ~cle: of the: m. The: r:at -kJay esuus ~cfe: is brokcn up mEometestlUS.. dîc:suusy proesuus. :wt estI'US. l'bis figure was adaptai â'omSmith. Frecman. Jnc:l SetH (1975).
.,.._.J

•
•
•
~IATERIALS AND METHOOS
Animais
Male and Female Sprague Dawley rats were provided by Charles River Laboratory Ltd.
(St. CODstan~ Que.) Male rats at each of the following intervals after birth 7, 15, 21, 29, 39, 49.
56, and 90 days were sacrificed 10 prepare tissue for light microscope immunocytochemical
analysis ofLRP-2 antibody localization in the efferent ducts and epididymis. Female rats at 7, 14,
21, 29, 39, 49, and 56 days were killed te evaluate the immunolocaJîzation of LRP-2 antibody in
the oviduets and uterus.
Vaginal Smear
In arder to detennine the estrus cycle stage of the perilpostpubertal females at 39, 49, and
56 days a vaginal smear was taken. Female rats aU were maintained in lightldark cycle and were
periodically exposed ta human presence for feeding. Four rats were sacrificed at each past
pubertal age (ie. 39.49,59 days). Stages of the estrus cycle were detennined in the rats: diestnls,
proestrus, estrus or metestrus by vaginal smear. This allows for the identification of vaginal
epithelial eells and., therefore, the stage of the estrus cycle. Vaginal cells were mixed with
wannecl distilled water pipetted iota the vagina of the rats. A drop of the mixture was mounted
onto a microscope slide for visualizatiOD. Diestrus is characterized by a large amount of smalt
round nucleated cells and leukocytes. Proestrus presents large, round nucleated epithelial ceHs.
Estrus is rich in comified epithelial cells. Metestrus is characterized by a mi.~re of cornified
epithelial cells and leukocytes (Wang et al., 1923). Once the stage of the estnls cycle was
detennined by vaginal smear, sa that one rat at each postpubescent stage demonstrated in one of
the four menstrual stages., the rats were perfused and dissected.
Perfusion and Dissection of the Male and FemaJe Rat Reproductive System
Perfusion was perfonned according te the technique described by Vitale et al. (1973).
FolloWÎDg anesthesia with pentobarbital (Somnitol~ MTC Pharmac:eutica1s, Hamilton, ON)
injeeted intraperitoneally, the abdominal wall of the rat was incised, and the lower part of the
abdominal aorta exposed and cleared ofoverlying tàt and connective tissue. The abdominal aorta
24

•
•
•
was chen grasped with a pair ofcurved forceps al its point of bifurcation. An 18-22 gauge needIe,.
connected via a polyethelene catheter to a bottle of lactated Ringer"5 set at a height of 130 cm
above the table,. was introduced into the lumen of the aorta in a rettograde direction above ifs
bifurcation. The organs quickly blanched<t as the blood was displaced by the lactated Ringer"s.
Following a quick rînse,. Bouin"s fixative was perfused" until the specimen had clearly contracted.
ln the case of the male" the reproductive tract from the testis ta the proximal part of the
urethra was excised. &ch epididymis was then cut along its long a.xis in order for each region ie.
efferent ducts.. initial segment, intermediate zone, proximal and distal caput, corpus and cauda to
be examined. In the femaIe, the tract was removed from the ovary to the distal end of the uterus
then placed iota Bouin"s solution for better fixation.
Licbt Microscope Immuno(ytochemistry
D~ltytiration: The tissues were dehydrated in graded ethanol; they were immersed io1o 500/0
ethanol for half an hour, theo transferred twice to 10% ethanol for five hours. Fmally, the tissues
were placed into 95% ethanol for one hour,. then in 100% etbanol for another hour.
EmbeddÙfg: The embedding process began with a solution of 50% ethanoU50% dioxane in
which the tissues remained ovemight at room temperature. They were transferred tluee tintes ta
100% dioxane for one hour each. The tissues were left in 500/0 dioxane/SO% paratlin at 58-S90C
ovemight. Lastly,. the tissues were in 100% paraffin at 58-S90C three times for one hour each.
Paraffin blacks contaîning the tissues were made and sent ta the Montreal Children·s Hospital for
sectioning.
Immunocytochemistry: Paratlin was removed with three successive solutions of 100% xylene, 5
minutes each. The tissues were then rehydrated with 1: 1 xylene/ethanol and graded ethanol
solutions (100%, 950/~ 700/0, 700/~ 10%, 700/~ 50%). The second 70% ethanol solution contained
1% hydrogen peroxide (2m1l200ml of solution) ta inactivate any endogenous peroxidase activity.
The third. 700/0 ethanol solution contained 1% lithium carbonate to neutraiize any residual picric
acid in the tissue. The tissues were then immersed in 300mM glycine for 5 minutes 10 block free
aldehyde groups. The sections were then blocked with 40m1 of 10% goat serum diluted in TBS
2S

• (pH 7.4~ 20mM Tris-base saline 0.1% bovine serum albumin) for 15 minutes at room temperature.
Anti..rabbit LRP..2 primary antibody (kindly provided by Dr. S. Argraves) (diluted 1:50) was
applied ta the tissues and incubated for 1 hour and 15 minutes al 33°C. The same was done with
anti..rabbit SGP-2 primary antibody. The sections were again blocked with 10% gOal serum in
TBS and left al room temperature for 5 minutes. Goat-anti-rabbit secondary antibody (diluted
1:250) was later applied ta the sections and incubated al 35°C for 45 minutes. The slides were
immersed in a solution ofdiaminobenzidine (0.25g ofDAB with 0.4g ofimidazole in TBS) for 10
minutes. They were subsequendy washed in two successive wells of distilled7 then in a solution
of0.1% methylene blue for 15 minutes and washed again in two wells ofdH20. The tissues were
tinally dehydrated in graded solutions of ethanol (50%, 70%~ 95%, 100o/~ 1: 1 ethanoUxylene,
xylene, xylene). The slides were pennounted and left to dry. Light micrographs were later taken
by a Zeiss photomicroscope ta visua1ize the localization of LRP-2.
•
• 26

•
•
•
RESULTS
Postnatal Developmentallmmunoloealization of LRP-2 in Efferent ODeu and Epididymis ofMale Rat
At each postnatal age from day 1 to day 90 the epithelial non-ciliated cells of the efferent
duets revealed an intense immunoreactive staÏn. This appeared at low magnification as a dense
band in the apical region of the non-ciliated cells. The supranuclear region also presented an
immunoperoxidase reaetio~ however this was granuJar in appearance (Platel; Fig A..D and Table
1)
ln comparison to the profuse reaction observed in the more cuboidal efferent duet cells
there was no reaction over the principal cells lining the proximal initial segment at ail ages (Plate
2; Fig A-F and Table 1). The principal cells of this region were undifferentiated until day 39.
Even wben spennatozoa was abundant in the lumen of the proximal initial segment (i.e. day 56)
the principal cells remained devoid ofanti LRP-2 antibody (Plate 2: Fig E).
The distal initial segment displayed a mueb different pattern of developmental
loca1ization of LRP..2 than the proximal segment. On days 7 and l5~ the undifferentiated
principal celis lining the distal segment were inununonegative (Plate 3: Fig A). On day 2l~
bowever. these undifferentiated cells exhibit a thin moderate immunoreactivity in the apical
region (Plate 2; Fig B). The stain becomes thicker in day 29, and increases ta a peak
immunoreactivity al day 39 (Plate 3 Fig C&D). At titis age the differentiating taU columnar cells
reveal dense staining in the apical regio~ and a sprayed localization of LRP-2 antibody in the
supranuclear region ofsome cells. Spermatozoa were first evident in the lumen of the distal initial
segment al day 49. Interestingly, on the same day, there was no immunoreactivity of LRP-2
antibodies~ in the principal ecUs. The absence of staining for LRP-2 antibodies was shown by
eacb specimen after 49 days (ie.~ 56 and 90 days)l' when luminal spennatozoal content was
increasing ( Plate 3; Fig. E&F and Table 1).
The intermediate zone was ooly tirst distinguisbable from the distal initial segment at day
39 because of structural differences between principal cells. The principal cells of the
27

•
•
•
intennediate zone were shorter and more columnar titan those of initial segment. More
convincingly7 the principal cells of intermediate zone revealed the presence of distinct apically
located vacuoles (1-3 per cell). At higher magnification the principal ceUs of the intennediate
zone were profusely immunoreactive al every postnatal age from day 397 ta adult age of 90 days
(Plate 4;Fig. A·D7 Table 1) .
Light microscope analysis of immunocytochemistry in the proximal caput showed a
slightly different pattern of localization of LRP·2. In 7 clay and 15 day tubules, the proximal
caput revealed DO staining for LRP-2 antibody (Plate 5; Fig. A). The epithelium lining the tubules
in the proximal caput of 21 day and 29 day epididymides aIthough still undifferentiated7 displayed
a moderate immunoreactivity in the MOst apical portion of the principal cell (Plate 5; Fig.B&C).
As in other regions, the principal cells of this region showed the bighest expression of LRP·2 at
clay 39 and day 41. At low magnificatio~ this appeared as a dense CODtinUOUS band loeated in the
apical region (Plate 5; Fig.D). Immunoperoxidase stain was also scattered in the supranuclear
region ofthe principal ceIls at day 39 and day 41. An intense immunoperoxidase staining reaction
produet was also observed in low and high magnifications of 49, 56, and 90 day proximal caput
regions (PlateS; Fig.E&F, and Table 1).
In contrast to the proximal capu~ the distal caput revealed a developmental localization7
which was dramatically diffèrent. LRP-2 was initially localized apically al postnatal day 21 (Plate
6; Fig.B). A similar pattern of moderate apical staining was seen al clay 29 prior to the complete
absence of LRP·2 antibody revealed in days 39 -90 (Plate 5; Fig.C·F, and Table 1).
The principal cells lining the epithelium of both the corpus and cauda regions revealed an
identical pattern of LRP-2 expression. Each segment became moderately stained in the apical
region al day 21 (Plate 7; Fig. B " Plate 8; Fig. B). The immunoreaetivity became more intense
in clay 29t when it appeated in the supranuclear region of the principal cells (Plate 7; Fig.C "
Plate 7; Fig.C). On day 397 when clear cells became evident, a dark band of immunoperoxidase
stain was observed in the corpus and cauda. It was also noted on day 39 that a blotched pattern
developed in the tubules ofthese regions. Certain principal cells revealed staining ooly a10ng their
28

• apical membraneT other principal cells showed staining throughout their entire supranuclear
regioDT while clear cells remaioed immunonegative (Plate 7; Fig. D, Plate 8; Fig. D). The same
intense pattern ofstain was observed for principal cells in the corpus and cauda al 41, 49T 56 days,
and in the adult 90 clay old rat (Plate 7; Figs.E&F, Plate 8; Figs. E&F).
•
• 29

