This article was downloaded by:[Fitzgerald-Bocarsly, Patricia] On: 27 December 2007 Access Details: [subscription number 789019860] Publisher: Informa Healthcare Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Immunological Investigations A Journal of Molecular and Cellular Immunology Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713597256 Image-Based Study of Interferongenic Interactions between Plasmacytoid Dendritic Cells and HSV-Infected Monocyte-Derived Dendritic Cells Nicholas J. Megjugorac a ; Evan S. Jacobs bc ; Alexander G. Izaguirre d ; Thaddeus C. George e ; Gunjan Gupta b ; Patricia Fitzgerald-Bocarsly bc a UMDNJ-New Jersey Medical School-University Hospital Cancer Center, Newark, New Jersey, USA b UMDNJ-New Jersey Medical School- Department of Pathology, Newark, New Jersey, USA c UMDNJ-Graduate School of Biomedical Sciences, Newark, New Jersey, USA d UMDNJ-Robert Wood Johnson Medical School, Piscataway, New Jersey, USA e Amnis Corporation, Seattle, Washington, USA Online Publication Date: 01 September 2007 To cite this Article: Megjugorac, Nicholas J., Jacobs, Evan S., Izaguirre, Alexander G., George, Thaddeus C., Gupta, Gunjan and Fitzgerald-Bocarsly, Patricia (2007) 'Image-Based Study of Interferongenic Interactions between Plasmacytoid Dendritic Cells and HSV-Infected Monocyte-Derived Dendritic Cells ', Immunological Investigations, 36:5, 739 - 761 To link to this article: DOI: 10.1080/08820130701715845 URL: http://dx.doi.org/10.1080/08820130701715845 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article was downloaded by:[Fitzgerald-Bocarsly, Patricia]On: 27 December 2007Access Details: [subscription number 789019860]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Immunological InvestigationsA Journal of Molecular and Cellular ImmunologyPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713597256
Image-Based Study of Interferongenic Interactionsbetween Plasmacytoid Dendritic Cells and HSV-InfectedMonocyte-Derived Dendritic CellsNicholas J. Megjugorac a; Evan S. Jacobs bc; Alexander G. Izaguirre d; ThaddeusC. George e; Gunjan Gupta b; Patricia Fitzgerald-Bocarsly bca UMDNJ-New Jersey Medical School-University Hospital Cancer Center, Newark,New Jersey, USAb UMDNJ-New Jersey Medical School- Department of Pathology, Newark, NewJersey, USAc UMDNJ-Graduate School of Biomedical Sciences, Newark, New Jersey, USAd UMDNJ-Robert Wood Johnson Medical School, Piscataway, New Jersey, USA
e Amnis Corporation, Seattle, Washington, USA
Online Publication Date: 01 September 2007To cite this Article: Megjugorac, Nicholas J., Jacobs, Evan S., Izaguirre, Alexander G., George, Thaddeus C., Gupta,Gunjan and Fitzgerald-Bocarsly, Patricia (2007) 'Image-Based Study of Interferongenic Interactions betweenPlasmacytoid Dendritic Cells and HSV-Infected Monocyte-Derived Dendritic Cells ', Immunological Investigations, 36:5,739 - 761To link to this article: DOI: 10.1080/08820130701715845URL: http://dx.doi.org/10.1080/08820130701715845
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Immunological Investigations, 36:739–761, 2007Copyright © Informa Healthcare USA, Inc.ISSN: 0882-0139 print / 1532-4311 onlineDOI: 10.1080/08820130701715845
LIMM0882-01391532-4311Immunological Investigations, Vol. 36, No. 5-6, October 2007: pp. 1–34Immunological InvestigationsImage-Based Study of Interferongenic Interactions between Plasmacytoid Dendritic Cells and HSV-Infected Monocyte-Derived Dendritic CellsInteractions of pDC with HSV-Infected MDDCN. J. Megjugorac et al.
Nicholas J. Megjugorac,1 Evan S. Jacobs,2,3 Alexander G. Izaguirre,4 Thaddeus C. George,5 Gunjan Gupta,2 and Patricia Fitzgerald-Bocarsly2,3
1UMDNJ-New Jersey Medical School-University Hospital Cancer Center, Newark,New Jersey, USA2UMDNJ-New Jersey Medical School- Department of Pathology, Newark, New Jersey,USA3UMDNJ-Graduate School of Biomedical Sciences, Newark, New Jersey, USA4UMDNJ-Robert Wood Johnson Medical School, Piscataway, New Jersey, USA5Amnis Corporation, Seattle, Washington, USA
Plasmacytoid dendritic cells (pDC) are well-known for their ability to produce largequantities of interferon-α (IFN-α) in response to viruses. In addition, pDC produceIFN-α in response to HSV-infected cells. We demonstrate that both tonsil and PBMCcontain pDC that respond to stimulation with HSV either in suspension or in tonsil tis-sue-fragment culture. We hypothesized that other DC subsets acquire virus in theperiphery and deliver the interferongenic signals to the pDC in the draining lymphoidtissue. As a model for pDC/myeloid DC interaction, we studied the interaction of pDCderived from blood with HSV-infected and uninfected monocyte derived dendritic cells(MDDC). Infected, but not uninfected, MDDC induced IFN-α in pDC. To further studypDC/infected MDDC interactions, we labeled MDDC with fluorescent cell trackersPKH67 or CFSE prior to infection with HSV and co-cultured with pDC. Cells were thenanalyzed using conventional and imaging flow cytometry. In addition, we infectedMDDC with a GFP-expressing HSV prior to co-culture with pDC. Using traditionalflow cytometry, we observed that pDC became fluorescent after co-incubation with
Authors Megjugorac and Jacobs contributed equally.Address correspondence to Dr. Patricia Fitzgerald-Bocarsly, UMDNJ – New JerseyMedical School, Department of Pathology and Laboratory Medicine, 185 South OrangeAve., Newark, NJ 07301; E-mail: [email protected]

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
740 N. J. Megjugorac et al.
uninfected or infected, fluorescently labeled MDDC, indicating that MDDC transferredfluorescent protein and membrane to pDC. By imaging flow cytometry, we observedformation of conjugates between pDC and MDDC as well as transfer and internaliza-tion of cellular components from the labeled MDDC by pDC, with preferential uptakefrom, and association with, infected vs. uninfected MDDC. These studies demonstratethat MDDC infected with HSV are able to stimulate IFN-α and chemokine productionby pDC through the transfer of cellular materials from the HSV-infected MDDC to thepDC. Together, these observations indicate that heterogeneous populations of DCinteract to generate an effective IFN-α response.
Keywords Plasmacytoid Dendritic Cells (PDC), MDDC, Imaging Flow Cytometry,HSV, IFN-α.
INTRODUCTION
Plasmacytoid dendritic cells (pDC) are recognized as one of the most potentsubsets of immune modulating cells in the body, effectively linking innate andadaptive immunity. In humans, this subset of CD123+, HLA-DR+, CD11c−,BDCA-2+ (CD303), BDCA-4+ (CD304) dendritic cells are found circulating inthe blood, where they represent 0.1–0.5% of PBMC, as well as in primary andsecondary lymphoid tissue including thymus, tonsil, spleen, lymph node andbone marrow. In addition, pDC have been described as either resident in, orrecruited to, non-lymphoid tissues such as lung and gut. pDC are best knownfor their production of massive amounts of IFN-α (up to 3–10 pg/cell) inresponse to enveloped viruses and synthetic TLR7 and TLR9 ligands(reviewed in (Fitzgerald-Bocarsly and Feng, 2007)).
These virus-responsive cells also produce inflammatory chemokinesCCL2-5 and CXCL10 in response to viral and CpG stimulation, inactivatedinfluenza virus, and CD40 ligation, and also express chemokine receptors thatallow them to be recruited to sites of inflammation (Megjugorac et al., 2004;Penna et al., 2002a, 2002b). We have demonstrated that supernatants fromactivated pDC chemoattract both activated T cells and natural killer cells(Megjugorac et al., 2004), and that IFN-α produced by pDC serves to furtheractivate these populations of cells. Close interaction of pDC with other popula-tions of cells is suggested by the observation that cell cultures containing pDCtend to form clusters of cells that disappear when HLA-DR+, IFN-α producingcells are removed (Feldman and Fitzgerald-Bocarsly, 1990).
Myeloid-derived dendritic cells (mDC, also known as “conventional” DC(cDC)) regulate immune responses through cell:cell interactions and cytokine/chemokine production, most notably IL-12. Although mDC, like pDC, circulatein low numbers in peripheral blood, a cell type that resembles mDC can begenerated in vitro by culturing human monocytes in the presence of GM-CSFand IL-4 for 7 days (Romani et al., 1996). These monocyte-derived dendriticcells (MDDC) serve as a model for immature blood mDC, and like blood mDC,

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 741
can be induced to become mature antigen presenting cells with such factors asIL-1β, LPS, and IFN-α (Romani et al., 1996). MDDC have greater endocyticactivity than pDC (Fanning et al., 2006), and have been shown to phagocytosecomponents from apoptotic cells via CD36 and αV-β3 and αV-β5 integrins(Rubartelli et al., 1997; Albert et al., 1998). Additionally, Barratt-Boyes andcolleagues have demonstrated that MDDC have the ability to internalizeantigen from the surface of live cells, a process known as “nibbling” (Harshyneet al., 2001, 2003). Antigens acquired from live or apoptotic cells by MDDCcan be effectively cross-presented to CD8+ T cells (Albert et al., 1998;Harshyne et al., 2001).
