Heredity 62 (1989) 169—176 The Genetical Society of Great llritain Received 24 May 1988 Genetic studies of laboratory reared Mytilus edulis. II. Selection at the leucine amino peptidase (Lap) locus A. R. Beaumont, C. M. Beveridge, E. A. Barnet and M. D. Budd School of Ocean Sciences, University College of North Wales, Menai Bridge, Gwynedd LL59 5EH, U.K. Offspring from several families and a mass mating of Mytilus edulis were cultured in the laboratory up to the juvenile stage under different temperature regimes. Mussels were genotyped at the leucine aminopeptidase (Lap) locus at the spat stage (post-metamorphosis) using cellulose acetate electrophoresis and at the juvenile stage using starch gel electrophoresis. Significant deviations from expected genotype frequencies were found in many cultures at both stages and certain genotypes were generally found to be favoured irrespective of temperature regime during culture. Estimates of relative fitnesses of the six common Lap genotypes suggest overdominance for the alleles Lap94 and Lap and semi-dominance for Lap98, but calculations based on this array of estimated fitnesses indicate that Lap98 would eventually become fixed in a panmictic population. Since a balanced Lap polymorphism exists in the wild we conclude that selection differentials at the Lap locus in laboratory cultures may be very different from those in nature, and suggest that nutritional constraints may account for some of these differences in selection. Also, in contrast to wild populations, the laboratory cultures showed a significant excess of Lap heterozygotes. INTRODUCTION The leucine aminopeptidase (Lap) locus which codes for an a-aminoacylpeptide hydrolase (Young et a!., 1979) in Mytilus edulis has been extensively studied in recent years (for reviews see Levinton and Koehn, 1976; Koehn, 1985; Hilbish and Koehn, 1985). Five Lap alleles have been detected, although only three are common in any one area (Levinton and Koehn, 1976). In UK waters and on the eastern coast of America, south of Cape Cod, Lap94, Lap96, and Lap98 are the commonest alleles (Koehn et a!., 1976; Skibinski et a!., 1983; Ahmad and Hedrick, 1985). In Long Island Sound, New York, there is a dine in the frequency of Lap94 that is associated with changes in salinity along the Sound. Under varying salinities, biochemical differences in Lap allo- zymes produce genotype specific rates of change in the free amino acid pool which is an important element in cell volume regulation (Hilbish et al., 1982). In a detailed study of this dine, Hilbish and Koehn (1985) concluded that a high degree of dominance existed for the biochemical and physio- logical phenotypic expression of Lap94 genotypes and that a selection model incorporating domin- ance gave a close fit to observed Lap genotype frequencies among natural populations in the dine. This paper reports on genotype specific mor- talities at the Lap locus in offspring from a number of families and a mass mating of M. edulis which were reared in the laboratory at different tem- peratures but at constant salinity. Evidence is pro. vided for selection at the Lap locus, and estimates are made of the relative fitnesses of Lap genotypes. MATERIALS AND METHODS Ripe M. edulis were collected in the Menai Strait, North Wales, UK, during 1984 and 1985 and cul- tures of larvae were reared as described by Beau- mont et a!. (1988). Larvae and juveniles from several families (Ml, M2, M3 and M4) and a mass mating (MM) were cultured under a number of temperature regimes. The temperature regime used for each culture is indicated in tables 1—4. Samples for electrophoresis were taken at the spat stage (500-1000 pm) in all cultures, and again at the

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Heredity 62 (1989) 169—176The Genetical Society of Great llritain Received 24 May 1988
Genetic studies of laboratory rearedMytilus edulis.II. Selection at the leucine aminopeptidase (Lap) locusA. R. Beaumont,C. M. Beveridge,E. A. Barnet andM. D. Budd
School of Ocean Sciences,University College of North Wales,Menai Bridge, Gwynedd LL59 5EH, U.K.
Offspring from several families and a mass mating of Mytilus edulis were cultured in the laboratory up to the juvenilestage under different temperature regimes. Mussels were genotyped at the leucine aminopeptidase (Lap) locus at thespat stage (post-metamorphosis) using cellulose acetate electrophoresis and at the juvenile stage using starch gelelectrophoresis. Significant deviations from expected genotype frequencies were found in many cultures at both stagesand certain genotypes were generally found to be favoured irrespective of temperature regime during culture. Estimatesof relative fitnesses of the six common Lap genotypes suggest overdominance for the alleles Lap94 and Lap andsemi-dominance for Lap98, but calculations based on this array of estimated fitnesses indicate that Lap98 wouldeventually become fixed in a panmictic population. Since a balanced Lap polymorphism exists in the wild we concludethat selection differentials at the Lap locus in laboratory cultures may be very different from those in nature, andsuggest that nutritional constraints may account for some of these differences in selection. Also, in contrast to wildpopulations, the laboratory cultures showed a significant excess of Lap heterozygotes.
