0014-2980/98/1212-4138$17.50 + .50/0 © WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1998 Influence of maternal antibodies on vaccine responses: inhibition of antibody but not T cell responses allows successful early prime-boost strategies in mice Claire-Anne Siegrist, Christy Barrios, Xavier Martinez, Christian Brandt, Monika Berney, Marco C ´ ordova, Jiri Kovarik and Paul-Henri Lambert W.H.O. Collaborating Center for Neonatal Vaccinology, Departments of Pathology and Pediatrics, University of Geneva Medical School, Geneva, Switzerland The transfer of maternal antibodies to the offspring and their inhibitory effects on active infant immunization is an important factor hampering the use of certain vaccines, such as measles or respiratory syncytial virus vaccine, in early infancy. The resulting delay in protec- tion by conventional or novel vaccines may have significant public health consequences. To define immunization approaches which may circumvent this phenomenon, experiments were set up to further elucidate its immunological bases. The influence of maternal anti- bodies on antibody and T cell responses to measles hemagglutinin (MV-HA) were analyzed following MV-HA immunization of pups born to immune or control BALB/c mothers using four different antigen delivery systems: live or inactivated conventional measles vaccine, a live recombinant canarypox vector and a DNA vaccine. High levels ( 5 log10) of maternal anti-HA antibodies totally inhibited antibody responses to each of the vaccine constructs, whereas normal antibody responses were elicited in presence of lower titers of maternal antibodies. However, even high titers of maternal antibodies affected neither the induction of vaccine-specific Th1/Th2 responses, as assessed by proliferation and levels of IFN- and IL-5 production, nor CTL responses in infant mice. On the basis of these unaltered T cell responses, very early priming and boosting (at 1 and 3 weeks of age, respectively) with live measles vaccine allowed to circumvent maternal antibody inhibition of antibody responses in pups of immune mothers. This was confirmed in another immunization model (tetanus toxoid). It suggests that effective vaccine responses may be obtained earlier in presence of maternal antibodies through the use of appropriate immunization strategies using conven- tional or novel vaccines for early priming. Key words: Maternal antibody / Immunization / Measles / Infant / Vaccine Received 28/7/98 Accepted 26/8/98 [I 18675] 1 Introduction In humans, maternal IgG Ab transferred from mother to fetus across the placental layer of the syncytiotropho- blast [1] play an essential role in infant protection against a number of viral and bacterial pathogens during a period of a few weeks to months, until their decline below protective thresholds. However, maternal Ab may inhibit infant Ab response to immunization, as shown more than 40 years ago [2]. In fact, this phenomenon is of considerable practical importance since it is prevent- ing the use of several vaccines during early infancy [3], when their protective effect would have a most signifi- cant impact on the related disease burden. It is the basis of the usual recommendation not to immunize against measles prior to the expected age of disappearance of maternal Ab around 12 months of age. Unfortunately, early susceptibility to measles and continued virus circu- lation in partly vaccinated populations still result in a sig- nificant measles morbidity and mortality in infants below 12 months [4, 5]. Inhibition of Ab responses by maternal Ab has also been observed in the context of other live vaccines [6]. Although maternal Ab are often not consid- ered as a problem for non-live infant vaccines, several 4138 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

0014-2980/98/1212-4138$17.50+.50/0 © WILEY-VCH Verlag GmbH, D-69451 Weinheim, 1998
Influence of maternal antibodies on vaccineresponses: inhibition of antibody but not T cellresponses allows successful early prime-booststrategies in mice
Claire-Anne Siegrist, Christy Barrios, Xavier Martinez, Christian Brandt, MonikaBerney, Marco Cordova, Jiri Kovarik and Paul-Henri Lambert
W.H.O. Collaborating Center for Neonatal Vaccinology, Departments of Pathology andPediatrics, University of Geneva Medical School, Geneva, Switzerland
The transfer of maternal antibodies to the offspring and their inhibitory effects on activeinfant immunization is an important factor hampering the use of certain vaccines, such asmeasles or respiratory syncytial virus vaccine, in early infancy. The resulting delay in protec-tion by conventional or novel vaccines may have significant public health consequences. Todefine immunization approaches which may circumvent this phenomenon, experimentswere set up to further elucidate its immunological bases. The influence of maternal anti-bodies on antibody and T cell responses to measles hemagglutinin (MV-HA) were analyzedfollowing MV-HA immunization of pups born to immune or control BALB/c mothers usingfour different antigen delivery systems: live or inactivated conventional measles vaccine, alive recombinant canarypox vector and a DNA vaccine. High levels ( G 5 log10) of maternalanti-HA antibodies totally inhibited antibody responses to each of the vaccine constructs,whereas normal antibody responses were elicited in presence of lower titers of maternalantibodies. However, even high titers of maternal antibodies affected neither the induction ofvaccine-specific Th1/Th2 responses, as assessed by proliferation and levels of IFN- + andIL-5 production, nor CTL responses in infant mice. On the basis of these unaltered T cellresponses, very early priming and boosting (at 1 and 3 weeks of age, respectively) with livemeasles vaccine allowed to circumvent maternal antibody inhibition of antibody responsesin pups of immune mothers. This was confirmed in another immunization model (tetanustoxoid). It suggests that effective vaccine responses may be obtained earlier in presence ofmaternal antibodies through the use of appropriate immunization strategies using conven-tional or novel vaccines for early priming.
Key words: Maternal antibody / Immunization / Measles / Infant / Vaccine
Received 28/7/98Accepted 26/8/98
[I 18675]
1 Introduction
In humans, maternal IgG Ab transferred from mother tofetus across the placental layer of the syncytiotropho-blast [1] play an essential role in infant protection againsta number of viral and bacterial pathogens during aperiod of a few weeks to months, until their declinebelow protective thresholds. However, maternal Ab mayinhibit infant Ab response to immunization, as shown
more than 40 years ago [2]. In fact, this phenomenon isof considerable practical importance since it is prevent-ing the use of several vaccines during early infancy [3],when their protective effect would have a most signifi-cant impact on the related disease burden. It is the basisof the usual recommendation not to immunize againstmeasles prior to the expected age of disappearance ofmaternal Ab around 12 months of age. Unfortunately,early susceptibility to measles and continued virus circu-lation in partly vaccinated populations still result in a sig-nificant measles morbidity and mortality in infants below12 months [4, 5]. Inhibition of Ab responses by maternalAb has also been observed in the context of other livevaccines [6]. Although maternal Ab are often not consid-ered as a problem for non-live infant vaccines, several
4138 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148

clinical studies reported an inhibitory influence of mater-nal Ab [7–10]. This phenomenon is thus recognized as achallenge for the development of new vaccines againstagents such as respiratory syncytial virus (RSV) or Strep-tococcus pneumoniae for which very early vaccineresponses would be desirable.