Although pDC are significantly less endocytic than MDDC, they are able tointernalize ligands such as FITC-dextran and viruses via endocytosis into acidiccompartments that contain TLR7 and TLR9, where viral nucleic acids signal forthe induction of IFN-α (Beignon et al., 2005; Fanning et al., 2006; Lund et al.,2003, 2004). Disruption of the endosomal pathway by chloroquine (a lysosomo-tropic weak base that raises endosomal pH) ablates IFN-α production by pDC,thus indicating that ligand internalization into endosomes is a vital step in theability of pDC to respond to viral stimulation (Feldman, 1994; Lebon, 1985).
pDC can be recruited to the skin and mucosal surfaces such as the lungand vagina (reviewed in (Fitzgerald-Bocarsly, 2002)), however, their numbersat these sites are relatively low in the absence of chemoattractive signals.Moreover, while it is known that activated pDC can be recruited to the lymphnode (Cella et al., 1999), as shown below, we have observed that pDC are resi-dent in a quiescent state (i.e., not expressing IFN-α) in tonsils and can bestimulated by HSV to produce IFN-α. We hypothesize that another cell bringsinterferongenic signals to these non-activated pDC in the secondary lymphoidtissue. Candidates for such viral couriers are cells within the myeloid DC lin-eage, which are known to acquire antigen and migrate to the draining lymphnodes. In this study, we have evaluated the interaction of HSV-infectedMDDC with pDC using a combination of traditional flow cytometric and imag-ing flow cytometric techniques. We report here that MDDC infected with HSVare able to stimulate IFN-α and chemokine production by pDC. Additionally,we demonstrate the preferential transfer of cellular components from HSV-infected MDDC vs. uninfected MDDC to pDC. Together, these results indicatethat heterogeneous populations of DC interact to generate an effective IFN-αresponse.
MATERIALS AND METHODS
Viruses. HSV-1 strain 2931 (originally obtained from Dr. Carlos Lopez,then of the Sloan-Kettering Inst., New York, NY) and Vesicular StomatitisVirus (VSV; originally obtained from Dr. Nicholas Ponzio of New Jersey Medi-cal School) were grown and titrated by plaque forming assay in VERO cells

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
742 N. J. Megjugorac et al.
(American Type Culture Collection, Manassas, VA). GFP-HSV (Foster et al.,1998) (originally obtained from Dr. K. Kousoulas of the School of VeterinaryMedicine, Louisiana State University, Baton Rouge), an HSV strain contain-ing an enhanced green fluorescent protein construct under an immediate earlypromoter was grown and titered as above. Sendai virus (SV; Sendai/Cantellstrain) was obtained from the ATCC. All virus stocks were stored at –70°Cuntil use. UV-irradiated HSV was obtained by exposing the above virus stockto 7800 J/cm2 in a UV Stratalinker 1800 (Stratagene, La Jolla, CA). This UV-treatment completely inhibited viral replication in permissive VERO cells.
Cell Lines. GM-0459A cells (National Institute of General MedicineSciences Human Genetic Mutant Cell Line Repository, Camden, NJ), a pri-mary fibroblast cell line trisomic for chromosome 21, were grown in DMEM(JHR Biosciences, Lenexa, KS) supplemented with 15% FCS (HyClone, Logan,UT),), 2mM L-glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin(DMEM, 15%). VERO cells were grown in DMEM-10% FCS.
Preparation of Peripheral Blood Mononuclear Cells (PBMC) and Mononu-clear Cells from Lymphoid Tissue. All human studies were approved by theIRB at UMDNJ–New Jersey Medical School. PBMC were isolated by Ficoll-Hypaque density centrifugation (Lymphoprep: Accurate Chemical and ScientificCo., Westbury, NY) from heparinized peripheral blood obtained via venipunc-ture with informed consent from healthy volunteers. PBMC were washedtwice with Hank’s Balanced Salt Solution (HBSS) (Life Technologies, GrandIsland, NY) and resuspended in RPMI 1640 media (Life Technologies) con-taining 10% FCS, 2mM L-glutamine, 100 U/mL penicillin, 100 μg/mL strepto-mycin and 25 mM HEPES (RPMI-10%) and enumerated electronically with aCoulter Counter Series Z1 (Coulter Electronics, Inc., Hialeah, FL). Non-reac-tive lymph nodes and tonsils were obtained from surgical specimens frompatients under an IRB-approved protocol. Mononuclear cell suspensions weremade from the lymphoid tissue by gentle teasing with forceps. In some experi-ments, tonsil was hand-cut into approximately 2 mm3 pieces; individual pieceswere placed in 100 μL of media in a flat-bottomed 96-well plate and HSV wasdirectly placed on the top of the fragments. The cultures were incubated at37oC for 18 hr, then supernatants were harvested for determination of antivi-ral activity as described next.
Enrichment of pDC. pDC for use in the ImageStream experiments werenegatively selected from PBMC with the Miltenyi Plasmacytoid Dendritic Cellisolation kit as described by the manufacturer, but with one modification:since high levels of purity were not required for the ImageStream experi-ments, but we did require high yields, one half of the recommended number ofmagnetic beads were used for the isolations. After the column separation,enriched pDC were counted as above and checked for purity by staining withanti-BDCA-2 and anti-CD123 expression followed by flow cytometric analysis.Purity under these non-stringent isolation conditions was typically about 70%.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 743
Preparation of MDDC. Monocytes were enriched from PBMC usingCD14-labeled microbeads (Miltenyi Biotec Inc., Auburn, CA) according to themanufacturer’s instructions. Positively selected monocytes evaluated for purityby flow cytometry (typically, ˜99% of cells were CD14+). Alternatively, mono-cytes were prepared by layering 1.5x108 PBMC on a 75 cm2 flask. After 1 hour,non-adherent cells were removed by washing with 1X PBS at roomtemperature. The resultant monocyte-enriched populations were cultured with15 mL of 1% autologous plasma in RPMI containing 500 IU/mL of IL-4 (R&DSystems, Minneapolis, MN) and 1000 IU/mL of GM-CSF (Sargramostim-Leuk-ine: Immunex Corp., Seattle, WA). The cells were fed with media containing500 IU/mL of IL-4 and 1000 IU/mL of GM-CSF on Days 2, 4, and 6. DC werecollected on day 7 and evaluated by flow cytometry (typically, 55-75% of cellswere HLA-DR+, CD3-, CD14-, CD16-, CD19-, CD20-, CD56-).
Protein and Membrane Labeling. For CFSE labeling, 1X106 cells wereincubated in 250nM CFSE (Molecular Probes) in 0.1% BSA in PBS for 15 min-utes and washed twice with 25 mL PBS. For PKH67 labeling, PKH67 wasdiluted 1:50 in ethanol, then 2 μL of the PKH67 solution was quickly added to250 μL of Diluent C which was immediately added to pelleted MDDC that hadbeen washed in Diluent C. After 5 minutes, 1 mL each of FCS and RPMI wereadded to the labeled MDDC, the cells were pelleted and were subsequentlywashed twice with RPMI-10%. Lower dye concentrations than recommendedby the manufacturer were utilized because of lower cell number (1–2 X 106
cells) and the ImageStream sensitivity of detection. In addition, to improvelabeling, cells were centrifuged in Diluent C prior to staining to eliminate thepossibility of carryover salts from PBS. Fluorescent-labeled cells were theninfected with HSV or left uninfected, as described above. In some experi-ments, MDDC were infected with GFP-HSV for 4 hr at an MOI of 5 prior toco-culture with pDC. Following co-culture, the cells were stained with anti-BDCA-2 and fixed with 1% PFA. Fixed samples were sent to Amnis for analysiswith the ImageStream.
Stimulation of PBMC and MDDC with Cell-Free HSV or HSV-InfectedCells. MDDC (1x106 cells/mL) in RPMI-10% FCS (RPMI-10%) were infectedwith either HSV or UV-inactivated HSV at a multiplicity of infection (MOI) of5 and incubated for 2 hours at 37°C, 5% CO2. The cells were then transferredto 50 mL conical tubes (Corning, New York) and washed twice with 10% FCSin RPMI. 1mL of the RPMI from the last wash was collected and stored at –70°C for later use. MDDC were resuspended at 2x106 cells/mL in RPMI-10%and added to PBMC (also at 2x106 cells/mL) in RPMI-10% at a 1:1 concentra-tion in 15mL conical tubes. The co-cultures were incubated for 24 hours at37°C, 5% CO2. After the incubation, supernatants were harvested and storedat -70°C for later evaluation for antiviral activity. For direct viral stimulationof PBMC at 2x106 cells/mL were stimulated with HSV-1 (MOI=1) overnight ina microtiter plate as described above.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
744 N. J. Megjugorac et al.
IFN Bioassay. IFN bioassays were performed using a cytopathic effectreduction assay with GM-0459A (GM) cells infected with VSV as the challeng-ing virus as previously described (Fitzgerald et al., 1982). An IFN-α referencestandard (National Institute of Allergy and Infectious Disease, Bethesda,MD-standard G-023-901-527) was used at 100 IU/mL.