INTRODUCTION
The leucine aminopeptidase (Lap) locus whichcodes for an a-aminoacylpeptide hydrolase(Young et a!., 1979) in Mytilus edulis has beenextensively studied in recent years (for reviews seeLevinton and Koehn, 1976; Koehn, 1985; Hilbishand Koehn, 1985). Five Lap alleles have beendetected, although only three are common in anyone area (Levinton and Koehn, 1976). In UKwaters and on the eastern coast of America, southof Cape Cod, Lap94, Lap96, and Lap98 are thecommonest alleles (Koehn et a!., 1976; Skibinskiet a!., 1983; Ahmad and Hedrick, 1985). In LongIsland Sound, New York, there is a dine in thefrequency of Lap94 that is associated with changesin salinity along the Sound. Under varyingsalinities, biochemical differences in Lap allo-zymes produce genotype specific rates of changein the free amino acid pool which is an importantelement in cell volume regulation (Hilbish et al.,1982). In a detailed study of this dine, Hilbish andKoehn (1985) concluded that a high degree ofdominance existed for the biochemical and physio-logical phenotypic expression of Lap94 genotypes
and that a selection model incorporating domin-ance gave a close fit to observed Lap genotypefrequencies among natural populations in thedine.
This paper reports on genotype specific mor-talities at the Lap locus in offspring from a numberof families and a mass mating of M. edulis whichwere reared in the laboratory at different tem-peratures but at constant salinity. Evidence is pro.vided for selection at the Lap locus, and estimatesare made of the relative fitnesses of Lap genotypes.
MATERIALS AND METHODS
Ripe M. edulis were collected in the Menai Strait,North Wales, UK, during 1984 and 1985 and cul-tures of larvae were reared as described by Beau-mont et a!. (1988). Larvae and juveniles fromseveral families (Ml, M2, M3 and M4) and a massmating (MM) were cultured under a number oftemperature regimes. The temperature regime usedfor each culture is indicated in tables 1—4. Samplesfor electrophoresis were taken at the spat stage(500-1000 pm) in all cultures, and again at the
170 A. R. BEAUMONT, C. M. BEVERIDGE, E A. BARNET AND M. D. BUDD
juvenile stage in some of the cultures. Techniquesfor cellulose acetate microelectrophoresis of spat,starch gel electrophoresis of juveniles and stainingfor Lap are detailed in Beaumont et a!. (1988).
Expected Lap genotype and allele frequenciesin the offspring from single families were calcu-lated from the known genotypes of the parents.Expected genotype and allele frequencies for themass mating were calculated on the assumption ofequal and random mixing of all gametes from the15 mussels (nine males, six females) spawned.Evidence for equal gametic contribution of allprogenitors in this mass mating are presented in aprevious paper (Beaumont et a!., 1988). Deviationsof observed from expected genotype and allelefrequencies were tested for significance using thex2 goodness of fit test.