Inhibition of Ag-specific Ab responses by pre-existing Abwas established long ago [11]. In contrast, the influenceof maternal Ab on Ag-specific T cell responses remainsincompletely defined in spite of numerous in vitro stud-ies. For example, certain mAb were shown to eitherinhibit [12–14] or potentiate [15, 16] in vitro proliferativeresponses of Ag-specific T cells, whereas others did notaffect T cell responses. One mAb was even shown toenhance the presentation of one T cell determinant byAg-specific B cells, while strongly suppressing the pre-sentation of a different T cell determinant [17]. mAbdirected against T cell epitopes presented by MHC classI molecules have also been described [18], suggestingthat they could interfere with CTL responses. In contrastto the effects of passively transferred Ab on adultimmune responses, fewer studies have been dealingwith the effects of maternal Ab on the induction of infantT cell responses in vivo and some controversial resultswere generated. Prolonged suppression of Ag-specificT cell proliferation by maternal Ab was reported uponimmunization of pups with malaria [19], rabies or Sendaivaccines [20] and interpreted as reflecting the inductionof regulatory T cells by maternal Ab. The in vivo transferof RSV-specific mAb prior to infection of neonatal micealso inhibited the induction of CTL responses, which incontrast remained unaffected in pups of RSV-immunemothers [21]. Last, the ability of DNA vaccines to induceimmunity despite the presence of maternal Ab wasrecently reported [22, 23] and attributed to the genericfeatures of DNA vaccines, although this approach wasnot successful in other immunization models [24–26]. Asa consequence of the diversity of results obtained inthese various studies, which probably reflects the influ-ence of a number of potentially confounding factors, aclear understanding of the immunological bases ofmaternal Ab-mediated inhibition of infant vaccineresponses has not yet emerged.
With the objective of further understanding these immu-nological phenomena, we have analyzed vaccine-induced antibodies, proliferation and cytokine produc-tion by Ag-specific Th1/Th2 cells and specific CTLresponses using various antigen delivery systems of acommon vaccine Ag, namely measles hemagglutinin(MV-HA). Immunization was carried out with either liveattenuated measles virus (infective but poorly replicativein mice) or with the same vaccine after inactivation, orwith a live recombinant canarypox vector expressing
MV-HA, or with an MV-HA DNA vaccine [27, 28]. Theseexperiments were performed in young pups born toimmune BALB/c mothers to take into account the con-text of the early life immunological immaturity, including(1) the assessment of physiological titers of maternal Ab,(2) the frequently observed preferential Th2 polarizationof early life responses to conventional vaccines, and (3)the potential influence of maternal Ab on the selection ofthe Ab repertoire. Eventually, experiments assessed thecapacity of new immunization strategies using conven-tional vaccines to escape from inhibition by maternal Ab.
2 Results
2.1 Inhibition of antibody responses to liveattenuated and novel canarypox-vectoredmeasles vaccines
Following immunization of adult female BALB/c micewith a standard live attenuated measles vaccine(Schwarz strain, MV-S) or with a novel recombinantcanarypox vector expressing MV-HA (ALVAC-HA) priorto mating, MV-HA IgG Ab were found at similar levels inimmunized mothers and in their 2-week-old pups(Fig. 1A–D). Comparative Ab subclass analyses indi-cated a similar distribution of MV-HA IgG1, IgG2a andIgG3 Ab in mothers and 2-week-old pups (not shown),and a lower level of IgG2b transfer (MV-S: 3.0 vs. X 2log10; ALVAC-HA: 3.6 vs. X 2 log10, respectively), con-sistent with the known lower affinity of the FcRn receptorfor IgG2b antibodies.
The influence of maternal Ab on Ab responses to a stan-dard live attenuated measles vaccine (MV-S) was mea-sured first. A progressive decline of MV-HA Ab wasobserved whether 2-week-old pups of MV-S immunemothers had been immunized with MV-S, or injectedwith PBS as controls (Fig. 1A). Ab disappearance at 12weeks of age confirmed the inhibition of vaccine-induced Ab responses in the offspring of immune moth-ers. This lack of active response contrasted with theinduction of significant and sustained Ab responses inpups born to nonimmune control mothers. The sameinhibition was observed when MV-S immunization wasperformed 48 h after i.p. transfer of MV-S immune serum(200 ? l), confirming that the interference of maternalimmunity with neonatal vaccine responses was due tothe maternofetal transfer of Ab rather than of Ag or ofcells of maternal origin. In contrast, when MV-S immuni-zation was delayed until maternal MV-HA Ab haddeclined to tenfold lower titers (4.5 log10), i.e. in 8-week-old pups of immune mothers, IgG Ab responses becamesimilar to those of pups of control mothers (Fig. 1B), withidentical isotype distribution (not shown). A prolonged
Eur. J. Immunol. 1998. 28: 4138–4148 Maternal antibodies do not inhibit T cell responses to vaccines 4139
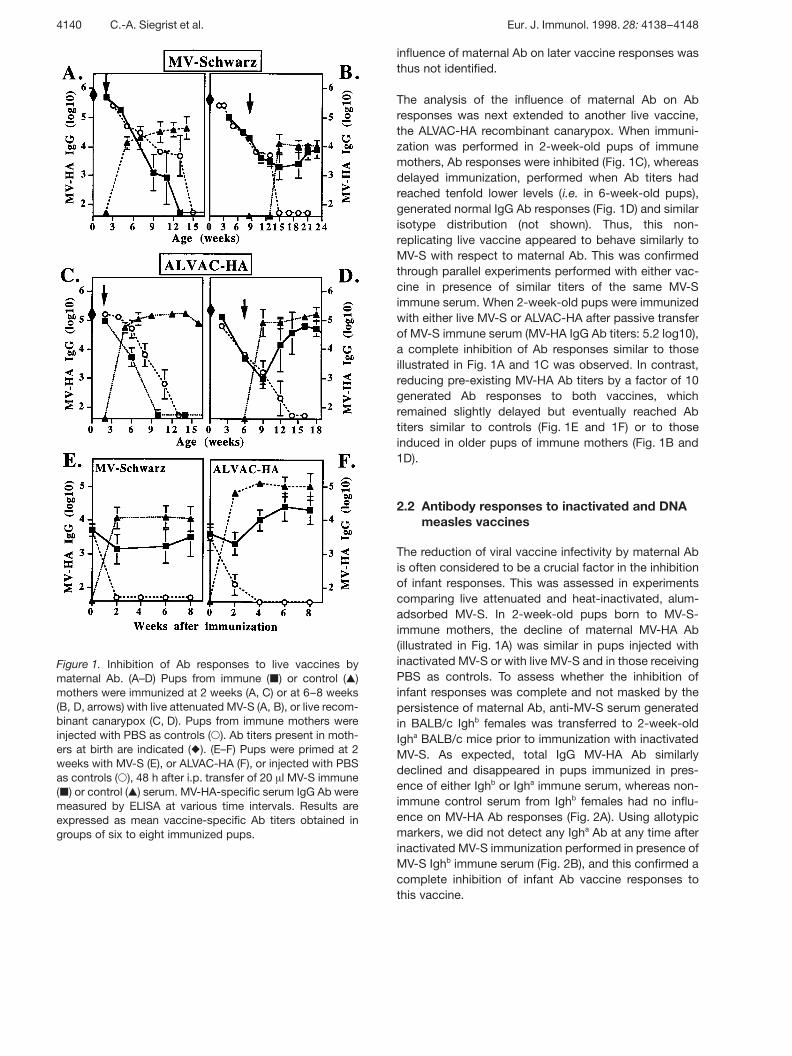
Figure 1. Inhibition of Ab responses to live vaccines bymaternal Ab. (A–D) Pups from immune ( | ) or control ( , )mothers were immunized at 2 weeks (A, C) or at 6–8 weeks(B, D, arrows) with live attenuated MV-S (A, B), or live recom-binant canarypox (C, D). Pups from immune mothers wereinjected with PBS as controls ( 1 ). Ab titers present in moth-ers at birth are indicated ( ! ). (E–F) Pups were primed at 2weeks with MV-S (E), or ALVAC-HA (F), or injected with PBSas controls ( 1 ), 48 h after i.p. transfer of 20 ? l MV-S immune( | ) or control ( , ) serum. MV-HA-specific serum IgG Ab weremeasured by ELISA at various time intervals. Results areexpressed as mean vaccine-specific Ab titers obtained ingroups of six to eight immunized pups.
influence of maternal Ab on later vaccine responses wasthus not identified.