Flow Cytometry - Surface Staining. After washing with 0.1% BSA(Sigma) in PBS, cells were resuspended in 0.1% BSA (Sigma) in PBS containing5% heat inactivated human serum and incubated with fluorochrome-conjugatedantibodies combinations designed to detect pDC (CD123-PE (BioLegend, SanDiego, CA, Clone 6H6) and HLA-DR-PerCP (Clone L243, BD Biosciences, SanDiego, CA), or CD123-PE-Cy5 (BD BioSciences, Clone 9F5) and BDCA-2 FITC(Miltenyi Biotec, Clone AC144). The cells were then washed and fixed in 1%paraformaldehyde (PFA, Fisher, Pittsburgh, PA). For determination of pDC vs.mDC in blood, tonsil and lymph node, cells were stained with a FITC-labeledlineage cocktail (BD BioSciences, Lin 1, containing anti-CD3, CD14, CD16,CD19, CD20 and CD56), anti-HLA-DR-PerCP (BD BioSciences, Lin 1), CD123-PE and CD11c-APC (BD BioSciences, Clone LHCL-3). Antibodies used for phe-notypic identification of MDDC were as follows: Anti-IgG2a, IgG1 (DAKO D/S,Denmark), CD11c. CD123, HLA-DR (BD BioSciences) and BDCA-2.
Intracellular Detection of IFN-a and Chemokines. PBMC were stainedfor intracellular cytokine or chemokine expression as described previously(Megjugorac et al., 2004). Briefly, cells were incubated with the indicatedstimuli for 4 hours at 37oC before addition of Brefeldin A (5 μg/mL) (Sigma)and a subsequent 2-hr incubation. Cells were washed with cold 0.1% BSA inPBS (Life Technologies), blocked with 5% heat-inactivated human serum, andstained with anti-CD123-PE (detected in Channel 2 with the FACS Calibur)and anti-HLA-DR-APC (Channel 4), for 20 min at 4°C, then washed and fixedwith 1% PFA in PBS at 4°C overnight.
The following day, cells were washed twice with PBS-2% FCS and perme-abilized with 0.5% saponin (Sigma) in PBS-2% FCS for 30 min at RT, and sub-sequently incubated with one of the following antibodies: biotinylated 293mAb to IFN-α (obtained from Gunnar Alm, Upsalla, Sweden), anti-MIP-1β(R&D Systems, Minneapolis, MN), MCP-1 (R&D) or biotinylated anti-IP-10(BD Pharmingen) for 30 min at room temperature. In the samples containingbiotinylated primary antibodies (IFN-α and IP-10), cells were then washedtwice with saponin buffer and incubated with Streptavidin-Quantum Red(Sigma) (Channel 3) for 30 min at room temperature. Finally, the cells werewashed and resuspended in 1% PFA and analyzed using the FACS Caliburflow cytometer with CellQuest Analysis software (BD Biosciences). Afterinitially gating side scatter vs. forward scatter to eliminate dead cells andneutrophils, pDC were gated in mixed lymphocyte populations by theirexpression of CD123 and HLA-DR. IFN-α was then detected within this pDCgate using Streptavidin-Quantum Red.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 745
ImageStream Data Acquisition and Analysis. For visualization of theinteraction between pDC and MDDC, we utilized the ImageStream multispec-tral imaging flow cytometer (Amnis Corporation Seattle, WA (Ortyn et al.,2006)). Enriched pDC (1–2 X 105 cells/sample) were co-cultured 1:1 with unin-fected or HSV-infected MDDC labeled with 5,6-carboxyfluorescein diacetatesuccinimidyl ester (CFSE, Molecular Probes, Eugene, OR) or with the PKH67Green Fluorescent Cell Linker Kit (Sigma, St. Louis, MO) in 5 mL round-bot-tom tubes for 4 hours at 37°C, 5%CO2 at a 1:1 ratio.
The fluorescence image-based method used for detecting cell contact andmembrane transfer relies on the spectral isolation of CFSE- or PKH67-labeled MDDC in conjugation with fluorescent labeled pDC (BDCA-2-PE)from images obtained using the ImageStream. In our initial studies, sam-ples were surface stained with both anti-BDCA-2-PE and anti-CD-123-PE-Cy5 to gate on pDC within the pDC enriched samples; however, while it isnecessary to dual-label the pDC to detect them in PBMC, we determinedthat we could identify the same population of pDC in enriched populationsbased on BDCA-2 labeling alone as was detected with the dual label (datanot shown). Therefore, further experiments were carried out in mixed pDCusing only BDCA-2 to identify the pDC. The data obtained were analyzedusing the IDEAS software (Amnis), which quantifies morphometric andphotometric parameters for each cell based on its imagery. All samples werecollected with 20mW of 488 nm and 60mW of 658 nm laser power, 50%height and 256 camera stages. Files of 10,000–20,000 events were collectedfor each sample.
Single color controls were used to create a compensation matrix that wasapplied to all experimental files to correct for spectral crosstalk. Compensateddata in Figures 5–7 were analyzed as follows. First, pDC-containing eventswere identified (R1) on the basis of positive staining for BDCA and an areaconsistent with one dendritic cell (250–1000 pixels). Singlet pDC are smaller(lower SSC area) and dimmer in PKH67/CFSE/GFP (green) intensity (R2)compared to pDC clumps or pDC-MDDC conjugates (R4). The pDC with greenintensity (R3) are candidates for having internalized MDDC-derived cellularmaterial. To quantify these cells, we used the internalization score, which is aratio of green intensity inside the cell to the total green intensity (the higherthe score the greater the concentration of intensity inside the cell). In order toprovide invariance to cell size and to focus the measurement on the brighterportion of green image, only the mean of the upper quartile pixel intensities isused to compute the ratio. The ratio is then weighted by the relative peakintensities inside and in the membrane to capture dim, tiny spots. Finally, theratio is mapped on a log scale to increase the dynamic range to values betweenminus infinity and infinity.
First, the cell is divided into 2 regions, one that defines inside (I) of the celland the other that is outside this mask (B). Next, the mean intensity of the

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
746 N. J. Megjugorac et al.
upper quartile of the pixels in each region is determined. The internalizationscore is then computed as follows:
RESULTS
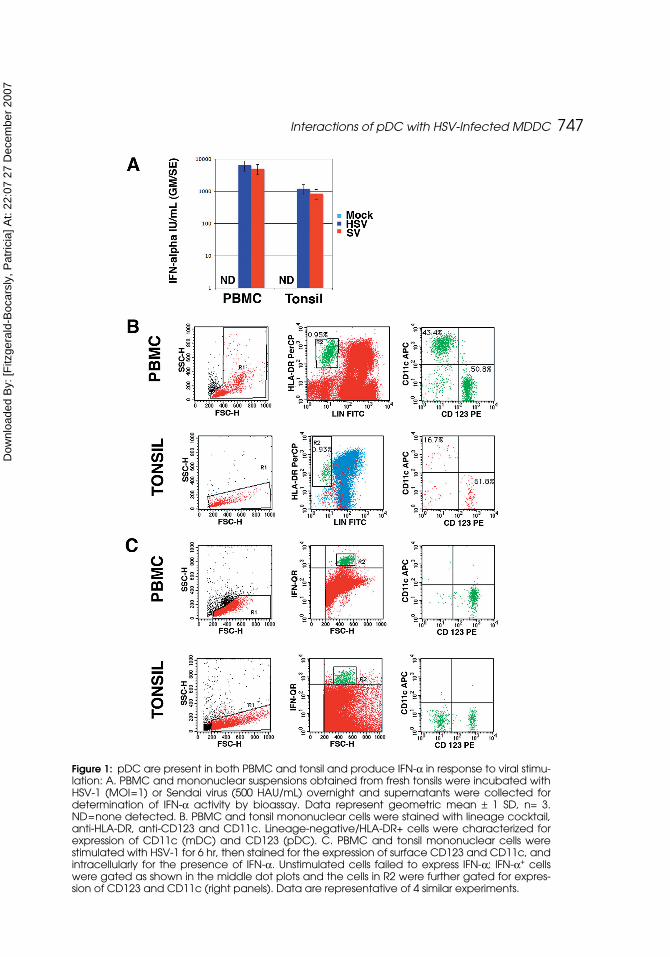
PBMC and tonsil both contain pDC that respond to HSV and Sendai Virus(SV) with IFN-α production PBMC and mononuclear cells obtained by disrup-tion of tonsil were stimulated with HSV-1 or SV for 18 hr, then supernatantswere harvested and tested for IFN-α bioactivity. Neither PBMC nor tonsillarmononuclear cells produced IFN-α in the absence of stimulation, but both popu-lations responded with robust IFN production in response to both HSV and SV(Figure 1A). pDC and mDC can be readily detected in peripheral blood as lin-eage negative, HLA-DR+ cells that are CD123hi/CD11c- and CD123lo/CD11c+
((Feldman et al., 2001) and Figure 1B). Likewise, pDC and mDC were readilydetected in the mononuclear cells obtained after tonsillar disruption (Figure 1B).
To determine the phenotype of the cells that responded to HSV stimulationwith IFN-α production, PBMC and tonsillar mononuclear cells were stimulatedwith HSV, then stained intracellularly for IFN-α. The IFN-α+ cells were thengated for the expression of CD123 and CD11c. CD123+ but not CD11c+ cellsfrom both PBMC and tonsil mononuclear cells that were stimulated with HSVexpressed IFN-α (Figure 1C), whereas unstimulated cells did not (data notshown). Interestingly, there was also a CD123- population of IFN-α-positivecells detected in the tonsil, but, as our emphasis was on pDC vs. mDC, we havenot yet characterized these cells. However, these cells may represent either adistinct IFN-α producing population or pDC that have down-regulated theirCD123. In addition, we stimulated small (1–2 mm3) sections of tonsil directlywith HSV, and these tissue fragments released no IFN-α in the absence ofHSV, but produced between 100 and 3000 IU/mL of IFN-α (mean 889 IU/ml;n=9 fragments for one of two tonsils tested) in the presence of HSV.
