REVIEW In Vitro Blood–Brain Barrier Models—An Overview of Established Models and New Microfluidic Approaches ANETTE WOLFF, 1 MARIA ANTFOLK, 1 BIRGER BRODIN, 2 MARIA TENJE 1,3 1 Lund University, Department of Biomedical Engineering, Lund, Sweden 2 University of Copenhagen, Department of Pharmacy, Copenhagen, Denmark 3 Uppsala University, Department of Engineering Sciences, Uppsala, Sweden Received 14 October 2014; revised 5 December 2014; accepted 5 December 2014 Published online 28 January 2015 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/jps.24329 ABSTRACT: The societal need for new central nervous system (CNS) medicines is substantial, because of the global increase in life expectancy and the accompanying increase in age-related CNS diseases. Low blood–brain barrier (BBB) permeability has been one of the major causes of failure for new CNS drug candidates. There has therefore been a great interest in cell models, which mimic BBB permeation properties. In this review, we present an overview of the performance of monocultured, cocultured, and triple-cultured primary cells and immortalized cell lines, including key parameters such as transendothelial electrical resistance values, permeabilities of paracellular flux markers, and expression of BBB-specific marker proteins. Microfluidic systems are gaining ground as a new automated technical platform for cell culture and systematic analysis. The performance of these systems was compared with current state-of-the-art models and it was noted that, although they show great promise, these systems have not yet reached beyond the proof-of-concept stage. In general, it was found that there were large variations in experimental protocols, BBB phenotype markers, and paracellular flux markers used. It is the author’s opinion that the field may benefit greatly from developing standardized methodologies and initiating collaborative efforts on optimizing culture protocols. C 2015 Wiley Periodicals, Inc. and the American Pharmacists Association J Pharm Sci 104:2727–2746, 2015 Keywords: BBB; in vitro models; membranes; blood–brain barrier; BBB on a chip; cell culture; CNS INTRODUCTION The pharmaceutical industry is faced with one of the greatest challenges of the modern age—developing medicines that can reach the brain. One of the incentives is the fact that the total cost of the neurovascular health care system in Europe was estimated in 2010 at 800 billion €PPP in direct and indirect costs, 1 with an estimated 85% increase in costs for dementia care by 2030, according to Alzheimers Disease International. 2 These numbers can only be expected to increase with the age- ing population. The challenge lies in the existence of the highly effective blood–brain barrier (BBB) that strictly controls what molecules are allowed to pass and reach the brain. Drug screen- ing is very expensive and time-consuming in the current drug development process, where in vivo screening is applied at the preclinical test stage. Much research effort is therefore directed toward the development of functional BBB in vitro models, which allows the speed by which new drugs are made avail- able to increase. Already in 1885, Paul Ehrlich found that dyes injected into the circulatory system stained all organs in the mammalian Abbreviations used: BAEC, bovine aortic endothelial cells; BCEC, brain capil- lary endothelial cells; BBEC, bovine brain endothelial cells; PBEC, porcine brain endothelial cells; HBEC, human brain endothelial cells; HBMECs, human brain microvascular endothelial cells; HBVEC, human brain vascular endothelial cells; hCMEC, human cardiac microvascular endothelial cells; MBEC, murine brain endothelial cells; NPC, neural progenitor cells; RBCECs, rat brain capillary endothelial cells; RBECs, rat brain endothelial cells; RBMECs, rat brain mi- crovascular endothelial cells; TEER, transendothelial electrical resistance; TJ, tight junction. Correspondence to: Anette Wolff (Telephone: +46-709948974; Fax: +46-46222 4527; E-mail: [email protected]) Journal of Pharmaceutical Sciences, Vol. 104, 2727–2746 (2015) C 2015 Wiley Periodicals, Inc. and the American Pharmacists Association body, except for the brain and the spinal cord. 3 On the basis of this work, in 1900, the term BBB or really bluthirnshranke, was coined by Berlin-based Max Lewandowsky. 4 One of Ehrlich’s students went on, in 1913, to show that the effect could be seen in the reverse situation as well, by injecting trypan blue into the cerebrospinal fluid and observing that it did not spread outside of the central nervous system (CNS). 5 It then became apparent that the brain is protected by a very effective barrier shown later to be the result of tight connections between the cerebral endothelial cells. 6 The BBB can be characterized as a highly effective and se- lective barrier in the interface between the blood of the brain microvasculature and the brain tissue and is crucial for achiev- ing a normal function of the CNS. Small lipophilic gases, such as O 2 and CO 2 may diffuse freely across the BBB, but tight junc- tions (TJs) restrict paracellular fluxes of hydrophilic molecules. Various nutrients and large molecules are actively transported across the cellular membranes through transporter proteins or receptor-mediated endocytosis. Recent excellent reviews in the field has dealt with only mi- crofluidic systems, 7 had a more biological approach, 8–11 or a focus on drug permeability and applications. 12,13 This review intends to compare the various different models designed to imitate the BBB. Papers have been selected based on influence on the field, as well as model development, and little focus has been given to application-oriented studies. Current state-of-the art BBB in vitro research models are presented both with re- spect to the cell cultures used and the engineering of the cell culture setup and its integration. This is, to our knowledge, the first time an all-encompassing view is taken on the research where equal efforts have been made to analyze the biological as well as the technical aspects of the complete in vitro systems. Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 2727

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REVIEW
In Vitro Blood–Brain Barrier Models—An Overview of EstablishedModels and New Microfluidic Approaches
ANETTE WOLFF,1 MARIA ANTFOLK,1 BIRGER BRODIN,2 MARIA TENJE1,3
1Lund University, Department of Biomedical Engineering, Lund, Sweden2University of Copenhagen, Department of Pharmacy, Copenhagen, Denmark3Uppsala University, Department of Engineering Sciences, Uppsala, Sweden
Received 14 October 2014; revised 5 December 2014; accepted 5 December 2014
Published online 28 January 2015 in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/jps.24329
ABSTRACT: The societal need for new central nervous system (CNS) medicines is substantial, because of the global increase in lifeexpectancy and the accompanying increase in age-related CNS diseases. Low blood–brain barrier (BBB) permeability has been one of themajor causes of failure for new CNS drug candidates. There has therefore been a great interest in cell models, which mimic BBB permeationproperties. In this review, we present an overview of the performance of monocultured, cocultured, and triple-cultured primary cells andimmortalized cell lines, including key parameters such as transendothelial electrical resistance values, permeabilities of paracellular fluxmarkers, and expression of BBB-specific marker proteins. Microfluidic systems are gaining ground as a new automated technical platformfor cell culture and systematic analysis. The performance of these systems was compared with current state-of-the-art models and it wasnoted that, although they show great promise, these systems have not yet reached beyond the proof-of-concept stage. In general, it wasfound that there were large variations in experimental protocols, BBB phenotype markers, and paracellular flux markers used. It is theauthor’s opinion that the field may benefit greatly from developing standardized methodologies and initiating collaborative efforts onoptimizing culture protocols. C© 2015 Wiley Periodicals, Inc. and the American Pharmacists Association J Pharm Sci 104:2727–2746,2015Keywords: BBB; in vitro models; membranes; blood–brain barrier; BBB on a chip; cell culture; CNS
INTRODUCTION
The pharmaceutical industry is faced with one of the greatestchallenges of the modern age—developing medicines that canreach the brain. One of the incentives is the fact that the totalcost of the neurovascular health care system in Europe wasestimated in 2010 at 800 billion €PPP in direct and indirectcosts,1 with an estimated 85% increase in costs for dementiacare by 2030, according to Alzheimers Disease International.2
These numbers can only be expected to increase with the age-ing population. The challenge lies in the existence of the highlyeffective blood–brain barrier (BBB) that strictly controls whatmolecules are allowed to pass and reach the brain. Drug screen-ing is very expensive and time-consuming in the current drugdevelopment process, where in vivo screening is applied at thepreclinical test stage. Much research effort is therefore directedtoward the development of functional BBB in vitro models,which allows the speed by which new drugs are made avail-able to increase.
Already in 1885, Paul Ehrlich found that dyes injected intothe circulatory system stained all organs in the mammalian
Abbreviations used: BAEC, bovine aortic endothelial cells; BCEC, brain capil-lary endothelial cells; BBEC, bovine brain endothelial cells; PBEC, porcine brainendothelial cells; HBEC, human brain endothelial cells; HBMECs, human brainmicrovascular endothelial cells; HBVEC, human brain vascular endothelial cells;hCMEC, human cardiac microvascular endothelial cells; MBEC, murine brainendothelial cells; NPC, neural progenitor cells; RBCECs, rat brain capillaryendothelial cells; RBECs, rat brain endothelial cells; RBMECs, rat brain mi-crovascular endothelial cells; TEER, transendothelial electrical resistance; TJ,tight junction.
Correspondence to: Anette Wolff (Telephone: +46-709948974; Fax: +46-462224527; E-mail: [email protected])
Journal of Pharmaceutical Sciences, Vol. 104, 2727–2746 (2015)C© 2015 Wiley Periodicals, Inc. and the American Pharmacists Association
body, except for the brain and the spinal cord.3 On the basis ofthis work, in 1900, the term BBB or really bluthirnshranke, wascoined by Berlin-based Max Lewandowsky.4 One of Ehrlich’sstudents went on, in 1913, to show that the effect could be seenin the reverse situation as well, by injecting trypan blue into thecerebrospinal fluid and observing that it did not spread outsideof the central nervous system (CNS).5 It then became apparentthat the brain is protected by a very effective barrier shownlater to be the result of tight connections between the cerebralendothelial cells.6
The BBB can be characterized as a highly effective and se-lective barrier in the interface between the blood of the brainmicrovasculature and the brain tissue and is crucial for achiev-ing a normal function of the CNS. Small lipophilic gases, suchas O2 and CO2 may diffuse freely across the BBB, but tight junc-tions (TJs) restrict paracellular fluxes of hydrophilic molecules.Various nutrients and large molecules are actively transportedacross the cellular membranes through transporter proteins orreceptor-mediated endocytosis.
Recent excellent reviews in the field has dealt with only mi-crofluidic systems,7 had a more biological approach,8–11 or afocus on drug permeability and applications.12,13 This reviewintends to compare the various different models designed toimitate the BBB. Papers have been selected based on influenceon the field, as well as model development, and little focus hasbeen given to application-oriented studies. Current state-of-theart BBB in vitro research models are presented both with re-spect to the cell cultures used and the engineering of the cellculture setup and its integration. This is, to our knowledge, thefirst time an all-encompassing view is taken on the researchwhere equal efforts have been made to analyze the biologicalas well as the technical aspects of the complete in vitro systems.
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 2727

2728 REVIEW
This work intends to contribute to a stronger interdisciplinaryapproach to BBB research.
The Neurovascular Unit
The neurovascular unit is the five cell types that constitutethe BBB: endothelial cells, astrocytes, pericytes, neurons, andmicroglia (Fig. 1).
Although the different cell types of the BBB have beenidentified,14,15 their interdependence and individual roles arestill not fully understood. Here, we list the current level of un-derstanding of the roles of the different cell types: the endothe-lial cells in the brain are specialized BBB cells that form themicrovessels permeating the brain. They differ from regularendothelial cells in four aspects:
1. An absence of fenestrae.2. A continuous basement membrane that is shared with
pericytes, as have been discovered in recent years.16–18
3. A high complexity of TJs.4. Limited pinocytic vesicular transport.
The barrier is tight for polar compounds because of theirlow lipid permeability, and the presence of extremely tight TJs,leaving only very small polar molecules crossing the barrier.19
The transport pathways available for larger molecules are theparacellular and transcellular pathways.20 The paracellularpathways are passive, thus relying solely on the concentrationgradient and the permeability, which is very low for macro-molecules. The transcellular pathways are energy dependentand substance specific.
Tight junctions are structures assisting the endothelialcells in accomplishing the tightness of the BBB. They com-prise at least three different types of transmembrane pro-teins; claudins, occludins, and junctional adhesion molecules.21
Claudin and occludin have both been shown to be paramount tothe selective paracellular permeability.22,23 Three specific trans-
membrane proteins are primarily present in cells with effectivebarrier functions, such as the BBB endothelial cells. They are:ZO-1, claudin-5, and occludin, where ZO-1 is needed as an an-chor point for the claudin and occludin proteins24 and is nec-essary for TJ formation.25 Because of the specificity of thesetransmembrane proteins to the BBB, they are often used asmarkers for successful BBB formation.