The analysis of the influence of maternal Ab on Abresponses was next extended to another live vaccine,the ALVAC-HA recombinant canarypox. When immuni-zation was performed in 2-week-old pups of immunemothers, Ab responses were inhibited (Fig. 1C), whereasdelayed immunization, performed when Ab titers hadreached tenfold lower levels (i.e. in 6-week-old pups),generated normal IgG Ab responses (Fig. 1D) and similarisotype distribution (not shown). Thus, this non-replicating live vaccine appeared to behave similarly toMV-S with respect to maternal Ab. This was confirmedthrough parallel experiments performed with either vac-cine in presence of similar titers of the same MV-Simmune serum. When 2-week-old pups were immunizedwith either live MV-S or ALVAC-HA after passive transferof MV-S immune serum (MV-HA IgG Ab titers: 5.2 log10),a complete inhibition of Ab responses similar to thoseillustrated in Fig. 1A and 1C was observed. In contrast,reducing pre-existing MV-HA Ab titers by a factor of 10generated Ab responses to both vaccines, whichremained slightly delayed but eventually reached Abtiters similar to controls (Fig. 1E and 1F) or to thoseinduced in older pups of immune mothers (Fig. 1B and1D).
2.2 Antibody responses to inactivated and DNAmeasles vaccines
The reduction of viral vaccine infectivity by maternal Abis often considered to be a crucial factor in the inhibitionof infant responses. This was assessed in experimentscomparing live attenuated and heat-inactivated, alum-adsorbed MV-S. In 2-week-old pups born to MV-S-immune mothers, the decline of maternal MV-HA Ab(illustrated in Fig. 1A) was similar in pups injected withinactivated MV-S or with live MV-S and in those receivingPBS as controls. To assess whether the inhibition ofinfant responses was complete and not masked by thepersistence of maternal Ab, anti-MV-S serum generatedin BALB/c Ighb females was transferred to 2-week-oldIgha BALB/c mice prior to immunization with inactivatedMV-S. As expected, total IgG MV-HA Ab similarlydeclined and disappeared in pups immunized in pres-ence of either Ighb or Igha immune serum, whereas non-immune control serum from Ighb females had no influ-ence on MV-HA Ab responses (Fig. 2A). Using allotypicmarkers, we did not detect any Igha Ab at any time afterinactivated MV-S immunization performed in presence ofMV-S Ighb immune serum (Fig. 2B), and this confirmed acomplete inhibition of infant Ab vaccine responses tothis vaccine.
4140 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148
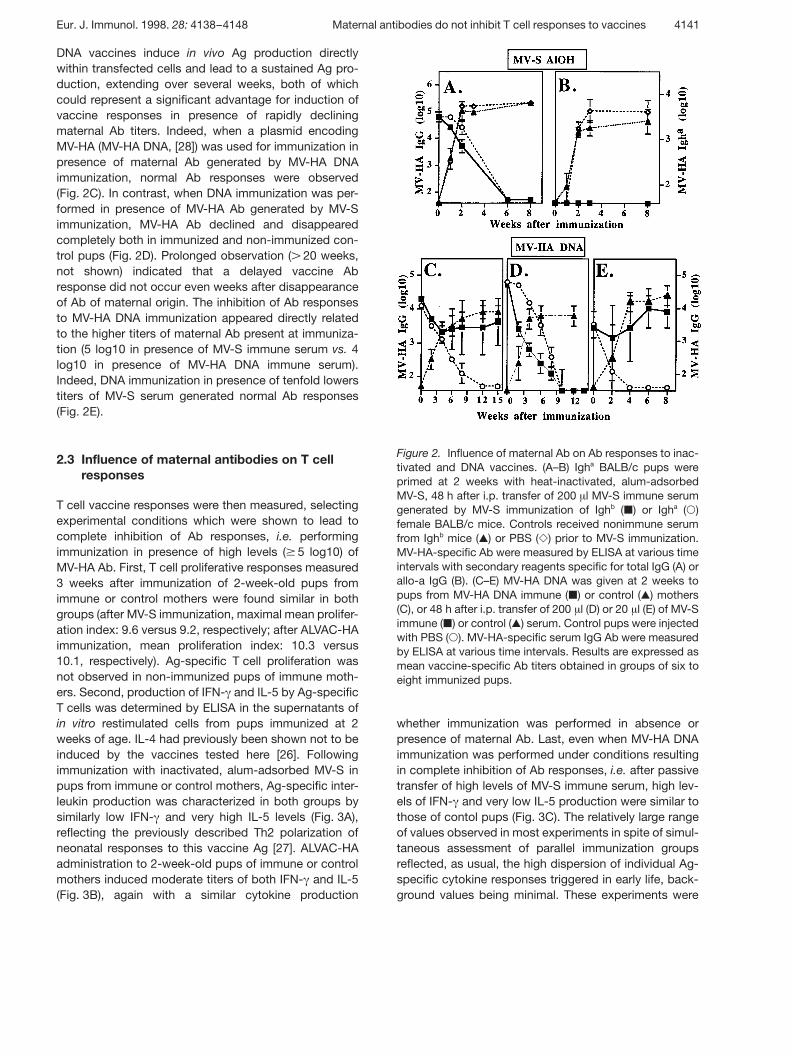
Figure 2. Influence of maternal Ab on Ab responses to inac-tivated and DNA vaccines. (A–B) Igha BALB/c pups wereprimed at 2 weeks with heat-inactivated, alum-adsorbedMV-S, 48 h after i.p. transfer of 200 ? l MV-S immune serumgenerated by MV-S immunization of Ighb ( | ) or Igha ( 1 )female BALB/c mice. Controls received nonimmune serumfrom Ighb mice ( , ) or PBS ( Y ) prior to MV-S immunization.MV-HA-specific Ab were measured by ELISA at various timeintervals with secondary reagents specific for total IgG (A) orallo-a IgG (B). (C–E) MV-HA DNA was given at 2 weeks topups from MV-HA DNA immune ( | ) or control ( , ) mothers(C), or 48 h after i.p. transfer of 200 ? l (D) or 20 ? l (E) of MV-Simmune ( | ) or control ( , ) serum. Control pups were injectedwith PBS ( 1 ). MV-HA-specific serum IgG Ab were measuredby ELISA at various time intervals. Results are expressed asmean vaccine-specific Ab titers obtained in groups of six toeight immunized pups.
DNA vaccines induce in vivo Ag production directlywithin transfected cells and lead to a sustained Ag pro-duction, extending over several weeks, both of whichcould represent a significant advantage for induction ofvaccine responses in presence of rapidly decliningmaternal Ab titers. Indeed, when a plasmid encodingMV-HA (MV-HA DNA, [28]) was used for immunization inpresence of maternal Ab generated by MV-HA DNAimmunization, normal Ab responses were observed(Fig. 2C). In contrast, when DNA immunization was per-formed in presence of MV-HA Ab generated by MV-Simmunization, MV-HA Ab declined and disappearedcompletely both in immunized and non-immunized con-trol pups (Fig. 2D). Prolonged observation ( G 20 weeks,not shown) indicated that a delayed vaccine Abresponse did not occur even weeks after disappearanceof Ab of maternal origin. The inhibition of Ab responsesto MV-HA DNA immunization appeared directly relatedto the higher titers of maternal Ab present at immuniza-tion (5 log10 in presence of MV-S immune serum vs. 4log10 in presence of MV-HA DNA immune serum).Indeed, DNA immunization in presence of tenfold lowerstiters of MV-S serum generated normal Ab responses(Fig. 2E).