PBMC Produce IFN-a in Response to HSV and UV-HSV Stimulated MDDCThe data shown in Figure 1 clearly demonstrate that both peripheral
blood and tonsil contain pDC that are capable of responding to HSV-1 with
Internalizationa
app
where am
m m
m Mean I
i
b
i
i b
i
=−
∗⎛⎝⎜
⎞⎠⎟
=+
=
log , ,1
nntensity of upper quartile pixels in Im Mean Intensity of upper qb
,= uuartile pixels in B
p Peak Intensity of upper quartile pixels in Ii
,= ,,
p Peak Intensity of upper quartile pixels in Bb =
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 747
Figure 1: pDC are present in both PBMC and tonsil and produce IFN-α in response to viral stimu-lation: A. PBMC and mononuclear suspensions obtained from fresh tonsils were incubated withHSV-1 (MOI=1) or Sendai virus (500 HAU/mL) overnight and supernatants were collected fordetermination of IFN-α activity by bioassay. Data represent geometric mean ± 1 SD, n= 3.ND=none detected. B. PBMC and tonsil mononuclear cells were stained with lineage cocktail,anti-HLA-DR, anti-CD123 and CD11c. Lineage-negative/HLA-DR+ cells were characterized forexpression of CD11c (mDC) and CD123 (pDC). C. PBMC and tonsil mononuclear cells werestimulated with HSV-1 for 6 hr, then stained for the expression of surface CD123 and CD11c, andintracellularly for the presence of IFN-α. Unstimulated cells failed to express IFN-α; IFN-α+ cellswere gated as shown in the middle dot plots and the cells in R2 were further gated for expres-sion of CD123 and CD11c (right panels). Data are representative of 4 similar experiments.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
748 N. J. Megjugorac et al.
robust IFN-α production. Similar results were found with cells from humanspleen and lymph nodes (data not shown). While in some instances the pDCresiding in the tissues may be directly exposed to free virions, we reasonedthat they are much more likely to be exposed to virus in the context of aninfected cell carried from the site of infection to the secondary lymphoid tis-sue. MDDC serve as an easily attainable model of the myeloid DC that arelikely candidates to carry virus to draining lymphoid tissue.
MDDC can be infected with HSV-1; they express GFP from GFP-HSV-1within 4–6 hr of infection (see Figure 7), and were found to productively trans-mit the virus to permissive VERO cells (data not shown). In contrast pDC failto express GFP-HSV-1 even after 24 hr of culture (data not shown). We havealso previously demonstrated that although pDC produce high levels of IFN-αin response to HSV-1, MDDC produce low or undetectable levels of IFN-α(Izaguirre et al., 2003). The ability of pDC, but not MDDC, to respond to HSV-1challenge by producing IFN-α provided us a method by which to investigatethe communication between MDDC and pDC and to test the hypothesis thatHSV-infected MDDC could transmit interferongenic signals to pDC.
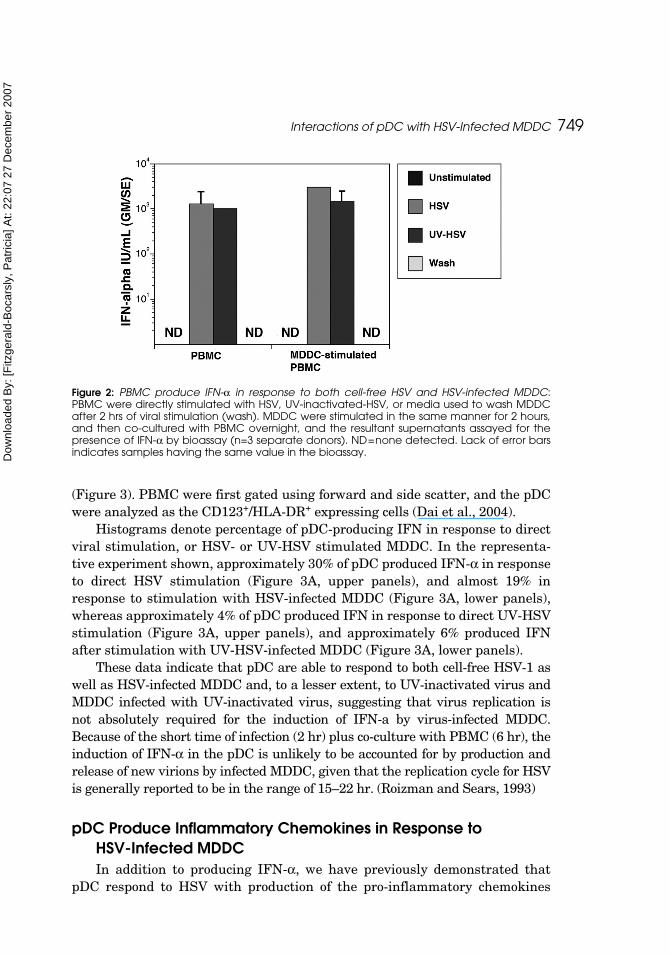
MDDC were either mock-infected or HSV-infected for two hours, washedvigorously, and co-cultured with PBMC overnight at a ratio of 1:5(MDDC:PBMC). To ensure that any IFN-α production by PBMC would not bedue to the release of infectious virions by MDDC, aliquots of the virus wereUV-irradiated to prevent viral replication (resulting in complete loss of GFPexpression and infectivity, data not shown). PBMC were also stimulateddirectly with HSV or UV-HSV as controls, and a portion of the media used towash the infected MDDC was retained to see if it contained any HSV thatmight directly stimulate PBMC. The resultant supernatants were assayed forthe presence of IFN-α via bioassay (Figure 2).
PBMC produced no IFN-α in response to media or wash media controls,but did produce similar amounts of IFN-α in response to direct stimulation byHSV or irradiated virus. HSV or UV-HSV-infected MDDC elicited similarIFN-α production in PBMC as compared to direct stimulation with virus, indi-cating that infected MDDC could deliver an interferongenic signal to the pDC.It should be noted that the production of IFN-α in response to HSV-MDDCwas not MHC-restricted in that autologous as well as heterologous infectedMDDC were able to induce IFN-α production in pDC (data not shown).
pDC Are Responsible for IFN-a Production by HSV-Infected MDDCpDC are known to be the predominant cells among PBMC that produce
IFN-α in response to HSV-1 (Siegal et al., 1999). To determine whether pDC arealso responsible for the IFN-α production to HSV-infected MDDC, PBMC werestimulated with either HSV-infected MDDC or UV-HSV-stimulated MDDC for6 hours and analyzed for IFN production via intracellular flow cytometry
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 749
(Figure 3). PBMC were first gated using forward and side scatter, and the pDCwere analyzed as the CD123+/HLA-DR+ expressing cells (Dai et al., 2004).
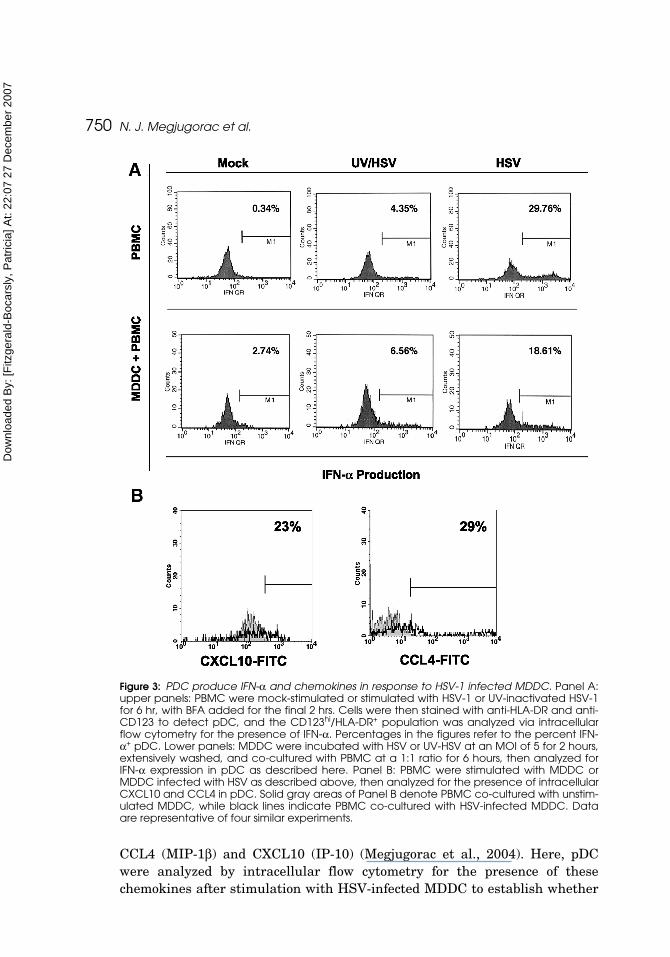
Histograms denote percentage of pDC-producing IFN in response to directviral stimulation, or HSV- or UV-HSV stimulated MDDC. In the representa-tive experiment shown, approximately 30% of pDC produced IFN-α in responseto direct HSV stimulation (Figure 3A, upper panels), and almost 19% inresponse to stimulation with HSV-infected MDDC (Figure 3A, lower panels),whereas approximately 4% of pDC produced IFN in response to direct UV-HSVstimulation (Figure 3A, upper panels), and approximately 6% produced IFNafter stimulation with UV-HSV-infected MDDC (Figure 3A, lower panels).