Astrocytes are characteristically star-shaped glial cellsfound only in the CNS and they perform different tasks in thebrain, one of them being to give biochemical aid to the endothe-lial cells that form the BBB.26 It has been found that astrocytessecrete factors are needed for BBB function27,28 and assist inregulating transport across the capillaries.29
Pericytes cover 22%–33% of a capillary,30 varying betweenmicrovessel type,31 seemingly correlating with the degree oftightness of the endothelial junctions,32 and pericyte deficiencyis known to increase permeability of the BBB.33 Although it hasbeen suggested that pericytes affect the BBB phenotype,34 it isbelieved that they instead inhibit the expression of moleculesthat increase vascular permeability35 and induce polarity to theastrocyte end feet that surrounds the microvessels,31 resultingin a tightening of the barrier. Pericytes also synthesize elementsnecessary for the differentiation of the BBB36 and are involvedin transport across capillary walls.31
Neurons are the main functional constituents of the brain.They need to be protected against the fluctuations inherentin the mammalian system, for example, temperature fluctua-tions, variations of O2 or CO2 levels, or chemical concentrationvariations, which is the reason for the existence of the BBB.Although the exact mechanism is unknown, Minami37 has re-cently shown that the presence of neurons increases the barriereffects, suggesting that neurons, or communication to the bar-rier, are also a crucial part of the BBB.
The final group of cells that form the neurovascular unitare microglia, which are responsible for clearing debris andhandling apoptotic cells in the brain. They are found inthe perivascular space but will not be discussed further, as
Figure 1. The overview of the neurovascular unit, with the basal lamina denoted as BL. Reproduced from Abbott et al.14 with permission fromElsevier B.V.
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2729
activated macrophages are one of the hallmarks of an infectedbrain17 and have been shown to drastically reduce barrier ef-fectiveness when activated.38
BBB Models
The oldest and best-established method to study the transportof molecules across the BBB is to perform in vivo experiments.The main advantage of in vivo experiments is that it allowsfor studies of the brain in its natural environment. As it isunethical to perform these studies on the human brain, it ne-cessitates the use of laboratory animals. The versatility of theresults is however questionable and it has been shown that ap-proximately 50% of results obtained from animal models cannotbe translated into correct human responses.39 To overcome theethical issues, in vitro models based on cell cultures are applied.The most commonly used methods to determine the quality ofthe in vitro models, is to measure the transendothelial electri-cal resistance (TEER), and to measure the permeability of spe-cific marker substances. Additionally, cell-type-specific mark-ers, such as ZO-1, Claudin-5, Occludin, and von Willebrandsfactor, are targeted for immunostaining to ensure the presenceof the correct cell phenotypes. The different analysis techniquesare described briefly below.
Transendothelial electrical resistance is a quantitative mea-surement of the resistance over the cell layer and the cell cul-ture membrane. Several factors affect the resistance, such ascell origin and the level of confluency of the cells. The measure-ment is performed by connecting electrodes to either side of thecell culture membrane and measuring the momentary resis-tance upon an applied potential (Fig. 2). The in vivo reportedvalues of the resistance across the BBB40–42 is 1500–8000 �cm2,and a value of 150–200 �cm2 is considered the lowest functionallimit for in vitro models.43
The method of measuring TEER differs between groups, be-cause of the various instruments available. “Chopstick” elec-trodes, Endohm cup electrodes, and CellZscope culture cham-bers may give readings that differ. Furthermore, althoughTEER is a relatively simple way of measuring barrier tightness,it is not enough to judge the selectivity of the barrier. Compar-isons of barrier tightness between in vitro models cultured in
Figure 2. The original in vitro Transwell setup. Endothelial cellsgrown on a Transwell membrane. A 4-electrode system is used to mea-sure the resistance over the cells and the membrane. The resistanceover an empty membrane is then subtracted from the cell culture re-sistance.
different laboratories should therefore ideally be accompaniedby comparisons of size selectivity of passively diffusing com-pounds and TJ protein profiles, as the composition of TJ pro-teins will be crucial for the permeation process (for references,see Haseloff et al.44).
Sucrose and mannitol are the most commonly used com-pounds for permeability studies, where the in vivo permeabil-ity has been noted to be approximately 3 × 10−8 and 1.4 ×10−7 cm/s, respectively.45 In vitro models are generally consid-ered useful if the permeabilities are within a factor 100 of the invivo values. Permeability studies often do not reflect the pres-ence of transporter proteins, but the authors have chosen to notgo into detail regarding active transport and efflux proteins.Investigations of this character are highly specific for each sub-stance evaluated, and are beyond the scope of this work. For anin-depth discussion on active transporter and efflux proteins,see the review article by Cardoso et al.10
The three methods to investigate the BBB (to measureTEER, to analyze permeabilities, and to immunostain forknown markers) may thus provide enough information thatconfluence uniformity and tightness of the model barrier canbe estimated.
In Vitro Models
In 1974, the first documented successful culture of bovine cere-bral cortex endothelial cells on nylon sieves was published.46
The most common platform of in vitro models used todayis created using Transwells (Corning Inc., Corning, NY) andmost publications are reported using 10 :m thick polystyreneor polycarbonate membranes with 400 nm pores with a108 pores/mm2 pore density.28,47–54
The models can be developed using one, two, or more celltypes, as described below. The endothelial cells are usuallygrown in the upper (luminal) compartment of the Transwellin cell-specific growth medium. Additional cells, such as astro-cytes or pericytes, are normally cultured on the lower (ablumi-nal) side of the membrane (Fig. 3).
The major challenge with the development of successful invitro models is to identify and culture pure cell types that re-sult in high TEER values and low permeabilities. Althoughthere is some use for nonbrain-derived endothelial cell culturesto establish a new model system, one should strive for usingbrain-derived endothelial cells in a live setup. Brain-derivedendothelial cells will retain some of their BBB phenotypic ex-pressions, whereas those derived from the kidney or gut willgive a tight barrier but will inherently differ widely in trans-port and efflux protein expression.
Preferably, human cells should be used to minimize species-specific responses, though they are not often used, as immor-talized human cell lines do not create a sufficiently tight bar-rier. In addition, primary cells of human origin often stemfrom biopsies of diagnosed patients giving rise to the ques-tion whether the cells are suitable when extracted from pos-sibly diseased tissue.55–57 Shusta and coworkers48,58–60 circum-vent this issue by instead using human pluripotent stem cells.As stem cell-derived brain endothelial cells are difficult toobtain, immortalized human cell lines such as human car-diac microvascular endothelial cells (hCMEC)/D3 are normallychosen for a human model. The cell line human brain mi-crovascular endothelial cells (HBMECs) has been shown to
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2730 REVIEW
Endothelial cellsAstrocytesNeurons/Pericytes
d
a
b c
Figure 3. The different kinds of in vitro models. (a) A monocultureof endothelial cells. (b) A contact coculture of endothelial cells andastrocytes or pericytes. (c) A noncontact coculture of endothelial cellsand astrocytes or pericytes. (d) A triple culture of endothelial cells withastrocytes in contact and pericytes or neurons in a noncontact position.
give the best barrier properties for permeability studies usingTranswells.61
Immortalized animal-derived cell lines are most commonlyused, although most of these models do not reach TEER valuesabove 300–500 �cm2. Barrier properties are commonly betterin rat than mouse models, but are outperformed by porcineand bovine models.50,62–68 Franke et al.69 have however shownthat it is possible to increase the TEER to near-in vivo lev-els by adding hydrocortisone to the culture, which is now acommon approach. Their maximum reported TEER value was1800 �cm2 with a low permeability for sucrose of 0.2–1.8×10−6
cm/s. There are definite advantages to using immortalized ani-mal cell lines—number one being the relative ease with whichthey are cultured.
Additionally, there is a growing interest in using primaryanimal cells to increase the fidelity of the in vitro models, al-though this is a time-consuming and sensitive process.70–76 Themajor drawback of this approach is that primary cells culturedex vivo lose some of their phenotypic expressions with each pas-sage as a result of the lack of exposure to physiological factors.This results in a leaky barrier with low TEER values and highpermeability of marker substances. The negative effects can beminimized by using primary cells within the first couple of pas-sages, preferably using the first passage, and all reports studiedin this review article are performed on cells prior to passage 10.
In the following sections, we will review the current workusing immortalized cell lines and primary cell cultures, andcompare the BBB model system qualities (as defined by re-ported TEER, permeability, and cell type purity) when mono-,
co-, and triple-cultures are applied. For the interested reader,we refer you to the reviews of Gumbleton and Audus,77 Deliet al.,13 and Abbott et al.8 for in-depth discussions about pri-mary cells and cell lines in in vitro studies.
Monoculture
One of the most widely used BBB models is developed by cul-turing endothelial cells alone in a Transwell (Fig. 3a). Hawkinset al.78 established that the microvascular endothelial cellsform the basis of the BBB by showing that frogs have a function-ing BBB, even without astrocyte assistance. Thus, the monocul-tures of endothelial cells have been a reasonable model despiteits simplicity and have been published recently,76,79,80 some-times assisted by the use of astrocyte-conditioned medium, asthis has been shown to increase barrier function.27 Another ap-proach to increase the tightness of a monoculture of endothelialcells is to culture the endothelial cells on a glia-conditioned ex-tracellular matrix, as presented by Hartmann et al.81 However,as established by Patabendige et al.,82 sometimes a monocul-ture can nonetheless be an informative choice when one is in-terested in a robust model with which to perform permeabilitystudies.
Monocultures have seen a revival upon the introduction ofmicrofluidic cell culture systems, which are often limited in thenumber of different cell types they can manage. By integratingthe cells into a microfluidic network, it is possible to intro-duce shear stresses to the cells through the fluid flow. Dane-man et al.27 showed that shear stress, together with astrocyte-conditioned medium can increase the TEER of primaryHBMECs by a factor of 3, from 500 to 1500 �cm2. Similarwork has been presented by Griep et al.83 where they used thehuman cerebral microvascular endothelial cell line hCMEC/D3and showed a 300% increase in TEER when shear stresses wereintroduced (Fig. 4). Cucullo et al.84 cultured hCMEC/D3 undera pulsating flow and could report on an increase in TEER val-ues of close to 20 times compared with static cultures, furtherindicating the importance fluid flow and shear stresses play onthe permeability of the BBB.
Prabhakarpandian et al.85 created a microfluidic chip with amicrocirculation two-compartment chamber in an effort to in-troduce shear stress, whereas at the same time keeping chipmanufacturing simple. Rat brain endothelial cell (RBEC) lineRBE4 was cultured with astrocyte-conditioned medium for theexperiments.85 Low permeability for dextran was presented,
Figure 4. The microsystem developed by Griep et al.83 to mimic the invivo situation of shear stress that affects the microvessels made from aPDMS chip with a Transwell membrane insert. In (a), numbers denote(I) top part, (II) Transwell membrane, (III) bottom part, and (IV) thePt electrodes. Image (b) gives the assembled device and (c) a photo ofthe BBB chip. Reproduced from Griep et al.83 with permission fromSpringer US.
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2731
but unfortunately no TEER values were reported. Dextran isavailable as quite large molecules at 3–5 kDa, so it is not im-mediately clear to see the effectiveness of the barrier in thiswork. This may be attributed to a higher focus of the articleon chip manufacture, and their proof-of-concept approach. Thefield of integrated systems is still young and more publishedwork where the main focus is on the microsystem engineeringcan be expected, though results from monocultures should gen-erally be regarded with caution because of the lack of astrocyteinduction.
Cocultures
In most common coculture models, endothelial cells are grownon the luminal side of a Transwell membrane, whereas astro-cytes, pericytes, or neurons are grown either on the bottomof the well (noncontact) or directly on the abluminal side ofthe membrane (contact) (Figs. 3b and 3d). The contact modelallows the nonendothelial cells to affect the endothelial cellssolely by close-range association, such as by processes throughthe membrane.86 The noncontact model ensures that any ef-fect the nonendothelial cells have on the endothelial cells isstrictly through chemical gradients. Thus, separate and im-portant information on the fundamental understanding of theBBB is provided by the two different methods. A more detaileddescription follows, where additional references are found inTable 1 below.
Endothelial Cells and Astrocytes
In 1980, it was recognized that many BBB features could beat least partly reestablished by coculturing endothelial cells to-gether with astrocytes.124 This work has been extremely influ-ential in the research field and astrocytes are the most commoncell type that endothelial cells are cocultured with.125 Typically,TEER values are increased up to a factor 2 in astrocyte cocul-tures compared with monocultures of endothelial cells.68 If thisis modified to a contact model, the improvement can give asmuch as nine times higher TEER values than the monolayer inprimary cultures.72,87 In addition, astrocytes have been shownto be central to the vasodilation of microvessels, which plays amajor part of the BBB.126
Harris and Shuler127 investigated the effect of the thick-ness of the cell culture membrane between the endothelialcells and the astrocytes. They developed a 1-:m thin silicon ni-tride membrane with 2-:m-wide pores where they cocultured ahuman cerebromicrovascular endothelial cell line (SV-HCEC)with astrocytes.127 The performance of the cell culture scaf-fold was evaluated by observing the cell density on the novelmembrane in relation to an identical culture on traditionalTranswell membranes. The number of cells was higher on thetreated silicon nitride membranes, but unfortunately no TEERvalues, or other quantifiable factors were reported, so it is dif-ficult to judge the barrier performance.