2.3 Influence of maternal antibodies on T cellresponses
T cell vaccine responses were then measured, selectingexperimental conditions which were shown to lead tocomplete inhibition of Ab responses, i.e. performingimmunization in presence of high levels ( n 5 log10) ofMV-HA Ab. First, T cell proliferative responses measured3 weeks after immunization of 2-week-old pups fromimmune or control mothers were found similar in bothgroups (after MV-S immunization, maximal mean prolifer-ation index: 9.6 versus 9.2, respectively; after ALVAC-HAimmunization, mean proliferation index: 10.3 versus10.1, respectively). Ag-specific T cell proliferation wasnot observed in non-immunized pups of immune moth-ers. Second, production of IFN- + and IL-5 by Ag-specificT cells was determined by ELISA in the supernatants ofin vitro restimulated cells from pups immunized at 2weeks of age. IL-4 had previously been shown not to beinduced by the vaccines tested here [26]. Followingimmunization with inactivated, alum-adsorbed MV-S inpups from immune or control mothers, Ag-specific inter-leukin production was characterized in both groups bysimilarly low IFN- + and very high IL-5 levels (Fig. 3A),reflecting the previously described Th2 polarization ofneonatal responses to this vaccine Ag [27]. ALVAC-HAadministration to 2-week-old pups of immune or controlmothers induced moderate titers of both IFN- + and IL-5(Fig. 3B), again with a similar cytokine production
whether immunization was performed in absence orpresence of maternal Ab. Last, even when MV-HA DNAimmunization was performed under conditions resultingin complete inhibition of Ab responses, i.e. after passivetransfer of high levels of MV-S immune serum, high lev-els of IFN- + and very low IL-5 production were similar tothose of contol pups (Fig. 3C). The relatively large rangeof values observed in most experiments in spite of simul-taneous assessment of parallel immunization groupsreflected, as usual, the high dispersion of individual Ag-specific cytokine responses triggered in early life, back-ground values being minimal. These experiments were
Eur. J. Immunol. 1998. 28: 4138–4148 Maternal antibodies do not inhibit T cell responses to vaccines 4141
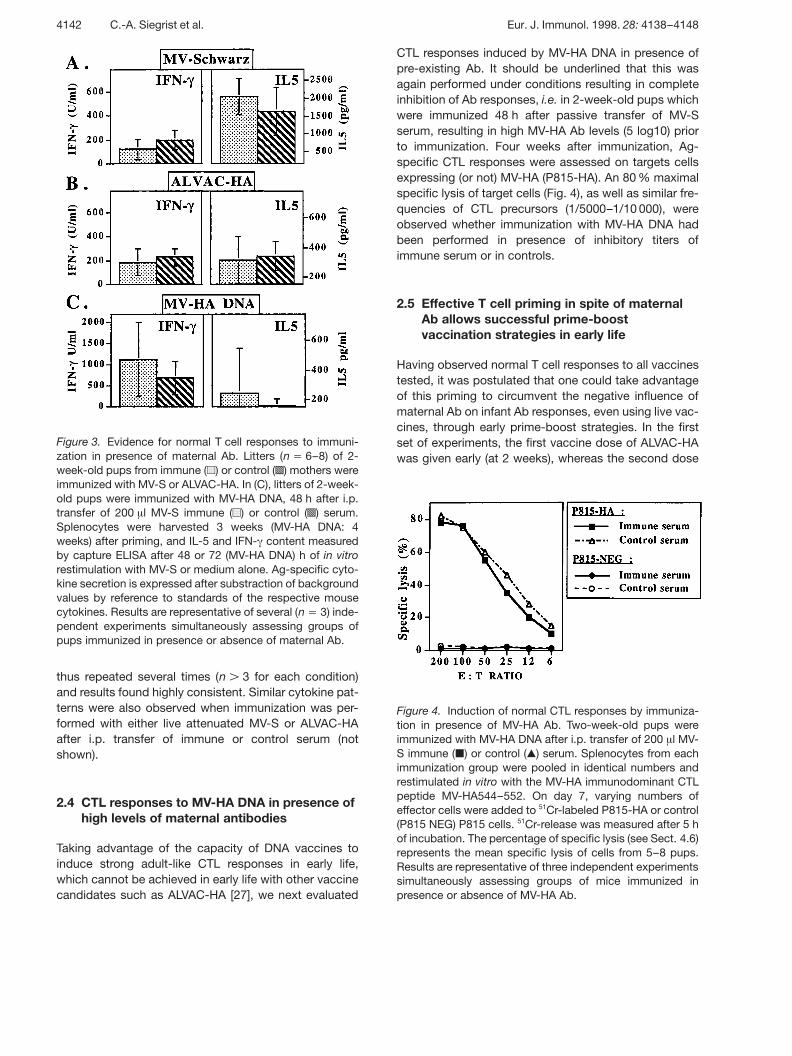
Figure 3. Evidence for normal T cell responses to immuni-zation in presence of maternal Ab. Litters (n j 6–8) of 2-week-old pups from immune ( ⁄ ) or control ( L ) mothers wereimmunized with MV-S or ALVAC-HA. In (C), litters of 2-week-old pups were immunized with MV-HA DNA, 48 h after i.p.transfer of 200 ? l MV-S immune ( ⁄ ) or control ( L ) serum.Splenocytes were harvested 3 weeks (MV-HA DNA: 4weeks) after priming, and IL-5 and IFN- + content measuredby capture ELISA after 48 or 72 (MV-HA DNA) h of in vitrorestimulation with MV-S or medium alone. Ag-specific cyto-kine secretion is expressed after substraction of backgroundvalues by reference to standards of the respective mousecytokines. Results are representative of several (n j 3) inde-pendent experiments simultaneously assessing groups ofpups immunized in presence or absence of maternal Ab.
Figure 4. Induction of normal CTL responses by immuniza-tion in presence of MV-HA Ab. Two-week-old pups wereimmunized with MV-HA DNA after i.p. transfer of 200 ? l MV-S immune ( | ) or control ( , ) serum. Splenocytes from eachimmunization group were pooled in identical numbers andrestimulated in vitro with the MV-HA immunodominant CTLpeptide MV-HA544–552. On day 7, varying numbers ofeffector cells were added to 51Cr-labeled P815-HA or control(P815 NEG) P815 cells. 51Cr-release was measured after 5 hof incubation. The percentage of specific lysis (see Sect. 4.6)represents the mean specific lysis of cells from 5–8 pups.Results are representative of three independent experimentssimultaneously assessing groups of mice immunized inpresence or absence of MV-HA Ab.
thus repeated several times (n G 3 for each condition)and results found highly consistent. Similar cytokine pat-terns were also observed when immunization was per-formed with either live attenuated MV-S or ALVAC-HAafter i.p. transfer of immune or control serum (notshown).