These data indicate that pDC are able to respond to both cell-free HSV-1 aswell as HSV-infected MDDC and, to a lesser extent, to UV-inactivated virus andMDDC infected with UV-inactivated virus, suggesting that virus replication isnot absolutely required for the induction of IFN-a by virus-infected MDDC.Because of the short time of infection (2 hr) plus co-culture with PBMC (6 hr), theinduction of IFN-α in the pDC is unlikely to be accounted for by production andrelease of new virions by infected MDDC, given that the replication cycle for HSVis generally reported to be in the range of 15–22 hr. (Roizman and Sears, 1993)
pDC Produce Inflammatory Chemokines in Response to HSV-Infected MDDCIn addition to producing IFN-α, we have previously demonstrated that
pDC respond to HSV with production of the pro-inflammatory chemokines
Figure 2: PBMC produce IFN-α in response to both cell-free HSV and HSV-infected MDDC:PBMC were directly stimulated with HSV, UV-inactivated-HSV, or media used to wash MDDCafter 2 hrs of viral stimulation (wash). MDDC were stimulated in the same manner for 2 hours,and then co-cultured with PBMC overnight, and the resultant supernatants assayed for thepresence of IFN-α by bioassay (n=3 separate donors). ND=none detected. Lack of error barsindicates samples having the same value in the bioassay.
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
750 N. J. Megjugorac et al.
CCL4 (MIP-1β) and CXCL10 (IP-10) (Megjugorac et al., 2004). Here, pDCwere analyzed by intracellular flow cytometry for the presence of thesechemokines after stimulation with HSV-infected MDDC to establish whether
Figure 3: PDC produce IFN-α and chemokines in response to HSV-1 infected MDDC. Panel A:upper panels: PBMC were mock-stimulated or stimulated with HSV-1 or UV-inactivated HSV-1for 6 hr, with BFA added for the final 2 hrs. Cells were then stained with anti-HLA-DR and anti-CD123 to detect pDC, and the CD123hi/HLA-DR+ population was analyzed via intracellularflow cytometry for the presence of IFN-α. Percentages in the figures refer to the percent IFN-α+ pDC. Lower panels: MDDC were incubated with HSV or UV-HSV at an MOI of 5 for 2 hours,extensively washed, and co-cultured with PBMC at a 1:1 ratio for 6 hours, then analyzed forIFN-α expression in pDC as described here. Panel B: PBMC were stimulated with MDDC orMDDC infected with HSV as described above, then analyzed for the presence of intracellularCXCL10 and CCL4 in pDC. Solid gray areas of Panel B denote PBMC co-cultured with unstim-ulated MDDC, while black lines indicate PBMC co-cultured with HSV-infected MDDC. Dataare representative of four similar experiments.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 751
the infected MDDC were able to stimulate this function of pDC as well.MDDC were infected for 2 hr, vigorously washed, and co-cultured with PBMCfor 6 hours prior to intracellular analysis of pDC for production of CCL4 andCXCL10. As demonstrated in the representative histograms presented in Fig-ure 3 B, HSV-infected MDDC induced chemokine production in pDC, with23% of cells producing CXCL10, and 29% producing CCL4. These percentagesare consistent with those seen upon direct HSV stimulation [Megjugoracet al., (2004) and data not shown].
Cellular Components from HSV-Infected MDDC Associate with pDC as Demonstrated by Traditional and Imaging Flow CytometryAs demonstrated here, interaction of pDC with HSV-infected pDC results
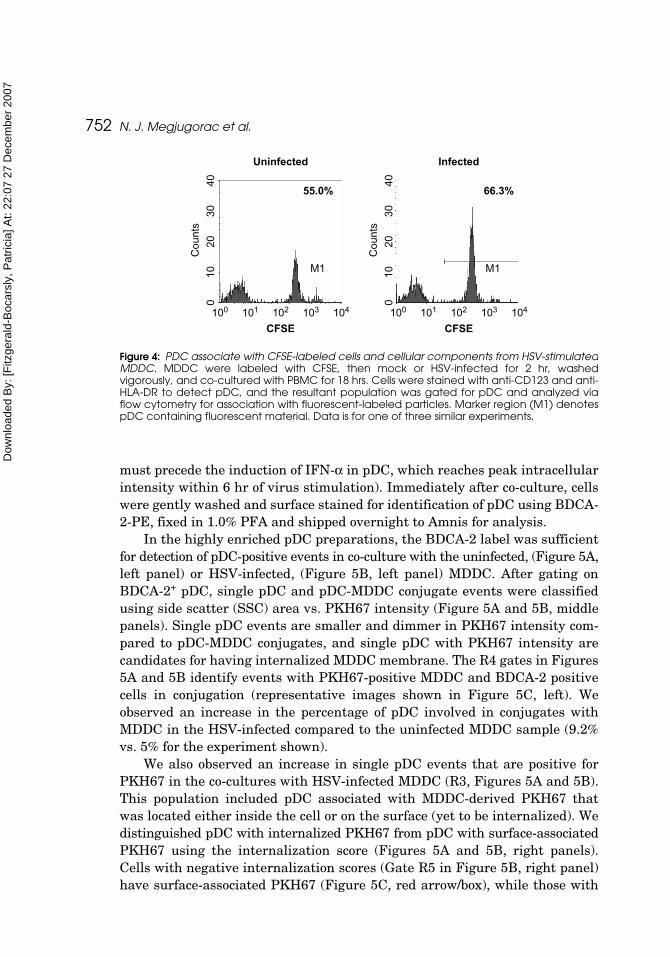
in the production of IFN-α and chemokine production in pDC. For induction ofIFN-α by cell-free HSV, it is known that HSV is internalized into the endo-somes where its DNA signals for IFN-α production through TLR-9 (Hochreinet al., 2004; Krug et al., 2004; Lund et al., 2003). We were therefore interestedin learning whether cellular components from HSV-infected MDDC likewiseassociate with pDC. We first utilized traditional flow cytometry to look forassociation of cellular components from CFSE-labeled, HSV-infected MDDCwith pDC. MDDC were labeled with CFSE, a green fluorescent dye that bindsto cytoplasmic proteins, the cells were infected for 2 hours with HSV, washed,and cultured with PBMC for 18 hr, corresponding to the culture time used forgeneration of supernatant IFN-α, as described here. pDC were analyzed byflow cytometry, and the gated pDC were evaluated for CFSE fluorescencefrom the HSV-infected and uninfected MDDC (Figure 4).
Fluorescent material from both uninfected and HSV-infected MDDC wasfound to associate with the pDC, with a modest preferential association withthe HSV-MDDC over the uninfected MDDC at this time point. However, usingthis methodology, we could not distinguish between extracellular and intracel-lular association of the CFSE-containing cellular components with the pDC. Itcannot be ruled out that, at this 18 hr time point, there is non-specific uptakeof non-cell associated CFSE released from the CFSE-labeled cells as describedin Tario et al. (this issue).
To further investigate the potential for uptake of components frominfected and uninfected MDDC by pDC, we analyzed enriched pDC/MDDCco-cultures using the Amnis ImageStream which combines digital imagerywith traditional flow cytometry. For these studies, MDDC were labeled withPKH67, a green fluorescent lipophilic dye that labels the membranes of cells,and then infected for 2 hr with HSV. The cells were washed and co-cultured ata 1:1 ratio with the partially enriched pDC for 4 hr. (The choice of 4 hr ofco-culture was made because we reasoned that uptake of material by pDC
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
752 N. J. Megjugorac et al.
must precede the induction of IFN-α in pDC, which reaches peak intracellularintensity within 6 hr of virus stimulation). Immediately after co-culture, cellswere gently washed and surface stained for identification of pDC using BDCA-2-PE, fixed in 1.0% PFA and shipped overnight to Amnis for analysis.
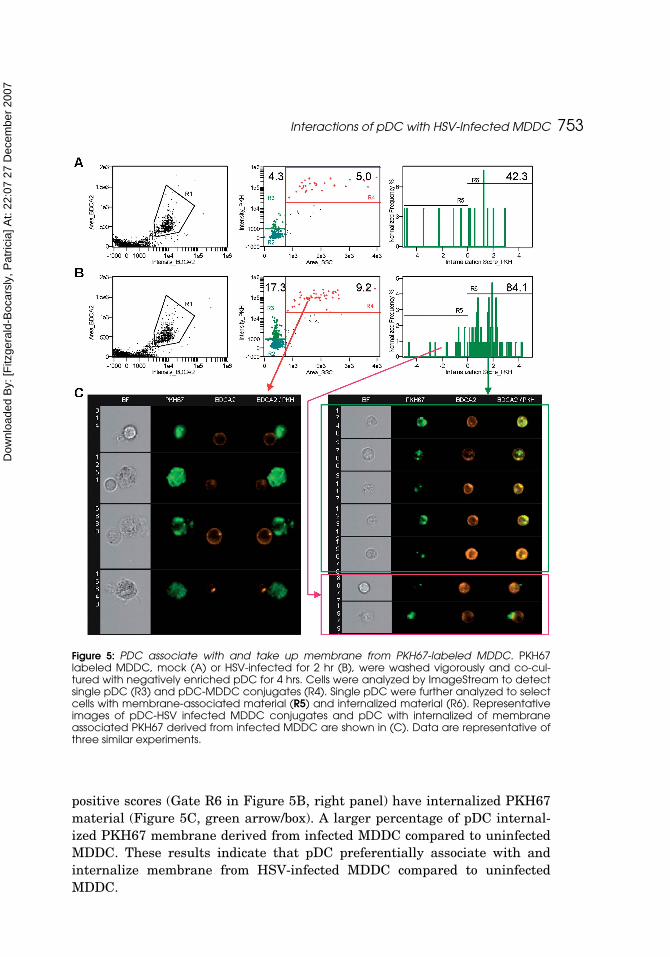
In the highly enriched pDC preparations, the BDCA-2 label was sufficientfor detection of pDC-positive events in co-culture with the uninfected, (Figure 5A,left panel) or HSV-infected, (Figure 5B, left panel) MDDC. After gating onBDCA-2+ pDC, single pDC and pDC-MDDC conjugate events were classifiedusing side scatter (SSC) area vs. PKH67 intensity (Figure 5A and 5B, middlepanels). Single pDC events are smaller and dimmer in PKH67 intensity com-pared to pDC-MDDC conjugates, and single pDC with PKH67 intensity arecandidates for having internalized MDDC membrane. The R4 gates in Figures5A and 5B identify events with PKH67-positive MDDC and BDCA-2 positivecells in conjugation (representative images shown in Figure 5C, left). Weobserved an increase in the percentage of pDC involved in conjugates withMDDC in the HSV-infected compared to the uninfected MDDC sample (9.2%vs. 5% for the experiment shown).