•
•
•
REGIONS 7DAY 15 DAY 21 DAY 29 DAY J9DAY 41 DAY 49 DAY 56 DAY 90DAY
E.D. ~ ++++ ~ ~ ~ ~.,.............~ -
~/.S. . . --1· .. . . .. .. -IJ)./.S. --/. ../- ~ ~ ~ +-r-r+ +/.. +/- ~/·
/.L - . - - ~ ~ ~ ~ ~
l/'ROX C.fPtff +/- --/- - - ~ -H-H- ~ ~ ~
'D1ST.CAPUT +/- +/- ++ ++ +/.. . - . -CORPUS +/. ++ - - ++- +-+-* ~ -C-tllDA +/. +/.. ++ -H-+ ~ ~
.,.........~ ~
Table 1: A developmental study of the regionaI expression ofLRP-2 in the efferent dUelsand along the length of the epididy~is. The number of positive signs represent theintensity of the immunoperoxidase reaction in the principal cells. The negative signrepresents lack of immunoperoxidase reaetion.E.O. P.I.S~ D.I.S~ LZ, P.Cap, D.Cap, Corp, Cau represe~ ,fferent duets, proximal anddistal initial segmentintermediate zone, proximal and distal caput, corpus~ proximal anddistal cauda respeaively.
30

e • •Developmental Localization ofLRP·2 in the Efferent Duets and Epididymis of Postnatal Rats Aged 7-90 Days
l'Id.••
lld., Sd.7d.
w.....
ID ED
Figure 6: Schematie representation of the localization of LRP-2 in the efferent ducts and in the various regions of the epididymis of postnatal ratsal increasing ages. The regions of the epididymis were divided on the basis of the height of the epithelium, the different ceU types, grossmorphology, the size of their tubules and by their immunocytochemical staining pattern of principal ceUs as determined by Iight microscopy. Thelabelled regions of E.D' t p.I.S. t IZ. P.C., D.C., Co., and Ca. represent the efferent duets, proximal and distal initial segmen~ intennediate zone,nrnvin\~1 ~ntl tfid~1 ....anllt t'nmllSo and cauda resoectivelv.

• Ligbt Microscope Postnatal Developmental Immunolocalization of LRP-% in the Oviduct,Uterus and Uterine Glands or Female Rats)
Postnatal day 29 was the earHest age al which LRP-2 was localized in the female
reproductive tract. A scarce~ granular immunostaining was revealed in the deep falds of the
proximal oviduct (Plate 9 Fig. A). The distal oviduc~ uterus, and uterine glands all showed a
profuse peroxidase staining ofLRP-2 antibody in the apical region ofthe ceUs (Plate 9 Fig 8-0).
Postnatal day 39 exhibited the mast abundant immunolabelling of LRP-2 antibody, ofany
other day in the study. The tall columnar secretery cel1s of the uterus demonstrated a strong
immunoreaction of LRP-2 antibody throughout each menstrual stage al titis age. Proestnls ofday
39 elicited LRP-2 antibody immunoreactivity in the apical regions of secretory ceIls in each
region. The noncilicated ceIls of the proximal oviduct clearly showed the granular appearance of
peroxidase label (Plate Il Fig. A). The staining in the distal oviduet was strong and appeared te
be deeper, in the supranuclear region of some eeUs. The uterus and uterine glands demonstrated
the granular appearance of LRP-2 antibody immunolabel concentrated a10ng the luminal
• membrane (Plate Il Figs 8-D).
Only minor developmental changes were observed during aduJtho~ from day 39 to day
56. The uterine glands, which on day 39 proestrus were intensely immunopositive (Plate Il Fig.
D), were completely devoid of LRP-2 antibody on day 49 Proestrus (Plate L5 Fig. D). On clay 56
proesttus, the uterus which on day 39 and day 49 exhibited a profuse expression of peroxidase
label~ aise became devoid of LRP·2 immunolabel (Plate 19 Fig. D). A similar disappearance of
LRP-2 antibody immunostaining oc:curred in the uterus in diestrus from day 39, when a positive
LRP-2 reaction was revealed (Plate 10 Fig. Cl, te day 56 when no peroxidase labeling was evident
in the uterus (Plate 19 Fig. Cl.
Estrus was not on1y the MOst immunoreaetive stage in tenns of intensity of staining, it
was also the only menstrual stage to demonstrate a consistently strong immunoreaction of LRP-2
antibody in each of the distal ovidu~ uterus and uterine glands, at each age after the onset of
puberty. At day 56 estnIs displayed a moderate, inununoreactivity in the uterus and uterine gland
•32

•
•
•
(Plate 20 Figs. C&O), when each ofthe other stages were immunonegative in these regions (Table
2, Plate 20).
Metestrus, was round ta be the estrous cycle stage which revealed the least amount of
immunostaining throughout each region, at every age in the study (Plates 13, 17, 21, Table 2).
Therefore following the tise in LRP-2 immunoreactivity at estrus, a decrease in LRP-2 expression
was demonstrated throughout the female reproductive~ at each age.
Regional differences in immunoreaetivity were evident, tbroughout the study. The distal
ovidu~ where fertilization OCCUlS, consistendy exhibited substantiallabeling of LRP-2 antibody.
The proximal oviduet demonstrated a weak ta moderate staining, along the apical aspect of the
non-eiliated cells, within the deep folds of this region. The uterus and uterine glands varied in the
immunoreaetivity of LRP-2 antibody labeling. As the female aged, it was apparent that
immunostaining of LRP-2 antibody diminished in the uterus and uterine glands, except in estrus
where it remained present throughout the study.
33

•
•
•
29 Do."Y 11
PROX. QVIDUCT .;-
OrST. OVID{;CT +---+
UTERUS ~
{Jl'ERlNE GLA.'ID ~
J9DAY DIESTRUS PROESTRUS ESTRVS JfETESTRUS
,PROX.OVIDt"CT ~ ++ +/- +/-OIST. OVIDtJCT ++ ~ - +r..lTERUS ~ ~ ~ ++l:-rERINE G~\lD - .;-.0...,... 1"+- -"'9 DAY DIESTRllS PROESTRUS ESTRUS .'4ETESTRUS
,PROX. OV1I)UCT - + ++ +DEST. OVIDUCT ~ ~ +-+ +UTERUS -- ~ +-0-+ -L1"ER!NE GLAND - - ~ .
56 DAY i DIESTRUS PROESTRVS ESTRUS .'ffETESTRUS
PROX. OVlDUCT ~ ~ ~ -DEST. OVIDUCT ~ -H- ~ ~
CTERUS - - +-- -L'TERINE GLAND . - +-- -
Table 2: A developmental study of the regional expression of LRP-2 in the proximalovidu~ distal ovidu~ uterus and uterine glands. The number of positive signs representthe intensity of the immunoperoxidase reaetion in the principal cells. The negative signrepresents Iack of immunoperoxidase reaaion.
34

•
•
•
DISCUSSION
Postnatal Df!1Ieiopmental LocaiiVlliotl of LRP-2 in the Efferent Ducts and Epididymis of tireMateRai
The immunostaining of LRP-2 antibody in the epithelium lining the tubules of the
efferent duets was intense at every postnatal age. The predoMinant ceU type of the efferent duets
is the non-eiliated cell, which reveals il highly developed endocytie apparatus al clay 49 (Henno et
al.~ 1992)~ presumably to endocytose spennatozoal proteins~ which are initially present in the
lumen of the etferent ducts on tbis day. The intense immunoreactivity of LRP-2 antibody in the
apical membrane of efferent duet tubules as early as day 7 suggests that spennatozoa have no
effect on LRP-2 expression in this segment. The dense staining observed al carly ages May resuIt
however~ from plasma androgens~ whieh were reported te he very high in newbom rats (Dohler
and Wuttke 1975~ Sun and Flickinger 1980)~ or by another testicular factor that remains to be
identified.
The consistent immunoreactivity of the non-ciliated cells of the efferent duets in contrast
te the complete immunoreactivity observed in its neighboring segment aise raises an important
point. In cach postnatal day of the study, the proximal initial segment displayed a total lack of
staining of LRP·2 antibody, despite exposure to the same levels of luminal factors as the efferent
duets. This distinct contrast May he related te a difference in function., carried out by the various
segments. For example~ it bas been proposed tbat the non-eiliated ceUs of the efferent ducts
endocytose testicular apo 1 wmch detaches from spennatozoa in the lumen of the duel (Herma et
aI.~ Water 1994). For this mechanism it is clear why LRP-2 is 50 heavilyexpressed in these cells.
The proximal initial segment seems ta be a transitional region between the effeœnt duets where
testicuJar apo 1 is endoeytosed~ and the distal initial segment, intermediate zone't and proximal
caput where epididymal apo 1 is mostly secreted (Henno et al., 1994). This may he the reasan for
the complete absence of immunostaining in this region. In consequence the absence of LRP·2
May favour the interaction between the epididymis apo 1 and the plasma membrane of luminal
spermatozoa.
35

•
•
•
The results of the study by Henno et al. (1994) which examined the apo l expression
during postnatal development of the epididymis and efferent ducts showed that the greatest
expression of apo l appeared in the distal initial segmen~ intennediate zone and proximal caput.
The immunocytochemical results of this study indicate that the highest expression of LRP-2 took
place in the same regions during the course of postnatal development (i.e. distal initial segmen~
intennediate zone~ and proximal caput). This apparent colocalization May provide evidence that
the high expression of LRP-2 in the intennediate zone, and proximal caput results from the
secretion and removal of apo J which is known te he abundant in these regions (Henno et al.
1994).
A region that revealed a pattern drastically ditTerent from any other was the distal caput.
This region is loeated between the proximal caput and the corpus~ and was devoid of staining al
every age after 39 days; when il's bordering regions revealed intense reaction produets. The
significance of the contrast in staining is unclear but it May be conneeted to the difTerent functions
carried out by the various segments. The distal caput region drains the contents of the
intermediate ZODe~ and proximal caput (sperma1Ozo~ secreted apo l and other epididymal
proteins) into the corpus and cauda where it will he stored until ejac:uJation. Agam, the reason for
the absence of LRP-2 in the distal caput is to favour a longer interaction between apo l and the
spermatozoa plasma membrane~ after it~s residence in the intermediate zone. Nevertheless~ the
significance ofthis interaction remains to be established.
It bas already been noted tbat tbere are secretory and endocytic mechanisms in the
intennediate zone and proximal caput which require high expression of both apo 1 and LRP..2.
The shorter epithelium of the more tenninal corpus and cauda is involved in spenn storage. In
these regions LRP-2 is consistently localized with intense immunoreaaivity from day 29 te day
90 (Le. adult age). The fact that sperm and the expression of apo 1 is not present in these regioDS
uotil day 56 (Henno et al. 1994), suggests that neither of these factors influence LRP-2
expression. However~ since the corpus and cauda are the regions where spenn accumulates until
36