RESULTS
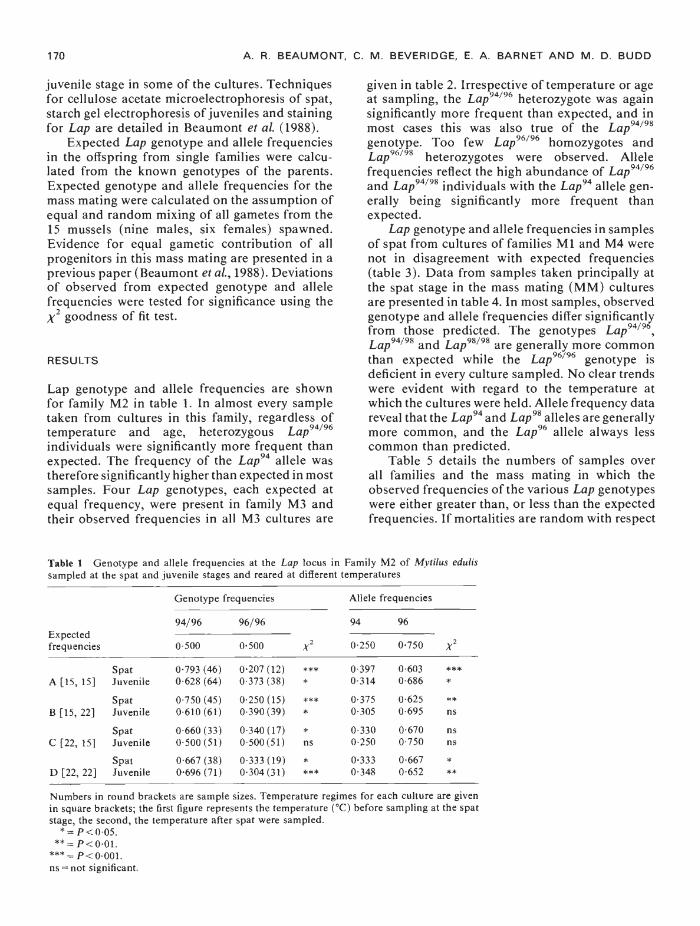
Lap genotype and allele frequencies are shownfor family M2 in table 1. In almost every sampletaken from cultures in this family, regardless oftemperature and age, heterozygous Lap94"96individuals were significantly more frequent thanexpected. The frequency of the Lap94 allele wastherefore significantly higher than expected in mostsamples. Four Lap genotypes, each expected atequal frequency, were present in family M3 andtheir observed frequencies in all M3 cultures are
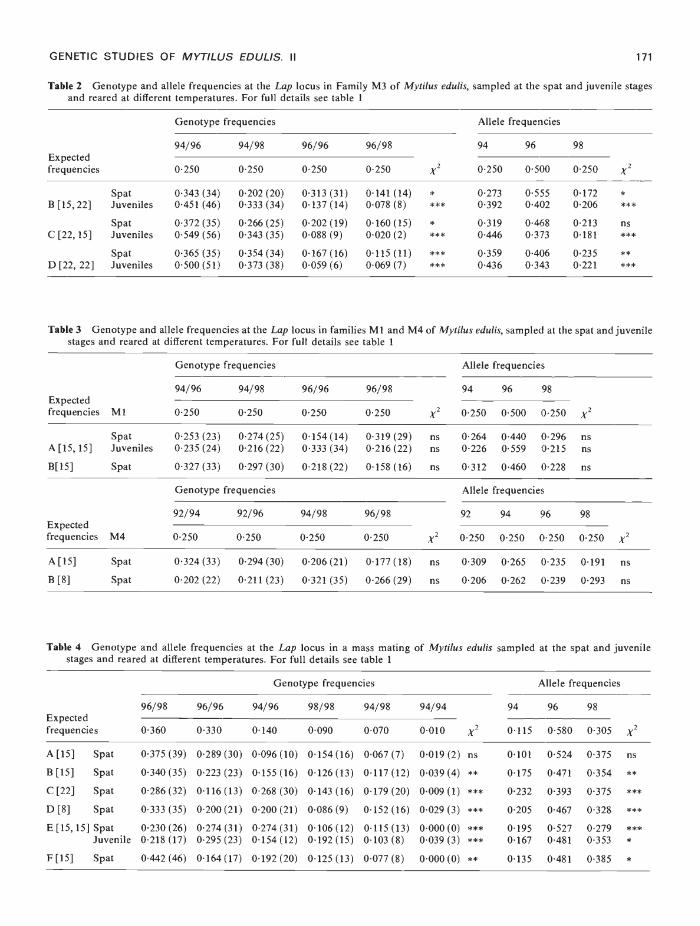
given in table 2. Irrespective of temperature or ageat sampling, the Lap94196 heterozygote was againsignificantly more frequent than expected, and inmost cases this was also true of the Lap94"98genotype. Too few Lap96"96 homozygotes andLap96195 heterozygotes were observed. Allelefrequencies reflect the high abundance of Lap9496and Lap94"98 individuals with the Lap94 allele gen-erally being significantly more frequent thanexpected.
Lap genotype and allele frequencies in samplesof spat from cultures of families Ml and M4 werenot in disagreement with expected frequencies(table 3). Data from samples taken principally atthe spat stage in the mass mating (MM) culturesare presented in table 4. In most samples, observedgenotype and allele frequencies differ significantlyfrom those predicted. The genotypes Lap94"96,Lap94"98 and Lap95"95 are generally more commonthan expected while the Lap96"96 genotype isdeficient in every culture sampled. No clear trendswere evident with regard to the temperature atwhich the cultures were held. Allele frequency datareveal that the Lap94 and Lap98 alleles are generallymore common, and the Lap96 allele always lesscommon than predicted.
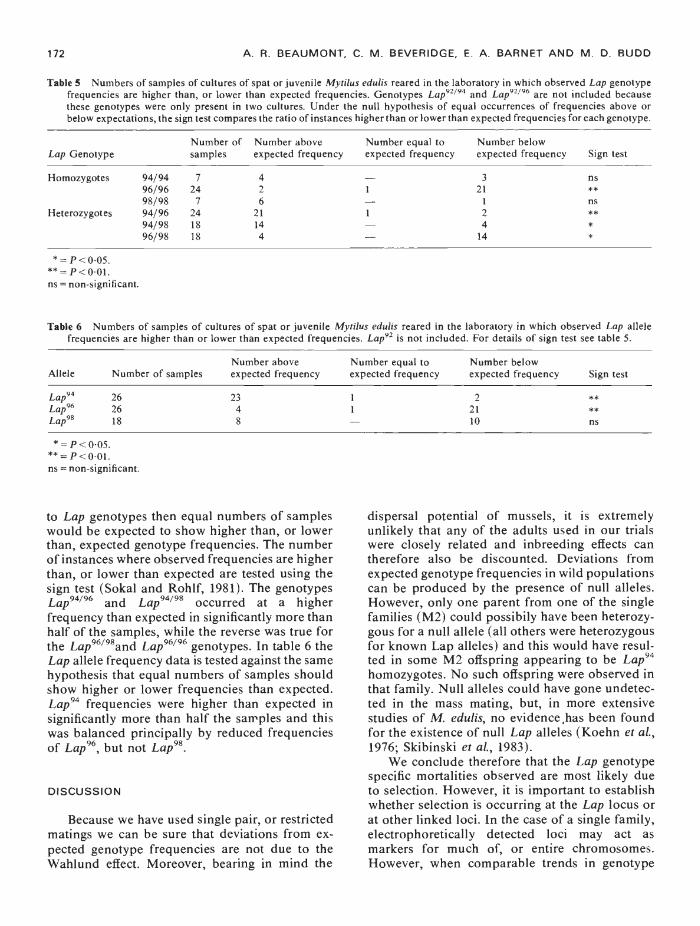
Table 5 details the numbers of samples overall families and the mass mating in which theobserved frequencies of the various Lap genotypeswere either greater than, or less than the expectedfrequencies. If mortalities are random with respect
Table I Genotype and allele frequencies at the Lap locus in Family M2 of Mytilus edulissampled at the spat and juvenile stages and reared at diflerent temperatures
Expectedfrequencies
Genotype frequencies Allele frequencies
94/96 96/96
x2
94
0250
96
0750 X20500 0500
A [15, 15]SpatJuvenile
0793 (46)0628 (64)
0207 (12)0373 (38)
****
03970314
06030686
****
B [15, 22]SpatJuvenile
0750 (45)0610 (61)
0250 (15)0390 (39)
****
03750305
06250695
**ns
C [22, 15]SpatJuvenile
0660 (33)0500 (51)
0340 (17)0.500 (51)
*ns
03300250
06700750
nsns
D [22, 22]SpatJuvenile
0667 (38)0696 (71)
0333 (19)0304 (31)
***'
03330348
06670652
***
Numbers in round brackets are sample sizes. Temperature regimes for each culture are givenin square brackets; the first figure represents the temperature ('C) before sampling at the spatstage, the second, the temperature after spat were sampled.