2.4 CTL responses to MV-HA DNA in presence ofhigh levels of maternal antibodies
Taking advantage of the capacity of DNA vaccines toinduce strong adult-like CTL responses in early life,which cannot be achieved in early life with other vaccinecandidates such as ALVAC-HA [27], we next evaluated
CTL responses induced by MV-HA DNA in presence ofpre-existing Ab. It should be underlined that this wasagain performed under conditions resulting in completeinhibition of Ab responses, i.e. in 2-week-old pups whichwere immunized 48 h after passive transfer of MV-Sserum, resulting in high MV-HA Ab levels (5 log10) priorto immunization. Four weeks after immunization, Ag-specific CTL responses were assessed on targets cellsexpressing (or not) MV-HA (P815-HA). An 80 % maximalspecific lysis of target cells (Fig. 4), as well as similar fre-quencies of CTL precursors (1/5000–1/10 000), wereobserved whether immunization with MV-HA DNA hadbeen performed in presence of inhibitory titers ofimmune serum or in controls.
2.5 Effective T cell priming in spite of maternalAb allows successful prime-boostvaccination strategies in early life
Having observed normal T cell responses to all vaccinestested, it was postulated that one could take advantageof this priming to circumvent the negative influence ofmaternal Ab on infant Ab responses, even using live vac-cines, through early prime-boost strategies. In the firstset of experiments, the first vaccine dose of ALVAC-HAwas given early (at 2 weeks), whereas the second dose
4142 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148
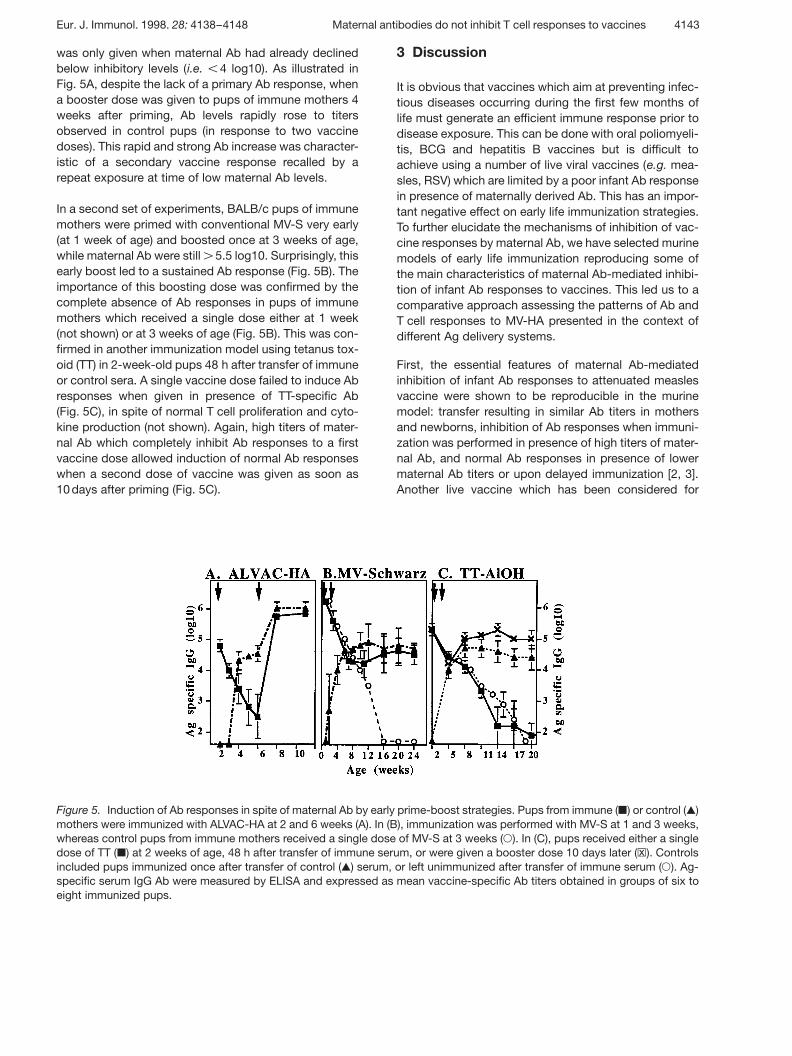
Figure 5. Induction of Ab responses in spite of maternal Ab by early prime-boost strategies. Pups from immune ( | ) or control ( , )mothers were immunized with ALVAC-HA at 2 and 6 weeks (A). In (B), immunization was performed with MV-S at 1 and 3 weeks,whereas control pups from immune mothers received a single dose of MV-S at 3 weeks ( 1 ). In (C), pups received either a singledose of TT ( | ) at 2 weeks of age, 48 h after transfer of immune serum, or were given a booster dose 10 days later ( ßx ). Controlsincluded pups immunized once after transfer of control ( , ) serum, or left unimmunized after transfer of immune serum ( 1 ). Ag-specific serum IgG Ab were measured by ELISA and expressed as mean vaccine-specific Ab titers obtained in groups of six toeight immunized pups.
was only given when maternal Ab had already declinedbelow inhibitory levels (i.e. X 4 log10). As illustrated inFig. 5A, despite the lack of a primary Ab response, whena booster dose was given to pups of immune mothers 4weeks after priming, Ab levels rapidly rose to titersobserved in control pups (in response to two vaccinedoses). This rapid and strong Ab increase was character-istic of a secondary vaccine response recalled by arepeat exposure at time of low maternal Ab levels.
In a second set of experiments, BALB/c pups of immunemothers were primed with conventional MV-S very early(at 1 week of age) and boosted once at 3 weeks of age,while maternal Ab were still G 5.5 log10. Surprisingly, thisearly boost led to a sustained Ab response (Fig. 5B). Theimportance of this boosting dose was confirmed by thecomplete absence of Ab responses in pups of immunemothers which received a single dose either at 1 week(not shown) or at 3 weeks of age (Fig. 5B). This was con-firmed in another immunization model using tetanus tox-oid (TT) in 2-week-old pups 48 h after transfer of immuneor control sera. A single vaccine dose failed to induce Abresponses when given in presence of TT-specific Ab(Fig. 5C), in spite of normal T cell proliferation and cyto-kine production (not shown). Again, high titers of mater-nal Ab which completely inhibit Ab responses to a firstvaccine dose allowed induction of normal Ab responseswhen a second dose of vaccine was given as soon as10 days after priming (Fig. 5C).
3 Discussion
It is obvious that vaccines which aim at preventing infec-tious diseases occurring during the first few months oflife must generate an efficient immune response prior todisease exposure. This can be done with oral poliomyeli-tis, BCG and hepatitis B vaccines but is difficult toachieve using a number of live viral vaccines (e.g. mea-sles, RSV) which are limited by a poor infant Ab responsein presence of maternally derived Ab. This has an impor-tant negative effect on early life immunization strategies.To further elucidate the mechanisms of inhibition of vac-cine responses by maternal Ab, we have selected murinemodels of early life immunization reproducing some ofthe main characteristics of maternal Ab-mediated inhibi-tion of infant Ab responses to vaccines. This led us to acomparative approach assessing the patterns of Ab andT cell responses to MV-HA presented in the context ofdifferent Ag delivery systems.