•
•
•
ejaculation., LRP-2 expression may be triggered by a luminal factor in anticipation of sperm
storage.
The most intense immunostaining ofLRP-2 antibody, in all regions., with the exception of
the proximal initial segment and the distal capu!:. occurred on postnatal day 39. On the same day,
the epithelial cells in each region resembled the columnar principal cells of the adult. It was also
noted by Henno et al.., (1992) that on day 39, the columnar epithelial cells possessed all the
structural femlles characteristic offully differentiated cells. funhermore, day 39 marked the tirst
appearance of large vacuoles in the intermediate zone, and clear cells in the capu!:. corpus and
cauda Therefore factors involved in the reguJation of structural difi'erentiation of ceIls, seem ta
he common to those influencing LRP-2 expression in the various epididymal regions.
In the present study, the expression of LRP-l in the efferent ducts and regions of the
epididymis prior ta the presence of spermatozo~ suggest that sperm docs not influence its
localization. While the effeet of androgens on the expression of LRP-2 in the epididymis is
unclear. there is evidence from this study, which suggests tbat they may indeed play a role. Scheer
and Robaire (1980) reported that plasma androgens increase dramatica11y by clay 35. which was
around the same rime that LRP-2 was highly expressed in the epididymal regions. Despite the
coloca1ization of LRP-2 and apo l in the same regions at certain postnatal ages, this research
demonstrates that LRP-2 expression is not solely dependent on the presence ofapo 1.
37

•
•
•
Postnatal Deve/opmental Loca/it,lltion 0/LRP·2 in the Oviduets and Uterus 0/the Female Rat
The immunolocalization of LRP-2 in the epithelium lining the female rat reproductive
trae~ namely the oviducts, uterus and uterine gIands~ also showed regionally-specific changes
which May be related ta age~ honnone levels~ and/or possibly ta the presence of luminal factors
such as apolipoprotein J.
Day 29 marked the first appearance of staining in the short columnar cells of the distal
oviduCl uterus and uterine glands while ooly a weak reaction was observed in the proximal
oviduct. Knudsen et al. (1974) showed that prior to day 29 the oviducts, and uteri were inactive.
Uterine glands were presen~ but Iacking mucoid secretion. It was also noted that around day 29 a
dramatic increase in activity, characterized by hypertrophy of aU tissues occurred; the lumen of
the uterus and oviduets dilared to sizes approaching that seen in estrus. Parker et al., (1976)
suggested a dramatic increase in estrogen al around day 30 based on an increased uterine weigh~
and changes in endometriaJ epithelium and strom~ whicb was attributed to the abundance of type
6-7 follicles" secreting large amounts of estrogen. These data suggest a possible relation between
the increase in ovarian activity, specifically the estrogen secretion by large follicles around day
29, and the first appearance ofLRP-2 at this age.
It bas been demonstrated that puberty in the male and female rat (signified by first
ovulation) occurs around day 39. Therefore LRP-2 expression wbich initially occurred on day 21
in the male and day 29 in the female seems ta be independent of the onset of puberty. However
day 39 was the most intensely immunoreactive age of LRP-2 antibody in bath the male and the
female. Despite the diffèrent estrus stages of the female rats in the 39 day group the developed
38

• columnar secretory cells of the uterus consistently demonstrated a profuse immunostaining of
LRP-2. The distal oviducts also revealed a strong LRP-2 reaction throughout the cycle, al this
age. Knudsen et al.~ (1974) reported that the increase in estrogen levels trom the secretion of
large follicles reached a peak al around Day 38 triggering ovulation. At this time serum F5H
showed a 2··,fold increase and LH rose simultaneously by as much as 40-fold~ then declined to
prepeak levels. Therefore the abundance of stain observed in the female reproductive tract al clay
39 is coïncident with the increase in sex honnones, which occurs in the female rat at this time.
Another interesting observation is that in the age groups studied after clay 39 the LRP-2
localization became more concentrated in the proximal and distal oviduct On the other hand~
LRP-2 immunoreactivity became completely absent in the uterus and uterine glands except in
estrus where it appeared in each region througbout adulthood. Blandau 3Ild Moncy (1944) found
the number of spenn cells entering the oviducts to be exceedingly small when compared to the
• number present in the uterine borns. The narrow distal portion of the oviduct seems to be
responsible for the low number of spenn cells that reach the site of fertilization (Gaddum-Rose,
(981). The fertilization of the ovum by strietly mature and mobile sperm cells takes place in the
distal oviduet, ooly during estrus. Therefore the pattern of LRP-2 expression specifically in the
oviduets~ as well as the uterus and uterine glands al estrus may fend support to a suggested spenn
maturing function of LRP-2 prior to the acrosome reaction.
The varying bonnone 1evels of the estrus cycle also appeaœd to have an influence on the
expression of LRP-2. In diestrus from day 39 to day 56 LRP-2 immunostaining was moderate in
the proximal and distal oviduct. A strong positive immunoreaetion was exhibited in the uterus al
day 39~ but was absent in the uterus and uterine glands for each of the age groups thereafter.
Diestrus is characterized by low levels of L~ F5H and progesterone, however estrogen levels
•39

•
•
•
begin ta rise. Proestrus revealed a similar loealization pattern as diestrus, but ofgreater intensity.
During proestrus, there is a ma~wn level ofLH preceding the ma"<.Îmum level ofestrogen.
Only at the end of proestnJs and during estrus will the female accept the male. This
notable increase in immunoreactivity during proestrus may he related ta the rise in bonnone levels
and enhanced receptivity. The possible effects of bonnone levels on LRP-2 localization are also
indicated by the contrast in the pattern of immunostaining observed between estnls and metestrus.
Estrus was the MOst immunoreactive stage in tenns of intensity of staining. Furthennore it was
also the only menstrual stage ta reveal a consistent positive immunoreaction ofLRP-2 antibody in
each ofthese regions ailer the onset ofpubeny. Estrus is the stage during which the female mates.
Estrogen levels begin ta decline from peak levels while progesterone levels begin to rise.
Conversely. during metestrus immunostaining of LRP-2 was we~ or completely absent
tbroughout each region at every postpubertal age in the study. During metesttus FSH and LH
levels are low, and esttogen and progesterone levels have declined to basal levels.
The presence of apo l and LRP-2 in specialized epithelia of the male and femaIe
reproductive tmet (Zheng et al., 1994; Kounnas et al., (994) evoked speculation that apo JILRP-2
binding could have an important role in the reproductive process, namely the maturation of
spennatozoa. The immunoeytochemical colocalization of apo J and LRP-2 in the etferent duets
and specifie regions of the epididymis were previously described in the maie rat during postnatal
development.
Studies of the postnatal developmental loca1ization of apo J in the proximal oviduet,
distal oviduct, uterus and uterine glands were also condueted. Preliminary results reveal that apo J
is present as early as day 21 in the uterus and uterine glands. In each age group after day 21, apo l
antibody is most pronounced in the uterine glands and less prominent in the uterine epithelium. A
consistently positive immunoreaetion is demonstrated however~ subtle differences are notable
40

•
•
•
between estrus cycle stages. The high degree ofapo 1 staining in the uterine glands suggests that
these structures are continuously secreting apo 1 throughout the cycle. This data lends more
support to the proposai that LRP-2 binding ofapo 1 may be involved in spenn maturation. In the
female, apo 1 was revealed in the glands of the uterus where it May he secreted in preparation for
spenn passage.
Apo J May altemate binding sites trom the spermatozoa membrane to the LRP-2 receptor
loca1ized in the uterus, distal and proximal oviduet in order to redistribute spenn plasma
membrane lipids and proteins, possibly giving sperm the ability to recognize and fertilize the cgg
or to develop the acrosome reaction.
Conclusion
In conclusion, light microscope immunocytochemistry was performed using polyclonal
antibody to LRP-2 on the efferent duets and epididymis of male rats (aged 7-90 days) and the
oviduct and uterus of female rats (aged 7-56 days). The objective was firstly to detennine the
pattern oflocalization ofLRP-2. This research also aimed to demonstrate the relation between the
changes in immunoreaetivity of LRP-2 and developmentai conditions in the male and female
reproductive system. This study confirmed the presence of the protein receptor LRP-2 in the
reproductive epithelia of the male and female reproductive tracts. Fwthennore during postnatal
development, it was demonstIated that the appearance of LRP-2, in both the male and female is
regionally specific, and that the expression of this receptor May he regulated by developmental
factors. In the maie these variations iDcluded age dependent luminal factors and androgen levels,
the presence of spennatozoa, and the pattem of postnatal developmentai localizatioD of apo 1 in
the male reproductive epithelia In the female, factors such ast age t hormone levels of the estrus
41

•
•
•
cycle., receptivity and the site of fertilizatio~and the postnatal developmentallocalization ofapo J
May also influence LRP-2 loca1ization.
42

•
•
•
Plate 1: Efferent Ouets at Postnatal Days 7, 21, 49, and 90ImmUDostained with LRP-l ..~ntibody
Figure 1 (A-D): Low power light micrographs of non-ciliated and ciliated cells lining the
tubules of the efferent duets in rats of varying postnatal ages. These euboidal eeUs were
immunostained with LRP-2 antibody. A strong immunoperoxidase reaetion produet is
evident at each age as a dense band in the apical region of the non-ciliated cells
(arrowheads). Ciliated eells (stars) were devoid ofimmunostaining.
Figure A: 7 day old (X300)
Figure B: 21 day old (XJ2S)
Figure C: 49 day old (X130)
Figure 0: 90 day old (X2S0)
43
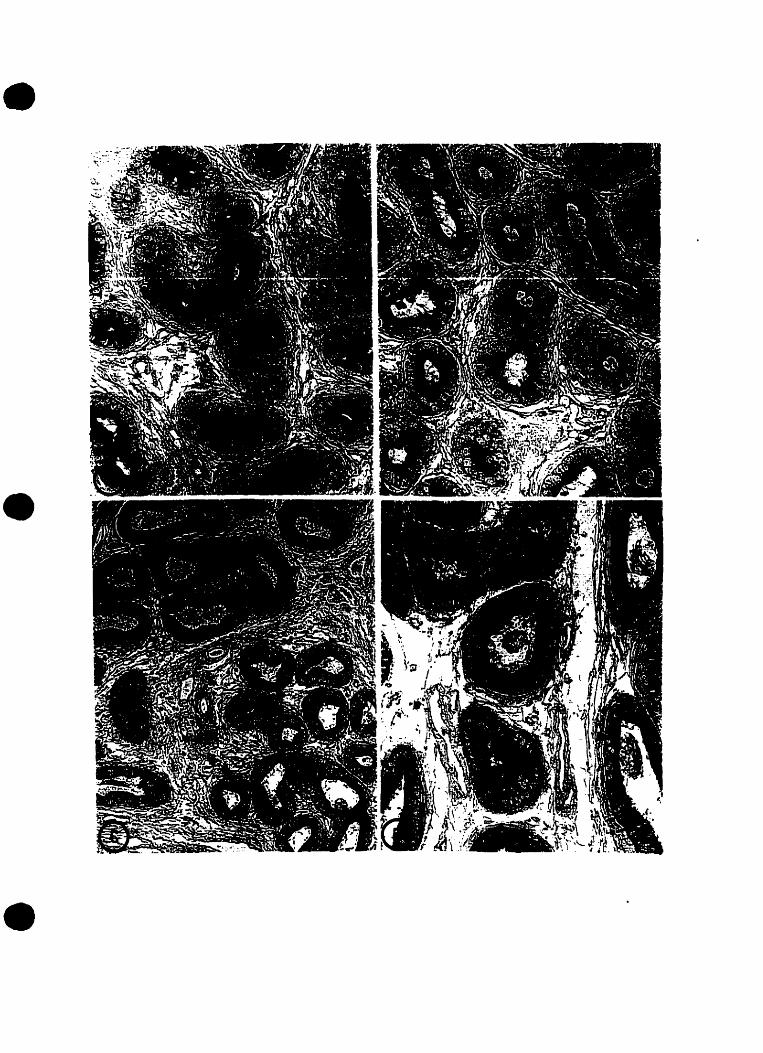
•
•
•