"= P<005.** P<0.01.
= P<0.001.ns = not significant.
GENETIC STUDIES OF MYTILUS EDULIS. II 171
Expectedfrequencies
B [15, 22]SpatJuveniles
C [22, 15]SpatJuveniles
D [22, 22]SpatJuveniles
Allele frequencies
Table 2 Genotype and allele frequencies at the Lap locus in Family M3 of Mytilus edulis, sampled at the spat and juvenile stagesand reared at different temperatures. For full details see table 1
Genotype frequencies
94/96 94/98
0-250 0250
0-343 (34) 0202 (20)0-451 (46) 0-333 (34)
0372 (35) 0266 (25)0549 (56) 0343 (35)
0365 (35) 0-354 (34)0500 (51) 0373 (38)
96/96 96/98
x2
94 96 98
0250 0500 0250 x20250 0250
0313 (31)0137 (14)
0141 (14)0078 (8)
****
02730392
0-5550402
01720206
****
0.202(19)0-088(9)
0160(15)0-020(2)
*** 0319
0-44604680373
02130181
ns***
0.167 (16)0059 (6)
0-115(11)0-069 (7)
******
03590436
04060-343
02350221
*****
Table 4 Genotype and allele frequencies at the Lap locus in a mass mating of Mytilus edulis sampled at the spat and juvenilestages and reared at different temperatures. For full details see table I
Genotype frequencies
96/98 96/96 94/96 98/98 94/98 94/94Expectedfrequencies 0-360 0-330 0-140 0-090 0-070 0-010 x2
A [15] Spat 0-375 (39) 0-289 (30) 0-096 (10) 0-154(16) 0-067 (7) 0-019 (2) ns
B[15] Spat 0-340(35) 0-223(23) 0-155(16) 0-12603) 0-11702) 0-039(4) **C[22] Spat 0-28602) 0-11603) 0-26800) 0-14306) 0-179(20) 0-0090) mD [8] Spat 0-333 (35) 0-200 (21) 0-200 (21) 0-086 (9) 0-15206) 0-029 (3) ***E [15, 15] Spat 0-230 (26) 0-274 (31) 0-274 (31) 0-106 (12) 0-115 (13) 0-000(0) ***
Juvenile 0-218(17) 0-295(23) 0-154(12) 0-192(15) 0-103(8) 0-039(3)
F[15] Spat 0-442(46) 0-16407) 0-192(20) 0-12503) 0-077(8) 0-000(0) **
Table 3 Genotype and allele frequencies at the Lap locus in families Ml and M4 of Mytilus edulis, sampled at the spat and juvenilestages and reared at different temperatures. For full details see table 1
Expected
frequencies Ml
Genotype frequencies Allele frequencies
94/96 94/98 96/96 96/98
x2
94
0250
96
0500
98
0250 x20-250 0-250 0-250 0-250
Spat 0-253 (23) 0-274 (25) 0-154(14) 0-319 (29) ns 0-264 0-440 0-296 nsA [15, 15] Juveniles 0-235 (24) 0-216 (22) 0-333 (34) 0-216 (22) ns 0-226 0-559 0-215 ns
B[l5] Spat 0-327 (33) 0-297 (30) 0-218 (22) 0-158 (16) as 0-312 0-460 0-228 ns
Expectedfrequencies M4
Genotype frequencies Allele frequencies
92/94 92/96 94/98 96/98
x2
92 94 96 98
0-250 0-250 0-250 0-250 0-250 0-250 0-250 0-250 x2
A [15] Spat 0-324 (33) 0-294 (30) 0-206 (21) 0-177 (18) ns 0-309 0-265 0-235 0-191 ns
B [8] Spat 0-202 (22) 0-211 (23) 0-321 (35) 0-266 (29) ns 0-206 0-262 0-239 0-293 ns
Allele frequencies
94 96 98
0-115 0-580 0-305 x2
0-101 0-524 0-375 ns
0-175 0-471 0-354 **
0-232 0-393 0-375 ***
0-205 0-467 0-328 ***
0-195 0-527 0-279 ***
0-167 0-481 0-353 *
0-135 0-481 0-385 *
172 A. R. BEAUMONT, C. M. BEVERIDGE, E. A. BARNET AND M. D. BUDD
Table 5 Numbers of samples of cultures of spat or juvenile Mytilus edulis reared in the laboratory in which observed Lap genotypefrequencies are higher than, or lower than expected frequencies. Genotypes Lap°2194 and Lap°2196 are not included becausethese genotypes were only present in two cultures. Under the null hypothesis of equal occurrences of frequencies above orbelow expectations, the sign test compares the ratio of instances higher than or lower than expected frequencies for each genotype.