Stanness et al.128 created the first microfluidic system usingthe morphological structure of capillaries to coculture a bovineaortic endothelial cells (BAEC) with a rat astrocyte glioma cellline (C6) under shear stress (DIV-BBB). Hollow fibers with en-dothelial cells cultured on the luminal side, and astrocytes onthe abluminal side, is suspended in a chamber (Fig. 5), withmedium pumped into the chamber. Although this system givesrelevant resistances and permeability results, it is limited infunction and application, as evaluated by Pooja and Cucullo.129
The DIV-BBB is the only system using hollow fibers, whereasthe following microfluidic systems use planar structures, whichis the conventional fabrication method in microengineering. Ifone uses planar structures, you do however risk loosing somefeatures inherent in the BBB by removing a physiological as-pect of the barrier.
Booth and Kim104 also approached the issue of the lack ofa combined shear stress, reported to play a role in the BBBphysiology,100 and dual cell layer setup. They used cut-outs fromTranswell membranes with luminally cultured mouse braincapillary endothelial cell (BCEC) line bEnd.3 and astrocytes incontact abluminally in a microfluidic chip of their own design.A dynamic coculture was shown to be the most effective forestablishing improved barrier functions, outperforming staticcoculture, and monoculture as well as a dynamic monolayer(Fig. 6).104 However, they report unexpectedly low TEER val-ues (never reaching more than 25 �cm2) for their monocul-ture Transwell setup where a TEER for bEnd.3 cells reaches100 �cm2, normally.54 This makes it difficult to comment on thequality of the in vitro model, although the effect of a dynamicenvironment is clearly demonstrated.
Endothelial Cells and Pericytes
In recent years, it has become clear that not only astrocytesplay a major role in the dynamic regulation of the BBB phe-notype in vivo and in vitro, but that also pericytes are veryinfluential. In vivo, it has been shown that pericytes play arole in angiogenesis.35 The effects of pericytes and astrocytesin vitro seem, however, to be strongly linked to cell type origin.Dohgu et al.130 presented a study on the pericyte productionof transforming growth factor-$ type I where they showed asignificant decrease in permeability for sodium fluorescein fora coculture of mouse BCEC and rat brain pericytes. Anotherstudy, using rat brain capillary endothelial cells (RBCECs) re-ports on a 100% increase in TEER to a final value of 200 �cm2
for cocultures with rat brain pericytes.111
The opposite is however reported by Hatherell et al.102 us-ing hCMEC/D3 cocultured with either astrocytes or pericytes ofhuman origin. Here, astrocyte contact resulted in an increasein TEER from 39 to 60 �cm2, whereas cocultures with pericytesresulted in an increase from 39 to 46 �cm2. Although these re-sistances are in line with other studies made with hCMEC/D3,the lack of reported permeabilities makes the study hard tocompare with others presenting higher TEER values.
Another feature that differs the actions of the astrocytes andthe pericytes is that astrocytes can affect the BBB also from adistance, whereas contact seems crucial for pericyte contribu-tion to barrier property.113 Surprisingly, for short-term hypoxicevents, pericytes seem to have a detrimental effect on barrierproperties, significantly lowering the TEER compared with amonoculture of RBE4 cells.49 However, under prolonged O2 de-privation astrocytes proved less efficient at preserving barrierfunction than did pericytes.49 This indicates a bearing for in-cluding pericytes when establishing a BBB model intended forischemic studies.
Triple-Culture
Although it is largely accepted that the endothelial cells formthe basis for the BBB structure, a higher level of resemblancebetween the in vitro model systems and in vivo can be achieved
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015
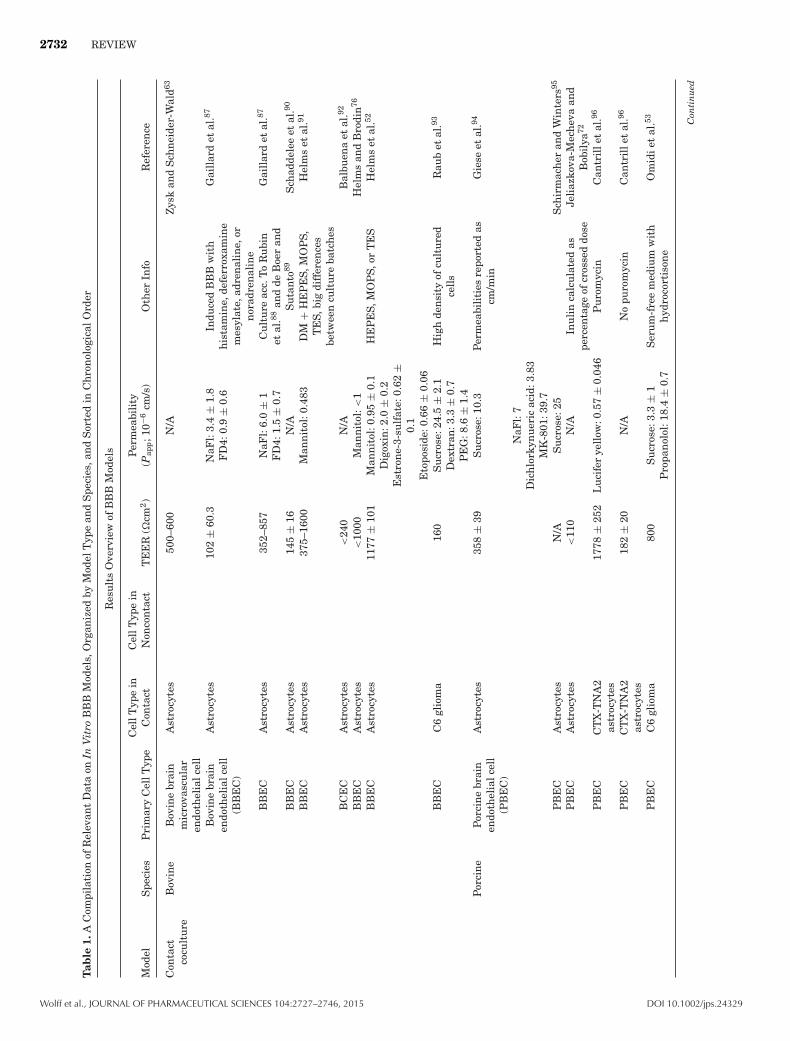
2732 REVIEW
Tab
le1.
AC
ompi
lati
onof
Rel
evan
tD
ata
onIn
Vit
roB
BB
Mod
els,
Org
aniz
edby
Mod
elT
ype
and
Spe
cies
,an
dS
orte
din
Ch
ron
olog
ical
Ord
er
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
Con
tact
cocu
ltu
reB
ovin
eB
ovin
ebr
ain
mic
rova
scu
lar
endo
thel
ialc
ell
Ast
rocy
tes
500–
600
N/A
Zys
kan
dS
chn
eide
r-W
ald63
Bov
ine
brai
nen
doth
elia
lcel
l(B
BE
C)
Ast
rocy
tes
102
±60
.3N
aFl:
3.4
±1.
8In
duce
dB
BB
wit
hh
ista
min
e,de
ferr
oxam
ine
mes
ylat
e,ad
ren
alin
e,or
nor
adre
nal
ine
Gai
llar
det
al.87
FD
4:0.
9±
0.6
BB
EC
Ast
rocy
tes
352–
857
NaF
l:6.
0±
1C
ult
ure
acc.
To
Ru
bin
etal
.88an
dde
Boe
ran
dS
uta
nto
89
Gai
llar
det
al.87
FD
4:1.
5±
0.7
BB
EC
Ast
rocy
tes
145
±16
N/A
Sch
adde
lee
etal
.90
BB
EC
Ast
rocy
tes
375–
1600
Man
nit
ol:0
.483
DM
+H
EP
ES,
MO
PS,
TE
S,bi
gdi
ffer
ence
sbe
twee
ncu
ltu
reba
tch
es
Hel
ms
etal
.91
BC
EC
Ast
rocy
tes
<24
0N
/AB
albu
ena
etal
.92
BB
EC
Ast
rocy
tes
<10
00M
ann
itol
:<1
Hel
ms
and
Bro
din
76
BB
EC
Ast
rocy
tes
1177
±10
1M
ann
itol
:0.9
5±
0.1
HE
PE
S,M
OP
S,or
TE
SH
elm
set
al.52
Dig
oxin
:2.0
±0.
2E
stro
ne-
3-su
lfat
e:0.
62±
0.1
Eto
posi
de:0
.66
±0.
06B
BE
CC
6gl
iom
a16
0S
ucr
ose:
24.5
±2.
1H
igh
den
sity
ofcu
ltu
red
cell
sR
aub
etal
.93
Dex
tran
:3.3
±0.
7P
EG
:8.6
±1.
4P
orci
ne
Por
cin
ebr
ain
endo
thel
ialc
ell
(PB
EC
)
Ast
rocy
tes
358
±39
Su
cros
e:10
.3P
erm
eabi
liti
esre
port
edas
cm/m
inG
iese
etal
.94
NaF
l:7
Dic
hlo
rkyn
uer
icac
id:3
.83
MK
-801
:39.
7P
BE
CA
stro
cyte
sN
/AS
ucr
ose:
25S
chir
mac
her
and
Win
ters
95
PB
EC
Ast
rocy
tes
<11
0N
/AIn
uli
nca
lcu
late
das
perc
enta
geof
cros
sed
dose
Jeli
azko
va-M
ech
eva
and
Bob
ilya
72
PB
EC
CT
X-T
NA
2as
troc
ytes
1778
±25
2L
uci
fer
yell
ow:0
.57
±0.
046
Pu
rom
ycin
Can
tril
let
al.96
PB
EC
CT
X-T
NA
2as
troc
ytes
182
±20
N/A
No
puro
myc
inC
antr
ille
tal
.96
PB
EC
C6
glio
ma
800
Su
cros
e:3.
3±
1S
eru
m-f
ree
med
ium
wit
hh
ydro
cort
ison
eO
mid
iet
al.53
Pro
pan
olol
:18.
4±
0.7
Con
tin
ued
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2733
Tab
le1.
Con
tin
ued
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
Hu
man
hB
ME
CA
stro
cyte
s55
–297
N/A
Kas
aet
al.97
hB
MV
EC
Ast
rocy
tes
260
±13
0S
ucr
ose:
16.7
±2.
5P
erm
eabi
liti
esre
port
edas
cm/m
inM
egar
det
al.98
Caf
fein
e:50
±15
Sta
vudi
ne:
83.3
±8.
3A
nti
pyri
ne:
100
±13
.3P
ropa
nol
ol:2
00±
16.7
HB
ME
CA
stro
cyte
s60
N/A
Zys
kan
dS
chn
eide
r-W
ald63
Hu
man
brai
nva
scu
lar
endo
thel
ialc
ell
Ast
rocy
tes
700
Su
cros
e:3.
16D
IV-B
BB
wit
hfl
owC
ucu
llo
etal
.99
Hu
man
brai
nen
doth
elia
lcel
l(H
BE
C)
Ast
rocy
tes
700
Su
cros
e:0.
005
DIV
-BB
Bw
ith
flow
.P
erm
eabi
liti
esre
port
edas
cm/m
in
Cu
cull
oet
al.10
0
Man
nit
ol:0
.016
D-G
luco
se:0
.3D
ilan
tin
:0.2
5L
-asp
:0.3
Mor
phin
e:0.
009
D-a
sp:0
.008
HB
EC
Ast
rocy
tes
100
Su
cros
e:0.
33D
IV-B
BB
no
flow
.P
erm
eabi
liti
esre
port
edas
cm/m
in
Cu
cull
oet
al.10
0
Man
nit
ol:3
3.3
D-G
luco
se:1
.2D
ilan
tin
:0.2
2L
-asp
:8.3
Mor
phin
e:1.
7D
-asp
:13.
3h
BM
EC
SV
G-A
22–2
6N
aFl:
7.22
±0.