•
•
•
Plate 2: Proximal Initial Segment at Postnatal Days 15. 21. 39. 49. 56and 90 ImmUDostained witb LRP-2 antibody
Figures (A-F): Light micrographs revealing the complete absence of staining for LRP-2
antibody in the region of the proximal initial segment. The principal cells (p) of this
region become strueturally differentiated at day 39 (Fig. Cl. Spermatozoa (arrowheads)
were initially present in the on day 49 and become immunopositive for LRP-2 staining
represent in the proximal initial segment tubules on day 90.
Plate A~ 15 day old (X 275)
Plate B: 21 clay old (X400)
Plate C: 39 day old (X400)
Plate D: 49 day old (X440)
Plate E: S6 day old (X27S)
Plate F: 90 day old (X300)
44

•
•
•

•
•
•
Plate 3: Distal Initial Segment at PostDatai Days 15, 11. 19. 39. 56 and90 Immunostained with LRP·l Antibody
Figures (A-F): Light micrographs illustrating the pattern LRP-2 localization in the distal
initial segment. The immunoreactivity increased from day 7 and day 15 (Fig. A) where it
wu negative to a peak of apical staining (arrowheads) at clay 39 (Fig. D). From day 49
through day 90. when sperm (curved arrows) was present, the principal cells become
immunonegative for LRP..2 staining.
Figure A: 15 day old (X520)
Figure B: 21 day old (X37S)
Figure C: 29 day old (X3S0)
Figure 0: 39 day old (X440)
Figure E: S6 day old (X3S0)
Figure F: 90 day old ('075)
45


•
•
•
Plate 4: Intermediate Zone at Postnatal Days 39, 49, 56, and 90ImmUDostained with LRP-2
Figures (A-D): Sections of the intermediate zones of 39, 49, 56, and 90 day old fats
immunostained with LRP-2 antibody. This illustrates the consistent intensity of apical
immunoreaetivity (arrowheads) in the intermediate zone, from day 39 (Fig. A), when it
fast reveals its characteristic vacuoles (arrows) ta day 90 (Fig D) when spermatozoal
content (curved arrows) is high and vacules resemble giant endosomes.
Figure A: 39 clay old (X375)
Figure B: 49 day old (X400)
Figure C: 56 day old ('050)
Figure 0: 90 clay old (X32S)
46

•
•
•

•
•
•
Plate 5: Proximal Caput at PostDatai Days 15, 11, 29, 39. 56 and 90Immunostained witb LRP·2 antibody
Figures A-F: Light micrographs illustrating the pattern of localization of LRP..2 antibody
in the proximal caput. At day l5 (Fig. A) the small tubules reveal an absence of
antibody. In days 21 and 29 (Fig. B&C), the still undifferentiated principal cells
demonstrated a well-defined immunoreactive staining pattern (arrowheads) along their
luminal membrane. From day 39 to day 90 the now differentiated principal cells
displayed a distinct band ofLRP..2 expression in their apical damain (Figs D-f).
Figure A: 15 day old (X600)
Figure B: 21 day old (X375)
Figure C: 29 day old (X400)
Figure 0: 39 clay old (X400)
Figure E: S6 day old (X375)
Figure F: 90 day old (X 275)
47

•
•
•

•
•
•
Plate 6: Distal Caput at Postnatal Days 15. 21. 39, 49, !6 and 90Immunostained witb LRP·1 antibody
Figures A-F: Tubules of the distal caput region of epididymides trom postnatal rats
ranging trom 15-90 days. The epithelial cells immunostained with LRP-2 antibody
demonstrate the drastic change in the pattern of localization between this region and the
more proximal region of the caput. The distal caput revealed a moderate apical
immunoreaction (arrowheads) on day 21 and day 29, then decreased significantly by day
39 Fig. B,C). The principal cells (p) were devoid of staining at every age after day 39,
when it's bordering regions revealed intense reaetÎon produets.
Figure A: 15 day old (XS60)
Figure B: 21 day old (X37S)
Figure C: 39 clay old (JOOO)
Figure 0: 49 day old (X480)
Figure E: 56 day old (X250)
Figure F: 90 day old (X275)
48


•
•
•
Plate 7: Corpus at Postnatal Days 7, 21. 29, 39, 49, and 90ImmuDostained witb LRP·2 Antibody
Figures A..F: Light micrographs revealing the developmental pattern of localization of
LRP..2 in the corpus region of the epididymis. Moderate apical immunostaining
(arrowheads) was apparent in the 21 day old rat (Fig.B). A heavier immunoreaetion was
observed in day 29, at which time the supranuclear region of some principal cells aIso
revealed the presence of stain (open arrowheads); (Fig. Cl. Epithelial clear cells (c),
identified by their pale appearance, tirst appeared in day 39 were immunonegative (Fig.
D). The blotched pattern of LRP-2 expression in principal cells , in addition to the
unreaetivity ofclear ceIls was evident trom day 39 to clay 90 (Fig. D-F).
Figure A: 7 day old (X400)
Figure B: 21 day old (X37S)
Figure C: 29 day old (X300)
Figure 0: 39 day old (X300)
Figure E: 49 day old (X2S0)
Figure F: 90 day old (X275)
49


•
•
•
Plate 8: Cauda at PostDataI Day! 15, 21, 29, 39, 49, and 90,ImmUDostained witb LRP·l antibody
Figures A-F: Low and high magnification light micrographs of tubules in the cauda
region of postnatal rats ranging in age from 15 days to 90 days, immunolabelled by
polyclonaJ LRP-2 antibody. The pattern displayed is identical to that of the corpus region
in each respect. Figure B (21 day old) reveals a thin unifonn apical reaetion
(arrowheads). The intensity increases and becomes localized more supranuclearly in
sorne principal cells of the 29 day old (open arrows);(Fig. C). At high magnification, the
39 day old cauda revealed the unstained clear cells (c), as weil as a distinct mottled
pattern ofstaining in the principal cells (Fig. D). The same pattem is displayed in lower
magnifications of49 and 90 clay old cauda (Figs.E&F).
Figure A: 15 day old (X400)
Figure B: 21 day old (X2S0)
Figure C: 29 clay old (X37S)
Figure D: 39 day old (X480)
Figure E: 49 clay old (X27S)
Figure F: 90 day old (X27S)
so

•
•
•

•
••
Plate 9: Proximal Oviduct, Distal Oviduct. Uterus. and Uterine Glandsat Postnatal Day 19 ImmUDostained witb LRP·1 Antibody
Figures A..D: Low and high power light micrographs of the prepubertal 29 day old
femaJe rat reproductive tract. This is the earliest postnatal day in which LRP-2
immunoreactivity was revealed during the study. The proximal oviduct demonstrates a
charaeteristic granular immunostaining of the non-ciliated cells (arrowheads; Fig. A).
The distal oviduet, uterus and uterine glands ail show immunopositive reaetion of LRP-2
antibody in the apical region ofthe cells (arrows; Figs 8·0).
Figure A: Proximal Oviduet (x130)
Figure B: Distal Oviduet (x200)
Figure C: Uterus (x250)
Figure 0: Uterine Glands (x640)
51


•
•
.'
Plate 10 : Proximal Oviduct, Distal Oviduct, Uterus, and UterineGlands at Diestrus of Postnatal Day 39 Immunostained withLRP-l Antibody
Figures A-D: Light photomicrographs orthe reproductive tract ofa 39 day old female rat
in diestnls (determined by vaginal smear). The immunoreaction in the nonciliated cells
of the proximal oviduet is granular in appearance (arrows~ Fig. A). The ratio of ciliated
cells is apparently lower than in estrus. The distal oviduet shows a consistent granular
reaction along the apical surface of secretory eells (arrowheads; Fig. B). A high power
micrograph of the uterus disp(ays the amorphous bead-like appearance of the reaetion of .
LRP-2 antibody in the apical region (Fig. Cl. ln sorne cells the reaetion is shown to be
more basal surrounding the nucleus laterally (open arrow). The uterine glands revea1 a
negative immunoreaction (Fig. D).
Figure A: Proximal Oviduet (;1(400)
Figure B: Distal Oviduet (x275)
Figure C: Uterus (x540)
Figure D: Uterine Glands (x300)
52

•
•

•
•
•
Plate Il: Proximal Oviduct. Distal Oviduct. Uterus, and UterineGlands at Proestrus of Postnatal Day 39 Immunostained withLRP·2 Aatibody
Figures A-D: Light micrographs of the reproductive tract of a 39 day old female rat in
Proestrus (determined by vaginal smear). A weak apical reaetion of the nonciliat~
secretory cells of the proximal oviduct is shawn (arrowheads; Fig. A). The ciliated cells
are clearly immunonegative for LRP-2 antibody (open arrowheads; Fig. A). The staining
in the distal oviduet reveals a positive reaetion., which appears granular in nature (Fig. B).
lmmunolocalization ofLRP-2 is clearly evident along the apical surface of the uterus and
uterine glands (arrows; Figs. C&D).
Figure A: Proximal Oviduet (x375)
Figure B: Distal Oviduet (x2S0)
Figure C: Uterus (,,400)
Figure 0: Uterine Glands (x400)
53

•

•
•
•
Plate 12 : Proximal Oviduct, Distal Oviduct. Uterus. aad UterineGlands at Estrus of Postnatal Day 39 Immunostained withLRP-2 Antibody
Figures A-D: Light photomicrographs ofthe reproductive tract ofa 39 day old female rat
in estrus (determined by vaginal smear). A weak immunoreaetion is demonstrated in the
nonciliated cells of the proximal oviduet (arrowheads), while the ciliated cells rernain
devoid of immunoperoxidase $lain (open arrowheads; Fig. A). The number of ciliated
cells in this region has drastically increased in order to facilitate ovulation. A moderate
apical reaction of LRP-2 antibody is evident in the distal oviduet (open arrows; Fig. B).
The uterus and the uterine glands shawn opening into the uterine space bath display
strong immunolabelling of LRP-2 along the luminal membrane of the ta11 columnar cells
(arrows; Figs. C&D).
Figure A: Proximal Oviduet (x125)
Figure B: Distal Oviduet (x150)
Figure C: Uterus (x32S)
Figure 0: Uterine Glands (:<300)
54