Lap GenotypeNumber ofsamples
Number aboveexpected frequency
Number equal toexpected frequency
Number belowexpected frequency Sign test
Homozygotes 94/9496/9698/98
724
7
4
2
6
—1
—
321
1
ns**fls
Heterozygotes 94/9694/9896/98
241818
21144
1
——.
24
14
****
9—P<005** = P<0•01.ns =non-significant.
Table 6 Numbers of samples of cultures of spat or juvenile My/i/us edulis reared in the laboratory in which observed Lap allelefrequencies are higher than or lower than expected frequencies. Lap92 is not included. For details of sign test see table 5.
Allele Number of samplesNumber aboveexpected frequency
Number equal toexpected frequency
Number belowexpected frequency Sign test
Lap94 26 23 1 2 9*
Lap°6 26 4 1 21 **Lap°8 18 8 10 ns
9=P<005.*9 =ns = non-significant.
to Lap genotypes then equal numbers of sampleswould be expected to show higher than, or lowerthan, expected genotype frequencies. The numberof instances where observed frequencies are higherthan, or lower than expected are tested using thesign test (Sokal and Rohlf, 1981). The genotypesLap94"96 and Lap94798 occurred at a higherfrequency than expected in significantly more thanhalf of the samples, while the reverse was true forthe Lap96198and Lap96796 genotypes. In table 6 theLap allele frequency data is tested against the samehypothesis that equal numbers of samples shouldshow higher or lower frequencies than expected.Lap94 frequencies were higher than expected insignificantly more than half the samples and thiswas balanced principally by reduced frequenciesof Lap96, but not Lap98.
DISCUSSION
Because we have used single pair, or restrictedmatings we can be sure that deviations from ex-pected genotype frequencies are not due to theWahiund effect. Moreover, bearing in mind the
dispersal potential of mussels, it is extremelyunlikely that any of the adults used in our trialswere closely related and inbreeding effects cantherefore also be discounted. Deviations fromexpected genotype frequencies in wild populationscan be produced by the presence of null alleles.However, only one parent from one of the singlefamilies (M2) could possibily have been heterozy-gous for a null allele (all others were heterozygousfor known Lap alleles) and this would have resul-ted in some M2 offspring appearing to be Lap94homozygotes. No such offspring were observed inthat family. Null alleles could have gone undetec-ted in the mass mating, but, in more extensivestudies of M. edulis, no evidence has been foundfor the existence of null Lap alleles (Koehn et a!.,1976; Skibinski et a!., 1983).
We conclude therefore that the Lap genotypespecific mortalities observed are most likely dueto selection. However, it is important to establishwhether selection is occurring at the Lap locus orat other linked loci. In the case of a single family,electrophoretically detected loci may act asmarkers for much of, or entire chromosomes.However, when comparable trends in genotype

GENETIC STUDIES OF MYTILUS EDULIS. II 173
frequencies at a locus are seen across progeny fromunrelated families and also in offspring from amass mating then that locus is less likely to beacting as a marker for more than a small portionof the chromosome tightly linked to the locus.Beaumont et a!. (1983, 1988) have reported datain which significant variations in genotypefrequencies at a number of loci occur betweenunrelated families of M. edulis. However, this isnot the case at the Lap locus in the culturesdescribed here where particular Lap genotypesoccur consistently at lower or higher frequenciesthan expected.
There is strong genetic, physiological and bio-chemical evidence for direct selection at the Laplocus in relation to salinity (Hilbish and Koehn,1985). However, the fact that Lap genotypes mayhave different fitnesses in relation to salinity doesnot rule out the possibility that linked loci may beinvolved in expressing the apparent differentialfItnesses at the Lap locus detailed here. Lap allelesor genotypes may impose no fitness constraints atoceanic salinity and therefore may be effectivelyneutral in situations other than at low salinity. Onthe other hand, it seems probable that fitness con-straints imposed by a locus, or loci closely linkedto Lap would have effects over a range of salinitiesrather than being restricted to high salinity. Indeedeffects are more likely under the stresses imposedby low salinity. It is difficult to reconcile this withthe strong biochemical and physiological evidencewhich points to direct selection at the Lap locusin low salinities.
One further possibility stems from the work ofDay et a!. (1982) who demonstrated that thealcohol dehydrogenase (Adh) locus is associatedwith a chromosome inversion polymorphism in theseaweed fly Coelopa frigida, such that consistentsignificant differences from Hardy-Weinbergexpectations recorded at the Adh locus are mostlikely due to the effect of linked loci within theinversion rather than to the Adh locus itself.Mussels do not possess polytene chromosomes andtherefore the presence of chromosomal inversionscan best be detected by observing bivalents duringmeiosis. As far as we know, no systematic searchhas yet been undertaken for evidence of inversionsduring meiosis in mussels although mitoticchromosomes are beginning to be studied in moredetail (Dixon and Clarke, 1982; Dixon et al., 1986).We cannot therefore rule out the possible associ-ation of the Lap locus with other linked loci withina chromosome inversion.
However, in the light of our earlier arguments,and in the absence of evidence of inversion poly-
morphisms, we feel that there is a high probabilitythat the apparent differences in fitness of Lapgenotypes observed in our experiments are directlyrelated to the Lap locus rather than to other closelylinked loci in general.