First, the essential features of maternal Ab-mediatedinhibition of infant Ab responses to attenuated measlesvaccine were shown to be reproducible in the murinemodel: transfer resulting in similar Ab titers in mothersand newborns, inhibition of Ab responses when immuni-zation was performed in presence of high titers of mater-nal Ab, and normal Ab responses in presence of lowermaternal Ab titers or upon delayed immunization [2, 3].Another live vaccine which has been considered for
Eur. J. Immunol. 1998. 28: 4138–4148 Maternal antibodies do not inhibit T cell responses to vaccines 4143

human immunization, the recombinant canarypoxALVAC-HA vaccine, behaved similarly and could notescape the inhibitory influence of high titers of anti-HAmaternal Ab. In fact, the high susceptibility of liveattenuated measles vaccines to the inhibitory influence ofmaternal Ab, which can affect the in vivo replication of theattenuated virus, led to the frequent understanding thatmaternal Ab only affect responses to live vaccines. How-ever, a complete inhibition of Ab responses tonon-live vaccine Ag, whether inactivated measles virus orTT, was also observed when immunization was performedin presence of high titers of maternal Ab. Thus, althoughthe immunogenicity of live replicating vaccines isexpected to be influenced by lower titers of neutralizingmaternal Ab as compared to non-replicating vaccineswith predefined Ag load [7–10], maternal Ab-mediatedinhibition of vaccine responses is clearly not restricted tovaccines subject to neutralization of infectivity.
Recent reports indicated that DNA vaccines were capa-ble of circumventing the influence of maternal Ab insome [22, 23], but not in other [24–26], experimental sys-tems. When compared to other Ag presentation systemsinvolving the same MV-HA protein, we found MV-HADNA to behave quite similarly in presence of maternalAb: depending on the level of maternal Ab titers at timeof immunization, MV-HA DNA was capable, or not, ofinducing normal Ab responses in the offspring. Thecapacity of DNA vaccines to escape from inhibition bymaternal Ab is therefore not a generic feature. We rathersuggest that the capacity of DNA vaccines to circumvent– or not – maternal antibody-mediated inhibition essen-tially depends on whether in vivo Ag production outlasts– or not – the persistence of inhibitory titers of maternalAb in the offspring. The duration of Ag production in vivoafter injection of a DNA vaccine was shown to depend onimmune-mediated destruction of APC [29]. This mayvary from a few weeks to several months depending onthe Ag [29], and is likely to directly contribute to thediversity of the reports using DNA vaccines in early life.Alternatively, maternal Ab persistence in the offspringalso varies, depending upon initial maternal Ab titers.Indeed, DNA vaccines were often shown to induce nor-mal Ab responses under conditions associated with rela-tively moderate levels of pre-existing Ab. This is the casein pups of mothers immunized with DNA vaccines, whichfrequently generate lower Ab responses than conven-tional vaccines, and in pups immunized at X 3 days oflife, when Ab transfer through suckling is not yet com-pleted. Thus, it is not the vaccine type, whether live,inactivated or DNA, which emerges as the main determi-nant of the influence of maternal immunity on Abresponses, but rather the relative amount of maternalantibodies to vaccine antigen.
The essential observation of this report is that the failureof both conventional and novel vaccines to induce Abresponses in presence of high levels of maternal Ab doesnot reflect a complete inhibition of the immune response.Vaccine-induced T cell proliferation and cytokine pro-duction, and CTL responses, are not affected even whenhigh titers of maternal Ab completely inhibit offspring Abresponses. This was shown after immunization in earlylife using attenuated measles or MV-HA expressingrecombinant viruses as well as inactivated measles virusor the MV-HA DNA vaccine. In addition, abrogation of Abresponses but evidence of unaltered T cell priming wasalso observed using a TT vaccine. This indicates thatclearing of immune complexes formed upon introductionof Ag into a host with pre-existing Ab does not lead toescape of Ag recognition by the immune system andsubsequent vacccine failure. At least some of theseimmune complexes are picked up by cells capable ofMHC-restricted presentation of antigenic peptides toT cells, without detectable negative effects of pre-existing Ab on Ag internalization, processing and pre-sentation. The preferential uptake of such immune com-plexes by non-professional APC was expected, whichmight have further enhanced the frequent Th2 polariza-tion of neonatal and early life responses. However, thefact that infant vaccine responses, following immuniza-tion with Ag delivery systems inducing (1) preferentialTh2 [MV-S, TT (not shown)], (2) mixed Th1/Th2 (ALVAC-HA) or (3) preferential Th1 (MV-HA DNA) responses, inpresence of inhibitory titers of maternal Ab were neitherassociated with any significant change in the productionof IFN- + nor IL-5 does not suggest an effect of maternalAb on the Th1/Th2 balance. Ag-specific T cell responsesfollowing immunization in presence of maternal Ab thusappear normal both quantitatively and qualitatively.
These observations of unaltered T cell responses differsomewhat from results obtained in in vitro experimentsshowing that Ag processing may be affected, either pos-itively or negatively, by specific Ab [12–17]. However, thisinterference with Ag processing was reported for high-affinity mAb capable of remaining tightly bound to Ageven at the acidic pH values found in endosomes, thuspreventing either Ag internalization or endosomal pro-cessing. It was not observed in vitro with Ab of loweraffinity, and does apparently not occur with polyclonalAb raised by maternal immunization in the weeks pre-ceding gestation (this report). Interference with Ag pre-sentation at the surface of APC, as shown in vitro with Abto MHC-peptide complexes, was not detectable either inour in vivo immunization models since successful activa-tion of both CD4 and CD8 T cells was demonstrated.However, one cannot exclude that such a phenomenonwould affect responses to some T cell epitopes.
4144 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148

Our observations are in agreement with one report ofunaffected CTL responses to RSV in pups of RSV-infected mothers [21], and with studies assessing theinfluence of passively acquired Ab on T cell proliferativeand CTL responses of adult animals, including measlesvaccines in primates [30]. Infant T cell responses raisedby measles vaccination in presence of various levels ofmaternal Ab have not yet been reported in humans, butsome clinical observations suggest a situation similar tothat of our murine neonatal immunization models. In chil-dren who did not seroconvert after measles immuniza-tion in presence of maternal Ab, either measles exposure[31] or a second vaccine dose given much later [32, 33]indeed leads to high IgG titers characteristics of second-ary responses. Measles is also relatively benign in thosevaccinated infants who did not seroconvert in presenceof residual maternal Ab [31], suggesting partial protec-tion related to an initial T cell priming, whether suchT cells directly contribute to viral control or accelerate Abresponses at time of exposure.
Thus, our experimental observations which identifyB cells as the essential targets of inhibition by maternalAb may have particular relevance for the design of exper-imental infant vaccination strategies aiming at the pre-vention of early infancy infectious diseases. The fact thata normal secondary response to live attenuated measlesand TT vaccines could be obtained by boosting miceonly 2 weeks after giving a priming dose at 1 week ofage, in presence of yet very high levels of maternal Ab,suggests the potential benefits of early prime-boostingstrategies for human vaccination. Should clinical studiesconfirm that infant T cell responses are not inhibited bymaternal immunization, early prime-boost strategiesmay help to solve vaccination problems related to thepersistence of maternal Ab.
4 Materials and methods
4.1 Mice
Adult BALB/c mice were purchased from IFFA-CREDO(L’Arbresle, France) and kept under SPF conditions. Breed-ing cages were checked daily for new births, and the day ofbirth was recorded as the day the litter was found. Pupswere kept with mothers until weaning at the age of 4 weeks.Two-week-old pups were selected (unless otherwise indi-cated) on the basis of their maximal acquisition of maternalAb through suckling compared to X 1-week-old pups (notshown), their sufficient size to allow bleeding for individualdetermination of acquired maternal Ab prior to immunizationand their yet immature immune system in response to vac-cine Ag [26].