.-
•
•
Plate 13 : Proximal Oviduct, Distal Oviduct. Uterus. and UterineGlands at l\letestrus or Postnatal Day 39 Immunostained\Vith LRP-2 Antibody
Figures A-D: High and medium power light micrographs of a 39 day old female rat in
metestrus (determined by vaginal smear). A weak immunoreaetion of LRP-2 antibody is
shown by the non-ciliated cells of the proximal oviduet (anowhead; Fig.A). The reaction
demonstrated in the distal ovidu~ and the uterus is apically located. (arrows; Figs.
B&C. Fig. D reveaJs a complete absence of staining for LRP-2 antibody in the uterine
glands.
Figure A: Proximal Oviduet (x540)
Figure 8: Distal Oviduet (x225)
Figure C: Uterus (x375)
Figure D: Uterine Glands (:<325)
55

•

•
•
•
Plate 14 : Proximal Oviduct, Distal Oviduct, Uterus, and UterineGlands at Diestrus of Postnatal Day 49 ImmuDostained withLRP-l Antibody
Figures A·D: Light micrographs of the reproductive tract of a 49 day old female rat in
diestrus (determined by vaginal smear). The blotched pattern of LRP-2 localization
appeared apicaJly and in the supranuclear region of nonciliated secretory cells of the
proximal oviduet (open arrowhead; Fig. A). In the distal oviduet immunoreaetivity in the
apical region appeared granular in nature (arrowheads; Fig. B). The tall columnar ceUs of
the uterus exhibited moderate staining along the luminal membrane (arrows; Fig. C)
while other ceUs aise displayed immunostaining along the lateral membrane (open
arrow). The uterine glands showed a complete absence ofperoxidase labeling (Fig. 0).
Figure A: Proximal Oviduet (:<520)
Figure B: Distal Oviduet (x300)
Figure C: Uterus (x400)
Figure 0: Uterine Glands (x27S)
56

" ;'.:""
.-..
- ,-~-
"- r,... ~,' ,'::.'Q :., ".,;~!~~;.
~'~. :, .~._.'1.'" .'",,'.... ;-:1 .:Ï1.rJar", .
, ..i~,· . •:. . ~..:'.'~.,'.:. ,•
...
,•
......... .
-....-,..
'....-......
.. -",.., - .

•
•
•
Plat! 15: Proximal Oviduct. Distal Oviduct. Uterus. and UterineGlands at Proestrus of Postnatal Day 49 ImmUDostained witbLRP·2 Antibody
Figures A-D: Light micrographs of the reproductive tract of a 49 day old female rat
during Proestrus (determined by vaginal smear). Fig. A demonstrates the scarce
appearance of peroxidase staining in the nonciliated cells of the proximal oviduet. The
secretory cells of the distal oviduet exhibit a moderate immunoreaction to LRP-2
antibody along their luminal membrane (arrowheads; Fig B). The contrast between the
immunopositive reaetion in the taU columnar cells of the uterus (open arrowheads) and
the lack of LRP-2 antibody stain in the uterine glands is shown in Figs. C&D. The
immunoperoxidase reaetion is also revealed along the basolateral aspect of severa! cells
of the endometrium (open arrows, Fig.D)
Figure A: Proximal Oviduet (x400)
Figure 8: Distal Oviduet (x27S)
Figure C: Uterus (x480)
Figure 0: Uterine Glands (x.J50)
57

•
•
• .' 1 \ .' .. ~, 1'.; t ••\ "~ ..... '- .
.. ";' ..-- :-
.....:~
-, .:'

•
•
•
Plate 16 : Proximal Oviduct, Distal Oviduct, Uterus. and UterineGlands at Estrus of Postnatal Day 49 Immunostained withLRP-Z Antibody
Figures A-D: High and medium magnification light micrographs of a 49 day oid female
rat during estrus. Fig. A is a high power contrast of the intense immunostairdng of the
nonciliated cells (arrowheads) and the immunonegative ciliated cells (open arrowheads).
The distal oviduet reveals abundant pero,odase labeling of LRP-2~ which forms a band
along the luminaJ surface of the cells (arrows; Fig.B). An intense immunoreaetion is
evident apically in the tangentia11y eut columnar cells of the endometrium (open arrows;
Fig. C). A blotched pattern of LRP·2 expression is revealed by the high power
micrograph of the uterine gland (large arrows; Fig. 0).
Figure A: Proximal Oviduet (xS20)
Figure B: Distal Oviduet (:<315)
Figure C: Uterus (x440)
Figure 0: Uterine Glands (x650)
S8

,
•
•
l -L
\, ~.: .2:~4,

e·
•
•
Plate 17 : Proximal Oviduct, Distal Oviduct, Uterus. and UterineGlands at Metestrus of Postnatal Day 49 Immunostainedwitb LRP-2 Antibody
Figures A..D: Light photomicrographs illustraring the pattern of localization throughout
the 49 day old female reproductive tract during metestrus. The proximal oviduct displays
a spony immunoreaction, which is darkest in the folded regions (arrows; Fig. A). A light
granular immunoreaction is shown aJong the apical aspect of the distal oviduet
(arrowheads; Fig. B). The uterus bas become devoid of staining for LRP..2 antibody.
Large vacuoles which are cbaracteristic of metestrus are aise shown (open arrowheads;
Fig. Cl. The uterine glands, which contain secretory produets (stars) are immunonegative
for LRP-2 antibody (Fig. D).
Figure A: Proximal Oviduet (x375)
Figure B: Distal Oviduet (x275)
Figure C: Uterus (x4oo)
Figure 0: Uterine Glands (voo)
59

•
•
•

•
•
•
Plate 18 : ProlÎmal Oviduct. Distal Oviduct. Uterus, and UterineGlands at Diestrus of Postnatal Day 56 Immunostaiaed withLRP·2 Antibody
Figures A-D (and iosert): Light micrographs illustratiog the pattern of localizatioo of
LRP·2 throughout the reproductive tract of a 56 day female rat duriog diestrus. The
proximal oviduet displays a spotty immunoperoxidase pattern in the foidee! regions
(arrows; Fig. A). The distal oviduet reveaJed an immunopositive reaetion along its apical
membrane as well as in the regions undergoing repair following metestrus (arrowheads
Fig. B). The transition between the well-stained shorter eells of the distal oviduet (large
arrows) and the unstained tall columnar cells of the uterus (open arrowheads) where
vacuoles are still present is shown in Fig. C. The insen (star) is a Iow power micrograph
of the same regio~ which clearly illustrates the transition from the distal oviduet to the
uterus. The uterine gJand tilled with mucoid fluids is immunonegative for LRP·2
labeling.
Figure A: Proximal Oviduet (x400)
Figure B: Distal Oviduet (x375)
Figure C: Uterus (xJ75)
Figure D: Uterine Glands (xS20)
60

•
•
•' ...\.. .
\
--~

•
•
•
Plate 19: Proximal Oviduct, Distal Oviduct. Uterus. and UterineGlands at Proestrus of Postnatal Day 56 ImmUDostained withLRP-l Antibody
Figures A..D: High and medium power magnification micrographs demonstrating the
immunoreactivity to LRP·2 antibody in the reproductive tract of a 56 day old female rat
during proestrus. Peroxidase labeling is present in smalt vesicle-like packages along the
luminal membrane of secretory non-ciliated cells (Fig. A). Figure B is a high power
image revea1ing the difference in merpholegy and immunoreactivity te LRP·2 between
the short, uneven secretory cells of the distal oviduet and the taIl columnar cells of the
uterus. The distal oviduet shows a modest reaetion of LRP..2 immuno..labeling
(arrowheads) while the narrow columnar cells of the uterus are devoid of staining (open
arrowheads; Fig. Cl. The uterine glands a1so demonstrate an absence of immunostaining
of LRP·2 antibody.
Figure A: Proximal Oviduet (xS40)
Figure B: Distal Oviduet (:<540)
Figure C: Uterus (:<.375)
Figure 0: Uterine Gland (x3 50)
61

•
•
•

•
•
•
Plate 10: Proximal Oviduct, Distal Oviduct, Uterus, andUterine Glands at Estrus o( Postnatal Day 56 Immunostainedwith LRP·2 Antibody
Figures A..D: Medium power magnification light micrographs of different parts of the
female rat reproductive tract at postnatal day 56 during estrus. The pattern of localization
of LRP..2 antibody was consistent throughout adulthood during this stage. A moderate
immunoreaction was displayed in the folds of the proximal oviduet (arrowheads; Fig. A).
The distal oviduet showed a light, apical pattern of peroxidase staining the secretory cells
(open arrowheads; Fig. 8). The uterus and uterine glands remained immunopositive
throughout estrus at each age after day 29 (arrows; Figs C&D).
Figure A: Proximal Oviduet (xJ25)
Figure B: Distal Oviduet (x375)
Figure C: Uterus (x440)
Figure D: Uterine Glands (xJ75)
62

•
•
•

•
•
•
Plate 21: Proximal Oviduc~ Distal Oviduc~ Uterus. and UterineGlands at Metestrus of Postnatal Day 56 Immunostainedwitb LRP·l Antibody
Figures A-D: Medium power light micrographs of regions of the 56 day old female rat
reproductive tract during metestrus. A weak granular immunoreaetion is shown in the
falds of the proximal oviduet (arrowheads; Fig. A). A strong reaetion is revea1ed.. along
the apical membrane in the columnar cells, in the distal oviduet (arrows; Fig. B). The tall
columnar cells of the uterus wim their charaeteristic large vacuoles are immunonegative
(open arrowheads; Fig. Cl. Similarly the uterine glands, which contain secretory produets
(star), are completely devoid of immunostaining for LRP·2 antibody (Fig. D).
Figure A: Proximal Oviduet (xISO)
Figure B: Distal Oviduet (:<125)
Figure C: Uterus (x375)
Figure 0: Uterine Glands (x32S)
63

•
•--.... """'iI'- ....
~•..$~.-.~ .. ' ..