The results clearly show that, in general, theLap94196, Lap94198 and Lap98198 genotypes are morefrequent, while the Lap96196 and Lap96198 are lessfrequent than expected. There is no suggestion inthe data that the temperature regime to which aculture was exposed had any significant effect ongenotype frequencies.
Using the genotype frequency data it is possibleto estimate approximate relative fitnesses for thesix common Lap genotypes. Estimates wereobtained by calculating for each genotype, themean percentage, over all families and the massmating, by which observed frequencies deviatedfrom expected frequencies. In the case of Lap94194,which was only present in the mass mating, themean proportion of Lap94'94 individuals (0.018+/ 0O10, 95 per cent CI) over all cultures was notsignificantly different from the expected propor-tion (0.01). This genotype was therefore given afitness of 1. The relative fitnesses of all six Lapgenotypes are given in Table 7.
Table 7 Relative fitnesses of six Lap genotypes estimated fromobserved frequencies in up to 24 cultures from four singlefamilies and one mass mating
Lap genotype 94/94 96/96 98/98 94/96 94/98 96/98
Relative fitness 1-000 0-701 1480 1406 1-350 0725
Hilbish and Koehn (1985) have proposed thata dominance model is most appropriate to explaintheir data on salinity related selection at the Laplocus in Long Island Sound. Individuals whichwere either homozygous or heterozygous for Lap94exhibited similar biochemical and physiologicalphenotypes that were equally less fit compared tonon-Lap94 genotypes. Furthermore, analyses ofheterozygote deficiencies in genotype frequencydata produced results which most closely fitted aselection model incorporating dominance. Therelative fitnesses at the Lap locus estimated fromour data would suggest overdominance rather thandominance, with regard to Lap94 and Lap96. Lap94and Lap96 heterozygotes had higher fitnesses thantheir respective homozygotes. On the other handthe Lap98 heterozygote classes were less fit thanthe Lap98198 homozygote genotype and the inter-mediate fitnesses of the two heterozygotes indicatessemi-dominance in the case of Lap98.

174 A. R. BEAUMONT, C. M. BEVERIDGE, E. A. BARNET AND M. ft BUDD
Theoretical considerations (Crow, 1986) sug-gest that, if polymorphisms are maintained byselection, then dominance or semi-dominance willlead ultimately to fixation of particular alleles.Only overall overdominance can produce a stableequilibrium between all alleles. The dominancereported by Hilbish and Koehn (1985) for Lap94at low salinity depends upon an annual influx ofrecruits from oceanic populations of mussels witha high frequency of Lap94. Without such imput,Lap94 would theoretically become lost in estuarinepopulations of mussels. Beaumont et al. (1988)have suggested that there may be selection for,rather than against Lap94 during the larval stageat low salinity and that ontogenetic changes infitness may be a feature in the maintenance of theLap polymorphism. However, there is no sugges-tion of significant ontogenetic differences in fitnessof Lap genotypes in the data presented here whichwere collected from cultures reared entirely atoceanic salinities.
Table 8 A model for selection at a locus with three alleles:A1, A2 and A2
Genotype A1A1 A2A, A1A3 A1A2 A1A3Fitness VV 'V13 'I2 I3
p p 2PIPIFrequency p
Using a standard model for selection at a locuswith three alleles (table 8, Crow, 1986) it is possibleto calculate what the frequencies of the three alleleswould be at equilibrium in a panmictic populationin Hardy—Weinberg equilibrium, assuming that therelative fitnesses proposed were applicable andwere constant. The equilibrium frequencies ofLap94 (Pi), Lap96 (P2) and Lap98 (p3) are calculatedas follows (Crow, 1986).
1) 1)3Pi = P2 and p3 =
= (14"2— 1''22)(l4'13 4"3)
—( '12 123)(4T13. "23)
D2=(W23- W3)( W12- W11)
-(W23 W13)( W12- W31)
D3=(W13- W1)(W23— W22)
-(W13- W12)( W23- W1)
and D1 + D2 + 1)3 = D. W1 = fitness of Lap94194,W12 = fitness of Lap94196 etc. A further quantity, 'T
T = VV1— 1"I3 — "13 — 'V33.
For an equilibrium to be stable, D1, 2 and D3must be positive numbers and T negative. Usingthe fitness values estimated here, the calculatedallele frequencies at equilibrium are Lap94 = 0632,Lap96=0332 and Lap98=0036, but although Tis a negative number, D1, D2 and D3 are alsonegative. In a panmictic population therefore,these relative fitnesses would lead eventually tofixation of one allele. In a stable equilibrium withthree alleles, the mean fitness of the populationshould exceed that of any homozygote. If such afit homozygote existed it would eventually becomefixed in the population. The mean fitness at equili-brium, W, can be calculated from the three allelemodel (table 8) using the expression:
W = p W11 + p W22 + p W33 + 2P1P2 W12
+ 2pp W13 + 2P2P3 W23,
where Pi, P2 and p are the allele frequencies atequilibrium. The calculated value for mean fitness(W) at the Lap locus is 1148 and this is lowerthan the fitness of the Lap98'198 homozygote (1480,table 7). The model therefore predicts that a pan-mictic population with these relative fitnesseswould lead to fixation of Lap98. Looking again atthe data used to estimate relative fitnesses, thegreatest error is likely to have occurred withestimating the fitness of the Lap9898 genotype.Firstly, the estimate is derived from the mass mat-ing cultures alone; no single families had thisgenotype. Secondly, Lap98198 was expected at alow frequency (0.09) in the mass mating progeny.Small differences between observed and expectednumbers will have a greater effect on percentagedifferences than in genotypes expected at higherfrequencies. The calculated relative fitness for theLap9898 genotype is probably, therefore an over-estimate.