We also observed an increase in single pDC events that are positive forPKH67 in the co-cultures with HSV-infected MDDC (R3, Figures 5A and 5B).This population included pDC associated with MDDC-derived PKH67 thatwas located either inside the cell or on the surface (yet to be internalized). Wedistinguished pDC with internalized PKH67 from pDC with surface-associatedPKH67 using the internalization score (Figures 5A and 5B, right panels).Cells with negative internalization scores (Gate R5 in Figure 5B, right panel)have surface-associated PKH67 (Figure 5C, red arrow/box), while those with
Figure 4: PDC associate with CFSE-labeled cells and cellular components from HSV-stimulatedMDDC. MDDC were labeled with CFSE, then mock or HSV-infected for 2 hr, washedvigorously, and co-cultured with PBMC for 18 hrs. Cells were stained with anti-CD123 and anti-HLA-DR to detect pDC, and the resultant population was gated for pDC and analyzed viaflow cytometry for association with fluorescent-labeled particles. Marker region (M1) denotespDC containing fluorescent material. Data is for one of three similar experiments.
Uninfected
55.0%
CFSE
Counts
010
2030
40
M1 M1
100 101 102 103 104
Counts
010
2030
40
100 101 102 103 104
CFSE
66.3%
Infected
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 753
positive scores (Gate R6 in Figure 5B, right panel) have internalized PKH67material (Figure 5C, green arrow/box). A larger percentage of pDC internal-ized PKH67 membrane derived from infected MDDC compared to uninfectedMDDC. These results indicate that pDC preferentially associate with andinternalize membrane from HSV-infected MDDC compared to uninfectedMDDC.
Figure 5: PDC associate with and take up membrane from PKH67-labeled MDDC. PKH67labeled MDDC, mock (A) or HSV-infected for 2 hr (B), were washed vigorously and co-cul-tured with negatively enriched pDC for 4 hrs. Cells were analyzed by ImageStream to detectsingle pDC (R3) and pDC-MDDC conjugates (R4). Single pDC were further analyzed to selectcells with membrane-associated material (R5) and internalized material (R6). Representativeimages of pDC-HSV infected MDDC conjugates and pDC with internalized of membraneassociated PKH67 derived from infected MDDC are shown in (C). Data are representative ofthree similar experiments.
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
754 N. J. Megjugorac et al.
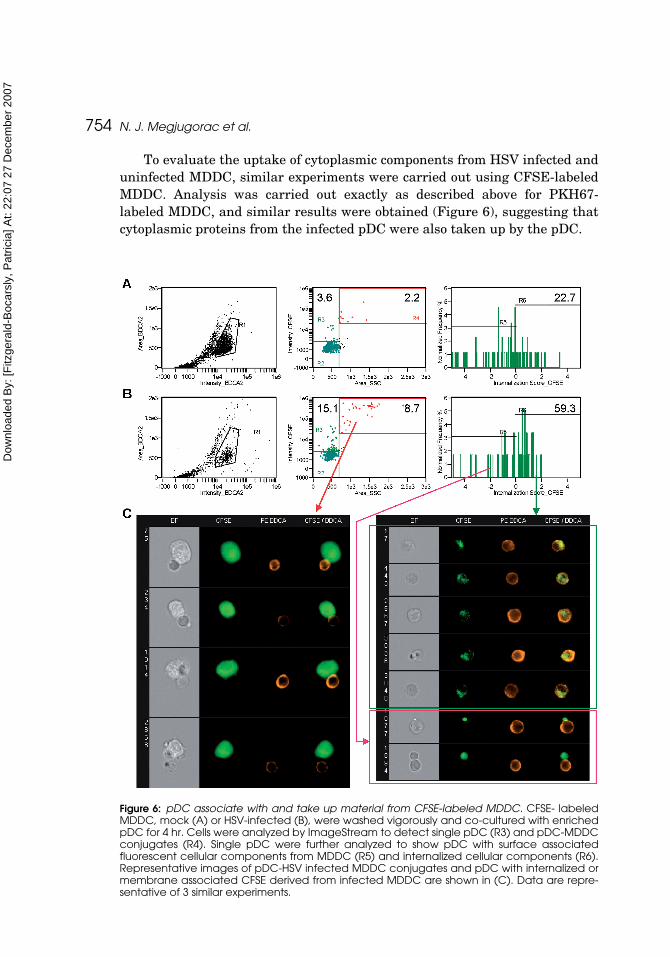
To evaluate the uptake of cytoplasmic components from HSV infected anduninfected MDDC, similar experiments were carried out using CFSE-labeledMDDC. Analysis was carried out exactly as described above for PKH67-labeled MDDC, and similar results were obtained (Figure 6), suggesting thatcytoplasmic proteins from the infected pDC were also taken up by the pDC.
Figure 6: pDC associate with and take up material from CFSE-labeled MDDC. CFSE- labeledMDDC, mock (A) or HSV-infected (B), were washed vigorously and co-cultured with enrichedpDC for 4 hr. Cells were analyzed by ImageStream to detect single pDC (R3) and pDC-MDDCconjugates (R4). Single pDC were further analyzed to show pDC with surface associatedfluorescent cellular components from MDDC (R5) and internalized cellular components (R6).Representative images of pDC-HSV infected MDDC conjugates and pDC with internalized ormembrane associated CFSE derived from infected MDDC are shown in (C). Data are repre-sentative of 3 similar experiments.
Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 755
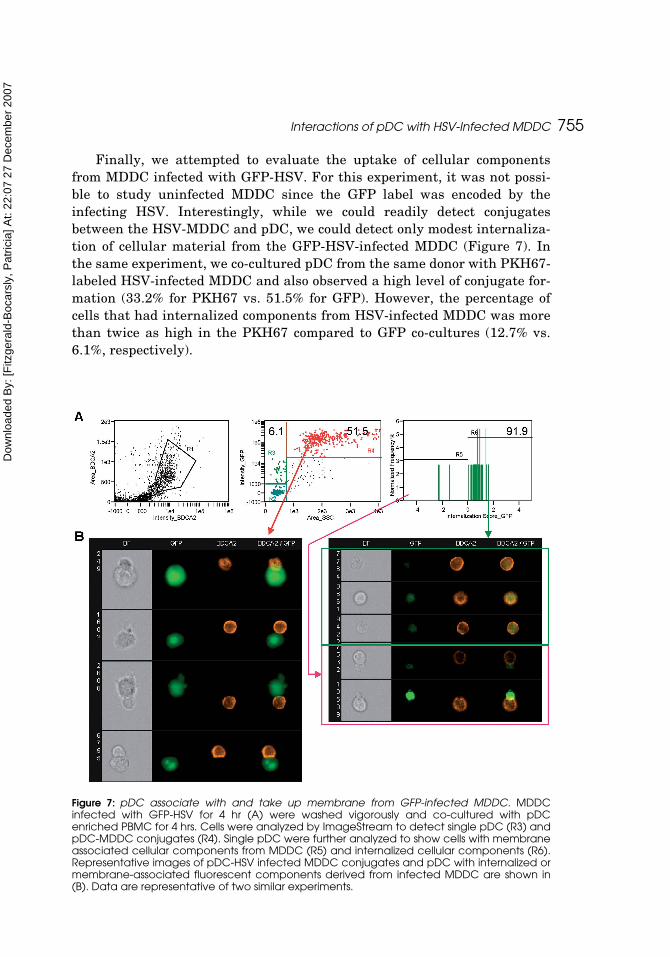
Finally, we attempted to evaluate the uptake of cellular componentsfrom MDDC infected with GFP-HSV. For this experiment, it was not possi-ble to study uninfected MDDC since the GFP label was encoded by theinfecting HSV. Interestingly, while we could readily detect conjugatesbetween the HSV-MDDC and pDC, we could detect only modest internaliza-tion of cellular material from the GFP-HSV-infected MDDC (Figure 7). Inthe same experiment, we co-cultured pDC from the same donor with PKH67-labeled HSV-infected MDDC and also observed a high level of conjugate for-mation (33.2% for PKH67 vs. 51.5% for GFP). However, the percentage ofcells that had internalized components from HSV-infected MDDC was morethan twice as high in the PKH67 compared to GFP co-cultures (12.7% vs.6.1%, respectively).