45ce
llZ
scop
e,n
ogr
owth
fact
ors
Eig
enm
ann
etal
.61
EC
V30
4A
stro
cyto
ma
1321
N1
<80
N/A
Tan
etal
.101
EaH
y929
Ast
rocy
tom
a13
21N
1<
20N
/AT
anet
al.10
1
hC
ME
C/D
3A
stro
cyte
sC
C-2
565/
SC
1810
60±
1N
/AH
ath
erel
let
al.10
2
BB
19S
VG
-A/H
BP
CT
<20
N/A
cell
Zsc
ope
Eig
enm
ann
etal
.61
TY
10S
VG
-A/H
BP
CT
<20
N/A
cell
Zsc
ope
Eig
enm
ann
etal
.61
hC
ME
C/D
3S
VG
-A/H
BP
CT
<20
NaF
l:6.
88±
0.52
cell
Zsc
ope,
no
grow
thfa
ctor
sE
igen
man
net
al.61
Con
tin
ued
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2734 REVIEW
Tab
le1.
Con
tin
ued
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
Mu
rin
eM
uri
ne
brai
nen
doth
elia
lcel
l(M
BE
C)
Ast
rocy
tes
N/A
NaF
l:25
±0.
2S
hay
anet
al.50
b.E
nd5
Ast
rocy
tom
a13
21N
1<
20N
/AT
anet
al.10
1
b.E
nd3
C6
glio
ma
130
Su
cros
e:19
.4±
2.6
Om
idie
tal
.53
Pro
pan
olol
:23.
4±
1.9
b.E
nd3
C6
glio
ma
<15
0S
ucr
ose:
16.4
±5.
2cA
mp
addi
tion
Om
idie
tal
.53
Pro
pan
olol
:23.
4±
5.9
b.E
nd3
Ast
rocy
tes
N/A
TA
MR
A:5
.9±
1.9
Lie
tal
.103
Dex
tran
10k:
0.78
±0.
24D
extr
an70
k:0.
16±
0.09
b.E
nd3
Ast
rocy
tes,
C8D
1A25
0N
/A:
BB
BB
ooth
and
Kim
104
b.E
nd3
Ast
rocy
tes,
C8D
1A25
N/A
Tra
nsw
ells
Boo
than
dK
im10
4
b.E
nd3
Ast
rocy
tes,
C8D
1A34
5N
/A:
BB
B,s
hea
rst
ress
opti
mal
at1.
2dy
n/c
m2
Boo
than
dK
im10
5
Rat
RB
EC
Ast
rocy
tes
N/A
Su
cros
e:2
DIV
-BB
Bpe
rmea
bili
ties
esti
mat
edfr
omgr
aph
sS
tan
nes
set
al.10
6
D-A
sp:2
L-A
sp:9
.3R
CE
CA
stro
cyte
sN
/AN
aFl:
4.7
Per
mea
bili
ties
repo
rted
ascm
/min
Kis
etal
.64
Eva
ns
blu
eal
bum
in:�
0.75
RB
EC
Ast
rocy
tes
N/A
Su
cros
e:7.
5±
1P
erm
eabi
liti
esre
port
edas
cm/m
inB
lasi
get
al.10
7
NaF
l:11
±1.
3D
ich
lork
ynu
ren
icac
id:1
1±
1.5
RB
EC
Ast
rocy
tes
130
±7
N/A
EC
GF
+20
%P
DS.
Per
mea
bili
ties
repo
rted
ascm
/min
Dem
euse
etal
.108
RB
ME
CA
stro
cyte
s40
0–45
0N
/AK
riza
nac
-Ben
gez
etal
.109
RB
ME
CA
stro
cyte
sN
/AS
ucr
ose:
9.9
Kri
zan
ac-B
enge
zet
al.11
0
Con
tin
ued
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2735T
able
1.C
onti
nu
ed
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
RB
EC
Ast
rocy
tes
<12
5N
aFl:
�4
App
roxi
mat
ion
sof
Pe
valu
esfr
omta
bles
Nak
agaw
aet
al.11
1
RB
EC
Ast
rocy
tes
<16
0N
aFl:
�4
Nak
agaw
aet
al.68
RB
EC
Ast
rocy
tom
a,13
21N
1<
20N
/AT
anet
al.10
1
Bov
ine
Bov
ine
len
sen
doth
elia
lcel
lP
eric
ytes
150
±17
0L
uci
fer
yell
ow:1
0.1
±2.
5E
ndo
thel
ialc
ells
deri
ved
from
hem
atop
oiet
icst
emce
lls.
Per
mea
bili
ties
repo
rted
ascm
/min
Cec
chel
liet
al.11
2
Hu
man
hB
ME
CP
eric
ytes
26±
0.16
N/A
cell
Zsc
ope
Eig
enm
ann
etal
.61
hC
ME
C/D
3H
BV
P46
±1.
1N
/AH
ath
erel
let
al.10
2
Rat
RB
EC
Per
icyt
es43
8±
75N
/A1:
mpo
res
Dem
euse
etal
.108
RB
EC
Per
icyt
es13
0±
7N
/A0.
45:
mpo
res
Dem
euse
etal
.108
RB
EC
Per
icyt
es97
.5±
8.5
N/A
Nor
mal
ized
valu
esfo
rpe
rmea
bili
tyH
ayas
hie
tal
.113
RB
EC
Per
icyt
es<
200
NaF
l:�
3.4
App
roxi
mat
ion
sof
Pe
valu
esfr
omta
bles
Nak
agaw
aet
al.11
1
RB
EC
Per
icyt
es<
230
NaF
l:�
4.1
Nak
agaw
aet
al.68
RB
E4
Per
icyt
es<
200
Su
cros
e:<
25P
erm
eabi
liti
esre
port
edas
cm/m
inA
lAh
mad
etal
.49
Dex
tran
:<3
Non
-con
tact
cocu
ltu
reB
ovin
eB
BE
CA
stro
cyte
sN
/AS
ucr
ose:
8.5
±0.
7P
erm
eabi
liti
esre
port
edas
cm/m
inH
amm
etal
.114
Inu
lin
:2.8
±0.
3B
BE
CA
stro
cyte
s66
1±
48N
/AD
ehou
cket
al.62
BB
EC
Ast
rocy
tes
500–
800
Su
cros
e:3.
8G
arbe
rget
al.11
5
Ala
nin
e:15
.2A
nti
pyri
ne:
54.0
AZ
T:5
.7C
affe
ine:
60.2
Cim
etid
ine:
4.0
Cyc
losp
orin
e:4.
6D
iaze
pam
:38.
0D
igox
in:3
.1L-D
opa:
16.3
Gly
cero
l:6.
8In
uli
n:0
.67
Lac
tic
acid
:9.3
Leu
cin
e:13
.0M
orph
ine:
22.0
Nic
otin
e:66
.0P
hen
ytoi
n:5
0.3
Ure
a:26
.3V
erap
amil
:23.
4V
inbl
asti
ne:
11.6
Vin
cris
tin
e:2.
3W
arfa
rin
:17.
9
Con
tin
ued
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2736 REVIEW
Tab
le1.
Con
tin
ued
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
BA
EC
C6
736
±0.
38S
ucr
ose:
0.08
8D
IV-B
BB
hol
low
fibe
rS
tan
nes
set
al.11
6
Th
eoph
ylli
ne:
1.88
Mor
phin
e:0.
05B
BE
CC
6<
200
Su
cros
e:<
10P
erm
eabi
liti
esre
port
edas
cm/m
inA
bbru
scat
oan
dD
avis
117
BA
EC
C6
<50
0D
IV-B
BB
Cu
cull
oet
al.11
8
BC
EC
C6
<30
00N
/AR
O20
–172
4(p
roin
flam
mat
ory)
Zen
ker
etal
.47
BC
EC
C6
2000
N/A
Zen
ker
etal
.47
BB
EC
C6
100
Su
cros
e:8.
1G
arbe
rget
al.11
5
Ala
nin
e:10
.1In
uli
n:0
.6L
euci
ne:
9.0
Vin
cris
tin
e:3.
7B
AE
CC
665
0±
26.5
Su
cros
e:5.
34±
0.23
DIV
-BB
BS
anta
guid
aet
al.11
9
Ph
enyt
oin
:1.4
6±
0.17
Por
cin
eP
BE
CA
stro
cyte
s21
4±
22N
/AG
iese
etal
.94
PB
EC
Ast
rocy
tes
<78
0M
ann
itol
:<1
CP
T-c
AM
P,R
O20
–172
4an
dh
ydro
cort
ison
etr
eate
dT
ran
swel
ls
Pat
aben
dige
etal
.82
PB
EC
C6
834
±13
6S
ucr
ose:
1.6
±1.
0S
eru
m-f
ree
med
ium
,wit
hsu
pple
men
tof
hyd
roco
rtis
one
Sm
ith
etal
.66
Pro
pan
olol
:18.
4±
0.7
PB
EC
C6
352
±34
N/A
Sm
ith
etal
.66
Hu
man
hB
ME
CA
stro
cyte
s<
500
N/A
Per
cen
tage
su
sed
for
perm
eabi
liti
esS
iddh
arth
anet
al.27
Hu
man
plu
ripo
ten
tst
emce
lls
Ast
rocy
tes
860
±26
0In
uli
n:0
.029
±0.
02H
um
anpl
uri
pote
nt
stem
cell
-der
ived
endo
thel
ial
cell
s
Lip
pman
net
al.59
Su
cros
e:0.
034
±0.
015
Glu
cose
:0.2
2±
0.07
Vin
cris
tin
e:0.
062
±0.
02C
olch
icin
e:0.
092
±0.
1P
razo
sin
:0.2
9±
0.04
Dia
zepa
m:1
.1±
0.5
Hu
man
prim
ary
brai
nen
doth
elia
lce
lls
Ast
rocy
tes
N/A
Gar
berg
etal
.115
Con
tin
ued
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329
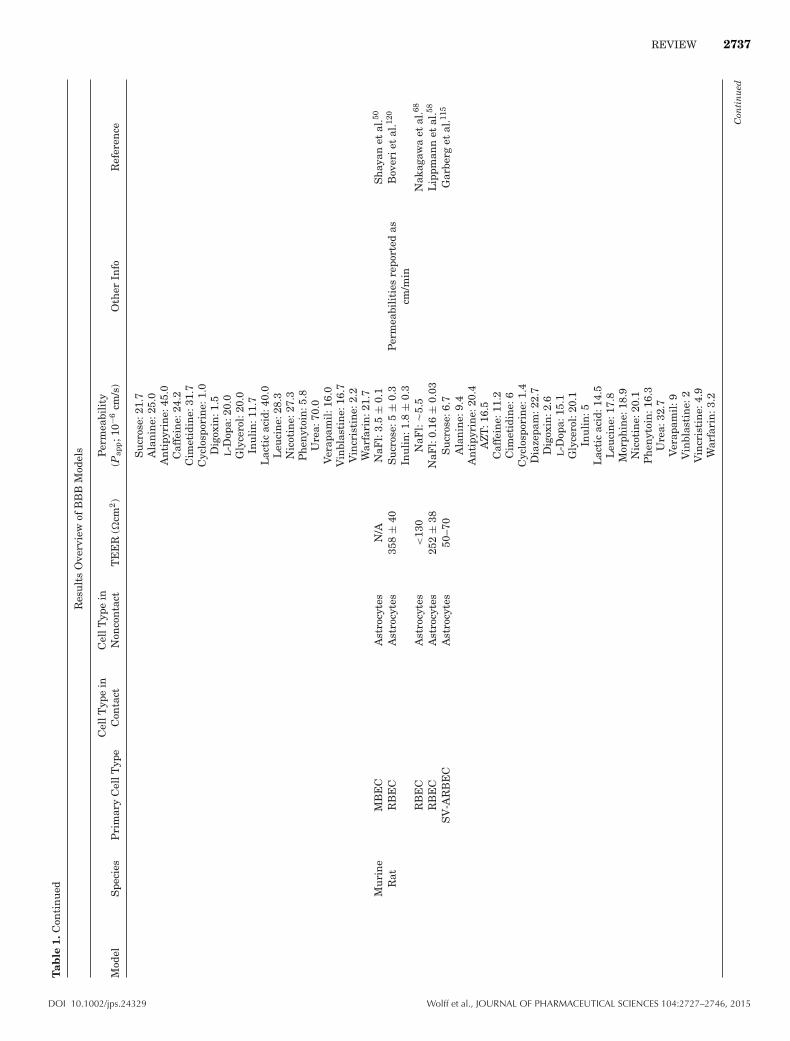
REVIEW 2737T
able
1.C
onti
nu
ed
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
Su
cros
e:21
.7A
lan
ine:
25.0
An
tipy
rin
e:45
.0C
affe
ine:
24.2
Cim
etid
ine:
31.7
Cyc
losp
orin
e:1.