Nevertheless, the analysis suggests that therelative fitnesses of Lap alleles observed in theselaboratory cultures would lead to fixation of Lap98.Because all three alleles are in fact present in localwild populations, the relative fitnesses estimatedfrom progeny reared in the laboratory must bedifferent from those in the wild if the polymorph-ism is indeed maintained by selection.
This is perhaps not a surprising conclusionsince laboratory conditions are obviously verydifferent from the situation in the wild. What isunexpected, however, is the extent of genotypespecific selection at the Lap locus in laboratory
is defined as
A2A3
w23
2P2P3
where

GENETIC STUDIES OF MYTILUS EDULIS. II 175
conditions which would be expected to be rela-tively non-stressful. One source of stress in labora-tory culture may be nutrition. Algal cells areprovided at higher than normal concentrations,but bacteria are very reduced in number and sea-water filtration systems tend also to reduce thelevels of free amino acids available for directuptake by larvae or adults (Manahan, 1983;Manahan and Stephens, 1983). The enzyme codedat the Lap locus, an a-aminoacyl peptide hydro-lase (EC 3.4.11.—) (Young eta!., 1979), is involvedin the cleavage of N-terminal aminoacids frompolypeptides and therefore in the composition ofthe cellular free aminoacid pool (Moore et a!.,1981). It is possible that a reduction in the amountof amino acids available for direct uptake by larvaland juvenile mussels interacts with the cellular freeaminoacid pool and therefore may be affected bygenotype specific differences in biochemicalactivity at the Lap locus.
Other studies of laboratory-reared singlefamilies and mass matings of bivalves have alsoimplicated direct or indirect selection at a numberof enzyme loci (Beaumont eta!., 1983, 1988; Adam-kewicz, 1984; Hvilsom and Theisen, 1984; Malletet a!., 1985, 1986; Gaflney and Scott, 1984). Ourresults suggest that selection at a locus in thelaboratory or in a hatchery condition may be verydifferent from that found in nature and that theextrapolation of genetic data obtained in this wayto the situation in the wild must be undertakenwith caution. It must also be borne in mind thatour data only deal with fitness to the juvenile stageand that changes in relative fitnesses may occur asthe mussels grow. However, where data are avail-able (tables 1 and 2), relative fltnesses of Lapgenotypes generally do not appear to changebetween spat and juvenile sampling within cul-tures, and, since mussels are sedentary andexperience no major ecological changes whilegrowing from juveniles to adults, we would notexpect to observe major changes in relativefitnesses of the Lap alleles during this period.
Further suggestive evidence that Lap genotypespecific mortalities in laboratory conditions maydiffer significantly from those in the wild comesfrom analysis of the overall deficiencies of Lapheterozygotes. In wild populations of M. edu!isand most other bivalves studied, Lap heterozygotesare generally deficient against Hardy-Weinbergexpectations (Koehn et aL, 1976; Skibinski et a!.,1983; Colgan, 1987) and a number of explanationshave been advanced to account for this observation(Zouros and Foltz, 1984; Colgan, 1987). TakingLap genotype data from 37 samples of single family
and mass mating cultures (excluding M4 where allLap genotypes were heterozygous, but includingcultures reared at varying salinities (Beaumont eta!., 1988), 27 samples showed an excess ofheterozygotes while only nine had too fewheterozygotes. According to the null hypothesisthat equal numbers of excesses and deficienciesare expected, the sign test confirms that there is asignificant excess of heterozygotes in these labora-tory cultures. This therefore contrasts with thedeficiencies of Lap heterozygotes displayed bywild populations. However, direct comparisonbetween heterozygote excesses caused by selectionin laboratory conditions and heterozygote deficien-ces in wild populations may not be entirely validsince the latter phenomenon may be caused byfactors other than selection (Zouros and Foltz,1984). Although there is an apparent anomaly atthe Lap locus, overall data from all loci studieddo show a significant deficiency of heterozygotesin samples taken at the juvenile stage (unpublisheddata). Thus the overall trend in laboratory culturesis similar to the situation in the wild and the Laplocus may be unusual in this respect.
Acknowledgements We are grateful to the Natural Environ-ment Research Council who funded this study under Grant no.GR3/5286 to A.R.B. We also thank Dr A. B. Yule for usefuldiscussions and an anonymous referee for helpful criticism ofthe manuscript.
REFERENCES
ADAMKEWICZ, L., TAUB, S. R. AND WALL, J. R. 1984. Geneticsof the clam Mercenaria mercenaria. II. Size and genotype.Malacologia, 25, 525-533.
AHMAD, M. AND HEDRICK, P. W. 1985. Electrophoretic vari-ation in the common mussel, Mytilus edulis: random associ-ation of alleles at different loci. Heredity, 55, 47—51.
BEAUMONT, A. R., BEVERIDGE, C. M. AND BUDD, M. D. 1983.Selection and heterozygosity within single families of themussel, Mytilus edulis (L.) Mar. Biol. Letts., 4, 151—161.