•
•
•
Original contributions to this thesis include:
1) Discovery of Gp 330 (Iater called LRP-2~ megalin) as the proximal tubule
glycoprotein involved in Heymann Nephritis (Kerjascki and Farquhar 1982).
2) Evidence proving the presence of LRP-2 on apical surfaces of epithelia specifically
within the male and female reproductive (Zheng et al. 1994).
3) Identification of LRP-2 as an endocytic receptor for the intemalization and
degradation ofapo J/clusterinl SGP-2 (Kounnas et al. 1995).
4) Demonstration that RAP inhibits binding of ligands to LRP-2 in the male
reproductive tract (Morales et al. 1996).
64

•
•
•
Abou..HaiI~ A. Androgen regulation of molecular farms of beta-D-glucuronidase in themouse epididymis: comparison with liver and kidney. 1. Androl. 1996; 17(3): 194-207.
Adamali. H.I., and Hermoy L. Apical and narrow ceUs are distinct eeU types differing intheir structure, distribution and funetions in the adult rat epididymis. 1. Androl. y 1996;17(3): 208-222.
Advis, 1.P.. Andrews, W.W., and Ojed~ S.R. Changes in ovarian steroidal andprostaglandin E responsiveness ta gonadotropins during the onset ofpubeny in thefemale rat. Endocrinology, 1979; 104(3): 653-659.
Aronowy BJ. y Lunel S.O., Brown, T.L., Harmony, lA.K. (1993). Apolipoprotein JExpression at Fluid-Tissue Interfaces: Potenùal Role in Barrier Cytoprotection. Proc.Natl. Acad Sei. USA. 90: 725-729.
Bedford, lM. Effects of dUel tigation on the fertilizing ability of spermatozoa tramdifferent regions ofthe rabbit epididymis. 1. Exp. Zool.• 1967; 166: 271-282.
Beisiegel, U., Weber, W., and Bengtsson-Olivecro", G. Proc. Nat/. Acad. Sc;. USA.,1991; 88: 8342·8346.
Beisiegel, U., Weber, W., Ihrke" G., Kea. 1., and Stanley, K.K. The LDL receptorrelated protein 1 LRP is an apolipoprotein E.binding protein. Nature., 1989; 341: 162164.
Biesmesderfer, D., Dekan. G., Aronson, P.S., Farquhar, M.G. (1992). AssemblyofDistinctive Coated Pit and Microvillar Microdomains in the Renal Brush Border.American Journal ofPhysi%gy, 262 (1 pt 2): F55-67.
Bloom W, Fawcen Dw. A Textbook ofHistololY. Toronto: W.B. SaundersCompany; 1968: 710- 712
Brooks DE. Androgen-Regulated EpididymaI Secretory proteins Associated with PostTesticular Sperm Development. Annals ofthe New York Academy ofSciences1987:513: 179-194.
Christensen, E.I., Gliemann, 1., and Moestrup, S.K. Renal tubule gp330 is a calciumreceptor for endocytic uptake ofprotein. 1. Histochem. Cytoche",., 1992; 40: 14811490.
Christensen, E.I., Nielsen, S., Moestrup, S.K., Borre, C., Maunsbacb, A.B., de Heer, E.,Roneo. P., Hammond., T.G." Verroust, P. (1995). Segmental Distribution orthe
65

•
•
•
Endocytosis Receptor gp330 in Renal Proximal Tubules. European Journal afCe/lBi%gy. 66 (-1): 349-364.
Cohen. lP., HotTer, A.P., Rose~ S. Carbonic anhydrase localization in the epididymisand testis of the rat. Biol. Reprod, 1976; 14: 339·346.
CoUard, M. W. & Griswol<L M.D. (1987). Biosynthesis and ~(olecular Cloning ofSulfated Glycoprotein-2 Secreted by Rat Sertoli CeUs. Biochemistry. 19: 5380-5389.
Cornwall, G.A., Orgebin-Crist, M-C.~ and Hanc, S.R. The CRES gene: a unique testis·regulated related to the cystatin family is highly restrieted in ilS expression to theproximal region ofthe mouse epididymis. /vIol. Endocrinology, 1992; 6(10): 1653-1664.
Cornwall, G.A., and HaM., S.R. Specialized gene expression in the epididymis: aminireview. ~ Alldrol., 1995; 16(5): 379-383.
Cummings., A.M.., and Yochim., J.M. Ditrerentiation ofthe uterus in preparation forgestation: a model for the action of progesterone. J. theor. Biol, 1994; 106: 353..374.
Cyr, D.G.., Robaire, B., and Hermo, L. Structure and turnover ofjunetional complexesbetween principal cells of the rat epididymis. Micro. Res. Teck, 1995; 30(1):54-66.
Cyr, O.G.., Hermo, L., and Robaire, B. Developmental changes in epithelial cadherinmessenger ribonucleic acid and immunocytochemicallocalization ofepithelial acdherinduring postnatal epididymal development in the rat. Endocrinology, 1993; 132: I11S1124.
de Silva HV,Harmony JAl(, Stuart WD, Gil C~ Robbins 1. Apolipoprotein 1: Structureand Tissue Distribution. Biochemisrry 1990:29: 5380-5389
de Silva HV, Stuart WD, Park WB, Mao SJT, Gil Cm, Wettereau JR. Busch SI, Hannony1AK. Purification and Characterization ofApolipoprotein 1. The Journal ofBiologicalChemistry, 1990.
Eddy AA, Fritz IB. Localization ofclusterin in the epimembranous deposits of passiveHeymann nephritis Antigenic Camplex. Annals ofthe New York Academy ofSciences1994;737: 96-113
Farquhar MG, Kerjascbki D, Lundstram M., Orlando RA. LRP·2 and RAP: TheHeymann Nephritis Antigenic Complex. Annois ofthe New York Academy ofSciences1994;737: 96-113
Flinckinger, C.J. Synthesis and secretion ofglycoprotein by the epididymal epithelium.J. A"droL t 1983; 4: 157-161.
66

•
•
•
Flinckinger, C.1. Radioautographic analysis ofthe secretory pathway for glycoproteins inprincipal cells of the mouse epididymis exposed to (3H) fucose. Bio. Reprod.. 1985~
32(2) : 377-389.
FraiIe.. B., ~[arti~ R., De ~liguel, M.P., Arenas, M.L.. Bethencou~ F.R... Peinado, f.,Paniagua.. R., and Santamaria.. L. Light and Electron microscopie immunohistochemicallocalization ofprotein gene produet 9.5 and ubiquitin immunoreaetivities in the humanepididymis and vas deferens. Bio. Reprod. 1996; 55(2): 291-297.
Franke~ AL, and Eik-Nes, K.B. Metabolism ofsteroids in the rabbit epididymis.Endocrinology. 1970; 87: 646-652.
fritz IB, Murphy B. Clusterin: lnsights ioto a Multifunetional Protein. TEM 1993; 4: 4145
Fri~ I.B., Burdzy, K., SetchelI, B., Blaschu~ O. (1983). Ram Rete Testis FIuid Containsa Protein (Clusterin) Which Influences Cell-Cell Interactions ln Vitro. Bi%gy ofReproduction, 28: 1173.. 1188.
Garr~ S.H., Garrett, lE., and Douglass, 1. In situ histochemical analysis ofregionspecifie gene expression in the adult rat epididymis. Atfol. Reprod. Dev., 1991; 30(1): 117.
Ghyselinck N.B., Dufaure, I., Lareyre, 1.l, Rigaudiere, N., Mattei, M.G., and Dufaure..J.P. Structural organization and regulation orthe gene for the androgen-dependentglutathione peroxidase-like protein specifie to the mouse epididymis. Mol.Endocrinology. 1993; 7(2): 258-72.
Gitlin, G. (1971). The Epithelium ofthe Vaginal (ntroitus in the Rat. The AnatomicalRecord 169(2): 201..206.
Gliem~ 1., Nykjaer, A., Peterse~ C.M., Jorggensen. K.E., Nielsen.. M., Andrease~P.A., Christense~ E.L, Lookene, A, Olïvecrona, G., Moestrup, S.K. (1994). TheMultiligand a2-Macroglobulin ReceptorlLow Density Lipoprotein Receptor RelatedProtein (a2MRILRP): Binding and Endocytosis ofFluid Phase and MembraneAssociated Ligands. Ann. NY. Acad. Sci.• 737: 20-38.
Griswold, M.D., Robens, K., Bishop, P. (1986). Purification and Charaeterization ofaSulfated Glycoprotein Secreted By Senoli CeHs. Biochemislry, 2S: 7265-7270.
Gude, W.D., Cosgrove, G.E., Hirsc~ G.P. Histologie,l Atlas ofthe Laboratol)' Mouse.New York: Plenum Press; 1982: 13-15 &. 60-63.
Hales, B.F., Hachey, C., and Robaire, B. The presence and longitudinal distribution ofthe glutathione S-transferases in rat epididymis and vas deferens. Biache",_ 1., 1980;189: 135-142.
67

• Hall, le., Pere~ F.M., and Kochins, J.G. Quantification and localization ofN-acetylbeta-D-hexosaminidase in the adult rat testis and epididymis. Bio. Reprod. 1996; 54(4):914-929.Hamilton.. D.W. Structure and funetion of the epithelium lining the ductuli etTerentes,ductuli epididymis and ductus deferens in the rat. In: Handbook ofPhysiology, Section7, Vol. V. Astwood E.B. and Greep R.O., Washington D.C" American PhysiologicalSociety, L975~ pp. 307-317.
Hebei, R., Stromberg, M.W. Anatomy orthe LaboratolY Rat. Baltimore: The Williamsand Wilkins Company; 1976: 76-78,
Hemeida.. N.A., Sac~ S.O., and McEntee, K. Duetuli efferents in the epididymis ofboar,goal, r~ bul~ and stallion. Alvl 1. Veto Res., 1978; 39: 1892-1900.
Herme, L" BaJin, K., and Robaire, B. Structural differentiation ofthe epithelial cells ofthe testicular excurrent duel system ofrats during postnatal development. Anat. Ree.,1992a; 233: 205-228.
•
•
Herme, L., Clermont, Y., and Morales, C.R. Fluid-phase and adsorptive endocytosis inciliated cells of the rat duetuli efferentes. AnaL Rec' t J985; 211: 285-294.
Hermo, L., and Morales.. C.R. Endocytosis in nonciliated epithelial cells ofthe duetuliefferentes in the rat. Am. J. AnaL. /984,' 171: 59-74.
Hermo, L. t Oka, R., and Morales, C.R. Secretion and Endocytosis in the malereproductive tract: a role in spenn maturation. lnt. Rev. eyta/ogy, 1994; 154: 104-109.
Hermo, L., BariD, K., and Oko, R. Developmental expression of immobilin in the ratepididymis. Anal. Ree., 1994b; 240: 86-103.
Herme. L., Spier, W., and Nadler, N.J. Role ofapical tubules in endocytosis innonciliated ceIls of the duetuli etTerentes ofthe rat: A kinetic analysis. Am. J. Anal.,J988a; 182: 107-119.
Herme, L., Wright, 1., Oko, R., and Morales, C.R. Role ofepithelial cells ofthe maleexcurrent dUel system ofthe rat in the endocytosis or secretion ofsulfated glycoprotein-2(Cluterin), Biol. Reprod., 1991; 44: 1113-1131.
Hermo, L. Structural features and funetions ofprincipal cells of the intermediatc zone ofthe epididymis ofadult rats. AnaL Rec., 1995; 242: 515-530.
Hermo.. L., Morales, C., and Oka, R. Immunocytochemicallocalization ofsulfatedglycoprotein-l (SGP-l) and identification of it'5 transcripts in epithelial cells oftheextratesticular dUel system ofthe rat. AnaL Rec., 1992c; 232: 401422.
68