Figure 7: pDC associate with and take up membrane from GFP-infected MDDC. MDDCinfected with GFP-HSV for 4 hr (A) were washed vigorously and co-cultured with pDCenriched PBMC for 4 hrs. Cells were analyzed by ImageStream to detect single pDC (R3) andpDC-MDDC conjugates (R4). Single pDC were further analyzed to show cells with membraneassociated cellular components from MDDC (R5) and internalized cellular components (R6).Representative images of pDC-HSV infected MDDC conjugates and pDC with internalized ormembrane-associated fluorescent components derived from infected MDDC are shown in(B). Data are representative of two similar experiments.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
756 N. J. Megjugorac et al.
DISCUSSION
Over the past several years, considerable progress has been made towardsunderstanding the pathways involved in IFN-α production by pDC inresponse to viral and synthetic stimuli. Toll-like receptors (TLR) expressed inthe endosomes of pDC confer the ability of these cells to respond with IFN-αproduction to DNA and RNA viruses, with DNA viruses such as HSV-1 signalingthrough TLR9 (Lund et al., 2003; Hochrein et al., 2004) and single-strandedviruses like influenza and HIV-1 signaling through TLR7 (Beignon et al.,2005; Lund et al., 2004; Wang et al., 2006). While it has been known for manyyears that cells in the peripheral blood [now known to be pDC which were firstdescribed in this context (Fitzgerald et al., 1982; Rönnblom and Alm, 1982;Trinchieri et al., 1977)] can produce IFN-α in response to virally infected cells,very little is known about this mechanisms underlying this process.
While myeloid dendritic cells, and in particular MDDC, are very efficientin antigen capture, we recently reported that pDC are also endocytic as dem-onstrated by their ability to take up FITC-dextran, albeit to a lesser extentthan MDDC (Fanning et al., 2006). In the current study, we have studied theinteraction of pDC with myeloid DC utilizing HSV-infected MDDC as ourmodel system. We have demonstrated that pDC are able to acquire surfaceand cytoplasmic material from HSV-infected MDDC and produce IFN-α andthe chemokines CXCL10 and CCL4 in response to these stimulating cells.
Although we could detect interaction of pDC and HSV-infected vs. unin-fected MDDC using traditional flow cytometry by differential labeling of the pDCwith fluorescent antibodies to pDC markers and the MDDC with CFSE, we werenot able to distinguish between conjugates of pDC and MDDC and actual uptakeby pDC of cellular components from the MDDC by this method. However, theseinitial studies provided evidence that the pDC/MDDC interactions could be effec-tively studied over extended periods of time using cell-tracker dyes.
Distinguishing between conjugation and internalization events could havebeen accomplished by fluorescent microscopy, but this technique is limited inthat it is highly qualitative, is extremely sensitive to fluorescent quenchingand does not allow for the statistical robustness of the large sample size thatcan be obtained by flow cytometry. Analysis of the interaction of pDC andMDDC by imaging flow cytometry carried out with the ImageStream allowedus to combine the statistical robustness of the flow cytometer with the imag-ing sensitivity of microscopy. Indeed, we observed that the pDC preferentiallyformed conjugates with the HSV-infected vs. uninfected MDDC and preferen-tially took-up cellular components from the infected vs. the uninfected cells.We noted considerable variability in percentages of pDC forming conjugateswith MDDC among donors, but there was always a higher percentages of con-jugates formed with the infected vs. uninfected MDDC (observed in 8/8donors, for which representative data is shown in Figures 5–7 ).

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 757
This variability may reflect true donor variability, or, possibly, disruptionof some of the conjugates during the antibody staining process for somedonors. The ImageStream analysis also allowed us to distinguish betweenfluorescent particulate cellular components that were internalized by the pDCvs. cellular components external to the pDC. For both infected and uninfectedMDDC, there was a higher degree of internalization of cellular componentsinto pDC rather than particulate material associating with the outside of thepDC, thus strongly implicating a phagocytic or endocytic event. We anticipatethat similar analyses will allow us in the future to carry out mechanistic stud-ies of interaction of infected MDDC with pDC to quantitatively assess the roleof specific cell-surface receptors and ligands in the interaction, uptake andinduction of IFN-α by pDC of virus-infected MDDC. Indeed, we are currentlyutilizing ImageStream analysis to evaluate interaction of pDC with a varietyof different virus-infected cells (Gupta et al., manuscript in preparation).
In the studies reported herein, we found that both the PKH67 and CFSEdyes were suitable for following uptake of cellular material from HSV-infectedpDC. However, as noted by Tario et al. (this issue), there is a possibility ofsecretion of CFSE-labeled proteins soon after labeling that can be taken upover time by non-labeled cells. This possibility should be formally ruled-out inour system in future studies utilizing CFSE-labeled MDDC that are allowedto stabilize for 24 hr prior to infection and co-culture with pDC, especially inthe 18 hr co-culture study presented in Figure 4. For the 4 hr co-cultureassays analyzed with the ImageStream, however, very similar results wereseen with either CFSE or PKH-labeled cells.
Our studies with HSV that expressed GFP upon infection of the MDDCwere not as informative as the CFSE and PKH67 cell tracker studiesdescribed above for several reasons. First, although the GFP in this virus isunder the control of an immediate, early promoter and is therefore expressedvery early in the HSV replicative cycle (Foster et al., 1998), we found that weneeded at least 4 hrs of infection for its unambiguous detection in the MDDC.Second, because the GFP is expressed as a viral gene product, there was noparallel control for uninfected MDDC. Finally, we observed less internalizationof cellular components from GFP-HSV-infected MDDC than from PKH67-labeled MDDC, despite similar levels of conjugate formation for the same pDCdonor. These results do not necessarily reflect a lower uptake of cellular mate-rial from the GFP-HSV-infected MDDC, but, rather, may reflect the quench-ing of GFP at the acid endosomal pH or degradation of the GFP by the acidicproteases.
Myeloid DC have been characterized for their ability to take up cellularcomponents from apoptotic cells. They have been shown to phagocytoseapoptotic bodies via CD36 and αV-β3 and αV-β5 (Albert et al., 1998; Rubartelliet al., 1997), utilizing mechanisms distinct from those employed by macrophages.MDDC have also been shown to sample surface components of neighboring

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
758 N. J. Megjugorac et al.
live cells using scavenger receptors and present them to T cells and elicitimmune responses, a process that has been termed “live cell nibbling”(Harshyne et al., 2001, 2003). MDDC as well as other cell types are also knownto constitutively generate and release exosomes, which are small (60–90 nm)membrane vesicles that mediate cellular interactions and can stimulate T cellresponses (Denzer et al., 2000; Raposo et al., 1996; Zitvogel et al., 1999).
Exosomes are formed by inward budding from the limiting membrane ofendosomal vesicles, and those derived from DC express high levels of MHCClass I and II, and the co-stimulatory molecule, CD86 (Denzer et al., 2000;Pan et al., 1985). Exosomes exhibit a higher level of organization than similarvesicles derived from cells undergoing apoptosis. The membrane blebs thatform apoptotic bodies are heterogeneous and contain an assortment of cytosolic,nuclear, and endoplasmic reticulum-derived proteins (Casciola-Rosen et al.,1994). However, membrane composition of exosomes and apoptotic bodies aresimilar in that the luminal portion of the vesicle faces outward, while the cyto-solic segment faces inward.
It is possible that the pDC are utilizing one or more of these mecha-nisms—live cell nibbling, or recognition of exosomes or apoptotic bodies—forthe uptake of cellular components from infected MDDC. The enhanced levelsof conjugates between pDC and HSV-infected MDDC supports the possibilitythat live cell nibbling may be occurring, while the extracellular association offluorescent particles with the exterior membrane of some pDC suggests thatsub-cellular fragments may also be recognized and taken-up by the pDC; fur-ther studies will be required to distinguish between these possibilities.
Our findings suggest that virally stimulated MDDC elicit a similarresponse from pDC as does direct viral stimulation, with the production ofhigh levels of IFN-α, CCL4 and CXCL10 exactly mirroring the cytokine profileof HSV-stimulated pDC (Megjugorac et al., 2004). It seems unlikely thatnewly formed virus particles transferred along with the MDDC componentsare responsible for the induction of IFN-α in the pDC since MDDC were onlyinfected with HSV-1 for 2 hr prior to the 4- hour co-culture with pDC, insuffi-cient time for production of new infectious progeny (Roizman and Sears,1993). In addition, MDDC “infected” with UV-inactivated HSV also inducedIFN-α in the pDC, arguing against an absolute requirement for viral replica-tion within the MDDC. Moreover, the wash media from the initial MDDCinfection was unable to induce IFN in PBMC, indicating that non-cell associ-ated HSV has been successfully removed from the cultures. Further studieswill be required to determine whether viral DNA associated with the ingestedcellular components or whole viral particles are responsible for the inductionof IFN-α in the pDC that sample HSV-infected pDC.
Physiologically, it makes sense that pDC are able to acquire interferogenicsignals from other cells as well as from cell-free virions. Our data indicatesthat pDC located in the tonsil (Figure 1) and in the spleen and lymph node

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 759
(data not shown) are not constitutively producing IFN-α when freshly isolatedbut are able to respond to viral stimulus with the production of IFN-α. It isplausible that these pDC, which traffic to the lymphoid organs through thehigh endothelial venules, await the delivery of interferogenic cells from mye-loid DC that either pick-up or are infected by virus and then travel to thedraining lymphoid tissue. Given the importance of IFN-α in signaling for Th1cell maturation and for pDC survival and mDC maturation, delivery of thisinterferongenic signal to pDC in the lymph node would be expected to result inthe localized production of IFN-α, leading to reciprocal myeloid DC activationand promotion of antigen presentation for cell-mediated immune responsesresulting in systemic viral clearance.
ACKNOWLEDGMENTS
The work presented in this manuscript was supported by grants R01 AI26806and 1S10RR022411 (for purchase of the Amnis ImageStream) to PFB from theNIH and a pre-doctoral fellowship to NJM from the New Jersey Commissionon Cancer Research.
REFERENCES
Albert, M. L., Pearce, S. F., Francisco, L. M., Sauter, B., Roy, P., Silverstein, R. L.,Bhardwaj, N. (1998). Immature dendritic cells phagocytose apoptotic cells viaalphavbeta5 and CD36, and cross-present antigens to cytotoxic T lymphocytes. J.Exp. Med. 188:1359–1368.