BEAUMONT, A. R., BEVERIDGE, C. M., BARNETT, E. A., BUDD,M. D. AND SMYTH-CHAMOSA, M. D. 1988. Genetic studiesof laboratory reared Mytilus edulis. 1. Genotype specificselection in relation to salinity. Heredity, 61, 389—400.
COLGAN, D. .. 1987. Deficits of heterozygotes in relation toselective interactions between loci in the musselsBra chidontes rostratus and Xenostrobus pulex. Heredity, 59,3 55—362.
CROW, J. F. 1986. Basic Concepts in Population, Quantitativeand Evolutionary Genetics. W. H. Freeman & Company,New York, 273 pp.
DAY, T. I-I., DOBSON, T., HILLIER, P. C., PARKIN, D. T. ANDCLARKE, B. 1982. Associations of enzymic and chromo.somal polymorphisms in the seaweed fly Coelopa frigida.Heredity, 48, 35-44.

176 A. R. BEAUMONT, C. M. BEVERIDGE, E. A. BARNET AND M. D. BUDD
DIXON, D. R.AND CLARKE, B. 1982. Sister chromatid exchange:a sensitive method for detecting damage caused byexposure to environmental mutagens in the chromosomesof adult Mytilus edulis. Mar. Biol. Let is., 3, 163—172.
DIXON, D. K., MCFADZEN, I. R. 15. AND SISLEY, K. 1986.Heterochromatic marker regions (nucleolar organisers) inthe chromosomes of the common mussel Mytilus edulis(Mollusca: Pelecypoda). J. exp. mar. Biol. Ecol., 97, 205—
212.GAFFNEY, P. M. AND SCOTT, T. M. 1984. Genetic heterozygosity
and production traits in natural and hatchery populationsof bivalves. Aquaculture, 42, 289—302.
1-JILBISH, T. J., DEATON, L. E. AND KOEHN, R. K. 1982. Effectof an allozyme polymorphism on regulation of cell volume.Nature, 298, 688—698.
HILBISH, T. J. AND KOEHN, R. K. 1985. Dominance in physio-logical phenotypes and fitness at an enzyme locus. Science,229, 52-54.
HVILSOM, M. M. AND THEISEN, B. F. 1984. Inheritance ofallozyme variations through crossing experiments with theblue mussel, Mytilus edulis L. Hereditas, 101, 1—7.
KOEHN, R. K. 1985. Adaptive aspects of biochemical and phy-siological variability. In Gibbs, P. E. (ed.) Proc. 19th Eur.Mar. Biol. Symp. Cambridge University Press, pp. 425-441.
KOEHN, R. K., MILKMAN, K. AND MIT'rON, J. B. 1976. Popula-lion genetics of marine pelecypods. IV. Selection, migrationand genetic differentiation in the blue mussel Mytilus edulis.Evolution, 30, 2-32.
LEVINTON, .1. S. AND KOEHN, R. K. 1976. Population geneticsof mussels. In Bayne, B. L. (ed.) Marine Mussels, TheirEcology and Physiology, Cambridge University Press, pp.3 57—384.
MALLET, A. L., ZOUROS, F,., GARTNER-KEPKAY, K. E. ANDFREEMAN, K. K. 1986. Genetics of growth in blue mussels:family and enzyme-heterozygosity effects. Mar. Duo!., 92,47 5-482.
MALLET, A. L., ZOUROS, E., GARTNER-KEPKAY, K. E., FREE-MAN, K. R. AND DICKIE, L. M. 1985. Larval viability andheterozygote deficiency in populations of marine bivalves:evidence from pair matings of mussels. Mar. Biol., 87,165— 172.
MANAHAN, D. T. 1983. Nutritional implications of dissolvedorganic material for laboratory culture of pelagic larvae.In Berg, C. J. Jr. (ed.) Culture of Marine Invertebrates.Selected Readings. Hutchinson Ross Pub. Co., Pennsyl-vania, USA, pp. 179-192.
MANAHAN, 0. T. AND STEPHENS, 0. C. 1983. The use of highperformance liquid chromatography to measure dissolvedorganic compounds in bivalve aquaculture systems. Aqua-culture, 32, 339-346.
MOORF., M. N., KOEI-IN, R. K. AND BAYNE, B. L. 1980. Leucineaminopeptidase (aminopeptidase-I), N-acetyl-fJ -hexos-aminidase and Iysosomes in the mussel, Mytilus edulis L.in response to salinity changes. I Exp. Zoo!., 214, 239-249.
SKIBINSKI, 0. 0. F., BEARDMORE. J. A. AND CROSS, T. F. 1983.Aspects of the population genetics of Mytilus (Mytilidae;Mollusca) in the British Isles. Biol. I Linn. Soc., 19, 137—183.
SOKAL, R. R. AND ROULI-, F. .t. 1981. Biometry, 2nd edn. W. H.Freeman and Company, San Francisco, USA, 859 pp.
YOUNG, P. J. W., KOEHN, R. K. AND ARNHEIM, N. 1979. Bio-chemical characterisation of "LAP" a polymorphicaminopeptidase from the blue mussel Mytilus edulis. Bio-chem. Genet., 17, 305—324.
zOUROS, E. AND FOI.TZ, o. w. 1984. Possible explanations ofheterozygote deficiency in bivalve molluscs. Malacologia,25, 583—591.
Related Documents