0D
igox
in:1
.5L-D
opa:
20.0
Gly
cero
l:20
.0In
uli
n:1
1.7
Lac
tic
acid
:40.
0L
euci
ne:
28.3
Nic
otin
e:27
.3P
hen
ytoi
n:5
.8U
rea:
70.0
Ver
apam
il:1
6.0
Vin
blas
tin
e:16
.7V
incr
isti
ne:
2.2
War
fari
n:2
1.7
Mu
rin
eM
BE
CA
stro
cyte
sN
/AN
aFl:
3.5
±0.
1S
hay
anet
al.50
Rat
RB
EC
Ast
rocy
tes
358
±40
Su
cros
e:5
±0.
3P
erm
eabi
liti
esre
port
edas
cm/m
inB
over
iet
al.12
0
Inu
lin
:1.8
±0.
3R
BE
CA
stro
cyte
s<
130
NaF
l:�
5.5
Nak
agaw
aet
al.68
RB
EC
Ast
rocy
tes
252
±38
NaF
l:0.
16±
0.03
Lip
pman
net
al.58
SV
-AR
BE
CA
stro
cyte
s50
–70
Su
cros
e:6.
7G
arbe
rget
al.11
5
Ala
nin
e:9.
4A
nti
pyri
ne:
20.4
AZ
T:1
6.5
Caf
fein
e:11
.2C
imet
idin
e:6
Cyc
losp
orin
e:1.
4D
iaze
pam
:22.
7D
igox
in:2
.6L-D
opa:
15.1
Gly
cero
l:20
.1In
uli
n:5
Lac
tic
acid
:14.
5L
euci
ne:
17.8
Mor
phin
e:18
.9N
icot
ine:
20.1
Ph
enyt
oin
:16.
3U
rea:
32.7
Ver
apam
il:9
Vin
blas
tin
e:2
Vin
cris
tin
e:4.
9W
arfa
rin
:3.2
Con
tin
ued
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2738 REVIEW
Tab
le1.
Con
tin
ued
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
RB
EC
C6
239
±51
Su
cros
e:11
.5±
3.3
Per
mea
bili
ties
repo
rted
ascm
/min
Bov
erie
tal
.120
Inu
lin
:4.3
±0.
3B
ovin
eB
ME
CP
eric
ytes
4450
±17
0N
/AN
PC
-der
ived
BM
EC
.M
odifi
edE
Cm
ediu
m(n
oex
ogen
ous
bFG
F)
Lip
pman
net
al.60
Rat
RB
EC
Per
icyt
es19
.8±
2.6
N/A
Nor
mal
ized
valu
esfo
rpe
rmea
bili
tyH
ayas
hie
tal
.113
RB
EC
Per
icyt
es<
150
NaF
l:�
4.5
Per
mea
bili
ties
repo
rted
ascm
/min
Nak
agaw
aet
al.68
Bov
ine
BM
EC
NP
Cs
2940
±80
0S
ucr
ose:
0.53
–0.5
7±
0.17
Tra
nsw
ellw
ith
NP
Cs
and
NP
C-d
eriv
edB
ME
CL
ippm
ann
etal
.60
BM
EC
NP
Cs
5160
±32
0N
/AN
PC
-der
ived
BM
EC
.M
odifi
edE
Cm
ediu
m(n
oex
ogen
ous
bFG
F)
Lip
pman
net
al.60
Rat
RB
EC
NP
Cs
110
±5
NaF
l:5.
5±
0.8
Wei
den
fell
eret
al.12
1
RB
EC
NP
Cs
246
±19
NaF
l:0.
170
±0.
05�
20%
fray
edT
JsL
ippm
ann
etal
.58
Tri
ple-
cult
ure
Rat
RB
E4
Ast
rocy
tes
Neu
robl
asto
ma
SH
-SY
5Y<
450
N/A
Bal
buen
aet
al.12
2
RB
EC
Ast
rocy
tes
Neu
ron
s26
8.67
N/A
Per
mea
bili
ties
inpe
rcen
tage
s,an
dre
lati
ven
um
bers
Xu
eet
al.12
3
Hu
man
hC
ME
C/D
3A
stro
cyte
sC
C-
2565
/SC
1810
Per
icyt
es44
±0.
9N
/AH
ath
erel
let
al.10
2
Rat
RB
EC
Ast
rocy
tes
Per
icyt
es�
270
NaF
l:�
3.5
App
roxi
mat
ion
sof
Pe
valu
esfr
omta
bles
Nak
agaw
aet
al.11
1
RB
EC
Ast
rocy
tes
Per
icyt
es�
260
NaF
l:�
4N
akag
awa
etal
.68
Hu
man
hC
ME
C/D
3H
BV
PA
stro
cyte
s42
±0.
8N
/AH
ath
erel
let
al.10
2
Con
tin
ued
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2739
Tab
le1.
Con
tin
ued
Res
ult
sO
verv
iew
ofB
BB
Mod
els
Mod
elS
peci
esP
rim
ary
Cel
lTyp
eC
ellT
ype
inC
onta
ctC
ellT
ype
inN
onco
nta
ctT
EE
R(�
cm2)
Per
mea
bili
ty(P
app;1
0−6
cm/s
)O
ther
Info
Ref
eren
ce
Rat
RB
EC
Per
icyt
esA
stro
cyte
s38
8±
18.8
NaF
l:�
4A
ppro
xim
atio
ns
ofP
eva
lues
from
tabl
esN
akag
awa
etal
.111
RB
EC
Per
icyt
esA
stro
cyte
s35
4±
15N
aFl:
3.9
±0.
2A
ppro
xim
atio
ns
ofP
eva
lues
offr
omta
ble
Nak
agaw
aet
al.68
Ver
apam
il:2
3P
razo
sin
:23
Qu
inid
ine:
7.3
Vin
cris
tin
e:6
Hyd
roco
rtis
one:
5.6
Su
lpir
ide:
4.5
Vin
blas
tin
e:3.
1C
imet
idin
e:3
Ate
nol
ol:2
.4E
pin
asti
ne:
1.9
Dig
oxin
:0.4
Pro
pan
olol
:200
0H
ydro
xyzi
ne:
1200
Caf
fein
e:50
0T
razo
don
e:33
0P
hen
ytoi
n:3
20A
nti
pyri
n:2
20C
arba
maz
epin
e:19
5Z
olpi
dem
:150
Mu
ltic
ult
ure
Rat
RB
E4
Ast
rocy
tes
and
peri
cyte
s28
2±
45S
ucr
ose:
<26
Per
mea
bili
ties
repo
rted
ascm
/min
AlA
hm
adet
al.49
Dex
tran
:<3
Wit
hex
tra
info
rmat
ion
ondr
ugs
and
any
devi
atio
nfr
omst
anda
rdpr
otoc
olis
give
nu
nde
r“o
ther
info
.”V
alu
esh
ave
been
reca
lcu
late
din
toth
esa
me
un
itfo
rbe
tter
com
pari
son
.
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015
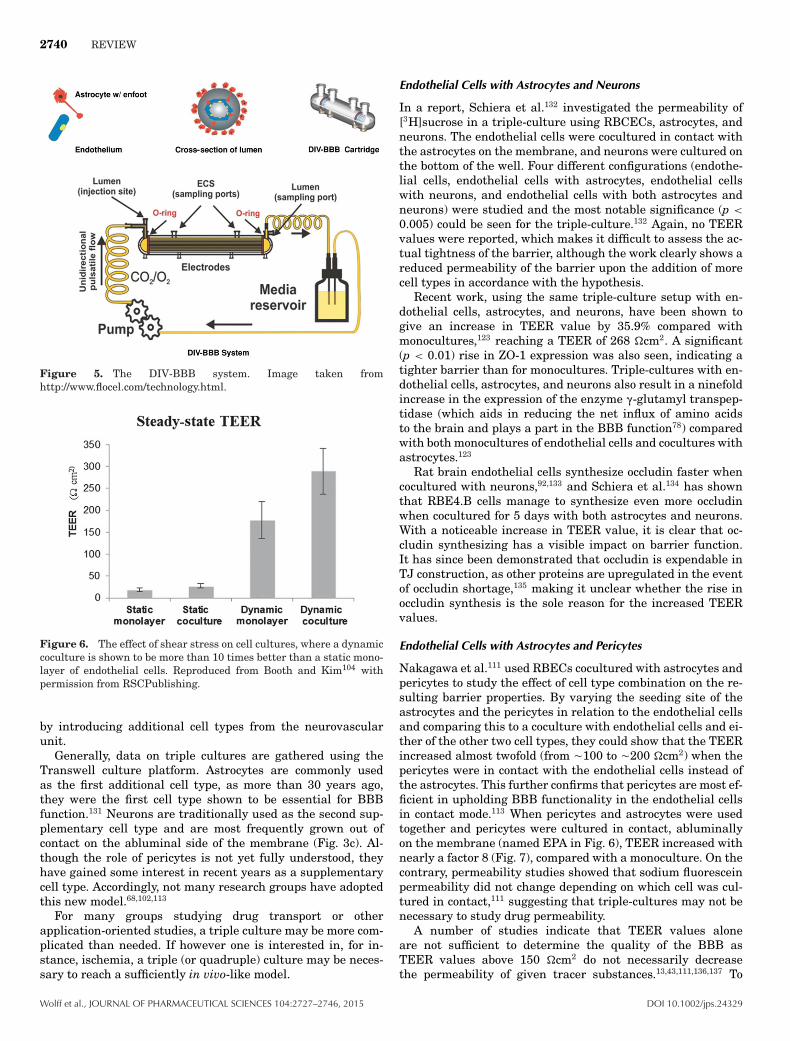
2740 REVIEW
Figure 5. The DIV-BBB system. Image taken fromhttp://www.flocel.com/technology.html.
Figure 6. The effect of shear stress on cell cultures, where a dynamiccoculture is shown to be more than 10 times better than a static mono-layer of endothelial cells. Reproduced from Booth and Kim104 withpermission from RSCPublishing.
by introducing additional cell types from the neurovascularunit.
Generally, data on triple cultures are gathered using theTranswell culture platform. Astrocytes are commonly usedas the first additional cell type, as more than 30 years ago,they were the first cell type shown to be essential for BBBfunction.131 Neurons are traditionally used as the second sup-plementary cell type and are most frequently grown out ofcontact on the abluminal side of the membrane (Fig. 3c). Al-though the role of pericytes is not yet fully understood, theyhave gained some interest in recent years as a supplementarycell type. Accordingly, not many research groups have adoptedthis new model.68,102,113
For many groups studying drug transport or otherapplication-oriented studies, a triple culture may be more com-plicated than needed. If however one is interested in, for in-stance, ischemia, a triple (or quadruple) culture may be neces-sary to reach a sufficiently in vivo-like model.
Endothelial Cells with Astrocytes and Neurons
In a report, Schiera et al.132 investigated the permeability of[3H]sucrose in a triple-culture using RBCECs, astrocytes, andneurons. The endothelial cells were cocultured in contact withthe astrocytes on the membrane, and neurons were cultured onthe bottom of the well. Four different configurations (endothe-lial cells, endothelial cells with astrocytes, endothelial cellswith neurons, and endothelial cells with both astrocytes andneurons) were studied and the most notable significance (p <
0.005) could be seen for the triple-culture.132 Again, no TEERvalues were reported, which makes it difficult to assess the ac-tual tightness of the barrier, although the work clearly shows areduced permeability of the barrier upon the addition of morecell types in accordance with the hypothesis.
Recent work, using the same triple-culture setup with en-dothelial cells, astrocytes, and neurons, have been shown togive an increase in TEER value by 35.9% compared withmonocultures,123 reaching a TEER of 268 �cm2. A significant(p < 0.01) rise in ZO-1 expression was also seen, indicating atighter barrier than for monocultures. Triple-cultures with en-dothelial cells, astrocytes, and neurons also result in a ninefoldincrease in the expression of the enzyme (-glutamyl transpep-tidase (which aids in reducing the net influx of amino acidsto the brain and plays a part in the BBB function78) comparedwith both monocultures of endothelial cells and cocultures withastrocytes.123
Rat brain endothelial cells synthesize occludin faster whencocultured with neurons,92,133 and Schiera et al.134 has shownthat RBE4.B cells manage to synthesize even more occludinwhen cocultured for 5 days with both astrocytes and neurons.With a noticeable increase in TEER value, it is clear that oc-cludin synthesizing has a visible impact on barrier function.It has since been demonstrated that occludin is expendable inTJ construction, as other proteins are upregulated in the eventof occludin shortage,135 making it unclear whether the rise inoccludin synthesis is the sole reason for the increased TEERvalues.