Beignon, A. S., McKenna, K., Skoberne, M., Manches, O., DaSilva, I., Kavanagh, D. G.,Larsson, M., Gorelick, R. J., Lifson, J. D., Bhardwaj, N. (2005). Endocytosis ofHIV-1 activates plasmacytoid dendritic cells via Toll-like receptor-viral RNA inter-actions. J. Clin. Invest. 115:3265–3275.
Casciola-Rosen, L. A., Anhalt, G., Rosen, A. (1994). Autoantigens targeted in systemiclupus erythematosus are clustered in two populations of surface structures on apo-ptotic keratinocytes. J. Exp. Med. 179:1317–1330.
Cella, M., Jarrossay, D., Facchetti, F., Alebardi, O., Nakajima, H., Lanzavecchia, A,Colonna, M. (1999). Plasmacytoid monocytes migrate to inflamed lymph nodes andproduce large amounts of type I interferon. Nat. Med. 5:919–923.
Dai, J., Megjugorac, N. J., Amrute, S. B., Fitzgerald-Bocarsly, P. (2004). Regulationof IFN regulatory factor-7 and IFN-alpha production by enveloped virus andlipopolysaccharide in human plasmacytoid dendritic cells. J. Immunol.173:1535–1548.
Denzer, K., Kleijmeer, M. J., Heijnen, H. F., Stoorvogel, W., Geuze, H. J. (2000). Exo-some: from internal vesicle of the multivesicular body to intercellular signalingdevice. J. Cell. Sci. 113 Pt 19:3365–3374.
Fanning, S. L., George, T. C., Feng, D., Feldman, S. B., Megjugorac, N. J., Izaguirre, A.G., Fitzgerald-Bocarsly, P. (2006). Receptor cross-linking on human plasmacytoiddendritic cells leads to the regulation of IFN-alpha production. J. Immunol.177:5829–5839.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
760 N. J. Megjugorac et al.
Feldman, M., Fitzgerald-Bocarsly, P. (1990). Sequential enrichment and immunocy-tochemical visualization of human interferon-a producing cells. J Interferon Res.10:435–446.
Feldman, S., Stein, D., Amrute, S., Denny, T., Garcia, Z., Kloser, P., Sun, Y.,Megjugorac, N., Fitzgerald-Bocarsly, P. (2001). Decreased interferon-a productionin HIV-infected patients correlates with numerical and functional deficiencies incirculating Type 2 dendritic cell precursors. Clin Imm 101:201–210.
Feldman, S. B., Ferraro, M., Zheng, H. M., N. Patel, N., Gould-Fogerite, S., andFitzgerald-Bocarsly, P. (1994). Viral induction of low frequency interferon-alphaproducing cells. Virology 204:1–7.
Fitzgerald, P. A., von Wussow, P., Lopez, C. (1982). Role of interferon in natural kill ofHSV-1 infected fibroblasts. J. Immunol. 129:819–824.
Fitzgerald-Bocarsly, P. (2002). Natural interferon producing cells: the plasmacytoiddendritic cells. Biotechniques 33:S16–S29.
Fitzgerald-Bocarsly, P., Feng, D. (2007). The role of type I interferon production bydendritic cells in host defense. Biochimie 89:843–855.
Foster, T., Rybachuk, G., Kousouas, K. (1998). Expression of the enhanced green fluo-rescent protein by herpes simplex virus type 1 (HSV) as an in vitro marker forvirus entry and replication. J. Virol. Meth. 75:151–160.
Harshyne, L. A., Watkins, S. C., Gambotto, A., Barratt-Boyes, S. M. (2001). Dendriticcells acquire antigens from live cells for cross-presentation to CTL. J. Immunol.166:3717–3723.
Harshyne, L. A., Zimmer, M. I., Watkins, S., Barratt-Boyes, S. M. (2003). A role forclass A scavenger receptor in dendritic cell nibbling from live cells. J. Immunol.170:2302–2309.
Hochrein, H., Schlatter, B., O'Keeffe, M., Wagner, C., Schmitz, F., Schiemann, M.,Bauer, S., Suter, M., Wagner, H. (2004). Herpes simplex virus type-1 induces IFN-alpha production via Toll-like receptor 9-dependent and -independent pathways.Proc. Natl. Acad. Sci. USA 101:11416–11421.
Izaguirre, A., Barnes, B., Amrute, S., Yeow, Y.-Z., Megjugorac, N., Dai, J., Feng, D.,Chung, E., Pitha, P., Fitzgerald-Bocarsly, P. (2003). Comparative analysis of IRFand IFN-alpha expression in human plasmacytoid and monocyte derived dendriticcells. J. Leuk. Biol. 74:1125–1138.
Krug, A., Luker, G. D., Barchet, W., Leib, D. A., Akira, S., Colonna, M. (2004). Herpessimplex virus type 1 activates murine natural interferon-producing cells throughtoll-like receptor 9. Blood 103:1433–1437.
Lebon, P (1985). Inhibition of herpes simplex virus type-1 induced interferon synthesisby monoclonal antibodies against viral glycoprotein D and by lysosomotropicdrugs. J. Gen. Virol. 66:2781–2786.
Lund, J., Sato, A., Akira, S., Medzhitov, R., Iwasaki, A. (2003). Toll-like receptor9-mediated recognition of Herpes simplex virus-2 by plasmacytoid dendritic cells.J. Exp. Med. 198:513–520.
Lund, J. M., Alexopoulou, L., Sato, A., Karow, M., Adams, N. C., Gale, N. W., Iwasaki, A.,Flavell, R. A. (2004). Recognition of single-stranded RNA viruses by Toll-likereceptor 7. Proc. Natl. Acad. Sci. USA 101:5598–5603.
Megjugorac, N., Young, H. A., Amrute, S., Olshalsky, S., Fitzgerald-Bocarsly, P. (2004).Virally stimulated plasmacytoid dendritic cells produce chemokines and inducemigration of T and NK cells. J. Leukoc. Biol. 75:504–514.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Interactions of pDC with HSV-Infected MDDC 761
Ortyn, W. E., Hall, B. E., George, T. C., Frost, K., Basiji, D. A., Perry, D. J.,Zimmerman, C. A., Coder, D., Morrissey, P. J. (2006). Sensitivity measurementand compensation in spectral imaging. Cytometry A 69:852–862.
Pan, B. T., Teng, K., Wu, C., Adam, M., Johnstone, R. M. (1985). Electron microscopicevidence for externalization of the transferrin receptor in vesicular form in sheepreticulocytes. J. Cell. Biol. 101:942–948.
Penna, G., Vulcano, M., Sozzani, S., Adorini, L. (2002a). Differential migration behaviorand chemokine production by myeloid and plasmacytoid dendritic cells. HumanImmunol. 63:1164–1171.
Penna, G., Vulcano, M., Roncari, A., Facchetti, F., Sozzani, S., Adorini, L. (2002b).Cutting edge: Differential chemokine production by myeloid and plasmacytoiddendritic cells. J. Immunol. 6673–6676.
Raposo, G., Nijman, H. W., Stoorvogel, W., Liejendekker, R., Harding, C. V., Melief, C. J.,Geuze, H. J. (1996). B lymphocytes secrete antigen-presenting vesicles. J. Exp.Med. 183:1161–1172.
Roizman, B., Sears, A. E. (1993). Herpes simplex viruses and their replication. In: Thehuman herpesviruses (Roizman, B., Whitley, R., Lopez, C., eds), pp 11–68. NewYork: Raven Press.
Romani, N., Reider, D., Heuer, M., Ebner, S., Kampgen, E., Eibl, B., Niederwieser, D.,Schuler, G. (1996). Generation of mature dendritic cells from human blood: Animproved method with special regard to clinical applicability. J. Immunol. Meth.196:137–151.
Rönnblom, L., Alm, G. V. (1982). Limiting dilution analysis of human peripheral bloodmononuclear leukocytes that react to human amnion cells and protect theseagainst viral challenge. Eur. J. Immunol. 12:437–441.
Rubartelli, A., Poggi, A., Zocchi, M. R. (1997). The selective engulfment of apoptoticbodies by dendritic cells is mediated by the alpha(v)beta3 integrin and requiresintracellular and extracellular calcium. Eur. J. Immunol. 27:1893–1900.
Siegal, F., Kadowaki, N., Shodell, M., Fitzgerald-Bocarsly, P., Shah, K., Ho, S,Antonenko, A., Liu, Y. J. (1999). The nature of the principal type 1 interferon-producing cells in human blood. Science 284:1835–1837.
Trinchieri, G., Santoli, D., Dee, R. R., Knowles, B. B. (1977). Anti-viral activity inducedby culturing lymphocytes with tumor-derived or virus-transformed cells. J. Exp.Med. 147:1299.
Wang, J. P., Liu, P., Latz, E., Golenbock, D. T., Finberg, R. W., Libraty, D. H. (2006).Flavivirus activation of plasmacytoid dendritic cells delineates key elements ofTLR7 signaling beyond endosomal recognition. J. Immunol. 177:7114–7121.
Zitvogel, L., Fernandez, N., Lozier, A., Wolfers, J., Regnault, A., Raposo, G.,Amigorena, S. (1999). Dendritic cells or their exosomes are effective biotherapies ofcancer. Euro. J. Cancer 35 Suppl 3:S36–38.

Dow
nloa
ded
By:
[Fitz
gera
ld-B
ocar
sly,
Pat
ricia
] At:
22:0
7 27
Dec
embe
r 200
7
Related Documents