•
•
•
Hermo. L., Adamali. R.L, Mahura~ D.• Grave~ R.A., and Trasler. J.M. BHexosaminidase gene expression and enzyme immunolocalization in the rat testis andepididymis. Mol. Reprod. Develop.• 1997; 46: 1-16
Hermo. L.• Oko, R., and Robaire, B. Epithelial cells of the epididymis show regionalvariations with respect to the secretion or endocytosis of immobilin as revealed by lightand electron microscope immunocytochemistry. Anal. Rec.. l992b; 232: 202-220.
Hermo L, Structural Features and Funetions ofPrincipal CeHs ofthe lntermediate Zone.Analomical Record 1995; 242: 545-550
Hermo L, Barin R, Robaire B. Structural Differentiation ofEpithelial Cells of theTesticular Excurrent System ofRats during Postnatal Development. Anatomical Record1992; 233: 205-228
Hermo L. Oko ~ Morales C. Secretion and Endocytosis in the Male Reproductive Tract:A Role in Sperm Maturation. International Review o/Cyr%gy 1994 Vol. 154;
Hermo L, Barin ~ Oka R. Developmental Expression of Sulfated Glycoprotein-2 in theEpididymis ofthe Rat. The Anatomical Record 1994; 240: 327-344
Hermo L., Wright J, Oko ~ Morales Cil. Role ofEpithelial Cells of the Male ExcurrentDuct System ofthe Rat in the Endocytosis or Secretion of Sulfated Glycoprotein-2(Clusterin). Biology ofReproduction 1991;44:1113-1131
H~ 1., Haman~ S., Myklebo~ O., Stanley, K.K. Surface location and high affinity forcalcium ofa 500-kd liver membrane protein closely related to the LDL..receptor sug8est aphysiological role as a lipoprotein receptor. Emba. J., 1988; 7: 41194127.
Hiesberger, T., Hodits, R., Ullrich, R., Exner, M. Receptor associated protein andmembers of the low density protein receptor family share a common epitope. J. Biol.Chem.. 1996; 271(46): 28792-28797.
Hinton BT. What does the epididymis do 50 and how docs it do it? Handbook 0/AnJro/ogy 1995: 18-20
Hinto~ B.T., and Setchell, B.P. Concentration and uptake ofcamitine in the ratepididymis: A micropuneture study. In: Camitine BiQsynthesis, Metabolisme andFunetion. Frankel, R.A., and McGarry, J.O., Eds., New York, 1980; pp.237-249.
Hinto~ B.T., and Palladino, M.A. Epididymal epithelium: its contribution to theformation ofa luminal fluid microenviroment. Micro. Res. Teck, 1995; 30(1): 67-81.
Hotter, A.P., and Hintol\ B.T. Morphological evidence for a blood-epididymis banierand the effects ofgossypol on it's integrity. Biol. Reprod. 1984; 30: 991-1004.
69

• Hoffer~ A.P., Hamilton., D.W., and Fawcett~ D.W. The ultrastrueture ofprincipal cellsand intraepithelial leukocytes in the initial segment of the rat epididymis. Anal. Rec.,1973; 175: 169-202.
HoIland, M.K., and Orgebin-eri~ M.C. Charaeterization and hormonal regulation ofprotein synthesis by the murine epididymis. Bio. Reprod., 1988; 38(2): 487-496.[gdo~ S.A, Hermo, L., Rosenthal, A., and Morales, C.R. Nonciliated cells of the ratefferent duets endocytose testicular sulfated glycoprotein-l (SGP-1) and synthesize SGP1 derived saposins. Anar. Ree., 1993; 235: 411-424.
Huang, 1., Makker, S.P. (1995). Role ofReceptor-Associated 39/45 kOa Protein inActive Heymann Nephritis. Kidney International, -17 (2): 432-441.
19doura, S.A, Morales, C.R., Henno, L. DifferentiaI expression ofcathepsins B and D intestis and epididymis ofadult rats. 1. Histochem. Cytochem., 1995; 43: 545-557.
Hia, K.Y., and Hess, R.A Structure and funetion orthe duetuli efferents: A Review.Micro. Res. Teck, 199-1; 29(6): 432-467.
•
•
James, R.W., Hochstrasser, D., Borghini, P., Martin, B. Arteriose/er. Thromb., 1991; Il:645-652.
leMe, O.• and Tschopp, J. Clusterin: the intriguiog guises ofa widely expressedglycoprotein. l1BS., 1992; 17: 154-159.
Jensen, P.H., Moestnlp, S.K., and Gliemann, 1. Purification ofthe human placental a2macroglobulin receptor. FEBS. Leu., 1989; 255: 275-280.
lunquiera LC, Cameiro 1, KeUey RD. Basic HistolQ&,Y. 7ed. NorwaJk: Appleton andLange;1992: 15-24&423-440
Kerjaschki D. Farquhar MG. The pathogenic antigen ofHeymann Nephritis is amembrane glycoprotein of the renal proximal tubule brush border. Cell Bi%gy1982;79:5557-5561
Knudse~ J.F., Castoit A, and Mahesh, V.B. Correlation of serum gonadotropins,ovarian and uterine histology in immature and prepubenal rats. Anat. Ree., 180: 497508.
Kounnas, M.Z., Argraves. W., Stricldan~ O. The 39-kDa Receptor associated proteininteraets with two members ofthe low density lipoprotein rec:eptor family, 12macroglobulin receptor and glycoprotein 330. J. Biol. Chem., 1992; 267(29): 2116221166.
70

•
•
•
KOUMas Ml, Chappell David A., Strickland Dudley K., Argraves Scott W.Glycoprotein 330, a Member ofthe Law Oensity Lipoprotein Receptor Family. BindsLipoprotein Lipase in Vitro. The Journal ofBi%gical Chemistry Vol. 268No.19;14177-14180
Kounnas MZ. Argraves WS, Strickland DK. The 39-kDa Receptor-associated Protein[nteraets with Two Members ofthe Law Density Lipoprotein Receptor Family, alMacroglobulin Receptor and Glycoprotein 330. The Journal ofBi%gica/ Chemistry1992;267:21162-21166
Kounnas, M.Z.. Loukinov~ E.B.. Stefansson. 5... Harmony, J.AK.. Brewer. B.H..StricJdand, O.K., Argraves, W.S. (1995). Identification ofGlycaprotein 330 as anEndocytic Receptor for Apolipoprotein J/Clusterin. The Journal ofBi%gical Chemistry,270: 1-6.
Kounnas Ml, Stefansson S, Loukinova E, Argraves KM, Strickland DK. Argraves WS.An Overview of the Structure and Funetion ofGlycoprotein 330, a Receptor Related tothe al- Macroglobulin Receptor. Annals ofthe New York Academy ofSciences1994;737: 114-123
Kowa!, R., Herz, 1., Goldstein J.l., Esser, V., and Brown, M.S. Law density lipoproteinreceptor-related protein Mediates uptake ofcholesterol esters derived tram apoprotein E...enriched lipoproteins. Proc. Natl. Acad. Sei. USA., 1989; 86: 5810-5814.
Krïeger, M., and Herz, 1. Structures and funetions ofmultiligand lipoprotein receptors:macrophage scavenger receptors and LDL receptor-related protein (LRP). Ann. Rev.Biochem., 1994; 63: 601-637.
LeI, O.A... Petrusz, P., and French., F.S. Purification and localization ofacidicepididymal glycoprotein (AEG): a sperm coating protein secreted by the rat epididymis.lnl. J. Androl., 1978; (Suppl. 2): 592-607.
Luca, M.E., Deeler., A.M., H08endoo~ P.C., Daha, M.R., Galceran. M., van Es, L.A., deHeer, E. (1995). Glomerulopathy Induced by a Single Monoclonal Autoantibody againstgp330. Nephrology, Dia/ysis, Transplantation, /0(4): 490-496.
Marquis, N.R., and Fritz, l.B. Etrects oftestosterone on the distribution ofcarnitine..acetylcamitine, and camitine acetyi-transferase in tissues ofthe reproductive system ofthe male rat. J. Biol Chem.• 1965; 240: 2197-2200.
Moestrup S~ Chrïstensen El., Nielsen S, Jorgenson IŒ, Bjom SE, Roigaard fi,Gliemann J. Binding and Endocytosis of Proteins Mediated by Epithelial LRP-2. Annoisofthe New York Academy ofSciences 1994;737:124-137
71

•
•
•
Moestrup, S. K., Chriestensen, E.I., Nielsen, S'7 Jorgensen, K.E., Bjo~ S.E., RoigaarcLH., and Glieman~ 1. Binding and endocytosis ofproteins mediated by epithelial gp330.,Ann. NY. Acad Sci., 1994; 737: 20-38.
Moestrup S~ : Biochimiea et Biophysica Acta LL97 (1994) L97·213
Moestrup, S.K... Nielsen., S., Andreasen, P.A., Jorgsen, K.E. J. Biol. Chem.. 1993; 268:16564-16570.
Moestrup, S.K., and Gliemann. 1. Analysis oflygand recognition by the purified al·macrogJobulin receptor (Iow density lipoprotein receptor·related protein): evidence thathigh affinity ofa2-maeroglobulin-proteinase complex is achieved by binding to adjacentreceptors. 1. Biol Chem. 266: L4011-14017.
Moore HD~ Hartman ID, Smith Ca. The Role of the Epididymal Epithelium in theDevelopment ofMammalian Sperm-Fertilizing Capacity in Vivo and in Vitro. Annois ofthe New York Academy ofSciences 1987; 513: 195-203
Moore, H.D., and Bedford, J.~f. The differential absorptive aetivity ofepitbelial cells ofthe rat epididymis before and after castration. AnaL Rec., 1979; 193: 313-j28.
Morales, C.R., Igdoura, S.A, Wosu, U.A, Boman, 1., Argraves, W.S. (1996). LowDensity Lipoprotein Reœptor-Related Protein-2 Expression in Efferent Duet andEpididymal Epithelia: Evidence for it's Role in Endocytosis od ApolipoproteinJ/Clusterin. Bi%gy ofReproduction (in press).
Nonogaki.. T., Noda.. Y., Marimoto, K., Shiotani, M., Mori, T., Matsuda, T., and Yoshida,O. Localization of CuZu-superoxide dismutase in the human male genital organs.Human Reprod.. 1994; 50: 502-509.
Orgebin-Crist, M.C. Sperm maturation in rabbit epididymis. JVature, 1967; 216: 816818.
Orgebin-Crist, M.C. Studies on the funetion ofthe epididymis. Bio/. Reprod., 1969; 1:155-175.
Orgebin-Crist, M.C. Androgens and epididymal funetioD. In: Pharmacol0iYe BiololY.and Clinical Application of Androsens. Shalender Shasin et al., Eds., Wiley-Liss, Ine.,1996; pp.27-38.
Orgebin-Crist, M.C., Danzo, 8.1., and Davies, 1. Endocrine control of the developementand maintenance ofsperm fertilizing ability in the epididymis. In: Handbook ofPhysioIQi,Y. Section 7, Vol V. Astwood E.B. and Greep R.O.~ Eds., Washington D.C.,American Physiological Society, 1975; pp.319-338.
72