Endothelial Cells with Astrocytes and Pericytes
Nakagawa et al.111 used RBECs cocultured with astrocytes andpericytes to study the effect of cell type combination on the re-sulting barrier properties. By varying the seeding site of theastrocytes and the pericytes in relation to the endothelial cellsand comparing this to a coculture with endothelial cells and ei-ther of the other two cell types, they could show that the TEERincreased almost twofold (from �100 to �200 �cm2) when thepericytes were in contact with the endothelial cells instead ofthe astrocytes. This further confirms that pericytes are most ef-ficient in upholding BBB functionality in the endothelial cellsin contact mode.113 When pericytes and astrocytes were usedtogether and pericytes were cultured in contact, abluminallyon the membrane (named EPA in Fig. 6), TEER increased withnearly a factor 8 (Fig. 7), compared with a monoculture. On thecontrary, permeability studies showed that sodium fluoresceinpermeability did not change depending on which cell was cul-tured in contact,111 suggesting that triple-cultures may not benecessary to study drug permeability.
A number of studies indicate that TEER values aloneare not sufficient to determine the quality of the BBB asTEER values above 150 �cm2 do not necessarily decreasethe permeability of given tracer substances.13,43,111,136,137 To
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2741
TE
ER
(cm
2 )
Days
0
2 3 4 5
TE
ER
(cm
2 )
0
5
10
0Days
a, b
a, b, c
a
a, b, c
a, b, c, f
b, c, d, e100
200
300
2 3 4
E00
EP0
EA0
E0P
E0A
EPA
EAP
0
400
000
0P0
0A0
5
Figure 7. The effect of different co- and triple-cultures on a BBB model using Transwells. With an inset graph showing that neither astrocytesor pericytes have any relevant resistances. E00, a monoculture of endothelial cells. E0A, noncontact coculture with astrocytes. EA0, contactcoculture with astrocytes. E0P, noncontact coculture with pericytes. EP0, contact coculture with pericytes. EAP, triple-culture with astrocytesin contact and pericytes in noncontact. EPA, triple culture with pericytes in contact and astrocytes in noncontact. Reproduced from Nakagawaet al.111 with permission from Springer US.
Figure 8. Graph of the correlation between permeability coefficientsof drugs in a triple culture in the EPA setup (in vitro), and the apparentpermeability coefficients of the same drugs in animal models (in vivo).Reproduced from Nakagawa et al.68 with permission from Elsevier B.V.
evaluate their previous work, Nakagawa et al.68 comparedthe absorption of 19 compounds with their known in vivovalues using the same seven models as in the 2007 exper-iment in a new study published in 2009. This study, witha larger number of trials, showed the highest TEER for thetriple-culture with the EPA setup, with a reduced perme-ability in regard to a monoculture of endothelial cells. Thegroup also showed that the expression of TJ proteins wasincreased by the presence of astrocytes or pericytes but in-creased most in the EPA model. Permeabilities of low Pe
substances remained low in the EPA model at 0.44–3.51×10−6 cm/s. When compared with in vivo values, a correlation ofR2 = 0.89 was found, (Fig. 8). When evaluated together, the two
studies68,111 give a good overall image of their model with manyof the parameters that have bearing on the tightness of thebarrier.
Hatherell et al.102 repeated Nakagawa’s experiment, usinghCMEC/D3 cocultured with human pericytes and astrocytes.Their obtained TEER values differed noticeably from Naka-gawa’s results with the endothelium/astrocyte culture result-ing in the highest TEER values, even compared with the twocombinations of triple cultures (Fig. 9). This effectively con-firms that even though two cell types may be similar in func-tion, they can still differ widely in application122 and thatspecies-specific parameters have substantial impact on barrierbehavior.39
Using RBE4, Al Ahmad et al.49 analyzed the stability oftriple cultures upon anoxic events. They showed that the pres-ence of astrocytes and pericytes in a triple-culture was theonly means of maintaining the original TEER values even48 h after oxygen deprivation. The triple-culture also gavethe lowest increase in permeability of sucrose (2.81 ± 0.81× 10−3 cm/s as opposed to 10.6 ± 1.28 × 10−3 cm/s for amonoculture) 48 h after the anoxic event. The mechanism bywhich the astrocytes and pericytes promoted the survival of theREB4 cells during anoxia was determined to be by caspase-3inhibition.
DISCUSSION ON CURRENT STATE-OF-THE-ART ANDNEW TRENDS
As mentioned before, endothelial cells from different speciesmay share many BBB attributes at a cellular or molecular levelbut they can still react differently in experimental setups.122
Given this, it is truly difficult to assess a model without testingit for all types of relevant endothelial cells; a model might workwell for one endothelial cell but not for another. Minor modifi-cations and additions to models can result in widely differentresults, as apparent in Table 1. In the author’s opinion, the
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2742 REVIEW
30
35
40
45
50
55
60
65
70
10987654321
TE
ER
(Ω
cm
2 )
Time (days)
A
B C
E FG H
D
*
hCMEC/D3 mono-cultivation (A) hCMEC/D3 and CC-2565 co-cultivation (B)
hCMEC/D3 and SC-1810 co-cultivation (C) hCMEC/D3 and HBVP co-cultivation (D)
hCMEC/D3, CC-2565, and HBVP tri-cultivation (E) hCMEC/D3, SC-1810, and HBVP tri-cultivation (F)
hCMEC/D3, HBVP, and CC-2565 tri-cultivation (G) hCMEC/D3, HBVP, and SC-1810 tri-cultivation (H)
Figure 9. The effect of co- and triple-cultures on hCMEC/D3 and twodifferent astrocyte cell lines (CC-2565 and SC1810) and one pericytecell line (HBVP) in three independent experiments. The asterisk marksthe most significant points compared with a monoculture (p < 0.001).Reprinted from Hatherell et al.102 with permission from Elsevier B.V.
field would benefit greatly from commonly accepted standardsfor permeability measurements, TEER, fluorescence, and otherquantitative evaluations.
Another area of the BBB in vitro research field with high po-tential is the development of models based on pluripotent stemcells, or neural progenitor cells (NPC). This approach couldenable human in vitro model systems with a much higher ex-pected prediction rate compared with animal-derived systems.In work from 2011, Lippmann et al.58 differentiated NPC forup to 12 days to reach a ratio of 3:1 of astrocytes and neu-rons (Fig. 10), which were then seeded on the bottom of aTranswell. The membrane cup with rat brain microvessel en-dothelial cells (RBMECs) was added to the well and TEER val-ues were monitored for 5 days. Their optimized NPC-derivedcultures increased membrane resistance by up to 200% com-pared with endothelial cell monoculture,58 raising TEER to avalue of approximately 250 �cm2. This work builds upon previ-ous research, where a controlled differentiation of NPC aimedat in vitro BBB models were presented already in 2007.121
The group has been further developing their work, and in2012 they showed that human pluripotent cells that were cod-ifferentiated with neural cells produced endothelial cells withBBB properties, such as TJ expression and presence of rel-evant nutrient transporters.59 The stem cells also respondedto astrocytes in cocultures, giving relevant TEER values(1450 �cm2) and a permeability of several compounds withinacceptable levels of the in vitro model. In 2014, additional im-provements were presented where TEER values greater than5000 �cm2 were achieved by first enhancing the BBB pheno-type in the HBMECs using retinoic acid.60 This is the highestpublished TEER value to date from any in vitro BBB model,indicating that triple-cultures are superior to mono- and cocul-tures with respect to obtaining high-barrier resistances. It is,
Figure 10. Immunofluorescent images of NPC cultured for 12 dayswith 10% fetal bovine serum (a) astrocytes labeled with GFAP (red) ornestin (green) (b) and neurons labeled for $III-tubulin (red) or nestin(green). Scale bars indicate 50 :m and nestin labels undifferentiatedNPC. Reproduced from Lippmann et al.58 with permission from WileyInc.
however, not completely clear whether triple-cultures are nec-essary to achieve reliable in vitro data because several otherreports are available (Table 1), where high TEER values arereached without using triple cultures. Using NPC instead ofmature astrocytes and neurons is, however, not a simple taskand the protocol for reaching the correct ratio of astrocytesand neurons is in itself a major undertaking. So far, only EricShusta’s group has worked with NPC cultures in this fashion.
For drug delivery studies, permeability is the most inter-esting performance factor to evaluate an in vitro model upon.Although many researchers are active in this field, few studiesuse the same test substances, making it hard to ascertain thetrue permeability ratio between the different models and celltypes. Even so, a BBB model with shear stress75 or a model withNPC-derived astrocytes and neurons100,116 gives the lowest re-ported permeabilities for both mannitol and sucrose. However,too little data exist regarding membrane permeability and pro-tein expression to claim superiority over other models. Lipp-mann et al.127 show a low permeability for sucrose at 0.57 ±0.12 × 10−6 cm/s in their coculture with cells derived from NPCand Cucullo et al.100 show sucrose at 0.29 × 10−6 cm/s with
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2743
their DIV-BBB, while also showing low-permeability values forsmaller molecules such as mannitol. In this particular compar-ison, the data clearly demonstrate how low permeabilities canbe found with relatively low TEER values and higher perme-abilities for a system with high TEER values, which makes itimpossible to correlate resistance to permeability. As microflu-idic devices today do not yet measure permeabilities, it is im-possible to evaluate them properly.
Most studies published to date on in vitro BBB research areperformed using the Transwell cell culture systems, but duringthe last 10 years, a major increase of interest in microchip BBBmodels has been seen.84,85,104,127,138,139 The motivation for this isthe fact that the capillaries in the microvasculature are as smallas 7–10 :m in diameter with an average intercapillary distanceof 40 :m140,141 and an average distance to the nearest neuronof only 10–20 :m.142 The standard semipermeable membraneof the Transwells has a thickness of 10 :m, which is more than300 times thicker than the naturally occurring basement mem-brane of the endothelial cells. A system developed in the samescale as the neurovascular unit is expected to more closely re-semble the in vivo situation and in turn result in in vivo-likebarrier properties. Most of the studies published on microfluidicBBB systems have not yet reached beyond the proof-of-conceptstage and the reported TEER values and permeability valuesare often of a lower standard than the Transwell-based sys-tems. It is likely several more articles in this highly interestingresearch area will be published in the coming years, with greatpotential for the BBB research field.
CONCLUSIONS
This review article has presented some of the most influentialand most recent work regarding in vitro BBB models to date.Monocultures, cocultures, and triple-cultures of the major con-stituent cell types of the neurovascular unit have been investi-gated to better understand the important aspects of achieving ahigh-fidelity model of the BBB. Different setups, ranging fromthe classical Transwells to more novel approaches using mi-crofluidics, have also been studied.
This review article further set out to evaluate the perfor-mance of cell-based in vitro systems of the BBB. It was foundto be very difficult to evaluate the performance of the plethoraof different models used because of the lack of coherence inthe research field. The authors therefore call for a more stan-dardized method to evaluate the performance of the differentin vitro systems developed as an effort to jointly bring the re-search field forward. It would also be interesting to see moreresearch groups taking an interdisciplinary approach to the is-sue as both the biological as well as the more technical aspectsof the culture platform are likely to be highly influential on thefinal in vitro performances.
REFERENCES
1. Olesen J, Gustavsson A, Svensson M, Wittchen H-U, Jonsson B. 2012.The economic cost of brain disorders in Europe. Eur J Neurol 19:155–162.2. Wimo A, Jonsson L, Bond J, Prince M, Winblad B. 2013.3. Ehrlich P. 1885. Das Sauerstoff-Bedurfniss des Organismus – einefarbenanalytische Studie.4. Lewandowsky M. 1900. Zur lehre der cerebrospinalflussigkeit.Zeitschrift Klin Medizin 40:480–494.