4.2 Vaccines
Live attenuated measles virus (MV-S, 5 × 105 CCID50 perdose, given with/without 0.25 mg of alum as indicated) andlive recombinant canarypox expressing MV-HA (vCp85,5 × 107 PFU per dose [27]) were obtained through PasteurMerieux Connaught. TT adsorbed to alum (AnatoxalR) wasreceived from the Swiss Serum Institute, Bern, Switzerland.The MV-HA DNA vaccine (100 ? g/dose) containing themembrane-bound MV-HA-encoding cDNA subcloned intothe pV1J plasmid [28] was kindly supplied by F. Wild (InstitutPasteur, Lyon, France). Endotoxin-free DNA was producedand purified as described [28].
4.3 Immunization procedures
Mice were immunized in groups of six to eight animals,either i.p. or i.m. (MV-HA DNA), as described [27, 28]. Con-trol mice were immunized simultaneously. Female BALB/cmothers were immunized twice at a 3 weeks interval prior tomating. Some mice were bled at regular intervals and serumaliquots pooled to obtain maternal MV-S/TT immune serum(MV-HA IgG Ab titers: 5.2 log10, TT IgG Ab titers: 5.8 log10).An i.p. transfer of 200 ? l of this maternal immune serum to 2-week-old mice was found to result in similar IgG Ab titersand isotype distribution in maternal immune serum and inpups (not shown).
4.4 Quantification of vaccine-specific antibodies
Mice were bled prior to immunization and at regular intervalsthereafter for the determination of Ag-specific Ab. SerumMV-HA and TT antibodies were measured by ELISA usingLtk-HA-transfected cells or TT-coated plates, as described[27]. Incubation was performed with serial serum dilutionsstarting at 1/100. Results were expressed by reference toserial dilutions of titrated serum pools from immunized adultmice. Ab titers below the cutoff of the assay were beinggiven an arbitrary titer of 1/2 the cutoff to allow calculation ofgeometric mean Ab titers.
4.5 Determination of T cells responses
Splenocytes were harvested 3 or 4 (MV-HA DNA) weeksafter immunization. They were incubated at 37 °C with vac-cine Ag or in DMEM-10 % FCS medium alone (control wells).Proliferation was assessed by determination of [3H]thymi-dine incorporation after 4 days of in vitro culture andexpressed by comparison to cells incubated in absence ofAg (stimulation index). Alternatively, cell supernatants werecollected after 48 h and 72 h for measurement of IL-5 andIFN- + content by capture ELISA [26], following the manufac-turer’s instructions (Pharmingen). Values for IL-5 and IFN- +were expressed by reference to a standard curve con-structed by assaying serial dilutions of the respective mouse
Eur. J. Immunol. 1998. 28: 4138–4148 Maternal antibodies do not inhibit T cell responses to vaccines 4145

cytokines. The assay cutoff was 50 pg/ml for IL-5 and 10 IU/ml (i.e. 80 pg/ml) for IFN- + . Ag-specific cytokine secretionwas obtained by subtracting the cytokine content of thesupernatants from splenocytes incubated with DMEMalone.
4.6 Generation of CTL and cytotoxicity assay
Splenocytes were harvested 4 weeks after MV-HA DNAimmunization. Identical numbers of splenocytes from DNA-HA-immune or control mice were pooled and cultured as abulk or under limiting dilution conditions, as described [28].Briefly, bulk cultures were tested on day 7 for Ag-specificlysis by adding varying numbers of effector cells to 51Cr-labeled HA-transfected or control P815 target cells (5 × 103).After 5 h of incubation at 37 °C, cell supernatants were har-vested for determination of 51Cr in a + -counter. The percent-age of specific lysis was calculated as [(experimental cpm −spontaneous cpm)/(total cpm − spontaneous cpm)] × 100.Spontaneous release and total release were determinedfrom target cells incubated with medium alone or after theaddition of 100 ? l 1 M HCl, respectively. For limiting dilutionanalyses, individual wells were tested at day 10. 51Cr-labeledP815-HA target cells (5 × 103) were added to each well for a5-h incubation at 37 °C. Cell supernatants were harvestedfor determination of 51Cr. Wells with a 51Cr-release higherthan the mean value + 5 SD of the radioactivity measured inthe supernatant of target cells alone were counted as posi-tive wells. CTL precursor frequencies, which represent themean frequency for the immune mice whose cells were ini-tially pooled, were determined by the intersection of theregression line of the frequency of negative wells with thecutoff frequency of 37 %.
4.7 Statistical analysis
Significance analysis between results obtained from variousgroups of mice was performed by using the Mann-WhitneyUtest. Probability values p G 0.05 were considered insignifi-cant.
Acknowledgments: This work was granted through theSwiss National Research Foundation, the W.H.O. GlobalProgramme for Vaccines and Immunization, the SandozFoundation and Pasteur-Merieux Serums et Vaccins. C. A.Siegrist was supported by the Swiss National Science Foun-dation (SCORE A), C. Barrios and X. Martinez by the W.H.O.Global Programme for Vaccines and Immunization, C.Brandt by the Deutsche Forschungsgemeinschaft, M. Cor-doba by the Swiss Confederation and J. Kovarik by the Hel-mut Horten Foundation. We thank Paolo Quirighetti forexcellent assistance in animal care.
5 References
1 Leach, J. L., Sedmak, D. D., Osborne, J. M., Rahill, B.,Lairmore, M. D. and Anderson, C. L., Isolation fromhuman placenta of the IgG transporter, FcRn, and locali-zation to the syncytiotrophoblast. J. Immunol. 1996.157: 337–332.
2 Osborn, J. J., Dancis, J. and Julia, J. F., Studies of theimmunology of the newborn infant. II Interference withactive immunization by passive transplacental circulat-ing antibody. Pediatrics 1952. 10: 328–334.
3 Albrecht, P., Ennis, F. A., Saltzman, E. J. and Krug-man, S., Persistence of maternal antibody in infantsbeyond 12 months: mechanism of measles vaccine fail-ure. J. Pediatr. 1977. 91: 715–718.
4 Burström, B., Aaby, P., Mutie, D. M., Kimani, G. andBjerregaard, P., Severe measles outbreak in WesternKenya. East Afr. Med. J. 1992. 69: 419–423.
5 Malfait, P., Jataou, I. M., Jollet, M. C., Margot, A.,De Benoist, A. C. and Moren, A., Measles epidemic inthe urban community of Niamey: transmission patterns,vaccine efficacy and immunization strategies, Niger1990 to 1991. Pediatr. Infect. Dis. J. 1994. 13: 38–45.
6 WHO Study Group on oral polio vaccine, Factorsaffecting the immunogenicity of oral poliovirus vaccine: aprospective evaluation in Brazil and the Gambia. J.Infect. Dis. 1995. 171: 1097–1106.
7 Booy, R., Aitken, S. J. M., Taylor, S., Tudor-Williams,G., Macfarlane, J. A., Moxon, E. R., Ashworth, L. A.,Mayon-White, R. T., Griffiths, H. and Chapel, H. M.,Immunogenicity of combined diphtheria, tetanus, andpertussis vaccine given at 2, 3, and 4 months versus 3,5, and 9 months of age. Lancet 1992. 339: 507–510.
8 Sarvas, H., Kurikka, S., Seppala, I. J., Makela, P. H.and Makela, O., Maternal antibodies partly inhibit anactive antibody response to routine tetanus toxoidimmunization in infants. J. Infect. Dis. 1992. 165:977–979.