•
•
•
Orlando, R.A, Kerjaschki, D., Kurihara., H., Biesmesderfer, D., and Farquhar, M.G.• ••••••••••_•••. Proc. LVat/. Acad Sei. USA., 1992; 89: 6698-6702.
Orlando, R.A., Exner, M.'t Czekay, R.P., Yamazaki, H. Identification ofthe secondcluster of ligand-binding repeats in megalin as a site for receptor-ligand interactions.Natl. Acad Sei. USA.. 1997; 4: 2368-2373.
Papp, S., Robaire, B., and Hermo, L. DevelopmentaJ expression of the glutathione Stransferase Y0 subunit in the rat epididymis using Hght microscopeimmunocytochemistry. Anal. Rec., 1994; 240: 345-357.
Parker, C.R., and Mahes~ V.B. Hormonal events surrounding the naturai onset ofpubeny in female rats. Biol. Reprod.. 1976; 14:347-353.
Pelletier, R.M. Freeze.fraeture study ofcell junetions in the epididymis and vas deferensafa seasonal breeder: the mink (muestela vison). Micro. Res. Tech.. 1995; 30(1): 37·53.
Pelliniemi, L.J., Dym, M., Gunsalus, G.L., Mutso, N.A., Bar~ C.W., and Fawcett,D.W. Immunocytochemicallocalization ofandrogen-binding protein in the malereproductive tract. Endocrinology, 1981; 108: 925-931.
Pietromonaco, S., KerjasehkL D., Binder, S., Ullrich, R., and Farquhar, M.G. Molecularcloning ofa cDNA encoding a major pathogenic domain ofthe Heymann nephritisantigen gp330. Proc. Natl. Acad Sci. USA., 1990; 87: 1811-1815.
Raja~ N., Sung, W.K., and Goodman, O.S. Localization ofcellular retinal bindingprotein maNA in rat testis and epididymis and ilS stage dependent expression during thecycle of the seminiferous epithelium. Bio. Reprod.. 1990; 43(5): 835-842.
Rambourg, A., Clermont., Y., Hermo, L., and Segretain.. O. Tridimensional architectureofthe Golgi apparatus ofnonciliated epitheliaJ cells ofthe duetuli efferentes in the rat:An electron microscope stereoscopie study. BioL Cell, 1987; 60: 103-116.
Rankin, T.L., Ong, D.E., and Orgebin-Crist, M.C. The 18-kDa mouse epididymal protein(MEP 10) binds retinaic acid. Bio. Reprod, 1992; 46(5): 767-771.
Raychowdhury ~ Niles JL, McCluskey RL Smith lA. Autoimmune Target in HeymannNephritis (5 a Glycoprotein with Homology ta the LDL Receptor. Science 1989;244:1163-1165
Rei~ B.L., and Clelan~ K.W. The structure and tùnetion ofthe epididymis. 1: Thehistology ofthe rat epididymis. Aust. J.looL. 1957; 5: 223-246.
Robaire~ B.~ and Henno, L. Efferent dudS, epididymis and vas deferens: Structure,funetions and their replation. In: The PhysiolQBY ofRe»roduetion. Knobil, E., andNeil~ 1. et al., Eds., Raven Press, New York, 1988; pp.999-10S0.
73

• Robaire~ B., and Viger, R.S. Regulation of epididymal epithelial cells funetions. Bio/.Reprod.. 1995; 52: 226-236.
Roberts KP. What are the components of the male reproductive system? Handbook ofAndrology 1995: 1-4
Rodman 1S, Seidman L. Farquhar MS. The Membrane Composition ofCoated Pits,Microvilli, Endosomes, and Lysosomes Is Distinctive in the Rat Kidney Proximal TubuleCell. The Journal ofCell Bi%gy 1986;102:77-87
Rosenberg NIE, Dvergsten J, Rotter Re. Clusterin: An enigmatic protein recruited bydiverse stimuli. J Lab Clin Med 1993: 121 :205-214
Ross, M.H., Romrell, L.J. Histoloa.v: A Text and Atlas, 2nd ed. Baltimore: Williams andWilkins; 1989: 684-695.
Saboori.. A.M. Nerve growth factor: biosynthetic produets of the mouse salivary glands.Charaeterization ofstable high molecular weight and 32,000-dalton nerve growth factors.Biochemistry. 1986; 25: 5565-5571.
Saito, A., Pietromonaco, S., Kwor-Chie~ A., and Farkuhar, M. Complete cloning andsequencing of rat gp3301"megalin", a distinctive member ofthe low density lipoprotein
• receptor gene family. Proc. Natl.. Acad Sei. USA.• 1994; 91: 9725-9729.
SiîterL J.E., Enrsu~ K.M.<t Moore, A., Hamilton, D.W. Identification ofosteopontin(QPN) mRNA and protein in the rat testis and epididymis, and on sperm. Mol. ReprodDev.• 1995; 40: 16-28.
Spomitz, U.M., Rinderknecht. B.P., Edelmann, A., Scheidegger, B., Cairoli, F. (1994).Ultrastrueture as a Buis for Dating ofRat Endometrium. Anatomical Record, 238(2):163-176.
Stefansso~ 5., ChappeIl, D., Argraves, K., and Strickland., D. Glycoprotein 330ILowdensity lipoprotein receptor related protein -2 mediates endocytosis oflow densitylipoproteins via interaction with apolipoprotein B100. 1. Biol.. Chem.• 1995; 270(33):19417-19421.
Stuart, W.O., Kro~ B., Jenkins, SB., and Harmony, I.A. Biochemistry.t 1992; 31:8553-8559.
•Stoffe1<t M.R. and Friess, A.E. Morphological characteristics ofboar efferent duetulesand epididymal duct. Micro.. Res. Tech.. , 1994; 29(6): 411-431.
Sun, E.L., and Flickinger, C.I. Morphological characteristics ofcells with apical nucleiin the initial segment ofthe adult rat epididymis. Anat. Ree., 1980: 196: 285-293•
74

•
•
•
Sun.. E.L., and Flickinger, C.J. Dt: dopment ofcell types and of regional differences inthe postnatal rat epididymis. Am. J. Allal.. 1979; 154: 27..55.
Sylvester SR, Morales C, Oka ~ Griswold~. Localization of Sulfated Glycoprotein-2(Clusterin) on Spermatazoa and in the Reproductive Tract of the Male Rat. Bi%gy ofReproduction 1991 ;45: 195-207
Syntin.. P., Dacheux., F., Dru~ X., Gatti, J.L., Okamura, N., and Daeheux., J.L.Charaeterization and identification ofproteins secreted in the various regians ofadultboar epididymis. Bio. Reprod., 1996; 55(5): 956-974.
Turner TT. Spermatazoa Are Exposed to a Complex Microenvironment as they Traversethe Epidiymis Annois ofthe New York Academy ofSciences 1987;513: 195..203
Turner, T.T. Spermatozoa are exposed to a complex microenviroment as they traversethe epididymis. Annal NY Acad. Sc;., 1991; 637: 364-388.
Ve~ IP., Hermo, L., and Robaire, B. ImmunocytochernicallocaJization ofglutathioneS-transferase y 0 subunit in the rat testis and epidiymis. 1. Andro/., 1994; 15: 415-434.
Veri, IP., Henno, L., and Robaire, B. lmmunocytochemieallocaJization of the Yfsubunit ofglutathione S-transferase P shows regional variation in the staining ofepithelial cells of the testis, efferent duets, and epididymis ofthe male rat. 1. Androl.,1993; 14: 23-44.
Vierula, M.E., Rankin, T.L., and Orgebin-Crist. Electron microscopieimmunolocalization ofthe 18 and 29 kilodalton secretory proteins in the mouseepididymis: evidence for differential uptake by clear cells. Micros. Res. Tech., 1995;30(1): 24-36.
Viger, R.S., and Robaire, B. Differential regulation ofsteady state 4-ene steroid Sa..reduetase messenger ribonucleic 8Cid levels along the rat epididymis. Endocrinology,1991; 128: 2401-2414.
Viger, R.S., and Robaire, B. Immunocytochemicallocalization of4-ene steroid 5 areduetase type 1 along the rat epididymis during postnatal development. Endocrinology,1995; 136(12): 5409-5415.
WaJker, lE., Iones, R., Moore, A., Hamilton, D.W., Han, L. Analysis of. majorandrogen-regulated eDNA clones ftom the rat epididymis. NIai. Cell Endocrinology,1990; 74(1): 61-68.
Wan& GR. (1923). Relation BetWeen Spontaneous Aetivity and the Oestrus Cycle in theWhite Rat. Comparative Psych%gy Monographs, 2: 1-27.
75

•
•
•
Willnow, T.E., Goldstei~ K., Orth, M.S., Brown, and Herz, J.. J. Biol Chem.• 1992;267:26172-26180.
Willnow. T., Hilpert, 1., Armstron& S., Rohlmann, A. Defective forebrain developementin mice lacking gp330/megalin. Proc. Nall. Acad Sei., USA., 1996; 93: 8460-8464.
Winer, M.A., Wadewitz, A.G., and Wolgemu~ D.J. Members ofthe rat gene familyexhibit segment -specifie patterns ofexpression in mouse epididymis. Nlol. Reprod Dev.1993; 35(1): 16-23.
Won~ P.Y.D., Au. C.L. and Ngai, H.K. Electrolyte and water transpon in ratepididymis; 115 possible raie in sperm maturation. lnt. J. Androl, 1978; Suppl.: 608628.
Zheng G, Bachinsky D~ Abbate M, Andres G, Brown D, Stamenkovic 1, Niles IL,McCluskey RT.LRP-2: Receptor and Autoantigen. Annals ofthe New York AcademyofSciences 1994;137:154-163
Zheng, G., Bachinsky, D.R., Stamenkovic, 1., Strickland, O.K., Brown, D., Andres, G.,McCluskey, R.T. (1994b). Organ Distribution in Rats ofTwo Members ofthe LowPensilY Lipoprotein Receptor Gene Family, gp330 and LRP/alpha 2MR. and theReceptor- Associated Protein (RAP). Journal ofHistochemistry & Cytoçh,mi3trY, -12(4):531-542.
76
Related Documents