5. Goldman EE. 1913. Vitalfarbung am zentralnervensystem:beitragzur physio-pathologie des plexus chorioideus und der hirnhaute. Ab-handlungen der Preuss. Akad. der Wissenschaften Philos Klasse 1.6. Reese TS, Karnovsky MJ. 1967. Fine structural localization ofa blood–brain barrier to exogenous peroxidase. J Cell Biol 34:207–217.7. Meyvantsson I, Beebe DJ. 2008. Cell culture models in microfluidicsystems. Annu Rev Anal Chem 1:423–449.8. Abbott N, Dolman D, Yusof S, Reichel A. 2014. In vitro models ofCNS barriers. Drug Deliv Brain.10:163–197.9. Aschner M, Fitsanakis VA, Paula A, Olivi L, Bressler JP. 2006.Blood–brain barrier and cell–cell interactions methods for establishingin vitro models of the blood–brain barrier and transport measurements.Methods Mol Biol 341:1–15.10. Cardoso FL, Brites D, Brito MA. 2010. Looking at the blood–brainbarrier: Molecular anatomy and possible investigation approaches.Brain Res Rev 64:328–363.11. Wilhelm I, Fazakas C, Krizbai IA. 2011. In vitro models of theblood-brain barrier. Acta Nerobiologiae Exp 71:113–128.12. Bicker J, Alves G, Fortuna A, Falcao A. 2014. Blood–brain barriermodels and their relevance for a successful development of CNS drugdelivery systems: A review. Eur J Pharm Biopharm 87:409–432.13. Deli Ma, Abraham CS, Kataoka, Y, Niwa M. 2005. Permeabilitystudies on in vitro blood–brain barrier models: Physiology, pathology,and pharmacology. Cell Mol Neurobiol 25:59–127.14. Abbott NJ, Patabendige AaK, Dolman DEM, Yusof SR, Begley DJ.2010. Structure and function of the blood–brain barrier. Neurobiol Dis37:13–25.15. Hawkins B, Davis T. 2005. The blood–brain barrier/neurovascularunit in health and disease. Pharmacol Rev 57:173–185.16. de Boer AG, Gaillard PJ. 2006. Blood–brain barrier dysfunction andrecovery. J Neural Transm 113:455–462.17. Chaudhuri JD. 2000. Blood brain barrier and infection. Med SciMonit 6:1213–1222.18. Bagley RG, Weber W, Rouleau C, Teicher BA. 2005. Pericytes andendothelial precursor cells: Cellular interactions and contributions tomalignancy. Cancer Res 65:9741–9750.19. Scherrmann JM. 2002. Drug delivery to brain via the blood–brainbarrier. Vascul Pharmacol 38:349–354.20. Pardridge W. 1999. Blood–brain barrier biology and methodology.J Neurovirol 5:556–569.21. Petty Ma, Lo EH. 2002. Junctional complexes of the blood–brainbarrier: Permeability changes in neuroinflammation. Prog Neurobiol68:311–323.22. Schneeberger EE. 2003. Claudin extracellular domains determineparacellular charge selectively and resistance but not tight junctionfibril architecture. Am J Physiol 284:1331–1333.23. Balda M, Flores-Maldonado C, Cereijido M, Matter K. 2000. Mul-tiple domains of occludin are involved in the regulation of paracellularpermeability. J Cell Biochem 78:85–96.24. Huber JD, Egleton RD, Davis TP. 2001. Molecular physiology andpathophysiology of tight junctions in the blood–brain barrier. TrendsNeurosci 24:719–725.25. Vorbrodt A, Dobrogowska D. 2003. Molecular anatomy of intercel-lular junctions in brain endothelial and epithelial barriers: Electronmicroscopist’s view. Brain Res Rev 42:221–242.26. Hurley R, Hayman A, Taber K. 2008. Astroglia: Not just glue. JNeuropsychiatry Clin Neurosci 20:iv–129.27. Siddharthan V, Kim YV, Liu S, Kim KS. 2007. Humanastrocytes/astrocyte-conditioned medium and shear stress enhance thebarrier properties of human brain microvascular endothelial cells.Brain Res 1147:39–50.28. Colgan OC, et al. 2008. Influence of basolateral condition on theregulation of brain microvascular endothelial tight junction propertiesand barrier function. Brain Res 1193:84–92.29. Zheng W, Aschner M, Ghersi-Egea JF. 2003. Brain barrier sys-tems: A new frontier in metal neurotoxicological research. Toxicol ApplPharmacol 192:1–11.
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2744 REVIEW
30. Kim JA, et al. 2006. Brain endothelial hemostasis regulation bypericytes. J Cereb Blood Flow Metab 26:209–217.31. Allt G, Lawrenson JG. 2001. Pericytes: Cell biology and pathology.Cells Tissues Organs 169:1–11.32. Lai CH, Kuo KH. 2005. The critical component to establish in vitroBBB model: Pericyte. Brain Res Rev 50:258–265.33. Armulik A. et al. 2010. Pericytes regulate the blood–brain barrier.Nature 468:557–561.34. Thanabalasundaram G, Schneidewind J, Pieper C, Galla H-J. 2011.The impact of pericytes on the blood–brain barrier integrity dependscritically on the pericyte differentiation stage. Int J Biochem Cell Biol43:1284–1293.35. Daneman R, Zhou L, Kebede Aa, Barres Ba. 2010. Pericytes are re-quired for blood–brain barrier integrity during embryogenesis. Nature468:562–566.36. Dore-Duffy P, Katychev A, Wang X, VanBuren E. 2006. CNS mi-crovascular pericytes exhibit multipotential stem cell activity. J CerebBlood Flow Metab 26:613–624.37. Minami M. 2011. Neuro–glio–vascular interaction in ischemicbrains. Yakugaku zasshi 131:539–544.38. Sumi N, et al. 2010. Lipopolysaccharide-activated microglia inducedysfunction of the blood–brain barrier in rat microvascular endothelialcells co-cultured with microglia. Cell Mol Neurobiol 30:247–253.39. Perel P, et al. 2007. Comparison of treatment effects between ani-mal experiments and clinical trials: Systematic review. BMJ 334:197.40. Crone C, Olesen SP. 1982. Electrical resistance of brain microvas-cular endothelium. Brain Res 241:49–55.41. Smith QR, Rapoport SI. 1986. Cerebrovascular permeability coeffi-cients to sodium, potassium, and chloride. J Neurochem 46:1732–1742.42. Butt AM, Jones HC. 1992. Effect of histamine and antagonists onelectrical resistance across the blood–brain barrier in rat brain-surfacemicrovessels. Brain Res 569:100–105.43. Reichel A, Begley DJ, Abbott NJ. 2003. An overview of in vitrotechniques for blood–brain barrier studies. Methods Mol Med 89:307–324.44. Haseloff RF, Dithmer S, Winkler L, Wolburg H, Blasig, IE. 2014.Transmembranal proteins of the tight junctions at the blood–brainbarrier: Structural and functional aspects. Semin Cell Dev Biol.[Epubahead of print]45. Ohno K, Pettigrew KD, Rapoport SI. 1978. Lower limits of cere-brovascular permeability to nonelectrolytes in the conscious rat. Am JPhysiol 235:H299–H307.46. Brendel K, Meezan, E, Carlson E. 1974. Isolated brain microves-sels: A purified, metabolically active preparation from bovine cerebralcortex. Science 185:953–955.47. Zenker D, Begley D, Bratzke H, Rubsamen-Waigmann H, vonBriesen H. 2003. Human blood-derived macrophages enhance barrierfunction of cultured primary bovine and human brain capillary en-dothelial cells. J Physiol 551:1023–1032.48. Weidenfeller C, Svendsen CN, Shusta EV. 2007. Differentiating em-bryonic neural progenitor cells induce blood–brain barrier properties.J Neurochem 101:555–565.49. Al Ahmad A, Gassmann M, Ogunshola OO. 2009. Maintainingblood–brain barrier integrity: Pericytes perform better than astrocytesduring prolonged oxygen deprivation. J Cell Physiol 218:612–622.50. Shayan G, Choi YS, Shusta EV, Shuler ML, Lee KH. 2011. Murinein vitro model of the blood–brain barrier for evaluating drug transport.Eur J Pharm Sci 42:148–155.51. Freese C, et al. 2014. A novel blood–brain barrier co-culture sys-tem for drug targeting of Alzheimer’s disease: Establishment by usingacitretin as a model drug. PLoS One 9:e91003.52. Helms HC, et al. 2014. An electrically tight in vitro blood–brainbarrier model displays net brain-to-blood efflux of substrates for theABC transporters, P-gp, Bcrp and Mrp-1. AAPS J 16:1046–1055.53. Omidi Y, et al. 2003. Evaluation of the immortalised mouse braincapillary endothelial cell line, b.End3, as an in vitro blood–brain barriermodel for drug uptake and transport studies. Brain Res 990:95–112.
54. Brown R, Morris A, O’Neil R. 2007. Tight junction protein expres-sion and barrier properties of immortalized mouse brain microvesselendothelial cells. Brain Res 1130:17–30.55. Claudio L, Raine CS, Brosnan CF. 1995. Evidence of persistentblood–brain barrier abnormalities in chronic-progressive multiple scle-rosis. Acta Neuropathol 90:228–238.56. Claudio L. 1995. Ultrastructural features of the blood–brain barrierin biopsy tissue from Alzheimer’s disease patients. Acta Neuropathol91:6–14.57. Najjar S, Pearlman DM, Devinsky O, Najjar A, Zagzag D. 2013.Neurovascular unit dysfunction with blood–brain barrier hyperperme-ability contributes to major depressive disorder: A review of clinicaland experimental evidence. J Neuroinflammation 10:142.58. Lippmann ES, Weidenfeller C, Svendsen CN, Shusta EV. 2011.Blood–brain barrier modeling with co-cultured neural progenitor cell-derived astrocytes and neurons. J Neurochem 119:507–520.59. Lippmann ES, et al. 2012. Derivation of blood–brain barrier en-dothelial cells from human pluripotent stem cells. Nat Biotechnol30:783–791.60. Lippmann ES, Al-Ahmad A, Azarin SM, Palecek SP, Shusta EV.2014. A retinoic acid-enhanced, multicellular human blood–brain bar-rier model derived from stem cell sources. Sci Rep 4:4160.61. Eigenmann D, et al. 2013. Comparative study of four immortal-ized human brain capillary endothelial cell lines, hCMEC/D3, hBMEC,TY10, and BB19, and optimization of culture. Fluids Barriers CNS10:33.62. Dehouck MP, Meresse S, Delorme P, Fruchart JC, Cecchelli R.1990. An easier, reproducible, and mass-production method to studythe blood–brain barrier in vitro. J Neurochem 54:1798–1801.63. Zysk G, Schneider-Wald B. 2001. Pneumolysin is the main inducerof cytotoxicity to brain microvascular endothelial cells caused by Strep-tococcus pneumoniae. Infect Immun 69:845–852.64. Kis B, et al. 2001. Adrenomedullin regulates blood–brain barrierfunctions in vitro. Neuroreport 12:4139–4142.65. Deli Ma, Abraham CS, Niwa M, Falus A. 2003. N,N-diethyl-2-[4-(phenylmethyl)phenoxy]ethanamine increases the permeability of pri-mary mouse cerebral endothelial cell monolayers. Inflamm Res 52Suppl1:39–40.66. Smith M, Omidi Y, Gumbleton M. 2007. Primary porcine brainmicrovascular endothelial cells: Biochemical and functional character-isation as a model for drug transport and targeting. J Drug Target15:253–268.67. Hutamekalin P, et al. 2008. Effect of nicotine and polyaromtic hy-drocarbons on cerebral endothelial cells. Cell Biol Int 32:198–209.68. Nakagawa S, et al. 2009. A new blood–brain barrier model using pri-mary rat brain endothelial cells, pericytes and astrocytes. NeurochemInt 54:253–263.69. Franke H, Galla HJ, Beuckmann CT. 1999. An improved low-permeability in vitro-model of the blood–brain barrier: Transport stud-ies on retinoids, sucrose, haloperidol, caffeine and mannitol. Brain Res818:65–71.70. Bowman PD, Ennis SR, Rarey KE, Betz aL, Goldstein GW. 1983.Brain microvessel endothelial cells in tissue culture: A model for studyof blood–brain barrier permeability. Ann Neurol 14:396–402.71. Miller D, Audus K, Borchardt R. 1992. Application of cultured en-dothelial cells of the brain microvasculature in the study of the blood–brain barrier. J Tissue Cult Methods 14:217–224.72. Jeliazkova-Mecheva VV, Bobilya DJ. 2003. A porcine astro-cyte/endothelial cell co-culture model of the blood–brain barrier. BrainRes Protoc 12:91–98.73. Bernas M, Cardoso F, Daley S. 2010. Establishment of primarycultures of human brain microvascular endothelial cells: A new andsimplified method to obtain cells for an in vitro model of the blood. NatProtoc 5:1265–1272.74. Abbott NJ, Dolman DEM, Drndarski S, Fredriksson SM. 2012. Animproved in vitro blood–brain barrier model: Rat brain endothelial cellsco-cultured with astrocytes. Methods Mol Biol 814:415–430.