9 Björkholm, B., Granström, M., Taranger, J., Wahl, M.and Hagberg, L., Influence of high titers of maternalantibody on the serologic response of infants to diphthe-ria vaccination at three, five and twelve months of age.Ped. Infect. Dis. J. 1995. 14: 846–850.
10 Englund, J. A., Anderson, E. L., Reed, G., Decker, M.D., Edwards, K. M., Pichichero, M. E., Steinhoff, M.C., Rennels, M. B., Deforest, A. and Meade, B., Theeffect of maternal antibody on the serologic responseand the incidence of adverse reactions after primaryimmunization with a cellular and whole-cell pertussisvaccines combined with diphtheria and tetanus toxoids.Pediatrics 1995. 96 (suppl): 580–584.
4146 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148

11 Uhr, J. W. and Baumann, J., Antibody formation. I. Thesuppression of antibody formation by passively adminis-tered antibody. J. Exp. Med. 1961. 113: 395–397.
12 Mills, K. H. G., Inhibitory effects of monoclonal anti-bodies to a synthetic peptide of influenza hemagglutininon the processing and presentation of viral antigens toclass II-restricted T-cell clones. Immunology 1988. 65:365–371.
13 Jemmerson, R., Johnson, J. G., Burrell, E., Taylor, P.S. and Jenkins, M. K., A monoclonal antibody specificfor a cytochrome c T cell stimulatory peptide inhibitsT cell responses and affects the way the peptide associ-ates with antigen-presenting cells. Eur. J. Immunol.1991. 21: 143–151.
14 Watts, C. and Lanzavecchia, A., Suppressive effect ofantibody on processing of T cell epitopes. J. Exp. Med.1993. 178: 1459–1463.
15 Celis, E., Zurawsky, Jr. V. R. and Chang, T. W., Regula-tion of T cell function by antibodies: enhancement of theresponse of human T-cell clones to hepatitis B surfaceantigen by antigen-specific monoclonal antibodies.Proc. Natl. Acad. Sci. USA 1984. 81: 6846–6850.
16 Schalke, B. C. G., Klinkert, W. E. F., Wekerle, H. andDwyer, D. S., Enhanced activation of a T cell line spe-cific for acetylcholine receptor (AChR) by using anti-AChR monoclonal antibodies plus receptors. J. Immu-nol. 1985. 134: 3643–3648.
17 Simitsek, P. D., Campbell, D. G., Lanzavecchia, A.,Fairweather, N. and Watts, C., Modulation of antigenprocessing by bound antibodies can boost or suppressclass II Major Histocompatibility Complex presentationof different T cell determinants. J. Exp. Med. 1995. 181:1957–1963.
18 Duc, H. T., Rucay, P., Righenzi, S., Halle-Pannenko, O.and Kourilsky, P., Monoclonal antibodies directedagainst T cell epitopes presented by class I MHC anti-gens. Int. Immunol. 1993. 5: 427–431.
19 Harte, P. G. and Playfair, J. H. L., Failure of malaria vac-cination in mice born to immune mothers. II. Induction ofspecific suppressor cells by maternal IgG. Clin. Exp.Immunol. 1983. 51: 157–164.
20 Xiang, Z. Q. and Ertl, H. C. J., Transfer of maternal anti-bodies results in inhibition of specific immune responsesin the offspring. Virus Res. 1992. 24: 297–314.
21 Bangham, C. R. M., Passively acquired antibodies toRespiratory Syncytial virus impair the secondary cyto-toxic T cell response in the neonatal mice. Immunology1986. 59: 37–41.
22 Hasset, D. E., Zhang, J. and Whitton, J. L., NeonatalDNA immunization with a plasmid encoding an internalviral protein is effective in the presence of maternal anti-bodies and protects against subsequent viral challenge.J. Virol. 1997. 71: 7881–7888.
23 Manickan, E., Yu, Z. and Rouse, B. T., DNA immuniza-tion of neonates induces immunity despite the presenceof maternal antibody. J. Clin. Invest. 1997. 100:2371–2375.
24 Wang, Y., Xiang, Z., Pasquini, S. and Ertl, H. C. J.,Effect of passive immunization or maternally transferredimmunity on the antibody response to a genetic vaccineto rabies virus. J. Virol. 1998. 72: 1790–1796.
25 Monteil, M., Le Potier, M. F., Guillotin, J., Cariolet, R.,Houdayer, C. and Eloit, M., Genetic immunization ofseronegative one-day-old piglets against pseudorabiesinduces neutralizing antibodies but not protection and isineffective in piglets from immune dams. Vet. Res. 1996.27: 443–452.
26 Le Potier, M. F., Monteil, M., Houdayer, C. and Eloit,M., Study of the delivery of the gD gene of pseudorabiesvirus to one-day-old piglets by adenovirus or plasmidDNA as ways to bypass the inhibition of immuneresponses by colostral antibodies. Vet. Microbiol. 1997.55: 75–80.
27 Barrios, C., Brawand, P., Berney, M., Brandt, C., Lam-bert, P. H. and Siegrist, C. A., Neonatal and early lifeimmune responses to various forms of vaccine antigensqualitatively differ from adult responses: predominanceof a TH2-biased pattern which persists after adult boost-ing. Eur. J. Immunol. 1996. 26: 1489–1496.
28 Martinez, X., Brandt, C., Saddallah, F., Tougne, C.,Barrios, C., Wild, F., Dougan, G., Lambert, P. H. andSiegrist, C. A., DNA immunization circumvents deficientinduction of TH1 and CTL responses in early life. Proc.Natl. Acad. Sci. USA 1997. 94: 8726–8731.
29 Davis, H. L., Brazolot Millan, C. L. and Watkins, S. C.,Immune-mediated destruction of transfected musclefibers after direct gene transfer with antigen-expressingplasmid DNA. Gene Ther. 1997. 4: 181–188.
30 van Binnendijk, R. S., Peolen, M. C. M., van Ameron-gen, G., de Vries, P. and Osterhaus, A. D. M. E., Pro-tective immunity in macaques vaccinated with live atten-uated, recombinant and subunit measles vaccines in thepresence of passively acquired antibodies. J. Infect. Dis.1997. 175: 534–542.
31 Cherry, J. D., Feigin, R. D., Schackelford, P. G., Hint-horn, D. R. and Schmidt, R. R., A clinical and serologicstudy of 103 children with measles vaccine failure.J. Pediatr. 1973. 82: 802–808.
Eur. J. Immunol. 1998. 28: 4138–4148 Maternal antibodies do not inhibit T cell responses to vaccines 4147

32 McGraw, T. T., Reimmunization following early immuni-zation with measles vaccine: a prospective study. Pedi-atrics 1986. 77: 45–48.
33 Stetler, H. C., Orenstein, W. A., Bernier, R. H., Herr-mann, K. L., Sirotkin, B., Hopfensperger, D., Schuh,R., Albrecht, P., Lievens, A. W. and Brunell, P. A.,Impact of revaccinating children who initially receivedmeasles vaccine before 10 months of age. Pediatrics1986. 77: 471–476.
Correspondence: Claire-Anne Siegrist, Centre for NeonatalVaccinology, C.M.U., 1 rue Michel-Servet, CH-1211 Geneva14, SwitzerlandFax: +41-22 7 02 57 46e-mail: [email protected]
4148 C.-A. Siegrist et al. Eur. J. Immunol. 1998. 28: 4138–4148
Related Documents