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329

REVIEW 2745
75. Navone SE, et al. 2013. Isolation and expansion of human andmouse brain microvascular endothelial cells. Nat Protoc 8:1680–1693.76. Helms HC, Brodin B. 2014. Generation of primary cultures ofbovine brain endothelial cells and setup of cocultures with rat astro-cytes. Methods Mol Biol 1135:365–382.77. Gumbleton M, Audus KL. 2001. Progress and limitations in theuse of in vitro cell cultures to serve as a permeability screen for theblood–brain barrier. J Pharm Sci 90:1681–1698.78. Hawkins R, O’Kane R, Simpson IA, Vina JR. 2006. Structure of theblood–brain barrier and its role in the transport of amino acids. J Nutr(1 Suppl):218S–226S.79. Zhang Y, Li C, Ye Y, Johnson K. 2006. Porcine brain microvesselendothelial cells as an in vitro model to predict in vivo blood–brainbarrier permeability. Drug Metab Dispos 34:1935–1943.80. Patabendige A, Skinner RA, Abbott NJ. 2013. Establishment ofa simplified in vitro porcine blood–brain barrier model with hightransendothelial electrical resistance. Brain Res 1521:1–15.81. Hartmann C, Zozulya A, Wegener J, Galla H-J. 2007. The impact ofglia-derived extracellular matrices on the barrier function of cerebralendothelial cells: An in vitro study. Exp Cell Res 313:1318–1325.82. Patabendige A, Skinner RA, Abbott NJ. 2013. Establishment ofa simplified in vitro porcine blood–brain barrier model with hightransendothelial electrical resistance. Brain Res 1521:1–15.83. Griep LM, et al. 2013. BBB ON CHIP: Microfluidic platform tomechanically and biochemically modulate blood–brain barrier function.Biomed Microdevices 15:145–150.84. Cucullo L, et al. 2008. Immortalized human brain endothelial cellsand flow-based vascular modeling: A marriage of convenience for ratio-nal neurovascular studies. J Cereb Blood Flow Metab 28:312–328.85. Prabhakarpandian B, et al. 2013. SyM-BBB: A microfluidic bloodbrain barrier model. Lab Chip 13:1093.86. Takeshita Y, et al. 2014. An in vitro blood–brain barrier model com-bining shear stress and endothelial cell/astrocyte co-culture. J Neurosci30:165–172.87. Gaillard PJ, et al. 2001. Establishment and functional character-ization of an in vitro model of the blood–brain barrier, comprising aco-culture of brain capillary endothelial cells and astrocytes. Eur JPharm Sci 12:215–222.88. Rubin LL, et al. 1991. A cell culture model of the blood–brain bar-rier. J Cell Biol 115:1725–1735.89. deBoer A, Sutanto W, Pardridge WM. 1998. Drug transport acrossthe blood–brain barrier: In vitro and in vivo techniques. Drug Deliv5:153–153.90. Schaddelee MP, et al. 2003. Functional role of adenosine receptorsubtypes in the regulation of blood–brain barrier permeability: Possibleimplications for the design of synthetic adenosine derivatives. Eur JPharm Sci 19:13–22.91. Helms HC, Waagepetersen HS, Nielsen CU, Brodin, B. 2010.Paracellular tightness and claudin-5 expression is increased in theBCEC/astrocyte blood–brain barrier model by increasing media buffercapacity during growth. AAPS J 12:759–770.92. Cestelli A, et al. 2001. Functional feature of a novel model of bloodbrain barrier: Studies on permeation of test compounds. J Control Re-lease 76:139–147.93. Raub TJ, Kuentzel SL, Sawada Ga. 1992. Permeability of bovinebrain microvessel endothelial cells in vitro: Barrier tightening bya factor released from astroglioma cells. Exp Cell Res 199:330–340.94. Giese H, Mertsch K, Blasig I. 1995. Effect of MK-801 and U83836Eon a porcine brain capillary endothelial cell barrier during hypoxia.Neurosci Lett 191:169–172.95. Schirmacher A, Winters S. 2000. Electromagnetic fields (1.8 GHz)increase the permeability to sucrose of the blood–brain barrier in vitro.Bioelectromagnetics 21:338–345.96. Cantrill CA, Skinner RA, Rothwell NJ, Penny JI. 2012. An immor-talised astrocyte cell line maintains the in vivo phenotype of a primaryporcine in vitro blood–brain barrier model. Brain Res 1479:17–30.
97. Kasa P, Pakaski M, Joo F, Lajtha A. 1991. Endothelial cells fromhuman fetal brain microvessels may be cholinoceptive, but do not syn-thesize acetylcholine. J Neurochem 56:2143–2146.98. Megard I, et al. 2002. A co-culture-based model of human blood–brain barrier: Application to active transport of indinavir and in vivo–invitro correlation. Brain Res 927:153–167.99. Cucullo L, Marchi N, Hossain M, Janigro D. 2011. A dynamic invitro BBB model for the study of immune cell trafficking into the centralnervous system. J Cereb Blood Flow Metab 31:767–777.100. Cucullo L, Hossain M, Puvenna V, Marchi N, Janigro D. 2011. Therole of shear stress in blood–brain barrier endothelial physiology. BMCNeurosci 12:40.101. Tan K, Dobbie M, Felix R. 2001. A comparison of the inductionof immortalized endothelial cell impermeability by astrocytes. Neuro-chemistry 12:1329–1334.102. Hatherell K, Couraud PO, Romero IA, Weksler B, Pilkington GJ.2011. Development of a three-dimensional, all-human in vitro model ofthe blood–brain barrier using mono-, co-, and tri-cultivation Transwellmodels. J Neurosci Methods 199:223–229.103. Li G, et al. 2010. Permeability of endothelial and astrocyte cocul-tures: In vitro blood–brain barrier models for drug delivery studies.Ann Biomed Eng 38:2499–2511.104. Booth R, Kim H. 2012. Characterization of a microfluidic in vitromodel of the blood–brain barrier (:BBB). Lab Chip 12:1784–1792.105. Booth RH, Kim H. 2012. A parallel array microfluidic blood–brainbarrier model for high-throughput quantitaion of shear stress effects. In16th International Conference on Miniaturized Systems for Chemistryand Life Sciences, 491–493.106. Stanness KA, et al. 1999. A new model of the blood–brain bar-rier: Co-culture of neuronal, endothelial and glial cells under dynamicconditions. Neuroreport 10:3725–3731.107. Blasig IE, Mertsch K, Haseloff RF. 2002. Nitronyl nitroxides, anovel group of protective agents against oxidative stress in endothelialcells forming the blood–brain barrier. Neuropharmacology 43:1006–1014.108. Demeuse P, et al. 2002. Compartmentalized coculture of rat brainendothelial cells and astrocytes: A syngenic model to study the blood–brain barrier. J Neurosci Methods 121:21–31.109. Krizanac-Bengez L, et al. 2003. Effects of transient loss of shearstress on blood–brain barrier endothelium: Role of nitric oxide and IL-6.Brain Res 977:239–246.110. Krizanac-Bengez L, et al. 2006. Loss of shear stress inducesleukocyte-mediated cytokine release and blood–brain barrier failure indynamic in vitro blood–brain barrier model. J Cell Physiol 206:68–77.111. Nakagawa S, et al. 2007. Pericytes from brain microvesselsstrengthen the barrier integrity in primary cultures of rat brain en-dothelial cells. Cell Mol Neurobiol 27:687–694.112. Cecchelli R, et al. 2014. A stable and reproducible human blood–brain barrier model derived from hematopoietic stem cells. PLoS One9:e99733.113. Hayashi K, et al. 2004. Effects of hypoxia on endothelial/pericyticco-culture model of the blood–brain barrier. Regul Pept 123:77–83.114. Hamm S, et al. 2004. Astrocyte mediated modulation of blood–brain barrier permeability does not correlate with a loss of tight junc-tion proteins from the cellular contacts. Cell Tissue Res 315:157–166.115. Garberg P, et al. 2005. In vitro models for the blood–brain barrier.Toxicol Vitr 19:299–334.116. Stanness K, et al. 1997. Morphological and functional character-ization of an in vitro blood–brain barrier model. Brain Res 771:329–342.117. Abbruscato TJ, Davis TP. 1999. Combination of hypoxia/aglycemiacompromises in vitro blood–brain barrier integrity. J Pharmacol ExpTher 289:668–675.118. Cucullo L, et al. 2002. A new dynamic in vitro model for the mul-tidimensional study of astrocyte-endothelial cell interactions at theblood–brain barrier. Brain Res 951:243–254.
DOI 10.1002/jps.24329 Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015

2746 REVIEW
119. Santaguida S, et al. 2006. Side by side comparison between dy-namic versus static models of blood–brain barrier in vitro: A perme-ability study. Brain Res 1109:1–13.120. Boveri M, et al. 2005. Induction of blood–brain barrier proper-ties in cultured brain capillary endothelial cells: Comparison betweenprimary glial cells and C6 cell line. Glia 51:187–198.121. Weidenfeller C, Svendsen CN, Shusta EV. 2007. Differentiationembryonic nerual progenitor cells induce blood–brain barrier proper-ties. J Neurochem 101:555–565.122. Balbuena P, Li W, Magnin-Bissel G, Meldrum JB, Ehrich M. 2010.Comparison of two blood–brain barrier in vitro systems: Cytotoxicityand transfer assessments of malathion/oxon and lead acetate. ToxicolSci 114:260–271.123. Xue Q, et al. 2013. A novel brain neurovascular unit model withneurons, astrocytes and microvascular endothelial cells of rat. Int JBiol Sci 9:174–189.124. DeBault LE, Cancilla PA. 1980. gamma-Glutamyl transpeptidasein isolated brain endothelial cells: Induction by glial cells in vitro. Sci-ence 207:653–655.125. Vastag M, Keseru GM. 2009. Current in vitro and in silico modelsof blood–brain barrier penetration: A practical view. Curr Opin DrugDiscov Dev 12:115–124.126. Zonta M, et al. 2003. Neuron-to-astrocyte signaling is centralto the dynamic control of brain microcirculation. Nat Neurosci 6:43–50.127. Harris SG, Shuler ML. 2003. Growth of endothelial cells on mi-crofabricated silicon nitride membranes for an in vitro model of theblood–brain barrier. Biotechnol Bioprocess Eng 8:246–251.128. Stanness K, Guatteo E, Janigro D. 1996. “A dynamic model of theblood–brain barrier “ in vitro”. Neurotoxicology 17:481–496.129. Naik P, Cucullo L. 2012. In vitro blood–brain barrier models: Cur-rent and perspective technologies. J Pharm Sci 101:1337–1354.130. Dohgu S, et al. 2005. Brain pericytes contribute to the in-duction and up-regulation of blood–brain barrier functions through
transforming growth factor-beta production. Brain Res 1038:208–215.131. Janzer RC, Raff MC. 1987. Astrocytes induce blood–brain barrierproperties in endothelial cells. Nature 325:253–257.132. Schiera G, Sala S, Gallo A. 2005. Permeability properties of athree-cell type in vitro model of blood–brain barrier. J Cell Mol Med9:373–379.133. Savettieri G, et al. 2000. Neurons and ECM regulate occludinlocalization in brain endothelial cells. Mol Neurosci 11:1081–1084.134. Schiera G, et al. 2003. Synergistic effects of neurons and astrocyteson the differentiation of brain capillary endothelial cells in culture. JCell Mol Med 7:165–170.135. Zlokovic BV. 2008. The blood–brain barrier in health and chronicneurodegenerative disorders. Neuron 57:178–201.136. Gaillard PJ, de Boer AG. 2000. Relationship between permeabilitystatus of the blood–brain barrier and in vitro permeability coefficientof a drug. Eur J Pharm Sci 12:95–102.137. Perriere N, et al. 2007. A functional in vitro model of rat blood–brain barrier for molecular analysis of efflux transporters. Brain Res1150:1–13.138. Griep LM, et al. 2012. BBB on chip: Microfluidic platform to me-chanically and biochemically modulate blood–brain barrier function.Biomed Microdevices 15:145–150.139. Neuhaus W, et al. 2006. A novel flow based hollow-fiber blood–brain barrier in vitro model with immortalised cell line PBMEC/C1–2.J Biotechnol 125:127–141.140. Duvernoy H, Delon S, Vannson J. 1983. The vascularization of thehuman cerebellar cortex. Brain Res Bull 11:419–480.141. Nicholson C. 2001. Diffusion and related transport mechanismsin brain tissue. Reports Prog Phys 64:815–884.142. Schlageter KE, Molnar P, Lapin GD, Groothuis DR. 1999. Mi-crovessel organization and structure in experimental brain tumors: Mi-crovessel populations with distinctive structural and functional prop-erties. Microvasc Res 58:312–328.
Wolff et al., JOURNAL OF PHARMACEUTICAL SCIENCES 104:2727–2746, 2015 DOI 10.1002/jps.24329
Related Documents











